Embed Size (px)
Citation preview

Eur. J. Biochem. 193,449-457 (1990) L FEBS 1990
Carbohydrate epitope structural elucidation by 'H-NMR spectroscopy of a new Mycobacterium kansasii phenolic glycolipid antigen Martine GILLERON ', Anne VENISSE ', Michel RIVIERE ', Philippe SERVIN' and Germain PUZO' ' Centre de Recherche de Biochimie et Genetique Cellulaires du Centre National de la Recherche Scientifique, Toulouse, France
Service de Resonance Magnetique NuclCaire i haut champ. Interaction moltculaire et reactivitk chimique et photochimique, Centre National de la Recherche Scientifique, Universitk Paul Sabatier, Toulouse, France
(Received February 9/July 9, 1990) - EJB 90 0138
The complete primary structure of the carbohydrate moiety of a new phenolic glycolipid antigen namely PheGl K-IV from Mycobacterium kansasii was successfully established from only one- and two-dimensional 'H-NMR data. Among the scalar two-dimensional techniques, correlated spectroscopy with a 45 O mixing pulse and phase- sensitive double-quantum-filtered correlated spectroscopy were selected, combined with two-dimensional dipolar techniques (nuclear Overhauser effect). These techniques using milligram of quantities native PheGl K-IV allowed the following monoacetylated tetrasaccharide to be proposed for its carbohydrate part:
4-O-Me-a-Manp-(l+ 3)-4-0-Ac-2-0-Me-a-Fucp-(l+3)-2-0-Me-cc-Rhap-(l+3)-2,4-di-0-Me-a-Rhap. The PheGl K-IV shares, with the other phenolic glycolipids isolated from M . kansasii (K-I, K-II), a common
core assigned to the lipid aglycone glycosylated by the monoacetylated trisaccharide part. It differs in the structure of the distal monosaccharide residue.
Most mycobacterial cell walls are endowed with species- specific immunoreactive glycolipids. From their structure, these glycolipids have been classified into three types: the phenolic glycolipids (PheGl), the trehalose-containing lipo- oligosaccharides and the glycopeptidolipids [l - 31. Whatever the glycolipid type, it has been established, in most cases, that the distal disaccharide residue of their oligosaccharide part contains the epitope. Its species-specificity results from the presence of partially 0-methylated sugars and, in some cases, of monosaccharides unique in nature [4 - 61. Thus, with the aim of determining the structure of these glycolipids, it appears that the critical step after their purification was the structural elucidation of their non-ubiquitous oligosaccharide part. This structural problem was solved using both sophisticated and modern analytical tools such as fast-atom-bombardment (FAB) mass spectrometry [7], plasma desorption [8], desorp- tion chemical ionization [9] for molecular mass assignments and FAB-tandem mass spectrometry [ 101 for oligosaccharide sequence determination, combined with one-dimensional (1 D)
Correspondence to G. Puzo, Centre de Recherche de Biochimie et de Genetique Cellulaires du Centre National de la Recherche Scientifique, 118 route de Narbonne, F-31062 Toulouse Cedex, France
Abbreviations. PheGl K-I, major phenolic glycolipid from Mycobacterium kansasii; PheGl K-11, PheGl K-111, PheGl K-IV, quantitatively minor phenolic glycolipids from Mycobacteriurn kansasii; COSY, two-dimensional 'H-NMR correlated spectroscopy; COSY-45, two-dimensional 'H-NMR correlated spectroscopy with a 45' mixing pulse; NOESY, two-dimensional 'H-NMR nuclear Overhauser effect spectroscopy; DQF-COSY, double-quantum- filtered correlated spectroscopy; FAB, fast atom bombardment; Fuc, fucose; Rha, rhamnose; P, pyranose; 2D, two-dimensional; lD, one dimensional; Hex, hexose.
'H-NMR spectroscopy and conventional oligosaccharide analysis. Recently, two-dimensional (2D) 'H-NMR tech- niques were successfully applied to peracetylated mycobacterial phenolic glycolipids to determine their carbo- hydrate structure [ll, 121. This approach had been previously developed by Rivikre [13] with the aim of determining the oligosaccharide structure of a minor phenolic glycolipid, PheGl K-I1 present in small amounts alongside the major PheGl K-I in the Mycobacterium kansasii cell wall.
From the literature it emerges that the combination of 2D techniques have proved useful for the structural elucidation of the oligosaccharide moieties of glycoconjugates. The potential utility of these techniques in obtaining this type of data was first explored in 1982 by the group of Prestegard and Hall [14, 1.51. Thereafter, new 2D techniques were developed and a selection of their successful applications for primary struc- tural elucidation of oligosaccharides are summarized in Dabrowski's review [16].
In the present report, phase-sensitive double-quantum- filtered-correlated spectroscopy (DQF-COSY), correlated spectroscopy with a 45" mixing pulse (COSY-45) and NOE spectroscopy (NOESY) were successfully applied for the structural elucidation of the carbohydrate part of an new phenolic glycolipid antigen (Phe-G1 K-IV) extracted from the M . kansasii cell wall. We demonstrate that the structure of PheGl K-IV differs from that of Phe-GI K-I [lo] by the distal monosaccharide residue: a 4-0-Me-a-Manp instead of the 2,6- dideoxy-4-O-Me-a-~-arabino-Hexp. This glycolipid was pre- viously purified from a crude M . kansasii lipid extract by HPLC using a 5-pm Spherimrb column eluted with a gradient of methanol in chloroform [17]. It was shown by ELISA that its carbohydrate epitope, assigned to the distal disaccharide residue, is different from that of K-I.
450
MATERIALS AND METHODS
Phenolic gl.ycolipid purification
The minor phenolic glycolipid of M . kansasii, Phe-G1 K-IV, was purified as previously described [17]. The native phenolic glycolipid (3 mg) was dried under vacuum, dissolved in C2HC13 (Spin et Techniques, Paris, France) and analysed in a 200mmx5mm NMR tube. For 2D NOESY exper- iments, the solution was degassed under vacuum after freezing the tube in liquid nitrogen.
