Embed Size (px)
Citation preview

1
Filières énergétiques lors
de l’exercice et de l’entraînement
Filières énergétiques lors
de l’exercice et de l’entraînement
Par Clément GALIEN
D’après les données deGeorges CAZORLA
Laboratoire Evaluation Sport Santé Université Victor SegalenBordeaux 2
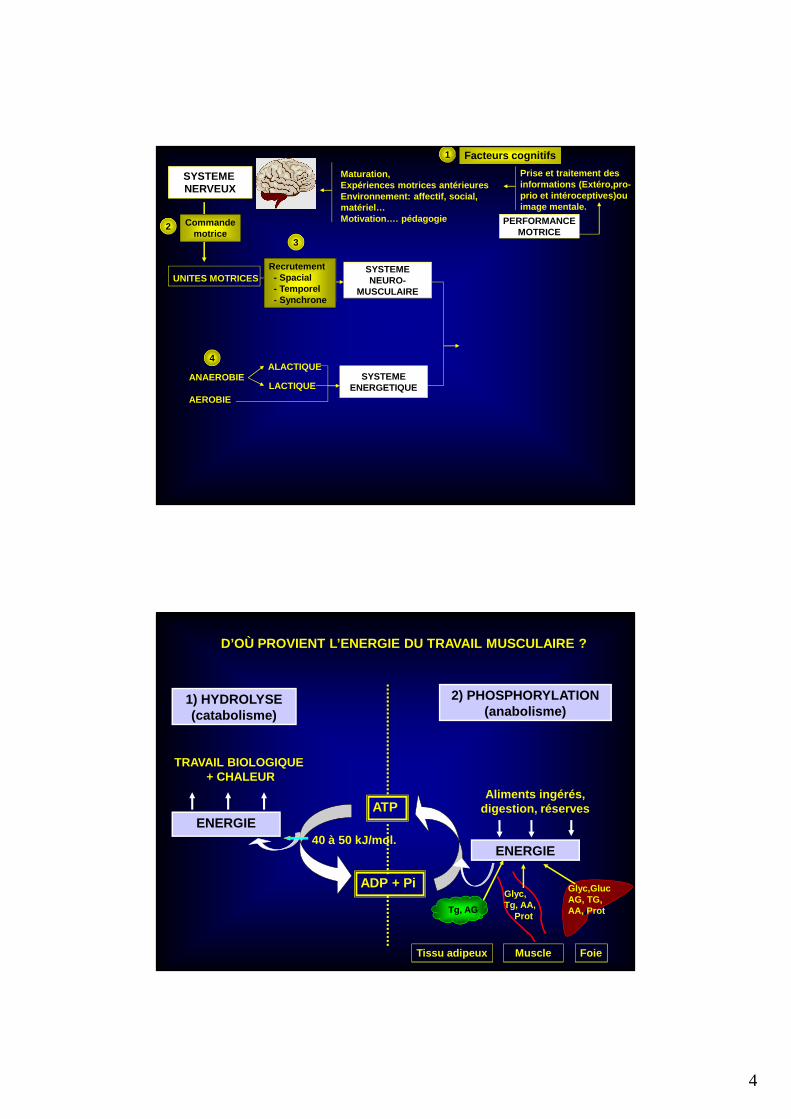
SYSTEME NERVEUX
UNITES MOTRICESSYSTEMENEURO-
MUSCULAIRE
PERFORMANCEMOTRICE
Maturation, Expériences motrices antérieuresEnvironnement: affectif, social, matériel…Motivation…. pédagogie
1
2 Commandemotrice
3
7
Prise et traitement des informations (Extéro,pro-prio et intéroceptives)ou image mentale.
Facteurs cognitifs
Recrutement- Spacial- Temporel- Synchrone

2
L’étude des facteurs physiologiques de la motricité del’enfant et de l’adolescent au cours de l’exercice se
heurte à des difficutés de trois ordres :
EthiqueEthique : Très peu de données recueilies expérimentalementchez les enfants surtout âgés de moins de 11 ans et les séniors.
MéthodologiqueMéthodologique :1- Difficile de distinguer ce qui revient, d’une part à la croissance, au niveau habituel d’ac tivité individuelle et d’autre part, à l’entraînement cont rôlé.
SubjectiveSubjective : L’enfant plus que l’adulte éprouve des difficutés à bien respecter les consignes et répugne à l’effo rt maximal voire supramaximal qui conduit à l’épuisement.
2 - Les études longitudinales sont très rares et ne sont qu’exceptionnellement comparatives à un groupe témoin.
CAPITALGENETIQUE(APTITUDEMOTRICE)
NIVEAU D’ACTIVITEET D’ENTRAINEMENT
ETAT DE SANTE
INTEGRATEUR
CAPACITEMOTRICE
DIETETIQUEFATIGUEMEDICAMENTSBLESSURES
PSYCHOLOGIE PHYSIOLOGIEBIOLOGIEBIOMECANIQUE
AGE
PRODUITPROCESSUS(FONCTIONNEMENT)
INTERACTIONS
PERFORMANCEMOTRICE
D’après Cazorla, 1999

3
CROISSANCECROISSANCE ++ MATURATIONMATURATION
Mesures Mesures biométriquesbiométriquesstandardiséesstandardisées
EVALUATION NORMATIVEEVALUATION NORMATIVE
Appréciation des stades atteints Appréciation des stades atteints dans les étapes critiques du dans les étapes critiques du développement ontogénétiquedéveloppement ontogénétique
OBSERVATIONS LONGITUDINALESOBSERVATIONS LONGITUDINALES
DEVELOPPEMENTDEVELOPPEMENTMOTEURMOTEUR
APPRENTISSAGESAPPRENTISSAGESMOTEURSMOTEURS
++
Caractérisés par les capacitésCaractérisés par les capacitéscognitives qui ne peuvent êtrecognitives qui ne peuvent êtreévaluées qu’au cours des difféévaluées qu’au cours des diffé--rents apprentissages. Les testsrents apprentissages. Les testsutilisés dépendent des contenusutilisés dépendent des contenusde ces apprentissagesde ces apprentissages
EVALUATION FORMATIVEEVALUATION FORMATIVE
Peut être caractérisé par les capaPeut être caractérisé par les capa--cités motrices évaluées par des cités motrices évaluées par des tests ou des batteries de tests stantests ou des batteries de tests stan--dardisésdardisés
EVALUATION NORMATIVE OUEVALUATION NORMATIVE OUNORMATIVONORMATIVO--CRITERIEECRITERIEE
COMPORTEMENT MOTEURCOMPORTEMENT MOTEUR
COMPORTEMENT MOTEURCOMPORTEMENT MOTEUR
Dépend du nombre et de la qualitéDépend du nombre et de la qualitédes apprentissages moteursdes apprentissages moteursantérieurs, des capacités motricesantérieurs, des capacités motriceset des capacités cognitives.et des capacités cognitives.ECHELLES D’APPRECIATION ET ECHELLES D’APPRECIATION ET TAXINOMIES PREALABLEMENTTAXINOMIES PREALABLEMENTDETERMINEES.DETERMINEES.OBSERVATION ORGANISEEOBSERVATION ORGANISEE
ENVIRONNEMENTENVIRONNEMENT
Caractérisé par des variables Caractérisé par des variables inconnues et constamment eninconnues et constamment enévolution aléatoire (ex: matchévolution aléatoire (ex: matchou situations pédagogiques ou situations pédagogiques contrôlées).contrôlées).
GRILLES D’OBSERVATION GRILLES D’OBSERVATION
++
CAPACITE D’ADAPTATIONCAPACITE D’ADAPTATION
Dépend de la “plasticité” des Dépend de la “plasticité” des comportements moteurs et descomportements moteurs et dessituations de l’environnementsituations de l’environnement
OBSERVATION ORGANISEE. OBSERVATION ORGANISEE. GodboutGodbout, 1988, 1988
4
SYSTEME NERVEUX
UNITES MOTRICESSYSTEMENEURO-
MUSCULAIRE
SYSTEMEENERGETIQUE
ANAEROBIE
AEROBIE
ALACTIQUE
LACTIQUE
PERFORMANCEMOTRICE
Maturation, Expériences motrices antérieuresEnvironnement: affectif, social, matériel…Motivation…. pédagogie
1
2 Commandemotrice
3
4
Prise et traitement des informations (Extéro,pro-prio et intéroceptives)ou image mentale.
Facteurs cognitifs
Recrutement- Spacial- Temporel- Synchrone
ENERGIE
ENERGIE
Aliments ingérés, digestion, réserves
1) HYDROLYSE (catabolisme)
2) PHOSPHORYLATION (anabolisme)
TRAVAIL BIOLOGIQUE + CHALEUR
ATP
ADP + Pi
D’OÙ PROVIENT L’ENERGIE DU TRAVAIL MUSCULAIRE ?
40 à 50 kJ/mol.
MuscleTissu adipeux Foie
Tg, AG
Glyc,Tg, AA,
Prot
Glyc,GlucAG, TG, AA, Prot
5
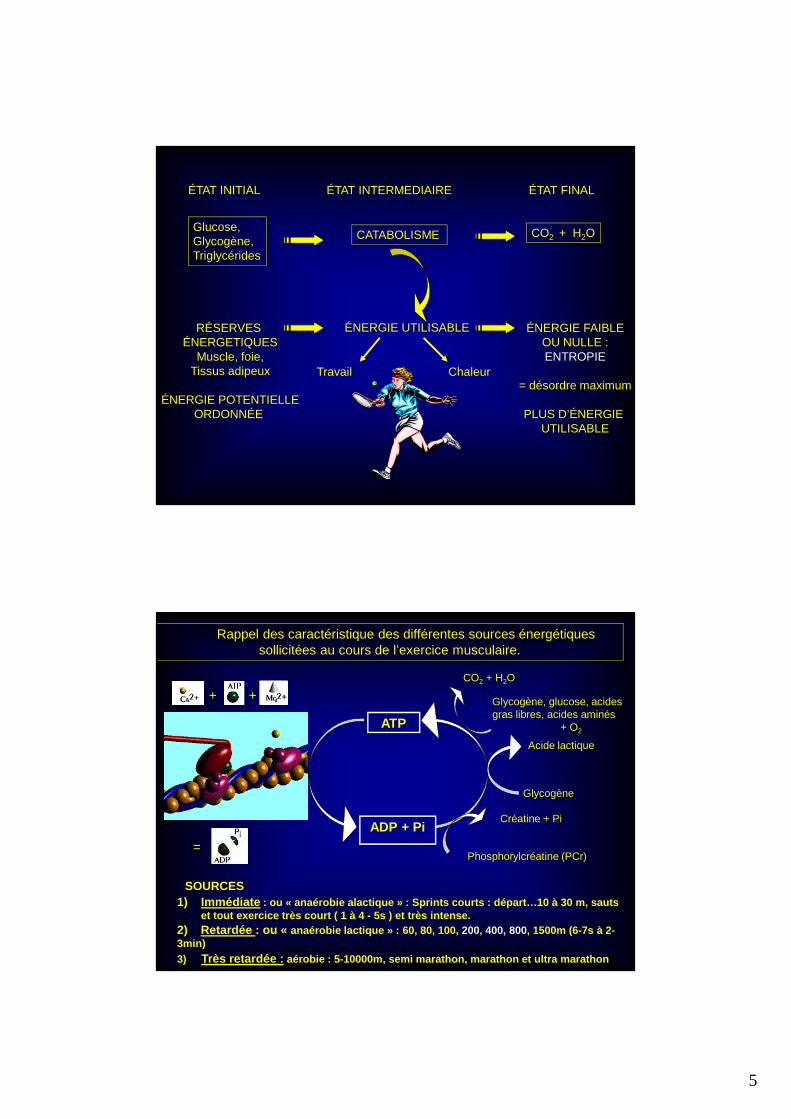
ÉTAT INITIAL ÉTAT INTERMEDIAIRE ÉTAT FINAL
Glucose,Glycogène,Triglycérides
CATABOLISME CO2 + H2O
ÉNERGIE UTILISABLERÉSERVES ÉNERGETIQUES
Muscle, foie,Tissus adipeux
ÉNERGIE POTENTIELLEORDONNÉE
ÉNERGIE FAIBLEOU NULLE :ENTROPIE
= désordre maximum
PLUS D’ÉNERGIE UTILISABLE
Travail Chaleur
ATP
ADP + PiCréatine + Pi
Phosphorylcréatine (PCr)
Rappel des caractéristique des différentes sources énergétiques sollicitées au cours de l’exercice musculaire.
SOURCES1) Immédiate : ou « anaérobie alactique » : Sprints courts : départ …10 à 30 m, sauts
et tout exercice très court ( 1 à 4 - 5s ) et très i ntense.
Glycogène
Acide lactique
2) Retardée : ou « anaérobie lactique » : 60, 80, 100, 200, 400, 800, 1500m (6-7s à 2-3min)
++
=
CO2 + H2O
3) Très retardée : aérobie : 5-10000m, semi marathon, marathon et ultr a marathon
Glycogène, glucose, acides gras libres, acides aminés
+ O2
6
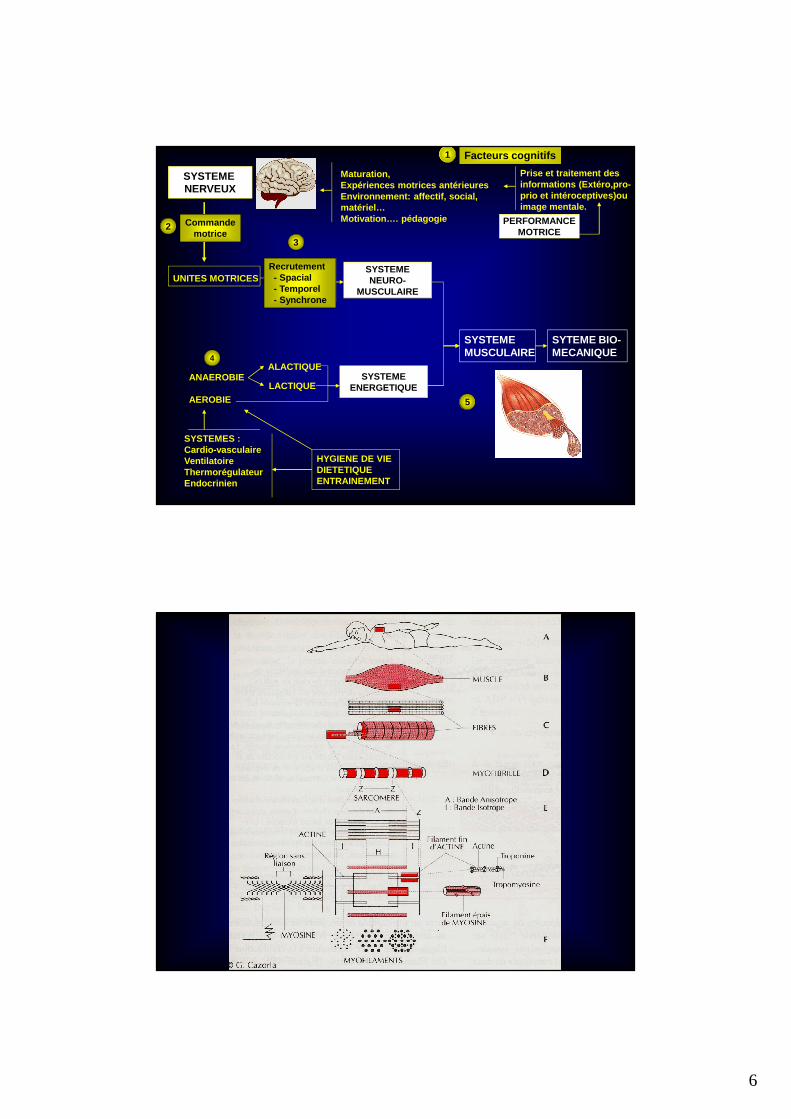
SYSTEME NERVEUX
UNITES MOTRICESSYSTEMENEURO-
MUSCULAIRE
SYSTEMEENERGETIQUE
ANAEROBIE
AEROBIE
ALACTIQUE
LACTIQUE
PERFORMANCEMOTRICE
Maturation, Expériences motrices antérieuresEnvironnement: affectif, social, matériel…Motivation…. pédagogie
SYSTEMES : Cardio-vasculaireVentilatoireThermorégulateurEndocrinien
HYGIENE DE VIEDIETETIQUEENTRAINEMENT
1
2 Commandemotrice
3
5
4
Prise et traitement des informations (Extéro,pro-prio et intéroceptives)ou image mentale.
Facteurs cognitifs
Recrutement- Spacial- Temporel- Synchrone
SYSTEMEMUSCULAIRE
SYTEME BIO-MECANIQUE

