Embed Size (px)
Citation preview

RAPPORT DE STAGE Assistante en organisation d’évènements
Mars 2016 – Octobre 2016
DUPONT MARIE Master Communication – D2P3
Enseignant référant : M.CAMILERIE
Superviseur de stage : Mme CASTEL
Entreprise d’accueil : SWIFT Ltd
Etablissement : Université Toulouse Capitole 1, 31000 Toulouse
Analyse de la répartition de bactéries codant le système dedétoxication TPMT (Thiopurine-S-méthyltransférase) en fonction dela présence de médicaments et des résistances conférées parl’expression de l’enzyme
Février- Juillet2018
BOUCHALIRayanMaster2IMHE– HMBA403
Directeurdestage:COURNOYERBenoit([email protected])
Co-encadrants:EmilieBOURGEOISBenjaminYOUENOU
Laboratoired’accueil:UMRd’écologiemicrobienne– équipeBPOE
SamuelLacaille ([email protected])


1
Remerciements
Je tiens tout d’abord à remercier mon directeur de stage, Monsieur Cournoyer Benoit
pour m’avoir accueilli dans son laboratoire, mais aussi pour son encadrement et ses conseils
précieux. Merci d’avoir été disponible pour répondre à mes questions et mes besoins à
n’importe quel moment. Je remercie également Monsieur Samuel Lacaille de m’avoir accueilli
au sein de son équipe ainsi que pour avoir encadrer ma formation. Merci à l’équiper d’égoutiers
de la ville de Lyon de m’avoir accueilli sur le terrain et de m’avoir expliqué leurs différentes
fonctions.
Je remercie aussi mes deux co-encadrants, Benjamin Youenou et Emilie Bourgeois,
pour m’avoir formé tout au long de mon stage et avoir répondu à toutes mes questions et pour
les discussions scientifiques ou non. Merci aux collaborateurs Jérome Labanowski (Univ.
Poitiers, UMR 7285) et Agnès Bouchez (INRA Thonon) qui ont rendu disponibles les jeux de
données physico-chimiques et de dosage de médicaments. Merci également pour votre
gentillesse et votre bonne humeur précieuse tout au long de mon stage. Merci également à
Sébastien Ribun et Laurence Marjolet pour leurs contributions dans l’acquisition et l’analyse
des données NGS, mais aussi pour tous leurs conseils techniques au laboratoire et leur
sympathie. Merci à Sylvie Elsen (CEA Grenoble, UMR 1036) pour m’avoir fourni les
constructions de P. aeruginosa PAO1.
Je remercie également toute l’équipe BPOE de m’avoir accueilli amicalement, mais
aussi pour l’ambiance qui régnait au sein du laboratoire.


3
Liste des abréviations utilisées
TPMT : Thiopurine Méthyltransferase
bTPMT : Thiopurine Méthyltransferase bactérienne
hTPMT : Thiopurine Méthyltransferase humaine
6-MP : 6-mercaptopurine
ADN : Acide Désoxyribonucléique
ARN : Acide Ribonucléique
ARNm : Acide Ribonucléique messagers
HGPRT : Hypoxanthine Phosphoribosylguanine transférase
PCR : Polymerase Chain Reaction
Se : Sélénium
Te : Tellurium
S : Soufre
DMTe : di-méthylTelluride
DMDTe : di-méthyldiTelluride
DMS : di-méthylSulfur
DMDS : di-méthyldiSulfur
H2S : Sulfure d’Hydrogène
NGS : New Generation Sequencing
Pb : Paire de bases
OTUs : Unité Taxonomique Opérationnelle
°C : Degrés Celsius
tpm : tour par minutes
mL : milliLitre
µL : microLitre
mM : milliMolaire
M : Molaire
ng : nanogramme
V : Volt
min : minute
UV : Ultra-Violets
LB : Luria-Bertani
BET : Bromure d’éthidium
nm : nanométre
CFU : Unité Formant Colonie
Step : Station D’épuration
CMI : Concentration minimales inhibitrice
EU : Eau usées


4
1. Introduction 1.1. Les risques liés au métier d’égoutier
Les égoutiers (ou agents d’assainissement) entretiennent et nettoient les canalisations ou transitent les
eaux usées (de différentes provenances, urbaines, hospitalières, industrielles …) de la source jusqu’à la
station d’épuration. Cela inclut les stations de relèvement et de pompage. De nombreuses études font
état d’une baisse significative de l’espérance de vie des personnes exerçant le métier d’égoutiers (jusqu’à
17 ans de moins que le reste de la population). En effet, une étude menée à Paris (sur une cohorte de
1182 sujets, actifs ou à la retraite) indique que la population d’égoutiers montre une surmortalité de 56%
(et jusqu’à 97% pour les tranches d’âge inférieures à 65 ans) par rapport à une population de référence
(Rapport d’étude INRS, 2004). Cette diminution de la durée de vie est très importante et il convient de
déterminer quels sont les facteurs clés pouvant l’expliquer. Les études épidémiologiques montrent que
certaines maladies affectent particulièrement les égoutiers, comme les cancers du foie (+85%, liés ou
non à la consommation d’alcool), de l’œsophage ou du poumon (+47%). Tous confondus, les cancers
présentent un excès de 82% chez la population d’égoutiers. Il existe aussi une forte prévalence de
maladies non malignes telles que des maladies cardiaques ou vasculaires, digestives (ulcère,
pancréatites), respiratoires (asthme, bronchopneumopathie), endocrinienne ou génito-urinaire. Il est à
noter que la population d’égoutiers montre un fort taux de morts violentes (suicides), pouvant être lié
aux conditions de stress et d’insalubrité, et est souvent affectée par des toux grasses et des diarrhées. La
prévalence de toutes ces maladies chez les égoutiers est supérieure à celle observées chez la population
de référence. Ces maladies peuvent s’expliquer par les conditions de travail mais aussi par l’exposition
des travailleurs à divers facteurs comme des polluants chimiques, des toxines ou des agents infectieux
présents dans les eaux usées.
Les toux grasses peuvent être dues à la présence d’endotoxines bactériennes (les lipopolysaccharides
(LPS) présents à la surface des membranes des bactéries Gram négatif) pouvant être projetées dans l’air
sous forme d’aérosols lors des curages (nettoyage à haute pression des réseaux d’eaux usées). La relation
entre symptômes respiratoires et endotoxine a déjà pu être mise en évidence (Smit et al., 2008). Ces
endotoxines sont à l’origine du syndrome des égoutiers, un état pseudo-grippal (fièvre, troubles
gastriques, frissons …). La forte prévalence de diarrhées n’est quant à elle toujours pas expliquée, mais
elles peuvent être dues à la présence de composés chimiques, de toxines, ou de micro-organismes
(bactéries, parasites …). La diversité microbienne présente dans les réseaux d’eaux usées demeurent
mal connues mais des études commencent à s’y intéresser. Certaines montrent qu’il est possible de
retrouver des bactéries pathogènes stricts des genres Salmonella, Shigella ou Campylobacter (Mara and
Horan, 2003). Certains agents pathogènes opportunistes comme Pseudomonas aeruginosa ou
Aeromonas caviae (Wheater et al., 1980 ; Monfort and Baleux, 1990) sont égalment détectés. La
présence de toxine bactérienne ou mycotoxine peut conduire à des allergies et de l’asthme. Des virus


5
peuvent également être présents dans ces milieux, principalement de la famille des adénovirus et des
norovirus, (pharyngites, pneumonies, conjonctivites) et des virus parainfluenzae (bronchiolite,
trachéites), ou des virus de l’hépatite A pouvant être responsables d’infections chez les égoutiers
(Montuori et al., 2009). Les taux d’anticorps anti-adénovirus et anti-parainfluenzae virus sont plus
élevés chez les travailleurs en contact avec les eaux usées, suggérant un contact étroit avec ces virus
(Altemyer et al, 1990). La présence de parasites dans les eaux usées a aussi été montrée, comme Giardia
intestinalis, causant de fortes diarrhées (Wallis et al., 1996).
En plus des différents agents microbiens pouvant causer plusieurs types de pathologies, les réseaux
d’eaux usées contiennent une grande diversité de polluants chimiques. Les propriétés et les
concentrations de ces polluants varient en fonction de l’origine des eaux usées (hôpital, industrie, zone
résidentielle …). Des études montrent que les eaux usées sont fortement contaminées par des polluants
chimiques dont certains peuvent avoir un impact sur la santé humaine (Harrison et al., 2006). Ainsi, les
eaux usées peuvent contenir des éléments traces métalliques toxiques (arsenic, cadmium, chrome,
cuivre, nickel, plomb, mercure …) ou des polluants organiques (pesticides, PCB, dioxines …). Chez
l’homme, les métaux lourds se stockent au niveau des os, du foie, des reins et du cerveau. Ils peuvent
induire des dysfonctions rénales, hépatiques, respiratoires, et certains métaux lourds montrent des effets
cancérigènes, tout comme certains pesticides (J.O. et al., 2007). Les PCB, à court terme, vont entrainer
des effets cutanés, oculaires et pulmonaires. Une exposition chronique des effets neuro-
comportementaux ont déjà pu être observés. Une exposition aux dioxines conduira à long termes à une
dégradation du système immunitaire, du système nerveux, du systèmes endocriniens et reproducteur
(Schantz, 1996). Ces molécules sont également cancérigènes. Il est à noter que, même à de faibles doses,
l’espace confiné que représente les canalisations concentrent les polluants, augmentant ainsi leurs effets
toxiques pour l’égoutier. De plus, les effets des cocktails de polluants sur la santé humaine ne sont pas
connus et demeurent peu étudiés. Ces molécules chimiques peuvent être à l’origine du nombre important
de cancers observé chez les égoutiers. En plus de ces polluants chimiques provenant d’une utilisation
par l’homme, on peut retrouver des molécules naturelles cancérogènes, comme les aflatoxines (produite
par des champignons). De plus, bon nombre de canalisations du réseau de la ville de Lyon sont
constituées d’amiantes. Il existe également un risque lié à la présence d’amiante. En effet, certaines
canalisations contiennent cette molécule toxique. Les égoutiers, en effectuant leurs tâches (se déplacer
dans le noir, les curages …), mettent en suspension dans l’air de grande quantité d’amiante. Cette
molécule peut déclencher l’amiantose, une maladie respiratoire, mais peut aussi être cancérigène et
favoriser le cancer du poumon (Hogdson and Darnton, 2000).
En dehors des risques infectieux/chimiques, les agents peuvent souffrir de troubles musculo-
squelettiques (TMS), une maladie professionnelle qui regroupent plusieurs troubles affectant les
muscles, les tendons ou les nerfs (Carayon et al., 1999). C’est la maladie professionnelle la plus
répandue dans les pays développés, notamment en France. Elle se caractérise par des douleurs, raideurs,


6
maladresses et une diminution de la force. Ces troubles sont dus à la position prise par les agents dans
les réseaux d’eau usées (diamètres des réseaux visitables : 1 à plus de 1,50 mètres, les agents doivent
progresser le dos courbé), mais sont aussi dus à la manipulation d’objets lourd (pioches, regards d’égouts
…) et à la répétition d’un même geste. Les TMS représentent un quart des effets négatifs observés sur
la santé de l’égoutiers.
Il faut ajouter à ces différentes pathologies le risque lié à la présence du soufre dont le sulfure
d’hydrogène (H2S). Le H2S provient de la dégradation bactérienne de composés organiques soufrés ou
de sulfates (ou autres formes comme celles citées ci-dessus) retrouvées dans les réseaux (Kadota and
Ishida, 1972) mais peut aussi être produit par des activités industrielles (notamment les tanneries et les
industries de caoutchouc). Cette molécule représente un risque de par son effet toxique pour l’homme.
Ce gaz peut conduire, en fonction des concentrations, a des intoxications aigues (troubles respiratoires,
irritations oculaires, céphalées, vertiges, conjonctivites), à des pertes de connaissance (à partir de 500
ppm) et même à la mort (> 1000 ppm) (Guidotti, 2015). La formation d’H2S est problématique dans les
réseaux d’eaux usées, en effet, des poches de gaz peuvent se former et seront libérées lors du curage,
exposant ainsi les agents. De plus, ce gaz, au contact de l’air, va former de l’acide sulfurique très corrosif
qui va dégrader le béton et la ferraille (Parker, 1951). Selon le ministère de la sante, des expositions
répétées peuvent être à l’origine d’intoxications chroniques (bronchites, irritations cutanées) même si
les effets à long termes demeurent inconnus. Les recommandation (CNAM R 420) préconisent des
expositions à un maximum de 5 ppm pour des expositions longues (8 heures) et 10 ppm pour des
expositions courtes (15 minutes). Une analyse des substances autorisées dans le traitement de l’eau
montre que plusieurs molécules soufrées peuvent être présentes dans les réseaux (selon la circulaire
2000/166 du 28 mars 2000 de la Direction Générale de la Santé, France). Les flocculants autorisés sont
le sulfate d’aluminium, chlorure d’aluminium, aluminate de sodium, polyhydroxychlorure d’aluminium,
polyhydroxychlorosulfate d’aluminium, polyhydroxychlorosilicate d’aluminium, chlorosulfate de fer,
et chlorure ferrique (ayant également un effet bactéricide). Les adjuvants de flocculation sont l’acide
sulfurique, le silicate de sodium, et la silice activée. Les désinfectants autorisés sont : le bisulfite de
sodium, le sulfate ferreux, le sulfite de sodium, l’hydrogénosulfite de sodium, le dioxyde de soufre, le
dioxyde de chlore, le chlore, le chlorure de sodium, l’hypochlorite de sodium ou calcium, l’ozone, le
peroxyde d’oxygène, sulfate de cuivre, et le permanganate de sodium. Ces molécules se retrouvent donc
dans les eaux usée, et certaines peuvent se montrer toxique comme le sulfate d’aluminium pouvant
favoriser l’apparition de la maladie d’Alzheimer (Walton, 2014).
Concernant les égoutiers de la ville de Lyon, les services de gestion des eaux usées classent les accidents
en 4 catégories : les chutes/glissades (25% des accidents déclarés), les troubles musculeux squelettiques
(25% des accidents), les coups/chocs (35 % des accidents) et des accidents divers (15 % des accidents)
de natures chimiques ou infectieuses. Les molécules chimiques et les différents micro-organismes
pouvant être retrouvés dans les eaux usées de la ville de Lyon restent largement méconnus, mais des


7
diarrhées et toux ont déjà été répertoriées, tout comme à Paris (Rapport d’étude INRS, 2004). Les alertes
gaz H2S (pour des concentrations supérieures à 10 ppm) sur les chantiers d’exploitation des réseaux
d’assainissement sont suivies depuis fin 2012, suite à une intoxication grave ayant eu lieu en surface, le
28 juin 2012, lors d’un curage (sources : ESX Lyon). Ainsi, plus de 163 alertes ont pu être détectées,
dont 31 pour lesquelles les agents ont été exposés et incommodés. En tout, il est répertorié plus de 120
intoxications (allant du mal de tête au malaise) dues au H2S ou à d’autres gaz de nature inconnue. Ajouté
au risque lié au H2S, les canalisations constituées d’amiante ne sont pour la plupart non répertoriées, ne
permettant pas de prendre les précautions nécessaires lors d’interventions dans ces réseaux.
Des observations faites par l’équipe de recherche d’accueil pour ce stage M2 suggèrent qu’une enzyme,
la thiopurine-S-methyltransférase (bTPMT) pourrait avoir une activité de détoxication des substances
soufrées, dont le H2S, et expliquées la présence de certaines formes pathogènes de l’Homme dans les
réseaux. Le spectre des substrats soufrés de la TPMT reste cependant à préciser.
1.2. La bTPMT et ses biais de répartition
La Thiopurine Méthyle Transférase (TPMT) est une enzyme cytoplasmique initialement découverte
chez l’Homme (Rémy, 1963). Cette enzyme permet la détoxication de la 6-mercaptopurine (6-MP ou
Thiopurine) qui est un analogue des purines. L’enzyme permet une méthylation du groupement thiol de
la 6-MP par un transfert du groupement méthyl de la S-adénosyl méthionine (SAM). Les Thiopurines
sont utilisées pour traiter les leucémies lymphoblastiques, les maladies inflammatoires, les maladies
auto-immunes ou pour éviter les rejets de greffons (Kandiel, 2005). Les 6-Mercaptopurines (6-MP)
entrent en compétition avec les purines lors de la synthèse de l’ADN chez les cellules en cours de
multiplication, et bloquent son élongation (Skipper, 1954). Ces molécules ont été découvertes en 1951
par Gertrude Elion (lauréat du prix Nobel pour cette découverte) lors de ces travaux sur l’inhibition de
la croissance de Lactobacillus casei par des analogues des purines (Elion et al., 1951).
Les effets cytotoxiques et immunosuppresseurs des Thiopurines peuvent être fortement
dommageables pour l’organisme en absence d’expression de la TPMT (Lennard, 1992). La TPMT est
exprimée dans tous les organes du corps humain à l’exception du cerveau et des poumons (Ford and
Berg, 2010), et contribue à l’efficacité des traitements en modulant la toxicité de la 6-MP. En effet, Une
fois dans l’organisme, la 6-MP interagira directement avec deux enzymes, l’Hypoxanthine
Phosphoribosylguanine transférase (HGPRT) et la TPMT. L’HGPRT produira de la Thioinosine
monophosphate à partir de la 6-MP, la forme active (inhibiteur de la synthèse de l’ADN) du médicament,
alors que la TPMT produira de la 6-MMP (6-MéthylMercaptopurine), une forme non-toxique pouvant
être éliminée par l’organisme (Baker, 2003). Les niveaux d’activités HGPRT et TPMT conditionnent le
succès des traitements à la 6-MP, et la cytotoxicité chez le patient. Certaines mutations du gène codant
la TPMT peuvent conduire à des changements d’affinité pour la 6-MP, ces modifications augmentent la
toxicité de cette molécule et peuvent conduire au décès des patients (McLeod et al., 2000). Des


