Embed Size (px)
Citation preview

ACADEMIE DE MONTPELLIER
UNIVERSITE MONTPELLIER IlSCIENCES ET TECHNIQUES DU LANGUEDOC
THE5E
pour obtenir le grade de
DOCTEUR DE L'UNIVERSITE MONTPELLIER Il
Discipline: Biochimie et Biologie MoléculaireFormation Doctorale: Parasitologie
Ecole Doctorale: Biologie Intégrative
présentée et soutenue publiquement
par
Diana VILLARREAL PENA
le 16 octobre 2003
Titre:
ETUDE DE L'EXPRESSION DIFFERENTIELLE DES GENES IMPLIQUES DANSLA RESISTANCE AU BENZNIDAZOLE CHEZ TRYPANOSOMA CRUZI,
L'AGENT DE LA MALADIE DE CHAGAS.
JURY
Mme Catherine Braun-Breton, Professeur-Université Montpellier Il PrésidentMme Paola Minoprio, Chef de Laboratoire - Institut Pasteur RapporteurMme Marie-Laure Dardé, Professeur - Université de Limoges RapporteurM. Philippe Nirdé, Chargé de Recherche - CNRS ExaminateurM. Serge Morand, Directeur de Recherche - CBGP ExaminateurM. Michel Tibayrenc, Directeur de Recherche - IRD Directeur de Thèse

A ma mère, Antonia
Pour m'avoir donné tout son amour,
Pour m'avoir montré toutes les vertus,
Pour avoir dédié toute sa vie à nous et aux autres
A mon père, Victor Manuel
J'espère pouvoir récupérer le temps perdu,
Ce n'est jamais trop tard pour recommencer.
A mon frère, J ainer
A meS sœurs, Nancy, Yolanda, Olga Lucia et Marcelo
Pour m'avoir encouragée et soutenue tout le temps.
A toute ma famille et celle de mon époux,
Pour leur soutien permanent au cours de ces années de thèse.
A mon époux, Juan Carlos
C'est grâce à toi que j'ai trouvé un sens à ma vie,
C'est grâce à toi que je suis ici et que j'ai réussi à
vaincre mes peurs, mes angoisses, mes doutes,...
Merci pour être toujours à mes côtés, dans les bons et mauvais jours.
Je tiens à toi.
ii

REMERCIEMENTS
Les travaux présentés dans ce mémoire ont été réalisés au Centre IRD de
Montpellier, au sein de l'équipe de "Génétique des Maladies Infectieuses, GMI
(Ex-CEPM)". C'est grâce à une bourse de thèse accordée par l'IRD dans le
contexte du Programme de Renforcement des Communautés Scientifiques du
Sud, que cette recherche a été possible.
Je tiens à remercier particulièrement le Docteur Michel TIBAYRENC qui m'a
permis de venir en France et de travailler au sein de son équipe de" recherche.
Ma reconnaissance va également aux membres du Jury: Catherine BRAUN
BRETON, Paola MINOPRIO, Marie-Laure DARDE, Philippe NIRDE et Serge
MORAND, qui ont accepté de juger ce travail et qui l'ont enrichi de leurs
commentaires et discussions.
Je voudrais aussi témoigner ma gratitude spécialement à Christian BARNABE et
Mallorie HIDE, qui ont accepté d'encadrer mon travail et m'ont donné leurs
temps et conseils, particulièrement précieux tout au long de ma thèse. Merci de
tout cœur également pour l'amitié que vous m'avez témoignée pendant ces
années. Un grand merci aussi à Anne-Laure BANULS pour sa disponibilité et ses
conseils toujours enrichissants.
Philippe NIRDE et Denis SERENO m'ont fait partager leurs connaissances et
leur expérience, ils m'ont donné des conseils pertinents qui ont enrichi
considérablement mon travail. Je les remercie très sincèrement.
Hi

Un grand merci à Bruno OURY, Jean-Loup LEMESRE et Françoise MATHIEU
DAUDE de l'équipe "Pathogénie des Trypanosomatidés" ainsi qu'à Christophe
BRUGIDOU, Jean-Paul BRIZARD, Benoît FARINAS, Christelle SIRE et
Marjolaine VENTELON de "GeneTrop", pour l'aide et tous les conseils qu'ils
m'ont apportés dans les moments opportuns.
Un grand merci à Jean-François GUEGAN "pour m'avoir enlevé une épine du
pied", à Marie Claude LECLERC pour son aide précieuse et à Frédéric THOMAS,
ils ont partagé mon quotidien et m'ont soutenue durant ce temps-là.
Merci aux amis et copains du laboratoire CEPM, spécialement à Loubna TAZI (tu
me manques beaucoup), à Hélène BROUTIN (qui a été toujours disponible pour
moi), à Bénédicte LAFAY (pour son constant encouragement), à Nadine MAURY
(pour son aide), à Sylvain GODREUIL (pour son intérêt),... pour votre amitié,
merci de tout cœur.
Merci à Alfredo et sa famille, à Alix et sa famille, à Marie et sa famille, à
Philippe et sa famille, à Sandra et Juan, à Nayani, à Philippe, Françoise et
Pierre, pour tous les moments de joie partagés.
Et finalement, merci à tous ceux qui ont eu la gentillesse de corriger mon
manuscrit.
Je n'oublierai jamais ma vie en France! ! !
iv

TABLE DE MATIERES
INTRODUCTION GENERALE xi
1 CHAPITRE 1. GENERALITES 1
1.1 MALADIE DE CHAGAS 11.1.1 Trypanosoma cruzi 21.1.1.1 Diversité génétique du parasite 21.1.1.2 Cycle de vie des parasites 41.1.2 Transmission 61.1.3 Symptômes et pathogénicité 71.1.4 Diagnostic 81.1.5 Traitement. 91.1.6 Chimiothérapie et ses perspectives 121.1 .7 Contrôle 12
1.2 STRUCTURE DU GENOME ET EXPRESSION GENETIQUE DE T. cruzi.. 131.2.1 Taille du génome de T. cruzi 131.2.2 Organisation du génome de T. cruzi 141.2.3 Composition du génome de T. cruzi en séquences répétées 151.2.4 Gènes et familles de gènes 161.2.5 Expression des gènes 161.2.5.1 Transcription 171.2.5.2 Mécanismes post-trancriptionnels 181.2.5.3 L'édition des ARl\Jm du kinétoplaste 21
1.3 COMPORTEMENT DES PARASITES FACE AUX MEDiCAMENTS 231.3.1 Sensibilité des parasites aux médicaments (Benznidazole et Nifurtimox) 231.3.2 Mécanismes de résistance chez d'autres Trypanosomatidés et autresparasites 261.3.3 Gènes candidats potentiellement intéressants chez T. cruzi 311.3.3.1 La Superoxyde Dismutase 311.3.3.2 Les Cystéine Protéases 351.3.3.3 P-Glycoprotéines 38
2 CHAPITRE II. MATERIEL ET METHODES .41
2.1 MATÉRIEL BIOLOGIQUE 41
2.2 METHODOLOGIE 42
2.2.1 CULTURE DES PARASITES 422.2.1.1 Cinétique de croissance des parasites 422.2.1.2 Préparation des culots parasitaires 432.2.2CARACTERISATIONS BIOCHIMIQUE ET MOLECULAIRE DES PARASITES
......................................................................................................432.2.2.1 Electrophorèse Isoenzymatique Multilocus (MLEE) 43 .2.2.2.2 Amplification de fragments d'ADN par amorces aléatoires (RAPD) 44
v

2.2.3 EVALUATION DE LA REPONSE DES EPIMASTIGOTES A LA PRESENCEDU BENZNIDAZOLE 462.2.3.1 Détermination de l'ICso par la micro-méthode MTT/PMS 462.2.3.2 Détermination de l'ICso par cytométrie de flux 482.2.3.3 Croissance des parasites sans/avec Benznidazole à l'ICso 482.2.3.4 Induction de la résistance au Benznidazole 48
2.2.4 ETUDE DE L'EXPRESSION· GENIQUE DIFFERENTIELLE PAR LATECHNIQUE DERIVEE DU RADES 502.2.4.1 Mise au point de la technique dérivée du RADES 512.2.4.2 Etude de l'expression génétique différentielle à l'ICso par RADES 552.2.4.3 Etude de l'expression génétique différentielle des stocks sauvages etrésistants au niveau thérapeutique 562.2.4.4 Etude de "expression génétique différentielle des stocks sauvages etrésistants à différents niveaux 562.2.4.5 Clonage et séquençage des bandes différentielles d'intérêt, issues des stocksrésistants et sauvages 57
2.2.5 ETUDES DES GENES SPECIFIQUES 612.2.5.1 Amplification du gène de ménage 622.2.5.2 Amplification des gènes spécifiques d'intérêt 63
2.2.6 METHODES D'ANALYSE DES DONNEES 652.2.6.1 Analyses statistiques 652.2.6.2 Analyses des séquences 66
3 CHAPITRE III. RESULTATS 68
3.1 CARACTERISATIONS BIOCHIMIQUE ET MOLECULAIRE DES PARASITES .. 683.1.1 Isoenzymes (MLEE) 683.1.2 RAPD 703.1.3 Analyses phylogénétiques 72
3.2 EVALUATION DE LA REPONSE DES PARASITES A LA PRESENCE DEBENZNIDAZOLE 73
3.2.1 Cinétiques de croissance des parasites 733.2.2 Sensibilité des parasites au Benznidazole 773.2.2.1 ICsodéterminée par la méthode MTI/PIVIS 773.2.2.2 ICso déterminée par cytométrie de tlux 783.2.3 Niveaux de résistance obtenus avec le Benznidazole 79
3.3 ETUDE DE L'EXPRESSION DIFFERENTIELLE PAR LA METHODE DERIVEEDU RADES 80
3.3.1 Etude de l'expression différentielle à l'ICso 803.3.2 Etude de l'expression différentielle au niveau de chimiorésistancethérapeutique 843.3.3 Etude de l'expression différentielle à différents niveaux de chimiorésistance. 863.3.4 Clonage et séquençage des bandes différentielles sélectionnées 87
vi

3.4 ANALYSES DE CORRELATION ENTRE VARIABLES 91
3.5 ETUDE DE L'EXPRESSION DES GENES SPECiFIQUES 923.5.1 fcsod-A et fcsod-B 933.5.2 fccp1 943.5.3 fcpgp2 95
4 CHAPITRE IV. DISCUSSION 97
4.1 COMPORTEMENT DES PARASITES VIS A VIS DU BENZNIDAZOLE 974.1.1 Sensibilité des parasites au Benznidazole 974.1.2 Comportement des parasites résistants versus parasites sauvages 100
4.2 ETUDE DE L'EXPRESSION DIFFERENTIELLE DES GENES 1034.2.1 RADES à l'ICSD sauvage et au niveau de chimiorésistance thérapeutique (50IJM) 1044.2.2 RADES à différents niveaux de chimiorésistance au Benznidazole 106
4.3 EXPRESSION DES GENES SPECIFIQUES 111
4.4 STABILITE DES GENOTYPES DES STOCKS 114
5 CONCLUSiON 117
6 PERSPECTiVES 118
7 REFERENCES BIBLIOGRAPHIQUES 120
8 ANNEXES 138
PUBLICA·rIONS 166
vii

LISTE D'ABREVIATIONS
A : Absorbance
ADN: Acide désoxyribonucléique
ADNe: ADN complémentaire
ANOVA : Analyse de variance
ARN : Acide ribonucléique
ARNm : ARN messager
BET : Bromure d'éthydium
CP : Cystéine protéase
DEPC : Diethyl pyrocarbonate
DMSO: Diméthyl sulfoxide
D.O. : Densité optique
DTU : Discrete Typing Unit (Unité de typage discrète)
ICso : Concentration inhibant 50% de croissance
IP : Iodure de Propidium
LIT : Milieu de culture Liver Infusion Tryptose
L30TC : Milieu de culture LIT modifié
min: Minute
MLEE : Multilocus Enzyme Electrophoresis (Electrophorèse isoenzymatique multilocus)
MTT/PMS : [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide] 1Phenazine methosulphate
NCBI : National Center for Biotechnology 1nformation
NNN : Milieu de culture I\JNN (de Noy, Mac Neal et Nicolle)
PCR: Polymerase Chain reaction (Réaction de polymérisation en chaîne)
PGP : P-Glycoprotéine
ORF : open reading frame (Cadre ouvert de lecture)
RADES: Random Arnplified Differentially Expressed Sequences (Amp.Hficationaléatoire de séquences exprimées)
RAPD : Random Amplified Polymorphie DNA (Amplification de fragments d'ADN paramorces aléatoires)
RFLP: Restriction Fragment Length Polymorphism (Polymorphisme de longueur desfragments de restriction)
rprn : Révolution par minute
5 : Seconde
SDS : Sodium Dodecil Sulfate
SLe: Splice~ leader ou séquence du miniexon de Trypanosoma cruzi
SOD : Superoxyde dismutase
TAE : Tampon Tris acétate EDTA
RT-PCR : Retrotranscription-Polymerase Chain reaction (Transcription inverse couple àune réaction de polymérisation en chaîne)
viii

GLOSSAIRE
ADN complémentaire: ADN simple brin, qui est une copie d'un ARN obtenue par unetranscription inverse.
ADN espaceur: Séquence d'ADN non transcrit, séparant les gènes à l'intérieur des unitésrépétées.
Amorce: Oligonucléotide qui, hybridé avec une matrice d'acide nucléique, permet à unepolymérase d'initier la synthèse du brin complémentaire.
ARN monocistronique: ARN ne comportant qu'une seule unité d'information génétique(cistron).
ARNm polycistronique : ARN messager contenant plusieurs cistrons, et donc codant pourplusieurs chaînes polypeptidiques distinctes.
ARN polycistronique : ARN comportant plusieurs unités d'informations génétiques (cistrons).
ARN précurseur: ARN représentant le produit de transcription primaire d'un gène.
Cadre de lecture: Ordonnancement des nucléotides en codons. Le cadre de lecture définitquel ensemble de 3 nucléotides est lu comme codon. Il est déterminé par le codon d'initiation etle codon de terminaison.
Cadre ouvert de lecture: Séquence contenant une série de triplets codant pour les acidesaminés, non interrompue par un codon de terminaison.
Cistron: Région du génome qui ne porte qu'une seule unité d'information génétique transcriteen ARN.
Clone, clonai, clonalité: dans un contexte de génétique des populations, une espèce estconsidérée comme clonale quand la descendance est génétiquement identique au progéniteur.Cette situation se rencontre, non seulement en situation de c10nalité mitotique "classique"(mode de reproduction habituel de nombreuses espèces bactériennes), mais également dansde nombreux cas de parthénogénèse, et dans certains cas de reproduction parautofécondation.
Coiffe: Courte séquence nucléotidique ajoutée par modification post-transcriptionelle, àl'extrémité 5' de l'ARNm chez les Eucaryotes.
Contrôle en cis : Régulation de l'expression génétique par les séquences d'ADN au voisinaged'un gène situé sur le même chromosome.
Contrôle en trans : Régulation de l'expression génétique qui s'exerce par l'intermédiaire d'unfacteur diffusible.
Epissage : Processus englobant l'excision des introns et la réunion des exons dans l'ARN.
Exon : Séquence du gène représentée dans l'ARNm mature.
Expression génétique: Ensemble de mécanismes qui assurent la transcription et la traductionde l'information génétique.
Extrémité 5' ou 3' terminale: Début ou fin d'une chaîne de polynucléotides (ARN ou ADN).
Famille de gènes: Ensemble de gènes ayant de grandes ressemblances fonctionnelles etstructurelles.
Gène: Segment d'acide nucléique comprenant les régions transcrites en ARN et les régionsrégulatrices adjacentes.
Gène de régulation: Gène dont la fonction essentielle est de contrôler le taux d'expressiond'un ou de plusieurs autres gènes.
Insert : Séquence d'ADN étranger introduite dans une molécule d'ADN donnée.
ix

Intron : Partie non codante du gène située entre deux exons.
Monocistronique : Se dit d'un ARNm ne possédant qu'un seul cistron et qui code donc pourune seule chaîne polypeptidique.
Plasmide: Molécule d'ADN extrachromosomique capable de se répliquer indépendamment.
Polyadénylation: Addition post-transcriptionelle d'un grand nombre de résidus dedésoxyadénosine monophosphate à l'extrémité 3' d'un ARN d'Eucaryote, y formant une queuepolyA.
Pré-ARN messager: ARN précurseur des ARNm.
Promoteur: Séquence d'ADN nécessaire à l'initiation de la transcription et le plus souventsituée en amont de la partie transcrite des gènes.
Séquençage: Détermination de l'ordre linéaire des composants d'une macromolécule.
Séquence codante : Partie d'un gène qui définit directement la séquence en acides aminés dela protéine correspondante.
Séquences répétées directes: Séquences identiques ou quasi identiques, présentes enplusieurs copies dans la même molécule.
Séquences répétées en tandem: Séquences répétées directes adjacentes.
Séquence unique: Séquence d'ADN qui n'existe qu'en un seul exemplaire dans le génome.
Signal de polyadénylation : Motif nucléotidique situé en aval de la partie codante d'un gène etqui définit la position où doit s'ajouter la séquence polyA sur l'ARNm.
Traduction: Processus de décodage de l'ARNm, conduisant à la synthèse d'une protéine.
Transcriptase inverse: Enzyme qui catalyse la formation d'ADNe à partir d'une matriced'ARI\lm. .
Transcription: Processus par lequel la séquence d'un gène est copiée en ARN.
Transposon: Fragment d'ADN susceptible de se déplacer d'un endroit du génome dans unautre.
Unité de transcription: Région du génome située entre un site d'initiation et un site determinaison de la transcription pour l'ARN polymérase.
Vecteur: Molécule d'acide nucléique dans laquelle il est possible d'insérer desfragments d'acide nucléique étranger, pour ensuite les introduire et les mantenir dansune cellule hôte.
x

INTRODUCTION GENERALE

Introduction Générale
INTRODUCTION GENERALE
La maladie de Chagas doit son nom au médecin brésilien Carlos R. J. das Chagas qui,
vers 1909, fut le premier à décrire l'agent causal de la maladie, le parasite
Trypanosoma cruzi. La complexité de cette maladie est en grande partie due à la
variation importante des caractéristiques biologiques des parasites. T. cruzi est un
protozoaire avec un cycle de vie long et complexe, comportant plusieurs stades, tant
chez le vecteur (des insectes appartenant à la sous-famille des triatominae) que chez
l'hôte vertébré (mammifères, l'homme inclus). D'autres aspects tels que la diversité des
formes cliniques ou le nombre important de vecteurs et de réservoirs impliqués,
montrent aussi la grande complexité de cette maladie.
La maladie de Chagas est présente dans presque toute l'Amérique latine avec des
manifestations et des caractéristiques épidémiologiques qui varient significativement
selon l'aire endémique. D'après les chiffres de l'OMS, il Yaurait aujourd'hui entre 16 et
18 millions d'individus parasités et au moins 100 millions de personnes exposées, ceci
dans 17 pays répartis sur toute la région d'Amérique Centrale et du Sud (WHO, 2002).
L'aire d'endémie s'étend du sud des Etats-Unis au nord de l'Argentine.
Cette maladie se développe lorsque les humains entrent en contact avec les foyers
d'infection en perturbant leur environnement, ce qui a pour conséquence que les
vecteurs infectés vont s'installer finalement dans les habitats péridomestiques et
domestiques. Chez l'homme, après l'infection, vient une période d'incubation suivie
d'une phase aiguë, caractérisée par des manifestations cliniques diverses. En l'absence
de traitement adéquat, les symptômes peuvent persister jusqu'à deux mois, avec des
taux de mortalité variables (2 à 8 %), surtout dans la population infantile (Rodrigues
Coura et de Castro, 2002).
Le traitement de la maladie de Chagas se limite actuellement à deux médicaments: le
Benznidazole et le Nifurtimox. Le premier semble agir sur la synthèse des protéines et
de l'ADN chez les parasites (Tacher, 1997), tandis que le deuxième aurait un effet à
travers la production de radicaux libres (Marr et Docampo, 1986). Les résultats obtenus
avec ces médicaments varient selon la phase de la maladie, la période et la dose du
traitement, ainsi qu'avec l'âge et l'origine géographique du patient. Bien que le
xi

Introduction Générale
Benznidazole et le Nifurtimox se soient montrés efficaces, surtout dans la phase aiguë
de la maladie (Rodrigues Coura et de Castro, 2002), l'apparition d'effets secondaires
indésirables chez les patients, ajoutée au manque de programmes médicaux adaptés,
ont limité le contrôle de la maladie dans les zones à risque. De plus, différents travaux
ont montré que parmi les stocks de T. cruzi, il existerait une variation très importante de
la sensibilité à ces deux médicaments (Neal et van Bueren, 1988 ; Toledo et al., 1997 ;
Revollo et al., 1998). Par ailleurs, l'existence de stocks naturellement résistants a été
déjà rapportée (Filardi et Brener, 1987).
Il semble que les propriétés biologiques du parasite soient expliquées (au moins en
partie) par l'existence de la grande diversité génétique décrite au sein des populations
naturelles de T. cruzi (Miles et al., 1981). En effet, plusieurs travaux ont permis de
corréler ces propriétés biologiques avec la diversité génétique in vitro (Laurent et al.,
1997; Revollo et al., 1998), dans le vecteur (De Lana et al., 1998) et in vivo (Toledo et
al., 2002, 2003). L'espèce Trypanosoma cruzi présente une structuration des
populations de type clonaI. Cette espèce n'est pas panmictique, les souches de T. cruzi
peuvent être considérées comme des clones qui échangent peu ou rarement des
informations génétiques (Tibayrenc et al., 1986 ; Tibayrenc et Ayala, 1988). Néanmoins,
des phénomènes occasionnels d'hybridation entre clones différents s'observent dans
les populations naturelles du parasite.
Différents mécanismes de résistance aux médicaments ont été décrits chez des
parasites tels que Leishmania, Plasmodium, Entamoeba et Trichomonas, entre autres.
Parmi les principaux mécanismes de résistance observés, le changement des
propriétés membranaires, la dégradation l'inactivation ou l'élimination du médicament,
la modification de la cible thérapeutique ou une réparation rapide des dommages
produits par les médicaments, ont été décrits (Borst et Ouellette, 1995 ; Ullman, 1995).
Chez les Trypanosomes, quelques mécanismes de résistance ont aussi été identifiés,
notamment dans l'espèce africaine Trypanosoma brucei, agent de la trypanosomose
humaine africaine. Ces mécanismes seraient apparemment impliqués dans
l'accumulation, l'absorption et l'élimination des médicaments (de Koning, 2001).
Néanmoins chez les autres espèces, et particulièrement chez T. cruzi, aucun
mécanisme jusqu'à présent, n'a été identifié comme étant associé à ce type de
xii

Introduction Générale
résistance. Ceci constitue un des obstacles majeurs pour le développement de
traitements efficaces contre la maladie de Chagas.
Parmi les différentes techniques qui ont été utilisées pour l'étude de l'expression
différentielle des gènes impliqués dans la résistance cellulaire aux médicaments, le
« Differentiai display » constitue l'une des techniques les plus répandues actuellement
(Wang et al., 2002 ; Higuchi et al., 2003). La technique RADES (pour Random Amplified
Differentially Expressed Sequences), est une méthode d'analyse moléculaire dérivée du
« Differentiai display». Cette technique permet d'obtenir un profil d'expression des
gènes à un moment donné, grâce à l'amplification aléatoire (RAPD) des produits de
rétro-transcription des ARN messagers. La technique RADES représente une technique
rapide, et qui, avec peu de matériel, donne des résultats reproductibles pOLIr l'étude des
gènes exprimés différentiellement chez les parasites, ceci avec un minimum d'effort en
terme de coût et de temps (Murphy et Pellé, 1994).
Face à la faible efficacité des médicaments actuels, ainsi qu'à la méconnaissance des
mécanismes impliqués dans la résistance aux drogues chez T. cruzi, les études sur la
résistance aux médicaments représentent une alternative importante pour le
développement de nouveaux médicaments, plus actifs et moins toxiques pour les
millions de personnes atteintes par cette maladie. C'est dans cette perspective que
nous nous sommes intéressés à l'étude de l'expression des gènes potentiellement
impliqués dans la résistance au Benznidazole chez T. cruzi. D'abord, nous avons induit
des résistances au Benznidazole in vitro en utilisant une pression médicamenteuse
continue. Ensuite, nous avons choisi deux approches pour aborder l'étude de la
résistance: une première approche en utilisant l'expression différentielle à l'aide d'une
technique dérivée du RADES, et une deuxième approche par l'étude des gènes·
spécifiques susceptibles d'être impliqués dans la résistance.
Notre hypothèse de travail était que le comportement du parasite face au Benznidazole
pourrait être en relation directe avec la structure c10nale des populations naturelles de
Trypanosoma cruzi, ou en d'autres termes que l'appartenance à un groupe génétique
donné puisse prédire le niveau de sensibilité d'un stock de T. cruzi au Benznidazole.
xiii

Introduction Générale
Par ailleurs, nous nous sommes demandé s'il y avait des différences dans l'expression
des gènes dans les stocks soumis a une condition de stress, comme par exemple au
contact bref du Benznidazole, et s'il existait des différences dans l'expression génique
entre les stocks induits à la dose de chimiorésistance thérapeutique, c'est-à-dire la dose
circulante de médicament chez un patient en cours de traitement? Les niveaux de
chimiorésistance acquis permettent-ils de mettre en évidence une sur-expression ou
sous-expression progressive de certains transcrits? en d'autres termes, s'il y avait des
différences dans l'expression des gènes à chacun des niveaux de chimiorésistance
acquis? Et enfin, nous nous sommes demandé quels sont les gènes qui pourraient être
impliqués dans le mécanisme de chimiorésistance de T. cruzi au Benznidazole?
Pour répondre à ces questions nos objectifs ont par conséquent été:
1. Etudier l'expression différentielle des gènes au niveau de sensibilité naturelle (ICso)
dans différents stocks représentatifs de la diversité génétique de l'espèce Trypanosoma
cruzi.
2. Comparer l'expression génique des parasites chimiorésistants ~u Benznidazole (au
niveau de chimiorésistance thérapeutique, 501JM) par rapport aux parasites sauvages.
3. Etudier les différences d'expression génique entre les parasites chimiorésistants au
Benznidazole (à différents niveaux de chimiorésistance) et les parasites sauvages.
4. Etablir quel sont les gènes potentiellement impliqués dans la résistance de T. cruzi
au Benznidazole, à partir de la caractérisation des séquences différentiellement
exprimées entre les stocks chimiorésistants et sauvages.
5. Analyser "expression de différents gènes candidats (fcsod-A, fcsod-B, fccp1 et fccp2)
potentiellement impliqués dans la chimiorésistance, chez les parasites chimiorésistants
au Benznidazole ainsi que chez les parasites sauvages.
xiv

GENERALITES

Généralités
1 CHAPITRE 1. GENERALITES
1.1 MALADIE DE CHAGAS
La maladie de Chagas est une parasitose humaine appelée aussi trypanosomose
américaine, provoquée par un protozoaire flagellé, appartenant au genre Trypanosoma:
Trypanosoma cruzi.
Entre 16 et 18 millions de personnes sont touchées en Amérique du Nord, en Amérique
Centrale et en Amérique du Sud oCl la maladie s'étend du sud des Etats-Unis au Nord
de l'Argentine. Environ 100 millions de personnes, soit 25% de la population
d'Amérique latine, sont exposées au risque de contracter la maladie (OMS, 1998).
Au Honduras par exemple, 1,8 millions de personnes vivent dans des zones
endémiques, dont 300 000 seraient infectées par la maladie de Chagas (Médecins sans
Frontières, 2003).
En Colombie, la maladie atteint 5% de la population et 20% sont exposés à l'infection.
On estime que chaque année 500 000 personnes sont infectées, parmi lesquelles 300
000 enfants. La mortalité pendant la phase aiguë de la maladie est estimée entre 10 et
15% (Guhl, 1998). En Colombie, le parasite est très fréquemment détecté tout le long
du bassin de la rivière Magdalena, dans la région du Catatumbo, dans la Sierra Nevada
de Santa Marta, dans le piémont de Llanos Orientales et dans la Serrania de la
Macarena. Les départements colombiens qui présentent une endémicité élevée
sont Santander, Norte de Santander, Cundinamarca, Boyacâ, Meta, Casanare, Arauca,
Tolima, Huila et Bolivar.
La maladie de Chagas est caractérisée par trois phases: une phase aiguë, de courte
durée et une phase chronique prolongée, séparées par une longue phase de latence
clinique, dite phase indéterminée. La phase chronique peut provoquer une cardiopathie
(méga-cœur), une viscéropathie (méga-œsophage, mégacôlon) ou une neuropathie
(voir chapitre 1.1.3).
1

Généralités
Cette maladie est essentiellement rurale et affecte les habitants des « ranchos » ou
« cafuas », aux murs de torchis (en boue et paille) et à toit de chaume. Mais elle sévit
également dans les quartiers défavorisés qui ceinturent les grandes villes d'Amérique
latine, nommés « favelas» ou « poblaciones » (Gentilini, 1995).
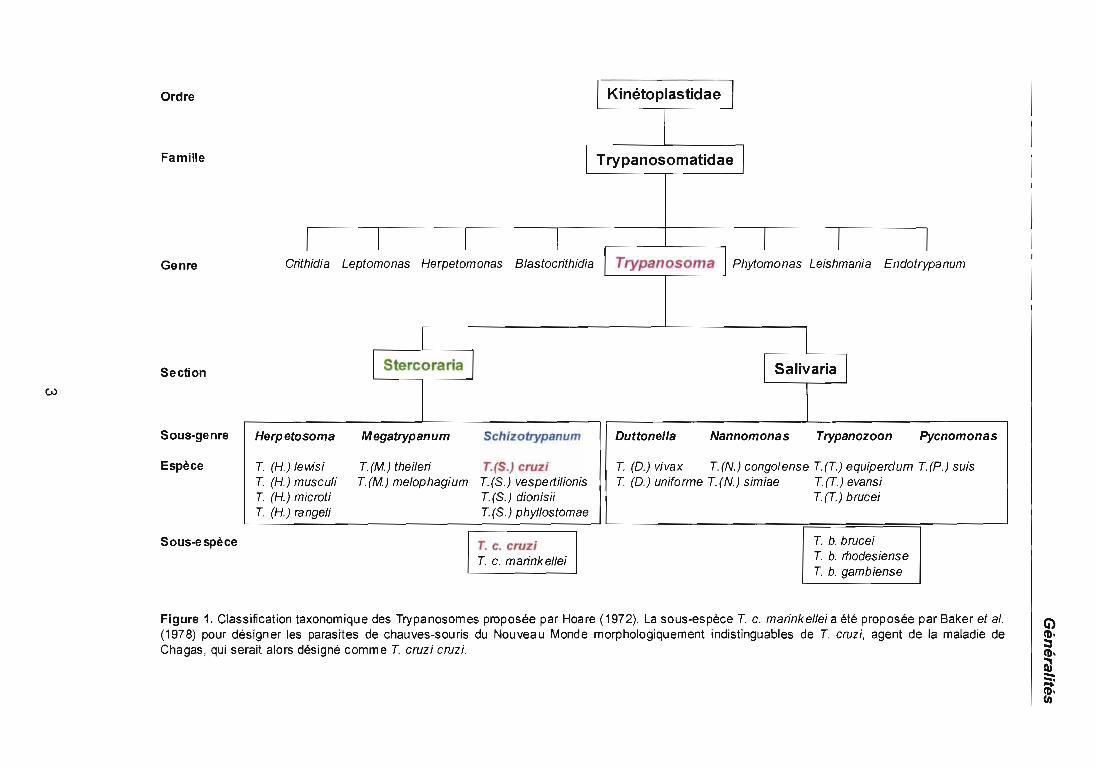
1.1 .1 Trypanosoma cruz;
L'agent étiologique de la maladie de Chagas est le parasite Trypanosoma
[Schizotrypanum] cruzi (Chagas, 1909), qui appartient au phylum des Kinetoplasta,
ordre Kinetoplastida, famille Trypanosomatidae, genre Trypanosoma, sous-genre
Schizotrypanum, voir figure 1. Ce parasite est caractérisé par un flagelle libre présent,
par exemple, chez les formes épimastigotes, un kinétoplaste volumineux, ovulaire ou
arrondi (voir plus loin description de cette structure), une taille de 4 ~m de large sur 20
~m de long et une forme de demi-lune.
1.1.1.1 Diversité génétique du parasite
Les populations naturelles de Trypanosoma cruzi montrent un polymorphisme
génétique considérable qui a d'abord été montré par l'utilisation des isoenzymes (Toyé,
1974 ; Miles et al., 1978). Tibayrenc et al. (1986) ont ensuite décrit 43 génotypes
différents en utilisant 15 systèmes isoenzymatiques pour 121 stocks issus de différents
environnements. Ces auteurs ont démontré que T. cruzi présente une structure des
populations de type essentiellement c10nale avec pas ou peu d'événements de
recombinaison. L'espèce n'est donc pas panmictique, les souches de T. cruzi peuvent.. " ." ~.
être considérées comme des clones échangeant peu d'information génétique.
Plus tard, il a été montré que l'espèce T. cruzi est très fortement structurée en deux
lignées phylogénétiques majeures (Tibayrenc 1995, 1996 ; Souto et al., 1996). La
première lignée est statistiquement plus présente en milieu sylvestre et la seconde en
milieu péri-domestique. Ces deux lignées ont été désignées par un comité d'experts T.
cruzi 1 et T. cruzi Il (Momen, 1999).
La lignée T. cruzi Il est elle-même structurée en cinq sous-groupes génétiquement
distincts lia, lib Ilc, IId et Ile, mais qui ont cependant des parentés génétiques entre
eux.
2
Ordre
Famille
1 Kinétoplastidae 1
1 Trypanosomatidae 1
Crithidia Leptomonas Herpetomonas BlastocrithidiaGenre
1 1 1 1 1 1 1
Trypanosoma J Phytomonas Leishmania Endotrypanum
Section 1 Stercoraria J 1 Salivaria 1
Sous-genre Herpetosoma Megatrypanum Schizotrypanum Duttonella Nannomonas Trypanozoon Pycnomonas
Espèce
Sous-espèce
T (H.) leVoAsiT (H) musculiT (H) microtiT (H.) ra ngeli
T (M.) theileriT(M.) melophagium
T.(S.) cruziT (S.) vespe rtilionisT (S.) dionisiiT (S.) phyllostomae
T. c. cruzlT c. marinkellei
T (O.) vivax T(N.) congolense T.(T) equiperdum T(P) suisT (O.) uniforme T(N) simiae T(T) evansi
T(T) brucei
T b. bruceiT b. rhodesienseT b. gambiense
Figure 1. Classification taxonomique des Trypanosomes proposée par Hoare (1972). La sous-espèce T c. marinkellei a été proposée par Baker et al.(1978) pour désigner les parasites de chauves-souris du Nouveau Monde morphologiquement indistinguables de T cruzi, agent de la maladie deChagas, qui serait alors désigné comme T cruzi cruzi.

Généralités
En effet, deux sous-groupes (lld et Ile) sont clairement des lignées hybrides qui ont
conservé beaucoup de caractères hétérozygotes à différents loci. Ces sous-group~s, du
fait des phénomènes d'hybridation, ne peuvent être considérés comme de réelles
clades, puisque certains ont deux ancêtres au lieu d'un seul. Tibayrenc (1998) a
proposé pour les désigner le concept de "Discrete Typing Unit" (DTU): ensemble discret
de génotypes génétiquement apparentés, caractérisables par un ou plusieurs
caractères génétiques ou "tags". T. cruzi apparaît ainsi subdivisé en un total de 6 DTUs:
l, lia, lib, Ile, Iid et Ile (Barnabé et al., 2000 ; Brisse et al., 2000).
D'autres techniques comme le RFLP (pour Restriction Fragment Length Polymorphism)
de l'ADN kinéstoplastique (Morel et al., 1980), le RAPD (pour Random Amplified
Polymorphie DNA, Tibayrenc et al., 1993 ; Brisse et al., 2000), la caractérisation du
rniniexon (McCarthy-Burke et al., 1989), l'ADN ribosomique (revue, Macedo et Pena,
1998), l'analyse des microsatellites (Oliveira et al., 1998, 1999 ; Macedo et al., 2001),
ont permis l'étude de la structure génétique du parasite. Toutes ces méthodes
moléculaires confirment la présence des deux lignées au sein de T. cruzi, et certaines
d'entre elles confirment la sous-structuration de la lignée II.
1.1.1.2 Cycle de vie des parasites
Comme beaucoup de parasitoses, la maladie de Chagas fait intervenir deux hôtes dans
son cycle parasitaire: un hôte intermédiaire, un insecte hématophage strict de la famille
des réduviîdés et de la sous-famille des triatominés (vecteur de la maladie), et un hôte
définitif, qui ne peut être qu'un mammifère (homme, et tous mammifères domestiques
ou sylvestres).
Les vecteurs se contaminent en prenant leur repas sanguin sur un homme ou un animal
parasité (1, figure 2). Dans le vecteur, les parasites passent successivement de la
forme trypomastigote (2) à celle d'épimastigote (3), cette transformation a lieu dans
l'intestin moyen de l'insecte. Ensuite, les épimastigotes se multiplient activement
jusqu'au moment où ils sont entraînés vers le rectum sous la forme de sphéromatigote,
donnant lieu, en fin de parcours, aux trypomastigotes métacycliques (4). Toute cette
évolution se déroule à l'intérieur de l'intestin du vecteur (Esquerdo-G6mez, 1996).
4

Généralités
Les formes métacycliques sont rejetées de l'intestin avec les fèces et se déposent sur
la peau et les muqueuses de leur hôte (5). C'est en se grattant suite à la piqûre, que
l'hôte fera pénétrer les parasites dans la circulation sanguine jusqu'à la fixation dans les
tissus (6). Le parasite peut également pénétrer par les muqueuses, en particulier la
muqueuse conjonctivale, en l'absence de tout grattage.
Figure 2. Schéma du cycle de vie du parasite Trypanosoma cruz; (d'aprèsTDR/Wellcome Trust, 2003).
Dans les cellules de l'hôte définitif, le parasite adopte la forme amastigote (7) ou
cryptomastigote. Il s'agit d'une forme intracellulaire sans flagelle au sein d'une vacuole
parasitophore. C'est sous cette forme que l'on trouve le parasite, par exemple, dans les
fibres musculaires (myocarde). Et c'est sous cette forme qu'il se reproduit chez son
hôte vertébré. En quelques heures, les amastigotes se multiplient par plusieurs cycles
de division cellulaire, puis se transforment en forme métacyclique. Les cellules infectées
éclatent sous la pression de la charge parasitaire et les parasites libérés vont infecter
d'autres cellules (cellules musculaires, cellules nerveuses, cellules du système réticulo
endothélial et macrophages). Dans le sang humain, les parasites libérés après
l'éclatement des cellules, sont sous forme trypomastigote (8), qui définit une forme
trapue du parasite avec un noyau allongé, un kinétoplaste volumineux sub-terminal et
5

Généralités
un ·~agelle court. Sous cette forme, le trypomastigote ne se multiplie pas (Esquerdo
G6mez, 1996).
Récemment, différentes morphologies de trypomastigotes ont été caractérisées,
démontrant qu'il existe plusieurs formes intermédiaires (Tyler et Engman, 2001). Enfin,
lors de son repas, le vecteur s'infecte en ingérant des formes trypomastigotes
présentes dans la circulation sanguine de l'hôte, bouclant ainsi le cycle parasitaire.
1.1.2 Transmission
La maladie de Chagas est provoquée par des parasites qui sont transmis à l'homme
par des insectes qui vivent proches des habitats humains : vinchucas, chipos, pitos,
chupones, dans les pays hispanophones, chupao et barbeiro au Brésil et "kissing bugs"
ou "assassin bugs" aux Etats-Unis.
Ces insectes vecteurs appartiennent à l'ordre des Hemiptera, à la famille des
Reduviidae et à la sous-famille des Triatominae. On en connaît 117 espèces réparties
en 14 genres, dont 13 genres (105 espèces) se rencontrent sur le continent américain.
Plus de la moitié d'entre elles ont été trouvées infestées par T. cruzi (Esquerdo-G6mez,
1996). On a décrit plus de 50 espèces dont 3 sont bien adaptées à l'habitat humain
(Triatoma infestans , Rhodnius prolixus et Panstrongylus megistus). Ce sont des
insectes hématophages stricts.
En Colombie, 20 espèces sont considérées comme vecteurs de T. cruzi. Parmi ces
espèces, celles qui ont le plus d'importance épidémiologique sont: Rhodnius prolixus,
Triatoma dimidiata, Triatoma maculata, Rhodnius robustus, Rhodnius brethesi,
Triatoma venosa et Rhodnius pallescens (Guhl, 1998). Les espèces T. venosa et T.
maculata sont bien adaptées en Colombie aux milieux domestiques, mais leur
importance dans la transmission de la maladie de Chagas est moind~e. L'espèce R.
prolixus reste sans doute le principal vecteur. Les régions à risque de transmission
vectorielle de T. cruzi en Colombie sont présentes dans l'annexe 1.
Les réservoirs animaux sont nombreux tant au niveau domestique (chiens, chats, rats
de grenier) que sauvage (rongeurs, tatous, sarigues, chauves-souris). En Colombie, au
moins une centaine de mammifères sylvestres jouant ·Ie rôle de réservoirs ont été
6

Généralités
décrits, parmi eux: Dasypus novemcintus, Didelphys marsupiales, Mus musculus,
Rattus rattus, Phyllostomus hastatus et plusieurs genres de singes comme les Saimiri,
Cebus, Ate/es et Alouatta. Parmi les animaux domestiques parasités on trouve les
chiens, les chats et les porcs (Guhl, 1998). Les oiseaux, en particulier les poules, sont
réfractaires à la maladie de Chagas. Cependant, les triatomes s'alimentent sur eux, et
les poules contribuent de ce fait indirectement à l'entretien des foyers chagasiques.
En plus du mode de transmission naturel par le vecteur, cette maladie peut également
se transmettre par transfusion sanguine (Schmunis et al., 1998), par transplantation
d'organes infectés (Zayas et al., 2002) et de la mère à l'enfant pendant la grossesse,
pendant l'accouchement ou pendant l'allaitement si la mère est infectée. Le passage
transplacentaire a lieu préférentiellement vers le 5ème mois de la grossesse.
Apparemment, l'infection de la mère n'a aucun effet sur sa fécondité, mais l'infection
transplacentaire peut provoquer un avortement ou un accouchement prématuré.
L'incidence de la transmission congénitale de la maladie de Chagas peut varier selon
les régions géographiques et peut atteindre 3,5% des nouveau-nés en Argentine,
10,5% au Brésil (à Bahia), 14,8% en Bolivie (à Santa Cruz) ou encore 18,8% au Chili
(Noireau, 1999).
1.1.3 Symptômes et pathogénicité
Après une semaine d'incubation, apparaissent les signes liés à la porte d'entrée des
parasites dans l'hôte humain. Si la manifestation est conjonctivale, on observe l'œil
enflé, avec un œdème (bipalpébral unilatéral) rose violacé, indolore, accompagné d'une
adénopathie pré-auriculaire satellite ou signe de "Romana" ; si elle est cutanée, on
observe une ulcération nécrotique du lieu d'infection ou "chagome" d'inoculation
(Gentilini, 1995).
La fièvre est un symptôme initial pratiquement constant. Elle dure entre un à deux mois
et oscille entre 38 et 39°C. La splénomégalie est fréquente chez les enfants. D'autres
signes généraux peuvent aussi apparaître, digestifs, respiratoires, cutanés ou nerveux
(méningo-encéphalite parfois mortelle).
La maladie peut évoluer de plusieurs façons: une forme aiguë qui peut se développer
rapidement, puis une phase chronique qui peut durer toute la vie et provoquer divers
7

Généralités
degrés de cardiopathie ou affecter les voies digestives (méga-œsophage ou
mégacôlon) ou la combinaison des deux ou encore affecter le système nerveux central
et périphérique. Lorsque la phase chronique commence, l'efficacité du traitement est
très discutée, car les symptômes tels que la faiblesse cardiaque ou des
dysfonctionnements sérieux de l'œsophage ou du côlon peuvent être irréversibles
(revue, Prata, 2001). Il existe aussi une forme indéterminée, qui se diagnostique soit
par dépistage systématique, soit en post-mortem. Globalement, 75% des patients ayant
survécu à la forme aiguë ne développent aucune symptomatologie clinique.
L'espérance de vie, pour les personnes atteintes par la maladie de Chagas, diminue de
9 années en moyenne (Médecins sans Frontières, 2003).
A partir de l'étude de la maladie de Chagas sur des modèles animaux, différents
mécanismes de pathogénicité au niveau du cœur ont été décrits, incluant l'inflammation
provoquée par des antigènes parasitaires spécifiques, la nécrose cellulaire du
myocarde induit par le parasite, le spasme microvasculaire et l'auto-immunité (Leon et
Engman, 2001). Quels que soient les mécanismes en cause, la conséquence principale
de la cardiopathie chagasique est une insuffisance cardiaque de très mauvais
pronostic.
Pour expliquer l'étiologie de la maladie, deux hypothèses ont été proposées. La
première, appelée hypothèse de la persistance, est la persistance du parasite dans des
sites spécifiques chez l'hôte infecté, ce qui provoquerait une réaction inflammatoire
chronique. La deuxième, appelée hypothèse de l'auto-immunité, est celle dans laquelle
l'infection de T. cruz; induit une réponse immune pour laquelle les propres tissus de
l'hôte sont cibles. Dans les deux cas, la pathologie, basée sur une réaction
immunologique, provoque la destruction des tissus et par conséquent les signes et les
symptômes cliniques de la maladie (Tarleton et Zhang, 1999; Zhang et Tartelon, 1999 ;
Tarleton, 2001).
1.1.4 Diagnostic
Dans les formes aiguës, les méthodes parasitologiques directes sont les plus adaptées,
en revanche, ellès sont négatives en phase chronique. Les méthodes employées dans
les zones endémiques sont l'analyse de sang au bout du doigt "goutte épaisse",
l'examen à l'état frais et les méthodes de concentration.
8

Généralités
On peut également cultiver le sang des hôtes dans du milieu NNN (de Novy, Mac Neal
et Nicolle) ou l'inoculer à divers animaux de laboratoire (souris ou cobayes). La
méthode la plus sensible est le xé~odiagnostic, dans lequel le patient est piqué par des
réduves saines au stade larvaire L3, dans les déjections desquelles on recherchera des
parasites de 20 jours à trois mois après la piqûre. On peut aussi isoler le parasite à
partir de ponction ganglionnaire, musculaire, conjonctival ou à partir d'un chagome, ou
chancre d'inoculation chagasique (Gentilini, 1995).
L'infestation par T. cruzi déclenche l'apparition d'anticorps de classe IglVI. Ceux-ci, au
fur et à mesure que l'infection s'installe, seront remplacés par ceux de la classe IgG.
Par conséquent, les patients présentant des anticorps IgM associés ou non à des
anticorps IgG spécifiques sont considérés comme étant en phase initiale aiguë et ceux
présentant uniquement des IgG sont considérés comme étant en phase indéterminée
ou chronique bien que dans ces deux phases, une immunosuppression puisse se
traduire par l'absence de production d'anticorps spécifiques (Esquerdo-Gomez, 1996).
Les méthodes les plus utilisées pour la détection d'anticorps sont la réaction de
déviation du complément (ROC), la réaction d'hémagglutination directe (AD et ME-2
AD) ou indirecte (HI), la réaction d'immunofluorescence directe (IFD) ou indirecte (IFI),
le test immunoenzymatique ELISA (pour Enzyme-Linked Immunosorbent Assay) et le
test d'immunocompétition.
D'autre part, il est possible de caractériser les clones naturels de T. cruzi, à partir de
leur ADN, sans mise en culture préalable, en utilisant la réaction d'amplification de
l'ADN par polymérisation en chaîne (PCR) (Moser et al., 1989 ; Brenière et al., 1992).
Pour détecter des parasites chez homme ou chez le vecteur il est également possible
d'utiliser des amorces spécifiques des ARN ribosomiques (Gonzalez et al., 1994) ou du
mini-cercle (Avila et al., 1991) (revue, Portela-Lindoso et Shikanai-Yasuda, 2003).
1.1.5 Traitement
Le traitement de cette maladie est basé sur l'utilisation de deux
médicaments nitrohétérocycliques: le Nifurtimox (Lampit®) et le Benznidazole
(Rochagan®! Radanil®).
9

Généralités
Le Nifurtimox est un 5-nitrofurane (3-methyl-4-(5'-nitrofurfurylideneamine) tetrahydro
4H-1 ,4-tiazine-1,1-di-oxide), Bayer 2502. Le Benznidazole est un 2-nitroimidazole (N
benzyl-2-nitro-1-imidazole-acetamide), RO 7-1051. La figure 3 présente la structure
chimique de ces deux médicaments.
L'activité antiparasitaire du premier (Nifurtimox) consiste en la réduction métabolique du
groupe nitro en radicaux nitroanions (R-N02') très réactifs qui réagissent avec
l'oxygène pour produire des radicaux libres comme l'anion superoxyde (02' ) et le
peroxyde d'hydrogène (H202) (Docampo et al., 1981 ; Docampo et Moreno, 1984, 1986;
Marr et Docampo, 1986).
Le Benznidazole (2-nitro-imidazole) agit en se liant de façon covalente aux
intermédiaires de la nitroréduction du parasite (Moreno et al., 1982 ; Masana et al.,
1984 ; Croft et al., 1997). Il est apparemment capable de produire aussi des radicaux
libres comme le O2' et H20 2 (Guilivi et al., 1988). Bien que le mode d'action de
Benznidazole ne soit pas complètement éclairé, il semble pouvoir inhiber la synthèse
des protéines et d'ARN chez le parasite (Polak et Richie, 1978), et peut-être,
endommager l'ADN (Tacher, 1997).
a
/-\N N
YN+
-' ~o o.
o
~NH
b
Figure 3. Structure chimique du Nifurtimox (a) et du Benznidazole (b).
10

Généralités
Trypanosoma cruzi ne possédant pas les mécanismes de détoxication des métabolites
de l'oxygène, il est plus sensible à ces radicaux libres que les cellules de l'hôte.
Néanmoins, chez l'hôte, ces médicaments sont également réduits en radicaux NO
. avec augmentation des anions oxygènes, peroxydes d'hydrogène et peroxydes de
lipides, phénomène qui pourrait ainsi expliquer les effets secondaires produits chez les
patients traités (Marr et Docampo, 1986).
Les effets secondaires produits par le Nifurtimox sont divers: anorexie, perte de poids,
altérations psychiques, excitabilité ou somnolence, troubles de type digestif (nausées,
vomissement et parfois colique abdominale et diarrhée) et aussi des complications
neurologiques telles que convulsions, paresthésies et polynévrites. Le Benznidazole
produit des effets importants comme des réactions d'hypersensibilité, dermatites et
éruptions cutanées, œdème généralisé, polynévrite, des troubles gastro-intestinaux,
fièvre, céphalées, vertiges, purpura thrombocytopénique pouvant s'accompagner de
leucopénie et d'agranulocytose entre autres (Brener, 1984 ; Marr et Docampo, 1986 ;
Teixeira et al., 1994 ; Khaw et Panosian, 1995 ; Cançado, 2002 ; Rodrigues Coura et de
Castro, 2002).
Le Nifurtimox, à la posologie de 8 à 10 mg/kg/jour, en trois prises orales pendant trois à
quatre mois est efficace en phase aiguë mais pratiquement sans effet sur les
viscéropathies chagasiques chroniques. Le Benznidazole à la posologie de 5 à 7,5
mg/kg/jour en deux à trois prises orales pendant trois à quatre semaines donne aussi
de bons résultats dans les formes aiguës. Mais suivant les souches de T. cruzi,
l'efficacité du médicament varie. Pour les patients de moins de 40 kg, la posologie peut
aller jusqu'au 12 mg/kg/jour de Nifurtimox et jusqu'au 7,5 mg/kg/jour de Benznidazole
pendant 30 à 60 jours (OPAS/OMS, 1998).
D'autre part, aucun de ces deux médicaments n'est considéré comme un traitement
parfaitement satisfaisant pour les raisons suivantes:
i) par manque d'efficacité dans la phase chronique,
ii) à cause des variations importantes observées entre les régions géographiques.
considérées, probablement dues à des souches de T. cruzi différentes,
iii) à cause d'un taux élevé d'abandon du traitement du fait des effets secondaires,
iv) à cause de la longue période de traitement (30-60 jours),
11

Généralités
v) à cause de souches naturellement résistantes aux médicaments, et enfin,
vi) à cause du besoin de contrôle des malades dans des structures médicales
adaptées.
1.1.6 Chimiothérapie et ses perspectives
Pendant des années, le traitement de la maladie de Chagas a consisté en l'utilisation
de Benznidazole et Nifurtimox, malgré leur efficacité discutée relative dans la phase
chronique de la maladie et leurs effets secondaires importants (Raether et Hanel, 2003)
et même néfastes dans bien des cas.
Il existe plusieurs molécules qui sont en voie d'expérimentation et qui pourraient être
utilisées contre la maladie de Chagas dans un proche avenir. Les molécules candidates
ont été abondamment décrites (Marr et Docampo, 1986 ; Marr, 1991 ; De Castro, 1993 ;
Khaw et Panosian, 1995 ; Croft, 1997 ; Croft et al., 1997 ; Amato Neto, 1999 ; Croft,
1999; Stoppani, 1999 ; Urbina, 1999a, 1999b ; Colley, 2000 ; Augustyns et al., 2001 ;
Docampo, 2001 ; Opperdoes et Michels, 2001 ; Cerecetto et Gonzalez, 2002 ; Gelb et
Hol, 2002 ; Gilbert, 2002; Rodrigues Coura et de Castro, 2002 ; Urbina, 2002).
Elles peuvent se répartir selon deux grandes catégories: les molécules naturelles et les
molécules synthétiques comme le Nifurtimox et le Benznidazole. Dans chacune de ces
catégories, certaines molécules sont actives sur des modèles expérimentaux et des
applications cliniques sont envisageables.
Une autre approche thérapeutique est à l'étude: l'immunothérapie à partir d'inhibiteurs
de la TcPA45 ou TcPRAG (proline racémase, Reina-San Martin et al., 2000), laquelle
est impliquée dans l'activation polyclonale de la réponse immune de l'hôte (Minoprio,
2001 ; Chamond et al., 2002,2003).
1.1.7 Contrôle
En 1991, les pays du Cône Sud d'Amérique latine (Argentine, Bolivie, Brésil, Chili,
Paraguay et Uruguay), puis en 1997 les pays Andins (Colombie, Equateur, Pérou et
Venezuela) et les pays d'Amérique Centrale (Belize, Costa Rica, El Salvador,
Guatemala, Honduras, Nicaragua et Panama) ont pris l'initiative de contrôler la
12

Généralités
transmission vectorielle de la maladie de Chagas en pulvérisant des insecticides dans
les maisons.
Cet objectif devrait être atteint d'ici à 2010. Néanmoins, la planification et la mise en
œuvre d'un programme international de lutte anti-vectorielle soulèvent un certain
nombre de problèmes d'éthique et de sécurité liés à chaque pays, particulièrement pour
ce qui concerne l'amélioration des conditions de vie et d'habitat des populations
concernées.
Les programmes de contrôle de vecteurs sont souvent coûteux pour les possibilités
économiques des pays touchés. En effet, l'éradication des vecteurs représente
seulement une partie du problème, car la prévention seule n'a aucun impact sur les
personnes déjà infectées. Un autre problème vient de la réinfestation de zones traitées
par des vecteurs sylvestres (Matias et al., 2003).
Les programmes de lutte anti-vectorielle devront être revus pour juger de leur efficacité.
De plus, ils doivent être complétés par le développement de nouvelles méthodes de
détection précoce ainsi que par le développement de nouveaux médicaments moins
toxiques et plus efficaces permettant de traiter des patients atteints de toutes les formes
cliniques (cardiopathie, viscéropathie et neuropathie chagasiques) et à tous les stades
de la maladie. De plus, les pays des zones endémiques devront continuer les
programmes de contrôle des vecteurs, y associer des activités de surveillance de la
transfusion sanguine, surveillance de la transmission congénitale de la maladie de
Chagas et enfin mettre en œuvre tout un programme de lutte contre la maladie elle
même (WHO, 2002).
1.2 STRUCTURE DU GENOME ET EXPRESSION GENETIQUE DE T. cruz;
1.2.1 Taille du génome de T. cruz;
Une caractéristique intéressante du parasite est la taille de son génome, qui peut varier
entre les différentes souches et même entre divers clones de la même souche
(McDaniel et Dvorak, 1993). Par exemple, la quantité totale d'ADN est estimée entre
125 et 280 fg par cellule pour différentes souches de T. cruzi. Elle inclue l'ADN du
kinétoplaste qui constitue de 16 à 30% du total de l'ADN génomique. Sans l'ADN
13

Généralités
kinétoplastique, la taille du génome est d'environ 100 à 200 Mb et la taille moyenne
d'un chromosome serait de 1,2 IVIb (Henriksson et al., 1996).
Des différences dans la taille et le nombre des chromosomes ou des changements
dans la ploïdie pour un ou plusieurs chromosomes (aneuploïdie) seraient responsables
de la variabilité du contenu d'ADN de T. cruzi (Henriksson et al., 1996 ; Gaunt et al.,
2003).
1.2.2 Organisation du génome de T. cruz;
L'organisation génétique du parasite comprend deux structures majeures: les
chromosomes nucléaires et le kinétoplaste.
Chez T. cruzi, les chromosomes ont une très faible condensation pendant la mitose et
ne peuvent pas être visualisés correctement, ce qui rend difficile l'étude directe du
caryotype (Vickerman et Preston, 1970 ; Belli, 2000). Mais, grâce à la technique de
séparation des chromosomes par électrophorèse en champs pulsés (Pulsed Field Gel
Electrophoresis, PFGE) le nombre total de chromosomes a pu être estimé. Il y aurait
environ 40 paires de chromosomes homologues, ce qui suggère que T. cruzi est une
espèce diploïde (Henriksson et al., 1995), soit 80 chromosomes au total. En fait, des
résultats expérimentaux récents conduisent à penser que T. cruzi est aneuploïde
(Gauntetal, 2003) _
Le kinétoplaste est un organite situé à la base du l1agelle (Henriksson, 1995). Il est
constitué par deux types d'ADN circulaire, les minicercles et les maxicercles,
concaténés entre eux en un réseau d'ADN unique qui a la forme d'un disque (Pérez
Morga et Englund, 1993 ; Zavala-Castro et al., 2002). Le nombre de minicercles est de
5 000 à 10 000 et le nombre de maxicercles est de 25 à 50 par kinétoplaste (Schenider,
2001 ).
Les minicercles sont organisés en quatre régions hypervariables séparées par quatre
régions constantes situées à intervalles réguliers de 90° autour du cercle (Degrave et
al., 1988). Les régions variables des minicercles ont été décrites comme étant les sites
de codage des ARNs guide (ARNg) impliqués dans l'édition des gènes des maxicercles
14

Généralités
(Sturm et Simpson, 1990 ; Avila et Simpson, 1995). Pour les régions conservées
aucune fonction spécifique n'a été décrite jusqu'à présent.
Les maxicercles sont les homologues fonctionnels des ADN mitochondriaux présents
chez les autres eucaryotes et codent pour l'ARN ribosomal et pour plusieurs enzymes
mitochondriales (Feagin, 2000). Chez L. tarentolae, chez C. fasciculata et chez T.
brucei, les maxicercles codent aussi pour des ARNg (Blum et al., 1990 ; van der Spek
et al., 1991 ; Benne, 1994).
En dehors des différentes familles d'ADN, chromosomique et kinétoplastique, il existe
aussi les familles habituelles d'ARN : les ARN messagers (ARNm), les ARN de transfert
(ARNt) et les ARN ribosomiques (ARNr).
Chez .Ies trypanosomes, tous les ARNm possèdent un miniexon en 5' ajouté par
épissage en trans et qui semble être important pour l'initiation de la traduction des
ARNm. Les unités transcriptionnelles sont organisées sous une forme polycistronique,
ressemblant aux opérons bactériens. L'ajout en 5' de ce miniexon sur chacun des
cistrons permet la formation de messagers individuels monocistroniques à partir d'un
précurseur plus long et à l'origine, polycistronique (voir chapitre 1.2.5.1 ).
1.2.3 Composition du génome de T. cruz; en séquences répétées
" existe différents types d'ADN répétés divisés principalement en deux classes, ceux
qui sont organisés en tandem et ceux qui sont dispersés dans le génome.
L'ADN répété en tandem a été décrit chez les eucaryotes mais il est pratiquement
inconnu chez les procaryotes. Il est divisé en ADN satellite, ADN minisateliite et ADN
microsatellite.
Parmi les ADN répétés, il existe deux éléments d'ADN répété avec un grand nombre de
copies, l'ADN satellite de 196 pb (Sloof et al., 1983) et le E13 répété (Requena et al.,
1992) qui forment respectivement 9% et 7% du génome du parasite.
D'autres éléments d'ADN répété d'abondance variable comme E12, E22, SIRE (Short
Interspersed Repeated Elements), RLE (Retroposon-Like Element), SRE (Short
15

Généralités
Repeated Elements), SREI-3 (Spacer Repetitive Elements), DGF ont été également
décrits chez T. cruz; (De Mendonça-Lima et Traub-Cseko, 1991 ; Requena et al., 1994,
1996 ; Vazquez et al., 1999, 2000), ainsi que d'autres transposons et éléments
similaires (Villanueva et al., 1991 ; Bhattacharya et al., 2002 ; Martin et al.,1995 ;
Verbisck et al., 2003).
1.2.4 Gènes et familles de gènes
Plusieurs gènes de T. cruz; sont présents en un grand nombre de copies. Certains
gènes de ménage ou « housekeeping genes» sont organisés en tandem avec un
nombre variable de copies, par exemple, les gènes qui codent pour les protéines de
choc thermique, les histones (H2A, H2B, H3), le miniexon, les protéines de Iigation au
Ca2+, la tyrosine aminotransférase, la cruzipaïne, le grp78 entre autres (Henriksson et
al., 1996). En général, ces gènes sont organisés en deux conglomérés apparemment
localisés dans les chromosomes homologues putatifs, à l'exception de la cruzipaïne,
pour laquelle les gènes sont organisés en quatre conglomères localisés dans
différentes bandes chromosomiques (Campetella et al., 1992).
En général, chez T.cruz; il y a des gènes présentant une seule copie, des gènes multi
copies organisés en tandem ainsi que des superfamilles de gènes (Agüero et al., 2000),
par exemple, la superfamille des transialidase-neuraminidases contient plusieurs sous
familles de gènes: SAPA, Tc13, gp85, CEA 160, FL160 et SA85-1.1 (Jazin et al., 1995;
Weston et al., 1999 ; Frash, 2000 ; Millar et Kahn, 2000).
1.2.5 Expression des gènes
A la différence des eucaryotes supérieurs, chez les trypanosomatidés la transcription
est unique pour divers aspects (Teixeira, 1998 ; Donelson et al., 1999). Tous les gènes
qui codent pour des protéines sont organisés en unités de transcription polycistroniques
(Graham, 1995). Les séquences codantes successives ne sont pas séparées par des
introns. Chaque transcrit est découpé en ARNm individuels par épissage en trans, à la
différence des eucaryotes dans lesquels l'épissage est réalisé en Gis. La régulation de
l'expression génétique se fait majoritairement au niveau post-transcriptionnel. Un des
mécanismes post-transcriptionnels découvert chez les trypanosomes est l'édition des
ARNm mitochondriaux (Benne et al., 1986).
16

Généralités
1.2.5.1 Transcription
A partir de l'initiation de la transcription en 5', chaque transcrit produit est une longue
molécule d'ARN (ARNm polycistronique précurseur ou pré-ARNm). A partir du
processus d'épissage en trans sont ajoutés les codons initiaux (coiffe miniexon ou
splice-Ieader en 5') et la queue poly (A) en 3' jusque là absents (Matthews et al., 1994).
Chez les trypanosomatidés, une séquence leader (SL) ou miniexon (Gibson et al.,
2000), possédant une structure de coiffe modifiée, est ajoutée à l'extrémité 5' des
ARNm précurseurs (Ullu et al., 1996 ; Teixeira, 1998). Ainsi, tous les ARNm de T. cruzi
portent en leur extrémité 5' les 39 nucléotides du miniexon (De Lange et al., 1984 ;
Gonzalez et al., 1985; Agabian, 1990 ; Perry et Agabian, 1991 ; Ullu et al., 1996).
Dans les cellules eucaryotiques, tous les ARNm à l'exception de ceux codant pour les
histones, possèdent une queue poly (A) à leur extrémité 3'. La queue poly (A) n'est pas
codée dans le génome et elle est produite de façon post-transcriptionnelle par un
clivage endonucléolytique au site de polyadénylation.
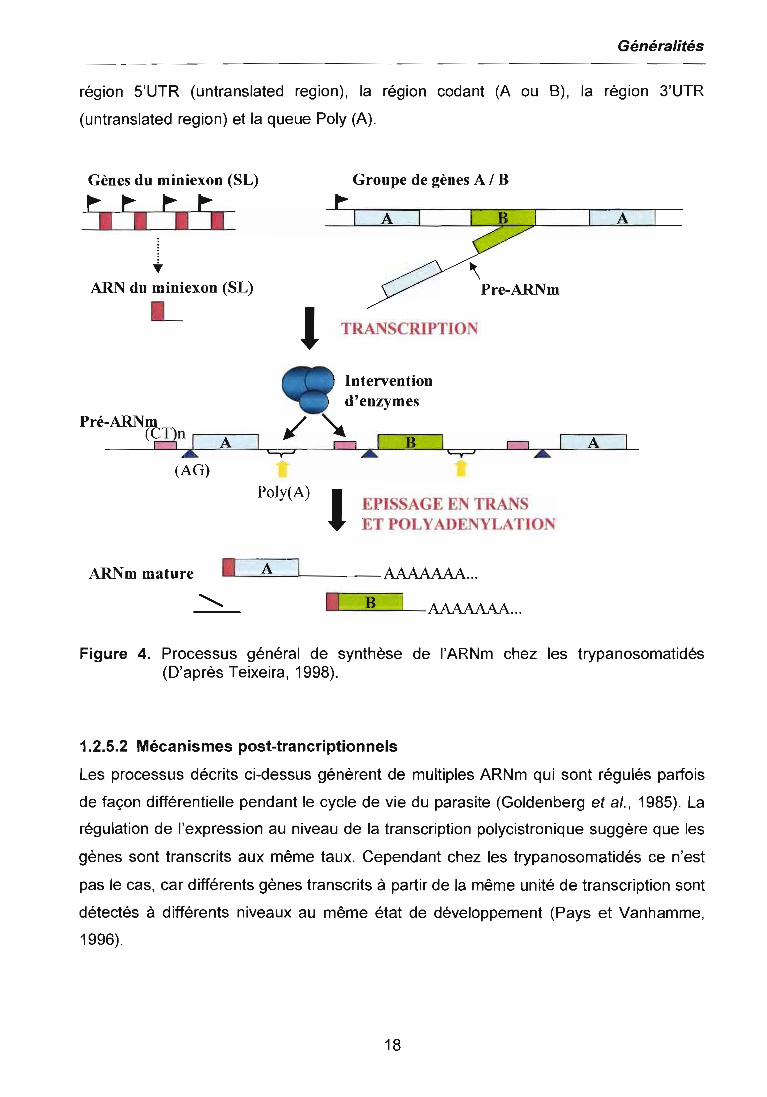
La figure 4 représente le processus général de synthèse de l'ARNm chez les
trypanosomatidés. A partir de deux exons qui proviennent de deux unités
transcriptionnelles différentes (généralement contenant des gènes en tandem), l'un des
exons représentant la partie 5' du futur ARI\lm mature (en rouge), est ajouté à un autre
exon (A ou B).
Dans le premier groupe de gènes en tandem, le gène A (en gris) est présent en deux
copies tandis que le gène B (en vert) est présent en une seule copie. Dans le deuxième
groupe (localisé dans un autre chromosome), il y a plusieurs copies en tandem du gène
qui code pour le miniexon ou "spliced leader" (SL). Pour le premier groupe de gènes, il
n'y a qu'un seul promoteur (drapeau noir) alors que pour le deuxième groupe, chaque
copie peut porter un promoteur.
Une fois transcrits, les pré-ARNm sont découpés par épissage en trans et polyadenilés.
Ces deux processus couplés ont besoin de signaux qui sont présents dans les régions
intergéniques: une séquence riche en polypyrimidine ou (CT)n (carrés roses), le site
d'addition du miniexon ou AG (triangles violets) et le site d'adénilation ou Poly (A)
(flèches jaunes). Enfin, les ARNm matures ont la séquence du miniexon (en rouge), la
17
Généralités
région 5'UTR (untranslated region), la région codant (A ou B), la région 3'UTR
(untranslated region) et la queue Poly (A).
Gènes du miniexon (SL)
...ARN du miniexon (SL)
L
Groupe de gènes A / B
A
Interventiond'enzymes
Pré-ARNm.. / '"(@~C==~A=~-,---I=~,..---_--L.~r=J--'--~I~B:.....---'-,I'--'-r---J.r=J--L~I_"-,,,A_L-
~ =r=' ~ =r=' ...(AG)
Poly(A) 1 EPI AGE EN TRAN
ET POLYADENVLATION
ARNm matu re l'----=-A-=------'- _
--=---.L-AAAAAAA...
Figure 4. Processus général de synthèse de l'ARNm chez les trypanosomatidés(D'après Teixeira, 1998).
1.2.5.2 Mécanismes post-trancriptionnels
Les processus décrits ci-dessus génèrent de multiples ARNm qui sont régulés parfois
de façon différentielle pendant le cycle de vie du parasite (Goldenberg et al., 1985). La
régulation de l'expression au niveau de la transcription polycistronique suggère que les
gènes sont transcrits aux même taux. Cependant chez les trypanosomatidés ce n'est
pas le cas, car différents gènes transcrits à partir de la même unité de transcription sont
détectés à différents niveaux au même état de développement (Pays et Vanhamme,
1996).
18

