Embed Size (px)
Citation preview

Cours Module L5-BH-05
Régulation de l’expression des génomes
Cours n°3
S. Bourgerie Université d’Orléans – UFR Sciences & Centre de Biophysique Moléculaire UPR4301

Plan du cours n°3Opéron arabinose
1- introduction – comparaison2- organisation3- régulateur AraC : fonctions4- répression catabolique5- fonctionnement : + - inducteur
Rappels sur terminaison de la transcription1- terminaison rho-indépendante2- terminaison rho-dépendante : structure-fonction de rho
Stratégies de modulation de la terminaison1- définition : antiterminaison-atténuation2- quelques exemples
Opéron tryptophane1- organisation2- régulation
mécanisme 1 : interaction répresseur-opérateurmécanisme 2 : atténuation transcriptionnelle

Opéron arabinoseUn exemple de mécanisme de régulation transcriptionnelle
positive & négative de l’activité de gènes

introduction
arabinose : sucre à 5 carbones.
opéron arabinose : 3 gènes structuraux, araBADcontrôlés par le régulateur général CRP et par une protéine spécifique AraC.
gènes structuraux : codent pour les enzymes qui permettent la transformation du L-arabinose en D-xylulose-5-phosphate.
C
C
HO
OHH
C HHO
C HHO
CH2OH
CH2OH
C O
C OHH
C HHO
CH2OH
CH2OH
C O
C OHH
C HHO
CH2 O P
O
O-
O-
CH2OH
C O
C HHO
C OHH
CH2 O P
O
O-
O-
L-arabinose L-ribuloseL-ribulose-5-phosphate D-xylulose-5-phosphate
Le D-xylulose-5-phosphate rejoint la voie des pentoses phosphate, productrice d'énergie pour la cellule.

The genes required for the uptake and catabolism of L-arabinose in Escherichia coli, the operon structures, their approximate locations on the 100-minute circulargenetic map, and the initial steps of arabinose catabolism showing the structures of the intermediatesand the steps catalyzed by the products of the araoperon geneproducts. AraC acts positively and negatively at pBAD, and negatively at pC.
*

Organisation de l’opéron arabinose
PC
tPBAD t
araC araB araA araD
Activateur/ Répresseur
I1 I2
AraC dimer
CRP
O2 O1

Le régulateur AraC
• Structure– homodimère– avec un domaine de fixation à l’ADN (côté C-ter.)– avec un site de fixation pour l’arabinose (activateur) (côté N-
ter.)
• Fonction- régulateur négatif
• se fixe à araO2 & araI1- régulateur positif
• se fixe à la fois aux demi-sites araO1 & araI1 & I2
- régule sa propre synthèse (autorégulation)• se fixe à araO1
- provoque la formation de boucles dans l’ADN
*

Répression Catabolite
(a)
Glucose
L-arabinose
Con
cent
ratio
n en
sucr
e
Nom
bre
de c
ellu
les
(b)
Glucose L-arabinose
Nom
bre
de c
ellu
les
Temps Temps

Effets de la source de carbone sur la transcription d’araBAD
Glucose Arabinose araBADfort faible réprimé
faible fort activéfort fort réprimé
absent absent réprimé
CRP
P t
Pt
araC araB araA araD
O2
I1 I2
O1
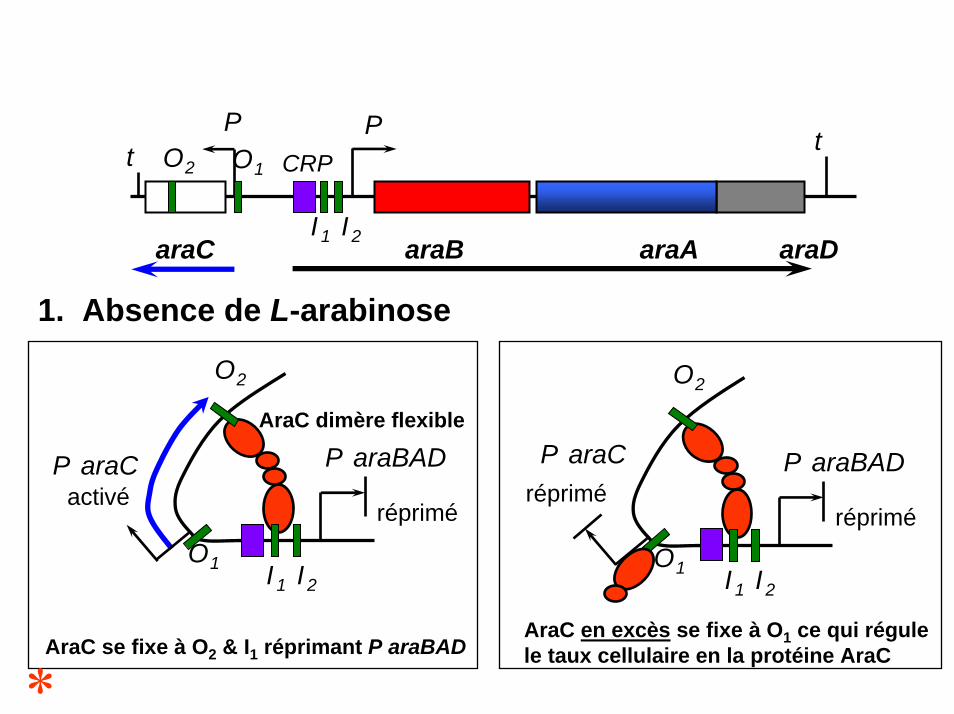
1. Absence de L-arabinose
AraC dimère flexible
O2
I1 I2
O1
répriméactivé
P araBADP araC
AraC se fixe à O2 & I1 réprimant P araBAD
O2
I1 I2
O1
répriméréprimé
P araBADP araC
AraC en excès se fixe à O1 ce qui régulele taux cellulaire en la protéine AraC
*

