Embed Size (px)
Citation preview

This article was downloaded by: [Dicle University]On: 08 November 2014, At: 19:13Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Acta Botanica Gallica: BotanyLettersPublication details, including instructions for authorsand subscription information:http://www.tandfonline.com/loi/tabg20
Etude des successions aprèsabandon des pratiquesculturales en BretagneJean-Claude Gloaguen a , Françoise Rozé a , JeanTouffet a , Bernard Clément a & Françoise Forgeard aa Laboratoire d'Ecologie végétale , URA CNRS 696,Complexe scientifique de Beaulieu, F-35042 , RennesCedexPublished online: 27 Apr 2013.
To cite this article: Jean-Claude Gloaguen , Françoise Rozé , Jean Touffet , BernardClément & Françoise Forgeard (1994) Etude des successions après abandon despratiques culturales en Bretagne, Acta Botanica Gallica: Botany Letters, 141:6-7,691-706, DOI: 10.1080/12538078.1994.10515230
To link to this article: http://dx.doi.org/10.1080/12538078.1994.10515230
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.

This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

Acta bot. GaUica, 1994, 141 (617), 691-706.
Etude des successions apres abandon des pratiques culturales en Bretagne
par Jean-Claude Gloaguen, Fran\(oise Roze, Jean Touffet, Bernard Clement et Fran\(oise Forgeard
Laboratoire d'Ecologie vegetale, URA CNRS 696, Complexe ~cientlfr.que de Beaulieu, F-35042 Rennes Cedex
(Manuscrit re~tu le 24 fovrier 1993; accepte le 28 mars 1994)
Resume.- Notre ~tude concerne les successions veg~tales dans les champs et prairies abandonn~es. La ~thodologie adopt~ est une approche diachro· nique et synchronique. Elle vise a ~tablir des s~ries dynamiques puis a caract~riser l'~volution des parametres &cologiques lors de Ia dynamique. Deux ~chelles spatiales et deux ~chelles temporelles sont utilis~s : l'echelle bocagere ou l'on peut suivre l'~volution de l'agencement spatial de Ia vegetation et de son recouvrement depuis quelques ~cennies et l'echelle parcellaire oil Ia dynamique v~g~tale est suivie depuis 3 ans dans des placettes et le long de lignes permanentes. Les processus dynamiques sont Ires rapides et mettent en jeu deux strat~gies, par croissance marginale a partir des haies et par croissance centrale. Divers s~narios de succession sont observ~s. qui different selon l'histoire des parcelles, leur etat au moment de !'abandon et leur environnement biotique. La plupart des theories concernant les successions v~g~tales peuvent s'appliquer a Ia recolonisation des parcelles abandonn~es.
Summary.- Our study is concerning wHh plant succession in neglected fields and grasslands, using diachronic and synchronic analysis. It is aiming to build dynamic series and to characterize the ecological parameters during these successions. Two spatial scales and two temporal scales is used : on one hand, the bocage landscape scale for long term analysis and on the other hand the parcel scale for short term studies on permanent plots and on perrna· nent lines. Dynamic processes occur very rapidly, using two strategies: by marginal growth from the hedges and by patch growth into the parcel. Various succession scenarios have been observed. They are depending on the parcel history (particularly its original state before abandonment) and also on the biotic environment of the parcel. Most of the plant succession theories can be applied to recolonization of neglected lands.
Key wor~ : bacterial biomass - cellulolysis · neglected field - nitrification -aerial photointerpretation · secondary successions.
©Societe botanique de France 1994. ISSN 1253·8078.
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

