Embed Size (px)
Citation preview

Nutr. Clin. M6tabol. 1995 ; 9 : 123-128
Etude des flux d'acides amin6s h l'6tat stationnaire dans le foie de rat isol6, perfus6
J e a n - P a s c a l De Band t 1, Co le t t e Rey 2, E l i sabe th Lasn ie r 1, 2 Raoul Poupon 2, J a c q u e l i n e G i b o u d e a u 1, Luc C y n o b e r 1, 2 '
1. Laboratoire de Biochimie A, CHU Saint-Antoine, Paris. 2. INSERM U 402, CHU Saint-Antoine, Paris.
R 6 s u m 6
Le foie de rat isol6 et perfus6 en recirculation est un module couramment utilis6 pour l'6tude des 6changes h6patiques d'acides amin6s (AA). Toutefois, la recir- culation entraine une 6volution des concentrations de substrats dans le milieu de perfusion en fonction du temps et ne permet pas d 'a t te indre un 6tat stationl naire. Le but de ce travail 6tait de d6finir des condi- tions d 'appor t en AA permettant de maintenir un 6tat stationnaire tout au long de la perfusion. Deux s6ries de foies ont 6t6 perfus6es : d a n s la pre- mibre (groupe t6moin), les AA ont 6t6 apport6s sous la forme d ' u n bolus unique ; d a n s la seconde, (groupe 6tat stationnaire) les AA ont 6t6 adminis- tr6s en bolus suivi d 'une perfusion d ' A A h vitesse constante pour compenser leur captation h6patique. Dans le groupe t6moin, on observe une r6duct ion significative de la concentration de la majorit6 des AA h l ' except ion du glutamate et de l 'orni th ine , lib~r~s par le foie, et des AA ~ chaine ramifi~e. Ceci s 'accompagne avec le temps d 'une r6duction signifi- cative des 6changes h6patiques de glycine (- 68 %), de proline (- 47 %), d 'alanine (- 46 %) et de gluta- mine (- 37 %). En revanche, la captation d 'arginine est prat iquement doubl6e en fin de perfusion. L ' a p p o r t d ' A A en perfusion continue permet d 'obtenir une stabilit6 des concentrations et des flux de la p lupa r t des AA. Ceci n ' e s t pas le cas pour l ' a rginine et la glutamine sans doute en raison de l 'effet induc teur de ces deux AA sur leur p ropre m6tabolisme.
Mots-cles : Foie isole perfus~, acides amin6s, 6tat station- naire.
Correspondance : L. Cynober, H6pital Saint-Antoine, Laboratoire de Biochimie A, 75571 Paris cedex 12. Travail en pattie financ~ par une bourse ESPEN-Ajinomoto et present6 au XIII e Symposium de la SFNEP, Marseille, decembre 1993. Regu le 30 septembre 1994, accepte apres revision le 2 mai 1995.
123

J.P. D E B A N D T
Introduction
Le foie de rat isol6 perfus~ est un module largement employ6 pour l '6 tude des fonct ions h6patiques [1]. Toutefois, les conditions exp6rimentales ddcrites dans la litt6rature sont extr~mement variables rant au point de vue de la nature du milieu de perfusion (pr6sence ou non de globules rouges ; apport variable en substrats : acides aminds, glucides . . . ) que du syst~me utilis6 (simple passage ou recirculation) [1]. Nous avons pr6cddemment d6fini, pour un syst~me en recirculation, des conditions de perfusion permettant d 'obtenir des 6changes h6patiques d 'acides amin6s se rapprochant de ceux observ6s in v ivo [2-4]. L'uti l isa- tion d ' un syst~me en recirculation est ndcessaire h la mesure pr6cise des 6changes hdpatiques. En effet, ceux-ci sont calculds, lors d 'une perfusion en simple passage, h partir des diff6rences de concentration entre veine porte et veines sus-h6patiques [5], ou, lors de perfusions en recirculation, ~ partir des variations de concentrat ion darts le milieu de perfusion [6]. Pour l '6tude de substrats 5 faible extraction comme les AA, la perfusion en recirculat ion amplifie les manifesta- tions du mdtabolisme h6patique et permet de mesurer plus commod6ment les 6changes tissulaires. Toutefois, un tel systbme, en entratnant une rdduction progressive de la concentration des AA dans le perfusat en raison de leur captation hdpatique, ne permet pas de maintenir l '6tat stationnaire. I1 en rdsulte une diminution progres- sive des 6changes h6patiques [7]. Dans le cadre de l '6tude du m6tabolisme h6patique, il devient alors dif- ficile de faire la part entre les modif icat ions intrin- sbques du m6tabolisme et celles induites par la simple variation des concentrations circulantes. Le but de notre travail a 6t6 de d6finir des conditions d' apport en AA permettant de maintenir un 6tat station- naire des concentrations et des 6changes. La m6thode retenue a 6t6 la technique du << clamp ~, c 'est-~-dire 1' apport d ' A A ~ une vitesse compensant leur utilisation.
