Embed Size (px)
Citation preview

27
3 Résultats
3.1 Introduction Le cycle de vie d’une plante abouti toujours à une multiplication des graines à partie d’une seule. Mais que se passe-t-il entre le semis et la formation des prochaines graines? . Nous savons que, pendant ce temps, la plante vit, elle augmente de taille, elle présente des changements morphologiques, physiologiques et biochimiques. Le poète scientifique Johann-Wolfgang Goethe dans l’ouvrage intitulé « La métamorphose des plantes » écrit en 1790, décrit le cycle de vie d’une plante idéal de la façon suivante: Dans le sec le noyau garde vie immobile, adonné à l’humidité douce, il se gonfle et se tend, et s’élève aussitôt de la nuit qui l’entoure; mais quand il apparaît, la forme en reste simple, dans les plantes aussi, c’est de l’enfant le signe. Tout aussitôt se dresse une pousse suivante, ajoute nœud à nœud, renouvelle la prime force, non, certes, toujours la même; car la feuille suivante est toujours, tu le vois, mieux formée, plus variée, plus étendues, plus échancrées,mieux séparées en pointes et parties, celles qui reposaient mêlées en l’organe inférieur. Enfin elle culmine en perfection précise […] Bientôt la forme montre de plus doux effets[…] La hampe de la tige se forme plus parfaite. Mais sans feuilles, très vite, le pédoncule s’élève et le regard découvre une merveille. Tout autour en un cercle, comptées et innombrables, la feuille plus petite se joint à sa semblable. Pressé autour de l’axe, le calice protecteur se révèle, lançant vers la suprême forme corolle colorées […] les plus tendres des formes apparaissent en double, destinées à s’unir.[…] Hymen descend sur eux et des vapeurs splendides versent de doux parfums vivifiant l’alentour. Des germes bien distincts s’enflent alors sans nombre, au sein de fruits renflés cachés et protégés, nature clôt l’anneau des forces
éternelles; mais en cycle nouveau tôt succède à l’ancien, pour que se continue à tout jamais la chaîne, et que tout et partie soient animés de vie.
3.2 Cycle de vie d’Arabidopsis thaliana
La taille moyenne d’une graine sèche d’Arabidopsis thaliana est de 0.3 mm x 0.5 mm comme il est montré dans la figure 4. Ces valeurs ont été obtenues en faisant une moyenne sur 5 graines du même écotype (Ws) récoltées en novembre 2000. Figure 4. Calcul de la surface de la graine sèche d’Arabidopsis thaliana écotype Ws. Les photographies avec la règle millimétrée ont été prises par le Dr M. Crèvecoeur du laboratoire d’Histologie et Cytologie Végétales de l’Université de Genève. Les figures 5 et 6 permettent d’avoir une vison générale du cycle de vie d’Arabidopsis thaliana. Dans nos conditions (voir Matériel) la plante nécessite environ 65 jours pour effectuer le cycle complet (obtention des nouvelles graines).
area ≈ 480110 pixelarea = 182267 pixel0.2mm*0.2mm=0.04mm2
Surface moyenne un mm2 de la graine sèche:
area = 480110/182267*0.04 ≈ 0.1mm2

28
Figure 5. Cycle de vie d’Arabidopsis thaliana (écotype Ws) en condition de 12 h de lumière et 12 h d’obscurité. Chaque photo est espacée par un intervalle d’une semaine. Dans la photo de fond, Arabidopsis a atteint l’âge de 50 jours.
29 jours
8 jours 15 jours
22 jours
36 jours
43 jours
cotylédonsradicule
1 cm
29
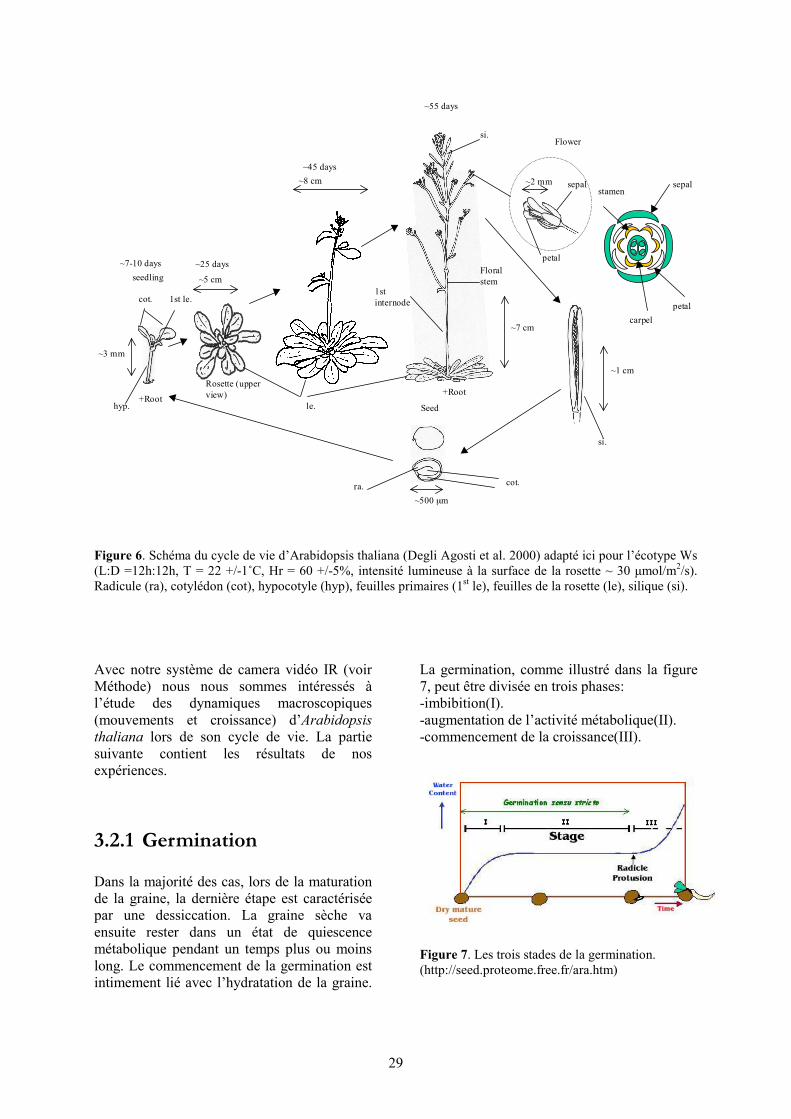
Figure 6. Schéma du cycle de vie d’Arabidopsis thaliana (Degli Agosti et al. 2000) adapté ici pour l’écotype Ws (L:D =12h:12h, T = 22 +/-1˚C, Hr = 60 +/-5%, intensité lumineuse à la surface de la rosette ~ 30 μmol/m2/s). Radicule (ra), cotylédon (cot), hypocotyle (hyp), feuilles primaires (1st le), feuilles de la rosette (le), silique (si). Avec notre système de camera vidéo IR (voir Méthode) nous nous sommes intéressés à l’étude des dynamiques macroscopiques (mouvements et croissance) d’Arabidopsis thaliana lors de son cycle de vie. La partie suivante contient les résultats de nos expériences.
3.2.1 Germination Dans la majorité des cas, lors de la maturation de la graine, la dernière étape est caractérisée par une dessiccation. La graine sèche va ensuite rester dans un état de quiescence métabolique pendant un temps plus ou moins long. Le commencement de la germination est intimement lié avec l’hydratation de la graine.
La germination, comme illustré dans la figure 7, peut être divisée en trois phases: -imbibition(I). -augmentation de l’activité métabolique(II). -commencement de la croissance(III). Figure 7. Les trois stades de la germination. (http://seed.proteome.free.fr/ara.htm)
~3 mm
~5 cm
~8 cm
~7 cm
~2 mm
~1 cm
~500 μmra. cot.
Seed
seedling
Rosette (upperview)
stamensepal
petalcarpel
si.
si.Flower
hyp.
cot.
le.
1st le.
Floralstem
1st internode
~7-10 days ~25 days
~45 days
~55 days
+Root+Root
sepal
petal

30
Figure 8. Schéma général des facteurs qui contrôlent la germination d’Arabidopsis (Bentsink et Koornneef 2002). GA=gibbérellines, ABA= acide abscissique, BR= brassinostéroides. La germination sensu stricto commence par l’imbibition et fini par la sortie de la radicule et l’élongation de l’axe de l’embryon (Bewley 1997). Une analyse protéomique de la germination pour Arabidopsis a permis de mettre en évidence des nouvelles protéines associées aux différents stades de la germination (Gallardo et al. 2001), tandis que une analyse génétique a mit en évidence l’existance de locus (ex. comatose) responsables de la réduction du potentiel de germination (Holdsworth et al. 2000). L’état des connaissances sur les mécanismes moléculaires impliqués dans la germination
d’Arabidopsis est résumé dans l’article écrit par Bentsink (voir figure 8). Des animation pour le stade III peuvent être trouvées sur les deux sites web suivants: http://www.bio.warwick.ac.uk/millar/Video/md5banw3.gif,http://sunflower.bio.indiana.edu/~rhangart/plantmotion/earlygrowth/germination/arabidopsis/atgermination.html. Nous savons que le stade III aboutit à la formation de la plantule, dont les cotylédons présentent un rythme circadien (Engelmann et al. 1992). Le moment de la naissance de ce rythme à notre connaissance n’a pas été caractérisé. Nous avons donc effectué une

31
expérience pour définir le commencement du rythme circadien des cotylédons d’Arabidopsis. Un support a été construit pour cette expérience (figure 9) sur lequel une graine, n’ayant pas subi de vernalisation précédemment, a été positionnée. Figure 9. Support pour l’étude de la germination Des photos digitales prises tous les 15 min ou 10 min nous ont permis de suivre la dynamique de croissance et des mouvements à partir du commencement du stade III (pour Arabidopsis il faut compter environ 3 jours après le commencement de l’imbibition). Nous avons ainsi obtenu des animations (voir CD: Germ1.avi et Germ2.avi). Le calcul de l’évolution de la surface a été obtenu à l’aide du programme area.m (voir 2.3.3.3.1 Programmes). Les résultats sont présentés dans la figure 10. Trois phases on été mise en évidence: -phase 1: croissance de la radicule. -phase 2: sortie des deux cotylédons de la graine. -phase 3: plantule complète, commencement du rythme circadien des deux cotylédons. Par nos observations nous pouvons dire que les cotylédons commencent à se lever verticalement pendant la nuit et à se positionner horizontalement pendant le jour,
une fois que l’hypocotyle se trouve en position verticale. Les changements ou décroissances rapides de la surface sont dû à des mouvements « rapides » de la radicule ou des cotylédons. Il est important de noter que la radicule a la capacité de pénétrer un petit peu à l’intérieur du filtre à charbon et sa position par rapport aux sources lumineuses de 950 nm peut varier. Ces événements impliquent une variation de la luminosité des pixels qui peuvent ne plus être détectables par la valeur du seuil (filtre) choisi dans le programme area.m.
Boite de Petriremplie d ’eau
Filtre de charbon qui mouilledans l ’eau et pemet de mantenirhumide le support sur lequel estposée la graine.
Support couvert d ’un filtre à charbon sur lequelse trouve la graine

32
Figure 10. Dynamique d’éclosion de la plantule d’Arabidopsis thaliana (voir CD Germ1.avi). La surface (mm2) a été calculée à l’aide du programme area.m. Intervalle de temps entre chaque photo=15 min.
0
0.2
0.4
0.6
0.8
1
0 1 2 3 4 5 6
1
23
Temps (jours)
mm2
Détachementdeuxième cotylédon
Mouvement circadien des cotylédons
Détachementpremier cotylédon
Enroulement de la racine
Sortie de la radicule
Enveloppe de la graine

33
3.2.2 Plantule Dans la littérature on trouve quatre types de dynamiques qui ont été décrites pour la plantule d’ Arabidopsis thaliana: 1) circumnutation de la radicule. 2) circumnutation de l’hypocotyle. 3) croissance circadienne de l’hypocotyle. 4) mouvement circadien des cotylédons. Nous n’avons pas effectué une expérience pour observer les mouvements de circumnutation de la radicule (figure 11 et CD root_movie.avi). Des mutants senestrogire (1-6C) et des mutants qui présentent les deux sens de rotation (aux-1) ont été trouvés pour Arabidopsis (Marinelli et al. 1997). Figure 11. Mouvement ondulatoire effectué par la radicule d’Arabidopsis thaliana posé sur une plaque d’agar incliné (30-60˚). http://www.oeb.harvard.edu/faculty/holbrook/projects/arabidopsis/arabidopsis.htm. Pour les mouvements de circumnutation et micronutation de l’hypocotyle (Schuster et Engelman 1997) nous avons effectué deux expériences, pour voir si l’écotype Ws présente aussi ce type de dynamique. Expérience 1(voir CD cotysoustraction.avi) -Total time = 24 h (durée de l’expériance). -Intervalle = 2 min (dt entre deux photos). -Stade de la plantule d’Arabidopsis: ~ 7 jours du semi. -Camera objectif: macro. Expérience 2 (figure 12) -Total time = 3 j (durée de l’expériance). -Intervalle = 5 min (dt entre deux photos).
-Stade de la plantule d’Arabidopsis: les deux premières feuilles commencent à êtres visibles (~ de 8 à 11 jours du semi). -Camera objectif: macro. Les images (vue du dessus) ont été traitées avec le programme centregravity.m (voir 2.3.3.3.1 Programmes) pour détecter les mouvements du centre de gravité (voir CD cotycentregravity.avi). Comme on peut le voir dans la figure 12 les déplacements du centre de gravité forment une maille difficilement compréhensible. Ce sont des très petits déplacements (amplitude maximale 0.5 mm) qui gravitent autour d’un attracteur. Ces mouvements sont affectés par le mouvement circadien des cotylédons (voir figure 12 graphique(b) coordonnée y). Pendant la phase diurne du quatrième jour (voir graphique a figure 13) nous avons remarqué des variations rapides de la surface des cotylédons. Cette variation est visible aussi dans la coordonnée x du graphique (b) de la figure 12. Par analyse plus détaillée de cette zone nous avons pu démontrer que ces variations correspondent surtout à des mouvements irréguliers avec des micronutations (terme inventé par Heathcote 1966) probables. Des mouvements de circumnutation de l’hypocotyle ont aussi été détectés (voir graphique (b) figure 13). Par l’analyse de fréquence (voir figure 21) de ces oscillations ultradiennes nous avons trouve une période d’environ 2h. Des études effectuées sur la circumnutation d’Arabidopsis ont observé que la majorité des hypocotyles se déplacent dans l’espace avec irrégularité et que seulement 10% effectuent des mouvements de circumnutation (Orbovic et Poff 1997). En plus il semble que la lumière bleue stimule les plantules à se courber vers la source de lumière (phototropisme), tandis que la lumière rouge aurait un effet d’inhibition sur la circumnutation (Orbovic et Poff 1997). Nous n’avons pas effectué d’autres expériences pour approfondir ce type de dynamique. Il est en tout cas important de ne pas oublier qu’à ce stade la structure de la plante est encore fragile et simple.