692 ACTA BOTANICA GALLICA
INTRODUCTION
La crise actuelle de l'agriculture, due a Ia fois a Ia politique agricole commune europeenne, a Ia surproduction, au vieillissement de Ia population et a l'exode rural, provoque un abandon accelere des terres agricoles. Mais celui-ci est tres difficile a chiffrer parce que les surfaces abandonnees, non rattachees a une exploitation, echappent au champ de Ia statistique agricolc (Mergoil et Roudie, 1991).
Ce processus atteint d'abord les zones les plus pauvres ct Ia Bretagne est particulierement touchee. La deprise agrieole s'y exprime par l'abandon de terres cultivees, mais surtout par l'absence d'entretien, Ia baisse d'utilisation et l'abandon de certaines formations vegetales exploitees de fa .. on plus ou moins extensive, telles que les prairies permanentes.
Le probleme du devenir de ces zones et de leur mode de gestion ou d'entretien necessite de mieux connaitre Ia structure, le fonctionnement et Ia dynamique des systemes ecologiques. II est done important d'evaluer les modifications des caracteristiques biologiques et eeologiques des zones en deprise.
Nous nous sommes interesses a l'etude des successions secondaires dans des parcelles qui etaient autrefois soit des champs cultives, soit des prairies permanentes. L'etude porte sur deux zones agricoles, l'une en bordure du camp militaire de Coetquidan (Morbihan) dans un environnement bocager et forestier de Bretagne interieure, l'autre a Frehcl (Cotes d'Armor) dans un environnement de Iandes littorales. Ces deux sites correspondent a deux contextes biogeographiques et paysagers deja reconnus comme des Clements majeurs en Bretagne, dans une analyse plus globale de Ia vegetation de Ia region (Gloaguen, 1988).
Nos travaux eoncernent essentiellement des parcelles abandonnees : les champs cultives evoluant vers des friches posteulturales, scion une nomenclature admise par les scientifiques (Blais et al., 1977 ; Clement et al., 1981 ; Delpech et al., 1985), les prairies fauchees ou (et) piiturees vers des prairies abandonnees.
L'objectif de cette etude est double. Le premier est de decrire et de tenter d'expliquer les divers scenarios de successions observes par l'abandon des pratiques culturales. II sera done egalement interessant de connaitre les consequences sur le sol, et notamment sur l'evolution de Ia matiere organique, de l'installation des principales especes impliquees dans les successions. Le second est de tenter une conceptualisation de ces successions en leur appliquant les principales theories, qu'elles soient anciennes, comme le concept autogenique de Clements (1916) ou le concept individualiste de Gleason (1926) ou, plus recentes, comme les modeles de competition de Connell et Slatyer (1977) ou lcs strategies adaptatives de Grime (1987).
METHODES
La methodologie adoptee pour !"etude des successions apres abandon des pratiques culturales est une approche a Ia lois synchronique et diachronique. Deux echelles spatia las. correspondant a deux echelles temporal· les. sont utilisees : l'echelle du paysage bocager (etude a long terme) et l'echelle parcellaire (etude a court et moyen terme).
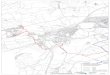
Deux methodes sont utilisees a l'echelle bocagine. o·une part, Ia dynamique de Ia vegetation, sur une cinquantaine d'hectares, a ate suivie grAce a l'interpretation de photographies aeriennes (sept missions en 1948, 1952, 1960, 1968, 1974, 1982 et 1986, dans Ia region de Coetquidan). D'autre part. toujours dans cette meme region, 45 releves de vegetation ont ate realises (Fig. 1 ). selon Ia methode de Braun-Bianquet, et traMs ensuite par une analyse factorielle des correspondances.
A l'echelle parcellaire, les methodes utilisees sont plus variees. L'etude des successions vegetales est faite a partir du suivi de lignes permanentes (deux en
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

J.-C. GLOAGUEN ET AL. 693
Fig. 1.- Localisation des stations etudiees et des releves. Secteur A, 2 releves ; secteur B, 7 releves ; secteur C, 21 releves ; secteur D, 7 releves ; secteur E, 3 releves ; secteur F, 2 releves ; secteur G, 3 releves.
Fig. 1.- Location of the study sites and of the plots. Area A, 2 plots ; area B. 7 plots ; area C, 21 plots ; area D, 7 plots ; area E, 3 plots ; area F, 2 plots ; area G, 3 plots.
!riches, trois en prairies abandonnees) lues annuellement par Ia methode des points quadrats (Canfield, 1941 ; Forgeard et Touffet, 1979). Par ailleurs, trois pet~es parcelles de 0,2 ha (une en friche, deux en prai· ries abandonnees) sont cartographiees avec precision taus les ans (methode des placettes permanentes, Van der Maarel, 1984). Dans l'une d'elles (une prairie abandonnee depuis Ia fin des annees 70, dans Ia region de
Coetquidan), les ligneux (Salix atrocinerea, Prunus spinosa et surtout Quercus robur) font l'objet de mesures biometriques annuelles : hauteur totale, circonference du tronc ~ 1 ,30 m, diametre du houppier.
Les relations vegetation-sol font appal c1 des methodes complementaires. La dllcomposition de Ia matiere organique est evaluee en raison de sa si"1>iicite par le coefficient de Jenny eta/. (1949), calcule c1 partir
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

694 ACTA BOTANICA GALLICA
de prelevements de litieres. l'activite cellulolytique est quantifiee en mettant de Ia cellulose ~ decomposer in situ (Roze, 1986). les biomasses bacteriennes sont mesurees par une methode de fumigation (Chaussod et a/., t 986). L'activtte minerallsatrice et nttrificatrice est appreciee gr~ce ~ des incubations in situ (lemee, 1967). Les analyses sont effectuees dans des parcelles correspondant aux principaux stades de Ia sene prairie-fourre (Fig. 7).
RESULTATS
I - Dynamique des groupements vegetaux a l'echelle bocagcre
1.1 - Evolution de Ia vegetation La zone du camp militaire de Coet
quidan presente un reel interet pour l'etude des successions ii long terme. En effet, les parcelles cultivees y ont etc progressivement soustraites ii ('agriculture, certaines depuis 1912. Dans
l'exemple choisi, l'examen des photographies aeriennes permet de distinguer 3 types de formations : les friches, les fourres et les forets.
En 1948 (Fig. 2), Ia structure bocagere primitive apparait encore tres nettement : les haies delimitent un reseau de parcelles de tailles et de formes differentes mettant en evidence un maillage heterogene. En 1968 (Fig. 3), ce maillage est beaucoup moins net. Les grandes parcelles soot toujours discernables, mais leur surface en friches s'est nettement reduite. Generalement, cette reduction est due ii une croissance marginale, ii partir des haies ou du massif forestier preexistants. Elle peut egalement etre due ii une croissance centrale, a l'interieur meme de Ia parcelle : c'est le cas de Ia seule parcelle en fourres en
Fig. 2.-les parcelles de Coetquidan en 1948 (l'echelle indique 100m) Fig. 2.- The parcels of Coetquidan in 1948 (the bar corresponds to 100m)
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