Mat6riel et m~thodes
M6thode de perfusion
Nous avons utilis6 des rats males Sprague-Dawley (Charles Rivers, Saint-Aubin-l~s-Elbeuf, France) de 220 + 17 g, nourris a d l i b i t u m (UAR A03, UAR, Vi l lemoisson-sur-Orge, France), adapt6s depuis au moins 7 jours au laboratoire en Cycle normal. Les animaux ont 6t6 mis ~ jeun la veille des exp6riences, avec libre accbs ~ l 'eau. Les foies, pr6pards selon la technique pr6c6demment ddcrite [2], ont 6t6 perfus6s par la veine porte, en recir- culation et ~ pression constante dans une enceinte ther- mostat6e ~ 37 °C. Nous avons utilis6, comme milieu de perfusion (volume 150 ml), un tampon Krebs-Ringer additionn6 d 'albumine (30 g/1 ; non d61ipidde, fraction
V, Calbiochem), de glucose (7,5 mmol/1) et de calcium (2 mmol/1) et contenant un m61ange d ' A A r6gulateurs de la prot6olyse (alanine, glutamine, leucine, ph6nyl- alanine, t ryptophane, mdthionine, proline, histidine) [8]. Le milieu de perfusion a 6t6 oxygdn6 par un m61ange 95 % 02/5 % CO 2 et le pH maintenu ~ 7,40.
Sch6ma experimental
Nous avons r6alis6 deux s6ries d 'exp6r iences (n = 6 dans chaque groupe) : un groupe t6moin recevant h t30 un bolus d 'une solution concentr6e d ' A A [7] et un groupe << 6tat stationnaire >> recevant le mSme bolus suivi d 'une perfusion continue d ' A A (0,2 ml/min) (tableau I). Les caract6ristiques de cette perfusion (composition et vitesse) ont 6t6 d6termin6es 5 partir des valeurs de captation mesur6es ~tans le groupe t6moin. Au cours de la perfusion, le d6bit portal, le d6bit biliaire et le pH ont 6t6 mesur6s toutes les 10-15 minutes. Des 6chantillons de perfusat ont 6t6 pr61ev6s aux temps 35, 45, 60, 75 et 90 minutes. Les roles ont 6t6 congel6s dans l 'azote liquide en fin de perfusion. Le sch6ma expdrimental est r6sum6 dans la figure 1.
Param~tres 6tudi6s
Les pr61~vements ont 6t6 conserv6s h - 20 °C jusqu 'au moment du dosage. Pour la mesure des AA, les 6chan-
Tab leau I : A p p o r t s d ' A A dans les d e u x sdr ies e x p d r i m e n t a l e s
Quantitd apport~e ~ t30 Vitesse de perfusion ~lmol/150 ml nmol/min/g*
ASP 25 9 THR 50 35 SER 25 29 ASN 25 34 GLU 25 0 GLN 128 129 PRO 25 63 GLY 25 32 ALA 25 107 VAL 50 20 CYS 25 10 MET 13 22 ILE 50 17 LEU 50 30 TYR 25 4 PHE 25 52 TRP 5 2 LYS 63 37 HIS 13 25 ARG 30 22 AAT ° 701 679
o : quantitd totale d'AA apportde. * : dans le groupe dtat stationnaire uniquement.
124

FLUX Ht~PATIQUES D ' A C I D E S AMIN[~S ~k L' t~TAT S T A T I O N N A I R E
Groupe t6moin
Groupe 6tatstationnaire
0 30 45 60 75
Acides amines Bolus d'acides antiprot6olytiques amin6s
0 30 45 60 75
Figure 1 : Sch6ma expdrimental. P : prdlkvement .
tillons ont 6t6 d6prot6inisds par l ' ac ide sulfosal icyl ique (50 mg/ml). Nous avons mesur6 les parambtres suivants : - AA par chromatographie d '6change d ' ions sur analy- seur B io t ron ic LC 5001 (Bio t ron ik , Munich , Al lemagne) ; - ur6e et g lucose par des m6thodes h l ' u r6ase et ~ la g lucose oxydase , automat is6es sur ana lyseur As t ra -8 Beckman (Beckman, Palo Alto, USA) ; - ammoniaque par la m6thode ~ la glutamate d6shydro- g6nase au tomat i sde sur ana lyseur Synchron CX-4 (Beckman) ; - activit6s aspartate aminotransf6rase (ASAT), alanine amino t rans f6 rase ( A L A T ) et lac ta te d6shydrog6nase (LDH), se lon les r e c o m m a n d a t i o n s de la Soci6t6 Fran~aise de B io log ie Cl in ique , sur ana lyseur Synchron CX-4. Les foies ont 6t6 pes6s en fin de perfusion, le poids sec et le contenu en eau ont 6t6 d6terminds apr~s dessicca- tion pendant 12 h ~ 80 °C.
