-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
1/15
Behavioral/Systems/Cognitive
Electrical Synapses and Synchrony: The Role of
Intrinsic Currents
Benjamin Pfeuty,1 German Mato,2 David Golomb,3,4 and David
Hansel1,51Laboratoire de Neurophysique et Physiologie du Systeme
Moteur, Centre National de la Recherche ScientifiqueUnite Mixte de
Recherche 8119, Universite
Rene Descartes, 75270 Paris Cedex 06, France, 2Comision Nacional
de Energia Atomica and Consejo Nacional de Investigaciones
Cientificas y Tecnicas,
Centro Atomico Bariloche and Instituto Balsiero, Universidad
Nacional de Cordoba, 8400 San Carlos de Bariloche, Argentina,
3Department of Physiology
and Zlotowski Center for Neuroscience, Faculty of Health
Sciences, Ben Gurion University of the Negev, Beer-Sheva 84105,
Israel, 4Mathematical
Biosciences Institute, Ohio State University, Columbus, Ohio
43210, and 5Interdisciplinary Center for Neural Computation, The
Hebrew University,
Jerusalem 91904, Israel
Electrical synapses are ubiquitous in the mammalian CNS.
Particularly in the neocortex, electrical synapses have beenshownto
connectlow-threshold spiking (LTS) as well as fast spiking (FS)
interneurons. Experiments have highlighted therolesof electrical
synapses in the
dynamics of neuronal networks. Here we investigate theoretically
how intrinsic cell properties affect the synchronization of
neurons
interactingbyelectricalsynapses.Numericalsimulationsofanetworkofconductance-basedneuronsrandomlyconnectedwithelectrical
synapses show that potassium currents promote synchrony, whereas
the persistentsodiumcurrent impedes it. Furthermore,
synchronyvaries with the firing rate in qualitatively different
ways depending on the intrinsic currents. We also study
analytically a network of
quadratic integrate-and-fire neurons. We relate the stability of
the asynchronous state of this network to the phase-response
function
(PRF),whichcharacterizes theeffect of small perturbations on
thefiring timingof theneurons.In particular, we show that
thegreater the
skew of thePRF towardthe firsthalf of theperiod, themorestable
theasynchronous state.Combining oursimulationswithour
analyticalresults, we establish general rules to predict the
dynamic state of large networks of neurons coupled with electrical
synapses. Our work
provides a natural explanation for surprising experimental
observations that blocking electrical synapses may increase the
synchronyof
neuronal activity. It also suggests different synchronization
properties for LTS and FS cells. Finally, we propose to further
test our
predictions in experiments using dynamic clamp techniques.
Key words: electrical synapses; conductance-based model;
synchrony; neuronal network model; intrinsic currents;
neocortex
IntroductionElectrical synapses are sites at which gap-junctions
bridge themembranes of two neurons. They have long been known to
existin invertebrates (Watanabe, 1958; Furshpan and Potter,
1959),but only recently has evidence of their ubiquity been
unequivo-cally found in the mammalian brain. Electrical synapses
arepresent in the inferior olive (Llinas and Yarom, 1986), the
hip-pocampus (Draghun et al., 1998; Venance et al., 2000), the
cere-
bellum (Mann-Metzer and Yarom, 1999), the locus
coereleus(Christieet al., 1989;Alvarez et al., 2002),
thestriatum(Kitaet al.,1990), the neocortex (Galarreta and Hestrin,
1999; Gibson et al.,
1999), the reticular thalamic nucleus (Landisman et al.,
2002),and between motoneurons (Kiehn and Tresch, 2002).
Experiments have revealed that electrical synapses are in-volved
in synchronizing neural activity (Draghun et al., 1998;Mann-Metzer
and Yarom, 1999; Beierlein et al., 2000; Perez-Velazquezand Carlen,
2000; Tamas et al., 2000; Deanset al., 2001;Hormuzdi et al., 2001).
In contrast to the results of these studies,it has been reported
recently that inspiratory motoneurons may
display more strongly synchronized activity in presence of
car-benoxolone (CBX), a blocker of electrical synapses, than in
thecontrol situation (Bou-Flores and Berger, 2001). Therefore,
inthis case, electrical synapses desynchronize neural activity.
Thedynamicsof networks of neurons interacting
viachemicalsynapses have been studied extensively (Golomb et al.,
2001).However, only a few theoretical studies have addressed the
dy-namics of networks in which neurons are coupled by
electricalsynapses. Models for pattern generation in the lobster
pyloricsystem have been investigated (Kepler et al., 1990; Abbott
et al.,1991; Meunier, 1992). Stable antiphase locking and
transitionsbetween in-phase and antiphase locking were demonstrated
inpairs of model neurons coupled by electrical synapses, and
itsimpact on rhythmogenesis was investigated (Sherman and Rin-zel,
1992; Cymbalyuk et al., 1994; Han et al., 1995). More re-
Received March 13, 2003; revised April 21, 2003; accepted April
22, 2003.
ThisworkwassupportedbyNorthAtlanticTreatyOrganizationPhysicalandEngineeringScienceand
Technology
CollaborativeLinkage Grant977683, Les Programmes
Internationauxde Cooperation ScientifiqueCentreNational
de la Recherche Scientifique (number 837), LAction Concertee
Incitative Neurosciences integratives et computa-
tionnelles (Ministere de la Recherche, France), and project
A99E01 from the Comision Asesora Cient fica de la
CooperacionArgentino-Francesa.Researchby
D.G.wassupportedbyIsraelScienceFoundationGrant657/01and by
the National Science Foundation, under Agreement 0112050. We
thank Y. Loewenstein, C. van Vreeswijk, B. Con-
nors,andY.YaromforstimulatingdiscussionsandB.Connors,Y.Loewenstein,andC.Meunierforcarefulandcritical
reading of this manuscript.
Correspondence should be addressed to Dr. David Hansel,
Laboratoire de Neurophysique et Physiologie du
Systeme Moteur, 45 Rue des Saints- Peres, 75270 Paris Cedex 06,
France. E-mail: dav [email protected]
paris5.fr.
Copyright 2003 Society for Neuroscience
0270-6474/03/236280-15$15.00/0
6280 The Journal of Neuroscience, July 16, 2003
23(15):62806294
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
2/15
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
3/15
(2003). For Ired Icred, the solution of Equation 6 diverges in
finite time.
This corresponds to the firing of an action potential. If one
supplementsEquation 6 with the condition that the variable vred is
reset to Vr V*immediately after vred reaches some threshold, VT,
from below, this
yields a reduced model that accurately describes the dynamics of
theneuron in the limit Iext
red3 Ic
red. In particular, the currentfrequencyrelationship( IFcurve)
of this model andthe neuronbehave similarly inthis limit, for any
value of the parameters Vr and VT. When Iext
red Icred is
not small, the reduced model is no longer an exact description
of theneuronal dynamics. However, one can fit the parameters so
that the re-duced model provides a good approximation of the IF
curve of theneuron. The difference, VT Vr, will be called the reset
depth.
It is convenient to rewrite the subthreshold dynamics of the
reducedmodel in terms of the dimensionless variables vred and I
red defined by:
vred A
Cred0vred V* (7)
Ired A
Cred2 I
red02, (8)
where 0 has the dimension of a time. This yields:
0dv red
dt v red
2 Iext
red Ic
red. (9)
The variable vred isresetto V r A(Vr V*)0 /Cred whenever it
reachesthe threshold value: V T A(VT V*)0 /Cred. Note that V r 0.
Asimilar model has been used to study networks of excitatory and
inhibi-tory neurons (Latham et al., 2000; Hansel and Mato,
2003).
The subthreshold dynamics (Eq. 9) need to be supplemented with
amodel for the suprathreshold part of the membrane potential
timecourse. Assuming that the width of the action potentials is
much smallerthan the interspikes, we represent their time course by
a -function ateach time a spike is fired. Therefore, the
(dimensionless) reduced mem-brane potential of the neuron can be
written:
V redt vredt
spikest tspike, (10)
where measures the integral over time of the suprathreshold part
of anaction potential.
The model definedby Equations9 and 10willbe called
theQIFmodel.To simplify notations of the reduced model, we will
drop the index(red) and the tildes.
Phase reduction in the weak coupling limit. Let us consider a
neuronfiring action potentials periodically with an interspike T. A
small andinstantaneous perturbation applied at time t after a spike
induces asmall change in the timing of the subsequent spikes. This
change, whichdepends on t, or equivalently on 2t/T, can be
characterized bya function, Z(), which measures the delay or the
advance induced inthe firing times after the perturbation. A
positive value ofZ() indicatesthat theperturbation advances
thesubsequent spikes. A negative value of
Z() indicates a delay. The effect of noninstantaneous weak
perturba-tions (such as spikelets) can be estimated by convolving
the phase-response function (PRF), Z, with the perturbation. It can
be shown thatthe dynamic behavior of a network of weakly
interacting neurons can becompletelydescribedin terms of the
response function in the frameworkof the phase reduction approach
(Kuramoto, 1984; Ermentrout and Ko-pell, 1986;Hanselet al., 1993,
1995; Golomband Hansel, 2000; Neltner etal., 2000; Lewis and
Rinzel, 2003). In this approach, a phase variable isassociated
witheach neuronin thenetwork. This variable,i(i 1, . . . ,
N), measures the time elapsed since the last action potential
fired byneuron i. For a network of identical neurons coupled with
electricalsynapses, one can show (see Appendix) that the phase
variables follow aset ofn first-order coupled differential
equations:
di
dt v
j1
n
Jiji j it, i 1, . . . , N (11)
(i j ) the phase coupling between neurons i and j given by:
i j g
20
2
Zu iVu j Vu idu .
(12)
Jij is the connectivity matrix, and i(t) is a white noise with
zero meanand variance
2
20
2
Zu 2du
with , the SD of the noise of the nonreduced dynamics.Numerical
integration. In the simulations of the conductance-
based model, the differential equations were integrated using
thesecond-order Runge-Kutta scheme with fixed-time step: t 0.01
msec. Averaged quantities (firing rate, CV, ) were com-puted over a
time period of 1 sec after discarding a transient of500 msec.