J.-C. GLOAGUEN ET AL. 695
Fig. 3.- Les parcelles de Coetquidan en t 968 (l'echelle indique t 00 m) Fig. 3.- The parcels of Coetquidan in 1968 (the bar corresponds to 100m)
1948. Les plus petites parceUes sont deja passees, en 20 ans, du stade de friche au stade forestier. En 1986 (Fig. 4), Ia foret a pris une importance considerable et le maiUage bocager n'apparait plus qu'ici et Ia. Certaines parceUes, a l'interieur du massif forestier' ont ete periodiquement remises en culture (ble, ma"is) entre 1968 et 1986 ; elles sont actuellement abandonnees.
En resume, l'etude des photographies aeriennes (de 1948 a 1986) a permis de mettre en evidence les processus suivants:
ll Ia surface des friches regresse regulierement au profit des forets, alors que celle des fourres commence par augmenter pendant une vingtaine d'annees avant de diminuer regulierement
jusqu'a nos jours ; 2/ les processus dynamiques sont tres
rapides : des parcelles en friches en 1948 sont recouvertes d'une vegetation forestiere en quelques decennies ;
3/ Ia dynamique forestiere a partir d'un reseau bocager met en jeu 2 strategies : par croissance marginate, a partir des haies ou du massif forestier preexistants, et par croissance centrale, a l'interieur meme de Ia parcelle.
1.2 - Dynamique des groupements vegetaux
L'analyse factorielle des correspondances des 45 releves (Le Bourbouac'h, 1989) met bien en evidence l'existence de 4 grands groupes (Tableau I) : les friches, les prairies abandonnees, les
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

696 ACTA BOTANICA GALLICA
Fig. 4.- Les parcelles de Coiltquidan en t 986 (l'~elle indique 100 m) Fig. 4.- The parcels of Coetquidan in 1986 (the bar corresponds to 100 m)
fourres et les forets. Alors que les friches sont bien caracterisees par un certain nombre d'especes, notamment les annuelles comme ]uncus bufonius ou AnagaUis arvensis, et des bisannuelles comme Centaurium umbeUatum, les prairies abandonnees ne possedent que tres peu d'especes qui leur soient infeodees : les hemicryptophytes Stachys offu:inalis et Potentilla erecta. Aucune espece ne caracterise les fourres alors que les especes strictement forestieres sont egalement abondantes, notamment les sciaphiles comme Circaea lutetiana.
Beaucoup d'especes sont communes a deux formations vegetales : friches et fourres, prairies abandonnees et fourres, fourres et forets. Ceci est bien visualise sur le tableau 1 et met en evidence le remplacement progressif des
especes des friches, par exemple, par celles des fourres, puis celles des fourres par celles des forets. En effet, les releves representent davantage des etats dynamiques de transition entre formations au sein de successions vegetales que des groupements vegetaux bien stabilises.
D'autres especes sont communes a trois ou meme quatre groupements : apparaissant rapidement, elles se maintiennent au cours des stades fourres, voire forets. Ceci est particulierement net pour les especes prairiales et explique le faible nombre d'especes caracteristiques des prairies abandonnees. Ces pra1r1es abandonnees, comme les fourres, sont neanmoins bien caracterisccs par leur composition floristique glohale.
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