34
Figure 12. Le graphique (a) montre les mouvements du centre de gravité (coordonnées x y) calculé à l’aide du programme centregravity.m en utilisant des images (vue du dessus) espacées de 5 min. Dans le graphique (b) on peut voir la corrélation entre les oscillations de l’évolution de la surface des cotylédons (voir figure 13) et les déplacements des coordonnées y et x. Les oscillations rapides de la coordonnée x correspondent à des mouvements de circumnutation de l’hypocotyle (pour plus de détails voir figure 13 et 20). En effet l’élongation de l’hypocotyle a été pendant longtemps reconnu comme étant particulièrement sensible aux facteurs endogènes et environnementaux. Des composés biochimiques tels que les gibbérellines ou les auxines sont souvent associés à une augmentation de l’élongation tandis que les cytokinines, l’acide abscissique et l’éthylène avec une inhibition de la croissance. Notre expérience nous a permis d’observer des variations rythmiques circadiennes de la vitesse de croissance de l’hypocotyle d’Arabidopsis pour l’écotype Ws. Comme on peut le voir dans la figure 13 l’hypocotyle croît surtout le jour (Δx) quand les cotylédons sont en position horizontale et ne croît quasiment pas la nuit. Par nos mesures nous avons pu estimer une vitesse maximale de 0.6 mm/h pour la croissance de l’hypocotyle. Des travaux effectués sur l’élongation de l’hypocotyle d’Arabidopsis (écotypes: Columbia gl1, C24 et La(er)) ont observé l’inverse: c’est quand les cotylédons se trouvent en position verticale (nuit) que la
vitesse d’élongation de l’hypocotyle est maximale (Dowson et Millar 1999). Des gènes responsables de la diminution de la période (toc1-1 mutant) ou qui inhibent l’arrestation de la croissance diurne de l’hypocotyle (elf3) permettent de confirmer l’hypothèse que l’horloge moléculaire circadienne qui contrôle le mouvement des cotylédons est impliqué aussi dans le contrôle de l’élongation rythmique de l’hypocotyle d’Arabidopsis thaliana (Dowson et Millar 1999). Comme le mouvement rythmique des cotylédons aussi le rythme d’élongation de l’hypocotyle fait partie des premier rythmes circadiens que l’on peut observer au cour du cycle de vie d’Arabidopsis thaliana. Le rythme circadien semble jouer un rôle très important pour l’économie de la plante. Dans la partie suivante nous allons essayer de suivre l’évolution du rythme circadien le long de la vie de la plante.
1.5
1.6
1.7
1.8
1.9
2
2.1
2.2
2.3
1 1.1 1.2 1.3 1.4 1.5 1.6
Coordonnée x (mm)
Coo
rdon
née y
(mm
)
1
1.5
2
2.5
3
3.5
4
4.5
0 1 2 3 4 5
Surf
ace d
es c
otyl
édon
s (m
m2 )
coor
donn
ées x
y (m
m) Surface des cotylédons
Coordonnée y
Coordonnée x
Attrac teur
Temps (jours)
1.5
1.6
1.7
1.8
1.9
2
2.1
2.2
2.3
1 1.1 1.2 1.3 1.4 1.5 1.6
a
b

35
Figure 13. Plantule vue latéralement et du dessus, du 9-12-02 (11:21h) au 14-12-02 (12:49) semis le: 2-12-02 (14:25h). La longueur de l’hypocotyle a été calculée avec l’outil mesure de Photoshop en utilisant des photos (vue latérale) espacées de 3h. Le Δx correspond à la croissance de l’hypocotyle en trois heures. Le calcul de la surface a été obtenu à l’aide du programme area.m, avec des photos (vue du dessus) espacées de 5 min. Dans cette figure nous pouvons voir la relation entre la vitesse de croissance de l’hypocotyle et la position des cotylédons. L’hypocotyle croît principalement pendant que les cotylédons se trouvent en position horizontale, cela correspond avec la phase lumineuse. Les mouvements rapides pendant la phase diurne du quatrième jour (graphique a), correspondent à des mouvements irréguliers et de circumnutation de l’axe de l’hypocotyle. Le graphique b montre la trajectoire d’un mouvement de circumnutation (Δx=0.25mm, Δy=0.12mm et Δt=2h10) qui se trouve à l’intérieur de la trajectoire totale (voir figure 12). Les résultats de l’analyse de fréquence de ces mouvements sont visibles dans la figure 21.
1
1.5
2
2.5
3
3.5
4
4.5
0 1 2 3 4 5
Long
er d
e l ’h
ypoc
otyl
e (m
m)
Temps (jours)
Surf
ace
des c
otyl
édon
s (m
m2 )
Δ x
2
2.02
2.04
2.06
2.08
2.1
2.12
2.14
2.16
1.1 1.15 1.2 1.25 1.3 1.35 1.4 1.45
Coo
rdo n
n ée s
y (m
m)
Coordonnées x (mm)
a
b

36
3.2.3 Rythme circadien La classification physique des rythmes est basée sur la longueur de la période d'oscillation. On distingue donc les rythmes : Domaine :...............................………..Période Ultradien :……………………………….<20 h Circadien :…………………..………24 +/- 4 h Infradien :………………………….……>28 h Circaseptien:…………………….…….7 +/- 3 j Circadiseptien :……………………...14 +/- 3 j Circavigintien :………………………21 +/- 3 j Circatrigintien :……………………...30 +/- 5 j Circannuel :………………….…….1 a +/- 2 m h = heures ; j = jours; m = mois ; a = année Parmi ces rythmes, les plus étudiés en biologie sont les rythmes circadiens (du latin circa: autour et dies: jour ) ou nycthéméraux. Les horloges circadiennes génèrent des rythmes dont la période est d’environ 24 heures. Elles sont présentes chez la plupart des organismes vivants et interviennent dans le contrôle de fonctions physiologiques variées. Alors que leur période est génétiquement déterminée, leur phase est sensible aux stimuli environnementaux tels que la lumière et la température, ce qui permet leur entraînement.
Figure 14. Schéma qui montre que les rythmes sont sous le contrôle d’une horloge interne sensible aux facteurs externes.
La nature intracellulaire des oscillateurs circadiens, et les mécanismes moléculaires (feedback) qui les régissent commencent à être décryptés (Cermakian 2001). Les approches génétiques développées chez la drosophile et plus récemment chez la souris ont conduit à la caractérisation de gènes d’horloge largement conservés au cours de l’évolution (gènes clock). Pour les végétaux les premiers modèles moléculaires capables d’expliquer partiellement ces phénomènes rythmiques sont apparus (McClung et al. 2002, Yonovsky et al. 2003). Nous connaissons actuellement quelques gènes dont l’activité oscille de façon rythmique (Millar 1999, Alabadi et al. 2002). Des premières tentatives d’analyse d’une partie du trancriptome pour Arabidopsis thaliana (Davis et Millar 2001) ont montré que nombreux gènes impliqués dans la transduction de signaux et dans diverses voies métaboliques sont réglés par des rythmes circadiens et/ou par la lumière.
3.2.3.1 Mouvement circadien des feuilles d’Arabidopsis thaliana
Les premiers écrits concernant la description des rythmes circadiens remontent au 4ème siècle avant J.-C. Durant les marches de l'armée d'Alexandre le Grand, le scribe Androsthènes décrivit le comportement nycthéméral des feuilles du tamarinier: celles-ci s'ouvraient le jour et se refermaient la nuit, mais ce rythme fut interprété comme une simple réponse passive à un environnement cyclique. Cette idée fut acceptée pendant de nombreux siècles durant. Nous connaissons actuellement quelques plantes dont les feuilles se lèvent et se baissent avec une rythmicité circadienne d’origine endogène: (ex. Phaseolus multiflorus Bünning 1967, Spinacia oleracea Degli Agosti 1985, Arabidopsis thaliana Engelman et al. 1992). De nombreuses autres plantes positionnent les feuilles horizontalement pendant le jour (surface maximale) et les lèvent ou les baissent pendant la nuit (voir Oxalis animation: http://sunflower.bio.indiana.edu/~rhangart/plantmotion/movements/leafmovements/oxalis/oxalis.html). Ce mouvement est le résultat du

37
changement de pression hydrostatique (turgescence) dans des régions plus ou moins caractérisées selon l’espèce. Pour Phaseolus vulgaris nous savons que le pulvinus (caractéristique des Fabacée) est l’organe qui engendre le mouvement des feuilles. Pour Arabidopsis thaliana les choses sont un peu moins claires. Nous n’avons pas de pulvini, mais comme le montre la figure 16 le mouvement des cotylédons peut être décomposé en deux angles (α, β), dont l’angle β est celui qui présente la plus grande variation. Par cette expérience nous avons pu démontrer que le mouvement circadien des cotylédons pour l’écotype Ws est principalement localisé dans la région entre le limbe et le pétiole. Il est possible que ces changements de l’angle des feuilles soient causés par une élongation asymétrique de la région que nous venons de caractériser (Millar 1999). Des animation sur ce type de mouvement peuvent être vues sur le web:http://www.dartmouth.edu/~rmcclung/MoreMovies.html ou dans le CD compilé pour ce travail (cotymov.avi). Ce rythme on le retrouve tout le long de la vie de la rosette (voir figure 17, 18 et 19 et dans le CD Circadien.avi). Des observations faites par d’autres auteurs sur les mouvements rythmiques des cotylédons ont pu mettre en évidence l’origine endogène de ce mouvement (Engelman et al. 1992, Degli Agosti et al. 2000). Nous avons effectué une expérience sur une rosette de 4 feuilles + 2 cotylédons jusqu’à 6 feuilles + 2 cotylédons pour voir si en lumière continue les feuilles se lèvent et se baissent toujours. Par analyse de l’évolution de la surface de la rosette vue du dessus nous avons obtenu le graphique 1 de la figure 20. Comme pour les cotylédons le mouvement est maintenu en lumière continue, mais nous observons l’apparition d’un rythme ultradien pendant l’hypothétique phase nocturne (voir CD feuilleultradien.avi). Déjà chez l’hypocotyle de la jeune plantule (voir figure 12 et 13 ) on avait rencontré des mouvements ultradien, mais pendant la phase diurne. Après avoir choisi la fenêtre d’intérêt à l’intérieur de notre signal et soustrait la courbe de tendance nous avons effectué une analyse de fréquence à l’aide de la première partie du programme timefreq.m (voir 2.3.3.3.1 Programmes). Ce programme effectue d’abord une analyse de Fourier du signal en question et
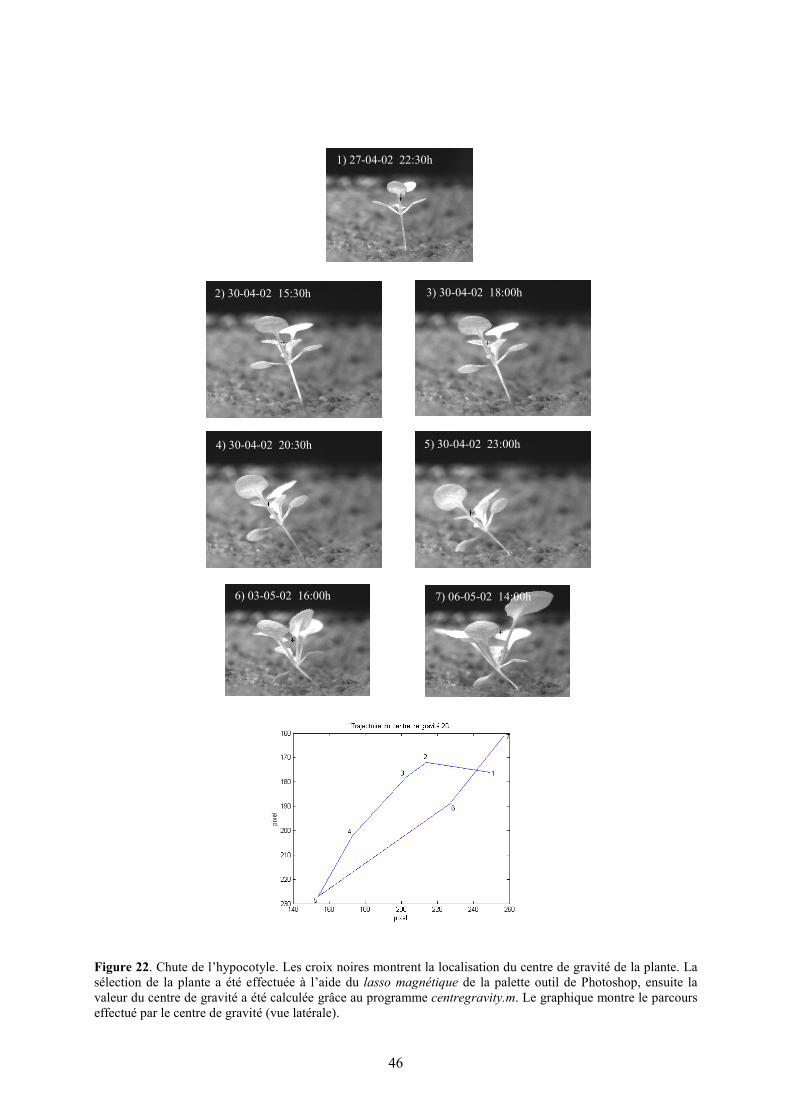
en suite par la sélection de/des fréquences plus importantes reconstruit un signal, ce qui permet une comparaison visuelle avec le signal original. Par nos mesures nous avons pu déterminer une période principale endogène de ~ 25,5h pour les mouvements circadiens des feuilles en lumière continue (voir graphique 2 figure 20). Cela signifie que la période endogène serait un peu plus grande du cycle jour nuit de 24h. Pour les oscillations ultradiennes des feuilles l’analyse a permit de déterminer une période de ~ 1.8h (voir figure 21). Des études effectuées sur les mouvements des feuilles d’Oxalis regnellii ont permit de détecter l’apparition de rythmes ultradiens, dont l’origine reste encore inconnue, avec un probable lien avec l’intensité lumineuse (Pedersen et al. 1993). Il est intéressant de remarquer que ces oscillations ultradiennes apparaissent et disparaissent avec une rythmicité circadienne. La présence de rythme ultradien avec une régulation cicadienne a déjà été observée dans d’autres plantes. La théorie du moment de singularité dit que si un traitement x est donné à un moment t il se peut qu’on observe l’arrêt d’un rythme. Des expériences faites sur les feuilles du haricot ont permit d’annuler le rythme circadien par un moment de singularité et observer une persistance du rythme ultradien (Kayali 1995). Les interconnections entre les rythmes reste encore un domaine obscur, mais sûrement de grand intérêt. Si l’on continue dans l’étude du cycle de vie d’Arabidopsis on constate qu’un changement des oscillations régulières est observable dans la figure 18. Comme il est indiqué dans la figure cet événement coïncide avec la chute de l’hypocotyle. Une vue latérale avec la dynamique du centre de gravité (croix noires) permet de mieux visualiser ce phénomène (figure 22). La chute dure environ 7 ½ h. La plante a besoin ensuite d’environ 7 jours pour retrouver un nouvel équilibre. La trajectoire du centre de gravité qui est visible sur le graphe de la figure 22, pourrait faire supposer que le centre retourne au même point de départ, une vérification avec la vue du dessus (pas illustré) permet de compléter l’information de la vue latérale. Nous avons donc pu conclure que la plante effectue une trajectoire en forme de demi-ellipse. Considérant la durée de temps (jours) de ce phénomène (Collings et al. 1998), nous pouvons supposer que le gravitropisme

