Embed Size (px)
Citation preview
Contrôle de l'activité métabolique (2 TSbc )
page 1
CONTRÔLE DE L'ACTIVITÉ ENZYMATIQUE IN-VIVO;notions de régulations métaboliques
La régulation d'une activité enzymatique peut s'expliquer par des phénomènes liés aux loisde la chimie ( loi d'action de masse, déplacement d'équilibre, compétitions d'enzyme au niveau d'unsubstrat commun) mais ces régulations n'ont pas alors une grande souplesse d'adaptation et nepermettent pas d'expliquer certains phénomènes comme:
- Toutes les enzymes ne fonctionnent pas en même temps dans une cellule- Les réactions enzymatiques sont réglées en fonctions des besoins de la cellule
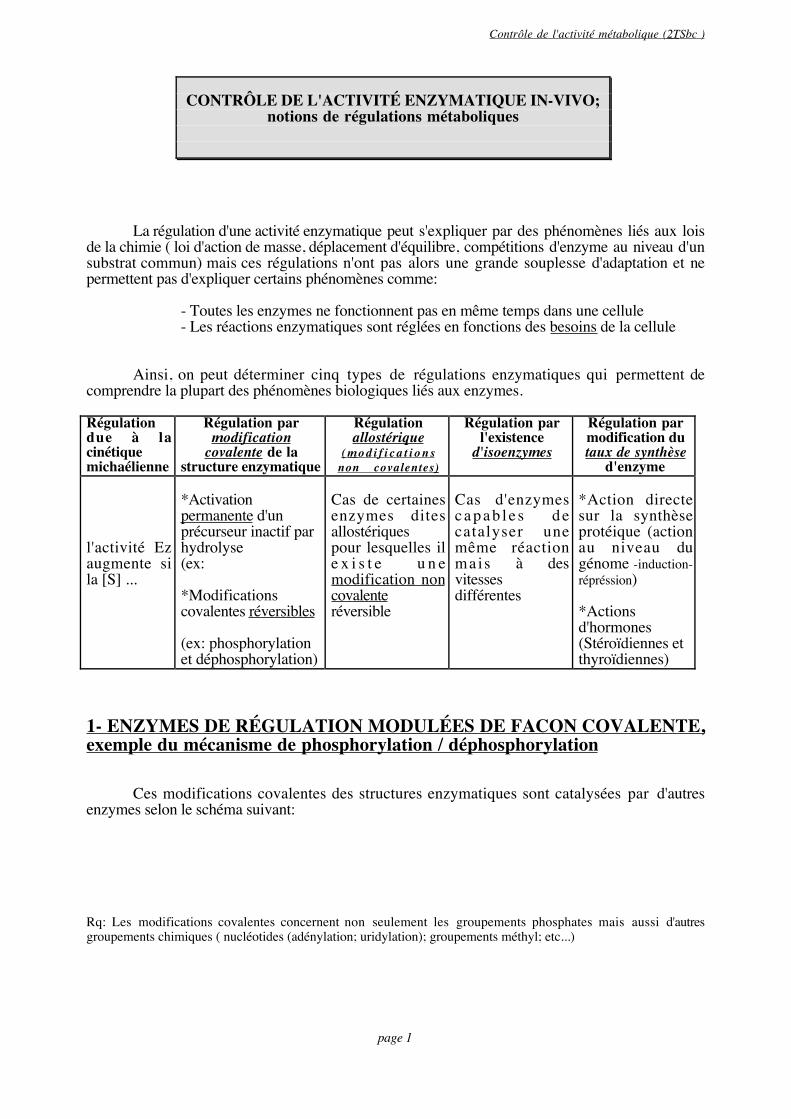
Ainsi, on peut déterminer cinq types de régulations enzymatiques qui permettent decomprendre la plupart des phénomènes biologiques liés aux enzymes.
Régulationdue à lacinétiquemichaélienne
Régulation parmodification
covalente de lastructure enzymatique
Régulationallostérique
( m o d i f i c a t i o n snon covalentes)
Régulation parl'existence
d'isoenzymes
Régulation parmodification dutaux de synthèse
d'enzyme
l'activité Ezaugmente sila [S] ...