J.-C. GLOAGUEN ET AL.
Tableau 1.- Repartition des principales especes dans les friches (I), les prairies (II), les fourres (Ill) et les forAts (IV).
Table 1.- Distribution of the main species in the fallow fields (1), the grasslands (II), the thickets (Ill) and the forests (IV).
FRICHES
FRICHES +
RlURRFS
PRAIRIES
PRAIRIES
+
+
RJRETS
RJRETS
ESPECES
COMMUNES
1 l.t'nlauruura /AitiDtlltllunt
Runl#'.-c ar.t!IDS~lln
}Urte'IIS bllfOIIiMS
Ana~tJJiis arwnsiJ Loliunt "tldlijlorum Arrllnta~Mrum bulbosum
l.in.iastriolo Cirsium tiiWIW
s .. u.n.,llok.umollis C_disus scoparia
Stacll.vsof!idntJiis
PotmlilltJer«fa
RanuttetdMS tiO"IS
Ft>stiiCOovirtd
Lolus ulisinosMS Str/lariaholosiH ConnpodiuntiMjiU
C ntldiU'H IWflltrdhS
Rumex aatosa Ulut>uroptii!US Sali.t otrocilw«J
Ajugar'f'IIJIU 1'irJ/anlVaicPia Gali,...apt'iN Smrbw.u.s rtitra ll~tJiteli..t
Dnula pubc!ll:rru
Bdulapmdula
CirctJ«Jbfttimta
Dr;mpt~is filix-mtU Brcrchypodiunt ~IWilicum
E W"*Y""" t'llfopiWIU
Ct~~ri'Uis ltiiiRI"nN Cm:vlus tn't'llana
lte.t oquifolitfnl Lonicn-a pn-rd_\'fflmlml
Carpinus bfttdHs
Frtnimu ~,-nl.fior Pmnustn'""" CaJ1tftftrmn·a
AclliiiH millrfoii,.,, l.cr1CQIIII¥mll"' l'ldRDTe Dac~lis tlnntmJia RanUttCubu rqwru llfllrNJ-IataHS
A~trtutis tmuis Ruhus p. frutiam•s Pnmus spint-sa CniiMJIUS ,.,..,R_\'na Quf'N:usrCJbur Ptt:'t'idiumaq11ilinuttt \',•rt~~~icarltanllJitdrvs
1'etuTium scnrodnt~ia
Ill IV
------- 1--- 1--- r--- 1--1-- 1--
--
1--
---------
2 - Consequences de Ia deprise a l'echelle parcellaire
La mise en place, depuis 3 annees, de lignes et placettes permanentes permet de faire quelques remarques.
11 La strate herbacee, qui evolue assez rapide-
697
ment dans les friches postculturales, est plus stable dans les prairies abandonnees.
2/ La cartographic des parcelles montre que le role de Ia croissance marginate a partir des haies est particulierement net pour un certain nombre d'especes comme Prunus spinosa, Pteridium aquilinum, ou du taxon Rubus gr.fruticosus. Cette derniere espece a ete, de plus, l'objet d'une cartographic detaillee le long d'une portion de haie de 10m dans Ia prairie de Coetquidan. Les calculs, par planimetric, mootrent une avancee moyenne d'environ 20 em par an du front du roncier. Cette progression correspond a une dynamique vegetative racinaire.
3/ Les ligneux, notamment les especes arborescentes comme Quercus robur, ont une croissance rapide et peuvent, lorsqu'ils sont assez denses, modifier en quelques annees Ia physionomie de Ia parcelle. Dans Ia prairie abandonnee de Coetquidan, le chene pedoncule a commence a s'installer au debut des annees 80. Les resultats des mesures effectuees sur cette espece, pendant trois annees consecutives, apparaissent dans le tableau 2 et Ia figure 5. Leur nombre est relativement stable. En fait, quelques-uns ont disparu pendant que quelques jeunes faisaient leur apparition, introduisant ainsi, en plus d'une dynamique vegetative, une dynamique de diaspores. Leur croissance en hauteur est relativement rapide, plus de
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

698 ACTA BOTANICA GALLICA
Tableau 2.- Quelques donnees biometriques concernant Quercus robur Table 2.- Some data on Quercus roburblometry
Hauteur (m) cwconterence Diametre Surface a t30cm (em) houppier (m) houppier (m2)
1989 1990 1991 1989 1990 1991 1989 1990 1991 1989 1990 1991
N 159 159 158 95 116 m1ni 02 0.3 04 05 05 maxJ 3.5 42 45 10 13
moyenne 1.42 177 209 318 4 78
E. st. 0.05 0.06 007 0.30 0 30 somme
(.st. : erreur standard
30 em par an en moyenne, alors que leur circonference a 1 ,30 m augmente annueUement de 1,6 a 1 ,8 em. Mais c'est au niveau de Ia surface des houppiers que l'accroissement est le plus spectaculaire: il passe de 81m2 en 1989 a 290m2 en 1991, ce qui correspond presque a un doublement annuel. Ceci illustre bien les importants changements physionomiques de cette prairie qui est en train d'evoluer rapidement vers un stade prHorestier et confirme les observations faites dans Ia structure bocagere par photointerpretation.
2.2 - Relations sol-vegetation La figure 6 montre que, par rapport
a Ia prairie prise comme temoin du Stade initial, l'implantation des groupements a Fougere aigle (Pteridium aquilinum), PruneUier (Prunus spinosa) et Ajonc (Ulex europaeus) entraine de nombreuses modifications de l'activite biologique des sols.
Dans le cas de l'installation d'un groupement a Fougere aigle, Ia vitesse de decomposition de Ia matiere organique est tres ralentie par rapport a Ia prairie initiate. L'activite cellulolytique est reduite. La biomasse bacterienne diminue et Ia nitrification devient quasiment nulle. Le pH des horizons de surface est de 4 alors qu'il etait de 6 dans Ia prairie temoin. Ces resultats tcmoignent
117 159 159 157 159 159 157 05 0.1 0.1 0.1 O.ot 001 001 16 2 22 3.5 314 3.80 962
660 0.68 096 1 35 0 51 093 1 85
0.33 004 004 006 0.05 007 015 81 52 148.12 290.18
d'une degradation du sol qui se traduit sur le terrain par un epaississement de l'humus acide de l'ordre de 10 em.
Dans le cas de Ia mise en place d'un fourre a PruneUier, Ia figure 6 montre que Ia decomposition de Ia litiere et Ia degradation de Ia cellulose se font plutot mieux qu'en prairie ; Ia biomasse bacterienne augmente. La nitrification est un peu moins importante mais Ia vegetation des fourres acides a Ia capacite d'absorber de l'azote ammoniacal (Roze, 1986). Globalement les potentialites du sol sont maintenues.
Dans le cas de }'implantation d'un groupement a Ajonc, Ia vitesse de decomposition est un peu plus lente qu'en prairie. La cellulolyse est equivalente dans les deux cas. La croissance rapide de cette Legumineuse ne se traduit pas par une augmentation de Ia biomasse bacterienne. La encore, les potentialites du sol semblent maintenues.
DISCUSSION
Les differents resultats obtenus, pour les successions vegetates, par les diverses methodes utilisees, et confortes par l'analyse experimentale de l'activite biologique des sols, permettent de preciser les divers scenarios de succession observes apres abandon des prairies culturales en Bretagne (Fig. 7).
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