ResultsSingle neuron and coupling properties of
theconductance-based modelFiring properties of single neuronsIn the
absence of persistent sodium and slow potassium currents(the
control model), our conductance-based model neuronfires tonically
for large enough external currents, Iext Ic 0.16A/cm 2. The
frequency of the discharge in response to a step ofcurrent is
plotted as a function of the step amplitude (IFcurve)in Figure 1A.
For Iext larger than but close to Ic, the minimumvalue of the
external current required for the neuron to fire, the
firing frequency can be arbitrarily small, because action
poten-tials appear at Ic through a saddle-node bifurcation [for a
defini-tion of a saddle-node bifurcation, see Strogatz (1994) and
Rinzel
Figure 1. IFcurves of the conductance-based model neuron. Each
panel corresponds to
adifferentsetofintrinsicconductancevalues.Thefiringratewascomputedfromthesteady-state
response of the neuronto stepsof constant external current of
different amplitudes.A, Controlcase: gK 9 mS/cm
2, gKs gNaP 0. B, gK 2.5 mS/cm2, gKs gNaP 0. Note that the
scale of the x-axis is one-half the scale used in A. The main
effect on the IFcurve of thereduction ofgK is multiplicative
(change in the gain). C, gK 9 mS/cm
2, gNaP 0.2 mS/cm2,
gKs 0.Themaineffectonthe
IFcurveofthepersistentsodiumissubtractive(changeintherheobase). D,
gK 2.5 mS/cm
2, gKs 0.2 mS/cm2, gNaP 0. The slow potassium current
reducesthegainoftheneuronandpreventslow-frequencyfiring.Italsolinearizesthe
IFcurve(compare with B).
6282 J. Neurosci., July 16, 2003 23(15):6280 6294 Pfeuty et al.
Electrical Synapses, Intrinsic Currents, and Synchrony
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
4/15
and Ermentrout (1998)]. Decreasing gK does not significantly
modify the resting potential and the rheobase of the neuron,
butit increases the gain of the IF curve (Fig. 1 B). The slow
potas-sium (IKs) and persistent sodium (INaP) currents modify the
on-setof firing, because these currents areactivated at rest. As
shouldbe expected, the persistent sodium current increases the
excit-ability of the neuron and reduces its rheobase, Ic (Fig. 1C).
For
gNaP 0.08 mS/cm2, the neuron is spontaneously active. As
shown in Figure 1 D, the main effects of adding IKs on the
IFcurve are a translation to the right, a linearization, and, if
gKs islarge enough, a suppression of the ability of the neuron to
fire atlow rates (because for large enough gKs, a subcritical Hopf
bifur-cationoccurs at the onset of firing[for a definition of a
subcriticalHopf bifurcation, seeStrogatz (1994)and Rinzeland
Ermentrout
(1998)].The response of the neuron to a step of external
current,
whose value is adjusted to give a firing frequency of 50 Hz,
isplotted in Figure 2 for different combinations of the
intrinsiccurrents. The action potentials are shown with a higher
temporalresolution in Figure 3C. In the control case (Fig. 2A; and
Fig. 3C,solid line), action potentials are narrow and are followed
by astrong afterhyperpolarization (AHP) that brings the
membranepotential 15 mV below its resting value. When gK is
decreased(Fig. 2 B), theresting potentialof theneuron does
notchange, butthe AHP is reduced and the membrane potential always
remainsabove rest. This conductance also controls the width of the
spikeand the depth of the AHP [Fig. 3C, compare the solid line
(gK
9 mS/cm
2
) with the dotted line (gK 2.5 mS/cm
2
)].The persistent sodium and the slow potassium currents
sig-nificantly modify the resting potential and the external
current
required to adjust the firing rate. In contrast, they only
slightlyaffect the depth of the repolarization, measured from
threshold,after the action potential (Fig. 2C,D). The size and
width of theaction potentials remain almost unchanged when the
conduc-tances of these currents are varied [Fig. 3C, compare the
dottedline with the dashed-dotted line (effect ofgKs) and the solid
linewith the dashed line (effect ofgNaP)].
Properties of the spikeletsWhen a neuron fires an action
potential, it induces a depolariza-tion, called a spikelet, in the
neurons that are connected to it. Thespikelets generated by a
neuron firing one isolated spike in re-sponse to a very brief but
strong pulse of current (Fig. 3A) orfiring tonically at 50 Hz (Fig.
3C), are shown in Figure 3, B and D,respectively, for different
parameters of the ionic currents.
In all four cases displayed, the width of the spikelets is
muchlarger than the width of the presynaptic spikes that
generate
them. This is because the spikelets are a filtered version of
thepresynaptic membrane potential profile.Thesize of thespikeletis
practically unaffected by thepresence
Figure 2.
Theresponseoftheneurontoastepofcurrent.Astepofcurrentof200msecwasinjected
into theneuron.No noisewas included. Themembranepotential beforeand
after the
currentinjectionisindicatedtotheleftofeachpanel.Inallcases,theexternalcurrentduringthe
stepwaschosentoobtainadischargerateof50Hz.A, Control
case.Externalcurrent,Iext, variesfrom0to1.10A/cm
2.Restingpotentialis 63mV.B,gK 2.5mS/cm
2,gKsgNaP 0. Iextvariesfrom0to0.48 A/cm 2. Note that
theconcavityof themembrane
potentialtimecourseisalwaysupwardincontrasttoA.C,gK 9mS/cm
2,gNaP 0.2mS/cm2,gKs 0.Theneuron
is hyperpolarized by the injection of a negative current, Iext
1.55 A/cm2, before the
current step, to prevent spontaneous firing. The step amplitude
is 1.0 A/cm 2. D, gK 2.5mS/cm2, gKs 0.2 mS/cm
2, gNaP 0. The external current varies from 0 to 4.88 A/cm2.
The resting membrane potential is more hyperpolarized than in B,
because of the slow potas-sium current that is activated at rest.
The (shifted) time course of the membrane potentialduringthe
dischargeis very similar tothe tracein B. Notethe
transienthyperpolarizationright
after the switching off of the current, which is caused by the
slow relaxation ofIKs back to itsresting value.
Figure3. Thespikeletsinducedfor differentcombinations of
ioniccurrents. Solidline,gK 9 mS/cm 2, gKs gNaP 0 (control case).
Dotted line, gK 2.5 mS/cm
2, gKs gNaP 0.Dashed line, gK 9 mS/cm
2, gKs 0, gNaP 0.2 mS/cm2. Dashed-dotted line, gK 2.5
mS/cm 2,gKs 0.2mS/cm2,gNaP 0.A, Anaction potentialgenerated by a
shortand strong
current pulse. The duration and the amplitude A of the pulse are
the same for all four traces: 1 msec and A 50 A/cm 2. The neuron
was hyperpolarized to prevent firing whenpersistent sodium was
present and to obtain the same initial membrane potential for all
four
parametersets.Thesolidlineand thedashedline overlap. B,
Spikeletinduced bya presynapticneuronfiringanactionpotentialasin A.
Thesynapticconductanceisg
0.005mS/cm2,whichcorrespondstoacouplingcoefficient(Amitaietal.2002)CC
5%.Thepostsynapticneuronhasthesameintrinsicpropertiesasthepresynapticneuron.C,Aconstantexternalcurrentisinjected
tomake theneuron fire at 50 Hz.The external current is Iext
1.10A/cm
2
forthe solid line,Iext 0.48A/cm2 for the dotted line, Iext
4.88A/cm
2 for the dashed-dotted line, andIext 0.55A/cm
2 for the dashed line. The subthreshold voltage of the neuron
when thepersistent sodium is added (dashed line) is below the
control (solid line), although the
persis-tentsodiumcurrentacceleratesthedepolarizationoftheneuron,becauseherewecomparethefiring
patterns for the same firing rate. The external current is
therefore smaller when thepersistent sodium is present than in the
control case. D, Spikelets induced by a presynaptic
neuron firing tonically, as in C. The postsynaptic and the
presynaptic neurons have the sameintrinsic properties.
Pfeuty et al. Electrical Synapses, Intrinsic Currents, and
Synchrony J. Neurosci., July 16, 2003 23(15):62806294 6283
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
5/15
of the persistent sodium (Fig. 3B,D, com-pare the solid line
with the dashed line), be-cause this current does not greatly
affect theshape of the action potential. The only no-ticeable
effect of the persistent sodium onthe spikelet is to slow down its
decay timecourse. This is because it reduces the input
conductance of the postsynaptic neuron (by50% at 76 mV).
The modulation of the spikelets by thedelayed rectifier
potassium current is pri-marily caused by changes in the
presynapticneuron voltage time course. When gK is re-duced, the
action potential is broader, andthis induces an increase in the
amplitudeand the width of the spikelets. Although theslow potassium
current does not greatly af-fect the width of the action potential,
it con-tributes to bringing the membrane of thepresynaptic neuron
transiently below itsholding potential. This explains why in
Fig-ure 3B the spikelet is narrower and displaysa faster and deeper
repolarization in thepresence ofIKs.
The spikelets generated by tonically fir-ing neurons at low
firing rates are similar tothose in Figure 3B (data not shown).
Forhigh-enough firing rates, the slow potas-sium saturates,
affecting the dynamics pri-marily as would a constant
hyperpolarizingcurrent. Therefore, its effect on the voltage traces
and on thespikelets becomes less pronounced (Fig. 3, compare B and
D).
Synchrony in the conductance-based network modelThe goal of this
section is to show how synchrony properties ofour conductance-based
network model depend on the intrinsiccurrents of the neurons.
Further understanding of the effectsdescribed in this section will
be provided in the next section byinvestigating analytically a
simplified and more abstract model.