Généralités
Le contrôle de la transcription se fait au niveau post-transcriptionnel, notamment au
niveau de l'épissage, de la polyadénylation, de la stabilité de l'ARN et au niveau de
l'élongation de la transcription (Matthews et al., 1994 ; Furger et al., 1997 ; Abuin et al.,
1999; Teixeira et DaRocha, 2003).
Chez les trypanosomatidés, différents modèles ont été sélectionnés pour l'étude des
mécanismes trancriptionnels et post-transcriptionnels qui contrôlent l'expression des
gènes. Par exemple, chez T. brucei, ont été utilisés comme modèle les gènes qui
codent pour des variants des glycoprotéines de surface VSG (Variant Surface
Glycoprotein) (revue, Vanhamme et Pays, 1995 ; Roditi, 1996) ainsi que les gènes qui
codent pour les PARP (Procyclic Acidic Repetitive Protein) (Graham, 1995).
Chez Trypanosoma cruzi, les gènes qui codent pour la tubuline (Ürményi et al., 1992),
pour l'amastine (Teixeira et al., 1994), et pour l'histone (Recinos et al., 2001) ont été
utilisés comme modèle pour étudier la régulation stade-spécifique de l'expression
génétique.
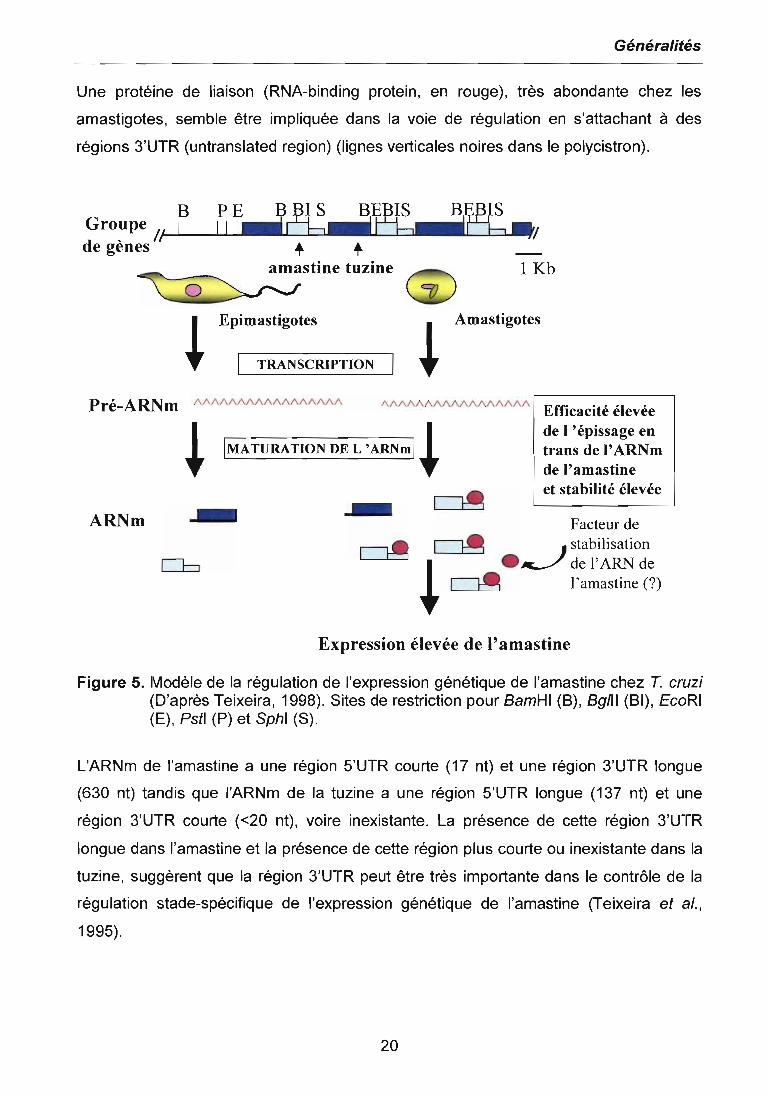
La figure 5 présente le modèle de la régulation de l'expression génique de l'amastine
chez T. cruzi. Le génome de T. cruzi contient plusieurs copies (8 à 23 copies, pour le
stock Tuhaluén) organisées en tandem du gène qui code pour l'amastine (en gris),
protéine très abondante à la surface des formes amastigotes. Ces gènes sont intercalés
avec d'autres gènes qui codent pour une autre protéine nommée tuzine (en bleue),
laquelle est moins abondante chez les amastigotes et a une fonction encore inconnue
(Teixeira etaI., 1994, 1999).
Le taux de transcription polycistronique est similaire chez les épimastigotes et chez les
amastigotes. Ce processus produit des pré-ARNm (en ligne ondulée rouge) qui doivent
être modifiés pour produire des ARNm matures qui codent pour les protéines amastine
et tuzine.
L'accumulation différentielle de ces deux protéines, amastine et tuzine, dans les deux
formes cellulaires du parasite dépendrait de l'efficacité de l'épissage en trans et de la
stabilité des ARNm dans les amastigotes.
19
Généralités
Une protéine de liaison (RNA-binding protein, en rouge), très abondante chez les
amastigotes, semble être impliquée dans la voie de régulation en s'attachant à des
régions 3'UTR (untranslated region) (lignes verticales noires dans le polycistron).
1 Kb
~ Amastigotes
... ...amastine tuzine
1 TRANSCRIPTION
Epimastigotes
Grou pe 1.j-l_-lJ......I.f.L::1:••i..L=1=••lr:=1=.de gènes
Facteur de
Efficacité élevéede l 'épissage entraos de l'ARNmde l'amastineet stabilité élevée
~
~ 1 stabilisation1 .~ del'ARNde..~ l'amastine (?)
/\/\/\/\/\/\/\/\1\/\1\/\/\1\/\/\/\
IMATURATION DE L 'ARNml~
ARNm
Pré-ARNm 1\1\1\I\I\/\/\/\/\/\I\/\/\/VV\/\
Expression élevée de l'amastine
Figure 5. Modèle de la régulation de l'expression génétique de l'amastine chez T. cruzi(D'après Teixeira, 1998). Sites de restriction pour BamHI (B), Bg/lI (BI), EcoRI(E), Pstl (P) et Sphl (S).
L'ARNm de l'amastine a une région 5'UTR courte (17 nt) et une région 3'UTR longue
(630 nt) tandis que l'ARNm de la tuzine a une région 5'UTR longue (137 nt) et une
région 3'UTR courte «20 nt), voire inexistante. La présence de cette région 3'UTR
longue dans l'amastine et la présence de cette région plus courte ou inexistante dans la
tuzine, suggèrent que la région 3'UTR peut être très importante dans le contrôle de la
régulation stade-spécifique de l'expression génétique de l'amastine (Teixeira et al.,
1995).
20

Généralités
Bien que les gènes de l'amastine et de la tuzine soient transcrits par la même
polymérase et dans le même pré-ARNm polycistronique, les ARNm matures de la
tuzine seront accumulés à la même concentration chez les deux formes parasitaires
(Le. épimastigotes et amastigotes) tandis que ceux de l'amastine seront faiblement
entassés chez les formes épimastigotes (Teixeira et al., 1995 ; Coughlin et al., 2000).
1.2.5.3 L'édition des ARNm du kinétoplaste
Un autre mécanisme post-transcriptionel important chez les trypanosomes est le
mécanisme d'édition des ARNm chez le kinétoplaste. Ce mécanisme d'expression du
génome mitochondrial a été étudié chez différents parasites, tels que T. brucei, T. cruzi,
L. tarentolae et C. fasciculata.
Les transcripts du maxicercle peuvent être modifiés par l'insertion, et en degré moindre,
par la suppression d'uridines (revue, Arts et Benne, 1996 ; Simpson, 1997 ; Stuart et al.,
1997 ; Estévez et Simpson, 1999 ; Schneider, 2001).
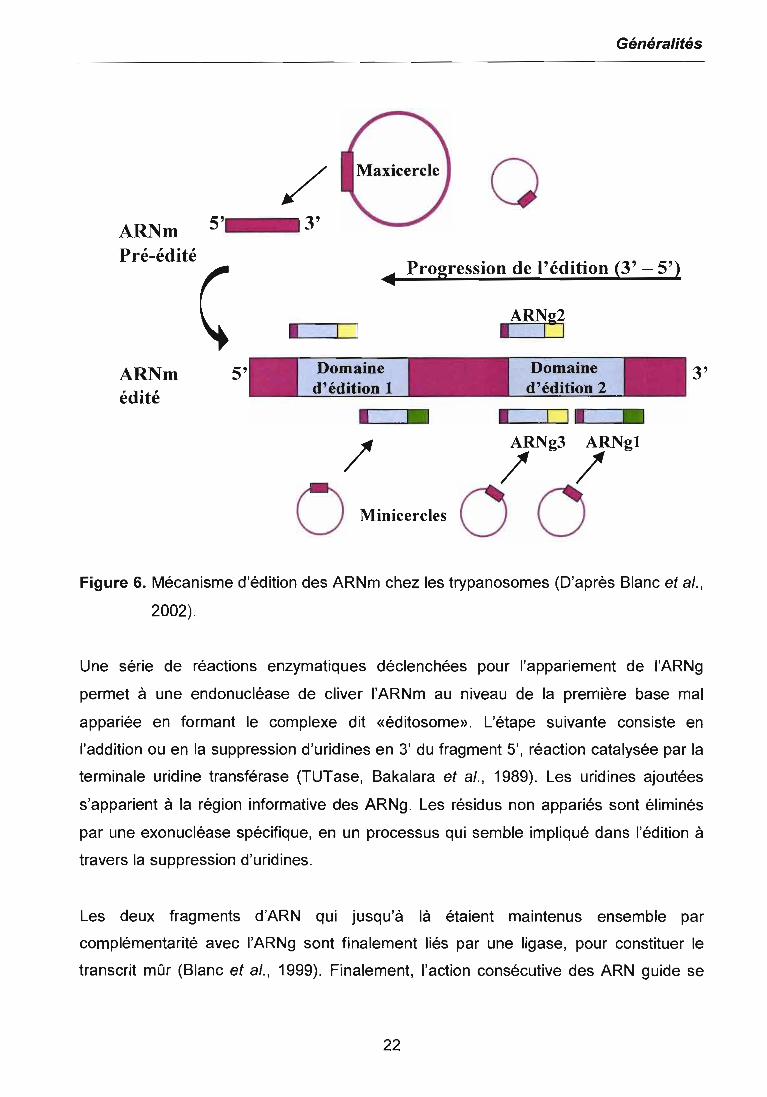
La figure 6 présente le mécanisme général d'édition des ARNm chez les trypanosomes.
Ce processus résulte d'une parfaite collaboration entre les gènes contenus dans les
maxicercles et les minicercles, les premiers apportant les ARN pré-édités et les
seconds apportant la majorité des ARN guides (ARNg). Les deux domaines d'édition
sur l'ARNm sont représentés en bleu, les régions non éditées, en mauve. Les ARNg ont
une séquence d'ancrage (en vert). Au fur et à mesure que l'ARNm est édité, une
nouvelle région d'ancrage est créée pour un deuxième ARNg (en jaune), ce qui fait que
l'édition opère avec une polarité 3'-5'.
L'édition des ARI\lm doit être un processus très précis, car l'addition ou l'insertion d'une
uridine pourrait changer le cadre de lecture provoquant une traduction incorrecte ou un
arrêt définitif. La clé de cette précision réside dans les ARN guides (Blum et al., 1990).
Ils possèdent une séquence complémentaire de la région éditée qui détermine le
nombre précis d'uridines à ajouter ou à supprimer. De plus, leur région 5' (séquence
d'ancrage) s'apparie au transcrit pré-édité tandis que leur région 3' (queue polyU)
semble être impliquée dans la stabilisation du complexe ARNm-ARNg. Ainsi, l'ARNg
interagit avec l'ARNm pré-édité en aval du site d'édition plus proche de l'extrémité 3' du
messager, via la séquence d'ancrage (en vert).
21
Généralités
... Progression de l'édition (3' - 5')
o/5'••_3'ARNm
Pré-édité
ARNm 5' Domaine Domaine 3'édité d'édition 1 d'édition 2
• • 1 1
l' ARNg3 ARNgl
l' l'0 Minicercles 0 0
Figure 6. Mécanisme d'édition des ARNm chez les trypanosomes (D'après Blanc et al.,
2002).
Une série de réactions enzymatiques déclenchées pour l'appariement de l'ARNg
permet à une endonucléase de cliver l'ARNm au niveau de la première base mal
appariée en formant le complexe dit «éditosome». L'étape suivante consiste en
l'addition ou en la suppression d'uridines en 3' du fragment 5', réaction catalysée par la
terminale uridine transférase (TUTase, Bakalara et al., 1989). Les uridines ajoutées
s'apparient à la région informative des ARNg. Les résidus non appariés sont éliminés
par une exonucléase spécifique, en un processus qui semble impliqué dans l'édition à
travers la suppression d'uridines.
Les deux fragments d'ARN qui jusqu'à là étaient maintenus ensemble par
complémentarité avec l'ARNg sont finalement liés par une ligase, pour constituer le
transcrit mûr (Blanc et al., 1999). Finalement, l'action consécutive des ARN guide se
22

Généralités
répète jusqu'à ce que l'ARNm soit complètement édité (revue, Blanc et al., 2002 ;
Simpson et al., 2003).
Le mécanisme d'édition des ARNm est post-transcriptionel et il est indispensable à la
production d'ARNm pour les unités Il et III de la cytochrome oxidase (COli et COIII,
respectivement), pour le cytochrome b (CYb) et pour la sous-unité six de l'ATPase
(ATPase6) (Bhat et al., 1991 ; Kim et al., 1994; Feagin, 2000) entre autres.
1.3 COMPORTEMENT DES PARASITES FACE AUX MEDICAMENTS
L'étude et la compréhension du comportement des parasites vis-à-vis des différents
médicaments sont primordiales pour développer une chimiothérapie plus spécifique et
mieux adaptée au niveau de sensibilité naturelle des souches. Compte tenu des liens
possibles entre sensibilité des clones et efficacité thérapeutique, il apparaît important de
pouvoir contrôler le suivi thérapeutique chez un même patient.
"D'autre part, la présence de parasites naturellement résistants ne doit être pas
négligée. La compréhension des mécanismes de résistance des parasites aux
médicaments devrait permettre la planification de nouvelles stratégies thérapeutiques
qui éviteraient l'apparition de résistances et la mise au point de nouvelles cibles
thérapeutiques. De plus, cette connaissance permettrait d'améliorer la chimiothérapie
existante par une utilisation mieux adaptée aux souches traitées.
1.3.1 Sensibilité des parasites aux médicaments (Benznidazole et Nifurtimox)
Le traitement des patients atteints de la maladie de Chagas par les médicaments
classiques (Benznidazole et Nifurtimox) engendre certaines variations dans la réponse
des patients (Stoppani, 1999). De plus, l'action de ces deux médicaments varie en
fonction de la sensibilité des parasites à ces molécules. Cette varia bilité de sensibilité
des différentes souches de T. cruzi a été décrite in vivo et in vitro par Hauschka (1949),
Bock et al. (1969), Habernock et Gonnert (1972), Andrade et al. (1975), Brener et al.
(1976), Filardi et Brener (1982), Andrade et al. (1985), Filardi et Brener (1987), Neal et
Van Bueren (1988), Revollo (1995), Toledo etai. (1997) et Revollo etai. (1998).
23

Généralités
La .sensibilité aux agents thérapeutiques est donc extrêmement inégale parmi les
souches de T. cruzi, dont un grand nombre sont naturellement résistantes aux
médicaments utilisés contre la maladie de Chagas (Haberkorn et Gonnert, 1972 ;
Brener et al., 1976 ; Filardi et Brener, 1987 ; Toledo et al., 1997). Par exemple, la
souche Colombiana a été décrite comme résistante aux nitrofuranes et aux dérivés 2
nitroimidazole (Andrade et Figueira, 1977 ; Andrade et al., 1977 ; Filardi et Brener,
1982).
La résistance induite par des médicaments ou chimiorésistance induite peut être
provoquée par un traitement prolongé (Andrade et al., 1977) et reste stable pendant
plusieurs passages successifs. " a été également démontré expérimentalement la
possibilité que se produise une résistance croisée (Pontes et Andrade, 1984). Il est
intéressant de noter que la chimiorésistance naturelle a été détectée dans un grand
nombre de souches isolées d'animaux réservoirs et de vecteurs sauvages dans
lesquels il n'y a pas eu de contact préalable avec des médicaments.
Un autre point important concerne l'origine géographique des souches': En effet, des
différences de niveau de sensibilité aux médicaments ont été retrouvées parmi diverses
souches qui provenaient de différents lieux géographiques (Filardi et Brener, 1987).
La résistance naturelle des souches aux dérivés nitrés est un facteur qui a d'ailleurs été
proposé pour expliquer les faibles taux d'efficacité thérapeutique chez certains patients
chagasiques (Filardi et Brener, 1987 ; Murta et al., 1998). Une autre explication possible
serait la faculté de ces souches à s'évader de la réponse par l'IL-2 et l'IFN-y pendant la
phase aiguë de la maladie (Muller et al., 1986 ; Michailowsky et al., 1998).
Les souches de T. cruzi ont été divisées en trois groupes sur la base de la sensibilité ou
de la résistance in vivo au Benznidazole : i) résistants, ii) partiellement résistants, et iii)
sensibles au traitement. Par exemple, les souches Colombiana, y et CL, correspondent
respectivement à cette classification (Filardi et Brener, 1987).
Les différentes réponses médicamenteuses des souches de T. cruzi ont suggéré
l'existence d'une diversité moléculaire entre les souches. Cette diversité a été étudiée
par les techniques isoenzymatiques (Andrade et al., 1983, 1992) mais aussi par
24

Généralités
amplification de fragments d'ADN par amorces aléatoires (RAPD), par sondes géniques
pour l'analyse de la P-glycoprotéine (TcPGP), la hypoxanthine-guanine phosphoribosyl
transférase (HGPRT), l'ARNr (ADNr) et le miniexon (MEX) (Murta et al., 1998).
Les RAPD et les isoenzymes ont montré une division des souches de T. cruzi en deux
groupes principaux, T. cruzi 1 et T. cruzi Il (Momen, 1999). De même, les sondes
géniques les ont divisées en deux groupes distincts. Pour l'ADNr et pour le MEX, cette
division est indépendante du phénotype de résistance, ce qui indiquerait qu'il n'existe
pas de relation évidente entre ces deux marqueurs et la sensibilité aux médicaments
(Murta et al., 1998). Il Y a une forte corrélation entre profils isoenzymatiques et RAPD
(Steindel et al., 1993; Tibayrenc et al., 1993 ; Barnabé et al., 2000 ; Brisse et al., 2000),
résultat corroborant l'hypothèse d'évolution c10nale prépondérante chez T. cruzi
(Tibayrenc et al., 1986).
La sensibilité des parasites aux médicaments est davantage en relation avec le
zymodème qu'avec l'origine géographique des souches (Murta et al., 1998).
Etant donné que le parasite T. cruzi montre une grande diversité biologique (Andrade,
1976 ; Melo et Brener, 1978 ; Dvorak et al., 1980 ; Garcia et Dvorak, 1982 ; Andrade et
al., 1983 ; Dvorak, 1984 ; Andrade et al., 1985 ; Sanchez et al., 1990), il a été proposé
que cette diversité biologique peut être expliquée en partie par la diversité génétique du
parasite (Miles et al., 1981).
Le modèle d'évolution c10nale proposé pour T. cruzi (Tibayrenc et al., 1986 ; Tibayrenc
et Ayala, 1988) prévoit une évolution non indépendante entre diversité génétique et
propriétés biologiques des clones naturels du parasite, ce qui inclut la sensibilité aux
médicaments.
La corrélation entre diversité génétique et biologique a été établie par plusieurs auteurs
(Laurent et al., 1997 ; De Lana et al., 1998 ; Toledo et al., 2002). Une concordance
entre diversité génétique et sensibilité in vitro et in vivo des parasites aux médicaments
a également été montrée (Revollo et al., 1998 ; Toledo et al., 2000 ; Martinez-Diaz et
al., 2001 ; Toledo et al., 2003).
25

Généralités
Récemment, la variation de sensibilité in vivo au Benznidazole de clones dérivés de la
souche Colombiana et de la souche Berenice-78, respectivement résistante et sensible,
a été explorée. La souche Colombiana et les clones dérivés d'elle-même ont montré
des niveaux de résistance très similaires (Camandaroba et al., 1999 ; Veloso et al.,
2001 ; Camàndaroba et al., 2003), tandis que les clones dérivés de la souche Berenice
78 ont montré différents niveaux de sensibilité (Veloso et al., 2001). La persistance et le
niveau de résistance des clones doivent être prises en compte pour la prescription
médicamenteuse.
De manière surprenante, il existe très peu de travaux dans la littérature décrivant le
processus d'induction de résistance aux médicaments chez T. cruzi. Un tel modèle
permettrait pourtant de mieux comprendre les mécanismes moléculaires de la
résistance. Différents travaux ont permis d'obtenir des souches chimiorésistantes au
Nifurtimox (Lampit®) in vitro (Dvorak et Howe, 1977 ; Nozaki et al., 1996), et des
souches chimiorésistantes au Benznidazole in vitro (Nirdé et al., 1995) et in vivo dans
des souris (Murta et Romanha, 1998). Récemment, des souches chimiorésistantes aux
azolés (Fluconazole et Ketoconazole) ont été obtenues (Buckner et al., 1998). Mais
malheureusement jusqu'à maintenant, aucun de ces travaux n'a permis de comprendre
les mécanismes de résistance de T. cruzi aux médicaments.
1.3.2 Mécanismes de résistance chez d'autres Trypanosomatidés et autres
parasites
Dans l'ensemble du règne animal, les mécanismes connus de résistance aux
médicaments sont très nombreux et complexes (Edwards, 1993 ; Johnson, 1993 ;
Upcroft et Upcroft, 1993 ; Bolhuis et al., 1997 ; van Veen et Konings, 1997 ; Ouellette et
Kündig, 1997 ; van Veen et Konings, 1998 ; White et al., 1998 ; Le Bras, 1999 ; Wolfger
et al., 2001 ; Teodori et al., 2002).
Les principaux mécanismes généraux impliqués dans la résistance aux médicaments
sont :
i) le changement dans les propriétés des membranes cellulaires qui provoque une
variation du flux de molécules par altération des transporteurs,
ii) l'inactivation du médicament,
26

Généralités
iii) des modifications quantitatives et qualitatives des cibles thérapeutiques au travers
de mutation, de surexpression, d'amplification génique ou de la recombinaison
mitotique (Flintoff et Essani, 1980 ; Endicott et Ling, 1989 ; Coons et al., 1990;
Katakura et al., 1991 ; White et al., 1998).
Mécanismes de chimiorésistance chez les Leishmania
Des études sur des souches de Leishmania (Duel/eUe et al. ,1990; Duel/ette et
Papadopoulou, 1993; revue, UI/man, 1995) et Plasmodium (revue, UI/man, 1995), chez
qui la résistance a été induite par pression médicamenteuse, ont montré que la
résistance était associée à la présence de gènes de multirésistance (MDR) qui codent
pour des P-glycoprotéines de membrane (PGP) impliquées dans la régulation du flux
des molécules.
CeUe multirésistance aux médicaments a également été observée dans des lignées de
cellules tumorales, ainsi que dans des pathologies parasitaires produites par
Entamoeba histolytica et par Trichomonas vaginalis (revue, Ullman, 1995).
Dans un autre modèle parasitaire, les Leishmania, le développement de modèles in
vitro a permis l'étude des mécanismes de résistance, en utilisant des mutants résistants
aux dérivés pentavalents de l'Antimoine, comme le Stibogluconate de sodium
(Pentostam®, Grogl et al., 1989) et le Meglumine antimoniate (Glucantime®, Ferreira
Pinto et al., 1996) ainsi que des mutants aux antifolates (Méthotrexate), à la
Pentamidine (Serena et al., 1997) et à l'Arsénite de sodium (Dey et al., 1994),
médicaments couramment utilisés en clinique humaine comme vétérinaire.
Certaines résistances chez des parasites pourraient donc être expliquées par la
modulation du transport trans-membranaire des molécules toxiques. Chez Leishmania,
deux classes de gènes pgp ont été décrites (revue, Ullman, 1995 ; Ross et Sutherland,
1997).
La première classe correspond aux pgpA : ItpgpA et ImpgpA, de L. tarentolae et L.
major respectivement, qui sont amplifiés après sélection des parasites par des
médicaments hydrophiles (Ouel/ette et al., 1990).
27

Généralités
Ces gènes ont été caractérisés et identifiés comme étant présents dans un ADN
circulaire extra-chromosomique (Garvey et Santi, 1986) appelé cercle H. Le cercle H
confère la résistance pour une série de médicaments qui n'ont pas de relation entre
eux, Méthotrexate, Arsénite, Primaquine ou Terbinafine. Quatre autres gènes pgp ont
été également identifiés dans le cercle H chez L. tarentolae : ItpgpB, /tpgpC, ItpgpD et
ItpgpE, lesquels hybrident avec une sonde dérivée du gène ItpgpA (Gamarro et al.,
1994).
La surexpression de ItpgpE, dans un mutant de L. tropica sélectionné par le
Méthotrexate, est davantage associée au réarrangement du gène qu'à l'amplification du
gène. Le mutant possède ainsi un nouveau phénotype qui lui confère une résistance à
l'Arsénite, Vinblastine, Puromycine et Ooxorubicine, tandis que les gènes ItpgpB, /tpgpC
et ItpgpD ne sont pas associés à la sensibilité du parasite aux médicaments. D'autres
études montrent que d'autres gènes sont impliqués en plus des pgp. Dans le cercle H, il
Y a un autre gène additionnel, le gène Itdh, qui code pour une déhydrogénase chez L.
tarentolae ou le gène ptr1 qui code pour une ptéridine réductase (PTR1) chez L. major,
tous les deux impliqués dans la résistance au Méthotrexate (Bello et al., 1994).
La seconde classe de gènes pgp correspond aux gènes MOR qui confèrent un
phénotype similaire au MOR de cellules humaines. Ces gènes, Idmdr1 et lemdr1, ont
été amplifiés et surexprimés dans des lignées sélectionnées par la Vinblastine, chez L.
donovani (Henderson et al., 1992) et chez L. enriettii (Chow et al., 1993). Cette dernière
a de plus montré une résistance croisée à la Puromycine et aux anthracyclines.
Des essais de transfection ont montré que la résistance au Méthotrexate est contrôlée
en partie par la PTR1, une déshydrogénase réductase à chaîne courte (Papadopoulou
et al., 1992 ; Callahan et Beverley, 1992), tandis que la résistance à l'arsénite est sous
le contrôle du transporteur MOR: la PGPA (Callahan et Beverley, 1991 ; Papadopoulou
et al., 1994).
En général, au moins trois mécanismes différents sont associés à la résistance de
Leishmania au Méthotrexate :
i) l'amplification de la dhfr-thymidylate synthétase (dhfr-ts), qui fait partie du cercle
appelé cercle R (Beverley et al., 1984),
28

Généralités
ii) l'amplification de différent locus du cercle H, par exemple le gène Itdh, qui confère la
résistance aux antifolates (Papadopoulou et al., 1992), et
iii) la réduction de l'entrée du médicament (Ellenberger et Beverley, 1987a).
Récemment, différentes études ont montré chez des mutants de L. tarentolae
sélectionnés pour leur résistance à l'arsénite ou à l'antimonite, la présence d'une autre
pompe d'efflux non-pgpA (Dey et al., 1994; Papadopoulou et al., 1996) laquelle
reconnaît des métaux conjugués à divers thiols, incluant le glutathion (GSH) (Dey et al.,
1996) et la trypanothione (TSH) (Mukhopadhyay et al., 1996). Chez les mutants
résistants à l'arsénite, la TSH a été trouvée très augmentée, tandis que les niveaux de
cystéine et de GSH étaient moins élevés.
En résumé, Leishmania résiste à l'action toxique de médicaments à travers plusieurs
mécanismes tels que l'amplification de gènes cibles (dhfr-ts, nagt, impdh, odc), de
gènes qui codent pour des pompes putatives (ldmdr1 et ItpgpA) ou de gènes impliqués
dans des voies métaboliques alternes (ltdh) (revue, Ouellette et Papadopoulou, 1993).
Mais Leishmania est capable également de résister aux effets toxiques en diminuant
l'accumulation du médicament, par exemple pour le Méthotrexate (Ellenberger et
Beverley, 1987b; Kaur et al., 1988), l'arsénite et l'analogue de la formycine B
(Iovannisci et al., 1984).
Mécanismes de chimiorésistance chez les Trypanosomes africains
Les médicaments d'élection contre T. brucei rhodesiense et T. b. gambiense, agents
étiologiques de la maladie du sommeil (ou trypanosomose humaine africaine), sont
respectivement la suramine et la Pentamidine. Le Mélarsoprol et l'Eflornithine (DFMO)
sont également utilisés pour le traitement de la maladie du sommeil (Denise et Barrett,
2000).
Carter et Fairlamb (1993) ont montré que les souches de T. b. brucei au Mélarsoprol
ont deux transporteurs de nucléosides (P1 et P2), l'un des deux (P2) possède une
spécificité pour l'adénosine, l'adénine et le Mélarsoprol. Celui-ci est perdu ou modifié
lorsque le parasite devient résistant au melarsenate de sodium. Les parasites résistants
importent alors les purines à travers le transporteur restant (P1) qui a une spécificité
pour l'adénosine et l'inosine.
29

Généralités
Chez T. b. rhodesiense une résistance spontanée au DFMO a été rapportée dans
différents isolats cliniques qui n'ont pourtant pas eu de contact préalable avec le
médicament (Bachi, 1993 ; Bachi et al., 1993). Les isolats résistants contiennent moins
de S-adénosyl méthionine intracellulaire (AdoMet), AdoMet synthétase et AdoMet
décarboxylase que les trypanosomes sensibles (Ross et Sutherland, 1997).
Phillips et Wang (1987) ont décrit des mutants résistants à la DFMO, dans lesquels le
seul mécanisme qui explique la résistance observée est l'altération de l'accumulation du
médicament. De plus, il n'y a pas eu d'amplification génique qui permet la résistance en
augmentant la quantité de protéine cible (ODC, ornithine décarboxylase). Ces résultats
montrent également que les protéines transporteurs de DFMO sont perdues dans les
mutants résistants, et que probablement d'autres protéines sans affinité au DFMO sont
surexprimées, inhibant ainsi la diffusion simple de ce médicament en réduisant la
surface cellulaire disponible.
La principale étude chez T. congolense, parasite responsable d'une maladie du bétail,
montre que l'isometamidium radiomarqué C4C), est augmenté chez les parasites
sensibles (Sutherland et al., 1992), tandis que les parasites résistants ont été
caractérisés par une élimination rapide du médicament (Sutherland et Holmes, 1993).
Ainsi, l'accumulation du médicament diffère entre les parasites résistants et les
parasites sensibles à l'Isometamidium.
D'autres études ont montré que les parasites T. brucei résistants ont tendance à moins
absorber le médicament que les parasites sensibles (Frommel et Balber, 1987 ; Carter
et al., 1995). Cependant pour des clones de T. b. brucei résistants à la Pentamidine,
cette réduction de l'absorption du médicament n'a pas pu être démontrée (Berger et al.,
1995).
La réduction de l'absorption des médicaments est considérée comme le résultat d'une
importation réduite ou d'une exportation augmentée. La balance eflluxlinflux dans les
parasites résistants et sensibles est probablement due à la présence de divers
transporteurs. Une de ces classes de transporteurs sont les ABC transporteurs, qui
permettent le transport de substrats à travers les membranes cellulaires en utilisant
l'énergie de l'ATP. Récemment, chez T. brucei trois ABC transporteurs ont été décrits
30

Généralités
(Maser et Kamisky, 1998), mais leur possible rôle dans la résistance du parasite n'a pas
encore été démontré.
1.3.3 Gènes candidats potentiellement intéressants chez T. cruz;
Compte tenue des stratégies développées par d'autres modèles parasitaires
chimiorésistants, il m'est apparu logique de rechercher par analogie, des gènes
impliqués soit dans les processus de modification de la pénétration des médicaments
dans le parasite, soit des processus de détoxification cellulaire, soit enfin la mise en
place de mécanismes d'échappement cellulaire.
Dans ce cadre, il apparaissait opportun de nous intéresser à l'expression de gènes:
i) soit dans les mécanismes de détoxication des parasites (comme les gènes de la
superoxyde dismutase, SaD),
ii) soit dans les mécanismes de différenciation cellulaire (comme les gènes de la
cystéine protéase majeure),
Hi) ou encore dans le mécanisme de transport de métabolites à travers la membrane
cellulaire (comme les gène de la P-glycoprotéine).
1.3.3.1 La Superoxyde Dismutase
L'enzyme SOO participe aux processus de détoxication des cellules en catalysant la
réaction de transformation d'ion superoxyde formé au cours du transport des électrons
sur la chaîne respiratoire mitochondriale vers l'oxygène moléculaire. Etant donné que
l'action des médicaments utilisés pour le traitement de la maladie de Chagas est
réalisée à travers la production de radicaux libres de l'oxygène, nous proposons
d'étudier l'expression des gènes (tcsodA et tcsodB) correspondants aux deux SaD
décrites chez T. cruzi (FeSOOA et FeSODB), avec le but d'établir leur possible
participation à la détoxication aux radicaux libres produits lors du processus d'induction
de la résistance au Benznidazole.
L'enzyme superoxyde dismutase (SaD, EC1.15.1.1) est un composant important de
défense antioxydant tant chez les eukaryotes que chez les procaryotes. Sa fonction est
de détruire les radicaux libres (Le. anion superoxyde) qui sont produits normalement
dans les cellules et qui sont notamment toxiques pour les systèmes biologiques.
31

Généralités
Un radical libre est un atome ou une molécule qui contient un électron non apparié.
Cette espèce instable a tendance, soit à accepter un autre électron, soit à transférer cet
électron sur une autre molécule. Ce phénomène provoque une réaction d'oxydation en
chaîne qui peut causer l'oxydation définitive de molécules importantes telles que les
lipides, les phospholipides des membranes, les protéines et même les acides
nucléiques (Oocampo et Moreno, 1986 ; Marr et Oocampo, 1986 ; revue, Nordberg et
Arnér, 2001). Du point de vue clinique, les radicaux superoxyde auraient un rôle à jouer
dans le processus d'inflammation (Nathan, 2003), de cancérogenèse (Nagano et al.,
1989 ; revue, Hussain et al., 2003) et dans le vieillissement (revue, Sohal et
Weindruch, 1996 ; Beckman et Ames, 1998). Les radicaux libres pourraient participer
également aux processus physiopathologiques des maladies neurodégénératives
comme la sclérose latérale amyotrophique, la maladie de Parkinson et la maladie
d'Alzheimer entre autres (Beai, 1995 ; Jenner, 2003 ; Luchsinger et al., 2003).
Normalement, le radical superoxyde est produit au niveau des mitochondries, au cours
du transfert d'électrons dans la chaîne respiratoire mais également dans d'autres
réactions biochimiques participant à la genèse du stress oxydatif (e.g. mécanisme de
défense via les cellules immunitaires comme les monocytes-macrophages et les
polynucléaires neutrophiles lorsqu'ils sont activés pour détruire des micro-organismes,
des macromolécules étrangères ou des cellules en décomposition des tissus
nécrotiques qui ont été captés par endocytose).
Un certain nombre de médicaments, parmi lesquels ceux qui sont utilisés contre T. cruzi
(Benznidazole et Nifurtimox) produisent aussi des radicaux libres qui vont induire un
stress oxydatif chez le parasite et chez le patient (Oocampo et Moreno, 1986 ; Marr et
Oocampo, 1986; Tocher, 1997).1
La SaD est une métallo-enzyme comprenant soit du cuivre ou du zinc (SaD
cytosolique) soit du manganèse (SaD mitochondriale) ou encore du fer. Ces métaux
confèrent à l'enzyme son activité antioxydante.
32

Généralités
Fonction de la SOD
La SaD ne possède qu'une seule fonction catalytique connue: la dismutation du radical
superoxyde (a2--) en peroxyde d'hydrogène et en oxygène moléculaire selon la
réaction présentée (1) :
Chez les bactéries, ce mécanisme de détoxication est réalisé en deux étapes. Il
consiste en un transfert d'électron du radical superoxyde vers un accepteur final qui est
le peroxyde d'hydrogène. L'ion métallique constitue l'élément l'intermédiaire du
transfert. La SaD constitue donc une molécule tampon entreposant temporairement
l'électron indésirable en provenance du radical toxique (1.1) pour le redistribuer, dans
un deuxième temps (1.2), au peroxyde d'hydrogène qui lui, sera rapidement neutralisé
par les catalases de la cellule.
Classes de SOD
Il existe trois familles de SaD distinctes, qui sont distinguées par le type de groupe
prosthétique: Cu/Zn, Fe, Mn et Ni. Les Fe-SaD et Mn-SaD constituent une seule
famille structurelle (Bannister et al., 1987). Elles ont une structure d'homodimères ou
d'homotétramères.
La Fe-SaD est présente dans les cellules anaérobies strictes, dans les aérobies
facultatives (seule ou avec le Mn-SaD), dans le cytosol de cyanobacteries, dans les
chloroplastes des plantes et chez les protozoaires (Grace, 1990). Les Fe- et Mn-SaD
présentent une spécificité absolue pour le métal (Le. chez E. colI), tandis que chez
d'autres organismes (Le. P. shermaniJ) l'enzyme est active avec n'importe quel métal.
Chez l'homme, il existe la SaD Cu/Zn au niveau cytoplasmique et la SaD Mn/Fe au
niveau mitochondrial.
33

Généralités
Structure des SOD
La structure cristalline de la SOD a été résolue par Parker et al. (1987). Les Fe-SOD et
Mn-SOD bactériennes sont composées de deux domaines avec 6 hélices a majeures et
3 brins b-antiparallèles. Le site actif de ces métallo-protéines est situé à l'interface des
deux sous-unités entre les deux domaines.
Le cofacteur métallique (fer ou manganèse) est lié dans le site actif par 5 ligands, par
exemple, trois résidus histidine et un résidu aspartate d'une même sous-unité et par
une molécule en provenance du solvant (Beyer et al., 1991).
Les SOD chez T. cruz;
Les médicaments utilisés normalement contre les parasites induisent le stress oxydatif
par la formation de radicaux nitrés qui vont générer des radicaux libres de l'oxygène. Le
parasite T. cruzi est également exposé aux intermédiaires toxiques qui sont générés
par le système immunitaire, et qui conduisent à la production d'oxyde nitrique (NO') par
la NO synthétase (Vespa et al., 1994; Saeftel et al., 2001). L'oxyde nitrique, en
combinaison avec l'anion superoxyde va produire le peroxynitrite, un oxydant puissant,
qui tue T. cruzi de manière dose dépendante (Denicola et al., 1993).
T. cruzi possède deux types de défenses antioxydantes, celles faisant intervenir les
défenses enzymatiques et celles faisant intervenir les défenses non enzymatiques.
L'activité SaD chez T. cruzi a été décrite pour la première fois par Boveris et Stoppani
(1977). Depuis, deux gènes homologues aux Fe-SOD ont été isolés chez ce parasite
(Temperton etai., 1996; Ismail etai., 1997).
A partir de l'analyse comparative des séquences en acides aminés chez d'autres
organismes, il a été suggéré que les gènes de la SOD de T. cruzi étaient plutôt en
relation avec des Fe-SOD et Mn-SaD. Ceux deux types d'enzymes peuvent se
distinguer selon les substitutions localisées dans le site actif ou à sa proximité.
Deux Fe-SaD distinctes (FeSaDA et FeSODB) ont été isolées chez T. cruzi à partir
d'une banque d'ADNc (Ismail et al., 1997). La FeSODA a une séquence de 804 pb et
code pour 212 acides aminés. Cette enzyme est constitutivement exprimée à tous les
stades de développement parasitaire sous la forme d'un transcrit de 1,2 kb. La Fe-
34

Généralités
SODA partage 55% d'homologie de séquence avec celle de L. chagasi (Paramchuk et
al., 1997). Au contraire, la FeSODB, qui code pour 196 acides aminés est plus
exprimée dans les formes épimastigotes que dans les formes trypomastigotes ou
amastigotes. La Fe-SODB partage 62% d'homologie de séquence avec celle de L.
chagasi. De plus, dans les formes épimastigotes, elle est plus abondante que la forme
Fe-SODA (Ismail et al., 1997).
1.3.3.2 Les Cystéine Protéases
Les cystéine protéases sont impliquées dans la différenciation cellulaire, mais
également dans d'autres mécanismes cellulaires majeurs comme par exemple, la
virulence ou l'échappement des parasites à la réponse immune de l'hôte. Compte tenu
du rôle essentiel des cystéine protéases dans la biologie cellulaire de T. cruzi, nous
nous sommes intéressés également à l'expression de cette enzyme avec le but d'établir
son possible rôle dans la défense du parasite et dans la chimiorésistance au
Benznidazole.
Il existe 5 grands types de protéases, enzymes qui coupent des liaisons peptidiques,
classées selon leur mécanisme d'action et l'acide aminé au site actif (cystéine, sérine,
thréonine, aspartate) ou un atome métallique directement impliqué dans ce mécanisme.
Chez les trypanosomatidés, plusieurs types de protéases sont décrits, mais nous nous
sommes intéressés plus particulièrement aux protéases à thiol.
Protéases à sérine et protéases à thiol (cystéine protéases)
Ces protéases fonctionnent selon un schéma général commun, dans lequel l'enzyme
fixée (au site de reconnaissance) à la protéine la coupe en libérant sa partie C
terminale, tandis que la partie N-terminale acyle l'enzyme par le groupe carboxylique
qui vient d'être libéré. L'hydroxyl de la sérine et le SH de la cystéine sont déprotonés,
transférant leur proton à un noyau d'histidine (qui lui-même est activé par un aspartate
dans le cas des protéases à sérine). La différence essentielle réside dans le
groupement intermédiaire acylé : sérine dans le premier cas (X=O) et cystéine dans le
second (X=S).
35

Généralités
Les cystéine protéases chez T. cruzi
Les protéases de T. cruz; ont été purifiées pour la première fois par Rangel et
al. (1981a). Elles étaient présentes aux stades épimastigotes, trypomastigotes et
amastigotes (Rangel et al., 1981 b).
Une autre protéase similaire a été localisée dans les lysosomes de T. cruz; (Bontempi
et al., 1989). Cette enzyme a été décrite comme une glycoprotéine (GP57/51, Murta et
al., 1990) constituant un antigène majeur de T. cruz; (Scharfstein et al., 1986). Cette
protéase a été supposée participer au dommage tissulaire produit chez l'hôte. Cette
enzyme appartient à la superfamille des papaïne cystéine protéases (Cazzulo et al.,
1989,1990 ; Murta et al., 1990).
La séquence complète du gène de la cystéine protéase majeure de T. cruz; (tccp1) a
été reportée par Eakin et al. (1992). Un autre étude a montré que les gènes étaient
organisés en tandem Uusqu'à 130) et qu'il y avait au moins six copies dans le génome
du parasite (Campetella et al., 1992). Le nom de cruzipaïne (ou cruzaïne) a été suggéré
pour cette protéase majeure en raison des similitudes de séquences avec la papaïne
protéase.
L'expression simultanée de plusieurs de ces gènes produit un mélange d'isoformes
(Cazzulo, 1999). Les isoformes dits cruzipaïne où cruzaïne ou GP57/51 (type l, selon la
classification de Coombs et Mottram;' 1997) différent à la fois dans le domaine C
terminale, dans la séquence ainsi que dans le patron de N-glycosylation. Le domaine C
terminal contient de nombreuses modifications post-transcriptionnelles et est
apparemment responsable de l'hétérogénéité retrouvée (Campetella et al., 1992 ; Eakin
et al., 1992 ; Parodi et al., 1995 ; Cazzulo et al., 1997; Martinez et al., 1998).
Par contre, les isoformes nommées cruzipaïne2 (type 1) ont une différence marquée
dans la région catalytique (Lima et al., 1994). De plus, la spécificité et l'affinité des
cruzipaïne2 pour leurs inhibiteurs (Lima et al., 2001) diffèrent avec celles des autres
isoformes cruzipaïne (Serveau et al., 1996,1999).
36

Généralités
Différentes études ont montré que l'expression de la cruzipaïne est plus élevée chez les
formes épimastigotes, bien que le niveau d'ARNm soit similaire dans toutes les formes
parasitaires (Campetella etai., 1990; Bonaldo etai., 1991a ; Tomas et Kelly, 1996).
Chez T. cruzi, le gène qui code pour une cystéine protéase type III ou « cathepsin B
Iike» a été décrit (Garcia et al., 1998 ; 1'J6brega et al., 1998). Les analyses par
Southern blot indiquent que le gène (tccb) existe en une seule copie. Le domaine de
cette protéase a une homologie de 62% et de 60% avec les protéases « cathepsin B
Iike » de Leishmania mexicana (Bart et al., 1995) et Leishmania major (Sakanari et al.,
1997), respectivement.
Fonction
Chez T. cruzi, les cystéines protéases ont été impliquées dans plusieurs processus
cellulaires comme les processus d'invasion cellulaire, de virulence, de modulation de la
réponse immune de l'hôte et dans la différenciation cellulaire des parasites (Souto
Padron et al., 1990 ; Caler et al., 1998 ; Mottram et al., 1998 ; Cazzulo, 1999 ; revue,
Sajid et McKerrow, 2002).
D'après plusieurs auteurs (Meirelles et al., 1992 Franke de Cazzulo et al., 1994;
Cazzulo, 1999), la cruzipaïne pourrait participer:
i) à la digestion Iysosomiale des protéines exogènes ou endogènes du parasite,
ii) à la protection contre la réponse immune de l'hôte, à travers la destruction du
fragment Fc des immunoglobulines liées aux antigènes respectifs, en produisant un
fragment F(ab')2 incapable d'activer le complément,
iii) à la pénétration des formes trypomastigotes dans les cellules de l'hôte, car les
inhibiteurs des protéases inhibent ce processus,
iv) à différents points du cycle de différenciation cellulaire.
Inhibiteurs de Cystéine protéases
Etant donné que les cystéine protéases jouent un rôle important dans la survie des
parasites (Bonaldo et al., 1991 b ; Meirelles et al., 1992 ; Harth et al., 1993), plusieurs
auteurs les ont proposées comme cibles thérapeutiques potentielles (revue, McKerrow
et al., 1993, 1999).
37

Généralités
De plus, leurs inhibiteurs comme les aryl urées (Du et al., 2000), le diazométhane, la
fluoroacétone (Harth et al., 1993), la phénylalanine et l'homophénylalanine liés à la
fJuorométhylacétone (McKerrow et al., 1995 ; revue, McKerrow, 1995, 1999 ; Engel et
al., 1999; Selzer et al., 1999 ; Caffrey et al., 2000 ; Cazzulo, 2002) pourraient servir de
base pour produire des nouveaux médicaments contre la maladie de Chagas.
Les structures cristallines des formes actives et inactives de ces enzymes ont été
obtenues dans le but de mieux comprendre les différents aspects de la spécificité
(McGrath et al., 1995). La connaissance de ces structures permettra également la mise
au point d'inhibiteurs plus sélectifs (Huang et al., 2003).
1.3.3.3 P-Glycoprotéines
Les transporteurs ABC (pour « ATP-binding cassette»), dont font partie les P
glycoprotéines, participent aux mécanismes de multi-résistance aux médicaments anti
cancéreux chez homme. Ces protéines ont été également observées chez différents
parasites chimiorésistants, ce qui suggère qu'elles pourraient être aussi impliquées
dans la chimiorésistance au Benznidazole chez T. cruzi.
Tous les organismes, des micro-organismes jusqu'à l'homme, possèdent des
mécanismes très polyvalents pour résister aux effets toxiques des drogues (van Veen
et Konings, 1997, 1998). Ces organismes possèdent des protéines membranaires qui
transportent des molécules actives à travers les barrières cellulaires et qui sont
capables de réguler la cytotoxicité des médicaments, par exemple, en diminuant leur
concentration intra-cytoplasmique (Lewis, 1994 ; Balzi et Goffeau, 1994 ; Barst et
Ouellete, 1995).
Ces transporteurs sont divisés en deux types:
i) les transporteurs ABC (pour « ATP-binding cassette»), qui utilisent l'énergie fournie
par l'hydrolyse de l'ATP pour expulser les médicaments hors des cellules (revue,
Higgins, 1992), dont les P-glycoprotéines.
ii) les transporteurs secondaires, qui régulent la sortie des médicaments à travers
l'échange d'ions (Paulsen et al., 1996)
38

Généralités
Structure
L'unité de base d'un transporteur ABC est constituée de 4 modules, deux
membranaires (TMDs, transmembrane domains ou MSDs, membrane-spanning
domains), chacun constitué de six segments transmembranaires en configuration a
hélice séparés par boucles hydrophiles, et de deux domaines (NBDs, nucleotide
binding domains) de liaison et d'hydrolyse d'ATP qui fournissent l'énergie nécessaire à
la translocation du substrat au travers de la membrane. Les NBD sont extrêmement
conservés dans les diverses espèces. Ce prototype de transporteur ABC peut être codé
par un seul gène ou par des gènes séparés (parfois organisés en opéron) (revue,
Higgins, 1992 ; Kerr, 2002).
Les P-glycoprotéines ont été décrites chez plusieurs organismes parmi lesquels
différents parasites comme Entamoeba histolytica (Descoteaux et al., 1992),
Plasmodium falciparun (Foote et al., 1989 ; Wilson et al., 1989), Leishmania tarento/ae,
Leishmania major et Leishmania donovani (Ouellette et al., 1990 ; Henderson et al.,
1992 ; Hendrickson et al., 1993). De manière intéressante, ces transporteurs
parasitaires sont fortement homologues aux transporteurs de métazoaires impliqués
dans la résistance de certains cancers aux chimiothérapies (résistance MOR) (revue,
Légaré et al., 1994 ; Ullman, 1995 ; van Veen et Konings, 1998).
Les P-glycoprotéines chez T. cruzi
La présence de séquences homologues aux gènes des P-glycoprotéines associés à la
résistance aux métaux lourds chez Leishmania, a été décrite chez T. cruzi
(Dallagiovanna et al., 1994). L'analyse de la séquence du gène tcpgp1 montre un ORF
de 3105 pb qui code pour une protéine de 1035 acides aminés de masse moléculaire
estimé de 115 KDa (Torres et al., 1999). La séquence complète du gène tcpgp2 qui
code pour une autre P-glycoprotéine similaire a été rapportée par Dallagiovanna et al.
(1996). L'analyse de la séquence du gène montre un ORF de 4602 pb, qui code pour
un polypeptide de 1534 acides aminés de masse moléculaire approximative de 169
KDa.
L'analyse d'hybridation moléculaire a montré que le gène tcpgp2 est présent dans le
génome du parasite en une seule copie et que celui-ci est localisé dans un seul
chromosome de 900 kb. L'analyse des séquences a démontré qu'il existe deux sites
39

Généralités
accepteurs pour l'épissage en trans à -5 et -11 nucléotides en amont du codon
d'initiation prédit. En plus, cette analyse a montré que le site accepteur n'est pas
constitué par une séquence riche en polypyrimidine ou (CT)n comme chez d'autres
gènes du parasite.
Apparemment, le gène tcpgp2 est transcrit également à tous les stades parasitaires,
mais il existerait une accumulation différentielle des ARNm chez les formes
épimastigotes et amastigotes, phénomène qui pourrait être dû à la régulation post
transcriptionnelle de l'expression soit au niveau du processus de maturation du pré
ARNm, soit au niveau du transport ou de la localisation de l'ARN ou enfin au niveau de
la stabilité de l'ARNm.
La séquence des acides aminés prédite pour cette P-glycoprotéine était plus en relation
avec celle des P-glycoprotéines LtpgpA, LtpgpB et LtpgpE de L. tarento/ae , YCF1 des
levures et MRP des humains qu'avec les CFTR (pour cystic fibrosis transmembrane
regulator) humains et les MOR protéines.
Fonction
Dans les cellules de mammifères, le phénotype de résistance croisée aux médicaments
anticancéreux (MOR, pour multidrug resistance) est associé à l'amplification et/ou la
surexpression des gènes mdr qui codent pOLir les transporteurs membranaires P
glycoprotéines. Bien que les P-glycoprotéines aient un rôle dans les mécanismes de
résistance ch'éz Leishmania, Plasmodium et chez d'autres organismes, chez T. cruzi ce
rôle n'a pas pu être mis en évidence jusqu'au maintenant (Murta et al., 2001).
40

MATERIEL ET METHODES
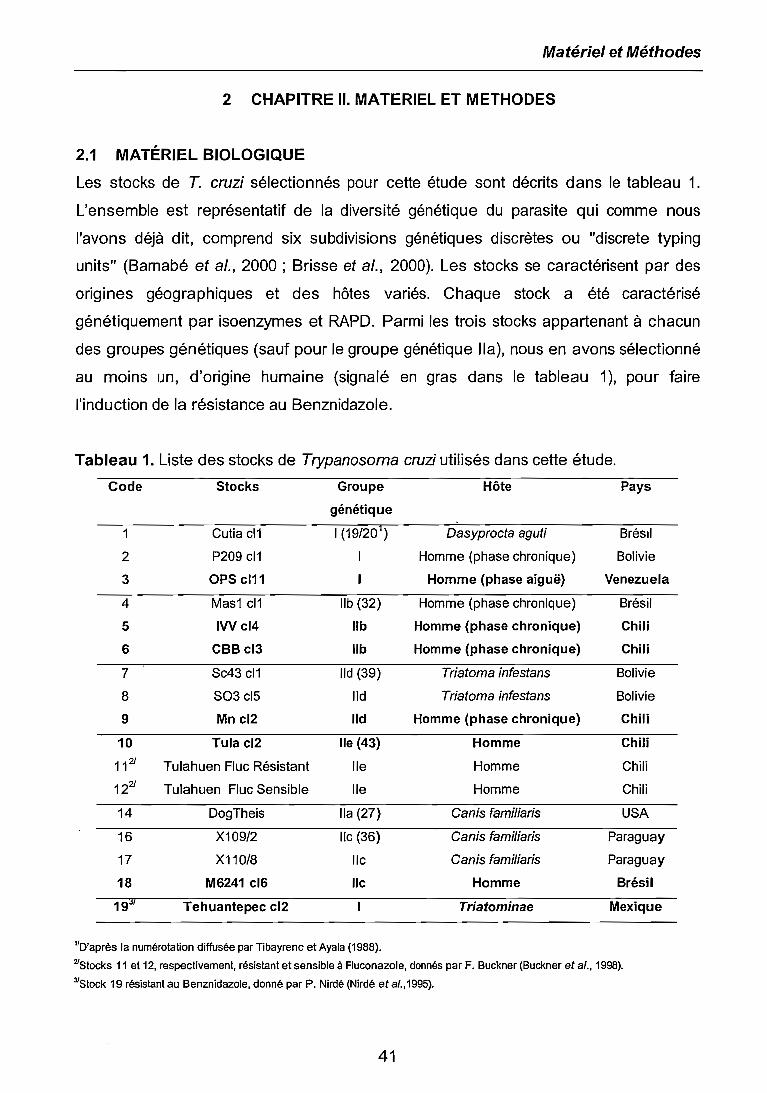
Matériel et Méthodes
2 CHAPITRE Il. MATERIEL ET METHODES
2.1 MATÉRIEL BIOLOGIQUE
Les stocks de T. cruzi sélectionnés pour cette étude sont décrits dans le tableau 1.
L'ensemble est représentatif de la diversité génétique du parasite qui comme nous
l'avons déjà dit, comprend six subdivisions génétiques discrètes ou "discrete typing
units" (Barnabé et al., 2000 ; Brisse et al., 2000). Les stocks se caractérisent par des
origines géographiques et des hôtes variés. Chaque stock a été caractérisé
génétiquement par isoenzymes et RAPD. Parmi les trois stocks appartenant à chacun
des groupes génétiques (sauf pour le groupe génétique lia), nous en avons sélectionné
au moins un, d'origine humaine (signalé en gras dans le tableau 1), pour faire
l'induction de la résistance au Benznidazole.
Tableau 1. Liste des stocks de Trypanosoma cruzi utilisés dans cette étude.
Code Stocks Groupe Hôte Pays
génétique
1 Cutia cl1 1(19/20') Dasyprocta aguti Brésil
2 P209 cl1 1 Homme (phase chronique) Bolivie
3 OPS cl11 1 Homme (phase aiguë) Venezuela
4 Mas1 c11 lib (32) Homme (phase chronique) Brésil
5 IWcl4 lib Homme (phase chronique) Chili
6 CBB cl3 lib Homme (phase chronique) Chili
7 Sc43 cl1 IId (39) Triatoma infestans Bolivie
8 S03 c15 IId Triatoma infestans Bolivie
9 Mn cl2 IId Homme (phase chronique) Chili
10 Tula cl2 Ile (43) Homme Chili
112/ Tulahuen Flue Résistant Ile Homme Chili
122/ Tulahuen Flue Sensible Ile Homme Chili
14 DogTheis lia (27) Canis familiaris USA
16 X109/2 IIc (36) Canis familiaris Paraguay
17 X110/8 Ile Canis familiaris Paraguay
18 M6241 cl6 Ile Homme Brésil
193/ Tehuantepec cl2 Tria tominae Mexique
l/D'après la numérotation diffusée par Tibayrenc et Ayala (1988).
21Stocks 11 et 12, respectivement, résistant et sensible à Fluconazole, donnés par F. Buckner (Buckner et al., 1998).
31Stock 19 résistant au Benznidazole, donné par P. Nirdé (Nirdé et al.,1995).
41

Matériel et Méthodes
2.2 METHODOLOGIE
2.2.1 CULTURE DES PARASITES
Les épimastigotes de T. cruzi ont été cultivés à 2rC dans le milieu LIT (Liver Infusion
Tryptose, annexe 2) en présence de sérum de veau fœtal décomplémenté (10% en
concentration finale).
Les épimastigotes chirniorésistants ont été cultivés dans le milieu de culture LIT modifié
auquel nous avons ajouté 30% de milieu TC-1 00 (L30TC, annexe 2).
2.2.1.1 Cinétique de croissance des parasites
Une cinétique de croissance est effectuée, pour chacun des 16 stocks de T. cruzi, par
comptage sur cellule de Thoma.
Méthode de Comptage direct
Après dilution au 1/10 de la culture parasitaire dans une solution de PBS 1X stérile, une
goutte est déposée sur la cellule de Thoma et le comptage est réalisé au microscope.
Pour chaque stock, trois comptages journaliers ont été effectués sur une période de 15
jours.
Méthode de comptage par cytometrie de flux
Pour les stocks sauv~ges ~t· ceux qui ont subi l'induction de la résistance au
Benznidazole, les courbes de croissance ont été réalisées à l'aide d'un cytomètre de
flux, à la fin de l'induction et en absence de Benznidazole.
Une courbe étalon a été faite pour ajuster ensuite toutes les conditions de mesure aux
parasites. Après la dilution et la vérification de la concentration parasitaire à 1x1 06
épimastigotes mr1 à l'aide de la cellule de Thoma, les épimastigotes morts sont colorés
avec une solution de iodure de propidium (IP) dans du PBS 1X. L'analyse cytométrique
est réalisée dans un cytomètre de flux FACSCalibur (Becton Dickinson) connecté à un
ordinateur Power PC Macintosh et équipé d'un laser réglé sur 488 nm.
Les courbes de croissance ont été réalisées pendant 15 jours, en faisant une lecture
par jour. La viabilité des épimastigotes est établie en mesurant la taille (Forward Scatter
FSC) et la «granulosité}} des parasites (Side Scatter SSC). Le calcul de la
42

Matériel et Méthodes
concentration parasitaire (épimastigotes mr1) est réalisé avec la formule suivante: [(x/2
- 136) / 6,436] x 104, dont x correspond au nombre mesuré de parasites par unité de
temps (1 : 46 min).
Détermination du temps de doublement
Le temps de doublement des parasites a été calculé à partir de données issues des
courbes de croissances réalisées par cytométrie de flux. Le calcul du temps de
doublement (Td) est réalisé à l'aide de la formule suivante:
Td=24*Ln(2)/((Ln(Xmax)-Ln(Xmin))/(Ymax-Ymin)) , avec:
Xmax: Jour correspondant à la fin de la phase de croissance exponentielle.
Xmin : Jour correspondant au debout de la phase de croissance exponentielle.
Ymax : Concentration parasitaire à la fin de la phase de croissance exponentielle.
Ymin : Concentration parasitaire au debout de la phase de croissance exponentielle.
2.2.1.2 Préparation des culots parasitaires
3,5 ml de suspension parasitaire sont centrifugés à 2800 g à 4°C pendant 20 min. Les
culots sont ensuite lavés deux fois dans du PBS 1X stérile et centrifugés à 2800 g dans
les mêmes conditions. Un troisième lavage est effectué dans 10 ml de PBS 1X stérile et
les culots sont ensuite transférés dans des microtubes de 2 ml, préalablement pesés.
Après une nouvelle centrifugation à 15000 g (à 4°C), le surnageant est éliminé et les
culots pesés puis congelés à -80°C dans les microtubes étiquetés. Les culots
parasitaires ont été utilisés soit pour l'extraction des enzymes, soit pour celle de l'ADN
ou encore pour celle des ARNm.
2.2.2 CARACTERISATIONS BIOCHIMIQUE ET MOLECULAIRE DES PARASITES
2.2.2.1 Electrophorèse Isoenzymatique Multilocus (MLEE)
Tous les stocks ont été régulièrement caractérisés par la méthode d'électrophorèse
enzymatique multilocus (MLEE) d'après le protocole modifié par Ben Abderrazak et al.
(1993) et Barnabé et al. (2000), avec le but de contrôler leur stabilité et/ou l'absence de
contamination. Le nom, le code et l'abréviation des 19 systèmes enzymatiques étudiés
dans ce travail sont présentés dans l'annexe 3.
43

Matériel et Méthodes
La caractérisation isoenzymatique a été réalisée au moins une fois par an pour tous les
stocks utilisés, et une fois avant toute expérimentation (par exemple, avant de
commencer le processus d'induction de la résistance au Benznidazole).
Préparation des extraits isoenzymatiques
Les parasites sont lysés dans la glace en présence d'un volume égal de stabilisateur
hypotonique (Ditbiotréitol 2 mM, Acide amino-n-caproïque 2 mM, EDTA 2 mM). Après
20 min d'incubation dans la glace, la suspension est centrifugée à 15000 g à 4°C
pendant 10 min. Le surnageant enzymatique est prélevé et conservé à -SO°C jusqu'à
son utilisation.
Electrophorèse
Les électrophorèses ont été réalisées sur des plaques d'acétate de cellulose (Helena
System). Les plaques sont imbibées pendant 20 min dans le tampon de migration
adéquat. L'excès de tampon sur les plaques est éliminé par séchage avec du papier
absorbant et les échantillons sont déposés sur ces plaques à l'aide d'un peigne à 12
puits (Helena System). Le gel d'acétate est ensuite déposé dans la cuve
d'électrophorèse. Une plaque de verre fait pression sur la plaque d'acétate afin
d'assurer un passage régulier du courant. La migration s'effectue dans la pellicule de
tampon à la surface du gel. Les conditions de migration (tampon utilisé, durée et
voltage) sont indiquées dans l'annexe 4.
2.2.2.2 Amplification de fragments d'ADN par amorces aléatoires (RAPD)
Le Random Amplified Polymorphie DNA (Welsh and McClelland, 1990), est une
technique dans laquelle l'ADN extrait est amplifié par PCR en utilisant une amorce
unique de 10 bases dont la séquence est déterminée aléatoirement. Les fragments
produits par cette amplification sont séparés par électrophorèse en gel d'agarose
concentré. Le gel, coloré au bromure d'éthidium est ensuite photographié sous UV. La
reproductibilité des RAPD pour les bandes majeures est très satisfaisante au
laboratoire en respectant les paramètres expérimentaux. Pour la lecture, les bandes
faibles ou des bandes inconstantes d'un essai à l'autre pour un même échantillon ne
sont pas prises en compte.
44
---

Matériel et Méthodes
Extraction de l'ADN total
L'extraction de l'ADN total est réalisée d'après le protocole récent proposé par Brisse et
al. (2000). Le culot parasitaire décongelé est remis en suspension dans 400 !JI de
solution de lyse (Tris-HCI 10 mM, pH 8,0 ; EDTA 1 mM). Après homogénéisation à la
pipette, du SDS est ajouté à 0,5% en concentration 'finale. Le culot est ensuite traité par
5 !Jg/ml finaux de RNase A pendant 2h à 3rc. Enfin, la protéinase K est ajoutée
(200!Jg /ml finaux) et la solution est placée au bain-marie à 55°C pendant la nuit. Deux
extractions au phénol puis au chloroforme/alcool isoamylique (24:1) sont ensuite
réalisées.
L'ADN présent dans la phase aqueuse est alors précipité à l'aide de 2,5 volumes
d'éthanol absolu froid (-20°C) en présence de 300 mM d'acétate de sodium (pH 5,2). Le
précipité d'ADN est immédiatement récupéré à l'aide d'une pipette Pasteur
préalablement stérilisée et refroidie. L'extrémité de la pipette, sur laquelle est enroulé
l'ADN, est placée dans un nouveau tube.
Après séchage pendant 20 min sur la paillasse, l'ADN est repris dans un volume d'eau
distillée stérile correspondant au poids du culot parasitaire initial (1!J1 pour 1mg). La
concentration de l'ADN total est alors voisine de 1!Jg/!JI.
Le dosage de l'ADN est réalisé par spectrophotométrie à 260 nm. La concentration de
l'ADN est estimée selon la relation 1DO = 50 !Jg/ml. L'ADN est dilué par paliers
successifs jusqu'à une concentration finale de 10 ng/!JI. Cette solution peut être
conservée à 4°C ou à -20°C jusqu'à son utilisation.
Amplification de l'ADN
L'ADN est amplifié dans un volume de 60!J1 final selon le protocole standard suivi dans
le laboratoire (Tibayrenc et al., 1993) avec 6 !JI de tampon 10X ; dNTPs à 100 JJM
chacun ; 1,2 !JI de l'amorce décamérique à 10 !JIVI ; 0,9U de Taq polymérase
(Boehringer Mannhein) et 20 ng d'ADN du stock à étudier. Les séquences des amorces
utilisées (Operon Technologies, Inc., Alameda, Etats Unis) sont répertoriées dans
l'annexe 5.
45

Matériel et Méthodes
L'amplification est réalisée dans un thermocycleur PTC-100 (MJ Research Inc., USA).
Le programme d'amplification comprend 45 cycles (1 min à 94°C, 1 min à 36°C et 2 min
à 72°C) suivis d'une étape d'élongation finale de 7 min à 72°C.
Les produits d'amplification ont été séparés par électrophorèse sur gel d'agarose à
1,6% dans du tampon TAE (Tris Acétate EDTA) 0,5X. Dix microlitres d'échantillon sont
mélangés à 2 !JI de tampon de charge (0,25% bleu de bromophénol, 0,25% xylène
cyanol, 40% w/v de sucrose) avant dépôt. Après l'électrophorèse, les gels sont colorés
par incubation 30 min dans un bac de TAE 0,5X contenant du bromure d'éthydium
(BET, 0,5!Jg/ml). Le gel est ensuite visualisé sous lumière ultraviolette (254 nm) et
photographié.
2.2.3 EVALUATION DE LA REPONSE DES EPIMASTIGOTES A LA PRESENCE DU
BENZNIDAZOLE
2.2.3.1 Détermination de l'ICSD par la micro-méthode MTT/PMS
La détermination de l'ICsD de chaque stock a été réalisée par la micro-méthode MTT [3
(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide] / PMS [phenazine
methosulfate] (Muelas-Serrano et al., 2000) à un temps de croissance identique pour
tous les stocks.
La corrélation entre nombre de parasites (en phase Log) et leur viabilité mesurée par
MTI/PMS est déterminée préalablement pour vérifier que les épimastigotes étaient au
début de la phase de croissance exponentielle (Muelas-Serrano et al., 2000). Ensuite,
l'effet du Benznidazole est déterminé après 24, 48, 72 et 96 h d'incubation pour
déterminer le meilleur temps d'évaluation.
Le principe de la méthode enzymatique consiste à déterminer la viabilité cellulaire en
utilisant le cofacteur NADPH produit que par les mitochondries des cellules vivantes,
pour réduire le sel jaune de tétrazolium en cristaux pourpres de formazan, dont la
quantité dépend du nombre des parasites viables. Les cristaux sont ensuite solubilisés
et la densité optique de la solution est lue à 575 ± 25 nm.
46

Matériel et Méthodes
Première étape: Culture des épimastigotes en présence de Benznidazole
Le Benznidazole (Ro 07-1051, F. Hoffmann - La Roche S.A) est préparé d'abord dans
du DMSO à une concentration de 10mM. Ensuite il est dilué à une concentration de
500IJM dans 0,25% de DMSO. Les concentrations de travail utilisées ont été: 50, 100,
150,200, 500 et 2000 IJM (solutions 10X, dans le milieu de culture).
Quatre-vingt dix rnicrolitres d'une dilution de parasites en phase exponentielle
(1 x1 06epis/ml) sont déposés par puits dans une plaque de 96 puits.
Pour chaque traitement concentration de médicament, 10 1-11 de solution de
Benznidazole (1 OX) sont déposés dans six puits pour tester la répétabilité du traitement,
puis les plaques sont incubées pendant 72 h à 2rC.
Deuxième étape: Evaluation de l'effet du Benznidazole à l'aide du test MTT
La solution de MTT/PMS (Mn 2,5 mg/ml; PMS 0,22 mg/ml) est ajoutée (10IJI/puits) et
les plaques sont incubées à 2rC pendant un période de 1 à 4 h jusqu'à ce que les
parasites ne bougent plus. Puis 1001-11 d'une solution de lyse (SDS1 0%, HCI 0,01 N) sont
ajoutés dans chaque puits et les plaques sont à nouveau incubées 30 min à
température ambiante sous agitation et à l'abri de la lumière. Finalement, l'absorbance
est mesurée à 570 nm dans un spectrophotomètre Multiskan.
Le calcull'ICso est effectué à partir de deux déterminations indépendantes en utilisant la
formule proposée par Hills (Hills et al., 1986; Serena et Lemesre, 1997) :
Log (ICso) =Log (X1) + [(Y1-ycl2) / (Y1-Y2)] [Log(x2)-Log(xtJ}. avec:
X1 : Concentration 1 de Benznidazole
X2 : Concentration 2 de Benznidazole
Yo: Valeur d'absorbance du contrôle sans Benznidazole
Y1 : Valeur d'absorbance à la concentration 1
Y2: Valeur d'absorbance à la concentration 2
Parmi tous les traitements, la valeur de Y1 est déterminée comme celle correspondant à
la valeur d'absorbance immédiatement au dessus de 50% par rapport au contrôle (Yo).
47

Matériel et Méthodes
De même, la valeur de Y2 est déterminée comme celle correspondant à la valeur
d'absorbance immédiatement au dessous de 50% par rapport au contrôle (Yo).
2.2.3.2 Détermination de l'ICso par cytométrie de flux
Afin de vérifier les valeurs d'lCso obtenues par la méthode de MTT/PMS, nous avons
déterminé la valeur d'ICso pour les stocks 3, 6 et 9 (voire tableau 1) en utilisant la
cytométrie de flux d'après le protocole proposé par Vergnes et al. (2002).
Neuf cents microlitres d'une dilution de parasites en phase exponentielle (1 x1 06epis/ml)
sont déposés par puits dans une plaque de 12 puits. Pour chaque traitement (Le.
concentration de médicament), 100 1-11 de la solution de Benznidazole (10X) sont
déposés dans chacun de trois puits (Le. répétition). Puis les plaques sont incubées
pendant 72, 96, 120 et 144 h à 2rC.
Après la période d'incubation correspondante, les épimastigotes morts sont colorés
avec une solution (1 mg/l-ll) de iodure de propidium (IP) dans du PBS 1X. L'analyse
cytométrique est ensuite réalisée dans un cytomètre de flux FACSCalibur (Becton
Dickinson). La viabilité des épimastigotes et le calcul de la concentration parasitaire
sont déterminés comme décrit précedemment (voir chapitre 2.2.1.1). L'ICso est
determinée pour le pourcentage d'inhibition de la croissance des parasites en présence
de Benznidazole par rapport à la croissance du témoin sans le médicament.
2.2.3.3 Croissance des parasites sans/avec Benznidazole à l'ICso
Une fois déterminée l'ICso, chacun des stocks sont cultivés en milieu LIT en présence
de Benznidazole à une concentration égale à l'ICso correspondant, pendant 72 heures à
2rC. La culture se fait parallèlement en l'absence de Benznidazole (groupe témoin).
Ensuite, les stocks sont récoltés et les culots obtenus sont utilisés pour l'étude de
l'expression génique différentielle en l'absence et en présence de Benznidazole (à
l'ICso).
2.2.3.4 Induction de la résistance au Benznidazole
L'induction de la résistance des parasites au Benznidazole est faite d'après le protocole
proposé par Nirdé et al. (1995). L'obtention de mutants résistants est réalisée en
48

Matériel et Méthodes
augmentant progressivement la concentration de médicament dans le milieu de culture.
Ceux-ci seront utilisés pour l'étude de l'expression génique différentielle.
L'induction de la résistance aux médicaments est un processus très long dû au fait que
la vitesse de croissance des parasites diminue et la mortalité augmente, il y a donc
moins de chances d'avoir assez de parasites pour pouvoir augmenter la dose jusqu'à la
concentration désirée. En conséquence, nous avons décidé de fixer un niveau-seuil
d'au moins 501JM, qui est la dose circulante de Benznidazole chez un patient en cours
de traitement (Raaflaub et Ziegler, 1979 ; Raallaub, 1980). Cette concentration
correspond au niveau de chimiorésistance thérapeutique.
Induction de la chimiorésistance jusqu'au niveau thérapeutique
Après leur caractérisation par isoenzymes et leur stabilisation, les six stocks
sélectionnés (stocks 3, 5, 6, 9, 10, et 18, voir tableau 1) pour suivre l'induction jusqu'au
niveau de résistance thérapeutique, ont été cultivés en présence et en l'absence de
Benznidazole (groupe témoin).
Pour l'induction de la chimiorésistance, les stocks ont été cultivés dans le milieu de
culture L30TC (annexe 2) contenant le Benznidazole (deux à dix fois concentrée)
pendant au moins dix semaines jusqu'à la stabilisation de la résistance. Ensuite, la
concentration de Benznidazole est augmentée et une nouvelle stabilisation est réalisé,. '
et ainsi de suite, jusqu'à l'obtention des mutants résistants. Les concentrations finales
de Benznidazole utilisées pour le processus d'induction de la résistance jusqu'au
niveau thérapeutique ont été les suivantes: 1, 2, 4, 8, 16, 20, 25, 30, 40 et 50IJM
(correspondant à dix niveaux de stabilisation en total).
A chaque niveau de résistance obtenu, les stocks résistants et sauvages (au même état
de croissance cellulaire) sont récoltés et cryoconservés dans l'azote liquide.
Induction de la chimiorésistance à différents niveaux pour un seul stock
Nous avons ajouté à notre étude le stock 19, qui a été induit en présence de
Benznidazole aux concentrations suivantes: 10, 15, 20, 50, 70, 90 et 100 IJM
(correspondant à sept niveaux de stabilisation en total). Le stock 19 résistant, induit par
Nirdé et al. (1995) a été cultivé en présence de 200IJM de Benznidazole.
49

