-
2
EETTUUDDEE BBIIBBLLIIOOGGRRAAPPHHIIQQUUEE
-
3
I. GENERALITES SUR LES TIQUES DES BOVINS.
I.1. Classification et biologie des tiques.
I.1.1. Classification (Camicas & Morel, 1977 ; Morel,
2000).
Les tiques sont des arthropodes appartenant la classe des
Arachnides et la sous-classe des Acariens ; celles qui parasitent
les bovins sont des tiques dures (sous-ordre des Ixodina), par
opposition aux tiques molles (sous-ordre des Argasina) (figure
1).
Leurs diffrences morphologiques avec les autres Acariens sont
:
la prsence dun rostre ou hypostome provenant de la runion de
deux pices antroventrales de la basis en un organe unique, dont la
longueur varie selon les genres et constitue lun des lments
essentiels la diagnose de lespce : lorsque le rostre est nettement
plus long que large on parle de tique longirostre, lorsquil est
plus ramass on parle de tique brvirostre, sa base, de forme
variable selon lespce, est appele capitulum ;
leur grande taille, par rapport aux Acariens en gnral (les
adultes jeun
mesurent de 1,5 15 mm) ;
lexistence dune cuticule souple extensible et susceptible de
croissance lors de la rpltion, en relation avec le comportement
alimentaire trs volu.
Figure 1 : Systmatique des tiques daprs Camicas & Morel
(1977).
Dermacentor
embranchement
sous-embranchement
classe
sous-classe
superordre
sous-ordre
ordre
superfamille
famille
tribu
genre
Arthropodes
Chlicrates
Arachnides
Acariens
Anactinatrichoida
Ixodida
Ixodina (tiques dures)
Ixodoidea
Ixodidae (prostriata)
Amblyommidae (metastriata)
Haemaphysalini
Haemaphysalis
Amblyommini
Amblyomma
Rhipicephalini
Hyalomma Rhipicephalus
Dermacentorini Boophilini
Boophilus
Ixodinae
Ixodes
Argasina (tiques molles)
-
4
I.1.2. Cycles biologiques.
Comme pour tous les Acariens, le cycle biologique des tiques
comporte quatre tapes volutives :
- luf ; - la larve (hexapode) ; - les stades nymphaux
(protonymphe, deutonymphe inconstante, tritonymphe) ; - ladulte
(mle ou femelle).
Chez les Ixodiformes, on appelle stases les diffrentes formes
spares par des mtamorphoses vraies et stades, les diffrentes formes
spares par une mue de croissance. Mais les Ixodids, qui sont
hmatophages, ne prennent quun unique repas de sang chaque stase, ce
dernier tant ncessaire lvolution et la ponte des ufs, aussi stases
et stades sont-ils synonymes (Prez-Eid & Gilot, 1998).
Les tiques tant des ectoparasites intermittents, il existe trois
types de cycles en fonction
du nombre dhtes intervenant (Morel, 2000) (figure 2) :
les cycles trixnes (ou triphasiques), o il y a un changement
dhte entre chaque stase, sont les plus frquemment rencontrs chez
les espces parasitant nos ruminants : il y a alors trois phases
parasitaires spares par deux phases terre, o se passent les
pupaisons ; les tiques passent la majeure partie de leur vie dans
lenvironnement et les facteurs climatiques entranent lalternance de
priodes dactivit et de diapause ;
les cycles dixnes (ou diphasiques), o les trois stases voluent
sur deux htes
individuellement diffrents : dans la premire phase, la larve
gorge mue sur lhte et la nymphe qui en provient se refixe proximit
; par la suite, la pupaison nymphale a lieu sur le sol et les
adultes se fixent un nouvel hte ;
les cycles monoxnes (ou monophasiques), o toutes les stases se
succdent
sur un unique vertbr abord par la larve, sont rares : il ny a
qune phase parasitaire et seuls la ponte, lincubation et les
dplacements des larves en qute dun hte se passent sur le sol, la
dure du cycle sen voit ainsi raccourcie. Ce type de cycle est
laboutissement dune slection adapte des conditions microclimatiques
difficiles (Morel, 1979).
Ces cycles sont schmatiss sur la figure 2.
De plus, la slectivit des tiques envers leurs htes est variable
et suivant la similitude ou la diffrence des tropismes manifests
aux diverses stases, on rencontre trois types de cycles (Bourdeau,
1993a ; Morel, 2000) :
les cycles monotropes rsultent dune mme slectivit dans le choix
de lhte
toutes les stases ; les cycles ditropes concernent les tiques
dont la slectivit des primagos est
diffrente (plutt des petits mammifres, oiseaux ou reptiles) de
celle des adultes (plutt des grands mammifres) ;
les cycles tlotropes voient les primagos se gorger sur les
vertbrs disponibles
(ils sont ubiquistes) tandis que les adultes se gorgent plutt
sur les grands mammifres (ils sont slectifs).
-
5
L = Larve A = Adulte = Mtamorphose N = Nymphe * = Stase
gorge
Figure 2 : Types de cycles en fonction du nombre dhtes
intervenant, daprs Mehlhorn & Walldorf (1988) et Collectif,
Ecole Nationale Vtrinaire de Nantes
(2000).
Hte 3
Ponte L uf Environnement
*
Hte unique
Ponte* AN*L
Hte 2Hte 1
uf
Environnement
L L* N N* + *
uf L L* N N* * A
Hte 1 Hte 2
Environnement
Ponte
Cycle trixne :
L - L* N - N* + *
Cycle dixne :
L - L* N - N* + *
Cycle monoxne :
-
6
I.1.3. Particularits biologiques.
a) La vie libre.
La vie libre des tiques est fortement lie aux facteurs
climatiques : la temprature est le
facteur dynamique essentiel dorganogense et dactivit tandis que
lhumidit est un important facteur de survie qui caractrise le
biotope. Aussi, concernant le milieu de vie, les espces de tiques
se divisent-elles en deux groupes (Morel, 2000) :
les tiques endophiles (ou pholophiles) vivent dans des habitats
trs spcialiss
en raison de conditions microclimatiques qui y rgnent, elles
infestent lhte dans son gte (terriers, nids), sy reproduisent et se
dplacent peu ;
les tiques exophiles nont pas dhabitat aussi spcialis, lhte est
infest suite un afft sur la vgtation et selon le terrain et la
vgtation, on distingue des tiques xrophiles, hygrophiles,
thermophiles
Notons ce sujet, que de nombreuses espces de tiques sont la fois
endophiles aux
stases larvaire et nymphale et exophiles la stase adulte
(Levasseur, 1993). La phase de vie libre est riche en facteurs de
rgulation des populations de tiques : la
temprature et lhygromtrie tant les principaux (Prez &
Rodhain, 1977). A ces facteurs climatiques ou abiotiques rendant la
dure du cycle fort variable, viennent sajouter les prdateurs (avec
parmi les vertbrs : les oiseaux principalement, les herbivores et
les rongeurs et parmi les arthropodes : les araignes, les fourmis
et larves de Lpidoptres ) et les hyperparasites (protozoaires,
bactries) dont le rle reste difficile estimer (Milne, 1949 ; Morel,
1974).
b) La vie parasitaire. Les tiques tant hmatophages, elles se
fixent en gnral sur les zones peau fine de
lhte. La fixation procde dabord dune action mcanique :
lhypostome senfonce peu peu dans leffraction cutane dtermine par
les mouvements des chlicres et de leurs crochets. La seconde phase
est la scrtion du cment : la scrtion salivaire se solidifie en
manchon. Cette substance permet une fixation trs solide de la
tique. La prise de sang se fait ensuite de manire indirecte dans la
poche hmorragique rsultant de la rupture de capillaires lors de la
pntration des pices buccales dans lpiderme (acariens telmophages)
(Prez-Eid & Gilot, 1998). Le repas de sang comporte deux phases
essentielles, surtout marques chez les femelles : une phase de
gorgement lent et progressif, au cours de laquelle les femelles
sont fcondes, puis une phase rapide qui dure gnralement de un trois
jours, au cours de laquelle la tique grossit beaucoup (Bourdeau,
1993a).
Concernant les mles adultes, ceux des Ixodids ne se fixent pas
et ne prennent pas de
repas de sang, tandis que ceux des Amblyommids prennent trs peu
de sang. Laccouplement des adultes a lieu parfois au sol (certains
Ixodids), le plus souvent sur lhte : il a alors lieu pendant le
gorgement grce aux phromones dagrgation et sexuelles produites au
cours du repas sanguin (Morel, 2000). La femelle peut commencer son
repas avant dtre fconde mais ne le termine quaprs fcondation (pause
prprandiale) (Prez-Eid & Gilot, 1998 ; Morel, 2000).
-
7
I.2. Actions pathognes des tiques chez les bovins.
Les scrtions salivaires des tiques jouent un rle majeur dans
leurs actions pathognes du fait de la transmission dagents
pathognes ou de linjection de composants toxiques pouvant affecter
le mtabolisme gnral ou causer des paralysies (Cupp, 1991).
Ainsi nous distinguons des actions pathognes directes et des
actions indirectes.
I.2.1. Action pathogne directe.
Laction phlogogne et mcanique (Morel, 2000). Il sagit dune
raction gouverne par des mcanismes inflammatoires non
spcifiques
au cours de la premire infestation. La poche de lyse lextrmit du
manchon de cment est entoure dune zone ddme, o les structures
cellulaires disparaissent peu peu et o se produisent des
extravasations et des ruptures de capillaires. La fixation simple
est prurigineuse, voire douloureuse par tiraillement des tissus
lss. Aprs le dpart de la tique, la lsion ncrotique demeure indure,
prurigineuse et chaude et peut exsuder pendant plusieurs mois. Des
complications bactriennes sont possibles par abcdation. Au cours
des rinfestations, il existe souvent des phnomnes dhypersensibilit
(notamment basophile). Au fur et mesure, un certain degr de
rsistance de lhte peut mme stablir, les ractions tissulaires sont
alors plus violentes et plus prcoces, avec un dme considrable sans
rupture de capillaires, cela peut ainsi limiter la charge
parasitaire.
Le rle anmiant (Prez-Eid & Gilot, 1998). Il affecte les
animaux qui sont porteurs de plusieurs dizaines, voire centaines de
tiques.
Les tiques ont laptitude concentrer le sang au fur et mesure
quelles le prlvent : on considre que le volume prlev est au moins
le triple de celui que traduit le volume des tiques en fin de
repas. Ces spoliations sont majores par laction anticoagulante des
scrtions salivaires et peuvent engendrer des anmies graves, surtout
en zones tropicales.
Le rle toxique (Prez-Eid & Gilot, 1998). Des toxines
slectives vis--vis du tissu nerveux, issues de femelles en
cours
dovogense (cas dcrits dIxodes ricinus chez des moutons dEurope
occidentale), entranent des paralysies lorsque la fixation a lieu
prs dun rameau nerveux. Citons galement les cas de dishydroses
rencontres chez les ruminants dAfrique Australe, parasits par
Hyalomma truncatum.
I.2.2. Action pathogne indirecte : le rle de vecteur.
Les tiques sont des arthropodes hmatophages haut pouvoir
vectoriel : des agents pathognes peuvent tre acquis durant les
divers stades du cycle de vie, tre ensuite transmis de manire
trans-ovarienne (de la femelle sa progniture) et trans-stasiale
(entre les diffrentes stases) pour enfin se retrouver chez un hte
sensible (Cupp, 1991).
Selon la maladie, elles jouent alors un simple rle de vecteur,
ou sont, en plus, le
rservoir dagents pathognes (Tissot-Dupont, 1998). Un lment
galement capital pour le transfert des germes est le partage
trophique entre htes domestiques et htes sauvages (Gilot et al.,
1987).