M6thodes de calcul
Les 6changes h6patiques de substrats (F) ont 6t6 calcu- 16s ~ l ' a ide des formules suivantes : - d a n s le groupe t6moin :
F = Ct2" V 2 - Ch "V]
(t 2 - t l ) • p - d a n s le groupe 6tat stationnaire :
F = Ct2" V2 - Ct] • V 1 (t2 _ t l ) • P - Vp
o~ Ct 1 et Ct 2 sont les concentra t ions d ' u n m6tabol i te dans le milieu de perfusion aux temps t 1 et t 2 respective- ment, V 1 et V 2, le volume du mil ieu de perfusion aux m~mes temps (afin de tenir compte des variations li6es aux pr61~vements, h l '6vaporat ion et, darts le groupe 6tat stationnaire, ~ la perfusion continue d 'ac ides amin6s) ; P l e poids de foie frais, et Vp la vi tesse de per fus ion pour I ' A A consid6r6. Une valeur n6gative de F traduit une captation nette, et une valeur positive une lib6ration.
9O
P
9O
Perfusion continue d'acides amines
Statistiques
Les r6sultats sont pr6sent6s sous la forme moyenne _+ 6cart-type. Les r6sultats ont 6t6 analys6s au moyen de tests non param6tr iques : test T de W i l c o x o n pour s6ries appari6es et test U de Mann-Whi tney pour s6ries non appari6es, ~t l ' a i d e du log ic ie l PSCM (Deltasoft , Grenoble, France). Les diff6rences ont 6t6 consid6r6es comme s ignif ica t ives pour une valeur de p inf6rieure ou 6gale ~ 5 %.
Resul ta ts
La viabili t6 h6patique, estim6e par la mesure des acti- vit6s ASAT, A L A T et LDH et par le d6bit portal, et le ddbit bi l iaire sont comparables entre les deux s6ries de perfusion (tableau II). Dans le groupe t6moin, les concentrat ions de la majo- rit6 des A A ( tab leau III) d6cro i s sen t r6gul ibrement avec le temps, except6 ce l les des ac ides amin6s cha~ne ramifide (AACR) qui restent stables et celles du
Tableau H : Inf luence de l ' apport d ' A A sur la viabili t6 hdpatique
T~moin Etat stationnaire (n = 6) (n = 6)
ASAT (UI/1)* 63 _+ 37 59 _+ 12 ALAT (UI/1)* 28 _+ 22 55 _+ 33 LDH (UU1)* 153 + 149 110 _+ 70 D6bit portal (ml/min/g de foie frais) 3,44 + 0,26 3,85 + 0,35 D6bit biliaire (pl/min/g de foie frais) 0,477 _+ 0,069 0,497 _+ 0,102 Contenu en eau (%) 74,6 _+ 1,5 74,2 + 2,2
Les r~sultats sont prdsent~s sous la forme moyenne + dcart- type. * activitds enzymatiques mesur~es gz t90
125
J.P. D E B A N D T
g l u t a m a t e et de l ' o r n i t h i n e q u i a u g m e n t e n t . C e c i s ' a c c o m p a g n e d ' u n e r6duc t i on p r o g r e s s i v e de la cap ta - t i on g l o b a l e de s A A ( f i g u r e 2), s i g n i f i c a t i v e p o u r la g l u t a m i n e , l ' a l a n i n e , la g l y c i n e et la p r o l i n e ( t a b l e a u IV). Pa r a i l leurs , la c a p t a t i o n d ' a r g i n i n e et la l i bd ra t ion d ' o r n i t h i n e s o n t s i g n i f i c a t i v e m e n t a u g m e n t d e s en f in d ' e x p 6 r i e n c e ( t ab l eau IV) .
Tableau I I I : Variat ions des concentrat ions d ' A A dans
le mil ieu de per fus ion au cours du temps.
T6moin Etat stationnaire (n = 6) (n = 6)
t35 t90 t35 t90
ASP 146_+10 1 3 0 _ 1 4 * 192_+l l 183_+15 THR 307 _+ 26 256 _ 38 * 337 + 26 361 _+ 22 SER 163 _+ 14 96 _+ 24 ** 190 + 22 198 _+ 38 ASN 144_+ 14 62_+ 17 ** 118_+ 11 143_+20 * GLU 254 _+ 28 452 _+ 32 ** 290 _+ 23 449 + 25 ** GLN 781_+97 5 1 9 + 9 1 ** 1195_+104 848_+224** PRO 340 _+ 21 233 _+ 51 * 350 _+ 23 388 _+ 36 GLY 210 _+ 37 168 _+ 41 ** 230 _+ 20 257 _+ 33 ALA 478_+91 258_+ 101"* 551_+34 556_+76 VAL 301_+23 295_+23 315_+ 11 341_+31 CYS 48_+5 3 6 + 5 ** 55_+4 60_+8 MET 127 _+ 10 72 _+ 16 ** 137 _+ 6 131 -+ 16 ILE 294_+26 291_+ 19 317_+ 19 337-+20 LEU 560 _+ 39 556 -+ 39 550 _+ 24 581 _+ 35 TYR 157 _+ 15 132 _+ 13 * 162 _+ 6 170 _+ 18 PHE 199_+20 101_+27 ** 223_+10 215_+34 ORN 10_+4 61_+39 * 7 + 3 7 1 + 3 5 ** LYS 328 -+ 23 254 _+ 36 ** 341 -+ 22 359 _+ 36 HIS 165_+32 98_+32 ** 160_+5 150-+25 ARG 153 _+ 23 57 _+ 34 ** 151 _+ 12 87 _+ 26 ** AAT 5197_+ 407 4167_+ 416"* 5847_+ 268 5869_+ 475
Les rdsultats (moyenne -+ dcart-type) sont exprimds en pmol/l. * p < 0,05 et ** p < tgo vs t35 (test T de Wilcoxon). AAT : concentration globale des acides amines.
