Embed Size (px)
Citation preview

COMPTE-RENDU TP CELOE AVRIL 2016
LUCAS JEAN-JACQUES & BRICE PICHARD
GROUPE 31

1/17
Sommaire
Les unicellulaires eucaryotes Illustration des grandes fonctions physiologiques des eucaryotes : ................................................................................................................. 2
Liste et tableau récapitulatif ................................................................................................................................................................................... 2
Différentes fonctions assurées par les unicellulaires .............................................................................................................................................. 3
Illustration d’interaction chez des unicellulaires eucaryotes : ......................................................................................................................... 6
Le lichen : symbiose entre le mycète Xanthoria et l’algue unicellulaire Trebouxia .............................................................................................. 6
Parasitisme : Néréis par Grégarines ....................................................................................................................................................................... 6
Utilisation pratique .............................................................................................................................................................................................. 7
La complexification des organismes Mise en place de colonies : ................................................................................................................................................................................... 8
Exemple du Volvox ................................................................................................................................................................................................ 8
Exemple de Nostoc ............................................................................................................................................................................................... 10
Position des choanoflagellés : ............................................................................................................................................................................ 11
Complexification des thalles d’algues unicellulaires : .................................................................................................................................... 12
Différents thalles .................................................................................................................................................................................................. 12
Fonctions des thalles ............................................................................................................................................................................................ 13