P t
Pt
araC araB araA araD
O2
I1 I2
O1 CRP
I1 I2
induit(activé)
P t O2 O1
P araBAD
2. + Inducteur L-arabinose+ CRP-AMPc
AraC se fixe à I2 & I1 activant P araBAD
*
En résumé
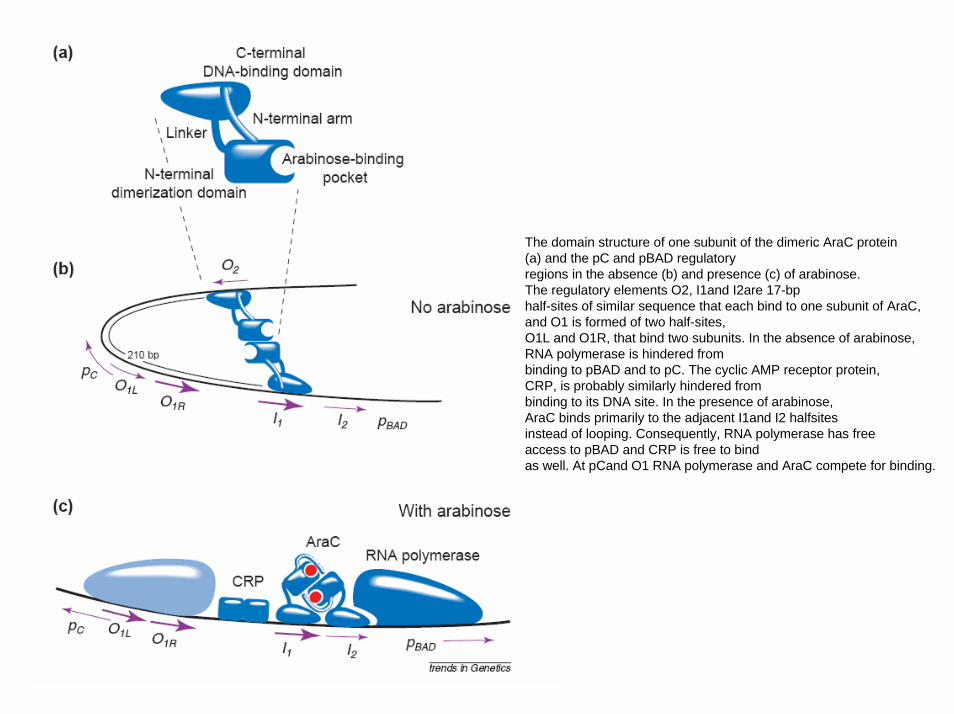
The domain structure of one subunit of the dimeric AraC protein(a) and the pC and pBAD regulatoryregions in the absence (b) and presence (c) of arabinose. The regulatory elements O2, I1and I2are 17-bphalf-sites of similar sequence that each bind to one subunit of AraC, and O1 is formed of two half-sites,O1L and O1R, that bind two subunits. In the absence of arabinose, RNA polymerase is hindered frombinding to pBAD and to pC. The cyclic AMP receptor protein, CRP, is probably similarly hindered frombinding to its DNA site. In the presence of arabinose, AraC binds primarily to the adjacent I1and I2 halfsitesinstead of looping. Consequently, RNA polymerase has free access to pBAD and CRP is free to bindas well. At pCand O1 RNA polymerase and AraC compete for binding.

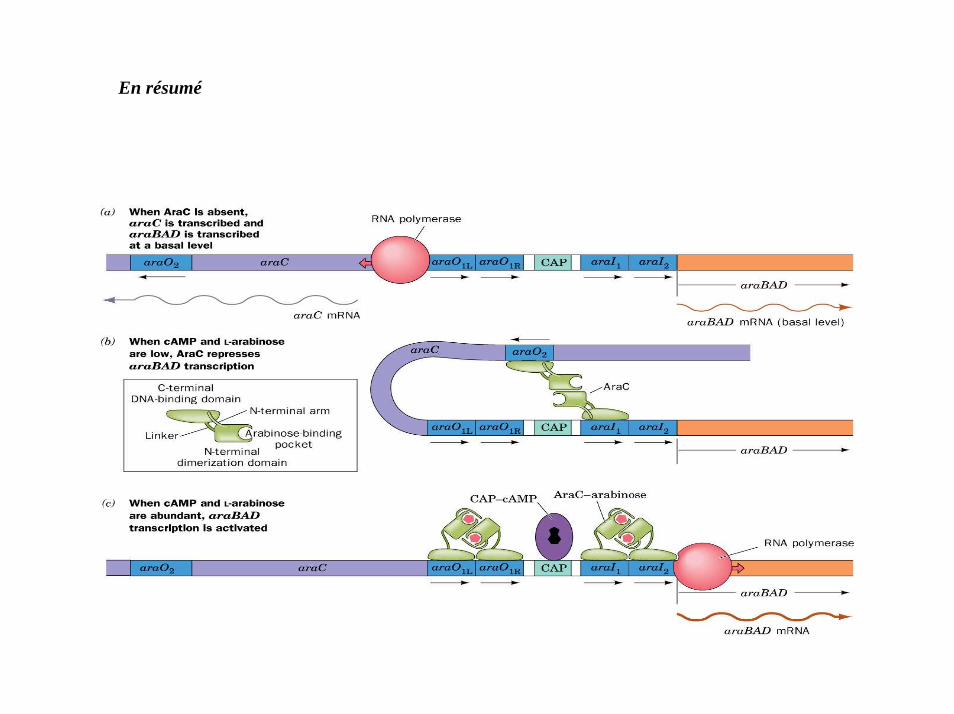
1. L’arabinose est un régulateur positif de la transcription.
2. En l’absence d’arabinose, AraC se fixe aux sites régulateurs I1 & O2 ce qui réprime la transcription de l’opéron AraBAD.
3. Le taux d’AraC est autorégulé, lorsqu’un excès d’AraC se fixe en O1
4. Quand l’arabinose se fixe à AraC, AraC se fixe à I1 & I2.
5. Quand le Glucose est absent, le taux d’AMPc s’élève et le complexe cAMP-CRP se fixe sur uneséquence adjacente à I1.
Il y a expression de l’opéron AraBAD.
En résumé
*

3 différences importantes existent entre les opérons lac et ara :
1- la protéine AraC peut fonctionner à la fois comme répresseur et activateur pour l’expressiondes gènes araBAD;
2- la protéine AraC régule également sa propre synthèse, réprimant sa propre transcription.
3- l’opéron ara constitue un exemple de régulation « menée » à distance du fait de la formation d’une boucle dans l’ADN