D a n s le g r o u p e << 6ta t s t a t i o n n a i r e >~, les c o n c e n t r a t i o n s des A A res ten t , d a n s l ' e n s e m b l e , s t ab les ( t ab l eau l l I ) . Le s c o n c e n t r a t i o n s de g l u t a m a t e et d ' o r n i t h i n e a u g - m e n t e n t e n r a i s o n de l e u r p r o d u c t i o n p a r le f o i e ( t a b l e a u IV) . E n r e v a n c h e , m a l g r 6 la p e r f u s i o n con t i - nue , o n o b s e r v e u n e d i m i n u t i o n s i g n i f i c a t i v e de la c o n c e n t r a t i o n de g l u t a m i n e et d ' a r g i n i n e ( t ab l eau II i) . E n t e r m e s de f l u x h 6 p a t i q u e , il e x i s t e u n e r d d u c t i o n s i gn i f i c a t i ve de la c a p t a t i o n de g l u t a m i n e avec le t e m p s et u n e a u g m e n t a t i o n s i g n i f i c a t i v e de la c a p t a t i o n d ' a r g i n i n e et de la l i b6 ra t i on d ' o r n i t h i n e ( t ab l eau IV). L a c o m p a r a i s o n e n t r e les d e u x g r o u p e s ( t a b l e a u I V ) i nd ique que les f lux de g l u t a m a t e et de g l u t a m i n e sont s i g n i f i c a t i v e m e n t p lu s f a i b l e s d a n s le g r o u p e t 6 m o i n tou t au l o n g de la p e r f u s i o n , t and i s que p o u r de n o m - b r e u x A A la r 6 d u c t i o n de la c ap t a t i on ne d e v i e n t s igni - f i c a t i v e q u ' e n f in d ' e x p 6 r i e n c e . P a r a l l ~ l e m e n t , e n f in
Tableau I V : Variat ions des f l u x d ' A A
en fonc t ion du temps
T6moin Etat stationnaire (n = 6) (n = 6)
p6riode p6riode p~riode p~riode initiale finale initiale finale t35 - t45 t75 - t90 t35 - t45 t75 - t90
ASP -6_+7 -7_+5 - 1 0 + - 7 -11_+7 THR -21_+10 -12.+ 10 -18-+14 - t 9 . + 1 6 SER - 2 1 e 8 - 1 5 + 3 -20-+10 -22 .+10 ASN 2 1 e l 0 -,17_+9 -16_+16 -20_+10 GLU 27 + 16 44 .+ 21 66 _+ 19 ££ 83 + 16 £ GLN -85_+35 -52_+26 * -238_+70 ££ - 1 6 3 + 7 2 **££ PRO -49_+28 -26_+11 * ~-35_+26 - 3 7 + 1 4 GLY -22_+4 - 7 . + 5 * " - 3 0 + 1 8 - 1 3 + 1 2 ALA - 6 9 + 2 5 - 3 7 + 7 * -88_+32 -79_+18 ££ VAL - 1 4 + 1 3 - 2 . + 5 -11 .+12 -10_+12 CYS -7_+3 -3_+2 -10_+4 - 5 . + 3 MET - 1 5 + 6 -11_+5 - 1 6 + 8 -20_+5 ££ ILE -11__.14 - 2 . + 3 - 4 + 1 5 -7 .+12 LEU - 2 0 + 2 6 -5_+11 - 6 . + 1 6 -11_+24 TYR -3_+8 -11_+ l l 2_+8 -1_+7 PHE -34_+ 14 -16_+6 -37_+ 19 - 4 4 + 9 ORN 3_+2 22.+11 * 5.+4 45.+35 * LYS - 2 4 + 1 2 -16_+12 -21_+16 -26_+13 HIS - 1 4 + 1 1 - 1 8 . + 7 -16_+13 -30_+11 £ ARG -16_+9 -32__.17 * 19_+13 - 7 3 + 5 0 * AAT -414_+175 -223_+88 * -517 .+212 -449_+229 £
Les rdsultats (moyenne -+ dcart-type) sont exprimds en nmol/min/g de foie frais. Les valeurs ndgatives correspondent g~ une captation et les valeurs positives & une libdration. AAT : flux global des acides aminis. * p < 0,05 et ** p < 0,01 t75 - t90 vs t35 - t45 (test T de Wilcoxon). £ p < 0,05 et ££p < 0,01 dtat stationnaire vs tdmoin (test U de Mann- Whitney).
