Embed Size (px)
Citation preview

Sciences fondamentales
DOI of or1Departme
Ing�eni�erie tissu2Departme
Linkoping, Lin3Departme
Linkoping, Su
CorrespondExperimentalLinkoping, Su
Ann Vasc SurgDOI: 10.1016/� Annals of V�Edit�e par ELS
Fibroblastes dermiques humains: une sourcecellulaire potentielle pourl’endoth�elialisation des proth�eses vasculaires
Lisa K. Karlsson,1 Johan P.E. Junker,1 Magnus Grenegard,2 Gunnar Kratz,1,3 Linkoping,
Su�ede
Introduction: R�ecemment, les travaux de recherche concernant les sources de cellules utilisa-bles en ing�enierie tissulaire vasculaire se sont multipli�es. De pr�ec�edentes �etudes rapportaientque des cellules potentiellement multipotentes �etaient retrouv�ees dans le tissu conjonctif cutan�e.De meme, des r�esultats pr�eliminaires de notre groupe sugg�eraient que les fibroblastes dermi-ques humains auraient la capacit�e de modifier leur ph�enotype in vitro en un ph�enotype sem-blable aux cellules endoth�eliales. Comme premi�ere �etape dans l’utilisation de ces cellules ening�enierie tissulaire vasculaire, nous cherchions �a d�eterminer leurs capacit�es �a former une cou-che de cellules de type endoth�elial-like sur une matrice in vitro. De plus, nous �etudions lapossibilit�e d’ensemencer des fibroblastes dermiques sur une matrice, puis de d�ebuter secondai-rement l’induction vers un ph�enotype cellulaire endoth�elial-like.M�ethodes: Les cellules cultiv�ees dans un milieu fibroblastique classique ou dans un milieud’induction endoth�elial �etaient ensemenc�ees sur une matrice �a base de g�elatine. Afin d’�etudierl’organisation des cellules, des colorations �etaient r�ealis�ees de routine. La diff�erenciation �etaitconfirm�ee par Western blot et immunohistochimie par des anticorps dirig�es contre les mol�eculescommun�ement utilis�ees dans l’identification des cellules endoth�eliales.R�esultats et Conclusion: Nos r�esultats soutiennent l’id�ee que des fibroblastes dermiqueshumains diff�erenci�es en cellules de type endoth�elial-like avant ensemencement d�emontraientune ressemblance histologique avec des cellules endoth�eliales matures, alors que des fibroblastesensemenc�es dont la diff�erenciation endoth�eliale �etait induite secondairement croissaient en multi-couches. Cependant, l’expression de diff�erentes mol�ecules de surface indiquant un ph�enotypeendoth�elial �etait observ�ee aves les deux techniques. En conclusion, les r�esultats pr�esent�es danscette �etude indiquent que les fibroblastes dermiques humains diff�erenci�es suivant un ph�enotypeendoth�elial-like pourraient etre une nouvelle source d’endoth�elialisation pour proth�eses vasculaires.
iginal article: 10.1016/j.avsg.2009.03.007.
nt of Clinical and Experimental Medicine, Center forlaire, University Hospital of Linkoping, Linkoping, Su�ede.
nt of Medical and Health Sciences, University Hospital ofkoping, Su�ede.
nt of Plastic Surgery, University Hospital of Linkoping,�ede.
ance: Lisa Karlsson, MSc, Department of Clinical andMedicine, University Hospital of Linkoping, SE-581 85�ede, E-mail: [email protected]
2009; 23: 663-674j.acvfr.2010.03.001ascular Surgery Inc.EVIER MASSON SAS
INTRODUCTION
La pathologie vasculaire est un probl�eme majeur de
sant�e publique �a l’�epoque actuelle. Les m�ethodes
th�erapeutiques utilis�ees font souvent appel �a l’utili-
sation de proth�eses autologues ou synth�etiques
(Dacron� ou polyt�etrafluro�ethyl�ene).1-3 Cepen-
dant, le fait que les sources de mat�eriel vasculaire
autologue soient limit�es ainsi que les probl�emes
d’occlusion des proth�eses synth�etiques encoura-
geaient la recherche d’alternatives th�erapeutiques
diff�erentes en ing�enierie tissulaire. L’objectif ultime
de l’ing�enierie tissulaire est de cr�eer des substituts
fonctionnels afin de remplacer les tissus perdus ou
endommag�es, en appliquant les principes de la
719

720 Karlsson et al. Annales de chirurgie vasculaire
biologie et de l’ing�eni�erie.4 Au cours de la d�ecennie
pr�ec�edente, diff�erentes techniques ont �et�e d�ecrites
pour la conception de proth�eses vasculaires.5-7 En
1998, L’Heureux et al.8 d�ecrivaient une m�ethode par
laquelle un vaisseau sanguin �etait compl�etement
g�en�er�e in vitro. Cependant, malgr�e les progr�es
r�ecents, il n’existe pas de greffon vasculaire id�eal
pour l’utilisation en pratique clinique.
Une alternative �a la conception d’un vaisseau
sanguin complet par l’ing�enierie tissulaire est
l’endoth�elialisation de proth�eses synth�etiques.
En 1984, Herring et al.9 publiaient la premi�ere�etude montrant les b�en�efices de l’ensemencement
de proth�eses vasculaires avec des cellules
endoth�eliales. Plusieurs �etudes ult�erieures mon-
traient que l’ensemencement de proth�eses vascu-
laires avec des cellules endoth�eliales avant
implantation fournissait une protection contre la
formation de thrombus et am�eliorait ainsi la survie
des greffons �a long terme.2,5,10-12 Cependant, obte-
nir des cellules endoth�eliales autologues humaines
peut s’av�erer difficile. Des cellules autologues ayant
une plasticit�e cellulaire, comme les cellules souches
m�esenchymateuses et h�ematopo€ı�etiques, �etaient
largement �etudi�ees comme source cellulaire alter-
native en ing�enierie tissulaire vasculaire.13,14
Cependant, il existe des obstacles majeurs contre
l’utilisation clinique de ces cellules du fait de leur
pr�el�evement et de leur culture.13,15,16
Au cours de ces derni�eres ann�ees, des populations
cellulaires ayant la capacit�e de modifier leur ph�eno-
type et de contribuer �a diff�erentes lign�ees distinctes
de leur tissu d’origine ont �et�e trouv�ees dans diff�erents
tissus.15-18 Un certain nombre d’�etudes rapportaient
que des cellules ayant un potentiel multipotent�etaient retrouv�ees dans le stroma conjonctif
cutan�e.19-23 Les r�esultats de notre �equipe indiquaient
que les fibroblastes dermiques humains, tout du
moins une de leurs sous-populations, pouvaient se
diff�erencier en plusieurs lign�ees m�esenchymateuses
comme l’os, le cartilage ou la graisse.24,25 Les fibro-
blastes existent en grand nombre dans la peau, et une
production cellulaire importante pouvait etre obte-
nue par des proc�ed�es mini-invasifs �a partir d’une
petite biopsie de peau. De plus, les r�esultats pr�elimi-
naires de notre groupe sugg�eraient que les fibro-
blastes dermiques humains avaient la capacit�e de se
diff�erencier suivant un ph�enotype endoth�elial-like
in vitro. La diff�erenciation survenait lorsque les
fibroblastes dermiques �etaient cultiv�es sur un milieu
de culture contenant du s�erum humain. Les cellules
endoth�eliales-like exprimaient le facteur von Wille-
brand (vWf), incorporaient des lipoprot�eines de basse
densit�e marqu�ees au fluorochrome, et formaient des
r�eseaux capillaires-like.
Notre objectif primaire �etait d’investiguer l’utilisa-
tion des fibroblastes dermiques humains diff�erenci�es
suivant un ph�enotype endoth�elial-like comme
source d’endoth�elialisation des proth�eses vasculai-
res. Nous utilisions une matrice �a base de g�elatine
porcine de type A hautement r�eticul�ee. D’anciennes
publications d�emontraient que la g�elatine favorise la
liaison et la croissance de plusieurs types cellulaires
chez le mammif�ere, et plus particuli�erement les cel-
lules endoth�eliales.26 La matrice �a base de g�elatine
utilis�ee dans cette �etude a �et�e pr�ealablement utilis�ee
pour guider la r�eg�en�eration tissulaire ainsi que pour
des th�erapies �a base de cellules in vitro et in vivo, et
ses propri�et�es indiquaient qu’elle pouvait etre
utilis�ee en ing�enierie tissulaire vasculaire.27-32 Le
deuxi�eme but de cette �etude �etait d’investiguer la
possibilit�e d’ensemencer des matrices avec des
fibroblastes dermiques humains puis de d�ebuter
secondairement l’induction en cellules de type
endoth�elial-like. Notre th�eorie �etait que cette pro-
c�edure pouvait raccourcir le temps de culture requis
pour l’endoth�elialisation des proth�eses vasculaires
par des fibroblastes dermiques humains.
MAT�ERIELS ET M�ETHODES
Isolement des cellules
Fibroblastes dermiques humains. Les fibroblastes
dermiques humains �etaient obtenus �a partir de sp�eci-
mens tissulaires (d�echets tissulaires) de patients en
bonne sant�e op�er�es en chirurgie plastique. Les tissus�etaient transport�es dans une compresse imbib�ee de
chlorure de sodium et trait�es dans les 24 heures.
