Embed Size (px)
Citation preview

Laboratoire de Physiologie Vegthale, Faculte des Sciences, Universite de Bretagne Occidentale, Brest, France
Contribution a l'etude de la physiologie des transports a longue distance du chlore 3°CI dans la plante
Long Distance Transport of Foliar Applied 36Cl
MICHEL PENOT et JACQUELINE GALLOU
Avec 11 figures
Res:u Ie 16 Mars 1977 . Acceptc Ie 8 J uin 1977
Summary
The long distance transport of foliar applied 36CI is preferentially acropetal although growing roots may also represent accumulative centers. This transport takes place in the phloem tissue, as is demonstrated by heat girdling or by cycloheximide pretreatment.
The long distance transport of 32p or 35S in isolated leaves is sensitive to phytohormone treatment (IAA - BAP - or GA3); that of a6CI is not. Therefore, transport of a6CI is not an hormone-directed transport.
However, redistribution of the element is dependent on the metabolism of the plant as a whole (sensitivity to light) and, more particularly, on the metabolism of the conducting tissues (reversible sensitivity of the phloem to local application of CHM).
Moreover, leaf age must be taken into account with respect to the clearing of the element from the source leaf.
This quite original physiology of long distance transport is discussed with regards to the role played by chloride in the plant.
Key words: Long distance transport, phloem, chloride, hormones.
Introduction
On sait que Ie chlore est un element mobile dans Ie phloeme depuis les resultats preliminaires de BUKOVAC et WITTWER (1957) et de PENOT (1964 a et b). Toutefois, Ie chlore presente, vis-a-vis des transports a longue distance, une physiologie originale. C'est ainsi que si Ie gradient de concentration cree au point d'apport (feuille donatrice = F.D.) influe sur la redistribution ulterieure de nombreux ele-
Abreviations: AlA: acide fJ indolylacetique; BAP: benzyl-aminopurine; CHM: cycloheximide; FAP: furfuryl-aminopurine (= kinetine); FD: feuille donatrice de radioelement; GAa: acide gibberellique.
z. Pjlanzenphysiol. Bd. 85. S. 201-214.1977.

202 MICHEL PENOT et JACQUELINE GALLOU
ments (P-S-Rb-Mo-Na), cette regie n'est pas SUI vie dans Ie cas du chlore, la quantite vehiculee restant toujours la meme pour une masse receptrice don nee (PENOT, 1972). Seule la masse du n!cepteur peut augmenter Ie vidage foliaire (PENOT, 1972).
C'est ainsi egalement que I'effet attractif developpe par la kinetine (furfurylaminopurine = FAP), s'exerce a l'egard de nombreux ions ou molecules organiques (MOTHES et a!., 1959; PENOT, 1965; MULLER et LEOPOLD, 1966) mais ne s'exprime pas dans Ie cas du chlore (PENOT, 1964 a).
Ce comportement assez original du chlore meritait donc un examen plus attentif de cette physiologie des transports a longue distance, d'autant plus que Ie chlore est un element dont les liens avec Ie metabolisme cellulaire restent obscurs, sauf peutetre en ce qui concerne la photosynthese (BOARDMAN, 1975).
Par contre, c'est un element dont les modalites de I'absorption cellulaire sont assez bien dHinies et dont on sait que cette absorption depend du metabolisme (necessite d'une depense d'energie - PITMAN et a!., 1974; JOHANSEN et LUTTGE, 1974, 1975).
C'est pour ces differentes raisons que nous avons essaye de repondre a un certain nombre de questions du point de vue de la physiologie des transports a longue distance de cet element, en la comparant, au besoin a des resultats obtenus avec des elements dont la mobilite liberienne est mieux connue, par exemple:
La migration du chlore est-elle de type oriente ou diffus? La redistribution est-elle essentiellement localisee au phloeme? Le transport est-il sensible a l'action de diverses phytohormones? La migration, dans Ie phloeme, est-elle dependante du metabolisme des tissus conducteurs?
- Le metabolisme propre de la F. D. influe-t-il sur la mobilite du chlore?
