Embed Size (px)
Citation preview

Croissance de la biomasse azotie du Phaeodactylum tricornutum (Bacillariophyceae) en cultures discontinues dialysante et non dialysante
PIERRE MARSOT Institut national de la recherche scientifique, 310, allde des Ursulines, Rimouski (Qudbec), Canada G5L 3AI
ALLAN D. CEMBELLA Conseil national de recherches du Canada, Institut des biosciences marines, 141 1, rue Oxford, Halifax
(~ouvelle-~cosse), Canada B3H 321
KHADIJA MOUHRI Laboratoire de biologie vkge'tale, Facult6 des sciences, Universite' Cadi Ayyad, Marrakech, Maroc
Requ le 13 novembre 1991
Revision reque le 1 1 mars 1992
Accepte le 23 mars 1992
MARSOT, P. , CEMBELLA, A. D., et MOUHRI, K. 1992. Croissance de la biomasse azotee du Phaeodactylum tricornutum (Bacillariophyceae) en cultures discontinues dialysante et non dialysante. Can. J. Microbiol. 38 : 945-952.
Les operations et les conditions particulikres de la technique de culture discontinue a dialyse determinent, chez la diatomee Phaeodactylum tricornutum, des comportements nutritionnels differents de ceux observes en culture non dialysante. Les mesures simultanees des concentrations de biomasse azotee et des nutriments azotes (NO, - + NO, -), du milieu de culture et du milieu nourricier, revklent que la plus grande partie de la biomasse en azote (>90%) d'une culture a dialyse est produite durant la croissance postexponentielle, quand les substrats nutritifs (NO,- + NO,-) sont en concentrations limitantes, inferieures a la constante de Michaelis-Menten. Prks de la moitie de cette biomasse est produite a la limite de diffusion de l'appareil, c'est-a-dire lorsque le transfert de masse des substrats, determinant l'activite d'absorption de la culture, atteint son maximum. Les cultures discontinues non dialysantes, dont la periode de croissance postexponentielle est courte, rkalisent leur biosynthkse en azote surtout durant la croissance active. Certains indices physiologiques touchant les activites metaboliques d'absorption et d'assimilation des nutriments azotes nous permettent de discerner des comportements nutritionnels distincts du P. tricornutum dans les deux types de culture et d'expliquer le niveau eleve de biomasse obtenu en culture a dialyse.
Mots clds : culture dialysante, diatomee, metabolisme de l'azote.
MARSOT, P., CEMBELLA, A. D., and MOUHRI, K. 1992. Croissance de la biomasse azotee du Phaeodactylum tricornutum (Bacillariophyceae) en cultures discontinues dialysante et non dialysante. Can. J. Microbiol. 38: 945-952.
The unique conditions associated with discontinuous (batch) dialysis culture of the diatom Phaeodactylum tricornutum elicit different nutritional responses from those observed in nondialysis culture. Simultaneous determinations of the nitrogen biomass concentration and of the nitrogen nutrients (NO, - + NO, -) in the culture chamber, as well as in the seawater nutrient medium at the entrance and exit of the dialyzer, revealed that nitrogen-biomass production in dialysis cultures is achieved mainly (>90%) during postexponential growth, when the concentration of nutrients is limiting (smaller than Michaelis-Menten constant). Almost half of this biomass is produced at the diffusion limit of the apparatus, i.e., when the mass transfer of nutrient substrates, which determines the total uptake activity of the culture, reaches a maximum. In contrast, in nondialysis discontinuous cultures, in which the postexponential growth phase is short, most of the total cellular nitrogen is accumulated during active growth. Certain physiological indices relating to the active uptake and assimilation of inorganic nitrogen are consistent with the different nutritional responses elicited by these two types of culture techniques and explain the high biomass levels obtained in dialysis culture.
Key words: dialysis culture, diatom, nitrogen metabolism.
Introduction L'emploi de cultures dialysantes en recherches ecophysio-
logiques nous a revel6 le fort potentiel productif des algues unicellulaires en milieu nature1 (Trainor 1965; Jensen et al. 1972). Les systemes dialysants utilises pour ces etudes s'appli- quent pour la plupart au mode d'operation continuous reser- voir - batch culture, tel que defini pour la culture de bacte- ries par Schultz et Gerhardt (1969) a cause des reserves illimitees du milieu nutritif (continuous reservoir = eau de mer) et de la croissance discontinue des algues (batch culture). Les cultures discontinues dialysantes se distinguent des cultures discontinues non dialysantes par une periode de croissance prolongee leur permettant d'atteindre une plus forte densite cellulaire meme avec un milieu pauvre en nutri- ments (Jensen et Rystad 1973; Skoglund et Jensen 1976). Printed in Canada / lmprime au Canada
L'extension de la croissance est d'autant plus importante que la capacite de diffusion de l'appareil envers les nutri- ments (PmAm/ V) (voir Liste des abrkviations et symboles) est elevee par rapport au taux de croissance maximum de l'algue (Marsot et al. 1981a, 1981b).
La concentration maximale de biomasse (X,,) d'une culture discontinue en general (dialysante ou non dialysante) est fonction de la quantite de substrat transforme, c'est-a- dire de la masse de nutriments disponibles et de la capacite d'assimilation et d'incorporation des nutriments destines a la croissance de l'algue. Pour une culture non dialysante (ND), X,,, est donc proportionnel a la concentration des nutriments presents dans le milieu de culture (C,,,) au depart (Spencer 1954) et au taux de conversion des nutri- ments en biomasse (Yx), d'oh X,, = CncYx. Pour ce qui
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
HIC
AG
O o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

CAN. J. MICROBIOL. VOL. 38, 1992
DIALYSEUR
FIG. 1. Schema du systkme de culture D discontinue illustrant l'apport des nutriments inorganiques (NO, - + NO, -) par diffu- sion (N,) et la production de la biomasse en azote (VXN). Adapt6 de Marsot et a[. (1991b). Cne, concentration en NO, - + NO, - de l'eau de rner a l'entree du dialyseur; ens, concentration en NO, - + NO, - de l'eau de rner ii la sortie du dialyseur; Cnc, con- centration en NO3 - + NO,- du milieu de culture; Vr, volume du reservoir nourricier (eau de rner); V, volume de la culture; Q,, debit d'eau de rner dans le dialyseur; XN, concentration de la biomasse en azote; N,, diffusion ou taux d'absorption des NO, - + NO2-.
concerne la culture dialysante (D), l 'apport des nutriments (exprime en grammes par jour) est continu, par consequent, X,, depend d e la capacite d e diffusion d e l'appareil
v et d'un facteur physiologique exprimant, selon une dimen- sion d e temps (par jour), l'inverse de l'activite metabolique d e survivance d e l'algue (1/ YE),
PmAm(Cne - Cnc) X m a x =
VYE
(Schultz et Gerhardt 1969). Une culture discontinue D peut ainsi obtenir, a u cours
d e son cycle d e croissance, une plus grande quanti te d e substrats nutritifs qu 'une culture N D et, par consequent, produire plus d e biomasse, en supposant que la valeur d e YE demeure faible. C'est gknkralement par ce concept d e bilan d e materiel que l'on s'explique theoriquement la forte biomasse obtenue en culture D (Sakshaug et Jensen 1978; Ney et al. 1981)' mais cette conversion substrat-biomasse des algues n'a pas vraiment ete evaluee.
