Embed Size (px)
Citation preview

BtOCHtMIE, 1976, 58, 377-379.
Distribution des acides gras dans les phospholipides de Micrococcus lysodeikticus.
M i c h ~ l e WELBY, M a r i e - A n t o i n e t t e LANEELLE et G i l b e r t LANEELLE ~ .
Centre de Recherche de Biochimie et G6ndtiqae Cellulaires du C.N.R.S., 118, route de Narbonne, 31077 Toulouse Cedex, France.
f14-11-1975).
INTRODUCTION.
La c o m p o s i t i o n en ac ides g ras des m e m b r a n e s bae td- rierrnes va r ie au eours de Ia cro.issance des cu l tu res , p a r exemple [1] les phosp ,hol ip ides de Eseherichia colt p r 6 s e n t e n t en t r e eux des d,iff6rences de c o m p o s i t i o n en ac ides g ras en p h a s e exponentiel l .e , d i f ferences qui d is- p a r a i s s e n t en phase s t a t i onna i r e .
Pen d ' 6 t u d e s on t por t6 sur 1,es bac t6 r i e s dons les- que l l e s les ae ides g ras r ami f i e s sont p r 6 p o n d 6 r a n t s , K'aned.a [2, 3] a mon t r~ que, dans Bacil lus subl i t is , les ac ides iso C15 se t r o u v e n t e s s e n t i e l l e m e n t en pos i - tion. 2, s~r le res te g l y c E r o p h o s p h a t e des p h o s p h o l i p i d e s , et 1.es ac ides iso et an te - i so C 17 sont p r i n c i p a l e m e n t en p o s i t i o n 1. C o n t r a i r e m e n t ~ ce qui a 5t6 observ~ p o u r E. coil, il n.'a pas t ronv6 de d.iffErences signifi- ea t ives d a n s la r 6 p a r t i t i o n des ac ides gras en t re les d ive r s types, de phospho],~pides.
N o u s avons 6tud.i~ 1.a r6pa r t i t i on des ac ides gras darts les p h o s p h o l i p i d e s &'une soue.he de Micrococcus lyso- deikt icus.
MATI~RIEL ET MI~THOD.ES.
Culture bactdrienne : La souc,he de Micrococeus lyso- deik t icus ut i l i s6e p o r t e le n u m 6 r o ATCC 4698-4, Les ce l lu les son t cu l t iv6es sous ag i t a t i on dans un mi l i eu pep ton6 [4] ; ,e.lles, sont r~coltdes au mi l i eu de la p h a s e e x p o n e n t i e l l e de cro issance .
I so l emen t des phosphol ip ides : Les ce l lu les en t i6res sont d61i~pid$es p a r q u a t r e e x t r a c t i o n s sucees.sives pa r le m~lange ch Io ro fo rme : -m6thano l 2/1 (v/v) et les phos - p h o l i p i d e s son.t pr~cipi t6s dans l'acdton-e b o u i l l a n t ; d o n s la f r a c t i o n .acdtono-so tuble il n ' y a pas de phos - pholipid.e d~celable p a r ch rom.a tograph ie en couche mince. Les p h o s p h o l i p i d e s sont pur i f i6s p a r c h r o m a t o - g raph ie su r co lonne d'acid.e s i l ie ique (Mal l inckrod t CCA) pr~alablerrren~ lav6 au m~thano l . La pu r i f i c a t i on est achevde p a r ,ch, romatogra :ph ie sur couche mince prEpa- r a t ive de 1 m m d ' 6 p a i s s e u r (Kiselgel PF254 - - M~erck) ; i ' d luan t est ,le me lange c h l o r o f o r m e - m d t h a n o l - e a u (6,5/ 2~5/4 v / v / v ) , Les: ch romatog l , aph i e s a n a l y t i q u e s sont effectu6es sur e o u c h e mince d,e Kieselgel H (Merck) a.vec le, m6m,e so lvant . Les phosp ,hol ip ides sont rEvEl6s p a r le r6actif , de D i t t m e r [5] ; la r h o d a m i n e B h 0,01 p. cent dans le p h o s p h a t e monosod, ique 0,25 M p e r m e t de r6v~]er t o u s l e s , l ip ides .