Synchrony is modified when intrinsic conductances are changed
Apair of conductance-based neuronsThe effect of intrinsic currents
on neuronal network dynamicscan first be demonstrated by studying
the dynamics of a pair ofneurons. Here, as an example, we show that
a persistent sodiumcurrent tends to promote antiphase locking,
whereas a potassium
current promotes in-phase locking. In Figure 4A, the traces
oftwo identical neurons (gK 9 mS/cm2; gKs gNaP 0) are
plotted over a time interval of 100 sec. Both neurons receive
anoisy input that makes them fire at 50 Hz. These traces
suggestthat the neurons fire action potentials in synchrony. This
is con-firmed by the cross-correlation of the traces computed over
alonger time interval (100 sec), which displays a strong peak
cen-tered around t 0 msec, as shown in Figure 4 B. In contrast,when
thepersistent sodiumconductanceis large enough(gNaP0.4 mS/cm2),
neurons tend to fire in antiphase (Fig. 4C), and thepeak of the
cross-correlogram is shifted to 10 msec (Fig. 4 D). If aslow
potassium current is added (gKs 0.15 mS/cm
2) whilekeeping the persistent sodium conductance the same,
antiphase
locking becomes unstable and neurons tend to fire in-phase
(Fig.4 E, F). Therefore, a slow potassium current is a promoter
ofsynchronous activity.
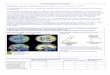
Large networks of conductance-based neuronsIn Figure 5 the
synchrony measure, , is plotted against the con-ductances gK, gKs,
and gNaP. Examples of raster plots of networkactivity are also
displayed in this figure, for particular values of
the intrinsic conductances (Fig. 5, insets). In each of the
panels,the external input (average deviation and SD) is kept
constant.Therefore, when the intrinsic conductances vary, the
average fir-ing rate of the neurons and the CV of their interspike
intervalschange. In particular, the firing rate decreases
(respectively in-creases) when the potassium (respectively the
persistent sodium)conductances increase (top right figures in each
of the panels inFigure 5).
In Figure 5A, the effect ofgK on the synchrony level is
shown(gKs gNaP 0). When gK is small enough (gK 4.5 mS/cm
2),is very close to zero. A detailed study shows that in this
region,is on the order of 1/N, where Nis the number of neurons
inthe network, and vanishes in the limit of a very large
network
(data not shown): non-zero values ofare attributable to
finitesize effects, and the network is asynchronous. For gK of
4.5mS/cm2, increases rapidly up to 0.35. For example, in theraster
plot shown in the right inset of Figure 5A (control situa-tion, gK
9 mS/cm
2), 0.34. This corresponds to a state inwhich the neurons fire
together within time windows of approx-imately one-third of the
period.
Figure 5B shows the effect of the slow potassium current forgNaP
0 and gK 2.5 mS/cm
2. For gKs 0, the neurons fireasynchronously. At gKs 0.06
mS/cm
2, starts to increase. Forlarge gKs, saturates at a value, 0.55,
that is substantiallylargerthan in thecontrol situation. This value
ofcorrespondstoa tight synchrony of the action potentials fired by
the neurons
(Fig. 5B inset on the right).In Figure 5C, is plotted against
gNaP for gK and gKs at theircontrol values (gK 9 mS/cm
2, gKs 0). Clearly, is a decreas-
Figure 4. Effect of persistent sodium and potassium currents on
the synchronization of a pair of neurons: phase versus
antiphasestability.Resultsfrom numericalsimulations. The
synaptic conductance isg 0.005mS/cm2
.TheSDofthenoiseis 0.3 mV/msec 1/2. In all of the simulations,
neurons were firing in antiphase at the beginning of the simulation
(data
notshown).Spiketrainswerecomputedwithatimebinof1msec.Thespikecross-correlograms(
B, D, F)werecalculatedfromspiketrainsrecorded over 100sec(after
discarding a transientof 5 sec)and were normalized tothe
expectednumber ofspikes duringthis time interval. Neurons fire at
an average firing rate of 50 Hz. A, B, Traces of the voltage of the
two neurons and thecross-correlograms of the spike trains for the
control parameters and Iext 1.08A/cm
2. Neurons tend to fire in-phase. C, D,
Persistentsodiumcurrentstabilizesantiphaselocking.ParametersaregK9mS/cm2,gNaP
0.4mS/cm
2,andgKs0. Iext1.38A/cm 2. E,
F,Slowpotassiumconductancedestabilizesantiphaselocking.ParametersaregK
9mS/cm
2,gNaP 0.4mS/cm2, and gKs 0.15 mS/cm
2. Iext 0.80A/cm2. Neurons tend to fire in-phase.
6284 J. Neurosci., July 16, 2003 23(15):6280 6294 Pfeuty et al.
Electrical Synapses, Intrinsic Currents, and Synchrony
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
6/15
ing function of gNaP. When gNaP is large enough (gNaP
0.1mS/cm2), the neurons fire asynchronously.
In the results presented in Figure 5, the firing rate of the
neu-rons is not controlled. Therefore, the origin of the variation
ofwith the intrinsic current conductances is not clear. It may
beattributable to the fact that, in general, synchrony depends on
the
firing rate for given intrinsic currents. It might also be a
result ofthe fact that dynamic properties of neurons with different
intrin-siccurrentsmay be differenteven if they have thesame
firingrate.
Dependence of synchrony on intrinsic currents at fixed firing
ratesTo clarify further how potassium and sodium currents affect
thesynchrony of the network, we performed another set of numeri-cal
simulations in which the external input and the noise levelwere
tuned to keep constant both the average firing rate of theneurons
and the CV of their spike trains when the conductances
of the intrinsic currents were changed. The results are shown
inFigure 6 (three panels on the left). In all of the simulations,
theaverage firing rate of the network was 50 Hz and the CV
0.12(Fig. 6, panels on the right).
The level of synchrony increases with the potassium
conduc-tances. For small-enough gK or gKs, the activity of the
network isweakly correlated. A detailed study of the dependence
ofwiththe network size reveals that for gK (respectively gKs)
smaller than
g*K 4 mS/cm2 (respectivelygKs smaller thang*Ks 0.075 mS/cm
2),ison the order of1/N(data not shown). Therefore in this
rangeof conductances, the network is in theasynchronous state.Atg*K
and
g*Ks, a transition to a synchronous state occurs, and beyond
thesevalues the network settles into a synchronous state. This
occurs al-
though the level of noise has been increased.In contrast to the
potassium conductances, which promotesynchrony, the persistent
sodium current impedes it, as shown in
Figure 5.
Effectofintrinsiccurrentsonthesynchronizationofalargenetwork.Resultsfromnumerical
simulations. Details on the simulation and the network parameters
are given in
Materials and Methods. The synaptic conductance is g 0.005
mS/cm2
. In all of the simula-tions, the SD of the noise was set at 0.6
mV/msec 1/2. The synchrony level, (left),
theaveragefrequency,andtheaverageCVoftheneuronaldischargesareplottedasafunctionofanintrinsicconductancethatisvaried.A,Theconductance,gK,varies;gKsgNaP
0.Theexternalcurrent is Iext 0.8A/cm
2. For gK g*K,increases and saturates. Inset, Raster plot
repre-senting the spiking times of 100 neurons for gK 9 mS/cm
2 (neurons are arrayed vertically,and the x-axis corresponds to
time where the scale denotes 25 msec). Panels on the right
indicate that the average frequency of the neurons decreases
when gK increases, because thegain of the IFcurve decreases. The
variability of the neuronal discharge varies slightly
andnonmonotonically in the investigated range ofgK. B, The
conductance, gKs, varies; gK 2.5mS/cm2,gNaP 0.The externalcurrent
is Iext 2A/cm
2.Inset,RasterplotsareforgKs 0andforgKs 0.1mS/cm
2.AtlargevaluesofgKs,aslightdecreaseoccurs,buttheactivityofthenetworkremainsstronglysynchronized.PanelsontherightindicatethattheaveragefiringrateoftheneuronsisstronglyreducedwhengKsvaries,primarilybecauseIKshasastrongsubtractiveeffect
on the IFcurve of the neurons (the rheobase increases with gKs). C,
The conductance,
gNaP,varies;gK 9mS/cm 2,gKs 0.Theexternalcurrentis Iext0.8A/cm
2.Thesynchronyis reduced when gNaP increases. Inset, Raster plot
for gNaP 0.2A/cm
2. Panels on the
rightindicatethattheaveragefiringrateoftheneuronsincreaseswith
gNaP,primarilybecauseofthe
subtractive effect ofINaP on the IFcurve of the neurons (the
rheobase is reduced when gNaPincreases).
Figure 6. The synchrony level depends on the intrinsic currents
when the frequency is keptconstant.Results
fromnumericalsimulations. Details on the parametersare givenin
Materials
and Methods. The synaptic conductance is g 0.005 mS/cm2
. The external current, Iext, andtheSDofthenoise,, werechangedto
control theaveragefiringrate andtheaverageCVof theneurons.In allof
thesimulations,theaveragefiring rate is50 Hz 10%,andtheCV
0.12.Ineach of the panels, the corresponding values ofIext andare
plotted versus the conductance
that is varied (panels to the right). A, The conductance, gK,
varies; gKs gNaP 0. B, Theconductance,gKs, varies;gK 2.5mS/cm
2,gNaP 0. C, The conductance,gNaP, varies;gK 9mS/cm 2, gKs
0.
Pfeuty et al. Electrical Synapses, Intrinsic Currents, and
Synchrony J. Neurosci., July 16, 2003 23(15):62806294 6285
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
7/15
Figure 6C. For large-enoughgNaP, synchrony is even destroyed.
Asharp transition to the asynchronous state occurs for gNaP
0.15mS/cm2.
Relying on the way intrinsic currents affect spikelet
modula-
tion (Fig. 3), one might naively expect that potassium
currentsthat reducethe size of the spikelets shouldalso
reducesynchrony,and that persistent sodium current, which increases
this size,should promote it. As a matter of fact, we found a trend
that wasexactly the opposite.