Materiel et methodes
Les travaux ont ete entrepris avec un materiel particulierement bien adapte a ce genre d'etudes, a savoir des rameaux de Tradescantia viridis, obtenus en culture clonale, en serres. Les plantes sont utilisees soit sous formes de rameaux isoles pourvus de 7 a 8 feuilles, bouture ou non sur Knop 1/2, soit sous forme de rameaux courts reduits a une seule feuille (v. PENOT, 1972).
Le chlore 36CI est apporte, en solution, apres piqlire a une F. D. a la dose et a la concentration sui vantes :
50 ,ul de KCI a 0,1 mM I-I, contenant 10 ,uCijml de 36CI. Ce depot, limite en volume, evite la formation d'un gradient hydrostatique eJeve, favorable a une migration toujours possible par Ie xyleme (PENOT, 1956).
La duree de migration varie entre 1 jour et 3 jours; elle se fait dans les conditions du laboratoire a la lumiere ou a l'obscurite. Les deplacements de la radioactivite sont mesures soit par la methode autoradiographique (Films Kodak-Regulix - duree d'exposition: 8 a 15 jours), soit par comptage au Compteur GEIGER-MULLER apres incineration du materiel en presence de carbonate de Na (BRUNEL, 194 8).
Quelques experiences compIementaires font appel a I'utilisation du 355 (S04Na2-10 a 20 ,uCijml) ou du 32p (NaH2P04 - 10 a 20 /lCi/I). Enfin quelques experiences de migration sont faites sur des feuilles isolees de Pelargonium (v. PENOT et BERAUD, 1977).
z. P/lanzenphysiol. Bd. 85. S. 201-214.1977.

Transport a longue distance du 3"CI 203
Resultats
1. La migration du 36Cl est-elle orientee? Influence des Recepteurs
a) Deplacement apres apport foliaire
Apres apport foliaire, on constate que Ie 35CI suit Ie mouvement de redistribution generalement observe chez Ie T. viridis pour de nombreux elements (P-S-Rb-Ca etc.) a savoir que la migration est essentiellement dirigee vers Ie bourgeon apical (Fig. 1) .
. -..................
\
.-t' .... ,I
Fig. 1: Migration comparee de deux radioeIements apportes par voie foliaire Ie 35S et Ie 36CI (Duree de migration: 3 j, en photoperiode normale). Noter la migration pre£erentiellement acropere dans les 2 cas.
b) Accumulation ou non accumulation tissulaire
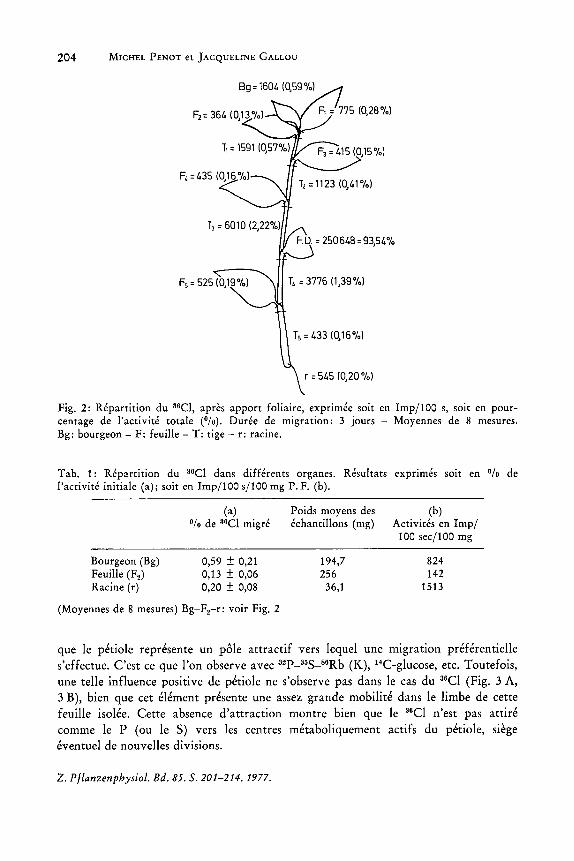
Des dosages quantitatifs (Fig. 2) confirment cela, en meme temps qu'ils indiquent (Tableau 1) que les zones en croissance (bourgeons ou racines) representent, a masse egale, des lieux d'accumulation par rapport aux tissus non en croissance.