Proton NMR was performed on a Bruker AM 300 WB instrument. Proton spectra were generally recorded at 50 "C and chemical shifts were referenced to external tetra- methylsilane. The I D spectrum was recorded by using 40" pulses with a recycle delay of 0.3 s and a 2.7-s acquisition time. 64 scans were accumulated. The spectral width used was 3 kHz, which. with a data memory of 16 K expanded to 32 K by zero filling, give a digital resolution of 0.1 8 Hz.
The standard 2D COSY, 2D phase-sensitive DQF-COSY, 2D NOESY pulse sequences supplied by Bruker were employed. 2D spectra (except DQF-COSY) are shown in the absolute value representation as contour plots. Unless stated otherwise, the 211 spectra were obtained with 256 x 1024 matrix data points over a spectral range of 1500-2200 Hz, which were expanded to 1024 x 2048 by zero filling. Reso- lution enhancement in both dimensions was performed by a non-shifted sine-bell function, except for 2D NOESY for which a shift of ;.r4 in the,fi and 7112 in the f2 dimension, and for DQF-COSY for which a shift of nil 6 in both dimensions were used. The parameters used in this work were as follows.
22) C'OSY4.5. 45 mixing pulse of about 2100 Hz spectral width, 64 scansit,, 256 experiments, 3s recycle delay, 2.1 Hz/ point digital resolution.
22) phase-sm.ritivr DQF-COSY. This experiment was per- formed using the time-proportional phase-increment method and the data arc displayed in the phase-sensitive method. The
H-1 OCH:
carbohydrate
H-1
methlne H
1 8 1 1 1 1 1 5.5 5.4 5.3 5.2 5.1 S d 4.9 4.*
parameters were 90'' mixing pulse, 1800 Hz spectral width, 128 scans]tl, 256 experiments, 1.2 s recycle delay, 3.52 Hzj point along Fl and 1.76 Hz/point along F2 digital resolution. Quadrature detections were used in both dimensions and the spectrum was not symetrised.
2 0 NOESY. (a) 2100 Hz spectral width, 64 scans/tl, 256 experiments, 1.2 s recycle delay, 2.1 Hz/point digital reso- lution, 1 s mixing time. (b) 256 x 2048 data matrix size expand- ed to 1024 x 4096 by zero-filling, 1500 Hz spectral width, 80 scans/tl, 256 experiments, 0.8 Hz/point digital resolution, 1 s mixing time.
The representation of PheGl K-IV in Fig. 5 was realized using an Alchemy I1 Tripos Associates (St Louis Mo.) software.
RESULTS
PheGl K-IV I D ' H - N M R studirs
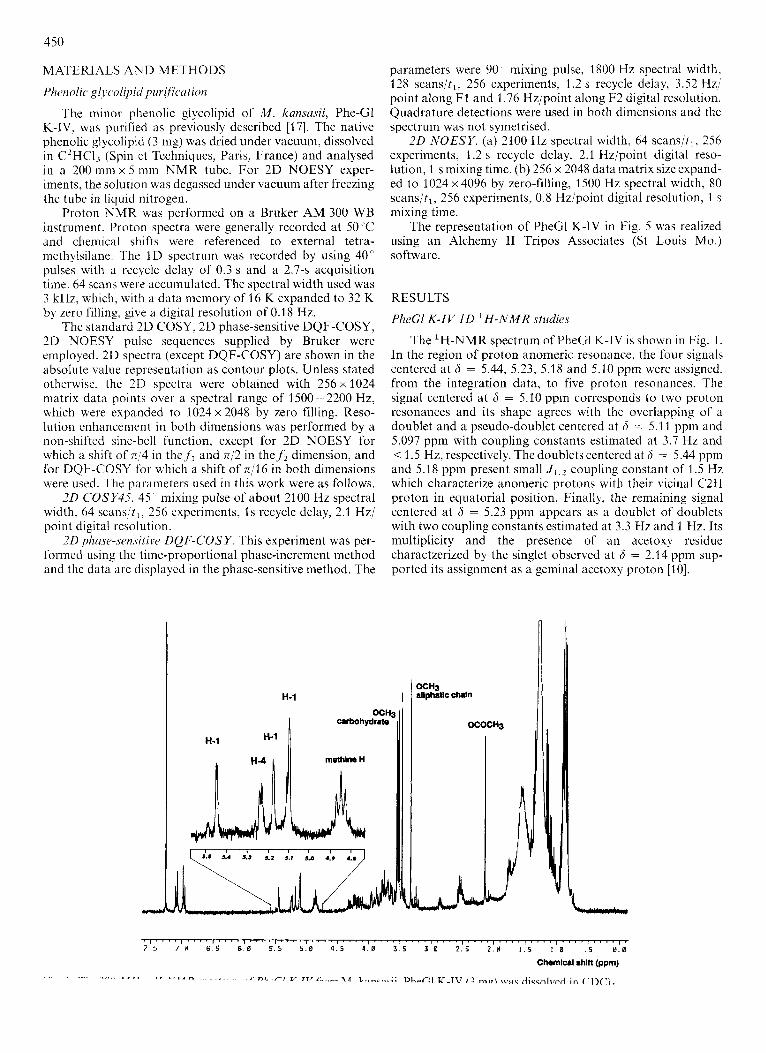
The 'H-NMR spectrum of PheGl K-IV is shown in Fig. 1. In the region of proton anomeric resonance, the four signals centered at 6 = 5.44, 5.23, 5.18 and 5.10 ppm were assigned, from the integration data, to five proton resonances. The signal centered at 6 = 5.10 ppm corresponds to two proton resonances and its shape agrees with the overlapping of a doublet and a pseudo-doublet centered at 6 = 5.1 3 ppm and 5.097 ppm with coupling constants estimated at 3.7 Hz and < 1.5 Hz, respectively. The doublets centered at 6 = 5.44 ppm and 5.18 ppm present small J 1 , 2 coupling constant of 1.5 Hz which characterize anomeric protons with their vicinal C2H proton in equatorial position. Finally, the remaining signal centered at 6 = 5.23 ppm appears as a doublet of doublets with two coupling constants estimated at 3.3 Hz and 1 Hz. Its multiplicity and the presence of an acetoxy residue charactzerized by the singlet observed at 6 = 2.14 ppm sup- ported its assignment as a geminal acetoxy proton [lo].
