Embed Size (px)
Citation preview

P/am Physiol. Biochmz.. 1998, 36 (3), 263-26X
Short paper
Accumulation of plastidial alanine-aminopeptidase in relation to plastid damage in cotyledons of dark-grown sugar beet seedlings
Abdelhak El Amrani ‘, Bilal Camara ‘, Jean-Pierre Gaudillcre ‘, Ivan Couee lY3*
’ Station de Physiologie VCg&tale, Institut National de la Recherche Agronomique, Centre de Recherches de Bordeaux, BP 81, 33883 Villenave d’Ornon cedex, France. ’ lnstitut de Biologie MolCculaire des Plantes. Centre National de la Recherche Scientifique. 12, rue du GCnCral Zimmer, 67084 Strasbourg cedex, France. ’ Present address: UnitC de Physiologie, Station d’Am&lioration des Arbres Forestiers, Institut National de la Recherche Agronomique, Centre de Recherches d’OrlCans, 45 160 Ardon, France. *Author to whom correspondence should be addressed (fax 33-238417879: e-mail [email protected])
(Received March 27, 1997; accepted June 19. 1997)
Abstract - During prolonged dark-growth of sugar beet (Beta ~r&~lris L.) seedlings, the etioplasts of cotyledons undergo a degenerative process shortly after the proplastid to etioplast transition. Two plastidial aminopeptidase activities were previously identified as early markers of this degenerative process which results in a decrease of greening capacity. One of these activities was shown to be due to one enzyme, plastidial alanine-aminopeptidase, which was purified to homogeneity and characterized. In the present study, the effects of dark development, light. and exogenous glucose on enzyme activity and protein level of plas- tidial alanine-aminopeptidase were investigated. Changes of enzyme activity during prolonged dark-growth were found to be correlated with changes of protein level. Significant increases of both activity and protein levels occurred at the onset of the degenerative process affecting etioplasts. By contrast, the activity and protein levels of plastidial alanine-aminopeptidase were negatively affected by light and by exogenous glucose. thus suggesting a type of expression that is related to prolonged dark- growth and etioplast ageing. 0 Elsevier. Paris.
Aminopeptidasel cotyledon I dark-growth I greening / plastid I protease I Beta vulgaris
ICL, isocitrate lyase/ pAla-AP, plastidial Ala-aminopeptidase
1. INTRODUCTION
Cotyledons of plants that exhibit epigeal germina- tion have often been used as an experimental system to investigate metabolic and developmental changes from a phase of het’erotrophic growth through pho- totrophic growth up to a stage of senescence. espe- cially because these changes take place in the same organ with limited cell division [3]. We described pre- viously the sequence of metabolic events [9] and the structural and bioche:mical modifications of proplas- tids and etioplasts [Sl in cotyledons during prolonged dark-growth of sugar beet seedlings. After proplastid- to-etioplast transition, etioplasts underwent a degener- ative process characterized by: (a) ultrastructural modifications affecting prolamellar bodies and pro- thylakoid membranes, (b) depletion of plastidial pro-
teins, (c) decrease in the capacity of carotenoid and chlorophyll accumulation. Protochlorophyllide oxi- doreductase protein decreased during prolonged dark- growth [8], in accordance with the repression of the two genes encoding protochlorophyllide oxidoreduc- tase, Pot-A and PorB, during prolonged etiolation of Arabidopsis thaliana seedlings [ 11. However, protein degradation also affected the small and large subunits of RUBISCO, and the a and p subunits of chloroplast ATPase [S] Thus, the decline of greening capacity seemed to reflect a general degradative process rather than a specific degradation of protochlorophyllide oxidoreductase. Remarkably, this type of dark devel- opment shared some similarities with the effects of far-red light on greening of Arabidopsis sp. seedlings whereby depletion of protochlorophyllide oxidoreduc- tase protein is accompanied by degenerative processes
Plant Phyriol. Biochem., 098 I -9428/98/03/C) Elaevier, Paris

264 A. El Amrani et al.
and the failure of plastids to accumulate prolamellar bodies [2].