Si cette accumulation se cons:oit c1airement dans Ie cas des elements metabolisables (P ou Spar exemple), incorpores au niveau des materiaux nouveaux, elle est moins facile d'interpretation dans Ie cas d'un ion dont Ie seul lien connu avec Ie metabolisme est celui deja evoque au niveau de la photosynthese.
c) Influence du petiole
Si l'on teste, sur des feuilles isolees, l'influence mobilisatrice de diverses phytohormones (MOTHES et aI., 1959; PENOT, 1965; PENOT et BERAUD, 1977), on s'apers:oit
Z. PJlanzenphysiol. Bd. 85. S. 201-214.1977.
204 MICHEL PENOT et ]ACQUELlNE GALLOU
Fs = 525 (0,19%) T, = 3776 (1,39%)
Ts = 433 (0,16%)
r = 545 (0,20 "to)
Fig. 2: Repartition du 36Cl, apres apport foliaire, exprimee soit en Imp/lOa s, soit en pourcentage de l'activite totale (0/0). Duree de migration: 3 jours - Moyennes de 8 mesures. Bg: bourgeon - F: feuille - T: tige - r: racine.
Tab. 1: Repartition du 36Cl dans differents organes. Resultats exprimes soit en Ofo de l'activite initiale (a); soit en Imp/lOa s/100 mg P. F. (b).
Bourgeon (Bg) Feuille (F2) Racine (r)
(a) Ofo de 36Cl migre
0,59 ± 0,21 0,13 ± 0,06 0,20 ± 0,08
(Moyennes de 8 mesures) Bg-F2-r: voir Fig. 2
Poids moyens des echan tillons (mg)
194,7 256
36,1
(b) Activites en Imp/
100 sec/lOa mg
824 142
1513
que Ie petiole represente un pole attractif vers lequel une migration preferentielle s'effectue. C'est ce que l'on observe avec 32p_35S_B6Rb (K), HC-glucose, etc. Toutefois, une telle influence positive de petiole ne s'observe pas dans Ie cas du 36CI (Fig. 3 A, 3 B), bien que cet element presente une assez grande mobilite dans Ie limbe de cette feuille isolee. Cette absence d'attraction montre bien que Ie 36CI n'est pas attire comme Ie P (ou Ie S) vers les centres metaboliquement actifs du petiole, siege eventuel de nouvelles divisions.
z. Pjlanzenphysiol. Bd. 85. S. 201-214.1977.

Transport a longue distance du 36CI 205
A B
p
Fig. 3 A et 3 B: Influence du petiole (p) sur la migration de 2 radioelements, Ie 32p (3 A) et Ie 36CI (3 B) dans Ia feuiIIe de Pelargcnium. Noter I'absence d'effet positif dans Ie cas du 36CI.
2. La redistribution se fait-elle essentiellement par Ie phloeme?
Pour tester ceLl, nous avons eu recours a 2 series d'experiences, la 1 ere en rapport avec des annelations thermiques, la seconde en relation avec I'influence de l'activite transpiratoire des feuilles.
a) Annelations thermiques
Si l'on detruit Ie phloeme a I'aide d'un jet de vapeur d'eau bouillante, on constate que la migration de 3
6CI est fortement diminuee si ce n'est totalement arr&tee au-dela de cette zone detruite (Fig. 4). Par contre, la voie ligneuse reste fonctionnelle pendant
"7 l···r_ .. ___ .....
... _-_ .... -.-
.... ---............ ---_ .•. :').
Te a
\ .......
\. '-./,: , \ I
I
a -,
a
Fig. 4: Arrh du transport du 36CI, apres ann elation des rameaux par jet de vapeur d'eau bouiIIante (a). Te = temoin non traite.