8
diagnostics PCR ont été développés pour identifier les mutations impactant le fonctionnement de la
TPMT humaine (hTPMT) (Oender et al., 2006).
La TPMT a émergé très tôt dans l’évolution des organismes. Elle est en effet retrouvée chez les
mammifères, les levures, et les bactéries (Krynetski and Evans, 2003). Son évolution est cohérente avec
celles de gènes marqueurs universels utilisés en phylogénie moléculaire dont ceux codant les ARN
ribosomiques (Favre-Bonte et al., 2005). La première observation d’une activité de méthylation des 6-
MP chez les bactéries a été effectuée par Remy (1967) mais la TPMT bactérienne (bTPMT) n’a été
découverte qu’en 1998 (Cournoyer et al., 1998). La bTPMT montre une distribution restreinte chez les
bactéries mais une occurrence chez plusieurs agents pathogènes du groupe des -protéobactéries comme
Legionella pneumophila, Pseudomonas aeruginosa, et Vibrio cholerae (Favre-Bonte et al., 2005). Chez
les bactéries, les substrats de la bTPMT demeurent cependant mal connus. Cette enzyme peut conférer
une résistance aux oxyanions à base de Sélénium (séléniate et sélénite) et Tellurium (tellurate et tellurite)
(Ranjard et al., 2002) mais d’autres substrats expliquent certainement sa conservation. Le Sélénium (Se)
est un métalloïde essentiel à la vie, il peut être un constituant de certains acides aminés, un cofacteur ou
une composante de plusieurs enzymes (Ranjard et al., 2003). Cependant, à forte dose, le Sélénium peut
conduire à des intoxications (Cui et al., 2017). Le sélénite (SeO32-) est toxique, aussi bien pour l’homme
ou l’animal (chez qui il peut causer perte de la vision, coliques, problèmes respiratoires, léthargies) que
chez les bactéries. En effet, de par ses propriétés oxydatives, le sélénite peut interagir avec les protéines
membranaires et provoquer un burst oxydatif de la cellule (Favre-Bonté et al., 2006). Le Tellurium (Te)
est également un métalloïde, cette molécule possède des propriétés chimiques proches du soufre. La
forme oxydée, le tellurite (TeO32-) est toxique pour les micro-organismes, et cela par plusieurs
mécanismes. Premièrement, à l’instar des formes toxiques du Sélénium, cette molécule a un fort pouvoir
oxydatif et peut provoquer un burst oxydatif de la cellule (Turner et al., 2001). Le Tellurium peut
également entrer en compétition avec le Sélénium et le Soufre (S) lors de la synthèse protéique, et donc
être incorporé dans les protéines, les rendant non fonctionnelles et induisant des dysfonctions cellulaires
(Prigent-Combaret et al., 2012). La bTPMT peut méthyler ces espèces toxiques en di-méthylSélénide
(DMSe) ou di-méthylDiSélénide (DMDSe) (Ranjard et al., 2003) et en di-méthylTelluride (DMTe) ou
di-méthyldiTelluride (DMDTe) (Cournoyer et al., 1998). Ces molécules volatiles sont non-toxiques.
Lors de ces analyses sur le Te/Se, une méthylation du Soufre par la bTPMT en di-méthylSulfur (DMS)
et di-méthyldiSulfur (DMDS) a également été observée (B. Cournoyer, Pers. Comm.).
L’affinité de la bTPMT pour les substrats soufrés pourrait donc être liée à un rôle « naturel » dans
la détoxication d’antimicrobiens contenant des atomes de soufre. Une possibilité est le sulfure
d’Hydrogène (H2S), un gaz toxique pour l’homme et les bactéries. L’H2S inhibe la cytochrome c
oxydase et donc la respiration cellulaire (Forte and Giuffrè, 2016). D’autres antimicrobiens pourraient
également être détoxiqués par la bTPMT comme certains antibiotiques, par exemple les sulfamides. En
effet, il a déjà été montré chez l’homme qu’une enzyme proche de la TPMT, la Thiol-S-


9
méthyltransférase, était capable de modifier le sulfure d’hydrogène (H2S) en méthanethiol (CH4S)
(Weisiger et al., 1980). Ces substrats de la TPMT pourraient expliquer certains biais de répartition
observés chez les bactéries dans les milieux pollués. Il est à noter que les « Gene Expression Omnibus
profiles » (GEOprofiles, www.ncbi.nlm.nih.gov/geoprofiles), des archives NCBI regroupant des
données publiées d’expression de gènes, suggèrent que le gène tpm (codant la bTPMT) serait induit en
condition de micro-aérophilie (Alvarez-Ortega and Harwood, 2007) et en présence de sulfates (Tralau
et al., 2007). Ces observations sont cohérentes avec les données d’activité de méthylation des
métalloïdes par la bTPMT. De plus, une relation entre l’expression du gène tpm et la formation de
biofilms (en présence de tobramycine) a été observée (Anderson et al., 2008), confortant l’hypothèse
que l’enzyme est exprimée lorsque les concentrations en oxygène sont faibles.
La première phase de ce travail a donc été d’étudier les biais de répartition des bactéries pouvant
exprimer la bTPMT dans des milieux fortement contaminés par des médicaments et pouvant également
conduire à une production de sulfure d’hydrogène (H2S), c’est à dire les réseaux unitaires. Les réseaux
unitaires sont chargés de collecter les eaux usées (domestiques ou hospitalières) et les eaux de pluies à
travers les mêmes canalisations. Les eaux collectées rejoignent ensuite une station d’épuration pour y
être traitées. Ces biais de répartition ont été étudiés par métabarcoding à partir de données de séquences
d’amplicons tpm qui ont été mis à disposition pour le projet. Ces amplicons tpm ont été produits à partir
d’ADN bactérien extraits de biofilms qui ont été exposés aux effluents hospitaliers ou domestiques de
réseaux d’eaux usées reliés à la station d’épuration de Bellecombe (Scientrier, Haute-Savoie, France).
Lors de la récupération des biofilms, des descripteurs du milieu ont été mesurés par l’INRA de Thonon
(Chonova et al., 2016) ainsi que les concentrations de plusieurs médicaments par Lenanowski et al.
(Pers. Comm., Université de Poitiers / Ecole nationale Supérieure d’Ingénieurs de Poitiers). Les relations
entre ces données et les espèces bTPMT des biofilms collectés ont été recherchées durant ce stage par
bio-informatique en s’inspirant de la trame d’analyses présentées par Bourgeois et al. (2017). Les OTU
tpm ont pu faire l’objet d’allocations taxonomiques. Ceci a permis d’identifier des espèces bTPMT
fortement dominantes dans les prélèvements ainsi que des espèces pathogènes comme Pseudomonas
aeruginosa et Aeromonas caviae. Une sélection d’espèces bTPMT par les effluents hospitaliers
(différentes des espèces de la file urbaine) a pu être mise en évidence. Les implications de la bTPMT
dans les biais de répartition observés restent à préciser, et ont été explorées dans la suite de ce travail.
La deuxième phase de ce stage a été de tester certaines hypothèses concernant les substrats de la
bTPMT et ses conditions d’expression. Pour ces travaux, une espèce modèle a été choisie suite à
l’analyse des données NGS des amplicons tpm obtenus de biofilms (phase 1). Les types tpm de P.
aeruginosa obtenus par NGS ont été comparés à une banque de données de types tpm d’isolats, et ainsi
permis d’observer l’occurrence de formes similaires à celles de génotypes cliniques de référence telles
que PA14 (Kukavica-Ibrulj et al., 2008), PAO1 (Stover et al., 2000), et une souche « hyper-virulente »
isolée chez des patients atteints de mucoviscidose, LESB58 (Kukavica-Ibrulj et al., 2008). Le génotype


10
PAO1 a été choisi pour les expérimentations en laboratoire dans un premier temps. Pour préciser les
résistances conférées par cette enzyme et autres propriétés chez cette souche, deux stratégies ont été
retenues : (1) construction d’une souche sur-productrice de la bTPMT et tests de croissance avec
différents anti-microbiens par la procédure omnilog combo (Kerangart et al., 2018) ; et (2) l’analyse
d’expression des transcrits tpm par qRT (quantitative reverse transcriptase) PCR en faisant varier les
conditions de croissance (taux d’oxygène, présence ou non de Tellurite). Les tests de croissance ont
permis d’observer un rôle de la bTPMT dans la résistance au Niaproof (un alkysulfate), à la colistine et
la tylosine (antibiotiques), au chlorure de Fer, à la Patuline (Mycotoxine) et au Bromure de Domiphène
chez P. aeruginosa.
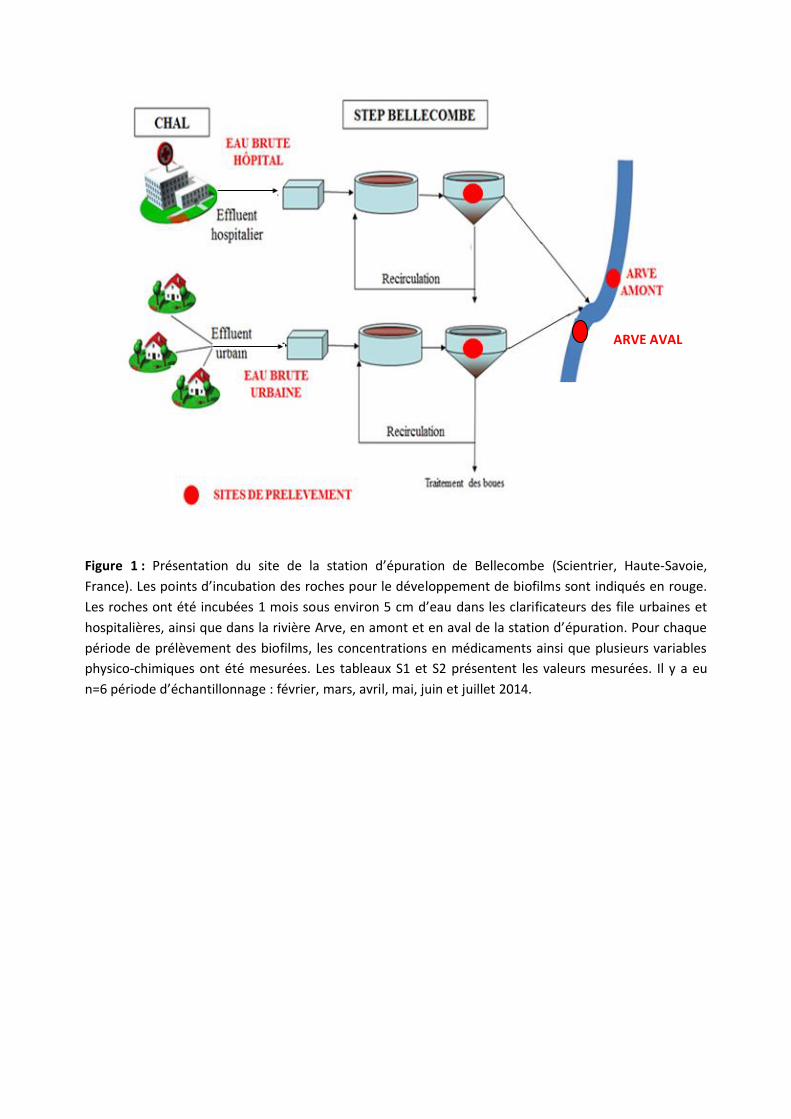
Figure 1 : Présentation du site de la station d’épuration de Bellecombe (Scientrier, Haute-Savoie,
France). Les points d’incubation des roches pour le développement de biofilms sont indiqués en rouge.
Les roches ont été incubées 1 mois sous environ 5 cm d’eau dans les clarificateurs des file urbaines et
hospitalières, ainsi que dans la rivière Arve, en amont et en aval de la station d’épuration. Pour chaque
période de prélèvement des biofilms, les concentrations en médicaments ainsi que plusieurs variables
physico-chimiques ont été mesurées. Les tableaux S1 et S2 présentent les valeurs mesurées. Il y a eu
n=6 période d’échantillonnage : février, mars, avril, mai, juin et juillet 2014.
ARVE AVAL

11
2. Matériels et méthodes 2. 1. Les bTPMT des effluents domestiques et hospitaliers 2.1.1. Présentation de la station d’épuration de référence L’étude implique des comparatifs entre les séquences tpm codant des bTPMT pouvant être retrouvées
dans les effluents hospitaliers et urbains de la station d’épuration de Bellecombe (WWTP) ainsi que
dans le milieu récepteur des eaux traitées (EU), c’est-à-dire la rivière Arve. Les points d’échantillonnage
de l’étude font partie du site Sipibel de la Zone Atelier bassin du Rhône (ZABR -
www.graie.org/zabr/index.htm) (Fig. 1). Ce site est localisé à Scientrier (Haute-Savoie, France), et a la
particularité de traiter séparément les eaux usées provenant de la filière domestique et de la filière
hospitalière. Cette station d’épuration reçoit les eaux usées du centre hospitalier Alpes Léman (CHAL)
ouvert depuis février 2012 (Chonova et al. 2018). Ces effluents hospitaliers arrivent à la station
d’épuration sans prétraitement (rapport Sipibel, 2014).
2.1.2. Echantillonnage et analyses chimiques
Cette étude des biais de répartition de séquences tpm est basée sur une analyse de biofilms se
développant sur roches de type amphibolite exposées (sur site) aux eaux usées ou de rivière. Ce choix a
été effectué pour permettre une co-accumulation de polluants chimiques et de micro-organismes, et ainsi
favoriser les phénomènes sélectifs pouvant conduire à une structuration de la diversité en relation avec
les concentrations en médicaments. Avant leur implantation sur site, les roches ont été brossées,
nettoyées à la javel 5% puis stérilisées à l’autoclave. Elles ont ensuite été disposées dans des nasses en
inox qui ont été immergées dans les clarificateurs des filières urbaine et hospitalière, ainsi que dans
l’Arve, en amont et en aval de la zone de rejet de la station d’épuration. Les points de prélèvements sont
décrits en Figure 1. Les roches ont été immergées durant 2 à 4 mois, en fonction de la vitesse de
formation des biofilms. Six campagnes d’échantillonnages ont été réalisées : février, mars, avril, mai,
juin et juillet 2014. Parallèlement, à chaque campagne d’échantillonnage, des paramètres physico-
chimiques et microbiologiques (Tab. S1) ont été mesurés, ainsi que certaines substances chimiques
comme des médicaments (Tab. S2). Quelques caractéristiques (Type d’utilisation) des molécules sont
décrites dans le tableau S6. Les concentrations en médicaments dans les biofilms ont été mesurées
comme décrit dans (Chonova et al., 2018) par Labanowski et al. (U. Poitiers), et les concentrations en
intégrons 1, 2 et 3 ont été mesurées selon (Marti et al., 2017) par Marjolet et al. (VetAgro Sup).
2.1.3. Extraction d’ADN génomique, amplification et séquençage du gène tpm
Les biofilms formés sur les roches ont été grattés à l’aide d’une spatule et d’une brosse à dent stériles et
à usage unique. L’ADN de ces biofilms a été extrait en utilisant le kit GenElute-LPA (Sigma-Alrich,
Saint-Louis, Missouri, Etats-Unis) en suivant les instructions du fournisseur. Les amplicons tpm produits
à partir de ces ADN ont été séquencés en utilisant la technique Miseq Illumina. tpm a été amplifié en
utilisant un mélange équimolaire des couples d’amorces ILMN-PTCF2/ ILMN-PTCR2 et ILMN-
PTCF2m/ ILMN-PTCR2m (Tab. S3) puis séquencé par Biofidal (Vaulx-en-Velin, France). Ces amorces
permettent de bien discriminer, entre autres, les espèces des genres Achromobacter, Aeromonas,


12
Dechloromonas, Nitrosomonas, Nitrospira, Pseudomonas, Xanthomonas, et Xanthobacter. Elles
donnent donc accès à la répartition d’espèces pathogènes de ces genres mais également d’espèces ayant
des propriétés métaboliques particulières comme la production de nitrate à partir de nitrite.
2.1.4. Tri des séquences et analyses de la diversité
Les séquences ont été traitées selon la méthode définie par J. Kozich (Kozich et al., 2013) en utilisant
le logiciel Mothur (Schloss et al., 2009). Les séquences ont été supprimées lorsqu’elles présentaient les
paramètres suivants : (1) plus courtes que 250 pb et plus longues que 270 bp, (2) peu d’identités avec
les séquences de la base de séquences de référence, (3) contenaient plus de huit homopolymères, (4)
contenaient des bases ambigües. Les séquences chimériques ont été supprimées en utilisant la
commande chimera.uchime (Edgar et al., 2011). Le nombre de séquences a été normalisé en réalisant
un ré-échantillonnage aléatoire. Les séquences d’ADN ont ensuite été regroupées en Unité
Taxonomique Opérationnelle (OTUs) lorsqu’elles présentaient 100% d’identité (Schloss and Westcott,
2011). Les OTUs ont été reliées à des groupes taxonomiques en utilisant la base de données
BD_TPM_Mar18_V1 (développée au laboratoire BPOE) lorsque la relation observée était soutenue par
une valeur P de boostrap > 80%. Cette base de données contient 917 séquences, 8 phylas, 15 classes, 36
ordres, 64 familles, 134 genres, 459 espèces et 80 sous-espèces.
Les analyses de la diversité ont été réalisées en suivant la trame proposée par (Marti et al., 2017). Les
calculs d’-diversité (Indice de Shannon, Evenness et Simpson) et de -diversité (matrice de Bray-
Curtis et UniFrac) ont été effectués en utilisant le logiciel Mothur, ainsi que les analyses de similarités
(test statistique Amova). Les différences observées sont considérées comme significatives lorsque la p-
value 0,05. A partir des affiliations des OTUs, les heatmaps ont été réalisées avec le logiciel Explicet.
Les ACP, les PCoA, les RDA et les Corrélogrammes ont été réalisés en utilisant le package Vegan du
logiciel R (https://cran.r-project.org/web/packages/vegan). La structure des communautés bactériennes
a été analysée en utilisant la matrice de distances UniFrac calculée à partir de la matrice OTUs générée
par Mothur. Les analyses de corrélations entre les communautés bactériennes présentes et les paramètres
mesurés à partir du milieu (physico-chimiques et concentrations en médicaments) ont été réalisées avec
le logiciel Cytoscape (http://www.cytoscape.org) en utilisant le test de corrélation de Spearman (1000
itérations) associé à une correction de Bonferroni (P-value = 0,05).
Les séquences de chaque OTU ainsi que leurs affiliations taxonomiques ont été extraites. Les séquences
des Pseudomonas aeruginosa ont été comparées à un alignement multiple de référence regroupant des
séquences « types » de l’espèce pour les grands sous-clades et complexes clonaux. Le logiciel ClustalX
a été utilisé pour la comparaison des séquences, puis un arbre phylogénétique illustrant les relations
observées a été produit avec le logiciel Mega6 en utilisant la méthode de Neighbor Joining.
2.2. Etude fonctionnelle de la bTPMT chez Pseudomonas aeruginosa
2.2.1. Liste des souches utilisées et conditions de culture
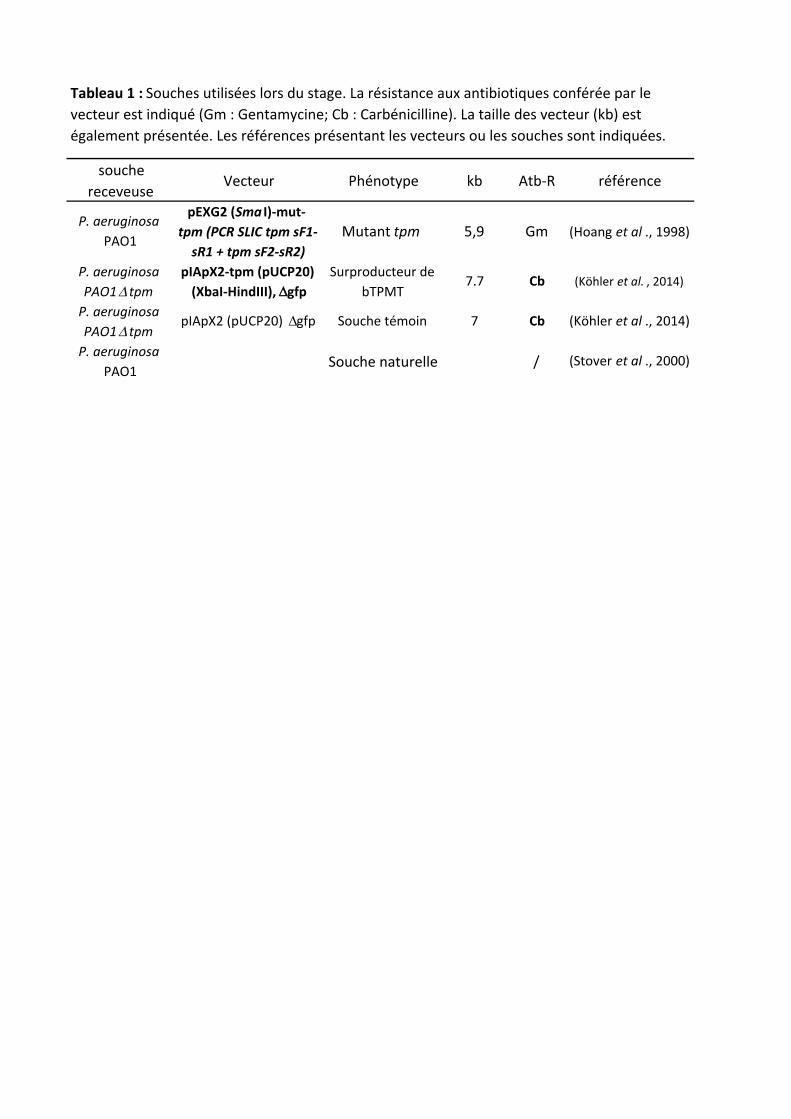
souchereceveuse
Vecteur Phénotype kb Atb-R référence
P.aeruginosaPAO1
pEXG2(Sma I)-mut-tpm(PCRSLICtpmsF1-sR1+tpmsF2-sR2)
Mutanttpm 5,9 Gm (Hoangetal .,1998)
P.aeruginosaPAO1D tpm
pIApX2-tpm(pUCP20)(XbaI-HindIII), Dgfp
SurproducteurdebTPMT
7.7 Cb (Köhleretal. ,2014)
P.aeruginosaPAO1D tpm
pIApX2(pUCP20)Dgfp Souchetémoin 7 Cb (Köhleretal .,2014)
P.aeruginosaPAO1
Souchenaturelle / (Stoveretal .,2000)
Tableau1:Souchesutiliséeslorsdustage.Larésistanceauxantibiotiquesconféréeparlevecteurestindiqué(Gm:Gentamycine;Cb:Carbénicilline).Latailledesvecteur(kb)estégalementprésentée.Lesréférencesprésentantlesvecteursoulessouchessontindiquées.