Le règne fongique, les Eumycètes : ................................................................................................................................................................... 13
Présentions d’organismes : ................................................................................................................................................................................... 13
Les fonctions vitales des eumycètes : .................................................................................................................................................................. 14
Champignon entre plante et animal : .................................................................................................................................................................... 15
Caractéristiques des organes végétaux types Embryophyte et comparaison aux ancêtres aquatiques : ................................................... 16
Sources ………………………………………………..……….…………………….17
2/17
Les unicellulaires eucaryotes
Les êtres unicellulaires eucaryotes semblent être les individus les moins complexes du domaine Eucaryote. Cependant ils n’en restent pas moins évolués
et répondent ainsi aux grandes fonctions physiologiques. Certains peuvent même entretenir des relations complexes avec d’autres espèces (notamment
pluricellulaires).
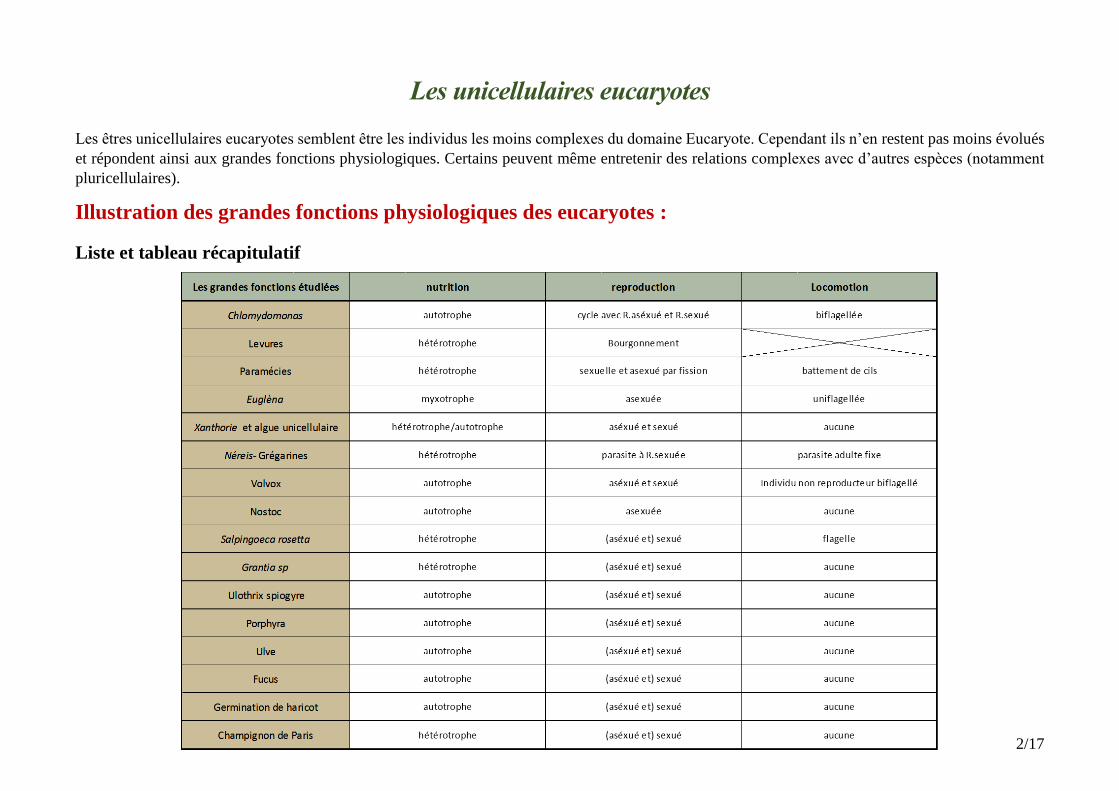
Illustration des grandes fonctions physiologiques des eucaryotes :
Liste et tableau récapitulatif
3/17
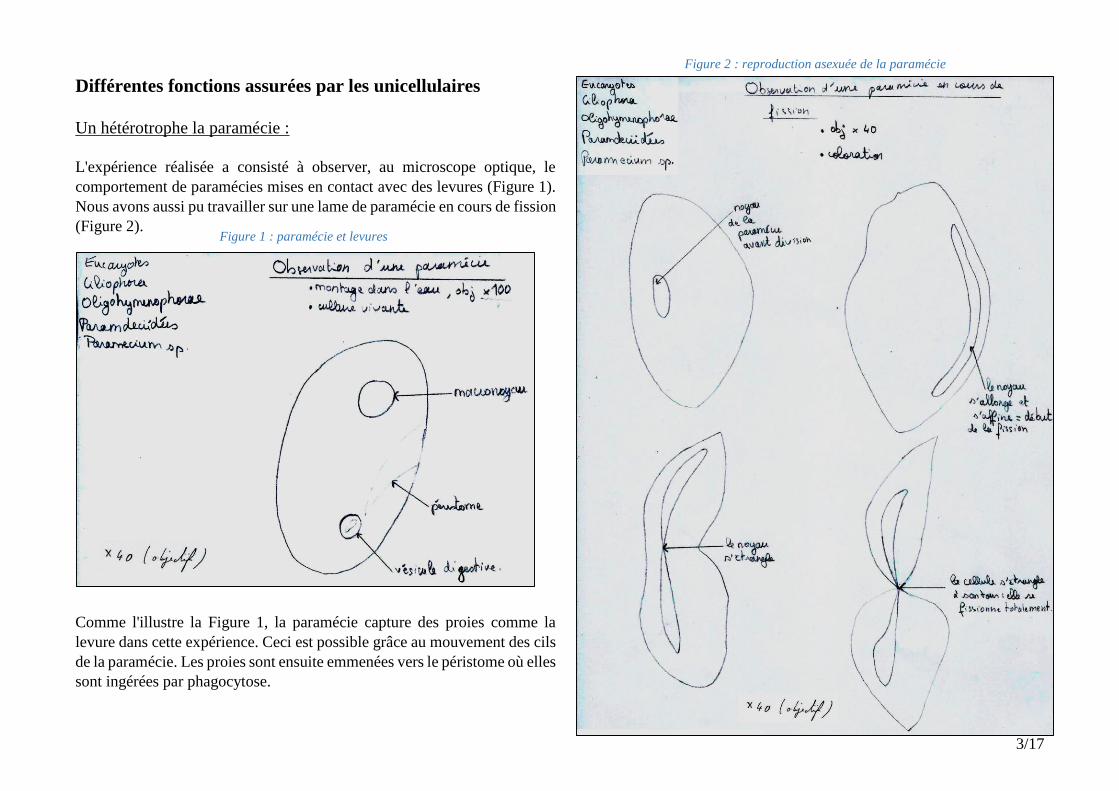
Figure 2 : reproduction asexuée de la paramécie
Figure 1 : paramécie et levures
Différentes fonctions assurées par les unicellulaires
Un hétérotrophe la paramécie :
L'expérience réalisée a consisté à observer, au microscope optique, le
comportement de paramécies mises en contact avec des levures (Figure 1).
Nous avons aussi pu travailler sur une lame de paramécie en cours de fission
(Figure 2).
Comme l'illustre la Figure 1, la paramécie capture des proies comme la
levure dans cette expérience. Ceci est possible grâce au mouvement des cils
de la paramécie. Les proies sont ensuite emmenées vers le péristome où elles
sont ingérées par phagocytose.

4/17
Figure 3 : chlomydomonas
Elles seront ensuite digérées dans les lysosomes de la paramécie. Ce protiste consomme donc des organismes vivants pour se nourrir. La paramécie
répond donc tout à fait à la définition de ce qu'est un hétérotrophe : un organisme vivant incapable de fabriquer sa propre matière organique et qui va la
puiser dans d'autres organismes hétérotrophes ou autotrophes.
De plus, la reproduction de la paramécie, schématisée sur la Figure 2, est remarquable. Ce schéma illustre une paramécie qui est en train de se scinder
progressivement en 2. Le noyau s’allonge devenant de plus en plus fin puis s'étrangle en deux parties jusqu'à que celles-ci ne soient presque plus reliées.