Firing rate affects synchrony differently when different
intrinsiccurrents are involvedWe also investigated in what ways the
network dynamic statedepends on the firing rate of the neurons. The
firing rate waschanged by varying the average external input, and
was plottedas a function of the frequency of the neurons for
several combi-nations of potassium and sodium conductances.
Figure 7 plots the synchrony measure, , versus the average
firing rate of the neurons for four sets of conductance
parametersandtwo levelsof noise.The qualitative behavior ofas a
functionof the firing rate depends on the intrinsic currents
involved.
As expected, is always larger for the smaller level of noise(
0.3 mV/msec1/2) (Fig. 7,circles) than for the largerone (0.6
mV/msec1/2) (Fig. 7, squares). In particular, for 0.6 mV/msec1/2,
the noise is strong enough to prevent synchrony in theentire range
of firing frequencies for gK 3.5 mS/cm
2 (Fig. 7B)and for gNaP 0.15 mS/cm
2 (Fig. 7C). This level of noise issufficient to destroy
synchrony as well in the control situation,but only at 20 Hz (Fig.
7A).
In thecontrol case,increases monotonously with thefiring
rate(except for a small decrease followedby a slight increase
between 50
and100Hzfor 0.3mV/msec
1/2
). In contrast, forsmallergK (e.g.,gK 3.5 mS/cm2), we find a
nonmonotonous variation of syn-
chrony: increases at low firing rates but decreases again for
firing
rates of20 Hz.Thenetworkstate becomesasynchronousforfiringrates
of60 Hz (Fig. 7B, squares). If the slow potassium current isadded
(gKs 0.15 mS/cm
2), again varies monotonously with thefiring rate (Fig. 7D).
Finally, with enough persistent sodium currentand not overly strong
noise (gK 9 mS/cm
2, with gNaP 0.15mS/cm2, gKs 0) (Fig. 7C, squares), varies
nonmonotonouslywith the firing rate, as in the reducedgK case (Fig.
7B).
Synchrony in networks of quadratic integrate-and-fireneurons
depends on the phase response functionThePRF,which characterizes
how neurons respondto small pertur-bations, is a key concept to
understand the relationship betweenintrinsic properties of neurons
and their collective dynamics(Kuramoto, 1984; Hansel et al., 1993;
van Vreeswijk et al., 1994;Kopell and Ermentrout, 2002). This
function depends on the excit-ability properties of the neurons and
therefore is determined by theintrinsic currents involved in their
dynamics. One expects that thedifferences in the dynamic behavior
of our conductance-based net-work model fordifferent sets of
parameters reflect thechanges in theexcitability properties of the
neurons. However, relying on numeri-calsimulations alone, itwould
verydifficult toestablish generalprin-ciplestorelatetheexcitability
properties andtheneuronal PRFto thesynchronization properties.
Below, we consider a network of QIFneurons (Eqs. 9, 10) fully
connected by electrical synapses. As weshow below, the qualitative
properties of the excitability of the QIFneurons crucially depend
on the parameters (the threshold and thereset voltages and the
external current). Thanks to its relative sim-plicity, this model
can be investigated using analytical techniques. Itreveals how, in
the framework of this model, one can relate thestability of
antiphase locking of a pair of neurons and thestability ofthe
asynchronous state of a large network to the shape of the PRF
oftheneurons.Subsequently, we showthat similarrules canbe appliedto
conductance-based models, and that they provide a unifiedframework
explainingthediversity of behaviorfoundin ournumer-
ical simulations of the conductance-based model.
The membrane potential and the PRF of QIF neuronsIn the
Appendix, it is shown that between two successive spikesthe
subthreshold membrane potential of a QIF neuron reads:
v(t)Iext Ic tantIext Ic0 tan1Vr
Iext Ic, (13)
where tmeasures the time elapsed since the first spike. The
firingperiod, T, is determined by the condition: v(T) VT.
Therefore:
T 0
tan1 VT
Iext
Ic
tan1 Vr
Iext
Ic
Iext Ic
. (14)
In the weak coupling limit,the dynamics of the network are
com-pletely determined by the phase-response function, Z()
(seeMaterials and Methods), which depends on three
parameters,namely, the firing frequency of the neurons (or
equivalently theexternal current Iext), thethresholdVT, and
thereset potential Vr.The response function of the QIF model can be
derived analyti-cally as shown in the Appendix. One finds:
Z 0
vT22
Iext Ic
. (15)
Changing the ratio Vr/VT for a fixed reset depth, VT Vrhas a
strong influence on the shape of the trajectory of the phase-
Figure 7. Synchrony varies with the firing rate in a way that
depends on the intrinsic cur-rents. Results from numerical
simulations. Details on the simulation parameters are given
inMaterialsandMethods.Thesynapticconductanceisg
0.005mS/cm2.Theaveragefiringrateof the neurons is varied by
changing the external current. In each of the panels, the results
aredisplayedfortwonoiselevels.Circles, 0.6mV/(msec)1/2.Squares,
0.3mV/(msec)1/2.
A, gK 9 mS/cm2, gKs gNaP 0. The external current was varied
between 0.15 and 2.7
A/cm 2 (100 Hz).B,gK 3.5 mS/cm2,gKs gNaP 0. The external
current, Iext,wasvaried
between 0.11 and 1.4 A/cm 2. C, gK 9 mS/cm2, gKs 0, gNaP 0.15
mS/cm
2. Iext wasvaried between 1.17and0.8A/cm 2. D,gK 3.5mS/cm
2,gKs 0.15mS/cm2,gNaP 0.
Iext was varied between 2.3 and 7.2 A/cm2. The firing rate
cannot be reduced to 20 Hz
because of the presence of the slow potassium cur rent (see also
Fig. 1 D).
6286 J. Neurosci., July 16, 2003 23(15):6280 6294 Pfeuty et al.
Electrical Synapses, Intrinsic Currents, and Synchrony
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
8/15
response function of the neuron, as shown in Figure 8. In
Figure8A, the membrane potential of a QIF neuron firing at 50 Hz
isplotted for three values of the ratio Vr/VTand a fixed reset
depth:VT Vr 3. For Vr/VT 0.1, the concavity of the subthresh-old
trajectory is always directed upward. In contrast, for Vr/VT 10, it
is always downward. For Vr/VT 1, the concavitychanges from upward
to downward.
It is easy to see that Z() 0 for all , and that it is with
onemaximum. Thelocationof themaximum ofZdepends on Vr, VT,and Iext.
This is depicted in Figure 8, where the voltage trace andthe
response function are plotted for different values of
theseparameters. For Vr/VT 1, Z() is symmetric and its maxi-mum is
always at . For Vr/VT 1, the maximum ofZislocated in the first half
of the firingperiod, whereas forVr/VT
1, it is located in the second half. The value of Z at the
maximumalso depends on Vr, VT. It has smaller values when Vr/VT
1.The larger the firing rate, the stronger the dependence of Z
onVr/VT(Fig. 8, compare B, for 50 Hz, and C, for 10 Hz).
A pair of identical QIF neurons without noiseWhen, in the
absence of noise, two identical neurons interact in asymmetric way,
they reach, at large time, a phase-locked state inwhich the two
neurons fire action potentials with a fixed phaseshift, . In the
Appendix, it is shown that is a solution to theequation:
0, (16)
wherethe function
() 1/2(() ()),and ()istheeffective phase coupling (see Materials
and Methods). Because ofthe symmetryof the systemand the
2periodicity of the function
, there are always at least two solutions tothis equation: 0 and
. The firstsolution corresponds to the state in whichthe two
neurons fire in-phase (in-phaselocking), the second one to the
state inwhich they fire in antiphase (antiphaselocking). Other
solutions with 0
may also exist. However, only solutions thatare stable can be
reached at large time.Therefore, out of all of the solutions
toEquation 16, only those that satisfy the con-dition (see
Appendix):
d
d 0 (17)
correspond to phase-locked states that thetwo neurons can
eventually reach startingfrom appropriate initial conditions. In
thefollowing, we focus on the stability of theantiphase state of a
pair of QIF neurons.
Stability conditions of antiphase lockingThe voltage trajectory
consists of threeparts: (1) a subthreshold part, during whichthe
membrane potential increases with timefrom a value Vr to a
threshold value VT; (2)a spike, modeled by a -function of
ampli-tude, and (3)a resetting, where potentialisinstantaneously
brought back to Vr. Then,as explained in the Appendix,
antiphaselocking of a pair of QIF neurons is stable if:
d
d gSsp Sr Ssub 0
(18)
with:
Ssp vdZ
d (19)
Sr VT Vr)Z() (20)
Ssub 1
20
2
Zdv
d d. (21)
Thefirstterm,Ssp, is thecontribution of thepresynapticspikesto
the destabilization of antiphase locking. The second term, Sr,
isthe contribution of the instantaneous reset of the membrane
po-tential. The last term, Ssub, corresponds to the effect of the
cou-pling between the two neurons when the presynaptic neuron
issubthreshold. These three terms are plotted in Figure 9A as
afunction of the ratio Vr/VT, for 1, VT Vr 3, and afrequencyv 50
Hz.
The terms Sr and Ssub (Fig. 9, dotted line and
dashed-dottedline, respectively) are invariant under the
transformationVr/VT 3 VT/Vr. That is why the corresponding curves
inFigure 8B are symmetric around Vr/VT 1 (note the logarith-mic
scale of the x-axis). Sr is always positive and therefore
desta-
bilizes antiphase locking. In contrast, Ssub is always negative
andleads to stabilized antiphase locking. The sum of the two terms
isalso plotted in Figure 9A (double-dotted-dashed line) to show
Figure 8. Voltage traces and phase-response functions of the QIF
neuron. The voltage traces and the phase-response func-tions were
computed using Equations 13 and 15, respectively. 0 10 msec. In A
and B, the external current is such that thefiring rate is 50 Hz.