Matériel et Méthodes
Le stock sauvage correspondant a été aussi cultivé sans Benznidazole et repiqué en
même temps que les parasites induits. A chaque niveau de résistance obtenu, les
stocks résistant et sauvages (au même état de croissance) sont récoltés et
cryoconservés dans l'azote liquide.
Stabilité de la résistance
Nous avons suivi le devenir du phénotype résistant après avoir stabilisé les stocks en
absence de Benznidazole. Pour cela, nous avons cultivé pendant au moins trois mois
tous les parasites chimiorésistants (au niveau thérapeutique et à différents niveaux pour
le stock 19) dans le milieu de culture L30TC sans Benznidazole. Ensuite, nous avons
soumis à nouveau les parasites aux concentrations de Benznidazole pour lesquelles ils
étaient devenus résistants.
2.2.4 ETUDE DE L'EXPRESSION GENIQUE DIFFERENTIELLE PAR LA
TECHNIQUE DERIVEE DU RADES
La technique utilisée dans ce travail (Amplification aléatoire de séquences exprimées)
est une technique dérivée de la technique RADES (pour Random Amplified
Differentially Expressed Sequences), ceci permet d'obtenir un profil d'expression des
gènes grâce à l'amplification aléatoire (RAPD) des produits rétro-transcripts des ARNm
(Murphyet Pellé, 1994).
La technique dérivée consiste à isoler et à rétrotranscrire l'ARNm des culots
parasitaires en ADN complémentaire. Ensuite, l'ADNe est amplifié avec un couple
d'amorces conservées (une amorce basée sur la séquence conservée du miniexon et
l'autre amorce un polydT) pour obtenir du matériel en quantité suffisante (ADNe) afin de
réaliser un grand nombre d'amplifications aléatoires. Une deuxième amplification est
faite à l'aide de la technique RAPD en utilisant des amorces décamériques.
La mise au point de la technique d'amplification aléatoire de séquences exprimées chez
T. cruzi a été faite en nous basant sur les travaux réalisés par Hide (1999) pour
l'analyse de l'expression génique différentielle chez les Leishmania.
50
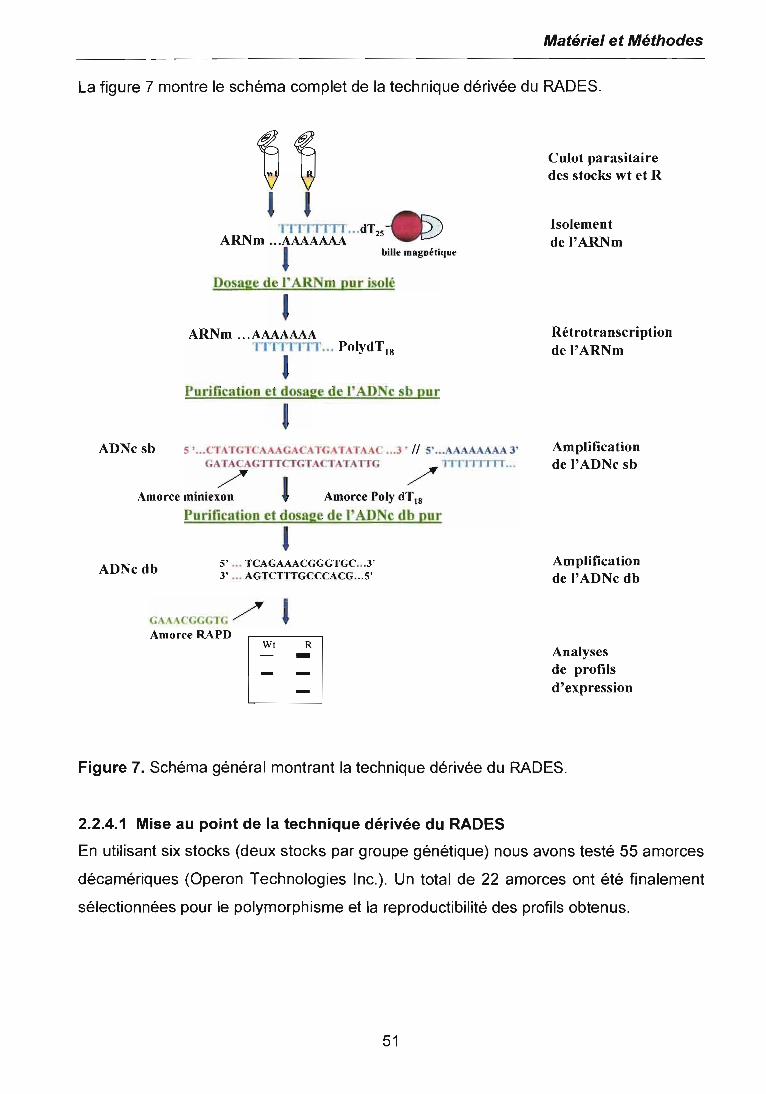
Matériel et Méthodes
La figure 7 montre le schéma complet de la technique dérivée du RADES.
~~1 1lTIIIIII ...dT2S•
ARNm ...AAAAAAA1 bille magnétique
DosaKc de l'AR m pur isolé
1
Culot parasitairedes stocks wt et R
Isolementde l'ARNm
ADNc sb 5 •••• . "T 1\ <- n;'A A A ••.3' Il 5' .••AA AAAAA J'
JO r .AGTITCfGT.'\CT T TrG / n Il 1 J 1 1 1. ..
Amorce minie~ 1 Amorce Poly dTI8
Purification et dosage de 1 AD c db pur
1ADNc db
Purification
s' TCAGAAACGGGTGC.•.3'3' AGTCTTTGCCCACG...5'
c sb pur
Rétrotranscriptionde l'ARNm
Amplificationde l'ADNc sb
Amplificationde l'ADNc db
G \('G GTG / 1Amo". RAPD 1 w. .: 1
- --Analysesde profilsd'expression
Figure 7. Schéma général montrant la technique dérivée du RADES.
2.2.4.1 Mise au point de la technique dérivée du RADES
En utilisant six stocks (deux stocks par groupe génétique) nous avons testé 55 amorces
décamériques (Operon Technologies Ine.). Un total de 22 amorces ont été finalement
sélectionnées pour le polymorphisme et la reproductibilité des profils obtenus.
51

Matériel et Méthodes
Extraction de l'ARN messager (ARNm)
L'ARNm est isolé à l'aide de billes magnétiques (Dynabeads® mRNA Direct kit, Dynal)
selon les conditions recommandées par le fournisseur. Deux cents microlitres de billes
(Dynabeads) sont mis en suspension dans un microtube de 1,7 ml. Les tubes sont
ensuite placés sur le portoir magnétique durant 30 s, puis le surnageant est enlevé. Les
billes sont resuspendues dans 200f.J1 de tampon de lyse (froid). Un ml de tampon de
lyse est ajouté au culot parasitaire congelé, puis mélangé par pipetage. Les tubes sont
mis ensuite sur la glace.
Pour diminuer la viscosité du lysat, l'échantillon est passé à travers une seringue de 1
ml (aiguille 21/22). Les tubes contenant le lysat sont centrifugés 30 s. Le surnageant de
la solution de billes est enlevé et le lysat parasitaire est ajouté sur les billes. Les tubes
sont ensuite agités et placés sur la roue 10 min à 25 tours/min. Ensuite, les tubes sont
placés sur le portoir magnétique 2 à 5 min (fixation du complexe billes-dT25-ARNm) puis
le surnageant est enlevé. Les billes sont rincées 2 fois avec 1000 f.J1 de tampon de
lavage A (froid) et 1 fois avec 1000 f.J1 de tampon de lavage B (à température ambiante).
Vingt-deux microlitres de tampon d'élution sont ajoutés puis les tubes sont placés 2 min
à 65°C, et immédiatement remis sur le portoir magnétique. Finalement, le surnageant
est récupéré (par aliquotes de 10 f.J1 d'ARNm) dans des microtubes (0,67 ml) propres.
Les ARNm récupérés sont alors dosés par spectrophotométrie. La rétrotranscription est
réalisée avec la moitié des ARNm obtenus (10f.J1) et le reste est conservé à -BO°C
jusqu'à son utilisation comme contrôle négatif des amplifications. Les billes sont ensuite
resuspendues dans 200 f.J1 de tampon de reconditionnement pour les régénérer et
pouvoir les utiliser à nouveau.
Les solutions utilisées pour l'extraction de l'ARNm sont répertoriées dans l'annexe 6.
Pour le protocole d'extraction des ARNm, le matériel en plastique est préalablement
traité à la RNase ZAP (Invitrogen) et le liquide est traité avec du DEPC (diethyl
pyrocarbonate).
52

Matériel et Méthodes
Dosage de l'ARNm
Le dosage de l'ARNm est réalisé par spectrophotométrie à 260 nm. La concentration de
celui-ci est estimée selon la relation 1DO = 40 I-Ig/ml et selon la formule:
(ng/l-ll) =O.026onmX facteur de dilution x 40.
Rétrotranscrip tion
La rétrotranscription permet de synthétiser l'ADN complémentaire simple brin (ADNc
sb) à partir de l'ARNm. Elle est réalisée grâce à une transcriptase reverse (AMV
reverse transcriptase, Promega) qui a une activité ARNase H lui permettant de
dégrader le brin d'ARNm matrice.
Pour la rétrotranscription, la première étape consiste à dénaturer les structures
secondaires de l'ARl\Jm et à hybrider celui-ci à l'amorce dT18. A cet effet, 10 1-11 de
ARNm (x ng) sont ajoutés à la moitié de la quantité (}'2 x ng) de dT18 selon le calcul
réalisé. Le mélange est incubé 5 min à 70 oC (dans le thermocycleur PTC-100, MJ
Research Inc.) et les tubes sont immédiatement mis sur la glace pour éviter la
renaturation des structures secondaires.
La seconde étape consiste à rétrotranscrire l'ARNm en ADN complémentaire simple
brin (ADNc sb) à l'aide du protocole suivant:
5 1-11 de ta.mpon de réaction (AMV RT 5X Reaction Buffer)
0,51-11 de Recombinant Rnasin® Ribonuclease Inhibiteur Promega (40 unités)
x 1-11 de dNTP mix (12,5 nmoi/l-lg d'ARNm)
x 1-11 de AMV Reverse Transcriptase Promega (15 unités/l-lg d'ARNm)
Ajuster le volume de réaction à 25 1-11 avec de l'eau miliQ stérile et traitée au DEPC.
On incube le mélange réactionnel 1 h à 42 oC (dans le thermocycleur PTC-100, MJ
Researeh Inc.). Après la rétrotranseription de l'ARNm en ADNe simple brin, l'ADNe sb
obtenu est purifié et quantifié au spectrophotomètre.
Purification de l'ADN simple brin
L'ADNc sb obtenu est purifié à l'aide du Kit QIAquick PCR Purification (Qiagen) selon
les conditions recommandées par le fournisseur. Cinq volumes du tampon PB sont
53

Matériel et Méthodes
ajoutés à un volume d'ADN, puis le mélange est transféré dans des tubes de 1,7 ml.
Après avoir mélangé et déposé la totalité du volume sur la colonne (placée sur le tube
collecteur), celui-ci est centrifugé 1 min à 15000 g. L'éluat est jeté et 750 IJI de tarnpon
PE sont ajoutés. La colonne est ensuite centrifugée 1 min à 15000 g. Il est
indispensable à cette étape de jeter l'éluat et de centrifuger à nouveau pour enlever
l'éthanol restant. Finalement, 50 IJI de tampon EB (Tris-HCI10mM, pH 8,5) sont ajoutés
à la colonne (placée sur un tube de 1,7 ml propre) pour éluer les ADNe sb purifiés.
Dosage de l'ADNe simple brin (ADNe sb)
Le dosage de l'ADNe sb est réalisé par spectrophotométrie à 260 nm. La concentration
de celui-ci est estimée selon la relation 1DO = 33 IJg/ml et selol1 la formule:
(ng/lJI) = O.026DnmX facteur de dilution x 33.
Amplification spécifique de l'ADNe simple brin
L'ADNe sb est amplifié avec deux amorces: l'une issue de la séquence conservée du
miniexon1 (SLc) de T. cruzi (Krieger et al., 1999) et l'autre (dT18), pour assurer une
quantité suffisante de matériel pour l'analyse RAPD.
Les réactions d'amplification de l'ADNe sb ont été mises au point sur les ADNe sb
obtenus à partir de parasites au même état de croissance cellulaire. Nous avons
modifié différentes conditions telles que la concentration des dl\lTPs, des amorces, de
la Taq polymérase et de l'ADN matrice. Les températures d'hybridation, le nombre de
cycles, les temps de dénaturation, d'hybridation et d'élongation ont été également
testés.
Les conditions optimales pOLir l'amplification ont été: l'amorce du rniniexon (SLc) à 16
IJM, l'amorce dT18 à 161JM, les dNTPs à 166,66IJM, 1U de Taq Polymerase (Boehringer
Mannhein), 10 IJI de tampon 10X et 100 ng d'ADNe dans un volume réactionnel de
1001J1.
1 Le miniexon ou Spliced Leader (SL) est une séquence conservée de 39 nucléotides présente àl'extrémité 5' de tous les ARNm matures des Trypanosomatidae (Ullu et al., 1996).
54

Matériel et Méthodes
Trente cinq cycles d'amplification ont été utilisés: 10 cycles à 61°C et 25 cycles à 45°C
(dénaturation 30 s, hybridation 1:30 min et élongation 3 min) et une étape d'élongation
finale à 72°C pendant 5 min. Cette amplification est réalisée dans le thermocycleur
PTC-100 (MJ Research Inc.).
Purification de l'ADN double brin (ADNe db)
Après l'amplification de l'ADNe simple brin en ADNe double brin (ADNe db), ce dernier
est purifié à l'aide du kit QIAquick PCR Purification (Qiagen) comme décrit ci-dessus.
Les ADNe db purs sont alors dosés par spectrophotométrie.
Dosage de l'ADNe double brin
Le dosage de l'ADNe db est réalisé par spectrophotométrie à 260 nm. La concentration
de l'ADN est estimée selon la relation 1DO = 50 I-lg/ml et selon la formule:
(ng/l-ll) =D.026onmx facteur de dilution x 50.
Amplification aléatoire de l'ADNe double brin par RAPD
L'amplification de l'ADNe db obtenu est réalisée selon le protocole standard de RAPD
suivi dans notre laboratoire (voir chapitre 2.2.2.2). Les amorces utilisées pour amplifier
"ADNe db ont été les mêmes que celles utilisées pour l'amplification de l'ADN total
(annexe 5).
Les produits d'amplification ont été séparés par électrop~orèse sur gel· d'agarose à
1,6% dans du tampon TAE (Tris Acétate EDTA) O,5X. Dix I-li d'échantillon sont
mélangés à 2 I-li de tampon de charge (0,25% bleu de bromophénol, 0,25% xylène
cyanol, 40% w/v de sucrose) avant dépôt. Après l'électrophorèse, les gels sont colorés
par incubation 30 min dans un bac de TAE 0,5X contenant du bromure d'éthydium
(BET, O,5I-lg/ml). Le gel est ensuite visualisé sous lumière ultraviolette (254 nm) et
photographié.
2.2.4.2 Etude de l'expression génétique différentielle à l'ICso par RADES
Les ARNm sont isolés à partir des culots parasitaires obtenus en absence ou présence
de Benznidazole à la concentration ICso (voir chapitre 2.2.3.3).
55

Matériel et Méthodes
Les ARI\lm sont quantifiés et rétrotranscrits en ADNc simple brin, puis amplifiés en
ADNc double brin à l'aide des amorces spécifiques (SLc et dT18) comme décrit
auparavant. Ensuite, ceux-ci sont réamplifiés par la technique RAPD en utilisant 22
amorces décamériques (voir chapitre 2.2.2.2).
Analyse des bandes différentielles à l'ICso
L'analyse des bandes différentielles est faite à partir des bandes issues de
l'amplification de l'ADNc db des stocks (16 stocks) en absence ou en présence de
Benznidazole à la concentration ICso spécifique de chaque stock. L'analyse se fait en
prenant en compte la reproductibilité des bandes (sur 2 PCR), la présence ou l'absence
de(s) bande(s), leur taille et leur intensité.
2.2.4.3 Etude de l'expression génétique différentielle des stocks sauvages et
résistants au niveau thérapeutique
Les ARNm sont isolés à partir des culots parasitaires obtenus des stocks sauvages et
résistants au Benznidazole au niveau de chimiorésistance thérapeutique (50IJM) pour
les stocks 3, 5, 6, 9, 10 et 18 (voir chapitre 2.2.3.4) au même état de croissance
cellulaire.
Les ARNm sont quantifiés et rétro-transcrits en ADNc simple brin puis amplifiés en
ADNc double brin à l'aide des amorces spécifiques (SLc et dT18), et réamplifiés par la
technique RAPD. L'analyse des bandes différentielles est faite par comparaison des
bandes issues de l'amplification de l'ADNc db des stocks sauvages et résistants, sur
des expériences réalisées en duplicat comme précédemment. L'analyse tient compte la
reproductibilité des bandes obtenues, de leur présence ou de leur absence, de leur
taille et de leur intensité.
2.2.4.4 Etude de l'expression génétique différentielle des stocks sauvages et
résistants à différents niveaux
Les ARI\lm sont isolés à partir des culots parasitaires obtenus du stock 19 sauvage et
chimiorésistant au Benznidazole à différents niveaux de résistance (15, 50, 100 et 200
IJM) au même état de croissance cellulaire.
56

Matériel et Méthodes
L'isolement, la rétrotranscription et l'amplification des ARNm sont réalisés comme il est
décrit précédemment, ainsi que l'analyse des bandes différentielles. Les bandes
différentielles sont alors sélectionnées pour le clonage et le séquençage.
2.2.4.5 Clonage et séquençage des bandes différentielles d'intérêt, issues des
stocks résistants et sauvages
Une fois réalisée l'analyse des bandes issues des produits RADES sur des stocks
résistants et sauvages, les bandes différentielles ont été isolées, clonées et
séquencées.
Isolement des bandes d'intérêt
Les bandes sélectionnées sont récupérées par découpage après séparation sur gel
NuSieve 1,5% et purifiées à l'aide du kit QIAquick Gel Extraction (Qiagen) selon les
conditions recommandées par le fournisseur.
Après 10 min d'incubation à 50°C avec le tampon QG (thiocyanate de guanidine), pour
faire fondre l'agarose, un volume d'isopropanol est ajouté et le mélange est alors
déposé sur la colonne (placée sur le tube collecteur). La colonne est ensuite centrifugée
1 min à 15000 g. Elle est ensuite rincée avec 500 1-11 de tampon QG et centrifugée à
nouveau pour enlever le l'agarose restant. Ensuite, 750 1-11 de tampon PE sont ajoutés
et la colonne est ,centrifugée à nouveau (1 min à 15000 g). L'éluat est jeté et la colonne
est centrifugée à nouveau pour enlever l'éthanol restant. Finalement, 50 1-11 de tampon
EB (Tris-HCI 10 mM pH 8,5) sont ajoutés à la colonne (placée dans un tube de 1,7 ml
propre) pour éluer l'ADN. Celui-ci est alors dosé par spectrophotométrie ou sur gel
d'agarose après coloration au BET et comparé aux standards de concentration connue.
Ligation dans le plasmide pGEM-T
La Iigation est réalisée en présence d'un mélange équimolaire de vecteur pGEM..T
(pGEM-T Easy Vector System l, Promega) et de la bande à cloner, dans un volume
final de 10 IJI. Le mélange réactionnel est composé de :
5 1-11 de tampon 2X
1 IJI de plasmide pGEM-T Easy vector (50ng/lJl)
1 1-11 d'enzyme de ligation T4 DNA Ligase (3 Weiss units/lJl)
57

Matériel et Méthodes
Ajuster le volume de réaction à 10 !JI avec de l'eau rniliQ stérile.
Le mélange est incubé une nuit à 4°C. Une réaction de contrôle de la qualité de la
Iigation est réalisée avec 2 !JI d'ADN témoin fourni avec le kit.
Transformation
La transformation est réalisée à partir de bactéries JM109 compétentes (Promega). Le
génotype de ces bactéries est recA1, endA1, gyrA96, thi, hsdR17 (rk-, mk+), re/A1,
supE44, II (lac-proAB), [F', traD36, proAB, laclqZ~15]. Les cellules sont conservées à
-80°C et décongelées sur la glace pour leur utilisation.
Pour la transformation, 2 !JI du mélange de Iigation sont déposés dans un tube
contenant 100!J1 de bactéries compétentes. Ensuite, les tubes sont légèrement agités
pour les mélanger et sont alors incubés pendant 20 min sur la glace. Un choc thermique
est appliqué par incubation 45-50 s au bain-marie à 42°C, puis les tubes sont remis
dans la glace pendant 2 min. Neuf cents microlitres de milieu SOC sont ajoutés et les
tubes sont incubés à 3rC pendant 1-3 h sous agitation (-150 rpm).
Etalement
Les étalements sont réalisés sur des boîtes de Petri contenant le milieu LB-agar
(annexe 7) avec Arnpicilline (50!Jg/ml finaux). Les boîtes sont conservées à 4°C et
préchauffées à température ambiante avant leur utilisation. Juste avant l'étalement,
200!J1 de bactéries transformées sont mélangés à 20 !JI de X-Gal (2%) et à 50 !JI d'IPTG
(0,1 M). Le mélange est étalé à l'aide d'une pipette Pasteur préalablement préparée à la
flamme en forme de râteau.
Les boîtes sont alors incubées en position inversée pendant 16-24h à 3rC. Puis, les
boîtes sont placées à 4°C pour bien révéler les colonies bleu non recombinantes et
faciliter la sélection des colonies blanches possédant les fragments d'intérêt.
Production de masse des colonies
Cinq à dix clones blancs sont sélectionnés, entourés et numérotés. Ceux-ci sont ensuite
prélevés à l'aide d'un cône stérile et plongés dans un tube de 15 ml contenant 3 ml de
58

Matériel et Méthodes
milieu de culture LB liquide avec ampicilline. Les tubes sont alors incubés une nuit à
3rC sous agitation.
Après une nuit d'incubation, il est possible à cette étape d'immortaliser les clones. Pour
cela, 200 ~I de la culture bactérienne sont mélangés à 200 ~I de milieu LB liquide avec
glycérol (50% v:v) dans des tubes Nunc. Les cellules transformées sont alors
conservées à -20°C.
Les tubes sont ensuite centrifugés à 2800 g pendant 2 min. Le surnageant est enlevé et
les culots conservés à -20°C jusqu'à l'extraction des plasmides recombinants.
Extraction des plasmides recombinants
L'extraction des plasmides recombinants est réalisée à l'aide du kit QIAprep Spin
Miniprep (Qiagen) selon les conditions recommandées par le fournisseur: Deux cents
cinquante microlitres de tampon P1 froid (Tris-HCI 50 mM pH8,O ; EDTA 10 mM, RNase
A 100 ~g/ml) sont ajoutés aux culots bactériens, puis le mélange est transféré dans des
tubes de 1,7 ml. Ensuite, 250 1-11 de tampon P2 (NaOH 200 mM, SDS 1%) sont ajoutés
et les tubes sont mélangés doucement par inversion (au moins 6 fois). 350 1-11 de
tampon N3 (hydrochloride de guanidine et isopropanol) sont ajoutés et les tubes sont
immédiatement agités par inversion pour éviter le phénomène de précipitation.
Les tubes sont ensuite centrifugés 10 min à 15000 g. Le surnageant e~t alors récupéré
et déposé sur la colonne (placée sur le tube collecteur). La colonne est ensuite
centrifugée 1 min à 15000 g. Celle-ci est ensuite rincée avec 500 1-11 de tampon PB
(hydrochloride de guanidine et isopropanol) et centrifugée (1 min à 15000 g). Ensuite,
750 1-11 de tampon PE sont ajoutés et la colonne est centrifugée (1 min à 15000 g).
L'éluat est jeté et la colonne est centrifugée à nouveau pour enlever l'éthanol restant.
Finalement, 50 1-11 de tampon EB (Tris-HCI 10 mM pH 8,5) sont ajoutés à la colonne
(placée sur un tube de 1,7 ml propre) pour éluer les plasmides recombinants.
Vérification de la taille des fragments clonés par la digestion des plasmides
Cette étape est une vérification de la présence du plasmide recombinant. Cinq
microlitres de la solution de plasmides recombinants sont digérés pendant 1 h à 3rC
dans un volume final de 20 ~I contenant 1 1-11 d'enzyme EcoRI (10U/I-II) et 21-11 de tampon
59

Matériel et Méthodes
10X (GIBCO). Les produits de digestion sont alors séparés par électrophorèse sur gel
d'agarose à 1,6% pour vérifier la taille du fragment inséré.
Les plasmides recombinants qui contiennent les fragments d'intérêt sont alors dosés
par spectrophotométrie ou sur gel d'agarose après coloration au BET et comparaison
avec des standards de concentration connue. Ceux-ci sont alors conservés à 4°C
jusqu'à leur utilisation.
Amplification des plasmides recombinants
Les réactions de séquence sont réalisées à l'aide du Kit ABI PRI8M® BigDye ™
Terminator v3.1 Cycle Sequencing Ready Reaction (Applied Biosystems) en suivant les
conditions recommandées par le fournisseur. Le mélange réactionnel est composé de :
8 pl de Terminator Ready Reaction Mix
4 1-11 d'amorce M13 (3,2 pmoles)
10-100 ng de plasmide recombinant (selon la taille du fragment cloné)
Ajuster le volume de réaction à 20 1-11 avec de l'eau déionisée.
Le programme d'amplification comprend 25 cycles (10 s à 96°c, 5 s à 50°C et 4 min à
60°C) suivis d'une étape indéfinie à 4°C. Cette amplification est réalisée dans un
thermocycleur PTC-100 (MJ Research Inc.).
Les produits obtenus sont purifiés par précipitation à l'éthanol, en les mélangeant à 64
1-11 d'éthanol pur et à 16 1-11 d'eau stérile. Ils sont incubés 15 minutes à température
ambiante, puis centrifugés 20 min à 15000 g. Le surnageant est enlevé et 250 1-11
d'éthanol à 70% sont ajoutés. Les tubes sont brièvement mélangés et centrifugés 10
min à 15000 g. Après avoir enlevé délicatement la totalité du surnageant, les tubes sont
laissés ouverts pour sécher complètement le culot (difficilement visible) et ensuite les
ceux-ci sont gardés à 4°C jusqu'au séquençage.
Séquençage
Nous avons utilisé un séquenceur monocapillaire automatique ABI PRISM™ 310
Genetic Analyzer (Applied Biosystems) pour réaliser le séquençage des fragments
clonés.
60

Matériel et Méthodes
Pour le chargement des échantillons sur l'appareil, les culots secs sont repris dans 201J1
de formamide HiDi. Les tubes sont passés au vortex et centrifugés brièvement avant de
transférer la totalité du volume dans des tubes de 0,2 ml. Puis les échantillons sont
dénaturés 4 min à 95°C et les tubes sont immédiatement remis à 4°C. Ensuite, le
séquenceur, préalablement calibré et préparé est chargé avec les échantillons
dénaturés.
2.2.5 ETUDES DES GENES SPECIFIQUES
Une des méthodes utilisées pour estimer le niveau d'expression des ARNm, est la RT
PCR, basée sur le principe de l'amplification par PCR (réaction de polymérisation en
chaîne) de l'ADNe issu de la transcription inverse l'ARNm d'intérêt. Cependant, la
réaction de PCR est un processus d'amplification exponentiel et, dans ces conditions,
de très faibles variations d'efficacité de la réaction produisent des modifications très
importants de la quantité de produit final.
Comme il est impossible de déterminer l'efficacité de la réaction PCR, qui peut même
varier d'un tube à l'autre dans une même expérience, il faut considérer que la RT-PCR
n'est pas une méthode quantitative. Néanmoins, pour pallier à ce problème, plusieurs
méthodes dérivées de la RT-PCR (RT-PCR relative, compétitive, comparative et RT
PCR en temps réel) ont été décrites, qui permettent Llne quantification fiable et
reproductible des molécules d'ARNm (Gilliland et al., 1990 ; Bouaboula et al., 1992 ;
Vidal,2000).
L'étude de l'expression des gènes candidats (tcpgp-2, tcsod-A, tcsod-B et tccp1) a été
réalisée par la méthode de RT-PCR relative, dans laquelle les ARNm rétrotranscripts
sont amplifiés par un couple d'amorces spécifiques correspondant aux gènes d'intérêt.
Pour comparer ensuite leur expression à celle d'un témoin, le gène de ménage2 de
l'actine a été utilisé pour la RT-PCR comme contrôle interne.
2Gène de ménage ou « Housekeeping gene» car sa synthèse est réalisé dans toutes les cellulesnucléées et il est nécessaire pour la survie de la cellule (Thellin et al.,1999).
61

Matériel et Méthodes
Dans ce travail, nous avons amplifié par PCR les produits obtenus après la RT-PCR. Le
protocole pour l'isolement et la rétrotranscription des ARNm ainsi que pour
l'amplification de l'ADNc sb en ADNc db (avec les amorces SLc et dT18) est décrit dans
le chapitre 2.2.4.1.
Pour l'amplification, les ADNc db sont dosés par spectrophotométrie comme décrit ci
dessus et aussi par une méthode semi-quantitative dite de «boîtes SET». Vingt cinq
millilitres d'agarose 1,5% dans du tampon TAE 0,5X sont chauffés au four à micro
ondes. Une fois l'agarose refroidi, 5 !JI de bromure d'éthydium pur (SET) sont ajoutés.
Le mélange est versé dans des boîtes de Petri et celles-ci sont laissées ouvertes 30
min à température ambiante pour permettre à l'agarose de se polymériser. A l'aide d'un
quadrillage, 1 !JI d'ADN standard de concentration connue est déposé sur l'agarose.
Une gamme de concentrations décroissantes d'ADN standard est utilisée: 200 ; 150 ;
100 ; 75 ; 50 ; 37,5 ; 25; 18,75 ; 12,5 et 9,375 ng/!J1. Ensuite, 1 !JI d'échantillon d'ADNc
est déposé et les boîtes sont laissées 15 min à température ambiante pour permettre à
l'ADN de se mélanger au SET. Les boîtes sont ensuite visualisées sous lumière
ultraviolette (254 nm), photographiées et enregistrées.
Les images capturées sont analysées à l'aide du logiciel Scion Image (NIH image,
www.scioncorp.com). pour ensuite calculer, grâce à une courbe étalon, la concentration
de l'ADN des échantillons par rapport à la gamme de concentrations standard. Une fois
le calcul effectué, tous les ADN sont dilués à une concentration donnée, ils sont à
nouveau dosés à l'aide de la boîte SET pour vérifier que tous les ADNc sont à la même
concentration.
2.2.5.1 Amplification du gène de ménage
Les ADNc sont alors amplifiés avec un couple d'amorces sélectionné à partir de
l'alignement des séquences connues du gène de l'actine de T. cruzi. L'expression de ce
gène est utilisée pour calibrer l'expression des ADNc et pouvoir ainsi quantifier, de
façon relative, l'expression des gènes spécifiques sélectionnés. Dans l'annexe 8, sont
présentés le nom, la longueur, le poids moléculaire, le % de GC et le Tm du couple
d'amorces sélectionné pour amplifier le gène de l'actine.
62

Matériel et Méthodes
Les réactions d'amplification du gène de l'actine ont été mises au point sur les ADN
d'un représentant de chacune des lignées de T. cruzi, isolés de parasites au même état
de croissance cellulaire. Différentes concentrations de dNTPs, d'amorces, de Taq
polymérase, d'ADN matrice ainsi que différents temps de dénaturation, d'hybridation et
d'élongation ont été testées.
Différents gradients de température ont également été réalisés dans un Mastercycler®
gradient (Eppendorf, Allemagne) pour sélectionner la température d'hybridation
optimale.
Nous avons également déterminé le nombre optimum de cycles qui permet l'obtention
de produits avant la phase de saturation (ou phase plateau de la réaction). Ensuite, les
amplifications ont été réalisées en utilisant l'ADNe db synthétisé à partir des ARNm
issus des stocks résistants et sauvages.
Les conditions optimales pour l'amplification du gène de l'actine sont: Pour un volume
réactionnel de 30 1-11, 3 1-11 de tampon 10X, l'amorce directe à 0,20 I-IM, l'amorce reverse
à 0,20 I-IM, les dNTPs à 150 I-IM chacun, 1,5 U de Taq Polymérase et 50 ng d'ADNe db.
L'amplification est réalisée dans un Mastercycler® gradient (Eppendorf, Allemagne)
avec un programme d'amplification comprenant 1 cycle 5 min à 94oC, 23 cycles
(dénaturation 30 s à 94oC, hybridation 1 min à 56°C et élongation 1 min à 72°C) et une
étape d'élongation finale à 72°C pendant 10 min.
Les produits d'amplification sont séparés par électrophorèse sur gel d'agarose à 2,2%
TAE 0,5x, colorés, photographiés et analysés comme décrit ci-dessus. Les images
capturées sont analysées à l'aide du logiciel Scion Image (NIH image), pour ensuite
calibrer à nouveau la quantité d'ADNe par rapport à l'expression du gène de l'actine.
2.2.5.2 Amplification des gènes spécifiques d'intérêt
A partir de l'alignement des séquences connues des gènes spécifiques de la P
glycoproteine-2 (tcpgp-2), des superoxyde dismutases (tcsod-A et tcsod-B) et de la
cruzipaïne-1 (tccp1) chez T. cruzi, à l'aide du logiciel Clustal X (Thompson et al., 1997)
nous avons sélectionné les zones d'homologie de séquences pour chaque gène. A
l'aide des logiciels Primer 3 (Rozen et Skaletsky, 2000) et Amplify 1.2 (Engels, 1992)
63

Matériel et Méthodes
nous avons sélectionné et testé des couples d'amorces capables d'amplifier
potentiellement les fragments souhaités pour chacun des gènes étudiés.
Le choix définitif a été basé notamment sur la longueur des fragments amplifiés mais
aussi sur d'autres critères comme le Tm et le % de Ge des amorces (annexe 8). Enfin,
nous avons sélectionné des couples d'amorces avec lesquelles nous avons obtenu des
produits amplifiés de 200, 302, 315 et 412 pb pour les gènes tcpgp-2, tcsod-A, tcsod-B
et tccp 1, respectivement.
La mise au point de l'amplification des gènes a été faite à partir de l'ADNe des parasites
dans un même état de croissance cellulaire. Différents gradients de température ont
été réalisés pour sélectionner la température d'hybridation optimale pour chaque gène.
Egalement, nous avons fait la détermination du nombre de cycles qui permet l'obtention
de produits avant la phase de saturation.
Après les mises au point, les amplifications sont réalisées en utilisant l'ADNe db
synthétisé à partir des ARNm issus des stocks résistants et sauvages. Les conditions
optimales pour l'amplification des gènes spécifiques ont été : dans un volume
réactionnel de 30 1-11, 3 1-11 de tampon 10X, l'amorce directe à 0,20 I-IM, l'amorce reverse
à 0,20 I-IM, les dNTPs à 150 pM chacun, 1,5U de Taq polymérase (Boehringer
Mannhein) et 50 ng d'ADNe db.. L'amplification est réalisée dans un Mastercycler®
gradient (Eppendorf, 'Allemagne) avec un programme d'amplification comprenant 1
cycle 5 min à 94°C, 23 cycles (tableau 2) et une étape d'élongation finale à 72°C
pendant 10 min.
Tableau 2. Programmes d'amplification des gènes spécifiques.
Conditions Tepgp-2 tcsod-A tes0 d-B tecp1
Dénaturation 5 min à 94°C
Dénaturation 30sà94°C 305 à 94°C 30 s à 94°C 30sà94°C
Hybridation 1 min à 65°C 1 min à 56°C 1 min à 50°C 1 minà71°C
Elongation 1 min à 72°C 1 min à 72°C 1 min à 72°C 1 min à 72°C
Nombre de cycles 23 23 23 23
Elongation finale 10 min à 72°C
64

Matériel et Méthodes
Les produits d'amplification sont séparés par électrophorèse sur gel d'agarose à 2,2%,
colorés, et photographiés comme décrit ci-dessus.
Les images capturées sont analysées à l'aide du logiciel Scion Image (NIH image), pour
comparer l'expression de gènes spécifiques entre les stocks résistants et sauvages.
2.2.6 METHODES D'ANALYSE DES DONNEES
2.2.6.1 Analyses statistiques
Les analyses phylogénétiques ont été réalisées à partir de données issues
d'isoenzymes, de RAPD et de RADES à l'ICso, en utilisant les méthodes de construction
d'arbre UPGMA (Unweighted Pair-Group Method with Arithmetic Averages, Sneath et
Sokal, 1973) ou NJ (Neighbor-Joining tree, Saitou et Nei, 1987) à l'aide du logiciel
«Genetic tools box» développé dans notre laboratoire.
Les distances génétiques de Jaccard (Jaccard, 1908), qui ont servi à la construction
des arbres, ont été calculées à l'aide du prograrnme Neighbor du PHYLIP « package»
[Felsenstein J. PHYLIP (Phylogeny Inference Package) version 3.57c. Seattle, WA:
University of Washington ,1995].
La distance de Jaccard se calcule selon la formule suivante:
o =1 - a / (a+ b + c)
dans laquelle:
a =nombre de bandes communes aux deux génotypes comparés
b =nombre de bandes présentes dans le premier génotype et absentes dans le second
c = nombre de bandes absentes dans le premier génotype et présentes dans le second
Les arbres sont visualisés à l'aide du logiciel Treeview (Page, 1996).
Les valeurs moyennes d'ICso entre stocks ont été comparées par le test ANOVA tandis
que la comparaison entre les groupes génétiques a été réalisée par le test de Fisher
65

Matériel et Méthodes
PLSD du programme Statview [StatVieW® software Web site fram SAS Institute Inc.
("SAS"). SAS Institute Inc., SAS Campus Drive, Cary, North Carolina 27513, USA,
2002].
La corrélation entre les valeurs d'ICso et les distances génétiques a été réalisée par le
test non paramétrique de Mantel (Mantel, 1967) à l'aide du logiciel ADE-4 (version
2001, Thioulouse et al., 1997).
2.2.6.2 Analyses des séquences
Les séquences brutes obtenues ont été corrigées à l'aide du logiciel Chromas 2.23
(Technelysium PtY Ltd, 1998-2002) afin d'éliminer les éventuelles incertitudes dans les
chromatogrammes.
L'alignement des séquences a été effectué soit par le programme Clustal X (Thompson
et al., 1997) soit par le programme Megalign (Expert Sequence Analysis Software,
version 5, DNASTAR) ou encore par I~ programme ALiGN «Query using sequence
data» (Pearson et al., 1997) sur le serveur GENESTREAM de l'IGH (Montpellier).
Les séquences nucléotidiques ont été utilisées pour la recherche des identités à travers
l'algorithme BLASTN (BLAST with eukaryotic genomes) sur le site NCSI (National
Center for Biotechnology Information, USA).
Le recherche d'homologies a été faite à l'aide de l'algorithme BLASTX (Altschul et al.,
1997) du NCBI et BLASTX du Parasite Genomes WU-Blast2 (Gish, 1996-2003)
d'EMBL-EBI (European Bioinformatics Institute, UK).
Les séquences nucléotidiques obtenues ont été comparées avec différentes banques
de données: EMBL nucleotide sequence database (Stoesser et al., 2003), Genbank
database du NCBI, DDBJ database (DNA database of Japan), EBI parasite database et
TIGR database (The Institute for Genomic Research).
Les séquences d'ADN ont également été traduites avec le logiciel «Six frame
translation of sequence du SCM Search Launcher » (Baylor College of Medicine HGSC)
66

Matériel et Méthodes
disponible sur le serveur « ExPASy (Expert Protein Analysis System) Molecular Biology
Server » de l'institut de bioinformatique suisse SIB (Gasteiger et al., 2003).
La recherche d'homologies a été également réalisée à partir de séquences des acides
aminés obtenues à l'aide de l'algorithme BLASTP sur le serveur ExPASy (SIB BLAST
Network service).
Les séquences des acides aminés obtenues ont été comparées avec différentes
banques de données: Swiss-Prot+TrEMBL+TrEMBL_NEW (Boeckmann et al., 2003).
67

RESULTATS

Résultats
3 CHAPITRE III. RESULTATS
3.1 CARACTERISATIONS BIOCHIMIQUE ET MOLECULAIRE DES PARASITES
Afin d'écarter la possibilité de contamination entre différents stocks de T. cruz; et de
vérifier leur stabilité, les parasites ont d'abord été caractérisés par isoenzymes et par
RAPD, puis au moins une fois par an par isoenzymes.
3.1.1 Isoenzymes (MLEE)
Nous avons vérifié le génotype de chacun des stocks en utilisant 19 systèmes
enzymatiques différents (correspondant à 22 locus) avant chaque manipulation. Les
stocks qui ont subi le processus d'induction de résistance ont été également
caractérisés après l'induction.
La figure 8, montre les profils spécifiques des groupes génétiques l, lib, IId et Ile pour
le locus enzymatique Gpi.
1 2 3 4 5 6 7 8 9 10 11 12"-y---J "-y---J "-y---J "-y---J
Il b Il d Il e
Figure 8. Electrophorèse de la Glucose-6-phosphate isomérase (GPI) de 12 des 16stocks de T. cruz; étudiés. L'ordre des stocks correspond à celui du tableau1. En bas, le groupe génétique correspondant.
La même analyse réalisée pour les stocks sauvages (wt) et pour les stocks qui ont subi
le processus d'induction de la résistance (R) jusqu'au niveau de chimiorésistance
68

Résultats
thérapeutique (50IJM) ne montre pas de différences d'expression entre les stocks
sauvages et les stocks résistants correspondants (figure 9).
yy yy yyyy yy yy3wt 3R 6wt 6R 9wt 9R 10wt 10R 5wt 5R 18wt 18R
Figure 9. Electrophorèse de la Glucose-6-phosphate isomérase (GPI) des stockssauvages (wt) et des stocks de T. cruzi chimiorésistants (R) au niveauthérapeutique (50IJM). L'ordre des stocks correspond à celui du tableau 1.
La figure 10 présente les profils isoenzymatiques de la GPI pour le stock 19 sauvage
(19wt) et pour le même stock à différents niveaux de chimiorésistance: 15IJM (R15),
50IJM (R50), 100IJM (R100) et 200IJM (R200). La comparaison entre les différents
profils montre clairement qu'il n'y a pas eu de changement au cours du processus
d'induction. De manière intéressante, pour le stock 19, au fur et à mesure que le niveau
de résistance augmente le niveau d'expression de cette enzyme augmente également.
\ A A Ay y yA j
y y
19wt R15 R50 R100 R200
Figure 10. Electrophorèse de la Glucose-6-phosphate isomérase (GPI) du stock 19sauvage (wt) et chimiorésistant correspondant à différents niveaux derésistance.
69