Des cultures primaires de fibroblastes humains�etaient isol�ees manuellement en diss�equant la cou-
che dermique des �echantillons cutan�es, puis les frag-
ments �etaient incub�es �a 37 �C pour la nuit sur un
milieu d’aigle modifi�e par Dulbecco (Dulbecco modi-
fied eagle medium, DMEM) contenant 2% de s�erum de
veau fœtal ( fetal calf serum, MCF), de la collag�enase
165 U/mL (type 1; Worthington, Freehold, New
Jersey, E-U), et de la Dispase� 2,5 U/mL (GIBCO
BRL, Life Technologies, Karlsruhe, Allemagne). Les
cellules �etaient trait�ees par incubation dans un fla-
con de culture cellulaire de 75 cm2 �a 37 �C avec 5%
de CO2 et 95% d’humidit�e dans un milieu de crois-
sance fibroblastique ( fibroblast growth medium, FGM;
DMEM contenant 10% de MCF, des antibiotiques, et
des antimycotiques [p�enicilline 50 U/mL et strepto-
mycine 50 mg/mL]). Les milieux �etaient chang�es
trois fois par semaine.
Clones de fibroblastes. Afin d’�eliminer le risque de
contamination cellulaire des cultures fibroblastiques
primaires, des fibroblastes dermiques normaux ainsi
que des clones de fibroblastes �etaient utilis�es. Pour

Vol. 23, No. 5, 2009 Les fibroblastes comme source de cellules endoth�eliales 721
produire les clones de fibroblastes, les cultures de
fibroblastes dermiques humains d�etach�ees du pla-
teau de culture lors des passages 1-4 par proc�ed�es
enzymatiques, en rincant les cellules dans de l’acide�ethyl�enediaminet�etraac�etique (EDTA) �a 0,25%
(v/v). Ensuite, les cellules �etaient incub�ees �a 37 �Cdans de l’EDTA �a 0,12% (v/v) et de la trypsine �a0,12% (v/v). La suspension cellulaire �etait ensuite
observ�ee en utilisant un microscope invers�e �equip�ed’un micromanipulateur (InjectMan NI 2; Eppen-
dorf, Hamburg, Allemagne), et d’une micropipette
(CellTram Air, Eppendorf). En utilisant le microma-
nipulateur, les cellules isol�ees �etaient transf�er�ees
dans des puits s�epar�es sur une plaque de culture �a24 puits. Dans les puits ou la confluence �etait atteinte,
les cellules �etaient trait�ees en utilisant des passages
cons�ecutifs jusqu’�a ce que des nombres ad�equats
soient obtenus pour les exp�eriences de diff�erencia-
tion. Le milieu �etait chang�e trois fois par semaine.
Cellules endoth�eliales humaines. Les cellules
endoth�eliales humaines �etaient isol�ees suivant le pro-
tocole d�ecrit par Jaffe et al.33 Bri�evement, des cordons
ombilicaux provenant de nouveaux-n�es bien por-
tants �etaient conserv�es �a 4 �C dans un tampon
phosphate salin (TPS). Les veines �etaient canul�ees,
rinc�ees au TPS, puis inject�ees avec une solution de
collag�enase 165 U/mL (type 1, Worthington). Apr�es
20 minutes d’incubation �a 37 �C, les vaisseaux �etaient
mass�es avec douceur et perfus�es vigoureusement
avec du TPS. La solution cellulaire �etait centrifug�ee et
le culot cellulaire �etait suspendu �a nouveau dans un
milieu de croissance pour cellules endoth�eliales
(Endothelial growth medium, EGM; DMEM contenant
30% de s�erum humain provenant de plusieurs don-
neurs, des antibiotiques, et des antimycotiques
(p�enicilline 50 U/mL et streptomycine 50 mg/mL avec
3,3 mM d’isobutylm�ethylxantine [Sigma, Stock-
holm, Su�ede] et 0,8 mg/mL de toxine chol�eriforme
[Sigma]). Les cellules �etaient cultiv�ees dans 75 cm2 de
flacons de culture impr�egn�es de g�elatine (g�elatine
2%) �a 37 �C avec 5% de CO2 et 95% d’humidit�e. Le
milieu �etait chang�e trois fois par semaine.
Coloration immunohistochimique des
cultures cellulaires primaires
Les ph�enotypes des cultures primaires des cellules
endoth�eliales, des fibroblastes dermiques normaux,
et des clones de fibroblastes �etaient caract�eris�es
en immunohistochimie, en utilisant du s�erum anti-
vWf, de la cadh�erine endoth�eliale vasculaire (ve-
cadh�erine), du CD31, et de l’oxyde nitrique
synth�etase endoth�eliale (NOS3). Toutes les
exp�erimentations �etaient r�ealis�ees �a temp�erature
ambiante sauf pr�ecision contraire. Bri�evement, les
cellules �etaient lav�ees pendant 3� 5 min dans du TPS
avant d’etre fix�ees par un tampon de para-
formald�ehyde neutre �a 4% (TPN) pendant 15 minu-
tes. Les cellules �etaient ensuite lav�ees dans du TPS 3�5 min, puis incub�ees pendant 45 minutes avec du
s�erum bloquant (albumine s�erique bovine 5%) con-
tenant 0,05% de Triton X-100 afin de perm�eabiliser
les membranes cellulaires. Du Triton X-100 �etait
ajout�e uniquement lors de la coloration des �epitopes
intracellulaires (vWf et NOS3). Ensuite, les cellules�etaient incub�ees pendant 1 heure avec des anticorps
primaires (vWf dilution 1:200, clone F8/86l, Dako
Cytomation, Glostrup, Danemark; ve-Cadh�erine
dilution 1:200, clone F-8, Santa Cruz Biotechnolo-
gies, Santa Cruz, Californie; CD31 dilution 1:50, clone
JC/70A, Abcam, Cambridge, RU; et NOS3 dilution
1:50, Santa Cruz Biotechnologies). Apr�es rincage
dans du TPS, les cellules �etaient incub�ees pendant 1
heure avec un anticorps conjugu�e secondaire Ale-
xaFluor488 (dilution 1:500, Invitrogen, Stockholm,
Su�ede). Finalement, les cellules �etaient rinc�ees 3� 5
min dans du TPS puis assembl�ees en utilisant le r�eactif
Prolong gold antifade (VWR, Stockholm, Su�ede)
contenant du 4’,6-diamidino-2-ph�enylindole (DAPI).
Les controles incluaient l’omission des anticorps
primaires. Les antis�erum primaires et secondaires
utilis�es dans cette �etude avaient �et�e utilis�es dans
plusieurs autres �etudes et test�es en consid�erant leurs
sp�ecificit�es et leurs r�eactivit�es crois�ees.34-39 Les�echantillons �etaient examin�es en utilisant des
microscopes �a lampes fluorescentes Olympus (Solna,
Su�ede) BX41 et IX51 (x40/0,75) munis de filtres
d’excitation appropri�es (U-MWIB2, BP 460-490
nm), puis les images �etaient captur�ees en utilisant un
appareil Olympus DP70 CCD.
Induction de la diff�erentiation
La diff�erentiation de cultures monocouches des
fibroblastes dermiques humains et des clones de
fibroblastes en cellules de type endoth�elial-like �etait
induite en modifiant le milieu de culture cellulaire
FGM en EGM, un milieu dans lequel les cellules
endoth�eliales matures maintiennent leur �etat de
diff�erentiation. La diff�erentiation �etait confirm�ee
apr�es 10 jours de culture par analyse en western
blot et analyse immunohistochimique selon le pro-
tocole d�ecrit ci-dessus pour la caract�erisation ini-
tiale des cellules endoth�eliales, des fibroblastes, et
des clones de fibroblastes.
Ensemencement des cellules sur les
matrices
Dans l’�etude pr�esent�ee, nous utilisions une matrice
bas�ee sur une matrice �a base de g�elatine porcine

722 Karlsson et al. Annales de chirurgie vasculaire
de type A hautement r�eticul�ee (Percell Biolytica,
Astorp, Su�ede). La matrice avait des pores inter-
connect�es avec une dimension interne moyenne
de 30 mm par pore, ce qui offrait des surfaces sub-
stantielles pour l’adh�esion cellulaire et facilitait la
signalisation paracrine. La matrice �etait pr�epar�ee
selon les recommandations du fabricant.
Bri�evement, la matrice d’environ 1 mm d’�epaisseur�etait hydrat�ee dans du TPS, coup�ee en pi�eces circu-
laires d’un diam�etre de 8 mm, et st�erilis�ee par un
autoclave (121 �C, 20 min, 15 psi). Les matrices
st�erilis�ees �etaient lav�ees au DMEM et plac�ees dans
des plaques de culture �a 24-puits.