A, The voltage trace of the neuron in response to a constant
injected current for three values of the ratioVr/VT. The reset
depth is the same in all three cases: VT Vr 3. Note the changes in
the concavity of the voltage
tracebetweenthespikeswhenVr/VTincreases.TheexternalcurrentisIext0.97for
Vr/VT 1andfor Vr/VT10,and Iext0.66 forVr/VT 0.1. B, The
phase-response function,Z,forthesamecasesasinA. C,Sameasin
B,butforafiringrateof10Hz. The phase-response changes less than in
B when Vr/VTvaries.
Pfeuty et al. Electrical Synapses, Intrinsic Currents, and
Synchrony J. Neurosci., July 16, 2003 23(15):62806294 6287
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
9/15
that their overall contribution is destabilizing except for
verysmall or very large values ofVr/VT.
Because the maximum ofZ moves from the first part of theperiod
to the second part of the period when Vr/VT crosses 1(Fig. 8), the
sign of the derivative ofZatalso changes from
negative to positive, and the term Ssp (dashed line) tends to
de-stabilize antiphase locking for Vr/VT 1. The sum of all
threeterms ofthe right sideof Equation18 isplotted inFigure 9A
(solidline). Antiphase locking is stable only if the phase at which
Zreaches its maximum is small enough.
The phase diagram for the stability of antiphase locking as
afunction of Vr/VT and the firing frequencies is displayed inFigure
9C. The stability of antiphase locking requires thatVr/VT be small
enough. Moreover, the range of values ofVr/VT where it is stable
increases with the frequency of theneurons. This is because the
contribution of the term Ssp in theright side of Equation 18 grows
with the firing rate and tends todominate the contribution of the
two other terms. In contrast, at
low frequencies, the response function weakly depends
onVr/VT(Fig. 8), and Ssp, which is proportional to the frequency,is
negligible. Moreover, Sr Ssub is positive except for very small
or very large values ofVr/VT. Therefore, for small
frequencies,the domain of stability of antiphase locking is very
narrow.
This analysis shows that for a pair of neurons, the stability
ofantiphase locking depends critically on the shape of the
phase-response function and in particular on the sign of the
derivativeof the response function at the mid-period. The more
skewedtoward the right (the second part of the firing period) the
re-
sponse function of the neurons, the more unstable
antiphaselocking becomes.
Large networks of weakly coupled QIF neuronsThe stability
analysis of the asynchronous state of a large networkof neurons
coupled all-to-all receiving a noisy input is also sim-plified if
one assumes weak coupling. Using the phase-reductionapproach, it
can be shown (see Appendix) that the asynchronousstate is stable
if:
n ngv ImZn n g ImZnvn) n22Z0
2 0.
(22)
For all integers, n 0, with Zn and vn the nth-Fourier compo-
nents of the function Z() and v(), respectively (Eqs. 13,
15)defined byZn 1/20
2Z()e 2ind and a similar equationfor vn. Iffor some n,n is
negative (respectively positive), it is saidthat the mode of order
n is stable (respectively unstable). For theasynchronous state to
be stable, the modes at any order must bestable. It is unstable
when at least one mode is unstable.
The first two terms in Equation 22 correspond to the effect
ofthe interaction. The first term represents the effect of the
spikesand can be written:
n nq
2 0
2
sinnZd. (23)
The second term, n ng Im(nZnvn), corresponds to the com-bined
effect of the reset of the membrane potential and its subse-quent
subthreshold evolution.
The sign of their sum depends on the parameters, Vr, VTandon the
frequency of the neurons, v. The last term in Equation
22corresponds to the effect of the noise. It is always positive.
There-fore, as should be expected, noise increases the stability of
theasynchronous state (because of the negative sign in front of
thisterm in Eq. 22). The stability of the asynchronous state
dependson the competition between the first two terms and the last
term.In particular, the sign ofn depends on the ratio
2/g. Note thatbecause of the factor n2 in the last term of
Equation 22, thestability of the modes increases rapidly with their
order.
We first consider the case of a noiseless network (i.e., with0).
In Figure 9B, we plotted 1 (dashed line) and 1 (dashed-dotted line)
against Vr/VT. The qualitative behavior of thesequantities is
similar to those ofSsp and Ssub Sr, respectively, fora pair of
neurons. In particular 1 is symmetric (in a semiloga-rithmic scale)
around Vr/VT 1. Moreover, 1 increasesmonotonically from negative
values to positive values whenVr/VTincreases, and it changes sign
at Vr/VT 1.This can beeasily explained. Indeed, for Vr/VT 1
(respectively, Vr/VT 1), the functionZ() is skewed
toward(respectively)where sin() 0 [respectively sin() 0];
therefore, for n 1, theintegral in Equation 23 is negative
(respectively positive) and1 0(respectively1 0). In
particular,becausefor Vr/VT 1,Z() is
symmetric around , the integral in Equation 23 vanishes forn 1
[becausesin() sin()]. The signof1 (solid line) isprimarily
determined by the sign of 1. It is negative for small
Figure 9. Stability of the asynchronous state of QIF networks at
weak coupling in the
ab-senceofnoise.ResultsfromtheanalyticalstudyoftheQIFmodel.Timeconstant
0 10msec.The size of the spikes is 1. A, A pair of identical
neurons. The quantity () is plotted(solidline). Thethree
contributions tothis quantity (Eq. 21)are also plotted:Ssub (dotted
line),
Sr (dashed-dotted line), and Ssp (dashed line). The
double-dotted-dashed line corresponds toSr Ssub. All of these
quantities were computed from Equations 21, 13, and 15. Note
thelogarithmicscaleonthex-axis.Theneuronsfireat50Hz.
B,Alargenetworkofall-to-allweaklycoupledidenticalneurons:thequantity1.Theasynchronousstateisstablewithrespecttothemode
n 1 if1 0. The quantities1 (dashed line) and1 (double-dotted-dashed
line)are also plotted. The firing rate of the neurons is 50 Hz. C,
The phase diagram of antiphaselocking fora pair of
identicalneuronsin theweakcouplinglimit.Notethe logarithmic scale
onthe x-axis. The solid lines separate the region in which
antiphase locking is unstable (in the
middle) from the regions in which it is stable (on the left and
in the small region on the right).TheparametersVr and
VTarevaried,whereastheresetdepthiskeptconstant: VT Vr 3.Theboundary
linesare defined by thecondition: () 0.
D,Thephasediagramforthestabilityof the asynchronous state in a
large network of all-to-all coupled neurons. The solid line is
theboundary between the domains in which the asynchronous state is
stable (to the left) andunstable (to the right).
6288 J. Neurosci., July 16, 2003 23(15):6280 6294 Pfeuty et al.
Electrical Synapses, Intrinsic Currents, and Synchrony
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
10/15
Vr/VTand becomes positivefor Vr/VT 1. Thereforethemoden
1isstableonlyifVr/VTis small enough.Similaranalysis canbeperformed
for the modes n 1. It reveals that ifVr/VT is not toolarge, the
mode n 1 determines the stability of the asynchronousstate.
The phase diagram for the stability of the asynchronous stateas
a function of Vr/VT and the firing frequency is plotted inFigure
9D. It is very similar to the phase diagram for the stability
of the antiphase state for a pair of neurons (Fig. 9C). The
onlyqualitative difference between these two phase diagrams is
thatfor a pair of neurons, antiphase locking is stable at low
firing ratesand very large Vr/VT, whereas for a large network, the
asyn-chronous state is unstable in that limit. Therefore, we can
con-clude that neurons coupled with electrical synapses will be
moreeasily synchronized if their phase-response functions are
skewedto the right than to the left.
The phase diagram for the stability of the asynchronous stateis
plotted in Figure 10 for different levels of noise. For
large-enough firing rates, the instability lines for 2/g 0 and
2/g0.01 are very close to each other. The distance between the
twolines increases when the frequency decreases. At some value,
*, the two lines separate completely. At *, the line for 2
/g0.01 changes direction and continues toward the right of
thephase diagram (large Vr/VT). In contrast, the line for
0continues toward the left of the diagram. In particular,
for2/g0.01, the asynchronous state is stable for any value
ofVr/VTforsmall-enough firing rates. As a matter of fact, one can
show ana-lytically that the asynchronous state is stable for any
value ofVr/VT if the firing rate is smaller than a critical value,
*
(2/g),which vanishes with 2/g. The size of the region in which
theasynchronous state is stable increases with the noise level, as
ex-pected. In particular,
*, and the frequency range in which the
asynchronous state is stable for all Vr/VT, increases with
2/g.
How synchrony depends on the ratio Vr/VT at fixed fre-
quencies can be deduced from the phase diagram in Figure 10.For
low and moderate noise levels (e.g., 2/g 0.01 and 2/g0.05), we find
three generic behaviors as a function of Vr/VT:
(1) At low firing rates, the asynchronous state is stable for
allVr/VT. (2) At high firing rates, the asynchronous state is
stablefor small Vr/VTand becomes unstable when Vr/VTincreases.(3)
In some intermediate range of firing rates, the stability of
the
asynchronous statevaries nonmonotonously with Vr/VT: whenVr/VT
increases from 0, the asynchronous state is first stable,then
unstable, and stable again for large Vr/VT. The size of
theintermediate region in which the asynchronous state is
unstabledecreases when the noise level increases.
Figure 10 also allows us to predict how the stability of
theasynchronous state depends on the frequency for fixed Vr/VT.For
small-enough Vr/VT, the asynchronous state is stable at allfiring
rates. The range of Vr/VT where this happens is largerwhen the
noise is stronger. For large-enough Vr/VT, the asyn-chronous state
is stable at low firing rates and becomes unstablewhen the firing
rate is large enough. If the noise is not too strong,the stability
of the asynchronous state varies nonmonotonously
in some intermediate range ofVr/VT 1. In this domain,
theasynchronous state is stable at low rates, loses stability when
therate increases, but becomes stable again at high firing
rates.