L'explication d u phenomkne doit egalement se trouver dans le comportement physiologique des algues associe aux processus de diffusion des metabolites cellulaires et a la crois- sance en milieu deficient en nutriments (Marsot et al. 1991 b). E n effet, il est fort probable que meme avec des quantites non limitantes d e nutriments, le X,, d 'une culture N D demeurera inferieur a celui d e la culture D a cause de l'accu- mulation d e metabolites auto-inhibiteurs (Pratt 1944). L'eli- mination des metabolites par la dialyse d'un milieu ameliore la condition physiologique d e l'algue (Prat t 1944)' mais son
effet sur le metabolisme nutritionnel reste a prkciser. Chez les batteries, o n reconnait que l'klimination des produits metaboliques (exemples : acetate, lactate) augmente le coef- ficient d e conversion d u glucose, YG (Landwall et Holme 1977). Nous presentons, dans ce travail, une etude d e la croissance et de l'activite d e conversion des nutriments azotes par la diatomee Phaeodactylum tricornutum en cultures D et ND. E n comparant ces deux modes de croissance, nous cherchons a identifier les conditionnements d e la diatomee qui contribuent, en culture D, a sa croissance en forte den- site cellulaire.
Materiel et mCthodes L'algue utilisee pour ces experiences est la diatomee Phaeodacty-
lum tricornutum. La souche provient de la collection du Woods Hole Oceanographic Institute, Woods Hole, Mass. Le montage et le mode d'operation des cultures discontinues dialysantes ont deja ete decrits dans Marsot et a[. (1991a). Le systkme est sche- matise a la figure 1. I1 est compose de deux circuits separes l'un de l'autre par une ou plusieurs membranes dialysantes (ligne poin- tillee) : (i) le circuit nourricier, par lequel l'eau de rner arrive a une concentration Cne en NO3- + NO2-, et en ressort avec la con- centration ens; le debit de l'eau de rner (Q,) y est ajuste a 134'4 L d - '; (ii) le circuit culture circulant en sens inverse au debit de 233 mL . min - ', griice a un bullage constant d'air (sans CO,) au debit de 300 mL .min - '; celui-ci contient une densite de biomasse cellulaire X et une concentration Cnc en NO,- + NO,-. Le volume total de culture (v) est de 628 mL. La chambre de crois- sance, de forme cylindrique, posskde un faible diamktre (3'2 cm) de f a ~ o n a rkduire le coefficient d'extinction lumineuse des cul- tures. Le systkme emploie un dialyseur a fibres creuses non bio- degradable~ (modkle B2-200, Toray Industries, Inc., Japon) dote d'une aire de dialyse (A,) de 15 600 cm2. La constante de per- meabilitk (P,A,) de ces membranes envers les NO, - + NO, - est evaluee a 56 160 cm3 - d - '. La P,A, est obtenue au cours de la croissance du P. tricornutum en culture D, en appliquant l'equa- tion derivee de la formule sur la diffusion moleculaire (Marsot et a[. 1991a):
La salinite de l'eau de rner variait entre 24 et 28 %O et le nitrate (NO, -) constituait la principale source d'azote pour la culture ( > 98%).
Nos analyses ont porte sur deux cultures ND (depourvues de dialyseur) : l'une a ete realisee sans aeration, l'autre a beneficie d'une aeration (air sans CO,) d'une intensite comparable a celle de la culture D (300 mL . min - '). Les cultures ND ont utilise le milieu nutritif enrichi f/2 de Guillard et Ryther (1962). Toutes les cultures ont Cte realisees a 13,5 + 1°C dans des conditions axe- niques. Les cultures ont ete exposees a des intensites d'eclairement saturantes. La culture D, qui a servi a l'etude des variations de biomasse et des nutriments azotes, recevait un eclairement de 85 pE . m s - ' . Les cultures ND et D utilisees pour l'analyse des acides amines intra- et extra-cellulaires etaient exposees a un Cclai- rement de 70 pE . m s - '. Les autres conditions de croissance sont decrites aux figures et au tableau correspondants. Le denom- brement cellulaire est effectue au microscope, sur une lame a puits de type Palmer-Maloney. Le carbone et l'azote cellulaires sont mesures sur l'analyseur CHN (Perkin-Elmer 2400) aprks une lyophilisation de 48 h des echantillons de culture. Les cellules ont ete recueillies sur filtre en fibres de verre Gelman (type A/E) pre- alablement chauffe a 400°C, 24 h. L'analyse des nutriments NO,- + NO,- de l'eau de rner (C,, et ens) et du milieu de culture (en,), est effectuee par une methode colorimetrique (Strickland et Parsons 1972)' a l'aide d'un appareil Technicon Autoanalyseur 11. Le taux de croissance, exprimant le nombre de
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
HIC
AG
O o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.
MARSOT ET AL.
EXPONENT l ELLE 0 1'"
0°C G - c -
2 +Jog- o i (I) - . w b w -
g I- 0.4 - g a n: +
m a 0-
0 n
x - 5.4 3 1 - n
w- Z x lo7 2; - 4.5 ~ i % LC_,
K A 5:: a j i a . -3.6 N i y o e 2 E 0 5 a A .