\ J L

451
By analogy with the PheCl K-I ‘H-NMR spectrum [lo], the five singlets located in the region 3.4-3.6 ppm are as- signed to five methoxy groups located on the oligosaccharide part while the singlet at 6 = 3.33 ppm (methoxy group of the aliphatic chain), the multiplet at 4.85 ppm (methine protons) and the two doublets centered at 6 = 6.95 ppm and 7.10 ppm (aromatic protons) characterize a diacylated phenol- phthiocerolic aglycone.
Thus, a monoacetylated tetrasaccharidic structure with five methoxy residues can be proposed for the oligosaccharide part of PheGl K-IV.
PheCl K-JV 2 0 COSY-45 and 2 0 phase-sensitive DQF-COSY H-NMR studies
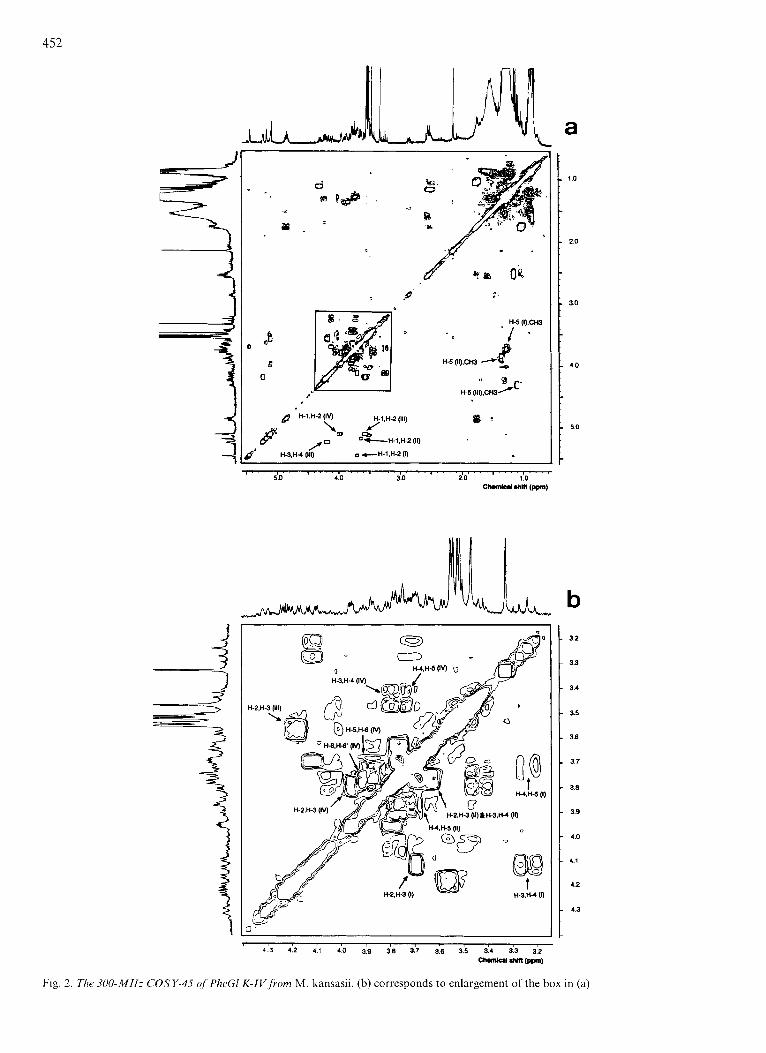
The monosaccharide unit composition of PheGl K-IV and the acetoxy residue location were unambiguously determined using combined COSY-45 and phase-sensitive DQF-COSY analysis of the native glycolipid. A representative spectrum of the COSY-45 is shown in Fig. 2.
Beginning at 6 = 5.44 ppm with the deshielded C1H anomeric proton, we found a cross-peak showing a con- nectivity to the C2H resonance at 6 = 3.68 ppm. From this C2H, a connectivity to the C3H resonance at 6 = 4.11 ppm was established. The C3H signal multiplicity observed in the 1D spectrum is a doublet doublets of (J3,4 = 9.6 Hz, J 2 , 3 = 3.7 Hz) supporting an C2H in equatorial configuration and both C2H and C4H in axial configuration. Continuation of this process allowed the localization of the C4H resonance at 6 = 3.21 ppm. It appears in the I D spectrum as a triplet (J3,4 = J4,5 = 9 Hz) proving that C3H and CSH are trans- diaxial towards C4H. Likewise, C5H was found at 6 = 3.69 ppm. The juncture connectivity end indicated a resonance at 6 = 1.28 ppm in agreement with the chemical shift of a methyl group from a 6-deoxyhexopyranosyl residue. Thus, from this data, it can be deduced that the anomeric proton at 6 = 5.44 ppm belongs to a rhamnopyranosyl (I) residue.