Fig
. 5.
-B
iom
etrie
des
chl
lnes
plld
on
cule
s : h
aute
ur (
en m
), ci
rcon
fere
nce ~
1,30
m (
en e
m)
et s
urfa
ce d
es h
oupp
iers
(en
rri'
). Le
s no
mbr
es a
ppar
aiss
ant
en a
bsci
ssa
des
diag
ram
mes
indi
quen
t le
s va
leur
s su
perie
ures
des
cla
sses
(pa
r ex
ampl
e 0,
5 po
ur Ia
cla
sse
0-0
,5;
1 po
ur Ia
cla
sse
0,5-
1 ..
. ).
Fig
. 5.
-Q
uerc
us r
obur
bio
met
ry :
hei
ght
(m).
1,30
m c
ircum
fere
nce
(em
). ca
no
py
area
(rr
i').
The
abs
ciss
a nu
mbe
rs i
ndic
ate
the
uppe
r va
lues
of
the
clas
ses
(for
ex
ampl
e, 0
,5 f
or th
e 0-
0,5
cla
ss;
1 fo
r th
e 0,
5-1
clas
s ..
. ).
~ 0 Cl ~ c m
z Ill
-1
~ r- "' :8
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

700 ACTA BOTANICA GALLICA
Fig. 6.- Modification de l'activitll biologique des sols apres dllprise Fig. 6.- Modnication of the biological activ~y of the soils after neglecting
Les friches et les prairies abandonnees peuvent etre envahies par Ia Ronce (Rubus gr. fruticosus) et Ia Fougere (Pteridium aquilinum), Ia premaere espece etant plus frequcnte en friches, Ia seconde en prairies. De meme, alors que les friches sont plutot colonisees par le Genet (Cytisus
scoparius), les prairies le sont plutot par I'Ajonc (Ulex europaeus). Quel que soil le scenario, friches et prairies abandonnees evoluent progressivement vers le fourre prHorestier a Prunellier (Prunus spinosa) et Aubepine (Crataegus monogyna), remplace a son tour par Ia foret.
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

J.-C. GLOAGUEN ET AL. 701
FORET
Fig. 7.- Les d~fllrents scllnarios de succession Fig. 7.- The d~ferent succession scenarios
Cette installation de Ia foret peut etre tres rapide : parfois moins de vingt ans (Fig. 2, 3 et 4), mais ce n'est pas toujours le cas. Certains scenarios sont caracterises par un net ralentissement (voire meme un blocage tcmporaire) des processus dynamiques se caracterisant par l'apparition de paliers successionnels (sucession en plateau de Whittaker et Levin, 1977). C'est le cas notamment des peuplements tres denses de Cytisus scoparius, Ulex europaeus, Rubus gr. fruticosus et surtout de Pteridium aquilinum. Ces processus ont cgalemcnt etc reconnus dans des successions en region mediterraneenne (Debussche et al., 1982, 1985, 1992).
Divers mecanismes peuvent expliqucr un tel ralentissement au cours de Ia succession ; d'unc part, un phcnomcnc
physique d'encombrement de l'espace qui est du, soit a l'importance de Ia biomasse aerienne (effet de masse du a Ia densite des individus, a leur hauteur, a leur forme ... ), soit a l'importance de Ia litiere (ou d'un mat fibreux en prairie) qui isole les plantules du sol et accroit leur sensibilite aux contraintes, notamment Ia secheresse ; d'autre part, un phcnomene chimique de modification des processus pedologiques, lie au ralentissement de Ia decomposition de Ia matiere organique, et qui est du notamment a une diminution de l'activite biologique du sol.
C'est ce qui apparait lors de l'installation d'un groupement a Pteridium aquilinum, avec comme consequence une degradation du sol. lnversement, Ia mise en place d'un fourre a Prunus
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