38
pourrait être impliqué dans cette dynamique. En effet la croissance dirigée de certains organes (tige, racine) selon le sens de la force de gravitation, suppose qu'il existe un ou des systèmes pouvant percevoir le stimulus gravitropique au sein de la plante (Chen et al 1999). Dans les tiges, il semble que le stimulus gravitropique soit perçu au niveau de cellules présentes dans l'endoderme de la zone d'élongation de la tige (Fukaki et al 1998). Ces cellules, appelées statocytes tout comme celles contenues dans la coiffe racinaire, contiennent de volumineux grains d'amidons qui sédimentent sous l'effet de la pesanteur, et qui sont donc aussi appelés statolithes. Cette sédimentation déclencherait l'activation de transporteurs membranaires actifs de l'auxine. L'action de ces transporteurs créerait un gradient d'auxine entre la face inférieure et supérieure de la tige. Les concentrations d'auxine présentes dans la tige correspondent, par opposition à ce qui existe dans la racine, à la zone où une augmentation de concentration d'auxine augmente l'élongation cellulaire. Des autres modèles de perception de la gravité par les plantes, font intervenir le rôle du cytosquelette (Buluska et Hasenstein 1997), les changements des zones de plus grande tension de la membrane plasmique qui induisent une activation des strech activated ion channel , ou les variation de la polarité du mouvement cytoplasmique (Staves 1997). Des gènes dont la mutation affecte la réponse gravitropique ont été identifiés (Masson et al. 2002). Dans le cas des tiges, tout comme pour les racines, de nombreuses incertitudes demeurent quant aux voies de transduction et de transmission intervenant dans la réponse gravitropique. Le phototropisme pourrait aussi être impliqué, en effet les feuilles retrouvent leur position parallèle par rapport à la source de lumière. Nous n’avons pas effectué des expériences ultérieures pour caractériser de façon plus détaillée la dynamique de la chute. Un autre sujet dont nous nous sommes intéressées lors de l’étude du cycle de vie d’Arabidopsis est illustrée dans la figure 23. Comme nous l’avons déjà vu, le rythme circadien des feuilles est présent tout le long de la vie de la rosette. A l’aide des animations nous avons pu voir que les feuilles diminuent l’amplitude de leur mouvement avec l’âge. Nous avons donc essayé d’analyser plus en
détail la durée de vie de ce mouvement rythmique d’un des cotylédons et des premières quatre feuilles. Premièrement, nous avons constaté que le mouvement rythmique circadien apparaît lors- que la feuille atteint une certaine taille. Deuxièmement, (bien visible dans la courbe des oscillations rythmiques du cotylédon1) nous observons après un certain temps, une diminution de l’amplitude du mouvement. On peut donc schématiser la vie d’une feuille dans les trois phases théoriques suivantes: -phase 1: croissance sans mouvements. -phase 2: croissance avec mouvement -phase 3: diminution du mouvement et de la croissance. Des études sur le mouvement rythmique des feuilles primaires de l’épinard (Degli Agosti et Greppin 1988) on permit d’observer la disparition du mouvement a 4 ½ semaines, c’est-à-dire lorsque les feuilles secondaires sont dans leur phase de croissance maximale et qu’apparaissent déjà les feuilles tertiaires. Cette observation a permit de formuler l’hypothèse que la disparition du mouvement serait probablement du à un empêchement physique lié à l’anatomie de la plante (rosette). Dans la figure 23 on peut observer que la diminution du mouvement du cotylédon (1) correspond avec l’apparition de la troisième et quatrième feuille. Cette observation pourrait alors renforcer l’hypothèse de l’empêchement physique vu que l’axe de croissance de ces feuilles est très proche à l’axe de croissance des deux cotylédons (voir image figure 23). L’architecture de la rosette joue très probablement un rôle important dans sa dynamique. Dans la majorité des plantes à fleur les feuilles sont disposées en spirale, et l’angle entre chaque nouvelle feuille et la feuille précédente est très souvent proche à l’angle de Fibonacci de 137.5 ° (Jean 1994).
Spiral (n+3)
Spiral ( n+5) Spiral génératrice

39
Figure 15. Schéma d’une coupe transversale d’un méristème apical avec les trois spirales dessinées (Callos et Medford 1994). La disposition radiale des feuilles est connue sous le terme de phyllotaxie (phyllo = feuilles, taxie = ordre). La figure 15 illustre la distribution des bourgeons foliaires autour du méristème apical. Trois spirales sont mises en évidence, la spirale génératrice représente l’ordre de formation des organes. Pour Arabidopsis le sens de rotation de la spirale génératrice est maintenu le long du développement végétatif et peut être horaire ou antioraire en égale fréquence (Callos et Medford 1994). Les évènements qui déterminent le sens ne sont actuellement pas encore connus. Des études récentes (Reinhardt et al. 2000) ont permis de mettre en évidence la nécessité de l’auxine dans la régulation de l’initiation de la position radiale des organes latéraux des plantes. Nous n’avons pas fait de coupes transversales au niveau du méristème apical, mais les mesures de l’angle entre chaque nouvelle feuille effectuées par d’autres auteurs (voir figure 15a) confirment la proximité à l’angle de Fibonacci pour la rosette d’Arabidopsis.
Figure 15a. Coupe transversale d’un méristème apical d’une plante d’Arabidopsis âgée de 30j. L’angle P5-P6 et P6-P7 est approximativement de 137.5 degrés. P= primordia foliaire. (Callos et Medford 1994).

40
Figure 16. Décomposition du mouvement circadien des cotylédons d’Arabidopsis thaliana écotype Ws.
1
1.2
1.4
1.6
1.8
2
2.2
2.4
0 2 4 6 8 10 12 14 16 18 20 22 24Surf
ace
des c
otyl
édon
s (m
m2 )
Temps (heures)
α et α1 = angle entre l’hypocotyle et le pétiole du cotylédon.β et β1 = angle entre le pétiole du cotylédon et le limbe du cotylédon.
β 1
α 1β
α
90100110120130140150160170180
0 2 4 6 8 10 12 14 16 18 20 22 24
α β
angl
e
Temps (heures)
75
95
115
135
155
175
195
0 2 4 6 8 10 12 14 16 18 20 22 24
α 1 β 1
angl
e
Temps (heures)

41
Figure 17. Évolution de la surface du 5ème au 16ème jour après le semis (programme utilisé: area.m). Intervalle de temps entre chaque photo=2h 30min. Jour et heure du semis: 10-04-2002 à 13h30.
1
3
5
7
9
11
13
15
17
19
5 6 7 8 9 10 11 12 13 14 15 16Temps (jours)
Surf
ace
(mm
2 )

42
Figure 18. Évolution de la surface du 16 ème au 26 ème jour après le semis (programme utilisé: area.m). Intervalle de temps entre chaque photo=2h 30. Jour et heure du semis: 10-04-2002 à 13h30.
15
35
55
75
95
115
135
155
15 16 17 18 19 20 21 22 23 24 25 26 27
Chute del’hypocotyl
Temps (jour)
Surf
ace
(mm
2 )

43
. Figure 19. Évolution de la surface du 26 ème au 41 ème jour après le semis (programme utilisé: area.m). Intervalle de temps entre chaque photo=2h 30. Jour et heure du semis: 10-04-2002 à 13h30.
0
2
4
6
8
10
12
14
16
18
26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41
Temps (jours)
Surf
ace
(cm
2 )

44
Figure 20. La courbe du premier graphique représente l’évolution de la surface de la rosette âgée de ~20j calculée avec le programme area.m. A partir du troisième jour nous avons substitué la phase nocturne avec une phase diurne. La persistance du rythme circadien en condition constante de température, humidité et lumière confirme son origine endogène. L’analyse de Fourier (voir timefreq.m) du signal original entre le troisième et septième jour auquel nous avons soustrait la courbe de tendance nous a permit de détecter une période principale de 25.5h. Dans le deuxième graphique nous avons superposé le signale original et le signal reconstruit grâce à la période principale. Cela nous permet de vérifier visuellement si l’oscillation de 25.5h correspond bien au signal original. Intervalle de temps entre chaque photo=15min.
Signal reconstruitpériode = 25.5h
Signal original
2729
3133
3537
3941
4345
0 1 2 3 4 5 6 7 8Temps (jours)
Surf
a ce
(mm
2 )

45
Figure 21.Mouvements ultradiens des feuilles de la rosette (a,b,c) et de l’hypocotyle (d,e,f) d’Arabidopsis. Les signaux originaux auquels nous avons soustrait la courbe de tendance se trouvent dans les graphiques a et d. Les résultats de l’analyse de Fourier (voir timefreq.m) sont présenté dans les graphiques b et e. Nous avons détecté une période principale de ~1.8h. Dans les graphiques (c,f) nous avons superposé le signal reconstruit avec la période principale (courbe rouge) et le signal original (a,d), cela permet de conclure que la période de ~1.8h correspond bien à notre signal original. Intervalle de temps entre chaque photo pour (a,b,c)=15min. Intervalle de temps entre chaque photo pour (d,e,f)=5min.
Période principale ≅ 1.8 h Période principale ≅ 1.8 h
a
b
d
e
c f
46
Figure 22. Chute de l’hypocotyle. Les croix noires montrent la localisation du centre de gravité de la plante. La sélection de la plante a été effectuée à l’aide du lasso magnétique de la palette outil de Photoshop, ensuite la valeur du centre de gravité a été calculée grâce au programme centregravity.m. Le graphique montre le parcours effectué par le centre de gravité (vue latérale).
1) 27-04-02 22:30h
2) 30-04-0215:30h
3) 30-04-02 18:00h
4) 30-04-02 20:30h 5) 30-04-02 23:00h
6) 03-05-02 16:00h 7) 06-05-02 14:00h
1) 27-04-02 22:30h
2) 30-04-02 15:30h 3) 30-04-02 18:00h
4) 30-04-02 20:30h 5) 30-04-02 23:00h
6) 03-05-02 16:00h 7) 06-05-02 14:00h

47
Figure 23. Le graphe montre l’évolution du rythme circadien pour un cotylédon et les premières quatre feuilles. La sélection des feuilles a été effectue à l'aide de Photoshop pour le calcul de la surface nous avons utilisé le programme area.m. Il est important de ne pas oublier que la position (angle) de la camera par rapport à l’objet observé peut affecter les amplitudes des mouvements en les diminuant ou en les amplifiant. Intervalle de temps entre chaque photo=2h 30. Jour et heure du semi: 10-04-2002 à 13h30.
cotylédon1feuille 1
feuille 2
feuille 3
feuille 4
feuille 5cotylédon2
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Temps (jours)
Surf
ace
des f
euill
es (p
ixel
s)
cotylédon1
feuille 1
feuille 2
feuille 3
feuille 4

48
3.2.3.2 Croissance rythmique des feuilles d’Arabidopsis thaliana
Une grande partie de la biomasse terrestre est produite par les végétaux. S’intéresser à la dynamique de croissance des feuilles est donc un point très important pour mieux comprendre le comportement de la biomasse végétale. Les observations antérieures (voir 3.1.3.1 Mouvement circadien des feuilles d’Arabidopsis thaliana) nous ont permis de constater que les feuilles se lèvent la nuit et se baissent le jour avec une rythmicité circadienne endogène. Pendant ces oscillations les feuilles augmentent aussi de taille. Le fait d’avoir simultanément de la croissance et du mouvement rend l’analyse de la croissance plus compliquée. Nous avons donc décidé de bloquer le mouvement des feuilles avec une bande de parafilm (voir figure 24). Quelques expérimentations ont été nécessaires pour obtenir le bon protocole. En effet si la bande de parafilm est trop courte une fois que les feuilles ont atteint une taille suffisamment grande elles se repliaient en assumant une forme concave. La majorité de ces changements de formes réversibles nécessitent environ 2h pour arriver au maximum de leur courbure, mais un cas où la feuille s’est repliée en 1h a aussi été observé (voir figure 25a). Ces « brusques » changements de forme apparaissent surtout pendant que la feuille se trouve en condition de lumière continue (voir figure 25). Nous avons vu que pour les feuilles d’Arabidopsis la zone où s’effectue la grande partie du mouvement se trouve entre le pétiole et le limbe (voir figure 16). La figure 26 montre la différence entre une mauvaise fixation et une bonne fixation. Pour une bonne fixation il suffit en effet de laisser sortir un petit peu la pointe de la feuille. Cela explique l’apparition du mouvement circadien si la bande de parafilm est trop courte. La bande de parafilm à été étiré enfin de diminuer son épaisseur. Un positionnement adequat des sources de lumière infrarouges (15 W) permet de diminuer la réflexion du parafilm, ce qui est un avantage pour l’analyse d’image. Les plantes que nous avons utilisées ont été semées et cultivées au phytotron dans les conditions environnementales suivantes: température 20°C (±1), humidité 70% (±5), 12h lumière (tube néon Daylight 6000 lux) 12h d’obscurité. A l’âge de 22 jours les plantes ont été
transférées dans la chambre de mesure où ce trouve le dispositif de mesure (voir figure 27). Un intervalle de temps de 15 min entre chaque photo a été utilisé pour suivre la dynamique de croissance des feuilles d’Arabidopsis thaliana écotype Ws. Figure 24. La photo montre la méthode de fixation des feuilles utilisées pour l’étude de la dynamique de croissance des feuilles d’Arabidopsis. Figure 25. Le calcul de la surface (mm2) a été effectué avec le programme area.m. Les diminutions de la surface sont dues au repliement de la feuille et à un faible mouvement rythmique circadien que la feuille arrive a effectuer si la bande de parafilm n’est pas suffisamment large.
Fixation de la feuille
0
10
20
30
40
50
60
70
80
90
100
0 1 2 3 4 5 6 7 8
Repliement en formeconcave de la feuille
Temps (jour)
mm
2