Résultats
L'annexe 9 présente les données MLEE (22 locus isoenzymatiques) pour les 16 stocks
étudiés.
3.1.2 RAPD
Nous avons également déterminé les profils RAPD des 16 stocks en utilisant 22
amorces décamériques (dans deux amplifications indépendantes). Les profils
d'amplification obtenus pour chaque amorce ont été étudiés indépendamment. Pour
une amorce donnée, chaque niveau de bande est codé spécifiquement par
comparaison entre l'échantillon total (16 stocks). L'annexe 10 présente les données
déterminées par RAPD (22 amorces) pour les 16 stocks étudiés.
Les profils d'amplification obtenus avec l'amorce R1 montrent un patron spécifique pour
chaque groupe génétique (figure 11). L'amorce A2 présente un polymorphisme moindre
(figure 12) et sépare les lignées génétiques 1(DTU 1) et Il (DTUs lib, IId, Ile, lia et IIc).
En général, toutes les amorces RAPD ont révélé des profils polymorphes, avec une
grande variabilité d'une amorce à l'autre. Par exemple, l'amorce N4 (figure 13) présente
un polymorphisme moindre par rapport à l'amorce N15 (figure 14).
1 2 3 4 5 6 7 8 9 10 11 12 14 18 - PM
Figure 11. Profils RAPD obtenus avec l'amorce R1 pour 14 des 16 stocks de T. cruziétudiés. L'ordre des stocks correspond à celui du tableau 1. En bas, lesgroupes génétiques correspondants. PM : Marqueur de poids moléculaire(Marker 3 : ÀDNAI EcoRI + Hindlll).
70
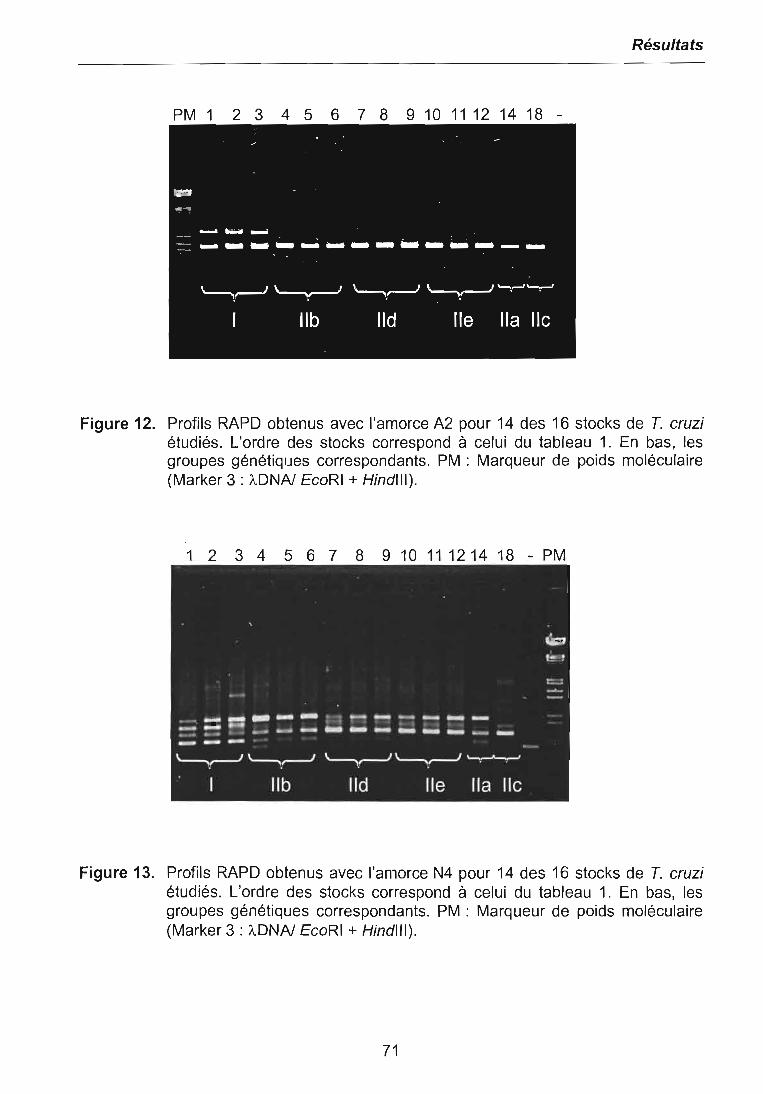
Résultats
PM 1 2 3 4 5 6 7 8 9 10 11 12 14 18 -
Figure 12. Profils RAPD obtenus avec l'amorce A2 pour 14 des 16 stocks de T. cruziétudiés. L'ordre des stocks correspond à celui du tableau 1. En bas, lesgroupes génétiques correspondants. PM : Marqueur de poids moléculaire(Marker 3 : À.DNN EcoRI + Hindlll).
1 2 3 4 5 6 7 8 9 10 111214 18 - PM
Figure 13. Profils RAPD obtenus avec l'amorce N4 pour 14 des 16 stocks de T. cruziétudiés. L'ordre des stocks correspond à celui du tableau 1. En bas, lesgroupes génétiques correspondants. PM : Marqueur de poids moléculaire(Marker 3 : À.DNN EcoRI + Hindi Il).
71
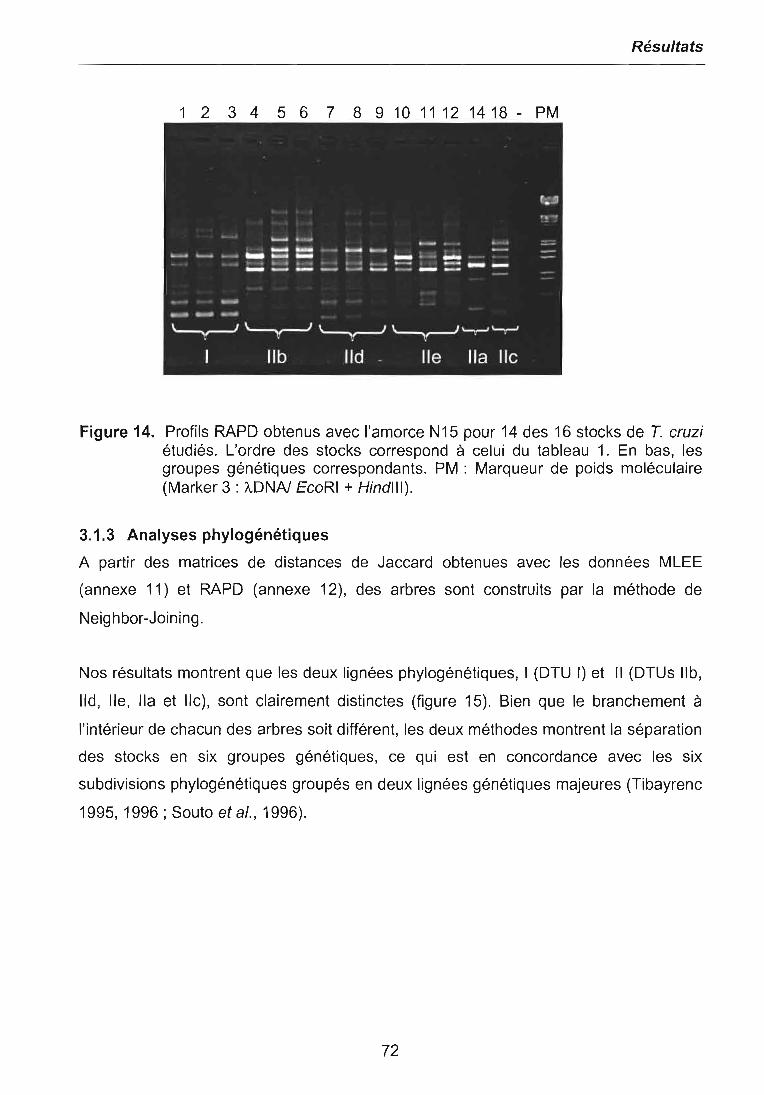
Résultats
1 2 3 4 5 6 7 8 9 10 11 12 14 18 - PM
Figure 14. Profils RAPD obtenus avec l'amorce N15 pour 14 des 16 stocks de T. cruziétudiés. L'ordre des stocks correspond à celui du tableau 1. En bas, lesgroupes génétiques correspondants. PM : Marqueur de poids moléculaire(Marker 3 : ÀDNA/ EcoRI + Hindlll).
3.1.3 Analyses phylogénétiques
A partir des matrices de distances de Jaccard obtenues avec les données MLEE
(annexe 11) et RAPD (annexe 12), des arbres sont construits par la méthode de
Neighbor-Joining.
Nos résultats montrent que les deux lignées phylogénétiques, 1(DTU 1) et Il (DTUs lib,
Ild, Ile, lia et Ile), sont clairement distinctes (figure 15). Bien que le branchement à
l'intérieur de chacun des arbres soit différent, les deux méthodes montrent la séparation
des stocks en six groupes génétiques, ce qui est en concordance avec les six
subdivisions phylogénétiques groupés en deux lignées génétiques majeures (Tibayrenc
1995, 1996 ; Souto et al., 1996).
72
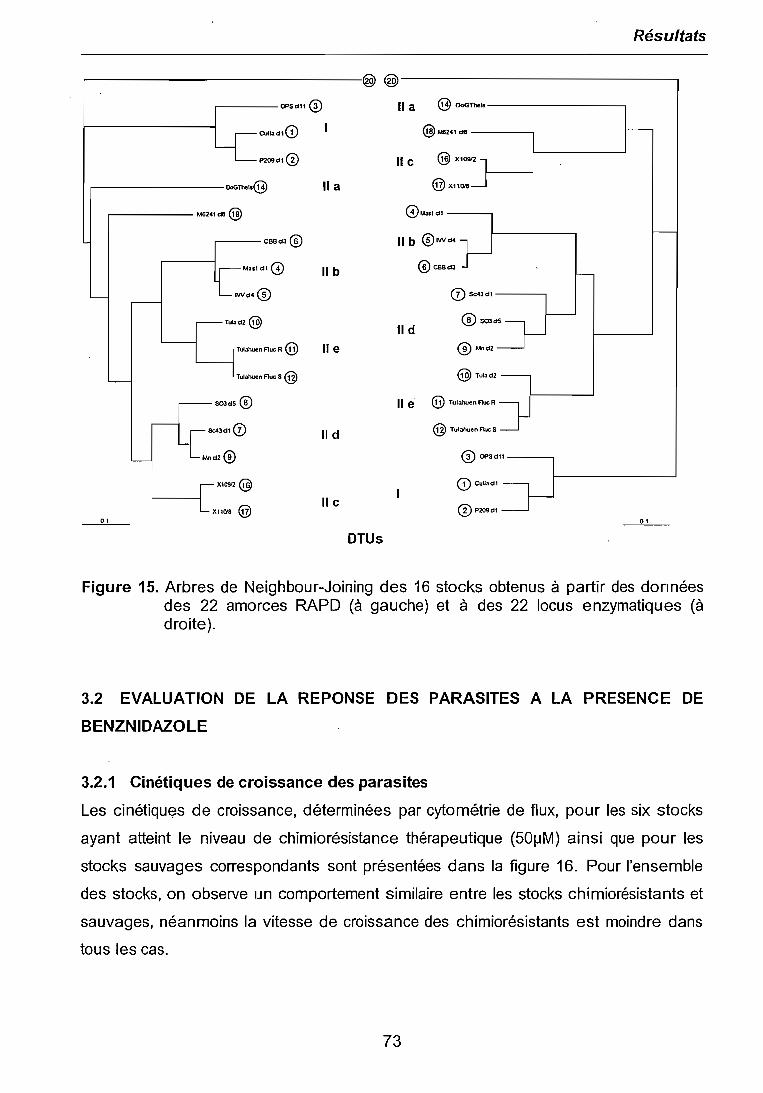
Résultats
@@>
OPSdll 0 lia @ OoGTheb
Culiad1G) @M6201d6
P209d'@ Ile @ Xl09/2
OoGTho'S@ lia @Xl1018
M0201,," @ @Mal1d1
CB8d3@ Il b @lWdO
MU:1d1@ lib @CBBd3
lWdO® CD Sc43dl
T.... d2@
IIdTutahuen Flue R @ Ile @Mnd2
Tulahuen Flue: S @ @ TuI.d2
S03dS@ Ile @ TulahuenFlucR
ScA3d·0 IId @ TulahuenRucS
Mnd2@ o OPSdll
XIO'l'2@ o Culiadl
XllC>8 @Ile o P209dl
o. o.
Drus
Figure 15. Arbres de l\Jeighbour-Joining des 16 stocks obtenus à partir des donnéesdes 22 amorces RAPD (à gauche) et à des 22 locus enzymatiques (àdroite).
3.2 EVALUATION DE LA REPONSE DES PARASITES A LA PRESENCE DE
BENZNIDAZOLE
3.2.1 Cinétiques de croissance des parasites
Les cinétiques de croissance, déterminées par cytométrie de flux, pour les six stocks
ayant atteint le niveau de chimiorésistance thérapeutique (50~M) ainsi que pour les
stocks sauvages correspondants sont présentées dans la figure 16. Pour l'ensemble
des stocks, on observe un comportement similaire entre les stocks chimiorésistants et
sauvages, néanmoins la vitesse de croissance des chimiorésistants est moindre dans
tous les cas.
73
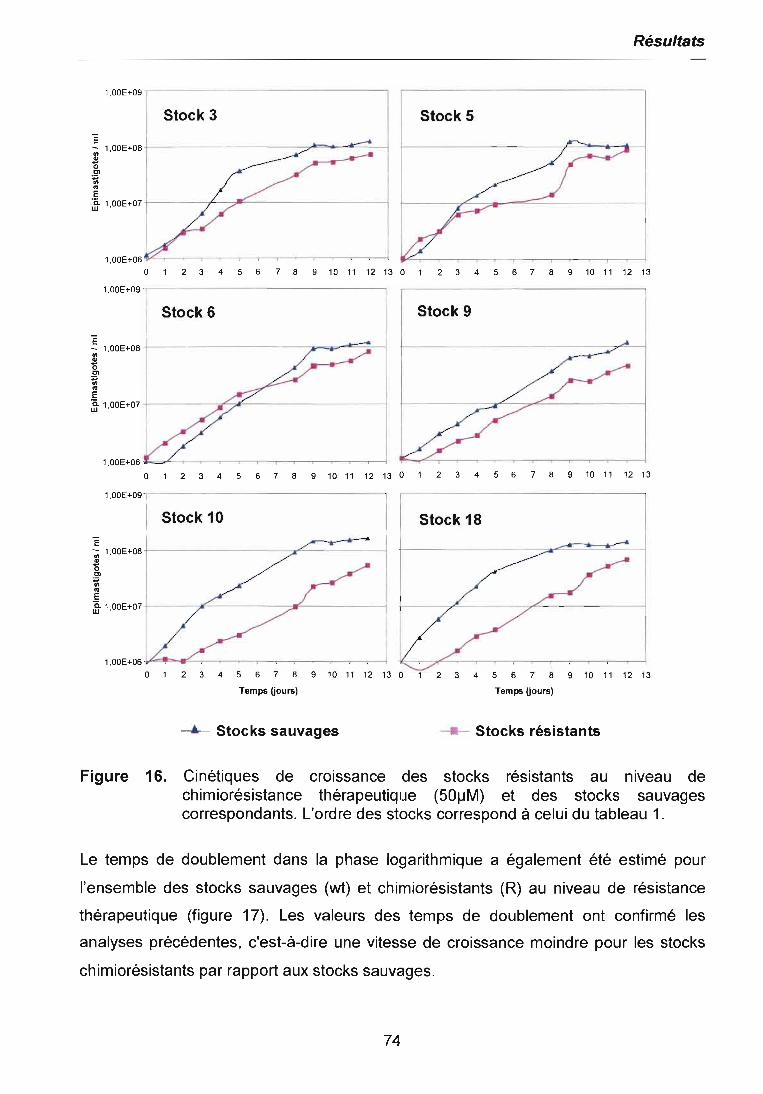
Résultats
1,OOE+09-.---- ----------.,
Stock 3 Stock 5
1,OOE+Ofi~--~-------'-----'-~~-'---~-'----'
o 1 2 3 4 5 fi 7 6 9 10 11 12 13 0 1 2 3 4 5 fi 7 6 9 10 11 12 13
1,OOE+09 .
Ë";;; 1 ,00E+OaCIl
oCl
~..E;fi- 1,OOE+07+----------t'---~-------______;
Stock 6 Stock 9
1.00E+Ofi ~./--r----,.---r---r---,-__..__,.__r .______I
o 1 2 3 4 5 fi 7 6 9 10 11 12 13 0 1 2 3 4 5 fi 7 6 9 10 11 12 13
~ 1.00E+Oa -I·-------",-.-;o~=:::.::o~ 1<II..~ 1,00E+07 t
1,OOE+09'1--------------
Stock 10 Stock 18Ë";;; 1,OOE+06·l--------
~Cl
~..E;fr 1,OOE+07-o.-·---7I'----------,;~----___l
1,OOE+Ofi -"k<,.....~.....----...,.---;- ---_--,----,----,----,----1
o 1 2 3 4 5 fi 7 6 9 10 11 12 13 0 1 2 3 4 5 6 7 6 9 10 11 12 13
~~~~ ~~~~
~ Stocks sauvages -- Stocks résistants
Figure 16. Cinétiques de croissance des stocks résistants au niveau dechimiorésistance thérapeutique (50IJM) et des stocks sauvagescorrespondants. L'ordre des stocks correspond à celui du tableau 1.
Le temps de doublement dans la phase logarithmique a également été estimé pour
l'ensemble des stocks sauvages (wt) et chimiorésistants (R) au niveau de résistance
thérapeutique (figure 17). Les valeurs des temps de doublement ont confirmé les
analyses précédentes, c'est-à-dire une vitesse de croissance moindre pour les stocks
chimiorésistants par rapport aux stocks sauvages,
74
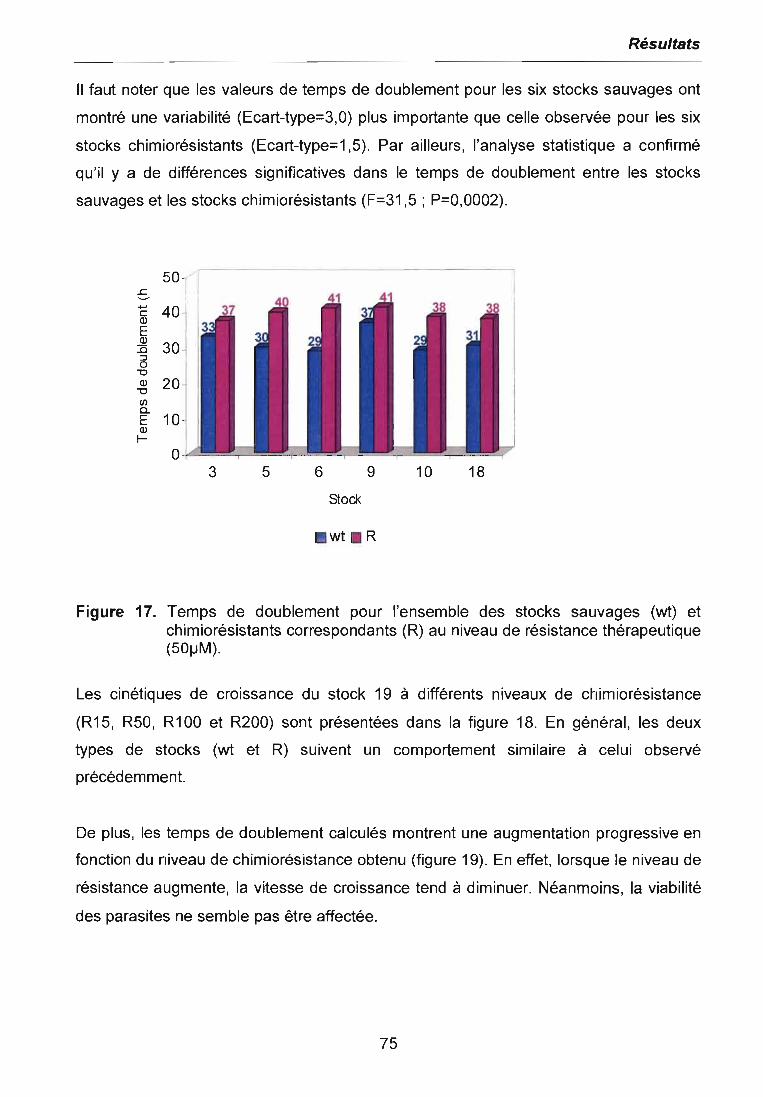
Résultats
Il faut noter que les valeurs de temps de doublement pour les six stocks sauvages ont
montré une variabilité (Ecart-type=3,O) plus importante que celle observée pour les six
stocks chimiorésistants (Ecart-type=1,5). Par ailleurs, l'analyse statistique a confirmé
qu'il y a de différences significatives dans le temps de doublement entre les stocks
sauvages et les stocks chimiorésistants (F=31,5 ; P=O,0002).
~.....--cQ)
EQ)
::i5:::J0"0Q) 20"0CIla.EQ)
1-
03 5 6 9
Stock
wt. R
10 18
Figure 17. Temps de doublement pour l'ensemble des stocks sauvages (wt) etchimiorésistants correspondants (R) au niveau de résistance thérapeutique(50IJM).
Les cinétiques de croissance du stock 19 à différents niveaux de chimiorésistance
(R15, R50, R100 et R200) sont présentées dans la figure 18. En général, les deux
types de stocks (wt et R) suivent un comportement similaire à celui observé
précédemment.
De plus, les temps de doublement calculés montrent une augmentation progressive en
fonction du niveau de chimiorésistance obtenu (figure 19). En effet, lorsque le niveau de
résistance augmente, la vitesse de croissance tend à diminuer. Néanmoins, la viabilité
des parasites ne semble pas être affectée.
75
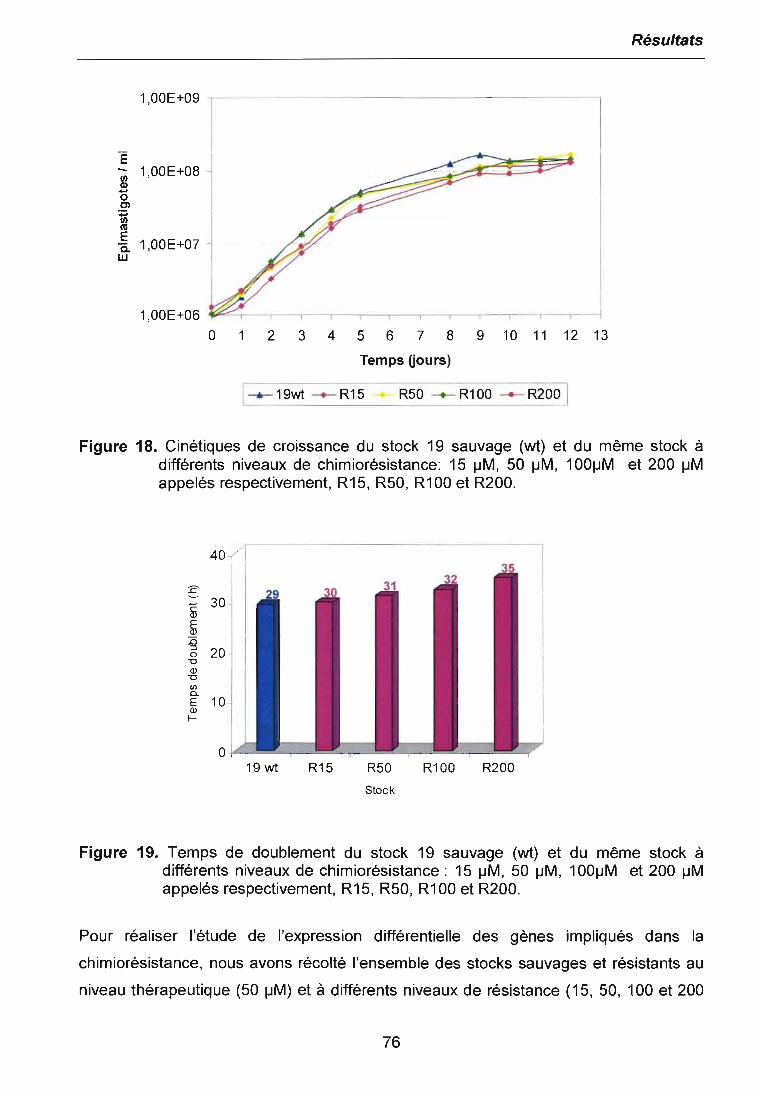
Résultats
1.00E+091
E l- 1,00E+08:g....oC)
~l'Cl
E;fr 1,00E+07 l
2 3 4 5 6 7 8 9 10 11 12 13
Temps üours)
1,00E+06 ......=-,---,-----------,-----,---~_r_-,_.____ , T
o
l--.-19wt -R15 R50 -R100 -~ool
Figure 18. Cinétiques de croissance du stock 19 sauvage (wt) et du même stock àdifférents niveaux de chimiorésistance: 15 IJM, 50 IJM, 100IJM et 200 IJMappelés respectivement, R15, R50, R100 et R200.
40-1 1
"P
ë 30Q)
EQ)
:0~ 200
""0Q)
""0(J)
a.E 10Q)
1-
019 wt R15 R50 R100 R200
Stock
Figure 19. Temps de doublement du stock 19 sauvage (wt) et du même stock àdifférents niveaux de chimiorésistance: 15 IJM, 50 IJM, 100IJM et 200 IJMappelés respectivement, R15, R50, R100 et R200.
Pour réaliser l'étude de l'expression différentielle des gènes impliqués dans la
chimiorésistance, nous avons récolté l'ensemble des stocks sauvages et résistants au
niveau thérapeutique (50 IJM) et à différents niveaux de résistance (15, 50, 100 et 200
76
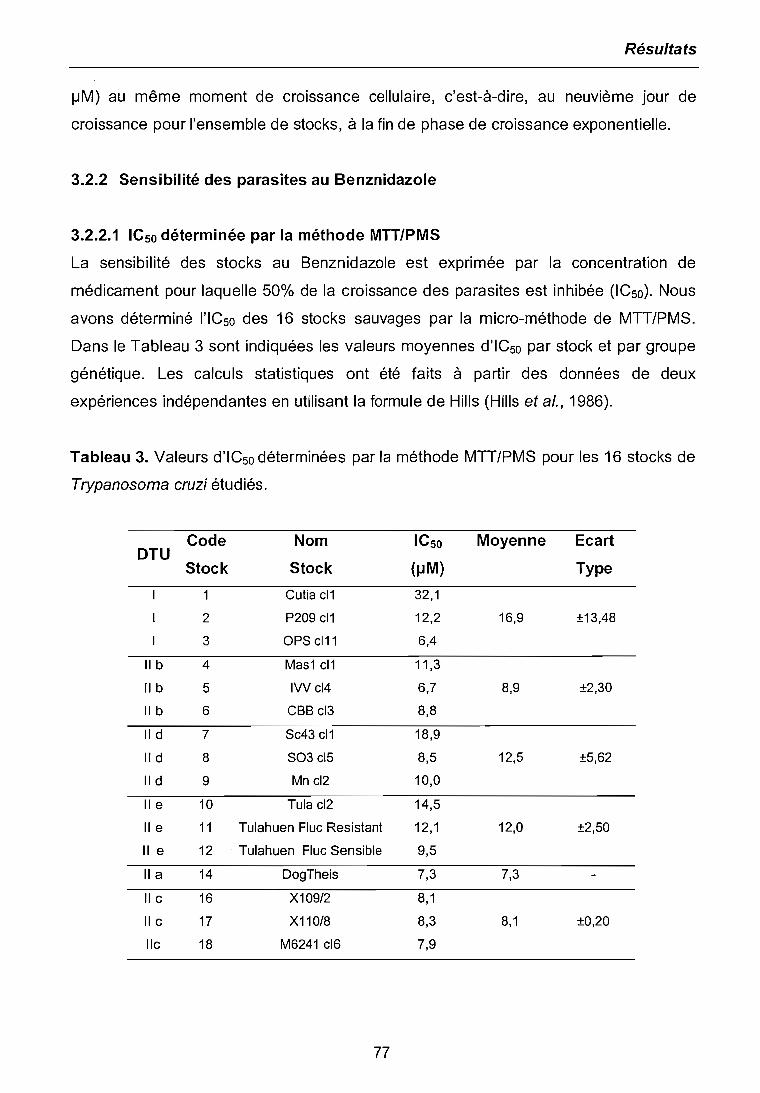
Résultats
!JM) au même moment de croissance cellulaire, c'est-à-dire, au neuvième jour de
croissance pour l'ensemble de stocks, à la fin de phase de croissance exponentielle.
3.2.2 Sensibilité des parasites au Benznidazole
3.2.2.1 ICso déterminée par la méthode MTT/PMS
La sensibilité des stocks au Benznidazole est exprimée par la concentration de
médicament pour laquelle 50% de la croissance des parasites est inhibée (ICso). Nous
avons déterminé l'ICso des 16 stocks sauvages par la micro-méthode de MTT/PMS.
Dans le Tableau 3 sont indiquées les valeurs moyennes d'lCso par stock et par groupe
génétique. Les calculs statistiques ont été faits à partir des données de deux
expériences indépendantes en utilisant la formule de Hills (Hills et al., 1986).
Tableau 3. Valeurs d'lCso déterminées par la méthode MTT/PMS pour les 16 stocks de
Trypanosoma cruz; étudiés.
Code Nom ICso Moyenne EcartDTU
Stock Stock (1-11\11) Type
Cutia el1 32,1
2 P209 el1 12,2 16,9 ±13,48
3 OPS el11 6,4
Il b 4 Mas1 c11 11,3
lib 5 IWel4 6,7 8,9 ±2,30
Il b 6 CSS c13 8,8
Il d 7 Se43 c11 18,9
IId 8 S03 c15 8,5 12,5 ±5,62
Il d 9 Mn c12 10,0
Il e 10 Tula c12 14,5
Ile 11 Tulahuen Flue Resistant 12,1 12,0 ±2,50
Il e 12 . Tulahuen Flue Sensible 9,5
lia 14 DogTheis 7,3 7,3
Ile 16 X109!2 8,1
Il e 17 X110!8 8,3 8,1 ±0,2ü
Ile 18 M6241 el6 7,9
77

Résultats
La comparaison statistique des valeurs d'lCso (ANOVA) entre les différents stocks n'a
pas révélé de différences significatives entre les différents groupes génétiques.
Néanmoins, nous avons classé le comportement des stocks en trois groupes similaires
hypothétiques.
Dans un premier groupe, on trouve le DTU 1qui diffère énormément des autres groupes
génétiques, ensuite un deuxième groupe qui regroupe les trois clades lib, IId et Ile, et
finalement un troisième groupe constitué par les DTU lia et Ile.
La comparaison entre les différents groupes génétiques a été réalisée par le test PLSD
de Fisher. L'analyse a confirmé qu'il n'y a pas de différences significatives des valeurs
moyennes d'ICso, ni entre les différents DTUs ni entre les deux lignées génétiques (1 et
Il ).
Une matrice de distances inter-stock calculée (différence entre les valeurs absolues)
sur les données /Cso pour les 16 stocks étudiés (annexe 13) a été réalisée dans le but
d'établir une possible corrélation entre la sensibilité des parasites au Benznidazole et
leur appartenance à un groupe génétique (voir chapitre 3.4).
3.2.2.2 ICso déterminée par cytométrie de flux
Pour trois stocks (3, 6 et 9) l'ICso a été également déterminée par cytométrie de "!lux,
dans le but de vérifier les valeurs obtenues par la micro-méthode MTT/PMS. L'ICso est
déterminée à 72, 96, 120 et 144 h après l'ajout de Benznidazole (tableau 4).
Pour l'ensemble des stocks, on observe que l'ICso diminue au fur et à mesure que le
temps d'exposition au Benznidazole augmente.
Nous avons ensuite comparé les valeurs d'lCso déterminées après 72 h d'exposition au
Benznidazole par les deux méthodes, MTT/PMS et cytométrie de flux (tableau 5).
78

Résultats
Tableau 4. Valeurs d'IC5D déterminées par cytométrie de flux à différents temps pour 3
des 16 stocks de Trypanosoma cruzi étudiés.
Stock
3
6
9
ICSD (IJM)
72 h 96 h 120 h 144 h
7,8 7,07 6,03 5,49
10,2 8,9 7,02 6,07
10,3 9,25 7,51 6,83
Tableau 5. Valeurs d'IC5D déterminées par les deux méthodes IV1TT/PMS et cytométrie
de flux, pour 3 des 16 stocks de Trypanosoma cruzi étudiés, à 72h.
ICso (IJM)
DTU StockMTT/PMS Cytométrie de flux
1 3 6,4 7,8
Il b 6 8,8 10,2
Il d 9 10,0 10,3
3.2.3 Niveaux de résistance obtenus avec le Benznidazole
L'induction de la résistance au Benznidazole a été réalisée avec six stocks (stocks 3, 5,
6,9, 10 et 18) jusqu'à l'obtention de chimiorésistance au niveau thérapeutique (50IJM).
Pour le stock 19, l'induction de la chimiorésistance a été réalisée jusqu'à 1OOIJM. Nous
avons rajouté à notre étude le stock 19 rendu chimiorésistant à 200IJM par Nirdé et al.
(1995).
Les stocks sauvages correspondants ont été cultivés en même temps en l'absence de
Benznidazole (groupe témoin).
Pour ces 6 stocks, l'étude de l'expression différentielle a été réalisée en considérant
uniquement le niveau de chimiorésistance thérapeutique, tandis que pour le stock 19,
quatre niveaux de chimiorésistance, 15R, 50R, 1OOR et 200R, ont été étudiés.
79

Résultats
Nous avons constaté que pour l'ensemble des stocks chimiorésistants, la résistance au
Benznidazole s'est avérée stable jusqu'à trois mois pour les niveaux étudiés et jusqu'à
six mois et demi pour un niveau intermédiaire (16f.lM), dans un milieu de culture sans
Benznidazole.
Notre étude montre qu'il est possible d'induire la résistance au Benznidazole de clones
appartenant à 5 des 6 groupes génétiques préalablement décrits. De plus, le niveau
maximal de résistance obtenu a été quatre fois supérieur à la dose du médicament
circulant chez un patient en cours de traitement, ceci pour le stock 19.
3.3 ETUDE DE L'EXPRESSION DIFFERENTIELLE PAR LA METHODE DERIVEE
DU RADES
L'étude de l'expression différentielle par la technique dérivée du RADES a d'abord été
réalisée au niveau de sensibilité naturelle des parasites au Benznidazole (à l'ICsD de
chacun des stock). Après avoir obtenu les stocks chimiorésistants, nous avons réalisé
l'étude au niveau thérapeutique (50 f-lM) pour six stocks et à différents niveaux (15, 50,
100 et 200f-lM) pour le stock 19.
Les bandes différentielles qui pourraient être associées à la résistance au Benznidazole
ont été sélectiol)nées à partir de l'analyse de l'ensemble des bandes RADES des six
stocks résistants au niveau thérapeutique (R50) et du stock 19 à différents niveaux de
résistance (R15, R50, R100 et R200). Certaines bandes différentielles de stocks
sauvages ont également été retenues. Pour l'ensemble des figures l'ordre des stocks
correspond à celui du tableau 1.
3.3.1 Etude de l'expression différentielle à l'ICso
L'amplification aléatoire de séquences exprimées (technique dérivée du RADES), a été
réalisée à l'aide de 22 amorces décamériques sur les 16 stocks en l'absence et en
présence de Benznidazole à la dose ICsD de chaque stock. L'ensemble des données
obtenues par la technique RADES en l'absence et en présence de Benznidazole à l'ICsD
est présenté respectivement dans les annexes 14 et 15. Les matrices des distances
correspondantes sont présentées dans les annexes 16 et 17.
80

Résultats
Les culots parasitaires des stocks cultivés en l'absence et en présence de
Benznidazole à l'IC50 (spécifique de chaque stock), ont été utilisés pour l'isolement de
l'ARNm, qui a été rétrotranscrit et amplifié initialement à l'aide des amorces SLc et dT18
et ensuite par amorces décamériques.
A partir des profils RADES, nous avons trouvé des amorces qui génèrent des profils
monomorphes et des amorces qui donnent des profils polymorphes (figure 20).
Néanmoins, la plupart d'entre elles montrent bien des profils polymorphes (A2, A3, A4,
A7, A9, A10, B1, B5, B11, B18, N4, N15, N18, R1, R2, R8, R10, U13, U16 et U19).
Parmi les 22 amorces testées, nous avons trouvé sept amorces qui ont révélé des
polymorphismes en rapport avec la présence du médicament, tandis que sept ont été
liées à l'absence de médicament. Ces amorces ont finalement permis l'amplification de
18 bandes différentielles liées soit à l'absence (9), soit à la présence (9) de
Benznidazole (tableau 6). Les bandes différentielles liées à la présence de
Benznidazole ont été retrouvées chez les stocks 3+, 4+, 7+, 11+ et 12+. Les bandes
différentielles liées à l'absence de Benznidazole, ont été trouvés chez les stocks 2, 3, 4,
7 et 11.
PM 7 7+ 8 8+ 9 9+ 1010+ 1111+1212+ - 7 7+ 8 8+ 9 9+ 10 10+ 11 11 + 12 12+ - PM
a b
Figure 20. Exemples de profils d'amplification RADES des stocks en l'absence et enprésence (+) de Benznidazole à l'IC5o. a, profils monomorphes (amorce F13);b, profil polymorphe (amorce A3). PM: Marqueur de poids moléculaire(Marker 3 : À-DNA/ EcoRI + Hindlll).
81

Résultats
Tableau 6. Bandes différentielles obtenues à l'ICso.
Benznidazole Stock Bande Amorce3+ 4 F133+ 2,4 N44+ 2 Ag
Présent 7+ 1 8511+ 1 81812+ 1,4 A212+ 1 A4
2 4 N182 4 U133 5 N44 2 A34 4 81
Absent 7 4 R111 4 A1011 4 N411 5 U13
+ : Stock cultivé en présence de Benznidazole à son ICso correspondant.
Les figures 21 à 23 présentent les profils obtenus pour cinq amorces différentes (N4,
A2, B18, R1 et U13).
1 1+ 2 2+ 3 3+ 4 4+ 5 5+ 6 6+ - PM 7 7+ 8 8+ 9 9+ 10 10+1111+1212+ - PM
Figure 21. Profils d'amplification RADES des stocks en l'absence et présence (+) deBenznidazole à l'ICso avec l'amorce N4. La présence de bandesdifférentielles est indiquée par des carrés rouges tandis que l'absence desbandes correspondantes est signalée par des flèches vertes. PM : Marqueurde poids moléculaire (Marker 3 : ÀDNAI EcoRI + Hindlll).
82
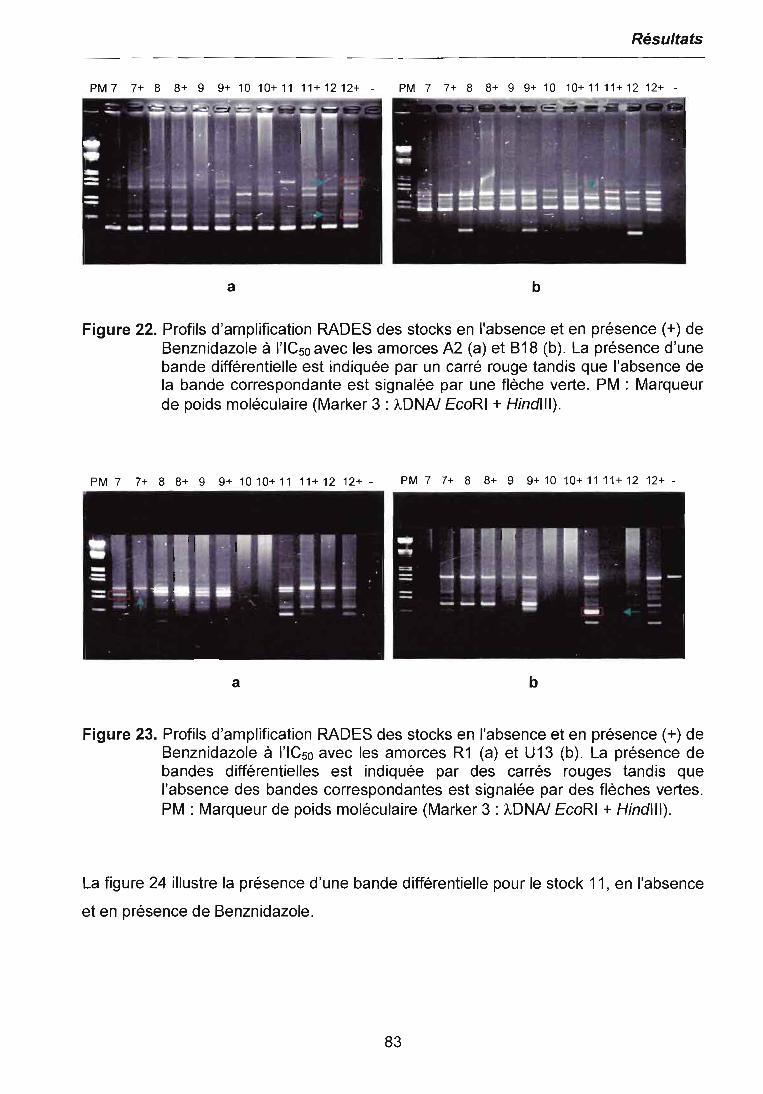
Résultats
PM7 7+ 8 8+ 9 9+ 10 10+1111+1212+ - PM 7 7+ 8 8+ 9 9+ 10 10+1111+1212+ -
a b
Figure 22. Profils d'amplification RADES des stocks en l'absence et en présence (+) deBenznidazole à l'ICSD avec les amorces A2 (a) et B18 (b). La présence d'unebande différentielle est indiquée par un carré rouge tandis que l'absence dela bande correspondante est signalée par une flèche verte. PM : Marqueurde poids moléculaire (Marker 3 : ADNAI EcoRI + Hindlll).
PM 7 7+ 8 8+ 9 9+ 1010+1111+1212+ - PM 77+ 8 8+ 9 9+1010+1111+1212+-
a b
Figure 23. Profils d'amplification RADES des stocks en l'absence et en présence (+) deBenznidazole à l'ICsD avec les amorces R1 (a) et U13 (b). La présence debandes différentielles est indiquée par des carrés rouges tandis quel'absence des bandes correspondantes est signalée par des flèches vertes.PM : Marqueur de poids moléculaire (Marker 3 : ADNAI EcoRI + Hindlll).
La figure 24 illustre la présence d'une bande différentielle pour le stock 11, en l'absence
et en présence de Benznidazole.
83

11 11+ 11 11+
Résultats
a b
Figure 24. Bandes différentielles RADES obtenues après la culture du stock 11 enl'absence de Benznidazole pour l'amorce N4 (a) et en présence (+) deBenznidazole à l'IC5D pour l'amorce B18 (b). La présence d'une bandedifférentielle est indiquée par des carrés rouges tandis que l'absence d'unebande correspondante est signalée par des flèches vertes.
3.3.2 Etude de l'expression différentielle au niveau de chimiorésistance
thérapeutique
Parmi les 22 amorces utilisées, seulement 8 (A2, A3, A7, 81, 85, N4, N18 et R2) ont
montré des bandes différentielles entre les stocks sauvages et les stocks
chimiorésistants au niveau thérapeutique (50IJM). Nous avons trouvé dix bandes
différentielles, parmi lesquelles trois sont exprimées seulement pour les stocks
chimiorésistants, quatre sont exprimées seulement pour les stocks sauvages et les
trois restantes sont exprimées tant pour les stocks sauvages que pour les stocks
chimiorésistants, mais surexprimées pour ces derniers (tableau 7). Les stocks qui ont
montré la présence de bandes différentielles sont les stocks 3, 5, 6 et 10, tandis que les
stocks 9 et 18 n'ont pas montré de différences d'expression entre chimiorésistants et
sauvages.
84

Résultats
Tableau 7. Bandes différentielles RADES obtenues à partir de l'ensemble des stocks
chimiorésistants au niveau thérapeutique (50IJM) et sauvages correspondants.
Bandes différentielles
Présentes chez les parasites résistants
Présentes chez les parasites sauvages
Présentes chez les parasites résistants et sauvages,
mais surexprimées chez les résistants
*Bandes sélectionnées pour le clonage et séquençage.
Stock
6R
10R
10R
5wt
5wt
6wt
10wt
3R
3R
6R
Bande
3
1
3*
1
3
4
4*
1*
3
Amorce
A3
B1
R2
B5
N18
N4
R2
A2
Al
Al
La figure 25 illustre des exemples de bandes différentielles qui sont exprimées soit par
le sauvage soit par le résistant ou présentes dans les deux, mais surexprimées pour le
stock chimiorésistant.
3wt 3R 5wt 5R 6wt 6R 9wt gR 10wt 10R 18w1 18R PM PM 3wt 3R 5wt SR 6wt 6R 9wt gR 10wt 10R 18w118R PM
a b
Figure 25. Profils d'amplification RADES avec les amorce R2 (a) et A7 (b) pourl'ensemble des stocks chimiorésistants au Benznidazole au niveauthérapeutique R (50 IJM) et sauvages correspondants (wt). La présence desbandes différentielles est indiquée par des carrés rouges tandis quel'absence ou la sous-expression des bandes correspondantes est signaléepar des flèches vertes. PM: Marqueur de poids moléculaire (Marker3 :ÀDNAI EcoRI + Hindlll).
85
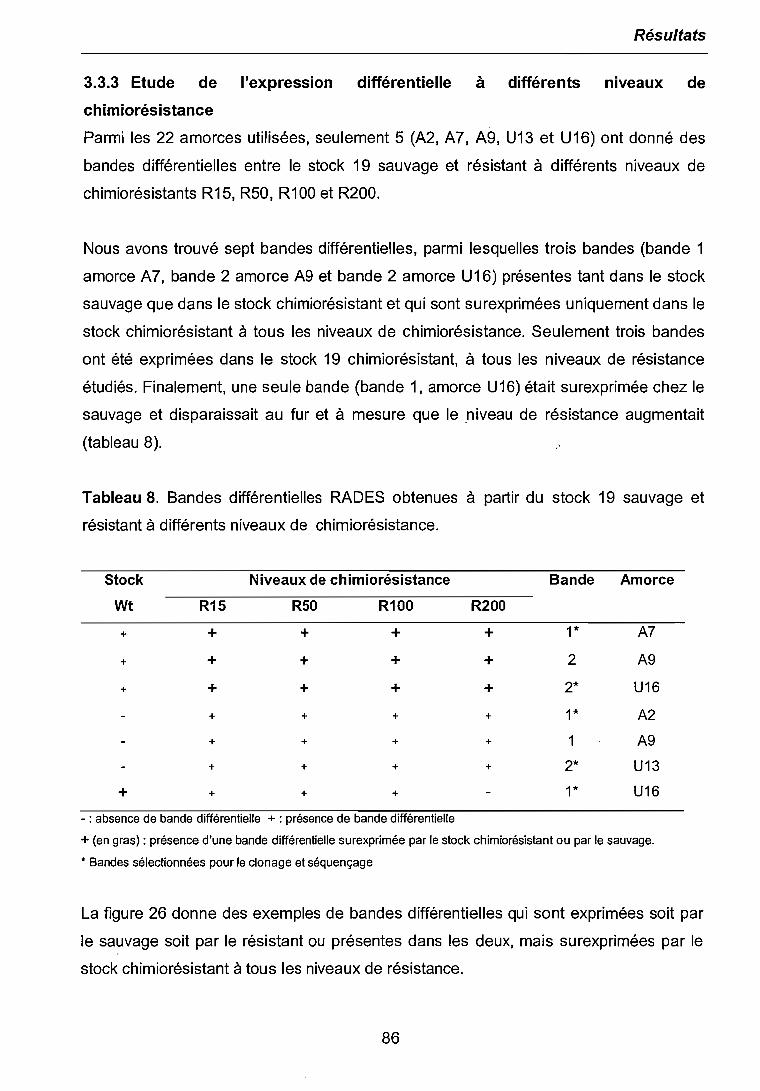
Résultats
3.3.3 Etude de l'expression différentielle à différents niveaux de
chimiorésistance
Parmi les 22 amorces utilisées, seulement 5 (A2, A7, A9, U13 et U16) ont donné des
bandes différentielles entre le stock 19 sauvage et résistant à différents niveaux de
chimiorésistants R15, R50, R100 et R200.
Nous avons trouvé sept bandes différentielles, parmi lesquelles trois bandes (bande 1
amorce A7, bande 2 amorce Ag et bande 2 amorce U16) présentes tant dans le stock
sauvage que dans le stock chimiorésistant et qui sont surexprimées uniquement dans le
stock chimiorésistant à tous les niveaux de chimiorésistance. Seulement trois bandes
ont été exprimées dans le stock 19 chimiorésistant, à tous les niveaux de résistance
étudiés. Finalement, une seule bande (bande 1, amorce U16) était surexprimée chez le
sauvage et disparaissait au fur et à mesure que le ,niveau de résistance augmentait
(tableau 8)."
Tableau 8. Bandes différentielles RADES obtenues à partir du stock 19 sauvage et
résistant à différents niveaux de chimiorésistance.
Stock Niveaux de chimiorésistance Bande Amorce
Wt R15 R50 R100 R200
+ + + + + 1* Al
+ + + + + 2 Ag
+ + + + + 2* U16
+ + + + 1* A2
+ + + + 1 Ag
+ + + + 2* U13
+ + + + 1* U16
- : absence de bande différentielle +: présence de bande différentielle
+ (en gras) : présence d'une bande différentielle surexprimée par le stock chimiorésistant ou par le sauvage.
* Bandes sélectionnées pour le clonage et séquençage
La figure 26 donne des exemples de bandes différentielles qui sont exprimées soit par
le sauvage soit par le résistant ou présentes dans les deux, mais surexprimées par le
stock chimiorésistant à tous les niveaux de résistance.
86

Résultats
PM 19w1 R15 R50 R100 R200 PM 19w1 R15 R50 R100 R200 PM 19W1 R15 R50 R100 R200
abc
Figure 26. Profils d'amplification RADES avec les amorces A7 (a), A9 (b) et U16 (c)pour le stock 19, sauvage et chimiorésistant à différents niveaux derésistance (R15, R50, R100 et R200). La présence des bandesdifférentielles est indiquée par des carrés rouges tandis que l'absence ou lasous-expression des bandes correspondantes est signalée par des flèchesvertes. PM: Marqueur de poids moléculaire (Marker 3 :ÀDNAI EcoRI +Hindi Il).
3.3.4 Clonage et séquençage des bandes différentielles sélectionnées
Nous avons isolé, cloné et séquencé les bandes différentielles tant au niveau
thérapeutique qu'à différents niveaux, représentant tous les cas de figure, c'est-à-dire, i)
bandes exprimées seulement par les stocks sauvages ou par les stocks
chimiorésistants, ii) bandes exprimées pour les deux mais surexprimées
préférentiellement par les stocks chimiorésistants (dans ce dernier cas, nous avons
également cloné la bande correspondante du stock sauvage). Les bandes différentielles
choisies sont présentées dans le tableau 9.
87
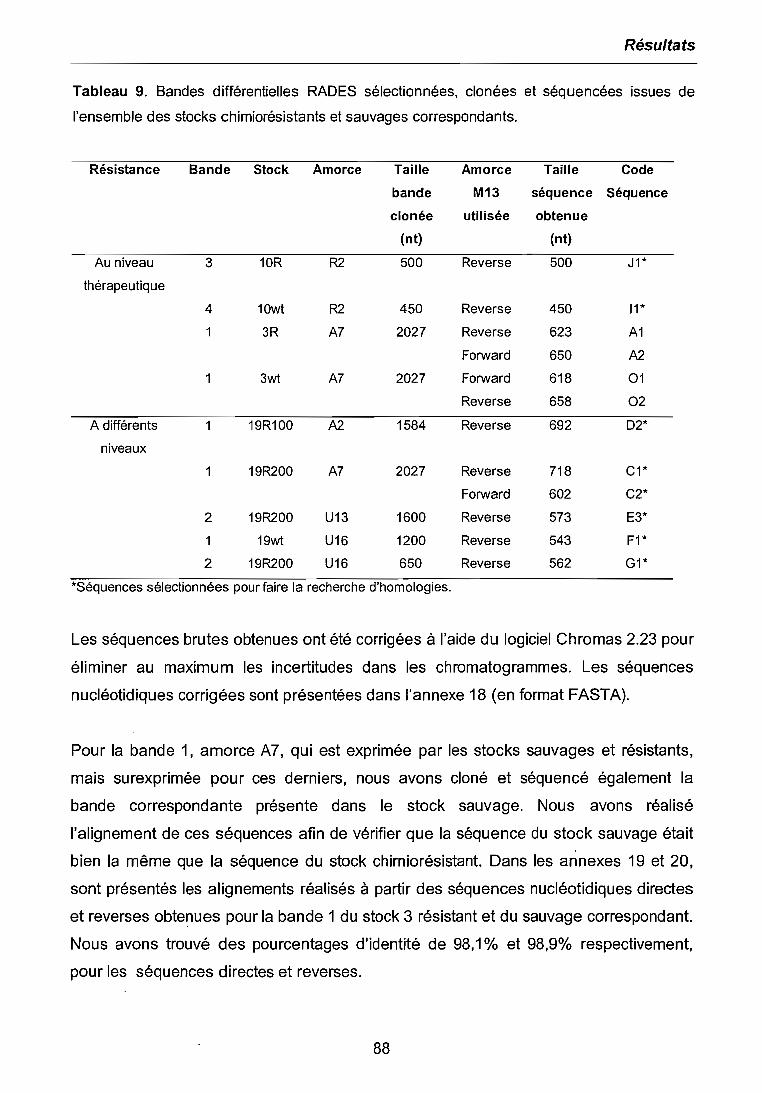
Résultats
Tableau 9. Bandes différentielles RADES sélectionnées, clonées et séquencées issues de
l'ensemble des stocks chimiorésistants et sauvages correspondants.
Résistance Bande Stock Amorce Taille Amorce Taille Code
bande M13 séquence Séquence
clonée utilisée obtenue
(nt) (nt)
Au niveau 3 10R R2 500 Reverse 500 J1*
thérapeutique
4 10wt R2 450 Reverse 450 11*
1 3R A7 2027 Reverse 623 A1
Forward 650 A2
1 3wt A7 2027 Forward 618 01
Reverse 658 02
A différents 1 19R100 A2 1584 Reverse 692 D2*
niveaux
1 19R200 A7 2027 Reverse 718 C1*
Forward 602 C2*
2 19R200 U13 1600 Reverse 573 E3*
1 19wt U16 1200 Reverse 543 F1*
2 19R200 U16 650 Reverse 562 G1*
*Séquences sélectionnées pour faire la recherche d'homologies.
Les séquences brutes obtenues ont été corrigées à l'aide du logiciel Chromas 2.23 pour
éliminer au maximum les incertitudes dans les chromatogrammes. Les séquences
nucléotidiques corrigées sont présentées dans l'annexe 18 (en format FA8TA).
Pour la bande 1, amorce Al, qui est exprimée par les stocks sauvages et résistants,
mais surexprimée pour ces derniers, nous avons cloné et séquencé également la
bande correspondante présente dans le stock sauvage. Nous avons réalisé
l'alignement de ces séquences afin de vérifier que la séquence du stock sauvage était
bien la même que la séquence du stock chirniorésistant. Dans les annexes 19 et 20,
sont présentés les alignements réalisés à partir des séquences nucléotidiques directes
et reverses obtenues pour la bande 1 du stock 3 résistant et du sauvage correspondant.
Nous avons trouvé des pourcentages d'identité de 98,1 % et 98,9% respectivement,
pour les séquences directes et reverses.
88

Résultats
Nous avons également aligné les séquences nucléotidlques directes et reverses
provenant de la bande 1 des stocks 3 et 19 chimiorésistants (annexes 21 et 22), car à
partir de la comparaison des poids moléculaires de chacune de ces deux bandes, nous
avons pensé qu'elles pourraient correspondre à la même bande. En effet, tant les
séquences directes comme les séquences reverses de la bande 1, provenant de
différents stocks chimiorésistants (3 et 19), ont des pourcentages d'identité très forts
(97,5% et 99,5%, respectivement).
Nous avons recherché les homologies des séquences nucléotidiques sélectionnées à
l'aide des algorithmes BLASTN et BLASTX du NeBI (BLAST with eukaryotic genomes).
Les valeurs de probabilité E ~1-5 et E~1-4 ont été considérées comme significatives,
respectivement pour BLASTN et BLASTX.
Les séquences d'acides aminés correspondantes (annexe 23) déduites à l'aide du
programme « Six Frame Translation of Sequence» sur le site BeM Search Launcher,
ont également été utilisées pour faire la recherche d'homologies protéiniques à l'aide de
l'algorithme BLASTP sur le site ExPASy Molecular Biology Server.
Le tableau 10 présente les résultats les plus importants concernant les homologies (en
utilisant BLASTN, BLASTX et BLASTP) rencontrées pour les bandes différentielles
RADES obtenues à partir des stocks chimiorésistants et sauvages.
Bien que les séquences nucléotidiques obtenues correspondent bien aux séquences
décrites chez T. cruzi (homologies BLASTN), seulement deux séquences d'acides
aminés correpondantes (D2 et F1) ont montré des scores importants (BLASTP) avec
des protéines déjà décrites dans les banques de données. A partir des résultats
obtenus pour chacune des séquences, nous avons sélectionné ceux qui ont donné le
score le plus fort et la valeur de probabilité E la plus significative, bien que ces deux
séquences aient montré des identités avec d'autres protéines hypothétiques et
putatives avec des scores plus faibles.
89
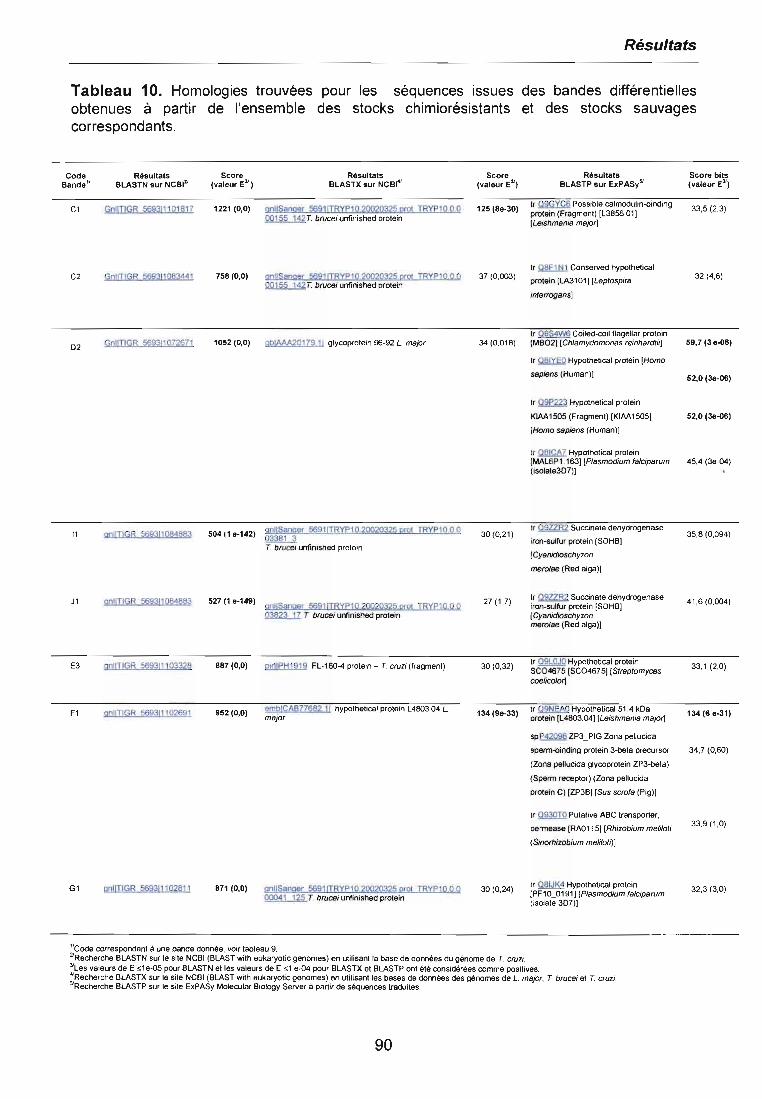
Résultats
issues des bandes différentielleschimiorésistants et des stocks sauvages
Tableau 10. Homologies trouvées pour les séquencesobtenues à partir de l'ensemble des stockscorrespondants.
Code Résultats Score Résultats Score Résultats Score bitsBande11 BLASTN sur NeBI" (valeur e) BLASTX sur NCBr" (valeur E") BLASTP sur ExPASy" (valeur E")
C1 GnlmGR 569311101817 1221 (0,0) gnllSq.MI1r 56911TRYP10 20020325 prat TRYPIO 0 0 125 (Se-3D)tr~ Possible calmodulin-binding
33,5 (2,3)prolein (Fragmenl) [L3856.01]
00155 142T. brucei unfinished protein[Leishmania major)
C2 Gn!!TIGR 569311083441 758 (0,0) "ni/Sanger 5691!TRYP10 29020325 prol TRYP10.0 0~T. brucei unfinished protein
37 (0,003)Ir~ Conserved hypolhetical
protein lLA3101] [Leptospire
inte"ogens]
32 (4,6)
02 GoilTIGR 569311 072671 1052 (0,0) gb~179 1J glycoprotein 96-92 L major 34 (0,018)tr gBS4W6 Coiled-coil f1egellar protein[MB02] [Chlamydomonas reinherdtit]
Ir Q§Jïlill Hypolhelical protein [Homo
sapiens (Humen)J
59,7 (3 e-08)
52,0 (30-06)
tr~ Hypolhelical protein
K1AA1505 (Fragment) [KIAA15051
[Homo saplens (Human))
52,0 (3e-06)
Ir~ Hypothelicel protein[MAL8Pl.I63] [Plesmodium falciperum(isolate307)J
45,4 (30-04)
Il gnllnGR 569311()84883 504 Il e-142) gnllSanger 569t1TRyplO 20020325 prol TRYP10 0 030(0,21)
Ir~ Succinale dehydrogenase35,8 (0,094)03381 3 iron-sulfur prolein [SOHBI
T. bruoei unfinished protein[Cyanidioschyzon
merolee (Red alga)J
Jl anliTIGR 569311084819 527 (1 e-149) 27 (1,7) Ir~ Succinete dehydrogenase 41,6 (0,004)gnllS!lQlJel 569HTRYP10.20020325 \!fol TRYP10.00 iron-sulfur prolein (SOHBI03823 17 T. orucei unfinished prolein [Cyanidioschyzon
maro/ae (Red alga))
E3 QnlmGR 569311103328 887 (0,0) pjf!!PH1919 FL-160-4 protein - T cruzi (fragmenl) 30 (0,32)Ir 09LOJO Hypothetical protein
33,1 (2,0)SC04675 (SC04675] (Streptomycescoe/icolorj
F1 onlmGR 569311102691 952(0,0) PmbICAB77682 11 hypothelical protein L4803.04 L. 134 (ge-33) Ir~ Hypothelical 51.4 kOa 134 (6 a-31)major prolein [L48D3.04] [Leishmenie majorj
SpP42098 ZP3_PIG Zona pellucida
sperm-binding protein 3-beta precursor 34,7 (0,60)
(Zona pellucida glycoprotein ZP3-bele)
(Sperm receptor) (Zona pellucida
protein C) [ZP3BJ (Sus scrofa (Pig)1
Ir Qll;ïQIQ Pulalive ABC lransponer,
permease [RA0115] [Rhizobium meliloti33,9 (1,0)
(Slnarhizobium melilot/]]
G1 gnlmGR 5693111 02811 871 (0,0) gnllSaooar 5!591ITRYP10 20020325 0f9! TRVPlO 0 0 30 (0,24) Ir Q81JK4 Hypothetical prolein 32,3 (3,0)00041 125 T. Orucei unfinished prolein [PF10_0191) [Plasmodium 'elciperum
(isolate 307)}
lICode correspondant à une bande donnée, voir tableau 9.21Recherche BLASTN sur le site NCBI (BLAST with eukaryotic genomes) en utilisant la base de domées du génome de T. cruzi."Les valeurs de E s1 e-OS pour BLASTN et les valeurs de E s1 e-04 pour BLASTX et BLASTP ont été considérées comme positives."Recherche 8LASTX sur le sile NCBI (BLAST wi1h eukaryotic genomes) en utilisant les bases de domées des génomes de L. major, T. brucei et T. cruzi.51Recherche BLASTP sur le sile ExPASy Molecular Biology Server à panir de séquences traduites.
90

Résultats
La première séquence, 02 (stock 19 chimiorésistant à 100 IJM, bande 1 obtenue avec
l'amorce A2) correspond à une protéine flagellaire de Ch/amydomonas reinhardtii (E=3
e-08). L'alignement entre ces deux séquences a montré un pourcentage d'identité de
27% sur 159 acides aminés (annexe 24).
La deuxième séquence, F1 (stock 19wt, bande 1 obtenue avec l'amorce U16)
correspond à une protéine hypothétique de L. major (E=6 e-31). L'alignement entre ces
deux séquences a montré un pourcentage d'identité de 40% sur 169 acides aminés
(annexe 25).
De manière intéressante, nous avons trouvé (en utilisant BLASTX) que la séquence C1
(stock 19 chimiorésistant, bande 1 obtenue avec l'amorce A7) a une identité de 37%
(sur 188 acides aminés) avec une protéine qui est en train d'être séquencée chez T.
brucei (E=8 e-30). De plus, elle a montré une identité faible, en utilisant BLASTX et
BLASTP, avec une possible calmoduline de L. major.
Les séquences 1 et J (bandes 4 et 3, respectivement, amorce R2) ont été exprimées de
manière différentielle par le stock 10. Bien que les deux séquences nucléotidiques
n'aient montré que 46,4% d'identité (sur 450 nucléotides), l'alignement des séquences
d'acides aminés prédites ont montré une identité de 74,5% (sur 146 acides aminés). De
manière intéressante, les deux séquences ont montré une identité faible avec une
succinate déshydrogénase de Cyanidioschyzon mere/ae (une algue rouge). Elles ont
montré également la présence d'un domaine conservé FrdB (COG0479, Succinate
déshydrogénase/ fumarate réductase, sous-unité protéique Fe-S). De plus, la recherche
d'ESTs (pour Expressed Sequence-Tags) chez T. cruzi a donné un score de 209 avec
un niveau de significativité de 1,9 e-25 avec le TENU2670, ceci étant similaire à la
succinate déshydrogenase Fe-S (dhdB) des levures (Porcel et a/., 2000).
3.4 ANALYSES DE CORRELATION ENTRE VARIABLES
Afin d'établir des relations entre la sensibilité des différents stocks au Benznidazole
(ICso) et leurs distances génétiques (MLEE, RAPD, RADES) nous avons construit des
matrices de distances qui ont été comparées par des analyses de corrélation. Les
91

Résultats
matrices de distances pour les données MLEE, RAPD, ICso, RADES (en absence et en
présence de Benznidazole à l'ICso) sont présentées respectivement dans les annexes
11,12,13,16 et 17.
Les analyses de corrélation ont été effectuées par le test non-paramétrique de Mantel
entre distances génétiques (MLEE, RAPD et RADES) et distances biologiques (iCso)
sur les 16 stocks étudiés (tableau 11). Les valeurs de corrélations entre les différents
types de distances génétiques (MLEE, RAPD et RADES) ont été très hautement
significatives. En revanche, les corrélations entre toutes ces distances génétiques et les
distances biologiques (ICso) ont été beaucoup plus faibles et non significatives.
Tableau 11. Résultats des différents tests de Mantel entre distances génétiques
(RAPD, MLEE et RADES, en absence et en présence de Benznidazole à l'ICso) et
sensibilité des parasites au Benznidazole (IC50) pour l'ensemble de 16 stocks étudiés.
RADES en RADES enICso RAPD MLEE présence de absence de
Benznidazole BenznidazoJeCoef. Coef. Coef. Coef. Coef.
Correl. P Correl. P Correl. P Correl. P Correl. P
ICso
RAPD 0,101 0,229
MLEE 0,201 0,158 0,729 0,0001***
RADESen
-0,063 0,593 0,822 0,0001*** 0,465 0,0023***présence
RADESen
-0,044 0,684 0,789 0,0001*** 0,677 0,0001*** 0,789 0,0001***absence
Niveau de significativilé 1%
3.5 ETUDE DE L'EXPRESSION DES GENES SPECIFIQUES
L'étude de l'expression des gènes spécifiques a été réalisée à travers l'amplification de
produits obtenus par la technique RT-PCR en utilisant les amorces SLc et dT18•
92

Résultats
Afin d'analyser les possibles différences d'expression entre stocks sauvages et
résistants des gènes candidats étudiés (tcsod-A, tcsod-B, tccp1 et tcpgp2) , nous avons
préalablement établi les conditions optimales de leur amplification en utilisant le gène
constitutif de l'actine, ceci pour calibrer la quantité d'ADNe avant l'amplification des
gènes candidats aux conditions optimales pour chacun.
La figure 27 présente le profil d'amplification du gène de l'actine standardisé pour
l'ensemble des stocks résistants et sauvages. Les bandes obtenues après PCR des
différents stocks ne montrent pas de différence d'expression évidente pour un même
niveau de chimiorésistance, sauf pour le stock 6wt pour lequel la bande a été très
faible, problème peut être lié à la technique d'amplification ou à l'échantillon.
PM 3wt 3R 5wt 5R 6wt 6R 9wt 9R 10wt 10R 18wt 18R PMl9wtR15 R50 R100 R200
334 pb
Figure 27. Profils d'amplification du gène actine pour les stocks sauvages (wt) etchimiorésistants au niveau thérapeutique (R) et à différents niveaux dechimiorésistance pour le stock 19 (R15, R50, R100 et R200). L'ordre desstocks correspond à celui du tableau 1. PM : Marqueur de poids moléculaire(100 bp DNA Ladder Plus).
3.5.1 tcsod-A et tcsod-B
L'amplification des gènes tcsod-A et tcsod-B a été mise au point en utilisant l'ADN total
et l'ADNe des stocks étudiés. La figure 28 illustre les produits d'amplification obtenus
avec l'ADNe pour les deux gènes.
La taille des fragments amplifiés des gènes tcsod-A et tcsod-B est bien en accord avec
la taille attendue (302 pb et 315 pb, respectivement). Pour les deux gènes, il n'y a pas
eu de différence significative d'expression entre les stocks sauvages et résistants tant
93
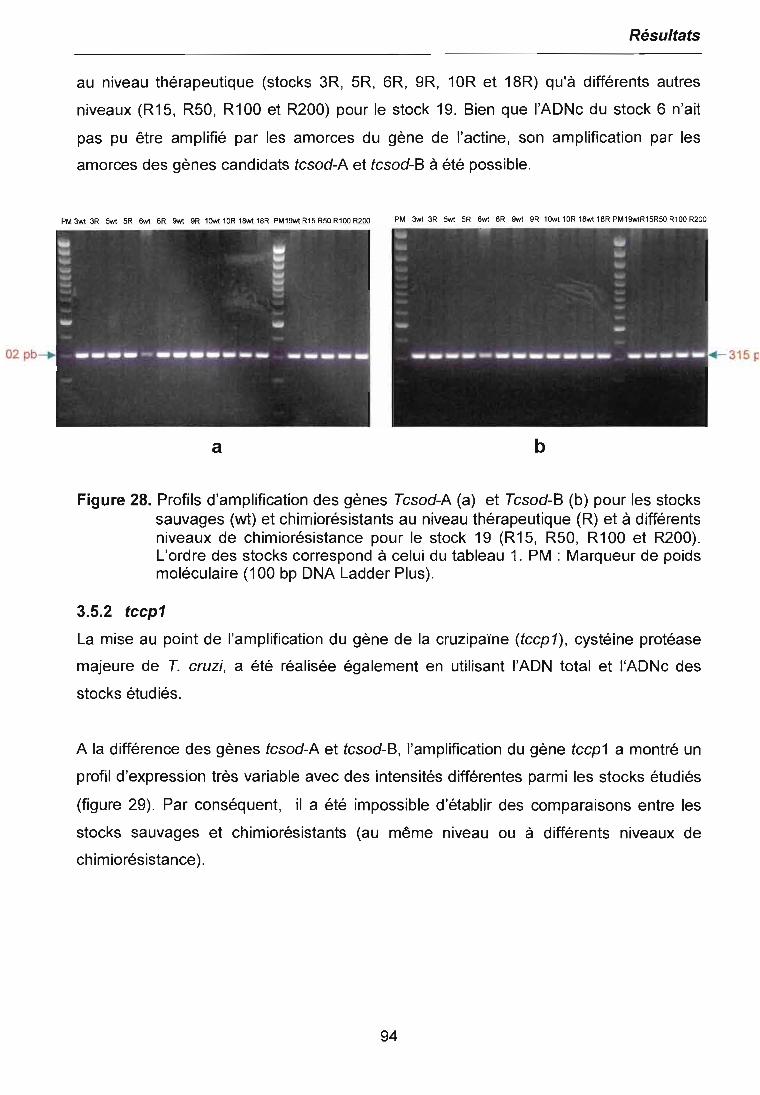
Résultats
au niveau thérapeutique (stocks 3R, 5R, 6R, 9R, 10R et 18R) qu'à différents autres
niveaux (R15, R50, R100 et R200) pour le stock 19. Bien que l'ADNe du stock 6 n'ait
pas pu être amplifié par les amorces du gène de l'actine, son amplification par les
amorces des gènes candidats tcsod-A et tcsod-B à été possible.
PM 3wt 3R Swt 5R 6wt 6R 9wt gR lOwt 10R 18wt 1eR PMl9wt R15 R50 Rl00 R200 PM 3wt 3R 5wt 5R 6wt 6R 9wt gR lOwt 10R lewt 1eR PMl9wtR15R50 R100 R200
02 pb .-3151=
a b
Figure 28. Profils d'amplification des gènes Tcsod-A (a) et Tcsod-B (b) pour les stockssauvages (wt) et chimiorésistants au niveau thérapeutique (R) et à différentsniveaux de chimiorésistance pour le stock 19 (R15, R50, R100 et R200).L'ordre des stocks correspond à celui du tableau 1. PM : Marqueur de poidsmoléculaire (100 bp DNA Ladder Plus).
3.5.2 tccp1
La mise au point de l'amplification du gène de la cruzipaïne (tccp1), cystéine protéase
majeure de T. cruzi, a été réalisée également en utilisant l'ADN total et l'ADNe des
stocks étud iés.
A la différence des gènes tcsod-A et tcsod-B, l'amplification du gène tccp1 a montré un
profil d'expression très variable avec des intensités différentes parmi les stocks étudiés
(figure 29). Par conséquent, il a été impossible d'établir des comparaisons entre les
stocks sauvages et chimiorésistants (au même niveau ou à différents niveaux de
chimiorésistance).
94
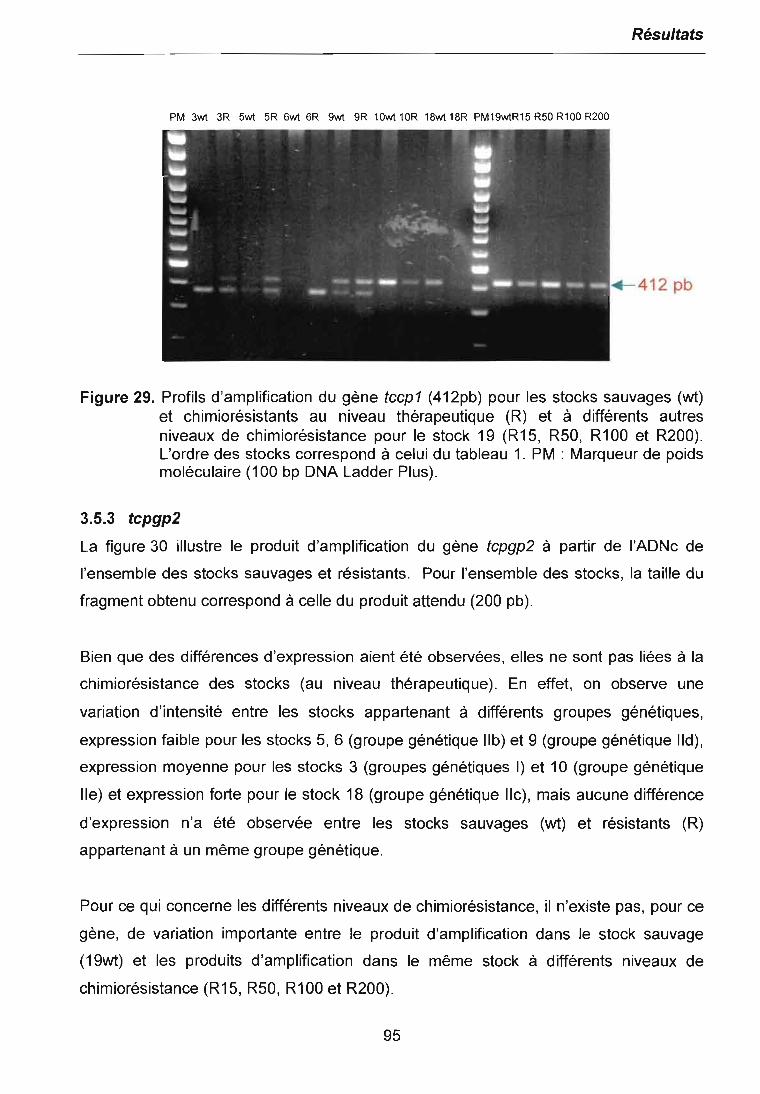
Résultats
PM 3wt 3R 5wt 5R 6wt 6R 9wt 9R 10wt 10R 18wt 18R PM19wtR15 R50 R100 R200
+-412 pb
Figure 29. Profils d'amplification du gène tccp1 (412pb) pour les stocks sauvages (wt)et chimiorésistants au niveau thérapeutique (R) et à différents autresniveaux de chimiorésistance pour le stock 19 (R15, R50, R100 et R200).L'ordre des stocks correspond à celui du tableau 1. PM : Marqueur de poidsmoléculaire (100 bp DNA Ladder Plus).
3.5.3 tcpgp2
La figure 30 illustre le produit d'amplification du gène tcpgp2 à partir de l'ADNe de
l'ensemble des stocks sauvages et résistants. Pour l'ensemble des stocks, la taille du
fragment obtenu correspond à celle du produit attendu (200 pb).
Bien que des différences d'expression aient été observées, elles ne sont pas liées à la
chimiorésistance des stocks (au niveau thérapeutique). En effet, on observe une
variation d'intensité entre les stocks appartenant à différents groupes génétiques,
expression faible pour les stocks 5, 6 (groupe génétique lib) et 9 (groupe génétique IId),
expression moyenne pour les stocks 3 (groupes génétiques 1) et 10 (groupe génétique
Ile) et expression forte pour le stock 18 (groupe génétique Ile), mais aucune différence
d'expression n'a été observée entre les stocks sauvages (wt) et résistants (R)
appartenant à un même groupe génétique.
Pour ce qui concerne les différents niveaux de chimiorésistance, il n'existe pas, pour ce
gène, de variation importante entre le produit d'amplification dans le stock sauvage
(19wt) et les produits d'amplification dans le même stock à différents niveaux de
chimiorésistance (R15, R50, R100 et R200).
95
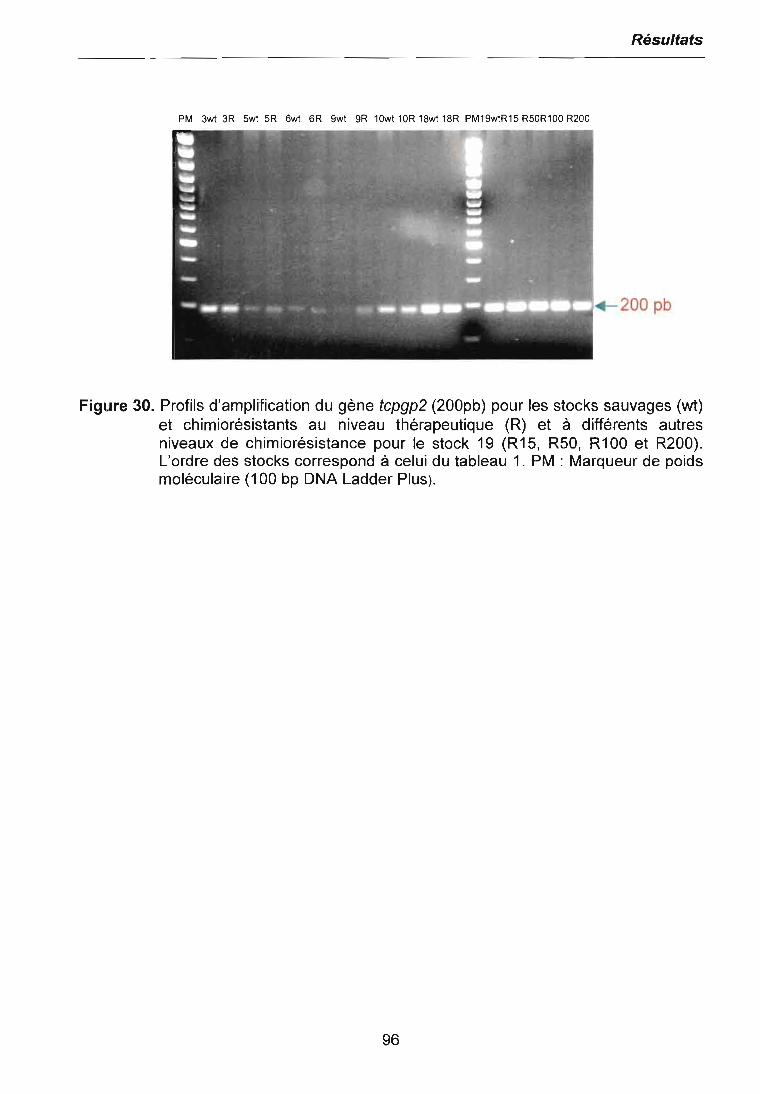
Résultats
PM 3wt 3R 5wt 5R 6wt 6R 9wt 9R 10wt 10R 18wt 18R PM19wtR15 R50R100 R200
200 pb
Figure 30. Profils d'amplification du gène tcpgp2 (200pb) pour les stocks sauvages (wt)et chimiorésÎstants au niveau thérapeutique (R) et à différents autresniveaux de chimiorésistance pour le stock 19 (R15, R50, R100 et R200).L'ordre des stocks correspond à celui du tableau 1. PM : Marqueur de poidsmoléculaire (100 bp DNA Ladder Plus).
96

DISCUSSION

Discussion
4 CHAPITRE IV. DISCUSSION
4.1 COMPORTEMENT DES PARASITES VIS A VIS DU BENZNIDAZOLE
4.1.1 Sensibilité des parasites au Benznidazole
La réponse des patients au traitement de la maladie de Chagas est très variable. Cette
variabilité a été attribuée en partie à la sensibilité des parasites aux médicaments, ainsi
qu'aux interactions entre les parasites et la composante immunologique de l'hôte
(Michailowsky et al., 1998). En effet, beaucoup d'auteurs ont décrit des sensibilités
différentes des parasites vis-à-vis des médicaments utilisés pour le traitement de la
maladie de Chagas (Brener et al., 1976; Filardi et Brener, 1982 ; Andrade et al., 1985 ;
Filardi et Brener, 1987 ; Neal et van Bueren, 1988 ; Toledo et al., 1997).
De plus, une association entre sensibilité des parasites aux médicaments et origine
géographique des souches a également été rapportée (Andrade et al., 1985 ; Filardi et
Brener, 1987). En effet, dans certaines zones du Brésil où la maladie avait été
récemment introduite, neuf souches ont été décrites comme particulièrement
résistantes au Nifurtimox et au Benznidazole. Dans d'autres zones du Brésil et de
l'Argentine, les souches ont montré des niveaux de sensibilité plus importants (98 à
100% de guérison chez des animaux infectés). Cette variation dans la sensibilité des
parasites aux médicaments en rapport avec l'origine géographique des souches
pourrait expliquer les taux de guérison plus élevés en Ârgentine comparés à ceux du
centre et du sud du Brésil (Filardi et Brener, 1987). La résistance naturelle des souches
aux dérivés nitrés est aussi un facteur qui a été proposé pour expliquer les faibles taux
de récupération chez certains patients chagasiques.
Concernant le Benznidazole, nous avons pu observer des variations de
chimiosensibilité naturelle des souches allant de 1 à 5 fois au sein d'une même DTU.
Nous confirmons les observations rapportées par Andrade (Andrade et al., 1985). En
effet, dans leurs études ces auteurs montrent que la chimiothérapie au Benznidazole et
au Nifurtimox chez des souris infectées par différents types de souches de T. cruzi a
présenté une grande gamme de sensibilité. Les souches dites de type l, caractérisées
par leur rapidité d'infestation chez la souris, une parasitémie et une mortalité élevées
vers le 9ème ou 10ème jour et Lin macrophagotropisme, ont présenté une grande
97

Discussion
sensibilité (50 à 100% de guérison avec le Benznidazole et 35 à 87% avec le
Nifurtimox). Les souches de type Il, caractérisées par Llne parasitémie augmentant du
12éme au 20éme jour de l'infestation, un faible taux de mortalité et un tropisme
myocardique, ont présenté une sensibilité moyenne à élevée et surtout très inégale en
fonction des souches. Les souches de type III, caractérisées par une parasitémie à
installation lente, atteignant des niveaux élevés seulement 20 à 30 jours après
l'inoculation, et par un tropisme pour les muscles squelettiques, ont présenté une forte
résistance à ces deux médicaments, avec un taux de guérison entre 0 et 20%
seulement. Cependant dans cette étude, aucune donnée n'était disponible concernant
l'accessibilité de la drogue dans les différents compartiments tissulaires, ni sur les taux
réels de drogue dans les différents tissus, en particulier ceux où étaient localisés les
parasites, une lacune qui pourrait moduler l'interprétation des résultats de ces auteurs.
Le modèle d'évolution clonale prépondérante des populations naturelles de T. cruzi
(Tibayrenc et al., 1986 ; Tibayrenc et Ayala, 1988), prédit une évolution non
indépendante entre diversité génétique et diversité biologique du parasite, ce dernier
paramètre pouvant inclure la sensibilité aux médicaments. Aussi, différents travaux ont
été réalisés dans le laboratoire ou en collaboration avec d'autres équipes, dans le but
de vérifier cette hypothèse de travail (Laurent et al., 1997 ; De Lana et al., 1998 ;
Revollo et al., 1998; Toledo et al., 2002, 2003).
Des corrélations très significatives ont en effet été trouvées entre distances génétiques
et les propriétés biologiques suivantes:
i) les cinétiques de croissance des épimastigotes in vitro,
ii) la différentiation d'épimastigote en trypomastigote,
iii) la parasitémie maximale in vivo chez la souris,
iv) l'infectivité et la mortalité chez la souris (Laurent et al., 1997 ; Toledo et al., 2002).
v) La transmissibilité par Triatoma infestans (De Lana et al., 1998)
vi) La sensibilité in vitro et in vivo aux médicaments comme le Nifurtimox et le
Benznidazole (Revollo, 1995; Revollo et al., 1998 ; Toledo et al., 2003).
Une constance dans tous ces résultats est que si les différences entre DTU sont
statistiquement très significatives, la variabilité intra-DTU est très forte, y compris pour
les études portant sur la sensibilité aux drogues in vitro et in vivo. Sur ce point, les
98

Discussion
résultats de notre équipe diffèrent sensiblement de ceux d'autres équipes (Andrade et
al., 1983), qui ont trouvé une concordance beaucoup plus stricte entre types génétiques
et propriétés biologiques du parasite.
Notre étude a montré une variation importante des valeurs ICsD (déterminées par la
technique de MTT/PMS) dans la plupart des groupes génétiques, la DTU 1(T. cruzi 1)
étant la plus variable. La comparaison entre distances génétiques de Jaccard issues de
l'analyse des trois marqueurs MLEE, RAPD et RADES, et distances estimées à partir
des valeurs ICSD n'a pas montré de corrélation significative. Par conséquent, la
sensibilité des parasites au Benznidazole, mesurée par ICsD avec la méthode
MTI/PMS, ne semble pas avoir de lien direct avec le génotype clonai du parasite. Ceci
paraît être en contradiction avec les études précédentes de notre équipe sur la
chimiosensibilité des souches de T. cruzi, en particulier celles qui portaient sLir
l'association entre diversité génétique et sensibilité des parasites au Benznidazole
mesurée par la méthode de la thymidine tritiée in vitro (Revollo, 1995 ; Revollo et al.,
1998).
Dans le but de comprendre pourquoi, avec le même paramètre biologique, une
corrélation avec les distances génétiques peut être ou non obtenue, nous avons
comparé les valeurs d'ICsD déterminées par notre méthode utilisant le MTT/PMS à
celles déterminées par Revollo (1995) utilisant la thymidine tritiée, ceci sur neuf stocks
communs aux deux études. Bien que les valeurs absolues d'ICsD ne soient pas les
mêmes entre les deux méthodes (MTI/PMS vs. thymidine tritiée), les valeurs relatives
vont dans le même sens. Dans les deux études, les valeurs moyennes d'ICsD pour T.
cruzi 1 sont plus élevées que pour celles de T. cruzi II. Par ailleurs, l'analyse de
corrélation entre les valeurs obtenues par les deux méthodes s'est avérée significative
(r=0,608, P=0,033).
Étant données les différences observées et l'absence de corrélation entre diversité
génétique et ICsD (déterminée par MTI/PMS), nous avons utilisé une troisième méthode,
plus directe, la cytométrie de flux, pour vérifier la validité de nos résultats. La cytométrie
de flux permet en effet la mesure, qualitative et quantitative, des caractéristiques
physiques et biochimiques des cellules et de ce fait, constitue une technique de
référence pour la mesure de la viabilité des parasites. Les analyses de corrélation ont
99

Discussion
montré une similarité importante entre les valeurs d'ICso obtenues par cytométrie de flux
et celles obtenues par la micro-méthode MTT/PMS, ce qui constitue une validation de
nos résultats.
Outre la méthode utilisée (MTI/PMS vs. thymidine tritiée), le protocole utilisé avant la
mesure de l'ICso, le nombre de stocks par groupe génétique ainsi que le nombre de
groupes génétiques sont autant de facteurs qui peuvent expliquer les différences entre
nos résultats et ceux de Revollo (1995). De plus, il est difficile de savoir si nos résultats
sont dus à l'absence réelle de corrélation entre diversité génétique et diversité
biologique (ICso) ou à cause d'une erreur statistique de type II. Ce type d'erreur
(impossibilité de rejeter l'hypothèse nulle due à la perte de puissance du test) pourrait
être favorisé par le faible nombre de stocks (trois) par groupe génétique utilisé dans
notre étude. Les deux hypothèses du reste ne s'excluent pas et peuvent contribuer
toutes deux à nos résultats. Même si ces derniers sont partiellement explicables par un
manque de puissance des tests, ils montrent cependant que la concordance entre
groupes c10naux (DTU) et sensibilité in vitro aux médicaments anti-chagasiques n'est
pas stricte. Des séries statistiques beaucoup plus fortes, rendues difficiles par la
lourdeur des protocoles expérimentaux impliqués, seront nécessaires pour trancher ce
débat de façon fiable.
Il est très probable de toute façon que l'importante variabilité de sensibilité aux drogues
que nous avons mise en évidence est d'origine génétique. Mais nos résultats suggèrent
que l'évolution clonale n'est pas le seul paramètre en cause pour rendre compte de ce
résultat. Ceci du reste n'est aucunement incompatible avec les résultats des autres
chercheurs de notre équipe ou des équipes associées (Laurent et al., 1997 ; Revollo et
al., 1998 ; Toledo et al., 2002, 2003).
4.1.2 Comportement des parasites résistants versus parasites sauvages
Pendant le processus d'induction de la résistance des parasites au Benznidazole, nous
avons observé une diminution de la vitesse de croissance des parasites devenus
résistants par rapport aux parasites sauvages. Ce phénomène a été mis en évidence
par des cinétiques de croissance et par l'analyse du temps de doublement réalisé sur
l'ensemble des stocks chimiorésistants, tant au niveau de la "résistance thérapeutique"
(pour six stocks) qu'à d'autres niveaux de résistance (pour le stock 19). Nous
100

Discussion
confirmons ici sur une plus grande échelle une observation réalisée avec d'autres
approches technologiques et un nombre plus restreint de souches (Nirdé et al., 1995).
Cette diminution de vitesse de croissance pourrait être expliquée par des changements
métaboliques impliquant des modifications dans le cycle cellulaire du parasite, ou à des
modifications chromosomiques induites par une interaction directe du médicament sur
l'ADN (Tocher, 1997).
La résistance s'est avérée stable en l'absence de médicament au moins pendant trois
mois après l'acquisition du niveau thérapeutique ou au-delà pour le stock 19, et au
moins pendant six mois et demi pour un niveau intermédiaire (16~M). Cette stabilité a
déjà été montrée pour des souches qui ont subi le processus d'induction au
Benznidazole soit in vitro (Nirdé et al., 1995), soit in vivo (Murta et Romanha, 1998)
mais aussi à d'autres molécules comme les dérivés azolés in vitro (Buckner et al.,
1998). Nous avons également constaté que la résistance n'est pas perdue après la
congélation de très longue durée (7 ans) des parasites dans l'azote liquide, phénomène
particulièrement intéressant qui rend possible l'étude dans le temps de parasites induits
à n'importe quel niveau de résistance. La stabilité du phénotype résistant, démontrée
dans différentes études, serait un argument qui irait à l'encontre de l'hypothèse selon
laquelle la résistance serait due à l'amplification d'un élément extra-chromosomique
circulaire, car, selon La Flamme et al. (1996), cet élément est théoriquement perdu en
absence de pression médicamenteuse.
Il apparaissait aussi important d'explorer une large gamme de résistance afin
d'observer des modifications possibles des mécanismes de défense du parasite en
fonction de l'importance de la pression médicamenteuse. C'est ainsi que nous avons
étudié des niveaux de résistance de 100 ~M, soit une concentration deux fois
supérieure à la dose circulante chez un patient en cours de traitement (Raaflaub et
Ziegler, 1979 ; Raaflaub, 1980) et 200 ~M.
Les travaux réalisés par Buckner et al. (1998) montrent que les stocks résistants au
Fluconazole présentent une résistance croisée avec d'autres dérivés azolés comme le
Kétoconazole, le Miconazole et l'itraconazole, mais pas de résistance croisée avec
l'Amphotéricine 8 ou le Benznidazole. Ces résultats indiquent que le mécanisme de
résistance au Fluconazole n'est pas spécifiquement lié à ce médicament mais plutôt
101

Discussion
aux dérivés azolés en général. Le Benznidazole inhiberait la synthèse d'ARN et des
protéines (Polak et Richie, 1978) alors que les dérivés azolés, structurellement différent
du Benznidazole, agiraient sur la biosynthèse des stérols. Ainsi le mécanisme de
résistance au Benznidazole devrait être différent de celui des dérivés azolés.
D'autre part, les souches résistantes au Benznidazole sélectionnées in vivo (Murta et
Romanha, 1998) présentent une résistance croisée avec d'autres médicaments
(Nifurtimox, MK-436' et Mégazole) ce qui pourrait suggérer la participation d'une
glycoprotéine du type Pgp-170. Il serait donc très intéressant de déterminer le
comportement des parasites chimiorésistants vis-à-vis d'autres familles
médicamenteuses. En dehors d'une glycoprotéine (séquence F1) potentiellement
impliquée dans le processus de chimiorésistance de T.cruzi, il n'existe pas, à ce jour,
de mécanismes connus de résistance chez T.cruzi impliquant une glycoprotéine.
Différentes hypothèses peuvent expliquer les facteurs qui modulent le processus de
sensibilité ou de résistance des parasites au Benznidazole, tels que des erreurs dans la
synthèse d'ADN ou dans la réplication des parasites (Mac Daniel et Dvorak, 1993). Une
autre hypothèse, proposée par Veloso (Veloso et al., 2001) pour expliquer la variation
de résistance des isolats C et D de la souche Be-78, est que les mutants résistants sont
le résultat d'échanges génétiques (Stothard et al., 1999 ; Gaunt et al., 2003). Selon ces
auteurs, la propagation c10nale des parasites pourrait prédominer dans les cycles de
transmission impliquant l'homme, tandis que les échanges génétiques permettraient