13
Toutes les souches d’E. coli et P. aeruginosa utilisées et leurs caractéristiques sont présentés dans le
Tableau 1. Toutes les souches ont été cultivées à 37°C sur gélose ou bouillon LB additionné ou non de
200 µg/mL de Carbénicilline. Les souches ont été conservées sur milieu gélosé à 4°C pendant quelques
semaines ou à -80°C dans leur milieu de culture additionné de 18% de glycérol. Les sur-producteurs
PAO1 de la bTPMT et la souche témoin ont été construits durant le stage.
Pour la construction d’une souche PAO1 sur-productrice de bTPMT, les cellules compétentes ont été
préparées à partir d’1mL d’une culture de nuit de P. aeruginosa PAO1tpm (Tab. 1 ; Fig. S5). Les
cellules ont été centrifugées à 13000 tpm pendant 30 secondes à température ambiante. Le culot a été
resuspendu dans 1mL de MgCl2 0,1 M froid. Cette solution a été centrifugée à 13000 tpm pendant 30
secondes puis le culot a été resuspendu dans 1 mL de TG (CaCl2 75 mM, MgCl2 6 mM et 15% de
glycérol) froid. Cette solution a été laissée sur glace pendant 10 minutes, puis centrifugée à 13000 tpm
pendant 30 secondes à température ambiante. Le culot a été resuspendu dans 200 µL de TG froid. 100
µL de cellules compétentes ont été mises en présence de 73 ou 219 ng de plasmide pIApX2 natif ou
contenant le gène tpm. Ce mélange a été laissé sur glace pendant 25 minutes et a ensuite été transféré
rapidement à 39 °C (choc thermique). Puis, 500 µL de LB ont été ajoutés, et les cellules ont été incubées
à 37°C pendant 1 heure sous agitation de 200 rpm. 100 µL et 500 µL de cette solution ont été étalés sur
gélose LB contenant 200 µg/mL de carbénécilline.
Pour valider les transformants, des cribles PCR ont été appliqués sur colonie. Une colonie a été mise
dans 200 µL d’H2O Ultra Pure (UP), les tubes ont été chauffés dans un bain marie sec Dry Bath System
(StarLab, Hambourg, Allemagne) à 100°C pendant 5 minutes, puis mis au congélateur à -20°C pendant
2 minutes afin de lyser les cellules. Le lysat a été centrifugé à 13000 tpm pendant 30 secondes à
température ambiante pour culoter les débris cellulaires. 2 µL des lysats bactériens ont été mis dans 23
µL de mix PCR contenant les amorces PTCF2 et PTCR2 (Tab. S3) amplifiant un fragment de 250 pb
du gène tpm. La taille des amplicons PCR a été vérifiée par électrophorèse (voltage = 100 V ; durée de
migration = 30 min) avec un gel à 2% d’agarose contenant 25 µg de bromure d’éthidium. Le gel a été
révélé aux UV à 302 nm.
Pour valider la surproduction de l’enzyme, P. aeruginosa PAO1, P. aeruginosa PAO1tpm-
pIApX2::tpm (sur-producteur) et P. aeruginosa PAO1tpm pIApX2 (témoin sans tpm) ont été incubé
en LB en absence ou en présence de 7,5 µg/mL de K2TeO3. La croissance a été arrêtée en phase
exponentielle (t=5h) avec une solution de RNAlater® (AmbionTM, Carlsbad, Etats-Unis) suivant le
protocole fourni par le fabriquant. L’ARN total de ces cultures a été extrait avec le kit RNeasy Mini
kit® (Qiagen, Hilden, Allemagne) en suivant les instructions du fournisseur. L’intégrité de l’ARN a été
vérifiée sur un gel à 1,5% d’agarose contenant 25 µg de BET. L’ARN a été quantifié en mesurant
l’absorbance à 260 nm au Nanodrop-One (Ozyme, Montigny-le-Bretonneux, France). L’ARN obtenu a
été traité avec de la Turbo DNase-freeTM Kit (Ambion, Carlsbad, Etats-Unis) en suivant les
recommandations du fabriquant pour un traitement rigoureux. 1,5 µL d’enzyme ont été utilisés. La
transcription inverse a été réalisée à l’aide du kit iScriptTM cDNA synthesis kit (Bio-rad, Marne-la-

14
Coquette, France) en suivant les instructions du fabricant. Une PCR sur le gène tpm a ensuite été réalisée
en utilisant les amorces PTCF2/PTCR2 (Tab. S3). Les amplicons PCR ont été vérifiés sur gel comme
indiqué précédemment.
2.2.2. Etude du rôle de la bTPMT dans la résistance de P. aeruginosa a divers antimicrobiens
2.2.2.1. Tellurite
Des suspensions (gouttelettes de 10 µL avec une concentration d’environ 105 CFU/mL) de P.
aeruginosa PAO1 (sauvage), P. aeruginosa PAO1tpm-pIApX2::tpm (surproducteur de bTPMT) ou P.
aeruginosa PAO1tpm pIApX2 (témoin) ont été déposées sur un milieu gélosé LB contenant 1, 5, 10,
20, 40, 80 ou 100 µg/mL de K2TeO3. Les boites ont été incubées à 37°C pendant 24 heures. Le nombre
de gouttelettes déposé montrant une croissance a été compté pour chaque construction testée.
2.2.2.2. Antibiotiques
Les antibiogrammes ont été réalisés en suivant les recommandations de la Société Française de
Microbiologie (SFM) pour l’analyse des résistances chez P. aeruginosa. Les antibiotiques (n=16)
utilisés (par la méthode du disque) et leurs concentrations sont décrits dans le Tableau 2. Les souches
P. aeruginosa PAO1 (sauvage), P. aeruginosa PAO1tpm-pIApX2::tpm (surproducteur de bTPMT) ou
P. aeruginosa PAO1tpm pIApX2 (témoin) ont été utilisées.
2.2.3.3. Autres substrats antimicrobiens
Les croissances des souches P. aeruginosa PAO1tpm-pIApX2::tpm (surproducteur de bTPMT) et P.
aeruginosa PAO1tpm pIApX2 (témoin) exposées à une large palette d’antimicrobiens ont été testées
par la technologie Omnilog microarray (OM) (Biolog, Bernay, France). Ces tests sont basés sur la
réduction par la respiration bactérienne du tétrazolium violet présent dans le milieu en formazan qui
donne une couleur violette. Cela permet de quantifier la croissance bactérienne au cours du temps
(Kérangart et al., 2018). Ces tests sont réalisés en plaque 96 puits contenant une large diversité
d’antimicrobiens (24 molécules par plaque, 240 molécules au total ; Tab. S4) et permettant une
exposition à quatre concentrations différentes (très faible, faible, moyenne, forte). Les inoculums et la
préparation des plaques ont été réalisés en suivant les recommandations du fournisseur. Les plaques ont
été incubées à 37°C pendant 48 heures, les aires sous la courbe de croissance ainsi que les valeurs
maximales ont été récupérées via les logiciels « Parametric » et « Kinetic » (Biolog, Bernay, France).
Les données ont été normalisée en calculant le ratio Aire mesurée/Aire maximal observée. Les ratios
obtenus sont indicatifs de la sensibilité de la souche face à l’antimicrobien présent dans le puit : (1) <0,5
= sensible ; (2) >0,5 = résistante.
3.Résultats
3.1. Les bTPMT des réseaux d’eaux usées domestiques et hospitalières
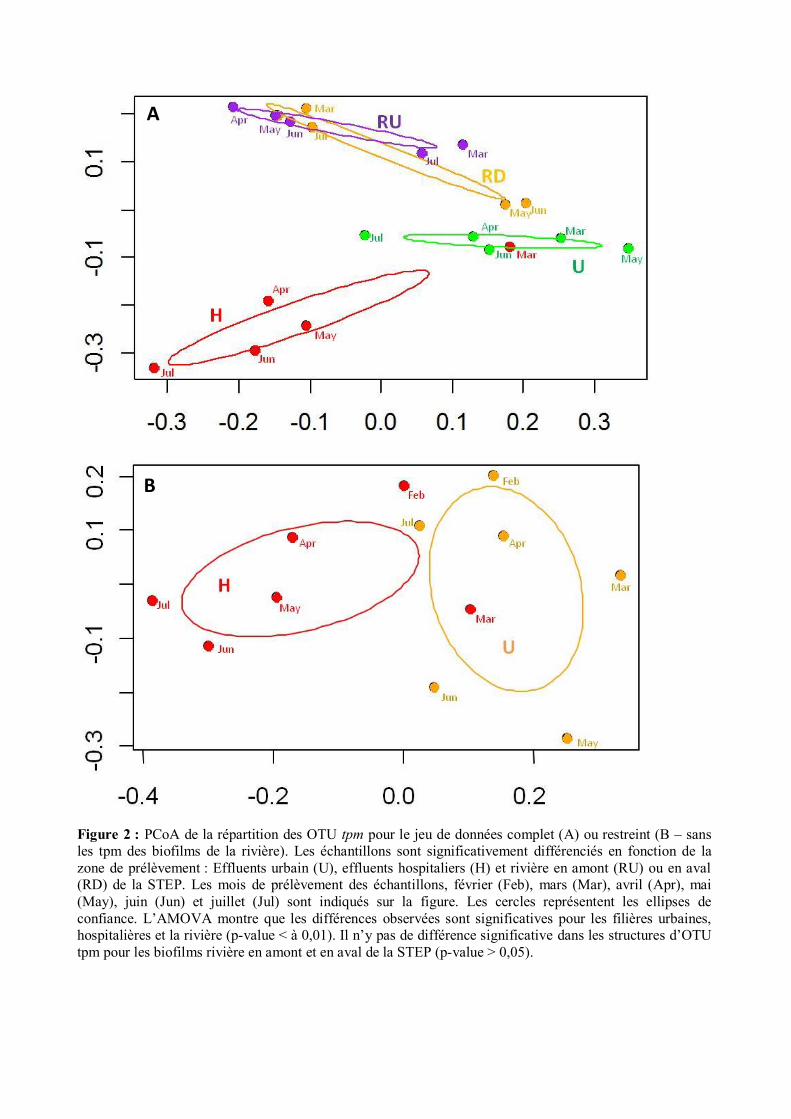
Figure 2 : PCoA de la répartition des OTU tpm pour le jeu de données complet (A) ou restreint (B – sans les tpm des biofilms de la rivière). Les échantillons sont significativement différenciés en fonction de la zone de prélèvement : Effluents urbain (U), effluents hospitaliers (H) et rivière en amont (RU) ou en aval (RD) de la STEP. Les mois de prélèvement des échantillons, février (Feb), mars (Mar), avril (Apr), mai (May), juin (Jun) et juillet (Jul) sont indiqués sur la figure. Les cercles représentent les ellipses de confiance. L’AMOVA montre que les différences observées sont significatives pour les filières urbaines, hospitalières et la rivière (p-value < à 0,01). Il n’y pas de différence significative dans les structures d’OTU tpm pour les biofilms rivière en amont et en aval de la STEP (p-value > 0,05).
A
B

15
3.1.1. Descriptions générales des données de séquençage tpm à partir de biofilms
Deux jeux de données de séquences d’amplicons tpm ont été utilisées pour inférer l’incidence des
paramètres de terrain et des polluants chimiques sur la structure génétique de la communauté bactérienne
codant le gène tpm : (1) un jeu de données restreint aux effluents urbains et hospitaliers, et (2) un jeu de
données incluant les séquences du point (1) mais complétées de données NGS tpm des biofilms de l’Arve
en amont ou en aval de la STEP. Entre 6211 et 30922 reads ont été obtenus par échantillon.
L’échantillons d’Avril de la filière hospitalière et de juillet de la filière urbaine contenaient les plus
faibles nombres de reads (6211 et 8308), impactant le nombre de séquences pouvant être utilisées pour
certaines analyses. Le Tableau 3 indique le nombre de séquences tpm obtenues par prélèvement pour la
totalité de l’étude, et ayant fait l’objet d’un ré-échantillonnage aléatoire. Ainsi, pour (1), un total de 6211
séquences tpm ont été retenues pour les analyses suivantes, et pour (2) un total de 8867 séquences tpm.
Des analyses d’-diversité ont été réalisées sur les jeux de données produits après ce ré-échantillonnage
aléatoire. Les courbes de raréfaction obtenues montrent que les échantillons prélevés dans les filières
urbaines ou hospitalières étaient plus faibles en nombre de reads que les échantillons prélevés dans la
rivière (Wilcoxon : p-value < 0,05) (Fig. S1). Par contre, le mois de prélèvement n’a pas impacté le
nombre de reads (Kruskal-Wallis : p-value > à 0,05). Les indices de richesse (Shannon, Simpson,
Evenness) sont indiqués dans le Tableau 3. Ils font ressortir une diversité plus élevée dans les biofilms
de la filière urbaine, par rapport à ceux de la filière hospitalière, ce qui est confirmé par un test de
Wilcoxon (p-value < 0,05). La richesse observée dans la rivière est similaire en amont et en aval du
point de rejet des eaux traitées (Wilcoxon : p-value > 0,05).
3.1.2. Structure génétique des communautés de bactéries codant le gène tpm dans les biofilms
Deux types d’analyse ont été effectuées pour comparer la diversité des séquences tpm des biofilms en
fonction de l’origine des prélèvements (filière hospitalière de la STEP ou urbaine ou rivière) et des
périodes d’échantillonnage : (a) le NMDS (non-metric multidimensional scaling) qui permet, par
analyse de rang, d’évaluer la proximité des structures en OTUs (sans considérer les nombres de reads)
entre paires de profils (et groupes) (Oksanen et al., 2018); cette méthode nécessite l’utilisation d’une
matrice de dissimilarités de type Bray-Curtis qui permet de comparer les abondances d’OTUs entre
paires d’échantillons (Beals, 1984) et (b) la PCoA (Principal Coordinates Analysis - metric
multidimensional scaling) impliquant une comparaison des distances entre patron d’OTUs prenant
également en considération le nombre de reads par OTU ; cette méthode nécessite l’utilisation d’une
matrice de distance et celle-ci a été calculée par approche UniFrac (Lozupone and Knight, 2005). La
comparaison des ordinations obtenues par approches NMDS (Fig. S2) et PCoA (Fig. 2) a montré de
fortes similitudes. Seules les représentations graphiques dérivées des analyses PCoA seront donc
décrites dans la suite du rapport. La matrice UniFrac (basée sur les pourcentages de dissimilarités entre
les échantillons) utilisée pour ces analyses est présentée dans le Tableau S5. Les tests AMOVA (analyse
de variances) ont montré une différenciation significative entre les structures des biofilms des files
hospitalières et urbaines de la STEP, ainsi qu’avec les ordinations inférées pour les biofilms de la rivière
(p-value = 0,01). La différenciation entre les structures d’OTUs des biofilms des files hospitalières et