d ' e x p 6 r i e n c e , la p r o d u c t i o n d ' u r 6 e e s t s i g n i f i c a t i v e - m e n t p lus i m p o r t a n t e d a n s le g r o u p e 6ta t s t a t i o n n a i r e que dans le g r o u p e t 6 m o i n ( r e s p e c t i v e m e n t 385 -+ 103 n m o l / m i n / g et 235 + 153 n m o l / m i n / g , p = 0 ,02) . E n r e v a n c h e , les f lux de g lucose ( - 43 _+ 76 n m o l / m i n / g et - 78 _+ 139 n m o l / m i n / g t dmo in ) et d ' a m m o n i a q u e ( - 8 -+ 9 n m o l / m i n / g e t - 19 _+ 21 n m o l / m i n / g t 6 m o i n ) n e son t pas d i f f6 ren t s en t re les deux g roupes .
Discussion
En s i tua t ion n o n s t a t ionna i re , il ex i s t e lors de p e r f u s i o n en r e c i r c u l a t i o n u n e d i m i n u t i o n p a r a l l ~ l e des c o n c e n - t r a t i ons et des 6 c h a n g e s h d p a t i q u e s d ' A A qui r e n d dif- f i c i l e l ' i n t e r p r d t a t i o n de s r d s u l t a t s d ' 6 c h a n g e s h6pa t iques . Nos rdsu l ta t s i n d i q u e n t q u ' u n 6tat s t a t ion-
126
FLUX HI~PATIQUES D'ACIDES AMINI~S A L'I~TAT STATIONNAIRE
6000
:i 5000, m 4000-
3000
2000
0 lOO0
• 0 ~ 30
i - 1 0 0 - O
"~ - 3 5 0 -
,~ - 6 0 0 -
~ -850
[] Etat d'dquilibre
• Tdmoin
I I ! I I I
40 50 60 70 80 90 T e m p s (rain.)
B [] Tdmoin £
[] Etat d'dquilibre
~ , 3 5 - 4 5 4 5 - 6 0 6 0 - 7 5 o~
0 Pdriodes de m e s u r e
7 5 - 9 0
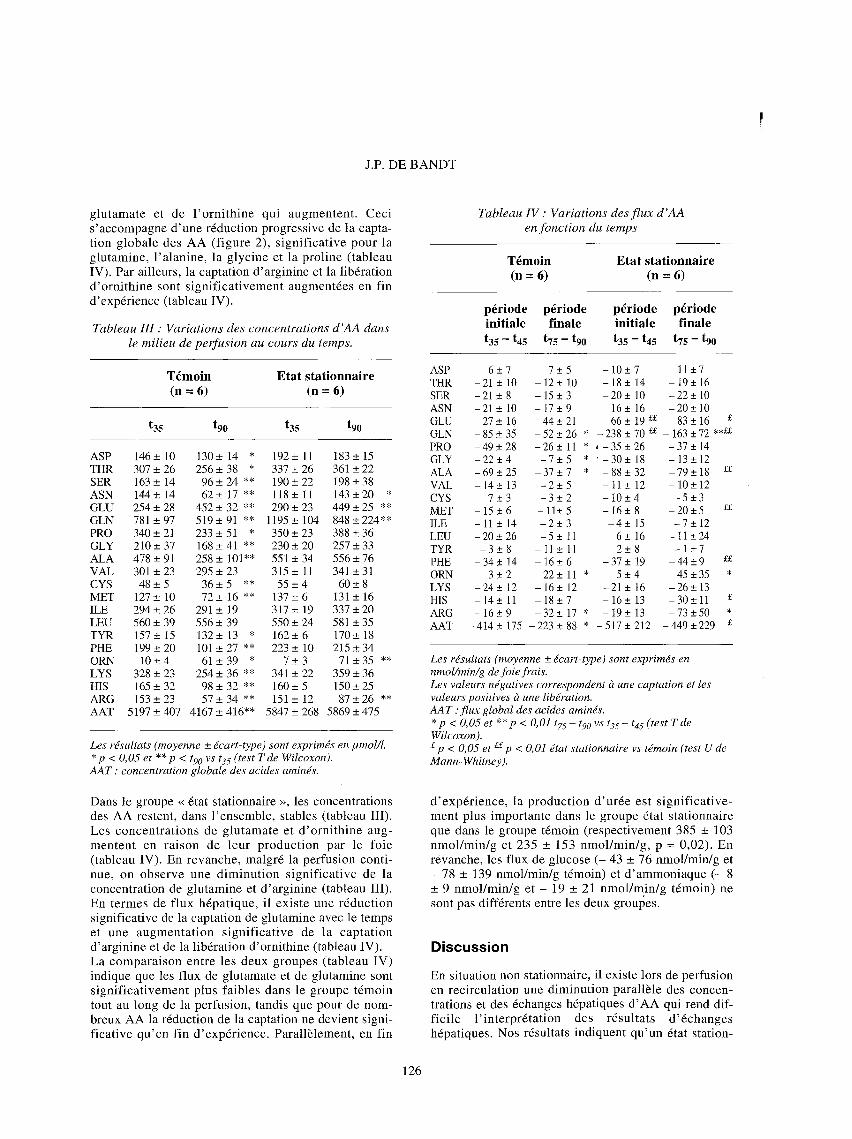
Figure 2 : Influence de l ' apport d 'AA sur le f lux global des AA en fonct ion du temps. A : variations de la concentration totale clans le perfu- sat : **p< 0,01 tgo vs t35. B : variations de la captation globale : *p< 0,05 t75 - t90 vs t35 - t45 ; £ p< 0,05 groupe dtat stationnaire vs groupe tdmoin.