Les matrices �etaient ensemenc�ees avec (1) des cel-
lules endoth�eliales cultiv�ees dans de l’EGM (avant et
apr�es ensemencement), (2) des fibroblastes et des
clones de fibroblastes cultiv�es dans du FGM (avant
et apr�es ensemencement), (3) des fibroblastes
diff�erenci�es en types cellulaires endoth�elial-like culti-
v�es dans de l’EGM (avant et apr�es ensemencement),
et (4) des fibroblastes et des clones de fibroblastes
cultiv�es dans du FGM (avant ensemencement). La
diff�erentiation de ces cellules �etait induite 24 heures
apr�es ensemencement en changeant le milieu de
culture par de l’EGM �a la place du FGM. Les cellules
endoth�eliales �etaient ensemenc�ees �a une densit�e de
200,000/cm2, alors que les fibroblastes dermiques
humains et les clones de fibroblastes �etaient
ensemenc�es �a une densit�e de 20,000/cm2.
Coloration �a l’h�ematoxylinee�eosine et
immunohistochimie des cellules
ensemenc�ees sur les matrices
Les cellules cultiv�ees sur des matrices pendant 10
jours �etaient lav�ees pendant 3 � 5 minutes dans
du TPS, imm�ediatement fix�ees dans du NBP 4%
pendant 24 heures, puis d�eshydrat�ees dans de
l’�ethanol-xyl�ene, et fix�ees dans de la paraffine. Des
coupes de 7 mm d’�epaisseur �etaient r�ealis�ees au
microtome RM 2255 (Leica Microsystem, Helsing-
borg, Su�ede). Les coupes �etaient d�eparaffin�ees,
color�ees �a l’h�ematoxylinee�eosine, et color�ees en
immunohistochimie. Pour l’analyse immunohisto-
chimique, un kit Vectastain Elite ABC (Immuno-
kemi, Jarfalla, Su�ede) �etait utilis�e pour d�etecter les
anticorps primaires. Bri�evement, les coupes �etaient
incub�ees avec du s�erum bloquant pendant 20 minu-
tes pour �eviter la liaison de prot�eines non sp�ecifi-
ques, avant d’etre incub�ees avec des anticorps
primaires dirig�es contre le vWf (dilution 1:20, clone
F8/86; Dako Cytomation), la ve-cadh�erine (dilution
1:50, clone F-8; Santa Cruz Biotechnologies), le
CD31 (dilution 1:50, clone JC/70A; Abcam); le
NOS3 (dilution 1:50, Santa Cruz Biotechnologies),
et le B2 (dilution 5 mg/mL; Sigma-Aldrich,
St. Louis, Missouri, E-U) pendant une heure.
Avant incubation des coupes avec les anticorps
dirig�es contre le CD31 ou le NOS3, deux �etapes
visant �a retrouver les antig�enes �etaient r�ealis�ees. Les
coupes color�ees avec des anticorps anti-CD31�etaient incub�ees avec une solution epsine 0,5%
pendant 10 minutes �a 37 �C. Les coupes color�ees
avec des anticorps anti-NOS3 �etaient incub�ees
pendant 20 minutes dans une solution de
r�ecup�eration cible (Dako Cytomation) pr�echauff�ee �a95 �C. Ensuite, les coupes �etaient rinc�ees pendant 3
� 5 minutes dans du TPS et incub�ees avec un anti-
corps secondaire biotinyl�e pendant 30 min. Les�echantillons �etaient lav�es �a nouveau et incub�es avec
le r�eactif Vectastain Elite ABC pendant 30 min.
Apr�es lavage dans du TPS pendant 3 � 5 min, les
anticorps fix�es �etaient localis�es avec le kit Vector VIP
(Immunokemi). Toutes les coupes �etaient contre-
color�ees au DAPI et enduites de g�elatine de glyc�erol
(Merck, Darmstadt, Allemagne). Les controles ne
contenaient pas d’anticorps primaire.
Analyse Western Blot
Un Western blot �etait r�ealis�e pour confirmer la
pr�esence de marqueurs endoth�eliaux dans les fibro-
blastes diff�erenci�es suivant un ph�enotype cellulaire
de type endoth�elial-like. Les prot�eines des cellules
cultiv�ees pendant 10 jours dans les flacons de
culture cellulaire et les cellules cultiv�ees sur les
matrices �etaient utilis�ees. Le milieu de culture cellu-
laire �etait retir�e, et les cellules �etaient lav�ees par
deux fois avec du TPS froid. Les cellules cultiv�ees
dans les flacons �etaient d�etach�ees en utilisant un
grattoir cellulaire, alors que les matrices �etaient
d�ecoup�ees en petits cubes puis trait�ees au sonica-
teur. Les suspensions cellulaires �etaient centrifug�ees�a 200 � g pendant 5 min, et le culot cellulaire �etait
incub�e avec un tampon de lyse pendant 30 minutes
sur de la glace. Les lysats cellulaires �etaient incub�es �a15000 � g pendant 5 minutes �a 4 �C, et les sur-
nageants �etaient collect�es et conserv�es �a e20 �Cavant l’analyse suivante. La teneur totale en pro-
t�eines �etait d�etermin�ee �a l’aide du Bio-Rad Protein
Assay avec de l’albumine s�erique bovine comme
standard (Bio-Rad Laboratories, Stockholm, Su�ede).
Les �echantillons utilis�es pour le Western blot�etaient bouillis �a 95 �C pendant 5 minutes apr�es
avoir ajout�e un tampon d’�echantillon de Laemmli
(Bio-Rad Laboratories) et du b-mercapto�ethanol.
L’�electrophor�ese �etait r�ealis�ee dans des gels ready
Tris/HCl 7,5% (Bio-Rad Laboratories) selon les
m�ethodes habituelles. Des montants �equivalents
d’�echantillons (30 mg de prot�eines pour les cellules

Vol. 23, No. 5, 2009 Les fibroblastes comme source de cellules endoth�eliales 723
cultiv�ees dans des flacons de cultures cellulaires et
60 mg de prot�eines pour les cellules cultiv�ees sur
les matrices) �etaient charg�es par ligne. Les gels�etaient �equilibr�es dans un tampon de transfert pen-
dant 15 minutes. Les prot�eines �etaient transf�er�ees
sur des membranes de nitrocellulose (Bio-Rad Labo-
ratories) et incub�ees dans une solution bloquante
(lait sec 5% dans 0,05 M de Trizma [pH 7,5], 0,15
M de NaCl, et du Tween 0,05%) pendant 90 minutes�a temp�erature ambiante. Ensuite, les membranes�etaient incub�ees pour la nuit �a 4 �C avec des anti-
corps primaires dirig�es contre: le vWf (1:100, clone
F8/86; Dako Cytomation) ou le NOS3 (dilution
1:200, Santa Cruz Biotechnologies) dilu�e dans un
tampon de dilution (lait sec 0,1 % dans 0,05 M de
Trizma [pH 7,5], 0,15 M de NaCl, et du Tween
0,05%). Apr�es cela, les membranes �etaient lav�ees
et incub�ees pendant une heure �a temp�erature
ambiante avec des anticorps secondaires conjugu�es�a la peroxydase de radis dilu�es �a 1:5000 dans un
tampon de dilution (lait sec 0,1% dans 0,05 M de
Trizma [pH 7,5], 0,15 M de NaCl, et du Tween
0,05%). Apr�es plusieurs lavages, les bandes �etaient
r�ev�el�ees par chemiluminescence (Amersham
ECL� Western Blotting System; GE Healthcare,
Uppsala, Su�ede). Des plaquettes humaines (40 mg
de prot�eine) �etaient utilis�ees comme controles
pour la d�etection de vWf.
R�ESULTATS
Caract�erisation initiale des ph�enotypes
cellulaires
Afin d’�eliminer le risque de contamination par
diff�erents types cellulaires au sein des cultures cellu-
laires primaires, les ph�enotypes des cellules
endoth�eliales, des fibroblastes dermiques, ainsi que
des clones de fibroblastes �etaient caract�eris�ees en
immunohistochimie, avant ensemencement des cel-
lules sur les matrices. Au cours de la culture primaire
monocouche, les cellules endoth�eliales d�emontraient
une immunocoloration positive lors de l’utilisation
d’anticorps dirig�es contre le vWf, la ve-cadh�erine, le
CD31, et le NOS3 (Fig. 1A, E, I, M), alors que cela
n’�etait pas le cas avec les fibroblastes dermiques
(Fig. 1B, F, J, N) ou les clones de fibroblastes
(r�esultats non pr�esent�es) cultiv�es sur du FGM, lors-
que le meme protocole exp�erimental �etait utilis�e.
La modification ph�enotypique des fibroblastes en
cellules de type endoth�elial-like �etait induite par
culture de fibroblastes dermiques normaux ainsi
que des clones de fibroblastes dans de l’EGM pen-
dant 10 jours. Avant ensemencement cellulaire sur
les matrices, la modification ph�enotypique �etait
confirm�ee en immunohistochimie par l’utilisation
d’anticorps primaires dirig�es contre le vWf, la
ve-cadh�erine, le CD31, et le NOS3. Une majorit�ede cellules d�emontraient une immunor�eactivit�e �ala fois cytoplasmique et extracellulaire au vWf
(Fig. 1C, D) ainsi que des localisations cyto-
plasmiques de NOS3 (Fig. 1O, P), alors qu’une
immunocoloration moins prononc�ee �etait d�etect�ee
lorsque du s�erum anti-ve-cadh�erine �etait utilis�e(Fig. 1 G, H). Les fibroblastes ou les clones de
fibroblastes dont la diff�erenciation en cellules de
type endoth�elial-like �etait induite ne d�emontraient
pas d’immunor�eactivit�e lors de la coloration des
anticorps anti-CD31 (Fig. 1 K, L). Afin de confirmer
les r�esultats de l’immunocoloration, nous avons�egalement r�ealis�e une analyse western blot. Cette
derni�ere confirmait nos pr�ec�edents r�esultats en
d�emontrant la pr�esence de vWf au sein des cellules
endoth�eliales et des fibroblastes diff�erenci�es en cel-
lules de type endoth�elial-like (Fig. 2).