A ce moment l’organisme se scinde en deux à son tour au niveau de cette zone d'étranglement du noyau, emportant une partie du noyau dans chaque
future cellule « fille ». Enfin, l’organisme se fissionne totalement en deux exemplaires comportant chacun une partie du noyau et du compartiment
cytoplasmique de la cellule « mère ». La paramécie réalise donc une reproduction asexuée par fission binaire : aucun matériel génétique n'est échangé.
Cependant, lorsque le milieu est carencé en nutriments, la paramécie va échanger un de ses micros noyaux (obtenus par méiose puis mitose) avec celui
d'une autre paramécie. Ces micros noyaux vont fusionner dans la cellule (« fécondation ») puis l'unique micro noyau ainsi obtenu va se diviser par mitose
en huit micro noyaux dont 4 deviendront des macros noyaux. Enfin la cellule (c'est-à-dire la paramécie), réalise deux mitoses afin d'obtenir quatre
nouvelles paramécies comportant chacune un micro noyau et un macro noyau. Ce mécanisme est celui d'une reproduction sexuée, le mélange génétique
engendré augmente ainsi les chances d'adaptation aux conditions du milieu.
Un autotrophe la Chlamydomonas :
Nous avons observé des chlamydomonas vivantes au microscope optique
(Figure 3).
Chlamydomonas est une algue unicellulaire verte. On peut noter la présence
d'un unique gros chloroplaste qui occupe tout l'espace cellulaire (ce qui
explique la couleur verte qui est donc due à la chlorophylle) comme illustré
sur la Figure 3. Il s'agit donc d'un organisme autotrophe, c'est-à-dire qui
produit sa propre matière organique à partir de CO2, d'H2O et d'énergie
lumineuse : elle réalise la photosynthèse.
Cet organisme réalise la mitose pour se reproduire de manière asexuée. De
même que pour la paramécie, si les conditions du milieu sont mauvaises, deux
cellules de polarité sexuelle différente peuvent fusionner pour former un
zygote qui, par mitose, va générer quatre cellules haploïdes. C'est la
reproduction sexuée.
5/17
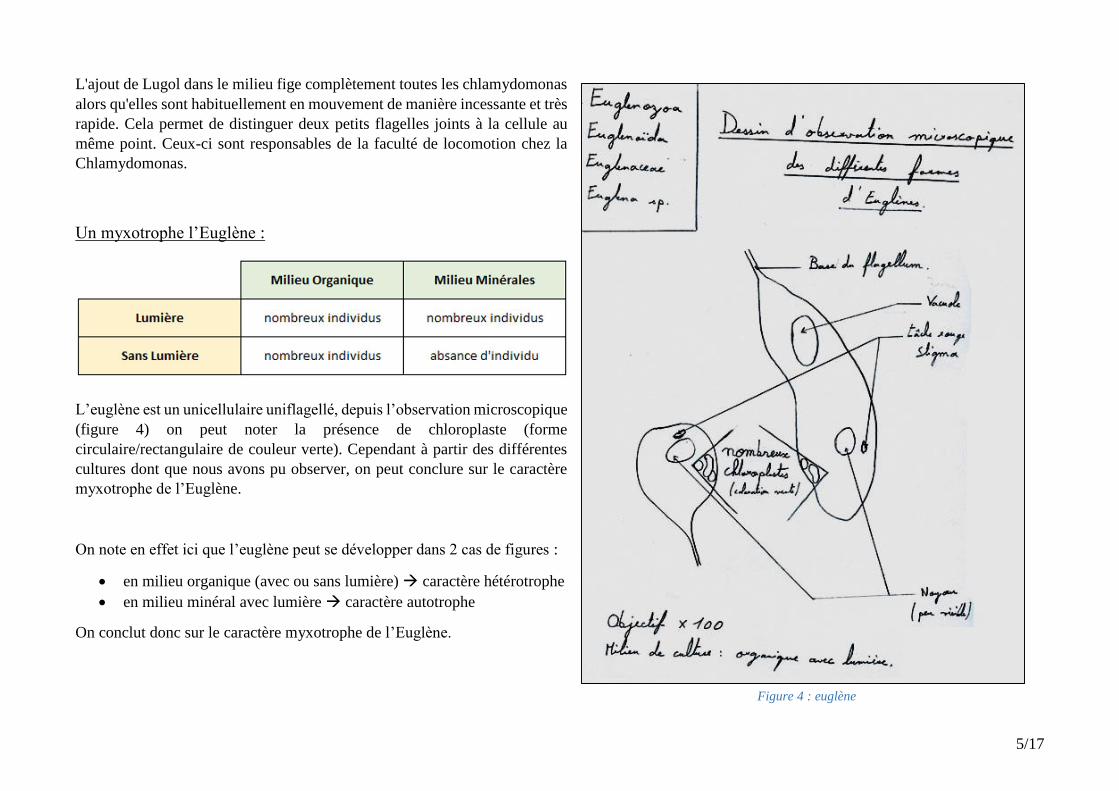
Figure 4 : Euglène observation avec lumière et MO
Figure 4 : euglène
L'ajout de Lugol dans le milieu fige complètement toutes les chlamydomonas
alors qu'elles sont habituellement en mouvement de manière incessante et très
rapide. Cela permet de distinguer deux petits flagelles joints à la cellule au
même point. Ceux-ci sont responsables de la faculté de locomotion chez la
Chlamydomonas.