naire peut ~tre obtenu pour la plupart des AA dans des conditions de << clamp aminoaciddmique >>. Les diffdrences d'dchanges h6patiques observdes entre les deux groupes ne sont pas lides h des altdrations de la viabilitd hdpatique. Le faible ddbit biliaire observ6 dans les deux groupes, compard aux valeurs obtenues lors d'exp6riences en l 'absence d 'AA (de l 'ordre de 0,95 pl/min/g), est imputable ~t l 'effet cholestatique connu des AA [9]. Darts le groupe tdmoin, la rdduction de la concentration totale en AA (- 20 %) traduit une diminution significa- tive de la concentration de la majorit6 des AA l'exception du glutamate et de l'ornithine, libdrds par le
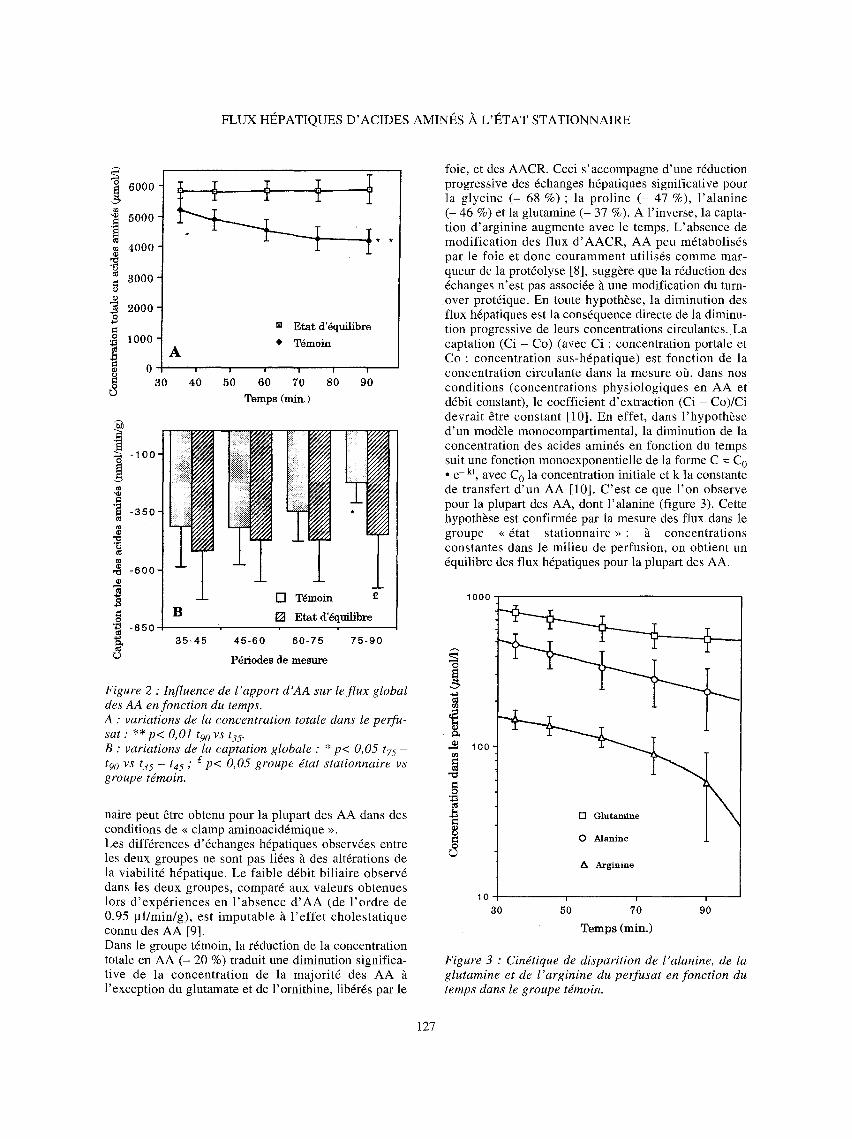
foie, et des AACR. Ceci s'accompagne d'une r6duction progressive des 6changes hdpatiques significative pour la glycine (- 68 %) ; la proline (- 47 %), l 'alanine (- 46 %) et la glutamine (- 37 %). A l'inverse, la capta- tion d'arginine augmente avec le temps. L'absence de modification des flux d 'AACR, AA peu m6tabolisds par le foie et done couramment utilis6s comme mar- queur de la prot6olyse [8], suggbre que la r6duction des 6changes n'est pas associ6e h une modification du turn- over prot6ique. En toute hypothbse, la diminution des flux h6patiques est la cons6quence directe de la diminu- tion progressive de leurs concentrations circulantes. La captation (Ci - Co) (avec Ci : concentration portale et Co : concentration sus-hdpatique) est fonction de la concentration circulante dans la mesure off, dans nos conditions (concentrations physiologiques en AA et d6bit constant), le coefficient d'extraction (Ci - Co)/Ci devrait ~tre constant [10]. En effet, dans l 'hypothbse d'un modNe monocompartimental, la diminution de la concentration des acides aminds en fonction du temps suit une fonction monoexponentielle de la forme C = C o • e - k t , avec C o la concentration initiale et k la constante de transfert d 'un AA [10]. C'est ce que l 'on observe pour la plupart des AA, dont l'alanine (figure 3). Cette hypothbse est confirm6e par la mesure des flux dans le groupe << 6tat stationnaire >> : h concentrations constantes dans le milieu de perfusion, on obtient un dquilibre des flux hdpatiques pour la plupart des AA.