Evaluation de l’organisation et de la
diff�erentiation des cellules cultiv�ees sur
les matrices
Les matrices �etaient ensemenc�ees avec (1) des cellu-
les endoth�eliales (Fig. 3A), (2) des fibroblastes et des
clones de fibroblastes cultiv�es dans du FGM
(Fig. 3B), (3) des fibroblastes diff�erenci�es en cellules
de type endoth�elial-like cultiv�ees dans de l’EGM
(Fig. 3C), et (4) des fibroblastes et des clones de
fibroblastes induits �a se diff�erencier d’abord en cel-
lules de type endoth�elial-like apr�es ensemencement
sur matrice (Fig. 3D). Afin d’�evaluer l’organisation
et la migration des cellules cultiv�ees sur matrices,
des coupes d�eparaffin�ees �etaient color�ees avec du
DAPI et de l’h�ematoxylinee�eosine. De plus, la
diff�erenciation des fibroblastes et des clones de
fibroblastes en cellules de ph�enotype endoth�elial-
like �etait confirm�ee par western blot et immunoco-
loration par des anticorps primaires dirig�es contre le
vWf, la ve-cadh�erine, le CD31, le NOS3, et le B2.
Migration des cellules dans les pores des
matrices
La coloration au DAPI r�ev�elait que tous les types cel-
lulaires �etaient capables de migrer dans les pores des
matrices. Les fibroblastes et les clones de fibroblastes
cultiv�es dans du FGM (avant et apr�es ensemence-
ment) migraient d’avantage par rapport aux cellules
endoth�eliales ou les fibroblastes diff�erenci�es en cel-
lules de type endoth�elial-like avant ensemence-
ment. Cependant, la migration cellulaire la plus
importante �etait retrouv�ee chez les fibroblastes,
dont la diff�erentiation en cellules de type
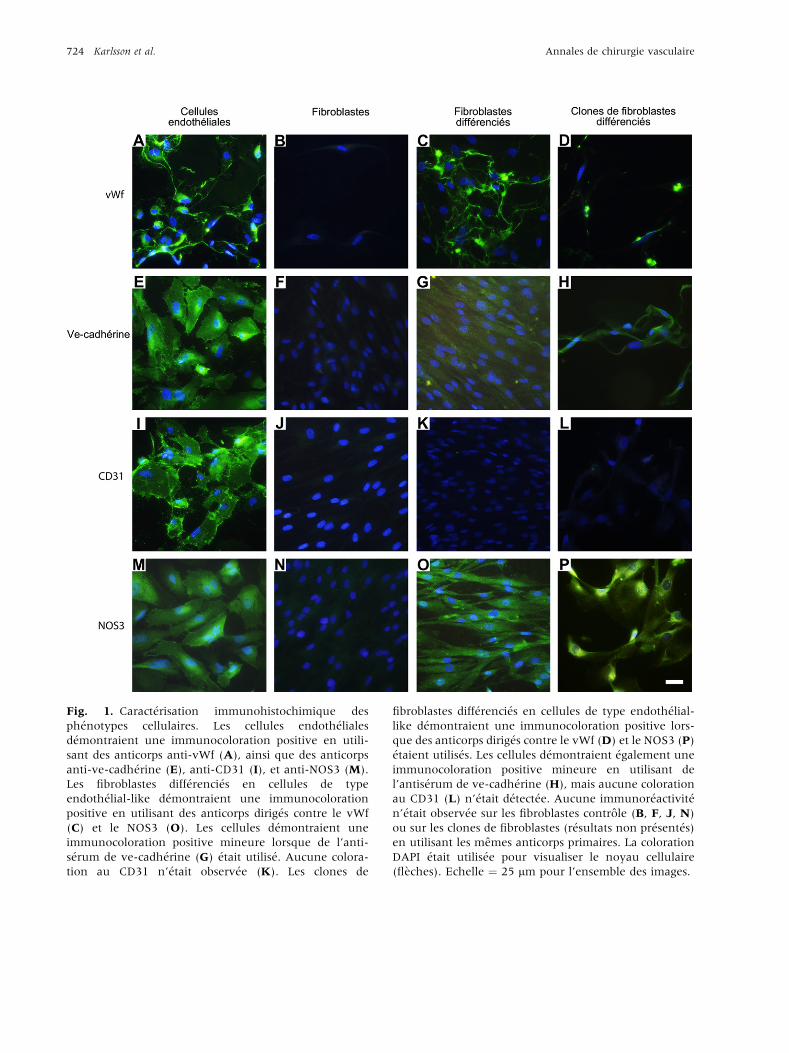
Fig. 1. Caract�erisation immunohistochimique des
ph�enotypes cellulaires. Les cellules endoth�eliales
d�emontraient une immunocoloration positive en utili-
sant des anticorps anti-vWf (A), ainsi que des anticorps
anti-ve-cadh�erine (E), anti-CD31 (I), et anti-NOS3 (M).
Les fibroblastes diff�erenci�es en cellules de type
endoth�elial-like d�emontraient une immunocoloration
positive en utilisant des anticorps dirig�es contre le vWf
(C) et le NOS3 (O). Les cellules d�emontraient une
immunocoloration positive mineure lorsque de l’anti-
s�erum de ve-cadh�erine (G) �etait utilis�e. Aucune colora-
tion au CD31 n’�etait observ�ee (K). Les clones de
fibroblastes diff�erenci�es en cellules de type endoth�elial-
like d�emontraient une immunocoloration positive lors-
que des anticorps dirig�es contre le vWf (D) et le NOS3 (P)
�etaient utilis�es. Les cellules d�emontraient �egalement une
immunocoloration positive mineure en utilisant de
l’antis�erum de ve-cadh�erine (H), mais aucune coloration
au CD31 (L) n’�etait d�etect�ee. Aucune immunor�eactivit�en’�etait observ�ee sur les fibroblastes controle (B, F, J, N)
ou sur les clones de fibroblastes (r�esultats non pr�esent�es)
en utilisant les memes anticorps primaires. La coloration
DAPI �etait utilis�ee pour visualiser le noyau cellulaire
(fl�eches). Echelle ¼ 25 mm pour l’ensemble des images.
724 Karlsson et al. Annales de chirurgie vasculaire

Fig. 2. Analyse western blot du vWf. Les cellules
endoth�eliales, les fibroblastes, et les fibroblastes
diff�erenci�es en cellules de type endoth�elial-like �etaient
analys�es en utilisant un western blot avec des anticorps
primaires dirig�es contre le vWf apr�es 10 jours de culture
dans des flacons. Un montant �equivalent de prot�eines
totales �etait analys�e dans tous les types cellulaires.
Vol. 23, No. 5, 2009 Les fibroblastes comme source de cellules endoth�eliales 725
endoth�elial-like �etait induite apr�es ensemencement
sur matrice.
Cellules endoth�eliales cultiv�ees sur
matrices
Les coupes color�ees �a l’h�ematoxylinee�eosine et au
DAPI r�ev�elaient que les cellules endoth�eliales culti-
v�ees pendant 10 jours sur matrice �etaient organis�ees
suivant une monocouche confluente. Pendant toute
l’exp�erimentation, les cellules endoth�eliales
d�emontraient une immunocoloration positive au
vWf (Fig. 4A), �a la ve-cadh�erine (Fig. 5A), au
CD31 (donn�ees non pr�esent�ees), au NOS3 (Fig. 6A),
et au B2 (Fig. 7A). De plus, l’analyse en western blot
confirmait nos pr�ec�edents r�esultats en d�emontrant
la pr�esence de NOS3 dans ces cellules (Fig. 8).
Fibroblastes et clones de fibroblastes
cultiv�es sur matrices
Les fibroblastes et les clones de fibroblastes cultiv�es
dans du FGM (avant et apr�es ensemencement) for-
maient des couches cellulaires continues, jusqu’�aune �epaisseur de trois couches cellulaires, lorsqu’ils�etaient cultiv�es sur matrice pendant 10 jours. Ces
cellules ne d�emontraient aucune immunocolora-
tion au vWf (Fig. 4B, C), �a la ve-cadh�erine
(Fig. 5B, C), au CD31 (donn�ees non pr�esent�ees), au
NOS3 (Fig. 6B, C) en utilisant le meme protocole
exp�erimental. Cependant, les fibroblastes et les
clones de fibroblastes color�es avec un s�erum anti-
r�ecepteur B2 �a la bradykinine d�emontraient une
faible immunocoloration (Fig. 7B, C). L’analyse
western blot confirmait nos pr�ec�edents r�esultats en
d�emontrant l’absence de NOS3 dans les fibroblastes
dermiques normaux (Fig. 8).