702 ACTA BOTANICA GALLICA
spinosa ne modifie pas les potentialites du sol. II en est de meme lors de l'instaUation d'un groupement a Ulex europaeus. Cependant, il est frequent, notamment lorsque le peuplement est tres dense, que I'Ajonc vieillisse sans que d'autres especes ligneuses ne puissent s'instaUer : le passage au fourre preforestier et a Ia foret peut alors etre tres long.
Dans d'autres types de scenarios, tres rapides eux aussi, les stades dont nous venons de parler sont absents : le passage des friches ou des prairies abandonnees a Ia foret se fait alors directement par l'installation d'especes arborescentes. Alors que les Chenes, notamment Quercus robur, colonisent indifferemment les friches ou les prairies, les Bouleaux (Betula pubescens, B. pendula) sont plutot abondants en friches et les Saules (Salix atrocinerea) en prairies. La croissance rapide de ces arbres (voir Fig. 5 pour Quercus robur) transforme en quelques annees un paysage de friches ou de prairies en un paysage preforestier, voire forestier.
Les successions, qu'elles soient primaires ou secondaires, ont fait l'objet de nombreuses theories depuis le debut du siecle.
Le concept autogenique de Clements (1916) s'applique bien aux prairies abandonnees : les especes en place creent des conditions favorables a l'installation de nouvelles especes, par reaction ou modification du site (ccci apparait dans lc tableau 1).
Egler (1954) ajoutc a cc concept cclui de Ia composition floristique initiate (il nc nic d'aillcurs pas le concept autogcniquc qu'il rcmplacc par cclui de rclais floristiqucs) ; travaillant sur les successions sccondaircs, notammcnt apres abandon cultural, il attribuc un role primordial au stock de graincs au sol. Ce concept s'appliquc bien aux champs abandonnes de Brctagne, cc qui corrcs-
pond a une interaction indirecte entre organismes vis-a-vis de ressources lirnitees, il s'agit d'un processus competitif par exploitation (Connell, 1990). Mais il est evident qu'une partie non negligeable des diaspores, notamment ceUes des especes ligneuses, resulte d'apports exterieurs, notamment dans le contexte bocagcr. II s'agit de processus competitifs par interference, c'est-a-dire d'une interaction directe entre individus qui recherchent activement ce que d'autres utiliscnt dans un pool commun de ressources (Connell, 1990; Debussche et Lepart, 1992).
Pour bien comprendre ces successions' il est necessaire de bien connaitre Ia biologic des especes : pour le concept individualiste des successions (Gleason, 1926 et 1927), les proprietes de Ia vegetation dependent des proprietes des individus qui Ia composent. Ainsi, lcs proprietes des ecosystemes sont uniquement fonction des caracteres physiologiques et ecologiques des cspeces presentes comme, par exemple, leur capacite de resistance aux contraintes, leur pouvoir de dissemination ou leur aptitude a Ia competition pour les ressources du milieu (Drury et Nisbet, 1973 ; Noble et Slatyer, 1980).
Grime (1977, 1979 et 1987) distingue, scion leurs strategies adaptatives, trois types d'especes : les especes S, tolerantes aux contraintes physiques de l'environncment ou stress, lcs especes R ou ruderalcs, qui profitcnt des perturbations, et les especes C, competitives, dans lcs milieux potentiellcment riches. Dans lcs successions secondaires resultant de Ia deprise agricole (Fig. 8), les cspeccs de type R ou R-S (friches et prairies) sont remplacees par des especcs de type C-R ou C-S-R (fourres), cllcs-memcs rcmplacees par des especcs de type C-S ou C (forets). Deux voies principales sc differencient (Fig. 9), en liaison avec l'etat initial de Ia parcelle
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

J.-C. GLOAGUEN ET AL. 703
c
It s I R
I S
Fig. 8.- Pos~ion des especes dans le triangle R-C-S (Grime, 1977). R: espece rudllrale; C: espece compet~ive; S : espece tolllrante au stress. IC, IR. IS : intensite croissants (salon Ia lleche) de Ia competitiv~e. de Ia ruderalitll. de Ia tolerance au stress. Ra : Rumex acetosella ; Ca : Cirsium atvense ; HI : Ho/cus lanatus ; So : Stachys officina/is ; Pe : Potentilla erecta ; At : Agrostis tenuis ; Cs : Cytisus scoparius ; Ue : Ulex europaeus ; Bp : Betula pubescens ; AI : Rubus fruticosus ; P1 : Pteridium aquilinum ; Ps : Prunus spinosa ; Cm : Grataegus monogyna ; Sa : Salix atrocinerea ; Sn : Sambucus nigra ; Fe : Fraxinus excelsior; Pa : Prunus avium; Or : Quercus robur; C1 : Castanea sativa ; Cl : Circaea lutetiana ; Of : Dryopteris filix-mas ; He : Hedera helix.
Fig. 8.- Species pos~ion in the R-C-S model (Grime, 1977). R: ruderal species; C: competitive species; S: stress-tolerant species. IC, IR, IS : increasing intensity (arrow) of competitiveness, ruderality and stress tolerance.
(champ ou prairie). C'est, dans le cas du champ abandonne, que Ia vitesse de colonisation est Ia plus rapide, pour deux raisons essentielles : d'une part, parce que ('invasion par lcs especes potcnticllcs de Ia foret peut intcrvcnir des Ia phase initiale de colonisation qui, d'autre part, est facilitee par Ia richcssc du milieu correspondant a d'ancicnncs zones cultivees. En prairie abandonncc, Ia lecture des ligncs permancntcs montre que Ia vitesse de colonisation est plus lente, avec des phases de ralcntisscment
(voir me me de bloc age temporaire) de Ia succession, en liaison avec des phenomenes d'encombrement (effet de masse, production importantc de litiere) auxqucls s'ajoutent des potentialites nutritives infericures sur lcs sols moins fertiles des prairies mcsophiles ou plus souvent mesohygrophilcs.
Scion leur strategic de colonisation, deux groupcs d'especes se distinguent : les espcces a dispersion clonale et les cspcces a dispersion par semences. Les premieres cnvahissent gcneralcmcnt les
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