A
E E - ~ , ~ E E ! 5 3 W ; ~ ; I o ~ . ~ 2 (I) (I) : a -1.sy ( I ) - r Z g lo4 g m - 0.9 i
- 0 - lo3
TEMPS (jours)
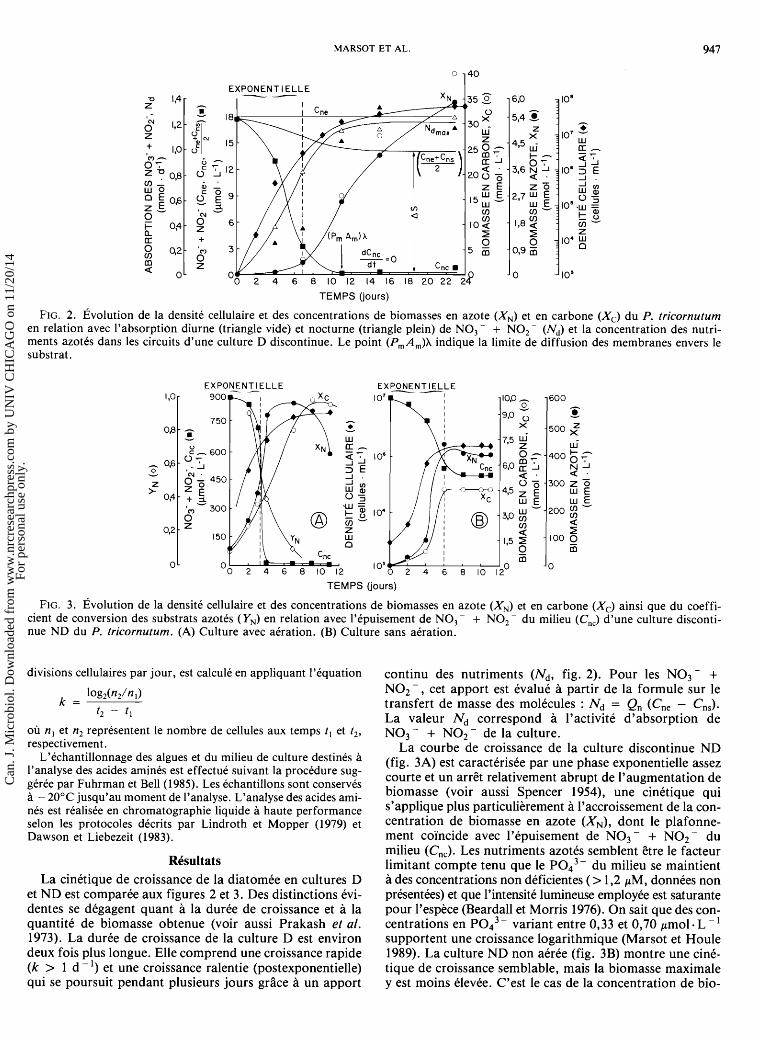
FIG. 2. ~volu t ion de la densite cellulaire et des concentrations de biomasses en azote (XN) et en carbone (X,) du P. tricornutum en relation avec l'absorption diurne (triangle vide) et nocturne (triangle plein) de NO, - + NO,- (N,) et la concentration des nutri- ments azotes dans les circuits d'une culture D discontinue. Le point (PmAm)X indique la limite de diffusion des membranes envers le substrat.
EXPONZNTJELLE - 900
750 - n
8 w
2 - 600 -0- - i $ < 450
E - + A
lo 300
150
'0 2 4 6 8 10 12
TEMPS (jours)
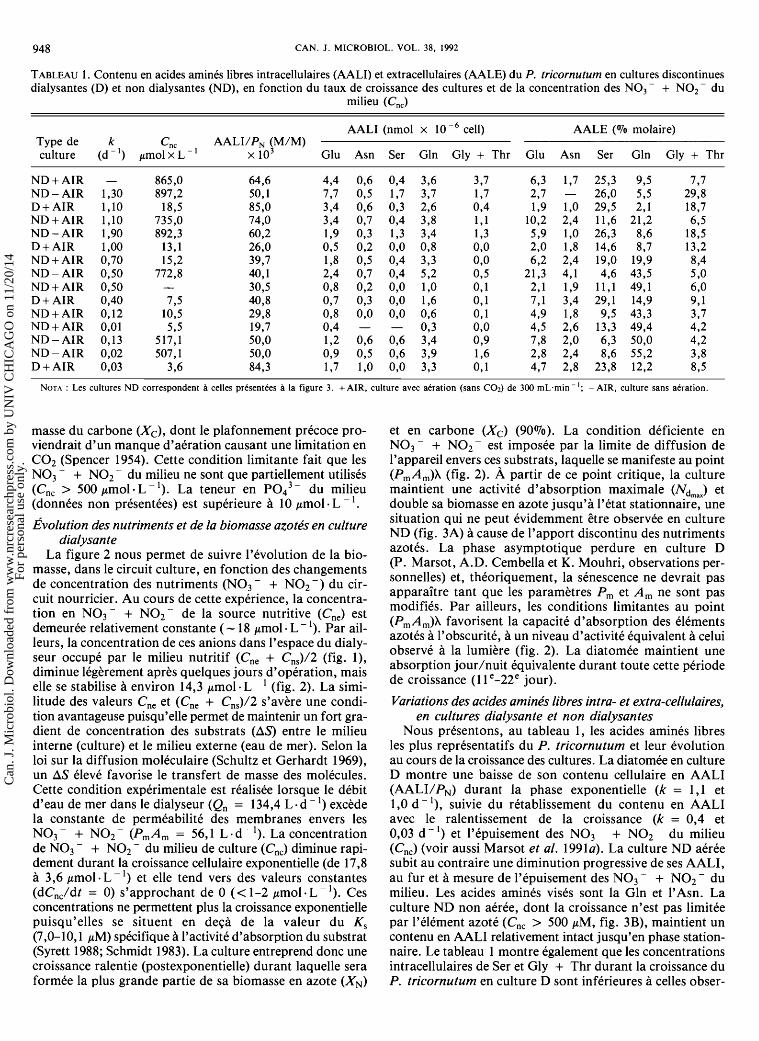
FIG. 3. ~volu t ion de la densite cellulaire et des concentrations de biomasses en azote (XN) et en carbone (X,) ainsi que du coeffi- cient de conversion des substrats azotes (Y,) en relation avec l'epuisement de NO, - + NO, - du milieu (C,,,) d'une culture disconti- nue ND du P. tricornutum. (A) Culture avec aeration. (B) Culture sans aeration.
divisions cellulaires par jour, est calcule en appliquant l'equation
ou n, et n, representent le nombre de cellules aux temps t, et t,, respectivement .
L'echantillonnage des algues et du milieu de culture destines a l'analyse des acides amines est effectue suivant la procedure sug- geree par Fuhrman et Bell (1985). Les echantillons sont conserves a - 20°C jusqu'au moment de l'analyse. L'analyse des acides ami- nes est realisCe en chromatographie liquide a haute performance selon les protocoles decrits par Lindroth et Mopper (1979) et Dawson et Liebezeit (1983).