Hydrolyse enzyma t ique des phosphol ip ides : Nous avon:s, e m p l o y d u n e p r 6 p a r a t i o n c o m m e r c i a l e (Koch Light) de p h o s p h o l i p a s e A v e n a n t du ven in de Crotalus
<> A qui t ou t e c o r r e s p o n d a n c e doi t ~tre adress6e .
adamanteus . Nous avons ut i l is~ ies cond i t i ons de r6ac- t ion de O,kuyam.a et N ~ j i m a [6], en s u i v a n t F a v a n c e - m e n t de la r6ac t ion p a r c h r o m a t o g r a p h i c en t o u c h e mince. Les p r o d u i t s de r6ac t ion sont s6par6s pa r chro- m a t o g r a p h i c sur co lonne d 'ac ide s i l ic ique et c h r o m a t o - g r a p h i c s sur couche mince. L es ac ides g ras l ib6r6s de la p o s i t i o n 2 du g lycero l son t est~rifi~s p a r le d iazo- m 6 t h a n e ; les ly so - p h o s p h o l i p i d e s f o r m 6 s s:ont s~po- nifi~s ct leu.rs ac ides gras ex t r a i t s et est~rifi6s p o u r d ~ t e r m i n e r la c o m p o s i t i o n en ac ides gras en p o s i t i o n 1 su r 1.e glyc6rol ,
P a r ai11~eurs, c h a q u e phosph,ol ip ide est saponifi~ et sa c o m p o s i t i o n en ac ides g ras t o t a u x (pos i t ion 1 A- 2) est d6 te rmin6e .
Analyse des acides gras : Les es te rs m 6 t h y l i q u e s des ac ides gras on t 6t6 ident if i6s pa r c h r o m a t o g r a p h i c en p h a s e gazeuse sur co Ionne cap i l l a i r e SCOT de 15 m charg6e de s i l icone OV 17, su r co lonne de 2 m ~ gar - n i ssage ~ 1'0 p. cen t de DEG,S, et sur eo lonne de 1 m
ga rn i s sage ~ 5 p. cent de s i l icone SE 30. Les a n a l y s e s q u a n t i t a t i v e s .de r o u t i n e on t en su i t e 6t~ effectu6es sur la eo lonne de sill.cone SE 30.
Preparat ion de la membrane : Los eellul 'es sont sus- p e n d u e s clans un. t a m p o n Tr is HC1 0,06 M h pH 7,6 : on a j o u t e .environ 0,2 mg de ] y s o z y m e p a r ml de sus- pens ion et on la i sse ag i r env i ron 30 minu te s . Les m e m - b r a n e s sont i sol~es p a r c e n t r i f u g a t i o n f r a e t i o n n 6 e ; la p r 6 p a r a t i o n est f i na l emen t cen t r i fug6e deux lo is 20 m.inute~ h 15 000 g.
Es t ima t ion des proport ions des phosphol ip ides : La cu l t u r e est effeetu6e en p resence de 1 mCi de 32P-or tho- p h o s p h a t e de s o d i u m (CEA-France) d a n s 50. ml de mi l ieu . Au t o u r s d.e Ia c ro issance , on f a i t une d iza ine de p r61bvements du mi l i eu de cul~ture de 0,8 ml chacun , que l 'on a j o u t e /~ 3 ml du m61ange m 6 t h a n o l - e h l o r o - f o r m e 2¢i (v/v) ; apr6s 1 heure de contac t , on f o r m e deux p h a s e s l i q u i d e s pa r a d d i t i o n de 1 ml de ch loro- f o r m e et 1 ml d ' e au ; la phase a q u e u s e est 61imin6e et la ph.ase c h l o r o f o r m i q u e est lavEe deux l o i s pu i s Eva- por6e sous c o u r a n t d ' azo te ,
R ~ S U L T A T S .