Fibroblastes diff�erenci�es en cellules de
type endoth�elial-like avant
ensemencement sur matrices
Les coupes color�ees �a l’h�ematoxylinee�eosine et au
DAPI r�ev�elaient que les fibroblastes diff�erenci�es en
cellules de type endoth�elial-like avant ensemence-
ment croissaient suivant une monocouche concen-
trique, similaire �a ce qui est observ�e au sein des
cellules endoth�eliales. Les cellules d�emontraient
une immunocoloration positive au s�erum anti-vWf
(Fig. 4E). Cependant, toutes les cellules n’expri-
maient pas ce marqueur. De plus, les cellules
d�emontraient une immunocoloration positive �a la
ve-cadh�erine (Fig. 5E), au NOS3 (Fig. 6E), et au B2
(Fig. 7E) avec un type de coloration ressemblant �a ce
qui est vu dans les cellules endoth�eliales. Il n’�etait pas
vu d’immunor�eactivit�e lorsque des anticorps anti-
CD31 �etaient utilis�es (donn�ees non pr�esent�ees). De
plus, l’analyse western blot d�emontrait la pr�esence
de NOS3 dans ces cellules (Fig. 8).
Fibroblastes ensemenc�es sur matrices
dont la diff�erenciation �etait
secondairement induite en cellules
de type endoth�elial-like
Afin de diminuer le temps de culture n�ecessaire �al’endoth�elialisation de proth�eses vasculaires, des
fibroblastes dermiques humains �etaient ensemenc�es
sur une matrice et leur diff�erentiation en cellules de
type endoth�elial-like �etait induite 24 heures plus
tard. Au contraire des fibroblastes normaux cultiv�es
dans du FGM, ces cellules formaient une multicou-
che �epaisse lorsqu’elles �etaient cultiv�ees sur matrice.
Cependant, les clones de fibroblastes trait�es de la
meme facon ne formaient pas une multicouche
mais une monocouche confluente comparable aux
cellules endoth�eliales et aux fibroblastes diff�erenci�es
en cellules de type endoth�eliales avant ensemence-
ment. L’immunocoloration des fibroblastes et des
clones de fibroblastes ensemenc�es sur les matrices
puis induits �a se diff�erencier en cellules de type
endoth�elial-like d�emontraient des r�esultats similai-
res �a ceux observ�es au sein des fibroblastes
diff�erenci�es avant ensemencement. Les cellules
d�emontraient une immunocoloration positive au
vWf (Fig. 4D, F), �a la ve-cadh�erine (Fig. 5D, F), au
NOS3 (Fig. 6D, F), et au B2 (Fig. 7D, F). Les clones
d�emontraient un important degr�e d’immunocolo-
ration �a la ve-cadh�erine. Cependant, l’intensit�e de la
coloration retrouv�ee dans les fibroblastes avant
ensemencement �etait augment�ee et le type de
coloration �etait d’avantage similaire �a celui observ�edans les cellules endoth�eliales. Aucune
immunor�eactivit�e n’�etait observ�ee lorsque des
anticorps anti-CD31 �etaient utilis�es (donn�ees non
pr�esent�ees).
Aucune immunocoloration n’�etait observ�ee dans
les �echantillons incub�es sans anticorps primaires
(controles n�egatifs, donn�ees non pr�esent�ees).

Fig. 3. Sch�ema de l’�etude. Des cellules
endoth�eliales cultiv�ees dans l’EGM pen-
dant 10 jours �etaient ensemenc�ees sur des
matrices de g�elatine et cultiv�ees dans
l’EGM pendant 10 jours suppl�ementaires
(A). Les fibroblastes dermiques humains
et les clones de fibroblastes cultiv�ees dans
le FGM pendant 10 jours �etaient
ensemenc�es sur des matrices de g�elatine
et cultiv�es dans du FGM pendant 10 jours
suppl�ementaires (B). Les fibroblastes
dermiques humains �etaient induits �a se
diff�erencier en cellules de type
endoth�elial-like par culture dans de
l’EGM 10 jours avant ensemencement sur
les matrices de g�elatine. Les cellules
�etaient cultiv�ees dans l’EGM sur la
matrice pendant 10 jours suppl�ementaires
(C). Les fibroblastes dermiques humains
�etaient cultiv�es dans du FGM pendant 10
jours puis ensemenc�es sur les matrices de
g�elatine, la diff�erenciation en cellules de
type endoth�elial-like �etait induite par le
changement du milieu de culture, du
FGM �a l’EGM (D).
726 Karlsson et al. Annales de chirurgie vasculaire
DISCUSSION
Les r�esultats obtenus dans cette �etude r�ev�elent que
les fibroblastes dermiques humains diff�erenci�es en
cellules de type endoth�elial-like formaient une
monocouche lorsqu’ils �etaient cultiv�es sur des
matrices de g�elatine, avec des cellules exprimant
les marqueurs commun�ement utilis�es pour
caract�eriser les cellules endoth�eliales matures. Les
fibroblastes ensemenc�es sur les matrices dont la
diff�erenciation en cellules de type endoth�elial-like�etait induite secondairement d�emontraient une
immunocoloration positive meme si l’organisation
de ces cellules montrait d’avantage de similarit�eavec les fibroblastes dermiques normaux que les cel-
lules endoth�eliales.
De pr�ec�edentes �etudes montraient que l’impr�e-
gnation de proth�eses synth�etiques avec des compo-
sants naturels de la matrice extra-cellulaire, comme
le collag�ene, la fibronectine, ou meme la g�elatine,
am�eliorait l’adh�esion cellulaire.2,40-42 Dans cette�etude, nous utilisions une matrice bas�ee sur de la
g�elatine porcine, une matrice ayant diff�erents
caract�eres attrayants en ing�enierie tissulaire. Par
exemple, elle est biocompatible, biod�egradable (au
sein de substances non toxiques), facilement
fabriqu�ee, et relativement peu on�ereuse. Les
r�esultats obtenus dans l’�etude actuelle
d�emontraient que les cellules endoth�eliales, les
fibroblastes, et les fibroblastes diff�erenci�es en
cellules de type endoth�elial-like �etaient cultiv�es
avec succ�es sur cette matrice. Cette matrice avait
d�ej�a �et�e utilis�ee dans de nombreuses �etudes in vitro
et in vivo, d�emontrant de bons r�esultats sur
la viabilit�e cellulaire, la migration et la prolif�era-
tion.27-32 Nos constatations soutiennent ces
r�esultats.
La pr�esence de cellules contaminantes dans la
culture fibroblastique primaire pouvait possible-
ment conduire �a un r�esultat faussement positif
concernant les capacit�es de diff�erenciation. Nous
avons corrig�e ce probl�eme de plusieurs facons: pre-
mi�erement, les cellules endoth�eliales ne sont pas
favoris�ees par le FGM; deuxi�emement, elles
adh�erent rarement aux surfaces non impr�egn�ees;
troisi�emement, l’utilisation de clones de fibroblastes
r�eduit grandement la risque de r�esultats faussement
positifs r�esultant d’une contamination; enfin, avant
toute exp�erimentation, les ph�enotypes des fibro-
blastes et des clones de fibroblastes �etaient
confirm�es par immunohistochimie indirecte utili-
sant des anticorps dirig�es contre des mol�ecules sp�eci-
fiques des cellules endoth�eliales. Puisque ni les
fibroblastes, ni les clones de fibroblastes ne mon-
traient d’immunor�eactivit�e �a ces marqueurs, le
risque d’avoir des cellules contaminantes au sein
des cultures cellulaires utilis�ees dans ces
exp�erimentations est quasiment �elimin�e.