La cinetique de croissance de la diatomee en cultures D et ND est comparee aux figures 2 et 3. Des distinctions evi- dentes se degagent quant a la duree de croissance et a la quantite de biomasse obtenue (voir aussi Prakash et al. 1973). La duree de croissance de la culture D est environ deux fois plus longue. Elle comprend une croissance rapide (k > 1 d - l) et une croissance ralentie (postexponentielle) qui se poursuit pendant plusieurs jours grace a un apport
continu des nutriments (Nd, fig. 2). Pour les NO3 - + NO2-, cet apport est evalue a partir de la formule sur le transfert de masse des molecules : Nd = Q, (C,, - C,,). La valeur Nd correspond a l'activite d'absorption de NO3 - + NO2 - de la culture.
La courbe de croissance de la culture discontinue ND (fig. 3A) est caracterisee par une phase exponentielle assez courte et un arret relativement abrupt de l'augmentation de biomasse (voir aussi Spencer 1954), une cinetique qui s'applique plus particulierement a l'accroissement de la con- centration de biomasse en azote (XN), dont le plafonne- ment coi'ncide avec l'epuisement de NO3 - + NO2- du milieu (C,,). Les nutriments azotes semblent &re le facteur limitant compte tenu que le PO,'- du milieu se maintient a des concentrations non deficientes (> 1,2 pM, donnees non presentees) et que l'intensite lumineuse employee est saturante pour l'espece (Beardall et Morris 1976). On sait que des con- centrations en ~ 0 , ~ - variant entre 0,33 et 0'70 pmol . L - ' supportent une croissance logarithmique (Marsot et Houle 1989). La culture ND non aeree (fig. 3B) montre une cine- tique de croissance semblable, mais la biomasse maximale y est moins elevee. C'est le cas de la concentration de bio-
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
HIC
AG
O o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.
948 CAN. J. MICROBIOL. VOL. 38, 1992
TABLEAU 1 . Contenu en acides amines libres intracellulaires (AALI) et extracellulaires (AALE) du P. tricornutum en cultures discontinues dialysantes (D) et non dialysantes (ND), en fonction du taux de croissance des cultures et de la concentration des NO, - + NO, - du
milieu (Cnc)
AALI (nmol x 10 -' cell) AALE (% molaire) Type de k Cnc AALI/PN (M/M) culture (d - l) pmol x L - x lo3 Glu Asn Ser Gln Gly + Thr Glu Asn Ser Gln Gly + Thr
ND + AIR ND - AIR D + AIR ND +AIR ND -AIR D + AIR ND + AIR ND - AIR ND + AIR D + AIR ND + AIR ND + AIR ND - AIR ND -AIR D + AIR
NOTA : Les cultures ND correspondent a celles presentees a la figure 3. +AIR, culture avec aeration (sans C02) de 300 mL.min- ' ; -AIR, culture sans ahation.
masse du carbone (Xc), dont le plafonnement prkcoce pro- et en carbone (Xc) (90%). La condition dkficiente en viendrait d'un manque d'akration causant une limitation en NO3 - + NO2 - est imposke par la limite de diffusion de C 0 2 (Spencer 1954). Cette condition limitante fait que les l'appareil envers ces substrats, laquelle se manifeste au point NO3 - + NO2- du milieu ne sont que partiellement utilisks (PmAm)X (fig. 2). A partir de ce point critique, la culture (Cnc > 500 pmol L - I) . La teneur en ~0~~ - du milieu maintient une activitk d'absorption maximale (Ndm,) et (donnkes non presentees) est supkrieure a 10 pmol . L - '. double sa biomasse en azote jusqu'a l'ktat stationnaire, une
~volution des nutriments et de la biomasse azotks en culture dialysan te
La figure 2 nous permet de suivre l'kvolution de la bio- masse, dans le circuit culture, en fonction des changements de concentration des nutriments (NO3 - + NO2-) du cir- cuit nourricier. Au cours de cette expkrience, la concentra- tion en NO3- + NO2- de la source nutritive (Cne) est demeurke relativement constante ( - 18 pmol L - I). Par ail- leurs, la concentration de ces anions dans l'espace du dialy- seur occupk par le milieu nutritif (Cne + Cns)/2 (fig. I), diminue lkgkrement aprks quelques jours d'opkration, mais elle se stabilise a environ 14,3 pmol L - ' (fig. 2). La simi- litude des valeurs Cne et (Cne + Cns)/2 s'avkre une condi- tion avantageuse puisqu'elle permet de maintenir un fort gra- dient de concentration des substrats (AS) entre le milieu interne (culture) et le milieu externe (eau de mer). Selon la loi sur la diffusion molkculaire (Schultz et Gerhardt 1969), un AS klevk favorise le transfert de masse des molkcules. Cette condition expkrimentale est rkaliske lorsque le dkbit d'eau de mer dans le dialyseur (Q, = 134,4 L. d - ') excede la constante de permkabilitk des membranes envers les NO3 - + NO2 - (P,A, = 56,l L d - '1. La concentration de NO3 - + NO2 - du milieu de culture (Cnc) diminue rapi- dement durant la croissance cellulaire exponentielle (de 17,8 a 3,6 p m o 1 - L ' ) et elle tend vers des valeurs constantes (dCnc/dt = 0) s'approchant de 0 (< 1-2 pmol L - I). Ces concentrations ne permettent plus la croissance exponentielle puisqu'elles se situent en d e ~ a de la valeur du K, (7,O-10,l pM) spkcifique a l'activite d'absorption du substrat (Syrett 1988; Schmidt 1983). La culture entreprend donc une croissance ralentie (postexponentielle) durant laquelle sera formke la plus grande partie de sa biomasse en azote (XN)
situation qui ne peut kvidemment Etre observke en culture ND (fig. 3A) a cause de l'apport discontinu des nutriments azotks. La phase asymptotique perdure en culture D (P. Marsot, A.D. Cembella et K. Mouhri, observations per- sonnelles) et, thkoriquement, la sknescence ne devrait pas apparaitre tant que les paramktres Pm et A, ne sont pas modifies. Par ailleurs, les conditions limitantes au point (P,A,)X favorisent la capacitk d'absorption des klkments azotks a l'obscuritk, a un niveau d'activitk kquivalent a celui observe a la lumikre (fig. 2). La diatomke maintient une absorption jour/nuit kquivalente durant toute cette pkriode de croissance (1 1 e-22e jour).