0
{D
o o
1 0 0 0
100
Glutamine
0 Alanine
A Arginine
1 0 1 ' , , ,
30 50 70 90
T e m p s (min. )
Figure 3 : Cindtique de disparition de l'alanine, de la glutamine et de l 'arginine du per fusat en fonct ion du temps dans le groupe tdmoin.
127

J.P. D E B A N D T
En revanche , dans les deux groupes , l ' 6qu i l i b r e des concentrat ions et des flux n ' e s t obtenu ni pour l ' a rg i - nine ni pour la glutamine : - Dans le cas de la glutamine, la captation (tableau IV) et, pour le g roupe t6moin, la pente de la courbe de d6cro i s sance de la concen t ra t ion darts le per fusa t (figure 3) diminuent avec le temps. Ceci doit 6tre inter- prdt6 en tenant compte des particulari t6s de la r6gula- t ion du m6tabo l i sme h6pat ique de la g lu tamine [11]. Les t ravaux de Sies et Hauss inger [12] indiquent une adaptat ion du flux hdpatique de glutamine en fonction de la concent ra t ion por ta le : pour un apport 61ev6, la captation de cet acide amin6 augmente et inversement. Ainsi , darts l e groupe tdmoin, la consommat ion hdpa- tique de glutamine, en rdduisant la concentration circu- lante de cet AA, est 5 l 'o r ig ine de la r6duction du flux en fonction du temps. A l ' inverse , darts le groupe 6tat s ta t ionnai re , l ' a p p o r t accru de cet A A entrMne une augmenta t ion s igni f ica t ive de la capta t ion pendant le ddbut de la p6r iode d ' 6 t u d e (t35 - t45 ). Toutefo is , la vitesse d ' appor t de glutamine par la perfusion continue a 6td ca lcul6e par r appor t au f lux mesurd dans le groupe t6moin ; darts ces condi t ions , l ' appo r t suppld- menta i re de cet A A (129 nmol /min /g ) ne compense que par t i e l l ement sa capta t ion h6patique ( - 238 +- 70 nmol /min /g) . Ceci condui t ~ une rdduct ion de la concen t ra t ion c i rcu lan te (848 _+ 224 lamol/l ~ t90 vs 1195 _+ 105 ~mol/1 ~ %5, P < 0,001) et donc ~ une dimi- nution progressive de la captation.