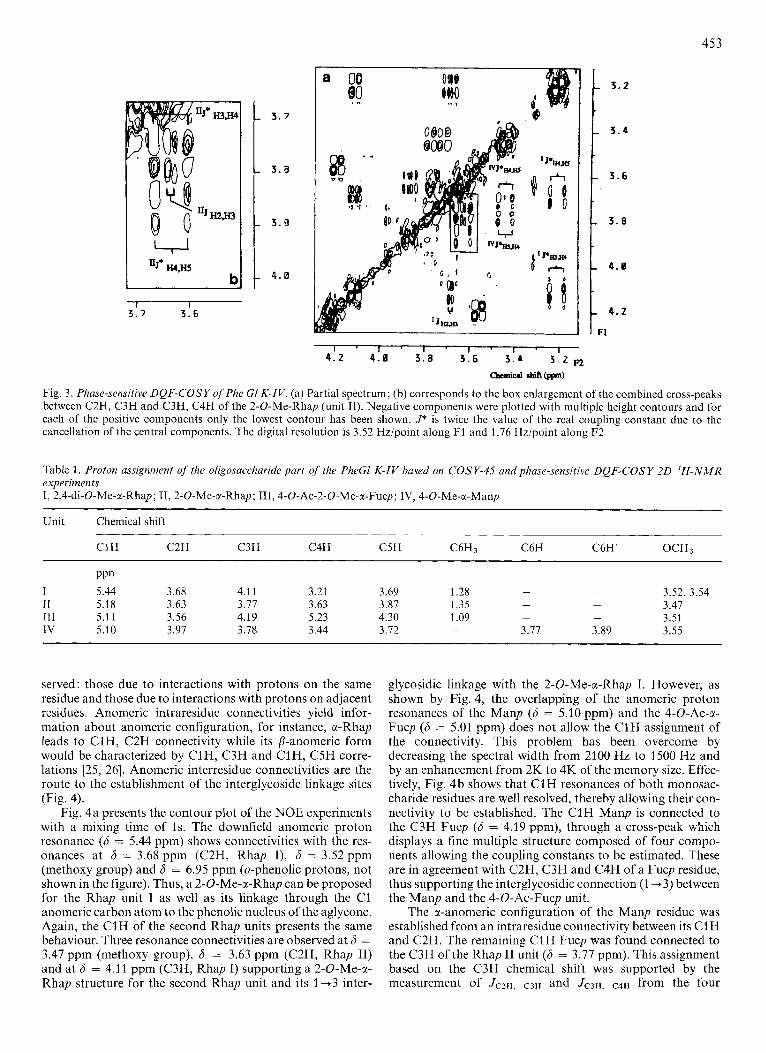
In a similar fashion, starting with the C1H resonance at 6 = 5.18 ppm (unit II), a connectivity was found with the equatorial C2H at 6 = 3.63 ppm and with the C3H resonance at 6 = 3.77 ppm. However, in neither case did the 1D spec- trum allow measurement of their coupling constants. More- over, ambiguities also arise for the chemical shift assignment of the C4H resonance, probably because of the overlapping of C2H, C3H and C3H, C4H cross peaks (Fig. 2). Thus, the structural elucidation of the second monosaccharide unit was achieved using 2D phase-sensitive DQF-COSY experiments [23] (Fig. 3). Again, the cross-peak allowing the C2H,, con- nectivity with the C3H appears complex. However, from its fine structure, i t can be clearly discerned (Fig. 3b) that it is composed of a combination of two cross-peaks allowing the connectivities between the C2H, C3H and C4H resonances. Along the F2 axis, the combined cross-peak is composed of four columns from which the 4 Hz and 18 Hz values can be measured. The cross-peak defined by the active coupling constant of 4 Hz allow the C3H resonance assignment at 6 = 3.77 ppm and thus its arrangement in axial configuration. The 18-Hz value (measured along the F2 axis) which is the sum of the active J3,4 and passive J4,5 coupling constants with J3,4 = J4,5 = 9 Hz characterizes the C4H in the axial configuration. This 18-Hz value can be explained by the cancellation of the phase and anti-phase of the central components as illustrated by the fine structure of the C3H, C4H cross-peak from the rhamnopyranosyl unit I (Fig. 3). This assumption is supported by the active coupling constant of 9 Hz measured along the
F1 axis. So, the unequivocal assignment and the coupling constants of all the resonances of unit I1 allow its identification as a rhamnopyranosyl residue.
The proton assignments which defined the third monosac- charide unit arise from the COSY-45 spectrum and are summarized in Table 1. This unit is characterized by the C1H resonance at 6 = 5.11 ppm as a doublet with an estimated J1,2 of 3.7 Hz. This value agrees with an a-anomer and its C2H in axial configuration. This statement is supported by the C3H resonance at 6 = 4.19 ppm as a doublet of doublets with two coupling constants of 10.3 Hz and 3.3 Hz. The main feature of the proton chemical shifts of this unit concerns the downfield displacement of its C4H proton to 6 = 5.23 ppm. Such a deshielding effect can be explained by the location on the C4 of the native acetoxy residue previously noticed (6 = 2.14ppm). Moreover, this C4H resonates as a doublet of doublets with coupling constants of 3.3 Hz and < 1.5 Hz characteristic of the C3H, C4H and C5H Fucp proton ar- rangements [13]. Despite the absence of the C4H, C5H Fucp cross-peak, this C5H resonance, which is a reporter group of a Fucp residue, can be easily recognized in the 1D spectrum at 6 = 4.29 ppm [19]. This C5H resonance correlates with a 6-deoxy group at 6 = 1.09. Thus, the chemical shifts and coupling constants are convincing evidence for the identifi- cation of a 4-0-Ac-a-Fucp unit.
The remaining monosaccharide unit (IV) was charac- terized from the COSY-45 (Fig. 2) and the phase-sensitive DQF-COSY (Fig. 3). Its C1H (6 = 5.10 ppm) was found con- nected to the C2H resonance at 6 = 3.97 ppm. Its equatorial position was established from the small J l , z (< 1.5 Hz). The C4H and CSH resonances were easily identified from COSY- 45, but owing to their signal overlap with other resonances, it was impossible to determine their coupling constants in the 1D spectrum. This data was obtained from the phase-sensitive DQF-COSY (Fig. 3). Analysis of the fine structure of the C3H, C4H and C4H, CSH cross-peaks (Fig. 3) suggests coup- ling constants JC3H, C4H = JC4H, C5H of 9 Hz, thereby in- dicating that C5H and C3H are in trans di-axial position with respect to C4H. The C2H, C3H and C4H chemical shifts (Table l), multiplicities, and coupling constants support a third rhamnopyranosyl residue. However, from the COSY- 45, C5H was connected to a proton which resonated at 6 = 3.77 ppm itself connected through a typical rectangular shape of C6H/C6H’ cross-peak [20] to another proton with a chemi- cal shift of 3.89 ppm (Fig. 2). The similar chemical shifts of C5H (6 = 3.72 ppm) and C6H (6 = 3.77 ppm) (Table 1) agree with the absence of the C5H, C6H cross-peak hidden under the diagonal peaks. This three-spin system composed of C5H, C6H and C6H’ characterized a hexopyranosyl unit, the structure of which was assigned to a Manp [21, 221.