49
Figure 25a. Ce graphique illustre le repliement en forme concave de la feuille d’Arabidopsis thaliana écotype Ws. Une fois atteint un maximum de courbure négative la feuille retourne lentement à une forme plus aplatie. Ce mouvement est donc réversible. Figure 25b. Ce graphique montre la dynamique de croissance de la surface d’une feuille d’Arabidopsis thaliana écotype Ws obtenu avec le programme area.m en sélectionnant toute la feuille ou une partie de la feuille. Cela montre que les oscillations rythmiques circadiennes de la croissance sont visibles aussi si le calcul de l’évolution de la surface n’est effectué qu’avec une partie de la feuille (sélection partielle).
35
40
45
50
55
60
65
0 1 2 3 4 5 6
Surf
ace
(mm
2 )
Temps (heures)
Repliement en forme concavede la feuille(courbure négative)
Retour à une forme plus aplatie
10000
20000
30000
40000
50000
60000
70000
80000
0 1 2 3 4 5 6
Temps (jours)
Surf
ace
feui
lle (p
ixel
s)
Dynamique de lacroissance de lafeuille totale
Dynamique de lacroissance d ’une partie de la feuille

50
Figure 26. Les quatre photos illustrent la différence entre une fixation qui permet d’éviter la courbure de la feuille (bonne fixation) et une fixation qui ne le permet pas. Les chiffres indiquent la taille de la feuille en mm2 calculé avec le programme area.m. au commencement et à la fin de l’experience. Des enregistrements d’une durée de 7 jours en condition de 12h de lumière et 12h d’obscurité ou de lumière continue nous ont permis, suite au traitement informatique du lot d’images (voir 2.3.3.3.1 Programme: area.m), d’obtenir les courbes des graphiques a et b de la figure 27. Pour ces courbes la valeur de la taille initiale de la feuille à été ajustée à 0. Figure 27. Photo du dispositif de mesure qui se trouve dans une chambre à environnement contrôlé (voir Matériel et Méthode). En voulant être le moins invasif possible nous avons décidé de ne pas couper les feuilles de la rosette. En choisissant une feuille à l’intérieur
de la rosette autour de laquelle les feuilles sont très petites, le problème du chevauchement des feuilles peut être réduit. Il est en tout cas inévitable que de temps en temps les feuilles autour de la feuille photographiée recouvrent une partie de la feuille analysée. A cause de ce problème le calcul de l’évolution de la surface foliaire a été effectué qu’avec la partie libre de la feuille en question. Une vérification sur la même feuille, qui permettait la sélection totale de sa surface et une sélection partielle, a permit de conclure que la dynamique présente la même allure (voir figure 25b). Résultats et discussion A partir des deux courbes (graphique c et d figure 28) obtenues en faisant la moyenne des courbes des graphiques a et b (voir figure 28) nous avons obtenu les deux courbes de vitesse de croissance (voir graphique e et g figure 28). La figure 29 contient les résultats de l’analyse de fréquence faite à partie des 5 derniers jours des deux courbes de la vitesse de croissance. Cette analyse a été effectuée à l’aide de la première partie du programme timefreq.m (voir 2.3.3.3.1 Programmes). Le fait de ne pas observer des décroissances dans les courbes de croissance et que la vitesse de croissance est quasiment jamais négative, prouve que notre méthode a bien permis d’annuler le mouvement rythmique circadien des feuilles. Le maintien des oscillations rythmiques de la croissance même en lumière continue prouve que la croissance des végétaux est très probablement sous le contrôle d’un rythme circadien endogène. Grâce à une analyse de Fourier (voir Figure 29) nous avons pu détecter deux périodes (voir périodes encerclées) d’environ 24h. La valeur du power correspond à leur importance au niveau du signal analysé. Nos signaux ne présentent pas suffisamment d’oscillation pour que l’analyse de Fourier fournisse un résultat précis. La méthode de régression, dans ces cas, permet de mieux déterminer la valeur de la période principale, en partant de la supposition que notre signal oscillatoire n’est pas engendré par des phénomènes de battement d’ondes. Cette méthode consiste à résoudre une équation : Acos(2πt/T+Phi) à trois inconnues : A(amplitude), T(période) et Phi (phase) pour lesquels la somme de la variance entre le signal originale et le signal théorique est minimale.
mauvaise fixation bonne fixation
CRLgi 100
CRLgi 755
CRLrn 100
CRLrn 768
Feuille en formeconcave
65.4 mm2
150.8 mm2
73.5 mm2
128.4 mm2

51
Figure 28. Croissance des feuilles de la rosette d’Arabidopsis thaliana écotype Ws du 22ème au 29ème jour après le semis. Les graphiques de gauche contiennent les données enregistrées en 12h de lumière et 12h d’obscurité, tandis que pour celles de droite, après deux jour, l’expérience se déroule en lumière continue. Les graphiques c et d ont été obtenus en calculant la moyenne des courbes des graphiques a et b. Les graphiques e et f contiennent la courbe de vitesse de croissance calculée à partir des graphiques c et d. Intervalle de temps entre chaque photo =15 min, programme informatique utilisé pour l’analyse de l’évolution de la surface: area.m.
-5
0
5
10
15
20
25
30
35
40
45
50
55
0 1 2 3 4 5 6 7
0
10
20
30
40
50
60
70
80
0 1 2 3 4 5 6 7
0
20
40
60
80
100
120
140
0 1 2 3 4 5 6 7
-5
0
5
10
15
20
25
30
35
40
45
50
55
0 1 2 3 4 5 6 7 8
0
10
20
30
40
50
60
70
80
0 1 2 3 4 5 6 7 8
0
20
40
60
80
100
120
140
0 1 2 3 4 5 6 7 8
Temps (jour)
mm
2
Temps (jour)Temps (jour)
mm
2m
m2
Temps (jour)
Temps (jour)Temps (jour)
mm
2 /jo
urm
m2
mm
2 /jo
ura b
c d
e f

52
Figure 29. Analyse de fréquence pour la vitesse de croissance des feuilles d’Arabidopsis thaliana écotype Ws effectuée avec la première partie du programme timefreq.m. Les graphiques a et b correspondent respectivement au données des graphiques e et f de la figure 28 à partir du deuxième jour. Les graphiques c et d contiennent toutes les fréquences (période/cycle), trouvées par analyse de Fourier de la vitesse de croissance. Avec la période plus importante (power maximal) nous avons reconstruit un signal oscillatoire qui a été superposé au signal original, le résultat est visible dans les graphiques e et f.
a b
c d
e f

53
Figure 29a. Le graphique a contient la courbe de la vitesse moyenne de la croissance des feuilles d’Arabidopsis thaliana écotype Ws en régime photopériodique L:D (12:12) à laquelle on soustrait la tendance, et une courbe théorique avec une période (T) de 23.7h calculé avec la méthode de régression. Le graphique b contient la courbe de la vitesse moyenne de la croissance des feuilles d’Arabidopsis thaliana écotype Ws en lumière continue à laquelle on soustrait la tendance, et une courbe théorique avec une période (T) de 25h calculé avec la méthode de régression. A l’aide de la méthode de régression nous avons trouvé une période de ~24h pour la vitesse de croissance des feuilles en régime photopériodique L :D (12 :12) et de ~25h pour les feuilles en lumière continue (voir figure 29a). Par cette technique nous avons donc pu mettre en évidence la présence d’un rythme
circadien endogène de ~25h pour la croissance des feuilles d’Arabidopsis thaliana écotype Ws Par superposition des deux courbes de la vitesse de croissance moyenne (voir figure 30), deux différences principales sont observables. La première, mit en évidence par le rectangle dans la figure 30, correspond à la première nuit illuminée pour les feuilles en lumière continue.
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0 1 2 3 4 5 6 7
Temps (jours)
mm
2 /he
ures
a
-0.2
-0.1
0
0.1
0.2
0.3
0.4
1 2 3 4 5 6 7 8
Temps (jours)
mm
2 /he
ures
b lumière continue
L:D (12:12)
25h
23.7h

54
Figure 30. Superposition des deux courbes de la vitesse de croissance des graphiques e et f de la figure 28. Le rectangle met en évidence la région de plus grande différence entre les deux courbes. Cette région correspond pour la deuxième courbe (graphique f figure 28) à la première nuit illuminée. Pendant cette phase les feuilles en lumière continue ne présentent pas la croissance normale de la phase nocturne mais une croissance réduite. La deuxième différence que nous avons pu remarquer, c’est que les feuilles en lumière continue ne présentent pas les pics d’une durée de ~ 2h 15 min observables au commencement de chaque journée (enclenchement des néons) pour la vitesse de croissance moyenne des feuilles en régime photopériodique de 12h de lumière et 12h d’obscurité (voir graphique e de la figure 28). L’enclenchement de la lumière pourrait donc être la cause des pics que nous observons. Des repliements du bord de la feuille ont aussi été observés en lumière continue (voir figure 25a). L’enclenchement des néons n’est donc pas le seul facteur qui pourrait induire la courbure de la feuille. L’origine du repliement des feuilles est actuellement encore inconnue. Des études effectuées sur le contrôle génétique de la courbure de la surface foliaire on mis en évidence l’existence d’un gène (CINCINNATA) dont la mutation affecte la taille, la courbure et la forme de la feuille de Antirrhinum (Nath et al. 2003). Une diminution de la croissance dans la région marginale par rapport à la région centrale des feuilles induit une courbure positive. En contraste quand la région centrale croit plus lentement la feuille assume une courbure négative (voir figure 31). Les mécanismes qui engendrent ces changements de courbure de la surface foliaire sont encore inconnus.
Figure 31. Le disque uniformément coloré de gris correspond à une surface Gaussienne avec une courbure de zéro. Une réduction de la croissance dans la région marginale (zone foncé) par rapport à la région centrale des feuilles induit une courbure positive. En contraste quand la région centrale croit plus lentement la feuille assume une courbure négative (Nath et al. 2003). En règle générale pour la rosette d’Arabidopsis se sont les feuilles plus âgées qui présentent une courbure plus importante. Il n’est pas fréquent de trouver dans la littérature botanique des études sur la dynamique de croissance des feuilles, à notre connaissance une telle étude n’a jamais été faite pour les feuilles d’Arabidopsis. Tsukaya dans son article résume l’états des connaissances sur la régulation du développement des feuilles d’Arabidopsis (Tsukaya 2002). Des gènes qui régulent la longueur (ROTUNDIFOLIA3) ou des gènes qui régulent la largeur (ANGUSTIFOLIA) de la feuille ont été trouvés chez Arabidopsis (Tsukaya 2002). Le fait que la croissance des feuilles s’arrête a une certaine taille, montre bien que la feuille a une structure déterminée. Dans un article apparu dans Pour la Science (Silk et Erickson 1980) les auteurs proposent de comparer la croissance d’une plante et l’écoulement d’un fluide. Dans les deux cas nous avons une dynamique globale engendrée par la somme de dynamiques locales. En mesurant la composante de la vitesse et la divergence de la vitesse pour 250 points sur une feuille de Xanthium ils ont obtenu une carte de distribution des taux élémentaires relatifs à l’augmentation de surface. Cette carte montre une croissance plus rapide à la base de la feuille (Silk et Erickson 1980). Les feuilles sont donc des structures beaucoup plus dynamiques de ce que l’on croit. Les repliements concaves ou convexes et les changements de forme que l’on peut observer lors du cycle de vie traduisent la nature
courburepositive
courburenégative
-5
0
5
10
15
20
25
30
35
40
45
50
55
0 1 2 3 4 5 6 7 8
Temps (jour)
mm
2 /jo
ur

55
plastique et en même temps, structuré de cet organe. A priori on ne peut pas exclure une probable régulation rythmique circadienne d’une très faible courbure de la surface foliaire. Nous pouvons donc formuler les deux hypothèses suivantes pour expliquer les changement rythmiques de croissance que nous observons: -hypothèse 1: les feuilles croissent avec une croissance isotropique sous le contrôle d’un rythme circadien endogène. -hypothèse 2: les feuilles croissent avec une croissance anysotropique sous le contrôle d’un rythme circadien endogène. La deuxième hypothèse nous parait plus plausible. En conclusion nous pouvons dire que les feuilles d'Arabidopsis thaliana écotype Ws croissent plus pendant la nuit que le jour et maintiennent ce rythme en condition de lumière continue. Cela prouve bien l’existence d’un rythme circadien endogène. Il est donc fort possible que l’horloge circadienne qui régule le mouvement circadien des feuilles soit aussi impliquée dans la régulation de la croissance circadienne des feuilles.
3.2.3.3 Circumnutation Introduction Les premières recherches expérimentales sur les mouvements révolutifs remontent au début du XIXème siècle, mais c’est Charles Darwin (Darwin et Darwin 1880) qui a véritablement ouvert la voie aux travaux ultérieurs. Dans son ouvrage « The power of movement in plants » il décrit toute une série de mouvements. Les figures 20 et 21 illustrent deux exemples de circumnutation observé par Darwin. Figure 32. Brassica oleracea: circumnutation de l’hypocotyle pendant 10 heures 45 minutes. L’image est réduite environ 4 fois par rapport à l’image original
Figure 33. Quercus (American sp.): circumnutation de la jeune tige, tracé sur un verre horizontal, de 12.50h 22 Février. à 12.50h 24 Février. De nombreuses plantes présentent des mouvements de circumnutation de l’hypocotyle, tige, feuilles, racines, voici une petite liste: -Brassica oleracea (hypocotyle) -Githago segetum (hypocotyle) -Gossypium (hypocotyle) -Oxalis rosea (cotylédons et hypocotyl) -Primula sinensis (cotylédons et hypocotyle) -Citrus aurantium (Orange) (épicotyle) -Dioscorea batatas (tige volubile) -Vicia faba (Fève) (radicule, épicotyle,tige) -Helianthus annuus (hypocotyle) -Lathyrus nissolia (tige de la jeune plantule) -Phaseolus vulgaris (tige volubile) -Helianthus annus (hypocotyle) Le terme circumnutation est composé de deux parties: circum: autour, et nutation: changement de direction d’un organe végétal en cours de croissance (petit Robert 1973). Pour la plantule d’Arabidopsis nous avons vu que la radicule et l’hypocotyle présentent des mouvements de circumnutation (voir 3.1.2 Plantule). Comme cela est illustré dans les figures 5 et 6 (voir 3.2 Cycle de vie d’Arabidopsis thaliana) dans nos conditions environnementales (voir Matériel) avec une photopériode de 12h de lumière et 12h d’obscurité, une tige centrale (hampe florale) apparaît environ 35 jours après le semis. Des études effectuées au sein du Laboratoire de Physiomatique de Genève avec des capteurs de croissance sur la dynamique de croissance