Nos r�esultats soutiennent l’id�ee que les fibroblas-
tes dermiques humains diff�erenci�es en cellules de
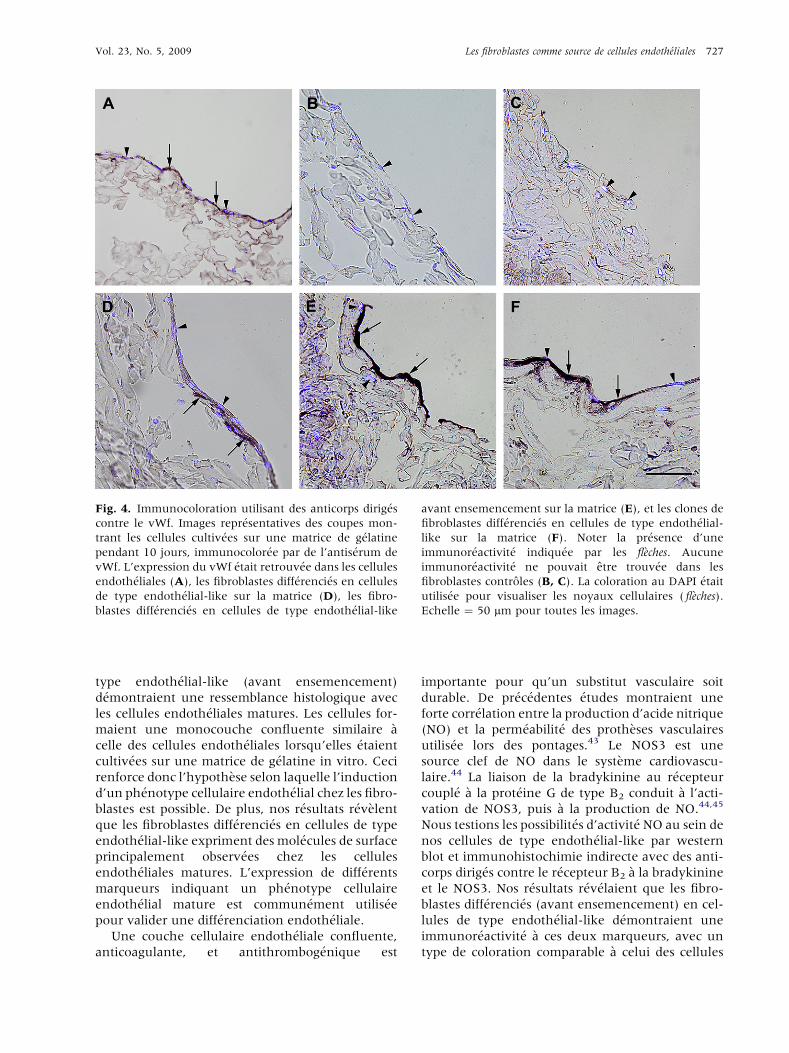
Fig. 4. Immunocoloration utilisant des anticorps dirig�es
contre le vWf. Images repr�esentatives des coupes mon-
trant les cellules cultiv�ees sur une matrice de g�elatine
pendant 10 jours, immunocolor�ee par de l’antis�erum de
vWf. L’expression du vWf �etait retrouv�ee dans les cellules
endoth�eliales (A), les fibroblastes diff�erenci�es en cellules
de type endoth�elial-like sur la matrice (D), les fibro-
blastes diff�erenci�es en cellules de type endoth�elial-like
avant ensemencement sur la matrice (E), et les clones de
fibroblastes diff�erenci�es en cellules de type endoth�elial-
like sur la matrice (F). Noter la pr�esence d’une
immunor�eactivit�e indiqu�ee par les fl�eches. Aucune
immunor�eactivit�e ne pouvait etre trouv�ee dans les
fibroblastes controles (B, C). La coloration au DAPI �etait
utilis�ee pour visualiser les noyaux cellulaires ( fl�eches).
Echelle ¼ 50 mm pour toutes les images.
Vol. 23, No. 5, 2009 Les fibroblastes comme source de cellules endoth�eliales 727
type endoth�elial-like (avant ensemencement)
d�emontraient une ressemblance histologique avec
les cellules endoth�eliales matures. Les cellules for-
maient une monocouche confluente similaire �acelle des cellules endoth�eliales lorsqu’elles �etaient
cultiv�ees sur une matrice de g�elatine in vitro. Ceci
renforce donc l’hypoth�ese selon laquelle l’induction
d’un ph�enotype cellulaire endoth�elial chez les fibro-
blastes est possible. De plus, nos r�esultats r�ev�elent
que les fibroblastes diff�erenci�es en cellules de type
endoth�elial-like expriment des mol�ecules de surface
principalement observ�ees chez les cellules
endoth�eliales matures. L’expression de diff�erents
marqueurs indiquant un ph�enotype cellulaire
endoth�elial mature est commun�ement utilis�ee
pour valider une diff�erenciation endoth�eliale.
Une couche cellulaire endoth�eliale confluente,
anticoagulante, et antithrombog�enique est
importante pour qu’un substitut vasculaire soit
durable. De pr�ec�edentes �etudes montraient une
forte corr�elation entre la production d’acide nitrique
(NO) et la perm�eabilit�e des proth�eses vasculaires
utilis�ee lors des pontages.43 Le NOS3 est une
source clef de NO dans le syst�eme cardiovascu-
laire.44 La liaison de la bradykinine au r�ecepteur
coupl�e �a la prot�eine G de type B2 conduit �a l’acti-
vation de NOS3, puis �a la production de NO.44,45
Nous testions les possibilit�es d’activit�e NO au sein de
nos cellules de type endoth�elial-like par western
blot et immunohistochimie indirecte avec des anti-
corps dirig�es contre le r�ecepteur B2 �a la bradykinine
et le NOS3. Nos r�esultats r�ev�elaient que les fibro-
blastes diff�erenci�es (avant ensemencement) en cel-
lules de type endoth�elial-like d�emontraient une
immunor�eactivit�e �a ces deux marqueurs, avec un
type de coloration comparable �a celui des cellules

Fig. 5. Immunocoloration utilisant des anticorps dirig�es
contre la ve-cadh�erine. Images repr�esentatives des cou-
pes montrant les cellules cultiv�ees sur une matrice de
g�elatine pendant 10 jours, immunocolor�ee par de l’anti-
s�erum de ve-cadh�erine. Une immunocoloration positive
�etait d�etect�ee dans les cellules endoth�eliales (A), les
fibroblastes diff�erenci�es en cellules de type endoth�elial-
like sur la matrice (D), les fibroblastes diff�erenci�es en
cellules de type endoth�elial-like avant ensemencement
sur la matrice (E), et les clones de fibroblastes diff�erenci�es
en cellules de type endoth�elial-like sur la matrice (F).
Noter la pr�esence d’une immunor�eactivit�e indiqu�ee par
les fl�eches. Aucune immunor�eactivit�e n’�etait retrouv�ee
dans les fibroblastes controles (B, C) en utilisant le meme
antis�erum. La coloration au DAPI �etait utilis�ee pour
visualiser les noyaux cellulaires ( fl�eches). Echelle ¼ 50 mm
pour toutes les images.
728 Karlsson et al. Annales de chirurgie vasculaire
endoth�eliales. De plus, la pr�esence de NOS3 au sein
de ces cellules �etait confirm�ee en utilisant une
analyse western blot. Ces r�esultats indiquent que les
fibroblastes diff�erenci�es en cellules de type
endoth�elial-like peuvent servir de couche cellulaire
fonctionnelle se substituant aux cellules
endoth�eliales. Cependant, d’autres �etudes investi-
guant les propri�et�es h�emostatiques non throm-
bog�eniques et h�emostatiques de la couche cellulaire
sont requises avant l’utilisation de ces cellules en
clinique.
Un inconv�enient de la plupart des proth�eses vas-
culaires issues de l’ing�enierie tissulaire est le temps
requis entre l’isolement cellulaire et l’implantation
proth�etique. Dans le but de r�eduire les temps de
culture n�ecessaires, nous recherchions une
possibilit�e d’ensemencer des fibroblastes dermiques
normaux sur une matrice puis de d�ebuter
secondairement la diff�erenciation en induisant un
ph�enotype cellulaire endoth�elial-like. Nos r�esultats
r�ev�elaient que les fibroblastes diff�erenci�es en cellu-
les de type endoth�elial-like sur matrice exprimaient
le vWf, la ve-cadh�erine, le NOS3, et le B2. Cepen-
dant, l’organisation des cellules r�ev�elait en fait plus
de similarit�es avec les fibroblastes dermiques nor-
maux qu’avec les cellules endoth�eliales, avec une
multicouche de cellules se d�eveloppant �a la surface
de la matrice. Ceci pourrait etre expliqu�e par le fait
que les fibroblastes ensemenc�es en premier puis
diff�erenci�es secondairement formaient d�ej�a des
structures avec une croissance de type ‘‘fibroblaste-
like’’ avant que le ph�enotype soit modifi�e. Quelque
peu en contradiction avec cela, il y a le type de
croissance des clones de fibroblastes diff�erenci�es sur
matrice, qui d�emontrait une morphologie mono-
cellulaire. De plus, les clones montraient �egalement
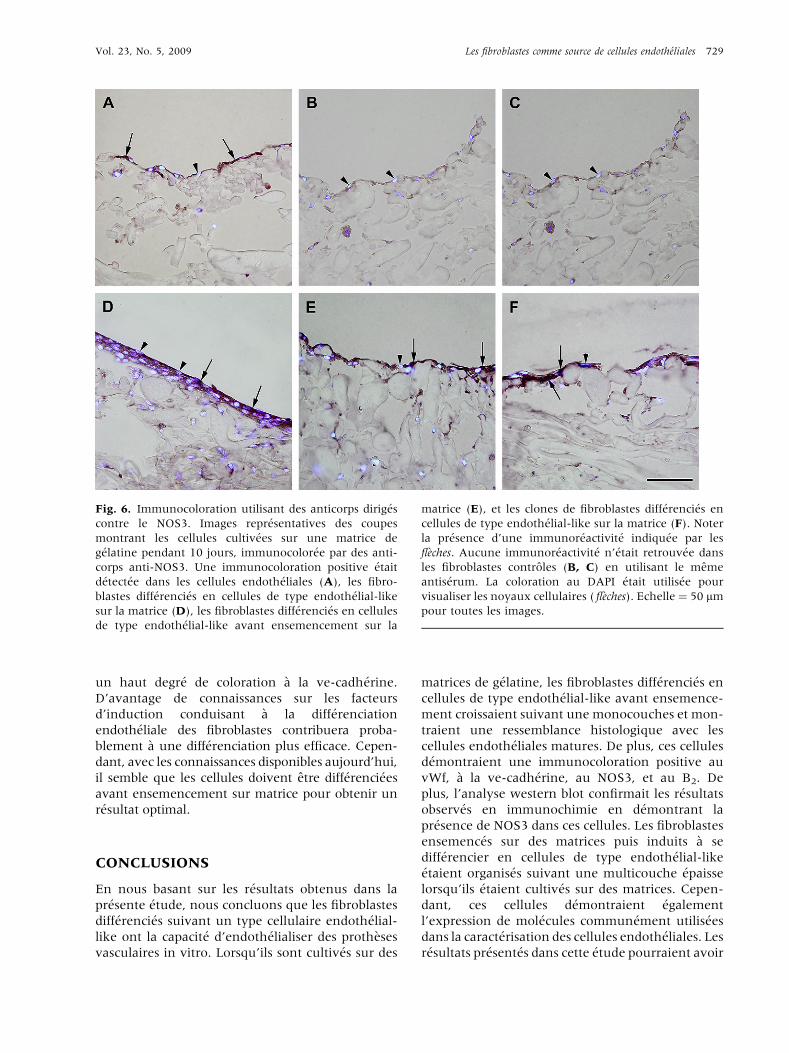
Fig. 6. Immunocoloration utilisant des anticorps dirig�es
contre le NOS3. Images repr�esentatives des coupes
montrant les cellules cultiv�ees sur une matrice de
g�elatine pendant 10 jours, immunocolor�ee par des anti-
corps anti-NOS3. Une immunocoloration positive �etait
d�etect�ee dans les cellules endoth�eliales (A), les fibro-
blastes diff�erenci�es en cellules de type endoth�elial-like
sur la matrice (D), les fibroblastes diff�erenci�es en cellules
de type endoth�elial-like avant ensemencement sur la
matrice (E), et les clones de fibroblastes diff�erenci�es en
cellules de type endoth�elial-like sur la matrice (F). Noter
la pr�esence d’une immunor�eactivit�e indiqu�ee par les
fl�eches. Aucune immunor�eactivit�e n’�etait retrouv�ee dans
les fibroblastes controles (B, C) en utilisant le meme
antis�erum. La coloration au DAPI �etait utilis�ee pour
visualiser les noyaux cellulaires ( fl�eches). Echelle ¼ 50 mm
pour toutes les images.