Nous nvoquerons ici que des agents pathognes transmis par les
tiques des bovins et
prsents en Europe tempre.
-
8
I.2.2.1. Les hmoparasites des bovins.
Les hmoparasites transmis par les tiques aux bovins en Europe
sont des protozoaires appartenant aux genres Babesia et Theileria.
Ils sont responsables de manifestations cliniques dans le cadre dun
syndrome hmolytique avec hyperthermie. La piroplasmose bovine la
plus cosmopolite en France est transmise par Ixodes ricinus et due
Babesia divergens. La maladie a en outre t dcrite chez l'Homme, en
cas d'immunodpression (notamment chez des individus splnectomiss)
(Gorenflot et al., 1998). Les autres hmoparasites, les espces de
tiques vectrices et la localisation gographique sont dtaills en
annexe I : il sagit de Babesia bigemina, Babesia bovis, Babesia
major, Theileria annulata et le complexe Theileria orientalis
(Euzby & Rancien, 1966 ; Brocklesby & Barnett, 1972 ;
Euzby, 1979 ; Joyner & Donnely, 1979 ; Malbert et al., 1988 ;
Gray, 1991 ; Bourdeau, 1993b ; Elfassy, 1993 ; Uilenberg, 1995 ;
Hillyard, 1996 ; Prez-Eid & Gilot, 1998 ; LHostis &
Seegers, 2002 ; LHostis, comm. pers.).
I.2.2.2. Les infections bactriennes.
Parmi les infections bactriennes transmises aux bovins par les
tiques, citons les rickettsioses (bactries de lordre des
Rickettsiales) comme lanaplasmose (Anaplasma marginale),
lehrlichiose (Anaplasma phagocytophilum, anciennement nomm
Ehrlichia phagocytophila, avant le changement de nomenclature en
2001) et la fivre Q (Coxiella burneti) (Collin, 2003 ; Joncour,
2003 ; Liz, 2003). LEhrlichiose et la fivre Q sont dailleurs des
zoonoses (Brouqui, 2003), tout comme la maladie de Lyme (Borrelia
burgdorferi, afzelii & garini en Europe) et la tularmie
(Francisella tularensis) qui peuvent tre transmises lHomme par
certaines tiques infestant communment les bovins. Les dtails
concernant ces agents pathognes sont retranscrits en annexe I
(Macaigne & Prez-Eid, 1991 ; Hillyard, 1996 ; Prez-Eid &
Gilot, 1998 ; Tissot-Dupont, 1998 ; Mage et al., 2001 ; Collin,
2003 ; Joncour, 2003 ; Liz, 2003).
I.2.2.3. Les arbovirus.
Les ruminants sont galement sensibles certains virus transmis
par les tiques comme les agents du louping-ill entranant une
encphalomylite chez le mouton ou dencphalites tique, qui sont aussi
les agents de zoonoses. Pour plus de dtails, se rfrer lannexe I
(Hillyard, 1996 ; Prez-Eid & Gilot, 1998 ; Tissot-Dupont,
1998).
-
9
II. CONNAISSANCES SUR LES ESPECES DE TIQUES DES BOVINS PRESENTES
EN FRANCE.
Parmi les centaines despces dIxodidae existantes, seules
quelques unes parasitent communment les bovins. Mais nous
nvoquerons ici que celles qui sont rpertories en France, dont voici
la liste (Morel, 1981 ; Soulsby, 1982, Bourdeau, 1993a ; Hillyard,
1996) :
Ixodes ricinus ; Dermacentor marginatus & Dermacentor
reticulatus ; Rhipicephalus bursa, Rhipicephalus sanguineus &
Rhipicephalus turanicus ; Haemaphysalis punctata, Haemaphysalis
concinna & Haemaphysalis inermis ; Hyalomma detritum scupense,
Hyalomma marginatum marginatum.
En gras, les espces autochtones des bovins, a priori les plus
frquentes.
Des cls de diagnose permettent de prciser le genre : cf. annexe
III.
II.1. Ixodes ricinus, Linn, 1758.
Il sagit de la tique des bovins la plus frquente et la plus
tudie en France. Elle est prsente quasiment sur lensemble du
territoire franais et affectionne particulirement les bovins.
II.1.1. Caractristiques morphologiques (Hillyard, 1996).
Les lments importants de la diagnose dIxodes ricinus sont ports
en annexe IVa.
Il sagit dune espce longirostre avec un sillon anal
semi-circulaire, en avant de lanus
(prostriata). Le bord postrieur du corps nest pas festonn.
Lcusson dorsal chitinis (ou scutum) est de forme hexagonale et ne
prsente ni
ornementation, ni yeux (ou ocelles). La hanche I possde une
longue pine externe, visible sur la face ventrale.
La terminologie morphologique utilise dans le texte est
rcapitule en annexe IIa & IIb.
II.1.2. Cycle biologique dIxodes ricinus.
II.1.2.1. Cycle volutif.
a) Les diffrentes tapes.
Ixodes ricinus ralise un cycle triphasique (figure 2), tlotrope
(Euzby & Rancien,
1966). Larves, nymphes et adultes se gorgent chacun une fois sur
un hte diffrent. Lensemble du cycle est schmatis sur la figure
3.
-
10
Figure 3 : Cycle dIxodes ricinus daprs Mehlhorn & Walldorf,
1988.
-
11
La copulation des adultes se produit le plus souvent sur lhte :
les mles recherchent les femelles sans faire de repas sanguin.
Pourtant, Graf (1974) a rcolt un grand nombre de femelles fcondes
dans la nature. En fait, des phromones sexuelles mises par les
femelles semblent attirer les mles et ainsi expliquer la copulation
dans lenvironnement (Graf, 1975). Quoi quil en soit, les femelles
ne peuvent pas se gorger compltement sans intervention de la
copulation (Graf, 1974). Une fois fcondes, les femelles terminent
leur repas sanguin, puis se dtachent, tombent sur le sol et pondent
leurs ufs (de 500 2000) avant de mourir, au terme dune priode de
prponte (digestion du sang et maturation des ovules) pouvant
atteindre deux mois. Les mouvements latraux de la femelle libre
sont trs restreints, et lorsquelle se laisse tomber sur un tapis
vgtal non protecteur vis--vis de la dessiccation, elle peut mourir
avant la ponte (Milne, 1946). La priode de ponte stale en gnral de
mai juillet sous nos latitudes et lclosion des larves prend six
huit semaines (Euzby & Rancien, 1966 ; Prez & Rodhain,
1977).
Aprs son closion, la larve ne se met en qute dun hte
(micromammifre) quau bout
de deux semaines, temps ncessaire pour durcir son tgument et
perdre une certaine quantit deau (Prez & Rodhain, 1977). Aprs
un repas de trois sept jours, elle ne se laisse pas tomber au
hasard, mais prfrentiellement dans le terrier de son hte. Puis,
grce un phototropisme positif, la mue larvaire a lieu lextrieur et
donne une nymphe exophile (Graf et al., 1978). De mme, la nymphe
recherche un hte (stase ubiquiste) sur lequel elle effectue son
repas sanguin. Aprs ce repas, elle tombe sur le sol pour muer en
adulte.
b) La dure des diffrentes tapes. La dure du cycle est variable
et dpend surtout de la temprature. Les dures des
diffrentes tapes, obtenues par diffrents auteurs et relates dans
le tableau I, ci-dessous, montrent lexistence dun cycle court
temprature tempre (15 20C environ) et dun cycle long temprature
plus basse (Aeschlimann, 1972).
Tableau I : Dure des diffrentes tapes du cycle dIxodes
ricinus
daprs Aeschlimann, 1972.
Pomerantzev (in Aeschlimann, 1972) Aeschlimann
(1972) Brossard
(in Aeschlimann, 1972) Cycle court Cycle long Cycle court
(17-21C) Cycle court (Dures en jours)
Proviposition 4 27 3 22 8 17 Embryognse 25 400 Environ 30 31 47
22C
Prnutrition larvaire 10 570 16 21 Repas larvaire 3 6 5 2 3
Postnutrition larvaire 28 426 30 55 21 25 (Mue) 26C
Prnutrition nymphale 10 540 24 72 21 Repas nymphal 3 6 4 7 2
6
Postnutrition nymphale 56 360 Jusqu 100 24 37 (Mue)
Prnutrition de la 10 810 30 21 26CRepas de la 6 14 6 14 6 11
Dans des conditions de temprature et dhygromtrie dfavorables, le
cycle peut subir
de forts ralentissements et certains auteurs parlent mme de
diapause. Mme si tous les stades dIxodes ricinus sont exophiles, il
leur arrive de devenir endophile et de se retirer dans les
anfractuosits du sol, lors de priodes dfavorables. Ces adaptations
permettent notamment Ixodes ricinus de passer sans dommage les mois
dhiver (Aeschlimann, 1972).
-
12
Quoi quil en soit, le dveloppement de cette espce est trs lent
et exige, selon les latitudes et les conditions climatiques, de
deux six annes pour saccomplir (Gardiner et al., 1981). En France,
ce dveloppement seffectue en moyenne en trois ans, soit un an par
stase (Euzby & Rancien, 1966).
Au bilan de ce cycle biologique, nous retiendrons que la priode
de vie parasitaire est
trs courte par rapport la longueur de la vie libre dans le
milieu extrieur. Les 18 jours passs sur lhte sont bien peu au
regard des 500 1000 jours dans lenvironnement (Milne, 1950 ; Lees
& Milne, 1951). Cela implique une grande rsistance au jene
(plus dun an), au froid (survie des individus jeun aprs quatre
jours 8C) et aux chaleurs estivales (Euzby & Rancien, 1966 ;
Prez & Rodhain, 1977).
II.1.2.2. Spectre dhtes.
En corollaire cette tude du cycle biologique, dtaillons
maintenant le spectre dhte des diffrentes stases dIxodes
ricinus.
Tout dabord, la gamme dhtes dIxodes ricinus est trs grande.
Certains sont habituels,
dautres ne sont quaccidentels ; certains sont responsables du
transport distance des tiques et dautres jouent un rle de
maintenance de par leur abondance (micromammifres) (Prez &
Rodhain, 1977).
Ainsi, de nombreuses espces danimaux sauvages ou domestiques
constituent des
htes potentiels. Les adultes, les nymphes et les larves
parasitent communment des mammifres de taille moyenne grande. Les
nymphes et les larves se nourrissent aussi sur des petits
mammifres, de nombreuses espces doiseaux et mme des reptiles
(Hillyard, 1996).
Lespce est frquemment retrouve sur lHomme, notamment au stade
nymphal et, plus
encore au stade adulte (Gilot et al., 1979), mais tous les
stades peuvent se fixer (Aeschlimann, 1972) et de nombreux cas
dinfestations par des larves ou des nymphes passent en fait
inaperus (Gilot & Marjolet, 1982).
Slectivit des larves et des nymphes : Elles parasitent
principalement les rongeurs, les insectivores, les Mustlids ainsi
que
les oiseaux, notamment les Galliformes et les Passriformes
(Morel, 1980). Mme si les larves prfrent les micromammifres, de
nombreuses espces dhtes sont dcrites, comme pour les nymphes qui
sont rputes plus ubiquistes.
Slectivit des adultes : Par rapport aux immatures, les adultes
ont une slectivit bien plus restreinte : leurs
htes seront de faon prdominante, les onguls sauvages ou
domestiques et secondairement les carnivores de grande taille.
Parmi les onguls domestiques, ils ont une prfrence pour le Mouton,
la Chvre et le Bovin. On les rencontre aussi sur le Cheval, le
Chien, le Chat et le Furet (Roman & Lipstein, 1945 ; Arthur,
1948). Au final, la prsence effective dIxodes ricinus semble
dpendante des cervids et du Sanglier en milieu bois et des
ruminants domestiques en milieu rural ouvert (Morel, 1981). Ce
constat nous conduit voquer la distribution gographique de cette
espce.