l'émergence de souches présentant de nouvelles propriétés biologiques liées à la
virulence ou à la résistance aux médicaments.
Murta et Romanha (1998) ont étudié une population de parasites de T. cruzi résistants
au Benznidazole après clonage. Tous les clones dérivés de la souche résistante ont
présenté un niveau de résistance important après un traitement bref au Benznidazole,
tandis que tous les clones dérivés d'une population sauvage ont présenté une forte
sensibilité au médicament. Ceci suggère que la résistance des souches puisse être en
relation avec la proportion des clones sensibles/résistants dans une population naturelle
et que le traitement initial pourrait éliminer les clones sensibles laissant ainsi la place à
des clones résistants, dans le cas où ceux-ci soient relativement nombreux. Il serait
possible de vérifier expérimentalement (in vitro puis in vivo) cette hypothèse en
102

Discussion
mélangeant, dans différentes proportions, une population résistante avec sa
contrepartie sensible dont on sait que la vitesse de croissance est plus rapide.
4.2 ETUDE DE L'EXPRESSION DIFFERENTIELLE DES GENES.
La génomique s'est particulièrement développée et s'impose pour exploiter les données
de séquençage des gènes afin d'étudier leur fonctionnement et leur régulation. De
nouvelles technologies, visant l'étude du transcriptome, sont apparues pour quantifier
les ARN messagers présents dans les échantillons biologiques. Bien que les ARNm ne
constituent qu'une étape de l'expression des gènes, leur abondance est souvent
corrélée à l'activité des protéines (Lockhart et Winzeler, 2000).
La méthode classique de "Differentiai display" utilise la technique PCR pour amplifier
les produits de rétrotranscription des ARN, avec des amorces aléatoires (Liang et
Pardee, 1995 ; revue, McClelland et al., 1995 ; Matz et Lukyanov, 1998). Cette
technique peut être utilisée pour détecter des transcrits qui sont différentiellement
exprimés dans des cellules exposées à différents environnements (Silva et al., 1999)
comme des parasites (Ferreira et al., 1999, 2002), à différents programmes de
développement chez T. cruzi (Diaz et al., 2000) ou des parasites soumis à des
conditions de stress médicamenteux (Thelu et al., 1994). En outre, cette technique est
très largement utilisée pour l'étude de l'expression différentielle des gènes impliqués
dans la chimiorésistance des cellules cancérigènes (Yamamoto et al., 2001 ; Parekh et
al., 2002 ; Wang et al., 2002 ; Higuchi et al., 2003).
Diverses adaptations ont été apportées à cette méthode, en particulier pour l'étude des
trypanosomatidés. La technique RADES est une technique dérivée de la "Differentiai
display" (Murphy et Pellé, 1994). Cette technique permet d'obtenir un profil d'expression
des gènes grâce à l'amplification aléatoire (RAPD) des produits rétro-transcripts des
ARNm.
La technique RADES a un très grand éventail d'avantages, par exemple, elle est
réalisable avec un nombre illimité d'amorces, avec également un nombre illimité de
combinaison entre elles. La technique est utilisable avec peu de cellules (théoriquement
avec une seule cellule), elle permet le clonage et le séquençage rapide de séquences
103

Discussion
différentiellement expnmees (Murphy et Pellé, 1994). Cette technique a permis
d'identifier des gènes différentiellement exprimés pendant le cycle de vie du parasite T.
brucei brucei (Murphy et Pellé, 1994), des gènes impliqués dans la mort cellulaire
programmée induite par la Concanavaline A chez T. brucei rhodesiense (Welburn et al.,
1999). Récemment, l'étude de l'expression différentielle des gènes impliqués dans le
développement chez Leishmania a été réalisée à partir d'une technique dérivée du
RADES (Hide, 1999).
Notre étude est la première concernant l'analyse de l'expression génique de T. cruzi
vis-à-vis d'un médicament (le Benznidazole) par une approche différentielle, en utilisant
une technique dérivée du RADES. Cette technique nous a permis d'étudier l'expression
des gènes dans le cas d'une sensibilité naturelle (correspondant à l'IC5o sauvage
propre à chaque stock), ou dans le cas de résistance ind uite à la dose thérapeutique
(SO IJM) et à différents niveaux de chimiorésistance pour le stock 19.
4.2.1 RADES à l'ICso sauvage et au niveau de chimiorésistance thérapeutique (SO
IJM)
L'étude RADES de l'IC5o de chaque stock, a été réalisée dans le but d'identifier les
changements éventuels dans l'expression des gènes du parasite entre son niveau de
sensibilité naturelle et le niveau de résistance thérapeutique (SOIJM) ou des niveaux
plus élevés de résistance (> SOIJM). A partir de cette étude d'expression, nous avons
trouvé 18 bandes différentielles qui ont été exprimées"par différents stocks, soit en
présence de Benznidazole, soit en son absence.
Il est intéressant de noter que les bandes différentielles trouvées à l'IC5o ne sont
associés ni à une DTU particulière, ni à l'une des deux lignées principales de T. cruzi.
Par exemple, certaines bandes différentielles sont présentes dans un stock mais pas
dans les deux autres stocks du même groupe génétique. Ceci semble aller à l'encontre
de notre hypothèse de travail selon laquelle les bandes différentielles seraient liées au
génotype spécifique des subdivisions génétiques de T. cruzi. Cette hypothèse ne tient
pas compte de l'influence de l'origine géographique du stock évoqué par Andrade et al.
en 1985 ou Filardi et Brener en 1987, ni de l'origine de l'hôte définitif. Cette distribution
apparemment aléatoire des bandes différentielles suggère que chaque souche réagit de
manière particulière, indépendamment de son appartenance à une DTU donnée, ce qui
104

Discussion
risque de compliquer considérablement l'étude de la résistance aux médicaments chez
T. cruzi.
Bien qu'il n'y ait pas de lien apparent entre l'expression différentielle et le groupe
génétique, l'expression génique globale (Le. l'ensemble des bandes RADES obtenues),
tant en absence qu'en présence de médicament est très fortement corrélée à la
diversité génétique du parasite, décrite par MLEE ou RAPD. En effet, les arbres
phylogénétiques construits (figure 31) avec l'ensemble de données RADES, en
absence ou en présence de Benznidazole à l'IC5o, groupent les stocks dans les six
même groupes génétiques (DTUs).
20+ lia 14
12+20
10+ Ile 12
7+ IId 7
IId9+ 10
Ile3+ 11
1+ 18
2+ Ile 16
4+ 17
5+ lib 3
6+
14+ lia 2
18+ 4
16+ Ile lib17+ 6
0.1 01
DTUsFigure 31. Arbres de Neighbour-Joining regroupant les 16 stocks de T. cruzi, obtenus à
partir des données RADES des 22 amorces, en présence (+) deBenznidazole à l'IC5o (à gauche) et en absence de Benznidazole (à droite).
Il est intéressant d'observer que l'expression génique globale des parasites en
présence ou non d'une stimulation brève de Benznidazole (à l'IC5o propre de chaque
stock), est directement liée à la structuration génétique de l'espèce T. cruzi. Ceci
montre que la structuration clonale de T. cruzi a un fort impact même sur les gènes
exprimés, c'est-à-dire soumis à sélection naturelle. Ceci confirme un résultat obtenu
105

Discussion
dans notre laboratoire par Tarrieu (2000), qui a observé que la phylogénie déduite de la
séquence du gène Tc52 chez T. cruzi (un gène impliqué dans la pathogénicité du
parasite) était très similaire à la phylogénie gobale de l'espèce, telle qu'établie par
MLEE ou RAPD. Au contraire, la sensibilité naturelle des parasites (exprimée par leurs
valeurs d'ICso) n'est pas corrélée à la structure génétique des parasites, mais apparaît
plutôt comme une caractéristique intrinsèque de chaque stock.
Il est également intéressant de relever que les bandes expnmees différentiellement
lorsque les souches sont exposées brièvement à une concentration de Benznidazole
équivalente à l'ICso sauvage, ne correspondent pas aux bandes exprimées
différentiellement par les stocks chimiorésistants thérapeutiques.
Tout se passe comme si le parasite avait naturellement un mécanisme efficace de
résistance aux drogues. Lors d'un stress médicamenteux, il serait capable de
développer des mécanismes encore plus efficaces lui permettant de résister à ce stress
temporaire. Puis, pour un état de chimiorésistance élevée et stabilisée, il pourrait être
capable de mettre en place de nouveaux mécanismes encore plus actifs, et ceci de
façon durable. Ce qui est alors remarquable, c'est que tous les parasites ne mettent pas
en place le même mécanisme pour répondre à une situation d'urgence.
4.2.2 RADES à différents niveaux de chimiorésistance au Benznidazole
Il est intéressant d'observer que la moitié des bandes différentielles du stock 19 est
surexprimée pour tous les niveaux de chimiorésistance. L'une des bandes est
spécifique de la DTU 1 (bande 1 de l'amorce Al), et elle est également surexprimée
dans le stock 3 chimiorésistant, lequel appartient au même groupe génétique que le
stock 19 (DTU 1). Cette bande est apparemment uniquement présente dans la lignée 1
(Stocks 3 et 19 chimiorésistants) et toujours absente dans la lignée Il (stocks 5, 6, 9, 10
et .18). Des analyses complémentaires seront nécessaires pour confirmer cette
observation originale, pouvant aboutir à la sélection d'un marqueur de résistance propre
à une DTU donnée.
Une autre bande commune aux deux stocks 3 et 19 chimiorésistants (la bande 1 de
l'amorce A2) est à la fois exprimée différentiellement dans le stock 19 chimiorésistant et
simplement surexprimée dans le stock 3 chimiorésistant.
106

Discussion
La disparition progressive de la bande 1 de l'amorce U16, au fur et à mesure que le
niveau de résistance augmente, est un phénomène également intéressant. Il suggère
une extinction progressive, dose-dépendante, donc d'une adaptation d'un mécanisme
moléculaire initialement présent chez le parasite. Au contraire, la plupart des bandes
différentielles qui sont de simples apparitions ou disparitions de bandes suggèrent une
modification plus marquée dans le métabolisme du parasite.
L'apparition ou la disparition de bandes RADES doit être interprété avec beaucoup de
prudence; les mécanismes sous-jacents sont probablement multiples et complexes, et
il convient ici de distinguer au moins trois cas de figures.
- Premier cas: un même stock montre des bandes différentielles quand il est soumis
au stress du médicament à la dose IC50 pour des stimulations brèves (sur 3
générations environ). Dans ce cas, des mutations du site de l'amorce RAPD ou de
grandes variations de séquences semblent improbables sur des temps aussi courts, et
le mécanisme d'apparition ou de disparition de bande repose essentiellement sur des
phénomènes de régulations géniques, vraisemblablement post-transcriptionnelles.
Néanmoins, chez les bactéries, des gènes mutateurs permettent une augmentation très
importante du nombre de mutations et la sélection de mutants après plusieurs
centaines de générations seulement (quelques jours). Les mutants qui sont les mieux
adaptés aux nou~elles conditions de stress, par exemple, les mutants sélectionnés
pendant l'exposition aux médicaments (Horst et al., 1999 ; Chopra et al., 2003) sont
ainsi rapidement sélectionnés. L'augmentation du taux de mutation paraît en effet être
l'une des stratégies les plus efficaces utilisée par les cellules tumorales, les bactéries et
les virus pour résister aux nouveaux médicaments (Taddei et al., 1997). Les gènes
mutateurs n'ont pas été encore décrits chez T. cruzi, cependant, plusieurs de ces
gènes, identifiés initialement chez Escherichia coli, ont été rencontrés chez d'autres
micro-organismes et chez l'homme.
- Deuxième cas: un même stock montre des bandes différentielles pour des
chimiorésistances stabilisées: ici, en plus des phénomènes de régulations géniques
interviennent probablement, des mutations, voire des remaniements génétiques
importants qui seraient la conséquence de l'exposition en continu du médicament. Ces
changements génétiques pourraient être sensiblement différents en fonction du génome
107

Discussion
de chaque souche, ce qui expliquerait que certaines bandes différentielles seraient
observées dans certains stocks et pas dans d'autres. On peut envisager ici la
participation de régulateur de la chromatine comme les HDAC (Histone Desacétylase)
qui interviennent dans le "silencing" comme c'est le cas dans la résistance de certains
cancers humains (Demirpence et al., 2002).
- Troisième cas: sur des stocks différents, existence de bandes différentielles
différentes quand les parasites sont soumis au stress du médicament à la dose ICso :
Dans ce cas, on ne sait pas à priori s'il s'agit vraiment d'une véritable expression
différentielle de gènes, qui suppose un génome identique, ou s'il s'agit de deux
génomes suffisamment distants pour montrer des différences RAPD. Néanmoins,
lorsque les différences concernent deux souches d'une même DTU, les génomes étant
plus proches, la probabilité d'observer une vraie expression différentielle augmente.
Dans ce dernier cas, des hybridations moléculaires de la sonde issue de la bande
d'intérêt avec l'ADN des deux génomes et avec les produits RAPD nous auraient
permis de déterminer quels étaient les vrais locus à expression différentielle et quels
étaient les locus différents. Ce travail extrêmement fastidieux n'a pas été fait pour des
raisons de temps.
Enfin, la surexpression de bandes dans un environnement donné, peut être attribuée à
des mécanismes de régulation post-transcriptionels intervenant par exemple au niveau
de l'épissage, de la polyadénylation ou de la stabilité de l'ARN (Matthews et al., 1994 ;
Furger et al., 1997 ; Abuin et al., 1999 ; Teixeira et DaRocha, 2003), ou à une
amplification de gènes (revue, Ullman, 1995).
A partir de l'ensemble des bandes différentielles, nous avons sélectionné, cloné et
séquencé 8 bandes représentatives de chacun des cas rencontrés, c'est-à-dire, soit des
bandes présentes seulement dans des stocks résistants mais pas dans les stocks
sauvages, soit des bandes présentes dans les stocks sauvages mais éteintes dans les
stocks résistants, soit des bandes présentes dans les deux mais surexprimées dans les
chimiorésistants. Les séquences nucléotidiques obtenues pour des bandes
sélectionnées ont été comparées à celles présentes dans les différentes banques de
données.
108

Discussion
Deux séquences seulement (02 et F1) ont montré une homologie importante avec des
séquences déjà décrites, avec un très fort score à des niveaux significatifs. D'autres
bandes avec des scores plus faibles ont néanmoins retenu notre attention (voir tableau
10).
La séquence 02 (bande 1 de l'amorce A2, stock 19 chimiorésistant) a montré une
identité de 27% avec une protéine flagellaire de Chlamydomonas reinhardtii. Cette
protéine est impliquée dans la conversion de forme du ·nagelle de cette algue qui passe
du type ciliaire au type flagellaire, cette conversion est dépendante du calcium (Ca2+)
(Tam et Lefebvre, 2002). A priori, il est surprenant d'observer qu'un tel organelle
cellulaire puisse être impliqué dans un mécanisme de chimiorésistance. Cependant, la
forme du flagelle de T.cruzi se modifie au cours de son cycle cellulaire, pour adopter
une forme particulière au stade trypomastigote puisqu'il est relié à la membrane
plasmique par une membrane ondulante. Par ailleurs, le Ca2+ est un second messager
intervenant dans la différenciation des trypanosomatidés (Lammel et al., 1996 ; Stojdl et
Clarke, 1996). De plus, plusieurs protéines de liaison au Ca2+ décrites chez le
trypanosomatidés, sont localisées dans le flagelle, organite qui semble être crucial pour
la transmission des signaux calcium (Godsel et Engman, 1999 ; Maldonado et al.,
1999). Il n'est pas inconcevable d'impliquer cette protéine dans le processus de
chimiorésistance puisqu'il a été démontré que la surexpression d'une protéine calcium
dépendante est impliquée dans des modèles de chimiorésistance cancéreuse (Parekh
et al., 2002).
La séquence F1 (bande 1 de l'amorce U16, stock 19 wt) a montré une identité de 40%
avec une protéine hypothétique de L. major. De plus, cette séquence a montré
également de faibles identités avec une glycoprotéine de liaison au sperme chez Sus
scrofa (porc) et avec un transporteur de type ABC décrit chez Sinorhizobium meliloti
(Barnett et al., 2001). Des études complémentaires devront être réalisées avec le but
d'établir la présence d'un autre transporteur ABC chez T. cruzi, ainsi que d'établir sa
possible participation dans la résistance du parasite au Benznidazole. Cette observation
pourrait suggérer que la Pgp-170 Iike (MOR protéine, Masanek et al., 2002) recherchée
chez T.cruzi possède une séquence très différente de. celles déjà décrites dans les
autres modèles, et suffisamment éloignée pour ne pas être mise en évidence par
immuno-histocytochimie ou par hybridation moléculaire.
109

Discussion
La séquence C1 (bande 1 de l'amorce Al, stock 19 chimiorésistant) a montré une
identité de 26% avec une possible calmoduline chez Leishmania major (Ivens et al.,
1998). L'alignement de la séquence C1 avec la calmoduline et les gènes calmodulin
ubiquitine associés (CUS) décrits chez T. cruzi a montré une identité de respectivement
15,4% et 20%. La calmoduline intervient dans le contrôle d'un grand nombre d'enzymes
à travers les ions calcium Ca2+. Parmi les enzymes modulées par la calmoduline,
existent les kinases et les phosphatases. Etant donné que les gènes CUS (pour
Calmoduline-Ubiquitine associated genes) sont essentiels à la survie de T. cruzi (Ajioka
et Swindle, 1996) on pourrait penser que cette protéine est surexprimée en conditions
de stress, comme c'est le cas lors de l'induction de la résistance. Néanmoins, le rôle
exact que jouent les gènes CUS reste inconnu. La calmoduline a quatre sites
fonctionnels de liaison au Ca2+ et présente des similarités avec d'autres protéines de
liaison au Ca2+, de type EF-hand. Nous avons cherché les motifs correspondants dans
la séquence de la protéine C1, mais nous n'avons pas trouvé ce type de motifs.
Cependant, nous avons trouvé un motif de liaison à l'ATP [A TPIGTP-binding site motif
A (P-Ioop)].
Il est intéressant de remarquer que les deux séquences D2 et C1 (correspondant à
deux bandes exprimées différentiellement par le stock 19 chimiorésistant, avec deux
amorces différentes A2 et Al), sont toutes les deux en relation avec des protéines de
liaison au Ca2+. Le Ca2
+ joue un rôle très Important dans la physiologie des organismes
supérieurs, et il est également impliqué dans la régulation de plusieurs processus
cellulaires comme la transcription génique ou les mécanismes complexes qui président
à la survie cellulaire (Brini et Carafoli 2000 ; Srini, 2003). Il est donç envisageable que le
stress produit lors du processus d'induction soit directement lié à la régulation du Ca2+
intracellulaire, impliquant ainsi l'activation de différents transporteurs de cet ion.
Pour les séquences 11 et J1, exprimées de manière différentielle par le stock 10, les
identités trouvées correspondent à une succinate déshydrogénase/fumarate réductase.
La succinate deshydrogenase (E.C. 1.3.99.1) est une enzyme essentielle pour la
respiration cellulaire. Cette enzyme est une 11avoprotéine Fe-S de la membrane interne
(qui fonctionne avec du FAD), correspondant au complexe Il de la chaîne respiratoire
mitochondriale. Le succinate est oxydé, par action de la succinate déshydrogénase, en
fumarate et les hydrogènes produits sont transmis au coenzyme Q. L'enzyme NADH-
110

Discussion
fumarate réductase est impliquée dans la réduction du fumarate en succinate.
Néanmoins, chez les trypanosomatidés, la production de succinate par cette voie n'a
pas été clairement démontrée (Tielens et van Hellemond, 1998, 1999). Des études
complémentaires devront être réalisées avec le but d'établir quel serait le possible rôle
de cette enzyme dans la résistance au Benznidazole.
4.3 EXPRESSION DES GENES SPECIFIQUES
Gènes tcsod-A et tcsod-B
Un grand nombre de médicaments, comme ceux qui sont utilisés pour le traitement de
la maladie de Chagas, exercent leur fonction à travers l'intermédiaire de radicaux libres
produits lors de leur propre métabolisme (Maya et al., 2003). Les parasites possèdent
différents systèmes de défense contre le stress oxydatif : la superoxyde dismutase ; la
thiol trypanothione, la trypanothione réductase, la tryparedoxyne et la trypanothione
peroxydase composants du système de la trypanothione peroxydase; l'ovothiol. " était
par conséquent logique de penser que pendant le processus d'induction de la
résistance au Benznidazole, les parasites pourraient surexprimer les gènes impliqués
dans la lutte contre le stress oxydatif, dont on sait qu'elles participent à l'inhibition de
l'apoptose chez les cellules eucaryotes.
A la différence des hôtes mammifères, T. cruzi est relativement déficient en
mécanismes de défense contre les métabolites de l'oxygène. Chez T. cruzi, la catalase
est complètement absente et la glutathione peroxydase a une activité déficiente
(Docampo et al., 1976 ; Boveris et al., 1980).
Chez d'autres organismes comme la bactérie Helicobacter pylori, il a été démontré que
le mécanisme de résistance au Métronidazole (un autre nitroimidazole) impliquait la
production de grandes quantités de superoxyde dismutase et de catalase (Smith et
Edwards, 1995). Pour ces auteurs, la résistance serait causée par la production
d'agents piégeurs "scavengers" ("éboueurs") de radicaux libres ou serait causée par
des altérations du métabolisme du pyruvate. Un mécanisme analogue pourrait être
possible chez T. cruzi chez qui des SODs avaient été déjà identifiées. C'est pour cette
raison que nous avons étudié l'expression des gènes tcsod-A et tcsod-B, qui codent
111

Discussion
pour les Superoxyde Dismutases A et B chez T. cruzi, afin d'établir son possible rôle
dans la résistance au Benznidazole.
Nos résultats montrent que ces gènes tcsod-A et tcsod-B ne sont apparemment pas
impliqués dans les mécanismes de résistance au Benznidazole des parasites pour les
niveaux de résistance analysés et pour les clones étudiés. Cela n'exclut nullement leur
participation dans la résistance pour d'autres stocks ou pour d'autres molécules comme
le Nifurtimox.
Pour nos stocks, il n'y a pas de différence d'expression des SOD entre les parasites
chirniorésistants et sauvages. L'effet du médicament ne se fait peut-être pas ici à
travers les intermédiaires de nitroréduction comme c'est pourtant le cas pour le
Nifurtimox (Viodé et al., 1999). Apparemment, le Benznidazole inhibe la croissance des
parasites à des concentrations qui ne stimule pas la production de 02-- et H202 (Moreno
et al., 1982). D'autres conditions expérimentales ont permis de montrer la production de
métabolites actifs, à partir de la réduction enzymatique du Benznidazole, qui sont liés
de façon covalente aux protéines (Masana et al., 1984). Récemment, une étude
réalisée avec des molécules dérivés du 2-nitroimidazole, du 5-nitroimidazole et des
nitrofuranes a montré que les nitrofuranes induisent le stress oxydatif tandis que les
nitroimidazoles agissent en réduisant le trypanothione (Maya et al., 2003). Quoi qu'il en
soit, même si les deux SOD jouent un rôle dans des mécanismes de détoxification,
elles ne sont pas surexprimées, dans nos conditions expérimentales, en cas de
résistance au Benznidazole.
Néanmoins, il est possible d'envisager d'étudier dans les stocks résistants, l'expression
de gènes impliqués dans le métabolisme du T~liol (tels la thiol trypanothione, la
trypanothione réductase, la tryparedoxyne et la trypanothione peroxydase), qui font
partie du système de défense du parasite et qui sont apparemment responsables des
différences de sensibilité (IV/oncada et al., 1989 ; Repetto et al., 1996 ; Maya et al.,
1997). De plus, le métabolisme du Thiol a été impliqué dans la résistance à l'arsénite
chez L. tarentolae (Dey et al., 1996 ; Mukhopadhyay et al., 1996) et au Glucantime chez
L. tropica (Arana et al., 1998).
112

Discussion
D'autre part, les radicaux libres produits lors du métabolisme des médicaments qui
possèdent des groupes nitroaromatiques sont très souvent impliqués dans les effets
toxiques chez les patients traités (Docampo et Moreno, 1984, 1986 ; Marr et Docampo,
1986). Ainsi, l'explication des effets secondaires produits par le Benznidazole reste
encore à élucider.
Pendant le processus d'induction de résistance, nous avons cultivé les parasites en
présence du Benznidazole en augmentant la surface d'échange avec l'oxygène de l'air
lorsque les boites de culture étaient mises en position horizontale. Dans ce cas, la
vitesse de croissance des parasites était plus importante, ce qui suggère un lien direct
entre l'oxygénation et la capacité des parasites à résister au médicament (résultats non
présentés). Cependant, la réduction du Benznidazole en condition aérobie est moins
importante qu'en condition anaérobie, ce qui expliquerait la survie des parasites dans
ces conditions. Le niveau général de 02~ produit pourrait donc être facilement éliminé
par la SOD du parasite (Docampo et al., 1984). D'autre part, l'enzyme SOD est
inductible, sa biosynthèse est augmentée par l'hyperoxygénation et par certains
toxiques comme le paraquat (un herbicide). Ce phénomène pourrait faire l'objet
d'études complémentaires.
Gène tccp1
Les cystéine protéases sont impliquées dans plusieurs processus cellulaires tels que
l'invasion cellulaire, la différentiation cellulaire, la cytoadhérence ainsi que la protection
des parasites contre la réponse immune de l'hôte. Nous avons étudié l'expression de ce
gène dans le but d'établir sa possible participation dans les mécanismes de défense et
résistance des parasites lors du processus d'induction. L'expression apparemment
aléatoire de ce gène dans les stocks sauvages et les stocks chirniorésistants ne permet
pas de tirer de conclusion de notre étude préliminaire. Cependant, l'importance de cette
enzyme dans la biologie cellulaire mériterait qu'une étude plus approfondie soit réalisée
chez les parasites chimiorésistants.
Gène tcpgp2
L'importance des P-glycoprotéines et des transporteurs trans-membranaires dans la
résistance aux médicaments a été amplement démontrée depuis la découverte de ce
type de protéines dans les cellules tumorales multirésistantes chez le hamster (Juliano
113

Discussion
et Ling, 1976). La résistance aux médicaments provient de la capacité des P
glycoprotéines à expulser de manière active les médicaments contre un gradient de
concentration, provocant ainsi une diminution du médicament au contact des cibles
dans les cellules.
Les P-glycoprotéines sont impliquées dans la résistance des cellules tumorales
humaines ainsi que dans la résistance des parasites comme Plasmodium, Leishmania
et Entamoeba, aux médicaments couramment utilisés. Chez T. cruzi, deux P
glycoprotéines (tcpgp1 et tcpgp2) ont été décrites, mais à ce jour aucune des deux n'a
été impliquée dans la résistance du parasite sur des parasites naturellement résistants
et sélectionnés in vivo (Murta et al., 2001). Dans notre étude, les résultats montrent que
le gène tcpgp2 n'est apparemment pas impliqué chez T. cruzi dans la chimiorésistance
induite in vitro par le Benznidazole. En effet nous n'avons pas observé de différence
d'expression entre les stocks sauvages et les stocks chirniorésistants tant au niveau
thérapeutique qu'à différents niveaux de chimiorésistance plus élevés. Cependant, la
présence d'un autre transporteur type MOR ne peut pas être exclue.
4.4 STABILITE DES GENOTYPES DES STOCKS
Des études préliminaires menées sur la diversité génétique de T. cruzi, avaient permis
d'établir une première classification des populations existantes en trois zymodèmes
principales prinCipaux nommés Z1, Z2 et Z3 (Miles et al., 1978). Dans un étude
postérieure, Tibayrenc et al. (1986) ont étudié un échantillon de 121 stocks afin
d'analyser la diversité génétique du parasite. Au total 43 génotypes c10naux furent
identifiés.
Des études plus récentes ont permis de clairement identifier deux lignées majeures au
sein de T. cruz; (Tibayrenc, 1995, 1996 ; Souto et al., 1996), qui ont été dénommées
par un comité d'experts T. cruz; 1et T. cruzi Il (Momen, 1999). Elles correspondent au
concept de Discrete Typing Unit (DTU) proposé par Tibayrenc (1998). Aucune
subdivision claire n'apparaît dans la DTU 1alors que la DTU Il est divisée en cinq DTU
additionnelles (DTUs lia, lib, IIc, IId et Ile) (Barnabé et al., 2000 ; Brisse et al., 2000).
114

Discussion
Dans notre étude, les analyses enzymatiques (MLEE) et moléculaires (RAPD) ont
permis de vérifier l'appartenance des stocks choisis pour cette étude aux six groupes
génétiques, répartis dans les deux lignées phylogénétiques majeures (1 et Il). Certains
arbres phylogénétiques construits à partir de ces données ont montré des hiérarchies
de branchement inter-DTUs légèrement différentes en fonction de la technique utilisée.
Néanmoins ces marqueurs ont toujours permis d'identifier les stocks ainsi que leur
appartenance à une DTU.
Nous n'avons pas observé de changement de profil isoenzymatique pendant les trois
années de culture des stocks, tant pour les stocks sauvages que pour les stocks
chimiorésistants, ce qui corrobore ce qui a été préalablement rapporté dans des
conditions similaires (Nirdé et al., 1995).
La Glucose-6-phosphate isomérase (GPI) n'a pas été étudiée dans notre étude du point
de vue de la résistance au Benznidazole, mais comme l'un des marqueurs des
différentes DTUs. En effet, la variabilité génétique de ce locus isoenzyma!jque en fait un
excellent "tag" spécifique de DTU (Barnabé, 2000 ; voir figure 8). Il est intéressant de
noter que lors des caractérisations enzymatiques du stock 19, nous avons pu observer
des différences d'expression de la GPI en fonction du niveau de résistance de ce stock.
En fait, plus le niveau de résistance est important, plus l'intensité de la bande est forte,
avec des pics d'expression très importants à 100 et 200 J.lM.
La GPI est une enzyme dimérique qui catalyse la conversion du D-glucose-6-phosphate
en D-fructose-6-phosphate (Achari et al., 1981 ; Smith et Doolittle, 1992). La GPI a été
impliquée dans plusieurs voies métaboliques telles que la glycolyse (chez les
organismes supérieurs), la gluconéogenèse (chez les mammifères), la biosynthèse des
hydrates de carbone (chez les plantes) et dans la voie Entner-Doudouroff (chez
certaines bactéries). Cette protéine multi-fonctionnelle a été également impliquée dans
différents processus comme étant un facteur autocrine de motilité (AMF, Watanabe et
al., 1996), comme facteur de différentiation et médiateur de la maturation (Xu et al.,
1996) et aussi comme neuroleukine (Gurney et al., 1986 ; Faik et al., 1988). Elle a
différents rôles à l'intérieur comme à l'extérieur des cellules. A l'intérieur du cytoplasme
elle catalyse la seconde étape de la glycolyse, tandis qu'à l'extérieur, elle peut agir
comme facteur de croissance des nerfs et comme cytokine. De plus, la déficience de
115

Discussion
cette enzyme a été impliquée dans l'anémie hémolytique et dans d'autres désordres
autosomiques récessifs (Baughan et al., 1968).
Les variations observées au niveau de l'expression de la GPI pourraient par
conséquent indiquer des changements au niveau métabolique liés à la chimiorésistance
au Benznidazole, dans laquelle la glycolyse pourrait être impliquée. Pour l'instant, nous
ne pouvons que constater ces variations d'expression en fonction du niveau de
résistance du stock 19 et des analyses complémentaires seraient nécessaires pour
éclaircir le rôle de cette enzyme dans la résistance aux médicaments chez T. cruzi.
116

CONCLUSION

Conclusion
5 CONCLUSION
L'un des résultats les plus remarquables est l'absence de lien avéré entre le niveau
de sensibilité des stocks de Trypanosoma cruzi et leur appartenance à un groupe
génétique donné. Par conséquent, la structuration c10nale des populations naturelles
de T. cruzi ne peut expliquer à elle seule l'ensemble des données obtenues sur la
chimiorésistance. Néanmoins nous ne pouvons pas exclure une erreur statistique de
type Il, c'est à dire l'impossibilité de rejeter l'hypothèse nulle (ici absence de lien
entre résistance et diversité génétique), par manque de puissance du test,
probablement lié à un effectif faible de stocks dans chaque DTU.
Quoiqu'il en soit, le niveau de sensibilité naturel au Benznidazole des ~lones de T.
cruzi s'est révélé être extrêmement variable, comme beaucoup d'autres paramètres
biologiques et génétiques de l'espèce, préalablement étudiés.
Les mécanismes de résistance au médicament sont très complexes et font appel
apparemment à plusieurs modifications métaboliques. Cette complexité pourrait
expliquer les différences importantes d'expression génique entre les différents
stocks, y compris ceux appartenant à un même groupe génétique. La méthode
RADES est une approche globale d'étude des différences d'expression génique. Elle
permet de montrer certaines différences d'expression, mais c'est un sondage
aléatoire, et dans le cas d'une espèce hypervariable comme T. cruzi, cet aspect
aléatoire pourrait avoir atteint ses propres limites. En d'autres termes, les différences
inter-stocks révélées par la méthode RADES ne sont pas forcément les plus
pertinentes et elles doivent être interprétées avec beaucoup de précaution.
Néanmoins, ces méthodes nous ont permis d'identifier plusieurs nouveaux gènes
probablement directement impliqués dans les mécanismes de résistance au
Benznidazole, et ceux-ci devraient faire l'objet d'études ultérieures.
De plus, il est intéressant de noter que la plupart des gènes candidats qui ont été
choisis dans ce travail pour leur implication potentielle dans la résistance aux
médicaments, n'ont de fait pas montré de différence d'expression entre stocks
sauvages et résistants.
117

PERSPECTIVES

Perspectives
6 PERSPECTIVES
La stratégie générale qui paraît la plus opportune pour continuer cette ligne de
recherche serait l'étude des principales molécules nouvellement identifiées et
impliquées dans la résistance au Benznidazole, ceci d'abord sur un seul stock, puis
étudier les modulations de l'expression de ces gènes au sein d'un échantillonnage
représentatif de la variabilité génétique de l'espèce Trypanosoma cruzi. Pour
chacune des protéines étudiées, il faudra tenir compte de l'existence possible
d'isoformes au sein d'un même stock mais aussi au sein d'une même DTU.
Au plan fonctionnel, on envisagera d'utiliser la technique du RNAi (ARN interférent)
qui permettra d'inhiber spécifiquement la production de la protéine d'intérêt afin de
vérifier sa participation dans l'acquisition du phénotype résistant.
Il est important de souligner ici que les formes résistantes obtenues dans cette étude
sont des formes épimastigotes du parasite, c'est-à-dire, des formes présentes chez
le vecteur et non chez l'hôte définitif. L'étude la plus pertinente serait de travailler
avec les formes trypomastigotes infectieuses ainsi qu'avec les formes intracellulaires
amastigotes du parasite, rencontrées chez l'homme. L'environnement différent entre
le vecteur et l'homme peut conduire à des réponses métaboliques différentes pour
un même phénotype résistant, bien que le phénotype résistant soit conservé au
cours de la transformation in vitro. Il conviendra de vérifier que les molécules
repérées chez les épimastigotes résistants sont bien les mêmes qui sont impliquées
chez les trypomastigotes et chez les amastigotes résistants in vivo qu'il faudrait
obtenir.
Toutes les protéines du parasite impliquées dans les mécanismes de
chimiorésistance sont des cibles thérapeutiques potentielles. De plus, bien qu'aucun
élément expérimental n'ait apporté la preuve de la sécrétion de ces protéines, cette
hypothèse ne peut être exclue et devra être explorée.
Des applications cliniques importantes peuvent découler de ces travaux, comme par
exemple la détection de la présence de souches résistantes au cours d'un traitement
chimiothérapeutique.
118

Perspectives
De plus des protéines détectées dans cette étude (protéine du flagelle, GPI,
Succinate déhydrogénase, Calmoduline-Iike, transporteur de type ABC), qui feront
l'objet d'investigations plus poussées, d'autres molécules se rapportant à
l'observation faite concernant l'interface air-milieu de culture ou concernant les
mécanismes de défense du parasite, devraient être étudiées. Ce sont les
cytochromes P450, la trypanothione réductase, l'ovothiol réductase, la TcPRAC ou la
Tc52.
Nous pensons que ces travaux préliminaires sur la chimiorésistance de T. cruzi
permettront de mieux comprendre le processus de résistance aux médicaments,
particulièrement complexe et important dans d'autres pathologies humaines, comme
les infections microbiennes ou en cancérologie, et pour lequel nous sommes
actuellement impuissants.
119

REFERENCES BIBLIOGRAPHIQUES

Références Bibliographiques
7 REFERENCES BIBLIOGRAPHIQUES
Abuin G., Freitas-Junior L.H.G., Colli w., Alves M.~I.M. et Schenkman S., (1999). Expression oftrans-Sialidase and 85-kDa Glycoprotein genes in Trypanosoma cruzi is differentially regulated atthe post-transcriptionallevel by labile protein factors. J. Biol. Chem. 274 :13041-13047.
Achari A., Marshall S.E., Muirhewad H., Palmieri RH. et Noltmann E.A., (1981). Glucose-6phosphate isomerase. Philos. Trans. R. Soc. Lond. B. Biol. Sei. 293(1063) :145-157.
Agabian, N., (1990). Trans splicing of nuclear pre-mRNAs. Cel/61 (7) :1157-1160.Agüero F., Verdun R.E., Frash A.C.C. et Sanchez 0.0., (2000). A random sequencing approach for
the analysis of Trypanosoma cruzi genome: General structure, large gene and repetitive DNAfamilies, and gene discovery. Genome Res. 10 :116-2005.
Ajioka J. et Swindle J., (1996). The calmodulin-ubiquitin (CUB) genes of Trypanosoma cruzi areessential for parasite viability. Mol. Biochem. Parasitol. 78 (1-2) :217-225.
Altschul S.F., Madden T.L., Schaffer A.A., lhang J., lhang l., Miller W. et Lipman D.J., (1997).Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic.Acids. Res. 25 :3389-3402.
Amato Neto V., (1999). Etiological treatment for infection by Trypanosoma cruzi. Mem. Inst. OswaldoCruz. 94 (Suppl 1) :337-9.
Ambudkar S.V. et Gottesmann M.M., (1998). ABC transporteurs: Biochemical, cellular and molecularaspects. Vol. 292. Academic Press, San Diego, CA, USA.
Andrade S.G., Figueira R.M., Carvalho M.L. et Gorini D.F., (1975). [Reaction of the Trypanosomacruzi strain to the experimental therapeutical response to Bay 2502 (results of long term treatment)].Rev. Inst. Med. Trop. Sao Paulo. 17(6) :380-389. (in Portuguese).
Andrade S.G., (1976). Tentative for grouping different Trypanosoma cruzi strains in some types. Rev.Inst. Med. Trop. Sao Paulo 18 :140-141.
Andrade S.G. et Figueira R.M., (1977). [Experimental study on the therapeutic action of Ro 7-1051on infections caused by different strains of Trypanosoma cruzl]. Rev. Inst. Med. Trop. Sao Paulo19(5) :335-41. (in Portuguese).
Andrade S.G., Andrade l.A. et Figueira RM., (1977). [Experimental study on the resistance of astrain of Trypanosoma cruzi to Bay 2502]. Rev. Inst. Med. Trop. Sao Paulo. 19(2) :124-129. (inPortuguese).
Andrade V., Brodskyn C. et Andrade S.G., (1983). Correlation between isoenzyme patterns andbiological behaviour of different strains of Trypanosoma cruzi. Trans. R. Soc. Trop. Med. Hyg.77(6):796-9.
Andrade S.G., Magalhaes J.B. et Pontes A.L., (1985). Evaluation of chemotherapy withBenznidazole and Nifurtimox in mice infected with Trypanosoma cruzi strains of different types. Bull.World Hea/th Organ. 63(4) :721-726.
Andrade S.G., Rassi A., Magalhaes J.B., Ferriolli Filho F. et Luquetti A.O., (1992). Specificchemotherapy of Chagas disease : a comparison between the response in patients andexperimental animais inoculated with the same strains. Trans. R. Soc. Trop. Med. Hyg. 86(6) :624626.
Arana F.E., Perez-Victoria J.M., Repetto Y., Morelia A.., Castanys S. et Gamarro F., (1998).Involvement of thiol metabolism in resistance to glucantime in Leishmania tropica. Biochem.Pharmacol. 56(9) :1201-1208.
Arts G.J. et Benne R., (1996). Mechanism and evolution of RNA editing in kinetoplastida. Biochim.Biophys. Acta 1307 :39-54.
Augustyns K., Amssoms K., Yamani A., Rajan P.K. et Haemers A., (2001). Trypanothione as atarget in the design of antitrypanosomal and antileishmanial agents. Curr. Pharm. Des. 7 :11171141.
Avila H., Sigman O.S., Cohen L.M., Millikan RC. et Simpson L., (1991). Polymerase chain reactionamplification of Trypanosoma cruzi kinetoplast minicircle DNA isolated from whole blood Iysates:diagnosis of chronic Chagas disease. Mol. Biochem. Parasitol. 48 :211-221.
Avila, H.A. et Simpson, L. (1995). Organization and complexity of minicircle-encoded guide RNAs inTrypanosoma cruzi. RNA 1 :939-947.
Bacchi C.J., (1993). Resistance to clinical drugs in African trypanosomes. Parasitol. Today 9(5) :190193.
120

Références Bibliographiques
Bacchi C.J., Garofalo J., Ciminelli M., Rattendi D., Goldberg B., McCann P.P. et Yarlett N.,(1993). Resistance to DL-alpha-difluoromethylornithine by clinical isolates of Trypanosoma bruceirhodesiense. Role of S-adenosylmethionine. Biochem. Pharmacol. 46(3) :471-481.
Bakalara N., Simpson AM. et Simpson L., (1989). The Leishmania kinetoplast-mitochondrioncontains terminal Uridylyltransferase and RNA ligase activities. J biol. Chem. 264(31) :1867918686.
Baker, J.R., Miles, M.A, Godfrey, D.G. et Barret, T.V., (1978). Biochemical characterization of somespecies of Trypanosoma (Schizotrypanum) from bats (Microchiroptera). Am. J. Trop. Med. Hyg.27:483-491.
Balzi E. et Goffeau A, (1994). Genetics and biochemistry of yeast multidrug resistance. Biochim.Biophys. Acta 1187 :152-162.
Bannister J.V., Bannister W.H. et Rotilio G., (1987). Aspects of the structure, function, andapplications of superoxide dismutase. CRC Crit Rev Biochem. 22(2) :111-80.
Barnabé C., Brisse S. et Tibayrenc M., (2000). Population structure and genetic typing ofTrypanosoma cruzi, the agent of Chagas disease: a multilocus enzyme electrophoresis approach.Parasitology 120 :513-526.
Barnett M.J., Fischer R.F., Jones T., Komp C., Abola AP., Barloy-Hubler F., Bowser L., CapelaD., Galibert F., Gouzy J., Gurjal M., Hong A et al., (2001). Nucleotide sequence and predictedfunctions of the entire Sinorhizobium meli/oti pSymA megaplasmid. Proc. Nat!. Acad. Sei USA.98(17) :9883-9888.
Bart G., Coombs G.H. et Mottram J.C., (1995). Isolation of Imcpc, a gene encoding a Leishmaniamexicana cathepsin-B-like cysteine proteinase. Mol. Biochem. Parasitol. 73(1-2) :271-4.
Baughan M.A., Valentine W.N., Paglia D.E., Ways P.O., Simons E.R. et DeMarsh a.B., (1968).Hereditary hemolytic anemia associated with glucosephosphate isomerase (GP!) deficiency - a newenzyme defect of human erythrocytes. Blood 32(2) :236-249.
Beai M.F., (1995). Aging, energy, and oxidative stress in neurodegenerative diseases. Ann. Neurol.38(3) :357-66.
Beckman K.B. et Ames, B.N., (1998). The free radical theory of aging matures. Physiol. Rev.78(2):547-581.
Belli 5.1., (2000). Chromatin remodelling during the life cycle of trypanosomatids. Int. J. Parasitol.30:679-687.
Bello AR., Nare B., Freedman D., Hardy L. et Beverley S.M., (1994). PTR1: a reductase mediatingsalvage of oxidized pteridines and methotrexate resistance in the protozoan parasite Leishmaniamajor. Proc. Nat!. Acad. Sei. USA. 91 (24) :11442-11446.
Ben Abderrazak S., Guerrini F" Mathieu-Daudé F., Truc P., Neubauer K., Lewicka K., Barnabe C.et Tibayrenc M., (1993). Isoenzyme electrophoresis for parasite characterization. In: Methods inMolecular Biology (ed. Hyde, J.E.). Humana Press: New York., pp 361-382.
Benne R., van den Burg J., Brakenhoff J.P., Sloof P., van Boom J.H. et Tromp M.C., (1986).Major transcript of the frameshifted coxll gene from trypanosome mitochondria contains fournucleotides that are not encoded in the DNA. CeI/46(6) :819-26.
Benne R., (1994). RNA editing in trypanosomes. Eur. J. Biochem. 221 :9-23Berger B.J., Carter N.S. et Fairlamb AH., (1995). Characterisation of pentamidine-resistant
Trypanosoma brucei brucei. Mol. Biochem. Parasitai. 69(2) :289-298.Beverley S.M., Coderre J.A, Santi D.V. et Schimke R.T., (1984). Unstable DNA amplifications in
methotrexate-resistant Leishmania consist of extrachromosomal circles which relocalize duringstabilization. CeI/38(2) :431-439.
Beyer W., Imlay J., et Fridovich 1., (1991). Superoxide dismutases. Prog. Nucleic Acid. Res. Mol.Biol. 40 :221-53.
Bhat G.J., Myler P.J. and Stuart K., (1991). The two ATPase mRNAs of Leishmania tarentolae differat their 3' ends. Mol. Biochem. Parasitai. 48(2) :139-150.
Bhattacharya S., Bakre A et Bhattacharya A, (2002). Mobile genetic elements in protozoan_parasites. J. Genet. 81 (2) :73-86 .
Blanc V., Alfonzo J.D., Aphasizhev R. et Simpson L., (1999). The mitochondrial RNA Iigase fromLeishmania tarentolae can join RNA molecules bridged by a complementary RNA. J. Biol. Chem.274(34) :24289-24296.
Blanc V., Farré J .C., Litvak S. et Araya A., (2002). Réécriture du matériel génétique: fonctions etmécanismes de l'édition de l'ARN. Medicine/Sciences 18 :181-192.
121

Références Bibliographiques
Blum B., Bakalara N. et Simpson L., (1990). A model for RNA editing in kinetoplastidmitochondria : « guide» RNA molecules transcribed from maxicircle DNA provide the editedinformation. Cell 60 :189-198.
Bock M., Gonnert R et Haberkorn A., (1969). Studies with Bay 2502 on animais. Bol. Chi!. Parasitol.24(1) :13-9.
Boeckmann B., Bairoch A., Apweiler R, Blatter M.-C., Estreicher A., Gasteiger E., Martin M.J.,Michoud K., O'Donovan C., Phan 1., Pilbout S. et Schneider M., (2003). The SWISS-PROTprotein knowledgebase and its supplement TrEMBl in 2003. Nuc/eic Acids Res. 31 :365-370.
Bolhuis H., van Veen H.W., Poolman B., Driessen A.J.M. et Konings W.N., (1997). Mechanisms ofmultidrug transporters. FEMS Microbiol. Rev. 21 :55-84.
Bonaldo M.C., D'Escoffier L.N., Salles J.M. et Godenberg S., (1991a). Characterization andexpression of proteases during Trypanosoma cruzi metacyclogenesis. Exp. Parasitol. 73 :44-51.
Bonaldo M.C., Scharfstein J., Murta A.C. et Goldenberg S., (1991 b). Further characterization ofTrypanosoma cruzi GP57/51 as the major antigen expressed by differentiating epimastigotes.Parasitol. Res. 77(7) :567-71.
Bontempi E., Martinez J., Cazzulo J.J., (1989). Subcellular localization of a cysteine proteinase fromTrypanosoma cruzi. Mol. Biochem. Parasitol. 33(1) :43-7.
Borst P. et Ouellette M., (1995). New mechanisms of drug resistance in parasitic protozoa. Annu.Rev. Microbiol. 49 :427-460.
Bouaboula M., Legoux P., Pessegue B., Delpech B., Dumont X., Piechaczyk M., Casellas P. etShire D., (1992). Standardization of mRNA titration using a polymerase chain reaction methodinvolving co-amplification with a multispecific internai control. J. Biol. Chem. 267 :21830 - 21838.
Boveris A. et Stoppani A.O., (1977). Hydrogen peroxide generation in Trypanosoma cruzi.Experientia. 33(10) :1306-1308.
Boveris A., Sies H., Martino E.E., Docampo R, Turrens J.F. et Stoppani A.O., (1980). Deficientmetabolic utilization of hydrogen peroxide in Trypanosoma cruzi. Biochem. J. 188(3) :643-648.
Brener Z., Costa C.A. et Chiari C., (1976). Differences in the susceptibility of Trypanosoma cruzistrains to active chemotherapeutic agents. Rev. Inst. Med. Trop. Sao Paulo 18(6) :450-455.
Brener Z., (1984). Recent advances in chemotherapy of Chagas disease. Mem. Inst. Oswaldo Cruz79 (suppl) :149-155.
Brenière S.F., Bosseno M.F., Revollo S., Rivera M.T. et Tibayrenc M., (1992). Direct identificationof T. cruzi natural clones in vectors and host blood by PCR technique. Am. J. Trop. Med. Hyg.46:335-341.
Brini M. et Carafoli E., (2000). Calcium signalling: a historical account, recent developments andfuture perspectives. Cell Mol. ure Sei. 57(3) :354-70.
Brini M., (2003). Ca(2+) signalling in mitochondria: mechanism and role in physiology and pathology.Cell Calcium. 34(4-5) :399-405.
Brisse S., Barnabé C. et Tibayrenc M., (2000). Identification of six Trypanosoma cruzi phylogeneticlineages by random amplified polymorphie DNA and multilocus enzyme electrophoresis. Int. J.Parasitol. 30 :35-44. .
Buckner F.S., Wilson A.J., White T.C. et van Voorhis W.C., (1998). Induction de la resistance toazoles drug in Trypanosoma cruzi. Antimicrob. Agents Chemother. 42(12) :3245-3250.
Caffrey C.R, Scory S. et Steverding D., (2000). Cysteine proteinases of trypanosome parasites:novel targets for chemotherapy. Curr. Drug Targets. 1(2) :155-62..
Caler E.V., Vaena de Avalos S., Haynes P.A., Andrews N.W. et Burleigh B.A., (1998).Olipeptidase B-dependent signaling mediates host cell invasion by Trypanosoma cruzi. The EMBOJournal 17(17) :4975-4986.
Callahan H.L. et Beverley S.M., (1991). Heavy metal resistance: a new role of P-glycoproteins inLeishmania. J. Biol. Chem. 266 :18427-18430.
Callahan H.L. et Beverley S.M., (1992). A member of the aldoketo reductase family confersmethotrexate resistance in Leishmania. J. Biol. Chem. 267 :24165-24168.
Camandaroba E., (1999). Caractezaçao clonai da cepa Colombiana do Trypanosoma cruzi: estudodo comportamento biol6gico, bioquimico e da resposta terapêutica dos clones isolados. Rev. Soc.Bras. Med. Trop. 32 (supl.l) :81-82.
122

Références Bibliographiques
Camandaroba E.L.P., Reis E.A.G., Gonçalves M.S., Reis M.G. et Andrade S.G., (2003).Trypanosoma cruzi: susceptibility to chemotherapy with benznidazole of clones isolated from thehighly resistant Colombian strain. Rev. Soc. Bras. Med. Trop. 36(2) :201-209.
Campetella O., Martinez J. et Cazzulo J.J., (1990). A major cysteine proteinase is developmentallyregulated in Trypanosoma cruzi. FEMS Microbiol. Lett. 55(1-2) :145-149.
Campetella O., Henriksonn J., Aslund L., Frasch A.C., Pettersson U. et Cazzullo J.J., (1992). Themajor cysteine proteinase (cruzipain) from Trypanosoma cruzi is encoded by multiple polymorphictandemly organized genes located on different chromosomes. Mol. Biochem. Parasitol. 50(2) :225234.
Cançado J.R., (2002).Long term evaluation of etiological treatment of chagas disease withbenznidazole. Rev. Inst. Med. Trop. Sao Paulo. 44(1) :29-37.
Carter N.C. et Fairlamb A.H., (1993). Arsenical-resistant trypanosomes lack an unusual adenosinetransporter. Nature 361 :173-176.
Carter N.S., Berger B.J. et Fairlamb A.H., (1995). Uptake of diamidine drugs by the P2 nucleosidetransporter in melarsen-sensitive and -resistant Trypanosoma brucei brucei. J. Biol. Chem.270(47):28153-7.
Cazzulo J.J., Couso R., Raimondi A., Wernstedt C. et Hellman U., (1989). Further characterizationand partial amino acid sequence of a cysteine proteinase from Trypanosoma cruzi. Mol. Biochem.Parasitol. 33(1) :33-41.
Cazzulo J.J., Cazzulo Franke M.C., Martinez J., Franke de Cazzulo B.M., (1990).Some kinetic properties of a cysteine proteinase (cruzipain) from Trypanosoma cruzi. Biochim.Biophys. Acta. 1037(2) :186-91.
Cazzulo J.J., Stoka V. et Turk V., (1997). Cruzipain, the major cysteine proteinase from theprotozoan parasite Trypanosoma cruzi. Biol. Chem. 378(1) :1-10.
Cazzulo J.J., (1999). La cruzipaina, cisteina proteinasa principal dei Trypanosoma cruzi. Secuencia yorganizaci6n gen6mica de los genes que la codifican. Medicina (Buenos Aires) 59(supl.lI) :7-10.
Cazzulo JJ., (2002). Proteinases of Trypanosoma cruzi: patential targets for the chemotherapy ofChagas disease. Curr. Top. Med. Chem. 2(11) :1261-71.
Cerecetto H. et Gonzalez M., (2002). Chemotherapy of Chagas' disease: status and newdevelopments. Curr. Top. Med. Chem. 2(11) :1187-213.
Chagas C., (1909). Nova tripanozomiaze humana. Estudos sobre a morfologia e 0 ciclo evolutivo doSchizotrypanum cruzi n. gen. n. sp., agente etiologico de nova entitade morbida do homem. Mem.Inst. Oswaldo Cruz 1 :159-218.
Chamond N., Coatnoan N. et Minoprio P., (2002). Immunotherapy of Trypanosoma cruzi infections.Curr. Drug Targets Immune Endocr. Metabol. Disord. 2(3) :247-254.
Chamond N., Gregoire C., Coatnoan N., Rougeot C., Freitas-Junior L.H., da Silveira J.F.,Degrave W.M. et Minoprio P., (2003). Biochemical characterization of proline racemases from thehuman protozoan parasite Trypanosoma cruzi and definition of putative protein signatures. J. Biol.Chem. 278(18) :15484-94.
Chopra 1., O'Neill A.J. et Miller K., (2003). The role of mutators in the emergence of antibioticresistant bacteria. Drug Resist. Updat. 6(3) :137-145.
Chow L.M.C., Wong A.K.C, Ullman B. et Wirth D.F., (1993). Cloning and functional analysis of anextrachromosomally amplified multidrug resistance-like gene in Leishmania enriettii. Mol. Biochem.Parasitol. 60(2) :195-208.
Colley D.G., (2000). Parasitic diseases: opportunities and challenges in the 21st century. Mem. Inst.Oswaldo Cruz. 95 (Suppl 1) :79-87.