We identified previously two plastidial aminopepti- dase activities as early markers of this degenerative process [8]. One of these markers, plastidial Ala-ami- nopeptidase activity, was shown to decrease upon exposure to light or addition of exogenous glucose, thus confirming the relationship with prolonged dark- growth and indicating a relationship with glucose limi- tation. We showed that this activity was due to one enzyme, pAla-AP, which was purified to homogeneity and characterized [lo]. Its kinetic and physico-chemi- cal characterization as a dimeric metallo-aminopepti- dase of 150 kDa showed that it was a novel enzyme, presumably differing from the previously described classes of plant aminopeptidases, such as hexameric Leu-aminopeptidases and monomeric aminopepti- dases [12].
These and other findings from the literature implied that plastids possess a variety of proteases. In this respect, earlier work on subcellular localization of activities [20, l!)] has been complemented by direct immunolocalization of pAla-AP in sugar beet [lo], of ATP-dependent proteases in Aruhidopsis sp. [28], and of a light-induced protease in barley [24]. These pro- teases are likely to play important roles in the biogene- sis, house-keeping functions and plasticity of plastids, especially in relation to light and carbohydrate status of the cell [24, ;I]. In the present study. the following effects were investigated: darkness, developmental state of the plastid, light, exogenous glucose. on enzyme activity and protein level of pAla-AP. Accu- mulation of pAla-AP is shown to be related to pro- longed dark-growth and etioplast ageing, whereas light and exogenous glucose decreased its steady-state level.
2. RESULTS AND DISCUSSION
2.1. Accumulation of pAla-AP in cotyledons during prolonged dark-growth
Sugar beet seedlings display the characteristic tem- poral pattern of post-germinative development of coty- ledons in epigeal plants. This pattern includes mobilization of lipid reserves with maximum activity between d 3 and d 5 [I 8, 91, gluconeogenesis with maximum activity at d 4 [ 18. 91, accumulation of nutrients in cotyledons and export to the hypocotyl with a maximum at d 6 [7], and establishment of pho- tosynthetic capacity with a maximum at d 6 18. 91, which, under illumination conditions, is followed by
autotrophic growth and subsequent senescence. Pro- longed dark-growth after d 6 results in a final stage of development that was characterized by modifications at the structural, metabolic and protein-expression lev- els, and ultimately led to plastids with decreased greening capacity [8, 91. Nearly all the parameters that were examined in our previous studies indicated that there was a pivotal event occurring around d 6 that led to etioplast degradation. We hypothesized that the expression of pAla-AP might be correlated with this event. In order to test this hypothesis, western blot analysis was performed with polyclonal antibodies raised against pAla-AP [I 01.