Vol. 23, No. 5, 2009 Les fibroblastes comme source de cellules endoth�eliales 729
un haut degr�e de coloration �a la ve-cadh�erine.
D’avantage de connaissances sur les facteurs
d’induction conduisant �a la diff�erenciation
endoth�eliale des fibroblastes contribuera proba-
blement �a une diff�erenciation plus efficace. Cepen-
dant, avec les connaissances disponibles aujourd’hui,
il semble que les cellules doivent etre diff�erenci�ees
avant ensemencement sur matrice pour obtenir un
r�esultat optimal.
CONCLUSIONS
En nous basant sur les r�esultats obtenus dans la
pr�esente �etude, nous concluons que les fibroblastes
diff�erenci�es suivant un type cellulaire endoth�elial-
like ont la capacit�e d’endoth�elialiser des proth�eses
vasculaires in vitro. Lorsqu’ils sont cultiv�es sur des
matrices de g�elatine, les fibroblastes diff�erenci�es en
cellules de type endoth�elial-like avant ensemence-
ment croissaient suivant une monocouches et mon-
traient une ressemblance histologique avec les
cellules endoth�eliales matures. De plus, ces cellules
d�emontraient une immunocoloration positive au
vWf, �a la ve-cadh�erine, au NOS3, et au B2. De
plus, l’analyse western blot confirmait les r�esultats
observ�es en immunochimie en d�emontrant la
pr�esence de NOS3 dans ces cellules. Les fibroblastes
ensemenc�es sur des matrices puis induits �a se
diff�erencier en cellules de type endoth�elial-like�etaient organis�es suivant une multicouche �epaisse
lorsqu’ils �etaient cultiv�es sur des matrices. Cepen-
dant, ces cellules d�emontraient �egalement
l’expression de mol�ecules commun�ement utilis�ees
dans la caract�erisation des cellules endoth�eliales. Les
r�esultats pr�esent�es dans cette �etude pourraient avoir
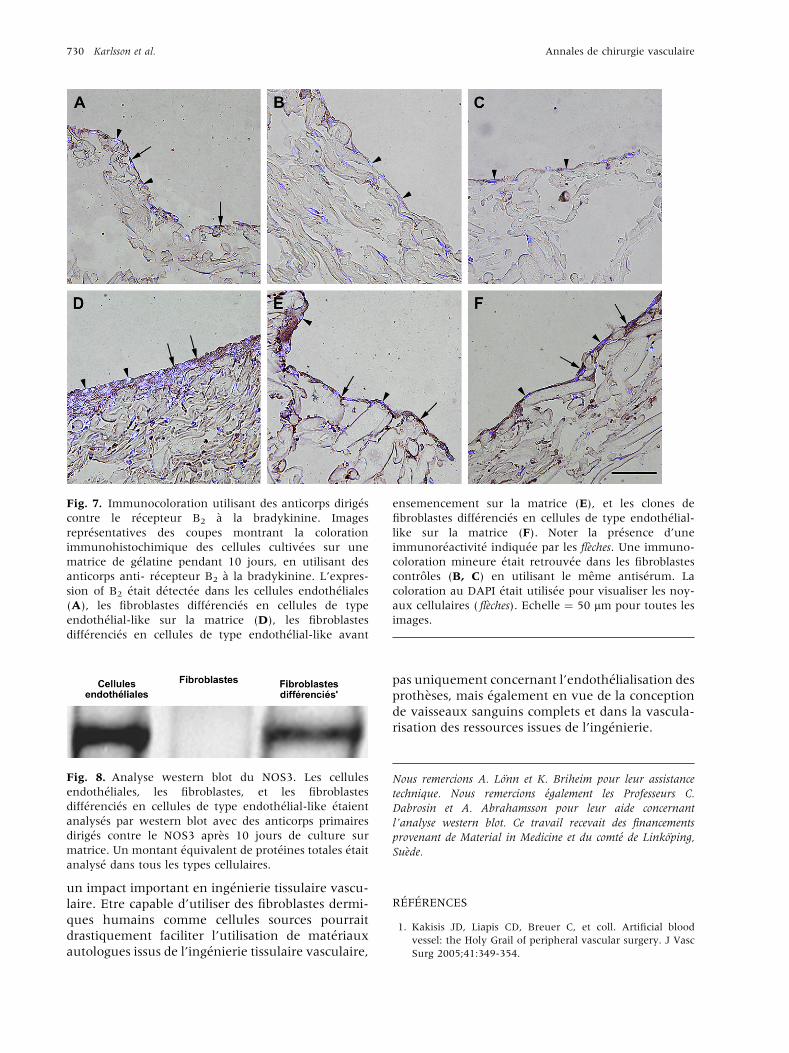
Fig. 7. Immunocoloration utilisant des anticorps dirig�es
contre le r�ecepteur B2 �a la bradykinine. Images
repr�esentatives des coupes montrant la coloration
immunohistochimique des cellules cultiv�ees sur une
matrice de g�elatine pendant 10 jours, en utilisant des
anticorps anti- r�ecepteur B2 �a la bradykinine. L’expres-
sion of B2 �etait d�etect�ee dans les cellules endoth�eliales
(A), les fibroblastes diff�erenci�es en cellules de type
endoth�elial-like sur la matrice (D), les fibroblastes
diff�erenci�es en cellules de type endoth�elial-like avant
ensemencement sur la matrice (E), et les clones de
fibroblastes diff�erenci�es en cellules de type endoth�elial-
like sur la matrice (F). Noter la pr�esence d’une
immunor�eactivit�e indiqu�ee par les fl�eches. Une immuno-
coloration mineure �etait retrouv�ee dans les fibroblastes
controles (B, C) en utilisant le meme antis�erum. La
coloration au DAPI �etait utilis�ee pour visualiser les noy-
aux cellulaires ( fl�eches). Echelle ¼ 50 mm pour toutes les
images.
Fig. 8. Analyse western blot du NOS3. Les cellules
endoth�eliales, les fibroblastes, et les fibroblastes
diff�erenci�es en cellules de type endoth�elial-like �etaient
analys�es par western blot avec des anticorps primaires
dirig�es contre le NOS3 apr�es 10 jours de culture sur
matrice. Un montant �equivalent de prot�eines totales �etait
analys�e dans tous les types cellulaires.
730 Karlsson et al. Annales de chirurgie vasculaire
un impact important en ing�enierie tissulaire vascu-
laire. Etre capable d’utiliser des fibroblastes dermi-
ques humains comme cellules sources pourrait
drastiquement faciliter l’utilisation de mat�eriaux
autologues issus de l’ing�enierie tissulaire vasculaire,
pas uniquement concernant l’endoth�elialisation des
proth�eses, mais �egalement en vue de la conception
de vaisseaux sanguins complets et dans la vascula-
risation des ressources issues de l’ing�enierie.
Nous remercions A. Lonn et K. Briheim pour leur assistance
technique. Nous remercions �egalement les Professeurs C.
Dabrosin et A. Abrahamsson pour leur aide concernant
l’analyse western blot. Ce travail recevait des financements
provenant de Material in Medicine et du comt�e de Linkoping,
Su�ede.