Coombs G.H. et Mottram J.C., (1997). Proteinases of trypanosomes and Leishmania. pp 177-197.ln: Trypanosomiasis and Leishmaniasis, biology and control. Hide G., Mottram J.C., Coombs G.H.,and Holmes, P.H. Eds. CAB International, Wallinford UK.
Coons T., Hanson S., Bitonti A.J., McCann P.P. et Ullman B., (1990). Alpha-difluoromethylornithineresistance in Leishmania donovani is associated with increased ornithine decarboxylase activity.Mol. Biochem. Parasitol. 39(1) :77-89.
Coughlin B.C., Teixeira S.M.R., Kirchhoff L.V. et Donelson ,I.E., (2000). Amastin mRNAabundance in Trypanosoma cruzi is controlled by a 3'Untranslated region position-dependent ciselement and an untranslated region-binding protein. J. Biol. Chem. 275(16) :12051-12060.
Croft S.L., (1997). The current status of antiparasite chemotherapy. Parasitology 114 Suppl :S3-15.
123

Références Bibliographiques
Croft s.L., Urbina J.A. et Brun R, (1997). Chemotherapy of human Leishmaniasis andTrypanosomiasis. In : Trypanosomiasis and Leishmania, biology and control. Hide G., Mottram J.C.,Coombs G.H. et Holmes P.M., eds. CAB International Wallinford. pp. 245-257.
Croft S.L., (1999). Pharmacological appraaches to antitrypanosomal chemotherapy. Mem. Inst.Oswaldo Cruz. 94(2) :215-20.
Dallagiovanna B., Castanys S. et Gamarro F., (1994). Trypanosoma eruzi: sequence of the ATPbinding site of a P-glycoprotein gene. Exp. Parasitol. 79 :63-67.
Dallagiovanna B., Gamarro F. et Castanys S., (1996). Molecular characterization of a Pglycoprotein-related gene tepgp2 gene in Trypanosoma eruzi. Mol. Bioehem. Parasitol. 75 :145-157.
De Castro S.L., (1993). The challenge of Chagas' disease chemotherapy: An update of drugsassayed against Trypanosoma eruzi . Acta Tropiea 53(2) :83-89.
Degrave, W., Fragoso, S.P., Britto, C., van Heuverswym, H., Kidane, G.Z., Cardoso, M.A.B.,Mueller, RU., Simpson, L. et Morel, C.M., (1988). Peculiar sequence organization of kinetoplastDNA minicircles from Trypanosoma eruzi. Mol. Bioehem. Parasitol. 27 :63-70.
De Lana M., da Silveira Pinto A., Barnabe C., Quesney V., Noel S. et Tibayrenc M., (1998).Trypanosoma eruzi: compared vectorial transmissibility of three major clonai genotypes by Triatomainfestans. Exp. Parasitol. 90(1) :20-25.
De Lange T., Berkvens T.M., Veeman H.J.G., Frasch A.C.C, Barry J.D. et Borst P., (1984).Comparison of the genes coding for the common 5' terminal sequence of messenger RNAs in threetrypanosome species. Nueleie Aeids Res. 12 :4431 .
De Koning H.P., (2001). Transporters in African trypanosomes: role in drug action and resistance.Int. J. Parasitol. 31 :512-522.
De Mendonça-Lima, L. et Traub-Cseko, Y. M., (1991). A new repetitive DNA sequence fromTrypanosoma eruzi. Mem.lnst. Oswaldo Cruz 86(4) :475.
Demirpence E., Semlali A., Oliva J., Balaguer P., Badia E., Duchesne M.J., Nicolas J.C. et PonsM., (2002). An estrogen-responsive element-targeted histone deacetylase enzyme has anantiestrogen activity that differs fram that of hydroxytamoxifen. Cancer Res. 62(22) :6519-6528.
Denise H. et Barrett M.P., (2000). Uptake and mode of action of drugs used against sleepingsickness. Bioehem. Pharmaeol. 61 (1) :1-5.
Denicola A., Rubbo H., Rodriguez D. et Radi R., (1993). Peroxynitrite-mediated cytotoxicity toTrypanosoma eruzi. Arch. Bioehem. Biophys. 304(1) :279-286.
Descoteaux S., Ayala P., Orozco E. et Samuelson, J., (1992). Primary sequences of two Pglycoprotein genes in Entamoeba histolyfiea. Mol. Bioehem. Parasitol. 54 :201-212.
Dey S., Papadopoulou B., Haimeur A, Roy G., Grondin K., Dou D., Rosen B.P. et Ouellette M.,(1994). High level,.arsenite resistance in Leishmania tarentolae is mediated by an active extrusionsystem. Mol. Bioehem. Parasitol. 67(1) :49-57.
Dey S., Ouellette M., Lightbody J., Papadopoulou B. et Rosen B.P., (1996). An ATP-dependent As(III)-giutathione transport system in membrane vesicles of Leismania tarentolae. Proe. Natf. Aead.Sei. USA 93 :2192-2197.
Diaz Anel A.M., Rossi M.S., Espinosa ..I.M., Guida C., Freitas F.A, Kornblihtt AR, Zingales B.,Flawia M.M. et Torres H.N., (2000). MRNA encoding a putative RNA helicase of the DEAD-boxgene family is up-regulated in trypomastigotes of Trypanosoma eruzi. J. Eukaryot. Mierobiol.47(6):555-60.
Docampo R, de Boiso J.F., Boveris A et Stoppani AO., (1976). Localization of peroxidase activityin Trypanosoma cruzi microbodies. Experientia 32(8) :972-975.
Docampo R, Moreno S.N.J., Stoppani AO.M., Leon W., Cruz F.S., Vil/alta F. et Muniz R.F.A,(1981). Mechanism of Nifurtimox toxicity in different forms of Trypanosoma cruzi. Biochem.Pharmaeol. 30(14) :1947-1951.
Docampo R et Moreno, S.N.J., (1984). Free radical metabolites in the mode of action ofchemotherapeutic agents and phagocytic cells on Trypanosoma cruzi. Rev Infect Dis. 1984 MarApr;6(2) :223-38.
Docampo R et Moreno S.N.J., (1986). Free radical metabolism of antiparasitic agents. FASEB
45(10) :2471-2476.
Docampo R, (2001). Recent developments in the chemotherapy of Chagas disease. Curr. Pharm.Design 7 :1157-1164.
124

Références Bibliographiques
Donelson J.E., Gardner M.J. et EI-Sayed N.M., (1999). More surprises from kinetoplastida. Proc.Nat!. Acad. Sei. USA 96 :2579-2581.
Du X., Hansell E., Engel J.C., Caffrey C.R., Cohen F.E. et McKerrow J.J., (2000). Aryl ureasrepresent a new c1ass of anti-trypanosomal agents. Chem. Biol. 7(9) :733-742.
Dvorak J.A. et Howe C.L., (1977). The effects of Lampit (Bayer 2402) on the interaction ofTrypanosoma cruzi with vertebrate cells in vitro. Am. J. Trop. Med. Hyg. 26 :58-63.
Dvorak J.A., Hartman D.L. et Miles M.A., (1980). Trypanosoma cruzi: correlation of growth kineticsto zymodeme type in clones derived from various sources. J. Protozool. 27(4) :472-474.
Dvorak J.A., (1984). The natural heterogeneity of Trypanosoma cruzi: biological and medicalimplications. J. Cel/ Biochem. 24(4) :357-371.
Eakin A.E., Mills A.A., Harth G., McKerrow J.H. et Craik C.S., (1992).The sequence, organization, and expression of the major cysteine protease (cruzain) fromTrypanosoma cruzi. J. Biol. Chem. 267(11) :7411-20.
Edwards 1., (1993). Nitroimidazole drugs-action and resistance mechanisms. II. Mechanisms ofresistance. J. Antimicrob. Chemother. 31 :201-210.
Ellenberger T.E. et Beverley S.M., (1987a). Biochemistry and regulation of folate and methotrexatetransport in Leishmania major. J. Biol. Chem. 262(21) :10053-10058.
Ellenberger T.E. et Beverley S.M., (1987b). Reductions in methotrexate and folate influx inmethotrexate-resistant lines of Leishmania major are independent of R or H region amplification J.Biol. Chem. 262(28) :13501-13506.
Endicott J.A. et Ling V., (1989). The biochemistry of P-glycoprotein-mediated multidrug resistance.Annu. Rev. Biochem. 58 :137-71.
Engel J.C., Doyle P.S. et McKerrow J.H., (1999). Efecto antiparasitario de inhibidores de cisteinproteasas in vitro e in vivo en la enfermedad de Chagas experimental. Medicina (Buenos Aires) 59(supl.ll) :171-175.
Engels B., (1992). Amplify for PCR experiments. University of Wisconsin, Genetics, Madinson, WI53706.
Esquerdo-G6mez, F., (1996). Maladie de Chagas. ln: Ripert. C. Epidémiologie des maladiesparasitaires. Tome 1. Protozooses. Editions Médicales Internationales Cahan Cedex. pp. 253-301.
Estévez A.M. et Simpson L., (1999). Uridine insertion/deletion RNA editing in trypanosomemitochondria - a review. Gene 240 :247-260.
Faik P., Walker J.I.H., Redmill A.A.M. et Morgan M.J. (1988). Mouse glucose-6-phosphateisomerase and neuroleukin have identical 3' sequences. Nature 332(6163) :455-457.
Feagin J.E., (2000). Mitochondrial genome diversity in parasites. International Journal for Parasitology30 :371-390.
Ferreira L.R., Silva AM., Michailowsky V., Reis L.F. et Gazzinelli R.T., (1999). Expression ofserum amyloid A3 mRNA by inflammatory macrophages exposed to membrane glycoconjugatesfrom Trypanosoma cruzi. J. Leukoc. Biol. 66(4) :593-600.
Ferreira L.R., Abrantes E.F., Rodrigues C.V., Caetano B., Cerqueira G.C., Salim AC., Reis L.F. etGazzinelli R.T., (2002). Identification and characterization of a novel mouse gene encoding a Rasassociated guanine nucleotide exchange factor : expression in macrophages and myocarditiselicited by Trypanosoma cruziparasites. J. Leukoc. Biol. 72(6) :1215-27.
Ferreira-Pinto K.C., Miranda-Vilela A.L., Anacleto C., Fernandes AP., Abdo M.C., PetrilloPeixoto M.L. et Moreira E.S., (1996). Leishmania (V.) guyanensis: isolation and characterization ofglucantime-resistant celilines. Cano J. Microbiol. 42(9) :944-9.
Filardi L.S. et Brener Z.A., (1982). Nitroimidazole-thiadiazole derivative with curative action inexperimental Trypanosoma cruzi infections. Ann. Trop. Med. Parasitol. 76(3) :293-297.
Filardi L.S. et Brener Z., (1987). Susceptibility and natural resistance of Trypanosoma cruzi strains todrugs used clinically in Chagas disease. Trans. R. Soc. Trop. Med. Hyg. 81 :755-759.
Flintoff W.F. et Essani K., (1980). Methotrexate-resistant Chinese hamster ovary cells contain adihydrofolate reductase with an altered affinity for methotrexate. Biochemistry 19(18) :4321-4327.
Foote S.J., Thompson J.K., Cowman AA. et Kemp D.J., (1989). Amplification of the multidrl1gresistance gene in some chloroquine-resistant isolates of P. falciparum. Cel/57 :921-930.
Frash A.C.C., (2000). Functional diversity in the Trans-sialidase and Mucin families in Trypanosomacruzi. Parasitol. Today 16 (7) :282-286.
125

Références Bibliographiques
Franke de Cazzulo B.M., Martinez J., North M .J., Coombs G.H et Cazzulo J.J., (1994). Effects ofproteinase inhibitors on the growth and differentiation of Trypanosoma cruzi. FEMS MicrobiologyLetters 124(1) :81-86.
Frommel T.O. et Balber A.E., (1987). Flow cytofluorimetric analysis of drug accumulation bymultidrug-resistant Trypanosoma brucei brucei and T. b. rhodesiense. Mol. Biochem. Parasitol.26(1-2) :183-91.
Furger A, Schürch N., Kurath U. et Roditi, 1., (1997). Elements in the 3' untranslated region ofProcyclin mRNA regulate expression in insect forms of Trypanosoma brucei by modulating RNAstability and translation. Mol. Cell. Biol. 17(8) :4372-4380.
Gamarro F., Chiquero M.J., Amador M.V., Légaré D., Ouellette M. et Castanys S., (1994). Pglycoprotein overexpression in methotrexate-resistant Leishmania tropica. Biochem. Pharmacol.47(11) :1939-1947.
Garcia E.S. et Dvorak JA. (1982). Growth and development of two Trypanosoma cruzi clones in thearthropod Dipetalogaster maximus. Am. J. Trop. Med. Hyg. 31 (2) :259-62.
Garcia M.P., N6brega O.T., Teixeira AR.L. Sousa M.V. et Santana J.M., (1998). Characterisationof a Trypanosoma cruzi acidic 30kDa cysteine protease. Mol. Biochem. Parasitol. 91 (2) :263-272.
Garvey E.P. et Santi D.V., (1986). Stable amplified DNA in drug-resistant Leishmania exists asextrachromosomal circles. Science 233 :535-540.
Gasteiger E., Gattiker A, Hoogland C., Ivanyi 1., Appel R.D. et Bairoch A., (2003).ExPASy: the proteomics server for in-depth protein knowledge and analysisNucleic Acids Res. 31 :3784-3788.
Gaunt, M. W.; Yeo, M.; Frame, 1.A.; Stothard, J.R.; Carrasco, H.J.; Taylor, M.C.; Solis M., S.;Veazey, P., Miles, G.A.J.; Acosta, N.; Rojas de A., A. et Miles, M.A (2003). Mechanism ofgenetic exchange in American trypanosomes. Nature 421 :936-939.
Gelb M.H. et Hoi W.G., (2002). Parasitology. Drugs to combat tropical protozoan parasites. Science297(5580) :343-4.
Gentilini, M., (1995). Trypanosomiase humaine Américaine ou Maladie de Chagas. ln: Médicinetropicale. Editorial Flammarion Médicine - Sciences. pp. 135-139.
Gibson W., Bingle L., Blendeman W., Brown J., Wood J. et Stevens J., (2000). Structure andsequence variation of the trypanosome spliced leader transcript. Mol. Biochem. Parasitol. 107 :269277.
Gilbert I.H., (2002). Inhibitors of dihydrofolate reductase in leishmania and trypansosomes. Biochim.Biophys. Acta 1587 :249-257.
Gilliland G., Perrin S., Blanchard K. et Bunn H.F., (1990). Analysis of Cytokine mRNA and DNA:Detection and Quantitation by Competitive Polymerase Chain Reaction. Proc. Nat!. Acad. Sci. USA.87(7) :2725-2729.
Gish W., (1996-2003). http://blast.wustl.edu.Godsel L.M. et Engman D.M., (1999). Flagellar protein localization mediated by a calcium
myristoyl/palmitoyl switch mechanism. EMBO J. 18(8) :2057-2065.Goldenberg S., Salles J.M., Contreras, V.T., Lima-Franco M.P., Katzin AM. , Colli, W. et Morel,
C.M., (1985). Characterization of messenger RNA from epimastigotes and metacyclictrypomastigotes of Trypanosoma cruzi. FEBS 180(2) :265-270.
Gonzalez A., Lerner T.J., Huecas M., Sosa-pineda B., Nogueira N. et Lizardi, P.M., (1985).Apparent generation of a segmented mRNA from two separate tandem gene families inTrypanosoma cruzi. Nucleic Acids Res. 13 :5789-5804.
Gonzalez, N., Galindo, 1., Guevara, P., Novak, E., Scorza, J.V., An"ez, N., Da Silveira, J.F. etRamirez, ,I.L., (1994). Identification and detection of T. cruzi using DNA amplification finger printobtained from the ribosomal intergenic spacer. J. Clin. Microbiol. 32 :153-158.
Grace S.C., (1990). Phylogenetic distribution of superoxide dismutase supports an endosymbioticorigin for chloroplasts and mitochondria. Ure Sci. 47(21) :1875-86.
Graham, S. V., (1995). Mechanisms of stage-regulated gene expression in kinetoplastida. Parasitol.Today 1(6) :217-224.
Grogl M., Oduola A.M., Cordero L.D. et Kyle D.E., (1989). Leishmania spp.: Development ofpentostam-resistant clones in vitro by discontinuous drug exposure. Exp. Parasitol. 69(1) :78-90.
Guhl, F. (1998). Estado actual dei control de la enfermedad de Chagas en Colombia. Curso Taller.Control de Tripanosomosis Americana y Leishmaniosis: Aspectos biologicos, genéticos y
126

Références Bibliographiques
moleculares. Universidad de los Andes. Centro de investigaciones en Microbiologia y ParasitologiaTropical-CIMPAT. Santafé de Bogota. pp.47-81.
Guilivi C., Turrens J.F. et Boveris A., (1988). Chemiluminescence enhancement by trypanocidaldrugs and by inhibitors of antioxidant enzymes in Trypanosoma cruzi. Mol. Biochem. Parasitol.30 :243-252.
Gurney M.E., Heinrich S.P., Lee M.R. et Yin H.S., (1986). Molecular cloning and expression ofneuroleukin, a neurotrophic factor for spinal and sensory neurons. Science 234 :566-573.
Haberkorn A. et Gonnert R., (1972). Animal experimental investigation into the activity of nifurtimoxagainst Trypanosoma cruzi. Arzneimittelforschung. 22(9) :1570-82.
Harth G., Andrews N., Mills A.A., Engel J.C., Smith R. et McKerrow J.H., (1993). Peptidefluoromethyl ketones arrest intracellular replication and intercellular transmission of Trypanosomacruzi. Mol. Biochem. Parasitol. 58(1) :17-24.
Hauschka T.S., (1949). Persistence of strain-specific behavior in two strains of Trypanosoma cruziafter prolonged transfer through inbred mice. J. Parasitol. 35 :593-599.
Henderson D.M., Sifri C.D., Rodgers M., Wirth D.F., Hendrickson N. et Ullman, B., (1992).Multidrug-resistance in Leishmania donovani is conferred by amplification of a gene homologous tothe mammalian mdr1 gene. Mol. Cell. Biol. 12 :2855-2865.
Hendrickson N., Sifri C.D., Henderson D.M., Allen T., Wirth D.F. et Ullman B., (1993). Molecularcharacterization of the Idmdr1 multidrug resistance gene from Leishmania donovani. Mol. Biochem.Parasitol. 60 :53-64.
Henriksson, J., (1995). Genomic variation in American trypanosomes. Acta Universitatis Upsaliensis,Uppsala, Sweden.
Henriksson, J., Porcel B., Rydâker M., Ruiz A., Sabaj V., Galanti N., Cazzulo J.J., Frasch A.C.C.et Pettersson U., (1995). Chromosome specific markers reveal conserved linkage groups in spiteof extensive chromosomal size variation in Trypanosoma cruzi. Mol. Biochem. Parasitol. 73 : 63-74.
Henriksson, J., Aslund, L. et Petterson, U., (1996). Karyotype variability in Trypanosoma cruzi.Parasitol. Today 12(3) :108-114.
Hide M., (1999). Etude de l'expression génique différentielle de différentes espèces de Leishmaniapar la technique d'amplification aléatoire de séquences exprimées. DEA de Parasitologie UniversitéMontpellier Il. 34p.
Higgins C.F., (1992). ABC transporters : from microorganismes to man. Annu. Rev. Cell. Biol. 8 :67113.
Higuchi E., Oridate N., Furuta Y., Suzuki S., Hatakeyama H., Sawa H., Sunayashiki-Kusuzaki K.,Yamazaki K., Inuyama Y. et Fukuda S., (2003). Differentially expressed genes associated withCIS-diamminedichloroplatinum (1l)resistance in head and neck cancer using differential display andcDNA microarray. Head Neck. 25(3) :187-193.
Hills M., Hudson C. et Smith, P.G. (1986). Global monitoring of the resistance of malarial parasitesto drugs : statistical treatment of micro-test data. Working paper 2.8.5 Informai consultation on theepidemiology of drug resistance of malaria parasites, Geneva, Switerzland.
Hoare C.A., (1972). The Trypanosomes of Mammals. A zoological monogragh. Blackwell ScientificPublications, Oxford. p. 749.
Horst J.P., Wu T.H. et Marinus M.G., (1999). Escherichia coli mutator genes. Trends Microbiol.7(1) :29-36.
Huang L., Brinen L.S. et Ellman J.A., (2003). Crystal structures of reversible ketone-based inhibitorsof the cysteine protease cruzain. Bioorg. Med. Chem. 11 (1) :21-29.
Hussain S.P., Hofseth L.J., Harris C.C., (2003). Radical causes of cancer. Nat. Rev. Cancer3(4):276-85.
lovannisci D.M., Kaur K., Young L. et Ullman B., (1984). Genetic analysis of nucleoside transport inLeishmania donovani. Mol. Cell. Biol. 4(6) :1013-9.
Ismail 5.0, Paramchuk W., Skeiky Y.A.W., Reed S.G., Bhatia A. et Gedamu L., (1997). Molecularcloning and characterization of two iron superoxide dismutase cDNAs from Trypanosoma cruzi.Mol. Biochem. Parasitol. 86 :187-197.
Ivens A.C., Lewis S.M., Bagherzadeh A., Zhang L., Chan H.M. et Smith D.F., (1998). A physicalmap of the Leishmania major Friedlin Genome. Genome Res. 8(2) :135-145.
Jaccard P., (1908). Nouvelles recherches sur la distribution florale. Bull. Soc. Vaudoise Sei. Nat.44 :223-270.
127

Références Bibliographiques
Jazin, E.E., Bontempi E.J., Sanchez 0.0., Aslund L., Henriksonn J., Frasch A.C. et Pettersson,U., (1995). Trypanosoma cruzi exoantigen is a member of a 160 kDa gene family. Parasitology110:61-69.
Jenner P., (2003). Oxidative stress in Parkinson's disease. Ann Neurol. 53 (Suppl 3) :S26-38.Johnson P.J., (1993). Metronidazole and drug resistance. Parasitol. Today 9(5) :183-186.Juliano RL. et Ling, V., (1976). A surface glycoprotein modulating drug permeability in Chinese
hamster ovary cell mutants. Biochim. Biophys. Acta 455 :152-162.Katakura K., Peng Y., Pithawalla R., Detke S. et Chang K.P., (1991). Tunicamycin-resistant variants
from five species of Leishmania contain amplified DNA in extrachromosomal circles of differentsizes with a transcriptionally active homologous region. Mol. Biochem. Parasitol. 44(2) :233-43.
Kaur K., Coons T., Emmett K. et Ullman B., (1988). Methotrexate-resistant Leishmania donovanigenetically deficient in the folate-methotrexate transporter. J. Biol. Chem. 263(15) :7020-7028.
Kerr 1.0., (2002). Structure and association of ATP-bindig cassette transporter nucleotide-bindingdomains. Biochim. Biophys. Acta 85536 :1-18.
Khaw M. et Panosian C.B., (1995). Human antiprotozoal therapy: past, present, and future. Clin.Microbiol. Rev. 8(3) :427-39.
Kim K.S., Teixeira S.M.R, Kirchhoff L.V. et Donelson J.E., (1994). Transcription and editing ofCytochrome Oxidase Il RNAs in Trypanosoma cruzi. J. Cell Biol. 269(2) :1206-1211.
Krieger M.A, Rodrigues A, Fumie S., Plazanet-Menut C. et Goldenberg S., (1999). Differentiaigene expression during Trypanosoma cruzi metacyclogenesis. Mem. Inst. Oswaldo Cruz. 94 (supl.1) :165-168.
La Flamme A.C., Buckner F.S., Swindle J., Ajioka J. et van Voorhis W.C., (1996). Trypanosomacruzi: expression of interleukin-2 utilizing both supercoiled plasmids and Iinear DNAs. Exp.Parasitol. 83(1) :159-63.
Lammel E.M., Barbieri M.A., Wilkowsky S.E., Bertini F. et Isola EL, (1996). Trypanosoma cruzi:involvement of intraceliular calcium in multiplication and differentiation. Exp. Parasitol. 83(2) :240249.
Laurent, J.P., (1994). Comparaison des propriétes biologiques de différents clones de Trypanosoma(Schizotrypanum) cruzi, Chagas, 1909, agent de la maladie de Chagas. Thèse de doctoratprésentée à l'Université de Montpellier Il, Sciences et Techniques du Languedoc, France pp. 86.
Laurent J.P., Barnabe C., Quesney V., Noel S. et Tibayrenc M., (1997). Impact of clonai evolutionon the biological diversity of Trypanosoma eruzi. Parasitology. 114 ( Pt 3) :213-218.
Le Bras J., (1999). Mécanismes et dynamique des chimiorésistances de Plasmodium faleiparum.Journée SPE du 13 octobre: «Génétique et maladies infectieuses dans l'environnement tropical ».Institut Pasteur, Paris.
Légaré O., Hettema E. et Ouellette, M., (1994). The P-glycoprotein-related gene family inLeishmania. Mol. Biohem. Parasitol. 68 :81-91.
Leon, J.S. et Engman, D.M., (2001). Autoimmunity in Chagas heart disease. Int. J. Parasitol. 31 :555561.
Lewis K., (1994). l'v1ultidrug resistance pumps in bacteria : variations on a theme. Trends Bioehem.Sei. 19 :119-123.
Liang P. et Pardee A.B., (1995). Recent advances in differential display. Curr. Opin. Immunol.7(2):274-280.
Lima AP.C.A., Tessier D.C., Thomas D.Y., Scharfstein J., Storer A.C. et Vernet T., (1994).Identification of new cysteine protease gene isoforms in Trypanosoma eruzi. Mol. Bioehem.Parasitol. 67(2) :333-338.
Lima AP.C.A., dos Reis F.C.G., Serveau C., Lalmanach G., Juliano L., Ménard R, Vernet T.,Thomas D.Y. , Storer AC. et Scharfstein J., (2001). Cysteine protease isoforms from T. eruzi,cruzipain2 and cruzain, present different substrate preference and suceptibility to inhibitors. Mol.Biochem. Parasitol. 114 :41-52.
Lockhart D.J. et Winzeler E.A., (2000). Genomics, gene expression and DNA arrays. Nature405(6788) :827-36.
Luchsinger J.A, Tang M.X., Shea S., Mayeux R., (2003). Antioxidant vitamin intake and risk ofAlzheimer disease. Areh Neurol. 60(2) :203-8.
Macedo, AM. et Pena, S.D.J., (1998). Genetic variability of Trypanosoma cruzi: Implications for thepathogenesis of Chagas disease. Parasitol. Today 14 (3) :119-124.
128

Références Bibliographiques
Macedo, A.M., Pimenta J.R., de Aguiar R.S., Melo A.I.R., Chiari E., Zingales B., Pena S.D.P. etOliveira, R.P., (2001). Usefulness of microsatellite typing in population genetic studies ofTrypanosoma cruzi. Mem. Inst. Oswaldo Cruz 96(3) :407-413.
Maldonado R.A., Mirzoeva S., Godsel L.M., Lukas T.J., Goldenberg S., Watterson D.M. etEngman D.M., (1999). Identification of calcium binding sites in the trypanosome flagellar calciumacyl switch protein. Mol. Biochem. Parasitol. 101(1-2) :61-70.
Mantel N., (1967). The detection of disease c1ustering and a generalized regression approach. CancerRes. 27 :209-220.
Martin F., Maraiion C., Olivares M., Alonso C. et L6pez, M.C., (1995). Characterization of a nonlong terminal repeat retrotransposon cDNA (LHc) from Trypanosoma cruzi: homology of the firstORF with the Ape Family of DNA repair enzymes. J. Mol. Biol. 247 :49-59.
Marr J.J. et Docampo R., (1986). Chemotherapy for Chagas' disease: a perspective of currenttherapy and considerations for future research. Rev. Infect. Dis. 8(6) :884-903.
Marr J.J., (1991). Purine analogs as chemotherapeutic agents in Leishmaniasis and Americantrypanosomiasis. J. Lab. Clin. Med.118 :111-119.
Martinez J., Henrinksson J., Ridâker M., Petterson U. et Cazzulo J.J., (1998). Polymorphisms ofthe genes encoding cruzipain, the major cysteine proteinase of Trypanosoma cruzi, in the regionencoding the C-terminal domain. FEMS Microbiology Letiers 159 :35-39.
Martinez-Diaz R.A., Escario J.A., Nogal-Ruiz J.J. et Gomez-Barrio A., (2001). Biologicalcharacterization of Trypanosoma cruzi strains. Mem. Inst. Oswaldo Cruz. 96(1) :53-9.
Masana M., de Toranzo E.G.D. et Castro J.A., (1984). Reductive metabolism and activation ofBenznidazole. Biochem. Pharmacol. 33(7) :1041-1045.
Masanek U., Stammler G. et Volm M., (2002). Modulation of multidrug resistance in human ovariancancer cell lines by inhibition of P-glycoprotein 170 and PKC isoenzymes with antisenseoligonucleotides. J. Exp. Ther. Oncol. 2(1) :37-41.
Maser P. et Kaminsky R., (1998). Identification of three ABC transporter genes in Trypanasamabrucei spp. Parasitol. Res. 84(2) :106-11.
Matias A., de la Riva J., Martinez E., Torrez M. et Dujardin J.P., (2003). Domiciliation process ofRhodnius stali (Hemiptera: Reduviidae) in Alto Beni, La Paz, Bolivia. Trop. Med. Int. Hea/th.8(3):264-8.
Matthews K.R., Tschudi C. et Ullu E., (1994). A common pyrimidine-rich motif governs trans-splicingand polyadenylation of tubulin polycistronic pre-mRNA in trypanosomes. Genes Dev. 8(4) :491-501.
Matz M.V. et Lukyanov S.A., (1998). Different strategies of differential display: areas of application.Nucleic. Acids. Res. 26(24) :5537-5543.
Maya J.O., Repetto Y., Agosin M., Ojeda J.M., Tellez R., Gaule C. et Morello A., (1997). Effects ofnifurtimox and benznidazole upon glutathione and trypanothione content in epimastigote,trypomastigote and amastigote forms of Trypanasoma cruzi. Mol. Biochem. Parasitol. 86(1) :101106.
Maya J.O., Bollo S., Nunez-Vergara L.J., Squella J.A., Repetto Y., Morello A., Perie J. etChauviere G., (2003). Trypanosama cruzi: effect and mode of action of nitroimidazole andnitrofuran derivatives. Biachem. Pharmacol. 65(6) :999-1006.
McCarthy-Burke, C., Taylor, Z.A., et Buck, G.A., (1989). Characterization of spliced leader genesand transcripts in Trypanosama cruzi. Gene 82 :177-189.
McClelland M., Mathieu-Daudé-é F. et Welsh J., (1995). RNA fingerprinting and differential displayusing arbitrarily primed PCR. Trends Genet. 11(6) :242-246.
McDaniel J.P. et Dvorak J. A., (1993). Identification, isolation, and characterization of naturallyoccurring Trypanosoma cruzi variants Mol. Biachem. Parasitai. 57(2) :213-222.
McKerrow J.H., Sun E., Rosenthal P.J. et Bouvier J., (1993). The proteases and pathogenicity ofparasitic protozoa. Annu. Rev. Microbiol. 47 :821-853.
McKerrow J.H., McGrath M.E. et Engel J.C., (1995). The cysteine protease of Trypanosoma cruzi asa model for antiparasite drug design. Parasitai. Taday 11 (8) :279-282.
McKerrow J.H., (1999). Development of cysteine protease inhibitors as chemotherapy for parasiticdiseases: insights on safety, target validation, and mechanism of action. Int. J. Parasitol. 29 :833837.
McKerrow J.H., Engel J.C. et Caffrey C.R., (1999). Cysteine protease inhibitors as chemotherapy forparasitic infections. Biaarg. Med. Chem. 7(4) :639-644.
129

Références Bibliographiques
McGrath M.E., Eakin A.E., Engel J.C., McKerrow J.H., Craik C.S. et Fletterick R.J., (1995). Thecrystal structure de cruzain : A therapeutic target for Chagas' disease. J. Mol. Biol. 247 :251-259.
Médecins sans Frontières, (2003). Drugs for I\legleted Diseases Iniciative (DNDi): Un espoir pourles malades oubliés. La maladie de Chagas. http://www2.paris.msf.org/
Meirelles M.N.L., Juliano L., Carmona E., Silva S.G., Costa E.M., Murta A.C. et Scharfstein J.,(1992). Inhibitors of the major cysteinyl proteinase (GP57/51) impair host cell invasion and arrestthe intracellular development of Trypanosoma eruzi in vitro. Mol. Bioehem. Parasitol. 52(2):175-184.
Melo R.C. et Brener Z., (1978). Tissue tropism of different Trypanosoma eruzi strains. J. Parasitol.64(3) :475-82.
Michailowsky V., Murta S.M., Carvalho-Oliveira L., Pereira M.E., Ferreira L.R., Brener Z.,Romanha A.J. et Gazzinelli RT., (1998). Interleukin-12 enhances in vivo parasiticidal effect ofbenznidazole during acute experimental infection with a naturally drug-resistant strain ofTrypanosoma eruzi. Antimicrob. Agents. Chemother. 42(10) :2549-56.
Miles, M.A, Souza, A., Povoa, M., Shaw, J. J., Lainson, R. et Toyé, P.J., (1978). Isozymicheterogeneity of Trypanosoma eruzi in the first autochthonous patients with Chagas' disease inAmazonian Brazil. Nature London 272 :819-821.
Miles M.A., Cedillos R.A., Povoa M.M., de Souza A.A., Prata A. et Macedo V., (1981). Do radicallydissimilar Trypanosoma eruzi strains (zymodemes) cause Venezuelan and Brazilian forms ofChagas' disease? Laneet. 1(8234) :1338-40.
Millar A.E. et Kahn S.J., (2000). The SA85-1.1 protein of the Trypanosoma eruzi trans-sialidasesuperfamily is a dominant T-cell antigen. Infect. Immun. 68(6) :3574-80.
Minoprio P., (2001). Parasite polyclonal activators: new targets for vaccination approaches? Int. J.Parasitol. 31 :588-591.
Momen H., (1999). Taxonomy of Trypanosoma cruzi: a commentary on characterization andnomenclature. Mem. Inst. Oswaldo Cruz 94(Suppl. 1) : 181-184.
Moncada C., Reppeto Y., Aldunate J., Letelier M.E. et Morello, A. (1989). Role of glutathione in thesusceptibilityof Trypanosoma eruzi to drugs. Comp Bioehem Physiol C. 94(1) :87-91.
Morel C., Chiari E., Camargo E.P., Mattei D.M., Romanha A.J. et Simpson L. , (1980). Strains andclones of Trypanosoma eruzi can be characterized by pattern restriction endonuclease products ofkinetoplast DNA minicircles. Proe. Nat!. Acad. Sei. U.S.A. 77 :6810-6814.
Moreno S.N., Docampo R, Mason R.P., Leon W., Stoppani A.O., (1982). Different behaviors ofBenznidazole as free radical generator with mammalian and Trypanosoma eruzi microsomalpreparations. Areh. Bioehem. Biophys. 218(2) :585-91.
Moser, D.R, Kirchoff, L.V. et Donelson, J.E., (1989). Detection of Trypanosoma eruzi by DNAamplification using the polymerase chain reaction. J. Clin. Mierobiol. 27 :1477-1482.
Mottram J.C., Brooks D.R et Coombs G.H., (1998). Roles of cysteine proteinases of trypanosomesand Leishmania in host-parasite interactions. Curr. Opin. Mierobiol. 1(4) :455-460.
Muelas-Serrano S., Nogal-Ruiz J.J. et G6mez-Barrio A, (2000). Setting of a colorimetric method todetermine the viability of Trypanosoma cruzi epimastigotes. Parasitol. Res. 86 :999-1002.
Mukhopadhyay R, Dey S., Xu N., Gage O., Lightbody J., Ouellette M. et Rosen B.P., (1996).Trypanothione overproduction and resistance to antimonials and arsenicals in Leishmania. Proe.Natl. Acad. Sei. USA. 93(19) :10383-10387.
Muller L.A., Anasco N. et Gonzalez Cappa S.M., (1986). Trypanosoma eruzi: isolate dependence inthe induction of Iytic antibodies in the mouse and rabbit. Exp. Parasitol. 61 (3) :284-293.
Murphy N.B. et Pellé R, (1994). The use of arbitrary primers and the RADES method for the rapididentification of developmentally regulated gene in trypanosomes. Gene 141 :53-61.
Murta A.C., Persechini P.M., Padron T. de S., de Souza W., Guimaraes J.A . et Scharfstein J.,(1990). Structural and functional identification of GP57/51 antigen of Trypanosoma eruzi as acysteine proteinase. Mol. Bioehem. Parasitol. 43(1) :27-38.
Murta S.M.F. et Romanha AJ., (1998). In vivo selection of a population of Trypanosoma eruzi andclones resistant to Benznidazole. Parasitology 116 :165-171.
Murta S.M., Gazzinelli RT., Brener Z. et Romanha AJ., (1998). Molecular characterization ofsusceptible and naturally resistant strains of Trypanosoma eruzi to benznidazole and nifurtimox.Mol. Bioehem. Parasitol. 93(2) :203-14.
Murta S.M.F., dos Santos W.G., Anacleto C., Nirdé P., Moreira E.S.A. et Romanha A.J., (2001).Drug resistance in Trypanosoma eruzi is not associated with amplification or overexpression of Pglycoprotein (PGP) genes. Mol. Bioehem. Parasitol. 117 :223-228.
130

Références Bibliographiques
Nagano T., Hirano T. et Hirobe M., (1989). Superoxide dismutase mimics based on iron in vivo. J.Biol. Chem. 264 :9243-9.
Nathan C., (2003). Immunology: Oxygen and the inflammatory cell. Nature 422 :675 - 676.Neal R.A et van Bueren J., (1988). Comparative studies of drug susceptibility of five strains of
Trypanosoma cruzi in vivo and in vitro. Trans. R. Soc. Trop. Med. Hyg. 82:709-714.Nirdé P., Larroque C. et Barnabé C., (1995). Drug-resistant epimastigotes of Trypanosoma cruzi and
persistance of this phenotype after differentiation into amastigotes. CR. Acad. Sci. Paris, Sciencesde la vie/Ure Science. 318 :1239-1244.
N6brega O.T., Santos S.M.A., Teixeira A.RL. et Santana J.M., (1998). Cloning and sequencing oftccb, a gene encoding a Trypanosoma cruzi cathepsin B-like protease. Mol. Biochem. ParasitaI.97 :235-240.
Noireau, F. (1999). La enfermedad de Chagas y sus particularidades epidémicas en Bolivia. In:Cassab JA, Noireau, F. et Guillem, G. (eds.), La enfermedad de Chagas en Bolivia.Conocimientos cientificos al inicio dei Programa de Control 1998-2002, Ministerio de Salud yPrevenci6n Social, OMS/OPS, IRD et IBBA, La Paz. pp. 17-47.
Nordberg J. et Arnér E.S.J., (2001). Reactive oxygen species, antioxidants, and the mammalianthioredoxin system. Free Radic. Biol. Med. 31(11) :1287-1312.
Nozaki, T., Engel, J.C. et Dvorak, J.A., (1996). Cellular and molecular biological analyses ofNifurtimox resistance in Trypanosoma cruzi. Am. J. Trop. Med. Hyg. 55(1) :111-117.
Oliveira RP., Broude N.E., Macedo A.M., Cantor C.R., Smith C.L. et Pena S.D.J., (1998). Probingthe genetic population structure of Trypanosoma cruzi with polymorphic microsatellites. Proc. Nat!.Acad. Sei. USA 95 :3776-3780.
Oliveira, R.P., Melo A.I.R., Macedo A.M., Chiari E. et Pena S.D.J., (1999). The population structureof Trypanosoma cruzi: expanded analysis of 54 strains using eight polymorphic CA-repeatmicrosatellites. Mem. Inst. Owaldo Cruz 94(suppl.l) :65-70.
OMS, (1998). Le conseil exécutif demande l'intensification des mesures prises en vue de l'éliminationde la maladie de Chagas et de la lèpre. Communiqué de presse OMS 113. Genève, Suisse.
OPAS/OMS, (1998). Tratamiento etiologico de la enfermedad de Chagas. Conclusiones de unaconsulta técnica. OPC/HCP/HCT/140/99, 32 pp. In : Rev. Patol. Trop. 28 :247-279, 1999).
Opperdoes F.R et Michels P.A.M., (2001). Enzymes of carbohydrate metabolism as potential drugtargets. Int. J. ParasitaI. 31 :482-490.
Ouellette M., Fase-Fowler F. et Borst P., (1990). The amplified H circle of methotrexate-resistantLeishmania tarentolae contains a novel P-glycoprotein gene. EMBO J. 9(4) :1027-1033.
Ouellette M. et Papadopoulou B., (1993). Mechanisms of drug resistance in Leihsmania. ParasitaI.Today9(5) :150-153.
Ouellette M. et Kündig C., (1997). Microbial multidrug resistance. Int. J. Antimicrob. Agents 8 :179187.
Page R D. M., (1996). TREEVIEW: An application ta display phylogenetic trees on personalcomputers. Comput. Appl. Biosci. 12(4) :357-358.
Papadopoulou B. Roy G. et Ouellette M., (1992). A novel antifolate resistance gene on the amplifiedH circle of Leishmania. EMBO J. 11 :3601-3608.
Papadopoulou B., Roy G., Dey S., Rosen B.P. et Ouellette M., (1994). Contribution of theLeishmania P-glycoprotein related gene ItpgpA to oxyanion resistance. J. Biol. Chem. 269 :1198011986.
Papadopoulou B., Roy G., Dey S., Rosen B.P., Olivier M. et Ouellette M., (1996). Gene disruptionof the P-glycoprotein related gene pgpA of Leihsmania tarentalae. Biochem. Biophys. Res.Commun. 224 :772-778.
Pararnchuk W.J., Ismail S.O., Bhatia A. et Gedamu, L., (1997). Cloning, characterization andoverexpression of two superoxide dismutase cDNAs from Leishmania chagasi: role inpathogenesis. Mol. Biochem. ParasitaI. 90 :203-221.
Parekh, H.K., Deng H.B., Choudhary K., Houser S.R. et Sirnpkins H., (2002). Overexpression ofsorcin, a calcium-binding protein, induces a low level of paclitaxel resistance in human ovarian andbreast cancer cells. Biochem. Pharmacal. 63(6) :1149-1158.
Parodi A.J., Labriola C., Cazzulo J.J., (1995). The presence of complex-type oligosaccharides at theC-terminal domain glycosylation site of some molecules of cruzipain. Mol. Biochem. ParasitaI.69(2):247-55.
131

Références Bibliographiques
Parker M.W., Blake C.C., Barra D., Bossa F., Schinina M.E., Bannister W.H., et Bannister J.V.,(1987). Structural identity between the iron- and manganese-containing superoxide dismutases.Protein Eng. 1 :393-7.
Paulsen I.T, Brown M.H. et Skurray RA., (1996). Proton-dependent multidrug efflux systems.Microbiol. Rev. 60 :575-608.
Pays, E. et Vanhamme, L., (1996). Developmental regulation of gene expression in Africantrypanosomes. ln: Smith D.F., Parsons, M., editors. Molecular Biology of Parasitic Protozoa. NewYork: IRL Press. pp. 88-114.
Pearson W.R., Wood T., Zhang Z., et Miller, W. (1997). Comparision of DNA sequences with proteinsequences. Genomics 46: 24-36.
Pérez-Morga, D. et Englund, P.T., (1993). The strucutre of replicating kinetoplast DNA networks. J.Cell Biol. 123 :1069-1079.
Perry K. et Agabian N., (1991). mRNA processing in the Trypanosomatidae. Experientia 47(2) :11828.
Phillips M.A. et Wang C.C., (1987). A Trypanosoma brucei mutant resistant to alphadifJuoromethylornithine. Mol. Biochem. Parasitol. 22(1) :9-17.
Polak A. et Richie R, (1978). Mode of action of the 2-nitroimidazole derivative Benznidazole. Ann.Trop. Med. Parasitol. 72(1) :45-54.
Pontes A.L. et Andrade S.G., (1984). [Reponse to chemotherapy with benznidazole and nifurtimox inmice inoculated with strains of T. cruzi isolated from previously treated animais]. In : Resumos doXX Congresso da Sociedade Brasileira de Medicina Tropical, Salvador-Bahia, 5-9 February, p 25.(in Portuguese).
Porcel B.M., Tran A.N., Tammi M., Nyarady Z., Rydaker M., Urmenyi T.P., Rondinelli E.,Pettersson U., Andersson B. et Aslund L., (2000). Gene survey of the pathogenic protozoanTrypanosoma cruzi. Genome Res. 10(8) :1103-7.
Portela-Lindoso A.A et Shikanai-Yasuda MA, (2003). [Chronic Chagas' disease: fromxenodiagnosis and hemoculture to polymerase chain reaction]. Rev. Saude Publica 37(1) :107-15.Portuguese.
Prata A., (2001). Clinical and epidemiological aspects of Chagas disease. The Lancet 1 :92-100.Pulido M., Martinez-Calvillo S. et Hernandez R, (1996). Trypanosoma cruzi rRNA genes: a
repeated element from the non-transcribed spacer is locus specific. Acta Tropica 62 :163-170.Raether W. et Hanel H., (2003). Nitroheterocyclic drugs with broad spectrum activity. Parasitol. Res.
90 :519-S39.Raaflaub J. et Ziegler W.H., (1979). Single-dose pharmacokinetics of the trypanosomicide
Benznidazole in man. Arzneim.-Forsch./Drug. Res. 29(10) :1611-1614.Raaflaub J., (1980). Multiple-dose kinetics of the trypanosomicide Benznidazole in man. Arzneim.
Forsch./Drug. Res. 30(12) :2192-2194.Rangel H.A., Araujo P.M., Repka D. et Costa M.G., (1981a). Trypanosoma cruzi: isolation and
characterization of a proteinase. Exp. Parasitol. 52(2) :199-209.Rangel H.A., Araujo P.M., Camargo I.J., Bonfitto M., Repka D., Sakurada J.K., Atta A.M., (1981 b).
Detection of a proteinase common to epimastigote, trypomastigote and amastigote of differentstrains of Trypanosoma cruzi. Tropenmed Parasitol. 32(2) :87-92.
Recinos RF., Kirchhoff L.V. et Donelson J.E., (2001). Cell cycle expression of histone genes inTrypanosoma cruzi. Mol. Biochem. Parasitol. 113(2) :21 5-222.
Reina-San Martin B., Degrave W., Rouget C., Cosson A., Chamond N., Cordeiro-Da-Silva A.,Arala-Chaves M., Coutinho A. et Minoprio P., (2000). A B-cell mitogen from a pathogenictrypanosome is a eukaryotic proline racemase. Nature Medicine 6(8) :890-897.
Requena J.M., Jimenez-Ruiz A., Soto M., Lopez M.C. et Alonso C., (1992). Characterization of ahighly repeated interspersed DNA sequence of Trypanosoma cruzi: its potential use in diagnosisand strain classification. Mol. Biochem. Parasitol. 51 (2) :271-280.
Requena, J.M., Martin F., Soto M., Lopez, M.C. et Alonso C., (1994). Characterization of a shortinterspersed reiterated DNA sequence of Trypanosoma cruzi located at the 3'·end of poly (Attranscript. Gene 146(2) :245-250.
Requena, J.M., Lapez, M.C. et Alonso, C., (1996). Genomic repetitive DNA elements ofTrypanosoma cruzi. Parasitol. Today 12(7) :279-283.
132

Références Bibliographiques
Repetto Y., Opazo E., Maya J.O., Agosin M. et Morello A., (1996). Glutathione and trypanothione inseveral strains of Trypanosoma cruzi: effect of drugs. Comp. Biochem. Physiol. B Biochem. Mol.Biol.. 115(2) :281-285.
Revollo S., (1995). Impact de l'évolution c10nale de Trypanosoma cruzi, agent de la maladie deChagas, sur certaines propriétés biologiques médicalement importantes du parasite. Thèse deDoctorat. Université de Montpellier Il. France. 209 p.
Revollo S., Oury B., Laurent J.P., Barnabé C., Quesney V., Carrière V., Noël S. et Tibayrenc M.,(1998). Trypanosoma cruzi: Impact of clonai evolution of the parasite on its biological and medicalproperties. Exp. Parasitol. 89 :30-39.
Roditi 1., (1996). The VSG-procyclin swith. Parasitol. Today 12 :47-49.Rodrigues Coura J. et de Castro S.L., (2002). A critical review on Chagas disease chemotherapy.
Mem. Inst. Oswaldo Cruz. 97(1) :3-24.Ross C.A et Sutherland D.V., (1997). Drug resistance in trypanosomatids. In : Trypanosomiasis and
Leishmaniasis, biology and control. Hide G., 1VI0ttram J.C., Coombs G.H. et Holmes P.M. CABInternattional Wallinford U.K. pp. 259-269.
Rozen S. et Skaletsky H. J., (2000). Primer3 on the WWW for general users and for biologistprogrammers. In: Krawetz S, Misener S (eds). Bioinformatics Methods and Protocols: Methods inMolecular Biology. Humana Press, Totowa, l'JJ, pp 365-386. http://wwwgenome.wi.mit.edu/genome_software/other/primer3.html
Saeftel M., Fleischer B. et Hoerauf A, (2001). Stage-dependent role of nitric oxide in control ofTrypanosoma cruzi infection. Infect. Immun. 69(4) :2252-2259.
Saitou N. et Nei M., (1987). The Neighbor-Joining Method: a new method for constructingphylogenetic trees. Mol. Biol. Evol. 4 :406-425.
Sajid M. et McKerrow, J.H., (2002). Cysteine proteases of parasitic organisms. Mol. Biochem.Parasitol. 120(1) :1-21.
Sakanari J.A., Nadler S.A, Chan V.J., Engel J.C., Leptak C. et Bouvier J., (1997). Leishmaniamajor: comparison of the cathepsin L- and B-Iike cysteine protease genes with those of othertrypanosomatids. Exp. Parasitol. 85(1) :63-76.
Sanchez G., Wallace A., Olivares M., Diaz N., Aguilera X., Apt W. et Solari A, (1990). Biologicalcharacterization of Trypanosoma cruzi zymodemes: in vitro differentiation of epimastigotes andinfectivity of culture metacyclic trypomastigotes to mice. Exp. Parasitol. 71(1) :125-33.
SAS Institute, (2002). StatVieW® software Web site from SAS Institute Inc.(lSAS"). SAS Institute Inc.,SAS Campus Drive, Cary, North Carolina 27513, USA. http://www.statview.com/copyrights.html.
Scharfstein J., Schechter M., Senna M., Peralta J.M., Mendonca-Previato L. et Miles M.A,(1986). Trypanosoma cruzi: characterization and isolation of a 57/51,000 m.w. surface glycoprotein(GP57/51) expressed by epimastigotes and bloodstream trypomastigotes. J. Immunol. 137(4):13361341.
Schenider A, (2001). Unique aspects of mitochondrial biogenesis in trypanosomatis. Int. J. Parasitol.31 :1403-1415.
Schmunis G.A., Zicker F., Pinheiro F. et Brandling-Bennettt, D., (1998). Risk for transfusiontransmitted infectious diseases in Central and South America. Emerg. Infect. Dis. 4(1) :5-11.
Selzer P.M., Pingel S., Hsieh 1., Ugele B., Chan V.J., Engel J.C., Bogyo M., Russell D.G.,Sakanari J.A. et McKerrow J.H., (1999). Cysteine protease inhibitors as chemotherapy: Lessonsfrom a parasite target. Proc. Nat!. Acad. Sci. USA 96 :11015-11022.
Sereno D. et Lemesre J.L., (1997). Axenically cultured forms as an in vitro model for investigation ofantileismanial agents. Antimicrob. Agents Chemother. 41 (5) :972-976.
Sereno O., Michon P., Brajon N. et Lemesre J.L., (1997). Phenotypic characterization of Leishmaniamexicana pentamidine-resistant promastigotes. Modulation of the resistance during in-vitrodevelopmentallife cycle. C. R. Acad. Sci 111.320(12) :981-7.
Serveau C., Lalmanach G., Juliano M.A, Scharfstein J., Juliano L., Gauthier F., (1996).Investigation of the substrate specificity of cruzipain, the major cysteine proteinase of Trypanosomacruzi, through the use of cystatin-derived substrates and inhibitors. Biochem. J. 313 ( Pt 3) :951-56.
Serveau C., Lalmanach G., Hirata 1., Scharfstein J., Juliano M.A et Gauthier F., (1999).Discrimination of cruzipain, the major cysteine proteinase of Trypanosoma cruzi, and mammaliancathepsins B and L, by a pH-inducible f1uorogenic substrate of trypanosomal cysteine proteinases.Eur. J. Biochem. 259(1-2) :275-80.
133

Références Bibliographiques
Silva AM., Pires E.G., Abrantes E.F., Ferreira L.R, Gazzinelli RT. et Reis L.F., (1999). Applicationof the differential display RT-PCR strategy for the identification of inflammation-related mousegenes. Braz. J. Med. Biol. Res. 32(7) :845-52.
Simpson L., (1997). The genomic organization of guide RNA genes in kinetoplastid protozoa: severalconundrums and their solutions. Mol. Biochem. Parasitol. 86 :133-141.
Simpson L., Sbieego S. et Aphasizhev R, (2003). Uridine insertion/delection RNA editing intrypanosome mitochondria : A complex business. RNA 9 :265-276.
Sloof P., Bos J.L., Konings A.F., Menke H.H., Borst P., Gutteridge W.E. et Leon W., (1983).Characterization of satellite DNA in Trypanosoma brucei and Trypanosoma cruzi. J. Mol. Biol.167(1):1-21.
Smith M.W. et Doolittle R.F. (1992). Anomalous phylogeny involving the enzyme glucose-6phosphate isomerase. J. Mol. Evol. 34(6) :544-545.
Smith M.A et Edwards, 0.1., (1995). The influence of microaerophilia and anaerobiosis onmetronidazole uptake in Helicobacter pylori. J. Antimicrob. Chemother. 36(3) :453-461.
Sneath P.H.A. et Sokal RR, (1973). Numerical taxonomy. The principe and practice of numericalclassification. San Francisco: Freeman.
Sohal RS. et Weindrueh R., (1996). Oxidative Stress, Calorie Restriction, and Aging, Science 273 :59-63.
Souto-Padron T., Campetella O.E., Cazzulo J.J. et de Souza W., (1990). Cysteine proteinase inTrypanosoma cruzi: immunocytochemical localization and involvement in parasite-host cellinteraction. J. Cel/. Sci. 96 ( Pt 3) :485-90.
Souto RP., Fernandes O., Maeedo AM., Campbell D.A. et Zingales, B., (1996). DNA markersdefine two major phylogenetic Iineages of Trypanosoma cruzi. Mol. Biochem. Parasitai. 83:141-152.
Steindel M., Dias Neto E., de Menezes C.L., Romanha A.J. et Simpson A.J., (1993). Randomamplified polymorphic DNA analysis of Trypanosoma cruzi strains. Mol. Biochem. Parasitol.60(1):71-9.
Stoesser G., Baker W., van den Broek A., Gareia-Pastor M., Kanz C., Kulikova T., Leinonen R,Lin Q., Lombard V., Lopez R, Maneuso R, Nardone F., Stoehr P., Tuli M.A., Tzouvara K. etVaughan R, (2003). The EMBL Nucleotide Sequence Database: major new developments.Nucleic Acids Res. 31 (1) :17-22.
Stojdl D.F. et Clarke M.W., (1996). Trypanosoma brucei: analysis of cytoplasmic Ca2+ duringdifferentiation of bloodstream stages in vitro. Exp. Parasitol. 83(1) :134-146.
Stoppani A.O.M., (1999). Quimioterapia de la enfermedad de Chagas. Medicina (Buenos Aires)59(supl.ll) :147-~65.
Stothard, J.R., Frê;lme, I.A. et Miles, MA, (1999). Genetic diversity and genetic exchange inTrypanosoma cruzi: dual drug-resistant "progeny" from episomal transformats. Mem. Inst. OswaldoCruz, Rio de Janeiro, 94 (Suppl. 1) :189-193.
Stuart K., Allen T.E., Heidmann S. et Seiwert 5.0., (1997). RNA editing in kinetoplastid protozoa.Microbiol. Mol. Biol. Rev. 61 (1) :105-120.
Sturm, N.R et Simpson, L., (1990). Kinetoplast DNA minicircles encode guide RNAs for editing ofcytochrome oxidase subunit III mRNA. Cel/61 :879-884.
Sutherland I.A., Mounsey A et Holmes P.H., (1992). Transport of isometamidium (Samorin) bydrug-resistant and drug-sensitive Trypanosoma congolense. Parasitology. 104 (Pt 3) :461-467.
Sutherland I.A. et Holmes P.H., (1993). Alterations in drug transport in resistant Trypanosomacongolense. Acta Trop. 54(3-4) :271-278.
Taddei F., Matie 1., Godelle B. et Radman M., (1997). To be a mutator, or how pathogenic andcommensal bacteria can evolve rapidly. Trends Microbiol. 5(11) :427-428.
Tam L.W. et Lefebvre P.A., (2002). The Chlamydomonas MB0210cus encodes a conserved coiledcoil protein important for f1agellar waveform conversion. Cel/ Moti!. Cytoskeleton 51 (4) :197-212.
Tarleton, RL. et Zhang, L. (1999). Chagas disease etiology: Autoimmunity or parasite persistence?Parasitol. Today 15(3) :94-99.
Tarleton, RL., (2001). Parasite persistence in the aetiology of Chagas disease. Int. J. Parasitol.31 :550-554.
Tarrieu F., (2000). Analyse de la diversité phylogénétique de Trypanosoma cruzi cruzi, agent de lamaladie de Chagas, par séquençage des gènes 18S et Tc52. Applications au diagnosticmoléculaire. Thèse de Doctorat d'Université, Université Montpellier Il. 181 p.
134

Références Bibliographiques
TDR/wellcome trust., (2003). Life-cycle of Trypanosoma cruzi. WHO/ TDR.http://www.who.inUtdr/diseases/chagas/lifecycle.htm
Teixeira, A.R, Calixto M.A. et Texeira ML, (1994). Chagas' disease: carcinogenic activity of theantitrypanosomal nitroarenes in mice. Mutat. Res. 305(2) :189-196.
Teixeira S.M.R., Rusell D., Kirchhoff L.V. et Donelson J.E., (1994). A differentially expressed genefamily encoding "amastin", a surface protein of Trypanosoma cruzi amastigotes. J. Biol. Chem.269(32) :20509-20516.
Teixeira S.M.R, Kirchhoff L.V. et Donelson J.E., (1995). Post-transcriptional elements regulatingexpression of mRI\lAs from the Amastinrruzin gene c1uster of Trypanosoma cruzi. J. Biol. Chem.270(38) :22586-22594.
Teixeira, S.M.R, (1998). Control of gene expression in Trypanosomatidae. Braz. J. Med. Biol. Res.31(12) :1503-1516.
Teixeira S.M.R., Kirchhoff L.V. et Donelson J.E., (1999). Trypanosoma cruzi: supression of Tuzingene expression by its 5'-UTR and spliced leader addition site. Exp. Parasitol. 93 :143-151.
Teixeira S.M. et DaRocha W.D., (2003). Control of gene expression and genetic manipulation in theTrypanosomatidae. Genet. Mol. Res. 2(1) :148-158.
Temperton N.J., Wilkinson S.R. et Kelly J.M., (1996). Cloning of an Fe-superoxidase dismutasegene homologue from Trypanosoma cruzi. Mol. Biochem. Parasitol. 76 :339-343.
Teodori E., Dei S., Scapecchi S. et Gualtieri F., (2002). The medicinal chemistry of multidrugresistance (MDR) reversing drugs. Il Farmaco 57 :385-415.
Thellin O., Zorzi W., Lakaye B. , De Borman B., Coumans B., Hennen G., Grisar T., Igout A. etHeinen E., (1999). Housekeeping gene as internai standards: use and Iimits. J. Biotechnol. 75(23):291-295.
Thelu J., Burnod J., Bracchi V. et Ambroise-Thomas P., (1994). Identification of differentiallytranscribed RNA and DNA helicase-related genes of Plasmodium falciparum. DNA Cell Biol.13(11):1109-1115.
Thioulouse J., Chessel D., Dolédec S. et Olivier J.M. (1997). ADE-4: a multivariate analysis andgraphical display software. Statistics and Computing 7(1) :75-83.
Thompson J.D., Gibson T.J., Plewniak F., Jeanmougin F. et Higgins D.G., (1997). The ClustalXwindows interface: flexible strategies for multiple sequence alignment aided by quality analysistools. Nucleic Acids Res. 24 :4876-4882.
Tielens A.G. et van Hellemond, J.J., (1998). Differences in energy metabolism betweenTrypanosomatidae. Parasitol. Today. 14(7) :265-271.
Tielens A.G. et van Hellemond, J.J., (1999). More differences in energy metabolism betweentrypanosomatidae - Reply. Parasitol. Today. 15(8) :347-348.
Tibayrenc M., Ward P., Moya A. et Ayala F.J., (1986). Natural populations of Trypanosoma cruzi,the agent of Chagas' disease, have a complex multiclonal structure. Proc. Nat!. Acad. Sei. USA.83(1):115-119.
Tibayrenc M. et Ayala F.J., (1988). Isozyme variability in Trypanosoma cruzi, the agent of Chagas'disease : genetical, taxonomical, and epidemiological significance. Evolution 42 :277-92.
Tibayrenc M., Neubauer K., Barnabé C., Guerrini F., Skarecky D. et Ayala F.J., (1993). Geneticcharacterization of six parasitic protozoa: parity between random- primer DNA typing andmultilocus enzyme electrophoresis. Proc. Nat!. Acad. Sei. USA 90(4) :1335-1339.
Tibayrenc, M., (1995). Population genetics of parasitic protozoa and other organisms. Adv. Parasitol.36 :47-115.
Tibayrenc, M., (1996). Towards a unified evolutionary genetics of microorganisms. Annu. Rev.Microbiol. 50 :401-429. .
Tibayrenc, M., (1998). Genetic epidemiology of parasitic protozoa and other infectious agents: theneed for an integrated approach. Int. J. Parasitol. 28(1) :85-104.
Tocher J.H., (1997). Reductive activation of nitroheterocyclic compounds. Gen. Pharmac. 28(4) :485487.
Toledo M.J.O., Guilherme A.L.F., da Silva J.C., de Gasperi M.V., Mendes A.P., Gomes M.L. et deAraujo S.M., (1997). Trypanosoma cruzi: chemotherapy with benznidazole in mice inoculated withstrains from Paranâ state and from different endemic areas of Brazil. Rev. Inst. Med. Trop. SaoPaulo. 39(5) :283-290.
135

Références Bibliographiques
Toledo M.J.O., de Lana M., Bahia M.T., Carneiro C.M., Alves C.F., Martins H.R., Tibayrenc M. etTafuri W.L., (2000). Evaluation of chemotherapy with Benznidazole and Itraconazole in miceinfected with different Trypanosoma cruzi clonai genotypes. Mem. Inst. Oswaldo Cruz95(suppl.lI) :314.
Toledo M.J.O., de Lana M., Carneiro C.M., Bahia M.T., Machado-Coelho G.L.L., Veloso V.M.,Barnabé C., Tibayrenc M. et Tafuri W.L., (2002). Impact of Trypanosoma cruzi clonai evolution onits biological properties in mice. Exp. Parasitol. 100(3) :161-72.