Terminaison de la transcription : quelques rappels

2 modes de terminaison de la transcription
• arrêt de la transcription quand l’ARN polyméraserencontre un terminateur
• arrêt de la polymérisation ; départ de l’ARNm ; dissociation de l’ADN.
• terminateurs formés d’une boucle dans l’ARN.• 2 types:
– terminaison facteur-indépendant– terminaison facteur-dépendant
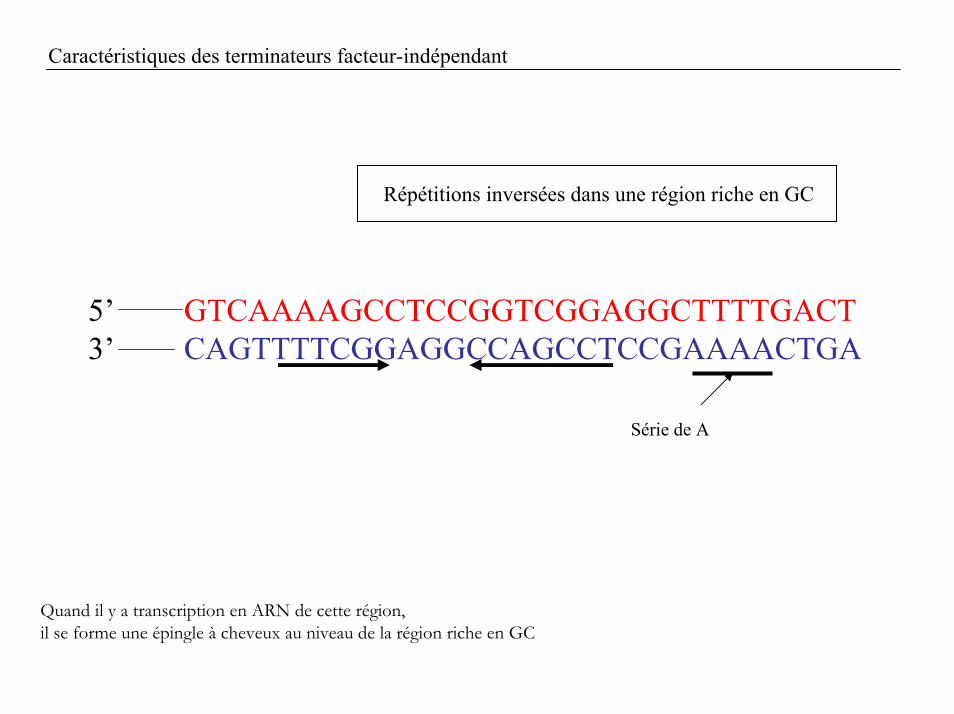
Caractéristiques des terminateurs facteur-indépendant
Répétitions inversées dans une région riche en GC
5’ GTCAAAAGCCTCCGGTCGGAGGCTTTTGACT3’ CAGTTTTCGGAGGCCAGCCTCCGAAAACTGA
Série de A
Quand il y a transcription en ARN de cette région, il se forme une épingle à cheveux au niveau de la région riche en GC
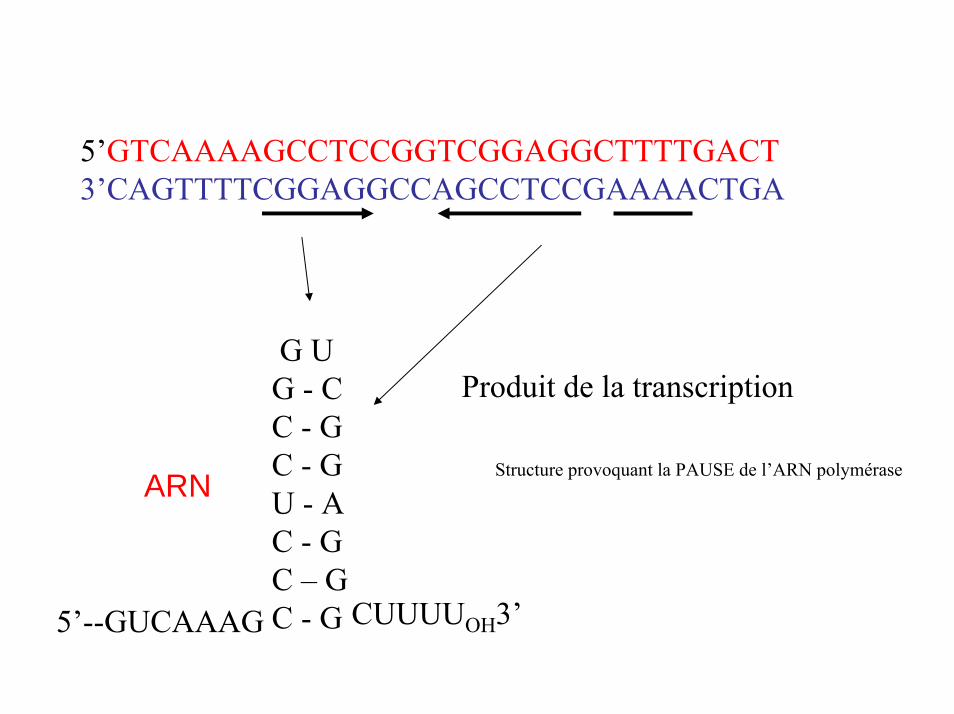
5’GTCAAAAGCCTCCGGTCGGAGGCTTTTGACT3’CAGTTTTCGGAGGCCAGCCTCCGAAAACTGA
Produit de la transcriptionG UG - CC - GC - GU - AC - GC – GC - G
Structure provoquant la PAUSE de l’ARN polyméraseARN
CUUUUOH3’5’--GUCAAAG

Séquences de deux terminateurs intrinsèques naturels λtR2 (A) et rrnBT2 (B) (lesflèches indiquent les sites de terminaison).
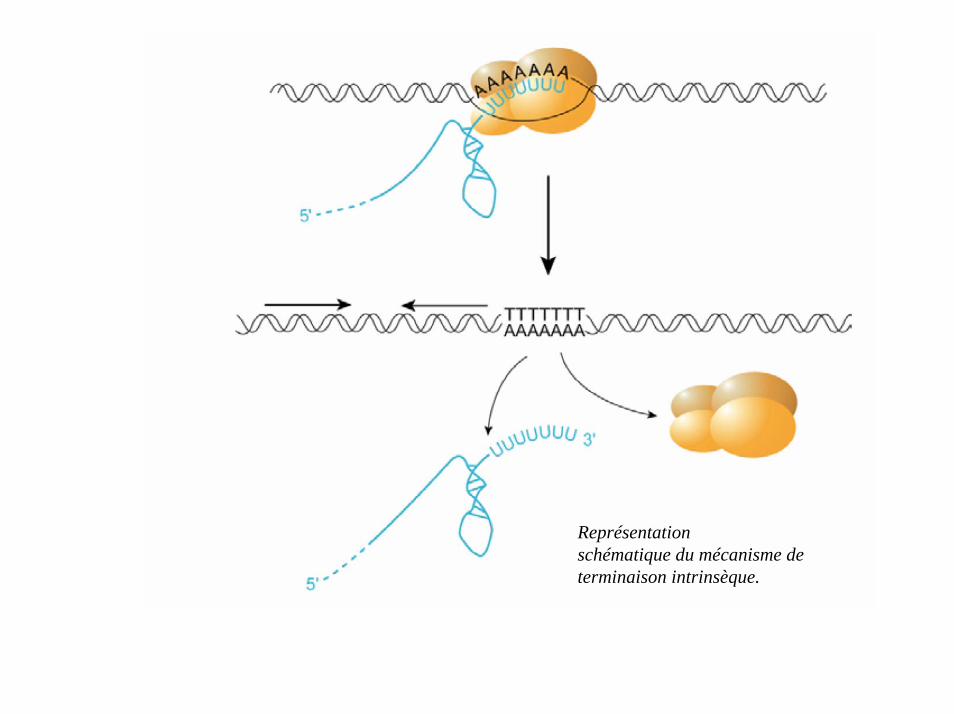
Représentationschématique du mécanisme determinaison intrinsèque.