7
Le sarcomère et la myofibrille
FONDEMENTS BIOENERGETIQUES DE L’EXERCICE ET DE L’ENTRAINEMENT
8
ENERGIE
ENERGIE
Aliments ingérés, digestion, réserves
1) HYDROLYSE (catabolisme)
2) PHOSPHORYLATION (anabolisme)
TRAVAIL BIOLOGIQUE + CHALEUR
ATP
ADP + Pi
D’OU PROVIENT L’ENERGIE DU TRAVAIL MUSCULAIRE ?
40 kJ/mol.
MuscleTissu adipeux Foie
TglyGlyc,Tgly
Glyc,Gluc
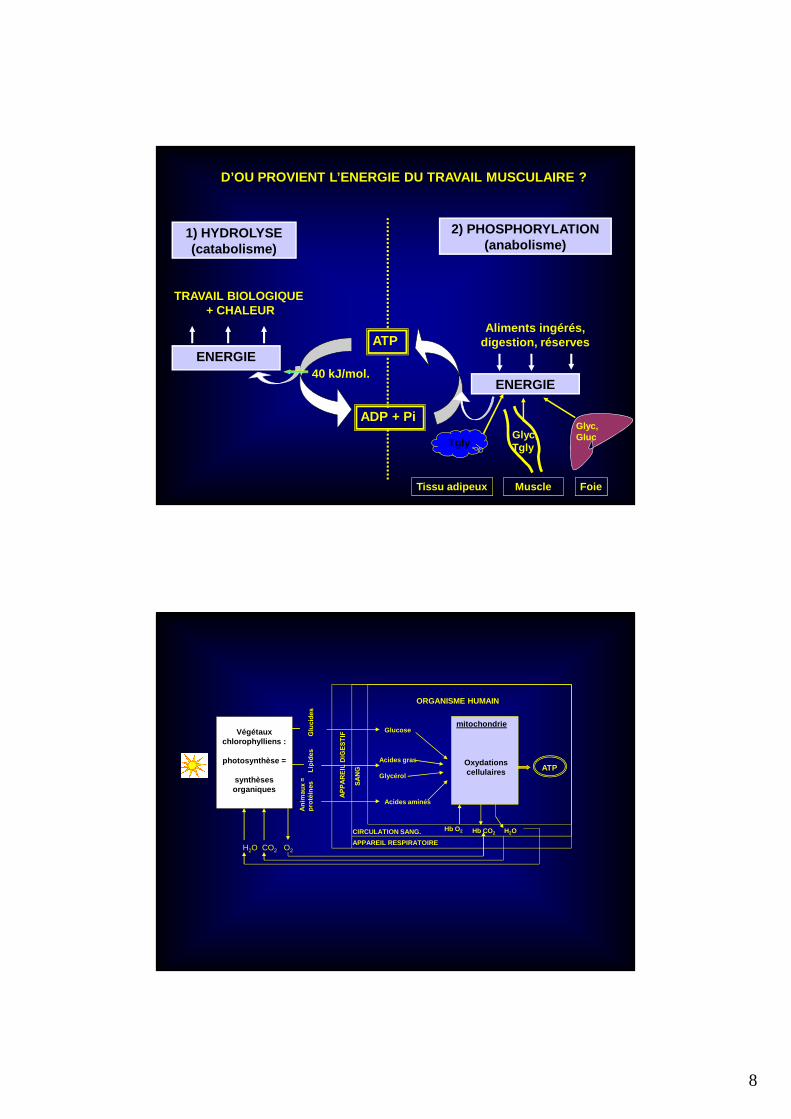
Végétauxchlorophylliens :
photosynthèse =
synthèses organiques
Glucose
Acides gras
Glycérol
Acides aminés
Oxydationscellulaires
mitochondrie
CIRCULATION SANG.
APPAREIL RESPIRATOIREH2O CO2 O2
Hb O2 Hb CO2 H2O
ATP
ORGANISME HUMAIN
9
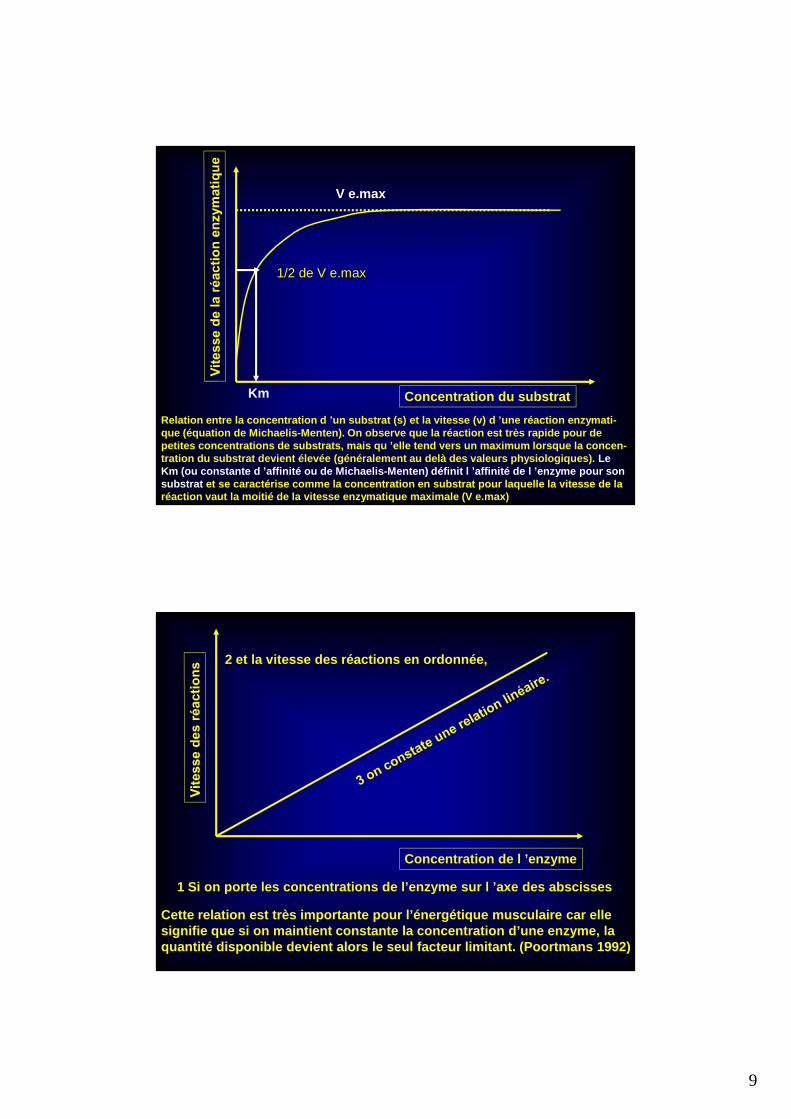
Relation entre la concentration d ’un substrat (s) e t la vitesse (v) d ’une réaction enzymati-que (équation de Michaelis-Menten). On observe que la réaction est très rapide pour de petites concentrations de substrats, mais qu ’elle t end vers un maximum lorsque la concen-tration du substrat devient élevée (généralement au delà des valeurs physiologiques). Le Km (ou constante d ’affinité ou de Michaelis-Menten) définit l ’affinité de l ’enzyme pour son substrat et se caractérise comme la concentration en substra t pour laquelle la vitesse de la réaction vaut la moitié de la vitesse enzymatique m aximale (V e.max)
Km Concentration du substrat
V e.max
1/2 de V e.max
Concentration de l ’enzyme
Cette relation est très importante pour l’énergétiq ue musculaire car elle signifie que si on maintient constante la concentra tion d’une enzyme, la quantité disponible devient alors le seul facteur l imitant. (Poortmans 1992)
1 Si on porte les concentrations de l’enzyme sur l ’ axe des abscisses
2 et la vitesse des réactions en ordonnée,

10
Conséquence...
hhhh or nous savons que, dans certains cas, les effets d e l’entraînement vont se traduire essentiellement par l’augmentation de la quantité d’une ou de plusieurs enzymes en présence.
hhhhautrement dit, plus on a d’enzyme(s), plus on pourra dégrader de substrat.
hhhhc’est précisément le cas des athlètes entraînés à l’endurance dont l’entraînement a augmenté le nombr e,la taille des mitochondries et par conséquent la co ncen-tration en enzymes oxydatives.
HYDROLYSE
ATP
ADP
Energie
Travail 25%
Chaleur 75%
Mécanique ( muscle )Circulation sanguineDigestion ChimiqueOsmotiqueSécrétions glandulairesProduction de tissuTransmission nerveuseet musculaire
Différentes formes de travail biologique que permet l’énergie libérée par l’hydrolyse de l’ATP

11
TRIPHOSPHATE
0 0 0 II II II
ADENOSINE 0 P ~ 0 P ~ 0 P 0H I I I OH OH OH DIPHOSPHATE MONOPHOSPHATE
0 0 0
II II II ADENOSINE - 0 - P ~ 0 - P - 0H ADENOSINE - 0 - P - 0H I I I 0H 0H 0H
ATPase
ATP + H2O ADP + P + H+ (+ énergie : ∆∆∆∆°’- 50 kJ/mol ) Réaction 1
1 2
Structure biochimique de l ’ATP
CARACTERISTIQUES DES SOURCES ENERGETIQUES
Chaque source énergétique se caractérise par :
• le délai d’apport optimum d’énergie,
• sa capacité ou énergie potentielle totale susceptib le d’être utilisée,
• sa puissance métabolique ou quantité maximale d ’éne rgie qu’elle peut fournir par unité de temps,
• son endurance ou pourcentage de la puissance énergé tique maximale qu’elle peut fournir pendant le plus long temps pos sible,
• son ou ses facteur(s) limitant(s),
• et la durée nécessaire pour reconstituer les réserv es utilisées ou/et
pour éliminer ou métaboliser les déchets et métaboli tes produits

12
EXERCICE TRES COURT ( < 6-7s)
et TRES INTENSE 140 - 250 % PAM)
et RECUPERATION
ATP
Créatine phosphokinase (CPK)
Energie de la PCr ∆G’ = - 77kJ / mol
ADP + Pi
Créatine + Pi
Phosphocréatine (PCr)
Caractéristique de la source énergétique immédiate constituée par le pool des phosphagènes (ATP + PCr) ou source
dite « anaérobie alactique ».
1) Immédiateanaérobie alactique
SOURCE
Sprints courts :départ…10 à 40 m,Tout exercice très court et très intense