704 ACTA BOTANICA GALLICA
c
R s
s
Fig. 9.- Les deux principaux scenarios de succession dans le triangle R-C-S (Grime, 1977). Voir fig. 8. I :!riches ; II : prairies abandonnees ; Ill : fourres ; IV : !orAls.
Fig. 9.- The two main succession scenarios in the R-C-S model (Grime, 1977). See fig. 8. I: fallow fields; II: neglected grasslands ; Ill : thickets ; IV : forests.
parcelles a partir des marges, scion deux tactiques (Clegg, 1978 ; Lovett Doust et Lovett Doust, 1982). Dans les tactiques de phalange, les modules sont courts, epais, serres, longuement persistants : Pteridium aquilinum en donne un bon exemple. Dans les tactiques de guerilla, les modules sont longs, fins, espaces, souvent a duree de vie courte : Rubus gr. fruticosus en est un exemple type. Les secondes, colonisatrices par semences, peuvent egalement se classer en deux groupes : les especes ancmochores (Betula pendula, B. pubescens), tres opportunistes, s'installant rapidement sur sol nu (apres perturbation par exemple) et les espcces zoochores (Quercus robur, Castanea sativa, Crataegus
monogyna, Prunus spinosa), possedant souvent des semences avec d'importantes reserves. Ces especes a dispersion par scmences sont avantagees en friches alors que les especes a dispersion clonale sont plutot avantagees en prairies.
Les interactions entre especes, et notamment Ia competition, jouent un role capital dans les processus de colonisation. Ceci a conduit Connell et Slatyer (1977) a proposer trois modeles qui permettent de mieux comprendre les mecanismes de Ia succession. Dans le modele de facilitation, les especes pionnieres, en modifiant l'environnement, permcttent l'installation et le developpement d'especes nouvelles. Ce modele reprend lcs deux concepts voisins de Ia
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

J.-C. GLOAGUEN ET AL. 705
succession autogenique (Clements, 1916) et des relais floristiques (Egler, 1954). Dans le modele de tolerance, les modifications du milieu dues aux especes pionnieres soot sans influence sur l'installation de nouvelles especes : seules interviennent leurs caracteristiques biologiques. Dans le modele d'inhibition, les especes pionnieres, en monopolisant l'espace ou les autres ressources, inhibent Ia croissance de certaines especes deja en place ou empechent l'installation d'especes nouvelles. Un quatrieme modele est signale par Lawton (1987): le modele de colonisation au hasard. L'installation se fait au hasard et seules les chances de survie des differentes especes interviennent sans qu'il y ait d'interaction entre elles. II est bien evident, comme le fait remarquer Miles (1987), qu'un de ces modeles ne peut a lui seul expliquer Ia succession observee. La colonisation par les especes ligneuses, quel que soit le stade de Ia succession, s'apparente au modele de tolerance, alors que l'apparition des especes sciaphiles dans les fourres et les forets illustre le modele de facilitation. La presence de Pteridium aquilinum ou de Cytisus scoparius en peuplement dense se traduit par un ralentissement, voire meme un blocage des processus dynamiques : ceci correspond bien au modele d'inhibition. Apres transport par
les oiseaux, par exemple, des germinations de chenes ou de chiitaigniers penvent apparaitre dans les prairies dans des perturbations en taches (Horn, 1976), sous l'action d'animaux comme les sangliers : le modele de colonisation au hasard s'applique bien dans ce cas, tout en sachant que l'apparence de distribution au hasard est liee au determinisme de transport et done au comportement des zoochores (Debussche et al., 1982 et 1985).
Parmi tous ces concepts theoriques evoques, Ia plupart peuvent etre transposes aux successions secondaires apres extensification ou abandon des pratiques culturales, mais il s'agit Ia de phenomenes tres complexes, tres variables dans l'espace et dans le temps. Aussi Ia recherche de theories generales de Ia succession (Van der Maarel, 1988) doivent-elles donner lieu a des precisions sur les mecanismes fonctionnels et sur Ia vitesse d'evolution des systemes presidant a l'organisation des successions. Ceci est particulierement net en systeme de deprise ou les situations de depart soot d'une extreme diversite : histoire des parcelles, etat lors de l'extensification ou de l'abandon, environnement biotique. II en resulte une grande diversite des processus dynamiques se traduisant par Ia grande variete des scenarios de recolonisation observes.
BIBLIOGRAPHIE
Blais R. eta/., 1977.- Dictionnaire d'agricuHure et des sciences annexes. La Maison Rustique. Paris, 219 p.
Canfield A.M .. 1941.- Application of the line intercept method in sampling range vegetation. J. For., 39, 388-394.
Chaussod R., B. Nicolardot & C. Catroux, 1986.- Mesure en routine de Ia biomasse microbienne des sols par Ia methode de fumigation au chloroforrne. Science du Sol, 2, 201-211.
Clegg L., 1978.- The morphology of clonal growth and ~s relevance to the population dynamics of perennial plants. Ph. D. Thesis, lniv. Wales.
Clement J.M. eta/., 1981.- Larousse agricola. Larousse, Paris, 1208 p.
Clements F.E., 1916.- Plant succession: an analysis of the development of vegetation. Carnegie Institute of Washington, Publ. 242,512 p.
Connell J.H. & R.O. Slatyer, 1977.- Mechanisms of succession : natural communities and their role in commun~y stabil~y and organization. Am. Nat .• 111. 1119-1144.
Connell J.H .• 1990.- Apparent versus "Real" competition in plants. In : J.B. Grace & D. Tilman. Acad. Press, 9-26.
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014