Interpretation of the simulation results of theconductance-based
networkOur analysis of the synchrony properties of QIF networks
showsthat the stability of the asynchronous states depends
crucially onthe shape of the phase-response function,Z(), and in
particularon the location of its maximum. This suggests that the
desyn-chronization effect of the persistent sodium and the
synchroni-zation effect of the potassium currents revealed by our
numericalsimulations may be related to the way these currents shape
the
phase-response functions of the neurons.The function, Z(), for
our conductance-based neuronmodel is plotted in Figure 11 for
different values of gK, gKs, and
Figure 10. Stability of the asynchronous (AS) state in a large
network of all-to-all
weaklycoupledQIFneuronsinthepresenceofnoise.Timeconstant
010msec.Thesizeofthespikesis 1. Thesolidlineseparates theregionin
whichthe asynchronousstateis stablefromthe
regioninwhichitisunstablefor2
/g 0.05.Thedashedlineisfor2
/g
0.01.Forcompar-ison,theinstabilityboundaryisalsoplottedforthenoiselesscase(dottedline).TheparametersVr
and VTare varied, but the reset depth is kept constant: VT Vr 3.
Note the logarithmicscale on the x-axis.
Figure 11. The phase-response functions depend on the intrinsic
currents. The
conductance-based model is defined in Materials and Methods. The
phase-response functionwas computed for the same four sets of
intrinsic conductances as in Figure 1 using XPPAUTsoftware
(Ermentrout, 2002). For each set of parameters, the external
current is such that thefiring frequency of the neuron is 50 Hz. A,
Control cases: gK 9 mS/cm
2, gNaP gKs 0.Externalcurrentis Iext 1.10A/cm
2. B,TheconductanceofthedelayedrectifierisreducedtogK 2.5
mS/cm
2 (gNaP 0, gKs 0 as in the control case). External current is
Iext 0.48A/cm 2. C, gNaP 0.2 mS/cm
2 [other currents as in the control (gK
9 mS/cm 2, gKs
0)].Externalcurrentis Iext 0.55A/cm
2. D,TheparametersareasinB,butgKs 0.2mS/cm2.
External current is Iext 4.88A/cm2.
Pfeuty et al. Electrical Synapses, Intrinsic Currents, and
Synchrony J. Neurosci., July 16, 2003 23(15):62806294 6289
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
11/15
gNaP. In Figure 11AC, Z() 0 for all , except in a very
tinyregion after the spike peak. In Figure 11 D, the region of
negative
Z() is larger. However, in all four cases, negative values
ofZ()are much smaller than positive values. This means that most
ofthe time, a small and brief depolarizing perturbation
advancessubsequent firing of action potentials.
In allof thecases presentedin Figure11, the functionZ()has
only one maximum, whose location depends on the
intrinsiccurrents. Comparison of Figure 11A with Figure 11 B shows
thatthe maximum ofZis shifted to the right when gK increases,
andthat the global shape of the phase-response function is
moreskewed to the right (respectively to the left) for large
(respectivelysmall) values of gK. The slow potassium current has an
effectsimilar to that seen for the delayed rectifier current. It
distorts thephase-response function toward the second half of the
firing pe-riod. Therefore, increasing the potassium conductances
has asimilar effect on the phase-response functionof the
conductance-based neuron as increasing Vr/VTdoes for the QIF
neuron. Incontrast, the persistent sodium current shifts the peak
of the re-sponse to the left and skews the shape of the
phase-responsefunction in that direction, as seen by comparing
Figure 11A withFigure 11C. Hence, increasing gNaP in the
conductance-basedmodel and decreasing Vr/VT in the QIF model have
the samequalitative effect.
We are now able to understand the results of our
numericalsimulations. In Figure 6, A and B, the network state is
asynchro-nous for small potassium conductances (both
delay-rectifier andslow-potassium) and becomes synchronous when
these conduc-tances are increased. A similar behavior is found in
the QIF net-work forv v
*(): theasynchronous state is stablewhen Vr/VT
is small, and the asynchronous state is destabilized when
Vr/VTincreases enough. This stems from the fact that the
qualitativechanges in the phase-response function of the
conductance-based neuron when gK increases and the QIF neuron
when
Vr/VTincreases are qualitativelysimilar. Conversely,
increasingthe conductance of the persistent sodium current has the
sameeffect on the phase-response function of the neuron as a
decreaseofVr/VT. This explains the results of Figure 6C.
The phase diagram of Figure 10 also explains the
differentbehaviors we found in our simulations for fixed-current
conduc-tances when the frequencyvaries (Fig. 7) in presence of
noise. Forinstance, in the control case, the response function is
skewed tothe right. This is equivalent to Vr/VT 1 for the QIF
model.This explains the monotonous transition from the
asynchronousstate at low firing rates to synchrony when the firing
rate in-creases, as shown in Figure 7, A and D. The
nonmonotonousbehavior in Figure7, B and C, can also beexplained. In
Figure 7C,
gK is smaller than in the control model. The corresponding
effec-tive value ofVr/VTis in the intermediate range, where the
asyn-chronous state changes stability nonmonotonously with the
fre-quency. A similar argument can account for the results in
Figure7B, but here the change in the phase-response function is
attrib-utable to the persistent sodium current.
DiscussionHow intrinsic currents affect synchrony in networks
ofneurons connected by electrical synapsesIn this work, we have
derived explicit rules for the stability of theasynchronous state
in networks of neurons interacting via elec-trical synapses. We
have shown analytically how the shape of the
PRF, which depends on the model parameters, determines
thestability of the asynchronous state in QIF networks. We
haveextended these results to provide a unified framework
explaining
the diversity of behavior found in our numerical simulationswhen
potassium and sodium conductances are changed.
One can understand intuitively how potassium and sodiumcurrents
shape the PRF. The current, IK, increases the refractori-ness of
the neuron; therefore, it reduces its responsiveness tosmall
depolarization after a spike. A similar effect, but strongerand
more lasting, occurs with IKs. This intuitively explains why
we found that largerconductances of these currents skew
thePRFtoward the second half of the firing period. Similar effects
havebeen found by Ermentrout et al. (2001) for M current and
AHPpotassium current. The current INaP is an inward current. It
isalready activated near rest, and depolarizing perturbations
am-plify this activation. Hence INaP increases the responsiveness
ofthe neuron after a spike and shifts the maximum of the
responsefunction towardthe first half of theperiod, as we have
found here.
We predict that potassium currents, like IK and IKs, tend
topromote synchrony of neurons coupled via electrical synapsesand
that, in contrast, INaP tends to oppose it. Our other predic-tions
concern the dependency of the synchrony level with thefiring rate.
We expect that these conclusions do not depend onthe particular
models we have chosen for the intrinsic currents.
Assumption of weak couplingThe stability of the asynchronous
state of a large network of all-to-allconnected QIFneurons can be
calculated analyticallyin theabsence of noise for any coupling
strength using the populationdensity method (Abbott and van
Vreeswijk, 1993; Hansel andMato, 2003). An alternative approach is
the phase-reductionmethod. Although it is only mathematically
justified for weakinteractions, it leads to nontrivial results that
remain valid in areasonable range of interaction strengths.
Moreover, it isstraightforward to study the effect of noise in this
framework.These were themotivations to usethe phase-reductionmethod
inthe framework of which all of the analytical results of this
paper
were derived.At finite coupling strength, an additional
instability of the
asynchronous state appears at low firing rates and small
Vr/VT.It corresponds to the impossibility of controlling low firing
rateswhen the coupling is too strong and the AHP is too small. This
isbecause for a small AHP, the recurrent electrical synapse
interac-tions tend to depolarize the neurons on the average, and
thisincreases their firing rates. This instability is similar to
the rateinstability in networks of excitatory neurons (Hansel and
Mato,2003).
A detailed analysis of the QIF network at finite
couplingstrength shows that in a broad range of coupling strengths,
thephase diagrams for two coupled neurons and for large
networks
are similar to those derived here, except for the additional
rateinstability line. A full report of this analysis will be
publishedelsewhere.
Simulation results were performed here for a coupling
con-ductance,g 0.005 mS/cm2. This corresponds to coupling
coef-ficients (Amitai et al., 2002) CC 5% and to a spikelet size
of0.5 mV (Fig. 3). We have verified that similar results have
beenfound for conductances four times larger (data not
shown).Therefore the conclusions of the present work are relevant
forelectrical synapse conductances in the physiological range
(Ga-larreta and Hestrin, 2001, 2002; Amitai et al., 2002).
Related works
Stable antiphase locking has been demonstrated in previous
stud-ies for a pair of neurons coupled via electrical synapses.
Shermanand Rinzel (1992) used numerical simulations to show that
this
6290 J. Neurosci., July 16, 2003 23(15):6280 6294 Pfeuty et al.
Electrical Synapses, Intrinsic Currents, and Synchrony
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
12/15
nonintuitive phenomenonoccurs in a simple
conductance-basedmodel. However, they did not relate it to the
intrinsic propertiesof their model. Antiphase locking was found by
Han et al. (1995)in simulations of coupled MorrisLecar oscillators.
For thismodel, they computed the effective phase interaction, ,
andshowed that their simulation results could be predicted from
it.Ourwork further clarifies theconditions of stableantiphase
lock-
ing in that it gives general qualitative criteria that the PRF
mustsatisfy, which allows us to predict how it depends on
intrinsiccurrents.
Phase locking of neurons connected by electrical synapses
hasbeen also investigated analytically (Chowand Kopell, 2000;
Lewisand Rinzel, 2003). In both studies, neurons were modeled in
thesubthreshold range as leaky integrators, but the spikes were
de-scribed in different ways. Chow and Kopell (2000) used a
phe-nomenological model to describe the time course of
spikes,whereas Lewis and Rinzel (2003), like us, modeled them with
a-function. Both studies found stable antiphase locking at
lowfiring rates and destabilization for large-enough firing rates.