Thus, the combination of DQF-COSY and COSY-45 ex- periments indicated that the oligosaccharide part of PheGl K- IV is composed of two Rhap (I and II), one 4-O-Ac-a-Fucp (111) and one Manp (IV) units.
PheGl K-JV NOESY studies
In order to continue the structal analysis, it was necessary to determine the type and site of the linkages between the monosaccharide units, the anomerlc configurations and the location of the five methoxy appendages. These data were obtained by NOESY experiments 123,241. In NOESY studies, significant connectivities can only be observed for interproton distances of less than 0.3 nm. Thus, two classes of cross-peak connecting anomeric resonances to other resonances were ob-
452
a
5.0 4.0 3.0 2.0 1 .o c- wrn (m)
I I
\
3.2
3.3
3.4
3.5
3.6
3.1
3.8
3.9
4.0
4.1
4.2
4.3
> 1 , , , , 1 , . , , , .
4 3 4 2 4 1 4 0 3 9 3 8 3 7 36 3 5 3 4 3 3 3 2
-1 .hl(l (m)
Fig 2 The 300-MHz COSY-45 of PheGl K-IV f rom M kansasn (b) corresponds to enlargement of the box in (a)
45 3
- 3.2
- 3 . 6
- 3.6
- 3 . 8
- 4.0
- 4.2
F1
9 =J* H4.m
b I I
3 . 7 3.6
3.7
3 . a
3 . 3
4 . 0
l ' l ' l . l . l * I 4 . 2 4 .0 3 . 8 3.6 3 . 6 3 . 2 p2
chrmiul shifI@pn)
Fig. 3. Phase-sensitive DQF-COSY ofPhe GI K-IV. (a) Partial spectrum; (b) corresponds to the box enlargement of the combined cross-peaks between C2H, C3H and C3H, C4H of the 2-0-Me-Rhap (unit IT). Negative components were plotted with multiple height contours and for each of the positive components only the lowest contour has been shown. J* is twice the value of the real coupling constant due to the cancellation of the central components. The digital resolution is 3.52 Hz/point along F1 and 1.76 Hz/point along F2
Table 1 . Proton assignment of the oligosaccharide part of the PheGl K-IV based on COSY-45 and phase-sensitive DQF-COSY 2D 'H-NMR experiments I, 2,4-di-O-Me-a-Rhap; 11, 2-0-Me-a-Rhap; III,4-O-Ac-2-O-Me-x-Fucp; IV, 4-0-Me-a-Manp
Unit Chemical shift __
C1H C2H C3H C4H C5H C6H3 C6H C6H' OCH3
PP" I 5.44 3.68 4.11 3.21 3.69 1.28 - - 3.52, 3.54
- 3.47 I1 5.18 3.63 3.17 3.63 3.87 1.35 -
- 3.51 IT1 5.1 1 3.56 4.19 5.23 4.30 1.09 -
IV 5.10 3.97 3.78 3.44 3.72 - 3.77 3.89 3.55
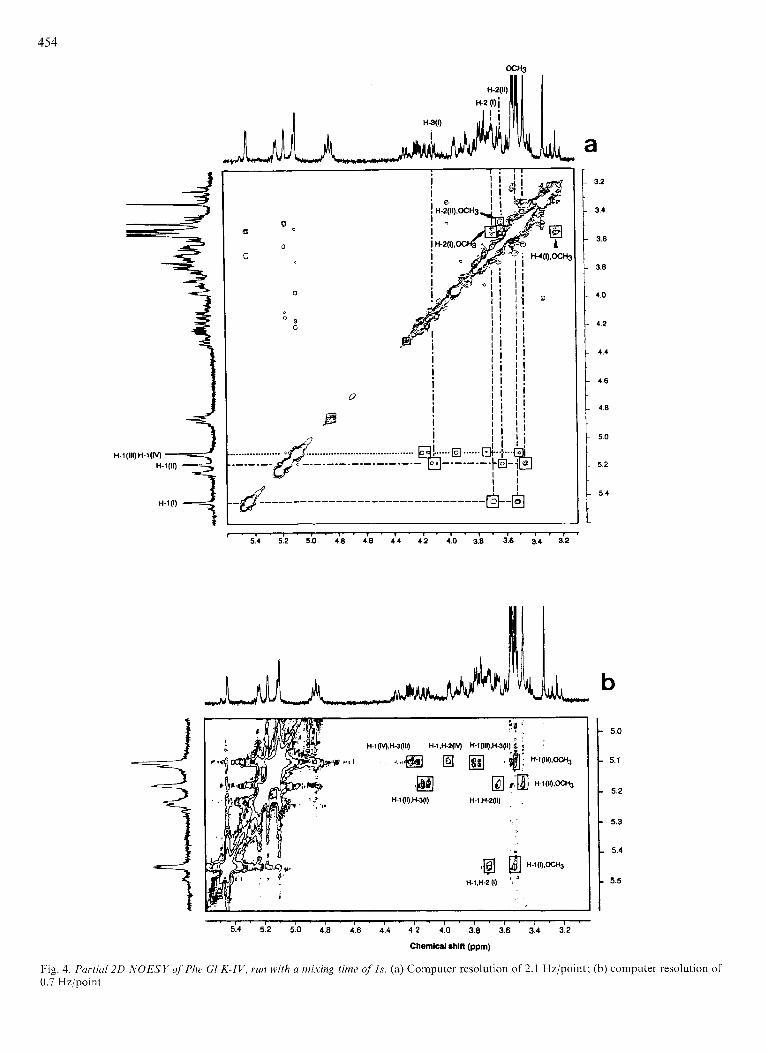
served: those due to interactions with protons on the same residue and those due to interactions with protons on adjacent residues. Anomeric intraresidue connectivities yield infor- mation about anomeric configuration, for instance, a-Rhap leads to CIH, C2H connectivity while its b-anomeric form would be characterized by ClH, C3H and ClH, C5H corre- lations [25, 261. Anomeric interresidue connectivities are the route to the establishment of the interglycoside linkage sites (Fig. 4).