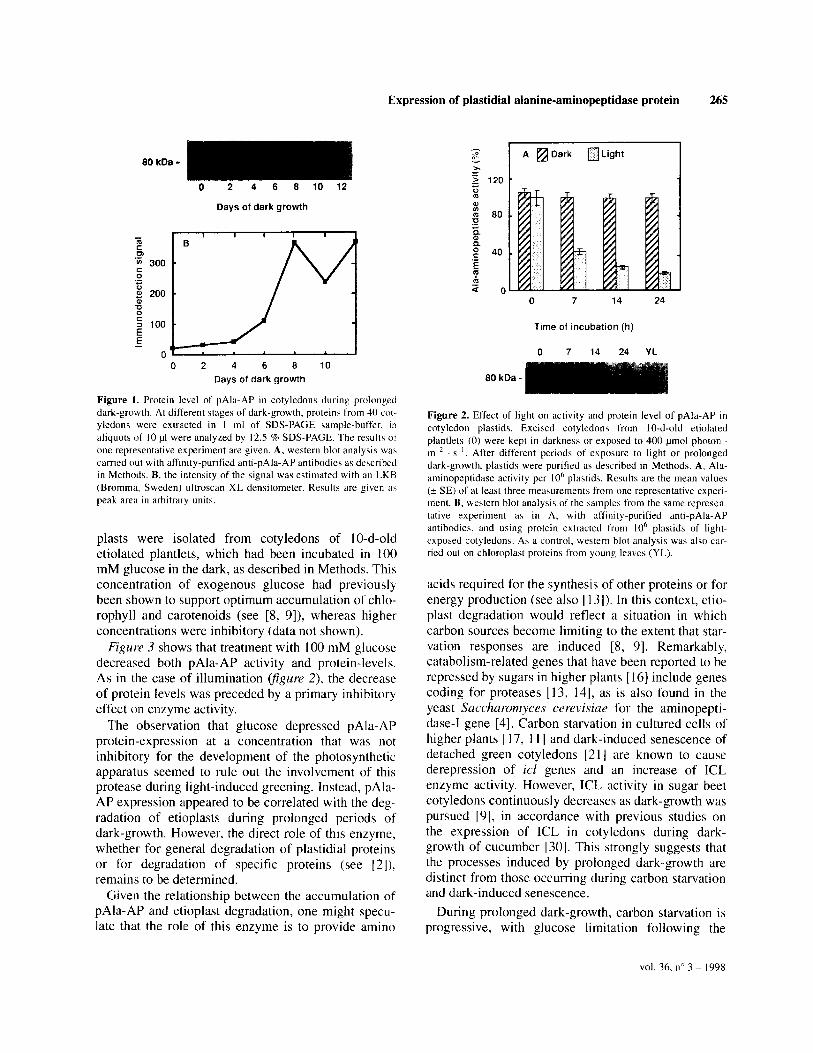
As shown infigure I, these antibodies indeed recog- nized the 80-kDa subunit of this enzyme [lo]. How- ever, for immunodetection of pAla-AP at different stages of development, the polyclonal antibodies were purified by affinity binding, as described in Methods. Using this purified antibody, a low level of pAla-AP was already detectable in dry seeds (figure I A). During dark-growth, pAla-AP protein levels then increased, in accordance with the increase of pAla-AP activity per plastid, as shown previously [8]. This apparent increase of pAla-AP signal between d 4 and d 8 was verified by scanning and quantification figure 1 B).
The relationship between pAla-AP expression and prolonged dark-growth was confirmed by testing the effect of light on pAla-AP protein level and activity. Plastids were isolated from cotyledons of IO-d-old eti- olated plantlets, which had been exposed to light for up to 24 h, as described in Methods. Figure 2 shows that light treatment caused a decrease in both pAla-AP activity and protein-level. Neither the activity nor the protein were detectable in chloroplasts of young leaves. However, the delay between the decrease in activity and the decline of protein level suggested that illumination primarily exerted a negative effect on enzyme activity on which degradation of pAla-AP and/or repression of its synthesis were later superim- posed.
2.2. Effect of exogenous glucose on the steady- state levels of pAla-AP
Exogenous sugars have been shown to maintain the greening capacity [2] or to facilitate the greening of dark-grown seedlings upon illumination [X, 91). How- ever, the phenomenon of sugar repression of chloro- plast development has also well been documented (see [29]). We therefore investigated the effect of exoge- nous glucose on the protein level of pAla-AP. Etio-
Expression of plastidial alanine-aminopeptidase protein 265
80 kDa -
Days of dark growth
0 2 4 6 8 10
Days of dark growth
Figure I. Protein level of pAla-AP in cotyledons during prolonged dark-growth. At different stages of dark-growth, protein\ from 40 cot yledons were extracted in 1 ml of SDS-PAGE sample-buffer, in
aliquota of IO pl were analy.zed by 12.5 % SDS-PAGE. The results ot one representative experiment are given. A, western blot analybis was
carried out with affinity-purified anti-pAla-AP antibodies as described in Methods. B, the intensity of the signal was estimated with an LKB
(Bromma, Sweden) ultroscan XL densitometer. Results are given a\ peak area in arbitrary unity.
plasts were isolated from cotyledons of IO-d-old etiolated plantlets, which had been incubated in 100 mM glucose in the dark, as described in Methods. This concentration of exogenous glucose had previously been shown to support. optimum accumulation of chlo-, rophyll and carotenoids (see [8, 9]), whereas higher concentrations were inhibitory (data not shown).