56
du premier entrenœud de la hampe florale d’Arabidopsis, ont permis de détecter une croissance rythmique circadienne endogène, avec l’auxine comme élément clé (Jouve 2000). Les courbes non lissées de cette étude, montrent des oscillations ultradiennes endogènes (période de 18 à 72 min) à l’intérieur de l’oscillation de base circadienne. A notre connaissance aucune étude cinématographique n’a été effectuée pour caractériser la dynamique des mouvements et de la croissance du sommet de la hampe florale d’Arabidopsis thaliana lors de son développement. La partie suivante contient les résultats de nos expériences. Matériel et Méthode Pour l’étude de la dynamique du sommet de la hampe floral d’Arabidopsis nous avons utilisé un intervalle de temps entre chaque photo de 15 min. Cet intervalle permet de visualiser des dynamiques ultradiennes. Les programmes compilés pour ce travail (voir 2.3.3.3.1 Programmes: centregravity.m, invlong.m, vitesseangle.m et timefreq.m) nous ont permis d’affiner notre recherche afin de mieux caractériser cette dynamique. La figure 34 (8 plantes en 12h de lumière et 12h d’obscurité) et la figure 37 (4 plantes en lumière continue) montrent les dynamiques espace-temps (x,y,t) obtenues suite au traitement des photos latérales avec le programme inlong.m. Le principe de fonctionnement de ce programme est très simple, la seule chose qu’il nécessite à la base, sont des images bien contrastées. D’abord il transforme l’image en 256 niveaux de gris en noir (0) et blanc (255) à l’aide d’un filtre. Ensuite en commencent par le haut il calcule la somme des pixels qui composent la première ligne, la deuxième, la troisième,…. Chaque fois que la ligne est composée seulement de pixels noirs la somme sera 0. Une fois que le programme rencontre le premier pixel blanc (somme≠0) il va descendre un certain nombre de lignes (ex. 25) qui correspondent environ à la hauteur de l’inflorescence finale. Cette opération permet d’atténuer les aberrations de la détection du sommet de la hampe de part les mouvements des feuilles de l’inflorescence. Enfin, à partir des pixels blancs qui se trouvent dans cette zone il va calculer un centre de gravité (x,y). Pour vérifier visuellement où se trouvent les coordonnées (x,y), le programme dessine à cet
endroit sur l’image originale un carré gris. Cette opération effectuée sur chaque image permet d’obtenir une animation (voir data.avi dans le programme). L’enchaînement des coordonnées x,y (voir invlongdata.txt dans le programme) permet de compiler l’animation de l’évolution de la trajectoire dans le temps en 2D (voir invlongdata.avi dans le programme). La coordonnée y correspond principalement à la croissance tandis que la coordonnée x au mouvement du sommet de la hampe floral. Les graphiques 35, 36 et 39 permettent de visualiser les trajectoires des coordonnes x et y dans le temps. Pour approfondir notre étude nous avons placé deux caméras perpendiculairement (90°±5°) l’une par rapport à l’autre afin d’avoir la dynamique vue du dessus et ainsi pouvoir reconstruire la trajectoire en 3D (voir figure 45). Enfin nous avons utilisé un dernier programme (voir 2.3.3.3.1 Programme : vitesseangle.m) qui permet de bien visualiser les changements de sens de rotation, calculer la vitesse angulaire, les déplacements du centre de gravité et les variation d’amplitude de la trajectoire, à partir des images vues du dessus traitées d’abord avec le programme centregravity.m. Les périodes des signaux oscillatoires détectées lors de cette étude sont ensuite caractérisées à l’aide du programme timefreq.m. Resultat et Discussion En compilant les films avec le lot d’images accumulées sur un arc de temps de ~ 6 jours, nous nous sommes aperçus que le sommet de la hampe florale d’Arabidopsis effectue des mouvements circulaires elliptiques et pendulaires, qui peuvent varier d’amplitude et sens de rotation dans le temps (voir CD circumH.avi et autres). C’est mouvements sont rassemblés sous le terme général de nutation ou plus spécifiquement pour les mouvements de rotation sous le terme de circumnutation. Nous n’avons pas trouvé dans la littérature des études effectuées sur ce type de mouvements pour la hampe florale d’Arabidopsis thaliana. Par l’observation des silhouettes des courbes (voir composante x figures 35,36 et 39), en règle générale, mais pas absolue, nous pouvons affirmer que la hampe florale parcourt des trajectoires plus amples pendant la nuit. Les composantes x de la figure 35 et du graphique f de la figure 36 montrent des fluctuations rythmiques circadiennes des amplitudes.

57
Figure 34. Dynamiques espace-temps (x,y,t) du sommet de la hampe florale d’Arabidopsis thaliana écotype Ws obtenues par traitement des images (vue latérale) avec le programme invlong.m. Les plantes des graphiques a (voir CD circumH.avi) et b ont été semées et cultivées dans les conditions environnementales de la chambre de mesure (voir Matériel). Les autres plantes ont été cultivées au phytotron: température 20°C (±1), humidité 70% (±5), 12h lumière (tube néon Daylight 6000 lux) 12h obscurité jusqu’au jour de la mesure. Les plantes c et d, e et f (voir CD Synchro.avi) et g et h ont été photographie en couple par la même caméra simultanément. Il est intéressant de remarquer qu’elle présente des similitudes dans la silhouette de la dynamique. Les chiffres le long des deux axes sont des mm. Les déplacements vers la droite de la trajectoire des graphiques c et g sont dus au phototropisme.
20
40
60
80
100
120
140
160
180
20 30 40 50 60 70 80 9020
40
60
80
100
120
140
160
20 30 40 50 60 70 80
20
40
60
80
100
120
140
160
180
40 50 60 70 80 90 10020
40
60
80
100
120
140
160
40 45 50 55 60 65 70
20
40
60
80
100
120
140
160
30 40 50 60 70 80 9020
40
60
80
100
120
140
160
20 30 40 50 60 70 80 90
10
20
30
40
50
60
70
80
90
100
110
20 30 40 50 60 70 8020
40
60
80
100
120
140
-100 -80 -60 -40 -20 0 20 40
Chute dela hampe
a b
c d
e f
g h

58
Figure 35. Décomposition des dynamiques espaces-temps de la figure 34 (graphiques a, b, c et d) dans les composantes x, y et t. La composante y correspond à la croissance de la hampe. Les décroissances sont dues aux mouvements de la hampe. Dans le graphique c pour la courbe de croissance la présence d’un rythme circadien est clairement visible. La composante x correspond au mouvement de la hampe, dans ce signal oscillatoire nous pouvons noter la présence d’une oscillation rythmique ultradienne qui varie d’amplitude dans le temps.
20
30
40
50
60
70
80
90
100
110
0 1 2 3 4 5 6 720
40
60
80
100
120
140
0 1 2 3 4 5 6 7
20
40
60
80
100
120
140
160
0 1 2 3 4 5 6 720
40
60
80
100
120
140
160
0 1 2 3 4 5 6 7
Temps (jour)Temps (jour)
mm
mm
Temps (jour)Temps (jour)
mm
mm
a b
c d
Chute de la hampe

59
Figure 36. Décomposition des dynamiques espaces-temps de la figure 34 (graphiques e, f, g et h) dans les composantes x, y et t. La composante y correspond à la croissance de la hampe. Les décroissances sont dues aux mouvements de la hampe. La composante x correspond au mouvement de la hampe, dans ce signal oscillatoire nous pouvons noter la présence d’une oscillation rythmique ultradienne qui varie d’amplitude dans le temps.
20
40
60
80
100
120
140
160
180
0 1 2 3 4 5 6 720
40
60
80
100
120
140
160
0 1 2 3 4 5 6 7
20
40
60
80
100
120
140
160
180
0 1 2 3 4 5 6 720
40
60
80
100
120
140
160
0 1 2 3 4 5 6 7
mm
mm
Temps (jour) Temps (jour)
Temps (jour) Temps (jour)
mm
mm
e f
g h

60
Figure 37. Dynamiques espace-temps (x,y,t) du sommet de la hampe floral d’Arabidopsis thaliana écotype Ws en lumière continue obtenu par traitement des images (vue latérale) avec le programme invlong.m. Les plantes ont été cultivées au phytotron: température 20°C (±1), humidité 70% (±5), 12h lumière (tube néon Daylight 6000 lux) 12 obscurités jusqu’au jour de la mesure. Les plantes abcd ont été photographie simultanément par deux caméras positionnées latéralement. La hauteur initiale de la hampe florale au commencement de l’expérience n’était pas pour toutes les quatre plantes la même, cette différence est visible le long de l’expérience (voir figure 38). Les chiffre le long des deux axes sont des mm. Le déplacement vers la gauche de la trajectoire de la hampe du graphique c est dû très probablement à du phototropisme. Figure 38. Photo prises au cinquième jour par la caméra (r) et la caméra (g). De gauche à droite: plante a (voir graphique a figure 37,39), plante b (voir graphique b figure 37,39), plante c (voir graphique c figure 37,39) et plante d (voir graphique d figure 37,39).
40
60
80
100
120
140
160
180
200
30 40 50 60 70 80 90 100 0
20 40 60 80
100 120 140 160 180
30 40 50 60 70 80 90
20 40 60 80
100 120 140 160 180 200 220 240
30 40 50 60 70 80 90 100 0
20 40 60 80
100 120 140 160 180 200
20 30 40 50 60 70 80
a b
c d
Caméra Caméra

61
Figure 39. Décomposition des dynamiques espaces-temps de la figure 37 dans les composantes x, y et t. La composante y correspond à la croissance de la hampe. Les décroissances sont dues aux mouvements de la hampe. La composante x correspond au mouvement de la hampe, dans ce signal oscillatoire nous pouvons noter la présence d’une oscillation rythmique ultradienne, qui varie d’amplitude dans le temps et qui se maintien même en lumière continue.
0
20
40
60
80
100
120
140
160
180
200
220
0 1 2 3 4 5 6 7 8 9
0
20
40
60
80
100
120
140
160
180
200
220
0 1 2 3 4 5 6 7 8 9
0
20
40
60
80
100
120
140
160
180
200
220
0 1 2 3 4 5 6 7 8 9
0
20
40
60
80
100
120
140
160
180
200
220
0 1 2 3 4 5 6 7 8 9
a b
c d
Temps (jour)Temps (jour)
Temps (jour) Temps (jour)
mm
mm
mm
mm

62
Figure 40. Séquence de photos espacées de 15 min permettant de voir un cycle de rotation entier. Figure 41. 12 courbes de croissance de la hampe florale d’Arabidopsis thaliana écotype Ws obtenues par l’analyse des images (vue latérale) avec le programme invlong.m. Les 4 courbes noires correspondent aux courbes de croissance de la figure 39 (plantes en lumière continue), tandis que les 8 courbes grises aux courbes de croissance des figures 35 et 36 (plantes en 12h de lumière et 12h d’obscurité). Nous avons obtenu une vitesse de croissance moyenne pour les plantes en lumière continue de 26.7 ± 0.6 mm/j et de 19.7 ± 3 mm/j pour les 8 plantes en régime photopériodique L:D 12:12.
Temps (jour)
Cro
issa
nce
ham
pe f l
ora l
e (m
m)
0
50
100
150
200
0 1 2 3 4 5 6 7

