Embed Size (px)
Citation preview

URPIGM Unite de recherche en Pathologie, Inununologie et Genetique Moleculaire
RAPPORT DE STAGE
presente par
Vinca LARDANS
Application a l'endocrinologie des Mollusques des techniques
de separation par HPLC, d'immunodosage et de PCR.
septembre 1991
FREMER

URPIGM Unité de recherche en Pathologie, Immunologie et Génétique Moléculaire
RAPPORT DE STAGE présenté par
VincaLARDANS
Application à l'endocrinologie des Mollusques des techniques
de séparation par HPLC, d'immunodosage et de PCR.
septembre 1991 .

1
REMERCIEMENTS
Je remercie Evelyne Bachère, responsable de l'URPIGM pour m'avoir accueillie dans son laboratoire, et Kristell Kellner qui a accepté de m'encadrer durant ce stage. Ce fut un plaisir de travailler en sa collaboration et je tiens à lui exprimer toute ma sympathie et ma reconnaisance pour les connaissances scientifiques qu'elle m'a inculquées, pour sa gentillesse et sa patience.
J'adresse mes remercie ments à toutes les autres personnes de "l'équipe" pou~ leur disponibilité, leur aide tout au long de mon stage et pour la rédaction du rapport, et pour leur bonne humeur.
Soyez tous assurés que je gardera i un excellent souvenir de ce passage par La Tremblade.

1
LISTE DES ABBREVIATIONS
ADN: acide désoxyribonucléique ADNe: adn complémentaire ARN: acide ribonucléique ARNm: arn messager ASP: activation de synthèses protéiques bp: paire de bases BSA: serum a lbumin bovine ("bovine sérum albumine") CCK: cholecystokinine CPG: controlled pore glass CPM: coups par minutes DEPC: diéthyl pyrocarbonate dA TP: désoxyadénosine triphosphate dCTP: désoxycytidine triphosphate dGTP: désoxyguanidine triphosphate DMT: diméthoxytrityl dNTP: désoxynucléotide triphosphate dTTP: désoxythimidine triphosphate DO: densité optique DPM: désintégrations par minute DTT: dithiothréitol ELISA:" enzyme linked immunosorbent assay" GC: ganglions cérébroïdes GuSCN: isothiocyanate de guanidine HPLC: chromatographie liquide haute performance MAB: anticorps monoclonal ("monoclonal antibody") MIPc: peptide de mollusque apparenté à la chaine C de l'insuline PAB: anticorps monoclonal ("polyclonal antibody") PBS: tampon phosphate salin ("phosphate buffer saline") PCR: polymerase cha in reaction PM: poids moléculaire PTTH: hormone prothoracicotropique ("prothoracicotropic hormone") TF A: acide trifluoroacétique ("trifluoroacetic acid") TEAA: acétate de triéthylammonium

SOMMAIRE
REMERCIEMENTS
LISTE DES ABREVIATIONS
INTRODUCTION 1
PREMIERE PARTIE: ETUDE DE PEPTIDES ACTIFS EXTRAITS DE GANGLIONS CEREBROIDES DE MOULES 3
MATERIEL ET METHODES 4
I- Fractionnement par HPLC de peptides extraits de ganglions cérébroïdes 4 II-Mesure de l'effet activateur de la synthèse protéique de cellules de moules 4
II -1 Préparation de suspensions de cellules dissociées de moule 4 II - 2 Estimation de la synthèse proté ique des fractions obtenues par HPLC 5 II -3 Tests d'inhibition par anticorps monoclonaux sur l'activité de l'extrait
brut de ganglions cérébroïdes. 5 III- Immunodosage 5
RESULTATS 7
I-Fractionnement de l'extrait de ganglions 7 II-Etude de l'effet des fractions obtenues par HPLC sur l'activation de la synthèse
protéique (ASP). 8 III- Etude de l'effet des MAB sur l'activité de l'extrait brut de ganglions cérébroïdes
de moules 9 IV- Caractérisation antigénique des fractions séparées par HPLC. 10
DISCUSSION - CONCLUSION 15
DEUXIEME PARTIE: RECHERCHE DE GENES D'HORMONES 17
MA TE RIELS ET METHODES 18
I-Extraction d'acides nucléiques 18 I - 1 Extraction de l'ADN génomique de moules entières 18 I -2 Préparation d'ADNe d'embryons 18 I -3 Préparation d'adne de ganglions cérébroïdes 19
II Choix et synthèse des amorces 19 II -1 Analyse des banques de séquences nucléiques et protéiques avec système
d'exploitation bisance 19 II -2 Synthèse et purification d'oligonucléotides 19
III- Amplification de séquences par PCR 20

RESULTATS
1- Préparation de l'ADN cible l - 1 Extraction de l'ADN génomiq ue de moules l - 2 Préparation d'ADNe d'embryons et de GC
II - Choix des amorces II - 1 Choix des amorces pour le gène récepteur d'insuline II -2 Choix des amorces pour le gène de prolactine II - 3 Choix des amorces pour le gène d'insuline
III - Synthèse et purification des oligonuclèotides IV- Amplification par PCR
lV- 1 Amplification de séquences de type insuline récepteu r lV-2 Amplification de séquences de type prolactine IV-3 Amplification de séq uences d'insuline
DISCUSSION - CONCLUSION
ANNEXES
BIBLIOGRAPHIE
21
21 21 21 21 21 23 23 24 25 25 27 28
31

INTRODUCTION
Chez les Vertébrés, des progrès très significatifs ont été réalisés concernant la caractérisation et la localisation d'hormones ou de neurotransmetteurs. Un grand nombre de substances ont déjà été purifiées et les séquences protéiques et nucléiques ont été caractérisées). En ce q ui concerne les Invertébrés, les travaux traitant de l'endocrinologie ont porté en premier lieu sur la classe des insectes. La facilité de réalisation d'expériences d 'ablation d'organes sécréteurs, de cautérisations ou de greffes a permis de mettre en évidence l'importance du contrôle hormonal dans la mue ou la métamorphose (Wigglesworth, 1970). Par la suite, des hormones protéiques telles que la bombyxine ont été purifiées puis séquencées (Adachi et al, 1989). Les techniques d'immununologie et de biologie moléculaire ont pe rmis de compléter ces recherches et d 'accéder à la séquence des gènes. Pour certaines hormones comme la prothoracicotropic hormone (PTTH) (Jhoti et al, 1987) ou la cholecystokinine (CCK) (De Loof et al, 1990), de très fortes homologies avec des peptides identifiés chez les Vertébrés ont été révélées.
Le domaine de l'endocrinologie des mollusques n'a été que plus récemment abordé, su r le modèle de la Lymnée Lymnaea stagnalis. Des organes endocrines ont été identifiés et cartographiés. Des gènes apparentés au gène d'insuline de Vertébrés (Smit et al, 1988; Smit, 1990» ou codant pour des hormones impliquées dans les comportements de ponte (Vreugdenhil et al, 1988) ont été identifiés dans les banques d'ADNe.
Chez les mollusques lamellibranches, aucune glande endocrine ni aucun orga ne neurohémal n'a été identifié. Cependant, Gabe (1955) et Lubet (1959) ont mis en évidence la présence de cellules neurosécrétrices au niveau des ganglions cérébro-pédieux (cérébroïdes) et viscéraux. Différents types de cellules ne urosécrétrices ont été observés selon des méthodes histologiques classiques et ultrastructurales (Mathie u, 1987), mais l'inaccessibilité des organes neurosécréteurs (enfermés dans la coquille) rend difficile toute expérimentation de type cautérisation, a blation ou greffe. C'est pourquoi l'utilisation de techniques immunologiques et la biologie moléculaire s'avèrent intéressantes.
L'URPIGM (Unité de Recherche en Pathologie, Immunologie et Génétique Molécula ire), en association avec le laboratoire de zoologie de l'unive rsité de Caen, a comme ncé une é tude dans le domaine de l'endocrinologie des mollusques et plus particulièrement la moule Mytilus.edulis. A l'heure actuelle, des cellules de ganglions cérébroïdes de moules ont été caractér isées antigéniquement grâce â des anticorps monoclonaux (MAB) spécifiques de types cellulaires. Le protocole d'hybridation lymphocytaire, qui avait été utilisé pour la préparation des anticorps il partir de souris immunisées avec des homogé néisats de ganglions cérébroïdes, est décrite en annexe 1. Ces réactifs ont conduit à l'identification antigénique de cellules potentiellement neurosécrétrices et à une approche cartographique des ganglions céréhoïdes en immunohistochimie. La mise a u point d'un test immunoenzymatique de type ELISA a permis de sélectionner des MAB spécifiques de déterminants antigéniques présents dans l'hémolymphe.
Dans le cadre de mon stage, deux aspects de l'étude ont été abordés:
- Un premier point, abordé lors d'un séjour de 15 jours dans le laboratoire de M. Mathieu de l'unive rsité de Caen, concerne la séparation de molécules peptidiques extraites de ganglions cérébroïdes de moules: Plusieures fractions moléculaires ont été identifiées sur la base de leu r PM, de leur activité sur la synthèse protéique, de leur réactivité vis-â-vis d'anticorps hétérologues spécifiques de différentes hormones ou de l'un des MAB spécifiques de ganglions.
1

Parallèlement, l'effet des MAB sur l'activation de la synthèse protéique a été étudié.
- Un deuxiéme aspect a consisté à rechercher dans le génome et dans les transcripts de la moule des séquences susceptibles de correspondre à des molécules impliquées dans les régu lations hormonales. Cette recherche a été réalisée selon la technique d'amplification par PCR (Polymerase Chain Reaction) schématisée en annexe 2, après avoir identifié des séquences consensus parmi celles établies pour des hormones ou des récepteurs d'hormones de différents groupes taxonomiques.
2

1
PRE MIE RE PARTIE ETUDE DE PEPTIDES ACTIFS EXTRAITS DE GANGLIONS CEREBROIDES
DEMOULES
Afin de contourner les obstacles posés par les caractéristiques anatomiques et physiologiques de la moule, un test biologique in uitro de dosage de l'activité de synthèse protéique a été mis au point par Lenoir et Mathieu (1986). Ce test, rapide et reproductible, permet de mesurer l'incorporation d'acides aminés marqués dans une suspension de cellules dissociées de bords de manteaux. Grâce à ce bioessai, J.Y. Toullec (1989) a montré l'effet activateur de la synthèse protéique (ASP) d'extrait brut de ganglions cérébroïdes (GC) de moules ajouté à la suspension de cellules de bords de manteaux. En outre, après séparation de l'extrait brut sur Sephadex G50, il a retrouvé l'effet ASP dans la fraction contenant des molécules d'environ 1000 Da.
Pour étudier la faisabilité de purification de la fraction active par HPLC, nous nous sommes attachés, dans un premier temps, à rechercher grâce au biotest, l'effet ASP de l'extrait de ganglions cérébroïdes de moule après séparation par HPLC sur une colonne protéine P AK 125.
Parallèlement à cette étude, la réactivité des différents MAB spécifiques de GC de moule ou de certains PAB hétérologues (anti-insuline, anti-somatostatine) a été mesurée par la technique d'immunodosage enzymatique (mis au point sur l'extrait brut de ganglions) sur les fractions issues de la séparation par HPLC des constituants des GC. De plus, l'effet inhibiteur de certains MAB sur l'ASP a été testé.
3

MATERIEL ET METHODES
I-Fractionnement par HPLC de peptides extraits de ganglions cérébroïdes
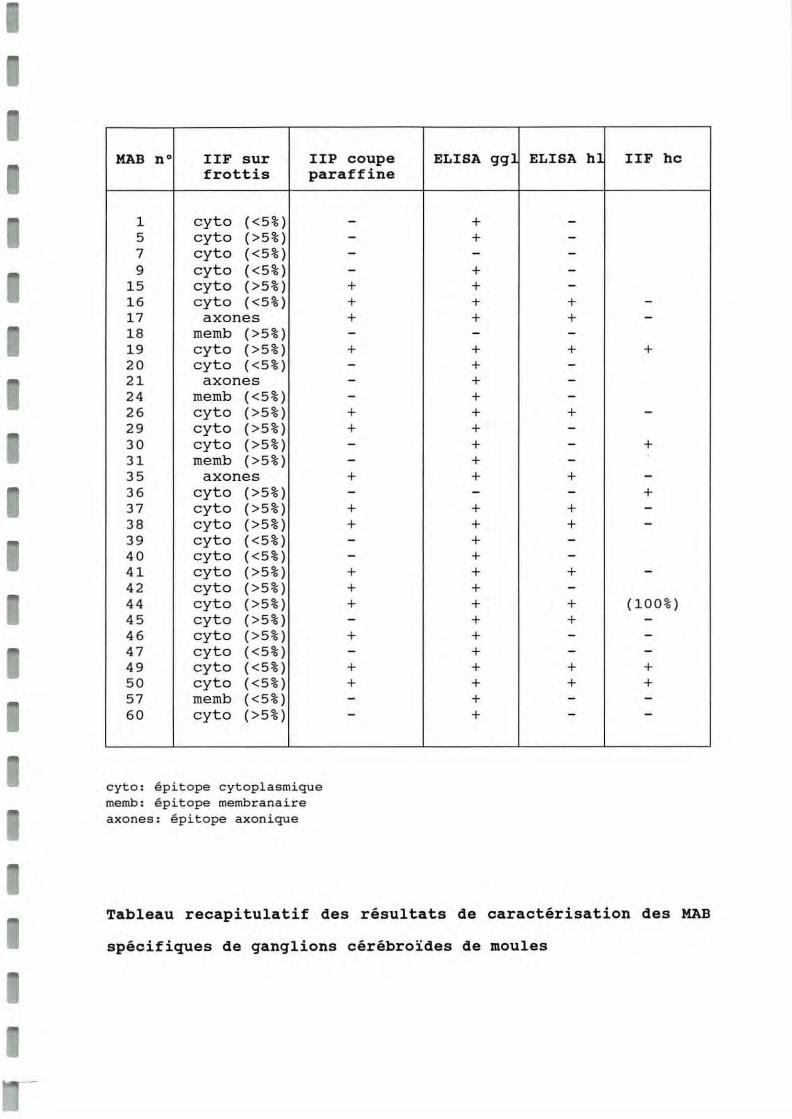
Les ganglions cérébroïdes de moule sont situés symétriquement le long des muscles rétracteurs antérieurs du byssus (annexe 3). Les ganglions sont repérés par observation à la loupe binoculaire et la microdissection s'effectue par section des connectifs cérébro-pédioviscéraux, des nerfs palléaux antérieurs et de la commissure intercérébrale. Cent ganglions dissequés sont dissociés à l'aide d'un homogénéiseur de tissus de type Dounce dans 1 ml d'acétate d'ammonium 10 mM. L'homogénéisat est centrifugé 10 mn à 1000 g afin d'éliminer les débris cellulaires et le surnageant est filtré sur filtre Millipore HV O,45p.m. L'équivalent de 2 ganglions cérébroïdes, soit 20 p.l de filtrat, sont injectés dans une colonne d'HPLC Protéine Pak 125, adaptée à la séparation des biopolymères protéiques dans une gamme de poids moléculaires compris entre 1000 et 30000 Da (structure linéaire). La séparation est réalisée en conditions isocratiques dans un tampon acétate d'ammonium 10 mM, à une pression de 0,9 mBar. Le débit de la colonne est de 500 p.l par minute. La séparation est contrôlée par mesure de la densité optique à 220 nm à la sortie de la colonne Protéine Pak. Le profil de DO est enregistré sur un traceur papier (déroulement: 0,5 cm/mn). Des fractions de 0,5 ml sont collectées à la sortie du système. Le volume mort entre le lecteur de DO et le collecteur est de 1 ml (soit un décalage de 1 cm sur la courbe). Les fractions ayant une absorbance significative à 220 nm sont retenues et lyophilisées.
II-Mesure de l'effet activateur de la synthèse protéique de cellules de moules
Le protocole utilisé a été mis au point par Lenoir et Mathieu (1986) et utilisé pour mesure r l'effet ASP d'un extrait de GC de moule.
11-1 Préparation de suspensions de cellules dissociées de moule
Les moules Mytilus edulis, de Basse- Normandie, sont placées en salle d'élevage en eau de mer filtrée. 48 heures avant l'expérimentation, des antibiotiques (pénicilline: 500000 U.l, auréomycine: 30 mg/!) sont ajoutés à l'eau de mer. L'ensemble du protocole est réalisé en conditions stériles sous hotte à flux laminaire. Le matériel utilisé est préalablement stérilisé. Le bord de manteau constitue une zone de croissance linéaire chez les Mollusques et lorsqu'il est spécifiquement prélevé, ne contient ni cellule germinale ni cellule de stockage (Bu bel, 1973).
10 à 15 g (poids frais) de bords de manteau sont prélevés dans 20 ml de milieu de Hanks stérile de composition suivante:
· 2,1717 g de milieu de Hanks 199 Boehringer · 4,38 g de NaCI · 0,187 g de KCl . 1,5 g de MgS04 · 0,275 g de CaCl2 · 1,195 g d'Hépès · eau distillée qsp 250 ml
pour 250 ml de milieu à 1100 mosm ajusté à pH 7,4 et filtré (O,22p.m).
Les bords de manteaux sont ensuite découpés en petits morceaux à l'aide d'un ciseau stérile. 20 mg de pronase (soit 0,1%) ajoutés au milieu permettent la dissociation des tissus (20 mn à
4