13
: Créatine (Cr) Phosphorylcréatine (PCr)
ATP ADP
Mg 2+
Phosphorylcréatine Kinase (PCK)
PCr + ADP Cr + ATP
Phosphorylcréatine Kinase (PCK)
ATP + H2OADP + Pi
Myosine ATPaseContraction :
Turnover de l’ATP
Réplétion de la PCr
Exercice : cycle du turnover de l’ATP par la PCr
Récupération : resynthèse de la PCr
Phosphagènes totaux ATP PCr (ATP +CP)Concentration musculaire mM.kg -1 de muscle 4 - 6 16 - 24 20 - 30 mM (masse musculaire totale) 128 - 192 512 - 768 640 - 960
Energie disponible kJ.kg -1 de muscle 0.16 - 0.2 5 0.67 - 1.00 0.84 - 1.25
kJ (masse musculaire totale) 5.36 - 8.04 21.43 - 32.15 27.13 - 40.19
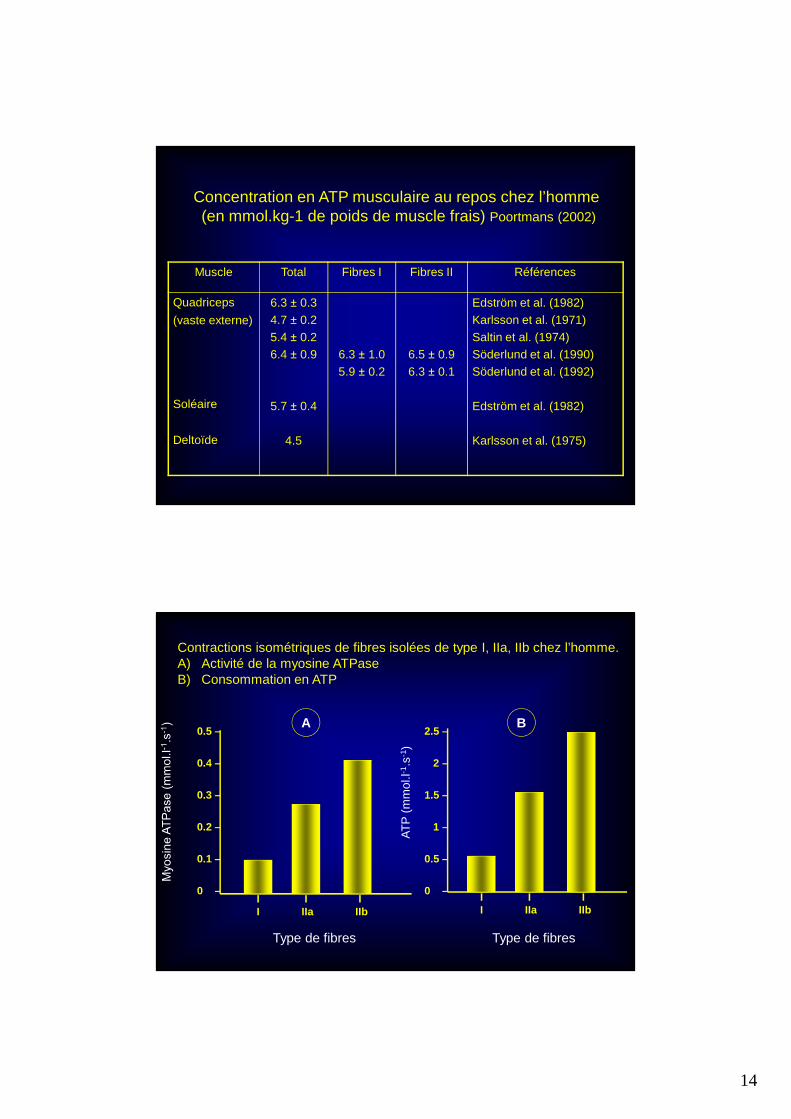
14
Muscle Total Fibres I Fibres II Références
Quadriceps
(vaste externe)
Soléaire
Deltoïde
6.3 ± 0.34.7 ± 0.25.4 ± 0.26.4 ± 0.9
5.7 ± 0.4
4.5
6.3 ± 1.05.9 ± 0.2
6.5 ± 0.96.3 ± 0.1
Edström et al. (1982)Karlsson et al. (1971)Saltin et al. (1974)Söderlund et al. (1990)Söderlund et al. (1992)
Edström et al. (1982)
Karlsson et al. (1975)
Concentration en ATP musculaire au repos chez l’homme (en mmol.kg-1 de poids de muscle frais) Poortmans (2002)
0.5 –
0.4 –
0.3 –
0.2 –
0.1 –
0 –
Type de fibres
I I II IIa IIb
I I II IIa IIb
Type de fibres
AT
P (
mm
ol.l-1
.s-1
)
2.5 –
2 –
1.5 –
1 –
0.5 –
0 –
Contractions isométriques de fibres isolées de type I, IIa, IIb chez l’homme.A) Activité de la myosine ATPaseB) Consommation en ATP
A B
15
100 –
90 –
80 –
70 –
60 –
100 –
90 –
80 –
70 –
60 –
50 –I I I I
Repos 2 6 16
•••
•
130 %
100 %85 %
[AT
P] (
en %
)
[AT
P] (
en %
)•
••
••
••
I I I I I I I I I I
0 55 65 75 85 95 100
Durée des exercices (en min) % VO2max
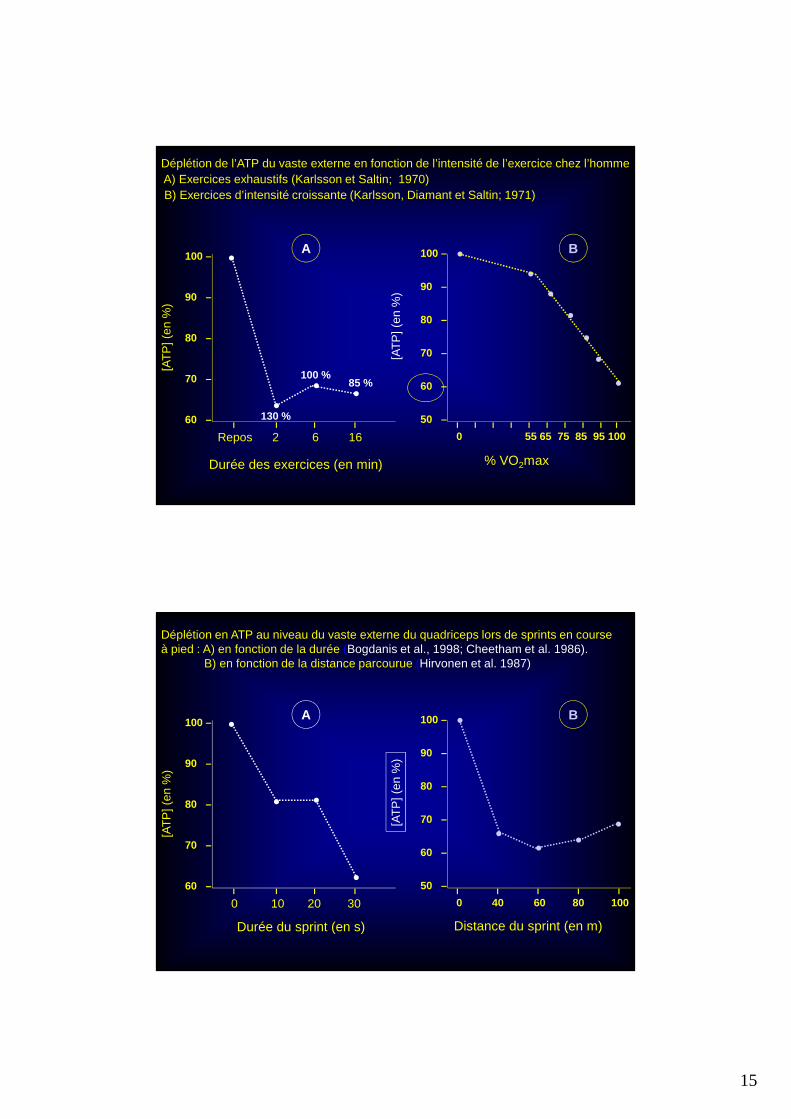
Déplétion de l’ATP du vaste externe en fonction de l’intensité de l’exercice chez l’homme A) Exercices exhaustifs (Karlsson et Saltin; 1970)B) Exercices d’intensité croissante (Karlsson, Diamant et Saltin; 1971)
A B
100 –
90 –
80 –
70 –
60 –
100 –
90 –
80 –
70 –
60 –
50 –I I I I
0 10 20 30
•
••
•
[AT
P] (
en %
)
[AT
P] (
en %
)
•••
•
•
I I I I I
0 40 60 80 100
A B
Durée du sprint (en s) Distance du sprint (en m)
Déplétion en ATP au niveau du vaste externe du quadriceps lors de sprints en course à pied : A) en fonction de la durée (Bogdanis et al., 1998; Cheetham et al. 1986).
B) en fonction de la distance parcourue (Hirvonen et al. 1987)

16
Les réserves en ATP musculaires sont faibles : 4 à 6 millimoles par kilogramme de
muscle frais.
Pour une personne de 70 kg le travail musculaire ne peut compter au total que
sur une réserve de 1.3 à 1.6 kJ, soit à peine l'éne rgie nécessaire pour parcourir :
1 m à 1 m 20 à une vitesse de course de 10 m/s soit 10 s au 100 m,
2 m 60 à 3 m 50 à une vitesse de course de 7,1 m/s soit 3 min 32 s au 1 500 m,
3 m 50 à 4 m 20 à une vitesse de course de 6,3 m/s soit 13 min 13 s au 5 000 m,
4 m 15 à 5 m 10 à une vitesse de course de 5,6 m/s soit 2 h 10 au marathon,
ou 7 m 80 à 9 m 60 à une vitesse de marche de 1,11 m/s soit 4 km/h c'est-à-dire à
une allure de promenade.
Voies métaboliquespour la synthèse de l’ATP
Fluxmmol.s -1.kg -1
Total disponiblemmol.kg-1 de muscle
ATP, PCr ADP, Cr 2.6 26
Glycogène lactate 1.4 60-75 (240 avec le foie)
Glycogène CO 2 0.51- 0.68 31000
Glucose CO 2 0.22
Acides Gras CO 2 0.24 (*)
Flux maximaux de production d’ATP ( P) à partir des différents substrats
disponibles dans le muscle (d’après Greenhaff et al.1993)
(*) selon Newsholme, 1993, les réserves en triacylglycérol de l’organisme
pourraient assurer l’apport énergétique nécessaire à un marathon de 119h !!
17
Reins
Pancréas
Muqueuseintestinale
Foie
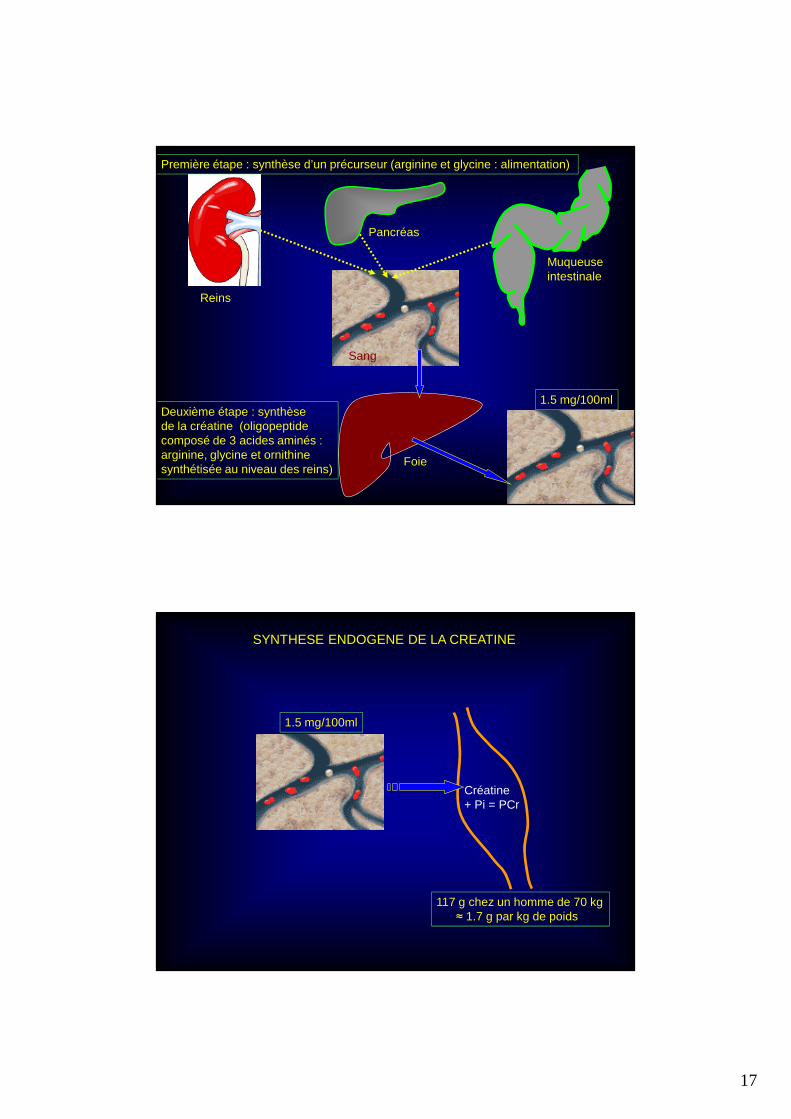
Première étape : synthèse d’un précurseur (arginine et glycine : alimentation)
Sang
Deuxième étape : synthèsede la créatine (oligopeptidecomposé de 3 acides aminés :arginine, glycine et ornithine synthétisée au niveau des reins)
1.5 mg/100ml
1.5 mg/100ml
117 g chez un homme de 70 kg≈ 1.7 g par kg de poids
Créatine + Pi = PCr
SYNTHESE ENDOGENE DE LA CREATINE