Thiscan be understood using Equations 1821 as follows. For a
pas-sive integrator,Z(),Z() 1/[I
extT] exp(T/2
m),whereT is
the firing rate, Iext is the external current, and m is the
neuronalmembrane time constant (Kuramoto, 1991; Hansel et al.,
1995).The value ofZdiverges exponentially with the period at all ,
asdo Ssub and Sr. Detailed analysis reveals that the most divergent
ofthese terms is Ssub 0. Hence, for low-enough firing rates,() 0.
The term Ssp is positive (Z() 0) and decreaseswith T, whereas Ssub
and Sr increase. Hence, for a large-enoughfrequency, () changes
sign. Therefore for passive integrators,antiphase locking is stable
at low rates and loses stability as thefiring rate increases. In
contrast, we have found that the QIFmodel behaves differently than
the LIF except in the limit of verylarge Vr/VT. Therefore our
results show that the predictions ofChow and Kopell (2000) and
Lewis and Rinzel (2003) may be
relevant only for neurons with strong potassium currents.
Relationship to experimentsElectrical synapses connect fast
spiking (FS) as well as low-threshold spiking (LTS) interneurons
(Kawaguchi and Kubota,1997; Gibson et al., 1999). Recently, the
synchrony level of pairsof LTS and FS cells (characterized by the
amplitude of the peak oftheir spikes cross-correlation) has been
studied as a function ofthe firing rate v(Mancilla et al., 2002).
It has been found that forFS cells, synchrony increases with v but
decreases for LTS cells.Our results may explain this difference if
one assumes that INaP isstronger in LTS than in FS cells and/or
that potassium conduc-tances are stronger in FS than in LTS cells.
This assumption is
compatible with thefact that FS andLTS cells have
differentfiringpatterns: in response to current injection, the
action potentials ofFS but not LTS cells are followed by a
pronounced AHP, whichmay reflect stronger potassium conductances in
FS than in LTScells. A test of this explanation would be to show
that the PRFs ofthese LTS and FS cells have different shapes.
The dynamic clamp technique (Sharp et al., 1993) makes
itpossible to artificially manipulate the intrinsic currents of
theneurons and to couple two neurons via artificial electrical
syn-apses. It provides a nice way of further testing our
predictions.Therefore, it enables systematic verification of the
effects of so-dium and potassium currents on the synchronization
propertiesof a pair of neurons (Fig. 4) and how they correlate with
their
PRF.Several studies in the mouse and the rat have reported
thepresence of electrical synapses in hypoglossal and other
inspira-
tory brainstem and phrenic motoneurons (Mazza et al.,
1992;Rekling and Feldman, 1998). Recently, it has been found
thatinspiratory motoneuron synchrony is modulated by
electricalsynapses: blocking these synapses increases their level
of syn-chrony (Bou-Flores and Berger, 2001). An explanation of
thesurprising findings of Bou-Flores and Berger (2001) is
suggestedby ourwork. Indeed, since strong INaP (Powers and Binder,
2003)
is present in these neurons, electrical synapses can reduce
thelevel of synchrony of their activity compared with the case
inwhich electrical synapses are blocked. Synchrony in the
lattersituation would be attributable, for example, to GABAergic
syn-aptic interactions between the neurons or to a
spatiallycorrelatedtime-dependent external input. As a matter of
fact, we have ver-ified that this effect occurs in numerical
simulations of our net-work model with GABAergic synapses added. A
detailed study ofthis phenomenon will be reported elsewhere.
AppendixThe IFcurve, the membrane potential, and the
phase-response function of the QIF neuronThe subthreshold membrane
potential, v(t), of a QIF neuronsatisfies the dynamic equation:
0dv
dt v2 Iext Ic. (24)
Integrating this differential equation, one finds that its
generalsolution is for Iext Ic:
vt Iext Ic tan tIext Ic0 , (25)where is a constant of
integration. The condition that at t 0the membrane potential of the
neuron is at its reset value, Vr,
determines . One finds:
tan1 VrIext Ic. (26)The function v(t) increases monotonously to
infinity. Therefore,after some time T, v(t) reaches the threshold,
VT. Atthattime, theneuron fires an action potential, and v(t) is
instantaneously resetto Vr. Subsequently, v(t) starts again to
increase until it againreaches the thresholdafter another time T.
Therefore, the neuronis firing periodically. The condition: v(T) VT
determines thefiring period. One finds:
T 0tan
1
VT
Iext Ic
tan1
Vr
Iext IcIext Ic . (27)The IFcurve of the QIF neuron is given byv
1/T(Iext). Whenthe subthreshold membrane potential, v(t) reaches
VT, the neu-ron fires an action potential represented by a
-function:
Vt vt spikes
(t tspike), (28)
where measures the integral over time of the suprathresholdpart
of an action potential.
The response function Z is the phase resetting curve of the
neuron (Kuramoto, 1984; Hansel et al., 1993; Rinzel and
Ermen-trout, 1998) in the limit of vanishing small perturbations of
themembrane potential. For the case of one-dimensional models,
it
Pfeuty et al. Electrical Synapses, Intrinsic Currents, and
Synchrony J. Neurosci., July 16, 2003 23(15):62806294 6291
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
13/15
can be written as dvd 1
(Kuramoto, 1991; Hansel et al.,
1995). Using Equation 24, one finds for the QIF model:
Z 0
vT22
Iext Ic
. (29)
Phase interaction for electrical synapsesThe phase coupling
function, (), between two identical neu-rons, i andj, interacting
synaptically depends on the phase differ-ence, i j , of the two
neurons and is given by (Kuramoto,1984; Hansel et al., 1993, 1995;
Kopell and Ermentrout, 2002):
i j 1
20
2
Zu iIsyn(u i, u j)du,
(30)
where Z is the phase-response function of the neurons andIsyn(i
, j) is the synaptic current coming from the presynapticneuron j to
the postsynapticneuroni. For electrical synapses, thiscurrent
is:
Isyn(i, j) gVi Vj. (31)
Phase locking of a pair of weakly interacting neuronsWhen, in
the absence of noise, two identical neurons interactweakly in a
symmetric way, they reach a phase-locked state atlarge time. This
is easilyshownin thephase-reduction framework(Hansel et al., 1993,
1995; Van Vreeswick et al., 1994; Kopell andErmentrout, 2002). The
phases, 1 and 2 , of the two neurons
satisfy the two coupled equations:
d1
dt v 1 2 (32)
d2
dt v 2 1. (33)
Subtracting these equations, one finds:
d
dt )2(), (34)
where is thephase shift between thetwo neurons, 12 ,
and
()
1
2(() ()). At large time, the phase shift, ,reaches a fixed point
(i.e., it is such that d/dt 0) and the twoneurons become locked
with a phase shift that satisfies thecondition:
0. (35)
Because of the symmetry of the system and the 2periodicity ofthe
function , there are always at least two solutions to
thisequation.One is the in-phase solution, 0,and the other is
theantiphase solution: . Other solutions with 0 mayalso exist.
However, only those that are stable can be reached atlarge time.
Linearizing Equation 34 around these solutions, onefinds that the
stability condition is:
d
d 0, (36)
where combining Equations 30 and 31:
g
20
2
ZuVu Vudu. (37)
Differentiating this function with respect to , one finds:
g
20
2
ZuVu du. (38)
The membrane potential V can be expanded in its
components(spike, reset, and subthreshold component) as:
V u v u Vr VTu vu, (39)
which allows us to rewrite ():
g
20
2
Zuu
Vr VT0
2
Zuu 0
2
Zuvu du. (40)Integrating by parts the first term and using the
properties of the-function, we finally obtain:
g v dZd (VT Vr)Z()
1
2
0
2
Zuvu du
. (41)
n neurons: stability of the asynchronous state for the
QIFnetwork with noiseThe stability analysis of the asynchronous
state can be investi-gated for a network of phase oscillators in
the presence of whitenoise (zero mean and variance D). For a
general phase-couplinginteraction, (), one can show (Kuramoto,
1984) that the asyn-chronous state is stable if for all n 0:
Real(n) 0, (42)
where we have defined:
n in0 n n2D, (43)
with n , the nth-Fourier component of the 2periodic
function():
n 1
20
2
e2in d. (44)
For a network of QIF neurons interacting with electrical
syn-apses (Eqs. 24, 28) the nth-Fourier component of phase
interac-tion has the form:
n gZnvn v, (45)
whereZn and vn are the nth-Fourier components of the
functionsZ() and v() defined in Equations 29 and 25. The variance
of
6292 J. Neurosci., July 16, 2003 23(15):6280 6294 Pfeuty et al.
Electrical Synapses, Intrinsic Currents, and Synchrony
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
14/15
the noise, D, depends on the average of the phase-response
func-tion,Z0 1/20
2Z()d andisgivenbyD 2Z02 (Kuramoto,
1984), where 2 is the variance of the white noise in the full
QIFmodel. This yields the stability condition for the
asynchronousstate:
ng v ImZn ng ImZnvn n2Z0
2 0 (46)
Note that Z0 depends on the parameters Vr, VTand on the
firingfrequency of the neurons. Therefore, the desynchronizing
effectof the noise depends on these parameters. In particular,
because
Z0 decreases when the firing rate increases, the noise is
moreefficient at stabilizing the asynchronous state at low firing
ratesthan at high firing rates.
ReferencesAbbott LF, van Vreeswijk C (1993) Asynchronous state
in networks of
pulse-coupled oscillators. Phys Rev E 48:14831490.Abbott LF,
Hooper SL, Marder E (1991) Oscillating networks: control of
burst duration by electrically coupled neurons. Neural
Comput3:487497.
Alvarez LF, Chow C, van Bockstaele EJ, Williams JT (2002)
Frequency-
dependent synchrony in locus coeruleus: role of electrotonic
coupling.Proc Natl Acad Sci USA 99:40324036.
Amitai Y, Gibson JR, Patrick A, Ho B, Connors BW, Golomb D
(2002)Spatial organization of electrically coupled network of
interneurons inneocortex. J Neurosci 22:41424152.
Beierlein M, Gibson JR, Connors BW (2000) A network of
electrically cou-pled interneurons drives synchronized inhibition
in neocortex. Nat Neu-rosci 3:904910.