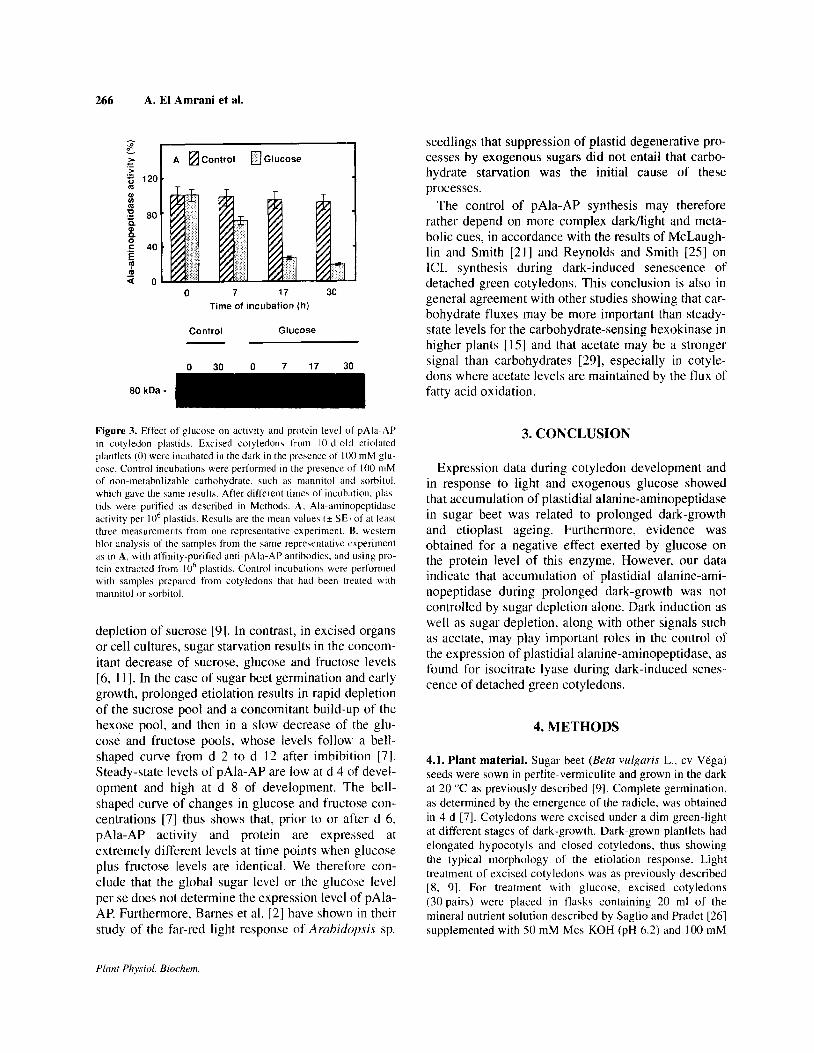
Figure 3 shows that treatment with IO0 mM glucose decreased both pAla-AP activity and protein-levels. As in the case of illurnination (figure 2), the decrease of protein levels was preceded by a primary inhibitory effect on enzyme activity.
The observation that glucose depressed pAla-AP protein-expression at a concentration that was not inhibitory for the development of the photosynthetic apparatus seemed to rule out the involvement of this protease during light-induced greening. Instead, pAla- AP expression appeared to be correlated with the deg- radation of etioplasts, during prolonged periods of dark-growth. However, the direct role of this enzyme, whether for general degradation of plastidial proteins or for degradation of specific proteins (see [2]). remains to be determined.
Given the relationship between the accumulation of pAla-AP and etioplasl degradation, one might specu- late that the role of this enzyme is to provide amino
2 A q Dark q Light 1
E :5 120
zi
24 m 80
0 x
i! .r 40
5 i a 0
0 7 14 24
lime of incubation (h)
0 7 14 24 YL
80 kDa -
Figure 2. Effect of light on activity and protein level of pAla-AP in
cotyledon plastids. Excised cotyledon\ from IO-d-old etiolated plantlets (0) were kept in darkness or exposed to 400 pmol photon m-’ ‘s ‘. After different periods of exposure to light or prolonged
dark-growth, plastids were purified as described in Methods. A, Ala- aminopeptidase activity per IO“ plastids. Results are the mean values
(2 SE) of at least three measurements from one representative experi- ment. B. western blot analysis of the samples from the same represen- tative experiment as in A. with affinity-purified anti-pAla-AP
antibodies. and using protein extracted from IO’ plastids of light- exposed cotyledons. As a control, western blot analysis was also car- ried out on chloroplast proteins from young leaves (YL).
acids required for the synthesis of other proteins or for energy production (see also [ 131). In this context. etio- plast degradation would reflect a situation in which carbon sources become limiting to the extent that star- vation responses are induced [8, 91. Remarkably, catabolism-related genes that have been reported to be repressed by sugars in higher plants [ 161 include genes coding for proteases [13, 141, as is also found in the yeast Saccharornyces cerevisiae for the aminopepti- dase-I gene [4]. Carbon starvation in cultured cells of higher plants [ 17. 111 and dark-induced senescence of detached green cotyledons [21] are known to cause derepression of id genes and an increase of ICL enzyme activity. However, ICL activity in sugar beet cotyledons continuously decreases as dark-growth was pursued [9], in accordance with previous studies on the expression of ICL in cotyledons during dark- growth of cucumber [30]. This strongly suggests that the processes induced by prolonged dark-growth are distinct from those occurring during carbon starvation and dark-induced senescence.