-
13
A titre indicatif, voici une liste non exhaustive dhtes sauvages
parasits par Ixodes ricinus (Macleod, 1932 ; Roman & Lipstein,
1945 ; Morel, 1961 ; Aeschlimann, 1972 ; Gilot & Aubert, 1985)
:
Pour les mammifres : Blaireau, Campagnol, Cerf laphe, Chat
sylvestre, chauve-souris,
Chevreuil, Ecureuil, Fouine, Genette, Hrisson, Hermine, Lapin,
Livre, Martre, mulots, Putois, Renard, Sanglier, Taupe
Pour les oiseaux : Chevalier combattant, Choucas, chouettes,
Corneille, Etourneau, Faisan, Faucon crcerelle, Fauvette tte noire,
Grive mauvis, Merle, Pluvier dor, Roitelet triple bandeau
Pour les reptiles : Lzard vert, Lzard des murailles, Lzard des
souches, Lzard vivipare
II.1.3. Distribution gographique.
Il sagit de la tique la plus commune de lEurope du nord-ouest et
son aire de rpartition
stend de lIslande lAsie Centrale, en passant par les montagnes
de lAtlas (Afrique du Nord) (Hillyard, 1996).
De nombreuses collectes dIxodes ricinus sont rpertories en
France, mais beaucoup
concernent des tiques prleves sur htes. Il parat donc difficile
de dcrire tous les habitats coloniss par lespce (Gilot et al.,
1975a).
Mais une bonne estimation de son aire de distribution peut tre
approche grce la
rpartition des cas de babsiose bovine dont elle est le vecteur :
la prsence de Babesia divergens est obligatoirement accompagne de
celle dIxodes ricinus. Pour ce faire, nous pouvons superposer les
cartes de rpartition de la babsiose bovine obtenues dans le cadre
des tudes de LHostis et al. (1995a) conduite en 1988-1989 et de
Devers (2002). Ces donnes sont issues de rponses de vtrinaires un
questionnaire national sur la babsiose bovine : la premire tude a
obtenu un total de 622 rponses (dont 467 foyers) sur 1480
questionnaires envoys et la seconde 605 rponses (dont 416 foyers)
sur 1443 questionnaires.
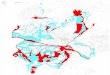
Les cartes de distribution de la babsiose ont t synthtises sur
la carte de la figure 4. Nous remarquons que lespce est trs
abondante en France et rpartie sur une grande
partie du territoire, lexception des zones sches du sud-est.
Dailleurs, toutes les tentatives de collecte au sud de la Drme se
sont avres infructueuses (notamment dans les forts des
Alpes-Maritimes), mme sil est probable que des lots coloniss y
persistent (Gilot et al., 1975b).
Le Limousin nchappe pas cette aire de rpartition. En effet,
lespce est trs largement
rpandue dans le sud-ouest et a notamment t collecte dans les
dpartements de : Charente, Charente-Maritime, Dordogne, Gers,
Gironde, Haute-Vienne, Landes, Puy-de-Dme et Vienne (Roman &
Lipstein, 1945 ; Lamontellerie, 1965 ; Morel, 1981). Un peu plus
lest, cest la seule espce collinenne pntrer dans les sries de ltage
montagnard humide du Massif Central et elle y a t collecte de 200
1150 m daltitude (Gilot et al., 1989).
-
14
Figure 4 : Synthse de la rpartition des foyers de babsiose
bovine en France partir de
deux enqutes auprs des vtrinaires, de LHostis et al. (1995a) et
Devers (2002).
II.1.4. Phnologie dIxodes ricinus.
II.1.4.1. Habitats coloniss.
La prsence dIxodes ricinus lintrieur de cette aire de
distribution nest pourtant pas uniforme et est corrle des facteurs
gographiques dune part (pluviosit, hydrologie, relief, sol et zones
phytogographiques) et des facteurs lis lactivit humaine dautre part
(Prez & Rodhain, 1977).
-
15
a) Les biotopes coloniss: A cause de ses besoins hygromtriques
levs (les trois stades volutifs ne survivent
que peu de jours si lhygromtrie baisse en dessous de 70 %), cest
surtout la prsence ou labsence dun couvert vgtal adquat qui rglera
la distribution gographique de cette espce (Milne, 1946 ;
Aeschlimann, 1972). Rappelons quIxodes ricinus est une tique
exophile tous les stades (mme si lendophilie est possible pour les
immatures).
Les associations vgtales les plus diverses pourront ainsi servir
de gte aux stades
volutifs. Les larves et les nymphes attendront dans les couches
basses de la vgtation, les adultes dans la strate plus leve. Cest
la fort de feuillus suffisamment claire, au sous-bois dense qui
semble la plus favorable cette espce (sylvo-exophilie)
(Aeschlimann, 1972 ; Gilot et al., 1975a). Pourtant, selon les
rgions tudies, le couvert vgtal le plus favorable varie : ainsi, au
Pays de Galles, Arthur (1948) observe quau sud les joncs sont plus
favorables que les fougres, tandis quau centre cest linverse. Les
plantes des ptures forts taux dinfestations classiquement dcrites
sont les suivantes (Macleod, 1939 ; Milne, 1944) : fougres, joncs,
carex, molinies et bruyres. A linverse, les tiques semblent fuir la
flore nitrophile : orties, Oseille sauvage (Euzby & Simon,
1984).
Ltude franaise de Dorchies (2001) a objectiv une relation
significative entre la
prsence de nymphes sur les ptures et les cinq plantes suivantes
:
- la Fougre aigle (Pteris aquilina) ; - le Chiendent rampant
(Agropyrum repens) ; - le Cynosure crtes (Cynosurus cristatus) ; -
le Lotier cornicul (Lotus corniculatus) ; - le Trfle blanc
(Trifolium pratense).
Il semble aussi que les tiques, qui ont elles-mmes des
mouvements trs limits, se
retrouvent prfrentiellement dans des niches cologiques riches en
mammifres (Joyner & Donnelly, 1979). A ce titre, la densit de
la faune sauvage parat essentielle dans le maintien et
lamplification des populations dIxodes ricinus et son rle est
suprieur celui jou par le btail domestique, car les Cervids sont
les htes prfrs des femelles adultes, les animaux domestiques ntant
quune alternative peu efficace et peu adapte (LHostis &
Seegers, 2002). Dailleurs, dans le sud-est de la France, Gilot
& Aubert (1985) remarquent que les densits maximales de
chevreuils concident avec les territoires les plus riches en Ixodes
ricinus, et Prez-Eid (1989) note une plus grande densit de tiques
adultes dans les zones forestires o les grands mammifres sauvages
se rassemblent.
b) Cas particulier de lcosystme prairial : I. ricinus est
essentiellement une tique de bois et fort, mais saccommode trs bien
de
certains types de pturages (LHostis, 1997). Les ptures des
bovins les plus risque semblent celles dont la clture est faite de
haies, ou qui sont adjacentes des bois, surtout quand les animaux
les utilisent comme abri temporaire pendant les grandes chaleurs de
la journe (Euzby & Rancien, 1966 ; Gern et al., 1982). De mme,
les prairies broussailleuses des bocages, avec buissons, fourrs,
roncires, friches ou marcages flore dense et varie sont des
biotopes propices linfestation (Famere et al., 1977).
Euzby & Simon (1984) notent un dfaut dhomognit dans la
rpartition spatiale des
tiques au sein des ptures. En effet, le pturage lui-mme, quelle
que soit la hauteur de son herbe, est souvent libre dI. ricinus,
hormis les bandes limitrophes de vgtation plus dense (Gern et al.,
1982). La capture dune tique en plein pturage reste donc
anecdotique par rapport aux collectes sur les bordures : 83,4 % des
tiques collectes par LHostis et al., (1995b) provenaient en effet
des bordures.
-
16
Enfin, trois catgories de prairies en fonction du risque de
contamination par I. ricinus ont t identifies par Mmeteau et al.
(1998) :
les parcelles fort risque dinfestation (96 %), qui sont situes
proximit dun
bois de feuillus ou de conifres ; les parcelles risque
dinfestation intermdiaire (39 %), qui sont bordes de haies
ou qui sont des prairies naturelles ; les parcelles faible
risque dinfestation (13 %), qui sont des prairies artificielles
loignes de tout bois.
c) La pdologie : Concernant la pdologie, les avis divergent.
Alors que Milne (1944) ne trouve aucune
influence de la composition chimique, de la texture et du pH du
sol, Arthur (1948) note que les sols infests ont un pH compris
entre 4,4 et 5,2 (sols acides), ce quil explique par une activit
bactrienne et une dcomposition des matires organiques du sol
interrompues, cela tant favorable au maintien du couvert vgtal
humide ncessaire la survie des tiques. Dans le sud-est de la
France, les biotopes I. ricinus correspondent des groupements lis
des sols dont les humus, bien structurs, sont de type mull (sols pH
neutre) calcique ou mull forestier et le rapport carbone sur azote
varie de 9,4 18,4 (Gilot et al., 1975b).
d) Le climat : Le climat qui influence la fois lhygromtrie, les
tempratures et la vgtation a, on
limagine, une influence majeure sur la distribution des tiques.
Tout dabord, Aeschlimann (1972) considre laltitude comme un facteur
limitant de premire importance ; il dcrit dans le Jura Suisse une
rarfaction de lespce de 800 1200 m et une absence au-del de 1500 m.
Cette constatation est dailleurs corrobore par son absence dautres
massifs montagneux comme le Mont Ventoux (Alpes) (Gilot, 1987).
Tandis quen France, I. ricinus est largement rparti sur le
territoire, il est plus rare au
centre de lEspagne et se retrouve cantonn aux zones climatiques
fraches et tempres humides des massifs montagneux (Encinas-Grandes,
1986). La temprature semble plutt contrler lactivit, alors que la
distribution et la densit des tiques sont grandement dpendantes de
lhumidit (Macleod, 1939 ; Milne, 1950). Au dbut du printemps (o
lhygromtrie du sol reste suffisante) ce sont les tempratures de 10
18C qui sont optimales et qui favorisent lactivit de lespce, alors
quen t, sa survie dpend fortement dune hygromtrie atmosphrique
suffisante (Gilot et al., 1975b).
II.1.4.2. Activit dIxodes ricinus.
a) Activit saisonnire.
i. Activit bimodale. Dans les pays dEurope tempre, la population
active dI. ricinus volue de manire
bimodale au cours de lanne (Edwards & Arthur, 1947 ; Arthur,
1948 ; Aeschlimann, 1972 ; Prez-Eid, 1989 ; LHostis et al., 1996b ;
Vassalo et al., 2000 ; Randolph et al., 2002) avec :
un important pic de printemps de mars juillet selon les rgions ;
un second pic dactivit moins important lautomne, de septembre
novembre.
-
17
Les adultes et les nymphes suivent la mme courbe dactivit au
printemps, tandis que les larves tardent entrer en activit de
quelques semaines (Milne, 1947). Les adultes et les nymphes ont une
activit importante partir de fvrier et les larves partir de juin
comme le montre la figure 5, ci-dessous (LHostis et al.,
1995b).