1
température ambiante sous agitation modérée). La suspension est filtrée sur de la toile à bluter stérile de 100 JJ.m de maille afin de retenir les agrégats et fragments non dissociés. Ceux-ci sont récupérés et subissent une nouvelle dissociation enzymatique suivie d'une filtration. L'ensemble des cellules est repris dans 50 ml de milieu puis centrifugé 4 mn à 200 g à température ambiante. Le surnageant est é liminé, le culot de cellules est repris dans 40 ml de milieu de Hanks et lavé une seconde fois, les lavages permettant l'élimination des enzymes de dissociation. Le culot est finalement repris dans le volume de milieu nécessaire à l'expérience (soit 0,5 ml de suspension cellulaire par tube).
II -2 Estimation de la synthèse protéique des fractions obtenues par HPLC
Chaque tube du test contient: - 500 Ji.l de suspension cellulaire (7 à 12 millions de cellules/ml) - 50 JJ.l d'une solution d'un mélange d'acides aminés marqués au carbone 14 (Dositek) soit
0.1 JJ.Ci par tube - 100 Ji.l des fractions à tester soit 0,6 équivalent ganglion (ou de milie u pour les témoins
négatifs)
Chaque essai est réalisé en trois exemplaires. Après incubation à l4°C sous agitation continue pendant 4 heures, les cellules sont précipitées par addition de 500 ,ul de TCA 10% froid (une nuit à 4°C). Les cellules de bords de manteau sont mises au culot par centrifugation pendant 10 mn à 1500 g et reprises dans 3 ml de TCA 10%. Ce lavage est répété 3 fois. Le culot de cellules est finalement solubilisé dans 500 ,ul de KOH 0,3 N pendant une heure à 40°C. Le comptage est réalisé sur un aliquot de 200 ,ul dans 5 ml de liquide scintillant à l'aide d'un compteur à scintillation (Packard 1600 CA).
II-3 Tests d'inhibition par anticorps monoclonaux sur l'activité de l'extrait brut de ganglions cérébroïdes.
Chaque tube contient: - 500 ,ul de suspension cellulaire -50 ,ul d'un mélange d'acides aminés marqués au carbone 14. - 10 JJ.l d'extrait brut préparé dans de l'acétate d'ammonium 10 mM, centrifugé (10 min,
1000 g) et filtré à 0,22 ,urn (soit un équiva lent ganglion) . - 90 ,ul de surnageant de culture de X MAB différents spécifiques de cellules
ganglionnaires.
Les essais sont réalisés en 4 exemplaires. Chaque tube est soumis au même protocole que précédemment cf II -2.
III-Immunodosage
L'activité des fractions vis-à-vis soit d'anticorps monoclonaux homologues spécifiques de cellules de ganglions cérébroïdes de moules, soit d'anticorps polyclonaux hétérologues a été mesurée selon la technique ELISA mise au point précédemment au laboratoire sur des extraits bruts de ganglions cérébroïdes ( annexe 1). Ce test est réalisé en microplaque de 96 puits. Les fractions d'HPLC lyophilisées sont reprises dans 100 ,ul de PBS (NaCl:8 g/1; KH2P04:0,2 g/l; NaHP04, 12H20:2,9 g/1; KC1:0,2 g/l),puis déposées dans les puits. L'étape de coating est réalisée
5

pendant une nuit à 4·C. La plaque est ensuite vidée et soumise à un lavage: - 3 fois 2001,[ de PBS Tween 20 1% pendant 3 mn à température ambiante . - 2 fois 200,,[ de PBS pendant 3 mn à température a mbiante.
La plaque est a lors saturée par une solution de PBS-BSA 5% pendant une heure à 37· C (100 l'l/puits) . Après un nouveau lavage, l'anticorps primaire est déposé dans la plaque (100 l'l/puits). Il s'agit soit:
• de MAB spécifiques des ganglions cérébroïdes de moules (surnageants dilués au 1/2; cf tableau en annexe 1)
• d'un anticorps polyclonal a nti-insuline humaine de cobaye (Biogenex laboratories; dilué au 1/5)
• d'un anticorps polyclonal anti -somatostatine (dilué au 1/500) • d'un anticorps polyclonal anti-MIPc (Molluscan Insulin like Peptide chain C, dilué au
1/1000 aimablement fourni par le Dr Van Minnen
Après 2 heures d'incubation à 37·C, la plaque subit un nouveau lavage. Un anticorps secondaire spécifique de l'anticorps primaire couplé â la péroxydase (100 l'l), et dilué dans un tampon PBS-sérum de porc 1% (au 1/4000 pour l'anticorps anti-souris, au 1/1000 pour les autres anti-anticorps primaires) sont ajoutés dans chaque puits. L'incubation d 'une he ure à 37· C est suivie d'un dernier lavage.
La révélation est obtenue par incubation de la plaque à température ambiante pendant 10 mn après addition de 100 ILl de la solution de chromogène suivante:
- 20 mg d'orthophényldiamine - 10 ml de tampon citrate- acétate 0,2 M, pH =5,6 - 10,,, d'H2ü2
L'absorbance à 492 nm est mesurée grâce à un spectrophotométre lecteur de plaques.
6
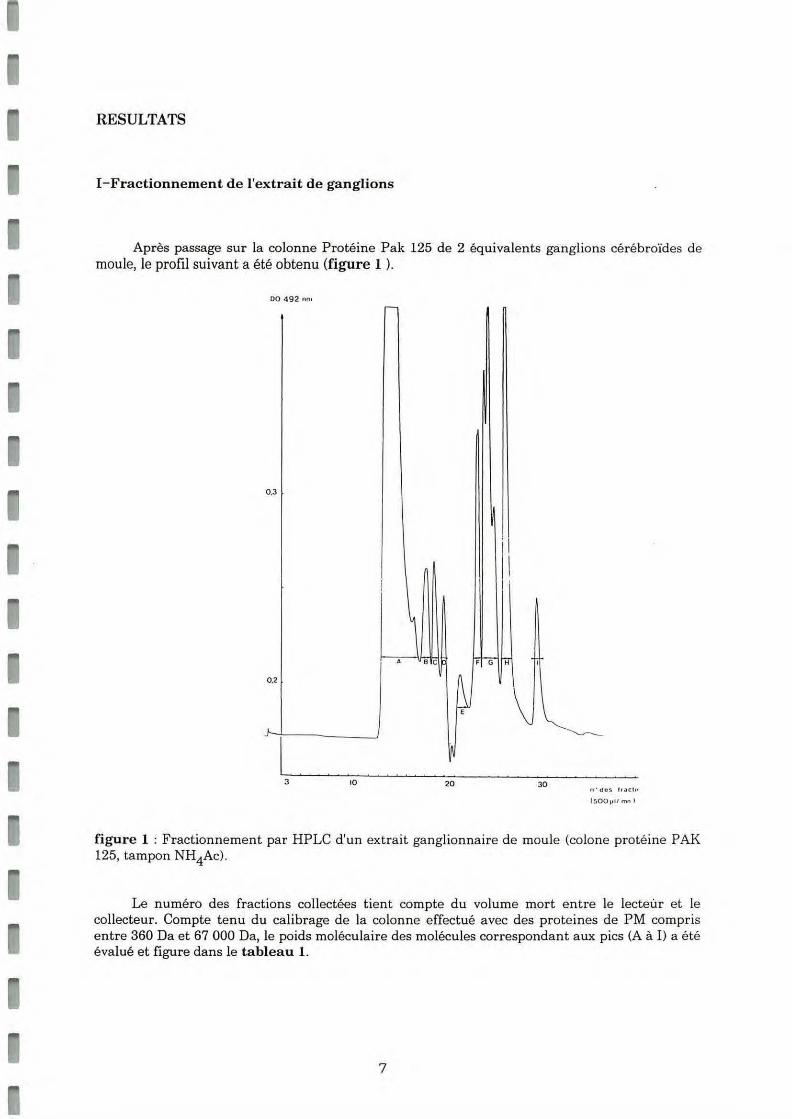
RESULTATS
!-Fractionnement de l'extrait de ganglions
Après passage sur la colonne Protéine Pak 125 de 2 équivalents ganglions cérébroïdes de moule, le profil suivant a été obtenu (figure 1 ).
DO 492 nm
0 .3
Il Il 1'
A B C
0 .2
3 10 20 30 n ·des fr ;ac t •~
(S00vll mn)
figure 1 : Fractionnement par HP LC d'un extra it ganglionna ire de moule (colone protéine P AK 125, tampon NH4Ac).
Le numéro des fractions collectées tient compte du volume mort entre le lecteùr e t le collecteur. Compte tenu du calibrage de la colonne e ffectué avec des proteines de PM compris entre 360 Da et 67 000 Da, le poids molécula ire des molécules correspondant aux pics (A à 1) a été évalué et figure dans le tableau 1.
7