*Activationpermanente d'unprécurseur inactif parhydrolyse(ex:
*Modificationscovalentes réversibles
(ex: phosphorylationet déphosphorylation)
Cas de certainesenzymes ditesallostériquespour lesquelles ile x i s t e u n emodification noncovalenteréversible
Cas d'enzymesc a p a b l e s d ecatalyser unemême réactionmais à desvitessesdifférentes
*Action directesur la synthèseprotéique (actionau niveau dugénome -induction-répréssion)
*Actionsd'hormones(Stéroïdiennes etthyroïdiennes)
1- ENZYMES DE RÉGULATION MODULÉES DE FACON COVALENTE,exemple du mécanisme de phosphorylation / déphosphorylation
Ces modifications covalentes des structures enzymatiques sont catalysées par d'autresenzymes selon le schéma suivant:
Rq: Les modifications covalentes concernent non seulement les groupements phosphates mais aussi d'autresgroupements chimiques ( nucléotides (adénylation; uridylation); groupements méthyl; etc...)

Contrôle de l'activité métabolique (2 TSbc )
page 2
1-1- L'exemple type du mécanisme de phosphorylation/déphosphorylation : la glycogènephosphorylase:
Cette enzyme est présente dans certains tissus animaux (muscle, foie) et catalysel'hydrolyse d'un polyoside de réserve: le glycogène pour former le glucose 1 phosphate qui pourraêtre utilisé par la cellule en entrant dans les réactions de la glycolyse.
La glycogène phosphorylase existe sous deux formes:- phosphorylase a (active)- phosphorylase b (inactive)
La phosphorylase a est un oligomère constitué par deux sous unités dimériques et chaquesous-unité est phosphorylée. On peut enlever les groupements phosphates par hydrolyse, cetteréaction est catalysée par une enzyme: la phosphorylase phosphatase et permet donc de former laphosphorylase b.
La réaction inverse est catalysée par une autre enzyme: la phosphorylase kinase.
Pour résumer les modifications covalentes on peut écrire:
Rq: l'action de la kinase est stimulée par deux hormones hyperglycémiantes: l'adrénaline et leglucagon
l'action de la phosphorylase est stimulée par une hormone hypoglycémiante: ....
+ La propriété essentielle de ce type d'enzyme est de pouvoir amplifier un signalchimique.

Contrôle de l'activité métabolique (2 TSbc )
page 3
1-2-L'amplification du signal de phosphorylation/déphosphorylation:
Toutes les enzymes peuvent apporter une amplification, c'est à dire une molécule d'enzymepeut catalyser la formation de milliers de molécules de produit à partir d'un substrat donné.Dans le cas de ces enzymes de régulation l'amplification est d'autant plus importante que desphénomènes hormonaux entrent en jeu.
La fixation d'une hormone (Adrénaline, glucagon) sur son récepteur membranaire, (respectivementmuscle et foie), déclenche une cascade d'évènements qui permettent d'amplifier le phénomène pouraboutir à l'activation massive de glycogène phosphorylase, ce qui entraîne donc la formation deglucose 1 phosphate en grande quantité.Ces phénomènes font intervenir un 2ème messager: l'AMP cyclique (voir document et voir cours debiologie cellulaire )
HORMONE 10-10 M
enzyme active
Kinase
AMPc 10-6 M
Adénylate cyclase
Produit(ex: Glucose 1 P)

Contrôle de l'activité métabolique (2 TSbc )
page 4
Rq: l'association kinase/phosphatase est une association très étroite !article Biofutur juillet/août 1998
2- REGULATION PAR TRANSITION ALLOSTERIQUES: LES ENZYMESALLOSTERIQUES;
2.1- Notion de rétro-inhibition:La rétro inhibition feedback négatif ou inhibition par le produit final) a été mis en évidence dans
de nombreux systèmes poly enzymatiques où le produit final de la réaction peut inhiberspécifiquement une enzyme se trouvant au début de la séquence réactionnelle.
ex: dans les séquences réactionnelles de synthèse d'un aminoacide , l'isoleucine à partir dela thréonine.
La première étape de la séquence réactionnelle est appelée étape déclenchante car une foisqu'elle a lieu toutes les autres se réalisent.Ce mécanisme permet d'assurer une économie en métabolites (si [P] ⁄, la vitesse diminue)
+ La rétroinhibition est donc un phénomène d'inhibition spécifique d'une enzymeplacée en début de séquence métabolique, par le produit final de séquence réactionnelle.
Ce type d'enzyme inhibé par le produit final de la réaction est appelé :....