During prolonged dark-growth, carbon starvation is progressive, with glucose limitation following the
vol. 36. Ilo 3 - I998
266 A. El Amrani et al.
e z. ?z I
A HControl 0 Glucose %
0 7 17 30
Time of incubation (h)
Control Glucose
130 kDa -
7 17 30
OYI Figure 3. Effect of glucose on activity and protein level of pAla-AP in cotyledon plastids. Excised cotyledon\ from IO-d-old etioluted
plantlets (0) were incubated in the dark in the presence of I00 mM glu- cose. Control incubations were performed in the presence of 100 mM of non-metabolizable carbohydrate, such as mannitol and sorbitol,
which gave the same results. After different times of incub.ltion, play- tids were purified as described in Methods. A, Ala-aminopeptidase
activity per IO6 plastids. Results are the mean value\ (i. SEI of at least three measurements from one representative experiment. K. western
blot analysis of the TampIes from the same representative experiment as in A, with aftinity-purified anti-pAla-AP antihodie\. and using pro- tein extracted from IO’ pla’;tids. Control incubation\ were perlormed with samples prepared from cotyledom that had been treated with
mannitol or sorbitol.
depletion of sucrose [9]. In contrast. in excised organs or cell cultures, sugar starvation results in the concom- itant decrease of sucrose, glucose and fructose levels [6, 1 I]. In the case of sugar beet germination and early growth, prolonged etiolation results in rapid depletion of the sucrose pool and a concomitant build-up of the hexose pool, and then in a slow decrease of the glu- cose and fructose pools, whose levels follow a bell- shaped curve from d 2 to d 12 after imbibition [7]. Steady-state levels of pAla-AP are low at d 4 of devel- opment and high at d 8 of development. The bell- shaped curve of changes in glucose and fructose con- centrations [7] thus shows that, prior to or after d 6, pAla-AP activity and protein are expressed at extremely different levels at time points when glucose plus fructose levels are identical. We therefore con- clude that the global sugar level or the glucose level per se does not determine the expression level of pAla- AP. Furthermore, Barnes et al. [2] have shown in their study of the far-red light response of Amhidopsis sp.
seedlings that suppression of plastid degenerative pro- cesses by exogenous sugars did not entail that carbo- hydrate starvation was the initial cause of these processes.
The control of pAla-AP synthesis may therefore rather depend on more complex dark/light and meta- bolic cues, in accordance with the results of McLaugh- lin and Smith [21] and Reynolds and Smith [25] on ICL synthesis during dark-induced senescence of detached green cotyledons. This conclusion is also in general agreement with other studies showing that car- bohydrate fluxes may be more important than steady- state levels for the carbohydrate-sensing hexokinase in higher plants [ 151 and that acetate may be a stronger signal than carbohydrates [29], especially in cotyle- dons where acetate levels are maintained by the flux of fatty acid oxidation.
3. CONCLUSION
Expression data during cotyledon development and in response to light and exogenous glucose showed that accumulation of plastidial alanine-aminopeptidase in sugar beet was related to prolonged dark-growth and etioplast ageing. Furthermore, evidence was obtained for a negative effect exerted by glucose on the protein level of this enzyme. However, our data indicate that accumulation of plastidial alanine-ami- nopeptidase during prolonged dark-growth was not controlled by sugar depletion alone. Dark induction as well as sugar depletion, along with other signals such as acetate, may play important roles in the control of the expression of plastidial alanine-amihopeptidase. as found for isocitrate lyase during dark-induced senes- cence of detached green cotyledons.
4. METHODS
4.1. Plant material. Sugar beet (Bela v~l~aris L.. cv VCga) seeds were sown in perlite-vermiculite and grown in the dark at 20 “C as previously described [9]. Complete germination, as determined by the emergence of the radicle, was obtained in 4 d [7]. Cotyledons were excised under a dim green-light at different stages of dark-growth. Dark-grown plantlets had elongated hypocotyls and closed cotyledons, thus showing the typical morphology of the etiolation response. Light treatment of excised cotyledons was as previously described [S, 91. For treatment with glucose, excised cotyledons (30 pairs) were placed in flasks containing 20 ml of the mineral nutrient solution described by Saglio and Pradet [26] supplemented with 50 mM Mes-KOH (pH 6.2) and 100 mM
Plant Physiol. Biochem.