63
Ces variations sont aussi observables dans les composantes x de la figure 39 en lumière continue. Il est donc possible que l’amplitude de la trajectoire soit sous le contrôle d’un rythme circadien endogène qui peut être plus ou moins clairement visible. Une régulation rythmique circadienne de l’amplitude de la circumnutation a déjà été observée (Johnsson 1997). Une explication claire de ce phénomène n’existe pas. Nous savons que la plasticité des tissus joue un rôle très important dans la détermination de l’amplitude de la trajectoire. Les vidéos compilées le long de ce travail, nous avons permit de constater que l’apparition des inflorescences latérales et d’inflorescences primaires auxiliaires (Jouve et al.1998) peut induire une diminution de l’amplitude de la trajectoire du sommet de la hampe florale. Nous avons aussi pu remarquer que ces inflorescences effectuent des mouvements rythmiques ultradiens (résultats non montré ici, observations personnelles à partir des vidéos). Par l’observations des variations des courbes de croissance il est possible d’apercevoir, dans celle qui ne sont pas trop affectées par les mouvements, la présence d’un rythme circadien de croissance (voir graphiques c et d figure 35). Ce rythme est aussi visible dans les plantes en lumière continue ce qui prouve son origine endogène (voir graphiques a et c figure 39). Nous pouvons donc dire que non seulement le premier entrenoeud de la hampe florale d’Arabidopsis montre un rythme circadien endogène (Jouve et al.1999) mais que la hampe entière croit avec un rythme circadien endogène, ce qui paraît une conséquence plutôt logique. Il est donc tout à fait possible qu’il existe une relation entre les variations rythmiques circadiennes de l’amplitude de la trajectoire et la croissance rythmique circadienne. En effet l’auxine qui semble être en facteur clé dans la régulation de la croissance circadienne (Jouve et al.1999) pourrait aussi être impliquée dans la variation des amplitudes, par la variation dans le temps de son gradient de concentration transversal (Friml et al.2002). Une autre hypothèse basée sur la variation de la vitesse du flux de l’eau à l’intérieur de la plante, pourrait aussi expliquer ces variations d’amplitudes.
En effet, il est connu que le mouvement de l’eau à l’intérieur de la plante est intimement lié aux flux de transpiration (flux d’eau entre les feuilles et l’air) et absorption (flux d’eau entre le sol et les racines). La régulation de ces deux flux détermine donc la vitesse du flux de la sève brute à l’intérieur de la plante. Nous savons que la vitesse de transpiration dépend de la température, de la lumière (Kinoshita 2001), du vent, du taux d’eau atmosphérique et de la disponibilité en eau du sol (Cornu 1998). Une régulation circadienne rythmique de l’ouverture des stomates a été détectée (Davis et Millar 2001 et Brinker et al. 2001). Cette régulation permet d’avancer l’hypothèse d’un flux rythmique de transpiration. Cela se traduit par des variations rythmiques de la pression hydrique à l’intérieur des vaisseaux conducteurs. Cette onde peut donc influencer le niveau de turgescence des cellules voisines. Il en résulterait donc une variation du volume des cellules qui composent les tissus et en conséquence une variation de l’amplitude du mouvement de circumnutation. Il faudrait étudier plus en détail la relation entre vitesse du flux et turgescence des cellules pour vérifier une telle hypothèse. Par analyse de Fourier (voir 2.3.3.3.1 Programme: timefreq.m) de nos signaux oscillatoires (voir composante x de la figure 35 et 36) nous avons pu établir un période d’oscillation moyenne de 3h ± 25min pour la hampe florale d’Arabidopsis thaliana écotype Ws en 12h de lumière et 12h d’obscurité (voir figure 40). En effectuant la même analyse (timefreq.m) avec les plantes en lumière continue (voir composantes x de la figure 39) la période moyenne obtenue était de 2h 25 ± 25 min. Par calcul des moyennes des amplitudes maximales et de la vitesse de croissance moyenne nous avons obtenu une amplitude maximale moyenne de 44 ±9.5mm et une vitesse de croissance moyenne de 19.7 ±3mm/j pour les plantes en 12h de lumière et 12h d’obscurité et de 54 ±9.5mm et 26.7 ±0.6mm/j pour les plantes en lumière continue (voir figure 41).
64
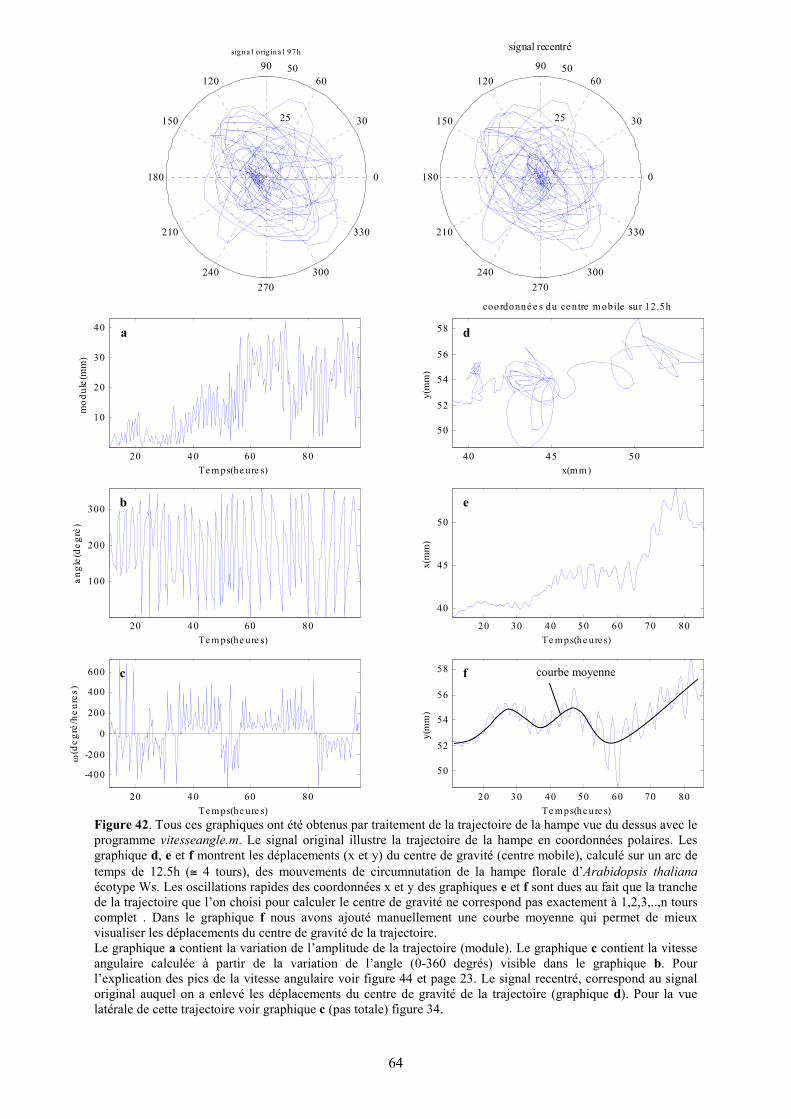
Figure 42. Tous ces graphiques ont été obtenus par traitement de la trajectoire de la hampe vue du dessus avec le programme vitesseangle.m. Le signal original illustre la trajectoire de la hampe en coordonnées polaires. Les graphique d, e et f montrent les déplacements (x et y) du centre de gravité (centre mobile), calculé sur un arc de temps de 12.5h (≅ 4 tours), des mouvements de circumnutation de la hampe florale d’Arabidopsis thaliana écotype Ws. Les oscillations rapides des coordonnées x et y des graphiques e et f sont dues au fait que la tranche de la trajectoire que l’on choisi pour calculer le centre de gravité ne correspond pas exactement à 1,2,3,..,n tours complet . Dans le graphique f nous avons ajouté manuellement une courbe moyenne qui permet de mieux visualiser les déplacements du centre de gravité de la trajectoire. Le graphique a contient la variation de l’amplitude de la trajectoire (module). Le graphique c contient la vitesse angulaire calculée à partir de la variation de l’angle (0-360 degrés) visible dans le graphique b. Pour l’explication des pics de la vitesse angulaire voir figure 44 et page 23. Le signal recentré, correspond au signal original auquel on a enlevé les déplacements du centre de gravité de la trajectoire (graphique d). Pour la vue latérale de cette trajectoire voir graphique c (pas totale) figure 34.
25
50
30
210
60
240
90
270
120
300
150
330
180 0
signa l origina l 97h
25
50
30
210
60
240
90
270
120
300
150
330
180 0
signa l norma lisé
20 40 60 80
10
20
30
40
mod
ule(
mm
)
Te m ps(he ure s)
20 40 60 80
100
200
300
angl
e(de
gré)
Te m ps(he ure s)
20 40 60 80
-400
-200
0
200
400
600
ω(d
egré
/heu
res)
Te m ps(he ure s)
40 45 50
50
52
54
56
58
coordonné e s du ce ntre m obile sur 12 .5h
y(m
m)
x(mm)
20 30 40 50 60 70 80
40
45
50
x(m
m)
Te mps(he ure s)
20 30 40 50 60 70 80
50
52
54
56
58
y(m
m)
Te mps(he ure s)
a
b
c
d
e
f
signal recentré
courbe moyenne
65
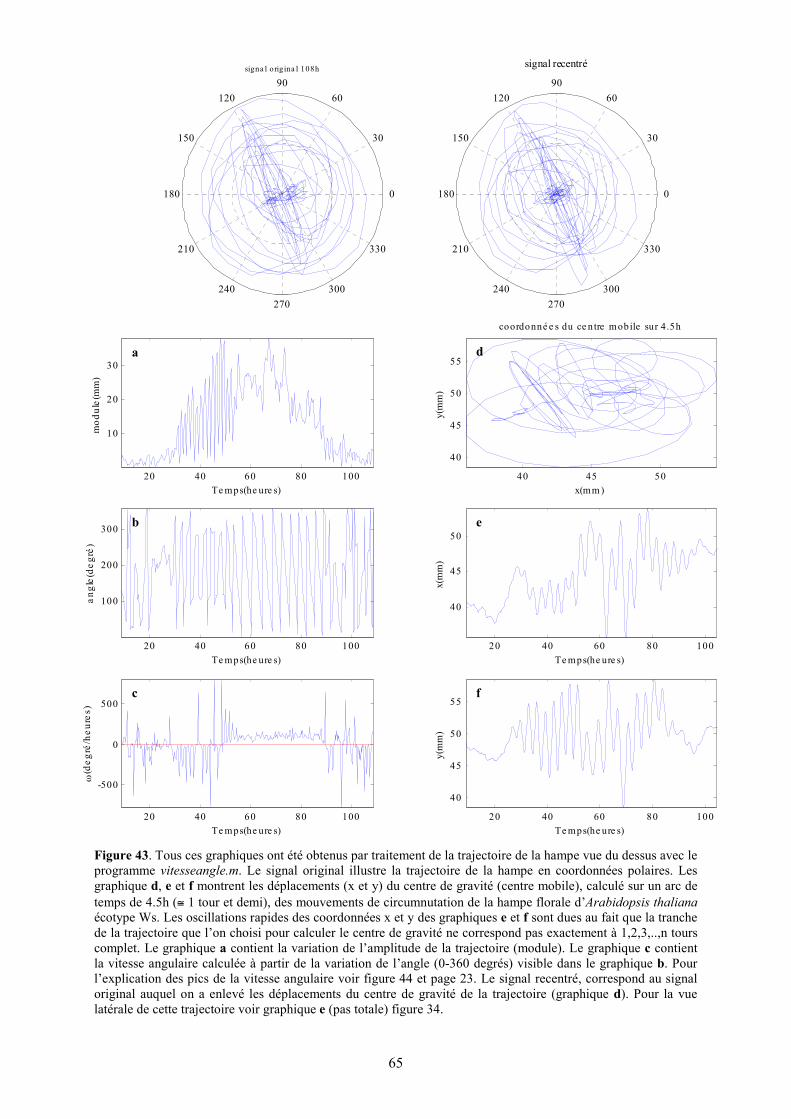
Figure 43. Tous ces graphiques ont été obtenus par traitement de la trajectoire de la hampe vue du dessus avec le programme vitesseangle.m. Le signal original illustre la trajectoire de la hampe en coordonnées polaires. Les graphique d, e et f montrent les déplacements (x et y) du centre de gravité (centre mobile), calculé sur un arc de temps de 4.5h (≅ 1 tour et demi), des mouvements de circumnutation de la hampe florale d’Arabidopsis thaliana écotype Ws. Les oscillations rapides des coordonnées x et y des graphiques e et f sont dues au fait que la tranche de la trajectoire que l’on choisi pour calculer le centre de gravité ne correspond pas exactement à 1,2,3,..,n tours complet. Le graphique a contient la variation de l’amplitude de la trajectoire (module). Le graphique c contient la vitesse angulaire calculée à partir de la variation de l’angle (0-360 degrés) visible dans le graphique b. Pour l’explication des pics de la vitesse angulaire voir figure 44 et page 23. Le signal recentré, correspond au signal original auquel on a enlevé les déplacements du centre de gravité de la trajectoire (graphique d). Pour la vue latérale de cette trajectoire voir graphique e (pas totale) figure 34.
30
210
60
240
90
270
120
300
150
330
180 0
signa l origina l 108h
30
210
60
240
90
270
120
300
150
330
180 0
signa l norma lisé
20 40 60 80 100
10
20
30
mod
ule(
mm
)
Te mps(he ure s)
20 40 60 80 100
100
200
300
angl
e(de
gré)
Te mps(he ure s)
20 40 60 80 100
-500
0
500
ω(d
egré
/heu
res)
Te mps(he ure s)
40 45 50
40
45
50
55
coordonné e s du ce ntre mobile sur 4.5h
y(m
m)
x(mm )
20 40 60 80 100
40
45
50
x(m
m)
Te mps(he ure s)
20 40 60 80 100
40
45
50
55
y(m
m)
Te mps(he ure s)
a
b
c
d
e
f
signal recentré

66
Figure 44. Ces graphiques ont été obtenus par le traitement de la trajectoire de la dynamique de la hampe florale d’Arabidopsis thaliana écotype Ws vue du dessus, avec le programme vitasseangle.m. Les graphiques a et c ont été obtenus par sommation de la différence des angles de la trajectoire. Par cette somme il est possible de voir clairement quand la plante change de sens de rotation. En effet si elle tourne en sens horaire (angle positif) la courbe croit tandis que si la plante tourne en sens anti-horaire (angle négatif) la courbe décroît. s indique les changements de sens de rotation (voir figure 45), tandis que les cercles mettent en évidence des région ou la trajectoire forme des 8. Si la hampe se déplace linéairement, vu qu’on n’observe pas de changements de l’angle, la courbe de la somme de la différence des angles sera horizontale. La variation de la vitesse angulaire calculée à partir des graphiques a et c est visible dans les graphiques b et d. Si la vitesse est positive la hampe tourne en sens horaire et vice-versa. Les grands pics de la vitesse angulaire sont causés par le passage très proche de la trajectoire au centre de gravité de la trajectoire (voir aussi remarques page 23). Ce phénomène est amplifié par le fait que nous avons un enregistrement discontinu de la trajectoire (Δt=15min) et donc l’angle entre deux points peut faire des variations très rapides. Dans la partie du graphique d ou la vitesse est assez stable (trajectoire circulaire) on peut estimer que la hampe tourne avec une vitesse de rotation de ~ 100degré/heure.
20 30 40 50 60 70 80 90
-1500
-1000
-500
0
500
1000
1500
2000
2500
som
me
angl
es(d
egré
)
Te m ps(he ure s)
se ns de rota tion (se uil 220 de gré s)
20 30 40 50 60 70 80 90
-1000
-500
0
500
ω(d
egré
/heu
res)
Te m ps(he ure s)
s s1 s2 s3s4 s5
10 20 30 40 50 60 70 80 90 100-2000
-1500
-1000
-500
0
500
1000
1500
2000
som
me
angl
es(d
egré
)
Te m ps(he ure s)
se ns de rota tion (se u il 220 de gré s)
10 20 30 40 50 60 70 80 90 100
-600
-400
-200
0
200
400
600
800
ω(d
egré
/heu
res)
Te m ps(he ure s)
s s1
déplacements« linéaires »
s2
a
b
c
d

67
Figure 45. Les dynamiques espaces-temps (x,z,t) des graphiques a et b ont été obtenues par le traitement des images (vue du dessus) avec le programme centregravity.m. Un petit bout de filtre de charbon noir (~1mm2) a été positionné au sommet de la hampe enfin de faciliter sa détection. A l’aide de l’animation de la trajectoire (voir dans le CD compilé pour ce travail: Armove.avi et Crmove450.avi) les changements de sens de rotation sont bien visibles. Les endroits où on a détecté un changement de sens de rotation sont indiqués par: s1, s2, s3, s4, s5 (s =switch). La figure 44 permet de mieux visualiser ces changements de sens de rotation et leur durée de temps. Comme est indiqué dans les premiers deux graphiques de cette figure, nous avons pu déterminer deux axes où la majorité des rotations se font en sens horaire (axe 2) ou en sens anti-horaires (axe 1). Dans l’animation Crmove450.avi, il est aussi possible de voir que si la tige tourne en sens horaire aussi l’axe 2 se déplace en sens horaire (voir figure 46) le contraire est aussi vrai pour l’axe 1. Une reconstruction de la trajectoire en 3D à été possible en positionnant les deux caméras à 90° (± 5) l’une par rapport à l’autre. En ayant les trois coordonnées x,y et z avec la fonction plot3 (x,y,z) sur Matlab, il est possible d’obtenir les graphiques en 3D. Le graphique c à été construit avec une partie de la coordonnée y du graphique e de la figure 36 et le graphique a de cette figure, tandis que le graphique d à été construit avec une partie de la coordonnée y du graphique c de la figure 35 et le graphique b de cette figure.
axe 2
axe 1
s1
s3
s2
s5
s4
end
axe 1 axe 2
s1
mm (x)
mm
(z)
mm (x)
mm
(z)
mm (x)mm (z) mm (x)mm (z)
mm
(y)
mm
(y)
a b
c d