16
urbaines de la STEP a été significative pour les deux jeux de données (avec ou sans les tpm de rivière)
(Fig. 2). Cependant, il n’y a pas eu différenciation des profils d’OTUs des biofilms entre l’amont et
l’aval du point de rejet des eaux traitées de la STEP (p value = 0,192). Cet ensemble « tpm biofilms
rivière » semble donc peu impacté par les bactéries tpm venant de la STEP.
3.1.3. Analyse des relations entre ordinations PCoA des profil d’OTU tpm des biofilms et leur composition chimique
L’effet des variables explicatives sur la structure génétique des profils tpm déduite par PCoA a été
étudié. Ces analyses ont été effectuées à partir des deux types de jeux de données (avec ou sans les
séquences tpm de rivière). Le jeu de données le plus complet en termes de séquences tpm présentait
moins de possibilités de confrontations avec les autres types de données (paramètres physico-chimiques,
concentrations en médicaments dans l’eau et dans les biofilms prélevés). Une première série de tests de
corrélation a été effectuée entre les concentrations en médicaments mesurées dans l’eau ou dans les
biofilms, pour préciser la redondance entre certaines variables (médicaments), pouvant avoir eu des
effets « confondants » sur la structuration des OTUs tpm dans les biofilms (Fig. S3). Ces analyses ont
permis d’observer une certaine complexité dans les relations entre les concentrations mesurées en
médicaments dans l’eau et dans les biofilms pour les molécules suivies dans l’étude. Cela a permis de
confirmer le caractère « intégrateur » des biofilms pour certaines des molécules analysées. Des analyses
plus exhaustives des flux de médicaments seraient nécessaires pour préciser la linéarité des relations
entre concentrations dans l’eau et les biofilms.
Pour cette raison, il a donc été décidé de focaliser la suite des analyses en utilisant uniquement les
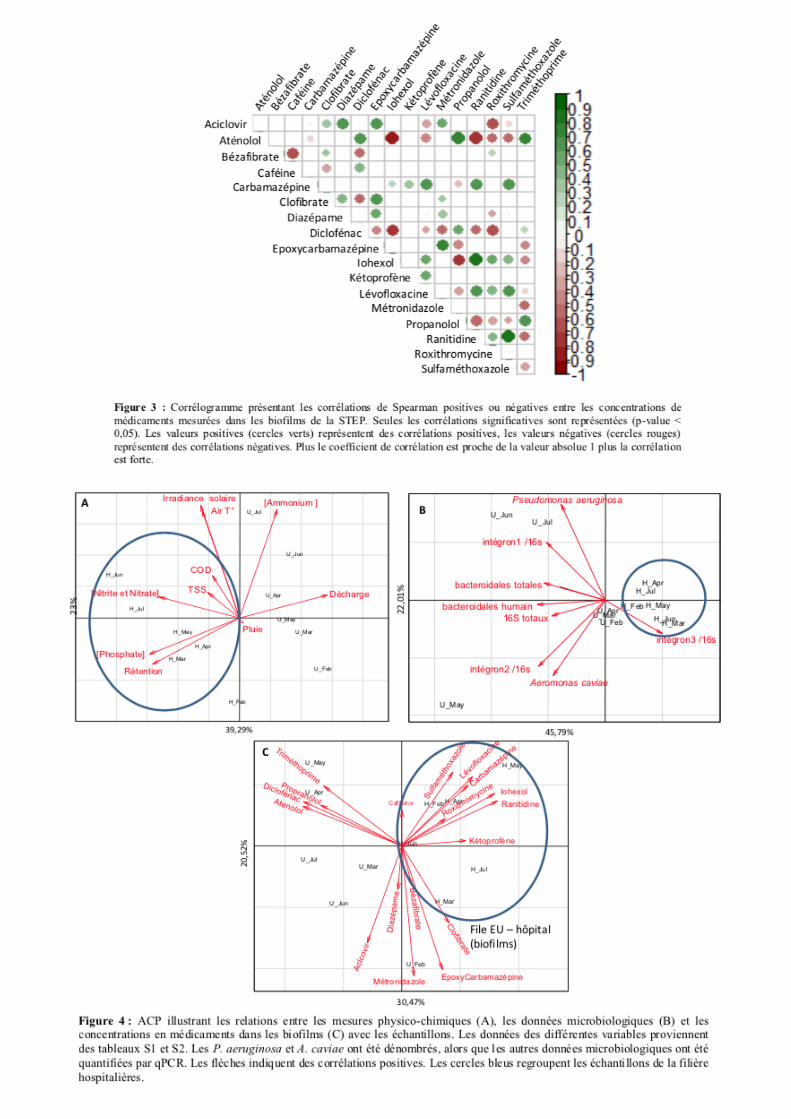
concentrations de médicaments mesurés dans les biofilms (Fig. 3). Des ACP ont permis d’observer des
co-occurrences entre médicaments dans les biofilms, et de confirmer que certains étaient principalement
(1) associés aux biofilms de la file hospitalière des EU : Sulfaméthoxazole, Lévofloxacine,
Carbamézépine, Roxithromycine, Ranitidine, Kétoprofène, Iohexol et Clofibrate, et (2) associés aux
biofilms de la file urbaine : Epoxycarbamazépine, Bézafibrate, Métronidazole, Aciclovir, Aténolol,
Diclofénac, Propanolol et Triméthoprime (Fig. 4). L’Iohexol est un produit de contraste triiodé
fortement lié, en termes d’utilisation, au milieu hospitalier. Le corrélogramme a permis de confirmer
une étroite relation entre cette molécule et les antibiotiques Lévofloxacine, Roxithromycine
Sulfaméthoxazole, et la Ranitidine. Une corrélation négative a été observée avec le Triméthoprime
(pourtant associé au Sulfaméthoxazole lors des traitements). En ce qui concerne la file urbaine, les
biofilms ont montré des corrélations positives entre les concentrations en Diclofénac, Caféine, Atenolol,
Propanolol et Diazépame. La ségrégation entre médicaments en fonction de la file d’épuration a donc
été confirmée (Fig. 3 & 4). Ceci a permis de conclure que les molécules suivantes présentaient une co-
occurrence significative dans les biofilms : (I) Aciclovir, Clofibrate, Diazépame, Epoxycarbamazépine,
Métronidazole, (II) Aténolol, Caféine, Diclofénac, Propanolol, Triméthoprime, (III) Iohexol,
Roxithromycine, Kétoprofène, Carbamazépine, Lévofloxacine, Ranitidine, et Sulfamethoxazole. Ces
co-occurrences seront à prendre en considération lors de l’interprétation des relations avec les structures
génétiques tpm des biofilms.
-0.4 -0.2 0.0 0.2
-0.3
-0.1
0.10.2
mds2[,1]
mds2[,2]
UH
Bezafibrate
Jul
Apr
Jun
May
Feb
Mar
May
Apr
Feb
Jun
Mar
Jul
-0.4 -0.2 0.0 0.2
-0.3
-0.1
0.10.2
mds2[,1]
mds2[,2]
UH
CaffeineB
Jul
Apr
Jun
May
Feb
Mar
May
Apr
Feb
Jun
Mar
Jul
-0.4 -0.2 0.0 0.2
-0.3
-0.1
0.10.2
mds2[,1]
mds2[,2]
UH
DiclofenacB
Jul
Apr
Jun
May
Feb
Mar
May
Apr
Feb
Jun
Mar
Jul
-0.4 -0.2 0.0 0.2
-0.3
-0.1
0.10.2
mds2[,1]
mds2[,2]
UHJul
Apr
Jun
May
Feb
Mar
May
Apr
Feb
Jun
Mar
Jul
-0.4 -0.2 0.0 0.2
-0.3
-0.1
0.10.2
mds2[,1]
mds2[,2]
UH
Jul
Apr
Jun
May
Feb
Mar
May
Apr
Feb
Jun
Mar
Jul
-0.4 -0.2 0.0 0.2
-0.3
-0.1
0.10.2
mds2[,1]
mds2[,2]
UH
LevofloxacineB
-0.4 -0.2 0.0 0.2
-0.3
-0.1
0.10.2
mds2[,1]
mds2[,2]
UH
Atenolol
-0.4 -0.2 0.0 0.2
-0.3
-0.1
0.10.2
mds2[,1]
mds2[,2]
UH
Propranolol
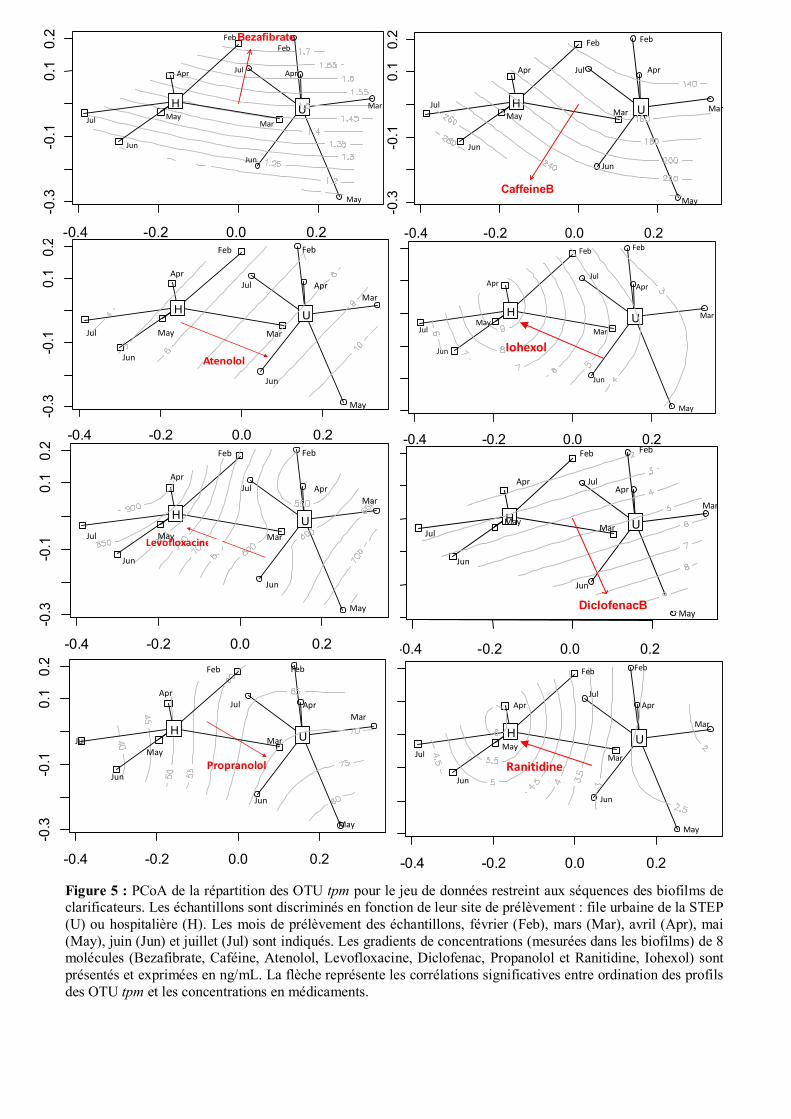
Figure 5 : PCoA de la répartition des OTU tpm pour le jeu de données restreint aux séquences des biofilms de clarificateurs. Les échantillons sont discriminés en fonction de leur site de prélèvement : file urbaine de la STEP (U) ou hospitalière (H). Les mois de prélèvement des échantillons, février (Feb), mars (Mar), avril (Apr), mai (May), juin (Jun) et juillet (Jul) sont indiqués. Les gradients de concentrations (mesurées dans les biofilms) de 8 molécules (Bezafibrate, Caféine, Atenolol, Levofloxacine, Diclofenac, Propanolol et Ranitidine, Iohexol) sont présentés et exprimées en ng/mL. La flèche représente les corrélations significatives entre ordination des profils des OTU tpm et les concentrations en médicaments.
Ranitidine
Iohexol
Jun
May
Mar
Feb
Apr Jul
Mar
Feb
Apr
May
Jun
Jul
Jun
May
Mar
Feb
Apr Jul
Feb
Apr
May
Jun
Jul Mar
Jun
May
Mar
Feb
Apr Jul
Feb
Apr
May
Jun
Jul Mar

17
Les relations entre concentrations de ces médicaments (mesurées dans les biofilms) et les ordinations
PCoA ont été explorées (Fig. 5). Des relations significatives ont été observée pour les molécules des
groupes II (Caféine, Atenolol, Diclofenac, Propanolol) et III (Iohexol, Levofloxacine, et Ranitidine).
Ces analyses montrent également de plus fortes concentrations en Carbone organique dissous (COD),
en matière solide en suspension (TSS), en Nitrites, Nitrates et Phosphate ainsi qu’une plus forte
occurrence de l’intégron 3 dans la filière hospitalière.
3.1.4. Allocation taxonomique des séquences tpm des biofilms
Les séquences tpm des biofilms ont été comparées en utilisant le package Mothur et un alignement
multiple regroupant les principales lignées tpm disponibles dans les banques de données. Ceci a permis
la classification d’environ 50% des séquences tpm de biofilms. La résolution, en termes de classification,
a été améliorée en réactualisant la banque de données via une recherche par blastN avec les principales
OTUs non-classées (n=100) du jeu de données. Ceci a permis de classer certaines OTUs dominantes au
sein du jeu de données dont l’Otu0001 (n=44049 reads), retrouvée dans les biofilms H_Apr-H_Jul-
H_Jun-H_May-U_Jun, qui a ainsi pu être reliée (74% de similarités – couverture de 96%) à un
endosymbionte non-isolé (sulfur-oxidizing symbionts of Riftia pachyptila - un ver tubicole géant) du
groupe des -protéobactéries. Cette inférence est une observation majeure lorsque mise en relation
avec les fonctions de détoxication de substances soufrées de la bTPMT. Pour illustrer les principales
inférences taxonomiques au niveau espèce, des représentations par « heat map » ont été réalisées (Fig.
6). Les séquences NGS des amplicons tpm des biofilms ont ainsi été classées principalement dans trois
genres : Herbaspirillum, Pseudomonas et Aeromonas. Les affiliations au niveau des espèces ont fait
ressortir Herbaspirillum aquaticum comme étant le taxon ayant le plus de séquences dans le jeu de
données (abondance relative de 12 à 81%). Cette espèce était davantage présente dans les effluents
hospitaliers et urbains que dans la rivière (test de Wilcoxon : p-value < 0,05). La présence de séquences
tpm de P. anguilliseptica était également récurrente (de 0,2 à 5%), mais leur nombre était plus abondant
dans la rivière (Wilcoxon : p-value < 0,01). Des différences dans la répartition des séquences tpm entre
les compartiments ont également été observées pour l’espèce candidatus Nitrospira defluvii avec une
occurrence élevée dans les biofilms des files urbaines et hospitalières mais une absence dans les biofilms
de la rivière (Wilcoxon : p-value < 0,001). Des tpm de type A. caviae ont été détectés dans les eaux de
la file hospitalière et dans la rivière, en aval de la STEP, comme P. aeruginosa mais qui était également
présent dans un échantillon de la file urbaine. Pour mettre en évidence les séquences tpm des taxons
pouvant être sélectionnées par les effluents hospitaliers, une figure présentant les ratios d’abondance
relative entre les biofilms des files d’eau usées hospitalière/abondance relative dans la filière urbaine en
fonction de l’abondance dans la filière hospitalière a été réalisée (Fig. 7). Cette analyse de répartition
des séquences tpm et taxons confirme une occurrence plus élevée de plusieurs espèces de Pseudomonas
dans les biofilms de la file hospitalière (Fig. 7). Les espèces observées peuvent être des formes
pathogènes opportunistes de l’homme comme Aeromonas sobria, Aeromonas caviae, Pseudomonas
putida et Pseudomonas flavescens ou de plantes comme Pseudomonas viridiflava. Les Pseudomonas

Figure 6 : Heatmaps illustrant les inférences taxonomiques déduites des séquences tpm obtenues des biofilms des files hospitalières et urbaines pour le jeu de données restreints (A – sans tpm des biofilms « rivière) ou avec les données concernant les biofilms de l’Arve en amont et en aval de la STEP (B). Les résultats sont présentés en abondance relative.
A
B
Abondance relative (%)
Abondance relative (%)

18
aeruginosa ressortent comme étant distribués de façon similaire dans les biofilms des files urbaines et
hospitalières. Ceci est en accord avec la heat map qui montrait une occurrence dans les deux files (Fig.
6). Une analyse phylogénétique a été réalisée pour préciser la proximité des types tpm de P. aeruginosa
du jeu de données NGS des biofilms avec les séquences décrites pour cette espèce dans les banques de
données. Cette analyse montre que deux des OTU présents dans les échantillons appartiennent aux sous-
clades PA14 (exoU+) et LESB58 (exoS+) (Fig. 8). Les OTU #1159 (22 reads), #2886 (5 reads), #7389
(2 reads) et #5321 (2 reads) des biofilms en aval de la zone de rejet de la STEP de la rivière ont montré
une proximité avec le clade PAO1 mais étaient différenciées. L’OTU #2886 (5 reads) était identique à
des tpm d’isolats cliniques et d’isolats de la ville (eaux urbaines).
3.1.5. Relation entre les abondances des séquences tpm et les concentrations en médicaments
Des corrélogrammes montrant les relations entre les OTUs tpm et les concentrations mesurées en
médicaments sont présentés en Figure 9. La présence de P. aeruginosa était peu corrélée aux
concentrations mesurées en médicament du groupe III (file hospitalière) (voir section 3.1.3.), et était
négativement corrélée aux molécules de la file urbaine. Une forte corrélation négative était observée
avec le triméthoprime malgré une résistance reconnue de cette espèce à cette molécule (Eliopoulos and
Huovinen, 2001). Les nombres de reads tpm de P. putida étaient corrélés à plusieurs concentrations en
médicaments de la file hospitalière, et étaient négativement corrélés à deux molécules (caféine et
diclofénac) liées à la file urbaine. Les nombres de reads tpm de P. flavescens étaient corrélés
positivement à plusieurs molécules liées à la file hospitalière, et ce type de tpm n’était pas lié à la
présence de Bézafibrate et Propanolol, des molécules de la file urbaine (Fig. 9). Ces résultats font
ressortir une cohérence entre répartition des médicaments et présence de certaines espèces de
Pseudomonas. Pour les espèces d’Aeromonas, des relations similaires ont été observées. La présence de
tpm d’A. caviae et A. sobria était plus fortement liée à la présence de molécules de la file hospitalière
(dont la Roxithromycine et la Lévofloxacine), que de la file urbaine (Fig. 9). Les nombres de reads tpm
d’A. allosaccharophila étaient plus fortement liés aux molécules de la file urbaine.
Les analyses faites avec le logiciel Cytoscape (Spearman –avec correction de Bonferonni) ont permis
d’observer des corrélations positives entre les dates de prélèvement des biofilms, et la température,
l’exposition solaire, ou la quantité pluie et les niveaux de décharge en eau traitée par la STEP. Des
corrélations négatives entre le temps de rétention des eaux et la quantité de pluie ont pu être observées
(non présentée).
3.2 . Rôle de la bTPMT dans la résistance aux antimicrobiens Les données de la section précédente ont fait ressortir plusieurs associations entre médicaments, file
d’épuration et présence de certaines bactéries arborant le gène tpm. L’importance de la bTPMT dans les
répartitions observées reste à préciser malgré une forte association avec une OTU ayant un
développement lié à la présence de molécules soufrées dont le sulfure d’Hydrogène. Pour explorer ces
relations, des manipulations génétiques ont été effectuées sur une souche modèle de l’espèce P.
aeruginosa ayant un gène tpm similaire à ceux retrouvés dans les eaux usées du site d’étude. Deux types