Fig. 4a presents the contour plot of the NOE experiments with a mixing time of Is. The downfield anomeric proton resonance (6 = 5.44 ppm) shows connectivities with the res- onances at 6 = 3.68 ppm (C2H, Rhap I), 6 = 3.52ppm (methoxy group) and 6 = 6.95 ppm (o-phenolic protons, not shown in the figure). Thus, a 2-0-Me-a-Rhap can be proposed for the Rhap unit I as well as its linkage through the C1 anomeric carbon atom to the phenolic nucleus of the aglycone. Again, the CIH of the second Rhap units presents the same behaviour. Three resonance connectivities are observed at 6 = 3.47 ppm (methoxy group), 6 = 3.63 ppm (C2H, Rhap 11) and at 6 = 4.1 1 ppm (C3H, Rhap I) supporting a 2-O-Me-a- Rhap structure for the second Rhap unit and its 1+3 inter-
glycosidic linkage with the 2-0-Me-a-Rhap I. However, as shown by Fig. 4, the overlapping of the anomeric proton resonances of the Manp (6 = 5.10 ppm) and the 4-0-Ac-a- Fucp (6 = 5.03 ppm) does not allow the C l H assignment of the connectivity. This problem has been overcome by decreasing the spectral width from 2100 Hz to 1500 Hz and by an enhancement from 2K to 4K of the memory size. Effec- tively, Fig. 4b shows that C1H resonances of both monosac- charide residues are well resolved, thereby allowing their con- nectivity to be established. The C1H Manp is connected to the C3H Fucp (6 = 4.19 ppm), through a cross-peak which displays a fine multiple structure composed of four compo- nents allowing the coupling constants to be estimated. These are in agreement with C2H, C3H and C4H of a Fucp residue, thus supporting the interglycosidic connection (I 4 3 ) between the Manp and the 4-0-Ac-Fucp unit.
The a-anomeric configuration of the Manp residue was established from an intraresidue connectivity between its C1 H and C2H. The remaining C1H Fucp was found connected to the C3H of the Rhap I1 unit (6 = 3.77 ppm). This assignment based on the C3H chemical shift was supported by the measurement of JCZH, C3H and JC3H, C4H from the four
454
- 3.2
- 3.4
- 3.6
- 3.8
- 4.0
- 4.2
- 4.4
a .L
1
- 4.8
- 5.0
- 5.2
- 5.4
-
' 5.4 5.2 5.0 4.8 4.8 4.4 4.2 4.0 3.8 3.6 3.4 3.2
a : . - 5.0
- 5.1
- 5.2
- 5.3
- 5.4
- 5.5
I ~ I ' I ' I ~ I ' I ' I ' 1 ' 1 ' I ' i ' l ' 5.4 5.2 5.0 4.8 4.6 4.4 4.2 4.0 3.8 3.6 3.4 3.2
Chemical rhnt (ppm)
Fig. 4. Purticrl20 -liOESY of'Phe GI K-IV, run with u mixing time o f l s . (a) Computer resolution of 2.1 Hz/point; (b) computer resolution of 0.7 Hzjpoinl

455
Fig. 5. Visualisation of the NOE contacts involving the Phe GI K-IV anorneric protons
components of the ClH, C3H cross-peak. The NOE contact between the C1H Fucp and the methoxy group at 6 = 3.51 ppm, agrees with the location of the methoxy group on the C2 of the Fucp unit. Thus, we can propose for this mono- saccharide unit 4-0-Ac-2-0-Me-a-Fucp structure.
As shown by Fig. 4a, other cross-peaks are observed al- lowing connectivities in which the anomeric protons are not involved. They result from NOE contacts between the methoxy groups (6 = 3.52 ppm, 6 = 3.47 ppm) localized on C2 of both Rhap and their respective C2H geminal protons. More interesting is the connectivity found between the unlocalized OCH3 group (6 = 3.45 ppm) and the C4H reso- nance of the Rhap I, supporting the 2,4-di-O-Me-a-Rhap structure.
From the five methoxy groups of the carbohydrate part, only one which resonated downfield (6 = 3.55 ppm) was still unlocalized by the NOE experiments. It can be linked either to the Rhap I1 C4 or on the C3, C4 and C6 of the distal Manp unit (the exclusion of C2 results from the absence of connectivity between C1H and OCH3).
From Table 1, it can be observed that the C2H chemical shifts of both Rhap residues are similar (6 = 3.68 ppm, RhapI; 6 = 3.63 ppm, RhapII) while their C4H (6 = 3.21 ppm, RhapI; 6 = 3.63 ppm, RhapII) show significant differences. This effect is in agreement with the absence of a methoxy group on the C4 of the RhapII since RhapI is a 2,Cdi-0-Me-a-Rhap unit. Thus, the methoxy group must be located on the distal Manp unit. From the comparative chemi- cal shift values analysis between the methoxy groups and of the C2H, C3H, C4H, C6H and C6H' of the 2,4-di-O-Me-a-~- Manp PheGl K-I1 residue 1211 (C2, OCH3, 6 = 3.46 ppm; C4, OCH3, 6 = 3.55ppm; C2H, 6 = 3.47ppm; C3H, 6 = 3.72 ppm; C4H, 6 = 3.30 ppm; C6H, 6 = 3.72 ppm; 6 C6H', 6 = 3.87 ppm) and those of the PheGl K-IV Manp unit (see Table l), it can be established that the remaining methoxy group is located on the C4 Manp unit. This assignment is in agreement with the C4 methoxy downfield resonance de- scribed by Vliegenthart et al. in the case of permethylated a-Manp and a-Rhap [27]. Thus, the resonance proximity of the C4H (6 = 3.44) and the methoxy group (3.55 ppm) of the 4-0-Me-a-Manp agrees with a cross-peak hidden by the diagonal cluster resonances on PheGl K-IV NOESY (Fig. 4).