18
Muscle Total Fibres I Fibres II Références
Quadriceps
(vaste externe)
Soléaire
Deltoïde
19 ± 1.514.5 ± 0.418.9 ± 1.116.3 ± 0.719.1 ± 0.3
17.7 ± 1.7
19.6
15.718.5 ± 0.517.0 ± 2.2
16.620.8 ± 1.019.2 ± 2.6
Edström et al. (1982)Karlsson et al. (1971)Saltin et al. (1974)Greenhaff et al. (1991) Söderlund et al. (1992)Tesch et al. (1993)
Edström et al. (1982)
Karlsson et al. (1975)
Concentration en PCr musculaire au repos chez l’homme (en mmol.kg-1 de poids de muscle frais) Poortmans (2002)
1 – Métabolisme de l’ATP et de la PCr
La concentration intracellulaire en ATP diminue au maximum de 50% après unexercice, aussi intense soit-il. Pour cela,
¤ Les réserves en PCr sont fortement mobilisées pour la resynthèse d’ATP,
¤ Le groupement phosphate de la PCr se lie à l’ADP ( ADP + PCr ⇒⇒⇒⇒ ATP + Cr),
¤ La concentration en PCr baisse alors très vite,
¤ Une déplétion totale n’est cependant très rarement atteinte ( ⇓⇓⇓⇓ ≈≈≈≈ 10 à 15%).
Ainsi ces facteurs peuvent limiter fortement la performanc e des exercicestrès intenses (sprint…) en terme de puissance métabolique (d ébit maximal).
19
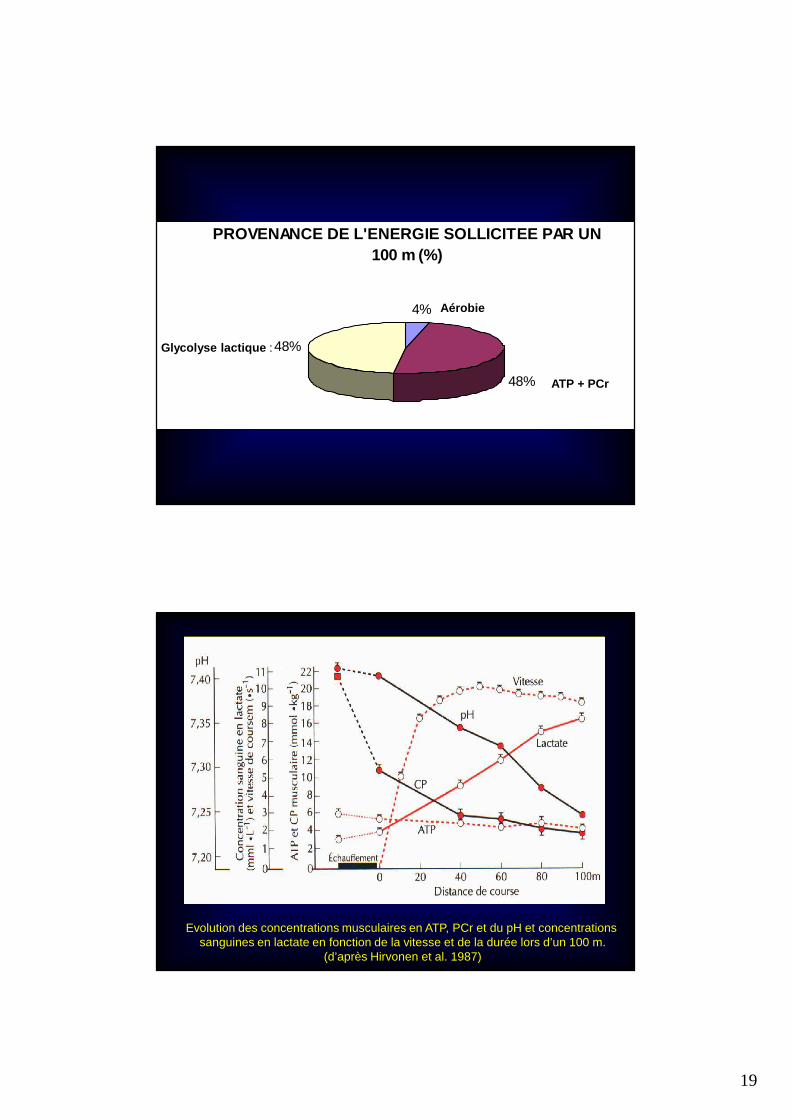
PROVENANCE DE L'ENERGIE SOLLICITEE PAR UN 100 m (%)
4%
48%
48%
Aérobie
ATP + PCr
Glycolyse lactique :
Evolution des concentrations musculaires en ATP, PCr et du pH et concentrations sanguines en lactate en fonction de la vitesse et de la durée lors d’un 100 m.
(d’après Hirvonen et al. 1987)
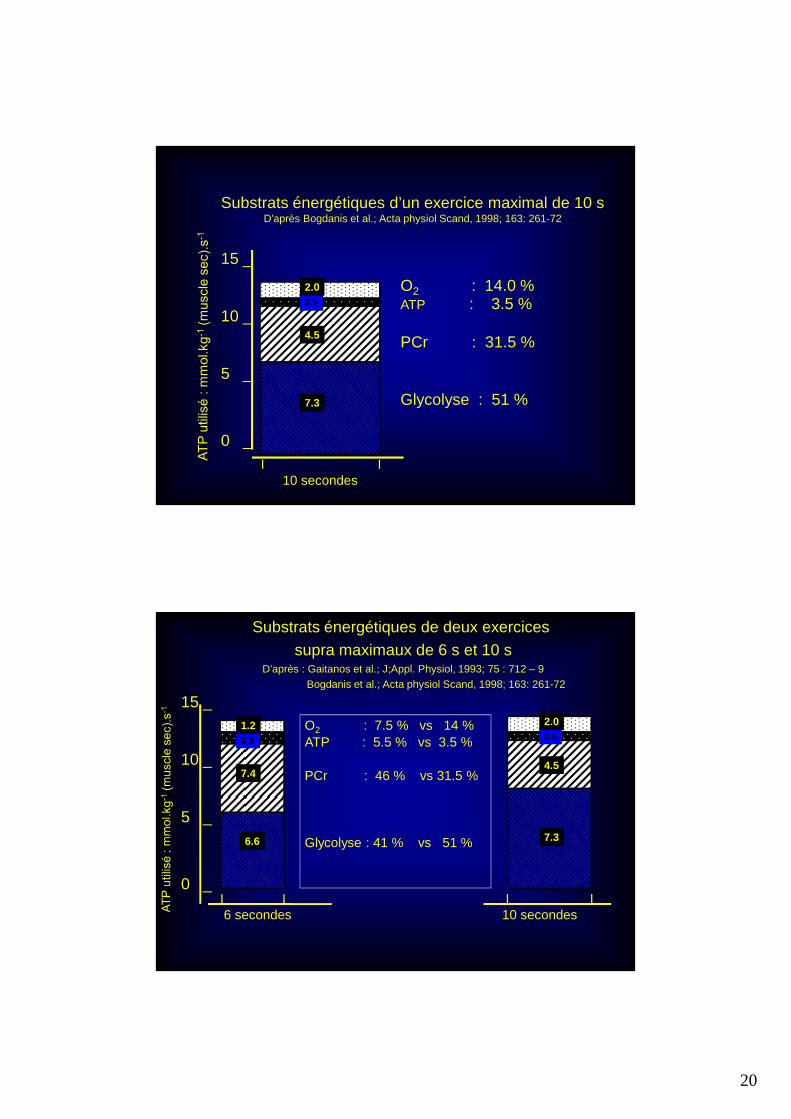
20
15 _
10 _
5 _
0 _
7.3
4.5
0.5
2.0 O2 : 14.0 %ATP : 3.5 %
PCr : 31.5 %
Glycolyse : 51 %
I I10 secondes
Substrats énergétiques d’un exercice maximal de 10 sD’après Bogdanis et al.; Acta physiol Scand, 1998; 163: 261-72
15 _
10 _
5 _
0 _
6.6
7.4
0.9
1.2 O2 : 7.5 % vs 14 %ATP : 5.5 % vs 3.5 %
PCr : 46 % vs 31.5 %
Glycolyse : 41 % vs 51 %
I I6 secondes
7.3
4.5
0.5
2.0
I I10 secondes
Substrats énergétiques de deux exercices supra maximaux de 6 s et 10 s
D’après : Gaitanos et al.; J;Appl. Physiol, 1993; 75 : 712 – 9Bogdanis et al.; Acta physiol Scand, 1998; 163: 261-72
21
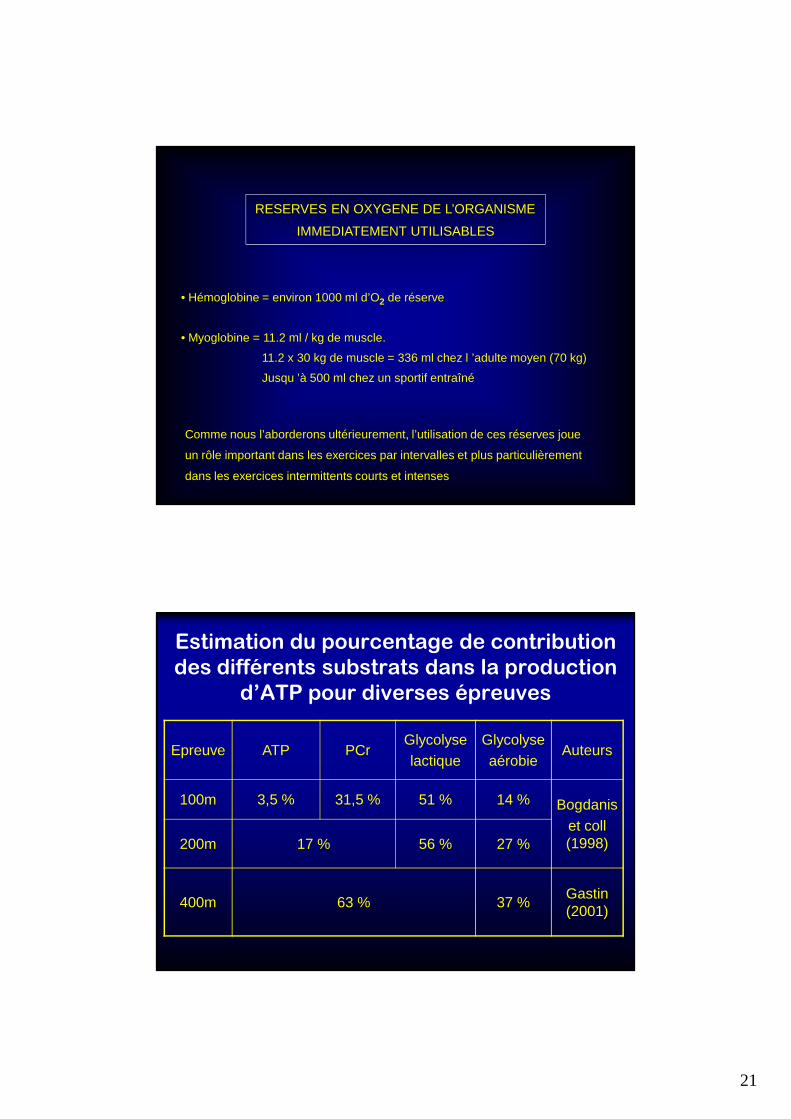
RESERVES EN OXYGENE DE L’ORGANISME
IMMEDIATEMENT UTILISABLES
• Hémoglobine = environ 1000 ml d’O2 de réserve
• Myoglobine = 11.2 ml / kg de muscle.
11.2 x 30 kg de muscle = 336 ml chez l ’adulte moyen (70 kg)
Jusqu ’à 500 ml chez un sportif entraîné
Comme nous l’aborderons ultérieurement, l’utilisation de ces réserves joue
un rôle important dans les exercices par intervalles et plus particulièrement
dans les exercices intermittents courts et intenses
Estimation du pourcentage de contribution
des différents substrats dans la production
d’ATP pour diverses épreuves
Epreuve ATP PCrGlycolyselactique
Glycolyseaérobie
Auteurs
100m 3,5 % 31,5 % 51 % 14 % Bogdaniset coll (1998) 200m 17 % 56 % 27 %
400m 63 % 37 %Gastin (2001)
22
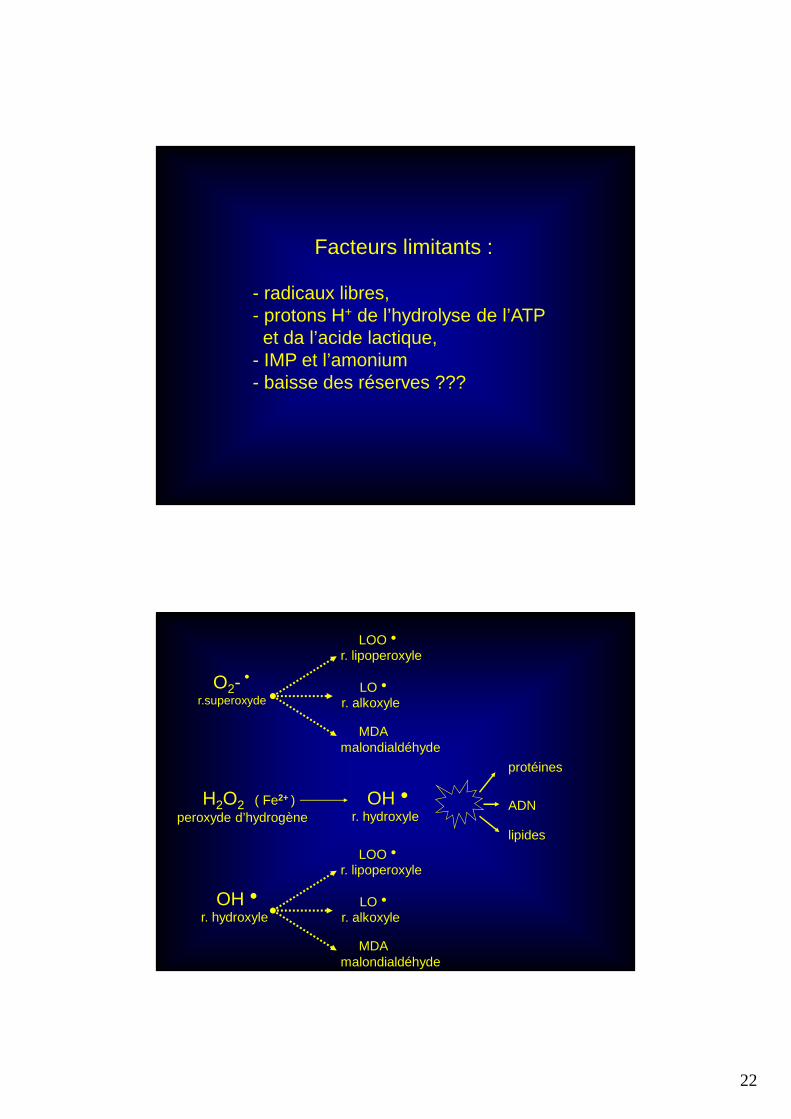
Facteurs limitants :
- radicaux libres,- protons H+ de l’hydrolyse de l’ATPet da l’acide lactique,
- IMP et l’amonium - baisse des réserves ???
O2-hr.superoxyde
LOOh
r. lipoperoxyle
LOh
r. alkoxyle
MDAmalondialdéhyde
H2O2 ( Fe2+ )peroxyde d’hydrogène
OHh
r. hydroxyle
protéines
ADN
lipides
OHh
r. hydroxyle
LOOh
r. lipoperoxyle
LOh
r. alkoxyle
MDAmalondialdéhyde

23
Figure 9 : Synthèse des phosphagènes après un exer cicecourt et intense. L’apport d’oxygène est indispen sablepour perm ettre la resynthèse de l’ATP dans les m ito chon-dries. Les m olécules d’ATP ains i form ées perm ettent elles-m êm es la resynthèse de la PCr. Harris et coll. (197 6)
20 _
16_
12 _
8 _
4 _
Occlusion = absence d ’ oxy-gène = pas de resynthèse desPhosphagènes (AT P + PCr).
Circulation sanguine normale
Exerciceépuisant
Repos Récupération
I I I I I I0 2min 4min 8min 12 min
90% en 4min
85% en 2min
70% en 50s
CINETIQUE DE LA RESYNTHESE DES PHOSPHAGENES
La PCr utilisée au cours de l ’exercice est reconsti tuée comme suit:
70 % en 30 s
84 % en 2 min
89 % en 4 min
97 % à 100 % en 6 à 8 min

24
• Comme l’ont démontré les travaux Quirstorff & al, 1992 ;Trump & al.,1996 ; Bogdanis & al.,1996, il est possible d ’améliorer la vitesse de resynthèse de la PCr entre plusieurs exercices courts et intenses grâce à un bon développement préalable de lacapillarisation et de la capacité oxydative des muscles sollicités.
Le développement de la capacité aérobie doit donc t oujours précéder l ’entraînement de la vitesse, de l’enduran ce de la vitesse et de la puissance musculaire.
• Après un exercice court et intense, la resynthèse de la phosphocréatine (PCr) à partir de nouvelles molécules d ’ATP, nécessite la présence d ’oxygène
SOURCED’ENERGIE
SUBSTRATS PRODUCTIOND’ATP
DELAI DEPRODUCTION
OPTIMALE
CAPACITE PUISSANCE ENDURANCE
IMMEDIATEPhosphagènes
ANAEROBIEALACTIQUE
ATP + PCr TRES FAIBLE1 PCr = 1 ATP
NUL TRES FAIBLE20 - 60 kJ65kJ (*)
TRES ELEVEE:250 à 530 kJ.min-1750 kJ (*)4 à 6 - 7 s
≤≤≤≤ 15 - 20sdépend du %de puissancemax (jamaisInférieurs à 95% de la puissanceMaximale)
(*) Sportif spécialiste entraîné et de haut niveau
RECAPITULATIF