Abondance relative H
Abondance H / Abondance U

19
de souches PAO1 ont été produites : (a) une souche mutante délétée du gène tpm, et (b) une souche sur-
productrice de la bTPMT exprimant constitutivement le gène tpm à partir d’un plasmide.
Ces souches ont été utilisées dans un premier temps pour évaluer l’incidence du gène tpm dans la
résistance au tellurite. La souche délétée du gène tpm (tpm) (témoin) et la souche sur-productrice de
bTPMT (tpm::pIApX2-tpm) ont été exposées à différentes concentrations de tellurite. Aucune
différence de résistance à ce métalloïde n’a pu être observée entre ces deux souches. En effet elles
montraient le même profil de résistance, avec une Concentration Minimale Inhibitrice (CMI) égale à 40
µg/mL de tellurite. Cependant, la souche sauvage P. aeruginosa PAO1 a montré une CMI plus élevée
de 80 µg/mL de tellurite, pour 42% des tests effectués. Ceci indique une probable acclimatation
fonctionnelle au tellurite. Une forte odeur métallique a toutefois été détectée à partir des boîtes de la
souche surproductrice de bTPMT. Cette odeur n’était pas détectable chez les cultures de la souche
(tpm). Dans tous les cas, les bactéries produisaient des cristaux noirs indicatifs d’une réduction du
tellurite en cristaux de Te. Ces analyses ont indiqué que le sur-producteur était bien fonctionnel. Ceci a
été confirmé par qRT-PCR avec la détection d’ARNm tpm chez le sur-producteur.
Suite à cette validation, la résistance envers différents antibiotiques (Tab. 2) et autres antimicrobiens
(Tab. S4) a été étudiée afin de préciser certains substrats de la bTPMT. Premièrement, la souche délétée
du gène tpm, la souche sur-productrice de bTPMT et la souche sauvage ont été exposés à des disques de
différents antibiotiques sélectionnés selon les recommandations de la Société Française de
Microbiologie pour l’étude de l’antibio-résistance. Sur les 16 antibiotiques testés, seule une différence
a été observée pour la Ticarcilline, de la famille des pénicillines, pour laquelle le surproducteur de
bTPMT et mutant tpm arborant le pIApX2 natif étaient résistants (Tab. 4). Cela était liée à l’expression
du gène bla de pIApX2 (Fig. S4). Sinon, les souches étaient sensibles à tous les antibiotiques testés dont
la Lévofloxacine présente dans les biofilms de la section précédente.
Dans un second temps, cette étude a été complétée par analyses Omnilog combo de plusieurs
antimicrobiens dont des éléments traces métalliques (Tab. S4). Sur les 240 molécules testées, 64 ont
montré un effet négatif sur la croissance des souches mutées (témoin) ou surproductrices de bTPMT.
Des différences de résistance entre ces souches ont pu être observées pour 7 molécules. La souche
surproductrice a montré une résistance plus élevée au chlorure de Fer (FeCl3.6H2O; un floculant
bactéricide pour le traitement des eaux usées et potables), au Niaproof (Détergent à base de soufre / sels
de sodium), à la colistine et la tylosine (Antibiotiques) (Fig. 10.A). Par contre, cette souche sur-
productrice était plus sensible que le mutant à la Patuline (Mycotoxine), au Bromure de Domiphène
(Antiseptique) et au 2,2'-Bipyridine (Bipyridine) (Fig. 10.B).
4. Discussions/Conclusions Le rôle de la bTPMT reste à préciser malgré des relations évidentes avec la détoxication de substances
soufrées et autres molécules à base de Se et Te. De plus, l’analyse des biais de répartition du gène tpm
codant cette enzyme a révélé une prévalence élevée chez des -protéobactéries pathogènes souvent

10,90,80,70,60,50,40,30,20,10-1 -0,9 -0,8 -0,7 -0,6 -0,5 -0,4 -0,3 -0,2 -0,1
Figure 9 : Corrélogrammes présentant les corrélations de Spearman positives ou négatives entre les concentrations mesurées en médicaments dans les biofilms et les nombres de séquences tpm pour certaines espèces de Pseudomonas (A) ou d’Aeromonas (B). Seules les corrélations significatives sont représentées (p-value < 0,05). Les cercles verts représentent des corrélations positives, les cercles rouges représentent des corrélations négatives. Plus le coefficient de corrélation est proche de la valeur absolue 1 plus la corrélation est forte. Le cercle rouge indique les molécules retrouvées principalement dans les biofilms de la file hospitalière, et le jaune, les molécules plutôt associées à la file urbaine. Les couleurs classent les molécules dans les 3 différents groupes (vert : I ; violet : II ; orange : III).
MedicamentsBiofilms
AciclovirAténololBézafibrateCaféineCarbamazépineClofibrateDiazépameDiclofénacEpoxycarbamazépineIohexolKetoprofèneLévofloxacineMétronidazolePropanololRanitidineRoxithromycineSulfaméthoxazoleTriméthoprime
AciclovirAténololBézafibrateCaféineCarbamazépineClofibrateDiazépameDiclofénacEpoxycarbamazépineIohexolKétoprofèneLévofloxacineMétronidazolePropanololRanitidineRoxithromycineSulfaméthoxazoleTriméthoprime
MedicamentsBiofilms
AciclovirAténololBézafibrateCaféineCarbamazépineClofibrateDiazépameDiclofénacEpoxycarbamazépineIohexolKetoprofèneLévofloxacineMétronidazolePropanololRanitidineRoxithromycineSulfaméthoxazoleTriméthoprime
AciclovirAténololBézafibrateCaféineCarbamazépineClofibrateDiazépameDiclofénacEpoxycarbamazépineIohexolKétoprofèneLévofloxacineMétronidazolePropanololRanitidineRoxithromycineSulfaméthoxazoleTriméthoprime
AciclovirAténololBézafibrateCaféineCarbamazépineClofibrateDiazépameDiclofénacEpoxycarbamazépineIohexolKétoprofèneLévofloxacineMétronidazolePropanololRanitidineRoxithromycineSulfaméthoxazoleTriméthoprime
MedicamentsBiofilms
AciclovirAténololBézafibrateCaféineCarbamazépineClofibrateDiazépameDiclofénacEpoxycarbamazépineIohexolKetoprofèneLévofloxacineMétronidazolePropanololRanitidineRoxithromycineSulfaméthoxazoleTriméthoprime
Coefficient de corrélation

20
présentes dans les eaux usées (Matyar et al., 2010). Pour mieux apprécier l’importance de la bTPMT,
deux stratégies ont été explorées lors de ce stage : (4.1) une analyse de la diversité génétique de cette
famille en fonction de l’origine des biofilms récoltés à partir d’une station d’épuration et d’un cours
d’eau, et (4.2) des tests de croissance avec des antibiotiques, des biocides, des substances chlorées, et
des éléments traces métalliques.
4.1. Les bTPMT des réseaux d’eaux usées domestiques et hospitalières
Tout d’abord, ces études ont confirmé, via l’utilisation du gène tpm comme marqueur de diversité, une
richesse bactérienne moins importante dans les effluents hospitaliers qu’urbains, comme déjà observé
(Stalder et al., 2014). Cela laisse à penser que ces milieux pollués par des molécules médicamenteuses
sélectionnent certaines OTUs tpm ; donc certaines espèces. Les profils OTUs tpm des biofilms de la
rivière en amont et aval de la STEP n’étaient pas différenciés. L’effet dilution des effluents de STEP est
certainement un phénomène explicatif. Dans un cours d’eau, les médicaments et biocides sont fortement
dilués limitant ainsi leur rôle dans la sélection des bactéries présentes dans un biofilm. Par contre, les
OTUs tpm présentes dans les biofilms des effluents urbains étaient différentes de celles des biofilms de
la file hospitalières. Il semble que cette différenciation entre files serait liée au différentiel dans le
contenu en substances médicamenteuses. Différentes OTUs tpm auraient été sélectionnées en fonction
de leur résistance et de leur capacité de détoxification des cocktails de médicaments présents dans ces
deux files d’épuration. De plus, des synergies entre molécules peuvent exister (Acar et al., 1975).
Comme observé lors de ce stage, toutes les molécules ne sont pas retenues de la même manière par les
biofilms, et ceci peut conduire à des cocktails très différents entre file d’épuration (Davies, 2003). La
pénétration des molécules au sein des biofilms dépend de leurs propriétés mais aussi de celles de la
matrice du biofilm. Les relations entre OTUs et ces molécules peuvent donc expliquer les différences
de structure des communautés de bactéries bTPMT+ au sein des biofilms de cette étude. Toutefois, il
est à noter que les volumes d’eau de la file hospitalière étaient beaucoup plus faibles que ceux de la file
urbaine. Ceci a donc induit un temps de résidence des eaux en cours d’épuration plus long dans le
clarificateur de la file hospitalière, pouvant impacter le développement des biofilms mais également les
temps d’exposition aux polluants chimiques.
L’espèce la plus retrouvée par inférences taxonomiques via l’analyse des séquences tpm, H. aquaticum
a souvent été retrouvée dans les milieux hydriques (Ding, 2004). Dans cette étude, cette espèce était
davantage présente dans les effluents hospitaliers et urbains que dans la rivière. Cependant, elle est
connue pour être sensible aux antibiotiques présents dans le milieux de prélèvement, notamment la
Lévofloxacine, le Triméthoprime et le Sulfaméthoxazole (Regunath et al., 2015). Les génotypes tpm
observés dans la file hospitalière pourraient être reliés à des formes résistantes aux antibiotiques. Ces
formes pourraient avoir émergé grâce à des acquisitions de gènes de résistance aux antibiotiques (GRA)
ou à la sélection de mutations génétiques. En effet, les hôpitaux sont des sources de multi-résistances
aux antibiotiques (Norrby, 1991). Des résistances aux antibiotiques ont déjà été décrites dans les STEP
(Chagas et al., 2011) ainsi que la dissémination d’intégrons qui sont des supports génétiques explicatifs
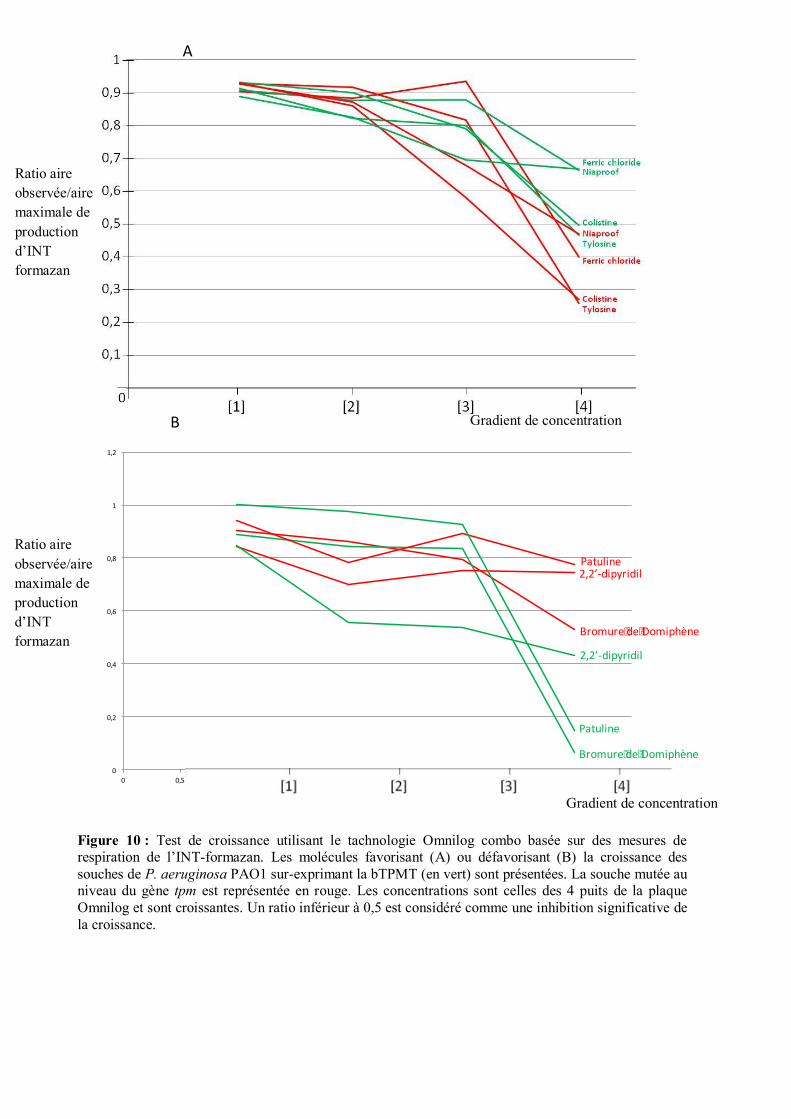
Figure 10 : Test de croissance utilisant le tachnologie Omnilog combo basée sur des mesures de respiration de l’INT-formazan. Les molécules favorisant (A) ou défavorisant (B) la croissance des souches de P. aeruginosa PAO1 sur-exprimant la bTPMT (en vert) sont présentées. La souche mutée au niveau du gène tpm est représentée en rouge. Les concentrations sont celles des 4 puits de la plaque Omnilog et sont croissantes. Un ratio inférieur à 0,5 est considéré comme une inhibition significative de la croissance.
A
B
0
0,2
0,4
0,6
0,8
1
1,2
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5
Patuline2,2’-dipyridil
Bromure�de�Domiphène
2,2’-dipyridil
Patuline
Bromure�de�Domiphène
Ratio aire observée/aire maximale de production d’INT formazan
Ratio aire observée/aire maximale de production d’INT formazan
Gradient de concentration
Gradient de concentration

21
d’une partie des disséminations rapides de ces résistances (Stalder et al., 2014). De plus, cette espèce a
déjà été associée à des bactériémies humaines (Regunath et al., 2015). La présence de tpm de candidatus
N. defluvii, peut par contre s’expliquer par un rôle fonctionnel majeur dans l’épuration via la
transformation du nitrite en nitrate durant le traitement des eaux usées (Lücker et al., 2010). Les tpm de
cette espèce ont été uniquement retrouvés dans les biofilms des files urbaines et hospitalières. Il est
également à noter que l’OTU tpm la plus abondante de la file hospitalière était clairement liée à un
groupe fonctionnel ayant un rôle dans la transformation des substances soufrées dont probablement les
sulfites et sulfure d’Hydrogène (H2S). Plusieurs espèces pathogènes opportunistes de l’homme arborant
le gène tpm semble avoir été favorisées par la file hospitalière, comme A. caviae, A. sobria, P. flavescens
ou P. putida. Cela peut être dû à une acquisition de GRA, ou aux plus fortes tolérances aux antibiotiques
présents dans les effluents hospitaliers. La présence de tpm d’A. caviae dans les biofilms de rivière, en
aval de la STEP, semble résulter de la non élimination de cette bactérie par le système de traitements de
la STEP. Cette bactérie a déjà été mise en évidence dans les eaux usées, et des défauts d’abattement ont
été reliés aux facteurs climatiques (Monfort and Baleux, 1990).
D’autres molécules liées au milieu hospitalier pourraient expliquer les biais de répartition des OTU tpm
observés avec la file urbaine. Par exemple, l’Iohexol (utilisé à l’hôpital) peut augmenter l’effet des
antibiotiques (Klessig et al., 2003). L’Iohexol est mal éliminé par les traitements conventionnels des
eaux usées (Papoutsakis et al., 2015). Cette molécule a donc été retrouvée dans les biofilms de la file
hospitalière. Il est à noter que l’Iohexol est un indicateur de la présence d’eaux usées d’origine
hospitalière. Pour le reste des polluants, les concentrations mesurées dans les deux types d’effluents
semblent dépendre de leur utilisation en amont, mais aussi de la capacité des biofilms à les concentrer.
4.2. Rôle de la bTPMT dans la résistance aux antimicrobiens Pour comprendre les phénomènes associés aux biais de répartition des bactéries TPMT+ observés dans
la section précédente, il sera essentiel d’approfondir l’étude des co-occurrences (ou déséquilibre de
liaison) entre tpm et des GRA comme dans le cas des espèces H. aquaticum, P. putida et A. caviae plus
fortement présentes dans les biofilms de la file hospitalière. Toutefois, il est également essentiel
d’explorer les résistances conférées directement par les bTPMT en fonction des cocktails chimiques
observés. Ces tests ont débuté par une validation d’une implication dans les phénomènes de résistance
au tellurite (te-r) en utilisant P. aeruginosa PAO1 comme souche modèle. Ces tests n’ont pas permis de
montrer de différence de sensibilité entre la souche surproductrice de bTPMT et le mutant délété du
gène tpm. Ceci est lié au fait que les bactéries te-r expriment un résistome complexe lorsqu’exposées au
tellurite incluant l’expression de plusieurs réductases (Cournoyer et al., 1998). Ces réductases réduisent
le tellurite en Te, une forme non-toxique. Ce Te s’accumule dans le périplasme, et donne une couleur
noire aux colonies (Prigent-Combaret et al., 2012). L’odeur d’ail métallique du DMTe a cependant été
détectée chez la souche surproductrice de bTPMT exposée au Tellurite. Cette odeur provient des gaz
émis lors de la méthylation du tellurite par la bTPMT sous la forme de DMTe et DMDTe non toxiques
(Cournoyer et al., 1998). L’antibiogramme de la souche P. aeruginosa surproductrice de bTPMT a, par