Caractéristiques d'une enzyme allostérique:
- est inhibée uniquement par le produit final de la réaction- le produit final n'agit pas comme un inhibiteur compétitif (structure du produit final très
différente de celle du substrat habituel de l'enzyme allostérique) il y a alors un effet allostérique (terme proposé par Monod, Jacob, Changeux).Cela signifie que l'action de l'inhibiteur sur une telle enzyme agit en dehors du site catalytique
- la cinétique enzymatique n'est pas de type cinétique Michaélienne
Définitions possibles d'une enzyme allostérique:

Contrôle de l'activité métabolique (2 TSbc )
page 5
2.2- Caractéristiques structurales et fonctionnelles des enzymes allostériquesLes enzymes allostériques ont en général un poids moléculaire plus élevé et sont plus
complexes et plus difficiles à purifier que les autres enzymes (allostérie= autre structure)
• les enzymes allostériques sont des ....
• les enzymes allostériques existent sous deux états conformationnel:
• il existe sur chaque protomère des sites de fixation pour les effecteurs (sitesallostériques).Si l'effecteur déplace l'équilibre en faveur de la conformation qui a la plus grande affinité pour lesubstrat on parle d'activateur allostérique ( coopérativité positive )Dans le cas inverse on parle d'inhibiteur allostérique ( coopérativité ..... )
L'effecteur allostérique se fixe de manière:
++
La fixation au site allostérique entraîne une légère modification de structure, c'est la transitionallostérique
2.3- La cinétique des enzymes allostériques: une cinétique non michaelienne* Étude d'une enzyme allostérique: l'isocitrate déshydrogénase:
Réaction:
• Étude la courbe vi = f([S]) (courbe A)vi
[S]Conclusion:
• Étude la courbe de Lineweaver-Burk

Contrôle de l'activité métabolique (2 TSbc )
page 6
1/vi
1/[S]
• Étude de la courbe vi = f([S]) en présence d'ADP à concentration élevée (courbe B)
*Cas général : coopérativité et modulation par les effecteurs allostériques
1- Coopérativité ou modulation par le substrat:
- Coopérativité positive: cinétique non michaélienne, courbe sigmoïde. La fixationde la 1°molécule de S facilite la fixation des autres. Une légère augmentation de [S] peutaugmenter très rapidement la vitesse. On considère dans ce cas que le substrat joue le rôle à la fois de substrat et d'effecteur allostériquepositif
- Coopérativité négative: on obtient une courbe plutôt aplatie
2- Modulation par les effecteurs allostériques:
La cinétique allostérique ne définit pas les termes Vm et Km comme en cinétiquemichaélienne. Les termes Vm et Km en cinétique allostérique doivent êtres envisagés avecprudence, ce sont en fait des constantes apparentes qui permettent de répartir les enzymes en diversgroupes:
- Enzymes du système V ( l'effecteur ne modifie que Vm)- Enzymes du système K ( l'effecteur ne modifie que l'affinité apparente de l'enzyme pour
le substrat, soit Km)- Enzymes du système mixte (l'effecteur modifie le deux paramètre)
2-1- Action d'effecteurs négatifs les inhibiteurs allostériques sont les .....d'une enzyme allostérique impliquée dans une séquence réactionnelle.
L'inhibiteur se fixe sur une enzyme allostériqueil s'ensuit une transition allostérique modifiant laconformation du site actif.
ENZYMES DU SYSTÈME K ENZYMES DU SYSTÈME V

Contrôle de l'activité métabolique (2 TSbc )
page 7
vi
[S]
vi
[S]
2-2- Action d'effecteurs positifs: l'enzyme allostérique qui régule une séquence métabolique estfréquemment activée par le produit final d'une autre séquence indépendante et parallèle.Lorsque l'activateur allostérique se fixe sur le site allostérique, il s'ensuit un changement deconformation permettant une liaison facilitée du substrat sur son site enzymatique.
Schéma:
ENZYMES DU SYSTÈME K ENZYMES DU SYSTÈME Vvi
[S]La transition allostérique provoque une modification dusite actif qui devient plus propice à la fixation de S, ona donc une augmentation de l'affinité de E pour S(baisse de Km sans modification de Vm)
vi
[S]L'activateur induit une augmentation de Vm sansmodifier le Km
La plupart des enzymes allostériques sont susceptibles d'être régulés par plusieurs effecteurs. La régulationla plus souvent rencontrée comprend à la fois
- l'activation par le substrat (phénomène de .....- l'inhibition par le ....- l'activation en parallèle
RQ: lorsque l'effecteur d'une enzyme allostérique est également son substrat, on parle d'effecteur homotrope. On parle d'enzyme allostérique homotrope quand elles sont régulés par le substrat.