Variations des acides aminks libres intra- et extra-cellulaires, en cultures dialysante et non dialysan tes
Nous prksentons, au tableau 1, les acides aminks libres les plus reprksentatifs du P. tricornutum et leur kvolution au cours de la croissance des cultures. La diatomke en culture D montre une baisse de son contenu cellulaire en AALI (AALI/PN) durant la phase exponentielle (k = 1,l et 1,O d - '), suivie du rktablissement du contenu en AALI avec le ralentissement de la croissance (k = 0,4 et 0,03 d - I) et l'epuisement des NO3 - + N O 2 du milieu (Cnc) (voir aussi Marsot et al. 1991a). La culture ND aeree subit au contraire une diminution progressive de ses AALI, au fur et a mesure de l'kpuisement des NO3 - + NO2 - du milieu. Les acides aminks visks sont la Gln et 1'Asn. La culture ND non aeree, dont la croissance n'est pas limitee par l'element azotk (Cnc > 500 pM, fig. 3B), maintient un contenu en AALI relativement intact jusqu'en phase station- naire. Le tableau 1 montre egalement que les concentrations intracellulaires de Ser et Gly + Thr durant la croissance du P. tricornutum en culture D sont infkrieures a celles obser-
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
HIC
AG
O o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.
MARSOT ET AL.
EXPONENT I E L L E
TEMPS (jours)
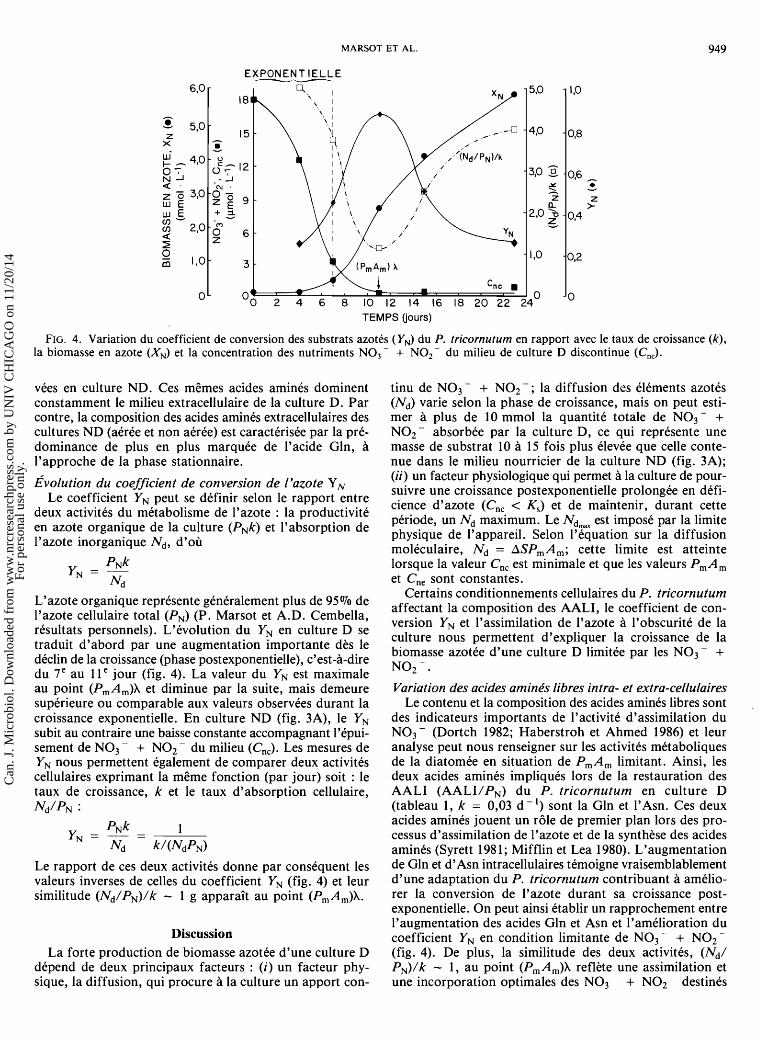
FIG. 4.. Variation du coefficient de conversion des substrats azotes (YN) du P. tricornutum en rapport avec le taux de croissance (k), la biomasse en azote (XN) et la concentration des nutriments NO3- + NO,- du milieu de culture D discontinue (C,,,).
vees en culture ND. Ces memes acides amines dominent tinu de NO3- + NO2-; la diffusion des elements azotes constamment le milieu extracellulaire de la culture D. Par (Nd) varie selon la phase de croissance, mais on peut esti- contre, la composition des acides arnines extracellulaires des me; a plus de 10 mmol la quantite totale de NO3 - + cultures ND (aeree et non aeree) est caracterisee par la pre- NO2 - absorbee par la culture D, ce qui represente une dominance de plus en plus marquee de l'acide Gln, a masse de substrat 10 a 15 fois plus elevee que celle conte- l'approche de la phase stationnaire. nue dans le milieu nourricier de la culture ND (fig. 3A);
~volu t ion du coefficient de conversion de l'azote Y, (ii) un facteur physiologique qui permet a la culture de pour-
Le coefficient YN peut se definir selon le rapport entre suivre une croissance postexponentielle prolongee en defi- cience d'azote (Cnc < K,) et de maintenir, durant cette deux activites du metabolisme de l'azote : la productivite periode, un Nd maimurn. Le Ndm, eSt par la limite en azote organique de la ( P ~ k ) et l 'absor~tion de physique de l'appareil. Selon l'equation sur la diffusion l'azote inorganique Nd, d'ou moleculaire, Nd = ASPmAm; cette limite est atteinte
L'azote organique represente generalement plus de 95% de l'azote cellulaire total (PN) (P. Marsot et A.D. Cembella, resultats personnels). L'evolution du YN en culture D se traduit d'abord par une augmentation importante dks le declin de la croissance (phase postexponentielle), c'est-a-dire du 7e au 1 l e jour (fig. 4). La valeur du YN est maximale au point (PmAm)X et diminue par la suite, mais demeure superieure ou comparable aux valeurs observees durant la croissance exponentielle. En culture ND (fig. 3A), le YN subit au contraire une baisse constante accompagnant l'epui- sement de NO3 - + NO2 - du milieu (Cnc). Les mesures de YN nous permettent egalement de comparer deux activites cellulaires exprimant la meme fonction (par jour) soit : le taux de croissance, k et le taux d'absorption cellulaire, Nd/PN :
Le rapport de ces deux activites donne par consequent les valeurs inverses de celles du coefficient YN (fig. 4) et leur similitude (Nd/PN)/k - 1 g apparait au point (PmAm)X.
lorsque la valeur Cnc est minimale et que les valeurs PmAm et Cn, sont constantes.