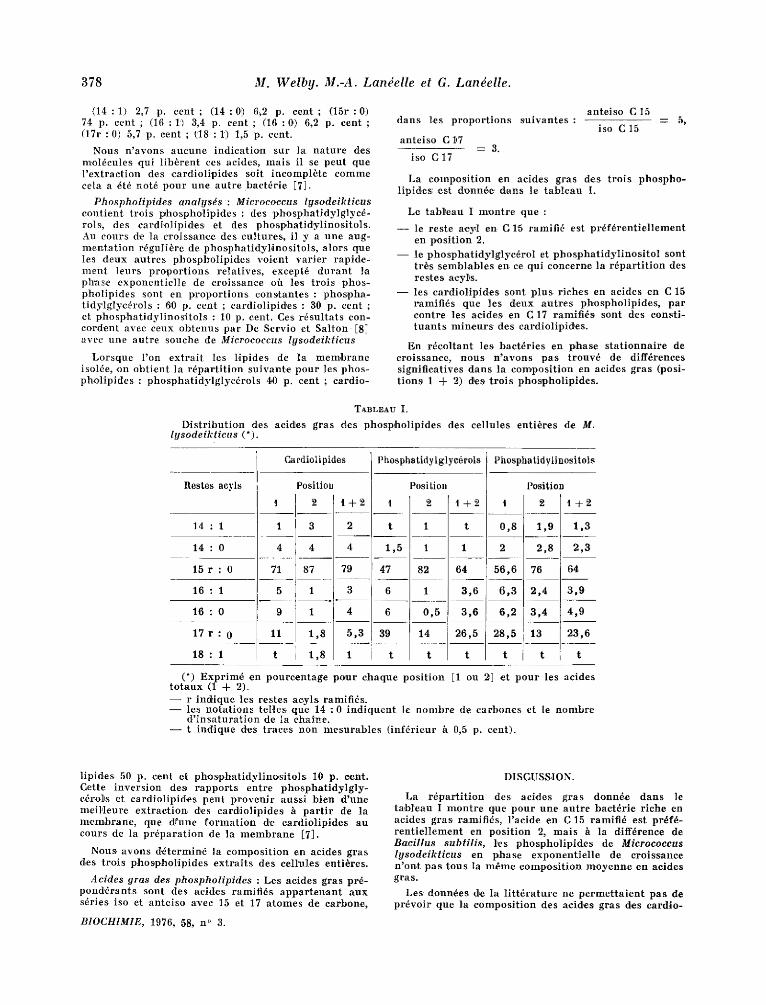
Micrococcus Igsodeikt icus f o u r n i t 7 p. cent de son p,oid, s sec en e x t r a i t Hpid ique . Les p h o s p h o l i p i d e s b r u t s r ep r~sen t en t 6 p. cen t du po ids sec de la baetErie , Les r~s idus d~lipid6s d.onnent pa r sapon i f i ca t ion env i ron 1 p. cent de l eu r po ids en ac ides gras ; l e u r c o m p o s i t i o n est proch,e de celle des r e s t e s aey l s des ca rd io l ip ides donn6s dans le t a b l e a u I :
3 7 8 M. WeIby . M.-A. Landel le et G. Landel le .
( 1 4 : 1 ) 2,7 p. cent ; (14:0,) 6,2 p. cent ; (15r :0) 74 p. cent ; ( 1 6 : 1 ~) 3,4 p. cen, t ; ( 1 6 : 0 ) 6,2 p. cent ; (17r :0) 5,7 p. cent ; t 1 8 : 1 ) 1,5 p. cent.
Nous n ' avons a u c u n e indicatio.n sur ]a n a t u r e d.es m oI6cules qui l ib6ren t ces acides, ma t s il se peut que l ' ex t r ac t ion des cardiol ipid.es soil incomplb te comme ceta a 6t4 not.6 pour une a u t r e bactdr ie [71.