From these data, the following structures can be advanced for the tetrasaccharide part of PheGl K-IV which is sum- marized in Fig. 6: 4-O-Me-a-Manp-( 1 + 3)-4-0-Ac-2-0-Me- a-Fucp-(1 + 3)-2- O-Me-a-Rhap-(l+3)- 2,4-di-O-Me-a-Rhap-l +aglycone.
DISCUSSION
In the present study, the primary structure of the carbo- hydrate part of a new phenolic glycolipid antigen isolated from the M . kansasii cell walls, in milligram quantities, was established from 'H-NMR analysis.
The results obtained confirm the application of the newer H-NMR pulse sequences for the structural elucidation of
unknown glycolipids. Although, it remains, of course, neces- sary to establish the absolute configuration of the constituent monosaccharides by conventional procedures.
In the recent past, two-dimensional 'H-NMR COSY and NOESY techniques have been successfully used in order to determine oligosaccharide structures [14 - 161. Analysis via 2D 'H NMR involve two types of correlated experiments: one allows the connection of vicinal protons through scalar coupling (COSY), the other correlates protons through space by NOE. The identification of the monosaccharide units was approached by conventional COSY experiments [23]. Starting from a given anomeric proton, we can assign the other protons which belong to the same monosaccharide unit; thereby, from their chemical shifts and coupling data, the structure of the monosaccharide can be elucidated. The success of this method depends on the fact that protons of each monosaccharide constitute a separate spin system. However, carbohydrate pro- tons which resonate independently in a small chemical shift range lower than 0.1 ppm led to cross-peaks hidden in the cluster peaks of the diagonal. As a consequence, it is impos- sible to define the connectivity paths. To overcome this prob- lem, COSY-45 was selected instead of COSY-90 even though this latter version is more sensitive. From the PheGl K-IV 2D COSY-45 spectrum, only two monosaccharide units, the 2,4- di-0-Me-Rhap (unit I) and 4-0-Ac-2-0-Me-Fucp (unit III), were identified. The impossibility of obtaining complete reso- nance assignments and couplings for the remaining monosac- charide units the 2-0-Me-a-Rhap (unit 11) and the 4-O-Me-a- Manp (unit IV) arise from the inherent complexity of the 'H- NMR spectrum. For instance, C2H and C4H from unit I1 resonate at 6 = 3.63 ppm while C3H and C5H of unit IV resonate at 6 = 3.78 ppm and 6 = 3.72 ppm (see Table 1). Thus, the connectivities of the C2H, C3H, C4H of the unit I1 and the C3H, C4H, C5H of unit IV give rise to superimposed cross-peaks (Fig. 3). Such a problem was solved by the use of two-dimensional phase-sensitive DQF-COSY sequence. This method has been successfully introduced by Wuthrich et al. in order to identify amino acids 1281, but also appeared useful for the elucidation of oligosaccharide structures [16]. Due to the improved resolution in phase-sensitive spectra, the fine structure of cross-peaks could be exploited to determine the coupling constants of the involved protons 116,281. Moreover, it is possible to discriminate between active and passive coup- ling by their in-phase and anti-phase splitting which appears in the cross-peak as positive and negative signals.Active coupling defines the coupling between the protons connected cross- peak, while passive coupling defines the coupling of one of these protons with any others [28]. From the fine structure of the combined cross-peaks C2H, C3H and C3H, C4H from the unit IT (Fig. 3), we were able to determine the active

456
S
K - I %OM. 2,6-dideoxy-4-e-Me~ -D-arabino-hexopyranosyl
CH,OH 2,4-di-O-Mea - -D-Manp
K - I1 wr OY.
K - I11 unknown
CH.OH K - IV wy OM. 4-O-Mea - -D-Manp
OH
Fig. 6 . Oligosuccharide structures uf’ the phenolic gljvolipids isolated from M. kansasii. The absolute configuration of PheGl K-IV was assumed to be similar to that of PheGl K-I
coupling constants of 4 Hz and 9 Hz in agreement with a Rhap structure. A similar process was applied for the remaining monosaccharide which was assigned to a Manp (Fig. 3).
By 2D NOESY experiments [24] with a mixing time of Is, the sequence of the tetrasaccharide was determined and the connection established as a unique (1 +3) interglycosidic link- age from interresidue NOE contacts. Also, convincing evi- dence was found for their cl-anomeric configurations (NOE contacts, C1 H, CZH), and the location of themethoxy residues (NOEcontacts: CIH, OCH3 and C2H, OCH3 for themethoxy at C2H; C4H, OCH3 for the methoxy at the C4 of the Rhap 11). Unfortunately, a limitation of this technique was found for the methoxy located at the C4 of the Manp residue due to the resonance proximity of this group and its geminal proton. This assignment results from the characteristic chemical shifts of C2 and C4 methoxy groups in the Manp and Rhap units (Table 1).
This study clearly confirms that ‘H-NMR spectroscopy using 2D NMR techniques with the sequence COSY-45, DQF-COSY and NOESY, with a mixing time of Is, is a powerful tool for the primary structure elucidation of PheGl K-IV carbohydrate in its native form. Other mixing times were checked in NOESY experiments, e.g. 250 ms and 500 ms. However. in both cases, compared to a 1-s mixing time, only the NOE contact between the C1H of the distal 4-0-Me-a- Manp and the C3H of the penultimate unit 4-0-Ac-2-0-Me- x-Fucp was missing.