Toledo M.J., Bahia M.T., Carneiro C.M., Martins-Filho O.A., Tibayrenc M., Barnabe C., TafuriW.L. et de Lana M., (2003). Chemotherapy with benznidazole and itraconazole for mice infectedwith different Trypanosoma cruzi clonai genotypes. Antimicrob. Agents Chemother. 47(1) :223-30.
Tomâs A.M. et Kelly J.M., (1996). Stage-regulated expression of cruzipain. the major cysteineprotease of Trypanosoma cruzi is independent of the level of RNA. Mol. Biochem. Parasitol. 76 :91103.
Torres C., Barreiro L., Dallagiovanna B., Gamarro F. et Castanys S., (1999). Characterization of anew ATP-binding cassette transporteur in Trypanosoma cruzi associated to LHc retrotrasposon.Biochim. Biophys. Acta 1489 :428-432.
Toyé, P. J., (1974). Isoenzyme variation in isolates of Trypanosoma cruzi. Trans. R. Soc. Trop. Med.Hyg. 68 :147.
Travis J., Potempa J., Maeda H., (1995). Are bacterial proteinases pathogenic factors? TrendsMicrobiol. 3(10) :405-7.
Tyler, K.M. et Engman, D.M., (2001). The life cycle of Trypanosoma cruzi revisited. Int. J. Parasitol.31 :472-481.
Ullman B., (1995). Multidrug resistance and P-glycoproteins in parasitic protozoa.J. Bioenerg. Biomembr. 27(1) :77-84.
Ullu E., Tschudi C. et Günzl A., (1996). Trans-splicing in trypanosomatid protozoa. ln: MolecularBiology of Parasitic Protozoa (Smith D. and Parsons M., eds.) pp115-133. Oxford University Press.Oxford. .'
Upcroft J.A. et Upcroft P., (1993). Drug resistance and Giardia. Parasitol. Today 9(5) :187-190.Urbina J.A., (1999a). Chemotherapy of Chagas' disease: the how and the why. J. Mol. Med.
77(3):332-338.Urbina J.A., (1999b). Parasitological cure of Chagas disease: is it possible? Is it relevant? Mem. Inst.
Oswaldo Cruz 94 (Suppl 1) :349-55.Urbina J.A., (2002). Chemotherapy of Chagas disease. Curr. Pharm. Des. 8(4) :287-95.Ürményi T., de Castro F.T., Carvalho J.F., de Souza W., Rondinelli E., (1992). Transcriptional and
post-transcriptional control of tubulin gene expression in Trypanosoma cruzi. DNA Cel! Biol.11(2):101-109.
van der Spek H., Arts G.J., Zwaal RR. et al., (1991). Conserved genes encode guide RNAs inmitochondria of Crithidia fasciculata. EMBO J. 10 :1217-1224.
Vanhamme L. et Pays E., (1995). Control of gene expression in trypanosomes. Microbiol Rev.59(2):223-40.
van Veen H.W. et Konings W.N., (1997). Multidrug transporters from bacteria to man: similarities instructure and function. Cancer Biology 8 :183-191.
van Veen H.W. et Konings W.N., (1998). The ABC family of multidrug transporters inmicroorganisms. Biochim. Biophys. Acta 1365 :31-36.
Vâzquez M., Lorenzi H., Schijman A.G., Ben-Dov C. et Levin M.J., (1999). Analysis of thedistribution of SIRE in the nuclear genome of Trypanosma cruzi. Gene 239 :207-216.
Vâzquez M., Ben-Dov C., Lorenzi H., Moore T., Schijman A.G. et Levin M.J., (2000). The shortinterspersed repetitive element of Trypanosma cruzi, SIRE, is part of VIPER, an unusualretroelement related to long terminal repeat retrotransposons. Proc. Nat!. Acad. Sei. USA 97 :21282133.
Veloso V.M., Carneiro C.M., Toledo M.J.O., de Lana M., Chiari E., Tafuri W.L. et Bahia M.T.,(2001). Variation in susceptibility to Benznidazole in isolates derived from Trypanosoma cruziparental strains. Mem. Inst. Oswaldo Cruz. 96(7) :1005-1011.
Verbisck N.V., Machado M.R, Engman D.M., Chiurillo M.A., Ramirez J.L., Araya J.E., MortaraRA et da Silveira, J.F., (2003). A novel reiterated family of transcribed oligo(A)-terminated,interspersed DNA elements in the genome of Trypanosoma cruzi. Mem. Inst. Oswaldo Cruz98(1) :129-133.
136

Références Bibliographiques
Vergnes B., Sereno O., Madjidian-Sereno N., Lemesre ~l-L. et Ouaissi A., (2002). CytoplasmicSIR2 homologue overexpression promotes survival of Leishmania parasites by preventingprogrammed cell death. Gene 296 :139-150.
Vespa G.N., Cunha F.Q., Silva J.S., (1994). Nitric oxide is involved in control of Trypanosoma cruziinduced parasitemia and directly kills the parasite in vitro. Infect. Immun. 62(11) :5177-82.
Vidal H., (2000). La RT-PCR et l'analyse de l'expression des gènes. Analyse de l'expression desgènes et Bioinformatique "Nouveaux développements, applications industrielles et biomédicales".Ecole Normale supérieure, Lyon.
Vickerman, K. et Preston, T.M., (1970). Spindie microtubules in the dividing nuclei of trypanosomes.J. Cell Sei. 6 :365-383.
Villanueva, M.S., Williams S.P., Beard C.B., Richards F.F. et Aksoy S., (1991). A new member of afamily of site-specific retrotransposons is present in the spliced leader RNA genes of Trypanosomacruzi. Mol. CeIl. Biol. 11(12) :6139-6148.
Viodé C., Bettache N., Cenas, N., Krauth-Siegel R.L., Chauvière G., Bakalara N. et Périé J. ,(1999). Enzymatic reduction studies of nitroheterocycles. Biochem. Pharmacol. 57(5) :549-557.
Wang X., Lan M., Shi Y.Q., Lu J., Zhong Y.X., Wu H.P., Zai H.H., Ding J., Wu K.C., Pan B.R. et JinJ.P., (2002). DifferentiaI display of vincristine-resistance-related genes in gastric cancer SGC7901cell. World J. Gastroenterol. 8(1) :54-59.
Watanabe H., Takehana K., Date M., Schinozaki T. et Raz A., (1996). Tumor cell autocrine motilityfactor is the neuroleukin/phosphohexose isomerase polypeptide. Cancer Res. 56(13) :2960-3.
Welburn S.C., Lillico S. et Murphy N.B., (1999). Programmed cell death in procyclic formTrypanosoma brucei rhodesiense - identification of differentially expressed genes during Con Ainduced death. Mem. Inst. Oswaldo Cruz. 94(2) :229-234.
Welsh J. et McClelland M., (1990). Fingerprinting genomes using PCR with arbitrary primers. NucleicAcids Res. 18 :7213-7218.
Weston O., Patel B. et van Voorhis W.C., (1999). Virulence in Trypanosoma cruzi infectioncorrelates with the expression of a distinct family of sialidase superfamily genes. Mol. Biochem.Parasitol. 98(1) :105-16.
White T.C., Marr K.A. et Bowden R.A., (1998). Clinical, cellular, and molecular factors thatcontribute to antifungal drug resistance. Clin. Microbiol Rev. 11 (2) :382-402.
Wilson C.M., Serrano A.E., Wasley A., Bogenschutz M.P., Shankar A.H. et Wirth D.F., (1989).Amplification of a gene related to mammalian mdr genes in drug-resistant Plasmodium falciparum.Science 244 :1184-1186.
Wolfger H., Mamnun Y.M. et Kuchler K., (2001). Fungal ABC proteins: pleiotropic drug resistance,stress response and cellular detoxification. Res. Microbiol. 152 :375-389.
WHO - World Health Organization, (2002). Control of Chagas Disease. Second Report of the WHOExpert Committee. Technical Report Series, No 905. WHO, Geneva. 115 p.
Xu W., Seiter K., Feldman E., Ahmed T. et Chiao J.W., (1996). The differentiation and maturationmediator for human myeloid leukemia cells shares homology with neuroleukin or phosphoglucoseisomerase. Blood 87(11) :4502-4506.
Yamamoto K., Okamoto A., Isonishi S., Ochiai K., Ohtake Y., (2001). Heat shock protein 27 wasup-regulated in cisplatin resistant human ovarian tumor cell line and associated with the cisplatinresistance. Cancer Left. 168(2) :173-81.
Zavala-Castro, J.E., Acosta-Viana, K., Baylon-Pacheco, L., Gonzalez-Robles, A., Guzmân-Marin,E. et Rosales-Encina, J.L., (2002). Kinetoplast DNA-binding protein profile in the epimastigotesform of Trypanosoma cruzi. Arch. Med. Res. 33(3) :250-256.
Zayas C.F., Perlino C., Caliendo A., Jackson O., Martinez E.J., Tso P., Heffron T.G., Logan J.L.,Herwaldt B.L., Moore A.C., Steurer F.J., Bem C., Maguire J.H., (2002). Chagas disease afterorgan transplantation - United States, 2001. MMWR Morb. Mortal. Wkly. Rep. 51(10) :210-212.
Zhang L, Tarleton RL., (1999). Parasite persistence correlates with disease severity and localizationin chronic Chagas' disease. Infect. Dis. 180(2) :480-6.
137

ANNEXES
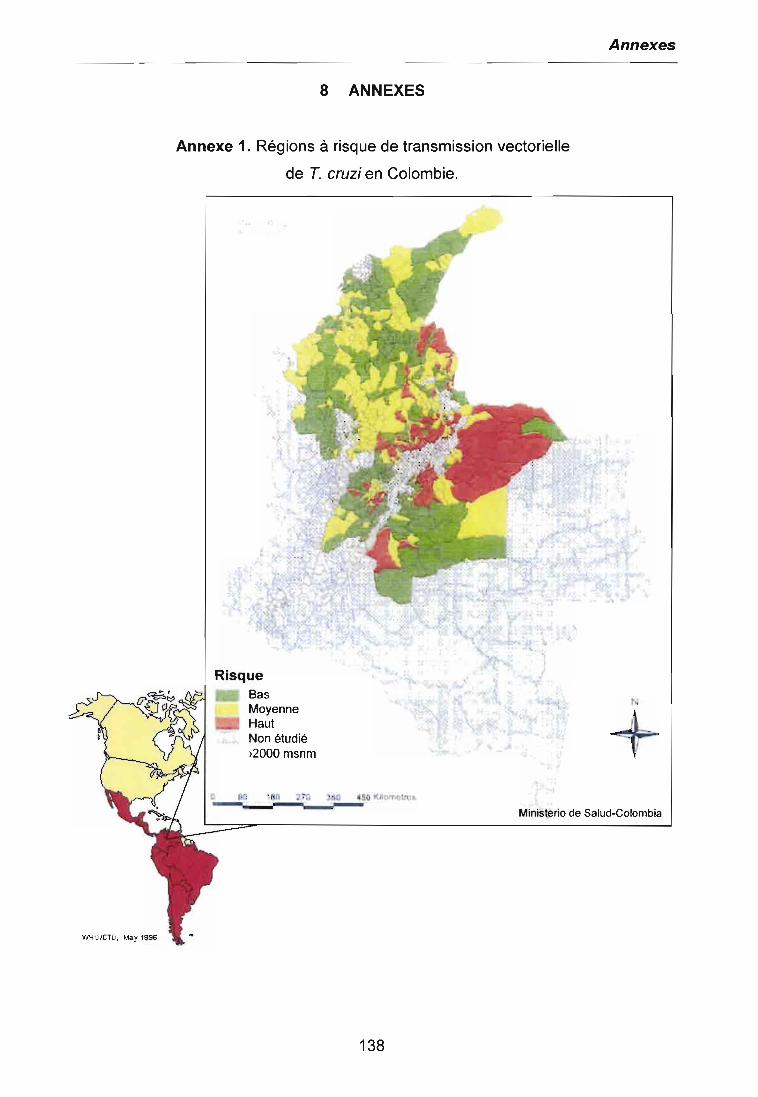
Annexes
8 ANNEXES
Annexe 1. Régions à risque de transmission vectorielle
de T. cruz; en Colombie.
WI-lO/CTC" May 1996
RisqueBasMoyenneHautNon étudié>2000 msnm
lia
+Ministerio de Salud-Colombia
138

Annexes
Annexe 2. Préparation des milieux de culture.
Milieu LIT (Liver Infusion Tryptose)
Pour 5 litres du milieu:
Réactifs
NaCI
Na2HP04
Tryptose
Extrait de levure
Infusion de foie
KCI
Eau déminéralisée
Ajuster le pH à 7,4 avec du NaH2P04 1M.
Stériliser à 120 oC pendant 30 min.
Poids (g)
25
37,5
25
15
15
2
qsq 5 litres
Ajouter 15 ml d'une solution d'Hémine à 10 mg/ml dans un mélange triéthanolamine
eau 1:1, filtrer sur 0,22 IJm.
La Gentamicine est ajoutée à une concentration finale de 50lJg/ml.
Ajouter le sérum de veau fœtal (SVF) à une concentration finale de 10%.
Milieu L30TC
Pour 500 ml du milieu:
Réactifs
Sérum de veau fœtal
TC-100 insect medium (Invitrogen)
Milieu LIT
Filtrer sur filtre 0,22 IJm avant son utilisation.
139
Volume (ml)
50
150
300

Annexe 3. Systèmes enzymatiq~es étudiés.
Annexes
Nom
Aconitase
Alanine amino-transférase
Diaphorase
Enzyme malique
Glucose-phosphate isomérase
Glucose-6-phosphate-déshydrogénase
Glutamate déshydrogénase
Glutamate oxaloacétate transaminase
Glycéraldéhyde-3-phosphate déshydrogénase
Isocitrate déshydrogénase
Leucine aminopeptidase
Malate déshydrogénase
Mannose-phosphate isomérase
Nucléoside hydrolase
Peptidase 1
Peptidase 2
Phosphoglucomutase
Phosphogluconate déshyd rogénase
Superoxyde dismutase
140
Code
E.C. 4.2.1.3
E.C. 2.6.1.2
E.C.1.6.99.1
E.C. 1.1.1.40
E.C. 5.3.1.9
E.C. 1.1.1.49
E.C.1.4.1.3
E.C.2.6.1.1
E.C.1.2.1.12
E.C.1.1.1.42
E.C.3.4.11.1
E.C.1.1.1.37
E.C. 5.3.1.8
E.C.3.2.2.1
E.C.3.4.11
E.C.3.4.13
E.C. 2.7.5.1
E.C. 1.1.1.44
E.C.1.15.1.1
Abréviation
ACON
ALAT
DIA
ME
GPI
G6PDH
GDH
GOT
GAPD
IDH
LAP
MDH
MPI
f\lhi
PEP1
PEP2
PGM
6PGDH
SOD
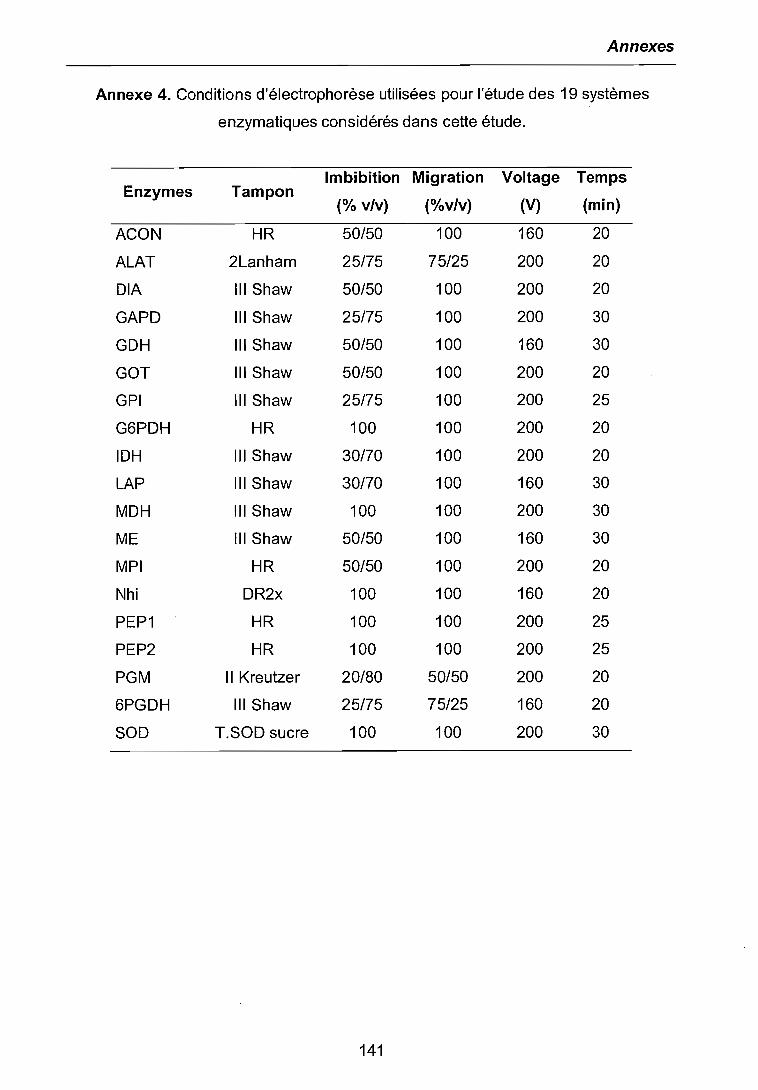
Annexes
Annexe 4. Conditions d'électrophorèse utilisées pour l'étude des 19 systèmes
enzymatiques considérés dans cette étude.
Imbibition Migration Voltage TempsEnzymes Tampon
(% v/v) (%v/v) (V) (min)
ACON HR 50/50 100 160 20
ALAT 2Lanham 25/75 75/25 200 20
DIA III Shaw 50/50 100 200 20
GAPD III Shaw 25/75 100 200 30
GDH III Shaw 50/50 100 160 30
GOT III Shaw 50/50 100 200 20
GPI III Shaw 25/75 100 200 25
G6PDH HR 100 100 200 20
IDH III Shaw 30/70 100 200 20
LAP III Shaw 30/70 100 160 30
MDH III Shaw 100 100 200 30
ME III Shaw 50/50 100 160 30
MPI HR 50/50 100 200 20
I\Ihi DR2x 100 100 160 20
PEP1 HR 100 100 200 25
PEP2 HR 100 100 200 25
PGM Il Kreutzer 20/80 50/50 200 20
6PGDH III Shaw 25/75 75/25 160 20
SOD T.SOD sucre 100 100 200 30
141
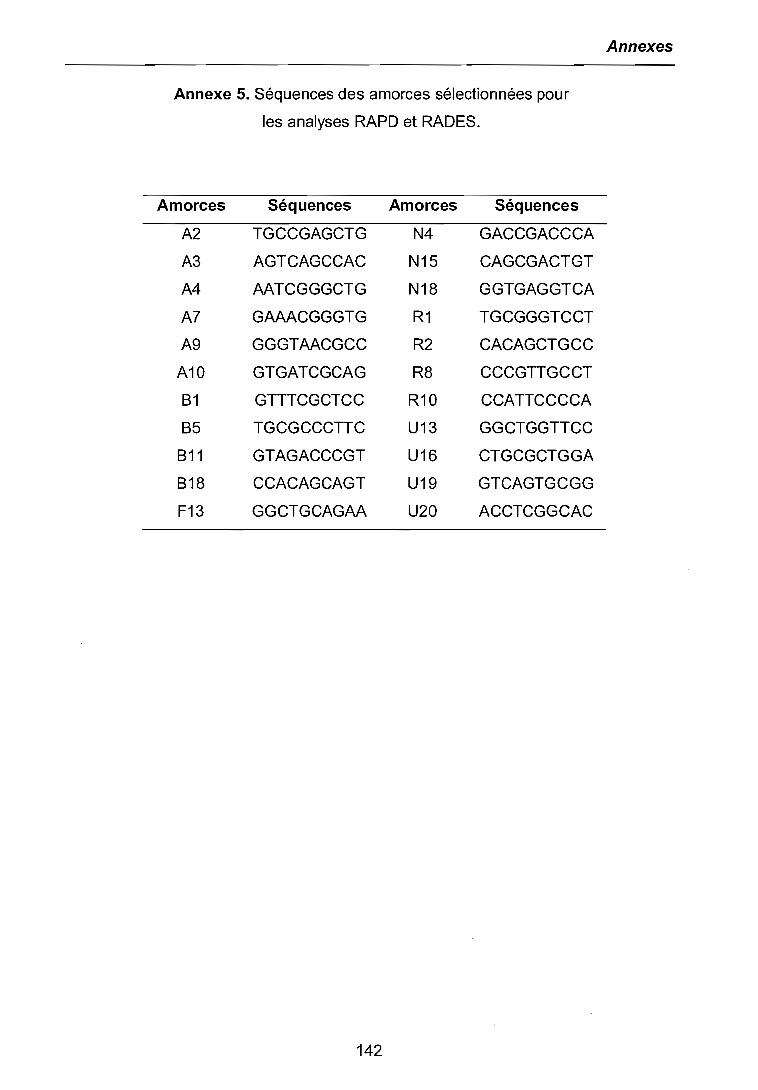
Annexe 5. Séquences des amorces sélectionnées pour
les analyses RAPD et RADES.
Amorces Séquences Amorces Séquences
A2 TGCCGAGCTG N4 GACCGACCCA
A3 AGTCAGCCAC N15 CAGCGACTGT
A4 AATCGGGCTG N18 GGTGAGGTCA
A7 GAAACGGGTG R1 TGCGGGTCCT
A9 GGGTAACGCC R2 CACAGCTGCC
A1ü GTGATCGCAG R8 CCCGTTGCCT
81 GTTTCGCTCC R1ü CCATTCCCCA
85 TGCGCCCTTC U13 GGCTGGTTCC
811 GTAGACCCGT U16 CTGCGCTGGA
818 CCACAGCAGT U19 GTCAGTGCGG
F13 GGCTGCAGAA U20 ACCTCGGCAC
142
Annexes

Annexes
Annexe 6. Solutions utilisées pour l'extraction de l'ARNm à l'aide du kit
Dynabeads® rnRNA Direct kit (Dynal).
Solutions
Tampon de lyse
Tampon de lavage A
Tampon de lavage B
Tampon d'élution
Tampon de reconditionnement
Tampon de stockage oligo (dT)z5
143
Composantes
Tris HCI100 mM, pH 7,5
LiCI500 mM
EDTA 10 mM, pH 8
Dithiothreitol (OTT) 5 mM
LiDS (SOS) 1%
Tris HCI10 mM, pH 7,5
LiCI 0,15 M
EDTA1 mM
LiDS 0,1 %
Tris HCI10 mM, pH 7,5
Liel 0,15 M
EDTA 1 mM
Tris-HC110 mM, pH 7,5
l\JaOH 0,1 M
Tris-HCI 250 mM, pH 7
EDTA 20 mM
Tween®-20 0,1%
Sodium azide 0,02%

Annexes
Annexe 7. Milieux pour la culture bactérienne.
Milieu LB liquide
Pour 1 litre du milieu:
Réactifs
Bacto-tryptone
Bacto-yeast extract
NaCI
Eau déminéralisée
Ajuster le pH à 7 avec NaOH.
Stériliser à 120 oC pendant 30 min.
Poids (g)
10
5
5
qsq 1 litre
L'ampicilline est ajoutée à une concentration finale de 50l-lg/ml, lorsque la solution
est à une température avoisinant 50°C.
LB solide
A 1 litre de milieu LB liquide ajouter 15 9 de Bacto-Agar.
Stériliser à 120 oC pendant 30 min.
L'ampicilline est ajoutée à une concentration finale de 50l-lg/ml, lorsque la solution
est à une température avoisinant 50°C.
Mélanger par inversion et couler immédiatement dans les boîtes de Petri. Conserver
les boîtes à 4°C pour un mois ou à température ambiante pour une semaine.
X-Gal (5-bromo-4-chloro-3-indolyl-,B-D-galactosidase)
Solution à 2% (p:v) dans du diméthylformamide.
Conserver à -20°C à l'abri de la lumière sous aluminium.
IPTG 0,1 M (Isopropyl-thio-galactosidase)
Dissoudre 0,239 9 dans 8 ml d'eau.
Ajuster le volume à 10 ml.
Filtrer sur filtre 0,22I-lm.
Aliquoter et conserver à -20°C.
144
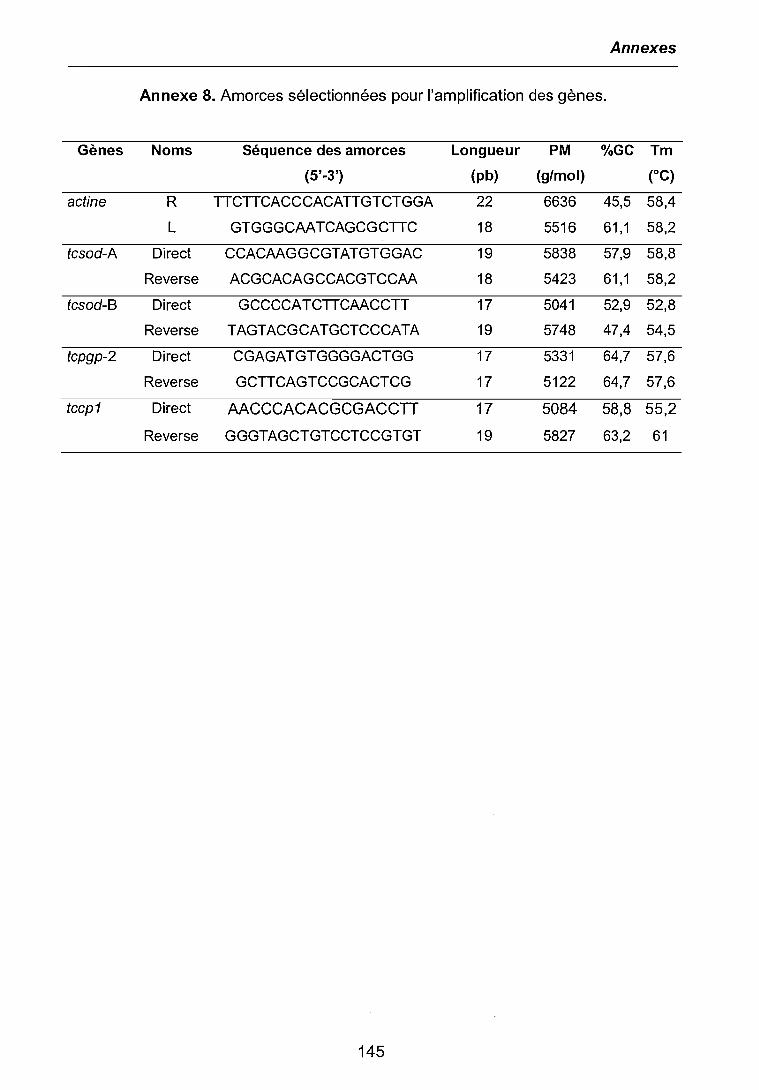
Annexes
Annexe 8. Amorces sélectionnées pour l'amplification des gènes.
Gènes Noms Séquence des amorces Longueur PM %GC Tm
(5'-3') (pb) (g/mol) (OC)
actine R TTCTTCACCCACATTGTCTGGA 22 6636 45,5 58,4
L GTGGGCAATCAGCGCTTC 18 5516 61,1 58,2
tcsod-A Direct CCACAAGGCGTATGTGGAC 19 5838 57,9 58,8
Reverse ACGCACAGCCACGTCCAA 18 5423 61,1 58,2
tcsod-B Direct GCCCCATCTTCAACCTT 17 5041 52,9 52,8
Reverse TAGTACGCATGCTCCCATA 19 5748 47,4 54,5
tcpgp-2 Direct CGAGATGTGGGGACTGG 17 5331 64,7 57,6
Reverse GCTTCAGTCCGCACTCG 17 5122 64,7 57,6
tccp1 Direct AACCCACACGCGACCTT 17 5084 58,8 55,2
Reverse GGGTAGCTGTCCTCCGTGT 19 5827 63,2 61
145
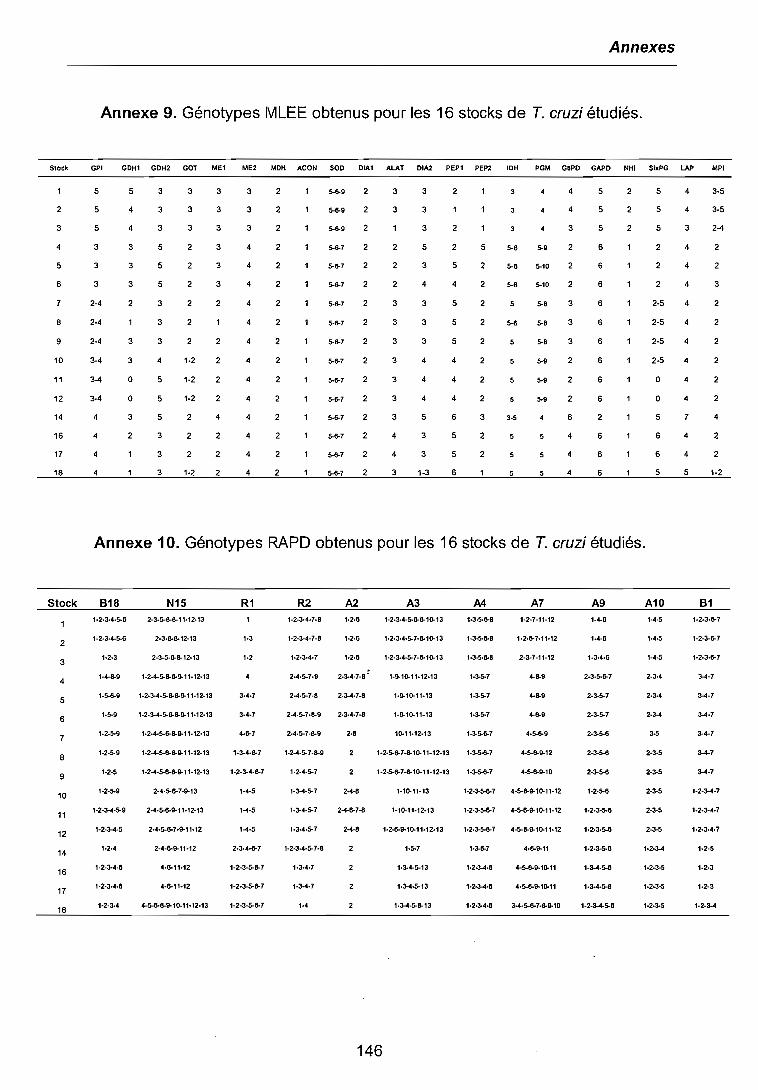
Annexes
Annexe 9. Génotypes IVILEE obtenus pour les 16 stocks de T. cruzi étudiés.
Stock GPI GDHI GDH2 GOT MEl ME2 MDH ACON SOD DIAI ALAT DIA2 PEPI PEP2 IDH PGM G6PD GAPD NH/ SlxPG LAP MPI
2
3
4
5
6
7
8
9
10
11
12
14
16
17
18
5
5
5
3
3
3
2-4
2-4
2-4
3-4
3-4
3-4
4
4
4
5
4
4
3
3
3
2
3
3
o
o
3
2
3
3
3
5
5
5
3
3
3
4
5
5
5
3
3
3
3
3
3
2
2
2
2
2
2
1-2
1-2
1-2
2
2
2
1-2
3
3
3
3
3
3
2
2
2
2
2
4
2
2
2
3
3
3
4
4
4
4
4
4
4
4
4
4
4
4
4
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
5-6-9
5-6-9
5-6-9
5-6-7
5-6-7
5-6-7
5-6-7
5-6-7
5·6-7
5-6-7
5-6-7
5·6-7
5·6-7
5-6-7
5-6-7
5-6-7
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
3
3
2
2
2
3
3
3
3
3
3
3
4
4
3
3
3
3
5
3
4
3
3
3
4
4
4
5
3
3
1-3
2
2
2
5
5
5
5
4
4
4
6
5
5
6
5
2
2
2
2
2
2
2
2
3
2
2
5-6
5-6
5-6
5-6
3·5
5-9
5-10
5-10
5-6
5-6
5-6
5-9
5-9
5-9
4
3
2
2
2
3
3
3
2
2
2
6
4
5
5
5
6
6
6
6
6
6
6
6
6
2
6
6
6
2
2
2
5
5
5
2
2
2
2-5
2-5
2·5
2-5
o
o
5
6
6
5
3
4
4
4
4
4
4
4
7
4
5
3-5
3-5
2-4
2
2
3
2
2
2
2
2
2
4
2
2
1-2
Annexe 10. Génotypes RAPD obtenus pour les 16 stocks de T. cruzi étudiés.
1-5-6-9 1·2·3-4·5-6-6-9-1'.12-13
1·5-9 1·2·3-4·5-6-6-9-11·12·13
1·2-5-9 1·2-4-5-6-6-9-11.12·13
1-2-5-9 1·2-4-5-6-6-9-11-12-13
1·2·5 1·2-4·5-6-6-9-11·12-13
1-2-5-9 2·4·5-6-7·9-13
1-2·3-4·5-9 2-4·5-6-11-11.12-13
1·2-3-4-5 2-4·5-6-7-11-11.12
1-2-4 2-4-6-11-11·12
1-2-3-4·6 4·6-11-12
1-2-3·4.6 4-6-11-12
1·2-3-4 4-5-6-6-11-1(1.11·12-13
2-4·5-7-6 2-3-4·7-6 1·9-1(1.11·13 1-3-5-7 4·6-9
2-4·5-7·6-9 2·3·4·7·8 1-9-1(1.11-13 1·3-5-7 4-6-9
2·4·5-7-6-9 2·6 1(1.11·12-13 1-3-5-6-7 4·5-6-9
1·2-4·5-7-6-9 1·2·5-6-7·6-1(1.11-12·13 1-3-5-6-7 4·5-6-9-12
1·2·4·5-7 1-2·5-6-7·6-1(1.11·12·13 1-3-5-6-7 4-5-6-11-10
1·3-4·5-7 2-4-8 1·1(1.11-13 1·2·3-5-6-7 4-5-6-11-1(1.11·12
1·3-4-5-7 2-4-6-7·8 1·1(1.11-12·13 1-2-3-5-6-7 4-5-6-11-1(1.11·12
1.3-4-5-7 2-4·6 1·2·6-11-1(1.11-12·13 1·2·3-5-6-7 4-5-6-11-1(1.11·12
1-2·3-4·5-7-6 1·5-7 1·3-6-7 4·6-11-11
1·3-4-7 1·3-4-5-13 1·2-3-4·6 4-5-6-11-1(1.11
1·3-4·7 1·3-4·5-13 1·2-3-4·6 4-5-6-11-1(1.11
1·4 1.3-4-5-6-13 1·2·3-4-6 3-4-5-6-7-6-11-10
Stock
2
3
5
6
7
8
9
10
11
12
14
16
17
18
818
1-2·3-4·5-6
1-2·3·4·5-6
1·2·3
1·4·6-9
N15
2·3-5-6-6-11·12·13
2·3-6-6-12-13
2-3-5·6-6-12·13
1-2-4-5-6-6-9-11·12·13
R1
1·3
1·2
3-4·7
3·4.7
4·6-7
1·3-4·6-7
1-2-3-4.6-7
1-4·5
1-4·5
1-4·5
2·3-4·6-7
1-2-3-5-6-7
1-2·3-5-6-7
1-2-3-5-6-7
R2
1-2-3-4·7·6
1·2-3·4·7·8
1·2·3·4·7
2·4·5-7·9
A21·2·6
1·2·6
1·2·6
2·3-4-7-6·
A3
1·2·3·4·5-6-6-1(1.13
1·2·3-4·5-7·6-1(1.13
1·2·3-4·5-7-6-1(1.13
1-9-1(1.11·12·13
A41·3-5-6-6
1-3-5-6-6
1·3-5-6-6
1·3-5-7
A7
1·2-7-11·12
1·2·6-7·11·12
2-3-7-11-12
4-6-9
A91·4-6
1-4·6
1·3-4·6
2·3-5-6-7
2·3-5-7
2-3-5-7
2·3·5-6
2·3-5-6
2·3-5-6
1·2·5-6
1·2·3-5-6
1·2·3-5-6
1·2·3-5-6
1·3-4·5-6
1·3-4-5-6
1·2·3-4-5-8
A101-4·5
1·4·5
1-4·5
2·3-4
2·3-4
2·3-4
3-5
2·3-5
2-3-5
2-3·5
2·3·5
2-3-5
1·2·3-4
1-2·3-5
1·2-3-5
1-2·3-5
81
1·2-3·6-7
1·2-3·6-7
1·2·3-6-7
3-4-7
3-4-7
3-4·7
3-4-7
3-4-7
3-4.7
1·2-3-4-7
1·2·3-4.7
1·2·3-4-7
1·2-5
1·2·3
1·2-3
1·2·3-4
146
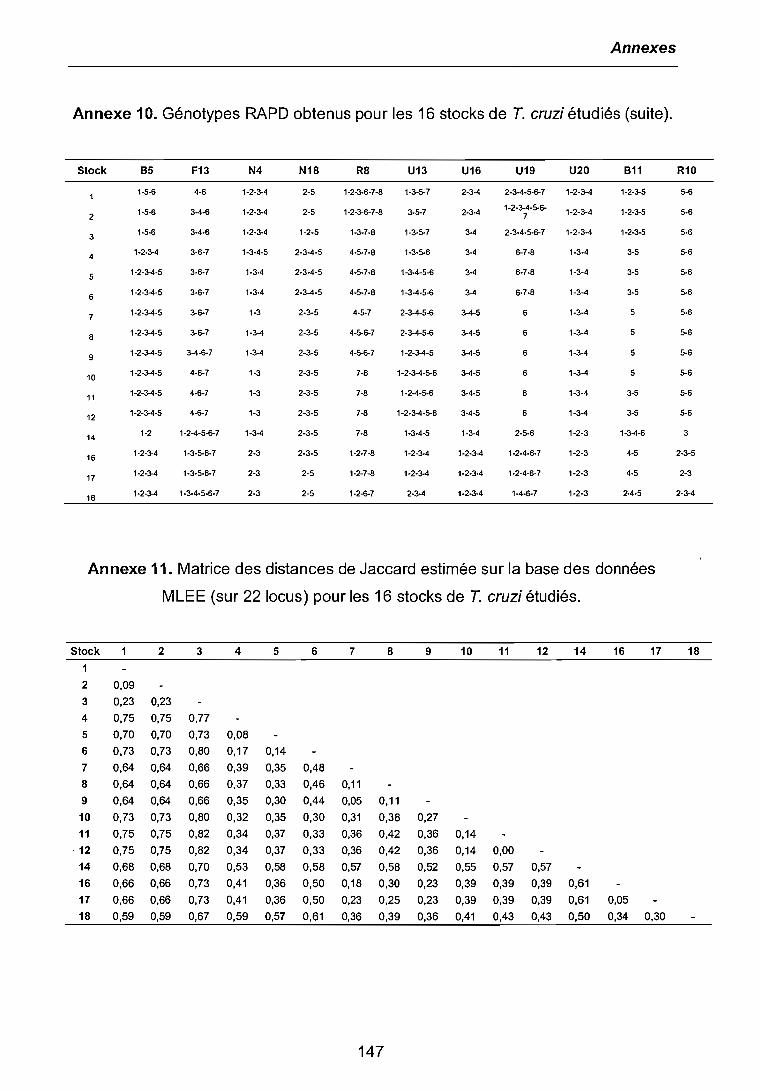
Annexes
Annexe 10. Génotypes RAPD obtenus pour les 16 stocks de T. cruzi étudiés (suite).
Stock 85 F13 N4 N18 R8 U13 U16 U19 U20 811 R10
1-5-6 4-6 1-2-3-4 2-5 1-2-3-6-7-8 1-3-5-7 2-3-4 2-3-4-5-6-7 1-2-3-4 1-2-3-5 5-6
1-5-6 3-4-6 1-2-3-4 2-5 1-2-3-6-7-8 3-5-7 2-3-41-2-3-4-5-6-
1-2-3-4 1-2-3-5 5-62 7
3 1-5-6 3-4-6 1-2-3-4 1-2-5 1-3-7-8 1-3-5-7 3-4 2-3-4-5-6-7 1-2-3-4 1-2-3-5 5-6
4 1-2-3-4 3-6-7 1-3-4-5 2-3-4-5 4-5-7-8 1-3-5-6 3-4 6-7-8 1-3-4 3-5 5-6
5 1-2-3-4·5 3-6-7 1-3·4 2-3-4-5 4-5-7-8 1-3-4-5-6 3-4 6-7·8 1-3-4 3-5 5-6
6 1-2-3-4·5 3-6-7 1-3-4 2-3-4-5 4-5-7-8 1-3-4-5-6 3-4 6-7-8 1-3-4 3-5 5-6
7 1-2-3-4-5 3-6-7 1-3 2-3-5 4-5-7 2-3-4-5-6 3-4-5 6 1-3-4 5 5-6
8 1-2-3-4-5 3-6-7 1-3-4 2-3-5 4-5-6-7 2-3-4-5-6 3-4-5 6 1-3-4 5 5-6
91-2-3-4-5 3-4-6-7 1-3-4 2-3-5 4-5-6-7 1-2-3-4-5 3-4-5 6 1-3-4 5 5-6
101-2-3-4-5 4-6-7 1-3 2-3-5 7-8 1-2-3-4-5-6 3-4-5 6 1-3-4 5 5-6
11 1-2-3-4-5 4-6·7 1-3 2-3-5 7-8 1-2-4-5-6 3-4-5 6 1-3-4 3·5 5-6
121-2-3-4-5 4·6-7 1-3 2-3-5 7·8 1-2-3-4-5-6 3-4-5 6 1-3-4 3-5 5-6
14 1-2 1-2-4-5-6-7 1-3-4 2-3-5 7-8 1-3-4-5 1-3-4 2-5-6 1-2-3 1-3-4-6 3
16 1-2-3-4 1-3-5-6-7 2-3 2-3-5 1-2-7-8 1-2-3-4 1-2-3-4 1-2-4-6-7 1-2-3 4-5 2-3-5
171-2-3-4 1-3-5-6-7 2-3 2-5 1-2-7-8 1-2-3-4 1-2-3-4 1-2-4-6-7 1-2-3 4-5 2-3
181-2-3-4 1-3·4-5-6-7 2-3 2-5 1-2-6-7 2-3-4 1-2-3-4 1-4-6-7 1-2-3 2-4-5 2-3-4
Annexe 11. Matrice des distances de Jaccard estimée sur la base des données
MLEE (sur 22 locus) pour les 16 stocks de T. cruzi étudiés.
Stock 2 3 4 5 6 7 8 9 10 11 12 14 16 17 18
1
2 0,09
3 0,23 0,23
4 0,75 0,75 0,77
5 0,70 0,70 0,73 0,08
6 0,73 0,73 0,80 0,17 0,14
7 0,64 0,64 0,66 0,39 0,35 0,48
8 0,64 0,64 0,66 0,37 0,33 0,46 0,11
9 0,64 0,64 0,66 0,35 0,30 0,44 0,05 0,11
10 0,73 0,73 0,80 0,32 0,35 0,30 0,31 0,38 0,27
11 0,75 0,75 0,82 0,34 0,37 0,33 0,36 0,42 0,36 0,14. 12 0,75 0,75 0,82 0,34 0,37 0,33 0,36 0,42 0,36 0,14 0,00
14 0,68 0,68 0,70 0,53 0,58 0,58 0,57 0,58 0,52 0,55 0,57 0,57
16 0,66 0,66 0,73 0,41 0,36 0,50 0,18 0,30 0,23 0,39 0,39 0,39 0,61
17 0,66 0,66 0,73 0,41 0,36 0,50 0,23 0,25 0,23 0,39 0,39 0,39 0,61 0,05
18 0,59 0,59 0,67 0,59 0,57 0,61 0,36 0,39 0,36 0,41 0,43 0,43 0,50 0,34 0,30
147

Annexes
Annexe 12. Matrice des distances de Jaccard estimée sur la base des données
RAPD (sur 22 amorces) pour les 16 stocks de T. cruzi étudiés.
Stock 2 3 4 5 6 7 8 9 10 11 12 14 16 17 18
1
2 0,08
3 0,15 0,16
4 0,62 0,63 0,59
5 0,60 0,60 0,58 0,11
6 0,60 0,60 0,58 0,12 0,02
7 0,63 0,62 0,62 0,32 0,30 0,28
8 0,57 0,56 0,57 0,34 0,30 0,29 0,13
9 0,55 0,55 0,53 0,36 0,33 0,33 0,18 0,07
10 0,55 0,58 0,56 0,42 0,39 0,39 0,26 0,27 0,26
11 0,52 0,55 0,53 0,37 0,37 0,37 0,29 0,30 0,29 0,10
12 0,52 0,55 0,52 0,38 0,38 0,38 0,30 0,29 0,27 0,09 0,06
14 0,59 0,60 0,58 0,56 0,53 0,54 0,57 0,50 0,47 0,52 0,50 0,48
16 0,54 0,53 0,55 0,63 0,63 0,63 0,58 0,53 0,50 0,51 0,50 0,49 0,43
17 0,54 0,53 0,55 0,65 0,65 0,65 0,61 0,56 0,53 0,54 0,53 0,51 0,44 0,03
18 0,56 0,54 0,57 0,64 0,65 0,65 0,58 0,52 0,49 0,55 0,54 0,52 0,49 0,20 0,18
Annexe 13. Matrice des distances estimée sur la base des données d'IC50 pour les
16 stocks de T. cruzi étudiés.
Stock 2 3 4 5 6 7 8 9 10 11 12 14 16 17 18
1
2 19,90
3 25,70 5,80
4 20,80 0,90 4,90
5 25,40 5,50 0,30 4,60
6 23,30 3,40 2,40 2,50 2,10
7 13,20 6,70 12,50 7,60 12,20 10,10
8 23,60 3,70 2,10 2,80 1,80 0,30 10,40
9 22,10 2,20 3,60 1,30 3,30 1,20 8,90 1,50
10 17,60 2,30 8,10 3,20 7,80 5,70 4,40 6,00 4,50
11 20,00 0,10 5,70 0,80 5,40 3,30 6,80 3,60 2,10 2,40
12 22,60· 2,70 3,10 1,80 2,80 0,70 9,40 1,00 0,50 5,00 2,60
14 24,80 4,90 0,90 4,00 0,60 1,50 11,60 1,20 2,70 7,20 4,80 2,20
16 24,00 4,10 1,70 3,20 1,40 0,70 10,80 0,40 1,90 6,40 4,00 1,40 0,80
17 23,80 3,90 1,90 3,00 1,60 0,50 10,60 0,20 1,70 6,20 3,80 1,20 1,00 0,20
18 24,20 4,30 1,50 3,40 1,20 0,90 11,00 0,60 2,10 6,60 4,20 1,60 0,60 0,20 0,40
148
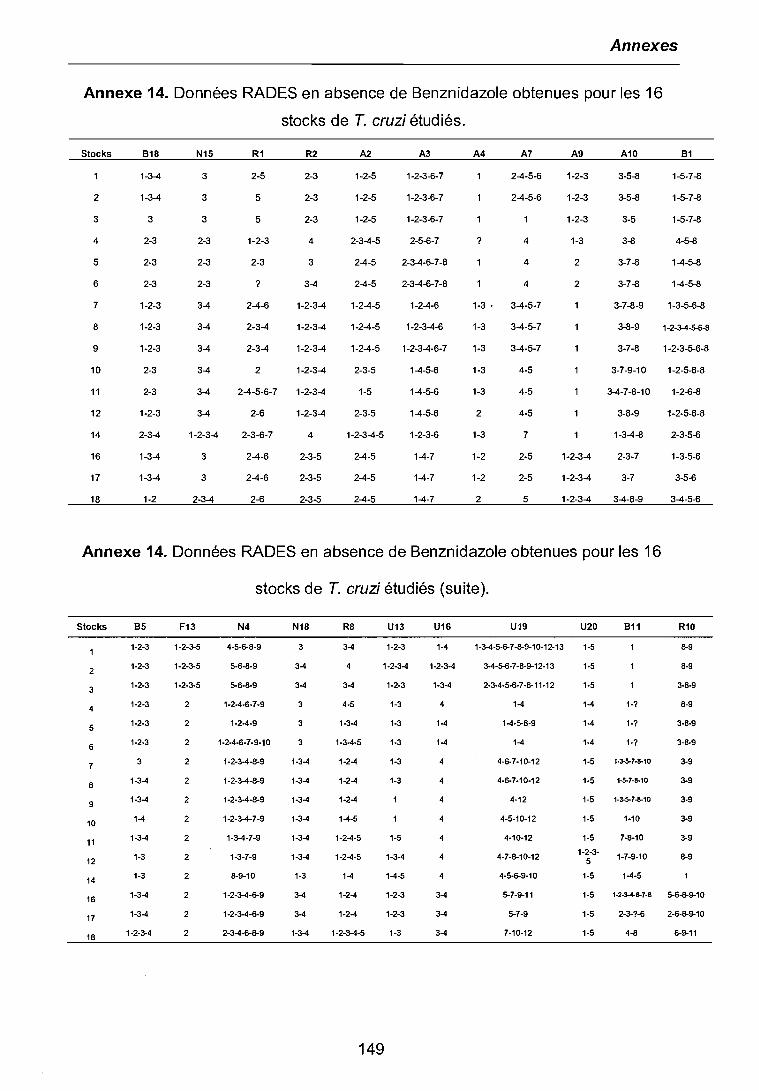
Annexes
Annexe 14. Données RADES en absence de Benznidazole obtenues pour les 16
stocks de T. cruzi étudiés.
Stocks
2
3
4
5
6
7
8
9
10
11
12
14
16
17
18
818
1-3-4
1-3-4
3
2-3
2-3
2-3
1-2-3
1-2-3
1-2-3
2-3
2-3
1-2-3
2-3-4
1-3-4
1-3-4
1-2
N15
3
3
3
2-3
2-3
2-3
3-4
3-4
3-4
3-4
3-4
3-4
1-2-3-4
3
3
2-3-4
R1
2-5
5
5
1-2-3
2-3
?
2-4-6
2-3-4
2-3-4
2
2-4-5-6-7
2-6
2-3-6-7
2-4-6
2-4-6
2-6
R2
2-3
2-3
2-3
4
3
3-4
1-2-3-4
1-2-3-4
1-2-3-4
1-2-3-4
1-2-3-4
1-2-3-4
4
2-3-5
2-3-5
2-3-5
A2
1-2-5
1-2-5
1-2-5
2-3-4-5
2-4-5
2-4-5
1-2-4-5
1-2-4-5
1-2-4-5
2-3-5
1-5
2-3-5
1-2-3-4-5
2-4-5
2-4-5
2-4-5
A3
1-2-3-6-7
1-2-3-6-7
1-2-3-6-7
2-5-6-7
2-3-4-6-7-8
2-3-4-6-7-8
1-2-4-6
1-2-3-4-6
1-2-3-4-6-7
1-4-5-6
1-4-5-6
1-4-5-6
1-2-3-6
1-4-7
1-4-7
1-4-7
A4
?
1-3 .
1-3
1-3
1-3
1-3
2
1-3
1-2
1-2
2
A7
2-4-5-6
2-4-5-6
4
4
4
3-4-5-7
3-4-5-7
3-4-5-7
4-5
4-5
4-5
7
2-5
2-5
5
A9
1-2-3
1-2-3
1-2-3
1-3
2
2
1-2-3-4
1-2-3-4
1-2-3-4
A1D
3-5-6
3-5-6
3-5
3-6
3-7-6
3-7-6
3-7-6-9
3-6-9
3-7-8
3-7-9-10
3-4-7-8-10
3-6-9
1-3-4-6
2-3-7
3-7
3-4-8-9
81
1-5-7-6
1-5-7-6
1-5-7-8
4-5-6
1-4-5-6
1-4-5-6
1-3-5-6-8
1-2-3-4-5-6-8
1-2-3-5-6-8
1-2-5-6-8
1-2-6-6
1-2-5-6-8
2-3-5-6
1-3-5-6
3-5-6
3-4-5-6
Annexe 14. Données RADES en absence de Benznidazole obtenues pour les 16
stocks de T. cruzi étudiés (suite).
Stocks
2
3
4
S
6
7
8
9
10
11
12
14
16
17
18
85
1-2-3
1-2-3
1-2-3
1-2-3
1-2-3
1-2-3
3
1-3-4
1-3-4
1-4
1-3-4
1-3
1-3
1-3-4
1-3-4
1-2-3-4
F13
1-2-3-S
1-2-3-S
1-2-3-S
2
2
2
2
2
2
2
2
2
2
2
2
2
N4
4-S·6-8-9
S-6-8-9
S-6-8-9
1-2-4·6-7-9
1-2-4-9
1-2-4-6-7-9-10
1-2-3-4-8-9
1-2-3-4·8-9
1-2-3-4-8-9
1-2-3-4-7-9
1-3-4-7-9
1-3-7-9
8-9-10
1-2-3-4-6-9
1-2-3-4-6-9
2-3-4-6-8-9
N18
3
3-4
3-4
3
3
3
1-3-4
1-3-4
1-3-4
1-3-4
1-3-4
1-3-4
1-3
3-4
3-4
1-3-4
R8
3-4
4
3-4
4·S
1-3-4
1-3-4·S
1-2-4
1-2-4
1-2-4
1-4-S
1-2-4-S
1-2-4-S
1-4
1-2-4
1-2·4
1-2-3-4-S
U13
1-2-3
1-2-3-4
1-2-3
1-3
1-3
1-3
1-3
1-3
1-S
1-3-4
1-4-S
1-2-3
1-2-3
1-3
U16
1-4
1-2-3-4
1-3-4
4
1-4
1-4
4
4
4
4
4
4
4
3-4
3-4
3-4
U19
1-3-4-S·6-7-8-9-10-12-13
3-4-5-6-7-8-9-12-13
2-3-4-S-6-7-8-11·12
1-4
1-4-5-8·9
1-4
4-6-7-10-12
4-6-7-10-12
4·12
4-S-10-12
4-10-12
4-7-8-10-12
4-S-6-9-10
5-7-9-11
5-7-9
7-10-12
U2D
1-S
1-S
1-S
1-4
1-4
1·4
1-S
1-S
1-S
1-S
1-S
1-2-3S
1-S
1-S
1-S
1-S
811
1-?
1-?
1·?
1·3-5-7·8-10
1·5-7·8-10
1-3-5-7-8-10
1·10
7-8-10
1-7-9-10
1-4-S
1·2·3-4-8-7·8
2-3-?-6
4-8
R1D
8-9
8·9
3-8-9
8-9
3-8·9
3-8-9
3-9
3-9
3·9
3-9
3-9
8·9
S-6-8-9-10
2-6·8-9-10
6-9-11
149
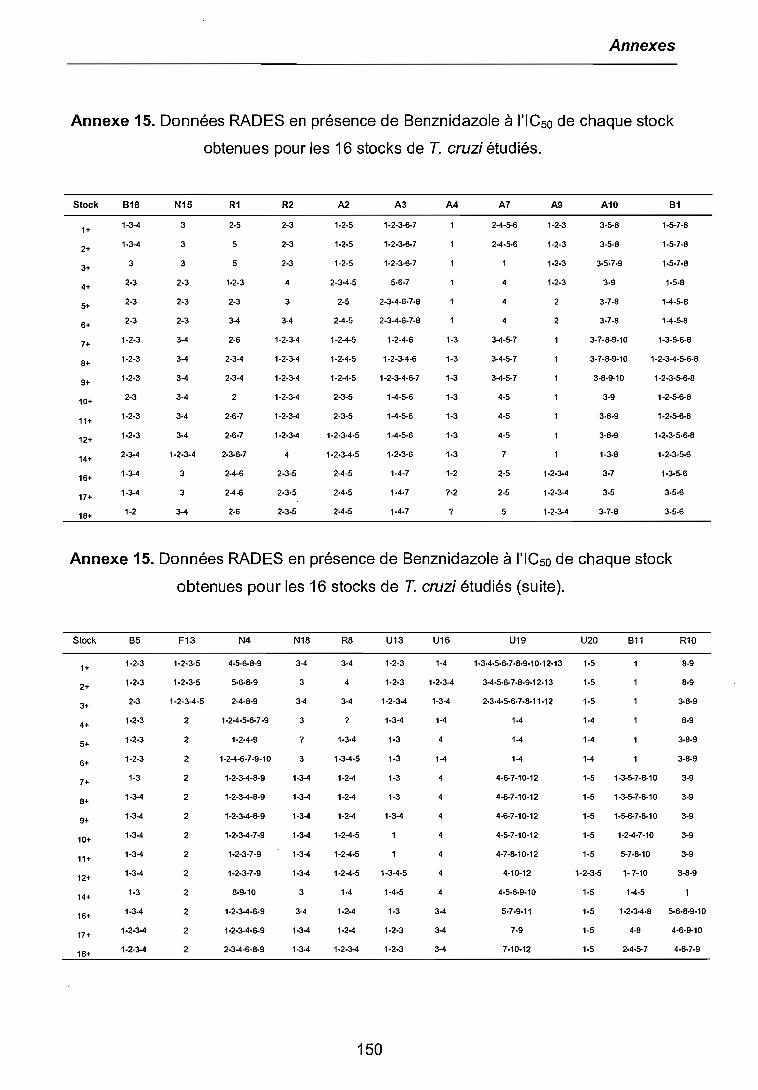
Annexes
Annexe 15. Données RADES en présence de Benznidazole à l'ICso de chaque stock
obtenues pour les 16 stocks de T. cruzi étudiés.
Stock
1+
2+
3+
4+
5+
6+
7+
6+
9+
10+
11+
12+
14+
16+
17+
18+
818
1-3-4
1-3-4
3
2-3
2-3
2·3
1-2-3
1-2-3
1-2-3
2·3
1·2·3
1·2-3
2-3-4
1-3-4
1-3-4
1-2
N15
3
3
3
2-3
2·3
2·3
3·4
3-4
3-4
3-4
3-4
3-4
1-2-3-4
3
3
3-4
R1
2-5
5
5
1'2-3
2-3
3-4
2-6
2-3·4
2-3-4
2
2-6-7
2-6-7
2-3-6-7
2-4-6
2-4-6
2·6
R2
2-3
2-3
2-3
3
3-4
1-2-3-4
1-2-3-4
1-2-3-4
1·2-3-4
1-2-3-4
1·2-3-4
2·3-5
2-3·5
2·3-5
A2
1-2·5
1-2-5
1-2-5
2-3-4-5
2-5
2-4-5
1-2-4-5
1·2·4-5
1-2-4-5
2-3-5
2-3-5
1·2-3-4-5
1·2-3-4·5
2-4-5
2-4-5
2-4-5
A3
1·2·3·6-7
1-2·3-6-7
1-2-3-6-7
5-6-7
2-3-4-6-7-8
2-3-4-6-7-8
1·2·4·6
1-2-3-4-6
1-2-3-4-6-7
1-4-5-6
1-4·5-6
1-4-5-6
1·2-3-6
1-4-7
1-4-7
1-4-7
A4
1-3
1·3
1·3
1-3
1-3
1-3
1·3
1-2
?-2
?
A7
2-4·5-6
2-4-5-6
3-4-5-7
3-4-5-7
3-4-5-7
4-5
4-5
4-5
7
2-5
2-5
5
A9
1-2-3
1-2-3
1-2-3
1-2-3
2
2
1-2-3-4
1·2-3-4
1·2-3-4
A10
3-5-8
3-5-6
3-5-7-9
3·9
3-7-8
3-7-8
3-7-8·9-10
3-7-8-9-10
3-8-9-10
3-9
3-8·9
3-8·9
1·3·8
3·7
3-5
3-7-8
81
1-5·7-8
1·5-7·6
1-5·7·6
1·5·8
1-4-5-6
1-4-5-8
1-3-5-6-8
1-2-3-4-5-6-8
1-2-3-5-6-8
1-2·5-6-8
1·2·5-6·8
1-2-3-5·6-8
1-2-3-5-6
1-3-5-6
3-5-6
3-5-6
Annexe 15. Données RADES en présence de Benznidazole à l'ICsode chaque stock
obtenues pour les 16 stocks de T. cruzi étudiés (suite).
Stock
1+
2+
3+
4+
5+
6+
7+
8+
9+
10+
11+
12+
14+
16+
17+
18+
85
1-2-3
1·2-3
2·3
1-2·3
1·2-3
1'2-3
1-3
1-3-4
1-3-4
1·3-4
1-3-4
1-3-4
1-3
1·3-4
1-2-3-4
1·2-3-4
F13
1-2-3-5
1-2-3-5
1-2-3-4-5
2
2
2
2
2
2
2
2
2
2
2
2
2
N4
4-5-6-8-9
5-6-8-9
2-4-8-9
1-2-4-5-6-7-9
1-2-4-9
1-2-4·6-7·9-10
1·2·3-4·8-9
1-2-3-4-8-9
1-2-3-4-8-9
1-2-3-4·7-9
1·2-3-7-9
1-2-3-7-9
8-9-10
1-2-3-4-6-9
1-2-3-4-6-9
2-3-4·6-8-9
N18
3-4
3
3-4
3
?
3
1-3-4
1-3-4
1-3-4
1·3-4
1-3-4
1-3-4
3
3-4
1-3-4
1-3-4
R8
3-4
4
3-4
?
1-3-4
1-3-4-5
1·2-4
1-2-4
1-2-4
1·2-4-5
1-2-4·5
1-2-4-5
1-4
1-2-4
1-2-4
1·2-3-4
U13
1-2-3
1·2-3
1-2-3-4
1-3-4
1-3
1-3
1-3
1-3
1-3-4
1-3-4-5
1-4-5
1-3
1-2-3
1·2·3
U16
1·4
1·2-3-4
1-3-4
1-4
3-4
3-4
3-4
U19
1·3-4·5·6-7·8-9-10·12·13
3-4·5-6-7-8·9·12-13
2·3-4·5·6-7-8-11-12
1-4
1-4
1-4
4·6-7·10-12
4-6-7-10-12
4-6-7-10-12
4-5-7-10-12
4-7-8-10·12
4-10-12
4-5-6-9-10
5-7-9-11
7-9
7-10-12
U20
1-5
1·5
1-5
1·4
1-4
1-4
1-5
1-5
1-5
1-5
1·5
1-2-3-5
1-5
1-5
1-5
1-5
811
1-3-5-7·8-10
1-3-5-7-8-10
1·5-6-7·8-10
1-2-4-7-10
5-7-8-10
1- 7-10
1-4-5
1-2-3-4-8
4-8
2-4·5-7
Rl0
8-9
8-9
3-8-9
8·9
3-8-9
3-8-9
3-9
3-9
3-9
3-9
3-9
3-8-9
5-6-8-9-10
4-6·9-10
4-6·7·9
150

Annexes
Annexe 16. Matrice des distances de Jaccard estimée sur la base des données RADES en
absence de Benznidazole (sur 22 amorces) pour les 16 stocks de T. cruzi étudiés.
Stock 2 3 4 5 6 7 8 9 10 11 12 14 16 17 18
1
2 0,12
3 0,20 0,17
4 D,52 D,58 0,60
5 0,42 D,52 0,49 0,33
6 0,47 D,54 D,51 0,27 0,10
7 D,56 D,58 D,59 D,57 D,53 D,54
8 D,54 D,56 D,57 D,54 D,51 D,52 0,09
9 D,56 D,57 D,59 D,56 D,50 D,52 0,14 0,09
10 0,60 0,62 0,61 D,53 D,53 D,54 D,3D 0,28 0,26
11 0,61 0,63 0,63 D,59 0,60 D,58 0,27 0,26 0,24 0,20
12 D,58 D,58 0,65 D,51 D,59 D,58 0,32 0,32 0,37 D,3D 0,32
14 0,62 0,63 0,67 D,56 0,61 0,61 0,44 0,42 0,43 0,49 0,48 D,53
16 D,50 0,49 D,53 0,65 D,57 0,60 0,45 0,47 0,46 D,54 D,54 D,52 0,62
17 D,51 D,50 D,54 0,65 D,57 0,61 0,47 0,49 0,47 D,55 D,56 D,54 0,62 0,08
18 0,60 0,62 0,63 0,61 D,59 0,60 0,45 0,43 0,49 D,53 D,53 0,43 0,60 0,35 0,36
Annexe 17. Matrice des distances de Jaccard estimée sur la base des données RADES en
présence de Benznidazole à l'IC5D (sur 22 amorces) pour les 16 stocks de T. cruzi étudiés.
Stock 1+ 2+ 3+ 4+ 5+ 6+ 7+ 8+ 9+ 10+ 11+ 12+ 14+ 16+ 17+ 18+
1+
2+ 0,11
3+ 0,24 0,26
4+ 0,47 D,51 D,51
5+ D,50 D,55 D,52 0,34
6+ 0,49 D,51 D,52 0,25 0,17
7+ D,53 D,57 D,56 D,59 D,52 D,54
8+ D,54 D,58 D,57 0,60 D,50 D,50 0,07
9+ D,53 D,57 D,55 D,57 D,53 D,53 0,11 0,05
10+ D,58 0,63 0,60 D,53 D,54 D,56 0,25 0,25 0,25
11+ D,58 0,62 0,63 D,58 D,57 D,58 0,22 0,22 0,22 0,12
12+ D,58 0,62 0,63 D,54 D,57 D,56 0,28 0,29 0,27 0,25 0,19
14+ 0,75 0,79 0,72 0,68 0,73 0,72 0,76 0,73 0,72 0,79 0,79 0,75
16+ 0,69 0,72 0,65 0,74 0,75 0,74 0,76 0,75 0,74 0,76 0,75 0,75 0,60
17+ 0,73 0,73 0,70 0,73 0,72 0,72 0,76 0,76 0,74 0,76 0,76 0,75 0,63 0,21
18+ 0,75 0,75 0,71 0,76 0,75 0,73 0,80 0,78 0,77 0,81 0,81 0,78 0,62 0,35 0,27
151

Annexes
Annexe 18. Séquences nucléotidiques obtenues.
Le code des séquences correspond à celui du tableau 9.
>Al (623 nt)GAAACGGGTGGGAGCACGAATACTCAGCGGATACGTCATGGGCGTTCAAACCGTGGTTATAGTGTTAGGTCGCACTCAGCGGAGCGTGGAGCAGCCGCAGGGGCGCAGGGGAAGACGTTGGGAACTTCGGGACGGTCAAGGCCATTCTCCCGTGCCGTGATGGATGCATATGCCTCACGTATACAAAGATGGTTCCGTAGGCTGGTTTTGGGAGGCGTCATGAGGTGCCGTGGGCTTCTGGTCATGCGCATTAGGGAAATTCTTGAATTGCAAAAACGTCTTTGGGAACTTCGTGTCCGTATGGCAAAACGTGTCATGTCCGAGGCGTTTAATACTTATGCCATAATTATTCATGGCGATTCCGAGATGAGAGGTATAAGGAAGTATAAACCTGAAGTAACATTGGTGAATGCATTTATGCGTGCAAAGAAGGACTGCGTTTTGAGGAGCGAAAAGCTGCATATTATTTACCAACGCGCGGTAAGGGTTATTGAACGAGCATGGACAAAGTGTGTCTTATTTCGTTCTATTCAACCAGCCTCTCTGGAGAACAAAATGCAATGGCTTCTCATCCAGCAAGAGGAGATTGAAAGGCGTGATTACATTCGTCAACGATTGTTATT
>A2 (650 nt)GAAACGGGTGTTACCAGCAACAATAAACAAGTTATCATCACCAGAGACACCAATTCCACGCATCAACAAAGCGTCGGTGGCAAATGCAAAGGCACACGTGCTTTTTTGAATTGCTGCCTGCTCTCTGTCACCTTTTTTCCTGCCTCTCCA.GCATTCCTCGTAGTCGCAGTACCTCGCTGAATAATCCATCCCAAAGCGGCAACCTCAGCATCGTACAAATCTTGCAATTTCAATCGGCGATGATACTCGCGCACTAAAAGACGCTCAATTTCATACTTGATGTCAAACTCACTTCCAAAGATCTTTATCGGAGCCCAATTCGTTTGTGTCTGGTCTGTAGGAGATGGGATTGGCGTGTGCAATTTGCCGGATGTCCGTGTCACTGAGGCCGAGCCTGTTCCCAGGAGAAAATGCAAAAATGCACGCCCTTTACTCCTACGAGAGACCATCAGCGATGCTAGCGGAGAGGAATCTGATGTAGTATTGTTACACGAGAAAGATTCAAAGACATCATCAGTGAGACCCATAAAGACGTAGAAATCCGCCCACGCCTTCCCCATGGAGGCCACGCTCACACTGAAGTCCATCGGCGAAAGAGTCCTTTCGCGGCTTTTCTTGTCGGCGCAGGGTGAGAAAGCACGCATTGACTG
>Cl (718 nt)GAAACGGGTGGGAGCACGAATACTCAGCGGAAACGTAATGGGCGTTCAAACCGTGGTTATAGTGTTAGGTCGCACTCAGCGGAGCGTGGAGCAGCCGCAGGGGCGCAGGGGAAGACGTTGGGAACTTCGGGACGGTCAAGGCCATTCTCCCGTGCCGTGATGGATGCATATGCCTCACGTATACAAAGATGGTTCCGTAGGCTGGTTTTGGGAGGCGTCATGAGGTGCCGTGGGCTCCTGGTCATGCGCATTAGGGAAATTCTTGAATTGCAAAAACGTCTTTGGGAACTTCGTGTCCGTATGGCAAAACGTGTCATGTCCGAGGCGTTTAATACTTATGCCATAATTATTCATGGCGATTCCGAGATGAGAGGTATAAGGAAGTATAAACCTGAAGTAACATTGGTGAATGCATTTATGCGTGCAAAGAAGGACTGCGTTTTGAGGAGCGAAAAGCTGCATATTATTTACCAACGCGCGGTAAGGGTTATTGAACGAGCATGGACAAAGTGTGTCTTATTTCGTTCTATTCAACCAGCCTCTCTGGAGAACAAAATGCAATGGCTTCTCATCCAGCAAGAGGAGATTGAAAGGCGTGATTACATTCGTCAACGATTGTTATTTCTCGTTGAGTGCCACCAGCAGTGTTACAATCACCCTCTTTAATGAAGGCTGTAATGCTGTGCGGAACACTGAATGCTGGTCTGGTATCATAACG
>C2 (602 nt)GAAACGGGTGTTACCAGCAACAATAAACAAGTTATCATCACCAGAGACACCAATTCCACGCATCAACAAAGCGTCGGTGGCAAATGCAAAGGCACACGTGCTTTTTTGAATTGCTGCCTGCTCTCTGTCACCTTTTTCCTGCCTCTCCAGCATTCCTCGTAGACGCAGTACCTCGCTGAATAATCCATCCCAAAGCGGCAACCTCAGCATCGTACAAATCTTGCAATTTCAATCGGCGATGATACTCGCGCACTAAAAGACGCTCAATTTCATACTTGATGTCAAACTCACTTCCAAAGATCTTTATCGGAGCCCAATTCGTTTGTGTCTGGTCTGTAGGAGATGGGATTGGCGTGTGCAATTTGCCGGATGTCCGTGTCACTGAGGCCGAGCCTGTTCCCAGGAGAAAATGCAAAAATGCACGCCCTTTACTCCTTCGAGAGACCATCAGCGATGCTAGCGGAGAGGAATCTGATGTAGTATTGTTTACACGAGAAAGATTCAAAGACATCATCAGTGAGATCCCATAAAGACGTAGAAATCCGCCCAACGCCTTCCCCATGGAAGGCCACGCTTCACACTGAAGTCCAATCGGCGAAAAA
>D2 (692 nt)TGCCGAGCTGCGTATGAAGGGGATGTACGTGAATGACGTACGCAAGAGGAAAGAGGAACTGGAGCAGCGATTGAGCGCCATGATGGATGCGTTTCAAAAGATCAAGACGGAACGTGGTCAGAAGGCTGCACAAATTCAGGCCATCACGCAGAAAATGACCGAAATGTCTGAGAAAAAGAAAATTGTTGAAAATGAGCTCGTTGTTCTTTGCCGCGAGTCTGCTTTAAAAGAAACGGAGTTGGTAAAGAAAAAACGTCAGGTGCAAGAACTTCGTCAGGTGTGCAAGAACCTCCGTATGGAAAAAAATCGACAACGTAAGTCTTTGGAGAAGGCGTTTGAGGATGAACGAAACGTTAAGATTAATGTGCGACGAACGAACGCACAGATTGCGGATCTTGAACAGGAGATGAATGGTGTGAAGCTGAGATACCATGACCTAATTGAGAAACGCAACTATGCCGGGGTCCAAGTACTTGGACCGCCAATGATGAACTCTGCGTGCTGTATGAACGGGTAAATGCGCAAGAAAAAGGCCATAAAATAATGGGTGTTCTCATGAACAAATGCCCGTGATGAGGAAAATACGGTACACTACGGAATTAAGCTTTGGCCGAATCTTTGCCCCTGAGGGTGGGACGTAGTTTCGACCGGTCTTATTTCCCAAAAGGTAAAAGGAAGTTTGGGAGGGAAAC
152

Annexes
>E3 (573 nt)GGCTGGTTCCCGTAAGGAAGTAAGAGAGTTTGCAGAGTGAGATTTCGCCCGACTAATCGCATCGCTCTCAAGTCGCAACCACGCGTGTCGCAGTGGCTGATGATGGGCAGTGATAAGTTCTGCCAGACGCCGCGGCAATTCCAGCAGCACCTCAGACAACATCTGCAACACCGCCGCCACCACAGGCAAGTTGGCCGTTGCATGCGCATCATCAACATGACTAGTGCCCTCGAGCCTCCGCATTTGGCACTTCAACGCCTCTGCACTGAGGCGCAATTGACGTGAGAGAGCGATCGGCAAGGCTCCTCCCGCCTGCACAAAACCAAGACCACACTCGGGACTCCCATCGCCACTCGACGGTGATGCGTCCATAGCTGCCCGTACGCTCGCCAAACACTTTAGCAGTAAATCTGTGTACATGCGAAGCTGATAAGCAGTTGGGGCCACGAGCAACATTGAAGCCCACACCCTGGAAGCCCTCCGGAAGACCCACGCCGCACCTTCTTCCCAAAAAGTGTTGCCCTTTGGATCCCAACTTCCAGCCAAGCCACCCAACTTCGGCCAACGTTTTAA
>Fl (543 nt)CTGCGCTGGAGAAGGGAAAGCATGACACTACGGTCCTTTAGAGCCCTTTCCACACGGTCCGCTGGTAACGTCGGCAGATGACCCGTGATAAACGCAAGCACATCAGAATCACTGCCAAAGTGCTCAACGACGCTCTGCATGACTTCTAGTGAAGCGTTTGGCAGGTAGCGGCGACGCATAGACTCCACCCTCGTCGCCTCGTTCGAGAAGAATAAGGGAGAACTTGCTGACCCGTTTTCATGCACCAGAGTATGGTCGGGACGCATAAAAATCATCTCATTTTGAATGGTCCAAATGTTACTCACGTTGTCCAGAAGAAGTGAAATGGAGCGGTCAAAAAAGCACGTCAGGAAAGGAGCCTTCTCACGTGGAATTAAATCGGCACCGTTACTGTCTTTTGGGGACCAATGCCAAAAGAGTCGGCCATGTATAGTCACTATACATGTCGGAACCTTCATATTTTGCGAAAGAAACACGACGTTGGCGTTCCCATTGAGTTCATGTGAAACACTGATAAATTTCCTTTCACCGCCGCCAACTTCC
>Gl (562 nt)CTGCGCTGGAGAAAAGGAGAATTTTTTCGGTGAAAAAATTTGAAAATCCTCCATAAAACTGCCAAGACGATCAATTTCCGCCTGAAGTGCTCTCAACTGCCGTTGCTGTTCCTCCACGGTATGAGAGAGCGTTTCAGTCATCTGCTTGAAATCCTCCTCCGTGTCGGATGTTGTTTTCTGAAGTAATTTCGGCGTGCTCTCGGTGGGCGGTTCATCGTCAGCAACAAGTGGATCCGTGATGGAAGGAATGGGCTCATCGCAGAGCGGGACATCTTGTAAGCCGTTGCTCTGCTTCTCGGAAAGGCAATTTTCAACATCCGGCAGCGCCTCAGCCTCCACTTGATGAACATTTGTTTTCGGCTCCTCATCGTTGTTTTTGTCACACTGCGTTGTTTCCTGCGTCCCCTTGTCGCCATACTCAAAAAGTTGTGCACGGTGTCTCTTCTGTTGGTCCCGATCGTTCATCCGTGAACATCATCTTGGGGATCCGCATTAAGTTCTTTTAGAACGCGGTCTCCCAAGAAACTCCCTCCCTCTGTGGCCGGTAAGTGGGCCCGCCTAA
>11 (450
nt)CACAGCTGCCGGTGCATGAGGCGCCAAGCACGCATTCATAAAGACCCTCCAGGCGATTTCTATTACTGCTGTGCACCTTCTCCTTGAGGTACCAACGGCTCACTGCGTATGGGTTGTTTCCAAAGCCTGGACGCTGTGTCAGAACGTGAAAAGCCGTCTTGCTGTTGTCTTTGGCGGTAAATACAGTCTCAGGCTTTGGTGCAGCGTTCTTTAATATACGACGATACATGAAGAGAGGGCTGCTGTACCTTGCAACCTCTCCTGTAAGCGGGGCCGAAGGCATATATATATATATTTCTCAAAGGGATTCTCTCTCTCTTTTTCTTTTTTCCCTACAACAGCTACCGGGCTATATATAAATATTTATATGTGTATTGCACGATTATTCGACTCCTCTCACTCTCCTTGGACTTCTCTTGGCAGCTGTGCAATCCACTAGTGGAATTCGCG
>Jl (500 nt)CACAGCTGCCCAGAGAAGTACAAGGAGGGTGAGAGGAGTCGAATAATCGTGCAATAAATATATAAATAAGTATTCATATAATAGCCTGGTAGCTGTTGTAGGAAAAAAAAGGAAAAAACAAGAGAGAGAGAGAGAGAGAATCCCTTTGAGAAATATATATATATATATATATGCCTTCGGCCCCGCTTACAGGAGAGGTTGCAAGGTACAGCAGCCCTCTCTTCATGTATCGTCGTATAATAAAGAATGCTGCACCAAAGCCTGAGACTGTATTTACCGCCAAAGACAACAGCAAGACGGCTTTTCACGTCCTGACACAGCGTCCGGGCTTTGGAAACAACCCATACGCGGTGAGCCGTTGGTACCTCAAGGAGAAGGTGCACAGTAGTAATAGAAATCGTCTGGAGGGTCTTTATGAATGCGTGCTTTGCGCCTCATGCACCGGCAGCTGTGAATCGAATTCCCGCGGCCGCCATGGCGGCCGGGAGCATGCGACGTCG
>01 (618 nt)GAAACGGGTGTTACCAGCAACAATAAACAAGTTATCATCCCCAGAGACACCAATTCCACGCATCAACAAAGCGTCGGTGGCAAATGCAAAGGCACACGTGCTTTTTTGAATTGCTGCCTGCTCTCTGTCACCTTTTTTCCTGCCTCTCCAGCATTCCTCGTAGTCGCAGTACCTCGCTGAATAATCCATCCCAAAGCGGCAACCTCAGCATCGTACAAATCTTGCAATTTCAATCGGCGATGATACTCGCGCACTAAAAGACGCTCAATTTCATACTTGATGTCAAACTCACTTCCAAAGATCTTTATCGGAGCCCAATTCGTTTGTGTCTGGTCTGTAGGAGATGGGATTGGCGTGTGCAATTTGCCGGATGTCCGTGTCACTGAGGCCGAGCCTGTTCCCAGGAGAAAATGCAAAAATGCACGCCCTTTACTCCTTCGAGAGACCATCAGCGATGCTAGCGGAGAGGAATCTGATGTAGTATTGTTACACGAGAAAGATTCAAAGACATCATCAGTGAGATCCATAAAGACGTAGAAATCCGCCCACGCCTTCCCCAATGGGAGGCCACGCTCACACTGAAAGTCCATCGGCGAAAAGAGTCCTTTCGCGGCTTTT
153

Annexes
>02 (658 nt)GAAACGGGTGGGAGCACGAATACTCAGCGGAAACGTAATGGGCGTTCAAACCGTGGTTATAGTGTTAGGTCGCACTCAGCGGAGCGTGGAGCAGCCGCAGGGGCGCAGGGGAAGACGTTGGGAACTTCGGGACGGTCAAGGCCATTCTCCCGTGCCGTGATGGATGCATATGCCTCACGTATACAAAGATGGTTCCGTAGGCTGGTTTTGGGAGGCGTCATGAGGTGCCGTGGGCTTCTGGTCATGCGCATTAGGGAAATTCTTGAATTGCAAAAACGTCTTTGGGAACTTCGTGTCCGTATGGCAAAACGTGTCATGTCCGAGGCGTTTAATACTTATGCCATAATTATTCATGGCGATTCCGAGATGAGAGGTATAAGGAAGTATAAACCTGAAGTAACATTGGTGAATGCATTTATGCGTGCAAAGAAGGACTGCGTTTTGAGGAGCGAAAAGCTGCATATTATTTACCAACGCGCGGTAAGGGTTATTGAACGAGCATGGACAAAGTGTGTCTTATTTCGTTCTATTCAACCAGCCTCTCTGGAGAACAAAATGCAATGGCTTCTCATCCAGCAAGAGGAGATTGAAAGGCCGTGATTACATTCGTCAACGAATGGTTATTTCTCGTTGAGTGCCACCAGCAGTGTTTACAATC
154
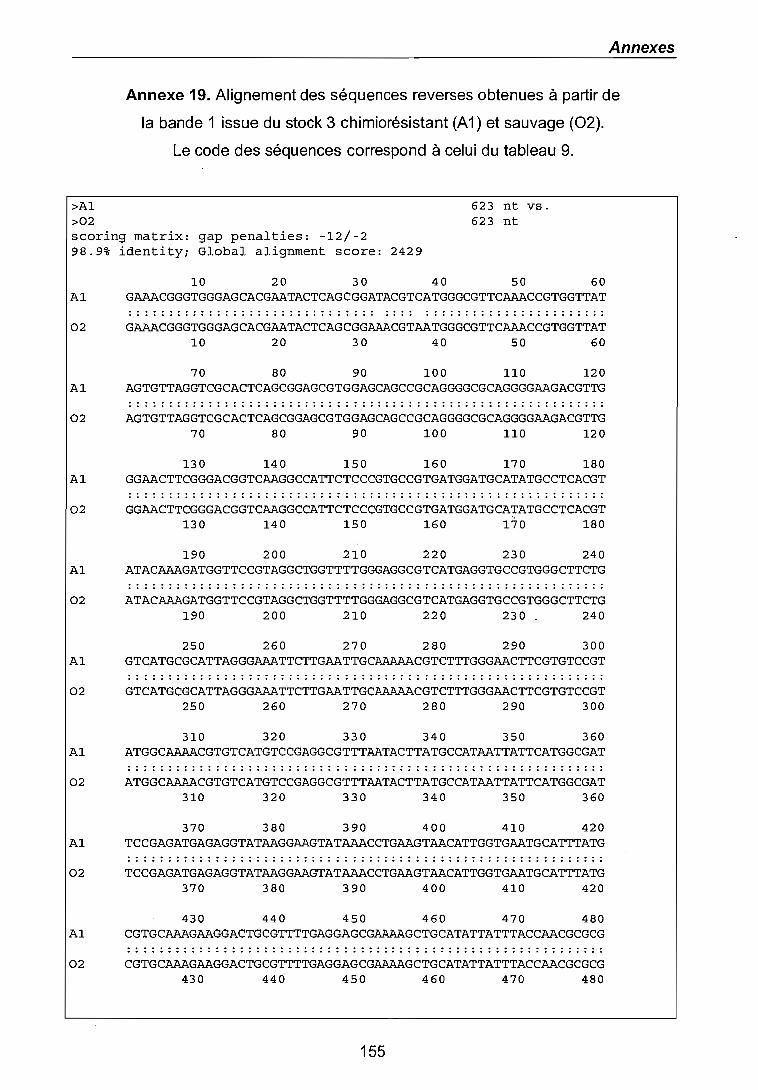
Annexes
Annexe 19. Alignement des séquences reverses obtenues à partir de
la bande 1 issue du stock 3 chimiorésistant (A1) et sauvage (02).
Le code des séquences correspond à celui du tableau 9.
>A1>02scoring matrix: gap penalties: -12/-298.9% identitYi Global alignment score: 2429
623 nt vs.623 nt
10 20 30 40 50 60Al GAAACGGGTGGGAGCACGAATACTCAGCGGATACGTCATGGGCGTTCAAACCGTGGTTAT
.. .. .. .. .. .. .. .. . .. . . . . . . . . . . .. . . .. .. . .. . . . ... . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..02 GAAACGGGTGGGAGCACGAATACTCAGCGGAAACGTAATGGGCGTTCAAACCGTGGTTAT
10 20 30 40 50 60
70 80 90 100 110 120Al AGTGTTAGGTCGCACTCAGCGGAGCGTGGAGCAGCCGCAGGGGCGCAGGGGAAGACGTTG
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. . .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. ..02 AGTGTTAGGTCGCACTCAGCGGAGCGTGGAGCAGCCGCAGGGGCGCAGGGGAAGACGTTG
70 80 90 100 110 120
130 140 150 160 170 180Al GGAACTTCGGGACGGTCAAGGCCATTCTCCCGTGCCGTGATGGATGCATATGCCTCACGT
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
02 GGAACTTCGGGACGGTCAAGGCCATTCTCCCGTGCCGTGATGGATGCATATGCCTCACGT130 140 150 160 170 180
190 200 210 220 230 240Al ATACAAAGATGGTTCCGTAGGCTGGTTTTGGGAGGCGTCATGAGGTGCCGTGGGCTTCTG
.................................................................................................... " .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
02 ATACAAAGATGGTTCCGTAGGCTGGTTTTGGGAGGCGTCATGAGGTGCCGTGGGCTTCTG190 200 210 220 230 240
250 260 270 280 290 300Al GTCATGCGCATTAGGGAAATTCTTGAATTGCAAAAACGTCTTTGGGAACTTCGTGTCCGT
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. . .. .... .. .. .. .. .. .. ........ .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .02 GTCATGCGCATTAGGGAAATTCTTGAATTGCAAAAACGTCTTTGGGAACTTCGTGTCCGT
250 260 270 280 290 300
310 320 330 340 350 360Al ATGGCAAAACGTGTCATGTCCGAGGCGTTTAATACTTATGCCATAATTATTCATGGCGAT
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ........ .. .. .. .. .. .. .. ...... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..02 ATGGCAAAACGTGTCATGTCCGAGGCGTTTAATACTTATGCCATAATTATTCATGGCGAT
310 320 330 340 350 360
370 380 390 400 410 420Al TCCGAGATGAGAGGTATAAGGAAGTATAAACCTGAAGTAACATTGGTGAATGCATTTATG
...................................................................................................................... . .. .. .. .. .. .. .. .. .. .. .. . . .. .. .. .. .. .. .. .. . .. .. .. .. . .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
02 TCCGAGATGAGAGGTATAAGGAAGTATAAACCTGAAGTAACATTGGTGAATGCATTTATG370 380 390 400 410 420
430 440 450 460 470 480Al CGTGCAAAGAAGGACTGCGTTTTGAGGAGCGAAAAGCTGCATATTATTTACCAACGCGCG
........................................................................................................................ .. . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ........ .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. ..02 CGTGCAAAGAAGGACTGCGTTTTGAGGAGCGAAAAGCTGCATATTATTTACCAACGCGCG
430 440 450 460 470 480
155

490 500 510 520 530 540Al GTAAGGGTTATTGAACGAGCATGGACAAAGTGTGTCTTATTTCGTTCTATTCAACCAGCC
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. " ..02 GTAAGGGTTATTGAACGAGCATGGACAAAGTGTGTCTTATTTCGTTCTATTCAACCAGCC
490 500 510 520 530 540
550 560 570 580 590Al TCTCTGGAGAACAAAATGCAATGGCTTCTCATCCAGCAAGAGGAGATTGAAAGGC-GTGA
........................................................................................................................ .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... ........02 TCTCTGGAGAACAAAATGCAATGGCTTCTCATCCAGCAAGAGGAGATTGAAAGGCCGTGA
550 560 570 580 590 600
600 610 620Al TTACATTCGTCAACGATTGTTATT
.. .. .. .. .. .. .. .... .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..02 TTACATTCGTCAACGAATGGT-TA
610 620
156
Annexes
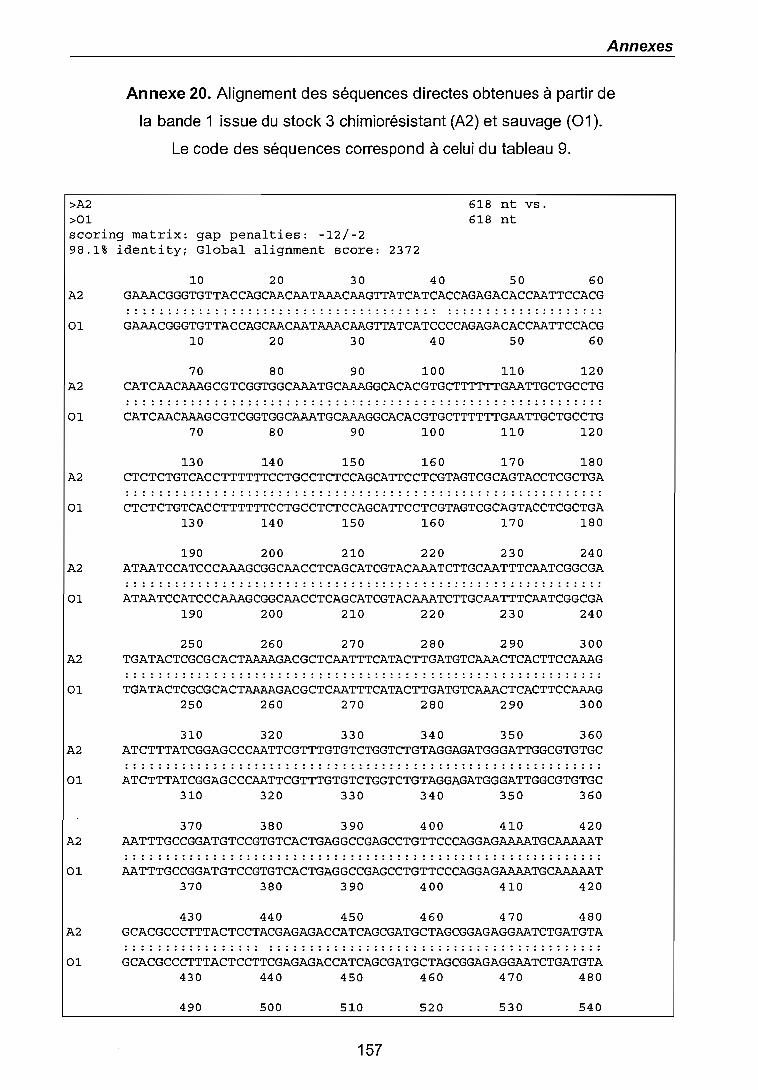
Annexes
Annexe 20. Alignement des séquences directes obtenues à partir de
la bande 1 issue du stock 3 chimiorésistant (A2) et sauvage (01).
Le code des séquences correspond à celui du tableau 9.
>A2>01scoring matrix: gap penalties: -12/-298.1% identitYi Global alignment score: 2372
618 nt vs.618 nt
10 20 30 40 50 60A2 GAAACGGGTGTTACCAGCAACAATAAACAAGTTATCATCACCAGAGACACCAATTCCACG
........................................................................................................................ .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..Dl GAAACGGGTGTTACCAGCAACAATAAACAAGTTATCATCCCCAGAGACACCAATTCCACG
10 20 30 40 50 60
70 80 90 100 110 120A2 CATCAACAAAGCGTCGGTGGCAAATGCAAAGGCACACGTGCTTTTTTGAATTGCTGCCTG
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .... .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
Dl CATCAACAAAGCGTCGGTGGCAAATGCAAAGGCACACGTGCTTTTTTGAATTGCTGCCTG70 80 90 100 110 120
130 140 150 160 170 180A2 CTCTCTGTCACCTTTTTTCCTGCCTCTCCAGCATTCCTCGTAGTCGCAGTACCTCGCTGA
.......................................................................................................................... .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
Dl CTCTCTGTCACCTTTTTTCCTGCCTCTCCAGCATTCCTCGTAGTCGCAGTACCTCGCTGA130 140 150 160 170 180
190 200 210 220 230 240A2 ATAATCCATCCCAAAGCGGCAACCTCAGCATCGTACAAATCTTGCAATTTCAATCGGCGA
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .... .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. ...... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
Dl ATAATCCATCCCAAAGCGGCAACCTCAGCATCGTACAAATCTTGCAATTTCAATCGGCGA190 200 210 220 230 240
250 260 270 280 290 300A2 TGATACTCGCGCACTAAAAGACGCTCAATTTCATACTTGATGTCAAACTCACTTCCAAAG
................................................................................................................... .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
Dl TGATACTCGCGCACTAAAAGACGCTCAATTTCATACTTGATGTCAAACTCACTTCCAAAG250 260 270 280 290 300
310 320 330 340 350 360A2 ATCTTTATCGGAGCCCAATTCGTTTGTGTCTGGTCTGTAGGAGATGGGATTGGCGTGTGC
.......................................................................................................................... .. .. .. . .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .... .... .. .. .. .. .. .. .. .. ...... .. .. .. .. .. .. .. ...... .. .. .. .. .. ..
Dl ATCTTTATCGGAGCCCAATTCGTTTGTGTCTGGTCTGTAGGAGATGGGATTGGCGTGTGC310 320 330 340 350 360
370 380 390 400 410 420A2 AATTTGCCGGATGTCCGTGTCACTGAGGCCGAGCCTGTTCCCAGGAGAAAATGCAAAAAT
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..Dl AATTTGCCGGATGTCCGTGTCACTGAGGCCGAGCCTGTTCCCAGGAGAAAATGCAAAAAT
370 380 390 400 410 420
430 440 450 460 470 480A2 GCACGCCCTTTACTCCTACGAGAGACCATCAGCGATGCTAGCGGAGAGGAATCTGATGTA
....................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..Dl GCACGCCCTTTACTCCTTCGAGAGACCATCAGCGATGCTAGCGGAGAGGAATCTGATGTA
430 440 450 460 470 480
490 500 510
157
520 530 540

A2 GTATTGTTACACGAGAAAGATTCAAAGACATCATCAGTGAGACCCATAAAGACGTAGAAA........................................................................................................................ .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
01 GTATTGTTACACGAGAAAGATTCAAAGACATCATCAGTGAGATCCATAAAGACGTAGAAA490 500 510 520 530 540
550 560 570 580 590A2 TCCGCCCACGCCTTCCCCAT--GGAGGCCACGCTCACACTGAA-GTCCATCGGCGAAA-G
Annexes
........................................ .. .. .. .. .. .. .. .. .. .. .. .. .. .. ...... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..01 TCCGCCCACGCCTTCCCCAATGGGAGGCCACGCTCACACTGAAAGTCCATCGGCGAAAAG
550 560 570 580 590 600
600 610A2 AGTCCTTTCGCGGCTTTTCTTG
...................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... ..
01 AGTCCTTTCGCGGCTTTT----610
158
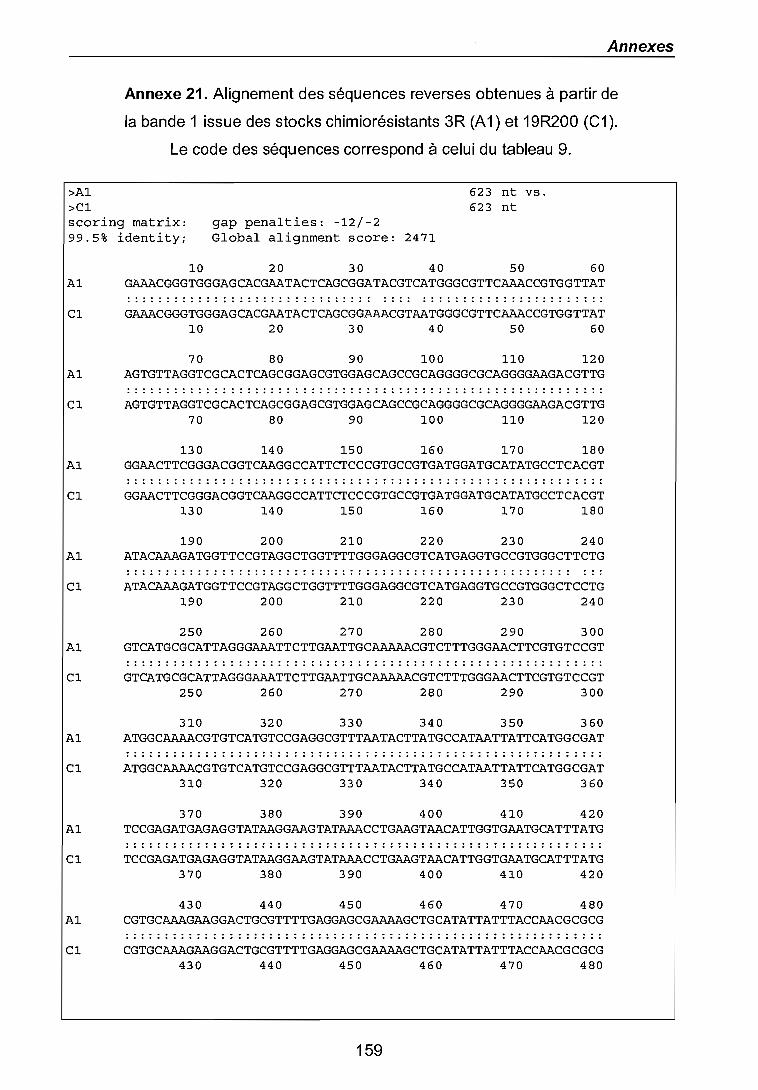
Annexes
Annexe 21. Alignement des séquences reverses obtenues à partir de
la bande 1 issue des stocks chimiorésistants 3R (A1) et 19R200 (C1).
Le code des séquences correspond à celui du tableau 9.
>A1>C1scoring matrix:99.5% identitYi
gap penalties: -12/-2Global alignment score: 2471
623 nt vs.623 nt
10 20 30 40 50 60Al GAAACGGGTGGGAGCACGAATACTCAGCGGATACGTCATGGGCGTTCAAACCGTGGTTAT
.. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..Cl GAAACGGGTGGGAGCACGAATACTCAGCGGAAACGTAATGGGCGTTCAAACCGTGGTTAT
10 20 30 40 50 60
70 80 90 100 110 120Al AGTGTTAGGTCGCACTCAGCGGAGCGTGGAGCAGCCGCAGGGGCGCAGGGGAAGACGTTG
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
Cl AGTGTTAGGTCGCACTCAGCGGAGCGTGGAGCAGCCGCAGGGGCGCAGGGGAAGACGTTG70 80 90 100 110 120
130 140 150 160 170 180Al GGAACTTCGGGACGGTCAAGGCCATTCTCCCGTGCCGTGATGGATGCATATGCCTCACGT
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
Cl GGAACTTCGGGACGGTCAAGGCCATTCTCCCGTGCCGTGATGGATGCATATGCCTCACGT130 140 150 160 170 180
190 200 210 220 230 240Al ATACAAAGATGGTTCCGTAGGCTGGTTTTGGGAGGCGTCATGAGGTGCCGTGGGCTTCTG
.. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ...... .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. ..
Cl ATACAAAGATGGTTCCGTAGGCTGGTTTTGGGAGGCGTCATGAGGTGCCGTGGGCTCCTG190 200 210 220 230 240
250 260 270 280 290 300Al GTCATGCGCATTAGGGAAATTCTTGAATTGCAAAAACGTCTTTGGGAACTTCGTGTCCGT
................................................................................................................ . .. . .. .. .. .. .. .. .. .. .. .. .. . .. . .. . . .. .. . .. .. .. .. .. .. .. .. . . . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. . ..Cl GTCATGCGCATTAGGGAAATTCTTGAATTGCAAAAACGTCTTTGGGAACTTCGTGTCCGT
250 260 270 280 290 300
310 320 330 340 350 360Al ATGGCAAAACGTGTCATGTCCGAGGCGTTTAATACTTATGCCATAATTATTCATGGCGAT
.................................................................................................................... .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. . .. . .. .. .. .. .. .. .. .. .Cl ATGGCAAAACGTGTCATGTCCGAGGCGTTTAATACTTATGCCATAATTATTCATGGCGAT
310 320 330 340 350 360
370 380 390 400 410 420Al TCCGAGATGAGAGGTATAAGGAAGTATAAACCTGAAGTAACATTGGTGAATGCATTTATG
........................................................................................................................ .. . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ......Cl TCCGAGATGAGAGGTATAAGGAAGTATAAACCTGAAGTAACATTGGTGAATGCATTTATG
370 380 390 400 410 420
430 440 450 460 470 480Al CGTGCAAAGAAGGACTGCGTTTTGAGGAGCGAAAAGCTGCATATTATTTACCAACGCGCG
....................................................................................................................... .. .. . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. . .. .. .. .. .. .. .. .. ..Cl CGTGCAAAGAAGGACTGCGTTTTGAGGAGCGAAAAGCTGCATATTATTTACCAACGCGCG
430 440 450 460 470 480
159

490 500 510 520 530 540Al GTAAGGGTTATTGAACGAGCATGGACAAAGTGTGTCTTATTTCGTTCTATTCAACCAGCC
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .. ........ .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. ..Cl GTAAGGGTTATTGAACGAGCATGGACAAAGTGTGTCTTATTTCGTTCTATTCAACCAGCC
490 500 510 520 530 540
550 560 570 580 590 600Al TCTCTGGAGAACAAAATGCAATGGCTTCTCATCCAGCAAGAGGAGATTGAAAGGCGTGAT
.......................................................................................................................... .. .... .. .. .. .. .. .. .. .. .. .. .. ...... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ...... .. .. .. .. .. ..
Cl TCTCTGGAGAACAAAATGCAATGGCTTCTCATCCAGCAAGAGGAGATTGAAAGGCGTGAT550 560 570 580 590 600
610 620Al TACATTCGTCAACGATTGTTATT
................................................ .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. ..
Cl TACATTCGTCAACGATTGTTATT610 620
160
Annexes

Annexes
Annexe 22. Alignement des séquences directes obtenues à partir de
la bande 1 issue des stocks chimiorésistants 3R (A2) et 19R200 (C2).
Le code des séquences correspond à celui du tableau 9.
>A2>C2scoring matrix:97.5% identity;
602 nt vs.602 nt
, gap penalties: -12/-2Global alignment score: 2259
10 20 30 40 50 60A2 GAAACGGGTGTTACCAGCAACAATAAACAAGTTATCATCACCAGAGACACCAATTCCACG
.......................................................................................................................... .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. .. .... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
C2 GAAACGGGTGTTACCAGCAACAATAAACAAGTTATCATCACCAGAGACACCAATTCCACG10 20 30 40 50 60
70 80 90 100 110 120A2 CATCAACAAAGCGTCGGTGGCAAATGCAAAGGCACACGTGCTTTTTTGAATTGCTGCCTG
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .... .. .. .. .. .. ..
C2 CATCAACAAAGCGTCGGTGGCAAATGCAAAGGCACACGTGCTTTTTTGAATTGCTGCCTG70 80 90 100 110 120
130 140 150 160 170 180A2 CTCTCTGTCACCTTTTTTCCTGCCTCTCCAGCATTCCTCGTAGTCGCAGTACCTCGCTGA
...................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..C2 CTCTCTGTCACCTTTTT-CCTGCCTCTCCAGCATTCCTCGTAGACGCAGTACCTCGCTGA
130 140 150 160 170
190 200 210 220 230 240A2 ATAATCCATCCCAAAGCGGCAACCTCAGCATCGTACAAATCTTGCAATTTCAATCGGCGA
.......................................................................................................................... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ....C2 ATAATCCATCCCAAAGCGGCAACCTCAGCATCGTACAAATCTTGCAATTTCAATCGGCGA
180 190 200 210 220 230
250 260 270 280 290 300A2 TGATACTCGCGCACTAAAAGACGCTCAATTTCATACTTGATGTCAAACTCACTTCCAAAG
.............................................................................. "4O" .... .. . . .. . .. . . . . . . .. . . . . .. . . . .. . . . . . . .. .. . .. .. . . . .. .. .. .. . .. .. .. . .. .. .. . . .. .. . .. . . .C2 TGATACTCGCGCACTAAAAGACGCTCAATTTCATACTTGATGTCAAACTCACTTCCAAAG
240 250 260 270 280 290
310 320 330 340 350 360A2 ATCTTTATCGGAGCCCAATTCGTTTGTGTCTGGTCTGTAGGAGATGGGATTGGCGTGTGC
• "4O 4O" "4O .404O .• "4O" 4O "4O" ..
C2 ATCTTTATCGGAGCCCAATTCGTTTGTGTCTGGTCTGTAGGAGATGGGATTGGCGTGTGC300 310 320 330 340 350
370 380 390 400 410 420A2 AATTTGCCGGATGTCCGTGTCACTGAGGCCGAGCCTGTTCCCAGGAGAAAATGCAAAAAT
· ... . . .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. . .. .. .. .. .. .. . .. .. .. .. .. .. .. .. .. .. . .. . .. .. .. .. .. .. .. .. ..
C2 AATTTGCCGGATGTCCGTGTCACTGAGGCCGAGCCTGTTCCCAGGAGAAAATGCAAAAAT360 370 380 390 400 410
430 440 450 460 470 480A2 GCACGCCCTTTACTCCTACGAGAGACCATCAGCGATGCTAGCGGAGAGGAATCTGATGTA
..........................................................................................................· .. . . . . . . . . .. .. . .. . .. ..C2 GCACGCCCTTTACTCCTTCGAGAGACCATCAGCGATGCTAGCGGAGAGGAATCTGATGTA
420 430 440 450 460 470
161

490 500 510 520 530A2 GTATTGTT-ACACGAGAAAGATTCAAAGACATCATCAGTGAGA-CCCATAAAGACGTAGA
. .. ~ .. .. .. .... .. .. .. .. .. ..C2 GTATTGTTTACACGAGAAAGATTCAAAGACATCATCAGTGAGATCCCATAAAGACGTAGA
480 490 500 510 520 530
540 550 560 570 580 590A2 AATCCGCCCA-CGCCTTCCCCATGGA-GGCCACGCT-CACACTGAAGTCCA-TCGGCGAA
.................................................................................................................. .. .. .. .. .. ..C2 AATCCGCCCAACGCCTTCCCCATGGAAGGCCACGCTTCACACTGAAGTCCAATCGGCGAA
540 550 560 570 580 590
600A2 AGAGTCCT
C2 AAA-----600
162
Annexes

Annexes
Annexe 23. Séquences des acides aminés déduits à l'aide du programme
"Six Frame Translation of Sequence" sur le site sel\ll Search Launcher.
Le code des séquences correspond à celui du tableau 9.
>C1, frame+1 (239 aa)ETGGSTNTQRKRNGRSNRGYSVRSHSAERGAAAGAQGKTLGTSGRSRPFSRAVMDAYASRIQRWFRRLVLGGVMRCRGLLVMRIREILELQKRLWELRVRMAKRVMSEAFNTYAIIIHGDSEMRGIRKYKPEVTLVNAFMRAKKDCVLRSEKLHIIYQRAVRVIERAWTKCVLFRSIQPASLENKMQWLLIQQEEIERRDYIRQRLLFLVECHQQCYNHPL**RL*CCAEH*MLVWYHN
>C2, frame-2 (200 aa)FSPIGLQCEAWPSMGKALGGFLRLYGISLMMSLNLSRVNNTTSDSSPLASLMVSRRSKGRAFLHFLLGTGSASVTRTSGKLHTPIPSPTDQTQTNWAPIKIFGSEFDIKYEIERLLVREYHRRLKLQDLYDAEVAALGWIIQRGTASTRNAGEAGKR*QRAGSNSKKHVCLCICHRRFVDAWNWCLW***LVYCCW*HPF
>D2, frame+2 (230 aa)AELRMKGMYVNDVRKRKEELEQRLSAMMDAFQKIKTERGQKAAQIQAITQKMTEMSEKKKIVENELVVLCRESALKETELVKKKRQVQELRQVCKNLRMEKNRQRKSLEKAFEDERNVKINVRRTNAQIADLEQEMNGVKLRYHDLIEKRNYAGVQVLGPPMMNSACCMNG *MRKKKAI K*WVFS *TNARDEENTVHYG IKLWPNLCP*GWDVVSTGLISQKVKGSLGGK
>E3, frame-1 (191 aa)LKRWPKLGGLAGSWDPKGNTFWEEGAAWVFRRASRVWASMLLVAPTAYQLRMYTDLLLKCLASVRAAMDASPSSGDGSPECGLGFVQAGGALPIALSRQLRLSAEALKCQMRRLEGTSHVDDAHATANLPVVAAVLQMLSEVLLELPRRLAELITAHHQPLRHAWLRLESDAISRAKSHSANSLTSLREPA
>F1, frame-2 (180 aa)EVGGGERKFISVSHELNGNANVVFLSQNMKVPTCIVTIHGRLFWHWSPKDSNGADLIPREKAPFLTCFFDRSISLLLDNVSNIWTIQNEMIFMRPDHTLVHENGSASSPLFFSNEATRVESMRRRYLPNASLEVMQSVVEHFGSDSDVLAFITGHLPTLPADRVERALKDRSVMLSLLQR
>G1, frame-3 (186 aa)RRAHLPATEGGSFLGDRVLKELNADPQDDVHG*TIGTNRRDTVHNFLSMATRGRRKQRSVTKTTMRSRKQMFIKWRLRRCRMLKIAFPRSRATAYKMSRSAMSPFLPSRIHLLLTMNRPPRARRNYFRKQHPTRRRISSR*LKRSLIPWRNSNGS*EHFRRKLIVLAVLWRIFKFFHRKNSPFLQR
>I1, frame-3 (149 aa)RIPLVDCTAAKRSPRRVRGVE*SCNTHINIYI*PGSCCREKRKRERESL*EIYIYMPSAPLTGEVARYSSPLFMYRRILKNAAPKPETVFTAKDNSKTAFHVLTQRPGFGNNPYAVSRWYLKEKVHSSNRNRLEGLYECVLGASCTGSC
>J1, frame+3 (166 aa)QLPREVQGG*EESNNRAINI*ISIHIIAW*LL*EKKGKNKRERERESL*EIYIYIYMPSAPLTGEVARYSSPLFMYRRIIKNAAPKPETVFTAKDNSKTAFHVLTQRPGFGNNPYAVSRWYLKEKVHSSNRNRLEGLYECVLCASCTGSCESNSRGRHGGREHATS
163

Annexes
Annexe 24. Alignement de la séquence des acides aminés déduite
de la séquence nucléotidique D2 avec la protéine flagellaire de C. reinhardtii.
tr Q8S4W6 Coiled-coil flagellar protein [MB02] [Chlamydomonas reinhardtii]
920 AA
align
Score = 59.7 bits (143), Expect = 3e-08Identities = 43/159 (27%), positives = 77/159 (48%), Gaps = 8/159 (5%)
D2 : 4
Sbjct: 461
D2 64
Sbjct: 521
D2 120
Sbjct: 577
RMKGMYVNDVRKRKEELEQRLSAMMDAFQKIKTERGXXXXXXXXXXXXMTEMSEKKKIVE 63R+K + + +++K ++E+ +R+ + +K +R TEM +K K+++RIKEVELAELKKIRKEVGRRIRDFEKLYDLVKNQRNKFVNLIQAASQSTTEMKDKLKVLQ 520
NELVVLCRESALKETELVKKKRQVQELRQVCKNLRMEKNRQRKSLEKAFEDERNVK---- 119NEL +L E +K+ L ++ Q LR+E R L F D++ VNELDILQNEVGIKDKLLQQQHTQHAANIAERDQLRVELGR----LGMVFRDKQAVVDEQI 576
INVRRTNAQIADLEQEMNGVKLRYHDLIEKRNYAGVQVL 158V + NA l E+EM +K +Y +IE RNY G+ ++
AEVDKLNAIINGCEKEMLRLKKQYELVIEARNYTGIMLI 615
164

Annexes
Annexe 25. Alignement de la séquence des acides aminés déduite
de la séquence nucléotidique F1 avec la protéine hypothétique de L. major.
tr Q9NEAO Hypothetica1 51. 4 kDa protein [L4803. 04] [Leishmania maj or] 460 AA
align
Score 134 bits (337), Expect = 6e-31Identities = 69/169 (40%), positives = 103/169 (60%), Gaps = 4/169 (2%)
F1: 5 GERKFISVSHELNGNANVVFLSQN--MKVPTCIVTIHGRLFWHWSP-KDSNGADLIPREK 61G K I+V E N N N+V + Q+ M+VP CIVT+HG++ + W+P + P K
Sbjct: 283 GGDKSIAVEIEFNPNFNMVMMDQSYHMRVPVCIVTVHGKMRYLWNPIRPETQQPTFPAGK 342
FI: 62 APFLTCFFDRSISLLLDNVSNIWTIQNEMIFMRPDHTLVHENGSASSPLFFSNEATRVES 121AP ++ FFDR+++L D +N W+I N+M+ MRPD +VH++G+ SSPLFF+N +R+E
Sbjct: 343 APMISTFFDRTLTLTWDGNTNAWSIANDMVLMRPDRAVVHDDGTPSSPLFFANTPSRIEQ 402
F1: 122 MRRRYLPNASLEVMQSVVEHFGSDSDVLAFITGHLPTLPADRVERALKD 170MRRR +P + +VM +V SD +V l L LP D++ D
Sbjct: 403 MRRRLMPGITADVMAVIVNATSSDEEVKQSIM-QLLMLPQDQLHAVASD 450
165

PUBLICATIONS

Publications
Poster présenté au Congrès: MEEGID-VI, 24-27 juillet 2002, Paris.
MOLECULAR ANALYSIS OF THE DIFFERENTIAL GENEEXPRESSION IN NATURAL SUSCEPTIBLE AND BENZNIDAZOLE
INDUCED RESISTANT Trypanosoma cruzi STRAINS
VILLARREAL P., Diana"; HIDE, Mallorie; BARNABE, Christian;T1BAYRENC, Michel and BANULS, Anne-Laure.