0
50
100
150
200
250
300
350
400
10.9
1
11.9
1
12.9
1
01.9
2
02.9
2
03.9
2
04.9
2
05.9
2
06.9
2
07.9
2
08.9
2
09.9
2
10.9
2
11.9
2
12.9
2
01.9
3
02.9
3
03.9
3
04.9
3
05.9
3
06.9
3
07.9
3
08.9
3
Mois
Nymphes et adultes
0
200
400
600
800
1000
1200
Larves
Nymphes et adultes Larves
Figure 5 : Variations mensuelles du nombre dIxodes ricinus
rcolts par la mthode du drapeau dans quatre levages de la Sarthe
pendant deux ans, daprs LHostis et al. (1995b).
ii. Activit unimodale. Le taux de dveloppement et la qute des
htes tant grandement dpendants de la
temprature, la dynamique saisonnire des populations de chaque
stase dI. ricinus connat des variations gographiques (Randolph et
al., 2002). Ainsi, le mode dvolution unimodal se rencontre surtout
dans des conditions climatiques moins favorables. Dans les pays
dEurope du nord, le seul pic dactivit a lieu au printemps ou en t
(Arthur, 1948). Enfin, les populations mditerranennes en bordure de
maquis ou en montagne sont actives en hiver, doctobre mai (Morel,
1981).
b) Activit journalire.
Durant la vie libre : Durant les priodes dactivit, les
diffrentes stases sont prsentes sur la vgtation
aussi bien le jour que la nuit. Cependant, le nombre dindividus
lafft et leur aptitude saccrocher un hte semblent trs infrieurs la
nuit. Ce comportement parat suivre les variations nycthmrales des
tempratures (Milne, 1947).
En effet, mme si les mouvements des tiques libres restent trs
limits, les mouvements
verticaux sur la vgtation sont importants. Ainsi, la descente
dune tique lafft amorce la fin dune priode dactivit et le dbut dune
priode de quiescence o lindividu reste au sol (Milne, 1945). La
nuit les descentes sont bien plus importantes que le jour, car les
tempratures sont infrieures, comme le montre la figure 6 (Lees
& Milne, 1951).
-
18
20
25
30
35
40
45
50
08:00 13:30 15:00 17:00 19:00 21:00 00:00 03:00 05:30 09:00
14:00 Heure
Nom
bre
de ti
ques
0
2
4
6
8
10
12
14
16
18
20 C
Nombre de tiques adultes en activit Temprature
Figure 6 : Activit quotidienne dIxodes ricinus, selon Lees &
Milne (1951). Concernant leur position sur les vgtaux, les tiques
lafft sont surtout prsentes sur
le tiers suprieur. Mais, le vent et la direction du soleil
influencent cette position, puisque les tiques saccrochent
majoritairement sur la face protge du support. Le soleil et le vent
acclrent la dessiccation et lalternance de priodes dactivit et de
priodes de quiescence permettant la tique de recouvrer leau
corporelle perdue. De plus, la rponse un stimulus est moins
efficace sous le vent, car les remous dair chaud servent
habituellement la dtection de lhte (Milne, 1948 ; Lees & Milne,
1951).
Durant la vie parasitaire : Malgr une activit majoritairement
diurne, linfestation des micromammifres par les
larves dI. ricinus se fait surtout entre 20 heures et 24 heures,
cela tant d aux rythmes dactivits nocturnes de ces htes dans la
nature. Par contre, le dtachement des larves est diurne (comme les
autres stases) et le sommet du pic de dtachement concide avec la
priode dactivit minimale de lhte, elles se dtachent donc surtout
dans son terrier (Graf et al., 1978).
Tempratures infrieures 10C Nuit
-
19
II.2. Les tiques du genre Dermacentor.
Les deux espces du genre Dermacentor vivant en France et
susceptibles de parasiter
les bovins sont D. marginatus et D. reticulatus. D. marginatus
est plutt une tique des ruminants, tandis que D. reticulatus est
plutt une tique des carnivores.
Signalons par ailleurs, que le D. niveus, Neumann, 1897 de la
littrature ancienne, correspond au
D. marginatus (Sulzer, 1776) de la nomenclature actuelle (Roman
& Sicart, 1957) et que lancien D. pictus (Hermann, 1804)
correspond au D. reticulatus (Fabricius, 1794) actuel (travaux
dArthur, 1960 in Lamontellerie, 1965).
II.2.1. Caractristiques communes au genre Dermacentor.
II.2.1.1. Caractristiques morphologiques communes.
Il sagit despces brvirostres (rostre court et large) et
mtastriata, le sillon anal tant postrieur lanus et en forme de
coupe. Le scutum prsente des ocelles et est orn de taches mailles
et panaches. Le trochanter I na pas dpine pointue large mais lpine
externe de la hanche I est prsente (Hillyard, 1996).
La terminologie morphologique utilise dans le texte
est rcapitule en annexe IIa & IIb.
II.2.1.2. Cycles volutifs des Dermacentor.
Les cycles volutifs de D. marginatus et D. reticulatus sont
identiques, avec seulement quelques variantes concernant la
slectivit des htes.
Leur cycle est qualifi de trixne (ou triphasique) comme celui
dI. ricinus mais est cette
fois-ci ditrope et pholo-exophile. Les immatures se gorgent
prfrentiellement sur des petits mammifres, tandis que les adultes
infestent les onguls ou les carnivores de grande taille (Morel,
2000).
Tandis que les adultes sont lafft sur de hautes herbes souvent
dans les prairies ou
en bordure de bois, les stases immatures sont exclusivement
endophiles et attendent les petits mammifres dans leur terrier
(Gilot & Pautou, 1983). Les adultes attendent leur hte une
hauteur variant de dix soixante centimtres. Ils tiennent leur
extrmit antrieure dirige vers le sol, et embrassent la tige vgtale
de leur troisime paire de pattes, les autres pattes tant tendues et
vibrantes au moindre dplacement (Gilot et al., 1974).
La femelle adulte se gorge de sang, le mle galement, mais en
moindre quantit (dure
du repas de sang de la femelle de six quinze jours).
Laccouplement des adultes a lieu sur lhte et une fois gorge, la
femelle se laisse choir et pond dans la partie superficielle du
sol. La larve hexapode se fixe sur un premier micromammifre, puis
se gorge, se laisse choir, et se transforme en nymphe. La nymphe a
le mme comportement que la larve et se transforme en adulte (Gilot
et al., 1974).
-
20
La ralisation du cycle ncessite entre une et deux annes
(Hillyard, 1996). Ce dernier est schmatis sur la figure 7,
ci-dessous :
Figure 7 : Cycle trixne ditrope de Dermacentor spp., daprs Morel
(2000).
II.2.1.3. Phnologie des Dermacentor.
Le genre est bien moins dpendant des biotopes forestiers quI.
ricinus et peut coloniser des biotopes ouverts ou mme proximits des
habitations (Gilot & Pautou, 1983). Les plantes qui servent de
support aux adultes sont gnralement des gramines, appartenant
diffrentes espces. Les adultes se fixent sur lappareil arien de la
plante (tige, feuilles ou pis). Dautres familles de plantes sont
possibles : ombellifres, composes, papilionaces Il peut aussi sagir
de plantes plus ligneuses comme de jeunes prunelliers ou mme des
branches mortes tombes terre (Gilot et al., 1974).
D. marginatus semble plus thermophile et moins hygrophile que D.
reticulatus (Gilot et
al., 1987). Lorsque, dans les bocages, les deux espces
coexistent, elles ont des saisons dactivit diffrentes, plus chaude
et plus sche pour D. marginatus. Et dans les paysages de campagnes,
D. reticulatus est associ aux formations denses et de bas-fonds,
tandis que D. marginatus se retrouve dans les pturages dcouverts
(Morel, 1981).
II.2.2. Dermacentor marginatus, Sulzer, 1776.
Cest une espce frquente chez les ruminants domestiques de notre
pays, surtout chez les ovins et les caprins, mais aussi chez les
bovins.
-
21
II.2.2.1. Caractristiques morphologiques (Hillyard, 1996).
Les principales diffrences morphologiques avec D. reticulatus
sont : - en vue ventrale : une pine externe de la hanche I qui est
plus courte et divergente ; - en vue dorsale : labsence dpine
latrale dirige postrieurement sur les pdipalpes.
Lensemble des critres de diagnose est rsum et schmatis sur la
planche en Annexe V.
II.2.2.2. Spectre dhtes.
D. marginatus se nourrit sur des mammifres de grande taille
comme les chvres, les bovins et les ovins. Les nymphes et les
larves infestent surtout les lapins, les petits rongeurs (comme
Apodemus sp.) et les oiseaux (Hillyard, 1996).
Elle semble nettement plus attire par les herbivores que les
carnivores, mme si elle a
t collecte sur Chien ou sur Blaireau par exemple (Gilot &
Aubert, 1985). Enfin, les cas de parasitisme humain sont loin dtre
rares (Gilot & Marjolet, 1982).
Voici une liste non exhaustive dhtes possibles (Lamontellerie,
1965 ; Gilot & Pautou, 1983 ;
Gilot & Aubert, 1985 ; Caeiro & Simes, 1991 ;
Estrada-Pea et al., 1992) : - pour les immatures : Eliomys
quercinus, Apodemus sylvaticus, Crocidura russula, Microtus
spp., Mus spp., Hrisson, Lapin de garenne, Rat - pour les
adultes : Homme, Chien, Cheval, Bovin, Chamois, Mouflon, Ovin,
Caprin, Cerf,
Sanglier, Chevreuil, Blaireau
II.2.2.3. Distribution gographique.
La distribution gographique de lespce semble trs lie la
rpartition des ruminants domestiques, qui sont ses htes principaux
; et la varit de leurs parcours, notamment en montagne (Alpages)
permet un essaimage considrable des populations de D. marginatus
(Gilot & Pautou, 1983).
D. marginatus est en fait plus thermophile et moins hygrophile
que D. reticulatus et parat
plus abondant dans les pays mditerranens (Morel, 1981 ; Gilot et
al., 1987). Il est en effet prsent en Espagne (Estrada-Pea et al.,
1992), en Italie (Principato et al., 1989), au Portugal (Caeiro
& Simes, 1991), en Grce (Papadopoulos et al., 1996) ou en
Algrie (Yousfi-Monod & Aeschlimann, 1986).
Des populations ont galement t signales en Allemagne de louest,
dans la valle du
Rhin, en Suisse, en Pologne et en Asie Centrale (Hillyard,
1996). En France, lespce est abondante mais est surtout signale
dans le sud-ouest et le midi
mditerranen (Roman & Sicart, 1957 ; Elfassy, 1993). Elle a
aussi t collecte dans dautres dpartements : Ain, Charente-Maritime,
Cte dOr, Deux-Svres, Haute-Vienne, Indre, Indre-et-Loire, Isre,
Nivre, Bas-Rhin, Rhne, Vienne (Enigk, 1947 ; Roman & Sicart,
1957 ; Callot, 1959 ; Morel, 1961 ; Lamontellerie, 1965).
Ainsi, la prsence de lespce dans le Limousin est possible, ce
qui a t confirm par sa collecte
en avril 1944, Limoges (Enigk, 1947). De plus, D. marginatus est
frquent dans le Massif Central (Gilot et al., 1989).
-
22
II.2.2.4. Phnologie de Dermacentor marginatus.
a) Biotopes coloniss. Les sries de vgtation o lon trouve D.
marginatus sont nombreuses : il sagit
notamment de la vgtation mditerranenne, surtout les tages de la
chnaie thermophile et de la vgtation subcontinentale chnaie
acidophile. D. marginatus habite des formations ligneuses et
forestires, mais supporte aussi bien des formations vgtales fermes
quouvertes et peut habiter les milieux ouverts arides sans strate
arbustive (pseudo-steppes), les milieux semi-ouverts (landes) et
les lisires de forts (Cotty et al., 1986). Il prfre les forts
clairires aux forts denses (Gilot & Pautou, 1983). Lespce parat
absente des milieux nettement artificialiss, contrairement D.
reticulatus (Gilot & Pautou, 1982).