~ G t a i n s conditionnements cellulaires du P. tricornutum affectant la composition des AALI, le coefficient de con- version YN et l'assimilation de l'azote a l'obscurite de la culture nous permettent d'expliquer la croissance de la biomasse azotee d'une culture D limitee par les NO3- + NO2 - . Variation des acides aminks libres intra- et extra-cellulaires
Le contenu et la composition des acides amines libres sont des indicateurs importants de l'activite d'assimilation du NO3 - (Dortch 1982; Haberstroh et Ahmed 1986) et leur analyse peut nous renseigner sur les activites metaboliques de la diatomee en situation de PmAm limitant. Ainsi, les deux acides amines impliques lors de la restauration des AALI (AALI/PN) du P . tricornutum en culture D (tableau 1, k = 0'03 d - ') sont la Gln et 1'Asn. Ces deux acides amines jouent un r81e de premier plan lors des pro- cessus d'assimilation de l'azote et de la synthese des acides amines (Syrett 198 1 ; Mifflin et Lea 1980). L'augmentation de Gln et d'Asn intracellulaires temoigne vraisemblablement d'une adaptation du P. tricornutum contribuant a amelio- rer la conversion de l'azote durant sa croissance post- exponentielle. On peut ainsi etablir un rapprochement entre 19augmentation des acides Gln et Asn e t I'amelioration du
Discussion coefficient YN en condition limitante de NO3 - + NO2 - La forte production de biomasse azotee d'une culture D (fig. 4). De plus, la similitude des deux activites, (Nd/
depend de deux principaux facteurs : (i) un facteur phy- PN)/k - 1, au point (PmAm)X reflkte une assimilation et sique, la diffusion, qui procure a la culture un apport con- une incorporation optimales des NO3 - + NO2 - destines
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
HIC
AG
O o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

950 CAN. J. MICROBIOL. VOL. 38, 1992
a la croissance de la diatomke. Un kcart entre ces deux une forte biomasse azotee ( - 1,5 mmol L - ') apres une valeurs, Nd/PN)/k > 1, indique au contraire que l'activitk d'absorption sert a d'autres fonctions cellulaires que celle de la croissance (voir aussi Laws et Wong 1978). La rkponse de la culture ND a une dkficience en NO3- + NO2- (culture akrke) se traduit par une baisse constante des acides intracellulaires Gln et Asn. Lorsque les NO3 - + NO2 - ne constituent pas le facteur limitant (culture ND non akrke), le contenu de ces AALI demeure inchangk. Ces resultats sont conformes a d'autres travaux selon lesquels une dkficience ou une privation en NO3 - ou NH4 + provoque un appau- vrissement en acides amines cellulaires Gln et Asn (Admiraal et al. 1986; Al-Amoudi et Flynn 1989).
Les faibles concentrations intracellulaires de Ser et Gly + Thr observkes en culture D (tableau 1) sont probablement attribuables a une meilleure klimination de ces metabolites. Les acides Ser, Gly et Thr representent en effet une forte proportion des acides aminks extracellulaires de la culture D (tableau I). Les acides Ser et Gly sont des produits de la photorespiration cellulaire associes au metabolisme du glycolate (Whittingham et Pritchard 1963), et une baisse de leur production et de leur accumulation cellulaires temoigne d'une photosynthese active (Halldal et Holmen 1979). L'on peut prksumer que la dialyse d'un milieu de culture contre un volume infini d'eau de mer modifie l'ktat de (( quasi- kquilibre >> des concentrations intra- et extra-cellulaires (Ignatiades et Fogg 1973)' notamment celles de molkcules non retenues par l'algue comme les acides Ser et Gly. Ce dkskquilibre de concentrations previendrait l'accumula- tion cellulaire de ces mktabolites et aurait un effet bknkfique sur l'activite genkrale de l'algue (Keys 1980).
Absorption de No3- + NO2- a l'obscurite' L'absorption nocturne de NO3- + NO2- par le
P. tricornutum semble un phknomene particulier a la culture D en milieu naturel (fig. 2). L'on sait qu'en culture ND (avec milieu enrichi) l'absorption a l'obscurite de NO3- + NO2 - reprksente moins de 10% de l'activitk diurne de cette diatomke (Raimbault et Mingazzini 1987). Nous pouvons dkduire que l'absorption nocturne contribue a l'klaboration de la biomasse en azote de la culture D, compte tenu du fait que le nitrate constitue la principale source d'azote (> 98%) et que le taux d'accumulation cellulaire de NO3 - + NO2 - est faible (< 17% de l'absorption nocturne; Marsot et al. 1992). I1 s'agit par conskquent d'une assimilation de l'azote qui implique, pour le moins, les deux premieres etapes de la reduction du nitrate.
La diatomee P. tricornutum est une espece estuarienne qui abonde dans la rkgion c6tiere oh ont kt6 effectuks nos prelevements d'eau de mer (48'32 ' de latitude nord, 68'32 ' de longitude ouest; voir Sinclair 1978). Les conditions eutro- phiques de son habitat naturel soulignent la capacite d'adap- tation de cette diatomee aux conditions oligotrophes du milieu de culture D (fig. 2, jours 11 a 22). La mkthode de culture D se przte donc a l'ktude de l'adaptabilitk trophique, puisqu'elle permet de maintenir une croissance continue (Marsot et al. 1991 b) ou discontinue des algues en dkficience de nutriments. Mzme si le P. tricornutum est reconnu comme une algue facile a cultiver en laboratoire, il est probable que nos observations puissent s'appliquer a d'autres especes estuariennes. La diatomke Skeletonema costatum, par exemple, montre une cinktique de croissance similaire en culture D (Marsot et al. 1983)' en produisant
phase de croissance prolongke. L'inference de nosr~sul tats a des especes ockaniques kvoluant en eaux oligotrophes et euhalines est plus difficile, quoique l'on puisse ktablir un lien entre l'assimilation a l'obscuritk du 14c02 dans ces eaux (Prakash et al. 1991) et l'assimilation nocturne des NO3 - + NO2 - en culture D durant la croissance post- exponentielle. Dans les deux cas, la croissance de l'algue est limitke par la diffusion de la ressource au niveau de son microenvironnement cellulaire.