Centre d'Eludes sur le Polymorphisme des Micro-Organismes (CEPM) UMR CNRS-IRD 9926911 avenue Agropolis, BP 64501, 34394 Montpellier Cede>< 5, France.•E-mail: [email protected]
CEPM
'Are the drug reslstance mechanlsms different among geneticgroups of T. cruzi?
'What are the genes involved on Trypanosoma cruz; drugreslstance?
ln absence of a suitable model to explain parasite resistance in thisspecies, some basic questions remains to be answered:
'Are lhere any differences ln gene expression belween reslslantand susceptible strains?
By the end of 80's the existence of natural resistance in T. cruziagainst drugs was demonstrated (Filardi and Brener, 1987). However,biochemical and moleeular mechanisms by which T, cruzi parasitesbecome resistant remains to be understood completely. Differentmodels have been proposed to explain drug resistance To resolve these questions we have proposed to study the differentialmechanisms in other parasites like: Leishmania, Plasmodium, E gene expression in T. cruzi parasites, Our objective is to analyse thehislolytica and T. vaginalis. As a result, specific molecular events gene expression as resull of the Benznidazole exposure using thesuch as gene amplification and/or over-expression have been Random Ampllfied Differentlally Expressed Sequences (RADES)observed to occur in association with parasite resistance. Further, technique (Murphy and Pellé,1994). Two levels of drug concentrationsome specifie genes like the rndr gene have been implicated in the are considered: the inhibitory concentration (IC,o) and the maximumresistance pathway. In spite of several efforts, expression of known human blood tolerance levels (50 ~M). Resistance-related geneputative resistance genes on resistant strains of T. cruz; couId not be expression is being studied using natural susceptible anddemonstrated so far (Nirdé el al., 1995; Murta el al., 2001). Benznidazole induced resistant slrains.
,..---------,
INTRODUCTIONChagas' disease represents one of the major human diseases ofconcem for economicaily baekward communities in tropical areas ofcentre and south America. Disease lrealment in endemie regions iscurrently limited to two drugs: the Benznidazole and the Nifurtimox.However, both these molecules are reported to exhibit not only a lowefficacy but also harmful and toxic effects on treated patients.
MLEE InalysisMETHODS
1 , , .. S 6 1 •• 10 ft 12
RADES Proliles
1Non DrugeXPTilion
1mRNA liolallon
cDNA s nthesis
:;. .
-- --- -
- ------
1Drug expositional IC60I,evel (+)
Plraslte's growlh ----. In vllro suscepllbilltykin lies delermlna~n (MTT/PMS)
parasilJs growth
818 •
Anllysis of speclflCpolymorphie bands
Isolalion andcloning
Biochemical and molecularParasi 's cullure .--__-GllW:W.lIt·WI.Ilm__,
RAPD Ina/ysis
BANDSEQUENCING
AND
IDENnFICAnON
r.bkt 1. ICso velues determined by MTI/PMSmicro-metnod (Mueras-Serrano et al.. 2000)
Ge...... IC N
gloup s..... !llM1 Mean t50
32.212.2 1~9t1354 A6.'
lib 1136.7 8.9123 a8.8
IId ,....5 12.515.62 810
Ile 10 14.511 12.1 1212.5'2 •.5
Il. 13 6.•14 7.3 7.5i 0.76 C15 63
Il c 18 •. 117 .3 ft.110.2 C1. 7.9
PRELI IHARY RESULTS AND PERSPECTIVESPreliminary results indicated a significant difference (p<O.OS) in the IC50s values in different T. cruzigenetic groups (Table 1.). Further, an important intra-group variation between strains of samegenetic group was evidenced. Stalistical comparison between genetic groups with negard to IC50levels. allowed to organise the differenl genetic groups in clusters (A, Band Cl.
Studied strains were exposed to the specific IC50 level and molecular analysis was canried out using22 RADES primers. Till date, 18 differential bands have been identified as related with resistant (9)and susceptible (18) phenotypes involving 6 different stocks of T. cruzi. The isolation andidentification of these RADES products are currenlly in progress.
Ailhough induction of drug resistance in vilro is a difficult process due to high mortality and slowgrowth of potential resistant parasites, a step by step methodology is being optimised in order toinduce a rapid stabilisation of drug-exposed parasites.
The next step, it is proposed to obtain RADES profiles from natural susceptible and induced resistantstrains. Then, differential bands will be isolated and sequenced in order to identify some putativegenes involved in T. cruzi Benznidazole resistance.
ln the future, developmenl of suitable melhods for detection of differentially expressed genes in T.cruzi will be important to achieve studies on drug resistance. Nevertheless further research effortsare needed to define a fitting molecular model to explain the natural resistance mechanism in thistropical parasite.
• Fllardi. L.S. & Brene" Z. (1987) T,.nsaclions of d'te Royal Society of Tropical MedlclMand Hyg~ne. 81:755-759.• MUNa-84>r,ano, 5..; Hog.I·Rulz. J.J.; G6me.z.-Sarrk). A. (2000) Par_Ilot Res. 10:089-1002.
REFERENCES, Mm."Y, N. a. & ..... R. (,...) Gen.'" ,&J.'"• Murta. S.M.F.;dos~. W.G.; AI\KWo. C.; Nk'de. P.; Morel,.. E.s.A; Rom..ha. A.J. (m()1) Mol a 8k1' P_.t\OI117:2:23-228• Nif. P.; lM'roque. C.; &.nIIb6. C. (1"5) C.R Ac.d. Sel Pan... Sdel"lOH de" vW.JfeSC"""" 31' :1~12'"
4: N6'h Inlemellonel Meeting on Moleculer Epldemloklgy end Ewlullonsry Genetlcs ln Infections Dlses!es', MEEGID-V1, 24-27 July 2002, Peris (Frvnœ)

Abstracts/Infection. Genetics and Evolution 2 (2003) 235-321 317
their drug regimen, resistance to 3TC was more frequentthan to AZT. Two of the three patients, with viruses resistantto PIs, acquired mutations associated with cross-resistance.Strikingly, 5 (39%) of the 13 patients developed resistancesto drugs which they never received (n = 3) or which theyreceived 18 or 36 months ago (n = 2). ddI/d4T pressure hadselected AZT-resistant viruses in four patients, and indinavirhad selected a nelfinavir-resistant virus in one patient.
Conclusion: In contrast to other reports from developing countries, our study showed that despite the presenceof many minor mutations at baseline, implementation ofHAART together with good c1inical, biological and logistical monitoring can reduce the emergence of resistant strainsin Africa.
Molecular analysis of the differential gene expression innatural susceptible and benznidazole induced resistantTrypanosoma cruzi strains
D. Villarreal*, M. Hide, C. Barnabé, M. Tibayrenc, A.-L.Bafiuls
Centre d'Etudes sur le Polymorphisme des MicroOrganismes (CEPM), UMR CNRS-IRD 9926, 911 AvenueAgropolis, B.P. 64501, 34394 Montpellier Cedex 5, France.
*Corresponding author. Tel.: +33-0467416232;fax: +33-0467416299.
E-mail address:[email protected].
Chagas disease caused by the parasite Trypanosoma cruziis one of the most important tropical diseases in central andSouth America. Almost 10 million people in 17 countriesare concemed. Although two drugs are currently used inChagas disease treatrnent (Benznidazole and Nifurtimox), itremains ineffective to chronic forms and further undesirabletoxic effects have been observed. In addition, occurrence ofnatural resistant T. cruzi strains in endemic zones, has beenrecently reported. In the last decade, research about drugresistance in parasitic organisms has become important.As consequence, sorne biochemical and genetic mechanisms have been proposed in sorne known parasites likePlasmodium, Leishmania, Trichomonas and Entamoeba.Nevertheless, mechanisms on drug resistance in T. cruziremain almost unknown. The aim of this work is to studythe differential gene expression in T. cruzi as result ofthe Benznidazole exposure. Eighteen naturally susceptiblestrains to Benznidazole drug, representing the overall genetic diversity of the parasite, were analyzed. Preliminarystrain characterization was carried out by both molecular (RAPD) and multi-enzyrnatic (MLEE) analyses. Geneexpression was studied using the Random Amplified DifferentiaIly Expressed Sequences (i.e. RADES), a techniquewhich aIlows to screen a widely mRNA population at onespecific developmental stage. The 50% inhibitory concentrations (i.e. ICsos) to Benznidazole were determined usingthe MTTIPMS micro-method. A preliminary comparativestudy on drug susceptibility was carried out at ICsos. Datashowed an important variation of the ICsos mean values
(7.5-16.9I-LM) between different strains as a result of drugexposure. After determination of the strain-specific ICsoslevels, a differential gene expression analysis was fulfilledusing the RADES technique. At present, nine differentialbands have been identified as a product of drug exposure(at the ICso level) in six of the T. cruzi strains. The isolationand identification of these RADES products are currently inprogress. The next step in this work will be to compare thedifferential gene expression between resistant versus susceptible strains. Accordingly, resistance is currently beinginduced using a continuous drug pressure protocol in sixrepresentative strains. We hope that the utilization ofmolecular tooIs Iike RADES will help to provide a more comprehensive picture of drug resistance mechanisms in T. cruzi.
Epidemiology of Yersinia pestis in Madagascar and theworld
A. Vogler l , F. Chan2, K. Smith l , S. Chanteau2 P. Worsham3,
E. Camiel4 , P. Keim 1.*1Biological Sciences Department, Northem Arizona Uni
versity, p.o. Box 5640, FlagstafJ, AZ 86011, USA.2Institut Pasteur De Madagascar, Antananarivo, Mada-
gascar.3 USAMRIID, Ft. Detrick, MD, USA.4Institut Pasteur, Paris, France.*Corresponding author. Tel.: + 1-928-523-1 078;
fax: + 1-928-523-0639.E-mail address:[email protected].
Yersinia pestis is a recently emerged pathogen of great interest due to its past influences on history and its potentialfor use as a bioterrorism weapon. Unfortunately, differentiation between strains has been very difficult since evendistinct biovar isolates are nearIy identical in gene sequencecomparisons. A recently developed MLVA (multi-IocusVNTR analysis) typing system for Y. pestis, encompassing43 VNTR marker loci, represents a robust and rapid typingmethod that can be used to answer a number of epidemiological and evolutionary questions. We have typed both aworldwide diversity set and an extensive set of isolates fromMadagascar. Our worldwide diversity study has confirmedthe classification of Y. pestis into three biovars but also hasidentified additional phylogenetic groups. We find additional pestoides mediaevalis and pestoides antiqua groupsthat are interrnediate between the traditional biovars and Y.pseudotuberculosis. The evolution of Y. pestis from Y. pseudotuberculosis appears to have been a stepwise process. Theisolates included from the Madagascar study were collectedover a large time period (1939 to present) and from aIlknown plague-endemic regions. MLVA demonstrated greatgenetic discrimination with nearly aIl the Madagascar isolates distinguishable. In addition, multi-Iocus phylogeneticanalysis identified major genetic types within the study. TheMLVA genotype data were analyzed in conjunction with aIlof the available temporal, spatial and victim demographicdata in order to understand plague transmission patterns.

Publications
Article soumis à la revue "Experimental Parasitology"
Lack of correlation between in vitro susceptibility to Benznidazole and phylogenetic
diversity of Trypanosoma cruzi, the agent of Chagas disease.
Diana Villarreala, Christian Barnabéa, Denis Serenob, Michel Tibayrenca•.
aGénétique des Maladies Infectieuses GMI (Ex-Centre d'Etudes sur le
Polymorphisme des Micro-organismes - CEPM) CNRS/IRD UMR No. 9926.
bpathogénie des Trypanosomatidés INSERMIIRD UMR No. 008, IRD, 911 avenue
Agropolis, BP 64501, 34394 Montpellier Cedex 5, France.
·Corresponding author: Génétique des Maladies Infectieuses GMI (Ex-Centre
d'Etudes sur le Polymorphisme des Micro-organismes - CEPM) CNRS/IRD UMR No.
9926, IRD, BP 64501,34394 Montpellier Cedex 5, France. Tel. +33 4 67 41 62 07.
Fax: +3346741 6299. E-mail: [email protected]
168

Publications
Abstract
Chagas disease remains an important health problem in Central and South
America. Nitroimidazole derivative drugs like Benznidazole are commonly used to
treat T. cruzi infection. Natural variation of drug susceptibility between various T.
cruzi stocks has been proposed as a possible expianation of treatment failure. Thus,
the aim of this work was to determine potential correlations between Benznidazole
susceptibility of different T. cruzi stocks and their genetic diversity. For this purpose,
sixteen naturally susceptible stocks representing the overall genetic diversity of the
parasite were analysed. Genetic characterisation was assessed by bath random
amplified polymorphie DI'JA (RAPD) and multilocus enzyme electrophoresis (N1LEE)
analyses. Drug activity was determined by two cornplementary methods, the MTT
PMS micro-methad and FACs analysis. The 50% inhibitory concentrations (ICsos)
were determined. Important variation of ICso values (7.3 IJM to 16.9 IJM) among
stocks belonging ta different discrete typing units (DTUs) was recorded. Further,
correlation analysis showed that natural susceptibility to Benznidazole in T. cruzi
expressed as ICso level was not related with its genetic structure represented by the
different DTUs. These results are discussed in relation with the proposed hypothesis
establishing a Iink between genetic diversity and biological behaviour in T. cruzi.
Index Descriptors and Abbreviations: MLEE, multilocus enzyme electrophoresis;
RAPD, random amplified polymorphie DNA; ICso• inhibitory concentration; DTU,
Discrete typing unit; DMSO, dimethyl sulfoxide; PI, Propidium lodide; NJ, Neighbor
Joining method; UPGMA, Unweighted Pair-Group Method with arithmetic averages.
169

Publications
Keywords: Chagas' disease; Trypanosoma cruz;; Drug susceptibility; Inhibitory
concentrations (ICsos); Benznidazole; Genetic diversity.
1. Introduction
Chagas' disease caused by the parasite Trypanosoma cruz; is one of the most
important tropical diseases in Central and South America. While 16 to 20 million
people in 17 countries are concerned (WHO, 1991), almost 100 million are exposed
to the infection. Since the beginning of the 70s, two drugs have been used for
treatment of Chagas disease, namely Benznidazole and l\Jifurtimox. Nevertheless,
response to treatment remains quite variable and sometimes ineffective. Moreover,
undesirable adverse reactions have currently been observed among treated patients
(Brener, 1984; Teixeira et aL, 1994; Khaw and Panosian, 1995; Rodrigues Coura and
de Castro, 2002).
Occurrence of naturally resistant T. cruz; strains have supposed to be one of
the most important factors explaining the low rates of cure on some treated chagasic
patients from endemic zones (Filardi and Brener, 1987; Toledo et aL, 1997). In fact,
occurrence of predominant strains, exhibiting different levels of susceptibility (in
specifie geographic zones), have been associated to high variability of c1inical results
concerning treated patients.
Furthermore, susceptibility to Benznidazole in T. cruz; has been associated to
different morphological and physiological characteristics of the parasite populations
(Andrade et aL, 1985). These results are supported by further extensive studies
revealing an important variation of susceptibility to drugs among T. cruz; strains
(Brener et aL, 1976; Filardi and Brener, 1987; Neal and van Bueren, 1988; Revollo et
aL, 1998; Veloso et aL, 2001).
170

Publications
By using either isoenzyme or RAPD, previous works (Tibayrenc et aL, 1986;
Tibayrenc and Ayala, 1988, 1999; Tibayrenc, 1998) have not found any evidence of
random mating in T. cruz; natural populations. Genetic exchanges in Trypanosomes
are apparently restricted. Nevertheless, although clonai propagation is predominant,
there is evidence that genetic recombination in T. cruz; occurs in natural populations
of the parasite, under the form of hybrid genotypes subsequently stabilized by clonai
propagation (Bogliolo et aL, 1996; Brisse et aL, 2003). Experiments have recently
shown that genetic recombination can be obtained in the laboratory (Gaunt et aL,
2003). Some biological properties Iike virulence or drug resistance would be spread
from wild reservoirs to human populations as result of new genetic recombination
occurred in undisturbed sylvatic hosts (Stothard et aL, 1999; Gaunt et aL, 2003).
Several reports have focused on the link between genetic diversity and
biological or c1inical characteristics in T. cruz; (Andrade, 1976; Dvorak et aL, 1980;
Dvorak, 1984; Andrade et aL, 1985; Laurent et aL, 1997; Revollo et aL, 1998; Toledo
et aL, 2002). However, the lack of an overall sampling covering the entire diversity
within T. cruz; species remains the major limitation of such studies (Revollo et aL,
1998). In the present report, different stocks representing a fair sample of the entire
genetic diversity in T. cruz; were studied in relation with their susceptibility to
Benznidazole. Further, genetic variability was scored using both isoenzyme and
RAPD methods. Relationship between drug susceptibility and genetic diversity was
analysed and possible implications with the clonai structure in this species was
discussed.
171

Publications
2. Materials and methods
2. 1 Biological Material
For this study sixteen stocks representing the whole genetic diversity of T. cruzi
were selected (Table 1). Epimastigote forms belonging to the different stocks were
cloned by micromanipulation method. Culture of epimastigotes was carried out at
2rC in LIT medium (Liver Infusion Tryptose) supplemented with FCS (10%). For
harvesting, cultured epimastigotes were centrifuged (2S00 9 during 20 min, 4°C) and
then washed twice in PBS solution. Final pellets were weighted and stored at -SO°C
up to characterisation.
2.2 Genetic characterisation of the stocks
Genetic characterisation was carried out by MLEE and RAPD analyses. A total
of r1ineteen different isoenzymatic systems corresponding to 22 polymorphie locus
were scored, namely: aconitate hydratase (E.C.4.2.1.3, ACON), alanine
aminotransferase (E.C.2.6.1.2, ALAT), NAD (P) H dehydrogenase (quinone)
(E.C.1.6.99.2, DIA), glyceraldehyde 3-phosphate dehydrogenase (phosphorylating)
(E.C.1.2.1.12, GAPDH), glutamate dehydrogenase (NAD+) (E.C.1.4.1.2, GDH-NAD),
glutamate dehydrogenase (NADP+) (E.C.1.4.1.4, GDH-NADP), aspartate
aminotransferase (E.C.2.6.1.1, GOT), glucose-6-phosphate 1-dehydrogenase
(E.C.1.1.1.49, G6PD), glucose-6-phosphate isomerase (E.C.5.3.1.9, GPI), isocitrate
dehydrogenase (NAD+) (E.C.1.1.1.42, IDH), leucyl arninopeptidase (E.C.3.4.11.1,
LAP), malate dehydrogenase (E.C.1.1.1.37, MDH), malate dehydrogenase
172

Publications
(oxaloacetate decarboxylating, (NADP+) (E.C.1.1.1.40, ME), mannose-6-phosphate
isomerase (E.C.5.3.1.8, MPI), purine-nucleoside phosphorylase (E.C.2.4.2.1, NHi),
tripeptide aminopeptidase (or peptidase B) (E.C.3.4.11.4, PEP-1, substrate: leucyl
leucyl-Ieucine), cytosol non-specifie dipeptidase (or peptidase A) (E.C.3.4.13.18,
PEP-2, substrate: leucyl-L-alanine), phosphogluconate dehydrogenase
(decarboxylating) (E.C.1.1.1.44, 6PGD), phosphoglucomutase (E.C.5.4.2.2, PGM),
superoxide dismutase (E.C.1.15.1 .1 , SOD).
Isoenzymatic analysis
Isoenzymes were revealed by electrophoresis in cellulose acetate plates
(Helena System) according ta previous pratocols (Ben Abderrazak et al.,1993 and
Barnabé et aL, 2000). Briefly, the parasites were Iysed in a hypotonie solution
(Oithiotreitol 2 mM, Acide amino-n-capraic 2 mM, EDTA 2 mM). Homogenised
samples were maintained 20 min on ice and then centrifuged (15000 9 during 10
min, 4°C). Soluble fractions were recovered and stored at -80°C until electrophoretic
analysis.
RAPD analysis
DNA was extracted fram Iysed cell following the pratocol suggested by Brisse et
al. (2000) with slight modifications. In short, cellular pellets were resuspended in 400
!JI Tris-HCI, 10 mM (pH 8.0), EDTA 1 mM with SOS ta a final concentration of 1%.
Samples were incubated for 2 h at 3rC in the presence of RNase A (5 IJg mr\ and
then overnight at 55°C in presence of Proteinase K (200 !Jg mr1). After two Phenol
173

Publications
extractions and two chloroform:isoamyl alcohol extractions, the DNA was ethanol
precipitated and resuspended in sterile water. DNA concentration was estimated by
spectrophotometry at 260 nm. Ali DNA extracts were stored at -BO°C until molecular
analysis.
For RAPD analysis a total of 22 different primers, giving reproductible bands,
corresponding to the A, B, F, N, Rand U kits from Operon technologies®, were used:
namely: A2, A3, A4, A7, A9, A10, B1, B5, B11, B1B, F13, N4, N15, N1B, R1, R2, RB,
R10, U13, U16, U19 and U20. PCR amplifications were carried out in a final volume
of 60 1-11 containing 100 pM each dNTP's, 10 I-IM primer, 0.9U Taq polymerase
(Roche), 61-11 Tampon 10X and 20 ng DNA. FortY five cycles (denaturation 1 min at
94°C, annealing 1 min at 36°C and elongation 2 min at 72°C) were followed by a final
elongation step of 7 min at 72°C. The amplifications were carried out in a PTC-100
Thermocycler (MJ Research). RAPD products were revealed by electrophoresis in
agarose gels (1.6 %).
IVILEE and RAPD analyses made it possible to attribute ail stocks to the 6
different genetic subdivisions or DTUs (Tibayrenc, 199B) identified until now in T.
cruzi (Barnabé et aL, 2000).
2.3 ICso determinations
For this study, Benznidazole drug (Ro7-1051) was supplied by Dr. R. Jakob- Roetne,
Dr. E.M. Gutknecht and P. Weber (F. Hoffmann - La Roche Ltd, Basel, Switzerland).
Parasite susceptibility to Benznidazole was measured as the ICsQ, corresponding to
the drug concentration which inhibits parasite growth by 50%. Inhibitory
concentration was assessed by two methods: MTI-PMS micromethod and f10w
cytometry. The drug powder was dissolved in DMSO to a final concentration of 10
174

Publications
mM, then dilutions were prepared in a LIT medium in order to obtain required
concentrations (50, 100, 150, 200, 500 and 2000~M).
To determinate by MTT-PMS method the inhibitory concentrations (ICsos
values) of T. cruzi epimastigotes to Benznidazole, we have previously determined the
linear correlation between absorbance values and numbers of parasites per ml in the
log-phase culture.
The MTT-PMS determination of the ICso values was assessed as previously
described by Mue/as-Serrano et al. (2000) with some modifications. Briefly, 90 !JI of
parasites in logarithmic growth phase were distributed in 96-well fiat bottomed plates
at a concentration of 1x1 06 epimastigotes mr1. Increased concentrations of
Benznidazole (10x) were displayed per weil and the plates were incubated during 72
h at 2rC. Each treatment (including controls) was repeated six time while each
experiment was performed at least twice. Plates were incubated for 4 h at 2rC after
adding 10~1 of a Mn (2.5 mg mr1) - PMS (0.22 mg mr1) solution. Formazan crystals
were dissolved in a SOS (10%) - HCI (0.01 N) solution, and then agitated for 30 min
at room temperature. Absorbance values were recorded at 570 nm in a Multiskan
spectrophotometer. ICso values were calculated following the formula stated by Hills
et al. (1986): log (ICso)= log (X1) + [(Y1-yo)/2)/(Y1-Y2)] [log(x2)-log(x1)] , where finding
two concentrations, X1 and X2, such that the parasite density, Y1, at concentration X1
(and ail lower concentrations) was more than half of the density found in the control,
Yo, and that the parasite density, Y2, at concentration X2 (and ail higher
concentrations) was less than a half of Yo.
Obtained values from MTT-PMS method were compared with f10w
cytofluorometric measures according to Vergnes et al. (2002). For that three stocks
(OPS c111, CBB c13 and Mn c12) belonging to different DTUs (l, lib et IId,
175

Publications
respectively; see Barnabé et aL, 2000) were analysed. Epimastigotes samples (1 x1 06
cells mr1) in exponential growing phase, were cultured in 24 cell-culture plates
containing Benznidazole at different concentrations. Plates were incubated at 2rc
during 72 h. Samples were stained using a PI solution in PBS (1X). Mean cell
number and viability was determined by f10w cytometric analysis, using a FACScan
flow cytometer (Becton Dickinson) interfaced to a digital Power Macintosh and
equipped with an argon-ion laser tuned to 488nm, as previously described (Vergnes
et aL, 2002). Briefly, epimastigote number and viability were calculated by
determining their specific forward (FSC) and side (SSC) Iight-scattering proprieties.
Following FSC (size) and SSC (granularity) gain adjustments. Then, the relative IP
fluorescence was collected and displayed using a logarithmic amplification (FL-2, FL
3). The quadrant statistics allowed us to determine the viable cells percentage.
2.4 Data analysis
Jaccard's genetic distances (Jaccard, 1908) between pairs of stocks were
calculated by using the software Genetic Toois Box. NJ method (Saitou and Nei,
1987) as weil as UPGMA method (Sneath and Sokal, 1973) were used to cluster the
genotypes, with the software Neighbor from the PHYLIP package [Felsenstein J.
PHYLIP (Phylogeny Inference Package) version 3.57c. Seattle, WA: University of
Washington ,1995]. Phylogenetic trees were visualised using the TREEVIEW
software (Page, 1996).
The IC5D average values per treatment (Le. stocks) were compared by an
ANOVA test while comparisons between genetic groups were performed using the
Fisher's PLSD test from the Statview program. Correlation between IC5D values and
176

Publications
genetic distances was tested by the Mantel test (Mantel, 1967) using the software
ADE-4 software (version 2001, Thioulouse et aL, 1997).
3. Results
Phy/ogenetic ana/ysis
Genetic distances calculated fram MLEE and RAPD data were highly
correlated (r=0.729, P<10-4), in spite of some branching differences (see Figure 1).
Indeed, the genetic groups IId and Ile are grauped separately depending on RAPD or
rvILEE analysis. Nevertheless, the two methods support the main subdivision within
T. cruzi, Iineage 1 and Iineage II. Additionally, RAPD and MLEE exhibited the same
c1uster organisation, which is in agreement with the six previously praposed
phylogenetic suddivisions organised into two main Iineages (Tibayrenc, 1995, 1996;
Barnabé et aL, 2000; Brisse et aL, 2000).
/Cso by MTT-PMS method
Data showed a significant correlation (r=0.9984, P<0.05) between the
absorbance values (as determined by spectrometry) and the number of parasites at
the log phase culture.
The ICsos for the 16 stocks determined by MTT-PMS micro-method, are
presented in Table 2. An important variation between stocks was observed in the
whole sampie as weil as within almost each genetic group. Comparisons of ICsos
mean values between ail genetic groups showed no significant difference. Further,
no signmcant difference was observed between lineage 1 and Iineage Il (P=0.1).
177

Publications
Moreover, there was no correlation between genetic distances (calculated
fram RAPD and MLEE data) and IC5D distances (i.e. IC5D differences between pairs of
stocks), as revealed by the Mantel test (Table 3).
Table 4 show the IC5D values determined for 3 stocks by using MTT-PMS
method, f10w cytometry and their comparison with [3H]-Thymidine method (Revollo,
1995) after 72 h of Benznidazole exposition.
4. Discussion
The genetic diversity of the parasite Trypanosoma cruzi was demonstrated
first by using MLEE and confirmed by other markers such as kinetoplastic DNA
minicircle patterns, DNA content analysed by pulsed field gel electraphoresis, DNA
fingerprinting,' RAPD, karyotyping, polymorphism in ribosomal RNA genes,
polymorphism in mini-exon genes, microsatellite analysis (reviewed in Macedo and
Pena, 1998) and then different biological characteristics were examined such a
virulence, grawth rates, in vivo and in vitro susceptibilities to drugs, antigenic profiles,
oxidative metabolism (Nozaki and Dvorak, 1993).
Departures from Hardy-Weinberg equilibrium and linkage disequilibrium have
constantly been found within the species, suggesting that T. cruzi undergoes
predominant clonaI evolution (Tibayrenc et al., 1986, 1993; Tibayrenc and Ayala,
1988), which explains the strong correlation found between RAPD and MLEE genetic
data. However, as already stated, events of recombination are present, even if they
are not frequent enough to brake the prevalent pattern of clonai population structure.
T. cruzi appears to be structured into two main Iineages, T. cruzi 1 and T. cruzi Il
(Momen, 1999), the last being sub-structured into five discrete typing units (DTU lia,
lib, IIc, Iid and Ile; Barnabé et aL, 2000) exchanging few genetic material between
178

Publications
them. Two of them (DTU Iid and Ile) are the product of ancient hybridization events
(Brisse et aL, 2000).
Many authors have explored possible links between genetic diversity and
biological diversity (Laurent et aL, 1997; De Lana et aL, 1998; Revollo et aL, 1998).
However, most of them have used only tree out of the six phylogenetic groups (DTU
l, lib and Ild) and besides most studies have largely involved T. cruzi strains from
widely dispersed geographic regions, rather than multiple samples from sympatric T.
cruzi populations in a single locality (Stothard et aL, 1999).
Recently, to verify the impact of T. cruzi clonai evolution on the biological
properties, fourteen parameters related to virulence and pathogenicity in mice were
assessed (Toledo et aL, 2002), the results showed a correlation between biological
differences and phylogenetic distances among T. cruzi clonai genotypes. The same
authors have further established clear links between phylogenetic distances and in
vivo drug susceptibility to Benznidazole and Itraconazole in mice for 3 out of 6 T.
cruzi DTUs (Toledo et aL, 2003).
Contrary to these in vivo experiments, our results demonstrated that when we
considered the overall DTU's, correlation between drug susceptibilities and
phylogenetic distances became no significant, at least for Benznidazole. This is
probably the consequence of the high intra-DTU variation of the IC5D values
surveyed, suggesting that for this parameter of in vitro drug susceptibility, the
subdivision into DTUs has no predictive value.
ln agreement with Revollo et al. (1998), we have found variability within each
genetic group. The discrepancy observed between the two studies is probably due to
the differences in the methodology. Brie'tly, in the present study two cornplementary
methods were used to detect live ceUs or dead cells. First, the MTI-PMS method is
179

Publications
based on the ability of viable cells ta MTT via dehydrogenase activity. Second,
propidium iodide stains dead cells. Propidium iodide stained cells have their plasma
membrane sa damaged that they are incapable of excluding propidium iodide and
thus are definitely dead. Interestingly, the ICSD values estimated by 110w cytometry
analysis corroborate results obtained by the MTT-PMS micro-method. During this
study, drug susceptibility was determined in cell culture having the same inoculum
and after a 72-hour contact with the drug. These conditions allowed us ta directly
compare drug susceptibility of the various stocks tested. These conditions were
slightly different from those of Revollo et al. (1998), where the time of contact and the
initial inocula depends on the capacity of cell ta proliferate. These differences in bath
incubation time and inocula make direct comparison of drug susceptibility more
tentative (Revollo, 1995). Moreover flow cytometry perform numerous quantitative
measurements on each individual cell within a large population, allowing a direct
measure of the cell viability (Kirk et aL, 1998, Moore et aL, 1999, Delgado et aL,
2001, Kamau et aL, 2001). Therefore, it can be considered as a standard method.
Nevertheless, when we compared ours results with Revollo' ones (1995), the
ICso values were correlated (r=0.608, P=0.033) for the same three genetic groups
previously surveyed by Revollo (l, lib and IId, respectively). At this point, it is difficult
ta know whether our negative results are due ta a real lack of Iink between T. cruzi
genetic diversity and in vitro drug susceptibility or ta a statistical type Il error
(impossibility ta reject the null hypothesis due ta a lack of power of the test). The
structure of our sample (only 3 stocks per DTU with a broad span of values within
each DTU) could favour ta a certain extent the risk of a type Il error. Still the fact
remains that we did not find an obvious link between classification into DTUs and in
vitro susceptibility ta Benznidazole. These results are not totally contradictory with
180

Publications
the results obtained by other researchers fram the same team (Laurent et aL, 1997;
Revollo et aL, 1998; Toledo et aL, 2002, 2003). As a matter of fact, for other
biological parameters, even if the statistical correlation between genetic distances
and biological values was highly significant, a strong intra-DTU variance was
constantly observed. Such a pattern can be explained by either phenotypic changes
or the complexity of the genetic mechanisms involved in the parameters surveyed or
a certain level of genetic exchange (Gaunt et aL, 2003). The three hypotheses are
not exclusive of each other. Other authors have reported some variations in
susceptibility to drugs (Veloso et aL, 2001) among stocks and with other parameters
such as parasitemia, virulence and pathogenecity in the progeny of T. cruzi strains
derived from parental stocks and maintained up to 7 years in dogs (Veloso et aL,
2003 in submission). In those cases, the observed variability could be interpreted as
a consequence of the phenotypic alterations induced by the maintenance of the
parasites in different host species.
ln the last decade research about drug resistance in parasitic organisms has
become important and some biochemical and genetic mechanisms have been
proposed in some known parasites Iike Plasmodium (Wilson et al., 1989),
Leishmania (Ouellette et Papadopoulou, 1993), Trichomonas and Entamoeba
(Ullman, 1995), Giardia doudenalis (Upcroft and Upcroft, 1993). Mechanisms
previously associated with decreased sensitivity or resistance to chemotherapy in
parasitic protozoa include decreased drug uptake, increased export to drugs,
decrease in drug activation, alteration of the targets enzyme to decrease drug
binding (Reviewed by Ullman, 1995) and recently, by upregulation of the secretory
pathway (Engel et aL, 2000). Nevertheless, mechanisms on Benznidazole resistance
in T. cruzi remain almost unknown.
181

Publications
5. Acknowledgements
This work was supported by the IRD Institute in the form of an scholarship for
the first author.
6. References
Andrade, S.G., 1976. Tentative for grouping different Trypanosoma cruzi strains in
some types. Revista do Instituto de Medicina Tropical de Sao Paulo 18, 140-141.
Andrade, S.G., Magalhaes, J.B., Pontes, AL., 1985. Evaluation of chemotherapy
with Benznidazole and Nifurtimox in mice infected with Trypanosoma cruzi strains
of different types. Bulletin of the World Health Organization 63,721-726.
Barnabé, C., Brisse, S., Tibayrenc, M., 2000. Population structure and genetic typing
of Trypanosoma cruzi, the agent of Chagas disease: a multilocus enzyme
electrophoresis approach. Parasitology 120, 513-526.
Ben Abderrazak, S., Guerrini, F., Mathieu-Daudé, F., Truc, P., Neubauer, K.,
Lewicka, K., Barnabé, C., Tibayrenc, M., 1993. Isoenzyme electrophoresis for
parasite characterisation. In: Hyde, ..I.E. (Ed), Methods in Molecular Biology.
Humana Press, New York, pp. 361-382.
Bogliolo, AR., Lauriapires, L., Gibson, W.C., 1996. Polymorphisms in Trypanosoma
cruzi: Evidence of genetic recombination. Acta Tropica 61,31-40.
Brener, Z., 1984. Recent advances in chemotherapy of Chagas disease. Mem6rias
do Instituto Oswaldo Cruz 79,149-155.
Brener, Z., Costa, C.A.G., Chiari, C.A, 1976. Differences in susceptibility of
Trypanosoma cruzi strains to active chemotherapeutic agents. Revista do Instituto
de Medicina Tropical de Sao Paulo 18, 450-455.
182

Publications
Brisse, S., Barnabé, C., Tibayrenc, M., 2000. Identification of six Trypanosoma cruzi
phylogenetic Iineages by random amplified polymorphie DNA and multilocus
enzyme electrophoresis. International Journal for Parasitology 30, 35-44.
Brisse, S., Henriksson, J., Barnabé, C., Douzery, E.J.P., Berkvens, D., Serrano, M.,
De Carvalho, M.R.C., Buck, G.A., Dujardin, J.C., Tibayrenc, M., 2003. Evidence
for genetic exchange and hybridization in Trypanosoma cruzi based on nucleotide
sequences and molecular karyotype. Infection, Genetics and Evolution 2(3), 173
183.
Buckner, F.S., Wilson, A.J., White, T.C., Van Voorhis W.C., 1998. Induction de la
resistance to azoles drug in Trypanosoma cruzi. Antimicrobial Agents and
Chemotherapy 42(12),3245-3250.
De Lana, M., Pinto, AS., Barnabé, C., Quesney, V., Noël, S., Tibayrenc, M., 1998.
Trypanosoma cruzi: compared vectorial transmissibility of three major clonaI
genotypes by Triatoma infestans. Experimental Parasitology 89, 1-5.
Delgado, G., Puentes, F., Moreno, A, Patarroyo, M.E., 2001. Flow cytometry, a
useful tool for detecting the lethal effect of Pentamidine on Leishmania viannia
complex promastigote forms. Pharmacological Research 44, 281-286.
Dvorak, J.A, 1984. The natural heterogeneity of Trypanosoma cruzi: Biological and
medical implications. Journal of Cellular Biochemistry 24, 357-371.
Dvorak, J.A, Hartman, D.L., Miles, M.A, 1980. Trypanosoma cruzi: correlation of
growth kinetics to zymodeme type in clones derived from various sources. Journal
of Protozoology 27,472-474.
Engel, J.C., Torres G., C., Hsieh, 1., Doyle, P.S., McKerrow, J.H., 2000. Upregulation
of the secretory pathway in cysteine protease inhibitor-resistant Trypanosoma
cruzi. Journal of Cell Science 113, 1345-1354.
183

Publications
Filardi, L.S., Brener, Z., 1987. Susceptibility and natural resistance of Trypanosoma
cruzi strains ta drugs used clinically in Chagas disease. Transactions of the Royal
Society of Tropical Medicine and Hygiene 81, 755-759.
Gaunt, M. W., Yeo, M., Frame, I.A., Stothard, J.R, Carrasco, H.J., Taylor, M.C., Salis
M. S., Veazey, P., Miles, G.A.J., Acosta, N., Rojas de A. A., Miles, M.A. 2003.
Mechanism of genetic exchange in American trypanosomes. Nature 421, 936
939.
Hills, M., Hudson, C., Smith, P.G. 1986. Global monitoring of the resistance of
malarial parasites ta drugs: statistical treatment of micro-test data. Working paper
2.8.5 Informai consultation on the epidemiology of drug resistance of malaria
parasites, Geneva, Switerzland.
Jaccard, P., 1908. Nouvelles recherches sur la distribution ·I~orale. Bulletin de la
Société Vaudoise de Sciences Naturelles Nat. 44, 223-270.
Kamau, S.W., Nunez, R, Grimm, F., 2001. Flow cytometry analysis of the effect of
Allopurinol and the dinitroaniline compound (Chloralin) on the viability and
proliferation of Leishmania infantun promastigotes. BMC Pharmacol. 1(1), 1.
Epub: http://www.biomedcentral.com/1471-2210/1/1.
Khaw, M., Panosian, C.B., 1995. Human Antiprotozoal Therapy: past, present, and
future. Clinical Microbiology Reviews 8, 427-439.
Kirk, S.M., Schell, RF., Moore, A.V., Callister, S.M., Mazurek, G.H., 1998. Flow
cytometric testing of susceptibilities of Mycobacterium tuberculosis isolates ta
Ethambutol, Isoniazid, and Rifampin in 24 hours. Journal of Clinical Microbiology
36,1568-1573.
184

Publications
Laurent, J.P., Barnabé, C., Quesney, V., l\Joël, S., Tibayrenc, M., 1997. Impact of
clonai evolution on the biological diversity of Trypanosoma cruzi. Parasitology
114,213-218.
Macedo, AM., Pena, S.D.J., 1998. Genetic variability of Trypanosoma cruzi:
Implications for the pathogenesis of Chagas disease. Parasitology Today 14,119
124.
lVIantel, N., 1967. The detection of disease c1ustering and a generalized regression
approach. Cancer Research 27, 209-220.
Momen, H., 1999. Taxonomy of Trypanosoma cruzi: a commentary on
characterization and nomenclature. Memôrias do Instituto Oswaldo Cruz
94(Suppl. 1), 181-184.
lVIoore, AV., Kirk, S.M., Callister, S.M., Mazurek, G.H., Schell, R.F., 1999. Safe
determination of susceptibility of Mycobacterium tuberculosis to antimycobacterial
agents by flow cytometry. Journal of Clinical Microbiology 37,479-483.
lVIuelas-Serrano, S., Nogal-Ruiz, J.J., G6mez-Barrio, A, 2000. Setting of a
colorimetrie method ta determine the viability of Trypanosoma cruzi
epimastigotes. Parasitology Research 86,999-1002.
Neal, R.A, van Bueren, J., 1988. Comparative studies of drug susceptibilities of five
strains of Trypanosoma cruzi in vivo and in vitro. Transactions of the Royal
Society of Tropical Medicine and Hygiene 82, 709-714.
Nozaki, T., Dvorak, J.A, 1993. Intraspecific diversity in the response of Trypanosoma
cruzi to environmental stress. Journal of Parasitology 79, 451-454.
Ouellette, M., Papadopoulou, B., 1993. Mechanisms of drug resistance in
Leishmania. Parasitology Today 9,150-153.
185

Publications
Page, R D. M., 1996. TREEVIEW: An application to display phylogenetic trees on
personal computers. Bioinformatics 12, 357-358.
Revollo, S., 1995. Impact de l'évolution c10nale de Trypanosoma cruzi, agent de la
maladie de Chagas, sur certaines propriétés biologiques médicalement
importantes du parasite. Thèse de Doctorat. Université de Montpellier Il, France.
209 p.
Revollo, S., Oury, B., Laurent, J.P., Barnabé, C., Quesney, V., Carrière, V., Noël, S.,
Tibayrenc, M., 1998. Trypanosoma cruzi: Impact of clonai evolution of the parasite
on its biological and medical properties. Experimental Parasitology 89, 30-39.
Rodrigues Coura J., de Castro, S.L., 2002. A critical review on Chagas disease
chemotherapy. Mem6rias do Instituto Oswaldo Cruz 97,3-24.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Molecular Biology and Evolution 4, 406-425.
Sneath, P.H.A., Sokal, RR, 1973. Numerical Taxonomy. The principle and practice
of numerical classification. In: Kennedy, D. and Park, RB. (Eds.). Freeman, San
Francisco, pp. 537.
Stothard, J.R., Frame, I.A., Miles, M.A., 1999. Genetic diversity and genetic
exchange in Trypanosoma cruzi: dual drug-resistant "progeny" from episomal
transformats. Mem6rias do Instituto Oswaldo Cruz 94 (Suppl. 1), 189-193.
Teixeira, A.R., Calixto, M.A., Teixeira, M.L., 1994. Chagas disease: carcinogenic
activity on the trypanosomal nitroarenes in mice. Mutation Research 305, 189
196.
Thioulouse J., Chessel D., Dolédec S., Olivier J.M., 1997. ADE-4: a multivariate
analysis and graphical display software. Statistics and Computing 7, 75-83.
186

Publications
Tibayrenc, IVI., 1995. Population genetics of parasitic protozoa and other organisms.
Advances in Parasitology 36, 47-115.
Tibayrenc, M., 1996. Towards a unified evolutionary genetics of microorganisms.
Annual Review of Microbiology 50,401-429.
Tibayrenc, M., 1998. Genetic epidemiology of parasitic protozoa and other infectious
agents: the need for an integrated approach. International Journal for Parasitology
28,85-104.
Tibayrenc, M., Ayala, F.J., 1988. Isozyme variability in Trypanosoma cruzi, the agent
of Chagas' disease: genetical, taxonomical, and epidemiological significance.
Evolution 42, 277-292.
Tibayrenc, M., Ayala, F.J., 1999. Evolutionary genetics of Trypanosoma and
Leishmania. Microbes and Infection. 1,465-472.
Tibayrenc, M., Neubauer, K., Barnabé, C., Guerrini, F., Skarecky, D., Ayala, F. J .,
1993. Genetic characterization of six parasitic protozoa : parity between random
primer DNA typing and multilocus enzyme electrophoresis. Proceedings of the
National Academy of Sciences USA 90(4), 1335-~339.
Tibayrenc, M., Ward, P., Moya, A, Ayala, F.J., 1986. Natural populations of
Trypanosoma cruzi, the agent of Chagas' disease, have a complex multiclonal
structure. Proceedings of the National Academy of Sciences USA 83, 115-119.
Toledo, M.J.O., Guilherme, AL.F., Silva, J.C., Gasperi, M.V., Mendes, AP., Gomes,
M.L., Araujo, S.M., 1997. Trypanosoma cruzi: chemotherapy with Benznidazole in
mice inoculated with strains from Paranâ state and from different endemic areas
of Brazil. Revista do Instituto de Medicina Tropical de Sao Paulo 39, 283-290.
Toledo, M.J.O., de Lana, M., Carneiro, C.M., Bahia, M.T., Machado-Coelho, G.L.L.,
Veloso, V.M., Barnabé, C., Tibayrenc, M., Tafuri, W.L., 2002. Impact of
187

Publications
Trypanosoma cruzi clonai evolution on its biological properties in mice.
Experimental Parasitology 100,161-172.
Toledo, M.J., Bahia, M.T., Carneiro, CM, Martins-Filho, O.A., Tibayrenc, M., Barnabé,
C., Tafuri, W.L., de Lana M., 2003. Chemotherapy with Benznidazole and
Itraconazole for mice infected with different Trypanosoma cruzi clonai genotypes.
Antimicrobial Agents and Chemotherapy 47, 223-230.
Ullman, B., 1995. Multidrug resistance and P-glycoproteins in parasitic protozoa.
Journal of Bioenergetics and Biomembranes 27, 77-84.
Upcroft, J.A, Upcroft, P., 1993. Drug resistance and Giardia. Parasitology Today 9,
187-190.
Veloso, -V.M., Carneiro, C.M., Toledo, M.J.O., de Lana, M., Chiari, E., Tafuri, W.L.,
Bahia, M.T., 2001. Variation in susceptibility to Benznidazole in iso/ates derived
from Trypanosoma cruzi parental strains. Mem6rias do Instituto Oswaldo Cruz 96,
1005-1011.
Vergnes, B., Sereno, D., lVIadjidian-Sereno, N., Lemesre, J-L., Ouaissi, A, 2002.
Cytoplasmic SIR2 homologue overexpression promotes survival of Leishmania
parasites by preventing programmed cell death. Gene 296, 139-150.
Wilson, C.M., Serrano, AE., Wasley, A, Bogenschutz, M.P., Shankar, A.H., Wirth,
D.F., 1989. Amplification of a gene related to mammalian mdr genes in drug
resistant Plasmodium fa/ciparum. Science 244, 1184-1186.
WHO-Wor/d Health Organization, 1991. Control of Chagas Disease. Technical
Report No.811. WHO, Geneva, Switzerland. pp. 150.
188
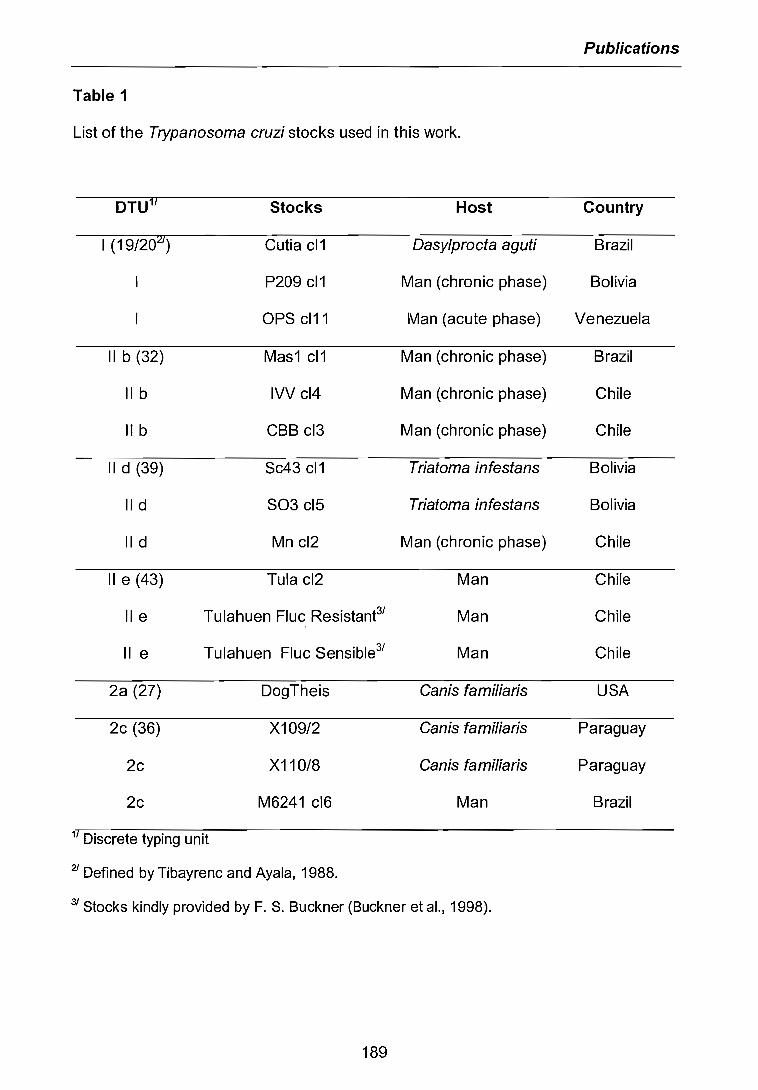
Table 1
List of the Trypanosoma cruzi stocks used in this work.
Publications
DTU 17 Stocks Host Country
1 (19/2027) Cutia c11 Dasylprocta aguti Brazil
P209 c11 Man (chronic phase) Bolivia
OPS c111 Man (acute phase) Venezuela
Il b (32) Mas1 c11 Man (chronic phase) Brazil
Il b IVV c14 Man (chronic phase) Chile
Il b CSB c13 Man (chronic phase) Chile
Il d (39) Sc43 cl1 Triatoma infestans Bolivia
Il d S03 c15 Triatoma infestans Bolivia
IId Mn cl2 Man (chronic phase) Chile
Il e (43) Tula c12 Man Chile
Ile Tulahuen Fluc Resistant3/ Man Chile
Il e Tulahuen Fluc Sensible3/ Man Chile
2a (27) DogTheis Canis familiaris USA
2c (36) X109/2 Canis familiaris Paraguay
2c X110/8 Canis familiaris Paraguay
2c M6241 cl6 Man Brazil
11 Discrete typing unit
2J Defined by Tibayrenc and Ayala, 1988.
31 Stocks kindly provided by F. S. Buckner (Buckner et aL, 1998).
189
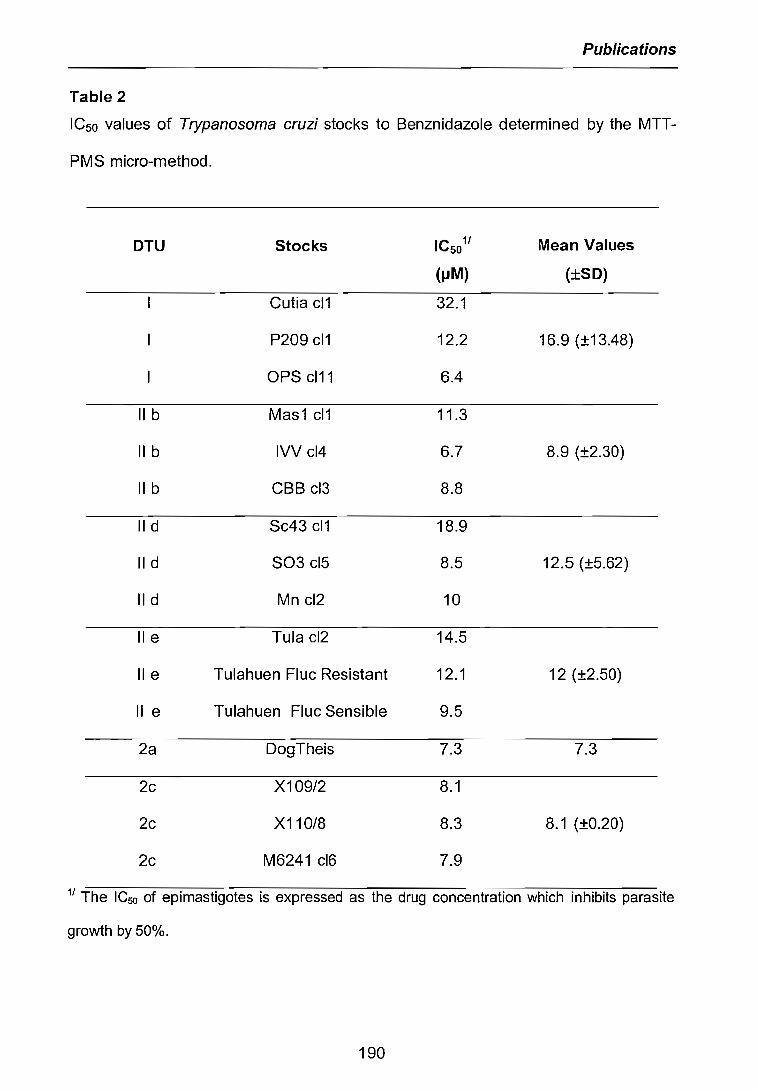
Publications
Table 2
IC50 values of Trypanosoma cruzi stocks to Senznidazole determined by the MTT-
PMS micro-method.
DTU Stocks ICso11 Mean Values
(~M) (±SD)
Cutia c11 32.1
P209 c11 12.2 16.9 (±13.48)
OPS c111 6.4
Il b Mas1 c11 11.3
Il b IVV c14 6.7 8.9 (±2.30)
Il b CSS c13 8.8
IId Sc43 c11 18.9
IId S03 c15 8.5 12.5 (±5.62)
IId Mn c12 10
Ile Tula c12 14.5
Ile Tulahuen Fluc Resistant 12.1 12 (±2.50)
Il e Tulahuen Fluc Sensible 9.5
2a DogTheis 7.3 7.3
2c X109/2 8.1
2c X110/8 8.3 8.1 (±0.20)
2c M6241 c16 7.9
11 The IC50 of epimastigotes is expressed as the drug concentration which inhibits parasite
growth by 50%.
190

Publications
Table 3
Correlation matrix determinate by a Mantel test between genetic distances (RAPD
and MLEE) and susceptibility to Benznidazole (IC5D values).
ICso
RAPD
MLEE
ICso
r = 0.101 P= 0.229
r = 0.201 P= 0.158
RAPD
r=0.729 P<10-4
MLEE
Table 4
IC5D values for three stocks determinated by MTT-PMS, flow fluorocytometry and
[3H]-Thymidine methods.
ICso (J,lM)
DTU Stock MTT-PMS Flow cytometry eH]-Thymidine
method method method!1
3 6.4 7.8 6.7
Il b 6 8.8 10.2 1.2
IId 9 10 10.3 1.9
1fdata fram Revollo, 1995.
191
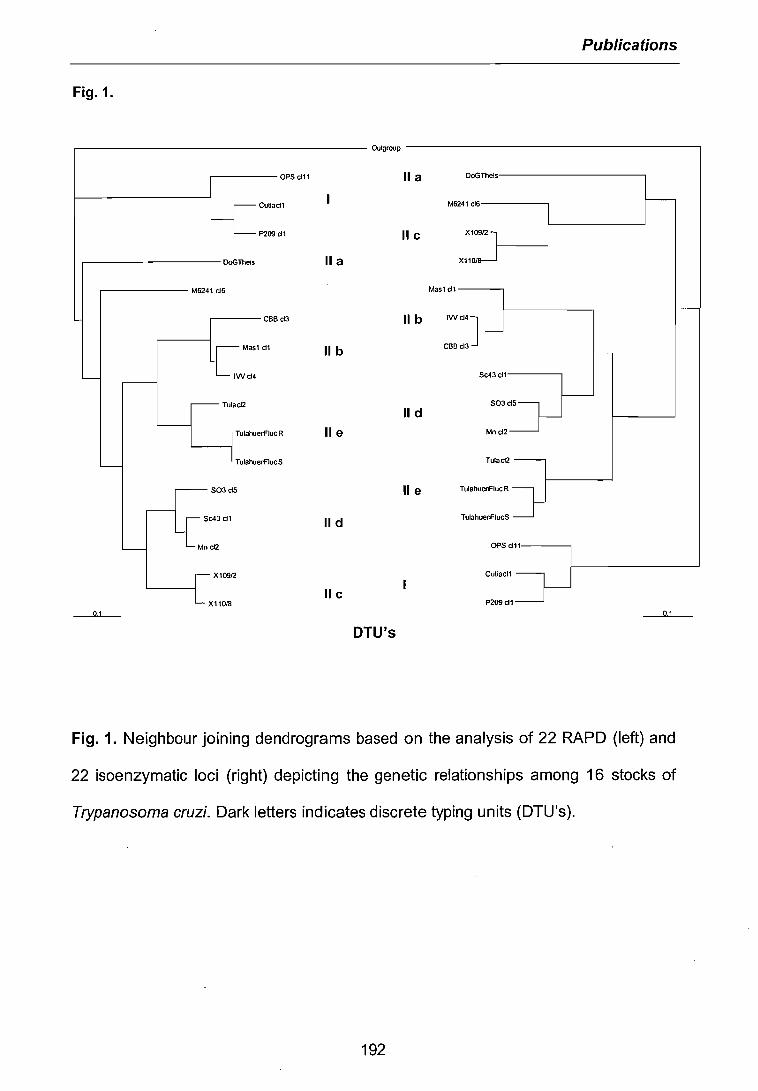
Publications
Fig.1.
Oulgroup
OPSdl1 lia DoGTheis
Culiadl M6241 cl6
P209 dl Ile Xl 0912
DoGTheis lia Xll018
MasldlM6241 d6
CBBd3 lib IWd4
Masl dl lib CBB d3
IWd4 Sc43 dl
Tulad2IId
TulahuerFJuc R Ile Mnd2
TulahuerFlucS Tulad2
S03d5 Ile Tulahued'lucR ]JSc43 dl IId
Tulahued'lucS
Mnd2 OPS dll
X 10912 Cutiacll
IleXll01B P209cll
01 01
DTU's
Fig. 1. Neighbour joining dendrograms based on the analysis of 22 RAPD (Ieft) and
22 isoenzymatic loci (right) depicting the genetic relationships among 16 stocks of
Trypanosoma cruzi. Dark letters indicates discrete typing units (DTU's).
192

TITRE en françaisEtude de l'expression différentielle des gènes impliqués dans la résistance au Benznidazole chezTrypanosoma cruzi, l'agent de la maladie de Chagas.RESUME en français
La maladie de Chagas ou trypanosomose américaine est provoquée par le parasiteTrypanosoma cruzi. Actuellement le traitement de cette maladie se limite à deux médicaments, leBenznidazole et le Nifurtimox. Des effets secondaires importants ainsi qu'une faible efficacité ontété rapportés chez les patients. Les variations du taux de guérison observées semblent être uneconséquence des différences de sensibilité des parasites aux médicaments. Il a été suggérél'existence d'un lien entre diversité génétique du parasite et comportement des souches face auxmédicaments. Plusieurs études de notre équipe ont montré un impact important de la structureclonale de T. cruzi sur la sensibilité aux drogues de ce parasite. L'objectif de cette étude a été decontinuer d'explorer ce résultat, et d'analyser l'expression différentielle des gènes grâce à unetechnique dérivée du RADES, sur un échantillon de stocks représentatif de T. cruzi, vis à vis deleur comportement face au Benznidazole ; ceci par deux approches: i) l'étude des stocks exposésà des concentrations de Benznidazole équivalentes au niveau de sensibilité naturelle (IC50), et ii)l'étude des stocks devenus résistants par pression médicamenteuse. L'expression génétique desstocks sauvages et exposés au Benznidazole à la dose IC50 , a été comparée à la structuregénétique des populations naturelles du parasite. Des bandes différentielles impliquées dans larésistance au Benznidazole ont été identifiées et les gènes correspondants ont été recherchés.Les gènes préalablement décrits comme potentiellement impliqués dans les mécanismes derésistance n'ont pas montré de différence d'expression. De plus, nous n'avons pas trouvé decorrélation significative entre le groupe génétique du parasite et sa sensibilité au Benznidazole,pas plus qu'entre les bandes différentielles et les groupes génétiques au sein de l'espèce T. cruzi.
TITRE en anglaisStudy of the differential expression of genes involved in Benznidazole resistance in Trypanosomacruzi, the agent of Chagas disease.RESUME en anglais. Chagas disease or American Trypanosomiasis is caused by Trypanosoma cruzi. Thetreatment of the disease is currently limited to two drugs: Benznidazole and Nifurtimox. Lowefficacy as weil as adverse reactions have been currently observed in patients. Variability of curerates might be due to differences in T. cruzi drug susceptibility. It has been suggested thepossibility of relationships between parasite genetic diversity and reactions of the strains withregards to drugs. Several studies from our team have shown a noticeable impact of T. cruzi clonaipopulation structure on this parasite's drug sensitivity. The aim of this study was to further explorethis result and to analyse the differential gene expression, by using a derived RADES technique,among a representative sample of T. cruzi stocks, in relation to Benznidazole exposures. Studieswere carried out both, i) at the natural susceptibility level (IC50), and (ii) at different resistancelevels (in resistant stocks induced by continuous drug pressure). Differentiai gene expression ofwild stocks and stocks submitted to Benznidazole exposure at the IC50 level, was compared to thegenetic structure of this parasite's natural populations. Differentiai bands involved in Benznidazoleresistance were identified and the corresponding genes were investigated. Genes previouslydescribed as probably involved in drug resistance mechanisms did not shown any differentialexpression. Furthermore, no significant correlation between genetic clustering and Benznidazolesusceptibility was found. A similar lack of relationship was also observed betwèen differentiallyexpressed bands and genetic groups within the species T. cruzi.
DISCIPLINEBiochimie et Biologie Moléculaire.MOTS-CLESMaladie de Chagas, Trypanosoma cruzi, Résistance, Benznidazole, ARNm, Expressiondifférentielle, RADES, RAPD, MLEE, Diversité phylogénétique.INTITULE ET ADRESSE DU LABORATOIRECentre de Recherche IRD (Institut de Recherche pour le Développement), UMR CNRS-IRD 9926,Génétique des Maladies Infectieuses GMI (Ex-CEPM), 911 avenue Agropolis, B.P. 64501, 34394Montpellier Cedex 5.





