nom du pic g a mme de poids moléculaire des molécules correspondantes
A 1 00 0 00 à 45 0 000 Da B 5 0 000 à 1 0 0 00 0 Da c 10 000 à 5 0 000 Da D 2 000 à 1 0 00 0 Da E 5 500 à 7 000 Da F 4 000 à 4 7 00 Da G 2 700 à 4 0 00 Da H 2 000 à 2 500 Da I 1 000 à 1 2 00 Da
Tableau 1: évaluation des poids moléculaires des différents pics de la courbe de fractionnement d'extrait ganglionnaire
La reproductibilité de la séparation a é té vérifiée par l'obtention de profils sembla bles avec p lusieurs lots d'extraits gan glionnaires.
II-Etude de l'effet des fraction s obtenues par HPLC sur l'activation d e la synthèse protéiq ue (ASP).
Les fractions collectées après HPLC ont été regroupées deux par deux et leur ASP sur des cellules de bords de manteaux a été mesurée par l' incorporation d'acides aminés marqués. Les valeurs numériques figurent dans le tableau présenté en annexe 4.
Ces résulta ts sont reportés sous forme d'histogramme sur la figure 2, sur laquelle est présenté également le profil d'HPLC d'extrait de ganglions.
8
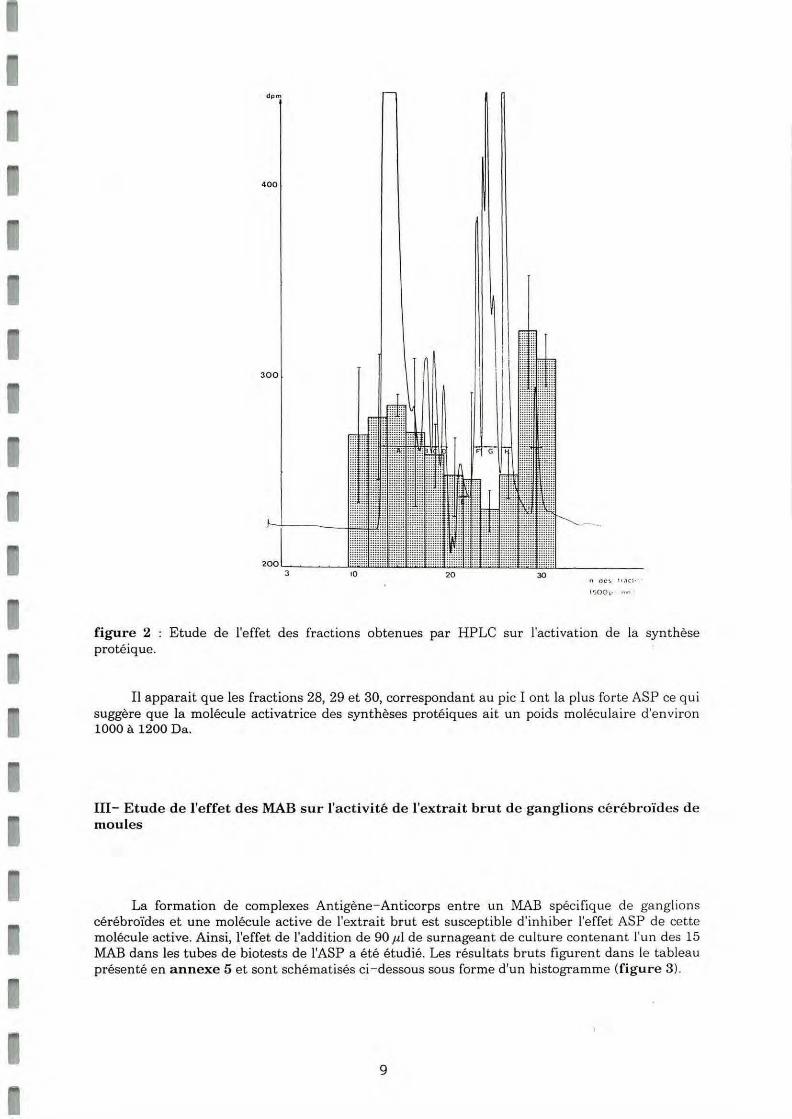
dp
400
300
10
l 500 p "'"
figure 2 : Etude de l'effet des fractions obtenues par HPLC sur l'activation de la syn t hèse protéique.
Il apparaît que les fractions 28, 29 et 30, correspondant au pic I ont la plus forte ASP ce qui suggère que la molécule activatrice des synthèses protéiques ait un poids moléculaire d 'environ 1000 à 1200 Da.
III- Etude de l'effet des MAB sur l'activité de l'extrait brut d e ganglions cérébroïdes d e moules
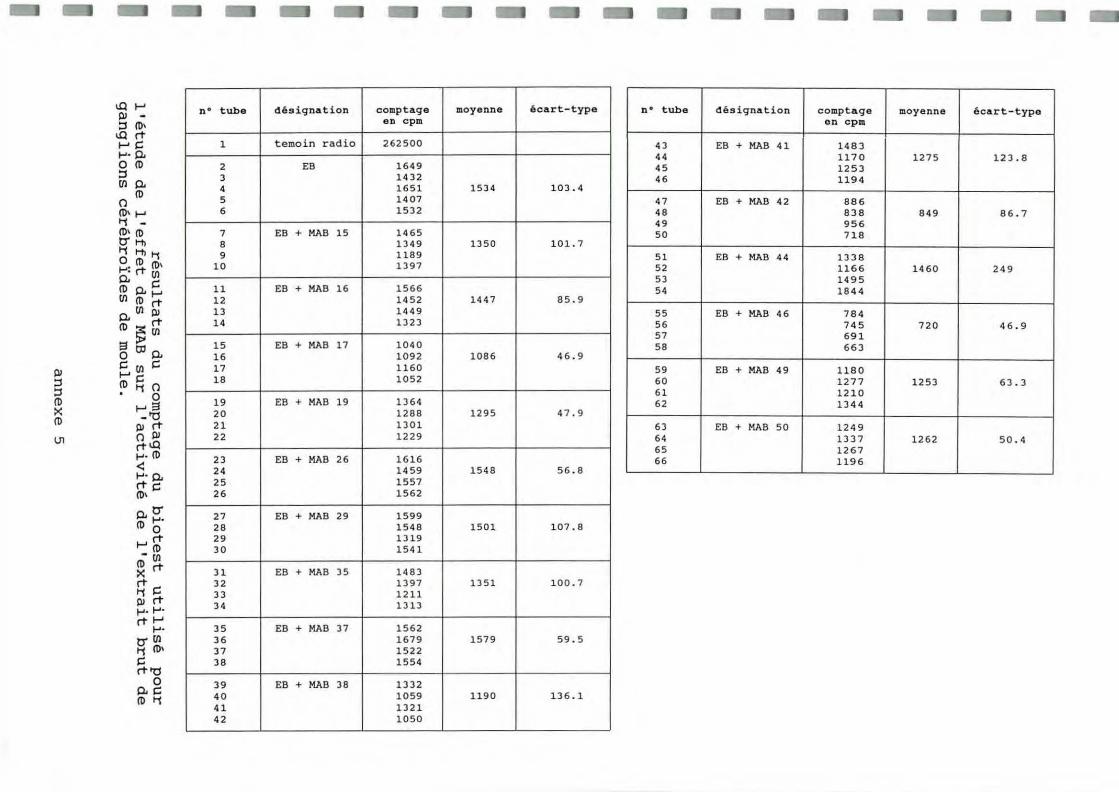
La formation de complexes Antigène- Anticorps entre un MAB spécifiq ue de ga nglions cérébroïdes et une molécule active de l'extra it brut est susceptible d 'inhiber l'effet ASP de cette molécule active. Ainsi, l'effet de l'addition de 90 .ul de surnageant de culture contenant l'un des 15 MAB dans les tubes de biotests de l'ASP a été étudié. Les résultats bruts figurent dans le tableau présenté en annexe 5 et sont schématisés ci-dessous sous forme d'un histogramme (figure 3).
9
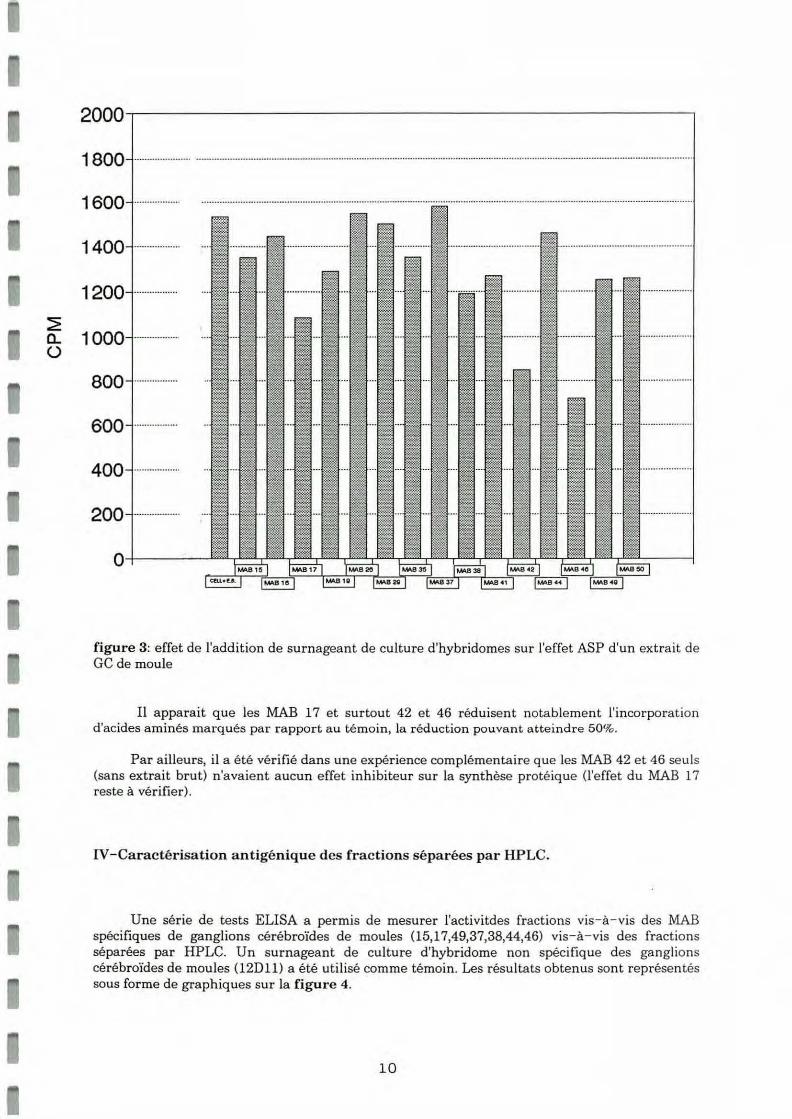
~ a.. ü
2000
1800 +····················· ·································································································································································································
1600
1400
1200
1000
800
600
400
200
0
figure 3: effet de l'addition de surnageant de culture d'hybridomes sur l'e ffe t ASP d'un extra it de GC de moule
Il apparaît que les MAB 17 et surtout 42 et 46 réduisent notablement l'incorporation d 'acides aminés marqués par rapport au témoin, la réduction pouvant atte indre 50%.
Par ailleurs, il a été vérifié dans une expérience complémentaire que les MAB 42 et 46 seuls (sans extra it brut) n 'avaient a ucun effet inhibiteur sur la synthèse protéique (l'effet du MAB 17 reste à vé rifie r).
IV-Caractérisation antigénique des fractions séparées par HPLC.
Une série de tests ELISA a permis de mesurer l'activitdes fractions vis-à-vis des MAB spécifiques de ganglions cérébroïdes de moules (15,17,49,37,38,44,46) vis-à- vis des fractions sépa rées par HPLC. Un surnagean t de culture d'hybridome non spécifique des ganglions cérébroïdes de mou les (12Dll) a été utilisé comme témoin. Les résultats obtenus sont représentés sous forme de graphiques sur la figure 4.
10
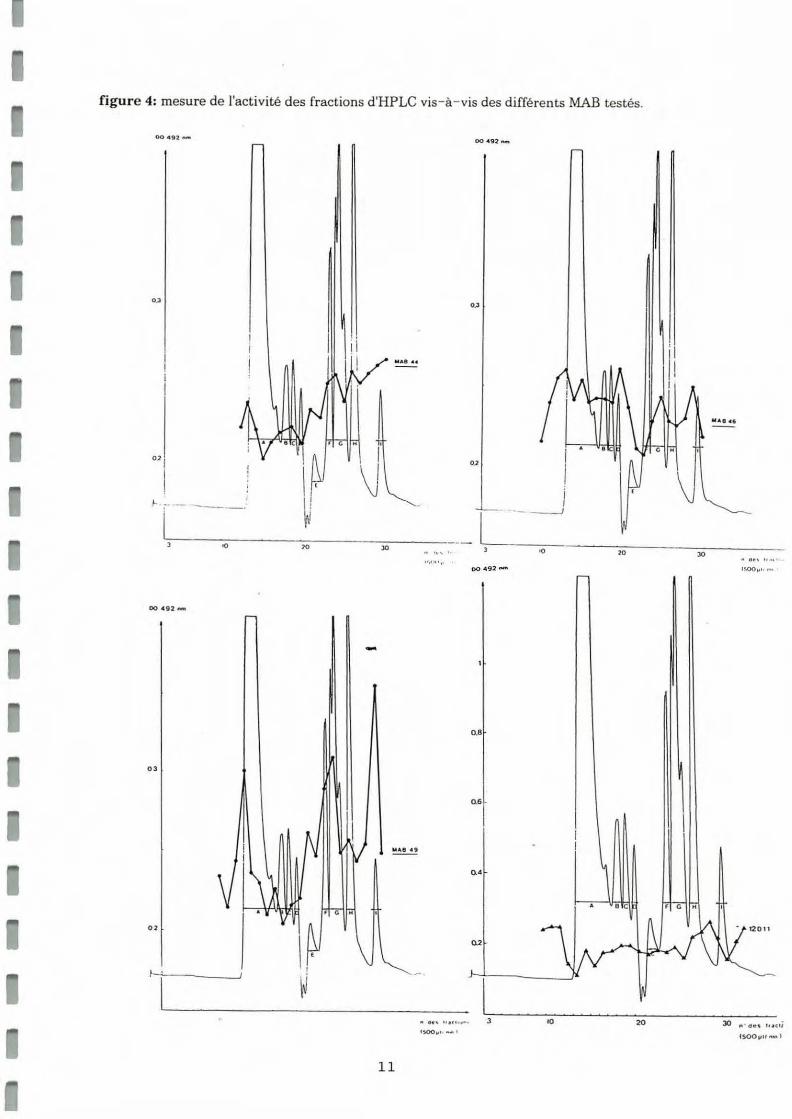
figure 4: mesure de J'activité des fractions d'HPLC vis- à - vis des différents MAB testés.
004Q2-
O,J
MA8 4C
. ( \j
}- ,-· -- ·----·
o() >O
00 492-
-
03
MAS ct
02
11
.........
•soo .... ..... .
00 4 92 .._
O,J
02
l---·--~.0~------~~~-------~---------00 4 9 2"'"'
0 .8
0 .6
0.4
Q.2
3 10 20
·~ ...... ..
30 n ' de~ h<IC1;
tsoo .., 11 """ '
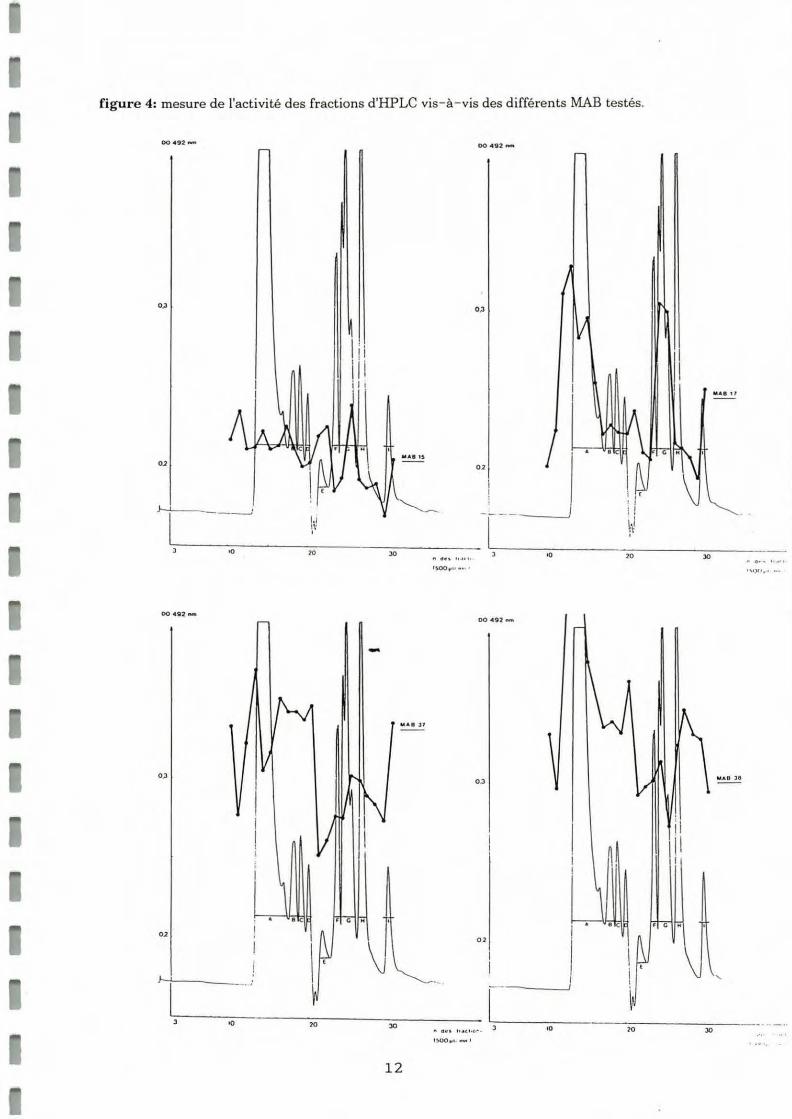
figure 4: mesure de l'activité des fractions d'HPLC vis-à- vis des différents MAB testés.
00 4 92-
O;J
02
1 ii 1 i 1 i , ,
0.3
02
Il 1
.... t7
~ ~'-;-----~~------~\~~------------------- L~--·------~------~~-------------tO 20 30 ., tO 20 JO
IS.OO.,u ...... ,
00 4V2 ,_ 00 492-
-
..... l7
O;J 0 .3
02 02
1 • f C H
~'---------· ·
~----~.0~------~20~--------~----------- IL- -·-________ __ tO
12
,. d~4 to.u.t .o··
lM>()"'·-·
.,.)(,,,, ....
MAB li
20 JO

Il apparait que:
• Le MAB 46 montre des faibles réactivités vis-à-vis d'antigènes de 7000 Da, 2500 Da el 1000 Da. Cette réactivité au niveau de la dernière fraction est à corréler avec les résultats obtenus lors de l'étude de l'effet ASP des fractions d'HPLC (pic d'activité vers 1000 Da) . Par ailleurs, il a été observé que le MAB 46 inhibe l'effet ASP de l'extrait brut (cf III).
• Le MAB 49 montre, en particulier, une réactivité très nette avec la fraction l, soit 1000 Da et moindre avec la fraction d'environ 3500 Da.
• Le MAB 44 montre un maximum de positivité au niveau de la zone l, avec une positivité moins marquée vers 3500 Da.
• Le MAB 38 montre deux zones de réactivité vis-à-vis de molécules de 7000 Da et 2000 Da.
• Le MAB 37 montre une zone de réactivité vis-à-vis d'antigènes de poids moléculaires compris entre 15000 et 7000 Da et une seconde au niveau de molécules de poids moléculaire inférieur à 1000 Da.
• Le MAB 17 réagit vis-à-vis de molécules de poids moléculaire voisin de 3500 Da ainsi que de molécules inférieures à 1000 Da.
• Le MAB 15 ne reconnait pas de molécule dans la zone des forts poids moléculaires, et montre 2 maximums de réactivité vis-à-vis de molécules d'environ 6500 Da et 2500 Da.
• Le MAB 12011 donne un signal faible et uniforme correspondant au bruit de fond .
Une autre serIe de tests ELISA a permis de mesurer l'activité des fractions vis-à-vis d'anti-insuline humaine, d'anti-MIPc de Lymnée et d'anti- somatostatine humaine (figure 4bis) .
• L'anti- MIPc montre un maximum de réactivité pour des molécules de poids moléculaires de 25000 Da, 15000 Da, 5500 Da.
• L'anti-somatostatine réagit vis-à- vis de molécules de poids moléculaires de 15000 Da et 3500 Da.
• L'anti- insu line présente un profil de réactivité simila ire à celui de l'anti - MIPc mais avec une intensité beaucoup plus faible .
13
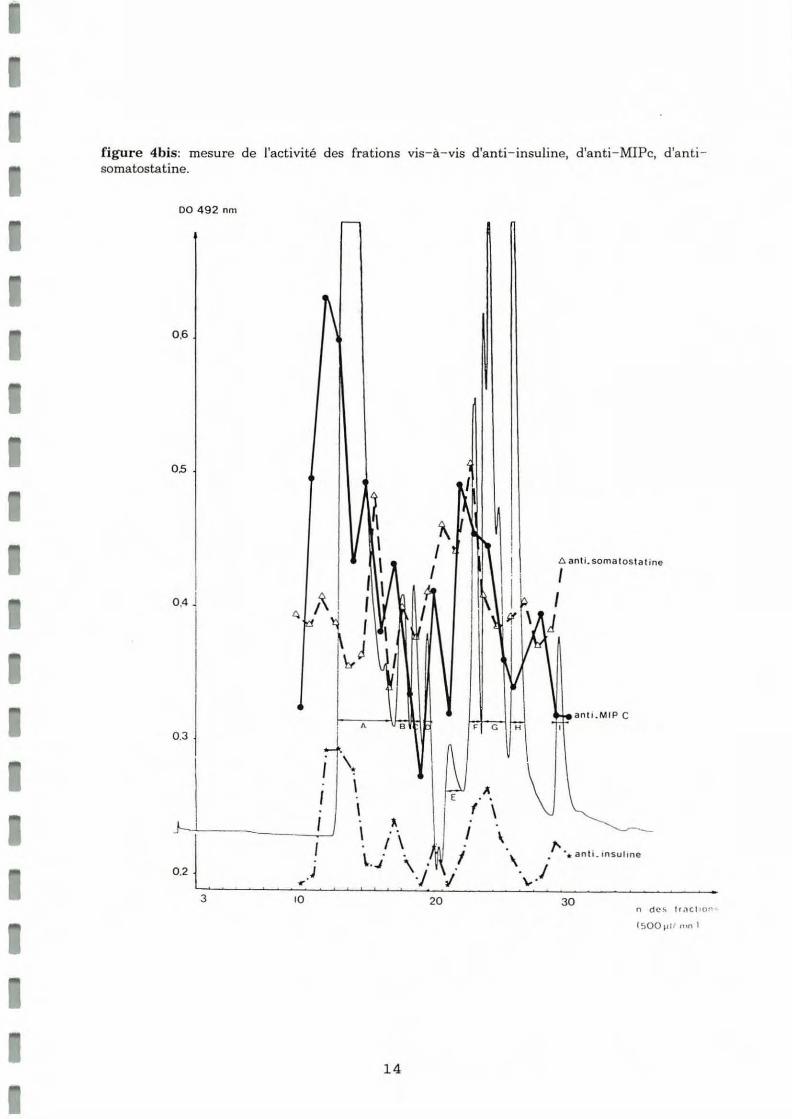
figure 4bis: mesure de l'activité des frations vis-à-vis d'ant i- insuline, d'anti- MIPc, d'antisomatostatine.
DO 4 92 nm
0 .6
0 .5
6 anli. somatostatine
1 0 .4 1
d
0 .3
\. f \ A
02 1'-::..._, .....__~__._--:"'-J_._f .............._ ........ \ __.. ~--'~'--\.__\_·/~· ~_...::.J_'_/_'_\ -'-~"~.J_·"~··_an-1 i_. •n-su-lm_: _ _ _
3 10 20 30
(S00 p l .' non l
14