Terminaison facteur-dépendant
• requiert des protéines qui interagissentavec la polymérase ou le transcript provoquant l’arrêt de la transcription
• E. coli possède 3 facteurs de terminaison :– Rho– Tau– NusA
• Rho (ρ) : mieux compris

Terminaison Rho-Dépendante
• Mode de terminaison qui ne fonctionne que sil’ARN produit n’est pas en cours de traduction (ribosomes absents)
• Rho est une ATPase ARN-dépendante; réalise l’hydrolyse de l’ATP si l’ARN estprésent
• Rho est une hélicase RNA-DNA (protéinehexamere); déroule le duplex ARN-ADN
• Un site de pause est nécessaire.

le terminateur de la transcription Rho
Protéine hexamérique ayant une affinité particulièrepour les régions riches en C de l’ARN
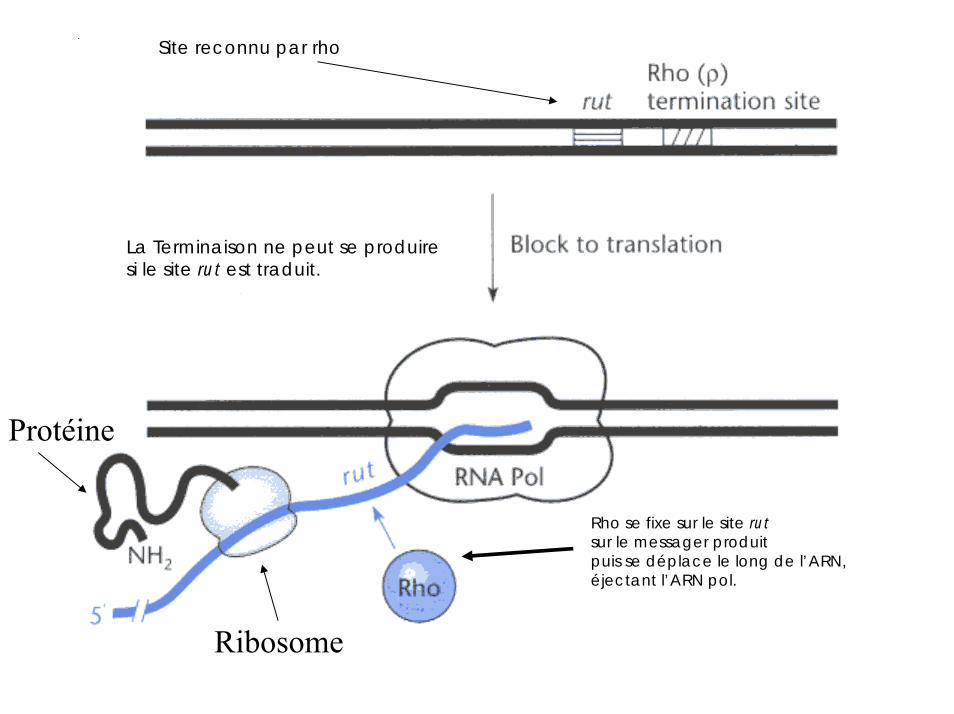
Site reconnu par rho
Ribosome
La Terminaison ne peut se produiresi le site rut est traduit.
Protéine
Rho se fixe sur le site rutsur le messager produitpuis se déplace le long de l’ARN, éjectant l’ARN pol.