R�EF�ERENCES
1. Kakisis JD, Liapis CD, Breuer C, et coll. Artificial blood
vessel: the Holy Grail of peripheral vascular surgery. J Vasc
Surg 2005;41:349-354.

Vol. 23, No. 5, 2009 Les fibroblastes comme source de cellules endoth�eliales 731
2. Kannan RY, Salacinski HJ, Butler PE, et coll. Current status
of prosthetic bypass grafts: a review. J Biomed Mater Res B
Appl Biomater 2005;74:570-581.
3. Hoenig MR, Campbell GR, Rolfe BE, et coll. Tissue-engi-
neered blood vessels: alternative to autologous grafts?
Arterioscler Thromb Vasc Biol 2005;25:1128-1134.
4. Langer R, Vacanti JP. Ing�eni�erie tissulaire. Science
1993;260:920-926.
5. Meinhart JG, Deutsch M, Fischlein T, et coll. Clinical auto-
logous in vitro endothelialization of 153 infrainguinal ePTFE
grafts. Ann Thorac Surg 2001;71:S327-S331.
6. Niklason LE, Gao J, Abbott WM, et coll. Functional arteries
grown in vitro. Science 1999;284:489-493.
7. Ratcliffe A. Ing�eni�erie tissulaire of proth�eses vasculaires.
Matrix Biol 2000;19:353-357.
8. L’Heureux N, Paquet S, Labbe R, et coll. A completely bio-
logical tissue-engineered human blood vessel. FASEB J
1998;12:47-56.
9. Herring M, Baughman S, Glover J, et coll. Endothelial
seeding of Dacron and polytetrafluoroethylene grafts: the
cellular events of healing. Surgery 1984;96:745-755.
10. Schmidt SP, Hunter TJ, Sharp WV, et coll. Endothelial cell-
seeded four-millimeter Dacron proth�eses vasculaires: effects
of blood flow manipulation through the grafts. J Vasc Surg
1984;1:434-441.
11. Swedenborg J, Bengtsson L, Clyne N, et coll. In vitro
endothelialisation of arteriovenous loop grafts for haemo-
dialysis. Eur J Vasc Endovasc Surg 1997;13:272-277.
12. Hoenig MR, Campbell GR, Campbell JH. Proth�eses vascu-
laires and the endothelium. Endothelium 2006;13:385-401.
13. Kim S, von Recum H. Endothelial stem cells and precursors
for ing�eni�erie tissulaire: cell source, differentiation, selec-
tion, and application. Tissue Eng B Rev 2008;14:133-147.
14. Riha GM, Lin PH, Lumsden AB, et coll. Application of stem
cells for ing�eni�erie tissulaire vasculaire. Tissue Eng 2005;11:
1535-1552.
15. Pittenger MF, Mackay AM, Beck SC, et coll. Multilineage
potential of adult human mesenchymal stem cells. Science
1999;284:143-147.
16. Zuk PA, Zhu M, Ashjian P, et coll. Human adipose tissue is a
source of multipotent stem cells. Mol Biol Cell 2002;13:
4279-4295.
17. Zuk PA, Zhu M, Mizuno H, et coll. Multilineage cells from
human adipose tissue: implications for cell-based therapies.
Tissue Eng 2001;7:211-228.
18. Caplan AI. Adult mesenchymal stem cells for ing�eni�erie
tissulaire versus regenerative medicine. J Cell Physiol
2007;213:341-347.
19. Young HE, Black AC, Jr. Adult stem cells. Anat. Rec. A
Discov. Mol Cell Evol Biol 2004;276:75-102.
20. Young HE, Duplaa C, Young TM, et coll. Clonogenic analysis
reveals reserve stem cells in postnatal mammals: I. Pluri-
potent mesenchymal stem cells. Anat Rec 2001;263:
350-360.
21. Bartsch G, Yoo JJ, De Coppi P, et coll. Propagation, expan-
sion, and multilineage differentiation of human somatic
stem cells from dermal progenitors. Stem Cells Dev 2005;14:
337-348.
22. Young HE, Steele TA, Bray RA, et coll. Human reserve plu-
ripotent mesenchymal stem cells are present in the connec-
tive tissues of skeletal muscle and dermis derived from fetal,
adult, and geriatric donors. Anat Rec 2001;264:51-62.
23. Chunmeng S, Tianmin C. Skin: a promising reservoir for
adult stem cell populations. Med Hypotheses 2004;62:
683-688.
24. Junker J, Sommar P, Skog M, et coll. Adipogenic, chon-
drogenic and osteogenic differentiation of clonally derived
fibroblastes dermiques humains. Cells Tissues Organs 2009
(in press).
25. Sommar P, Pettersson S, Ness C, Johnson H, Kratz G,
Junker JP, et coll. Engineering three-dimensional cartilage-
and bone-like tissues using fibroblastes dermiques humains
and macroporous gelatine microcarriers. J Plast Reconstr
Aesthet Surg 2009. March 28. [Epub ahead of print].
26. Bachetti T, Morbidelli L. Cellules endoth�eliales in culture: a
model for studying vascular functions. Pharmacol Res
2000;42:9-19.
27. Huss FR, Junker JP, Johnson H, et coll. Macroporous gela-
tine spheres as culture substrate, transplantation vehicle,
and biodegradable scaffold for guided regeneration of soft
tissues. In vivo study in nude mice. J Plast Reconstr Aesthet
Surg 2007;60:543-555.
28. Liu JY, Hafner J, Dragieva G, et coll. Autologous cultured
keratinocytes on porcine gelatin microbeads effectively heal
chronic venous leg ulcers. Wound Repair Regen 2004;12:
148-156.
29. Stover NP, Bakay RA, Subramanian T, et coll. Intrastriatal
implantation of human retinal pigment epithelial cells
attached to microcarriers in advanced Parkinson disease.
Arch Neurol 2005;62:1833-1837.
30. Fernandes AM, Fernandes TG, Diogo MM, et coll. Mouse
embryonic stem cell expansion in a microcarrier-based
stirred culture system. J Biotechnol 2007;132:227-236.
31. Del Guerra S, Bracci C, Nilsson K, et coll. Entrapment of
dispersed pancreatic islet cells in CultiSpher-S macroporous
gelatin microcarriers: preparation, in vitro characterization,
and microencapsulation. Biotechnol Bioeng 2001;75:
741-744.
32. Liu JY, Hafner J, Dragieva G, et coll. High yields of auto-
logous living dermal equivalents using porcine gelatin
microbeads as microcarriers for autologous fibroblasts. Cell
Transplant 2006;15:445-451.
33. Jaffe EA, Nachman RL, Becker CG, et coll. Culture of human
cellules endoth�eliales derived from umbilical veins. Identi-
fication by morphologic and immunologic criteria. J Clin
Invest 1973;52:2745-2756.
34. Denis CV. Molecular and cellular biology of von Willebrand
factor. Int J Hematol 2002;75:3-8.
35. Werner M, Kaloutsi V, Walter K, et coll. Immunohisto-
chemical examination of routinely processed bone marrow
biopsies. Pathol Res Pract 1992;188:707-713.
36. Kastenbauer S, Klein M, Koedel U, et coll. Reactive nitrogen
species contribute to bloodelabyrinth barrier disruption in
suppurative labyrinthitis complicating experimental pneu-
mococcal meningitis in the rat. Brain Res 2001;904:
208-217.
37. Noda M, Kariura Y, Amano T, et coll. Kinin receptors in
cultured rat microglia. Neurochem Int 2004;45:437-442.
38. Li JM, Shah AM. Intracellular localization and preassembly
of the NADPH oxidase complex in cultured cellules
endoth�eliales. J Biol Chem 2002;277:19952-19960.
39. Moriyama M, Kumagai S, Kawashiri S, et coll. Immuno-
histochemical study of tumour angiogenesis in oral squa-
mous cell carcinoma. Oral Oncol 1997;33:369-374.
40. Pankajakshan D, Krishnan VK, Krishnan LK. Vascular tissue
generation in response to signaling molecules integrated
with a novel poly(epsilon-caprolactone)-fibrin hybrid scaf-
fold. J Tissue Eng Regen Med 2007;1:389-397.
41. He W, Ma Z, Yong T, et coll. Fabrication of collagen-coated
biodegradable polymer nanofiber mesh and its potential for

732 Karlsson et al. Annales de chirurgie vasculaire
cellules endoth�eliales growth. Biomaterials 2005;26:
7606-7615.
42. Ma Z, He W, Yong T, et coll. Grafting of gelatin on electro-
spun poly(caprolactone) nanofibers to improve endothelial
cell spreading and proliferation and to control cell orienta-
tion. Tissue Eng 2005;11:1149-1158.
43. Amiel GE, Komura M, Shapira O, et coll. Engineering of
blood vessels from acellular collagen matrices coated with
human cellules endoth�eliales. Tissue Eng 2006;12:
2355-2365.
44. Dudzinski DM, Igarashi J, Greif D, et coll. The regulation
and pharmacology of endothelial nitric oxide synthase.
Annu. Rev. Pharmacol. Toxicol 2006;46:235-276.
45. Govers R, Rabelink TJ. Cellular regulation of endothelial
nitric oxide synthase. Am J Physiol Renal Physiol 2001;280:
F193-F206.





