25
CINETIQUE DE LA RESYNTHESE DES PHOSPHAGENES
La PCr utilisée au cours de l ’exercice est reconstituée comme suit:
70 % en 30 s
84 % en 2 min
89 % en 4 min
97 % à 100 % en 6 à 8 min
Evaluation de la capacité de la source des phosphagènes
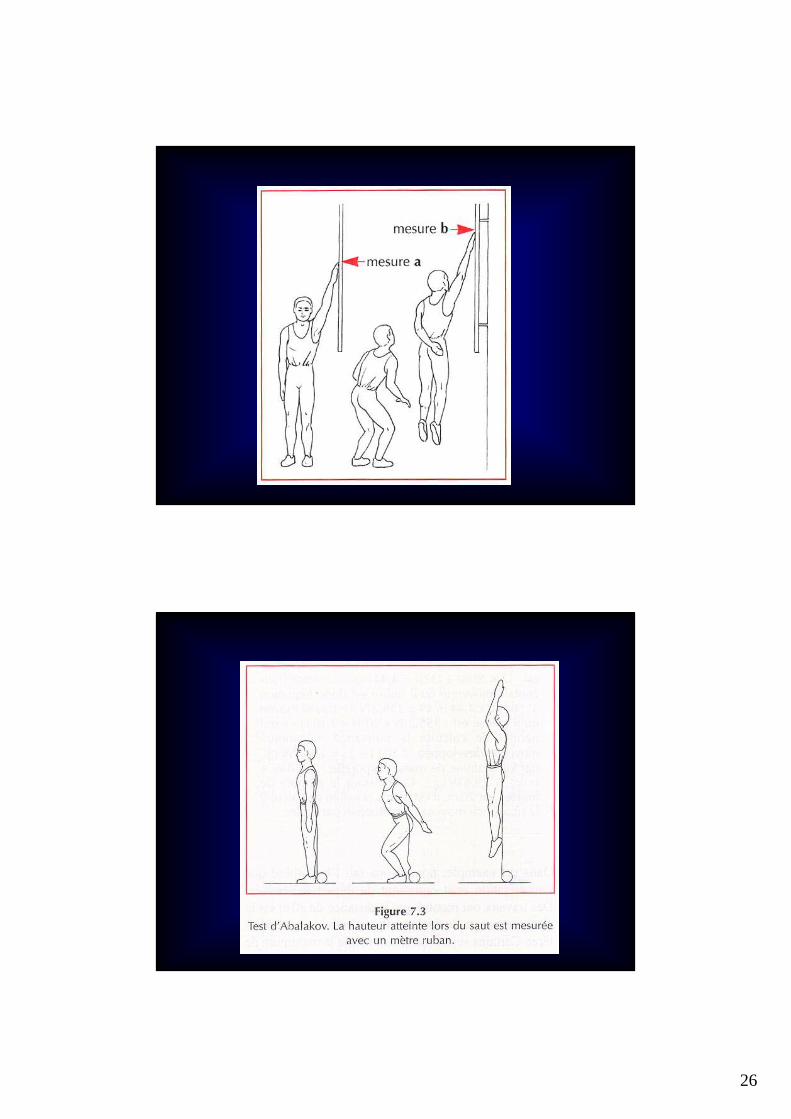
26
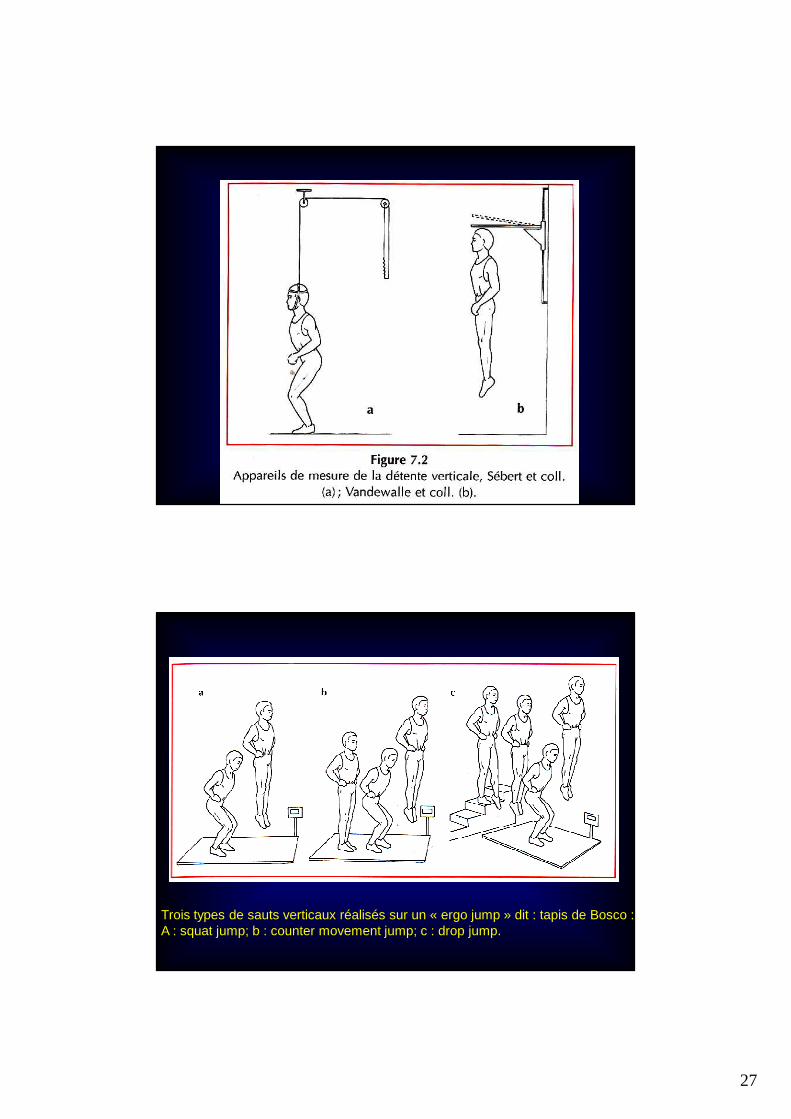
27
Trois types de sauts verticaux réalisés sur un « ergo jump » dit : tapis de Bosco :A : squat jump; b : counter movement jump; c : drop jump.

28
Evaluation de la vitesse sur des distances très courtes : 20, 30, 40m maximum

29
1- VITESSE:
10m; 20m; 30m
COMMENT CALCULER LA PUISSANCE A PARTIR
DE LA CONNAISSANCE DE LA VITESSE

30
Comme la puissance (P) est égale au produit de laforce par la vitesse :P = F . v (4)
Pour une distance donnée (d), il est possible d’estimer l’accélération horizontale moyenne (a) d’après la formule suivante :d = ½ a t2 (1)
donc a = 2d / t2 (2)
Connaissant l’accélération, la force moyenne depropulsion horizontale (Fh ) peut alors être calculée en utilisant la deuxième loi du mouvement de Newton:Fh = m . a (3)
• Prenons l’exemple d’un sportif qui pèse 80kg et qui parcourt20m lancé en 3s.
En utilisant l’équation (2), son accélération moyennehorizontale est : (2 x 20m) ÷ (3s)2 = 4.44 m/s2 .
• La force horizontale moyenne qu’il utilise est donc :(équation 3) : 80kg x 4.44m/s2 = 355.2N.
• Le travail moyen qu’il réalise est :355.2N x 20m = 7104 J ce qui permet de calculer
• la puissance mécanique moyenne développée :7104 J ÷ 3s = 2368 W ou par kg de masse corporelle : 2368W ÷ 80kg = 29.6 W/kg.
Connaissant le nombre de foulées sur 20m, il est mêmepossible de calculer la puissance moyenne développée parfoulée.

31
EXERCICES A DOMINANTE LACTIQUE: (400, 800m)
et RECUPERATION
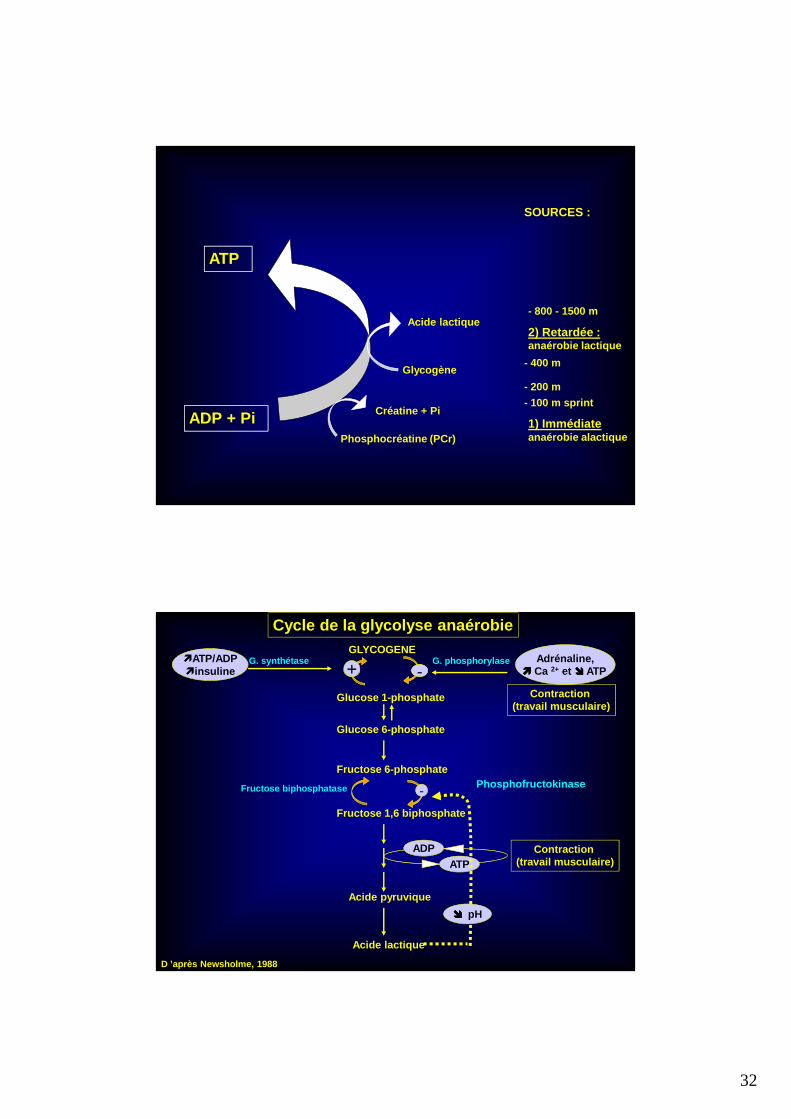
32
ATP
ADP + Pi Créatine + Pi
Phosphocréatine (PCr)1) Immédiateanaérobie alactique
SOURCES :
Glycogène
Acide lactique2) Retardée :anaérobie lactique
- 100 m sprint
- 200 m
- 400 m
- 800 - 1500 m
Cycle de la glycolyse anaérobieGLYCOGENE
+Adrénaline,
���� Ca 2+ et ���� ATP
Glucose 1-phosphate
Glucose 6-phosphate
G. phosphorylase
Fructose 6-phosphate
-
Fructose 1,6 biphosphate
Acide pyruvique
ADP
ATP
Acide lactique
D ’après Newsholme, 1988
Contraction (travail musculaire)
Contraction (travail musculaire)
PhosphofructokinaseFructose biphosphatase
G. synthétase
���� pH
�ATP/ADP�insuline --+
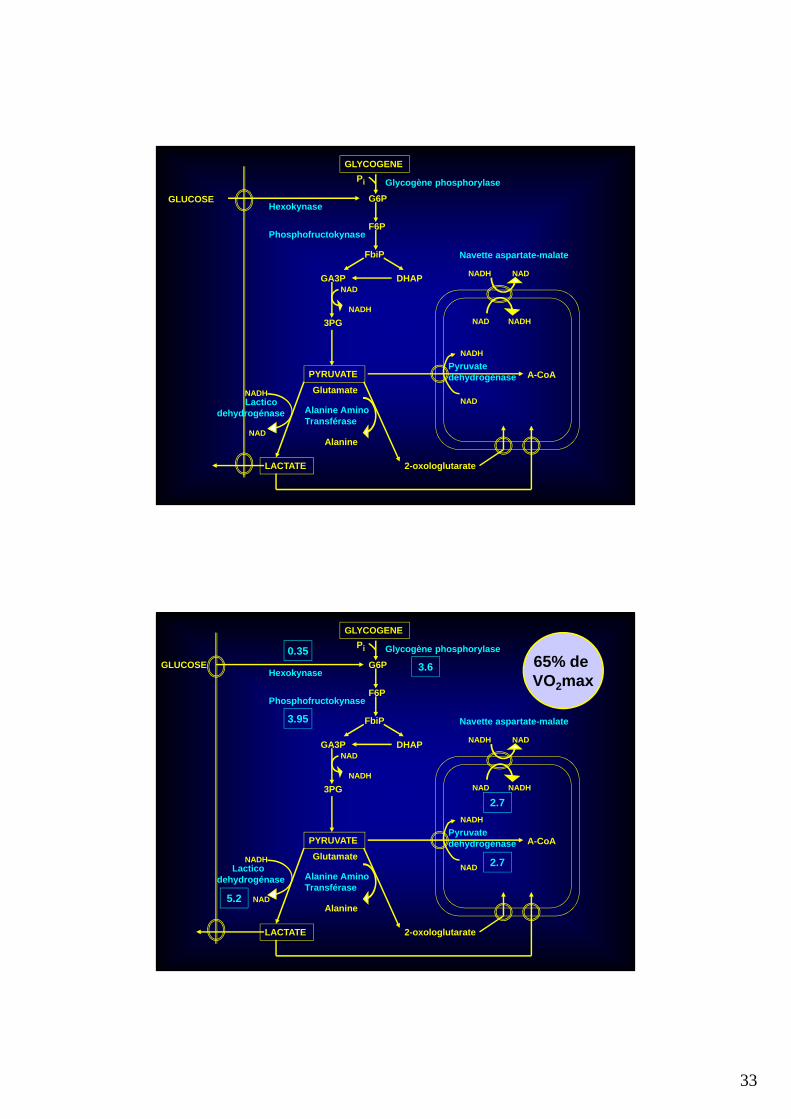
33
GLYCOGENE
Glycogène phosphorylase
G6P
Pi
F6PPhosphofructokynase
GLUCOSEHexokynase
FbiP
GA3P DHAPNAD
NADH
3PG
PYRUVATE
LACTATE
Glutamate
Alanine
Alanine AminoTransférase
2-oxologlutarate
NADH
NAD
NADH NAD
NAD NADH
Pyruvatedehydrogénase A-CoA
Navette aspartate-malate
Lacticodehydrogénase
NADH
NAD
GLYCOGENE
Glycogène phosphorylase
G6P
Pi
F6PPhosphofructokynase
GLUCOSEHexokynase
FbiP
GA3P DHAPNAD
NADH
3PG
PYRUVATE
LACTATE
Glutamate
Alanine
Alanine AminoTransférase
2-oxologlutarate
NADH
NAD
NADH NAD
NAD NADH
Pyruvatedehydrogénase A-CoA
Navette aspartate-malate
65% de VO2max
3.6
0.35
3.95
Lacticodehydrogénase
NADH
NAD5.2
2.7
2.7
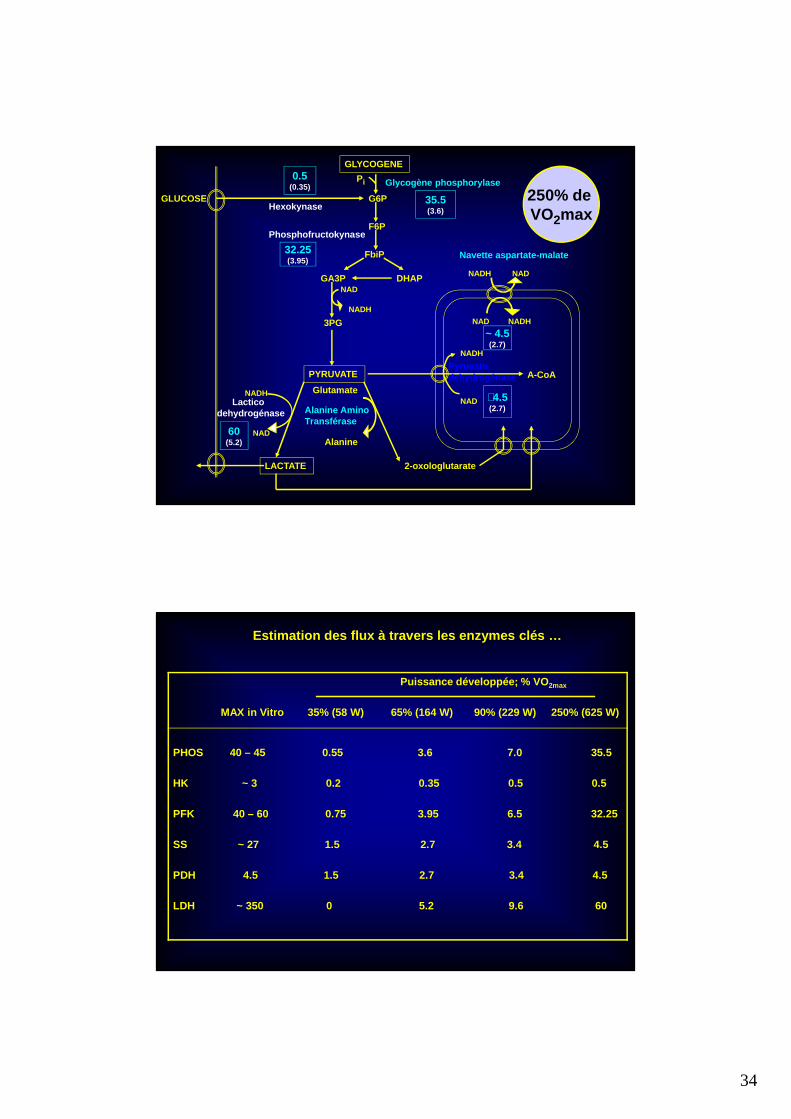
34
GLYCOGENE
Glycogène phosphorylase
G6P
Pi
F6PPhosphofructokynase
GLUCOSEHexokynase
FbiP
GA3P DHAPNAD
NADH
3PG
PYRUVATE
LACTATE
Glutamate
Alanine
Alanine AminoTransférase
2-oxologlutarate
NADH
NAD
NADH NAD
NAD NADH
Pyruvatedehydrogénase A-CoA
Navette aspartate-malate
250% de VO2max
35.5(3.6)
0.5(0.35)
32.25(3.95)
Lacticodehydrogénase
NADH
NAD60(5.2)
∼4.5(2.7)
~ 4.5(2.7)
Puissance développée; % VO 2max
MAX in Vitro 35% (58 W) 65% (164 W) 90% (229 W) 250% (625 W)
PHOS 40 – 45 0.55 3.6 7.0 35.5
HK ~ 3 0.2 0.35 0.5 0.5
PFK 40 – 60 0.75 3.95 6.5 32.25
SS ~ 27 1.5 2.7 3.4 4.5
PDH 4.5 1.5 2.7 3.4 4.5
LDH ~ 350 0 5.2 9.6 60
Estimation des flux à travers les enzymes clés …