Conclusion Le caractere limitatif des parametres de diffusion (P, et
A,) d'une culture D discontinue, dktermine une cinktique de croissance particuliere chez la diatomke P. tricornutum, diffkrente de celle observee en culture non dialysante. En culture D, la synthese de la biomasse azotee se rkalise plu- t6t durant une periode de croissance postexponentielle pro- longee, quand les nutriments azotes (NO, - + NO2-) du milieu de culture (C,,) sont dkficients (Cnc < K,). La moitie de cette biomasse est klaborke alors que l'appareil atteint sa limite de diffusion envers les molkcules concer- nkes. En culture ND, au contraire, la biomasse (XN) est surtout produite en phase exponentielle, lorsque Cnc est superieur au K,, et le rapide dkclin de la croissance (absence de pkriode de croissance prolongke) tkmoigne de l'apport discontinu des nutriments. Certains conditionnements cel- lulaires du P. tricornutum en culture D, durant la phase postexponentielle, nous permettent d'expliquer la croissance prolongke et le haut niveau de biomasse en azote obtenu. I1 s'agit (i) d'une augmentation du contenu des AALI res- ponsables de l'assimilation de l'azote (Gln et Asn); (ii) de l'klimination plus complete des AALI issus de la respira- tion cellulaire (Ser et Gly); (iii) de la similitude des activitks d'absorption cellulaire des NO3 - + NO2 - et du taux de croissance, et de l'amelioration du coefficient de conversion de l'azote (YN); (iv) de la capacitk d'assimilation des NO3 - + NO2 - par la culture a l'obscuritk, en situation de P, A, limitant.
Remerciements Nous dksirons remercier Madame Johanne Belzile pour
son assistance technique et Monsieur Juan Carlos Colombo pour l'analyse des acides amines. Cette recherche a kt6 rea- lisee griice aux appuis financiers du ministere des Approvi- sionnements et Services et de Pzches et Ockans Canada, de 1'Association des professeurs pour des programmes univer- sitaires internationaux et de 1'Agence canadienne de dkve- loppement international du Canada. Ce manuscrit est enre- gistrk au Conseil national de recherches du Canada sous le no 33016.
Admiraal, W., Peletier, H. , et Laane, R.W.P.M. 1986. Nitrogen metabolism of marine planktonic diatoms: excretion, assimila- tion and cellular pools of free amino acids in seven species with different cell size. J . Exp. Mar. Biol. Ecol. 98 : 241-263.
Al-Amoudi, O.A., et Flynn, K.J. 1989. Effect of nitrate - N incor- poration on the composition of the intracellular amino acid pool of N-deprived Tetraselmis marina. Br. Phycol. J . 24 : 53-61.
Beardall, J . , et Morris, I . 1976. The concept of light intensity adaptation in marine phytoplankton: some experiments with Phaeodactylum tricornutum. Mar. Biol. (Berl.), 39 : 377-387.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
HIC
AG
O o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

MARSOT ET AL. 95 1
Dawson, R., et Liebezeit, G. 1983. Determination of amino acids apd carbohydrates. Dans Methods of seawater analysis. 2e Cd. Editeurs : K. Grasshoff, M. Ehrhardt et K. Kremling. Verlag Chemie GmbH, Weinheim. pp. 319-340.
Dortch, Q. 1982. Effect of growth conditions on accumulation of internal nitrate, ammonium, amino acids and protein in three marine diatoms. J. Exp. Mar. Biol. Ecol. 61 : 243-264.
Fuhrman, J.A., et Bell, T.M. 1985. Biological considerations in the measurement of dissolved free amino acids in seawater and implications for chemical and microbiological studies. Mar. Ecol. Prog. Ser. 25 : 13-21.
Guillard, R.R.L., et Ryther, J.H. 1962. Studies of marine plank- tonic diatoms. I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. Can. J . Microbiol. 8 : 229-239.
Haberstroh, P.R., et Ahmed, S.I. 1986. Resolution by high pres- sure liquid chromatography of intracellular and extracellular free amino acids of a nitrogen deficient marine diatom, Skeletonema costatum (Grev.) Cleve, pulsed with nitrate or ammonium. J . Exp. Mar. Biol. Ecol. 101 : 101-117.
Halldal, P., et Holmen, A.T. 1979. The interrelationship between photosynthetic electron transport, glycolate excretion and amino acid metabolism in the blue-green alga Anacystis nidulans. Plant Cell Physiol. 20 : 757-763.
Ignatiades, L., et Fogg, G.E. 1973. Studies on the factors affect- ing the release of organic matter by Skeletonema costatum (Greville) Cleve in culture. J. Mar. Biol. Assoc. U.K. 53 : 937-956.
Jensen, A., et Rystad, B. 1973. Semi-continuous monitoring of the capacity of sea water for supporting growth of phytoplank- ton. J. Exp. Mar. Biol. Ecol. 11 : 275-285.
Jensen, A., Rystad, B., et Skoglund, L. 1972. The use of dialysis culture in phytoplankton studies. J. Exp. Mar. Biol. Ecol. 8 : 24 1 -248.
Keys, A.J. 1980. Synthesis and interconversion of glycine and serine. Dans The biochemistry of plants. Vol. 5. Amino acids and derivatives. Editeur : B.J. Mifflin. Academic Press, New York. pp. 359-374.
Landwall, P., et Holme, T. 1977. Removal of inhibitors of bacterial growth by dialysis culture. J . Gen. Microbiol. 103 : 345-352.
Laws, E.A., et Wong, D.C.L. 1978. Studies of carbon and nitrogen metabolism by three marine phytoplankton species in nitrate- limited continuous culture. J . Phycol. 14 : 406-416.
Lindroth, P., et Mopper, K. 1979. High performance liquid chromatographic determination of subpicomole amounts of amino acids by precolumn fluorescence derivatization with a-phthaldialdehyde. Anal. Chem. 51 : 1667-1674.
Marsot, P., et Houle, L. 1989. Excretion cellulaire et morphoge- nese de Phaeodactylum tricornutum (Bacillariophyceae). Bot. Mar. 32 : 355-367.
Marsot, P., Fournier, R., et Blais, C. 1981a. Culture a dialyse : emploi de fibres creuses dialysantes pour la culture massive de phytoplancton. Can. J . Fish. Aquat. Sci. 38 : 905-911.
Marsot, P., Fournier, R., et Blais, C. 198 1 b . Un nouveau procede de culture continue a dialyse pour le phytoplancton. Biotechnol. Lett. 3 : 689-694.