Bou-Flores C, Berger AJ (2001) Gap junctions and inhibitory
synapsesmodulate inspiratory motoneuron synchronization. J
Neurophysiol85:15431551.
Chow CC, Kopell N (2000) Dynamics of spiking neurons with
electricalcoupling. Neural Comput 12:16431678.
Christie MJ, WilliamsJT, NorthRA (1989) Electrical coupling
synchronizessubthreshold activity in locus coeruleus neurons in
vitro from neonatal
rats. J Neurosci 9:35843589.Cymbalyuk GS, Nikolaev EV, Borisyuk
RM (1994) In-phase and antiphaseself-oscillations in a model of two
electrically coupled pacemakers. BiolCybern 71:153160.
Deans MR, Gibson JR, Sellitto C, Connors BW, Paul DL (2001)
Synchro-nous activity of inhibitory networks in neocortex requires
electrical syn-apses containing connexin36. Neuron 31:477 485.
Draghun A, Traub RD, Schmitz D, Jefferys JGR (1998) Electrical
couplingunderlies high-frequency oscillations in the hippocampus in
vitro.Nature394:189192.
Erisir A, Lau D, Rudy B, Leonard CS (1999) Function of specific
K chan-nels in sustained high-frequency firing of fast-spiking
neocortical inter-neurons. J Neurophysiol 82:24762489.
Ermentrout B (1996) Type I membranes, phase resetting curves and
syn-chrony. Neural Comput 8:9791001.
Ermentrout B (2002) Simulating, analyzing, and animating
dynamical sys-tems: a guide to Xppaut for researchers and students
(software, environ-ments, tools). Philadelphia: Society for
Industrial and AppliedMathematics.
Ermentrout B, Kopell N (1986) Parabolic bursting in an excitable
systemcoupled with a slow oscillation. SIAM J Appl Math
46:233253.
Ermentrout B, Pascal M, Gutkin B (2001) The effect of spike
frequencyadaptation and negative feedback on the synchronization of
neural oscil-lators. Neural Comput 13:12851310.
French CR, Sah P, Buckett KJ, Gage PW (1990) A voltage-dependent
per-sistent sodium current in mammalian hippocampal neurons. J
GenPhysiol 95:11391157.
Furshpan EJ,Potter DD (1959) Transmissionat
thegiantmotorsynapsesofthe crayfish. J Physiol (Lond)
145:289325.
Galarreta M, Hestrin S (1999) A network of fast spiking cells in
the neocor-tex connected by electrical synapses. Nature
402:7275.
Galarreta M, Hestrin S (2001) Electrical synapses between
GABA-releasingneurons. Nat Neurosci 2:425433.
GalarretaM, Hestrin S (2002) Electricaland chemicalsynapsesamong
parv-
albumin fast-spiking GABAergic interneurons in adult mouse
neocortex.
Proc Natl Acad Sci USA 19:1243812443.
Gibson JR,Beierlein M, Connors B (1999) Twonetworksof inhibitory
neu-
rons electrically coupled. Nature 402:7579.
Golomb D, Hansel D (2000) The number of synaptic inputs and the
syn-
chrony of largesparse neuronal networks.Neural Comput
12:10951139.
Golomb D, Rinzel J (1994) Clustering in globally coupled
inhibitory neu-
rons. Physica D 72:259282.
Golomb D, Hansel D, Mato G (2001) Theory of synchrony of
neuronalactivity. In: Handbook of biological physics, Vol 4:
Neuro-informatics
and neural modeling (Gielen S, Moss M, eds), pp 887968.
Amsterdam:
Elsevier.
Han SK, Kurrer C, Kuramoto Y (1995) Dephasing and bursting in
coupled
neural oscillators. Phys Rev Lett 75:31903193.
Hansel D, Mato G (2003) Asynchronous states and the emergence of
syn-
chrony in largenetworksof interacting excitatoryand
inhibitoryneurons.
Neural Comput 15:156.
Hansel D, Sompolinsky H (1992) Synchrony and computation in a
chaotic
neural network. Phys Rev Lett 68:718721.
Hansel D, Mato G, Meunier C (1993) Phase dynamics for weakly
coupled
Hodgkin-Huxley neurons. Europhys Lett 23:367372.
Hansel D, Mato G, Meunier C (1995) Synchrony in excitatory
neural net-
works. Neural Comput 7:307337.
HormuzdiSG, Pais I, LebeauFEN, TowersSK, Rozov A, Buhl E,
WhittingtonMA, Monyer H (2001) Impaired electrical signaling
disrupts gamma
frequency oscillations in connexin 36-deficient mice. Neuron
31:487495.
Kawaguchi Y, Kubota Y (1997) GABAergic cell subtypes and their
synaptic
connections in rat frontal cortex. Cereb Cortex 7:476486.
Kepler TB, Marder E, Abbott LF (1990) The effect of electrical
coupling on
the frequency of model neuronal oscillators. Science
248:8385.
Kiehn O, Tresch MC (2002) Gap junctions and motor behavior.
Trends
Neurosci 25:108115.
Kita H, Kosaka T, Heizmann CW (1990)
Parvalbumin-immunoreactive
neurons in the rat neostriatum: a light and electron microscopic
study.
Brain Res 536:115.
Kopell N, Ermentrout GB (2002) Mechanisms of phase-locking and
fre-
quency control in pairs of coupled neural oscillators. In:
Handbook of
dynamical systems, Vol 2 (Fiedler B, ed), pp 555. Amsterdam:
Elsevier.Kuramoto Y (1984) Chemical oscillations, waves and
turbulence. New
York: Springer.
Landisman CE, Long MA, Beierlein M, Deans MR, Paul DL, Connors
BW
(2002) Electrical synapses in the thalamic reticular nucleus. J
Neurosci
22:10021009.
Latham PE, Richmond BJ, Nelson PG, Nirenberg S (2000) Intrinsic
dynam-
ics in neuronal networks. I. Theory. J Neurophysiol 83:808
827.
Lewis T, Rinzel J (2003) Dynamics of spiking neurons connected
by both
inhibitory and electrical coupling. J Comput Neurosci
14:283309.
Llinas R, Yarom Y (1986) Oscillatory properties of guinea-pig
inferior oli-
vary neurones and their pharmacological modulation: an in vitro
study.
J Physiol (Lond) 376:163182.
Mancilla JG, Lewis T,PintoDJ, RinzelJ, Connors BW (2002)
Firingdynam-
ics of single and coupled pairs of inhibitory interneurons in
neocortex.
28:840.13.Mann-Metzer P, YaromY (1999) Electrical coupling
interacts with intrinsic
properties to generate synchronized activity in cerebellar
networks of
inhibitory interneurons. J Neurosci 19:32983306.
Mazza A, Nunez-Abades PA, Spielmann JM, Cameron WE (1992)
Anatom-
ical and electronical coupling in developing genioglossal
motoneurons of
the rat. Brain Res 598:127137.
Meunier C (1992) The electrical coupling of two simple
oscillators: load and
acceleration effects. Biol Cybern 67:155164.
Neltner L, Hansel D, Mato G, Meunier C (2000) Synchrony in
heteroge-
neous networks of spiking neurons. Neural Comput
12:16071642.
Perez-Velazquez JL, Carlen PL (2000) Gap junctions, synchrony
and sei-
zures. Trends Neurosci 23:6874.
Pfeuty B, Golomb D, Mato G, Hansel D (2002) Potassium currents
modu-
late synchronization in networks of neurons connected with
electrical
synapses. 28:840.15.Powers RK, Binder (2003) Persistent sodium
and calcium currents in rat
hypoglossal motoneurons. J Neurophysiol 89:615624.
Pfeuty et al. Electrical Synapses, Intrinsic Currents, and
Synchrony J. Neurosci., July 16, 2003 23(15):62806294 6293
-
8/3/2019 Benjamin Pfeuty et al- Electrical Synapses and
Synchrony: The Role of Intrinsic Currents
15/15
Rekling JC, Feldman JL (1998) Bidirectional electrical coupling
between in-spiratory motoneurons in the newborn mouse nucleus
ambigus. J Neu-rophysiol 78:385 405.
Rinzel J, Ermentrout B (1998) Analysis of neural excitability
and oscilla-tions. In: Methods in neuronal modeling: from ions to
networks, Ed 2(Koch C, Segev I, eds), pp 251292. Cambridge, MA:
MIT.
Sharp AA, ONeil MB, Abbott LF, Marder E (1993) The dynamic
clamp:artificial conductances in biological neurons. Trends
Neurosci
16:389394.Sherman A, Rinzel J (1992) Rhythmogenic effects of
weak electrotonic cou-pling in neuronal models. Proc Natl Acad Sci
USA 89:24712474.
Strogatz S (1994) Nonlinear dynamics and chaos. Reading,
MA:Addison-Wesley.
Tamas G, Buhl EH, Lorincz A, Somogyi P (2000) Proximally
targetedGABAergic synapses and gap-junctions precisely synchronize
cortical in-terneurons. Nat Neurosci 3:366371.
Traub RD, Kopell N, Bibbig A, Buhl E, Lebeau FEN, Whittington M
(2001)Gap junction between interneuron dendrites can enhance
synchrony ofoscillations in distributed networks. J Neurosci
21:94789486.
Van Vreeswijk C, Abbott LF, Ermentrout GB (1994) When inhibition
notexcitation synchronizes neural firing. J Comput Neurosci
1:313321.
Venance L, Rozov A, Blatow M, Burnashev N, Feldmeyer D, Monyer
H(2000) Connexin expression in electrically coupled postnatal rat
brainneurons. Proc Natl Acad Sci USA 97:1026010265.
Watanabe A (1958) The interaction of electrical activity among
neurons oflobster cardiac ganglion. Jpn J Physiol 8:305318.
6294 J. Neurosci., July 16, 2003 23(15):6280 6294 Pfeuty et al.
Electrical Synapses, Intrinsic Currents, and Synchrony