68
La grande homogénéité de la vitesse de croissance pour les plantes en lumière continue pourrait être due ou faite que ces plantes ont été semées le même jour, qu’elles ont subi les mêmes déplacements depuis le phytotron au lieu de mesure et qu’elles se trouvaient exactement dans les mêmes conditions pendant la mesure vu qu’elles ont été photographiées en même temps. Il semble que le rapport entre la vitesse de croissance et la vitesse de rotation soit grossièrement linéaire (Baillaud 1962). En considérant l’ordre de grandeur des écarts type pour les amplitudes maximales moyennes et la vitesse de croissance moyenne nous pouvons supposer que la diminution de la période de rotation pour les hampes florales en lumière continue est probablement due à la faible augmentation de la vitesse de croissance plutôt qu’a des variations de l’amplitude. Par le traitement des images de la hampe vu du dessus avec le programme centregravity.m nous avons obtenu la trajectoire des graphiques a et b de la figure 45. Pour faciliter la détection du centre de la hampe (vue du dessus) nous avons positionné en petit carré noir (papier filtre de charbon 0.1mm2) au sommet de la hampe. Parfois le carré etait caché par une feuille. Dans ce cas nous avons dessiné sur les images un carré noir avec Photoshop. L’étude de cette projection a surtout été faite avec le programme vitesseangle.m, qui nous a permis d’obtenir les graphiques des figures 42, 43 et 44. Cette prospective nous a permis de mieux caractériser les changements de sens de rotation, les déplacements du centre de la trajectoire, les variations d’amplitude et la vitesse angulaire. Cette étude a été effectuée pour une question de temps seulement sur deux plantes. La figure 42 et les graphiques a et b de la figure 44 contiennent les résultats obtenus pour la première plante (voir graphique b figure 45 ou pour la vue latérale graphique c figure 34), tandis que la figure 43 et les graphiques c et d de la figure 44 contiennent les résultats obtenus pour la deuxième plante (voir graphique a figure 45 ou pour la vue latérale graphique e figure 34). Par cette étude, il semblerait que les plantes tournent plutôt en sens horaire qu’en sens anti-horaire et que la vitesse angulaire de rotation est approximativement de 100 degrés/heure pour les deux sens de rotation (voir figure 44). Des trajectoires en forme de 8 ont aussi été
détectées (voir courbes encerclé des graphiques a et c de la figure 44). Par l’observation des films (voir CD circumH.avi) et par la comparaison des courbes de la variation d’amplitude et du sens de rotation de la trajectoire (voir graphiques a de la figures 42 et graphiques a de la figures 44 ou graphiques a de la figures 43 et graphiques c de la figures 44) nous avons constaté que les changements de sens de rotation se produisent surtout lors d’une phase de faible amplitude de la trajectoire. Les graphiques e et f des figures 42 et 43 contiennent les déplacements des coordonnées x et y du centre de gravité des mouvements de circumnutation de la hampe florale. Les oscillations rapides (ultradiennes) sont dues au fait que le calcule du centre de gravité est toujours effectué sur une tranche de la trajectoire qui ne correspond pas exactement à 1,2,3…n tours complet. Ainsi le centre semble se déplacer à gauche et à droite avec un rythme ultradien qui correspond au rythme de rotation de la hampe. Une représentation plus correcte des déplacements du centre peut être obtenue en traçant la courbe moyenne (voir graphique f figure 42). Le sens de rotation et les déplacements du centre de gravité de la trajectoire de la hampe florale, semblent être reliés. Nous n’avons pas approfondi ce sujet mais il nous paraît en tout cas important de le mettre en évidence. Figure 46. Déplacement en sens horaire de l’axe 1, 2 et 3 (=grand axe de l’ellipse) et de la trajectoire de la hampe. Nous avons remarqué que quand la hampe tourne en sens horaire, si l’axe se déplace, il aura aussi plutôt une tendance à tourner en sens horaire (voir figure 46) et vice-versa. Nous avons aussi pu déterminer deux positions
10
20
30
40
50
60
70
80
90
100
0 10 20 30 40 50 60 70 80 90
1
2
3
~ 8h30 de trajectoire
mm (x)
mm
(z) start
end

69
du grand axe de l’ellipse (voir axe1 et axe2 des graphiques a et b de la figure 45) où la hampe effectue plutôt des rotations horaires (axe 2) ou anti-horaires (axe 1). En combinant la vue latérale avec la vue du dessus nous avons pu obtenir une représentation en 3D de la trajectoire de la hampe florale d’Arabidopisi thaliana (voir graphiques c et d de la figure 45). Mécanisme Le mouvement circulaire (circumnutation) résulte de l'existence d'un allongement inégal des génératrices successives de la tige (voir figure 47). Nous savons que chaque génératrice est composée d’une somme de cellules dont le volume peut varier selon la valeur de la pression osmotique. Des variations rythmiques de la pression osmotique ont été observées pour des cellules épidermiques d’une longueur d’approximativement de 100 μm (Millet et Badot, 1996).
Figure 47. Sur ce schéma du mécanisme de la circumnutation, seules les génératrices opposées sont regardées pour faciliter la compréhension mais il faut généraliser ce phénomène à toutes les génératrices. + : allongement important, - : allongement faible. (http://www.snv.jussieu.fr/bmedia/mouvements/nutation-analyse.html). Les mêmes auteurs ont démontré que la distribution asymétrique de la pression osmotique est reliée à une répartition asymétrique du K+ dans la zone de recourbement. La régulation des canaux K+ joue donc sûrement un rôle important dans le mécanisme de la circumnutation. En effet le blocage des canaux K+ inhibe la variation du volume cellulaire. Le calcium, connu pour son importance au niveau des régulations cellulaire (Malho 2000) est très probablement en ion impliqué dans la circumnutation vu que le blocage des canaux Ca2+ induit un
ralentissement de la vitesse de rotation (Millet et Badot, 1996). Une relation entre le mouvement révolutif et le métabolisme spécifique des hydrates de carbones (sucres) a aussi été mise en évidence. Des efforts produit pendant ces dernières décennies sur la relation entre la gravité et la circumnutation (Johnsson 1997) ont permis de mettre en évidence que l’effet de la gravité varie selon l’espèce. Le poids de la région apicale de la tige intervient inévitablement comme en frein dans les mouvements ascendants et accélère les mouvements descendants vu que la trajectoire dans aucun cas n’est horizontale. La circumnutation est très sensible aux différents stimuli physiques: vibration, toucher, température, champ magnétique (Ginzo and Décima 1995) et chimiques: lithium, éthylène, azote, jasmonate (Schuster 1996). La réorientation des microtubules corticaux peut être induite par différent signaux: hormones, lumière, gravité et stimulation physique (Wymer and Nick 1999). Des études in vivo sur la dynamique des microtubules corticaux induite par la gravité (Himmelspach et al. 1999) ont permis d’observer une réorganisation de la position transversale à la position longitudinale en ~ 40 min. Les microtubules corticaux comme composantes principales du cytosquelette jouent un rôle très important dans la morphogenèse des cellules. Des gènes (SPIRAL) dont la mutation change l’orientation des microtubules dans la cellule, induisent une modification au niveau du sens de rotation de la circumnutation (Furutani et al. 2000). L’auxine semble aussi être un élément impliqué dans le mécanisme de la circumnutation, en effet une forte activité auxinique a été détectée dans la partie mobile (Baillaud 1962). Des transporteurs transversaux (PIN3) ont été récemment identifiés (Friml et al. 2002) ce qui pourrait expliquer la rotation du gradient de l’auxine. Vu que dans la majorité des cas la période de rotation est comprise entre une et quatre heures (Baillaud 1962) il est fort probable que les mécanismes de base soient similaires pour toutes les plantes qui circumnutent.
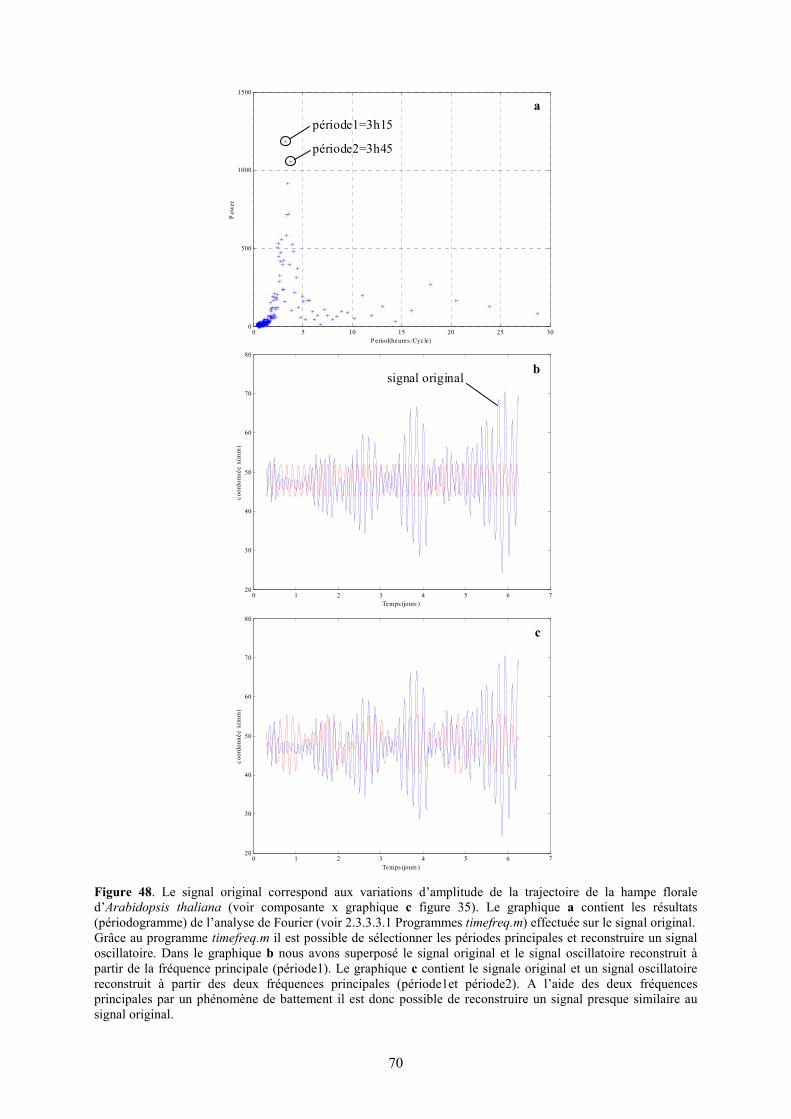
70
Figure 48. Le signal original correspond aux variations d’amplitude de la trajectoire de la hampe florale d’Arabidopsis thaliana (voir composante x graphique c figure 35). Le graphique a contient les résultats (périodogramme) de l’analyse de Fourier (voir 2.3.3.3.1 Programmes timefreq.m) effectuée sur le signal original. Grâce au programme timefreq.m il est possible de sélectionner les périodes principales et reconstruire un signal oscillatoire. Dans le graphique b nous avons superposé le signal original et le signal oscillatoire reconstruit à partir de la fréquence principale (période1). Le graphique c contient le signale original et un signal oscillatoire reconstruit à partir des deux fréquences principales (période1et période2). A l’aide des deux fréquences principales par un phénomène de battement il est donc possible de reconstruire un signal presque similaire au signal original.
0 1 2 3 4 5 6 720
30
40
50
60
70
80
Temps (jours )
coor
donn
ée x
(mm
)
0 1 2 3 4 5 6 720
30
40
50
60
70
80
Temps (jours )
coor
donn
ée x
(mm
)
0 5 10 15 20 25 300
500
1000
1500
Pow
er
P eriod(heures /Cycle)
période1=3h15
période2=3h45
signal original
a
b
c
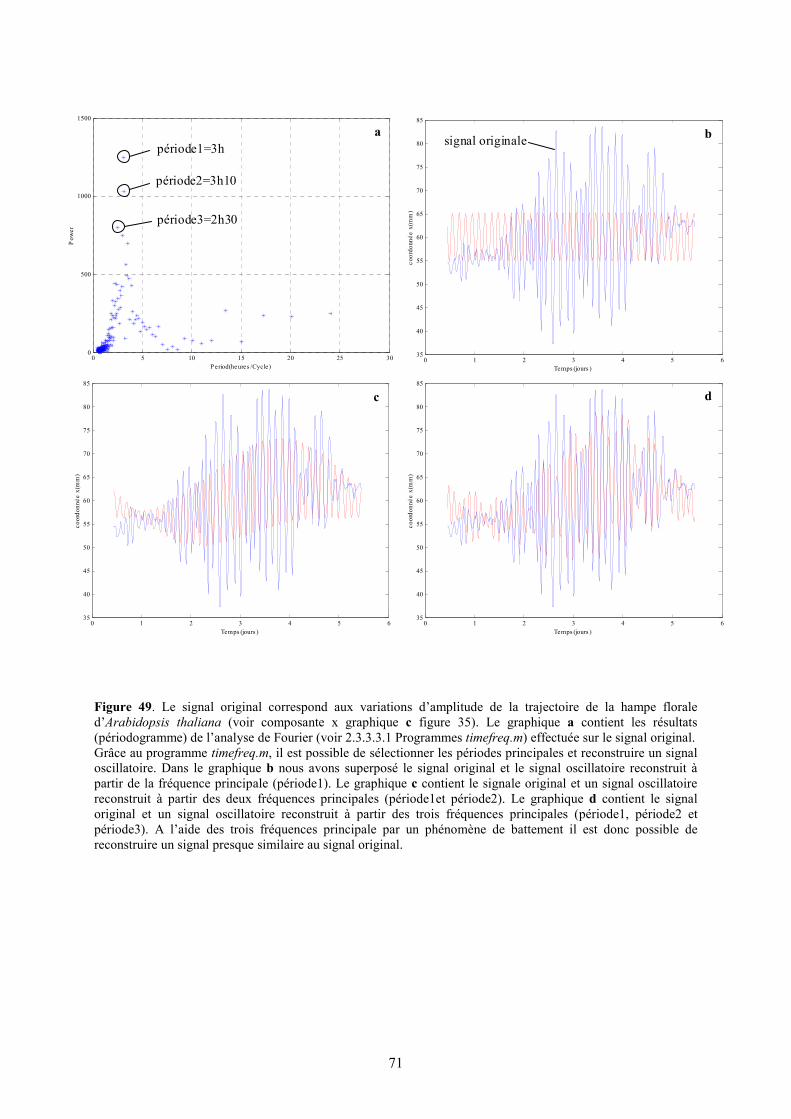
71
Figure 49. Le signal original correspond aux variations d’amplitude de la trajectoire de la hampe florale d’Arabidopsis thaliana (voir composante x graphique c figure 35). Le graphique a contient les résultats (périodogramme) de l’analyse de Fourier (voir 2.3.3.3.1 Programmes timefreq.m) effectuée sur le signal original. Grâce au programme timefreq.m, il est possible de sélectionner les périodes principales et reconstruire un signal oscillatoire. Dans le graphique b nous avons superposé le signal original et le signal oscillatoire reconstruit à partir de la fréquence principale (période1). Le graphique c contient le signale original et un signal oscillatoire reconstruit à partir des deux fréquences principales (période1et période2). Le graphique d contient le signal original et un signal oscillatoire reconstruit à partir des trois fréquences principales (période1, période2 et période3). A l’aide des trois fréquences principale par un phénomène de battement il est donc possible de reconstruire un signal presque similaire au signal original.
0 1 2 3 4 5 635
40
45
50
55
60
65
70
75
80
85
Temps (jours )
coor
donn
ée x
(mm
)
0 1 2 3 4 5 635
40
45
50
55
60
65
70
75
80
85
Temps (jours )
coor
donn
ée x
(mm
)
0 1 2 3 4 5 635
40
45
50
55
60
65
70
75
80
85
Temps (jours )
coor
donn
ée x
(mm
)
0 5 10 15 20 25 300
500
1000
1500
Pow
er
P eriod(heures /Cycle)
période1=3h
période2=3h10
période3=2h30
asignal originale b
c d