Le déplacement de Rho requiertl’hydrolyse de l’ ATP
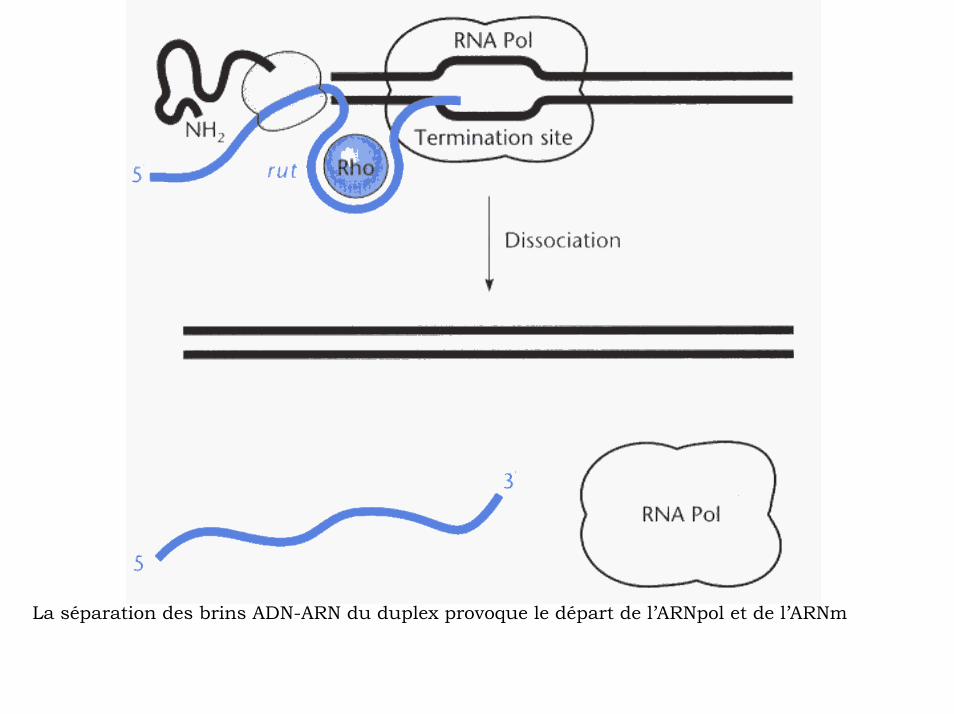
La séparation des brins ADN-ARN du duplex provoque le départ de l’ARNpol et de l’ARNm
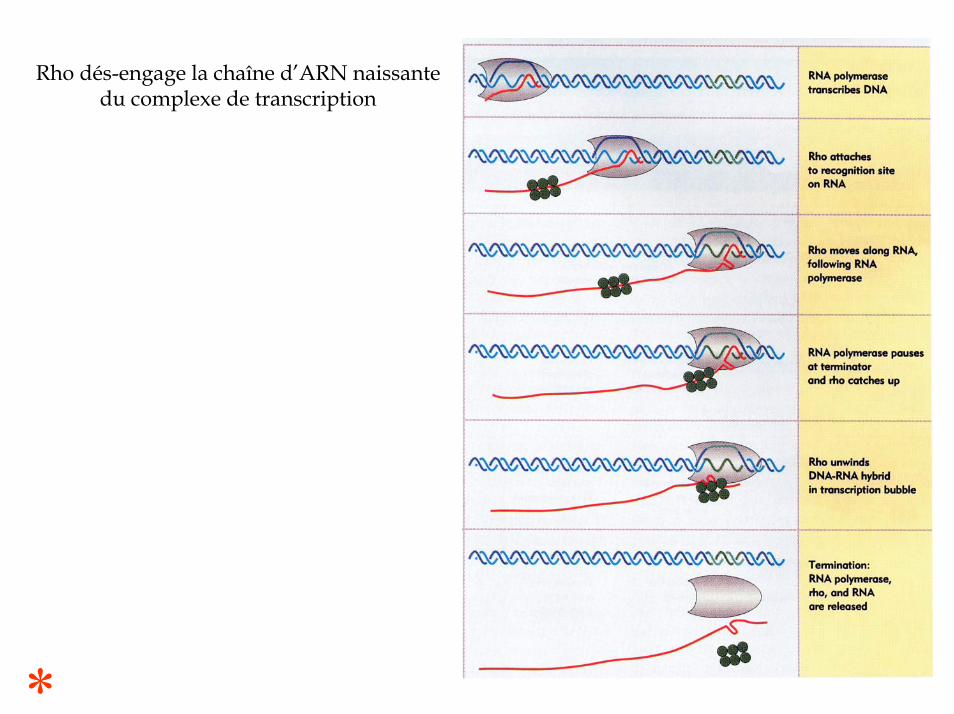
Rho dés-engage la chaîne d’ARN naissantedu complexe de transcription
*
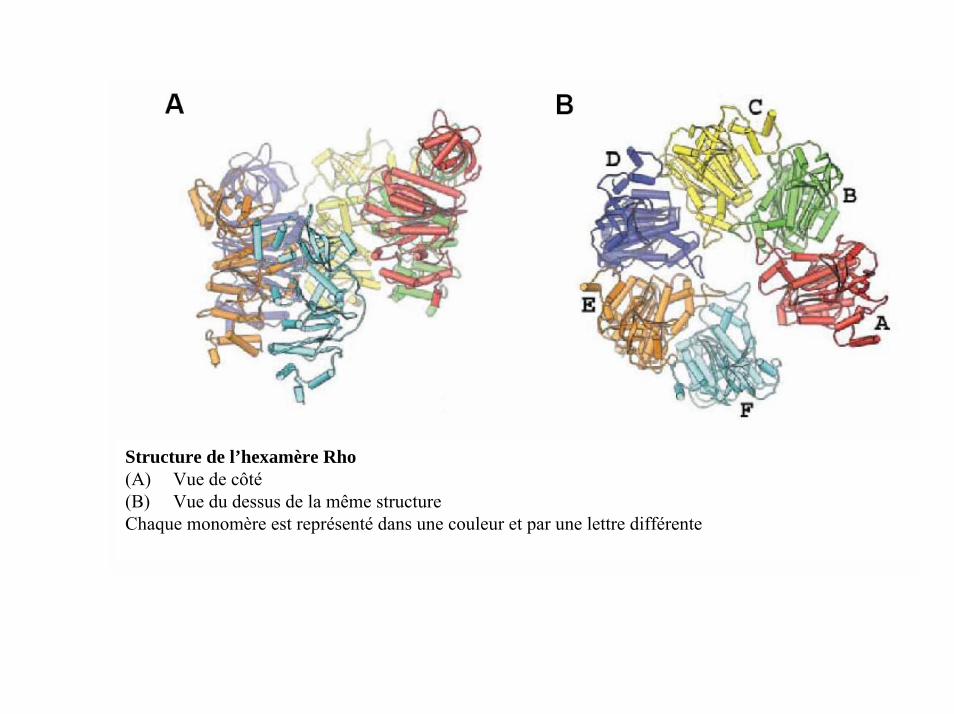
Structure de l’hexamère Rho(A) Vue de côté(B) Vue du dessus de la même structureChaque monomère est représenté dans une couleur et par une lettre différente

Modulation de la terminaison de la transcription
L’ensemble des mécanismes par lesquels la terminaison de la transcription est abolieou fortement diminuée est appelé antiterminaison.
Deux types de stratégies utilisées chez les bactéries pour obtenir l’antiterminaison :
1- effet sur l’ARN polymérase, la rendant « résistante » aux signaux de terminaison
ANTITERMINAISON GENERALE
Exemple : système d’antiterminaison par la protéine N du bactériophage λ
2- action localisée en masquant les signaux de terminaison eux-mêmes
ATTENUATION TRANSCRIPTIONNELLE
Mise en jeu d’interactions entre l’ARN polymérase et des effecteurs qui favorisent ou empêchent la formation des structures terminatrices.
*

ATTENUATION TRANSCRIPTIONNELLE
Nombreux exemples d’opérons régulés par atténuation transcriptionnelle
2 sous-classes d’atténuation transcriptionnelle selon la nature de l’effecteur mis en jeu dans le mécanisme :
1- un composant général de la machinerie cellulaire, le ribosome ou 2- une protéine régulatrice spécifique.

REGULATION IMPLIQUANT un couplage entre la TRANSCRIPTION ET LA TRADUCTION
Exemple 1 : les opérons de biosynthèse d’acides aminés chez les Entérobactéries
Exemple 2 : l’opéron pyrBI de biosynthèse des pyrimidines
REGULATION IMPLIQUANT une PROTEINE régulatrice spécifique
Exemple 1 : l’opéron trp de B. subtilis
Exemple 2 : l’opéron bgl d’E. coli et le gène sacB de B. subtilis

Opéron tryptophane (Trp) d’E. coliun exemple de la modulation de la terminaison de la
transcription par atténuation
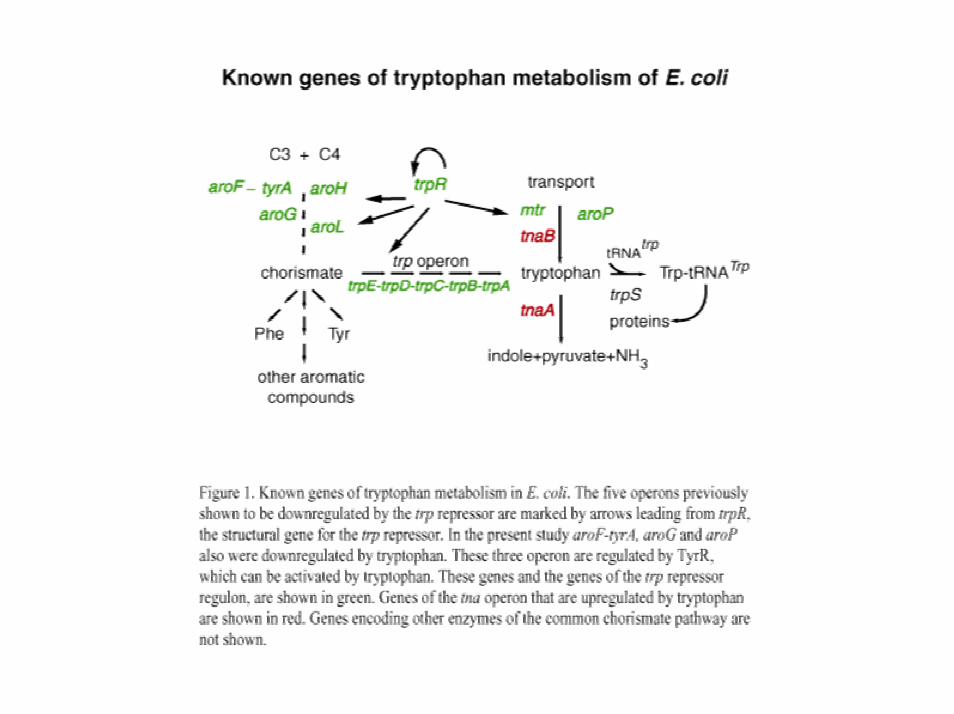