Un myxotrophe l’Euglène :
L’euglène est un unicellulaire uniflagellé, depuis l’observation microscopique
(figure 4) on peut noter la présence de chloroplaste (forme
circulaire/rectangulaire de couleur verte). Cependant à partir des différentes
cultures dont que nous avons pu observer, on peut conclure sur le caractère
myxotrophe de l’Euglène.
On note en effet ici que l’euglène peut se développer dans 2 cas de figures :
en milieu organique (avec ou sans lumière) caractère hétérotrophe
en milieu minéral avec lumière caractère autotrophe
On conclut donc sur le caractère myxotrophe de l’Euglène.

6/17
Illustration d’interaction chez des unicellulaires eucaryotes :
Le lichen : symbiose entre le mycète Xanthoria et l’algue unicellulaire Trebouxia
L’observation microscopique et les documents à disposition du lichen nous ont permis de mettre en évidence la relation de symbiose entre xanthoria et
L’algue Trebouxia. Dans cette relation le champignon apporte une niche écologique favorable à l’algue : protection en plus de l’apport en eau et ions.
L’algue quant à elle apporte des produits de la photosynthèse.
On note sur la reproduction que l’algue est défavorisée par rapport au champignon. En effet il y a deux types de reproductions utilisées mais une seule
autorisant la multiplication et dispersion de l’algue :
La reproduction sexuée : seul le champignon se reproduit de façon sexuée grâce à des hyphes fongiques sexuellement différenciés fusionnant et
donnant, à la surface du thalle, des structures en forme de boutons (les apothécies dans lesquelles des cellules particulières (les asques) vont
élaborer les ascospores. Les nouveaux hyphes « captureront » ensuite des algues pour élaborer la symbiose ;
La reproduction asexuée : se fait par déchirure du thalle et dispersion de fragments (contenant les deux partenaires).
Le lichen a été utilisé par l’homme dans la médecine traditionnelle, il était utilisé pour traiter diverses maladies comme la diarrhée. Aujourd’hui, bien
qu’encore utilisé en médecine, alimentation et industrie (parfumerie), il sert surtout de bio-indicateur pour, par exemple, le taux de pollution ou la stabilité
chimiques des sols.
Parasitisme : Néréis par Grégarines
Grégarines est un parasite (généralement spécifique) se logeant dans l’intestin d’invertébré, ici Néréis. Il se fixe grâce à sa tête dans la paroi de l’intestin
puis profite ensuite de cet environnement qui lui est favorable (forte concentration de nutriment et abri).

7/17
Utilisation pratique
Les eucaryotes unicellulaires sont exploités par l’homme dans différents domaines :
Alimentaire : depuis l’antiquité les unicellulaire notamment les levures sont
utilisées pour produire du vin, du pain ou de la bière grâce à la fermentation.
Récemment une entreprise : Euglena1 voudrait grâces au fort potentielle
nutritives des euglènes régler des problèmes de malnutrition et sous-nutrition
dans des pays tels que le Bengladesh. Cette entreprise commence aussi à
diversifier cette utilisation vers les biocarburants et la médecine.
Industrielle : du biocarburant à la parfumerie.
Médecine : utilisation comme compléments de vitamines, traitement
(pénicilline), homéopathie,…
Ecologie : en tant que bio-indicateur (cas du lichen et de certaines algues).
Ils ont aussi souvent un rôle majeur dans la décomposition et le
renouvellement de leurs milieux
1 https://www.euglena.jp/en/company/

8/17
Figure 5 : colonie de volvox
La complexification des organismes
Après nous être focalisés sur les êtres unicellulaires, il est légitime de se demander comment nous sommes passés d’êtres peu complexes à des êtres
composés de tissu et d’organes. Nous verrons donc au-travers de différents exemples la complexification des organismes de l’unicellulaire à l’organisme
complexe en passant par l’état colonial et les thalles.
Mise en place de colonies :
Exemple du Volvox
Après observation au microscope optique d'une colonie de
Volvox, illustrée par la Figure 5, on constate des
agglomérats de cellules au sein de la colonie. On observe
donc des colonies filles en formation. Celles-ci sont
obtenues par mitoses successives à partir d'une des
cellules constituant la colonie. La division cellulaire a
donc lieu à partir d'un seul génome : toutes les cellules de
la colonie ainsi formées auront un génome identique.