Z. PJlanzenphysiol. Bd. 85. S. 201-214. 1977.

206 MICHEL PENOT et JACQUELINE GALLOU
la duree de l'experience (maintien de la turgescence foliaire). En corollaire, Ie transport basi pete du 36CI est augmente dans Ie cas de cette annelation.
b) Influence de l'activite transpiratoire des feuilles
Si Ie xyleme joue un rMe important dans la redistribution du 36CI, Ie maintien des rameaux en atmosphere humide (100 % HR) devrait fortement diminuer ces transports en direction apicale par rapport ;\ ceux mesures sur des temoins main tenus en atmosphere normale (68 Ofo HR). Les resultats montrent (Fig. 5 A et 5 B) que les
Fig. 5: Migration comparee de 36Cl en atmosphere saturee (A) ou a 68 Ofo HR (B). On notera une legere diminution des translocations a 100 Ofo HR.
migrations restent importantes en atmosphere humide et sont donc bien l'expression d'une redistribution par voie liberienne. La diminution des transports enregistree en atmosphere humide traduit Ie fait qu'une «fuite» laterale existe entre Ie liber et Ie bois, en accord avec ce qui a dej;\ ete dit anterieurement (PENOT, 1964 b).
3. Transport du aGel et phytohormones
Nous avons dej;\ montre que la FAP pouvait exercer une action mobilis at rice specifique des ions (PENOT, 1964 a) Plus recemment, no us avons repris (PENOT et BERAUD, 1977) ce probleme du contr6le hormonal des transports ;\ longue distance qui reste encore un sujet de controverse actuelle (WARDLAW et MONCUR, 1976).
II nous paraissait donc interessant de mesurer, sur des feuilles isolees de Pelargonium, l'influence attractive de diverses phytohormones (AIA-BAP-GAs) dont on
z. PJlanzenphysiol. Ed. 85. S. 201-214.1977.

Transport a longue distance du 3BCI 207
sait qu'elles exercent une action posltlve a. l'egard de nombreux ions inorganiques (PENOT et BERAUD, 1977) en comparant l'influence de ces phytohormones vis-a.-vis du 3SS, du 32p et du 3BCl.
Le protocole experimental est Ie suivant: - une application de phytohormones est faite en solution aqueuse aux doses suivantes:
AlA (50 mg 1-1) BAP (20 mg I-l) GAs (25 mg 1-1);
une zone opposee de la feuille est traitee a. l'eau distillee et sert donc de temoin. Apres une periode d'attente de 3 jours, un dep8t de radioisotope est fait, a. egale
distance, des zones traitees. La periode de migration est de 3 a. 5 jours. Les autoradio-
A
SAP
S
c
Fig. 6: Influence comparee de diverses phytohormones sur la mohilite du 32p (A) du 355 (B) et du MCI (C). Autoradiographies apres une migration de 3 j. (v. texte).
z. P/lanzenphysiol. Bd. 85. S. 201-214. 1977.

208 MICHEL PENOT et JACQUELINE GALLOU
graphies montrent, dans tous les cas, un effect positif des phytohormones vIs-a-vis du 32P, du 35S, une action nulle vis-a-vis du 3sCl (Fig. 6). Le transport du 36Cl est done insensible a l'action mobilisatrice des 3 groupes de phytohormones testees.
4. Relation entre la migration de 36Cl et le metabolisme
La relation entre Ie transport a longue distance et Ie metabolisme peut &tre mise en evidence soit par une influence du facteur lumiere, soit par une action localisee d'un inhibiteur metabolique. C'est ce que nous avons tente de faire.