35
I . DES QUESTIONS QUI SE POSENT...
1) Acide lactique ou lactate... quelle différence ?
2) Quel est le devenir du lactate ?
3) Quelles significations accorder au(x) seuil(s) anaérobie(s) ?
3.1 - A quelle puissance le muscle produit-il du lactate ?
3.2 - Est ce l'absence d'oxygène qui entraîne la production du lactate musculaire ?
3.3 - Quelle validité accorder aux nombreuses techniques de détermination du seuil anaérobie ?
3.4 - Quel est le niveau de reproductibilité intra technique du seuil anaérobie ?
II. DES QUESTIONS QUI SE POSENT...(suite)II. DES QUESTIONS QUI SE POSENT...(suite)
�� La glycolyse aLa glycolyse a--tt--elle vraiment un mauvais rendement elle vraiment un mauvais rendement énergétique ?énergétique ?
�� EstEst--ce l’accumulation de lactate qui induit :ce l’accumulation de lactate qui induit :La fatigue musculaire ?La fatigue musculaire ?
et donneet donne : Des crampes ?Des crampes ?Des courbatures ?Des courbatures ?
36
Membrane cellulaire
ATP
ADP + Pi
+ Glycogène phosphorylase
+ Hexokinase Glycogène musculaire (à 6 carbones)
ADP + Pi
ATP
Glucose-6-phosphate
1
2
3
4
5
ATP
ADP + Pi
)
2 NAD
2 NADH 2
6
7
ADP + Pi
ATP
(2) Pyruvate (à 3 carbones chacun)
1 - Phased’amorçage :
- 1 ATPavec le glucose
2 - PhasePréparatoire :
- 1ATP
3 - Phase derembourse-ment et de
production :+ 4 ATP
Phosphofructokinase
Pyruvate kinase
8
9
10
Bilan :+ 2ATP avec le glucose+ 3 ATP avec le glycogène
Capillaire sanguin GLUCOSE CIRCULANT
à 6carbones
(2) lactate
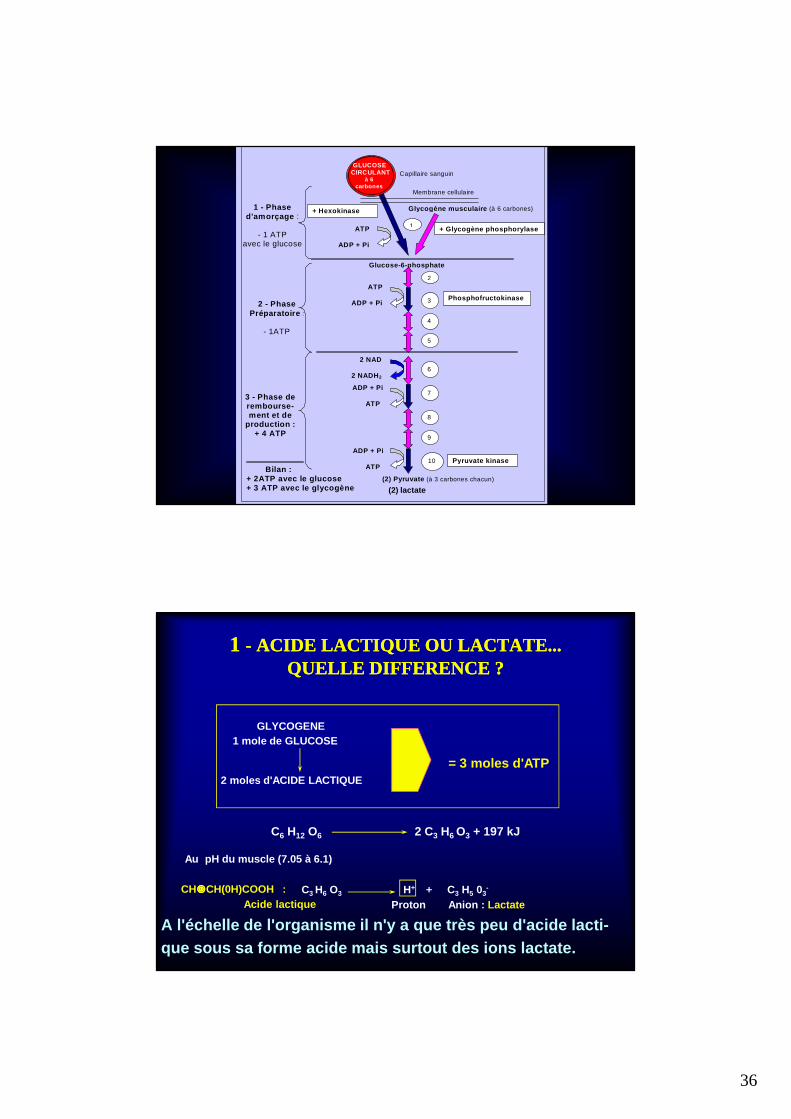
1 - ACIDE LACTIQUE OU LACTATE... QUELLE DIFFERENCE ?
1 - ACIDE LACTIQUE OU LACTATE... QUELLE DIFFERENCE ?
GLYCOGENE1 mole de GLUCOSE
2 moles d'ACIDE LACTIQUE
C6 H12 O6 2 C3 H6 O3 + 197 kJ
Au pH du muscle (7.05 à 6.1)
C3 H6 O3 H+ + C3 H5 03-
Proton Anion : Lactate
A l'échelle de l'organisme il n'y a que très peu d' acide lacti-que sous sa forme acide mais surtout des ions lacta te.
= 3 moles d'ATP
CH����CH(0H)COOH : Acide lactique

37
LACTATE ET EXERCICE COURT ET INTENSE
100m sprint……………environ 13-16 mmol/l200m sprint……………environ 18-20 mmol/l400,800 et 1500m……environ 22-26 mmol/l5000m…………………environ 13 mmol/l10000m………………..environ 8 mmol/l
1 mole de lactate = 23,5 kcal1 g ……………….= 0,26 kcal
... DONC : EST- IL MAUVAIS DE PRODUIRE BEAUCOUP DE LACTATE ?
... DONC : EST- IL MAUVAIS DE PRODUIRE BEAUCOUP DE LACTATE ?
• Une mole de glycogène permet la synthèse de 3 ATP et s’accompagne de la formation de 2 moles de lactate.
• Donc, plus le muscle produit de lactate par unité de temps, plus d’ATP ont été synthétisés,et donc plus important a été le travail musculaire.
• L’athlète qui réussit dans les disciplines courtes : 10 s à 9 min. est celui qui produit le plus de lactate par unité de temps (Lacour et Coll. 1991)
38
GLYCOGENE
CELLULE MUSCULAIRE
CAPILLAIRE SANGUIN
O2
PYRUVATE
LACTATE
Cycle de Krebs.
ADP ATP (36)CO2
CO2
ATP (+3)
ADP
H+
NADH
NAD
Chaîne des transporteurs d’électrons
H2
MITOCHONDRIE OXYDATION
MILIEU INTERSTITIEL
NAD
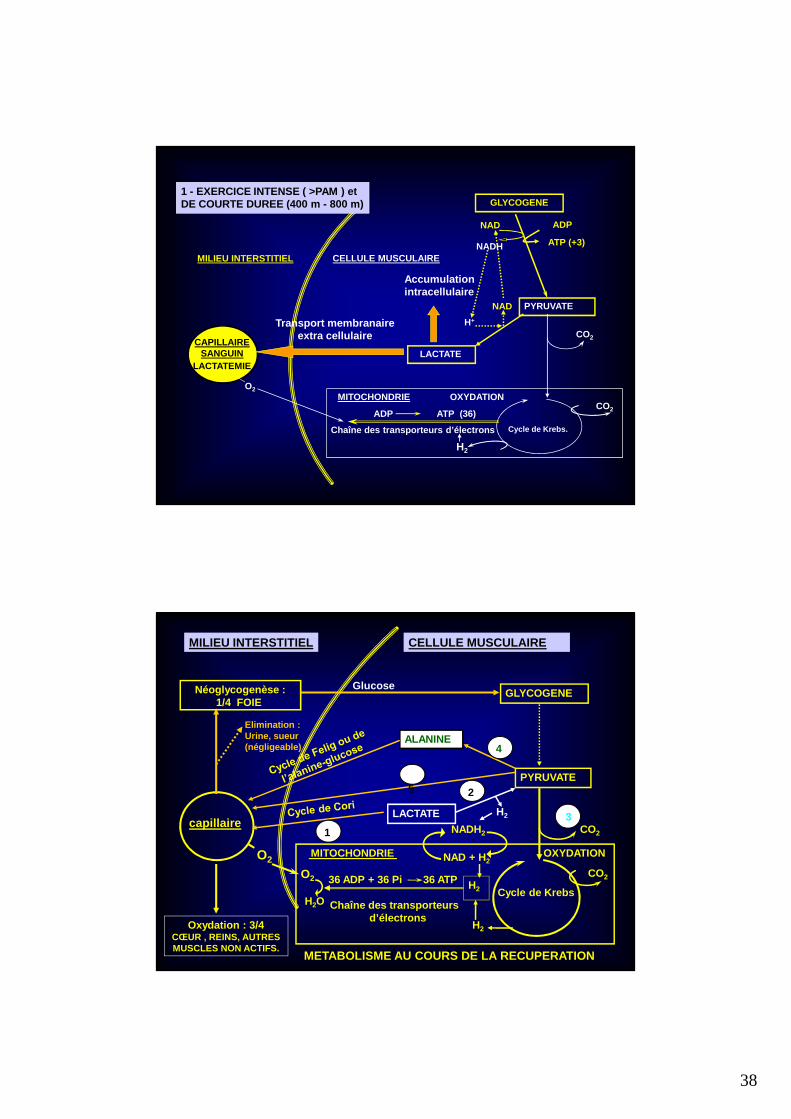
1 - EXERCICE INTENSE ( >PAM ) etDE COURTE DUREE (400 m - 800 m)
LACTATEMIE
Accumulationintracellulaire
Transport membranaireextra cellulaire
GLYCOGENE
CELLULE MUSCULAIRE
capillaire
O2
PYRUVATE
LACTATE
MITOCHONDRIE OXYDATION
Cycle de Krebs36 ADP + 36 Pi 36 ATP
CO2
CO2
H2
MILIEU INTERSTITIEL
NADH2
Chaîne des transporteurs d’électrons
H2
H2
NAD + H2
ALANINE
Néoglycogenèse : 1/4 FOIE
Glucose
Oxydation : 3/4CŒUR , REINS, AUTRES MUSCLES NON ACTIFS.
1
2
4
Elimination :Urine, sueur(négligeable)
3
5
O2
H2O
METABOLISME AU COURS DE LA RECUPERATION