DISCUSSION - CONCLUSION
Une première serle d'expériences a consisté à tester ferret activateur des synthèses protéiques des fractions obtenues après séparation d'extraits de ganglions cérébroïdes en HPLC sur colone protéine PAK 125. Le plus fort effet activateur de la synthèse protéique est obtenu avec les fractions correspondant à des molécules d'environ 1000 à 1200 Da. Ce résultat, confirmé par plusieurs essais, est tout à fait en accord avec les résultats de J.Y.Toullec (1990) qui a mis en évidence l'existence d'une hormone de croissance de Mytilus edulis, purifiée à partir d'extraits de ganglions sur Séphadex G 50 et sur Biogel P4 (acrylamide), et dont les caractéristiques sont les suivantes: un poids moléculaire de 1000 Da, sa nature peptidique, thermolabile, sensible à la trypsine, son caractère hydrophile.
Par ai lleurs, il a aussi montré que l'extrait ganglionnaire de moule stimule la croissance de cellules de coquilles Saint Jacques in vitro.
U ltérieurement, il sera intéressant de vérifier que la molécule de 1000 Da correspondant aux fractions 28, 29, 30 est bien la mème que celle mise e n évidence par J.Y.Toullec. Pour cela, la dénaturation par la chaleur, l'action de la trypsine seront testées; les caractères hormona l et hydrophile de la molécule seront recherchés. Par ailleurs, des enzymes prése ntes da ns les fractions d'HPLC d'extraits de ganglions, auraient pu entraîner une perte d'activité des molécules peptidiques. Une autre série d'expériences sera réalisée en présence d'un inhibiteur de protéase afin d'éviter toute dégradation enzymatique des molécules fractionnées (Leung, 1990).
Une seconde partie de l'étude a consisté à étudier l'effet de différents MAB sur l'activité de l'extrait brut de ganglions cérébroïdes. Ce biotest a montré que les MAB 42 et 46 bloquent jusqu'à 50% de ttincorporation d'acides aminés marqués par rapport aux essais réa lisés sans MAB. Ce résultat laisse suppose r que ces deux MAB reconnaissent une molécu le impliquée dans l'activation de la synthèse protéique. D'autres biotests seront donc réalisés dans les conditions suivantes: l'activité des fractions d 'HPLC sera testée avec ou sans les MAB 42 ou 46 et permettra de caractériser plus présicement ces molécules. Cette voie de recherche semble prometteuse dans la mesure où elle pourrait permettre d'envisager la purification pa r simple chromatographie d'affinité d'une molécule ayant un effet biologique in vitro.
Dans cette optique de purfication des antigènes spécifiques des MAB, l'immunoréactivité des fractions aprés sèparation sur une colone d'HPLC a été analysée avec les MAB (49,44, 38, 37, 15, 17) qui ont montré un bon seuil de positivité en ELISA sur les extraits bruts de ganglions l
ainsi que pour des anticorps anti - MIPc, anti-somatostatine, anti - insuline. Un signal de DO é levé pour une fraction reflétant la spécificité de l'anticorps pour l'antigène, la présence de max ima de positivité pour les MAB 17 et 49 devrait permettre de purifier par chromatographie d'affinité les molécules contenues dans l'extrait brut de ganglions. La majeure partie des MAB testés reconnaissent des molécules de haut poids moléculaires ce qui permet d'émettre 2 hypothèses:
• les MAB forment un complexe non spécifique avec des molécules de hauts poids moléculaires.
• les MAB reconnaissent de manière spécifique les précurseurs des molécules aya nt un effet biologique in vitro.
Il est intéressant de noter que la courbe réalisée avec un anticorps non spécifique d'antigène de moule 12D 11 ne présente pas de maximum de réactivité pour les molécules de haut poids moléculaires (de même, les MAB 46, 44 et 15 ne montrent pas de pics pour les molécules des premières fractions) . L'hypothèse de formation d 'un complexe non spécifique entre les MAB et des molécules agglutinantes de haut poids molécu laires ne permet donc pas d'expliquer la présence de pics en ELISA dans la zone de forts PM. Par ai lleurs, il est inté ressant de noter que
15

l'anti-MIPc (polyclonal anti- insuline- like de Lymnée), bien qu'il soit hété rologue présente un maximum de réactivité pour un pic aux e nvirons de 15000 Da qui pourrait correspond re à un précurseur de MIP (13.4 kDa). L'anti - MIPc et le MAB 17 ont des profils de réactivité proches. Le MlP a un PM de 6400 Da, mais lors du processing, il ne conserve que la chaine C et donc le MIP ne peut être reconnu par l'anticorps anti - MlPc spéci fique de la chaine C. Seul le précurseur pourra réagir avec l'anti -MIPc (Smit, 1990).
La deuxième hypothèse avancée semble donc la plus probable, il sera intéressant de tester par exemple le clivage à la trypsine des molécules contenues dans les premières fractions puis de visualiser une perte ou non de signal après un nouveau fractionnement de l'hydrolysat (Leung, 1990).
16

DEUXIEME PARTIE RECHERCHE DE GENES D'HORMONES
De nombreuses hormones poly pe pt idiques ont été identifiées, dont la majeure par tie chez les Ver tébrés. Les informations sur les séquences d'hormones peptidiques d'Invertébrés, homologues à des hormones de Verté brés sont restreintes. Nous avons choisi de travaille r sur les modèles d'insuline e t de récepteur d 'insuline qui ont été séquencés chez différents groupes d 'Invertébrés et de prolactine, séquencée uniquement chez des vertébrés:
. la structure de l'insuline, son mode d'action et la structure du gène sont bien con n us chez les Vertébrés mais ont a ussi fai t l'objet de recherches chez un cer tain nombre d'Inver tébrés tels que Bombyx morii (Kawakami et al , 1990) ou Lymnaea stagnalis (Smith et al, 1988) .
. le gène du récepteur d 'insuline, séquencé chez l'homme, la souris, le ra t, le porc, ma is aussi la drosophile, présen te une forte homologie dans la chaîne B au niveau du domaine tyrosine kinase .
. le gène de prolactine qui a été séquencé chez un grand nombre de Vertébrés (y compris le poisson ) et qui mon tre une forte homologie dans 2 régions, permet a insi la conception a isée d'une pa ire d'amorces. Le fa it que la présence de ce gène n'ait encore jamais été démon trée chez des Invertébrés, laisse une incertitude q uand à l'amplification possible de fragmen ts. Toutefois, des déterminants ant igéniques "prolacti n - like" ont été mis en evidence dans les cellules neurosécrétrices de cerveaux d'abeilles (Sch mit et al, 1990).
1 7

1
MATERIELS ET METHODES
!-Extraction d 'acides nucléiques
1-1 Extraction de l'ADN génomique de moules entières
Les moules, Mytilus edulis, originaires du Bassin de Marennes-Oléron sont maintenues en bac a u laboratoire. L'ADN génomique de moule, déjà disponible au laboratoire, a été préparé selon le protocole de Jeanpierre (1987). En bref, 3 moules sont broyées dans un milie u à base d'isothiocyanate de guanidine puis soumises à deux extractions successives à la protéinase K. Après trois extractions phénol-chloroforme, les acides nucléiques sont précipités à l'éthanol. La purification de l'ADN s'achève par un traitement enzymatique pa r la ribonucléase A (RNase, DNase free, Boehringer ). La quantité d 'ADN purifiée est évaluée par mesure de l'absorbance, le spectre de longueur d 'ondes variant entre 350 et 200 nm.
1-2 Préparation d 'ADNe d 'embryons
L'émission des gamètes est stimulée par injection de 200 .ul de KCl 0,5 M dans le muscle adducteur postérieur des moules maintenues indépendemment les unes des autres. Les ovocytes d'une moule et les spermatozoïdes de 2 moules sont ensuite mis en présence dans 750 ml d 'eau de mer sous agitation, à 16°C. La fécondation est observée au microscope après 10 mn. Les embryons sont lavés par passage sur un filtre de 100 .um. Les spermatozoïdes sont éliminés pa r filtration (45 .um). Les embryons sont rincés à l'eau de mer filtrée. L'incubation est poursuivie dans l'eau de mer à 16°C, une nuit, sous agitation. Les embryons au stade morula sont a lors récupérés par centrifugation (50 g, 5 mn).
Cette technique a utilisé le kit "SuperScript Preamplification System for first strand eDNA synthesis" (BRL): Les cellules sont lavées avec 1 ml de PBS, vortexées doucement pendan t 15 à 20 secondes, et centrifugées à 5000 g pendant 1 mn à température ambiante. Les cellules sont a lors dissociées par addition de 200 .ul d'isothiocyanate de guanidium 25 mM (GuSCN), et de mercapto-éthanol 25 mM; 5 .ug d'ARNt de levure sont ajoutés en tant que "carrie r". La dissociation est prolongée par passage de la solution dans l'aiguille fine (23 G) d'une seringue, jusqu'à ce qu'elle d evienne visqueuse. L'ARN est ensuite extrait par 240 .ul de phénol saturé en eau et 48 .ul de chloroforme/alcool isoamylique (49/1; v/v). Le mélange est vortexé rapidement et déposé sur de la glace pendant 15 mn. Après centrifugation (5000 g, 30 mn, 4°C), la phase aqueuse est transférée dans un microtube et l'ARN est précipité par 2 volumes d'éthanol absolu à -20°C pendant 60 mn puis centrifugé (5000 g, 30 mn, 4°C). Le culot est dissous dans 100 .ul de solution de GuSCN, précipité comme précédemment et repris dans 75 .ul d'eau distillée traitée a u diéthylpyrocarbonate (DEPC) et précipité à nouveau. L'ARN est réhydra té par lavage à l'éthanol 70 % (200 .uD, cen trifugé (5000 g, 10 mn, 4°C) et séché à température ambiante. Le culot est a lors redissous dans 13 .ul d'eau traitée au DEPC et conservé à - 20°C.
La transcription réverse d'ARNm en ADNe est amorcée en additionnant a ux 13 .ul obtenus ci- dessus, 1 .ul d'oligo dT. L'appariement sur la matrice polyA des ARNm a lieu à 70°C pendant 10 mn et le mélange est ensuite refroidi rapidement sur de la glace. La synthèse d' ADNe est obtenue par adjonction de transcriptase réverse "Superscript" (200 U/.ul), 0,1 M de dithiothréitol (DTT) , 10 mM de dNTP, de tampon de synthèse 10X, puis par incubation 10 mn à température ambiante suivie de 50 mn à 42°C. L'ARNm est désapparié du brin d 'ADNe par chauffage à 90°C pendant 5 mn puis est détruit en ajoutant 1 .ul de RNase H, 20 mn à 37°C.
18

I -3 Préparation d'ADNe de ganglions cérébroïdes
Vingt ganglions sont disséqués et dissociés dans un homogéné iseur de tissus de type Downce. Le protocole de préparation d'ADNe de ganglions cérébroïdes de moules est ide~tique à celui de préparation d'ADNe d 'embryons.
II Choix et synthèse des amorces
II -1 Analyse des banques de séquences nucléiques et protéiques avec système d'exploitation BISANCE
BISANCE est un système d'exploitation de banques de données (EMBL et GENBANK) où sont répertoriées les séquences déjà connues d'acides nucléiques et de protéines dont la mise à jour est faite régulièrement. Il permet la consultation et l'utilisation des banques de séquences ainsi que des données personnelles. Notamment, des programmes d'homologies (FASTA) et d 'alignement (KANEHISA) entre une ou plusieurs séquences sont accessibles et ont été utilisés pour identifier des séquences consensus. La recherche des portions en hairpin, des possibilités d'hybridation des oligonucléotides entre eux et les homologies de séquences entre les oligonucléotides et la banque, a également été effectuée .
II -2 Synthèse et purification d 'oligonucléotides
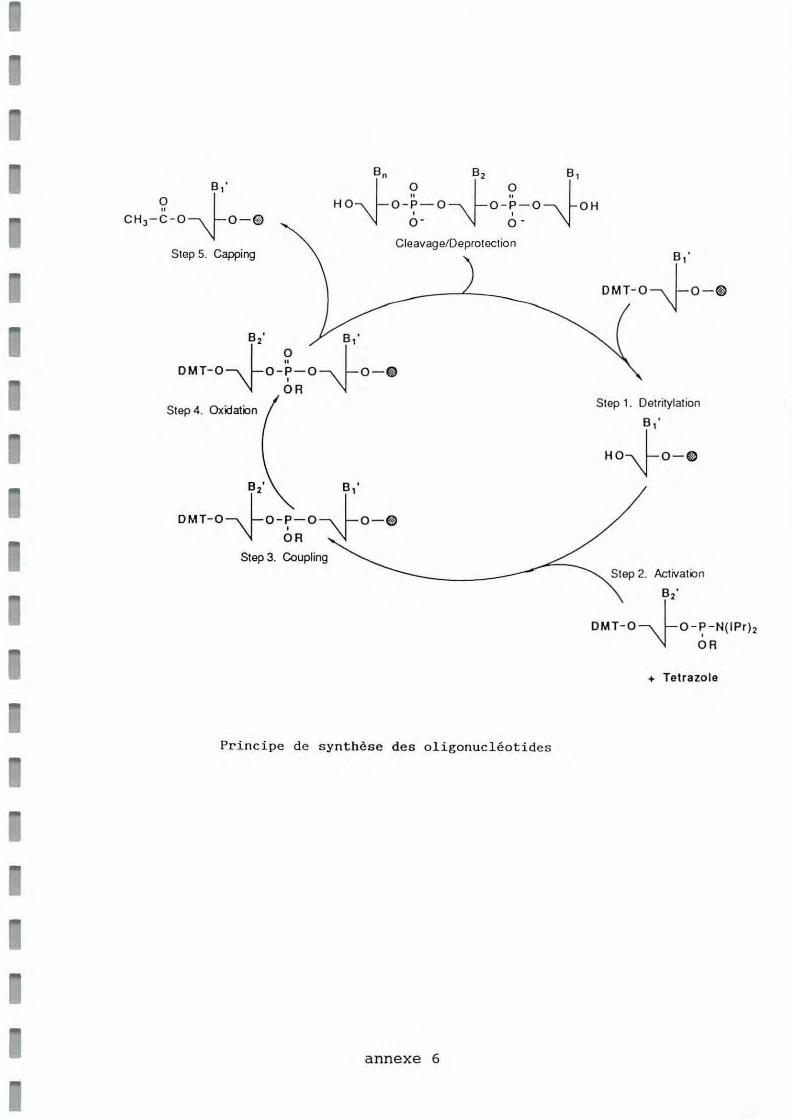
La synthèse d'ADN est un procédé cyclique (annexe 6). La première base, à l'extrémité 3', est déjà fixée sur un support solide CPG ("controlled pore glass"). Ce nucléotide est protégé en 5' par un groupement diméthoxytrityl (DMT). La première étape du cycle (étape de détritylation) va consister à enlever ce groupement protecteur par un traitement acide (acide trichloracétique). Le groupement hydroxyl en 5' est ainsi régénéré. Le nucléoside phosphoramidite correspondant à la deuxième base, après activation par le tétrazole (étape d' activation), vient réagir avec ce groupement 5' OH : une base supplémentaire est ajoutée à la chaîne oligonucléotidique croissante (étape de couplage). Enfin la nouvelle liaison internucléotidique est oxydée avec une solution d'iodine pour donner une liaison stable pentavalente (étape d'oxydation). Pour les chaînes qui n'ont pas réagi avec le nucléoside phosphoramidite, le groupement 5' OH est acétylé avec de l'anhydride acétique et du N - méthyl imidazole: il ne sera plus fonctionnel à la prochaine étape d 'addition. Cette étape de "capping" empêche la formation d'une population hétérogène de sousproduits n-1, facilitant la purification du produit final. Ces quatre étapes sont répétées successivement jusqu'à l'addition du dernier nucléotide en 5'. En fin de synthèse, l'oligonucléotide reste attaché de façon covalente à son support.
Les oligonucléotides ainsi obtenus subissent un traitement à l'ammoniaque (NH40H à 33 %) à température ambiante pendant 24 heures, qui les libère de leurs supports et enlève les derniers groupements protecteurs. L'ADN est alors biologiquement actif. (Beaucage et Caruthers, 1981; Mc Bride et Caruthers, 1983; Caruthers, 1989)
Les oligonucléotides peuvent ainsi être purifiés sur colonne Oligo- Pack (Millipore): 3 fois 5 ml d'acétonitrile sont injectés dans la colonne à l'aide d 'une seringue, puis pour compléter le processus d'équilibration, le même volume d'acétate de triéthylammonium (TEAA, 1 M, pH 7) est ajouté. Les oligonucléotides sont alors injectés doucement dans la colonne, et l'éluat récupéré est à nouveau introduit. L'éluat est ainsi injecté trois fois de manière à obtenir une fixation maximale des oligonucléotides couplés a u DMT. Afin d'éliminer les séquences trop courtes et les
19