a) Influence du facteur lumiere
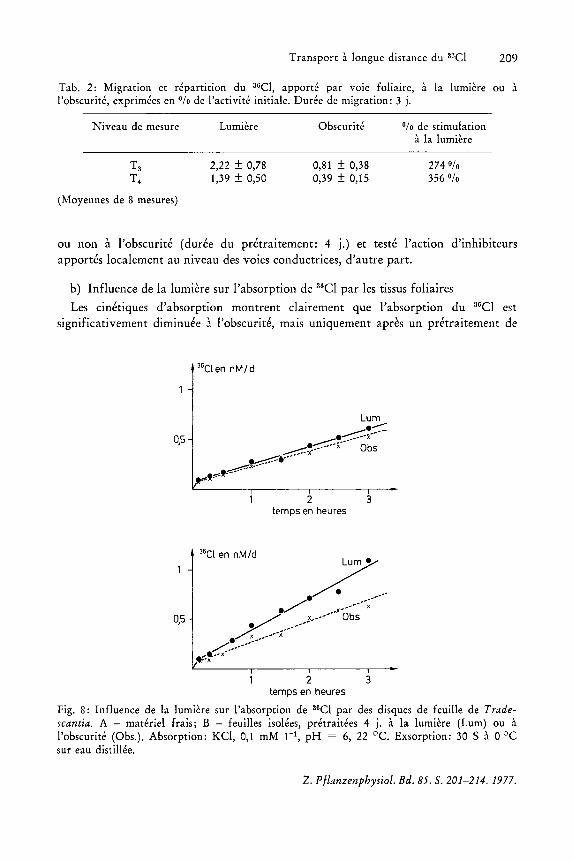
L'absence de lumiere au cours de la periode de transport diminue la redistribution de 36Cl et ceci d'autant plus que Ie materiel a subi un pretraitement de 8 j a l'obscurite (Fig. 7). Des mesures quantitatives confirment ces observations autoradiographiques (Tableau IT).
L 0, \ Fig. 7: Influence de la lumiere sur la migration de 36CI apporte a une feuille. O[ = materiel frais - Migration a l'obscurite - L = materiel frais - Migration a la lumiere. - O2 = pretraitement de 8 j. a I'obscurite - Migration a I'obscurite. Duree de migration: 3 j. dans tous les cas.
II est evident que cette methode ne laisse pas apparahre la part qui revient au metabolisme de la F. D., point de depart de tout transport a longue distance, et au met abo lis me du tissu conducteur Iui-meme.
Pour essayer de dissocier entre ces deux aspects, nous avons mesure, d'une part I'influence de Ia Iumiere sur l'absorption du 3
6CI par des disques de feuilles pretraitees
z. Pflanzenphysiol. Ed. 85. S. 201-214.1977.
Transport It longue distance du 3'CI 209
Tab. 2: Migration et repartition du a6CI, apporte par voie foliaire, a la lumiere ou a I'obscurite, exprimees enOfo de I'activite initiale. Duree de migration: 3 j.
Niveau de mesure
T3 T4
(Moyennes de 8 mesures)
Lumiere
2,22 ± 0,78 1,39 ± 0,50
Obscurite
0,81 ± 0,38 0,39 ± 0,15
% de stimulation ala lumiere
274 Ofo 356 Ofo
ou non a I'obscurite (durt~e du pretraitement: 4 j.) et teste l'action d'inhibiteurs apportes localement au niveau des voies conductrices, d'autre part.
b) Influence de la lumiere sur l'absorption de a6CI par les tissus foliaires
Les cinetiques d'absorption montrent clairement que I'absorption du 3GCI est significativement diminuee J. l'obscurite, mais uniquement apres un pretraitement de
36CI en nMI d
0,5
2 3 temps en heures
0,5
7 .. /e ....... .
• / .~x ... -.. x
~ } •••• , Obs
e •• ' x ..... - .. x"' .. -
/.~ ........ . ~~~ .... x"
36CI en nM/d
2 3 temps en heures
Fig. 8: Influence de la lumiere sur I'absorption de 3sCI par des disques de feuille de Tradescantia. A - materiel frais; B - feuilles isolees, pretraitees 4 j. a la lumiere (Lum) ou a I'obscurite (Obs.). Absorption: KCI, 0,1 mM r 1, pH = 6, 22°C. Exsorption: 30 SaO °C sur eau distillee.
z. PJlanzenphysiol. Bd. 85. S. 201-214.1977.

210 MICHEL PENOT et JACQUELINE GALLOU
4 j. des feuilles (Fig. 8), c'est-a-dire dans des conditions qui diminuent Ie potentiel energetique des tissus.
c) Influence de la cycloheximide sur Ie transport a longue distance
Nous avons deja montre que la cycloheximide (CHM), inhibiteur de la proteosynthese, pouvait exercer une action inhibitrice sur les transports a longue distance (PENOT et al., 1976). Ici encore, on constate que la CHM, a condition qu'elle ait ete appliquee en pretraitement pendant 48 h, arrhe, de fas:on tres nette, Ie transport du 36CI, au-dela de la zone traitee (Fig. 9). Toutefois cet arrh peut ~tre interprete comme un effet generalise et non comme un effet local au niveau des voies conductrices. C'est pour cette raison que nous montrons (Fig. 10) la reversibilite de
'.