22
la suite, été réalisé et comparé avec celui de la souche avec un tpm muté. Aucune différence n’a été
observée. La bTPMT ne peut donc expliquer les biais de répartition des séquences tpm en fonction de
la présence des antibiotiques testés et potentiellement présents dans les biofilms de l’étude.
D’autres molécules seraient donc liées aux changements de communautés bTPMT+. Pour identifier ces
molécules, les phenotypic microarrays de l’Omnilog ont été utilisés. Ainsi, la souche surproductrice a
montré une résistance plus élevée au chlorure de Fer (FeCl3.6H2O ; un floculant bactéricide pour le
traitement des eaux usées et potables), au Niaproof (détergent à base de soufre), à la colistine et la
tylosine (antibiotiques). Par contre, cette souche sur-productrice était plus sensible que le mutant à la
Patuline (mycotoxine), le Bromure de Domiphène (antiseptique) et au 2,2'-dipyridil (bipyridine). La
plus forte résistance au Niaproof de la souche surproductrice peut s’expliquer par des transformations
de la molécule par la bTPMT. En effet, le Niaproof contient un groupement sulfate dont l’atome de
Soufre pourrait être méthylé par la bTPMT. Ce type de réaction a déjà été observé mais sans préciser le
rôle de la bTPMT (Kiene, 1988). Des résistances des bactéries du genre Pseudomonas avait été
rapportées (Timothy, 1999). Ces bactéries pourraient dégrader ces molécules et les utiliser comme
source de Soufre et d’énergie pour leur croissance (Kahnert and Kertesz, 2000). Cette résistance au
Niaproof conférée par la bTPMT conforte l’idée d’un rôle dans la détoxication de substances soufrées
retrouvées dans les réseaux d’eaux usées.
Une analyse des substances autorisées pour la potabilisation de l’eau montre que plusieurs molécules
soufrées peuvent être utilisées (selon la circulaire 2000/166 du 28 mars 2000 de la Direction Générale
de la Santé, France). Seul l’Aluminium sulfate et le chlorure ferrique ont ainsi pu être testés par
l’approche Omnilog. La souche PAO1 sur-productrice de la bTPMT n’a pas montré de différence de
croissance avec le sulfate d’Aluminium. Par contre, une croissance plus forte a été observée en présence
de chlorure ferrique. Cette molécule présente un caractère bactéricide en raison des atomes de chlore.
Les raisons d’une modification de la sensibilité de P. aeruginosa face à cette molécule via la bTPMT
sont actuellement difficiles à établir mais pourraient être liées aux interactions Cl et S pouvant conduire
à la production de chlorosulfate de fer. La bTPMT agirait donc potentiellement sur des sous-produits
d’interactions chimiques entre les composantes du milieu, et diminuerait la toxicité de ces sous-produits
par méthylation du soufre.
D’autres molécules ont montré une toxicité moindre sur la souche sur-productrice de bTPMT de P.
aeruginosa. La Colistine est un médicament de dernier recours face à Pseudomonas aeruginosa,
Klebsiella pneumoniae et Acinetobacter baumannii (Berlana et al., 2005). C’est un antibiotique
polypeptidique du groupe des polymyxines E (Vidaillac et al., 2012). Il a été rapporté que la Colistine
peut être sulfométhylées par des mécanismes qui sont toujours mal connus (Li, 2004). Cet antibiotique
a été utilisé pour traiter des infections à Pseudomonas aeruginosa chez des patiaents atteints de
mucoviscidose (Schneider-Futschik et al., 2018). La présence d’une enzyme pouvant modifier
l’efficacité de la Colistine chez ces bactéries est donc très préoccupante mais le changement de
sensibilité induit par la bTPMT semble limité. La Tylosine est un antibiotique bactériostatique qui vient


23
se fixer sur le ribosome, inhibant ainsi la traduction (Gaynor and Mankin, 2003). Cet antibiotique est du
groupe des macrolides. Le mode d’action de la bTPMT permettant de modifier la sensibilité face à cet
antibiotique est difficile à inférer avec les informations actuelles. Il est cependant possible que la bTPMT
puisse modifier certaines interactions avec les ribosomes en méthylant certaines composantes. Un
mécanisme de résistance impliquant des méthyltransférases a déjà été observé chez l’actinomycete
Streptomyces fradia (Liu and Douthwaite, 2002).
Les souches PAO1 sur-productrices de bTPMT étaient plus sensibles à la Patuline (mycotoxine), au
Bromure de Domiphène (antiseptique) et au 2,2'-dipyridil (bipyridine) que le mutant délété du gène tpm.
La Patuline, une mycotoxine, a une forte affinité pour les groupements thiols (Puel et al., 2010), comme
la bTPMT qui méthyle le groupement thiol de la 6-MP. Il faut noter que la Patuline perd son effet
bactéricide lorsqu’elle interagit avec la cystéine, un acide aminé qui possède un groupement Thiol
(Lindroth and von Wright, 1990). Si la bTPMT méthyle le groupement Thiol de la cystéine, il ne pourrait
donc plus interagir avec la Patuline qui conserverait alors ses propriétés toxiques. Cela pourrait expliquer
le fait que la souche surproductrice de bTPMT résiste moins à la Patuline. Pour le Bromure de
Domiphène et la 2,-2’dipyridil, des modifications de leurs modes d’action via la bTPMT n’ont pu être
inférées en fonction des éléments actuellement disponibles dans la littérature.
Pour poursuive ces études, des analyses complémentaires seront nécessaires pour valider les
changements de résistance observés. Plusieurs pistes très intéressantes ont été identifiées dont des
transformations possibles via la bTPMT de désinfectants utilisés dans les filières de traitements des eaux
usées, et un rôle de la bTPMT dans la résistance aux Colistines et Tylosines. Des tests de résistances
(avec détermination des Concentrations Minimales Inhibitrices) par méthodes de cultures classiques
seront nécessaires pour valider ces résultats. Par la suite, il serait intéressant de rechercher les molécules
identifiées dans la 2e partie de ce projet au sein du site d’étude (STEP de Bellecombe).
Il sera ensuite nécessaire d’étudier les conditions d’expression du gène tpm chez ces bactéries. Ces
études pourraient débuter par des analyses en RT qPCR avec la souche P. aeruginosa PAO1 naturelle
poussant en conditions de micro-aérophilie (2% d’oxygène) et en présence de Tellurite, mais à des
concentrations assez faible (7,5 µg/mL) pour éviter la dégradation des ARNm. En effet à 40 µg/mL, les
propriétés oxydatives de ce métalloïde dégradent les transcrits (Prigent-Combaret et al., 2012). Des
analyses d’expression de tpm en présence d’autres substrats comme la Colistine seront également à
effectuer.


24
Bibliographie Acar J.F., Sabath L.D., and Ruch P.A. (1975). Antagonism of the antibacterial action of some penicillins by other penicillins and cephalosporins. J. Clin. Invest. 55, 446–453.
Alvarez-Ortega C., and Harwood C.S. (2007). Responses of Pseudomonas aeruginosa to low oxygen indicate that growth in the cystic fibrosis lung is by aerobic respiration. Mol. Microbiol. 65, 153–165.
Anderson G.G., Moreau-Marquis S., Stanton B.A., and O’Toole G.A. (2008). In Vitro Analysis of Tobramycin-Treated Pseudomonas aeruginosa Biofilms on Cystic Fibrosis-Derived Airway Epithelial Cells. Infect. Immun. 76, 1423–1433.
Baker D.E. (2003). Pharmacogenomics of azathioprine and 6-mercaptopurine in gastroenterologic therapy. Rev. Gastroenterol. Disord. 3, 150–157.
Beals E.W. (1984). Bray-Curtis Ordination: An Effective Strategy for Analysis of Multivariate Ecological Data. In Advances in Ecological Research, (Elsevier), pp. 1–55.
Berlana D., Llop J.M., Fort E., Badia M.B., and Jódar R. (2005). Use of colistin in the treatment of multiple-drug-resistant gram-negative infections. Am. J. Health-Syst. Pharm. AJHP Off. J. Am. Soc. Health-Syst. Pharm. 62, 39–47.
Chagas T.P.G., Seki L.M., Cury J.C., Oliveira J.A.L., Dávila A.M.R., Silva D.M., and Asensi M.D. (2011). Multiresistance, beta-lactamase-encoding genes and bacterial diversity in hospital wastewater in Rio de Janeiro, Brazil: Resistant bacteria in hospital sewage. J. Appl. Microbiol. 111, 572–581.
Chonova T., Keck F., Labanowski J., Montuelle, B., Rimet F., and Bouchez A. (2016). Separate treatment of hospital and urban wastewaters: A real scale comparison of effluents and their effect on microbial communities. Sci. Total Environ. 542, 965–975.
Chonova T., Labanowski J., Cournoyer B., Chardon C. et., al (2018). River biofilm community changes related to pharmaceutical loads emitted by a wastewater treatment plant. Environ. Sci. Pollut. Res. 25, 9254–9264.
Cournoyer B., Watanabe S., and Vivian A. (1998). A tellurite-resistance genetic determinant from phytopathogenic pseudomonas encodes a thiopurine methyltransferase: evidence of a widely-conserved family of methyltransferases. Biochim. Biophys. Acta 1397, 161–168.
Cui Z., Huang J., Peng Q., Yu D., Wang S., and Liang D. (2017). Risk assessment for human health in a seleniferous area, Shuang’an, China. Environ. Sci. Pollut. Res. 24, 17701–17710.
Davies D. (2003). Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2, 114–122.
Ding L. (2004). Proposals of Curvibacter gracilis gen. nov., sp. nov. and Herbaspirillum putei sp. nov. for bacterial strains isolated from well water and reclassification of Pseudomonas huttiensis, Pseudomonas lanceolata, Aquaspirillum delicatum and Aquaspirillum autotrophicum as Herbaspirillum huttiense comb. nov., Curvibacter lanceolatus comb. nov., Curvibacter delicatus comb. nov. and Herbaspirillum autotrophicum comb. nov. Int. J. Syst. Evol. Microbiol. 54, 2223–2230.
Edgar R.C., Haas B.J., Clemente J.C., Quince C., and Knight R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200.
Elion G.B., Hitchings G.H., and Vanderwerff H. (1951). Antagonists of nucleic acid derivatives. VI. Purines. J. Biol. Chem. 192, 505–518.
Eliopoulos G.M., and Huovinen P. (2001). Resistance to Trimethoprim-Sulfamethoxazole. Clin. Infect. Dis. 32, 1608–1614.
Favre-Bonte S., Ranjard L., Colinon C., Prigent-Combaret C., Nazaret S., and Cournoyer B. (2005). Freshwater selenium-methylating bacterial thiopurine methyltransferases: diversity and molecular phylogeny. Environ. Microbiol. 7, 153–164.
Favre-Bonté S., Ranjard L., Champier L., Cournoyer B., and Nazaret S. (2006). Distribution and genetic diversity of bacterial thiopurine methyltransferases in soils emitting dimethyl selenide. Biochimie 88, 1573–1581.
Ford L.T., and Berg J.D. (2010). Thiopurine S-methyltransferase (TPMT) assessment prior to starting thiopurine drug treatment; a pharmacogenomic test whose time has come. J. Clin. Pathol. 63, 288–295.
Forte E., and Giuffrè A. (2016). How bacteria breathe in hydrogen sulfide-rich environments.
Gaynor M., and Mankin A. (2003). Macrolide Antibiotics: Binding Site, Mechanism of Action, Resistance. Curr. Top. Med. Chem. 3, 949–960.
Kandiel A. (2005). Increased risk of lymphoma among inflammatory bowel disease patients treated with azathioprine and 6-mercaptopurine. Gut 54, 1121–1125.
Kerangart S., Cournoyer B., and Loukiadis E. (2018). C-source metabolic profilings of foodborne Shiga-toxin producing E. coli match serogroup differentiations and highlight functional adaptations. Int. J. Food Microbiol. 266, 324–336.
Kiene R. (1988). Dimethyl sulfide metabolism in salt marsh sediments. FEMS Microbiol. Lett. 53, 71–78.
Klessig H.T., Showsh S.A., and Sekorski A. (2003). The Use of Intradiscal Antibiotics for Discography: an in vitro Study of Gentamicin, Cefazolin, and Clindamycin: Spine 28, 1735–1738.
Kozich J.J., Westcott S.L., Baxter N.T., Highlander S.K., and Schloss P.D. (2013). Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 79, 5112–5120.
Krynetski E., and Evans W.E. (2003). Drug methylation in cancer therapy: lessons from the TPMT polymorphism. Oncogene 22, 7403–7413.
Kukavica-Ibrulj I., Bragonzi A., Paroni M., Winstanley C., Sanschagrin F., O’Toole G.A., and Levesque R.C. (2008). In Vivo Growth of Pseudomonas aeruginosa Strains PAO1 and PA14 and the Hypervirulent Strain LESB58 in a Rat Model of Chronic Lung Infection. J. Bacteriol. 190, 2804–2813.
Lennard L. (1992). The clinical pharmacology of 6-mercaptopurine. Eur. J. Clin. Pharmacol. 43, 329–339.
Li J. (2004). Pharmacokinetics of colistin methanesulphonate and colistin in rats following an intravenous dose of colistin methanesulphonate. J. Antimicrob. Chemother. 53, 837–840.
Lindroth S., and von Wright A. (1990). Detoxification of patulin by adduct formation with cysteine. J. Environ. Pathol. Toxicol. Oncol. Off. Organ Int. Soc. Environ. Toxicol. Cancer 10, 254–259.