1
groupements protecteurs, trois lavages sont effectués avec 5 ml d'hydroxyde d'ammonium (3%). Après un rinçage à l'eau bidistillée (3 fois 5 ml), un lavage avec 5 ml d'acide t r ifluoroacétique 2% (TFA) permet de détrityler la liaison de l'oligomère avec son support. Puis l'excès de TFA est enlevé par lavage à l'eau bidistillée (2 fois 5 ml). Enfin, le produit détritylé est élué à l'acétonitrile 40 % (3 fois 1 ml dans 3 tubes différents), évaporé après mesure de la DO et repris dans du TE .
III- AMPLIFICATION DE SEQUENCES PAR P.C.R. (POLYMERASE CHAIN REACTION)
Les amplifications par P.C.R. sont réalisées selon le protocole classique de Larzul (1989).
Les 50 ,ul de mélange réactionnel contiennent:
- 6 ,ul d 'ADN (1 ,ug) - 1 ,ul de chaque amorce, leur quantité est fonction de la longueur, (315 ng pour un
oligonucléotide de 25 mers) - x ,ul de MgC12 (de 1,5 à 5 mM en concentration finale) - 5 ,ul de tampon de réaction (10 X) (Tris- HCl 10 mM, pH=8,3; KCl 50 mM; MgC12 100 mM;
gélatine 0,01% (p/v)) - 1 ,ul soit 100 mM de chaque désoxynucléotide (dATP, dCTP, dGTP, dTTP) - 0,5 ,ul soit 2,5 unités d'enzyme Taq polymérase (Cetus) - eau stérile qsp 50 ,ul
Le mélange est recouver t de 3 gouttes d'huile de paraffine, puis soumis à des cycles d'amplification dans un bain thermostaté programmable (PHC II, Techne OS!).
Chaque cycle comprend:
- une dénaturation de l'ADN (incubation à 92°C, 2 mn) - une hybridation des amorces (la température d'incubation dépend de la Tm des amorces choisies, pendan t 2 mn) - une extention des deux brins d'ADN par l'ADN polymérase (incubation à 70°C, 2 mn)
Après le dernier cycle, une dernière incubation de 3 mn à 70°C permet l'élongation totale de tous les brins inachevés.
Les échantillons sont ensuite refroidis à température ambiante, l'huile de paraffine est enlevée par dissolution dans de l'étheroxyde. 8 ,ul de chaque échantillon et 2 ,ul de bleu de dépot sont déposés sur un gel d'agarose à 1 % soumis à une é lectrophorèse. Le marqueur de poids moléculaire utilisé est du DNA ladder de 1 kb (BRL) .
20

RESULTATS
1- Pré paration de l'ADN cible
1-1 Extraction de l'ADN génomique de moules
La quantité d 'ADN de moule obtenue pa r extraction à pa rtir de 3 anima ux a é té estimée sur la base de l'absorbance à 260 nm, (1 unité de DO équivaut à une concen tration d'ADN de 50 ).lglml) . La concentration de l'ADN purifié a été verifiée par le rapport des a bsorbances à 260 nm e t à 280 nm. La valeur de 1.87 confère à cette prépa ration d'ADN un caractère de pureté suffisant pour l'amplification de gènes.
1- 2 Préparation d'ADNe d'embryons e t de GC
Compte tenu de la faible quanti té de ma tériel de départ, l'estima tion de la concent ration en ADNe dans chaque tube n'a pas été réalisée; ma is la sensibilité de la réaction de PCR est telle q ue la quant ité de matériel cible n'est généra llement pas limitante.
II- Choix d es amorces
Pour les 3 gènes pris en considération (insuline- like, recepteur d'insu line et prolactine) des zones de bonne homologie ont é té iden t ifiées et des systèmes d 'amorces en ont été dédui ts. Dans le cas où le choix entre deux nucléotides semblait difficile, les deux possibilités ont été retenues. L'inosine a été utilisée dans le cas où 3 à 4 choix éta ient possibles.
Outre les homologies de séquences, le choix des amorces a été basé sur une ta ille d 'au moi ns 20 pa ires de base. La formule simplifiée suivan te peut s'appliquer pour le ca lcul de la température de de mi- dé na turatio n (Tm) dan s le cas d'a m orces de l'ordre de 20 mers.
Tm = 4 * (G + C) + 2 * (A + T)
II -1 Choix d es amorces pour le gè n e r écepteur d'insuline
Les gènes de récepte ur d'insuline séquencés chez de nombre ux Ve rtébrés mais aussi des Invertébrés de la classe des insectes comme la drosophile présentent d'importantes homologies -de l'ordre de 70% - a u niveau de la cha ine b et plus particulièrement dans le domaine fyros ine kinase. Dans l'optique d 'une recherche d 'amorces pour l'amplification par PCR, les zones de la chaine B de plusie urs espèces présentant toutes une forte homologie entre elles ont été a lignées selon le programme d 'alignement multiple de BISANCE. Le résultat de cet a lignement figure en annexe 7.
2 1

1
Pour chaque zone d'alignement, 2 régions susceptibles de fournir des amorces ont été sélectionnées en fonction:
. d'un pourcentage minimum d'homologie sur 20 à 25 nucléotides
. d'une très bonne conservation de l'extrémité 3' à partir de laquelle se fait l'élongation lors de l'amplification par PCR
. de l'absence de structures secondaires (hair-pin, Cariello; 1991)
. de l'absence d'homologie significative par rapport au reste de la banque
Deux paires d'amorces ernboitées ont ainsi été préparées (2 amorces 3', 2 amorces 5').
amorce position s équence 5 1 -+ 3 1 taille Tm en nt oc
38 5 1 GGGCAGGGATCCTTTGGIATGGT 23 76 c c T c
39 3 1 ATCTCCCAGAGIACCACICCAAAG T A
24 72
4 2 5 1 TTCCTCAAIGAAGCI CIGTCATGAA G G G G
25 74
43 3 1 ACGTCICGIGTCATCCCGAAGTC 23 72 TA T T A
Tableau 2: Caractéristiques des différentes amorces insulin- recepteur
La figure suivante (figure 5) indique les tailles entre les différentes paires d'amorces de la séquence codante de Drosophila melanogaster.
<D 0 M w w Ol w Ol ,.... N w ,... N N N N
;38 .42 43 • 39 .
630
373
497
506
Figure 5: Taille entre les différentes paires d'amorces de la séquence codante de Drosophila melanogaster
22

1
II -2 Choix des amorces pour le gène de prolactine
Aucune séquence a pparentée au gène de prolactine n'a été à l'heure actuelle étudiée chez des Invertébrés. Cependant, le gène de prolactine a été identifié chez de nombreuses espèces de Vertébrés non mammifères et pour lesquels la prolactine semble réguler d 'autres fonctions que la lactation.
Parmi les espèces pour lesquelles le gène a été séquencé, 2 zones d'homologies ont été mises en évidence annexe 8 . Une paire d'amorces a été sélectionnée dans ces zones d'homologies en fonction des mêmes critères que pour les amorces de récepteur d'insuline.
amor ce posit ion séquence 5 1 -+ 3' tai lle Tm en nt oc
44 5' ATICCIGAIGATAAGGAICAAGC 23 68 c A c A A
45 3 1 CGCAGGGATTcAAATAAGGTTGACA c c CCGC AAC
25 84
Tableau 3: Caractéristiques des amorces prolactine
La longueur de la séquence codante comprise entre les 2 amorces pour les espèces a lignées est de 350 paires de bases.
II - 3 Choix des amorces pour le gè ne d'insuline
Des gènes appar tenant à la superfamille de l'insulin- like ont été séquencés ces dernières années chez différents groupes d'Invertébrés.
* 5 gènes d 'insuline- like ont été séquencés chez la Lymnée (Smith et al., 1988). Leur expression semble être spécifique du type cellula ire et du stade de développement de l'animal,et est régulée par des stimuli externes et internes (Smith et al., 1990).
* Par ailleurs, chez les insectes, la séquence du gène de bombyxine, apparentée à l'insuline est aussi disponible (Kawakami et a l. ,1990).
*Enfin la séquence d 'insulin-like d'éponge (Sch wabe C. et al; 1989).
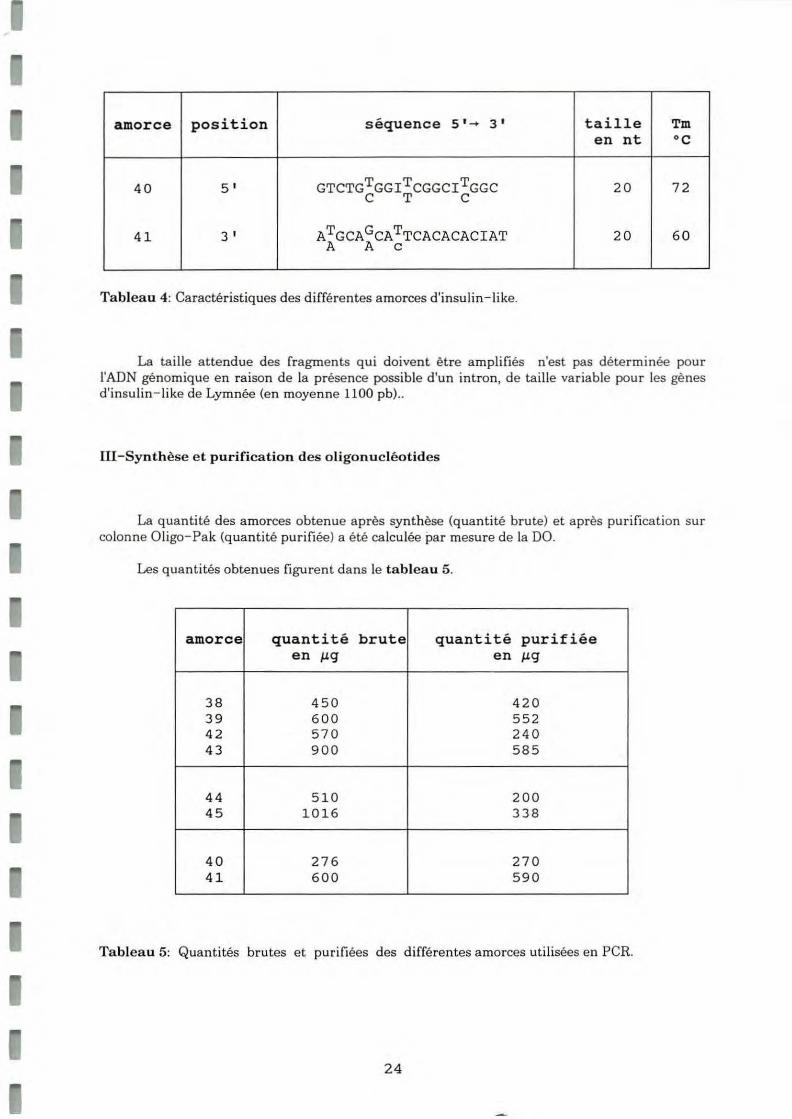
L'homologie entre toutes ces séquences est faible. Les deux zones les mieux conservées qui contiennent les résidus cystéines se trouvent a u niveau de la chaine B et de la cha ine A. Les alignements figurent en annexe 9. Dans chaque zone d'alignement une amorce a été choisie en fonction des mêmes critères que pour les amorces d'insuline récepte ur. Ces amorces figurent da ns le tableau 4.
23
1 ,
amorce position séquence s•- 3' taille Tm en nt oc
40 5' GTCTGTGGITCGGCITGGC C T C
20 72
41 3 1 ATGCAGCATTCACACACIAT 20 60 A A c
Tableau 4: Caractéristiques des différentes amorces d'insulin- like.
La taille attendue des fragments qui doivent être amplifiés n'est pas déterminée pour l'ADN génomique en raison de la présence possible d'un intron, de taille variable pour les gènes d'insulin-like de Lymnée (en moyenne 1100 pb) ..
III -Synthèse et purification des oligonucléotides
La quanti té des amorces obtenue après synthèse (quantité brute) et après purification sur colonne Oligo-Pa k (quantité purifiée ) a été calculée par mesure de la DO.
Les quantités obtenues figurent dans le tableau 5.
amorce quantité brute quantité purifiée en J.Lg en J.Lq
38 450 420 39 600 552 42 570 2 4 0 43 900 585
44 510 200 45 1016 338
40 276 270 41 600 590
Tableau 5 : Quantités brutes e t purifiées des différentes amorces utilisées en PCR.
24

~1
IV- Amplification par PCR
IV-1 Amplification de séquen ces de type ins uline r écepteu r
Une première expérience de PCR avec la paire d'amorces externes (38,39), résumée dans le tableau 6, a permis de visualiser à partir d'une matrice d'ADN génomique, une bande d'environ 750pb (figure 6).
tube ADN amorces MgCl2 température (mM) (oc)
0 0 38-39 1.5 60 1 génomique 38 - 39 1.5 60 2 génomique 38 - 39 2 60 3 génomique 38 -3 9 2.5 60 4 génomique 38 - 39 3 60
5 1 Kb ADN
Tableau 6 : Paramètres utilisés pour l'essai d'amplification avec les amorces insuline récepteur
0 1 2 3 4 5
figure 6: fragments amplifiés à partir d'ADN génomique (amorces 38-39 ;60°C)
Au cours d 'une seconde experience dont les paramétres figure dans le tableau 7, les amorces externes et internes ont été utilisées sur de l'ADN génomique. Les résul tats de cette expérience sont présentés en figure 7.
25

tube ADN amorces MgCl2 température (mM) ( oc )
0 0 38-39 2.5 55 1 génomique 38 - 39 2.5 55
2 ADNe embryon 38-39 2 . 5 55 3 ADNe ggl 38-39 2.5 55
4 génomique 42-43 1.5 55 5 génomique 42-43 2 55 6 génomique 42-43 2.5 55
7 1 kb leader
8 génomique 38-43 1.5 55 9 génomique 38-43 2 55
10 génomique 38-43 2 . 5 55
11 génomique 42-39 1. 5 55 12 génomique 42-39 2 55 13 génomique 42-39 2.5 55
Tableau 7: Paramètres utilisés pour l'essai d'amplification avec les amorces insuline récepteur.
5 10
figure 7: fragments amplifiés à partir d'ADN d'embryon, de ganglions et génomique (amorces 38-39, 42- 43, 38-43, 42- 39; 55°C) ·
26