72
Comme nous venons de le voir beaucoup d’éléments semblent être impliqués dans ce phénomène dont nous n’avons pas encore une explication. Il se peut que ces mouvements d’oscillations rythmiques soient sous le contrôle d’un rythme endogène (génétique) comme pour les mouvements circadiens où cette relation a été démontrée (voir 3.1.3 Rythme circadien). Nous ne voulons pas ici avancer d’hypothèses, mais deux observations nous semblent dignes de citation. Nous avons remarqué que les variations d’amplitude de nos signaux oscillatoires (voir coordonnées x des figures 34, 35 et 39) sont composées de plusieurs fréquences (voir graphiques a des figures 48 et 49) dont la fréquence principale oscille autour des 3h. Nous avons remarqué que le signale original peut être reconstruit approximativement par un phénomène de battement entre les deux ou trois fréquences principales (voir figure 48 et 49). Pendant la discussion sur la variation des amplitudes nous n’avons pas pris en considération ce phénomène, mais il se pourrait que ces fréquences légèrement décalées correspondent à des décalages par exemple entre le déplacement transversal de l’auxine, le changement de direction des microtubules, l’onde de potassium, calcium et de l’eau. Le fait que ces ondes soient en phase produirait donc des grandes amplitudes et vice versa quand elles sont déphasées. Une autre relation qui nous paraît intéressante, est la comparaison entre la rotation d’un tuyau d’arrosage qui peut être plus ou moins rapide selon la vitesse du débit de l’eau et la circumnutation. Ces deux mouvements ont des ressemblances. Par cette constatation, comme nous l’avons déjà mentionné pour le cas des variations d’amplitude de la trajectoire, il se pourrait que la vitesse de l’eau qui parcourt la hampe florale joue en rôle dans les mouvements de circumnutation.
3.2.3.3.1 Synchronisation La synchronisation est un phénomène que l’on rencontre souvent dans la nature. Si la vie existe, c’est en partie dû à la synchronisation entre des différents événements dans l’espace-temps. Très souvent un élément perturbateur (ondes électromagnétiques, ondes sonores, stress mécanique) peut synchroniser toute une série de cascades de réaction, à l’intérieur d’un
être vivant, qui peuvent se manifester dans un comportement macroscopique. Dans les expériences où nous avons filmé simultanément deux plantes âgées de ~ 36 jours, nous avons remarqué que les hampes florales, de longueur similaire, présentent une dynamique générale similaire (voir graphique c et d ou e et f ou g et h de la figure 34). Le moment de l’arrosage (différent pour chaque couple mais equivalent pour les deux plantes en couple) pourrait être la cause de l’induction de la synchronisation au niveau de la forme générale de la trajectoire de la hampe vue latéralement. Les plantes étaient arrosées au commencement de l’expérience et après trois ou quatre jours. Il est possible que l’heure de l’arrosage soit aussi en facteur important. Il serait donc necessaire de faire des expériences on prenant en considération plus précisément le moment et la quantité d’eau donnée aux plantes. Un deuxième niveau de synchronisation a aussi été observe au niveau du sens de rotation. Les hampes florales d’Arabidopsis thaliana écotype Ws, comme nous avons découvert, peuvent tourner dans les deux sens (voir 3.1.1.3 Circumnutation). Nous avons pas pu déterminer le ou les facteurs qui déterminent le choix d’un sens plutôt que de l’autre. Nous avons quand même pu constater que les changements de sens se produisent normalment pendant une phase de faible amplitude de la trajectoire (voir comparaison entre les changements du sens de rotation (graphiques a et c figure 44) et la variation de l’amplitude (graphique a de la figure 42 et graphique a de la figure 43). Figure 50. Cette figure montre les quatre types de synchronisation possibles pour deux hampes florales qui circumnutent.
synchronisation (parallélisme)
synchronisation (rapprochement et éloignement)
a
b
c
d

73
Théoriquement il y a quatre types de synchronisation pour des mouvements de circumnutation (voir figure 50). Comme est visible dans le film (voir CD Synchro.avi) les deux hampes florales à un certain moment bougent de façon très synchrone. Dans la figure 51 nous avons superposé la dynamique des mouvements le long de l’axe x (voir coordonnées x graphiques e et f figure 36) des deux hampes florales filmées simultanément, avec la variation du sens de rotation pour la hampe de gauche. La figure 51 montre le moment où commence la synchronisation parallèle (voir type c figure 50). Nous somme tombé tout à fait par hasard sur ce phénomène. Il est difficile d’avancer des hypothèses avec le peu de résultats que nous avons à disposition, il se pourrait que ce phénomène soit le fruit du hasard, il est en tout cas intéressant de remarquer que la
synchronisation dans la figure 51 se fait au commencement de la phase lumineuse. Dans la littérature nous avons trouvé un cas où les auteurs ont observé de façon tout à fait fortuite la synchronisation et la mise en phase du mouvement de deux tiges de la Fève suite à un contact (Millet 1970). Les auteurs ont donc proposé que la synchronisation observée était manifestement la conséquence d’un phénomène d’entrainement mécanique ce qui n’est pas le cas pour notre expérience vu que les tiges ne se sont pas touchées. En filmant deux autres couples de plantes, pour mieux caractériser la circumnutation de la hampe florale d’Arabidopsis et la synchronisation nous avons constaté la présence de synchronisations de courte durée (quelque rotation), mais nous n’avons pas rencontré une synchronisation aussi longe telle que l’ont peut voir dans la figure 51.
Figure 51. Synchronisation parallèle en sens oraire (voir type c figure 48) des mouvements de deux hampes florales d’Arabidopsis thaliana écotype Ws. La courbe de la somme des angles (voir graphique c figure 44) montre les changements de sens de rotation pour la hampe florale de gauche. Le phénomène de synchronisation et visible dans le film Synchro.avi (voir CD).
-2500
-2000
-1500
-1000
-500
0
500
1000
1500
2000
2500
3000
0 1 2 3 4 50
10
20
30
40
50
60
70
80
90
Temps (jour)
Som
me
angl
e (d
egré
)
mm
(x)
Commencement de la synchronisationtype c voir figure 48.
Hampe florale de gauche Hampe florale de droite Somme des angles

74
Pour l’instant nous ignorons la cause d’un telle phénomèn, « il s’agit là d’un domaine neuf et on est en droit d’espérer, d’une expérimentation systématique, des surprises intéressantes » (Millet 1970). Une communication chimique ou électromagnétique (Popp 1989) entre les plantes pourrait aussi être un facteur de synchronisation, actuellement encore mal connu. CONCLUSION Cycle de vie Le cycle de vie d’Arabidopis commence par le contact de la graine avec l’eau. C’est à ce moment que la graine se gonfle et son métabolisme change. Très rapidement apparaît la petite radicule (~3 jours après l’imbibition) qui va pénétrer dans le sol par des mouvements de circumnutation. L’hypocotyle se dresse pour atteindre sa position verticale et amener avec lui les deux cotylédons (~5 jours après l’imbibition) qui vont sortir en dernier de l’enveloppe protégeant la plantule pendant sa phase de dormance. Très tôt les deux cotylédons commencent à se lever et à se baisser comme les ailes d’un papillon (~6 jours après l’imbibition). C’est un mouvement rythmique circadien endogène qui peut être synchronisé par le cycle jour nuit issus de la rotation de la terre sur elle-même. La croissance de l’hypocotyle, l’ouverture des stomates, la synthèse de la chlorophylle,… fluctuent aussi avec une rythmicité circadienne. L’axe de l’hypocotyle effectue aussi des petits mouvements rythmiques ultradiens de rotation et de pendulation bien visibles pendant la phase de croissance de l’hypocotyle. L’apparition des premières feuilles ne se fait pas attendre (~9 jours après l’imbibition). Tout gentiment la spirale génératrice de la rosette se construit avec des nouvelles formes de feuilles qui apparaissent soigneusement distribués. Les feuilles les plus âgées diminuent l’amplitude de leurs mouvements rythmiques circadiens, maintenus par les nouvelles feuilles. A un certain moment, la rosette (~20 jours après l’imbibition) devient trop grande et lourde pour que l’hypocotyle puisse rester vertical, ainsi la plante tombe (~7h) sur un côté pour
ensuite retrouver un nouvel équilibre après ~7 jours. Pendant tout ce temps le rythme circadien endogène (~ 25h 30min) des mouvements des feuilles est toujours présent. Mais les feuilles ne font pas que ces mouvements. A l’intérieur de cette oscillation circadienne elles effectuent aussi des mouvements rythmiques ultradiens (~ 2h) de plus faible amplitude. En plus, leur expansion (croissance) est régulée par un rythme circadien endogène (~25h). Environ 36 jours après l’imbibition de la graine, une tige centrale (hampe florale) portant les jeunes fleurs apparaît. La hampe croît aussi avec un rythme circadien endogène et elle effectue aussi des mouvements de rotation et pendulation rythmiques (~3h) en sens horaire et anti-horaire qui ont tendance à présenter une variation circadienne de l’amplitude. En effectuant ces mouvements de circumnutation la hampe varie la position de la surface de ces feuilles par rapport à la source lumineuse. Ce qui induit une variation au niveau des taux photosynthétiques à l’intérieur de chaque feuille. Cette variation, comme les mouvements ultradien des feuilles de la rosette, pourrait être importante au niveau de l’optimalisation de la photosynthèse. Rapidement des inflorescences primaires et latérales apparaissent à la base des feuilles caulinaires et à la base de la hampe. Ces inflorescences présentent aussi des mouvements rythmiques ultradien. Pendant le développement de la hampe les bourgeons floraux se développent en fleurs qui vont ensuite se transformer en fruits (silique) à l’intérieur desquels des graines sont prêtes pour effectuer un autre cycle de vie. Interconnections rythmiques L’avantage de la méthode que nous avons utilisées, est qu’elle permet d’étudier simultanément la dynamique de différentes partie de la plante de façon non invasive et ainsi pouvoir étudier leur interconnections. Comme est illustré dans la figure 52, à l’aide de trois traitements d’images différents (area.m, long.m et invlong.m) nous avons pu détecter simultanément quatre dynamiques oscillatoires sur la même plante : mouvements rythmiques circadiens des feuilles de la rosette, croissance rythmique circadienne du premier entrenœud, mouvement de circumnutation du sommet de la hampe florale et la variation

75
rythmique d’amplitudes de ces mouvements. La survie d’un individu dépend du caractère plastique et de la synchronisation des rythmes entre eux. Le rythme de la respiration et le rythme cardiaque chez l’homme sont de très bons exemples pour comprendre l’importance de l’interconnexion des rythmes et leur plasticité. L’interconnexion entre les rythmes découle de relations très intimes et fondamentales. Dans le futur il sera donc très important et intéressant d’étudier les liens entre les rythmes. En effet ils jouent un rôle certainement important au niveau des relations entre les parties qui composent un individu, dans les relations entre être vivants différents et au niveau des relations des êtres vivant avec le cosmos. Par notre étude nous avons pu remarquer l’importance du rythme circadien au niveau du
cycle de vie d’Arabidopsis. Ce rythme est intimement lié au rythme de rotation terrestre qui détermine l’apparition et la disparition de la lumière du soleil. Nous savons que les plantes utilisent l’énergie lumineuse du soleil pour se construire (organismes autotrophes). L’importance d’une régulation rythmique circadienne à plusieurs niveau dans le cycle de vie de la plante, qui découle d’une co-évolution entre la plante et son environnement, montre l’importance des interconnections rythmiques entre le micro cosmos et le macro cosmos pour l’existence de la vie.
Figure 52. Détection de 4 rythmes sur la même plante d’Arabidopsis (voir CD circumH.avi) en régime photopériodique L:D (12:12). 1) Mouvements rythmiques circadiens des feuilles détectées avec le programme area.m. 2) Croissance rythmique circadienne du premier entrenœud détecté avec le programme long.m. 3) Mouvements de circumnutation du sommet de la hampe floral détecté avec le programme invlong.m. 4) Variations rythmiques de l’amplitude des mouvements de circumnutation détecté avec le programme invlong.m.
-10
-8
-6
-4
-2
0
2
4
6
8
10
0 20 40 60 80 100 120 140 1600
1
2
3
4
5
6
Temps (heures)
cm
mou
vem
ents
de
la h
ampe
(cm
)m
ouve
men
ts d
es fe
uille
s (cm
2 )
1 2 3 et 4

76





