Contexte biologique
Le tryptophane est un acide aminé– produit à partir de l’acide chorismique– nécessaire à la synthèse des protéines– peu fréquent dans les protéines– qui a besoin d’une régulation fine
Régulation à différents niveaux– activation/répression de la transcription– atténuation de la transcription– inhibition du produit final (feedback négatif)
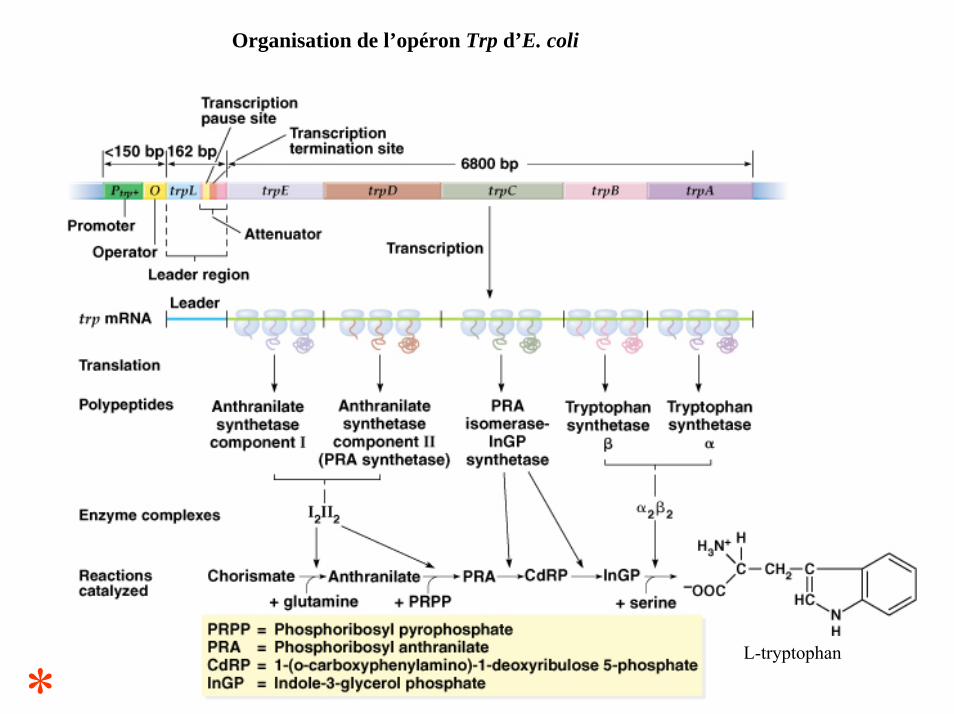
Organisation de l’opéron Trp d’E. coli
*L-tryptophan

Que sont les atténuateurs ?
Ils agissent afin de permettre à la transcription de se terminer de manière prématurée.
Leur cible se situe avant ou au sein de la région codante.
Quand ils sont non fonctionnels, la transcription se poursuit au-delà de la régiond’atténuation.
Quand ils sont fonctionnels, la transcription s’achève.

L’opéron Trp d’E. coli
• Si les acides aminés dont elle a besoin sont présents dans le milieu de culture, la bactérie E. coli “importera” ceux-ci avant d’en faire la synthèse.
Les gènes impliqués dans la biosynthèse des acides aminés sont réprimés : on parle d’opéron réprimé.
• Quand les acides aminés sont absents du milieu de culture, certains gènes sontexprimés et la synthèse des acides aminés se produit.

REGULATION de l’OPERON Trp (d ’E. coli)
Deux mécanismes mis en œuvre :
1- interaction répresseur- opérateur
2- terminaison de la transcription

Mécanisme n°1
Interaction répresseur – opérateur
Quand le tryptophane est présent, celui-ci se fixe au produit du gène trpRLa protéine TrpR se fixe à l’opérateur trp ce qui empêche la transcription de se faire.(la répression observée réduit le taux de transcription d’environ 70 fois)
La séquence des bases de l’opérateur trpla séquence presque palindromique est encadrée et sa région -10 est surlignée
CGAACTAGTTAACTAGTACGCAAGGCTTGATCAATTGATCATGCGTTC
-20 -10 +1
*

La fixation du tryptophane au répresseur trp altère sa structureUn déplacement de 0,8nm des hélices impliquées dans la reconnaissance permet au répresseurd’interagir avec l’ADN.
*

Un monomère comprend 107 acides aminés.Il est replié en 6 hélices α.Il forme un dimère stable en solution ;Les deux hélices reconnaissant l’ADN sont séparées de 3,4nm.
*

Mécanisme n°2
Terminaison de la transcription
La transcription est également contrôlée par atténuation, processus qui aboutit à la traductiond’un petit polypeptide
Quand les cellules sont privées de tryptophane, les gènes de l’opéron sont exprimés au taux le plus fort;Quand la privation en tryptophane est moins sévère, les gènes de l’opéron s’expriment à un niveau plus faibleque le maximum.
L’atténuation régule le niveau de transcription par un facteur de 8 à 10 et combiné avec le mécanisme 1(interaction répresseur-opérateur) il y a diminution de la transcription d’un facteur 560 à 700.
*
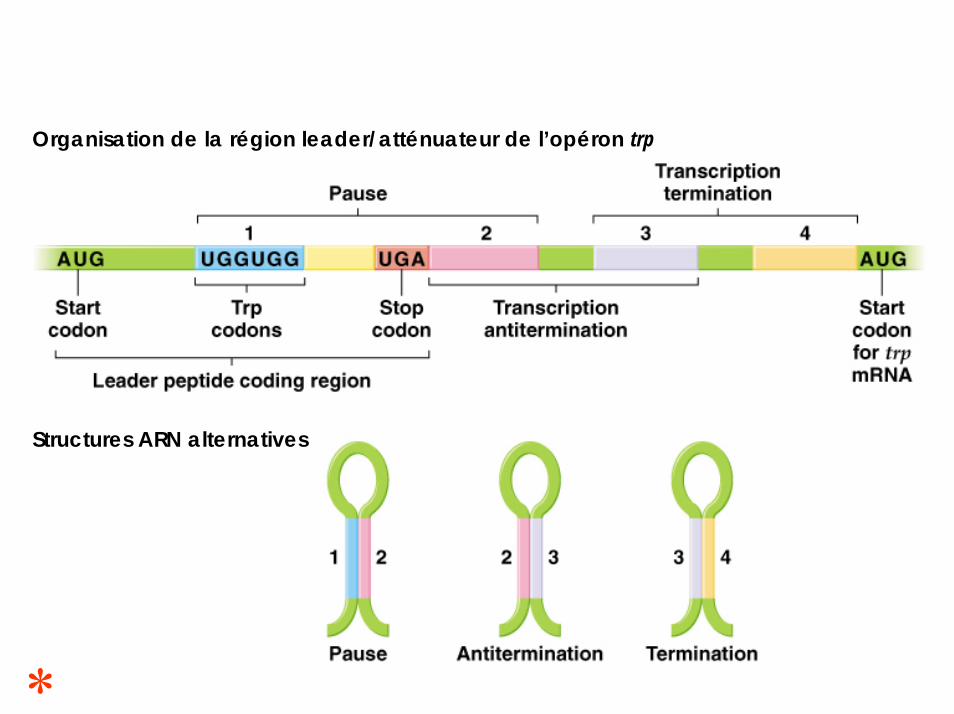
Organisation de la région leader/atténuateur de l’opéron trp
*
Structures ARN alternatives