Certaines espces herbaces ont une valeur indicatrice de porte
gnrale : cest le cas
du Brachypode penn (Brachypodium pinnatum) et du Brome dress
(Bromus erectus) (Gilot & Pautou, 1983).
Soulignons galement une affinit marque de D. marginatus pour les
landes,
notamment les landes Gent (Genista cinerea), Genvrier (Juniperus
communis, Juniperus oxycedrus), Ajonc dEurope (Ulex europaeus),
Argousier (Hippophae rhamnoides), Epines-vinettes (Berberidion) ou
Peuplier (Populus nigra), particulirement dans les valles des cours
deau (Gilot & Pautou, 1983). Dans les rgions de bocage, elle
est frquemment retrouve au cur des prairies ouvertes (Morel,
1981).
Lespce est gnralement prsente jusqu 800-1000 m daltitude mais
est susceptible
de sinstaller au dessus de 1000 m en mettant profit une
exposition favorable (Gilot & Pautou, 1983).
b) Le climat.
Du point de vue climatologique, les stations de collectes de
Gilot et al. (1989) dans le
Massif Central ne prsentent jamais des prcipitations trs leves.
D. marginatus manifeste donc des tendances la xrophilie. Les
tempratures annuelles des diffrents sites varient de 10C 14C. Malgr
une certaine thermophilie, dans le centre de lEspagne, lespce est
cantonne aux zones froides et semi-arides semi-humides des
montagnes (Encinas-Grandes, 1986).
Dans les zones o les tempratures sont trs favorables, les
populations peuvent se
dvelopper dans des biotopes relativement plus humides. Cette
diffrence dans les facteurs climatiques limitants, observe entre
deux rgions, illustre le phnomne nomm stnocie rgionale (Cotty et
al., 1986).
c) Activit saisonnire.
Lactivit des adultes se situe en climat tempr, de mars mai et
daot octobre ;
dans le Bassin Mditerranen, de fvrier mai et de septembre
novembre (Morel, 1981). Dans les Alpes, les immatures, qui sont
exclusivement endophiles, sont actifs en t (mi-
juin aot) et en dbut dautomne (jusqu la mi-septembre), tandis
que les adultes exophiles ont une activit plus verno-automnale avec
une vritable diapause imaginale (dpendante de la temprature et de
la photopriode) en t (Gilot & Pautou, 1983).
-
23
En Italie, Principato et al. (1989) retrouvent, dans les ptures
de moutons, ces deux pics dactivit, comme en France (figure 8,
ci-dessous).
0
10
20
30
40
50
janvie
r
fvrie
rma
rsav
ril mai
juin
juille
tao
t
septe
mbre
octob
re
nove
mbre
dce
mbre
Nombre deD. marginatus
rcolts
Figure 8 : Fluctuations mensuelles des populations adultes de D.
marginatus en Italie,
daprs Principato et al. (1989). Mais dans des sries trs
thermophiles, la priode dactivit est courte. Les larves sont
actives au printemps (mars avril) et les nymphes la fin de lt
(aot et septembre). Les adultes sont actifs approximativement de
fin septembre mars, quand la temprature descend sous les 15C de
moyenne et que les pluies reviennent (Estrada-Pea et al., 1992). En
Algrie, D. marginatus est une tique exclusivement hivernale
(janvier et fvrier), parasitant principalement les suids
(Yousfi-Monod & Aeschlimann, 1986).
II.2.3. Dermacentor reticulatus (Fabricius, 1794).
Elle semble au moins aussi importante que D. marginatus chez les
bovins en France.
II.2.3.1. Caractristiques morphologiques (Hillyard, 1996).
Contrairement D. marginatus, D. reticulatus possde : - en vue
ventrale : une longue pine externe parallle et colle la hanche I ;
- en vue dorsale : une pine latrale dirige postrieurement sur les
pdipalpes.
Lensemble des critres de diagnose est rsum et schmatis sur la
planche en annexe VIa.
II.2.3.2. Spectre dhtes.
Rappelons quil sagit dun cycle ditrope comme pour D. marginatus.
Les adultes infestent les chiens, les chevaux, bovins, ovins,
caprins, renards, livres et hrissons, tandis que les immatures qui
sont endophiles, se nourrissent sur des petits mammifres, Rongeurs
(mulots, campagnols), insectivores (musaraignes) et Mustlids ;
occasionnellement sur les oiseaux (Hillyard, 1996).
-
24
On le voit, les adultes infestent des grands mammifres,
principalement des Onguls domestiques ou sauvages, mais galement
des carnivores. Le Chien, qui D. reticulatus transmet Babesia
canis, est un hte trs important (Elfassy, 1993). Et parmi les
carnivores sauvages, cette tique parat prsenter une affinit lective
pour le Renard (Gilot & Aubert, 1985). Le parasitisme humain
existe, notamment au niveau du cuir chevelu (Vermeil & Puech,
1950).
II.2.3.3. Distribution gographique.
D. reticulatus est trs largement rpandu de lEurope lAsie
Centrale. Il est prsent en France, Espagne, Suisse, Grande
Bretagne, Belgique, Allemagne et Pologne. En Europe du nord-ouest,
lespce est rare ou localise quelques rgions (Hillyard, 1996). Cette
tique est en effet retrouve sur les Bovins des ctes du sud-ouest de
lAngleterre et du Pays de Galles (Campbell, 1950). Mais elle est
absente des pays qui bordent la Mditerrane comme lEspagne ou
lItalie (Gilot et al., 1987).
En France, contrairement D. marginatus, D. reticulatus est
thoriquement absent de la
Rgion mditerranenne sensu stricto, et sa distribution ne dpasse
pas la latitude de Montlimar (Elfassy, 1993). Cela nempchant pas
les nombreuses collectes de Gilot et al. (1974) dans le sud-est de
la France.
Concernant les dpartements proches du Limousin, lespce a t
collecte en : Charente,
Dordogne, Indre, Indre-et-Loire, Vienne (Roman & Lipstein,
1945 ; Roman & Sicart, 1957 ; Morel, 1961 ; Lamontellerie,
1965). Le Limousin fait donc partie intgrante de cette aire de
rpartition, comme en tmoignent les collectes en Creuse (adultes sur
Cerf laphe) ou en Haute-Vienne sous le nom de D. pictus, Hermann,
1804 (Enigk, 1947 ; Morel, 1961).
II.2.3.4. Phnologie de Dermacentor reticulatus.
a) Biotopes coloniss. Les terrains dans lesquels on retrouve D.
reticulatus peuvent tre classs comme
suit (Gilot et al., 1974) : Pour les biotopes ruraux :
les prairies ptures (comme D. marginatus) : souvent dans un
paysage bocager ; la prsence de haies en bordure, abritant de
nombreux micromammifres est en effet trs favorable ;
les haies : notamment celles avec Aubpine, Eglantier,
Cornouiller ou Orme champtre
les landes (tout comme D. marginatus) : elles juxtaposent des
plantes herbaces et des arbustes (Gent et Chtaignier) ;
les bois clairirs de chnaies (tout comme D. marginatus) : il
prfre les lisires et les sentiers o les gramines peuvent
pousser.
Pour les biotopes anthropiss :
les terrains vagues suburbains des grandes villes (contrairement
D. marginatus) : il sagit souvent danciennes prairies, du moment
que la strate herbace soit continue ; les fortes densits de chiens
en promenade et de micromammifres peut mme conduire une vritable
pullulation ;
les berges des rivires : lieux de promenade trs frquents.
-
25
Martinod & Joubert (1981) observent mme une focalisation de
lespce dans les zones priurbaines du Jura mridional, par rapport
aux zones rurales. Quoi quil en soit, D. reticulatus colonise les
zones rurales o la fort noccupe que des surfaces restreintes et peu
denses, do lattrait des zones bocagres (Martinod et al., 1984).
D. reticulatus se localise principalement dans les zones basses
des bassins fluviaux,
dans les formations denses, fourrs arbustifs ou galeries Aulne,
Peuplier et Saule, au bord des rivires (Morel, 1981). Et dans les
rgions thermophiles, lespce se cantonne aux zones dpressionnaires
situes dans laxe des rivires et prs des lacs (Gilot et al.,
1974).
Lespce nest pas adapte aux rgions montagneuses et semble absente
au-del de
700 ou 800 m daltitude ; elle est rare en rgions
submontagnardes, mais est frquente dans les zones collinennes
(Gilot et al., 1974 ; Martinod et al., 1984).
b) Activit saisonnire. Il sagit dune tique de saisons fraches et
humides. Lactivit des adultes en Europe
occidentale sobserve surtout doctobre dcembre et de fvrier
avril, tout en restant possible de septembre mai (Gilot, 1981).
En France, cest au printemps que lactivit est la plus forte
(mars, avril et mai). Il nest
cependant pas rare que D. reticulatus conserve une activit tout
au long de lanne. Dans le sud-est, mme si des individus sont rcolts
en dcembre et janvier ou en juillet
et aot, lactivit baisse nettement en hiver et en t. En automne,
lactivit est faible et remonte rapidement vers la fin fvrier,
jusquen mai, comme on peut le voir sur la figure 9, ci-dessous
(Gilot et al., 1974).
0
10
20
30
40
50
60
70
80
90
100
110
janv
ier
fvr
ier
mar
s
avril
mai
juin
juille
t
aot
sept
embr
e
octo
bre
nove
mbr
e
dce
mbr
e
Mois
Nombre d'individus
Eybens
Echirolles
Grenoble
Figure 9 : Rpartition mensuelle des rcoltes de D. reticulatus en
Isre (38),
daprs Gilot et al. (1974).
-
26
II.3. Les tiques du genre Haemaphysalis.
II.3.1. Haemaphysalis punctata, Canestrini & Fanzago,
1877.
Sa capacit coloniser de nombreux microhabitats en France et sa
slectivit vis--vis des bovins font delle une espce incontournable
dans notre tude.
II.3.1.1. Caractristiques morphologiques (Hillyard, 1996).
Il sagit de tiques brvirostres, mtastriata avec un sillon anal
postrieur lanus. Le scutum ne prsente ni ocelle, ni ornementation.
Le corps possde des festons. De plus, le trochanter I (2me article
des pattes I) possde une large pine mais la hanche I est simple et
ne possde pas dpine externe.
Lensemble des caractristiques morphologiques est rsum et
schmatis
sur la planche en annexe VII.
II.3.1.2. Cycle volutif de Haemaphysalis punctata.
a) Particularits de son cycle. On a affaire un cycle trixne,
tlotrope qui prsente de grandes analogies avec celui
dIxodes ricinus (Cotty et al., 1986). La ralisation du cycle
ncessite le plus souvent entre deux et trois annes. Mais dans
des conditions trs favorables la dure peut tre grandement rduite
: jusqu 131 jours (Herbert et al., 1981).
Les diffrentes stases sont exophiles (Cotty et al., 1986). Et
mme si lexophilie des
larves nest pas systmatique, elle a t prouve par la technique du
drapeau (Gilot, 1985). La mthode du drapeau (cf. chapitre III.2.2)
semble trs efficace dans la collecte de cette espce, mais donne
parfois des sex-ratios dsquilibrs. En effet, Herbert et al. (1981)
ont collect moins de mles que de femelles (sex-ratio de 1 : 1,56)
et mettent les hypothses quun mle pourrait copuler avec plusieurs
femelles ou que les femelles saccrocheraient mieux au drapeau. La
copulation a lieu le plus souvent sur le sol, car de nombreux
individus accoupls sont collects. La spermatogense du mle ne
ncessite donc pas de repas sanguin aprs le repas nymphal. Et aucun
des mles qui ont t dposs jeun sur des ovins ne se sont fixs, alors
que toutes les femelles se sont gorges et ont pondu des ufs fconds
(Herbert et al., 1981). Il scoule ensuite de 38 45 jours entre le
dbut de la ponte et lclosion des ufs (Lamontellerie, 1965).
b) Son spectre dhtes. Les stases immatures sont relativement
ubiquistes, avec une prfrence pour les petits
mammifres, tandis que les adultes ne parasitent que des
mammifres de grande taille, principalement des onguls (Elfassy,
1993).