/' A
\ . \
CHM SOmgll
B
Fig. 9 A et 9 B: Influence de la CHM (50 mg 1-1) sur Ie transport de 36Cl, apporte par voie foliaire. A - temoin sur eau distillee (e. d.); B - pretraitement a la CHM de 2 j. et action pendant la phase de migration. Duree de la migration: 2 j. A noter la migration a predominance basiphe en B.
l'inhibition, apres un pretraitement de 4 j. a la CHM, suivi d'une periode de bouturage de 3 semaines sur Knop 1/2. Le fait que Ie 36CI soit a nouveau transporte vers Ie bourgeon apical traduit bien Ie caractere reversible de l'action inhibitrice precedemment mise en evidence.
Z. P/lanzenphysiol. Bd. 85. S. 201-214. 1977.

Transport a longue distance du 36CI 211
. ---'---_ .. --, _ .. ----_.,
/ ,,, ............. ------~ -;.~
/ ... i ' .. ---\ ; ....... -:..-
V e.d
i \. \
CHM SOmg/l \ I
/ I : \ I
\/ A
~ \ ... _--. "--.. ...... - B ,
Fig. IDA et lOB: Caractere reversible de I'inhibition due a la CHM sur Ie transport du 36CI, apporte par voie foliaire. A - temoin sur eau distillce (e. d.) - r = racines; B - petraitement de 4 j. a la CHM, suivi d'un bouturage de 3 semaines. Comparer avec la figure 9. A noter que la migration redevient acrophe en B.
5. Transport et age de la feuille donatrice
Nous avons compare, sur des rameaux courts de T. viridis, munis d'une seule feuille (v. PENOT, 1972) la redistribution du 36CI en fonction de !'age de la F.D. (feuille jeune, adulte ou senescente). Les resultats indiquent (Fig. 11) que c'est la feuille jeune qui est la plus exportatrice de 36CI, contrairement au schema generalement admis, en particulier avec les assimilats (v. CRAFTS et CRISP, 1971; TURGEON etWEBB,1973).
Discussion et Conclusions
Ces resultats confirment, en particulier grace aux experiences d'annelation thermique et grice it l'emploi de la CHM, la mobilite, dans Ie phloeme, du 36Cl apporte par voie foliaire. Ils montrent en outre que la redistribution suit les regles generales auxquelles obeissent les differents elements nutritifs, c'est-a-dire que la migration est preferentiellement orientee vers les organes recepteurs (bourgeons, racines) qui peuvent meme representer des centres d'accumulation, tout comme on l'observe chez
Z. PJlanzenphysiol. Ed. 85. S. 201-214.1977.

212 MICHEL PENOT et JACQUELINE GALLOU
5
4
3
2 J I~l
I F.D. F.D. F.D.
jeune adulte senescente
Fig. 11: Redistribution de 3BCl apporte par voie foliaire en fonction de 1';1ge de la F. D. Resultats exprimes en % de l'activite totale. (Moyennes de 12 mesures.)
Ie m~me materiel, avec Ie 3:;S, Ie 32P, Ie 86Rb et m&me Ie <SCa (v. PENOT et aI., 1976; PENOT et BERAUD, 1977).
Toutefois, au niveau d'une feuille isolee (Fig. 6), a la difference de ce que nous observons avec d'autres ions ou molecules, la migration ne se fait pas vers des centres metaboliquement actifs (petiole ou centres attractifs induits par diverses phytohormones); ceci veut dire que la migration orientee (effet MOTHES) reste une caracteristique essentielle d'un type d'ion ou de molecule.
Cela pose la question de la signification physiologique du chlore et de sa migration preferentiellement acropete. On sait que cet element mineral est implique dans Ie metabolisme photosynthetique (BOARDMAN, 1975), mais a des doses qui n'expliquent pas les quantites de chlore presentes dans la cellule.