Traduction de la partie “leader” de l’ARNm : synthèse du peptide “leader”
L’atténuateur est un terminateur Rho-indépendant de la transcription !

Modèle moléculaire de l’atténuation
La région leader (trpL) se situe entre l’opérateur et la séquence du locus trpE.Dans cette région leader se trouve la séquence att (= atténuateur).
Cette séquence att comprend un codon start, deux codons Trp et un codon stop, ainsi que 4 régions formées de séquence pouvant s’apparier selon 3 structures secondaires alternatives
S’il y a appariement des régions 1-2, le signal formé correspond à une PAUSE.S’il y a appariement des régions 2-3, le signal formé correspond à une ANTITERMINAISONS’il y a appariement des régions 3-4, le signal formé correspond à une TERMINAISON.
Transcription et traduction sont étroitement couplées chez les procaryotes; ces deux processus se produisent simultanément.
L’appariement des régions 1 avec 2 de l’ARNm produit provoque la pause de l’ARN polymérase juste après queces régions aient été synthétisées.Cette pause est suffisamment longue pour que le ribosome se fixe à l’ARNm et démarre la traduction du messagerjuste derrière l’ARN polymérase.
*
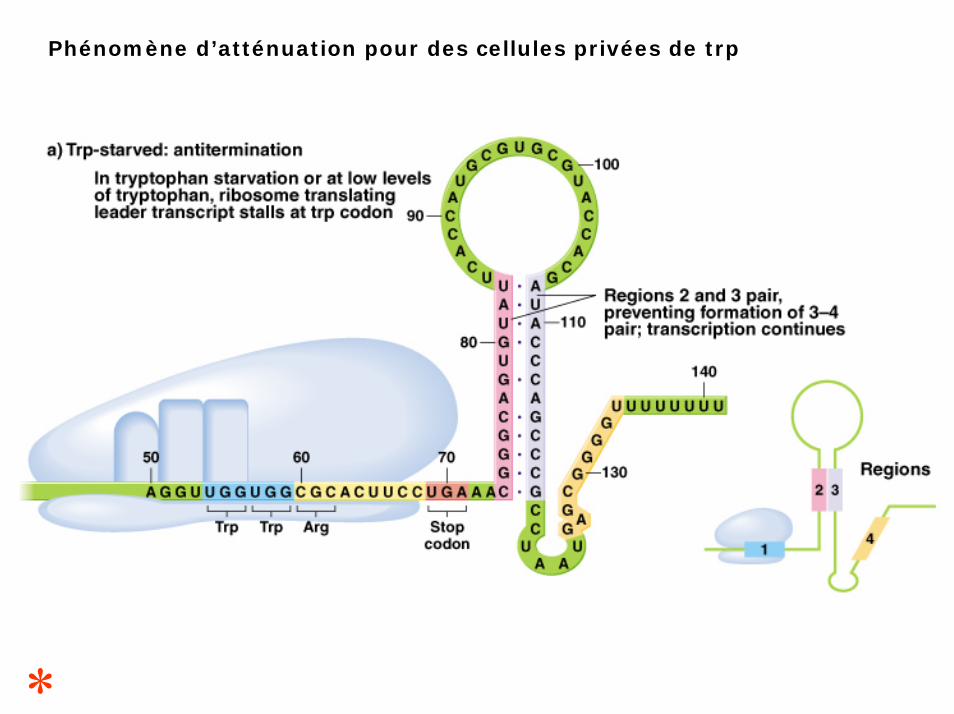
Phénomène d’atténuation pour des cellules privées de trp
*

Phénomène d’atténuation pour des cellules non-privées de trp
*

La position du ribosome joue un rôle important dans le phénomène d’atténuation
situation 1 :
le tryptophane est absent (ou en quantité insuffisante)1- les trp-ARNt sont indisponibles, le ribosome s’arrête aux codons trp ce qui couvre la région 1.2- la région 1 ne peut s’apparier avec la région 2, à la place la région 2 s’apparie à la région 3.3- la région 3 ne peut donc s’apparier à la région 4.4- l’ARN polymérase continue à transcrire l’ensemble de la séquence codantece qui permet la synthèse complète de l’ARNm.
situation 2 :
le tryptophane est abondant1- le ribosome ne s’arrête pas au niveau des codons trp; il continue à traduire la séquence leader, s’arrêtant au niveau de la région 22- la région 2 ne peut s’apparier avec la région 3; cette dernière s’apparie alors avec la région 4.3- cet appariement 3-4 constitue la séquence « atténuateur » et fonctionne comme signal determinaison4- la transcription s’achève avant que l’ARN polymérase atteigne les gènes permettant la synthèse du tryptophane
*

Le phénomène d’atténuation est un processus de régulation communà plusieurs opérons de biosynthèse des acides aminés