Marsot, P. , Leclerc, M., et Fournier, R. 1983. Aspect morpholo- gique et composition chimique de Skeletonema costatum (Bacillariophyceae.) croissant en milieu nutritif nature1 a l'aide d'un systeme de culture a fibres dialysantes. Can. J. Microbiol. 29 : 1235-1240.
Marsot, P., Cembella, A.D., et Colombo, J.C. 1991a. Intracellular and extracellular amino acid pools of the marine diatom Phaeodactylum tricornutum (Bacillariophyceae) grown on unenriched seawater in high-cell density dialysis culture. J . Phycol. 27 : 478-491.
Marsot, P., Cembella, A.D., et Houle, L. 1991b. Growth kinetics and nitrogen-nutrition of the marine diatom Phaeodactylum tricornutum in continuous dialysis culture. J . Appl. Phycol. 3 : 1-10.
Marsot, P., Mouhri, K., et Cembella, A.D. 1992. Assimilation du nitrate en cycles diurnaux par Phaeodactylum tricornutum. Plant Physiol. Biochem. 30. Sous presse.
Mifflin, B.J., et Lea, P.J. 1980. Ammonia assimilation. Dans The biochemistry of plants. Vol. 5. Amino acids and derivatives. ~ d i t e u r s : B. J . Mifflin. Academic Press, New York. pp. 160-202.
Ney, J.M., Conary, C.L., et Chapman, S.R. 1981. High density diatom production utilizing dialysis techniques. Aquaculture, 24 : 363-369.
Prakash, A., Skoglund, L., Rystad, B., et Jensen, A. 1973. Growth and cell-size distribution of marine planktonic algae in batch and dialysis cultures. J. Fish. Res. Board Can. 30 : 143-1 55.
Prakash, A., Sheldon, R.W., et Sutcliffe, W.H., Jr. 1991. Geographic variation of oceanic 14c dark uptake. Limnol. Oceanogr. 36 : 30-39.
Pratt, R. 1944. Studies on Chlorella vulgaris. IX. Influence on growth of Chlorella of continuous removal of chlorellin from the culture solution. Am. J. Bot. 31 : 418-421.
Raimbault, P., et Mingazzini, M. 1987. Diurnal variations of intra- cellular nitrate storage by marine diatoms: effects of nutritional state. J . Exp. Mar. Biol. Ecol. 112 : 217-232.
Sakshaug, E., et Jensen, A. 1978. The use of cage culture in studies of the biochemistry and ecology of marine phytoplankton. Oceanogr. Mar. Biol. Annu. Rev. 16 : 81 -106.
Schmidt, G. 1983. Note on the effect of high nitrate concentra- tion and light intensity on the growth and uptake rates of Phaeodactylum tricornutum (Bohlin) in culture. Bol. Inst. Oceanogr. 32 : 91 -98.
Schultz, J.S., et Gerhardt, P. 1969. Dialysis culture of micro- organisms: design, theory and results. Bacteriol. Rev. 33 : 1-47.
Sinclair, M. 1978. Summer phytoplankton variability in the lower St. Lawrence Estuary. J. Fish. Res. Board Can. 35 : 1 171-1 185.
Skoglund, L., et Jensen, A. 1976. Studies on N-limited growth of diatoms in dialysis culture. J. Exp. Mar. Biol. Ecol. 21 : 169-1 78.
Spencer, C.P. 1954. Studies on the culture of marine diatom. J. Mar. Biol. Assoc. U.K. 33 : 265-290.
Strickland, J.D.H., et Parsons, T.R. 1972. A practical handbook of seawater analysis. 2e ed. Bull. Fish. Res. Board Can. Bull. no 167.
Syrett, P.J. 1981. Nitrogen metabolism of mjcroalgae. Dans Physiological bases of phytoplankton ecology. Editeur : T. Platt. Can. Bull. Fish. Aquat. Sci. 210 : 182-210.
Syrett, P. J . 1988. Uptake and utilization of nitrogen compounds. Dans Biochemistry of the algae and cyanobacteria. Proceedings of the Phytochemical Society of Europe. Editeurs : L. J . Rogers et J.F. Gallon. Oxford University Press, Oxford. pp. 23-39.
Trainor, F.R. 1965. A study of unialgal cultures of Scenedesmus incubated in nature and in the laboratory. Can. J . Bot. 43 : 701 -706.
Whittingham, C.P., et Pritchard, G.G. 1963. The production of glycolate during photosynthesis in Chlorella. Proc. R. Soc. London B, 157 : 366-380.
Liste des abriviations et symboles AALE Acides amines libres extracellulaires. AALI Acides amines libres intracellulaires. A, Aire de la membrane de dialyse (cm2). C,, Concentration du substrat nutritif du milieu de culture
(mol . L - I). C,, Concentration du substrat nutritif du milieu nourricier,
a l'entree du dialyseur (mol . L - I). C,, Concentration du substrat nutritif du milieu nourricier,
a la sortie du dialyseur (mol L -I). D Dialysante. k Taux de croissance en divisions par jour (d - I). Ks Constante de demi-saturation (Michaelis-Menten) pour
l'absorption des NO, - + NO2 - (mol . L - I).
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
HIC
AG
O o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

CAN. J . MICROBIOL. VOL.
Non dialysante. V Transfert de masse des nutriments azotes (NO,- + Vr NO,-), equivalent du taux d'absorption des NO,- + X NO, - par la culture (mol d - I). x c Nombre de cellules au temps t. xmax Coefficient de permeabilite de la membrane envers un substrat (cm . d - I). XN Azote particulaire, c.-a-d. azote organique et azote YE inorganique cellulaires (mol). YG Debit du milieu nourricier (eau de mer) dans le dialyseur YN (L. d -I). Difference de concentration du substrat entre le milieu Y, de culture et le milieu externe (eau de mer). Temps.
Volume de culture (L). Volume du reservoir nourricier (eau de mer) (L). Concentration de la biomasse cellulaire (g . L - '). Concentration de la biomasse en carbone (mol . L -I). Concentration maximale de la biomasse cellulaire ( g . ~ - ' ) . Concentration de la biomasse en azote (mol L - I). Activite metabolique de survivance de l'algue (d -I). Coefficient de conversion du glucose. Coefficient de conversion des substrats azotes (NO, - + NO, - ) en biomasse azotee. Coefficient de conversion des substrats en biomasse cellulaire.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV C
HIC
AG
O o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.





