Lors d 'un troisième essai (tableau 8), la température d'hybridation des amorces a été augmentée afin d'avoir une amplification plus spécifique. Les résultats sont présentés en figure 8 .
tube ADN amorces MgC1 2 température (mM} ( 0 c)
0 0 38- 39 2 .5 57 1 génomique 38- 39 2 . 5 57 2 génomique 38 - 43 2.5 57 3 génomique 42-39 2.5 57 4 génomique 42-4 3 2.5 57 5 1 kb leader - ---- --- --
Tableau 8 : Paramètres utilisés pour l'essai d'amplification avec les amorces insuline- recepteur.
0 1 2 3 4 5
figure 8: fragments amplifiés à partir d 'ADN génomique (amorces 38- 39, 42- 39, 38- 43, 42- 43; 57°C) .
IV-2 Amplification de séquences de type prolactin e
Une première expérience réalisée avec la paire d'amorces 44- 45 (tableau 9) a permis de visualiser à partir d 'ADN génomique une bande à 360 pb (figure 9 )
27

tube ADN amorces MgCl2 température (mM) ( oc)
0 0 44-45 2.5 53 1 génomique 44-45 1. 5 53 2 génomique 44-45 2 53 3 génomique 44 - 45 2 . 5 53 4 génomique 44-45 3 53
5 1 Kb leader
Tableau 9 : Paramètres utilisés pour l'essai d'amplification avec les amorces prolactine.
0 1 2 3 4 5
figure 9 : fragments a mplifiés à partir d'ADN génomique (amorces 44-45; 53°C)
IV-3 Amplification d e séquen ces d'insuline
Dans un premier temps l'amplification à partir d'ADN génomique a été réalisée à 47°C. L'observation d'un smear dans tous les puits du gel nous a conduit à a mé liorer la spécificité en augmentant la tempé rature d'hybridation des a morces à 60°C (tableau lO).Les résultats sont présentés en figure 10 (47°C) et 11 (60°C).
28
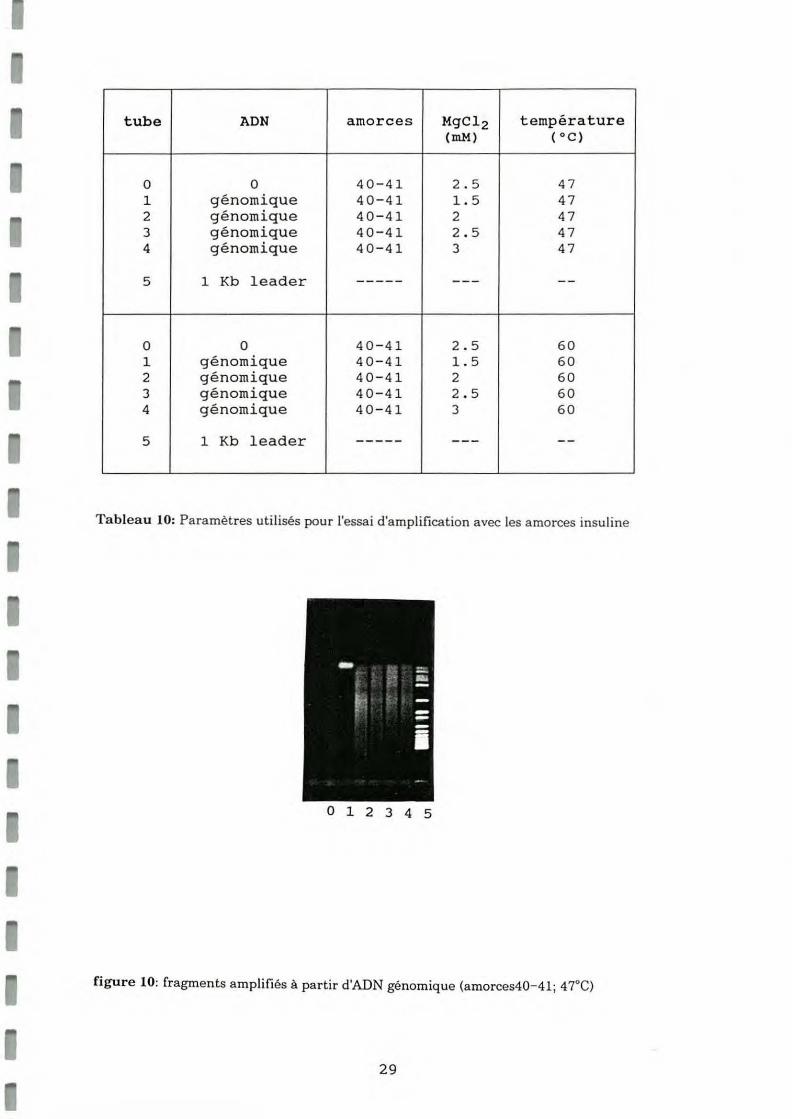
tube ADN amorces MgCl2 température (mM) (oc)
0 0 40-41 2 . 5 47 1 génomique 40-41 1.5 47 2 génomique 40-41 2 47 3 génomique 40-41 2.5 47 4 génomique 40-41 3 47
5 1 Kb leader ----- --- --
0 0 40-41 2 .5 60 1 génomique 40-41 1.5 60 2 génomique 40-41 2 60 3 génomique 40-41 2.5 60 4 génomique 40-41 3 60
5 1 Kb leader ----- --- --
Tableau 10: Paramètres utilisés pour l'essai d 'amplification avec les amorces insuline
0 1 2 3 4 5
figure 10: fragments amplifiés à partir d'ADN génomique (amorces40-41; 4rC)
29

501234
figure 11: fragments amplifiés à partir d'ADN génomique (amorces 40- 41; 60' C)
Tous les essais réalisés avec de l'ADNe de ganglions ou d'embryons comme matrice et les différentes paires d'amorces n'ont pas permis de visualiser les bandes d'interêL
30

DISCUSSION ET CONCLUSION
La stratégie retenue dans ce trava il a consisté à exploiter les homologies de séquences en t re les gènes d'une même famille caractér isés chez des espêces phylogénétiquement é loignées. Le trava il thêorique d 'alignement de sêquences a permis de déterminer des zones relativement conservées et de synthé tiser des amorces avec peu de dégénérescences. Dans le cas où 3 ou 4 possibilités devaient ê tre retenues, l'ut il isation d'inosine a été pré férêe à celle d'un mélange des 4 nucléotides pour e ffectuer la synthèse d 'amorces.
La taille des amorces est généralement de l'ordre de 20 nucléotides, ce qui correspond à une proba bilité d'appa riement complet sur une ma trice éga le à 10- 12. La spécificité théorique des amorces a été pa r a illeurs vérifiée par recherche d'homologies avec l'ensemble de la banque de sêquences nucléiques. En pratique, les amorces sont susceptibles de s'appa rier plus ou mOInS spécifiquement en fonction de leur température de demi- dénaturation (Tm) et de la concent ration en MgC12'
Les amorces nuclêotidiques syn thètisées suite a ux a lignements mult iples ont é té uti lisées pour rechercher les génes d'insuline, de prolactine et de récepteur d'insuline à parti r d 'ADN génômique de moule dans un premier temps.
La premlere série d'expériences réalisées avec les amorces 38 et 39, visant à mettre en évidence un gêne de récepteur d'insuline) a permis de visualiser une bande d'environ 750 bp. La taille a ttendue d'après les a lignements de sêq uences (cf annexe ) est de 630 bp. Afin d'explique r cette différence de tai lle, 3 hypothèses peu vent être avancées:
• Il est possible que la bande amplifiêe ne corresponde pas a u gène du récepteur d' insuline ma is soit due à une amplification non spécifique. Toutefois, les essais d 'amplification ont été réalisés à 60°c, température é levée qui implique une hybridation très spécifiq ue des amorces.
• La présence d'un intron dans ce gène pourra it explique r que la taille obtenue soit supér ieure à 630 bp.
• Les a lignemen ts étant effectués sur des espéces é loignêes des mollusques bivalves, il est possible que des dive rgences existent au niveau des gènes.
La seconde série d'expériences a été réalisée à 55°C avec les amorces externes et inte rnes. La bande d'environ 700 bp est retrouvée avec les amorces externes. Avec les amorces internes puis e n croisant les amorces, des smears contenant des bandes sont vis ibles. La te mpé rature d'hybridation a donc été augmentée à 57°C et les bandes a ppa raissen t plus nettement sur le gel. Ceci a tendance a confirmer la s ignification rêelle des ampli ficat ions. Cependant, un clonage et un sêq uençage seront nécesaires a fin de con fi rmer la nature d u gène amplifié.
En ce q ui concerne les amplifications réalisées avec les amorces prolactine une seule bande d 'environ 360 pb a été mise en évidence, les essais ayant été réa lisés dans des conditions re la tivement drastiques. Cette taille semble fa ible compte-tenu de la présence d'int rons dans les gènes de Vertébrés q ui ont servi de référence pour la prépa ra tion d'amorces. Mais l'é loignement phylogénétique ent re les espéces de référence et la moule est telle q ue l'on peut supposer q ue les structures des gènes sont différentes ainsi que la fonction des protéines pou r lesquelles ils codent. Là aussi, il sera nécessaire de cloner e t séquencer le fragment amplifié a fin d'avo ir une information précise sur sa nature.
3 1

Une troisième étude concerne l'amplification de fragments de type insuline. Smit et al a mis en évidence plusieurs gènes d'insulin- like chez la Lymnée, montrant une divergence assez importante tant d'un point de vue de la taille que d'un point de vue de la séquence nucléotidique, ce qui expliquerait la présence de plusieurs bandes après é lectrophorèse. Par a illeu rs, la difficulté d'alignements e ntre les espèces a conduit à inclure dans les amorces une proportion plus importante d'inosine pouvant entrainer des phénomènes d'amplification non spécifique. Le séquençage des fragments permettra de confirmer leur nature.
Tous les essais d'amplification réalisés à partir d'ADNe de ganglions ou d'embryons n'ont pas permis de visualiser de bandes. Il sera intéressant de réiterer les expériences afin de s'assurer de la qualité des ADNe.
Compte tenu de la relative facilité de réalisation de la PCR, celle - ci reste intéressante en dé pit de son côté aléatoire. Cette stratégie prospective a été déjà et devient de plus en plus souvent pratiquée que ce soit en endocrinologie (Sei no et al, 1990) ou dans d'autres domaines (Frye et al, 1989).
32

.~
BIBLIOGRAPHIE
ADACHI, N.F., S. TAKlYA, Y. SUZUKl, M. IWAMI, A KAWAKAMI, S.Y. TAKAHASHI, H. ISHIZAKl, H. NAGAZAWA AND A SUZUKl, 1989. cDNA structure and expression of bombyxin, an insulin-like brain secretory peptideof the silkmoth Bombyx mari. J . BIOL. CHEM., 264: 7681 -7685.
BUBEL, A., 1973. An electron microscope study of periostracum formation In sorne marIne bivalves. 1: The origin of the periostracum. Mar. Bio\., 20: 213-221.
BUCHER, I.J., P. LUBET, 1980. Etude de l'activité neurosécrétrice au cours du cycle sexuel de la moule (Mytilus edulis), mollusque lamellibranche. Bull. Soc. Zool. Fr., 105: 141-145.
CARIELLO, N.F., W.G. THILLY, J .A. SWENBERG, T.R. SKOPEK, 1991. Deletion mutagenesis du ring polymerase chain reaction: dependance on DNA polymerase. Gene, 99: 105- 108.
CARUTHERS, M.H., 1989. Synthesis of oligonucleotides and oligonucleotides analogues. In oligodesoxynucleotides. Ed. by Cohen J . S.: 7-11.
DE LOOF, A., L. SCHOOFS, 1990. Homologies between the amino acid sequences of sorne vertebrate peptidic hormones and peptides isolated from invertebrate sources. Comp. Biochem. Physiol., 95 B (3): 459- 468.
FRYE, R.A., C.C. BENZ, E . LIU, 1989. Detection of amplified oncogenes by differential polymerase chain reaction. Oncogene, 4: 1153- 1157.
GABE, M., Particularités histologiques des cellules neurosécrétrices chez quelques lamellibranches. C. R. Acad . Sei ., Paris, 240: 1810- 1812.
JEANPIERRE, M., 1987. A rapid method for the purification of DNA from blood. Nuc\. Ac. Res., 22:9611.
JHOTI, H., AN. Mc LEOD, T.L. BLUNDELL, H. ISHIZAKl, H . NAGASAWA, A. SUZUKl, 1987. Prothoracicotropic hormone has an insulin- like tertiary structure. Febs. Letters, 219: 419-425
KAWAKAMI, A., H. KATOOKA, T. OKA, A MIZOGUCHI, M. KlMURA- KAWAKAMI, T . ADACHI, M. IWAMI, H. NAGASAWA, A. SUZUKl, H . ISHIZAKl, 1990. Molecular cloning of the Bombyx mari prothoracicotropic hormone. Sciences, 247: 1333- 1335.
LARZUL, D., 1989. Une évolution dans le diagnostic: l'amplification enzymatique Ln vitro. Biofutur (janvier) : 45-49.
LENOIR, F. and M. MATHIEU, 1986. Utilisation de cultures de cellules dissociées dans l'étude du contrôle exercé sur la gamétogénèse chez la moule, Mytilus edulis (L.J. C. R. Acad. Sei., Paris,

303:523-528.
LEUNG, M., 1990. Opioïd biochemistry of Mytilus edulis. In Neurobiology of Mytilus edulis. Ed. by Stefano: 148-163.
LUBET, P., 1959. Recherches sur le cycle sexuel et J'emission des gamètes chez Izes mytilidés et les pectinidés. Rev. Trav. ISTPM, Paris, 23 (4) : 396-545.
LUBET, P. and J . ILLANES, 1980. Etude de J'activité neurosécrétrice au cours du cycle sexuel annuel de la moule Mytilus edulis. Bull. Soc. Zoo!. Fr. , 105 (1) : 141-145.
MATHIEU, M., 1987. Etude expérimentale des controles exercés par les ganglions nerveux sur la gamétogénèse et les processus métaboliquesassociés chez la moule Mytilus edulis (mollusque lamellibranche). Thèse de doctorat d'état de J'université de CAEN.
RANDALL, K.S., 1990. Amplification of DNA. in PCR protocols, a guide to methods and applications. Ed. by Berger S. L., Kimmel A. R : 13- 14.
ROSSIGNOL, J.M., 1989. Principe de la technique PCR Propriétés biologiques des ADN polymérases thermorésistantes et leur utilisation en PCR In colloque "culture in vitro" des gènes par PCR Applications aux domaines vétérinaires. Zoopole 14-15 déc. 1989.
SCHMIT, K.P., V. MAYER, C. HAUG, E.F. PFEIFFER, 1990. Ultrastructural loca lisation of prolactin- like antigenic determinants in neurosecretory cells of the brain of the honeybee (Apis mellifica). Horm. Meta. Res. 22: 413- 417
SEINO, S., M. SEINO, G.!. BELL, 1990. Human insulin recepteur gene. Partial sequence and amplification of exon by polymerase chain reaction. Diabetes, 39 (1): 123-128.
SMIT, A.B., 1990. The organization, neuronal expression and evolution of a family of insu lin related genes in the mollusc Lymnaea stagnalis. Thèse de doctorat de J'Un iversité Vrije d 'Amsterdam.
SMIT, A.B., RH.M. VREUGDENHIL, W.P .M. GERAERTS, J. KLOOTWIJK, J . JOOSSE, 1988. Growth-controlling molluscan neurons produce the precursor of an insu lin - related peptide. Nature, 331: 535- 538.
THORPE, A. and H. DUVE, 1984. Insulin and glucagon- like peptides in insects and molluscs. Molec. Physiology, 5: 235-260.
TOULLEC, J.Y., 1989. Recherches de substances hormonales actives sur la croissance d'invertébrés. Thèse de doctorat en sciences naturelles) université de Paris VI .
VREUGDENHIL, E ., J .F . JACKSON, T . BOUWMEESTER, A. SMIT, J. VAN MINNEN, H. VAN HEERIKHUIZEN, J. KLOOTWIJK, J . JOOSSE, 1988. Isolation, characterization and

1
evolutionary aspects of a eDNA clone encoding multiple peptides involved in the stereotyped egg laying be havior of a freshwater sna il Lymnea stagnalis. J. of Neurosc., 8 : 4184-4191.
WIGGLESWORTH, V.B., 1970. Insect h ormones. Edinburg: olivier and boyd. Univ. Revs. m Biol., 12