- Darts le cas de l ' a rginine , on observe une augmenta- tion de la captation (tableau IV) alors que la concentra- t ion dans le per fusa t d iminue ( tableau III) . Ceci impl iquera i t un effet ac t iva teur de l ' a rg in ine sur son propre mdtabolisme. Le transport intrahdpatique de cet AA, se fa isant p r i nc ipa l emen t par d i f fus ion pass ive [13], ne reprdsente pas une 6tape limitante de son mdta- bolisme. Les variations de flux sont donc lides ~ celles du gradient intra/extracellulaire d 'arginine, c 'es t-h-dire
son ca tabo l i sme . Dans l ' h6pa tocy te , l ' a rg in ine est hydro lysde de fa~on 6quimola i re en orni thine et ur6e par l 'arginase, enzyme prdsente en grande quantit6 darts le foie [14]. Par a i l leurs , l ' a rg in ine est un inducteur puissant de l 'ur6ogen~se par son effet activateur sur la N-ac6tylglutamate synth6tase, enzyme responsable de la synth~se du N-acdtylglutamate, lui-m~me activateur allost6rique obl igatoire de la carbamyl-phosphate syn- th6tase [14]. I1 est poss ib l e que, dans nos condi t ions exp6r imenta les , un cer ta in d61ai soit n6cessaire pour activer l 'ur6ogenbse, entrMnant en retour une activation ou une lev6e d ' inhibi t ion de l 'arginase. En conclusion, darts les conditions exp6rimentales d6fi- nies dans ce t ravai l , l ' 6 t a t s ta t ionnai re des 6changes hdpatiques de la plupar t des A A peut ~tre obtenu dans le foie de rat isol6 perfus6 en recirculation. Cet 6tat peut ~tre assimil6 ?~ la phase postprandiale rda- l isant un appor t pro long6 d ' A A par la veine porte. I1 faut souligner toutefois que darts notre 6tude, le niveau
de concen t ra t ion ~ l '6 ta t s ta t ionnai re (de l ' o r d r e de deux fois la concent ra t ion p h y s i o l o g i q u e des acides aminds chez le rat) a 6tfi fix6 de mani~re ?~ obtenir des flux h6patiques constants par rapport ~t nos exp6riences en simple bolus. Dans des condit ions non saturantes en AA, les flux Mpat iques 6tant fonction de la concentra- t ion c i rcu lan te des ac ides amin6s, il est pos s ib l e en faisant varier l ' appor t d ' A A de clamper les concentra- tions et les flux h diff6rents niveaux. Ceci permet par e xe mp le d ' d tud ie r la r6ponse hdpat ique ~ diff~rents appor t s en substrats ou d ' 6 v a l u e r l ' a c t i o n de mddia- teurs sur le mdtabol isme hdpatique sans avoir ~ prendre en compte l ' e f fe t propre des var ia t ions de concentra- tions dans le mil ieu de perfusion.
Bibliographie
1. Wolkoff AW, Johansen KL, Goeser T. The isolated fiver : pre- paration and application. Anal Biochem 1987 ; 167 : 1-14.
2. De Bandt JP, Cynober L, Ballet F, Coudray-Lucas C, Rey C, Giboudeau J. Amino acid metabolism in isolated per- fused rat liver. J Surg Res 1990 ; 49 : 8-13.
3. De Bandt JP, Lim SK, Cynober L, Ballet F, Rey C, Coudray-Lucas C et al. Amino acid flux in isolated per- fused rat liver : cell free buffer vs diluted rat blood. Clin Nutr 1990 ; 9 : 56 (r6sum6).
4. De Bandt JP, Cynober L, Rey C, Coudray-Lucas C, Lira SK, Poupon R et al. Effets des acides gras sur le m6tabo- lisme h6patique des acides amin6s dans le foie de rat isol6 et perfus6. Gastroentdrol Clin Biol 1992 ; 16 : 46-9.
5. Stoll B, Haussinger D. Functional hepatocyte heterogeneity. Vascular 2-oxoglutarate is almost exclusively taken up by perivenous, glutamine-synthetase containing hepato- cytes. Eur J Biochem 1989 ; 18l : 709-16.
6. Lacy WW. Effect of acute uremia on amino acid uptake and urea production by perfused rat liver. Am J Physiol 1969 ; 216 : 1300-5.
7. De Bandt JP, Lira SK, Plassart F, Coudray-Lucas C, Rey C, Poupon R et al. Independent and combined action of IL- 113, TNFa and glucagon on amino acid metabolism in isolated perfused rat liver. Metabolism 1994 ; 43 : 822-9.
8. Mortimore GE, Poso AR, Lardeux BR. Mechanism and regulation of protein degradation in liver. Diabetes Metab Rev 1989 ; 5 : 49-70.
9. Shattuck KE, Grinell CD, Rassin DK. Amino acid infusions induce reversible, dose-related decrease in bile flow in the isolated rat liver. JPEN 1993 ; 17 : 171-6.
10. Petit JY. Administration intraveineuse. In : Houin G Ed. Pharmacocin6tique. Paris : Ellipses, 1990 : 56-66.
11. Haussinger D. Glutamine metabolism in the liver : over- view and current concepts. Metabolism 1989 ; 38 : 14-7.
12. Sies H, Haussinger D. Hepatic glutamine and ammonia metabolism. In : Hanssinger D, Sies H Eds. Glutamine metabolism in mammalian tissues. Berlin : Springer Verlag, 1984 : 78-97.
13. Metoki K, Hommes FA. The uptake of ornithine and lysine by isolated hepatocytes and fibroblasts. Int J Biochem 1984 ; 16 : 833-6.
14. Meijer AJ, Lamers WH, Chamuleau RAFM. Nitrogen metabolism and ornithine cycle function. Physiol Rev 1990 ; 70 : 701-48.
128





