L'absence de flux oriente, stimule sous l'influence des phytohormones, laisse supposer qu'il n'y a pas synthese de transporteur, camme on pourrait Ie penser dans Ie cas des effets positifs obtenus avec d'autres elements (35
S_32p par exemple). Cette absence d'effet des phytohormones sur Ie transport du 36CI opposable a une
migration orientee vers Ie bourgeon apical no us conduit a penser que Ie r6le essentiel du CI est d'assurer Ie maintien des equilibres electrochimiques au niveau cellulaire.
Neanmoins l'absorption cellulaire du 36CI et sa redistribution a longue distance sont directement dependantes du metabolisme cellulaire; ceci est atteste par de
Z. Pflanzenphysiol. Bd. 85. S. 201-214.1977.

Transport 11 longue distance du 36CI 213
nombreux tra vaux en ce qui concerne Ie premier point (PITMAN et aI., 1974; J OHANSEN et LUTTGE, 1974, 1975), par une diminution importante des migrations a. l'obscurite (v. Fig. 7), en ce qui concerne Ie second point. Toutefois, il est difficile d'imputer la cause de cette diminution a. l'obscurite, soit a. un ralentissement des processus de secretion «active» au nive:lU de la feuille donatrice, soit a. une action localisee au niveau du metabolisme du tissu conducteur lui-meme.
L'action inhibitrice mise en evidence apres application de CHM au niveau des voies conductrices (Fig. 9), ainsi que Ie caractere reversible de cette inhibition (Fig. 10) ajoutent a. l'hypothese deja. avancee (PENOT et aI., 1976) d'une participation activite des proteines dans Ie fonctionnement du tube crible.
II est apparu egalement que trois facteurs pouvaient influer sur Ie taux de vidage d'une F. D. a. savoir: 1. L'age de la feuille, Ie taux d'exportation diminuant significativement au cours
de l'avancement en age; ceci est contraire a. ce que I'on observe habituellement avec les assimilats (TURGEON et WEBB, 1973), mais est con forme a. d'autres observations faites avec d'autres elements mineraux (Rb-Mo-P, v. PENOT, 1974 et 1976).
2. L'influence de la masse du recepteur rattachee a. la F. D. (PENOT, 1972). 3. Enfin l'influence d'un flux transpiratoire, consequence d'une fuite laterale du CI
depuis Ie phloeme vers Ie xyleme; une augmentation des translocations en cas d'activite transpiratoire elevee expliquerait ceia. (v. Fig. 5). Le vidage moindre de la feuille adulte ou de la feuille senescente par rapport a.
la feuille jeune (Fig. 11) montre la. encore la non-dependance du transport a. longue distance du CI vis-a.-vis d'un controle hormonal; il est, en effet admis qu'une richesse hormonale locale se traduit au niveau de la F. D. par une retention accrue de 32p
par exemple (PENOT, 1965) corollaire des effets attractifs deveioppees a. distance par diverse phytohormones (effet MOTHES).
Ceci nous confirme dans I'idee que la migration du CI n'est pas directement controlee par l'activite metaboEque des centres recepteurs actifs mais qu'elle represente plutot une mise en equilibre electrochimique d'origine secondaire.
Bibliographie
BOARDMAN, N. K.: Trace elements in photosynthesis. p. 199-212. In: Trace elements in soilplant-animal systems. NICHOLAS, D.]. D., A. R. EGAN, ed. - Academic Press (1975).
BRUNEL, A.: Traite pratique de chimie vcg,hale, Tome II, Ed. GEORGE FRERE - Tourcoing, 513 p. (1948).
BUKOVAC, M. ]. and S. H. WITTWER: Absorptio:J. and mobility of foliar applied nutrients. Plant Physiol. 32, 428-435 (1957).
CRAFTS, A. S. and C. E. CRISP: Phloem transport in plants. W. H. FREEMAN et Cie. - San Francisco, 481 p. (1971).
MACDoNALD, I. R. and A. E. S. MACKLON: Light-enhanced chloride uptake by wheat Laminae. Plant. Physiol. 56, 105-108 (1975).
MACDoNALD, I. R., A. E. S. MACHON and R. W. G. MACLEOD: Energy supply and lightenhanced chloride uptake in wheat Laminae. Plant Physiol. 56, 699-702 (1975).
z. P/lanzenphysiol. Ed. 85. S. 201-214.1977.