-
27
Slectivit des adultes : Les adultes sont avant tout des
parasites des bovins et des ovins (Hillyard, 1996).
Cependant, cette espce est aussi associe une faune sauvage
relativement riche. Les adultes parasitent des vertbrs de grande
taille, incluant les cerfs, chamois, chameaux, porcs, chiens,
chvres, nes et chevaux. Ils ont aussi t retrouvs sur des hrissons,
lapins et livres (Herbert et al., 1981 ; Rossi & Meneguz,
1989). Le Faisan (Phasianus colchicus) est le seul oiseau signal
comme hte pour les adultes (Herbert et al., 1981). LHomme est une
cible possible (Hillyard, 1996).
Slectivit des immatures : Larves et nymphes infestent surtout
les petits mammifres, comme les lapins, les livres,
les cureuils, les souris, les mulots sylvestres, les campagnols,
les taupes et les hrissons. Elles sont trs couramment collectes sur
des oiseaux (souvent sur des espces migratrices), comme les
corneilles, les pies, les faisans, les tourneaux, les moineaux, les
merles, les fauvettes, les bruants et les sternes. Des larves ont t
signales sur des lzards. Et les nymphes peuvent aussi sattaquer
occasionnellement des chvres, bovins, moutons, chevaux, chiens,
renards, belettes et blaireaux (Herbert et al., 1981 ; Hillyard,
1996).
II.3.1.3. Distribution gographique.
H. punctata est trouv dans toute lEurope, lIrlande excepte.
Cette tique est prsente de lEspagne la Scandinavie et lest, jusqu
lAsie Centrale. Elle a aussi t signale en Afrique du nord. En
Grande Bretagne, la plupart des populations habite le sud-est de
lAngleterre et la cte ouest du Pays de Galles (Campbell, 1950 ;
Hillyard, 1996).
Sa distribution gographique peut galement tre rvle par la
prsence de Babesia
major, dont elle est le seul vecteur prouv (Brocklesby &
Barnett, 1972 ; Brocklesby & Sellwood, 1973). Ainsi, en
Allemagne, la dcouverte de B. major a conduit Liebisch & Rahman
(1976) a suspecter sa prsence ; hypothse qui a t confirme par des
recherches au drapeau.
En Italie par exemple, 8,4 % des tiques (119 adultes sur 1410 au
total) collectes par
Cringoli et al. (2002) dans le sud et 11,7 % de celles collectes
par Principato et al. (1989) au centre, appartenaient cette
espce.
En France, lespce est largement rpartie, mais souvent dans des
petites zones
biotope bien dfini (Mage et al., 2001 ; LHostis & Seegers,
2002). La babsiose bovine B. major transmise par H. punctata a t
mise en vidence dans le Lot (Malbert et al., 1988) et en Charente
(LHostis et al., 1995a). Laire de distribution thorique de cette
tique inclurait en fait tout le territoire, except les Alpes et la
Corse. Mais les collectes sporadiques dans les Alpes laissent
entrevoir le rle possible des oiseaux migrateurs parasits dune part
et ladaptabilit de lespce divers microhabitats dautre part (Gilot
et al., 1979). De plus, la prsence de Theileria orientalis,
transmis par H. punctata a t mise en vidence en Corse, de mme que
dans une le Bretonne sous forme asymptomatique, H. punctata a
dailleurs t collecte Belle-Ile-en-Mer (LHostis, comm. pers ; Mage
et al., 2001 ; Blary, 2004).
Les donnes sont trop parses pour savoir si lespce est rellement
prsente en Limousin.
Toutefois, quelques collectes nous conduisent suspecter sa
prsence au moins de faon localise : il sagit de collectes dans
louest et le sud-ouest, notamment en Dordogne, en Gironde, dans
lIndre, dans les Landes et dans le Lot (Morel, 1961 ; Morel, comm.
pers. in Lamontellerie, 1965).
-
28
II.3.1.4. Phnologie dHaemaphysalis punctata.
a) Biotopes coloniss. Cette vaste aire de rpartition htrogne et
morcele laisse supposer une grande
plasticit cologique et une extrme adaptabilit : cette tique est
en effet capable de sacclimater au climat doux et humide de
lAngleterre et de persister dans les semi-dserts dEurope Centrale
(Gilot, 1985).
Les exigences cologiques de H. punctata sont trs voisines de
celles de D.
marginatus ; les deux espces sont msophiles et leurs
distributions concident en grande partie, en Eurasie tempre comme
dans le bassin mditerranen (Morel, 1981).
Lespce trouve son optimum dans ltage supramditerranen (notamment
ltage du
Chne pubescent, dans les landes Gent). Sur la bordure orientale
du Massif Central, elle colonise les biotopes allant de 50 600-800
m daltitude, avec une temprature moyenne de 11,3 12,5C et des
prcipitations assez abondantes variant de 1110 1400 mm par an. Elle
reste cependant assez localise (Gilot et al., 1989).
Comme Ixodes ricinus, H. punctata est prsent dans les zones
forestires peu denses et
les prairies (Dellac, 1999). Les plantes servant de support
lafft sont souvent des gramines (Ftuques ou brises intermdiaires),
plus exceptionnellement dautres plantes herbaces (Ombellifres :
carottes sauvages et Composes notamment). Des tiges ligneuses
peuvent galement convenir, notamment dans les paysages de landes
(rameaux latraux de gents, Genista cinerea, de genvriers, Juniperus
communis, voire daubpines, Crataegus monogyna). La tique se poste
quelques centimtres de lextrmit du support vgtal, le plus souvent
la tte dirige vers le bas. Des supports artificiels (poteaux en
bois) ont t signals (Gilot, 1985).
Comme nous lavons dit, lespce est vraisemblablement msophile
(Gilot, 1985). Mais
le phnomne de stnocie rgionale dj signal chez D. marginatus est
possible. En effet, alors que dans le Massif Central et les Alpes,
lespce est loin de se comporter comme une tique xrophile (Gilot,
1985 ; Gilot et al., 1989), en Suisse, Cotty et al. (1986)
constatent la fois une xrophilie lchelle de la rgion (msoclimat)
avec des prcipitations infrieures 700 mm par an et une msophilie
lchelle des diffrentes localits (microclimat). En fait, elle peut
coloniser des biotopes secs uniquement lorsquelle dispose de
conditions microclimatiques au niveau de la litire permettant une
hydratation rgulire. Lespce semble assez thermophile, comme D.
marginatus et la temprature est un facteur climatique limitant, mme
si elle colonise rarement les tages mditerranens thermophiles
(Gilot, 1985 ; Cotty et al., 1986).
b) Activit journalire.
Lactivit est grandement rduite la nuit, de minuit sept heures du
matin, du fait des tempratures basses. Lactivit nymphale est
maximale de neuf heures quinze heures et celle des adultes stend
beaucoup plus, comme le montre la figure 10 (Herbert et al.,
1981).
-
29
0
10
20
30
18 19 20 21 22 23 24 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
17Heures
Nombre de tiques
collectesNymphesAdultes
Figure 10 : Rpartitions horaires des collectes dHaemaphysalis
punctata au mois de mai,
au Pays de Galles, daprs Herbert et al. (1981).
c) Activit saisonnire. Sous climat tempr, les adultes sont
actifs de septembre dcembre et de fvrier
juin, les nymphes de mai aot, les larves de juin octobre (figure
11, ci-dessous). Sous climat mditerranen, les adultes ne se
rencontrent que doctobre avril (Morel, 1981). En Algrie, il sagit
dune tique hivernale (activit de novembre fvrier), trs localise
gographiquement (Yousfi-Monod & Aeschlimann, 1986).
J F M A M J Jt A S O N D
Figure 11 : Dynamique saisonnire dHaemaphysalis punctata, rvle
par la technique du
drapeau dans les Alpes, daprs Gilot (1985).
II.3.2. Autres tiques du genre Haemaphysalis.
Malgr la complexit et la diversit du genre Haemaphysalis, les
autres espces parasitant les bovins et prsentes en France sont trs
rares. Il sagit principalement dH. inermis Birula, 1895, dH.
concinna Koch, 1844 et dH. sulcata.
Activit imaginale
Activit nymphale
Activit larvaire
-
30
II.3.2.1. Haemaphysalis concinna Koch, 1844.
a) Spectre dhtes (Hillyard, 1996). Les htes les plus frquents
des adultes en Europe, sont les cervids, surtout le
Chevreuil. Mais ils peuvent aussi infester le Bovin, le Cheval,
le Chien, le Chat et le Livre. Les immatures sont retrouvs sur les
cervids, hrissons, rongeurs, reptiles et oiseaux,
notamment le Rouge-gorge (Erithacus rubecula), la Grive, la
Msange et le Bruant. Les nymphes parasitent galement lHomme.
b) Aire de rpartition.
H. concinna a t collecte en France, Allemagne et Pologne, mais
aussi en Chine ou au Japon. Lespce semble rsiduelle et cantonne aux
forts quasiment inaltres (Hillyard, 1996). Elle a aussi t collecte
dans le piedmont Italien sur Chevreuil (Rossi & Meneguz,
1989).
En France, la plupart des rcoltes provient des forts du Bassin
Parisien : Compigne,
Fontainebleau et Chantilly. Elle est aussi prsente dans le
centre du pays et la frontire Espagnole (Hillyard, 1996). En
Alsace, parmi les espces prsentes avec une faible probabilit, H.
concinna est la plus thermophile de toutes et serait la limite de
son aire de distribution (Prez, 1987).
La prsence dH. concinna dans le Limousin nest pas impossible,
mais compte tenu de sa raret,
il est trs peu probable de la dceler. Cela dit, des collectes
dans les dpartements de lIndre, Indre-et-Loire, Landes,
Haute-Vienne et Vienne (Morel, 1961 ; Lamontellerie, 1965)
confirment cette possibilit.
c) Phnologie dH. concinna (Hillyard, 1996).
H. concinna habite de vastes forts mixtes des climats temprs.
Elle semble frquemment associe au Htre (Bourdeau, 1993b). Toutes
les stases sont actives de mars juin, ainsi quen octobre. Les
immatures peuvent tre actifs durant de nombreux mois de lanne ; ils
sont frquents sur les micromammifres en juin et juillet. Les deux
sexes sont souvent trouvs cte cte sur lanimal et la copulation qui
a lieu sur lhte, se situe principalement en avril.
II.3.2.2. Haemaphysalis inermis Birula, 1895.
a) Spectre dhtes. Les adultes parasites les ruminants sauvages
(Chevreuil, Cerf), mais aussi les Bovins
domestiques (Rossi & Meneguz, 1989 ; Cringoli et al.,
2002).
b) Aire de rpartition. En janvier 1991, 60 adultes dH. inermis
(18 mles et 42 femelles) ont t rcolts,
jeun, par la technique du drapeau dans un rserve de chasse situe
une soixantaine de kilomtres au nord de Bordeaux (Macaigne &
Prez-Eid, 1991).
Lespce est galement cite dans les dpartements de la Vienne par
Nuttal et Warbuton (Lamontellerie, 1965), et de lIndre sur Cervus
elaphus (Morel, 1961).
En Italie elle a t identifie sur des ruminants sauvages du
Piedmont (Rossi & Meneguz, 1989). Et Cringoli et al. (2002) ont
rcolt 14 adultes (un mle et treize femelles) pour la premire fois,
dans le sud de lItalie, soit 0,9 % des 1410 tiques collectes sur
les bovins de 62 exploitations.