Le répresseur trp contrôle 3 opérons
Le répresseur est une protéine dimérique à structure hélice-tour-héliceLes sites opérateurs reconnus présentent une symétrie d’ordre 2.Les opérons contrôlés sont dans l’ordre décroissant de répression (du + vers le – réprimé)
trp > aroH > trpR
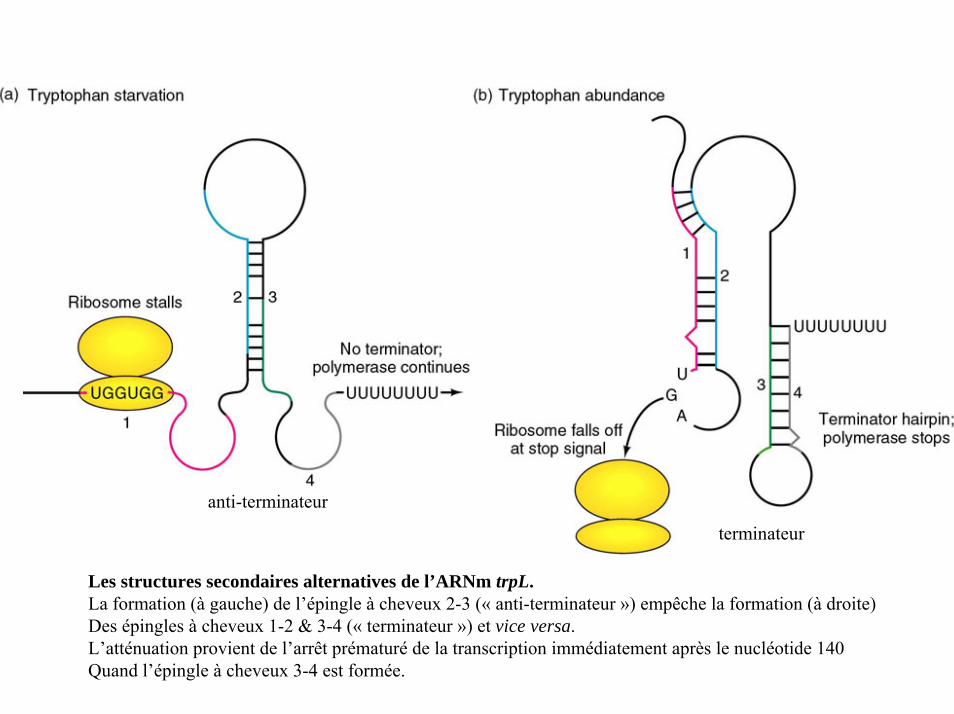
anti-terminateur
terminateur
Les structures secondaires alternatives de l’ARNm trpL.La formation (à gauche) de l’épingle à cheveux 2-3 (« anti-terminateur ») empêche la formation (à droite)Des épingles à cheveux 1-2 & 3-4 (« terminateur ») et vice versa.L’atténuation provient de l’arrêt prématuré de la transcription immédiatement après le nucléotide 140Quand l’épingle à cheveux 3-4 est formée.

Contrôle enzymatique du taux de tryptophane
production : l’anthranilate synthase est inhibée totalement par le tryptophaneinterconversion : la tryptophanase (tna) catalyse l’interconversion tryptophane-indole
Contrôle par le transport du taux de tryptophane
le produit d’aroP facilite le transport des acides aminés aromatiquesle produit de mtr facilite le transport du tryptophane (le gène mtr est
activé quand de la phénylalanine est présente et le tryptophane absent)le produit de tyrR module l’expression des gènes aroP et mtr.

Promoteurs de l’opéron tryptophane
Promoteur P1 (fort) : le répresseur actif se fixe à l’opérateur trp et couvre P1 ce qui exclut l’ARN pol.
Promoteur P2 (moins efficace) : situé à distance du locus trpD; gouverne la production des ARNm C, B & A.
La production des trpE & D est différenciée de celle de trpC, B & A
La dégradation des ARNm C, B & A est plus lente que celle des ARNm D & E.

Récapitulatif
• Répression par le métabolite terminal– TrpR : apo-répresseur– Tryptophane : co-répresseur
• Atténuation– Diminution de la transcription par arrêt prématuré– Action de la traduction sur la transcription
• « Feedback » négatif (action rapide)– Inhibition de l’enzyme 1 par le tryptophane
• Autorégulation négative– TrpR bloque sa propre transcription en absence de trp
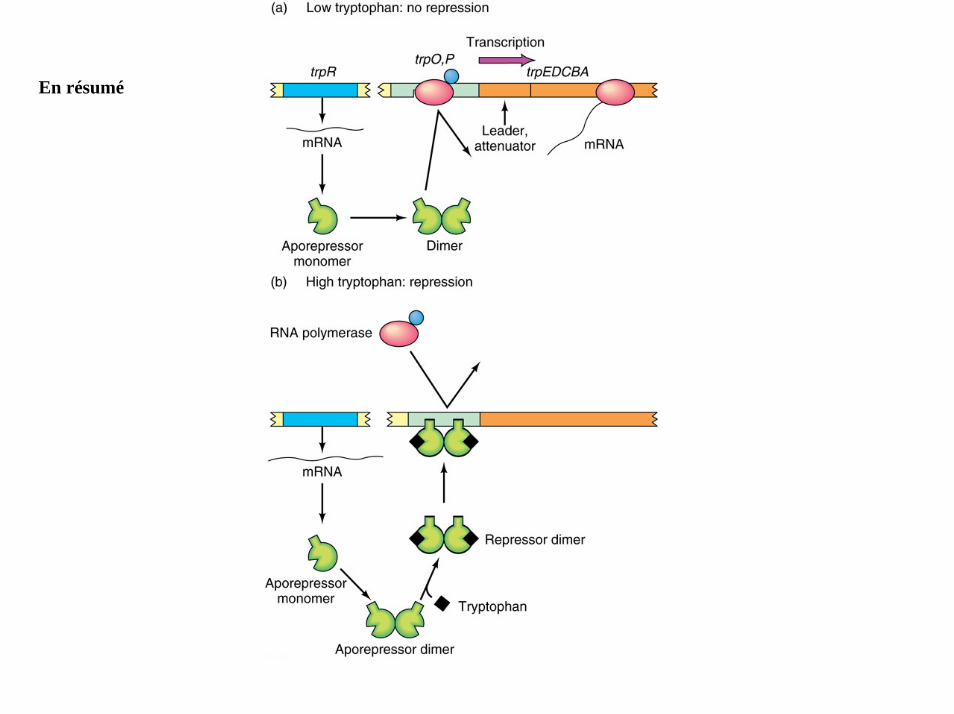
En résumé




![Quantum mechanics of nucleon interactions · 2017-08-31 · K1rl,rr19]. taqy coh¡tf'or¡¡ to thr relatl,vl,¡tlo two boity probtcr h¡ve bc¡a propoacd, by varlouE euthorl, but](https://img.pdfslide.fr/doc/110x75/5eb58328f2504b18ca4eb46d/quantum-mechanics-of-nucleon-interactions-2017-08-31-k1rlrr19-taqy-cohtfor.jpg)
![Formulation dynamique d’un mod ele de zone coh esive ...mentaire pour décrire l’état du contact. Cette variable, notée et initialement intro-duite dans [FRé 82], permet de](https://img.pdfslide.fr/doc/110x75/6089ff6ecd37e2599f25601a/formulation-dynamique-daun-mod-ele-de-zone-coh-esive-mentaire-pour-dcrire.jpg)