MAB spécifiques de ganglions cérébroïdes de moules
Géné rale ment, la prépa ration d'a nticorps monoclonaux spécifiques de neurohormones ou de cellules neurosécrétrices nIa été envisagée chez les Vertébrés qu'après purification des substances immunogéniques. Une a lternative à cette stratégie classique mais particulièrement lourde à mettre en oeuvre, a été récemment développée sur quelques modèles d 'insectes ou de gastéropodes et consiste à immuniser les souris avec des homogénéisats de tissus produisant les neurosécrétions, puis à immortaliser les lymphocytes producteurs d'anticorps spécifiques. Compte te nu du fait que, chez la moule, la plupart des cellules sécré trices sont localisées dans les ganglions cérébroïdes (Lubet et IlIanes, 1979), des anticorps monoclonaux ont été produits selon la technique d'hybridation lymphocytaire, contre des homogénéisats de ganglions e ntie rs (cf schéma) . Grâce à ces réactifs, une approche de cartographie des ganglions a é té réalisée selon des techniques immunohistochimiques:
I-lmmunohistologie sur frottis de cellules fixées
La caractérisation sur frottis de cellules ganglionnaires préalablement dissociées et fixées pa r cytocentrifugation sur des lames histologiques, permet d 'analyser la réactivité des anticorps vis-à-vis de l'ensemble des populations de cellules ganglionnaires, quelques soien t le ur localisation et leur représentation dans les ganglions. Da ns cette première phase de caractérisation, 32 anticorps, disponibles e n surnageant de culture, sur les 55 sélectionnés ont été étudiés et répartis en 5 catégories: 10 sont spécifiques de granula tions cytoplas miques da ns de ra res cellu les « 5%), 15 reconnaissent des gra nulations cytoplasmiques présentes dans une plus grande proportion de cellules; 4 sont spécifiques d'épitopes membranaires et pa rmi eux, 2 reconna issent une forte population de cellules; enfin 3 MAB ont montré des images t rès caracté ristiques de trajets axoniques.
2-Immunohistologie sur coupe de ganglions en paraffine
La cartographie anatomo-antigénique des ganglions a été e ntreprise en localisant les épitopes sur des coupes de ganglions inclus e n paraffine et a permis, pour plusie urs a ntico rps, de mettre en evidence des îlots cellula ires de méme spécificité.
3-Immunoenzymologie (ELISA)
Certains anticorps réagissant avec des granulatons dans des cellules de type neurosécréteur de ganglions sont susceptibles d'être spéci fiques de molécules synthétisées par ces cellules et sécrétées dans l'hémolymphe. Dans le but de détecter dans l'hémolymphe la présence de te lles molécules qui pourraient étre assimilables à des hormones ou à des cytokines, un test immu noenzymatique a é té mis au point sur des extraits ganglionna ires, puis a été testé su r la fraction sérique de l'hé molymphe (filtrée à O.22,im ).
La réactivité de ces anticorps positifs sur hé molymphe a été e nsuite precisee vis- à - vis d 'hémocytes selon un test d'immunoOuorescence indirecte pratiquée soit sur des hémocytes fixés et cytocentrifugés, soit sur des primocultures hémocytaires. Il s'avère que certains a nticorps monoclonaux positifs sur sérum filtré, ne donnent aucun signal sur hémocytes, alors que d 'autres réagissent également avec certaines populations d'hémocytes.
annexe 1

Immunis ation : injec tion intrapéritone ale des s u s pens ions de cellules ganglionnaires ( 5 00 ùl équiva lent 20 ganglions ).
Fusion pa r tra iteme n t c h i mi que (P.E . C.) .
S~ J ec t ion des hybr1domes par mi c r ocul t u rc e n mi li e u ~~ l er. Lif (H . A. T . )
Sél ec tion des hybr i d o mes séc r~ teurs
d'anticor ps sp~ci f i ques par i mmuno . fluo rescence i nd irec t e s ur fro tt is de cellul e s g a ng li o nna i res ( ) é qu iva l e nts gang l ions d i ssocié~ ~t cy tocentr i fug~ ~ ) .
Cl o nage des hybridomes pa r di l ut i o n l i mi te ( 0 . 3 hybr idome pa r pui t ) (cul ture in vitr·o l .
Prod u c t i o n de~ ant i cor~s mo noc l o nau• en a sc ite s ur souris ( c u ltu r e i n v i vo ) .
Cryo prés erva tio n en azo t e liquide .
lno cu l.at. Jon -v~loa< M .C . P . R . ~ .
rate 1)011 l~~cs lt.s• c cllu l <S
t" - ""0C•I ( .. . .. o ... • • • t f ..... . . . ... .. ... . .. ...... , ................. .
.... . • 1 "' . . . . . . .. . 1 1 ... . '~ • .. , ... ,
.. •• • ·oO- f • • ~ ,. '. • ... • ... .... "1
. .... '"' . ........ • l' . 0 • •
--- -·---- -------------- -- -··--r--·----- - -------- - ---- ---
1 100 1 ------- - -----------------
• OOO •• OO • O • O OOO • OO • O • O • O ••• OO • O • OOO OOOO • O • OO ••• 0 • • 0 • 0 0 • C 0 • G j 0 <> 00 • 0 • 00 • 0 OO • OO • O • OO • • <> O • O O • OO • O
Sché ma du p rotocol e d e p roduc t ion d ' anticorps mo n oc l o na ux s pé ci f i ques de ce llules gan g l i o nna i r e s d e moules.
1
MAB no IIF sur IIP coupe frottis paraffine
1 cyto (<5% ) 5 cyto (>5%) 7 cyto (<5% ) 9 cyto (<5% )
15 cyto (>5%) 16 cyto (<5%) 17 axones 18 rnernb (>5%) 19 cyto ( >5%) 20 cyto ( <5%) 21 axones 24 rnernb ( <5%) 26 cyto ( >5 %) 29 cyto ( >5%) 30 cyto ( >5 %) 31 rnernb ( >5% ) 35 axones 36 cyto ( >5 %) 37 cyto (>5% ) 38 cyto ( >5% ) 39 cyto ( <5% ) 40 cyto ( <5% ) 41 cyto ( >5% ) 42 cyto ( >5% ) 44 cyto ( >5% ) 45 cyto ( >5% ) 46 cyto ( >5% ) 47 cyto ( <5% ) 49 cyto ( <5% ) 50 cyto ( <5% ) 57 rnernb ( <5% ) 60 cyto ( >5% )
cyto: épitope cytoplasmique memb: épitope membranaire axones: épitope axenique
----+ + + -+ ---+ + --+ -+ + --+ + + -+ -+ + --
ELISA ggl ELISA hl IIF he
+ -+ -- -+ -+ -+ + -+ + -- -+ + + + -+ -+ -+ + -+ -+ - + + -+ + -- - + + + -+ + -+ -+ -+ + -+ -+ + (100%) + + -+ - -+ - -+ + + + + + + - -+ - -
Tableau recapitulatif des résultats de caractérisation des MAB
spécifiques de ganglions cérébroïdes de moules

1
Principe de la technique PCR
La technique d'amplification enzymatique par PCR (Larzul, 1989) est fondée sur le fonctionnement cyclique d'une ADN polymérase qui synthétise simultanement deux brins complémentaires par extension à partir de de ux amorces hybridées aux extrémités opposées de l'ADN cible. La synthèse se fait unidirectionnellement à partir de l'extrémité 3'0H libre de chaque amorce, la localisation de celles-ci délimitant la région à amplifier.
Chaque cycle d 'amplification est composé de trois étapes, permettant de réaliser successivement la dénaturation de l'ADN (95°C), l'hybridation des amorces (45 à 60°C) et l'extension des brins d'ADN (72°C) (cf schéma). Pour éviter l'addition d 'enzyme à chaque cycle, une ADN polymérase thermorésistante est utilisée: la "Taq ADN polymérase". En théorie, après n cycles d'amplification, à partir de chaque molécule cible originelle, 2n copies néosynthé tisées sont obtenues.
(tope oenoh .. rat•on
d e I'AON
- foq polymérose - Amorces {ollgonucl•olides ) - Tampon - Nucleolides _ d CTP
- d ATP - d TTP - d CTP
- AON ë amplif ier
ltope rt'tbr •ciot•on
des amorces
annexe 2
ftcpe polymer•sot c,r
de , .• 0 ,.,_

PRINCIPE DE 3 CYCLES SUCCESSIFS D'AMPLIFICATION ENZYMATIQUE (P.C.R.) .
1 CYCLE N" 1 (T AP( ><· 1 [ T AP( H' 2 (TAP( H· J
J - -À À L S tavc,.cc- c•Oic - - ~·"""""''""·"
:Jt"C\v'OI• O"" (i•Ot•Of'\ 0CJ O"""'"CC)
[ r :oo• • ~,.. ; .-c • • : ...-.c •"·c • • •~'" .... ,...,.-o1•-•
-
l -- "'== ~·· · __ c ..... :-......... z_.
r , , CI · :J'"' O~l U""O'C.Cl
1 CYCLE .-: 3
---.. aa;; - -..._ - b
= -. 0 - -9--- --L_ ={ -. - -t:=:
,.__
l b
"""""'1. W1l81\ - -b ~
J 6,..olo., .. o t.o- c,•Ot•O" o•~ ()n"\Qtc c: J Potr"•' oloon d e I"AON
" Huscte antérieur ~étractcur du byssus
G Ganglions cér~broidcs
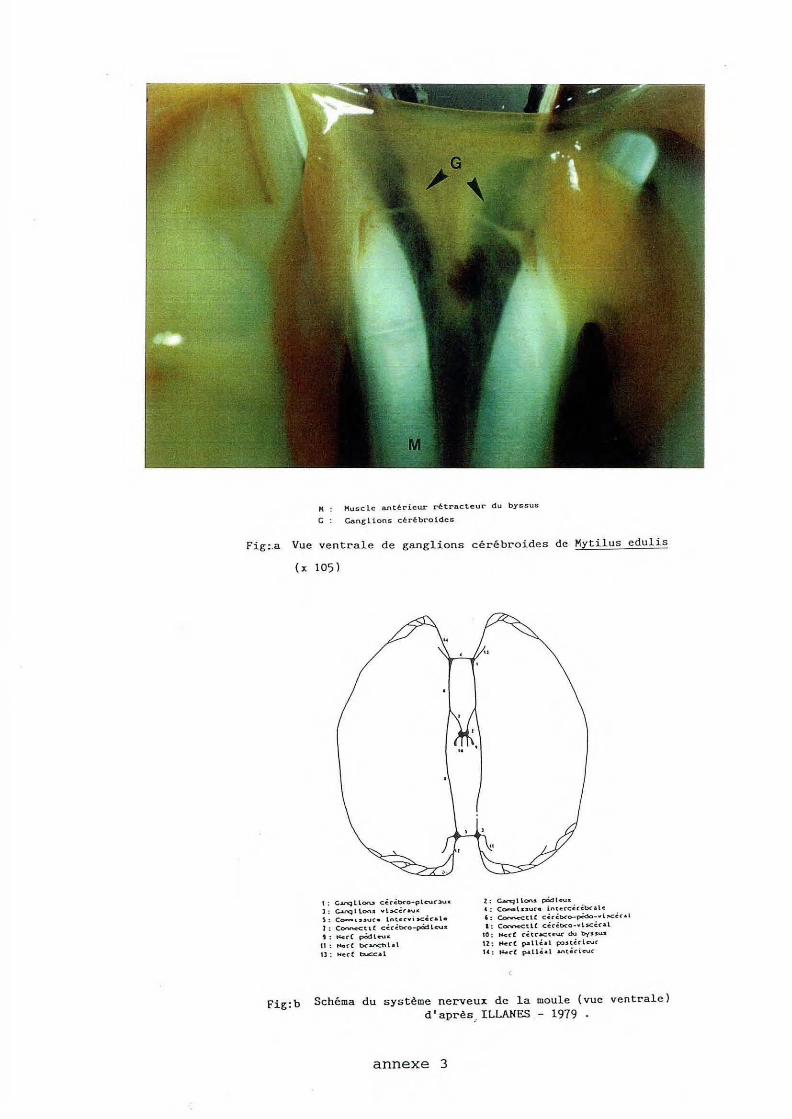
F'ig:.a Vue ventrale de ganglions cérébroides de Kytilus edulis
(x 105)
1 : G.u-qllOtU cécébeo- plc-uc-::aua l : C,.,.i"'Q l \.ocu v b.cér . ... u r:
i : Co-- \ ssuc• lntcCYÎ sc.ic.a l• 1: Conn<"Ctt l c.ëcC.b<"o-péd lc-u• t: t4e:r( pédlc-u.c. u: fo4.e(( b<'.lol"oC't''l.al 11: ~cc b.JC:c.al
2 : c...rq l lo~u pëd l eu c 4 : C~lssuC• intcrcérébc.llc ' ; ~tU: céréb<'o-pédo•"l !'Cér• l 1: COC"V"M:Ct\.C cêcé.bCo -v l .scC.c.al
'0 : H<cC cétt".IC'tC'UC ~ byssus 12: r-t-recC pallé..al p::utérlc:vc U: t~-oec( p.&ll•&l antécieuc
Fig: b Schéma du s ystème nerveux d e la moule (vue ventrale) d'après, I LLANES - 1979 .
anne xe 3
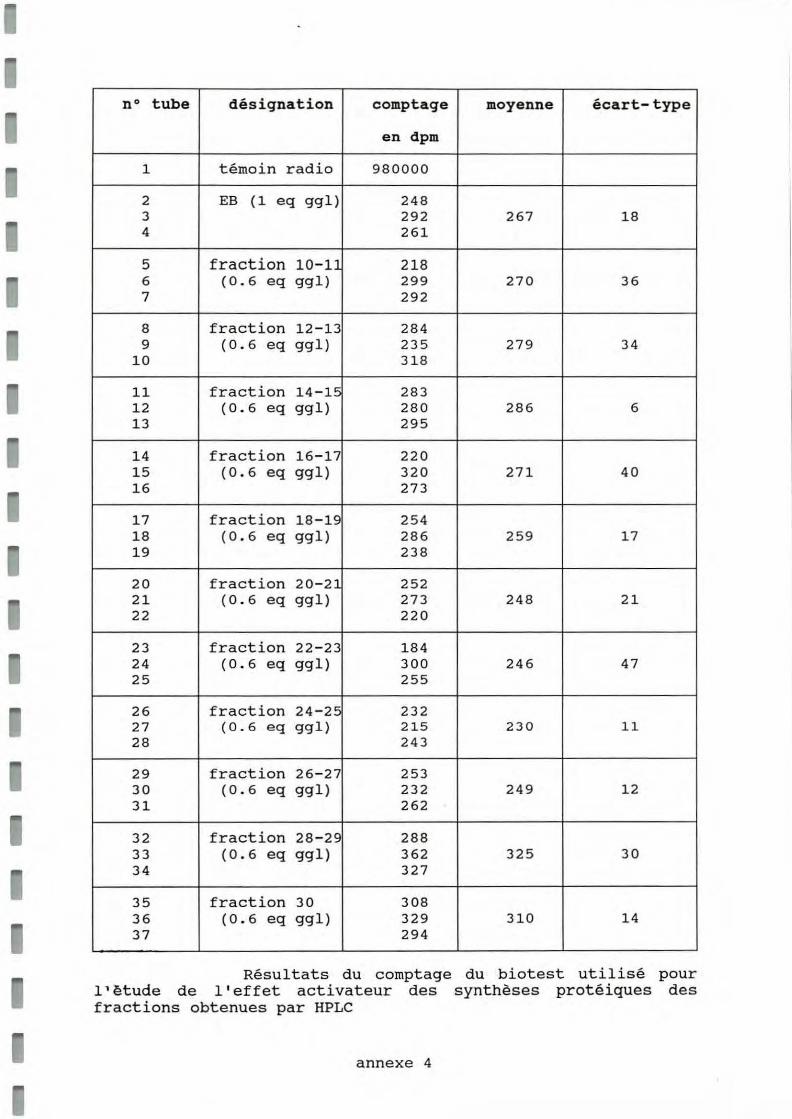
no tube désignation comptage moyenne écart-type
en dpm
1 témoin radio 980000
2 EB (1 eq ggl) 248 3 292 267 18 4 261
5 fraction 10-11 218 6 (0.6 eq ggl) 299 270 36 7 292
8 fraction 12-13 284 9 (0.6 eq ggl) 235 279 34
10 318
11 fraction 14-15 283 12 (0.6 eq ggl) 280 286 6 13 295
14 fraction 16-17 220 15 (0 . 6 eq ggl) 320 271 40 16 273
17 fraction 18-19 254 18 (0.6 eq ggl) 286 259 17 19 238
20 fraction 20-21 252 21 (0.6 eq ggl) 273 248 21 22 220
23 fraction 22 - 23 184 24 (0.6 eq ggl) 300 246 47 25 255
26 fraction 24-25 232 27 (0.6 eq ggl) 215 230 11 28 243
29 fraction 26-27 253 30 (0.6 eq ggl) 232 249 12 31 262
32 fraction 28-29 288 33 (0 . 6 eq ggl) 362 325 30 34 327
35 fraction 30 308 36 (0.6 eq ggl) 329 310 14 37 294
Résultats du comptage du biotest utilisé pour l 1 ètude de l ' effet activateur des synthèses protéiques des fractions obtenues par HPLC
annexe 4
-1.0 ...... PJ -~ (D\
n • tube d és ignation c omptage moyenne écart-t y pe en c pm
n • tube d ésigna t i on compta ge mo yenne é cart -type en cpm
I.Ort t-~e l temoin radio 262500 43 EB + MAB 41 1483 ...... o. 0 CD ~ mo.
CD () (D\ ...... li -(D\ (D O'Hl li Hl li 0 CD CD' 1-': rt (/) o. CD O. e Ul(D~ p,.UlPJ
ro~ijf ~Olo..
PJ e Ul e t-~e
~ CD li 0 ~ 0 CD ...., a x -'0 CD PJrt U1 () PJ
rti.O ..... co < ...... o. rte (D\
2 EB 1649 3 14 32 4 1651 153 4 103 . 4 5 1 407 6 1532
7 EB + MAB 15 1465 8 1349 1350 101.7 9 1189
10 1397
11 EB + MAB 16 1566 12 14 52 1447 85.9 1 3 1449 14 1323
15 EB + MAB 17 1040 16 1092 1086 46.9 17 1160 18 1052
19 EB + MAB 19 1364 20 1288 1295 47.9 21 1301 22 1229
23 EB + MAB 26 1616 24 1459 1548 56.8 25 1557 26 1562
44 1170 1275 123.8 45 1253 46 1194
47 EB + MAB 42 886 48 838 849 86.7 49 956 50 7 1 8
51 EB + MAB 44 1338 52 1166 1460 249 53 1495 54 1844
55 EB + MAB 46 784 56 7 4 5 720 46 . 9 57 69 1 58 663
59 EB + MAB 49 1180 60 1277 1253 63.3 61 12 1 0 62 1344
63 EB + MAB 50 1249 64 13 37 1262 50.4 65 1267 66 1196
t:r o. 1-'· CD 0
27 EB + MAB 29 1599 28 1548 1501 1 07.8
t-'rt CD -m
CDrt x
29 1319 30 1541
31 EB + MAB 35 1483
rte ~rt ...... 1-'· rtt->
1-'·
32 1397 1351 100.7 33 1211 3 4 1313
35 EB + MAB 37 1562
~~\ 36 1679 1579 59.5 37 1522
e 38 1554 rt'O
0 o.e CD li
39 EB + MAB 38 1332 40 1059 1190 136.1 4 1 1321 42 1050
1
If ..
8n O ~82 O ~81 HO 0-~-0 0 - ~-0 OH
o- o -
Step 5. Capping
Step 3. Coupling
Principe de s ynthèse d e s oligonucléotides
annexe 6
8,·
OMT-0~0-®
Step 1. Detritylation
s,·
HO~O-®
+ Tetrazole