214 MICHEL PENOT et JACQUELINE GALLOU
JOHANSEN, C. and U. LUTTGE: Respiration and photosynthesis as alternative energy sources for chloride uptake by Tradescantia albi/lora leaf cells. Z. Pflanzenphysiol. 71, 3, 189-199 (1974).
- - A comparison of potassium and chloride uptake by Tradescantia albi/lora leaf. Cells at different KCl concentrations. Aust. J. Plant Physiol. 2, 471-479 (1975).
MOTHES, K., L. ENGELBRECHT und O. KULAJEWA: Dber die Wirkung des Kinetins auf Stickstoffverteilung und Eiwei6synthese in isolierten BHittern. Flora 147, 445-464 (1959).
MULLER, K. and A. C. LEOPOLD: The mechanism of kinetin-induced transpOrt in corn leaves. Planta (Berl.) 68, 186-205 (1966).
PALLAGHY, C. K.: Electron probe microanalysis of potassium and chloride in freeze-substituted leaf sections of zea mays. Aust. J. bioI. Sci. 26, 1015-1034 (1973).
PENOT, M.: Recherches sur les voies de circulation des sels, dans les pi antes, d'apres des experiences d'isotopes radioactifs. Rev. gen. Bot. 63, 545-550 (1956).
- Mouvements liberiens du phosphore lies a l'apport de cinetine. C. R. Acad. Sci. (Paris) 256,488-490 (1963).
- Action comparee de la kinetine sur Ie mouvement liberien du 36Cl et du 32P. C. R. Acad. Sci. (Paris) 258, 2881-2883 (1964 a).
- Etude comparee des deplacements liberiens du chlore a6Cl et du phosphore 32P, chez Ie Tradescantia viridis. C. R. Acad. Sci. (Paris) 259, 868-870 (1964 b). Etude du role des appels moleculaires dans la circulation liberienne. Physiol., veg. 3, 41-89 (1965).
- Migrations liberiennes chez Tradescantia viridis. Relation entre les quantites migrees et l'apport initial aux feuilles. Physiol., veg. 10, 687-696 (1972).
- Modifications de la permeabilite et du transport du molybdate au cours du vieillissement foliaire. C.R. Acad. Sci. (Paris) 278, 2779-2782 (1974).
- Evolution de l'absorption et du tau x d'exportation du 86Rb, au cours de la senescence foliaire. C. R. Acad. Sci. (Paris) 282, 855-858 (1976).
PENOT, M. and J. BERAUD: Phytohormones et transport oriente au niveau de la feuille isolee de Pelargonium zonale. Competition phytohormonale. BioI. Plantarum (sous-presse) (1977).
PENOT, M., J. Y. FLOC'H et M. PENOT: Etude comparee de l'absorption et de la redistribution du 45Ca chez divers groupes de vcgetaux. Planta (Berl.) 129,7-14 (1976).
PITMAN, M. G., U. LUTTGE and A. LXUCHLI and E. BALL: Ion uptake to slices of barley leaves, and regulation of K content in cells of the leaves. Z. Pflanzenphysiol. 72, 75-88 (1974).
TURGEON, R. and J. A. WEBB: Leaf development and phloem transport in Cucurbita pepo. Transition from import to export. Planta 113, 179-191 (1973).
VAN STEVENINCK, R. F. M., B. BALLMENT, P. D. PETERS and T. A. HALL: Ultrastructural localization of ions. II. X-ray analytical verification of silver precipitation products and distribution of chloride in mesophyll cells of barley seedlings. Aust. J. Plant Physiol. 3, 359-365 (1976).
WARDLAW, 1. F. and L. MONCUR: Source, sink and hormonal control of translocation in wheat. Plant a 128, 93-100 (1976).
Prof. M. PENOT, Universite de Bretagne Occidentale, Laboratoire de Physiologie vegetale, Avenue Victor Le Gorgeu, F - 29283 Brest Cedex, France.
Z. P/lanzenphysiol. Bd. 85. S. 201-214. 1977.





