Phospholipides analys~s : Micrococcus lysodeikticus con t i en t t ro is phospho l ip ides : des p h o s p h a t i d y l g l y c d - rols, des cardiolipid,es, et des p h o s p h a t i d y l i n o s i t o l s . Au cours de ]a croissan,ce des cu l tu res , il y a une aug- m en.tation r6guli+re de phosphat idyl~inosi to ls , a lors que les d,eux a,utres phospbo l ip ides vo ien t va r i e r rap ide- m e n t l eurs p r o p o r t i o n s rel 'atives, exceptd d u r a n t ],a p.h'ase exponen t i e l l e de cro issance off les t ro i s phos- phol ip ides sont en p r o p o r t i o n s cons t an t e s : phospha - t idy lg lyc~ro l s : 6r0 p. cent ; cardiolipid, es : 30 p. cent ; et p ,hospha t idy l inos i to l s : 10 p. cent. Ces r6su l t a t s con- cord.ent avec ceux ob tenus par De Servio et S a l t o n [8] avec une au t re souche de Micrococcus lysodeikticus
Lorsque l 'on ex t r a i t les 1.ipides. de l a m e m b r a n e isolde, Oh ob t i en t la r d p a r t i t i o n su ivan te pou r les phos- pho l ip ides : p h o s p h a t i d y l g l y c 6 r o l s J~0 p, c e n t ; card io-
d a n s les p r o p o r t i o n s s .u ivan te s :
an te i so C 117
iso C 17 ----3.
an te i so C 15
iso C 15 = 5,
La compos i t i on en acides gras d.es t ro i s phospho - lipid, es .es¢ d'onn6e d, ans le t a b l e a u I.
Le tabl~eau I m,ontre que :
- - le res te a.cyl en. C 15 ramif i6 est p r~ f6 ren t i e l l emen t en pos i t ion 2.
- - le p h o s p b a t i d y l g l y c d r o l et p h o s p h a t i d y l i n o s i t o l sont tr+s semblab les en. ce qui concerne ]a r 6 p a r t i t i o n des res tes acyl~s.
- - les ca rd io l ip ides sont p l u s r iches en acides en C 15 ramif ids que les deux a u t r e s pkospho l ip ides , par cont re les ac ides en C 17 ramifies, sont des eons t i - t u a n t s min.eurs des cardiol ipiffes .
l~n rdco l t an t les bae tdr ies en phase s t a t i o n n a i r e de croissance, nous n ' avons pas t rouvd de dif fdrences s ignif icat ives dons la compos i t i on en acides gras (posi- t i o n s 1 + 2) t~es t ro i s phospho l ip ides .
TABLEAU I.
D i s t r i b u t i o n des ac ides gras des phospho l ip ides des ce l lu les ent i6res de M. lysodeikticus (*).
Restes aeyls
1 4 : 1
1 4 : 0
1 5 r : 0
1 6 : 1
1 6 : 0
1 7 r : 0
1 8 : 1
t
1
4
71
5
9
11
t
Cardiolipides
PositioD
2 . 1 + 2
Phosphatidy | glye~rols Phosphatidylinositols
Position Position
1 2 t + 2 I 2 1 + 2 . . . . . . . . _ _ _ _
t 1 t 0 , 8 1 , 9 1 . 3
47 82 64 5 6 , 6 76 64
t t t t l t I t
(*) Exprim,d en pou reen t age p o u r chaque pos i t ion [1 ou 2] et p o u r les acides t o t a u x (1 + 2'). - - r ind*ique les res tes ac.yls ramifids. - - les n o t a t i o n s te l les que 1 4 : 0 i n d i q u e n t le n o m b r e d'e ca rbones et Ie h o m b r e
d ' i n s a t u r a t i o n de Ia ehaine . - - t i nd ique d.es t races non m e s u r a b l e s ( in f6r ieur fi 0,5 p. cent) .
l ip ides 50 p. cent et phospha t idy l ino . s i t o l s I0 p. cent. Cette invers ion des r a p p o r t s en t re p h o s p h a t i d y l g l y - cdrol, s et ca rd io l ip ides peu t pro ven i r aussi b ien d ' une m eil, l eure e x t r a c t i o n d'es ca rd io l ip ides h p a r t i r de la m e m b r a n e , q, ue d~une f o r m a t i o n de ca rd io l ip ides an t o u r s de la p r d p a r a t i o n de la m e m b r a n e [7].