706 ACTA BOTANICA GALLICA
Debussche M., J. Escarre & J. Lepart, 1982.- Ornithochory and plant succession in mediterranean abandoned orchards. Vegetatio, 48, 255-266.
Debussche M., J. Lepart & J. Molina, 1985.- La dissemination des plantas a fruits charnus par las oiseaux : rOle de Ia structure de Ia vegetation et impact sur Ia succession en region mediterraneenne. Acta Oecologica, Oecol. Gener., 6, 65-80.
Debussche M. & J. Lepart, 1992.- Establishment of woody plants in mediterranean old fields : opportunity in space and time. Landscape Ecology, 6 (3), 133-145.
Delpech R., G. Duma & P. Galmiche, 1985.- Typologie des stations forestieres. Vocabulaire. lnst. Develop. forestier, 243 p.
Drury W.H. & I.C.T. Nisbet, 1973.- Succession. J. of the Arnold Arooretum, Harvard Univ., 54, 331-368.
Egler F.E., 1954.- Vegetation science concepts. 1. Initial floristic composition : a factor in old-field vegetation development. Vegetatio, 4, 412-417.
Forgeard F. & J. Toullet, 1979.- Las premieres phases de recolonisation vegetale apres incendie dans las pelouses et las Iandes de Ia region de Paimpont (llle-et-Vilaine). Bull. Soc. bot. Fr .. 126, Lett. bot., 473-485;
Gleason H.A., 1926.- The individualistic concept of the plant association. Bull. To"ey Bot. Club, 53, 7-26.
Gleason H.A., 1927.- Further views on the succession concept. Ecology, 8, 299-326.
Gloaguen J.C., 1988.- Etude phytosociologique des Iandes bretonnes (France). Lejeunia, 124, 1-47.
Grime J.P., 1977.- Evidence lor the existence of three primary strategies in plants and ns relevance to ecological and evolutionary theory. Am. Nat., 111, 1169-1194.Grime J.P., 1979.- Plant strategies and vegetation processes. John Wiley and Sons, Chichester, 222 p.
Grime J.P., 1987.- Dominant and subordinate co~nents of plant communities : implications for succession, stability and diversny. In : Colonization, Succession and Stabiltty. A.J. Gray, M.J. Crawler & P.J. Edwards (ads). Blackwell, Oxford, 413-428.
Hom H.S., 1976.- Succession. In: Theoretical Ecology, Principles and Applications. A.M. May (ad.), Saunders, Philadelphia, 187-204.
Jenny H., S.P. Gessel & F.T. Bingham, 1949.- Comparative study of decomposnion rates of organic matter in temperature and tropical regions. Soil Sci., 68, 419-432.
Lawton J.H., 1987.- Are there assembly rules for successional communities ? In : Colonization, Succession and Stabilny.A.J. Gray, M.J. Crawler & P.J. Edwards (eds). Blackwell, Oxford, 225-244.
Le Bourbouac'h F., 1989.- Dynamique de Ia vegetation suite a !'abandon des terres dans le camp de Colltquidan. D.E.A., Univ. Orsay et de Rennes, 95 p.Lemee G., 1967.- Investigations sur Ia mineralisation de !'azote et son evolution annuelle dans les humus lorestiers in situ. Oecol. Plant., 2, 285-324.
Lovett Doust L. & J. Lovett Doust, 1982.- The battle strategies of plants. New Scientist, 95, 81-84.
Mergoil G. & P. Roudie, 1991.- Friches et recensements de !'agriculture. Rev. Geographie Lyon, 66, 5-10.
Miles J., 1987.- Vegetation succession : post and present perceptions. In : A.J. Gray, M.J. Crawler & P.J. Edwards (eds). Blackwell, Oxford, 1-29.
Noble I.A. & R.O. Slatyer, 1980.- The use of vnal attributes to predict successional changes in plant communtties subject to recurrent disturbances. Vegetatio, 43,5-21.
Roze F., 1986.- Le cycle de !'azote dans les Iandes bretonnes. These d'Etat, Univ. Rennes, 292 p.
Dow
nloa
ded
by [
Dic
le U
nive
rsity
] at
19:
13 0
8 N
ovem
ber
2014