-
31
II.4. Les tiques du genre Rhipicephalus.
II.4.1. Caractristiques morphologiques du genre Rhipicephalus
(Hillyard, 1996).
Les espces du genre Rhipicephalus prsentent un sillon anal
contournant lanus en arrire (mtastriata). Le scutum possde des
ocelles sur les marges latrales, mais nest pas orn. La hanche I
possde une longue pine externe, mais le trochanter I nen possde
pas. Ce sont des tiques brvirostres : les pdipalpes sont plus
courts que la largeur du capitulum qui est hexagonal en vue
dorsale.
Les cls de diagnose de lannexe III permettent de prciser le
genre.
II.4.2. Rhipicephalus bursa, Canestrini & Fanzago, 1877.
Tous les stades de cette espce parasitent les bovins. Il sagit
dune tique autochtone, mais sa distribution en France reste floue.
Et malgr une rpartition un peu plus localise quI. ricinus, D.
marginatus, D. reticulatus ou H. punctata, elle constitue aussi une
espce majeure des bovins en France.
II.4.2.1. Caractristiques morphologiques (Snvet, 1937 ;
Bourdeau, 1993a ; Hillyard, 1996).
La hanche I des adultes possde une longue pine externe et
apparat donc bifide. Chez les nymphes, cette pine est absente. La
femelle possde de grandes aires poreuses sur la vue dorsale. Sur la
vue dorsale, le mle adulte possde un sillon marginal en arrire des
yeux et limitant le feston postrieur ; les sillons cervicaux sont
courts.
Lensemble des caractristiques morphologiques de lespce est rsum
et
schmatis sur la planche en annexe VIII.
II.4.2.2. Cycle volutif de Rhipicephalus bursa.
a) Son cycle. R. bursa a dvelopp un cycle particulier deux htes
(diphasique), monotrope et
exophile (Bourdeau, 1993b). Il y a suppression de la phase libre
entre la larve et la nymphe avec mue sur lorganisme hte (Dellac,
1999).
Un lment plus anecdotique est lobtention exprimentale de larves
parthnogntiques
de cette espce (Snvet, 1937).
b) Son spectre dhtes. Les immatures et les adultes infestent le
mme type dhte : les ruminants et
occasionnellement les Lporids ou lHomme (Bourdeau, 1993b). R.
bursa parasite frquemment les bovins et les moutons (Enigk, 1947).
Les quids (Cheval et ne), les chiens et les livres (parfois
abondamment infests) sont des htes possibles (Yeruham et al.,
1985).
-
32
II.4.2.3. Distribution gographique.
Laire de distribution de R. bursa stend du 31me au 45me parallle
nord, en particulier dans le bassin Mditerranen : on le trouve
notamment en Tunisie, dans les zones sub-humides et semi-arides
(Bouattour et al., 1999), au Portugal (Caeiro & Simes, 1991),
en Espagne, dans de rares rgions vgtation msomditerranenne
(Estrasa-Pea et al., 1992), en Algrie, en Grce, en Isral, en
Syrie
Dans le sud de lItalie, R. bursa est la tique la plus frquente :
65,5 % des 1410 tiques
rcoltes sur les bovins par Cringoli et al. (2002) dans 62
fermes, appartenaient cette espce.
En France, on connat mal sa rpartition, mais elle est signale
dans le sud et trs
rpandue en Corse, sa localisation ailleurs est imprcise (surtout
rgion mditerranenne) (Bourdeau, 1993b). Dans le sud-ouest, elle a t
collecte dans les Pyrnes Atlantiques ( Bayonne et ses environs,
Saint-Jean-de-Luz et Saint-Armou), en Dordogne et dans le
Lot-et-Garonne (Lamontellerie, 1965 ; Elfassy, 1993). Des tudes sur
lAnaplasmose bovine ont prouv sa prsence en Sane-et-Loire (Rossi
& Demeusois, 1952).
Il reste difficile de savoir si cette espce est prsente en
Limousin. Cependant, des populations
rduites, en liaison avec la prsence du btail, existent sous
climat tempr mridional, dans les formations ouvertes du sud du
Massif Central (Morel, 1981), comme en tmoigne les rcoltes en
Dordogne et dans le Lot (Elfassy, 1993). Dans le Massif Central,
lespce ne parat pas occuper de vastes espaces, mais Gilot et al.
(1989) sinterrogent sur la relle efficacit de la technique du
drapeau pour la dtection de cette tique.
II.4.2.4. Phnologie de Rhipicephalus bursa.
a) Biotopes coloniss. Les exigences cologiques de R. bursa sont
assez voisines de celles de H. punctata.
Son optimum se situe dans les sries submditerranennes du chne
pubescent. Certains de ces biotopes sont situs dans le
msomditerranen (Gilot et al., 1989).
Son habitat est constitu par les prairies herbeuses, les lisires
de forts, en particulier
dans les zones de collines et de montagnes basses (Bourdeau,
1993b). Lespce est signale dans des rgions arides, semi-arides et
sub-humides, comme en
Isral, o les tiques ont t trouves surtout entre les isohytes
400-600 mm, dans les rgions de collines, couvertes de garrigue.
Elles affectionnent particulirement les zones climat mditerranen
avec un t long et sec, un hiver froid et humide et des tempratures
annuelles comprises entre 15 et 20C (Liebisch & Zukari, 1976 ;
Yeruham et al., 1985).
Lactivit semble positivement corrle la temprature moyenne et
ngativement
corrle lhumidit relative. Il ny a pas de corrlation
significative entre les prcipitations mensuelles et lactivit de
cette tique (Papazahariadou et al., 1995).
b) Activit saisonnire. De manire gnrale, la dynamique saisonnire
de R. bursa est unimodale, avec un seul
pic dactivit : lactivit des adultes prend place en saison chaude
(de mars septembre avec un maximum en juin) et celle des immatures
stend doctobre mars (Morel, 1981).
-
33
Ainsi, en Italie, les adultes sont actifs, de dbut mai fin aot.
Les larves et les nymphes qui sattaquent volontiers aux animaux
domestiques sont actives du mois de novembre fin janvier (Enigk,
1947).
De mme, en Isral, les immatures ont t trouvs pendant les mois
dhiver (novembre
mars) alors que les adultes sont apparus en avril et ont persist
jusquau dbut de juillet, comme en tmoigne la figure 12 (Yeruham et
al., 1989). Cette dynamique saisonnire unimodale est aussi retrouve
en Algrie (Yousfi-Monod & Aeschlimann, 1986) et en Syrie
(Liebisch & Zukari, 1976).
0
10
20
30
40
50
60
70
80
90
10 11 12 1 2 3 4 5 6 7 8 9 Mois
Nombre de tiques
(moyennes / 6 animaux) Larves
Nymphes
Adultes
Figure 12 : Distribution saisonnire de Rhipicephalus bursa sur
les moutons dIsral,
daprs Yeruham et al. (1989).
II.4.3. Rhipicephalus sanguineus (Latreille, 1806).
Nous nvoquerons cette espce que brivement car sa slectivit
vis--vis du Chien est tellement grande que le parasitisme des
bovins semble anecdotique en France. Sauvage lorigine, cette tique
a diversifi une souche domestique adapte tous les stades au Chien.
Cependant, la large rpartition de cette espce sur le territoire
franais nous interdit docculter la possibilit du parasitisme du
btail.
Signalons demble que les tudes antrieures au travail de Morel
& Vassiliade (1962), qui
voquent R. sanguineus (Latreille, 1806), confondent en fait
trois espces diffrentes : R. sanguineus, R. turanicus et R.
pusillus (Gilot, 1984).
II.4.3.1. Cycle biologique de Rhipicephalus sanguineus.
a) Cycle volutif. Le cycle est triphasique. Il est infod au
Chien et laccouplement a lieu sur lhte. Il se
droule en moyenne en quatre mois et demi, mais peut se rduire 63
jours dans les conditions optimales. Dans les chenils du nord-ouest
de lEurope, la ralisation du cycle biologique prend facilement une
anne (Boudeau, 1993b ; Hillyard, 1996).
Les adultes jeun peuvent survivre trs longtemps : jusqu 19 mois
(Bourdeau, 1993b).
-
34
b) Spectre dhtes. Le Chien est lhte prfr de tous les stades de
R. sanguineus : 90 % des tiques
collectes proviennent de chiens. Les autres collectes sont
issues de bovins, chats, renards et de lHomme. Les immatures
parasitent aussi les lapins, livres, hrissons et cureuils
(Hillyard, 1996).
En fait, le parasitisme des bovins est surtout important dans
les pays africains o laire
de rpartition de lespce concide mieux avec les zones dlevage du
btail quen France. Dans notre pays, cest le Chat qui est le deuxime
hte le plus frquent, du fait de la frquentation des mmes lieux que
le Chien (Gilot, 1984). Le btail parasit est en gnral celui qui
cohabite avec des chiens, comme cest le cas en Algrie, o des
adultes ont t rcolts sur lchine et la tte de bovins pturant dans
des zones frquentes par les chiens (Yousfi-Monod & Aeschlimann,
1986)
Mme si le parasitisme des animaux sauvages est rare, R.
sanguineus a t trouv sur
des cervids (Caeiro & Simes, 1991). Les cas de parasitisme
humain ne sont pas rares (Gilot & Marjolet, 1982).
II.4.3.2. Distribution gographique.
Lhabitat originel de cette espce est le Bassin Mditerranen et
lAfrique, mais il sest tendu dautres rgions du monde en suivant la
population canine. Dans le centre et le nord de lEurope, les
climats froids empchent la colonisation de lenvironnement, mais des
populations localises peuvent survivre au contact du Chien. R.
sanguineus est signal en France, Grande-Bretagne, Belgique,
Hollande, Danemark, Norvge, Pologne et Suisse (Hillyard, 1996).
En France, R. sanguineus est abondant dans le midi Mditerranen
et le sud-ouest,
mais il a dj t reconnu responsable dinvasions domiciliaires
Lyon, dans la rgion Parisienne, dans le Finistre, lIlle-et-Vilaine
et Strasbourg (Couatarmanach et al., 1989).
Concernant le Limousin, des collectes ont t effectues dans des
dpartements assez proches,
comme la Dordogne, le Lot-et-Garonne ou la bordure orientale du
Massif Central (Lamontellerie, 1965 ; Gilot et al., 1989). Cela
dit, en dehors du climat mditerranen, signalons que lespce est
toujours lie une forte urbanisation (Gilot et al., 1989).
II.4.3.3. Phnologie de Rhipicephalus sanguineus.
a) Biotopes coloniss. Le cycle biologique seffectue dans des
milieux artificialiss. Le lieu classique de
dveloppement de R. sanguineus est lhabitat du Chien, certaines
parties du cycle pouvant mme se raliser dans les appartements
(Gilot, 1984). Pour que le cycle puisse se drouler, la temprature
moyenne doit tre suprieure 18C et lhygromtrie de lordre de 50 % au
moins. Les habitations assurent donc une reproduction optimale de
lespce (Bourdeau, 1993b).
Cette espce a dailleurs un comportement principalement endophile
synanthrope : elle
ne semble pas coloniser les terriers des carnivores sauvages,
mais peut les infester lors de leur passage dans les biotopes
anthropognes traditionnels comme la ferme et ses dpendances. La
technique du drapeau semble par ailleurs peu efficace pour sa
dtection (utilisation possible du CO2) (Gilot, 1984 ; Gilot &
Aubert, 1985).
-
35
Sur la bordure orientale du Massif Central, lespce est observe
de 50 600 m daltitude, dans des biotopes vgtation mditerranenne ou
plus rarement subcontinentale : mme si le climat mditerranen est le
plus favorable, le climat tendance continental peut tre colonis
(Gilot et al., 1989).