Nous avons d.dtermin,d la compos i t i on en aeides gras des t ro i s phosphoHpides e x t r a i t s des ee l lu les ent i6res .
Acides gras des phospholipides : Les aeides gras prd- p o n d d r a n t s sont des aci4es ramif ids a p p a r t e n a n t aux s6ries iso et an te i so avec I5 et 17 a t o m e s de carbone,
BIOCHIMIE, 1976, 58, n ° 3.
DISGUSS,ION.
La r d F a r t i t i o n des acides gras donnde dans le ta,bleau I m o n t r e q,ue p o u r une au t r e bactdr ie r iche en acides g ras ramifids, l ' ac ide en C 15 ramif id est prdf~- r e n t i e l l e m e n t en pos i t ion 2, ma t s h la diffdrenee de Bacillus subtilis, 1,es phospho l ip ides de Microcoeeus lysodeikticus en p,hase exponen t i e l l e de cro issance n'on.t pas tous la m~me compos i t i on m o y e n n e en aeides gras.
Les donn,6es d~e la l i t t 6 r a t u r e ne p e r m e t t a i e n t pas de p rdvo i r que l:a c o m p o s i t i o n des ae ides gras d~es card io-

Distribution d'acides 9ras ramifies. 379
l ip ides p o u v a i t 6 t re d i f f~rente de celle des p h o s p h a - t idy lg lyc6roIs . En effet, les p h o s p h a t i d y l g l y e ~ r o l s son t l~es p r 6 e u r s e u r s d i rec ts des c a r d i o l i p i d e s et il y a des v a r i a t i o n s i m p o r t a n t e s des p r o p o r t i o n s r e l a t i ve s de ees de ux p h o s p h o l i p i d e s oh ez Micrococcus lysodeikticus au eours d:,e la eroiss 'ance. De relies v a r i a t i o n s sont 6gale- m e n t obse rv6es ehez Diplococcus pneumoniae, d o n t la c o m p o s i t i o n en phosphol . ip ides es t vo i s ine [10].
I1, s e m b l e done q u e des bac t6 r i e s oil earc~iolipides et phosp,hatic~ylglyc~rols son t p r6pond~ran t s , mod i f i en t la p a t t i e pol~aire des l i p ides de l eur m e m b r a n e en a j u s - t an t les p r o p o r t i o n s de ces deux p h o s p h o l i p i d e s ; chez Micrococcus l!tsodeikticus eet aj~ustement pent , de pins , i m p l i q u e r une modi f i ca t ion d~e la c o m p o s i t i o n en ae ides gras de ces l ip ides , ee qui change la p a r t i e non po la i r e d~e la mendarane .
Remerciemen ts.
Ce t r a v a i l a b$n6fiei$ d 'un e o n t r a t de f o n r n i t u r e s de r ad io~ lSment s d~u C o m m i s s a r i a t /t l 'Energ ie A tomique .
R E F E R E N C E S .
1. De Servio, A. J. (1969) J. Bacferiol., 100, 1342-1349. 2. Kaneda , T. (1972) Biochim. Biophys. Acre, 280, 297-
305. 3. Kaneda , T, (1972) B:~ochim. Biophys. Acta, 270, 32-
39. 4. Sal~on, M. R. J. & Freer , J. M. (1.965) Bioehim. Bio-
phys. Acta, 107, 5,31-536. 5. Di t tmer , J. C. • Les ter , R. L. (1964) J. Lipid Res.,
5, 1,96-1~7. 6. O k u y a m a , H. & No i ima, S. {1965') J. Biochem.
(Japon) 57, 52,9-538. 7. Bertsch~ L. L., Bonsen , P. P. M. & Kornberg , A.
[1969) J. Bacteriol., 98, 75-81. 8. De Servio, A. J. & Sialton, M. R. J. (1973) Microbios,
8, 73-78. 9. De Servio, A. J. & Sal ton, M. R. J. (1971) Biochim.
Biophys. Acla, 23~J, 280-292. 10. T r o m b e , M. C. • Lan6el le , M. A., r 6 su l t a t s non
publ'i~s.
BIOCHIMIE, 1976, 58, n ° 3.