Expression of plastidial alanine-aminopeptidase protein 267
glucose, and incubated in the dark for the periods given in the figure legends. The flasks were bubbled continuously with air and solutions changed every 8 h.
4.2. Isolation of plastids. Plastids were isolated from cotyle- dons of dark-grown seedlings, and from light- or glucose- treated cotyledons of IO-d-old etiolated seedlings, as described in El Amrani et al. [8]. The number of plastids per volume unit was determined by visual counting in a haemocytometer.
4.3. Assay of aminopeptidase and analysis of proteins. Ala-aminopeptidase activity was assayed using L-Ala-:!- naphthylamide as substrate, as previously described 181. Pro- tein was determined by the method of Bradford [S] using ‘y- globulin as the standard. SDS-PAGE of proteins was carried out in a Mighty Small II (Hoefer. San Francisco, CA) elec- trophoresis unit in the presence of 0.1 % (w/v) SDS, essen- tially as described by O’Farrell [23].
4.4. Immunochemical studies. Polyclonal antibodies against purified pAla-AP were raised in Fauve de Bourgogne rabbits [IO]. Anti-pAla-AP antibodies were further purified by eluting the antibodies bound to nitrocellulose-fixed pAla- AP, as described by Sambrook et al. [ 271. Western blot analy- sis was carried out with the antiserum (diluted IOOO-fold) or with the preparation of affinity-purified antibodies (diluted IO-fold) as described previously [IO].
Acknowledgements. This work was partly funded by the Institut Technique FranGais de la Betterave Industrielle. We thank Mr Philippe Mire for raising the antibodies against plastidial alanine-aminopeptidase. We also wish to thank the reviewers for their thorough work and their advice.
REFERENCES
[l] Armstrong G. A., Runge S., Frick G., Sperling U., Ape1 K., Identification of NADPH:protochlorophyllide oxidoreductases A and B: a branched pathway for light-dependent chlorophyll biosynthesis in Arabidol-,- sis thaliana, Plant Physiol. 108 (1995) 15051517.
[2] Barnes S. A., Nishizawa N. K., Quagglo R. B., Whitc- lam G. C., Chua N-H., Far-red light blocks greening of Arabidopsis seedlings via a phytochrome A-medi- ated change in plastid development. Plant Cell 8 (1996) 601-615.
[3] Becker W. M., Leaver C. J., Weir E. M., Riezman H.. Regulation of glyoxysomal enzymes during germina- tion of cucumber. [. Developmental changes in coty,le- donary protein, RNA and enzyme activities durmg germination, Plant Physiol. 24 (1978) I -IS.
[4] Bordallo J., Suarez-Rendueles P.. Identification of reg- ulatory proteins that might be involved in carbon catabolite repression of the aminopeptidase-I gene of the yeast Sacchuromyw ccrevisiae. FEBS Lett. 376 (1995) 120-124.
[5] Bradford M. M., A. rapid and sensitive method for the quantification of microgram quantities of protein utiliz-
ing the principle of protein-dye binding, Anal. Bio- them. 72 (1976) 248-252.
[6] Brouquisse R., James F., Raymond I?, Pradet A., Study of glucose starvation in excised maize root tips, Plant Physiol. 96 ( 199 1) 6 19-626.
[7] El Amrani A.. Saglio P., Raymond I?, Nature and utili- zation of seed reserves during germination and het- erotrophic growth of young sugar beet seedlings. Seed Sci. Res. 2 (1992) I-8.
[ 81 El Amrani A.. Co&e I., Carde J.-P., Gaudilliire J.-P., Raymond P., Modifications of etioplasts in cotyledons during prolonged dark growth of sugar beet seedlings: identification of etiolation-related plastidial aminopep- tidase activities, Plant Physiol. 106 ( 1994a) 1555 1565.
[9] El Amrani A., Gaudillkre J.-P., Raymond I?. Carbohy- drate starvation is a major determinant of the loss of greening capacity in cotyledons of dark-grown sugar beet seedlings, Physiol. Plant. 90 (1994b) 56-64
[IO] El Amrani A., Suire C.. Camara B.. Gaudill&re J.-P., CouCe I., Purification and characterization of a novel aminopeptidase, plastidial alanine-aminopeptidase, from the cotyledons of etiolated sugar beet seedlings, Plant Physiol. 109 (1995) 87-94.