Néanmoins, si la colonie est placée dans un milieu où les
conditions trophiques sont défavorables, elle modifiera
son mode reproduction. La colonie produira des gamètes
grâce à la méiose à partir des cellules gonidiales,
présentent en périphérie de la colonie. Il y aura
fécondation entre deux gamètes issus de deux individus
différents : il y a donc une modification du génome.
Volvox pratique ainsi la reproduction sexuée afin
d'augmenter ses chances de se reproduire de manière
viable.
9/17
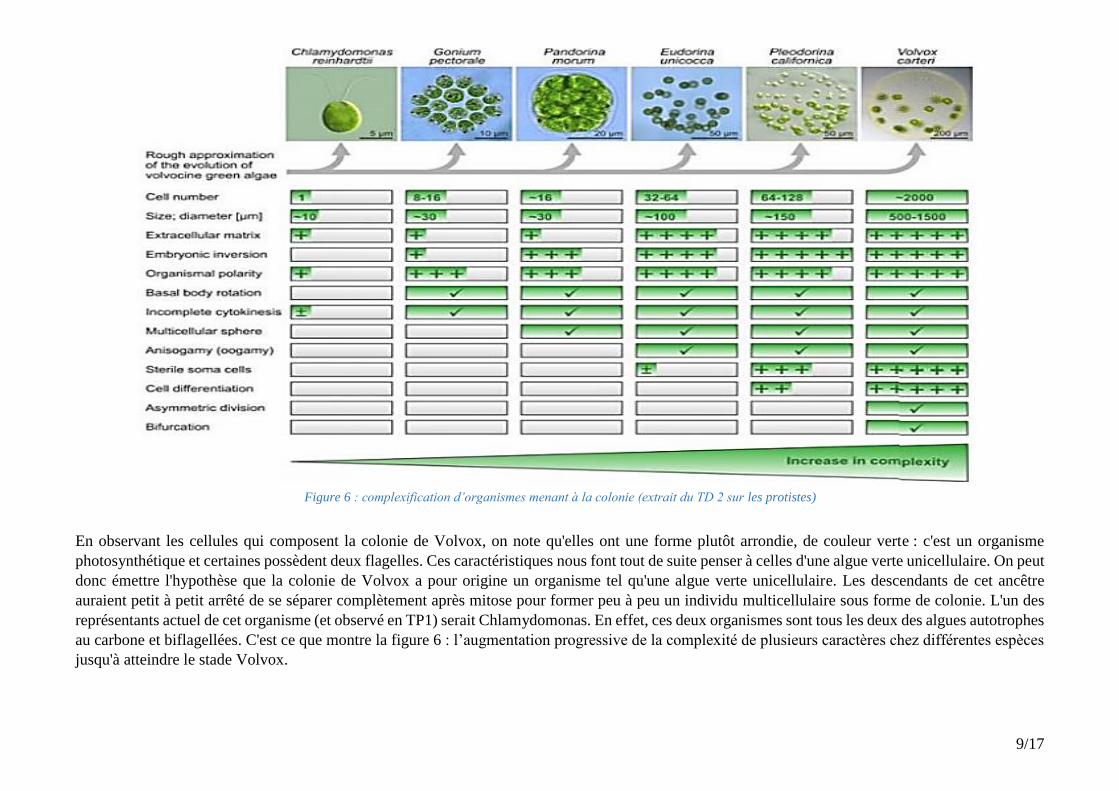
Figure 6 : complexification d’organismes menant à la colonie (extrait du TD 2 sur les protistes)
En observant les cellules qui composent la colonie de Volvox, on note qu'elles ont une forme plutôt arrondie, de couleur verte : c'est un organisme
photosynthétique et certaines possèdent deux flagelles. Ces caractéristiques nous font tout de suite penser à celles d'une algue verte unicellulaire. On peut
donc émettre l'hypothèse que la colonie de Volvox a pour origine un organisme tel qu'une algue verte unicellulaire. Les descendants de cet ancêtre
auraient petit à petit arrêté de se séparer complètement après mitose pour former peu à peu un individu multicellulaire sous forme de colonie. L'un des
représentants actuel de cet organisme (et observé en TP1) serait Chlamydomonas. En effet, ces deux organismes sont tous les deux des algues autotrophes
au carbone et biflagellées. C'est ce que montre la figure 6 : l’augmentation progressive de la complexité de plusieurs caractères chez différentes espèces
jusqu'à atteindre le stade Volvox.