Compared to the conventional methods, NMR offers sev- eral advantages: it is a non-destructive technique, which can be applied to samples of the order of 1 mg in their native form, and performed in a reasonable time. An obvious limitation of this technique concerns the absolute configuration assignment which must be achieved by conventional procedures.
The PheGl K-IV, a quantitatively minor phenolic gly- colipid, was isolated from the pathogenic mycobacteria M. knnsnsii. Its antigenicity was demonstrated by ELISA 1371. Beside PheGl K-IV, three other phenolic glycolipids were isolated; the major one, i.e. PheCl K-I [lo], and two others in less abundance, PheGl K-111 [I71 and PheGl K-I1 [21]. At present, the structures of K-I, K-I1 and K-IV have been estab- lished and are presented in Fig. 6. The structures show that M . kansasii phenolic glycolipids share a common core assigned to the monoacetylated trisaccharide unit. They differ by the structure of the distal monosaccharide unit: a 2,h-dideoxy- 4-O-Me-a-~-arahino-Hexp, a 2,4-di-O-Me-a-~-Manp and a 4- 0-Me-a-D-Manp for K-I, K-TI and K-IV, respectively.
In previous work, the PheGl K-IV antigenicity and the lack of PheGl K-IV recognition by the PheGl K-I antibodies in ELISA were established [17]. Such a specificity can now be supported by the structure of the distal PheGl K-IV monosac- charide unit.
We thank Dr Nuzillard (Reims, Francc) for hclpful discussion.

457
REFERENCES 1. Brennan, P. J. (1988) Microbial lipids (Ratledge, L. & Wilkinson,
S. G.. eds) vol. 1, pp. 203-298, Academic Press, London. 2. Brennan, P. J. (1984) in Microbiology (Leive, L. & Schlessinger,
D,, eds) pp. 366-375, American Society for Microbiology, Washington.
3. Puzo, G. (1990) Critical Rev. Microhioll7, 305 - 327. 4. Fournit, J. J., Rivibre, M., Papa, F. & Puzo, G. (1987) J . Biol.
5. Fournie, J . J., Rivibre, M. & Puzo, G. (1987) Eur. J . Biochem.
6. Hunter, S. W., Fujiwara, T., Murphy, R. C. & Brennan, P. J.
7. Riviere, M., Fournit, J. J., Vercellone, A. 81 Puzo, G. (1988)
8. Jardine, I., Scanlan, G., McNeil, A. & Brennan, P. J. (1 989) Anal.
9. Vercellone, A., Rivikre, M. & Puzo, G. (1989) Adv. Mass.
10. FourniC, 3 . J., Riviere, M. & Puzo, G. (1987) J . Biol. Chem. 262,
11. Vercellone, A. & Puzo, G. (1989) J . Biol. Chem. 264,7441-7454. 12. DaffC, M. & Servin, P. (1989) Eur. J . Biochem. 185, 157-162. 13. Riviere, M. (1 987) Doctorat d’UniversitC., Universite Paul
14. Prestegard, J . H., Koerner, T. A. W., Derman, P. C. & Yu, R. K.
Chem. 262, 3180-3184.
168, 181-183.
(1984) J . Biol. Chem:259, 9729-9734.
Biomed. Environ. Mass Spectrom 16, 275 -278.
Chem. 61,416-422.
Spectrom, l lB , 1350-1351.
3 1 74 - 31 79.
Sabatier, Toulouse.
(1982) J. Am. Chem. Soc. 104,4993-4995.
15.
16. 17.
18.
19.
20. 21.
22.
23.
24.
25.
26.
27.
28.
Bernstein, M. A. & Hall, L. D. (1982) J . Am. Chem. Soc. 104,
Dabrowski, J. (1989) Methods Enzymoll79, 122- 157. Gilleron, M., Venisse, A,, Rivibre, M.: Fournie, J. J. & Puzo, G.
Homans, S. W., Dweck, R. A., Fernandez, D. L. & Rademacher,
Vliegenthart, F. G. J., Dorland, L. & Harlbeck, H. V. (1983) Adv.
Berman, E. (1987) Eur. J . Biochem. 165, 385-391. Riviere, M., Fournie, J. J . & Puzo, G. (1987) J . Biol. Chem. 262,
Webb, G. A. (1982) Anual reports on N M R spectroscopy, Aca-
Derome, A. E. (1988) Modern N M R techniques for chemistry
Yu, R. K., Koever, T. A. W., Scansdala, T. N. & Prestegard, J.
Michon, F., Brisson, J. R., Roy, R., Ashton, F. E. & Jennings,
Romanowska, E., Romanowska, A,, Dabrowski, J. & Hauck, M.
Haverkamp, J., De Bie, M. J. A. & Vliegenthart, J. F. G. (1975)
Neuhaus, M., Wagner, G., Vasak, M., Kagi, J. H. R. & Wiithrich,
5553 - 5555.
(1990) Eur. J . Biochem. 189, 167- 173.
T. W. (1984) Biochim. Biophys. Actcl 798, 78-83.
Carhohydr. Chem. Biochem. 41,209 - 374.
14 879 - 14 884.
demic Press, London.
research (Baldwin, J. E., ed.) Pergamon Press, Oxford.
H. (1986) Chem. Phys. Lipids 42, 27-48.
H. J. (1985) Biochemistry 24, 5592-5598.
(1987) FEBS Lett. 211, 175-178.
Carhohydr. Res. 39, 201 -211.
K. (2985) Eur. J . Biochem. 151, 257-273.







![Elucidation de la structure de molécules chirales par ...[5] Pérez-Mellor, A. & Zehnaker, A. Virational irular dihroism of a 2,5‐diketopiperazine (DKP) peptide: Evidene for dimer](https://img.pdfslide.fr/doc/110x75/5ec12e8b1d9a931fa06bde99/elucidation-de-la-structure-de-molcules-chirales-par-5-prez-mellor-a.jpg)