Dans le centre de lItalie, Principato et al. (1989) ont trouv 70
% de R. sanguineus sur
les pturages herbeux de moutons transhumants. Sa prsence
abondante, dans les pturages durant tout lt jusquau dbut de
lautomne est lie la persistance, pendant ces mois, dun climat chaud
et sec, sujet la scheresse, et la prsence, outre dovins, de chiens
de berger infests.
b) Activit saisonnire. Dans le sud-est de la France, lactivit
des adultes semble maximale au printemps et au
dbut de lt (mois de juin). Cest en avril et en mai que les
infestations sont les plus fortes. Lactivit imaginale est
pratiquement nulle en hiver (Gilot, 1984).
En Italie, la dynamique saisonnire des adultes prsente deux pics
annuels : en avril
mai et en septembre octobre, mais elle reste la tique la plus
frquente en t. Les immatures ont t collects davril octobre
(Principato et al., 1989).
II.4.4. Rhipicephalus turanicus (Pomerantzev, Matikasvili,
Lototzki, 1940).
Cette espce semble beaucoup plus rare en France, que les espces
tudies prcdemment et il sagit dune espce dindividualisation rcente
(avant les travaux de Vassiliade & Morel, 1962, elle tait
confondue avec R. sanguineus). Cest une tique du btail, mais elle
est exclusivement mditerranenne (Gilot & Pautou, 1981).
II.4.4.1. Cycle biologique de Rhipicephalus turanicus.
a) Cycle volutif. R. turanicus prsente un cycle triphasique
ditrope, faisant se succder les repas des
immatures sur Rongeurs myomorphes de terrier et les repas des
adultes sur mammifres sauvages ou domestiques, de taille petite
(notamment le Hrisson), moyenne ou grande (Elfassy, 1993).
b) Spectre dhtes.
Slectivit des immatures : En France, la majorit des rcoltes a t
effectue sur les rongeurs et les insectivores :
Mus musculus, Apodemus sylvaticus, Rattus rattus, Pitymys
duodecimcostatus, Crocidura russula, Eliomys quercinus, Erinaceus
europaeus, Mustela nivalis
Slectivit des adultes : Voici le gradient daffinit dcroissante
dcrit classiquement :
1) artiodactyles (Bovin, Mouton, Chvre, Chameau, Porc, Sanglier)
; 2) carnivores (Chien, Renard) ; 3) prissodactyles (Cheval et ne)
; 4) rongeurs, insectivores (Hrisson) et lagomorphes (Livre).
-
36
Pourtant, en France, les bovins de la rgion mditerranenne ne
sont pas parasits car ils sont rares et quittent rarement ltable.
Seuls les taureaux camarguais semblent touchs. En revanche, ovins
et caprins sont parasits (Gilot & Pautou, 1981 ; Gilot et al.,
1989).
II.4.4.2. Distribution gographique (Gilot & Pautou,
1981).
Lespce est connue dans tous les pays bordant la Mditerrane. En
Europe, R. Turanicus ne semble pas pntrer dans les forts tempres de
feuillus
de la rgion holarctique. En France, lespce est absente du
domaine atlantico-europen et du domaine
mdioeuropen. Elle nest en fait connue que de la rgion
mditerranenne et les stations les plus septentrionales
correspondent la limite nord du Chne vert et de lOlivier (Gilot et
al., 1989).
Du fait de linadquation du climat, lespce est trs certainement
absente du Limousin.
II.4.4.3. Phnologie de Rhipicephalus turanicus.
a) Biotopes coloniss. Lespce colonise les biotopes mditerranens
constitus de groupements herbacs,
groupements landes ou groupements forestiers. Cest surtout une
espce de landes strate arbustive et herbace htrogne : landes Gent,
landes Romarin (Gilot & Pautou, 1981)
b) Activit saisonnire.
Lactivit des adultes est exclusivement printanire, tale du mois
de mars au mois de mai sur le btail ou sur la vgtation. Ils nont
jamais t collects en dautres saisons. Lactivit des larves et des
nymphes se situe en t, sur les rongeurs (Gilot & Pautou,
1981).
II.5. Les tiques du genre Hyalomma.
Ces espces seront tudies trs brivement du fait du nombre rduit
de leurs rcoltes et de leur caractre accidentel en France.
Il existe une trentaine despces de tiques appartenant au genre
Hyalomma, parasites
donguls en Eurasie et en Afrique (Bourdeau, 1993b). Mais ces
espces sont surtout exotiques et leur prsence en France est
rarissime. On peut cependant rencontrer H. detritum scupense
Schulze, 1919 (synonyme de H. scupense) et H. marginatum marginatum
Koch, 1844 dans le sud du Pays.
Il sagit de tiques mtastriata corps festonn, longirostres, avec
le deuxime article des
pdipalpes un peu plus long que le troisime. Leur taille est trs
importante en comparaison avec les espces habituelles dEurope. En
vue dorsale, le capitulum prend un aspect triangulaire, on
distingue des ocelles, mais le scutum est peu orn. Leur hanche I
possde une longue pine externe (Moulinier, 2003).
-
37
II.5.1. Hyalomma detritum scupense Schulze, 1919.
Certains auteurs comme Lamontellerie (1965) ou Macaigne &
Prez-Eid (1993) considrent H.
scupense Schulze, 1919 comme une espce part entire, trs peu
diffrente de lespce H. detritum Schulze, 1919. Morel, lui, ne
considre quune seule espce H. detritum Schulze, 1919 avec deux
sous-espces H. detritum detritum et H. detritum scupense.
Ainsi H. scupense et H. detritum scupense sont-ils
synonymes.
Cycle biologique. Le cycle est monoxne, alors que H. detritum
detritum a un cycle dixne (Macaigne &
Prez-Eid, 1993 ; Morel, 2000). Lclosion des larves a lieu de 45
68 jours aprs le dbut de la ponte (Lamontellerie, 1965). Cest
essentiellement une tique des bovins, mais elle parasite galement
les moutons et les chevaux (Couatarmanach et al., 1989).
Aire de rpartition. Sa rpartition gographique, mditerranenne,
couvre toute lAfrique au nord du Sahara,
mais aussi la Bulgarie, la Yougoslavie, la Grce, lEspagne et la
Turquie (Couatarmanach et al., 1989). Sa prsence dans le sud-ouest
de la France tait sujette discussion jusquaux travaux de Macaigne
& Prez-Eid (1993).
En France, il semble que les spcimens collects par
Lamontellerie, en Charente-Maritime, Gironde, Landes et
Basses-Pyrnes appartiennent bien lespce H. scupense (Lamontellerie,
1965 ; Macaigne & Prez-Eid, 1993). Un adulte a aussi t trouv
dans la Sarthe sur un cheval (Morel, 1961). Dans notre pays, sa
distribution est donc trs limite et restreinte au sud-ouest.
Elle a t observe sur un enfant, en juin 1985 Rennes. Il sagirait
cependant dun cas dimportation d un tapis achet en Algrie
(Couatarmanach et al., 1989).
Phnologie Dans le sud-ouest de la France, les adultes sont
actifs de fvrier avril et les nymphes
en fvrier et mars : il sagit donc dune espce de saison froide.
La ponte a lieu de fin mars fin mai, le maximum des pontes ayant
lieu fin mai (Lamontellerie, 1965). La vgtation colonise correspond
en principe aux tages du Chne pubescent et du Chne tauzin (Morel,
1981).
II.5.2. Hyalomma marginatum marginatum Koch, 1844.
Cette espce a pour synonyme Hyalomma savignyi (Gervais,
1844).
Cycle biologique (Hillyard, 1996). Il sagit dun cycle biologique
dixne. La larve se nourrit sur un hte, un oiseau en
gnral, et se mtamorphose en nymphe sur lanimal, la nymphe se
nourrit alors sur le mme hte. Les adultes parasitent un second hte,
le plus souvent un grand mammifre domestique (surtout les Onguls).
Les stades adultes nont jamais t signals en Europe du nord-ouest.
Les repas sanguins et le dveloppement des larves et des nymphes sur
les oiseaux durent de 12 26 jours. Ces tiques peuvent donc tre
transportes sur de longues distances, car le cycle tant dixne, les
larves restent sur lhte.
-
38
Rpartition gographique (Morel, 1981 ; Hillyard, 1996). H. m.
marginatum est prsent au sud de lEurope, au sud de lAsie et surtout
en Afrique.
En Europe du nord-ouest (Grande-Bretagne, France, Danemark et
Norvge), toutes les collectes font rfrence des immatures issus
doiseaux migrateurs et les adultes sont absents. En clair, lespce
nest pas tablie en France. Elle est en revanche prsente en Italie
(Principato et al., 1989) et au Portugal (Caeiro & Simes,
1991).
Phnologie (Liebisch & Zukari, 1976 ; Morel, 1981 ; Bouattour
et al., 1999). Les rcoltes en France ne sont qupisodiques, et
concident avec les arrivages
printaniers doiseaux migrateurs venus de lAfrique mditerranenne.
Cette espce reste donc une tique dimportation, incapable de raliser
lensemble de son cycle sur notre territoire. En Afrique, elle est
capable de sadapter de nombreux climats allant du dsert Saharien
avec moins de 100 mm de prcipitations par an aux hautes montagnes
avec des prcipitations suprieures 1000 mm par an. Elle y est active
au printemps et lautomne.
Au bilan de cet inventaire sur le territoire franais, nous
souhaitons insister sur son
caractre labile. En effet, il nest que le reflet dune situation
ponctuelle et susceptible dvoluer. Mme si les exigences des tiques
vis--vis des conditions microclimatiques et leur dpendance par
rapport aux htes disponibles restent des obstacles, les mouvements
du btail sur de longues distances (bateau, avion) et les htes
migrateurs intervenant dans le cycle sont des lments favorables aux
propagations. Les cas de dispersion anthropique de nouvelles espces
de tiques dun continent lautre sont autant dexemples dmonstratifs
(Rodhain, 1996 ; Morel, 2000).
Tout dabord, citons le cas dAmblyomma variegatum, tique dAfrique
tropicale qui sest
implante aux Antilles franaises et en Guyane suite limportation
de bovins sngalais. La colonisation a t trs lente mais sest
brutalement acclre ces 25 dernires annes. Et des cas de cowdriose
et de dermatophilose bovine en Guadeloupe ont conduit suspecter sa
prsence aux Carabes (Rodhain, 1996).
Plus proche de nous, lespce Boophilus annulatus, qui est une
tique dAfrique centro-
occidentale semble avoir colonis lEurope partir de la pninsule
Ibrique (Caeiro, 1999 ; Morel, 2000). En Sardaigne, lespce
reprsentait 0,7 % des tiques rcoltes par Todaro et al. (1999). En
Corse, B. annulatus est frquent, et mme si le reste de la France
mtropolitaine en est exempte, sa propagation est toujours possible
(Uilenberg, 2000, in Mage et al., 2001).
-
39
III. METHODES DETUDES DES POPULATIONS DE TIQUES.
III.1. Recherche des tiques sur les animaux.
III.1.1. Sur les animaux domestiques : exemple des bovins.
Mme si lactivit saisonnire des tiques est bien connue dans de
nombreuses rgions
dEurope de louest, peu dtudes se sont intresses la dynamique
saisonnire de linfestation des bovins eux-mmes. Quelques-unes ont
nanmoins t conduites au Pays-de-Galles (Edwards & Arthur, 1947
; Arthur, 1948 ; Evans, 1950), en Suisse (Gern & Brossard,
1986) et en France, dans le dpartement de la Sarthe (LHostis et
al., 1996a). La plupart de ces comptages sur bovins concernent
Ixodes ricinus et considrent un nombre restreint de bovin