[I I] Graham I. A., Denby K. J., Leaver C. J., Carbon catab- olite repression regulates glyoxylate cycle gene expres- sion in cucumber, Plant Cell 6 (1994) 761-772.
1121 Herbers K., Prat S., Willmitzer L., Functional analysis of a leucine aminopeptidase from Snlanum tuhemsrrm L., Planta 194 (1994) 230-240.
[ 131 James F., Brouquisse R., Pradet A., Raymond P., Changes in proteolytic activities in glucose-starved maize root tips, Plant Physiol. Biochem. 31 (1993) 845 856.
[14] James F., Brouquisse R.. Suire C., Pradet A., Raymond P., Purification and biochemical characteriza- tion of a vacuolar serine endopeptidase induced by glu- cose starvation in maize roots, Biochem. J. 320 (1996) 283-292.
115 1 Jang J. C., Sheen J., Sugar sensing in higher plants, Plant Cell 6 ( 1994) I665- 1679.
[I h] Koch K. E., Carbohydrate-modulated gene expression in plants, Annu. Rev. Plant Physiol. Plant Mol. Biol. 47 ( 1996) 509-540.
1171 Kudielka R. A., Theimer R. R., Derepression of glyox- ylate cycle enzyme activities in anise suspension cells, Plant Sci. Lett. 3 1 (1983) 237-244.
[ 181 Lawrence D. M., Halmer P.. Bowles D. J.. Mobilization of storage reserves during germination and early seed- ling growth of sugar beet, Physiol. Plant. 78 (1990) 42 I-429.
[ 191 Liu X. Q., Jagendorf A. T., Neutral aminopeptidases in the stroma of pea chloroplasts, Plant Physiol. 8 1 (1986) 603-608.
[20] Malek L., Bogorad L., Ayers A. R., Goldberg A. L., Newly synthesized proteins are degraded by an ATP- stimulated proteolytic process in isolated pea chloro- plasts. FEBS Lett. 166 (1984) 253-257.
[21] McLaughlin J. C., Smith S. M.. Metabolic regulation of glyoxylate-cycle enzyme synthesis in detached cucum-
vol. 36, no 3 - I998

268 A. El Amrani et al.
ber cotyledons and protoplasts, Planta 195 ( 1994) 22. 28.
[22] Mullet J. E., Chloroplast development and gene expres- sion, Ann. Rev. Plant Physiol. Plant Mol. Biol. 39 (1988) 475-502.
[23] O’Farrell P H., High resolution two-dimensional electro- phoresis of proteins, J. Biol. Chem. 250 (1975) 4007- 4021.
(241 Reinbothe C., Ape1 K., Reinbothe S., A light-induced protease from barley plastids degrades NADPH:pro- tochlorophyllide oxidoreductase complexed with chlo- rophyllide, Mol. Cell. Biol. 16 (1995) 6206-6212.
[25] Reynolds S .J., Smith S. M., Regulation of expression of the cucumber isocitrate lyase gene in cotyledons upon seed germination and by sucrose, Plant Mol. Biol. 29 (1995) 885896.
[26] Saglio P. H., Pradet A., Soluble sugars. respiration and energy charge during ageing of excised maize root tips, Plant Physiol. 66 (1980) 5 16-5 19.
[27] Sambrook J., Fritsch E. F., Maniatis T., Molecular clon- ing, a laboratory manual. Edition 2, Cold Spring Har- bor Laboratory Press, Cold Spring Harbor, New York 1989.
[28] Shanklin J., Dewitt N. D., Flanagan J. M., The stroma of higher plant plastids contains ClpP and ClpC, func- tional homologs of Escherichia coli ClpP and ClpA, an archetypal two-component ATP-dependent protease, Plant Cell 7 (1995) 1713-1732.
[29] Sheen J., Metabolic repression of transcription in higher plants, Plant Cell 2 (1990) 1027-1038.
[30] Weir E. M., Riezman H., Grienenberger J.-M., Becker W. M., Leaver C. J., Regulation of glyoxysomal enzymes during germination of cucumber: temporal changes in translatable mRNAs for isocitrate lyase and malate synthase, Eur. J. Biochem. 112 (1980) 469-477.
Plant Physiol. Biochem.





