1
CPIRRA
PMINSLR
HSIRRA
HSIRPR
MMINSR
M29014
[ IFREHER .OATCit)INSREC I.AU< RECEPTEUR D'INSULINE
DHINSLR HS IRPR HHIHSR H 29014 CPI RRA HSI RAA
DHIHSLR HS IRPR HMlHSR K29014 CPIRRA HS IRRA
DHINSLR H "lPR KJ•<utlSR !129014 CPIRRA HSIRRA
OHlNSLR HSIRPR HMINSR H29014 CPIRRA HSIRRA
DHIHSLR HSIRPR HMINSR K29014 C" "lAA H~ .1.RRA
2108 ATAC.ATCCCACACGATTCCCACCTCC'TCCCACACA.ACATCA.Tl'CACT"''CCCTCCAIC )04) -TACCTCCCCCACCACTCCCACCTCTCTCCACACAACATCACCCTCCTTCCACAC )049 - TACCTCCCCCACCACTCCCACCTCCCTCCACACAACATCACCCTTCTTCCACAC ))80 ATACCTTCCCCATCACTCCCACCTACCTCCACACAACATCACCCTCCTCCCACAC C 2898CTACATCCCTCATCAATCCCACCTCCCCCCCCACCACATCTCCATAATCCGGGAC 28 16 ---TCTCCCTCATCAATCCCACCTCCCTCCCCACCACATCTCCATAATCCCCCAA ....... ········ .. ... . ... . .. ..
CE )8 CCACCCATCCTTTCcCATCC CCACCCCTCCTTCCCCATCC CCACCCATCCTTTCCTATCC
TATCACCCTATCCTCAACTCCTTTCCACCCAATGCCCT TATCACCCCAATCCCACCCACATCATCAACCCTCACCC TATCAACCCAATCCCAACCATATCATCAACCCTCACCC TACCAACGCAATCCCAACCATATCATCAAGCCTCACCT ATCAC~CAAAACCACTTCAACCTCCACACCA
TATCACGCCCTCCCACCACCACTTCACCCTCGACACCA
CCAT-CCCCACTCTCCCATTAACACTCTCAACCAAAATCCTACCCATCCCCACCCAACCA ACACACCCCCCTC-CCCCTCAACACCCTCAACCACTCACCCACTCTCCCACACCCCATTC ACACACCCCTCTT-CCCCTTAACACTCTCAATCACTCACCCACTCTTCCACAACCCATCC ACACACCCCTCTT-GCCCTCAACACCCTCAATCACTCACCCACTCTTCCACAACCCATCC ATCCACCCCTCTC-GCCCTCAACACACTCAATCACCTCCCCACCCCTCCCCAACCCATTC CTCCACACCCCTC-CCCCTCAACACCCTCAATCACCTCCCCAGCCCACCCCAATCCATTC
AHORC CCTCACCCACCCCACCCTCA AACCAGTTCCATACCTATCATCTCCTAACATTCC CCTCAATCACCCCTCCCTCA AACCCCTTCACCTCCCATCACCTCCTCCGCCTCC CCTCAATCACCCATCACTCA AACCCATTCACCTCCCATCATCTCCTCCCCCTTC CCTCAATCACCCATCACTCA AACCCCTTCACCTCTCATCACCTCCTCCGCCTTC CCTCAACCAACCATCTCTCA AACCCATTCCACTCTCACCATCTCCTCCCTCTCC CCTCAACCAACCTTCTCTCA AAACCCTTCAACTCTCACCATCTCCTCCCTCTCC ... ....
TCCCTCTTTCTTCCACCCCTCACCCCCCTCTCCTCCTCATCCACCTAATC 2)96 TCGCAGTCCTCTCCAACCCCCACCCCACCCTCCTCCTCATCCACCTCATC ))) 0 TTCCCGTCCT A TCCAAACCACACCCAA TCCTCCTACTCA TCCAA TTCA TC ) ) l 6 TTCCCCTCCTCTCCAAACCCCACCCCACATTCCTACTCATCCAACTCATC )668 TCCCCCTCCTCTCTCACCCCCAACCAACTCTCCTCATCATCCAATTAATC )186 TCCCTGTCCTATCTCACCCCCACCCAACTCTCCTCATCATCCACTTAATC )101
[ I FREHER . BATCit 1 INSREC2. AU< RECEPTEUR 0' INSULINE
-----------------------------------------------------------------······---LIH1NSLI< IISIRPR HMINSR H29014 CPIRRA liSIRRA
DHI NSI.R HSIRPR HHIHSR 11Z.0014 Cri RRA HSIRRA
OHINS LR HSIRPR HM I NSR H29014 CPIRRA HSIRRA
OHIHSI.R HSIRPR HMIHSR K 29014 CPIRRA HSIRR!\
DHINSLR HSIRPR HMINSR :129014 CPIRRA ltSIRRA
Guinea
~sss 3 491 34!12 .!>fl4 3.34:5
CATCTTCCACCTCCAAAT CACCTCCCACCCACAAAC CACCTCCCACCTCCAAAC CACCTCCCACCTCCCAAC -ACCTCCCACCCCCCAAC - ATCTACCACCCOGCAAC
TCCATCCTTCCTCATCATTTCACCCTCAAAATTCC TCCATCCTCCCCCATCATTTTACTCTCAAAATTCC TCCATCCTTGCCCATCATTTTACTCTCAAAATTCG TCCATCCTTCCCCATCATTTTACTCTCAAAATCCC TCCATCCTCTCCCACCACTTCACTCTCAACA TCCATCCTCTCCCACCACTTCACCCTCAACATCCCC
AHDRCE 4 l ACTTTCCAATCACCCCTCACA ACTTTCCAATCACCACACACA ACTTTGCAATCACAAGGCACA ACTTTCCAATGACCACACACA ACTTTCCCATCACTCCCCATC ACTTCCCCATCACTC":CGCACC
TATCACACCCATTACTATCCCAACCCCACAAAACCCCTCCTCCCACTTCCCTCCATCCCA TATCAAACCCATTACTACCCCAAACCCCCCAACCCTCTCCTCCCTCTACCCTCCATCCCA TACCACACACATTACTATCCCAAACCCCCCAACCCACTCCTTCCTCTCACCTCCATCTCA T ACCACACACATTACTATCCCAAACCCCCCAACCCCTTCCTCCCCCTCACCTCCATCTCA TATCACACACACTATTACCCAAACCCTCCCAACCCACTCCTCCCCCTCCCCTCCATCCCT TATCACACACACTATTACCCCAACCCTCCCAACCCCCTCCTCCCCCTCCGCTCCATCCCC
CCCCACACCTTCCCACATCCTCTCTACTCTACTCCCACTCATCTATTCA CCCCACTCCCTCAACCATCCCCTCTTCACCACTTCTTCTCACATCTCC CCTCACTCCCTCAAGCATCCACTCTTTACTCCTTCTTCTCATATCTCCT CCCCIICTCCCTCAACCACGCACTCTTCACTCCTTCTTCCCACATC:TCr.r.rbrTT"CCCo CCTCACTCCCTCAAACATCCAATCTTCACCACCCACTCACATCTCTCGT CCCCIICTCCCTCAAACATCCCATCTTCACCACCCIICTCCCATGTCTCCT
CTTCTCTCCC GTCCTTTCCC CTCCTTTCcC CTCCTTTCCC CTCCTCTCCC CTACTCTCCC
TCCCCACCTTACCCCCTCACCCATACCACGCACTTTCCAACCACCAA TCIICCACCTTCCCACAACACCCTTIICCAAGCCCTCTCTAATCAACAC TCACTACCTTCCCTCACCAACCTTATCAACCCCTCTCTAATCAACAC TCACCACCCTCCCTCACCAACCTTACCAACCCCTCTCTAATCAACAC
ATTCTCACCCTCCCTCAACAACCCTATCACCCCCTCTCCAATCACCAA ATTCTCACCCTCCCACAACAACCCTACCACCCCCTCTCCAATCAGCAC . . . .. . . ..... .... .. ....... .
pig ins ulin receptor-related receptor (IRR) g e ne
o .me l a n og ast er insulin- like r eceptor mRNA, partial cds .
insulin receptor-related receptor (IRR) gene, 3 Hu man
Huma n mRNA for ins u lin receptor precursor
Mou se insulin receptor (IR) mlUIA, complete cds.
Rat insulin r e c e ptor alpha and beta s ubunits, comp l ete
annexe 7
3903
=3 586 b
end.
=5180 b
;:4167 b
c d s. =5397 b

(IFREMER.BATCH]PROALI.ALN PROLACTINE AMORCE 44 ~=====--======~===========================================================
CCPRL OKSPRL OKPROLA BTLACT HSPRLA HSPRLD HSPROL4 MGPLAC MMPRLHR1 MMPROLR SSPPLAC
250 TGCCACACTTCCTCTCTTCA 2 4 5 TGTCACACCTCCTCACTCCA 2 2 3 TGTCACACCTCCTCGCTCCA 329 TGCCATACCTCCTCCCTTC 2 2 8 TGCCACACTTCTTCCCTTGC 2 2 8 TGCCACACTTCTTCCCTTGC
8 6 TGCCACACTTCTTCCCTTGC 67 TGCCACACTTCCTCCTTAAC
3 0 5 TGCCCCACTTCTTCCCTGG 307 TGCCCCACTTCTTCCCTGG 2 71 TGCCACACCTCCTCCCTCT
** * ** ** ** *
TTCCCAATGACAAAGACCAAG TACCCAAGGACAAGGAGCAAG CACCCAAGGACAAGGAGCAAG CCCCGGAAGATAAAGAACAAG CCCCCGAAGACAAGGAGCAAG CCCCCGAAGACAAGGAGCAAG CCCCCGAAGACAAGGAGCAAG CTCCTGAAGATAAGGAGCAAA CACCTGAAGACAAGGAACAAG CACCTGAAGACAAGGAACAAG CGCCTGAAGACAAAGAGCAAG
** * ** ** ** ***
GAAA 301 CTTAGA 296 CTCAAG 274 CAACA- 379 CAACAG 279 CAACAG 279 CAACAG 137 CAGCAG 118 CTGAAA 356 CTGAAA 358 CAACAG 322
[IFREMER.BATCH]PROALII.ALN PROLACTINE AMORCE 45 ================~=====================================================--·=
CCPRL OKSPRL OK PRO LA BTLACT BTPRLB HSPRLA HSPRLD HSPROL7 MMPRLHR1 MMPROLR SSPPLAC AGPROA01 BTPROB2
584 579 560 669 732 568 568 TGCTCCACTGCCT 299 TGCTCCACTGCCT 645 CCATTCGCTGCCT 64 7 CCATTCGCTGCCT 611 TGCTCCACTGCCT 574 TGCTCCACTGCCT 3 17 TGCTCCACTGCCT
GCAGGGACTCCCACAAAATTGAC GCAGGGACTCCCACAAAATCGAC GCAGGGACTCCCACAAAATCGA GCAGGGATTCAAGCAAGATTGAC GCAGGGATTCAAGCAAGATTGAC GCAGGCATTCACATAAAATCGAC GCAGGCATTCACATAAAATCGAC GCAGGGATTCACATAAAATCGAC GCAGGGATTCCCACAAGGTTGAC GCAGGCATTCCCACAAGGTTGAC GCAGGGATTCACATAAGATTGAC GCAGGGATTCAAGCAAGATTGAC GCAGGGATTCAAGCAAGATTGAC
CCPRL OKSPRL OKPRLA BTLACT BTPRLB HSPRLA HSPRLD HSPRLOL7 MMPRLHR1 MMPRLHR SSPPLAC AGPROA01 BTPROB2 HSPROL4
MGPLAC
* **
ARNm de prolactine de carpe ARNm de prolactine de chum saumon ARNm de prolactine de saumon O.keta ARNm codant pour la préprolactine de taureau ARNm de prolactine de taureau exon 5 extrémité 3' de l'ARNm de prolactine humaine ARNm -de prolactine humaine ARNm de prolactine humaine, région 3 ', exon 5 ARNm de prolactin e de souris
ARNm de préprolactine de porc
extrémité 3' de l'ARNm de prolactine de bovin exon 3 et régions flanquantes du gène de
humaine ARNm de dinde
annexe 8

1
CHAINE B
ADNe lyrnnée GTG TGT GGC TCG GCT CTG GCC GAC CTC GTG
Bombyx TAC TGC GGG CGT CAC TTG GCT CGC ACT CTG
Eponge TTG TGC GGC TCG CAT GTA GTG GAA GCT ** **· * *
CHAINE A
ADNe lyrnnée AAC ATA GTG TGT GAA TGC TGC ATG AAA CCG TGC
Bombyx GGC ATC GTG GAT GAG TGT TGT CTC AGA CCC TGC
Eponge ATC GTG CAG GAG TCT ACG AGC GGC ATC TGC ** *** *** * ***
* : ide ntité entre les trois séquences . : ide ntité entre deux des trois séquences (dont une est la lyrnnée)
a nne x e 9





