39
DEVENIR DU LACTATE AU COURS DE LA RECUPERATION
LACTATEOXYDATION
4/5
GLYCOGENESE1/5
ELIMINATIONnégligeablePar :
• Les muscles squelettiquesLes fibres musculaires productricesLes fibres musculaires environnantes (navette)Les fibres musculaires d’autres territoires au repos
• Le myocarde 10 %
• Les reins < 10 %
Par :• Le foie- Cycle de Cori- Cycle de l’alanine-glucose
• Les reins
• Les muscles (indirectement ?)
Par :
• L’urine et la sueur
~
~
CINETIQUE DU METABOLISME DU LACTATE POST EXERCICE
Transformation du lactate après un exercice épuisan t de deux minutes
1- RECUPERATION PASSIVE:
50 % en environ 25 min75 % en environ 50 min88 % en environ 1h 15 min100 % en environ 1h 30 min
2- RECUPERATION ACTIVE (entre 40 et 60 % de VAM)
50 % en environ 6 min75 % en environ 12 min
100 % en environ 20 min

40
VITESSE DE DECROISSANCE DU LACTATE POST EXERCICE (en % du La max.min -1)
Auteurs Nature de l’exercice
Récupération passive
Récupérationactive (% VO 2max)
Mc Grail et al. (1978)
Bonen et Belcastro (1977)
Hermansen et Stenvold(1972)
Cazorla et al. (1984)
Ergocycle
Course
Course
Natation
2.0
2.5
2.8
2.73
2.9 (30)
4.5 (60 – 70)
4.8 (70)
5.3 (70 et libre)
2.2 EN CONSEQUENCE
Le lactate n’est donc pas un « déchet » ni
surtout « une toxine qui empoisonne le
muscle » mais bien une source énergétique
potentielle utilisable après, ou au cours
d’une récupération passive ou active.

41
2.3 QUEL EST LE DEVENIR DU LACTATE ?
LACTATE ENTRANT (LE)
(MUSCLE)
SANGSANG
LACTATE SORTANT (LS)
( OXYDATION, GLYCOGENESE )
• LE > LS : (accumulation)
• LE = LS : (Etat stable) 6-8 mmol.l-1
• LE < LS : (décroissance)
1 H 30
LACTATEMIE
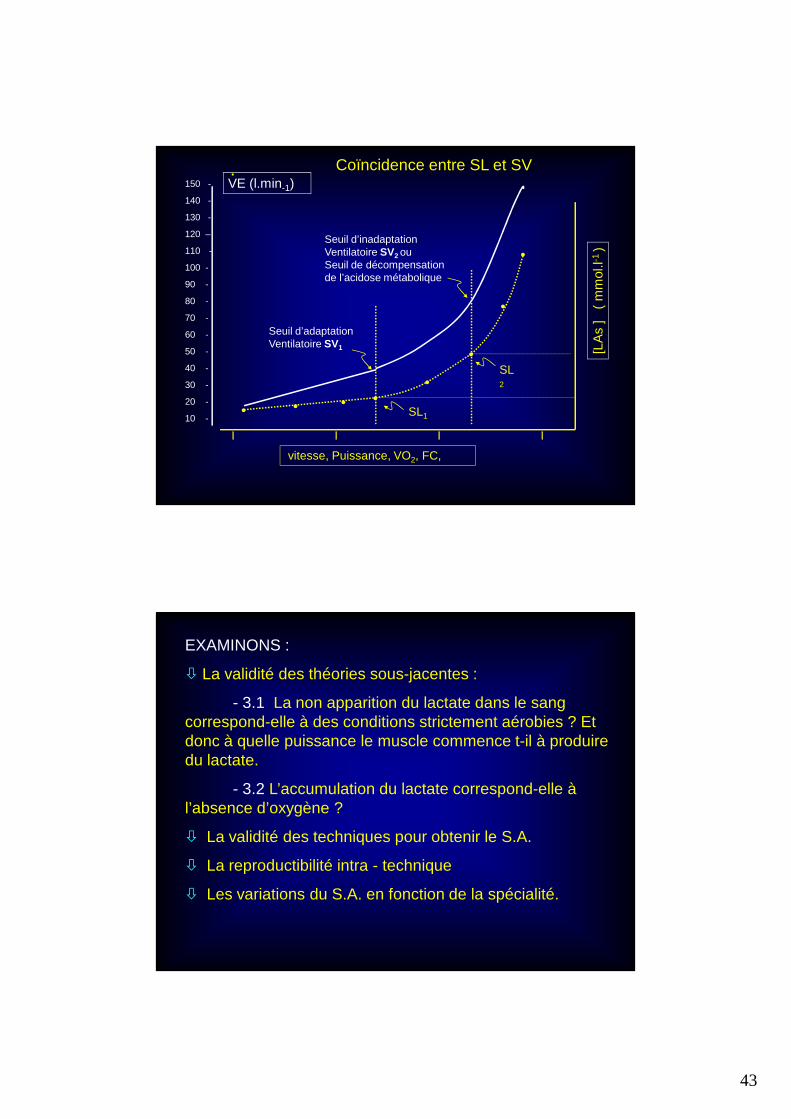
3. QUELLES SIGNIFICATIONS ACCORDER AU(X) SEUIL(S) ANAEROBIE (S.A.)
3. QUELLES SIGNIFICATIONS ACCORDER AU(X) SEUIL(S) ANAEROBIE (S.A.)
SEUIL = « Limite au delà de laquelle les conditions sont changées » (Larousse).
ANAEROBIE : sans air, donc sans oxygène.
42
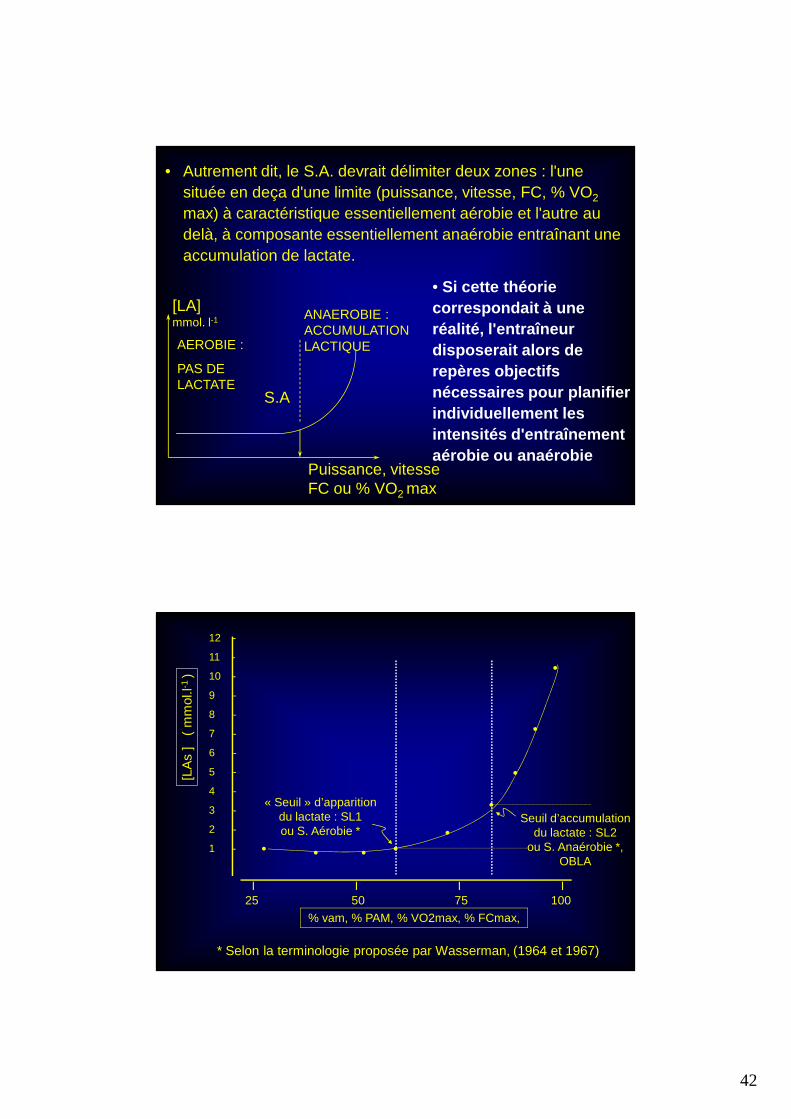
• Autrement dit, le S.A. devrait délimiter deux zones : l'une située en deça d'une limite (puissance, vitesse, FC, % VO2
max) à caractéristique essentiellement aérobie et l'autre au delà, à composante essentiellement anaérobie entraînant une accumulation de lactate.
• Si cette théorie correspondait à une réalité, l'entraîneur disposerait alors de repères objectifs nécessaires pour planifier individuellement les intensités d'entraînement aérobie ou anaérobie
Puissance, vitesseFC ou % VO2 max
AEROBIE :
PAS DE LACTATE
[LA]mmol. l-1
S.A
ANAEROBIE : ACCUMULATIONLACTIQUE
I I I I25 50 75 100
% vam, % PAM, % VO2max, % FCmax,
•
•
•••••
•
•
« Seuil » d’apparitiondu lactate : SL1ou S. Aérobie *
Seuil d’accumulationdu lactate : SL2
ou S. Anaérobie *,OBLA
12 -
11 -
10 -
9 -
8 -
7 -
6 -
5 -
4 -
3 -
2 -
1 -
[LA
s ]
( m
mol
.l-1 )
* Selon la terminologie proposée par Wasserman, (1964 et 1967)
43
I I I I
vitesse, Puissance, VO2, FC,
•
•
•••••
•
Seuil d’adaptationVentilatoire SV1
Seuil d’inadaptationVentilatoire SV2 ou Seuil de décompensationde l’acidose métabolique
[LA
s ]
( m
mol
.l-1 )
150 -
140 -
130 -
120 –
110 -
100 -
90 -
80 -
70 -
60 -
50 -
40 -
30 -
20 -
10 -
SL2
SL1
VE (l.min-1)•
Coïncidence entre SL et SV
EXAMINONS :
� La validité des théories sous-jacentes :
- 3.1 La non apparition du lactate dans le sang correspond-elle à des conditions strictement aérobies ? Et donc à quelle puissance le muscle commence t-il à produire du lactate.
- 3.2 L’accumulation du lactate correspond-elle à l’absence d’oxygène ?
� La validité des techniques pour obtenir le S.A.
� La reproductibilité intra - technique
� Les variations du S.A. en fonction de la spécialité.
44
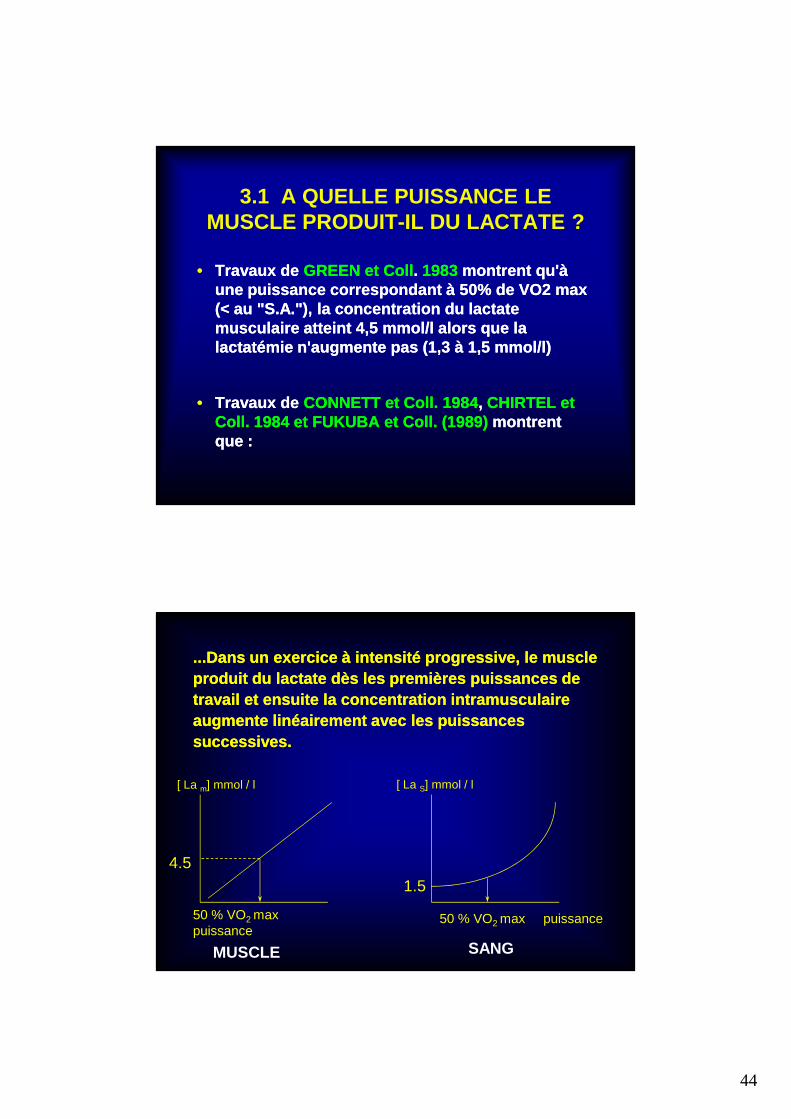
3.1 A QUELLE PUISSANCE LE MUSCLE PRODUIT-IL DU LACTATE ?
•• Travaux de Travaux de GREEN et CollGREEN et Coll .. 1983 1983 montrent qu'à montrent qu'à une puissance correspondant à 50% de VO2 max une puissance correspondant à 50% de VO2 max (< au "S.A."), la concentration du lactate (< au "S.A."), la concentration du lactate musculaire atteint 4,5 mmol/l alors que la musculaire atteint 4,5 mmol/l alors que la lactatémie n'augmente pas (1,3 à 1,5 mmol/l)lactatémie n'augmente pas (1,3 à 1,5 mmol/l)
•• Travaux de Travaux de CONNETT et Coll. 1984CONNETT et Coll. 1984 , , CHIRTEL et CHIRTEL et Coll. 1984 et FUKUBA et Coll. (1989) Coll. 1984 et FUKUBA et Coll. (1989) montrent montrent que :que :
...Dans un exercice à intensité progressive, le mus cle ...Dans un exercice à intensité progressive, le mus cle produit du lactate dès les premières puissances de produit du lactate dès les premières puissances de travail et ensuite la concentration intramusculaire travail et ensuite la concentration intramusculaire augmente linéairement avec les puissances augmente linéairement avec les puissances successives.successives.
4.5
[ La m] mmol / l
50 % VO2 max puissance
MUSCLE
[ La S] mmol / l
50 % VO2 max puissance
SANG
1.5