25
Liu M., and Douthwaite S. (2002). Resistance to the macrolide antibiotic tylosin is conferred by single methylations at 23S rRNA nucleotides G748 and A2058 acting in synergy. Proc. Natl. Acad. Sci. 99, 14658–14663.
Lozupone C., and Knight R. (2005). UniFrac: a New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 71, 8228–8235.
Lücker S., Wagner M., Maixner F., Pelletier E., Koch H et al. (2010). A Nitrospira metagenome illuminates the physiology and evolution of globally important nitrite-oxidizing bacteria. Proc. Natl. Acad. Sci. U. S. A. 107, 13479–13484.
Marti R., Bécouze-Lareure C., Ribun S., Marjolet L., Bernardin Souibgui C., Aubin J.-B., Lipeme Kouyi G., Wiest L., Blaha D., and Cournoyer B. (2017). Bacteriome genetic structures of urban deposits are indicative of their origin and impacted by chemical pollutants. Sci. Rep. 7.
Matyar F., Akkan T., Uçak Y., and Eraslan B. (2010). Aeromonas and Pseudomonas: antibiotic and heavy metal resistance species from Iskenderun Bay, Turkey (northeast Mediterranean Sea). Environ. Monit. Assess. 167, 309–320.
McLeod H.L., Krynetski E.Y., Relling M.V., and Evans W.E. (2000). Genetic polymorphism of thiopurine methyltransferase and its clinical relevance for childhood acute lymphoblastic leukemia. Leukemia 14, 567–572.
Monfort P., and Baleux B. (1990). Dynamics of Aeromonas hydrophila, Aeromonas sobria, and Aeromonas caviae in a sewage treatment pond. Appl. Environ. Microbiol. 56, 1999–2006.
Norrby S.R. (1991). Patterns of Multi-Resistance and Their Clinical Significance. Scand. J. Infect. Dis. 23, 1–70.
Oender K., Lanschuetzer C., Laimer M., Klausegger et al (2006). Introducing a fast and simple PCR-RFLP analysis for the detection of mutant thiopurine S-methyltransferase alleles TPMT*3A and TPMT*3C. J. Eur. Acad. Dermatol. Venereol. 20, 396–400.
Oksanen J., Blanchet F.G., Friendly M., Kindt R., Legendre P. et al. (2018). vegan: Community Ecology Package. Ordination methods, diversity analysis and other functions for community and vegetation ecologists. Version 2.5-1. URL https://CRAN.R-project.org/package=vegan.
Papoutsakis S., Afshari Z., Malato S., and Pulgarin C. (2015). Elimination of the iodinated contrast agent iohexol in water, wastewater and urine matrices by application of photo-Fenton and ultrasound advanced oxidation processes. J. Environ. Chem. Eng. 3, 2002–2009.
Prigent-Combaret C., Sanguin H., Champier L., Bertrand C. et., al (2012). The bacterial thiopurine methyltransferase tellurite resistance process is highly dependent upon aggregation properties and oxidative stress response: The TPMT Te R process. Environ. Microbiol. 14, 2645–2660.
Puel O., Galtier P., and Oswald I.P. (2010). Biosynthesis and toxicological effects of patulin. Toxins 2, 613–631.
Ranjard L., Prigent-Combaret C., Nazaret S., and Cournoyer B. (2002). Methylation of Inorganic and Organic Selenium by the Bacterial Thiopurine Methyltransferase. J. Bacteriol. 184, 3146–3149.
Ranjard L., Nazaret S., and Cournoyer B. (2003). Freshwater bacteria can methylate selenium through the thiopurine methyltransferase pathway. Appl. Environ. Microbiol. 69, 3784–3790.
Regunath H., Kimball J., Smith L.P., and Salzer W. (2015). Severe Community-Acquired Pneumonia with Bacteremia Caused by Herbaspirillum aquaticum or Herbaspirillum huttiense in an Immune-Competent Adult. J. Clin. Microbiol. 53, 3086–3088.
Rémy C.N. (1963). Metabolism of Thiopyrimidines and Thiopurines.
Remy C.N. (1967). Ribonucleotides and ribonucleosides as methyl acceptors for S-adenosylmethionine: (Amino- and thio-purine methyltransferases. Biochim. Biophys. Acta BBA - Nucleic Acids Protein Synth. 138, 258–275.
Schloss P.D., and Westcott S.L. (2011). Assessing and Improving Methods Used in Operational Taxonomic Unit-Based Approaches for 16S rRNA Gene Sequence Analysis. Appl. Environ. Microbiol. 77, 3219–3226.
Schloss P.D., Westcott S.L., Ryabin T., Hall J.R., Hartmann M., Hollister E.B. et al. (2009). Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 75, 7537–7541.
Schneider-Futschik E.K., Paulin O.K.A., Hoyer D., Roberts K.D. et., al (2018). Sputum Active Polymyxin Lipopeptides: Activity against Cystic Fibrosis Pseudomonas aeruginosa Isolates and Their Interactions with Sputum Biomolecules. ACS Infect. Dis. 4, 646–655.
Skipper H.E. (1954). Effects of 6-mercaptopurine on experimental tumors. Ann. N. Y. Acad. Sci. 60, 267–272.
Stalder T., Barraud O., Jové T., Casellas M., Gaschet M., Dagot C., and Ploy M.-C. (2014). Quantitative and qualitative impact of hospital effluent on dissemination of the integron pool. ISME J. 8, 768–777.
Stover C.K., Pham X.Q., Erwin A.L., Mizoguchi S.D., Warrener P., Hickey M.J. et al. (2000). Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 406, 959–964.
Timothy J. (1999). Compositions containing a high percent saturation concentration of antibacterial agent.
Tralau T., Vuilleumier S., Thibault C., Campbell B.J., Hart C.A., and Kertesz M.A. (2007). Transcriptomic Analysis of the Sulfate Starvation Response of Pseudomonas aeruginosa. J. Bacteriol. 189, 6743–6750.
Turner R.J., Aharonowitz Y., Weiner J.H., and Taylor D.E. (2001). Glutathione is a target in tellurite toxicity and is protected by tellurite resistance determinants in Escherichia coli. Can. J. Microbiol. 47, 33–40.
Vidaillac C., Benichou L., and Duval R.E. (2012). In Vitro Synergy of Colistin Combinations against Colistin-Resistant Acinetobacter baumannii, Pseudomonas aeruginosa, and Klebsiella pneumoniae Isolates. Antimicrob. Agents Chemother. 56, 4856–4861.
Weisiger R.A., Pinkus L.M., and Jakoby W.B. (1980). Thiol S-methyltransferase: suggested role in detoxication of intestinal hydrogen sulfide. Biochem. Pharmacol. 29, 2885–2887.
Effluents hospitaliers et stations d’épuration urbaines : caractérisation, risques et traitabilité-Presentation et premiers resultats. Rapport sipibel.
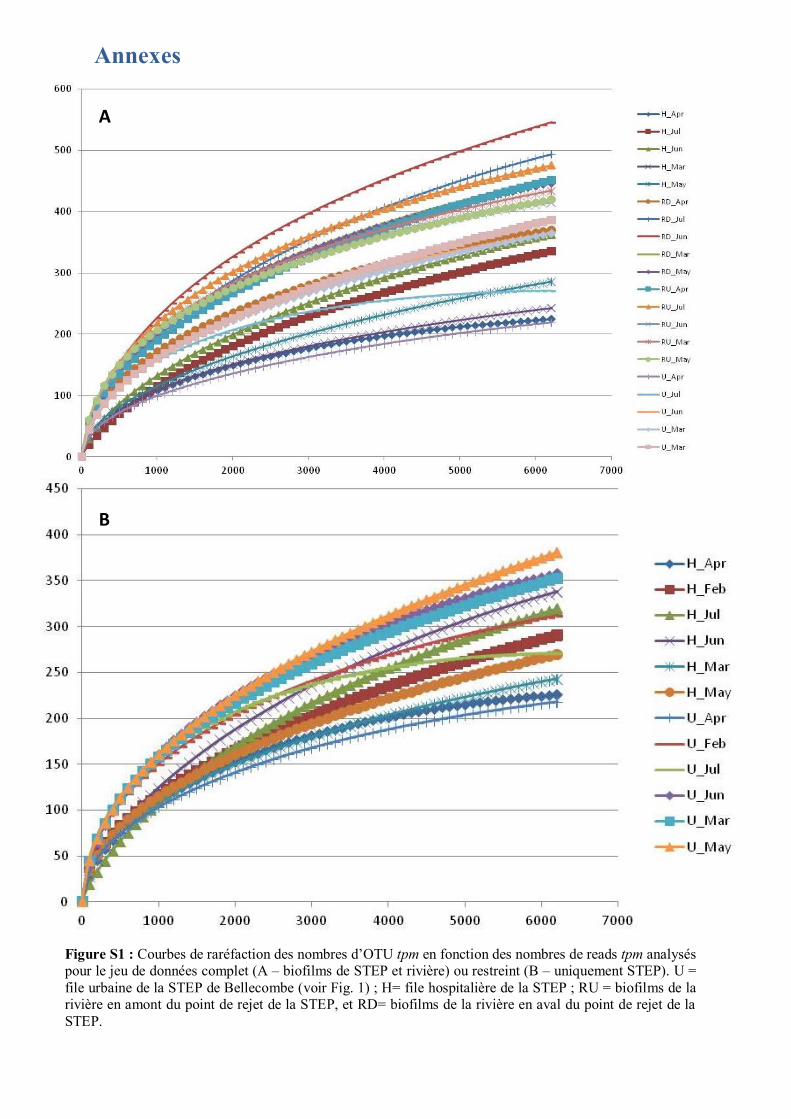
Annexes
Figure S1 : Courbes de raréfaction des nombres d’OTU tpm en fonction des nombres de reads tpm analysés pour le jeu de données complet (A – biofilms de STEP et rivière) ou restreint (B – uniquement STEP). U = file urbaine de la STEP de Bellecombe (voir Fig. 1) ; H= file hospitalière de la STEP ; RU = biofilms de la rivière en amont du point de rejet de la STEP, et RD= biofilms de la rivière en aval du point de rejet de la STEP.
A
B
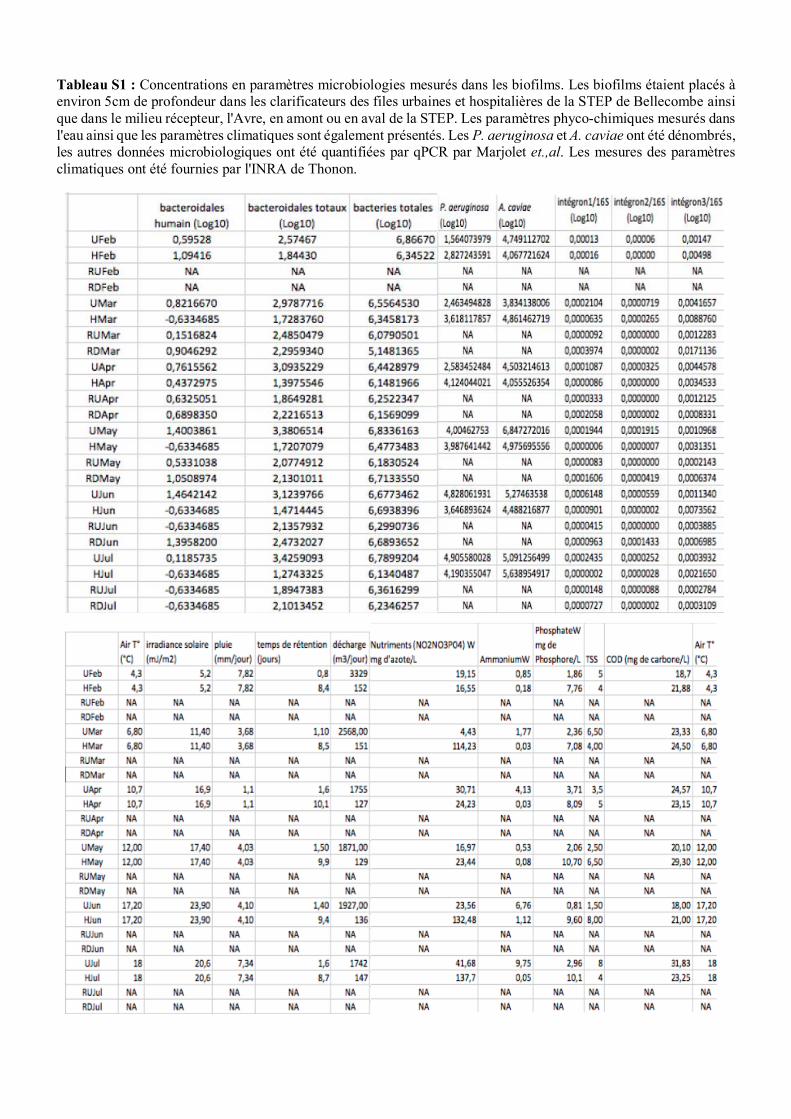
Tableau S1 : Concentrations en paramètres microbiologies mesurés dans les biofilms. Les biofilms étaient placés à environ 5cm de profondeur dans les clarificateurs des files urbaines et hospitalières de la STEP de Bellecombe ainsi que dans le milieu récepteur, l'Avre, en amont ou en aval de la STEP. Les paramètres phyco-chimiques mesurés dans l'eau ainsi que les paramètres climatiques sont également présentés. Les P. aeruginosa et A. caviae ont été dénombrés, les autres données microbiologiques ont été quantifiées par qPCR par Marjolet et.,al. Les mesures des paramètres climatiques ont été fournies par l'INRA de Thonon.
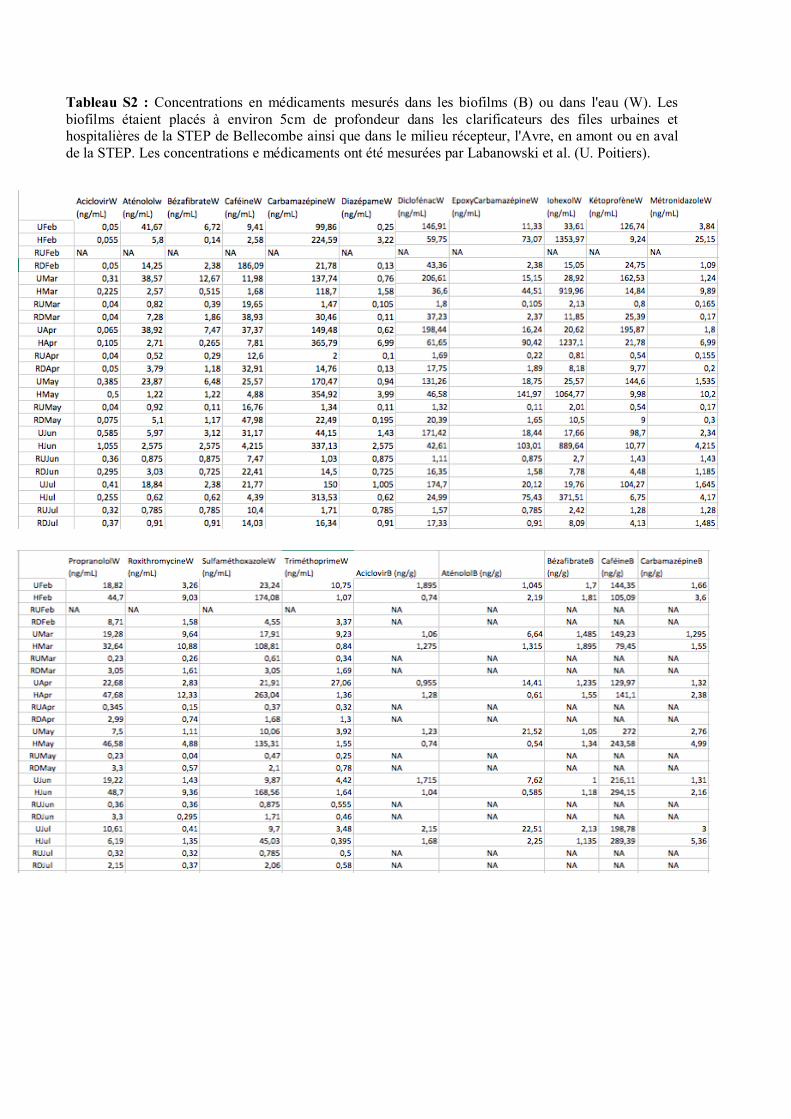
Tableau S2 : Concentrations en médicaments mesurés dans les biofilms (B) ou dans l'eau (W). Les biofilms étaient placés à environ 5cm de profondeur dans les clarificateurs des files urbaines et hospitalières de la STEP de Bellecombe ainsi que dans le milieu récepteur, l'Avre, en amont ou en aval de la STEP. Les concentrations e médicaments ont été mesurées par Labanowski et al. (U. Poitiers).

Suite du tableau S2 :
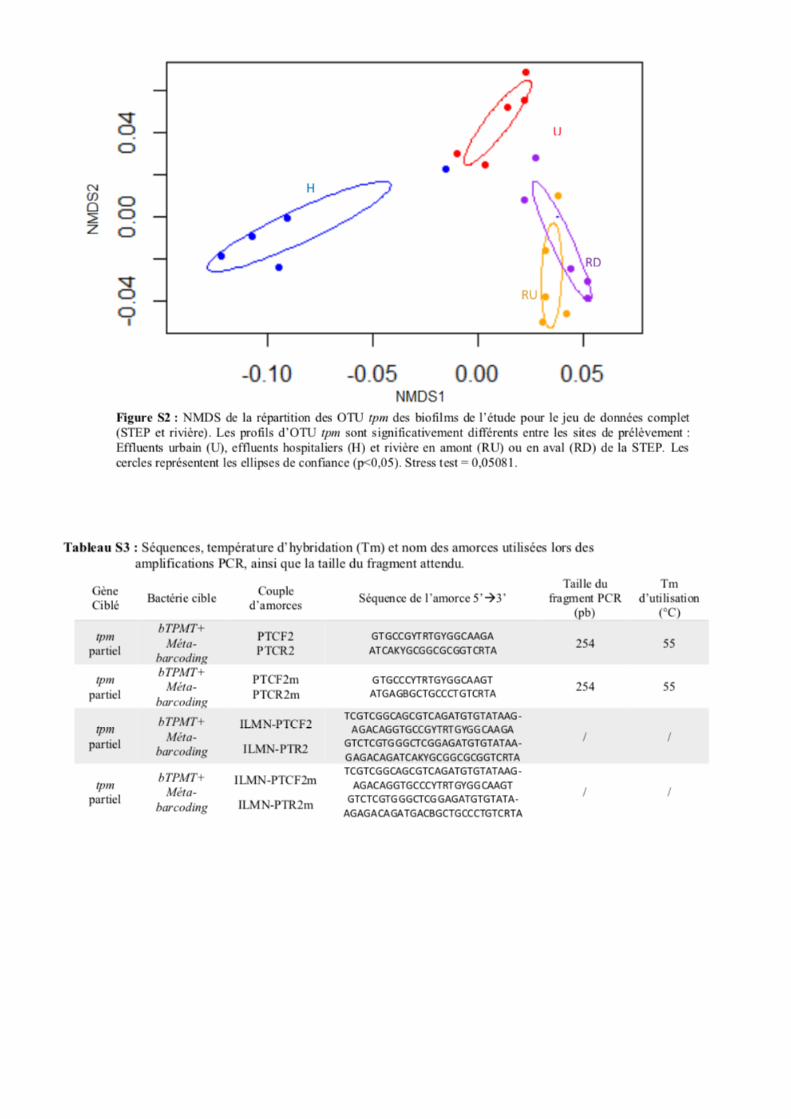
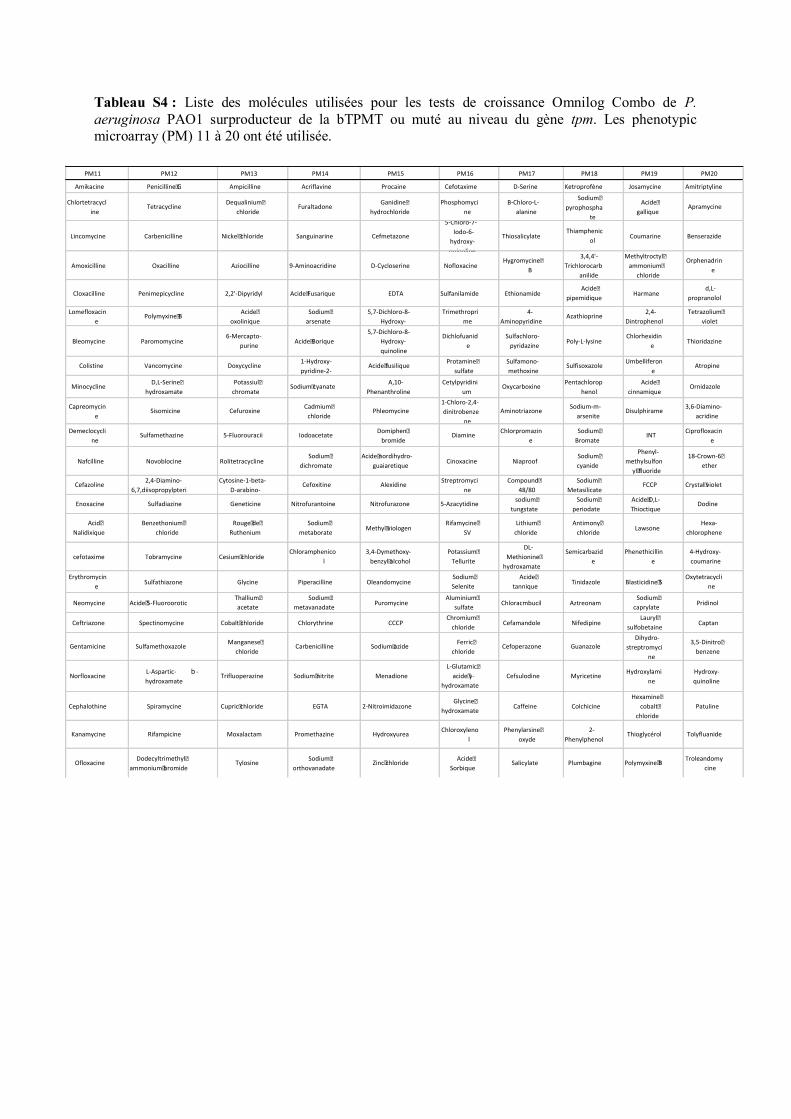
Tableau S4 : Liste des molécules utilisées pour les tests de croissance Omnilog Combo de P. aeruginosa PAO1 surproducteur de la bTPMT ou muté au niveau du gène tpm. Les phenotypic microarray (PM) 11 à 20 ont été utilisée.
PM11 PM12 PM13 PM14 PM15 PM16 PM17 PM18 PM19 PM20
Amikacine Penicilline�G Ampicilline Acriflavine Procaine Cefotaxime D-Serine Ketroprofène Josamycine Amitriptyline
Chlortetracycline
TetracyclineDequalinium�
chlorideFuraltadone
Ganidine�hydrochloride
Phosphomycine
B-Chloro-L-alanine
Sodium�pyrophospha
te
Acide�gallique
Apramycine
Lincomycine Carbenicilline Nickel�chloride Sanguinarine Cefmetazone
5-Chloro-7-Iodo-6-hydroxy-quinoline
ThiosalicylateThiamphenic
olCoumarine Benserazide
Amoxicilline Oxacilline Aziocilline 9-Aminoacridine D-Cycloserine NofloxacineHygromycine�
B
3,4,4'-Trichlorocarb
anilide
Methyltroctyl�ammonium�
chloride
Orphenadrine
Cloxacilline Penimepicycline 2,2'-Dipyridyl Acide�Fusarique EDTA Sulfanilamide EthionamideAcide�
pipemidiqueHarmane
d,L-propranolol
Lomefloxacine
Polymyxine�B Acide�oxolinique
Sodium�arsenate
5,7-Dichloro-8-Hydroxy-
Trimethroprime
4-Aminopyridine
Azathioprine2,4-
DintrophenolTetrazolium�
violet
Bleomycine Paromomycine6-Mercapto-
purineAcide�Borique
5,7-Dichloro-8-Hydroxy-quinoline
Dichlofuanide
Sulfachloro-pyridazine
Poly-L-lysineChlorhexidin
eThioridazine
Colistine Vancomycine Doxycycline1-Hydroxy-pyridine-2-
Acide�fusilique Protamine�sulfate
Sulfamono-methoxine
SulfisoxazoleUmbelliferon
eAtropine
MinocyclineD,L-Serine�
hydroxamatePotassiul�chromate
Sodium�cyanate A,10-Phenanthroline
Cetylpyridinium
OxycarboxinePentachlorop
henolAcide�
cinnamiqueOrnidazole
Capreomycine
Sisomicine CefuroxineCadmium�chloride
Phleomycine1-Chloro-2,4-dinitrobenze
ne
AminotriazoneSodium-m-arsenite
Disulphirame3,6-Diamino-
acridine
Demeclocycline
Sulfamethazine 5-Fluorouracii IodoacetateDomiphen�bromide
DiamineChlorpromazin
eSodium�Bromate
INTCiprofloxacin
e
Nafcilline Novoblocine RolitetracyclineSodium�
dichromateAcide�nordihydro-
guaiaretiqueCinoxacine Niaproof
Sodium�cyanide
Phenyl-methylsulfonyl�fluoride
18-Crown-6�ether
Cefazoline2,4-Diamino-
6,7,diisopropylpteriCytosine-1-beta-
D-arabino-Cefoxitine Alexidine
Streptromycine
Compound�48/80
Sodium�Metasilicate
FCCP Crystal�violet
Enoxacine Sulfadiazine Geneticine Nitrofurantoine Nitrofurazone 5-Azacytidinesodium�
tungstateSodium�
periodateAcide�D,L-Thioctique
Dodine
Acid�Nalidixique
Benzethonium�chloride
Rouge�de�Ruthenium
Sodium�metaborate
Methyl�viologenRifamycine�
SVLithium�chloride
Antimony�chloride
LawsoneHexa-
chlorophene
cefotaxime Tobramycine Cesium�chlorideChloramphenico
l3,4-Dymethoxy-benzyl�alcohol
Potassium�Tellurite
DL-Methionine�hydroxamate
Semicarbazide
Phenethicilline
4-Hydroxy-coumarine
Erythromycine
Sulfathiazone Glycine Piperacilline OleandomycineSodium�Selenite
Acide�tannique
Tinidazole Blasticidine�SOxytetracycli
ne
Neomycine Acide�5-FluorooroticThallium�acetate
Sodium�metavanadate
PuromycineAluminium�
sulfateChloracmbucil Aztreonam
Sodium�caprylate
Pridinol
Ceftriazone Spectinomycine Cobalt�chloride Chlorythrine CCCPChromium�chloride
Cefamandole NifedipineLauryl�
sulfobetaineCaptan
Gentamicine SulfamethoxazoleManganese�
chlorideCarbenicilline Sodium�azide Ferric�
chlorideCefoperazone Guanazole
Dihydro-streptromyci
ne
3,5-Dinitro�benzene
NorfloxacineL-Aspartic- b -hydroxamate
Trifluoperazine Sodium�nitrite MenadioneL-Glutamic�acide�y-
hydroxamateCefsulodine Myricetine
Hydroxylamine
Hydroxy-quinoline
Cephalothine Spiramycine Cupric�chloride EGTA 2-NitroimidazoneGlycine�
hydroxamateCaffeine Colchicine
Hexamine�cobalt�
chloridePatuline
Kanamycine Rifampicine Moxalactam Promethazine HydroxyureaChloroxyleno
lPhenylarsine�
oxyde2-
PhenylphenolThioglycérol Tolyfluanide
OfloxacineDodecyltrimethyl�
ammonium�bromideTylosine
Sodium�orthovanadate
Zinc�chloride Acide�Sorbique
Salicylate Plumbagine Polymyxine�B Troleandomycine
Tableau�S4�: �Liste�des�molécules�utilisées�pour�les�tests�de�croissance�Omnilog�Combo�de� P.�aeruginosa �PAO1�sur-producteur�de�la�bTPMT�ou�mutée�au�niveau�du�gène�tpm .�Les�phenotypic�microarray�(PM)�11�à�20�ont�été�utilisées.�
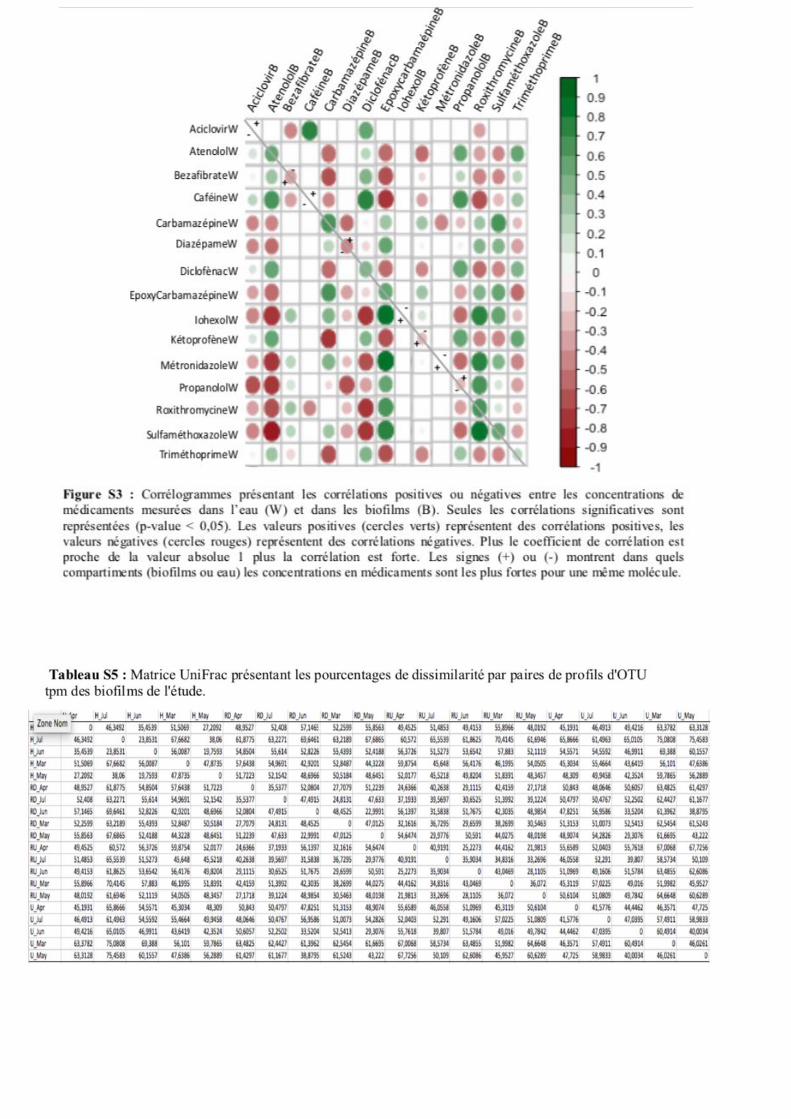