10/17
Figure 7 : colonie de nostoc
Exemple de Nostoc
L'observation de Nostoc au microscope optique nous permet de visualiser sa structure cellulaire : la colonie est organisée en longs filaments de cellules,
ce sont des trichomes. On note rapidement que certaines cellules, plus rares, sont différentes des autres : plus grandes, celles-ci possèdent une paroi plus
épaisse : c'est les hétérocystes. La colonie est donc formée de deux types cellulaires différenciés : les hétérocystes sont spécialisées dans le métabolisme
azoté, ils fixent l'azote atmosphérique
pour la fabrication des acides aminés
alors que les autres cellules sont
différenciées pour la photosynthèse. Ces
deux processus sont antagonistes (l’O2
est toxique pour la protéine de ce
métabolisme azoté) et ne peuvent se
produire au même moment dans la même
cellule. Cette séparation des tâches dans
des cellules distinctes permet donc au
Nostoc de les réaliser toutes les deux en
même temps, ce qui augmente le
rendement métabolique du trichome et lui
confère donc un avantage certain.
Cependant, chaque cellule est un individu
unique possédant son propre génome.
Cela met en avant un des inconvénients
de la multicellularité : si toutes les
cellules n'ont pas le même génome il
existe un risque de conflit génétique qui
peut à terme nuire à l'organisme (exemple
de l'amibe sociale).

11/17
Figure 8 : comparaison choanoflagellé et choanocyte de Grantia
Position des choanoflagellés :
On peut noter la structure particulière des choanoflagellés : une cellule pourvue
d'un flagelle entouré d'une collerette de microvillosités. En observant une coupe
de spongiaire au microscope, on note la présence de cellules différenciées sur les
parois de la cavité interne. Celles-ci ont des formes plus ou moins variables mais
elles possèdent surtout un flagelle. Ce sont donc des choanocytes, on sait aussi
qu'ils sont aussi munis d'une collerette de microvillosités pour piéger les particules
alimentaires. Les choanocytes sont donc très semblables à la cellule de
choanoflagellés.
De plus, les choanoflagellés peuvent former des colonies si les cellules filles ne
se détachent pas complètement après mitoses successives de la cellule mère.
Avec tous ses éléments, on peut donc supposer que la multicellularité chez les
métazoaires prend sa source chez un ancêtre commun avec les choanoflagellés.
Cette théorie est encouragée par l'analyse phylogénétique comme l'indique la
figure 8 et l’arbre ci-dessus.

12/17
Figure 9 : archéthalle
Figure 10 : nématothalle
Figure 11 : nématothalle avec des cellules regroupées en îlots
Complexification des thalles d’algues unicellulaires :
Différents thalles
Nos observations microscopiques nous ont permis d’observer la complexification des thalles.
Nous avons pu voir tout d’abord les archéthalles représentés par l’Euglène et Ulothix. Ces thalles sont simples et formés par des unicellulaires ou des
cellules semblables qui se divisent par bipartition ou deviennent des cellules reproductrices.
Ensuite les nématothalles avec Ulve et porphyra. Ces thalles sont formés par un filament rampant sur lequel sont joints des filaments dressés. L’ensemble
pouvant prendre l’apparence d’une lame comme dans le cas de nos deux sujets d’observations.
On retrouve le plus haut degré de complexité avec les cladothalles qui peuvent ressembler (du point de vue de la structure) à une tige feuillée. C’est le
cas ici de Fucus.

13/17
Fonctions des thalles
Les thalles offrent une optimisation de l’utilisation des ressources pour l’ensemble des cellules le composant, on se retrouve avec des avantages similaires
à la mise en place de colonies (voir Mise en place de colonies).
Les thalles forment aussi une adaptation aux niches écologiques, les ramifications ou la forme en lame permet d’augmenter la surface d’échange pour la
photosynthèse. La portance par l’eau peut aussi être assurée par des flotteurs : les ballonnets (fucus) alors que les crampons fixent le tout sur le substrat.
Le règne fongique, les Eumycètes :
Présentions d’organismes :
Le règne fongique est composé de différents organismes unis ou multicellulaires. Nous avons pu par exemple observer des cellules de levures :
champignons unicellulaires de forme arrondie, les Hyphes des xanthoria qui donnaient une forme aplanie avec plusieurs couches de cellules (ressemblant
à certains thalles d’algues) et pour finir des champignons de Paris avec une forme de « champignon » comme elle est désignée couramment.
Comme chez les algues on passe d’unicellulaires à une forme multicellulaire plus complexe (cladothalle), il en va de même pour les champignons. On
observe donc des ensembles de filaments donnant un mycélium, cette forme peu prendre une structure plus « classique » : le champignon avec un pied
et un chapeau. Néanmoins sur ces différentes formes on note que les cellules sont indifférenciées : il n’y a ni tissu ni organes observables.