Si l'effecteur n'est pas le substrat, il s'agit d'effecteur hétérotrope. L'enzyme hétérotope est modulé parun effecteur autre que le substrat qui agit donc sur un site différent.

Contrôle de l'activité métabolique (2 TSbc )
page 8
3- LES ISOENZYMES : plusieurs protéines aux activités enzymatiquesidentiques
Un autre mécanisme de régulation de l'activité métabolique fait intervenir les isoenzymes.
Définition: Les isoenzymes sont des formes moléculaires multiples existant chez une espècedonnée ou même dans une cellule donnée. Ces formes différentes d'une même enzyme s edistinguent par la composition en acides aminés et donc par leur pHi .
L'exemple historique encore d'actualité: la lactate déshydrogénase
La découverte des isoenzymes s'est faite après une électrophorèse de la LDH de lapin:
Conclusion:
5 bandes
pôle - M4
M3H
MH3H4
M2H2
pôle +
Répartition organique de la LDH:
M4 M3H M2H2 MH3 H4coeur + + + + + + + ++++++rein + + + + + + + + + + + + +
hématies + + + + + + + +cerveau + + + + + + + + + + + +
leucocytes + + + + + + + +muscle + + + + + + + + + + +
foie + + + + + + + + + +
Les isoenzymes de la LDH possèdent comme propriétés:- même masse molaire (134000)- 4 chaînes polypeptidiques (enzyme ...- constituent 5 combinaisons différentes- individuellement les chaînes M et H n'ont aucune activité ....
( Ces chaînes sont codées par deux gènes différents)- catalysent la ... réaction mais diffèrent par leur constantes cinétiques.
Réaction catalysée:
• L'isoenzyme H4 (=LDH1) possède par rapport à M4 (=LDH5):- un Km plus élevé pour le pyruvate et un Vm plus faible pour la réduction de ce dernier
cela signifie que: ....

Contrôle de l'activité métabolique (2 TSbc )
page 9
Quel est l'intérêt de ces formes isoenzymatiques ?
Il faut raisonner à l'échelle de l'organisme; sachant que le pyruvate est une molécule dumétabolisme qui peut avoir plusieurs destinées:
- soit il est dégradé en aérobie et entre alors dans le cycle du Citrate (= cycle de Krebs)- soit il est utilisé en absence d'oxygène et entre dans la voie fermentative (fermentation
lactique)
• Au niveau du muscle: il utilise le glucose en anaérobiose, il décompose donc le pyruvateen lactate grâce à la LDH
L'AFFINITÉ de l'enzyme (LDH 5 = ) pour le pyruvate doit être .....
• Au niveau du coeur: le coeur est un muscle particulièrement bien oxygéné, il est moinsintéressé par la fermentation lactique que les autres muscles.Ceci explique la faible affinité de la LDH1 pour le pyruvate à ce niveau.
Rq: - il existe de nombreuses isoenzymes, on peut citer la Créatine phosphokinase (CPK),l'a-amylase (pancréas et salive),les transaminases , etc..
- les isoenzymes ont un grand intérêt clinique, exemple de la LDH dans le diagnosticmédical.Le taux de LDH sérique est normalement faible mais au cours de certaines maladies (au niveau dufoie ou du coeur) les tissus lésés libèrent entre autre cette enzyme dans le sang où la concentrationva augmenter.
Contrôle de l'activité métabolique (2 TSbc )
page 10
Document: Document: à propos de la cinétique des enzymes allostériques ! à propos de la cinétique des enzymes allostériques !