>séquence d’ADN obtenue à partir d’une amorce interne tpm (pIApX2-F) : GGGCATCCGGWCCTGMTCCTGTACCTCCGCATTCGTGGGAAAAACCGCAAGGTGGCGACA ATCGG TCTAGAGGATCC GCACGCCCCACCCGGTGTTGTAGCGTGCTTACAGTTTGGCGATGAGGCGAACCTGCATCA CCAGGGCGCTACTGATGACCGTCAGTAGCCAGCGGGGTGAAATCAAAGTTGATGGCATTA TGTGTTCAGAAGGCCGGTTTATCAAGCAGCGCCGGGGTTTACTGGCCTCTGGAGGTTATT TGACCGCGACCACCGCGCCCTCGGGGATGGGAGCAGGGAGCGCAATGGCAGTGGCTCCGA GGCGGCGTGTGACTAACTGGCCGTTGCGCCTGACGCCAACTAGGTCGGATTCCGTACCGG CATCCACGGCTGCCGCCAGCCCTGCCGGCGGCGAGGCGATGAAGGCGTAGCTGGTGCCGG CGGCGATGTAGCCCTGAAGCCGCACTGGCTTGCGGAACCAGGCCGGTAGACCAAGAGCGG AGTCCGCCGGCGAACCGCTGAAACCGGGGTTGGCGTGTGCGTACTCCGCCAGCCTGGAGC GGTAGAGCAGCAAGCTCCTGGCCAGGGTGTCGACCTCGGCCAGTTCGGCGCTCGAGGTCG CGTGGTTGCTTTGTACACTCAGCCAGGTCCCGGTGATCAGCGCGAGCACCAGGACGATCC ACATCAGCGGCATATGGCAGTCCTTGAGTCGTGGCGACCTGCTAGTTCACGCAGGTAACG GTGACGTTGGTCGAGTCGGGGGTCTTGTAGGGCTCCGGATAGTTGAAGCCCGGGCCGATC TCGGTGATCAGGCCGAACGAAGTAGTTGGTGCCAAAGGTCTGGTTGAAGCCCACCGCGCA GAATCGGCCGCCGCAGGTGATCGCGTAGCGGACATAGTCGTAATACATCATGCCGCTGTC CTGGCCGTGCAACGTAMACCAGTCCTTGATC CCCGGGTACCGAGCTCAAATCGAATCATGTCATACCTGTTTCCTGGGGAAATTGTTATC GTCCAATTCCCACCCCAACCCGGAAACTAAAGGGTAAGCCGGGGGGGCCAAAGGAGGGA CCACCCCCATTATTTGGTTGGCWCACTGGCCCCTTTCTATCGGRAAACTGTCGTGCCCT ATGTTWAAAAAAATGCCAACCWCGGGGAAAGGGGTTTTATAGGGACTCTTCGATTACAA ACAGAACAAAACCATAGGGTTGGAGGGAAAAATATAAYATAGGGGRATGKTTTYAMTAG GGGGAGAGAAAAAATTTAGAAGCCAAAGAAAAAAAAAGTKTTGRTTTTTTCCCCCCGAA AATACAAACGGAAAMTAAAGAAAACCMDCCCAGCCCCATACATAATACTATMCGG Figure S4 : Carte et analyse des séquences du vecteur pIApX2::tpm pour valider le positionnement du gène tpm.
pIA (séquence du plasmide)
tpm
pX2 (Promoteur constitutif fort)
SmaI KpnI SacI
XbaI BamHI
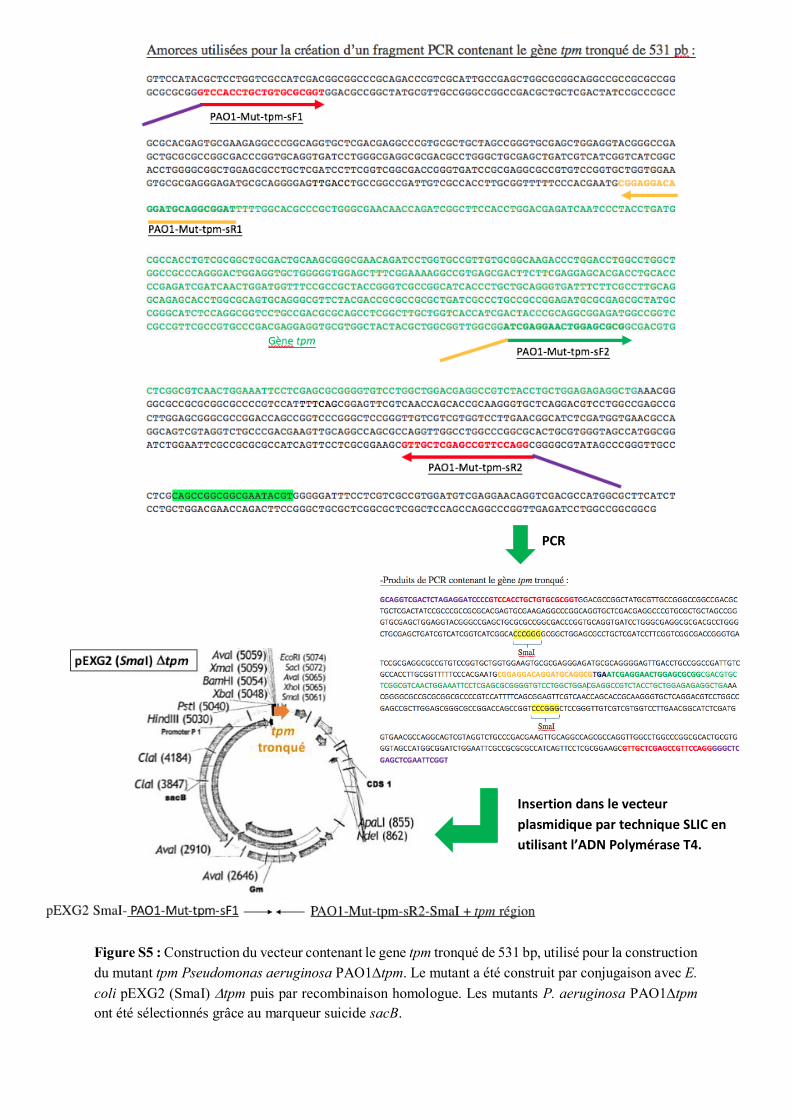
Figure S5 : Construction du vecteur contenant le gene tpm tronqué de 531 bp, utilisé pour la construction du mutant tpm Pseudomonas aeruginosa PAO1'tpm. Le mutant a été construit par conjugaison avec E. coli pEXG2 (SmaI) 'tpm puis par recombinaison homologue. Les mutants P. aeruginosa PAO1'tpm ont été sélectionnés grâce au marqueur suicide sacB.
PCR
Insertion dans le vecteur plasmidique par technique SLIC en utilisant l’ADN Polymérase T4.
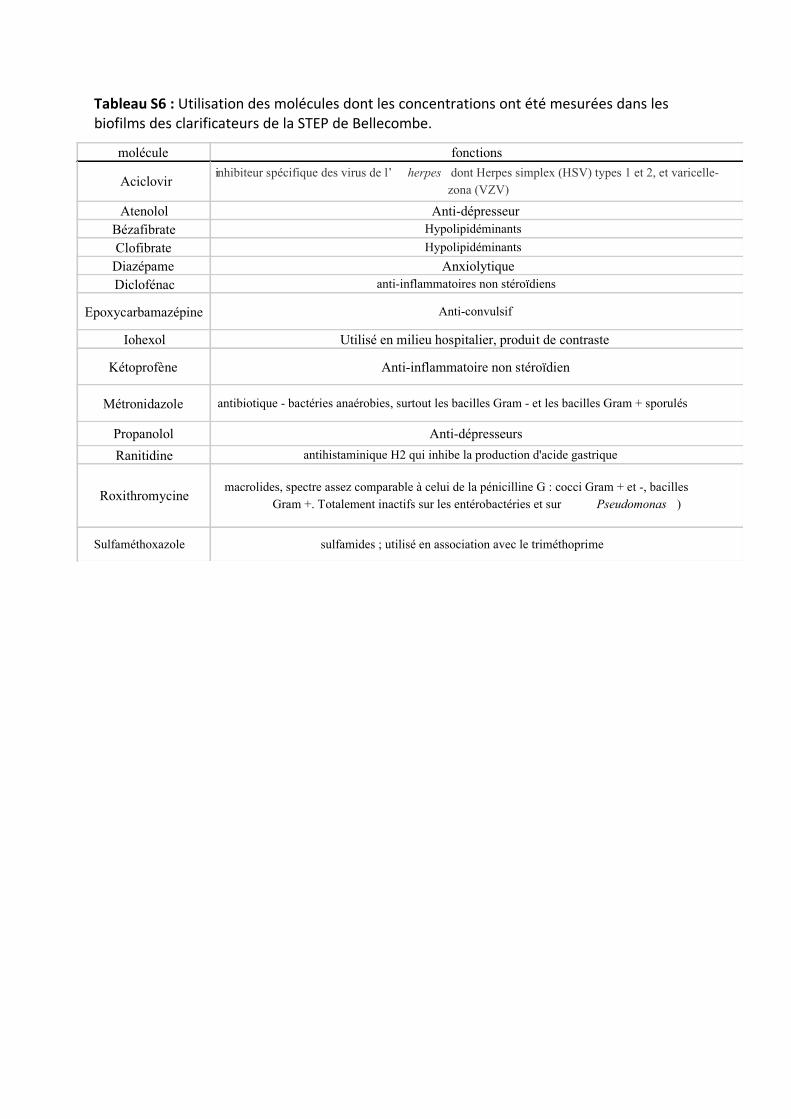
Tableau S6 : Utilisation des molécules dont les concentrations ont été mesurées dans les biofilms des clarificateurs de la STEP de Bellecombe.
molécule fonctions
Aciclovirinhibiteur spécifique des virus de l’ herpes dont Herpes simplex (HSV) types 1 et 2, et varicelle-
zona (VZV)
Atenolol Anti-dépresseurBézafibrate HypolipidéminantsClofibrate Hypolipidéminants
Diazépame AnxiolytiqueDiclofénac anti-inflammatoires non stéroïdiens
Epoxycarbamazépine Anti-convulsif
Iohexol Utilisé en milieu hospitalier, produit de contraste
Kétoprofène Anti-inflammatoire non stéroïdien
Métronidazole antibiotique - bactéries anaérobies, surtout les bacilles Gram - et les bacilles Gram + sporulés
Propanolol Anti-dépresseursRanitidine antihistaminique H2 qui inhibe la production d'acide gastrique
Roxithromycinemacrolides, spectre assez comparable à celui de la pénicilline G : cocci Gram + et -, bacilles
Gram +. Totalement inactifs sur les entérobactéries et sur Pseudomonas )
Sulfaméthoxazole sulfamides ; utilisé en association avec le triméthoprime

Résumé : Les effluents hospitaliers sont des milieux fortement pollués par des médicaments, antiseptiques,
détergents et biocides. Ces eaux représentent un cocktail favorable aux échanges génétiques entre
espèces bactériennes dont des formes pathogènes. Une enzyme bactérienne, la Thiopurine-S-Méthyl
transférase (TPMT), codée par le gène tpm, pourrait être impliquée dans la détoxication de certaines de
ces molécules, et expliquer l’occurrence de Pseudomonas aeruginosa et Aeromonas caviae dans ces
réseaux. Pour vérifier cette hypothèse, la diversité des gènes tpm dans des biofilms bactériens présent
dans les clarificateurs d’une station d’épuration a été comparée entre des files urbaines ou hospitalières.
Après amplification et séquençage NGS des amplicons tpm, l’occurrence des OTUs tpm a été confrontée
aux concentrations mesurées en médicaments. Ces OTUs ont pu être classées au sein de certains taxons
bactériens. Ceci a permis d’observer une association entre le nombre de séquences tpm affiliées aux
espèces P. putida et A. caviae au sein des biofilms de la file hospitalière, et la présence de certains
médicaments. Pour préciser le rôle de la TPMT dans la résistance ou tolérance aux antimicrobiens
présents, des tests de croissance ont complété cette étude. Ces tests ont permis d’observer une relation
entre la résistance à un biocide et un floculant avec effet bactéricide.
Abstract : Hospital wastewaters are highly contaminated by drugs, antiseptics, detergents and biocides. These
waters represent a supportive environment for genetic exchanges between bacterial species including
pathogens. A bacterial enzyme, the Thiopurine-S-Methyltransferase (TPMT), encoded by the tpm gene,
could be involved in the detoxification of some of these molecules. This can explain the Pseudomonas
aeruginosa and Aeromonas caviae occurrence. To verify this hypothesis, the diversity of tpm gene in
bacterial biofilm presents in a wastewater epuration station was compared. These biofilms have been
developed in the urban or hospital treatment files. After tpm amplicons amplification and NGS
sequencing, the tpm OTUs occurrence was compared with the measured drugs concentrations. These
OTUs could be classified within certain bacterial taxa. This allowed to observe an association between
tpm sequences number affilied with P. putida and A. caviae present in the hospital biofilms, and the
presence of certain drugs. To clarify the role of TPMT in antimicrobial resistance, growth tests
completed this study. These tests showed a relation between the resistance to a biocide and a flocculant
with a bactericidal effect.