14/17
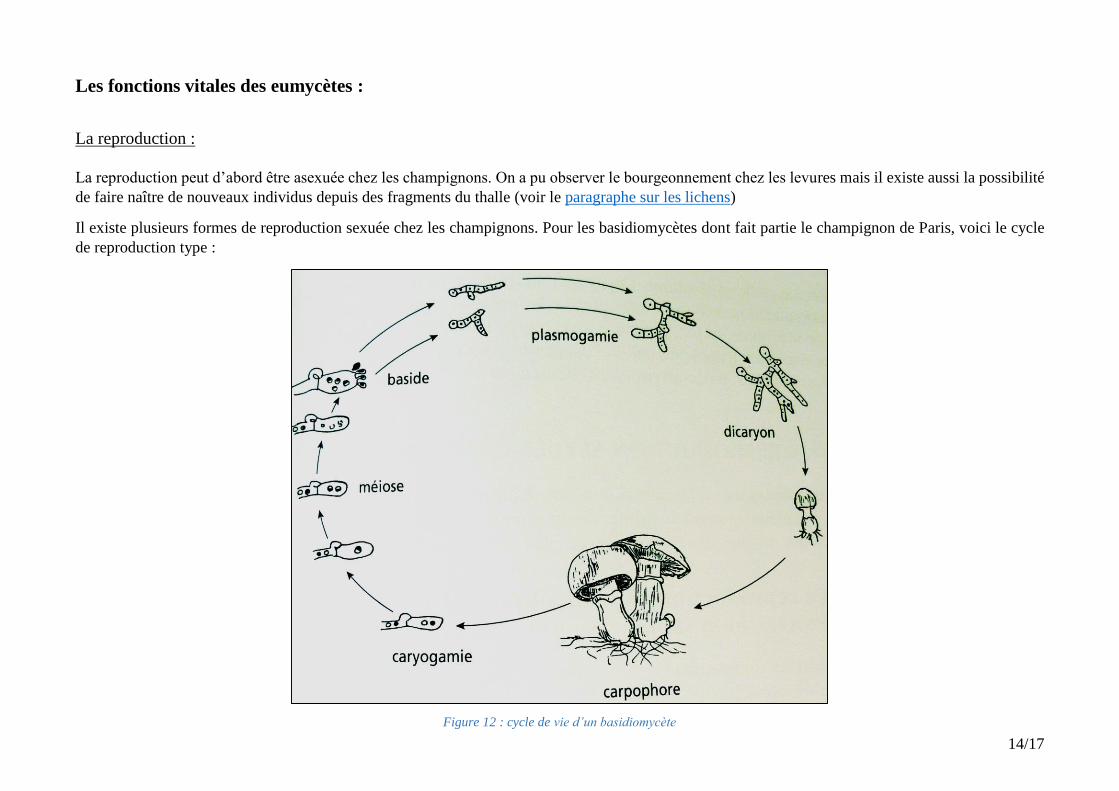
Figure 12 : cycle de vie d’un basidiomycète
Les fonctions vitales des eumycètes :
La reproduction :
La reproduction peut d’abord être asexuée chez les champignons. On a pu observer le bourgeonnement chez les levures mais il existe aussi la possibilité
de faire naître de nouveaux individus depuis des fragments du thalle (voir le paragraphe sur les lichens)
Il existe plusieurs formes de reproduction sexuée chez les champignons. Pour les basidiomycètes dont fait partie le champignon de Paris, voici le cycle
de reproduction type :

15/17
La nutrition :
Les champignons sont hétérotrophes, ils absorbent leurs nourritures directement dans leur milieu (par le mycélium pour les plus complexes).Ce sont
généralement des saprophytes décomposant les déchets organiques de leur milieu, on note donc qu’ils jouent un rôle majeur dans le renouvellement et
le nettoyage du sol (ils sont parmi les rares organismes avec quelques bactéries à pouvoir digérer la lignine).
Nous avons pu observer aussi la symbiose grâce au lichen Xanthoria. Il tire ici profit de l’apport en matière organique de leur symbiote. Cette association
peut se faire au niveau des racines des plantes avec les mycorhizes (endomycorhizes, ectomycorhizes ou ectendomycorhyses).
NB : La limite entre symbiose, commensalisme et parasitisme est mince, laissant ainsi de nouvelles possibilités de nutrition (sans retour bénéfique pour
l’hôte).
Champignon entre plante et animal :
Sur le plan macroscopique, le champignon pourrait comme nous l’avons vu s’apparenter aux plantes et même plutôt aux algues (en considérant les
champignons comme des thalles). Leur reproduction est d’ailleurs assez semblable aux « végétaux primaires ».
Cependant cette parenté est discutée sur le plan cellulaire et moléculaire. Ces organismes possédants à la fois des caractères apparentés aux animaux et
aux végétaux. Leur mode de nutrition est hétérotrophes et donc plutôt semblable aux animaux avec une molécule de réserve typique de ceux-ci : le
glycogène. On note néanmoins des structures types végétales comme la paroi ou la vacuole (mais il faut noter que la paroi est notamment en N-
acétylglucosamine (chitine) rapprochant l’organisme du règne animal).