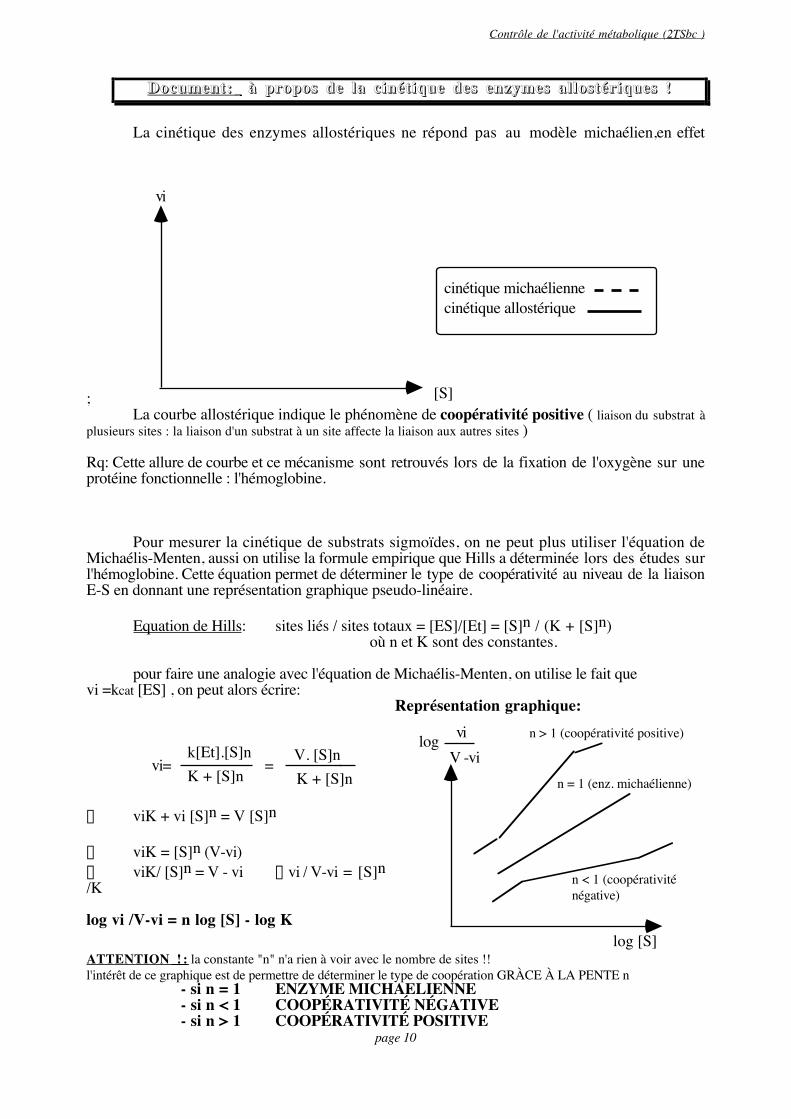
La cinétique des enzymes allostériques ne répond pas au modèle michaélien,en effet
;
vi
[S]
cinétique michaéliennecinétique allostérique
La courbe allostérique indique le phénomène de coopérativité positive ( liaison du substrat àplusieurs sites : la liaison d'un substrat à un site affecte la liaison aux autres sites )
Rq: Cette allure de courbe et ce mécanisme sont retrouvés lors de la fixation de l'oxygène sur uneprotéine fonctionnelle : l'hémoglobine.
Pour mesurer la cinétique de substrats sigmoïdes, on ne peut plus utiliser l'équation deMichaélis-Menten, aussi on utilise la formule empirique que Hills a déterminée lors des études surl'hémoglobine. Cette équation permet de déterminer le type de coopérativité au niveau de la liaisonE-S en donnant une représentation graphique pseudo-linéaire.
Equation de Hills: sites liés / sites totaux = [ES]/[Et] = [S]n / (K + [S]n)où n et K sont des constantes.
pour faire une analogie avec l'équation de Michaélis-Menten, on utilise le fait quevi =kcat [ES] , on peut alors écrire:
vi=k[Et].[S]nK + [S]n = V. [S]n
K + [S]n
Ò viK + vi [S]n = V [S]n
Ò viK = [S]n (V-vi)Ò viK/ [S]n = V - vi Òvi / V-vi = [S]n/K
log vi /V-vi = n log [S] - log K
Représentation graphique:
log viV -vi
log [S]
n > 1 (coopérativité positive)
n = 1 (enz. michaélienne)
n < 1 (coopérativité négative)
ATTENTION ! : la constante "n" n'a rien à voir avec le nombre de sites !!l'intérêt de ce graphique est de permettre de déterminer le type de coopération GRÀCE À LA PENTE n
- si n = 1 ENZYME MICHAELIENNE- si n < 1 COOPÉRATIVITÉ NÉGATIVE- si n > 1 COOPÉRATIVITÉ POSITIVE