45
� CONCLUSION : On ne peut conclure à une absence de production de lactate par le muscle à partir de l’absence de modification de la lactatémie en début d’exercice triangulaire.
Il n’existe donc pas de puissance «SEUIL» en deça de laquelle le muscle ne produit pas de lactate et au delà de laquelle il en produit.
� HYPOTHESEL’absence de modification de la lactatémie en début d’exercice triangulaire résulte probablement de l’interaction de 3 phénomènes :
1- Du gradient de concentration trop faible (Karlsson 1971)
2- Des transporteurs membranaires insuffisamment activés.
3- De la dilution du lactate dans un vaste espace extramusculaire (Zouloumian et Freund, 1981)
TRANSPORT TRANSPORT MEMBRANAIRE DU LACTATEDU LACTATE
• Il existe des protéines qui permettent le transport du lactate à travers
le sarcolemme : monocarboxylate tansporter : MCT1 et MCT4
• D'elles dépend la vitesse du passage du lactate musculaire :
milieu intracellulaire milieu extracellulaire <–> sang
• La vitesse du passage membranaire dépend :
1. du niveau de stimulation des transporteurs.
2. du nombre de transporteurs mis en jeu.
MCT1MCT1
MCT4MCT4
(Juel et al.,1991; Dermott et Bonen,1993;Pilegaard et al.,1993; Roth et Brooks,1993)

46
2- Le nombre des transporteurs recrutés dépend :
- du niveau d ’entraînement
- de la nature des fibres activées : les fibres rapi des présentent plus de MTC 4 et pratiquement pas de MTC 1,
- de l’âge des sujets (le nombre de transporteurs so llicités diminue avec le vieillissement).
1- Le niveau de stimulation dépend :
- du gradient pH entre les milieux intra et extra cellulaires
- du type d ’entraînement (l ’entraînement en intensité augmente la vitesse
du passage membranaire par rapport à l ’entraînement de longue durée)
(Juel et al.,1991;Dermott et Bonen,1993; Pilegaard et al.,1993; Roth et Brooks,1993; Brooks, 1999; Pilegaard, 1999; Bonen, 2000)
Glycémie ≤ 5.8 mmol.l-1 (1.05 g.l-1)Glycémie moyenne au repos et à jeun : 4.6 ± 0.4 mmol.l-1 (0.83 ± 0.O6 g.l-1)
Transport membranaire
GLUCOSE
SGLT (sodium-glucose co-transporter)SGLT2(rein)
SGLT2(intestin)
GLUT(5 isoformes)
GLUT-1 : cellules endothéliales des vais. Sanguins etdans tissus sensibles à l’insuline. Affinité élevée pour le glucose. Erythrocytes, cerveau,
GLUT-2 : cellules qui libèrent du glucose dans le sangfoie, pancréas, rein, placenta
GLUT-3 : action identique GLUT-1 Mais grande affinitépour le glucose. Cerveau, rein, placenta
GLUT-4 : Forte sensibilité à l’insuline. Muscle striés :squelettiques et cœur, tissu adipeux
GLUT-5 : Affinité pour le fructose mais nulle pour leglucose. Intestin grêle (surtout), rein, muscles striés, tissu adipeux, cerveau.

47
lactate
lactate
lactatepyruvate
NAD+NADH
C R
cytosol
mitochondrie
m.ext
m.int
LDHm
pyruvate
pyruvate
Transfert et oxydation du lactate cytoplasmique dan s la mitochondrie. D'après [Brooks, 1999 ], modifié.
C R = chaîne respiratoire.
m. ext = membrane externe de la mitochondrie; m. in t.= membrane interne de la mitochondrie
LDHc
3.2 EST-CE L’ABSENCE D’OXYGENE QUI ENTRAINE LA FORMATION ET L’ACCUMULATION DU LACTATE DANS
LE MUSCLE ?
3.2 EST-CE L’ABSENCE D’OXYGENE QUI ENTRAINE LA FORMATION ET L’ACCUMULATION DU LACTATE DANS
LE MUSCLE ?
��Localement dans la cellule musculaireLocalement dans la cellule musculaire
Les travaux de Connet et Coll. (1984) montrent aucun gradient de PO2
inférieur à 2 mm Hg alors que la PO2 minimale nécessaire pour assurer une
activité maximale de la phosphorylation oxydative est inférieure à 0.5, voire
0.1 mm Hg. (Chance et Quirstorff,1978)
� Il est fréquent de lire qu’« il y a formation de lactate en absence d’oxygène »
Ceci est exact mais ... dans un tube à essai !
��Globalement au niveau des muscles actifsGlobalement au niveau des muscles actifsLes travaux de Pirnay et Coll. (1972) montrent qu’au cours d’un exercice
maximal (= à VO2 max), la PO2 du sang veineux effluent ne s’abaisse pas
au dessous de 20 mm Hg.

48
EN CONSEQUENCE :EN CONSEQUENCE :
� Malgré sa production et son accumulation du lactate, le muscle squelettique qui travaille (même à puissance maximale = à VO2 max) N’EST EN HYPOXIE, ni globale-ment, ni localement.
� Il y a toujours plus d’oxygène que la quantité maximale susceptible d’être utilisée par le muscle.
� Ainsi l’hypothèse sous-jacente à la théorie du « Seuil anaérobie » selon laquelle le muscle produit du lactate car il est en hypoxie au delà d’une certaine puissance « seuil »n’est pas confirmée.
HYPOTHÈSES...HYPOTHÈSES...
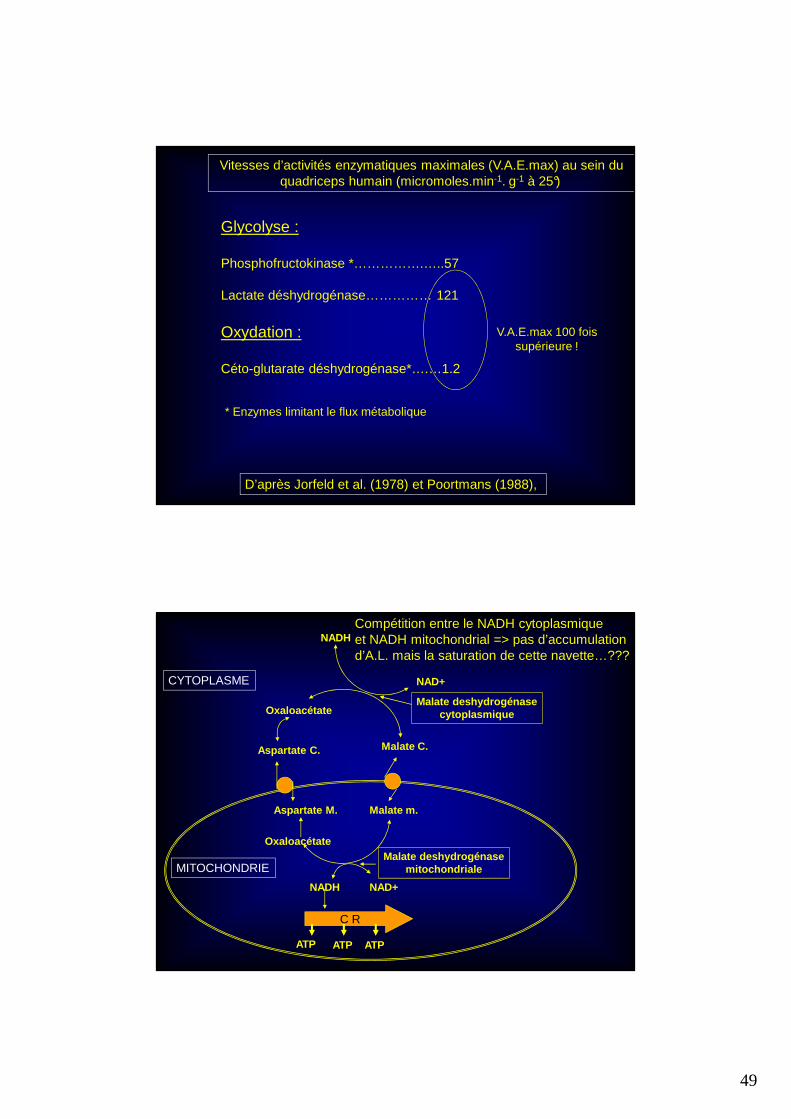
L’accumulation du lactate pourrait être due :
�A la différence entre l’activité enzymatique maximale de la lactate déshydrogénase (en amont) et celle de la céto-glutarate déhydrogénase (en aval) qui sont deux des enzymes limitant le flux métabolique respectivement : de la glycolyse et de l’oxydation.
� A la saturation de la navette aspartate - malate qui permet le transfert membranaire mitochondrial des protons H+.
� Au niveau d’activation des protéines permettant le transport transmembranaire du lactate vers le milieu extracellulaire.
49
Vitesses d’activités enzymatiques maximales (V.A.E.max) au sein du quadriceps humain (micromoles.min-1. g-1 à 25°)
Glycolyse :
Phosphofructokinase *…………….…..57
Lactate déshydrogénase…………… 121
Oxydation :
Céto-glutarate déshydrogénase*….…1.2
* Enzymes limitant le flux métabolique
D’après Jorfeld et al. (1978) et Poortmans (1988),
V.A.E.max 100 foissupérieure !
Malate m.
Oxaloacétate
Aspartate M.
Malate C.Aspartate C.
Oxaloacétate
NAD+
NADH
NADH NAD+
C R
ATP ATP ATP
Malate deshydrogénasemitochondriale
Malate deshydrogénasecytoplasmique
MITOCHONDRIE
CYTOPLASME
Compétition entre le NADH cytoplasmique et NADH mitochondrial => pas d’accumulationd’A.L. mais la saturation de cette navette…???
50
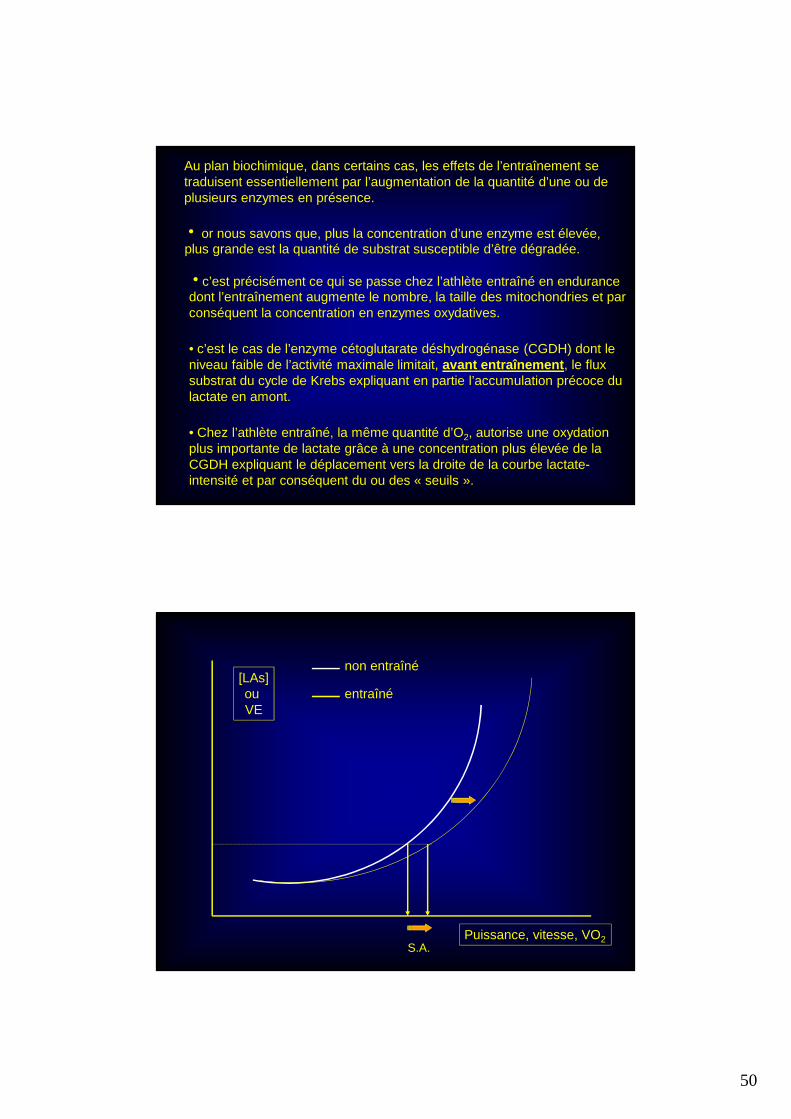
• Chez l’athlète entraîné, la même quantité d’O2, autorise une oxydation plus importante de lactate grâce à une concentration plus élevée de la CGDH expliquant le déplacement vers la droite de la courbe lactate-intensité et par conséquent du ou des « seuils ».
Au plan biochimique, dans certains cas, les effets de l’entraînement se traduisent essentiellement par l’augmentation de la quantité d’une ou de plusieurs enzymes en présence.
h or nous savons que, plus la concentration d’une enzyme est élevée, plus grande est la quantité de substrat susceptible d’être dégradée.
hc’est précisément ce qui se passe chez l’athlète entraîné en endurance dont l’entraînement augmente le nombre, la taille des mitochondries et par conséquent la concentration en enzymes oxydatives.
• c’est le cas de l’enzyme cétoglutarate déshydrogénase (CGDH) dont le niveau faible de l’activité maximale limitait, avant entraînement , le flux substrat du cycle de Krebs expliquant en partie l’accumulation précoce du lactate en amont.
[LAs]ou VE
Puissance, vitesse, VO2S.A.
non entraîné
entraîné








![La Premier Guerre Mondiale Atp[1][1]](https://img.pdfslide.fr/doc/110x75/55c0397abb61eb58208b45b6/la-premier-guerre-mondiale-atp11.jpg)




