16/17
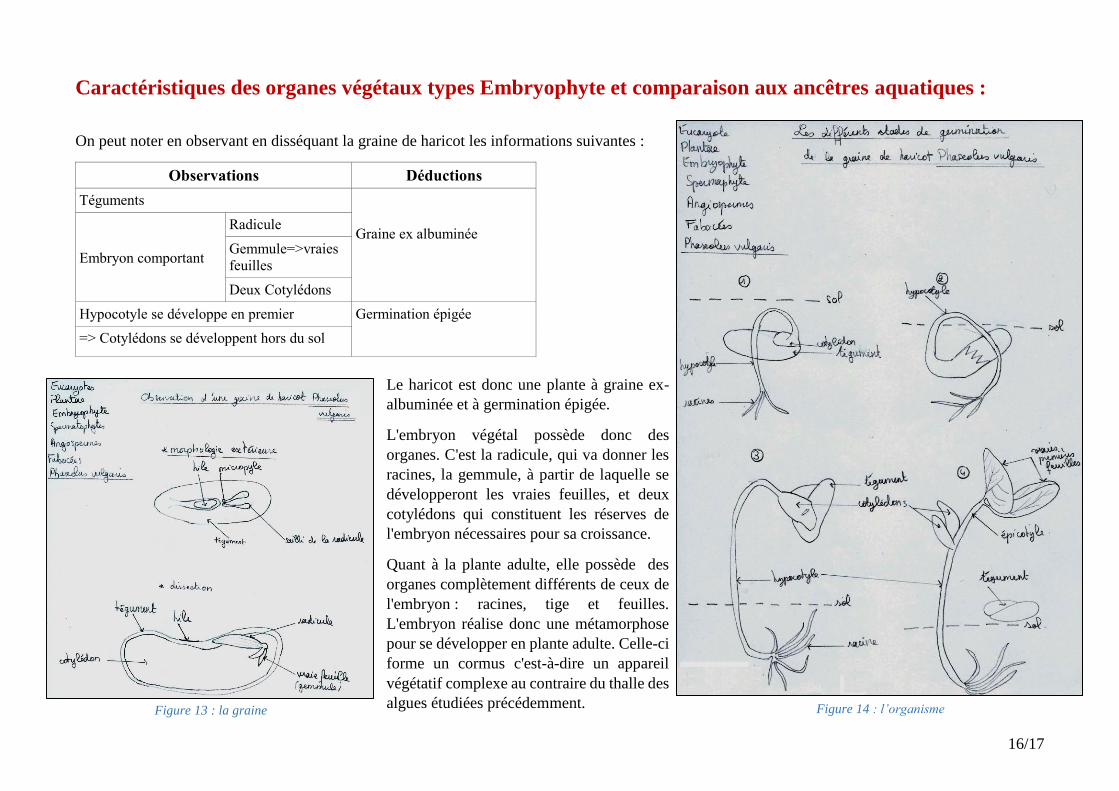
Figure 13 : la graine
Figure 14 : l’organisme
Caractéristiques des organes végétaux types Embryophyte et comparaison aux ancêtres aquatiques :
On peut noter en observant en disséquant la graine de haricot les informations suivantes :
Observations Déductions
Téguments
Graine ex albuminée
Embryon comportant
Radicule
Gemmule=>vraies
feuilles
Deux Cotylédons
Hypocotyle se développe en premier Germination épigée
=> Cotylédons se développent hors du sol
Le haricot est donc une plante à graine ex-
albuminée et à germination épigée.
L'embryon végétal possède donc des
organes. C'est la radicule, qui va donner les
racines, la gemmule, à partir de laquelle se
développeront les vraies feuilles, et deux
cotylédons qui constituent les réserves de
l'embryon nécessaires pour sa croissance.
Quant à la plante adulte, elle possède des
organes complètement différents de ceux de
l'embryon : racines, tige et feuilles.
L'embryon réalise donc une métamorphose
pour se développer en plante adulte. Celle-ci
forme un cormus c'est-à-dire un appareil
végétatif complexe au contraire du thalle des
algues étudiées précédemment.
17/17
Sources
En plus des divers documents mis à notre disposition sur moodle ainsi que les TDs de CELOE nous avons utilisé des sources supplémentaires :
Livre :
Biologie animale des protozoaires aux métazoaires épithélioneuriens, André BEAUMONT et Pierre CASSIER, 2004
Biologie végétal, Jean-Claude LABERCHE, 2001
Site internet :
http://www.digimages.info/listeoiseauxmonde/phylo/phylo.html
http://www.cvc.u-psud.fr/spip.php?article138
https://www.euglena.jp/en/company/
http://www.lesbeauxjardins.com/cours/botanique/6-lichens/usages.htm
__________________________________