Embed Size (px)
Citation preview

Document réalisé par H.Belhocine 2011/2012
1
H.Belhocine
Génétique humaine Rappels de cours et exercices corrigés
Cours destinés aux étudiants de première année de médecine et chirurgie
dentaire

Document réalisé par H.Belhocine 2011/2012
2
Hérédité Mendélienne
Résumé du cours I- MONOHYBRIDISME & DIHYBRIDISME :
Pour le monohybridisme il y a : 1 caractère, 1 gène, 1 locus, 2 allèles, 1 paire de chromosomes homologues Pour le dihybridisme il y a : 2 caractères, 2 gènes, 2 locus, 2 couples d’allèles, 1ou 2 paires de chromosomes homologues
I-1 Le monohybridisme I-1-1 Les proportions Mendéliennes : a- croisement de deux hétérozygotes :
A/a X A/a → 3/4 [A] + 1/4[a] b- Croisement test-cross : a/a (homozygote récessif) X individu de génotype inconnu, dans ce croisement la fréquence des individus de la descendance est
égale à la fréquence des gamètes de l’individu de génotype inconnu.
§ Le but du test- cross est la détermination du génotype de l’individu
testé, en déterminants ses gamètes.
c- bak-cross :
Croisement d’un individu avec l’un de ses parents, le but étant
la purification des lignées animales ou végétales, c'est-à-dire
l’obtention d’individus homozygotes.

Document réalisé par H.Belhocine 2011/2012
3
I-2 Le dihybridisme, gènes indépendants : Croisement de deux doubles hétérozygotes :
A/a B/b X A/a B/b → 9/16[AB]; 3/16[Ab]; 3/16[aB]; 1/16[ab] Chaque individu donnera 4 types de gamètes avec une fréquence de
¼ chacun
1/4 gamètes [AB]; 1/4 gamètes [Ab]; 1/4 gamètes [aB]; 1/4 gamètes [ab]. II - LISTE DE QUELQUES MALADIES AUTOSOMIQUES
Nom de la maladie
phénotype
allèle
caractéristiques
albinisme
Dépigmentation de
la peau et des
phanères
a
récessif
Peut être polygénique
phénylcétonurie
Présence de
phénylcétones
dans les urines,
retard mental
p
récessif
Dentinogenesis
imperfecta
Denture anormale,
coloration brune et
s’usent facilement
D
dominant
Létale à l’état homozygote
D/D
Brachydactylie
Doigts et orteils
courts
B
dominant
Létale à l’état homozygote
B/B
Progeria
Vieillissement
rapide conduisant
à une mort précoce
P
dominant
Néomutations

Document réalisé par H.Belhocine 2011/2012
4
Exercices Exercice N° 1 : monohybridisme
Un homme homozygote aux cheveux frisés épouse une femme
homozygote aux cheveux raides.
Sachant que le phénotype cheveux frisés est dominant par rapport au
phénotype cheveux raides.
a) Ecrire les génotypes et phénotypes des parents.
b) Donner les proportions phénotypiques de la 1ere et 2eme
générations
Solution 1 :☺
a) Le symbole pour chaque allèle est choisis en prenant la première lettre du phénotype dominant, on l’écrits en majuscule et la même lettre est écrite en minuscule pour le phénotype récessif. Frisé dominant, donc allèle F Raide récessif donc allèle f b) la première génération et le résultat du croisement de deux homozygotes F/F * f/f → 4/4 [F], soit 100% de phénotype dominant. La deuxième génération est le croisement de deux hétérozygotes F/f * F/f → 3/4 [F], 1/4[f]
__________________________________

Document réalisé par H.Belhocine 2011/2012
5
Exercice N° 2 : monohybridisme
Un homme atteint de dentinogenesis imperfecta épouse une femme à
denture normale.
a) donner les génotypes des parents.
b) quelle est la probabilité pour ce couple d’avoir des enfants
atteint de dentinogenesis ? Donner leur génotype.
Solution 2 :
a)Pour répondre à cette question, il faut savoir que la dentinogenesis imperfecta est un caractère contrôlé par un allèle autosomique dominant, létal (provoque la mort) à l’état homozygote. L’allèle de la dentinogenesis est donc ‘D’ et il n’existe pas de génotype D/D.
Le génotype de la femme est d/d et celui de l’homme ne peut être que D/d.
b) La probabilité d’avoir des enfants atteints est le résultat du croisement
D/d * d/d → 1/2[D] et 1/2[d] la probabilité est donc de ½ soit 50%
____________________________________

Document réalisé par H.Belhocine 2011/2012
6
Exercice N° 3 : monohybridisme
Soit un couple formé de deux hétérozygotes pour l’albinisme.
a) quelle est la probabilité que leur premier enfant soit albinos ?
b) quelle est la probabilité que leur deuxième enfant soit albinos, si
le premier était normal ?
c) quelle est la probabilité que les deux premiers enfants soient
albinos ?
d) quelle est la probabilité pour ce couple d’avoir 3 enfants tous
normaux ?
Solution 3 :
a) ils sont tout deux hétérozygotes A/a. La probabilité que le premier enfant soit albinos est de 1/4. b) La probabilité pour le deuxième enfant reste toujours ¼, car les probabilités ne sont pas liées. Tous les enfants ont la même probabilité d’être atteints quelque soit le phénotype des autres enfants. c) La probabilité d’avoir un enfant albinos est de 1/4, la probabilité d’avoir 2 enfants albinos est de (1/4)2 soit 1/16 ou 0,062. d) la probabilité d’avoir un enfant normal est de 3/4, la probabilité d’avoir les 3 enfants tous normaux et de (3/4)3, soit 27/64 ou 0,42.
_______________________________
Exercice N° 4 : dihybridisme, gènes indépendants
Un homme albinos avec des cheveux frisés épouse une femme qui a
des cheveux raides et une pigmentation normale. Leur premier
enfant est albinos aux cheveux raides.
a) Sachant que le phénotype cheveux frisés est dominant par
rapport au phénotype cheveux raides, donner les génotypes des
parents.
b) Donner les génotypes et phénotypes possibles pour les enfants
de ces parents et leurs pourcentages probables.

Document réalisé par H.Belhocine 2011/2012
7
Solution 4 : ☺
a) cheveux frisés F, cheveux raides f ; albinos a, pigmentation
normale A. Puisqu’ils ont un premier enfant avec un phénotype récessif [fa], la femme qui a un phénotype [FA] doit donc être hétérozygote F/f A/a et l’homme qui est récessif [fa] est f/f a/a. b) Le système branché donne les résultats ci-dessous
______________________________
Exercice N° 5 : dihybridisme, gènes indépendants
Un homme brachydactyle, à denture normale épouse une femme
atteinte de brachydactylie et de dentinogenesis.
a) écrire les génotypes des parents.
b) Quelle est, pour ce couple, la probabilité d’avoir un enfant sain
pour les deux caractères ? Justifier.
c) Quelle est, pour ce couple, la probabilité d’avoir un enfant atteint
pour les deux caractères ? Justifier.

Document réalisé par H.Belhocine 2011/2012
8
Solution 5:
a) génotype de l’homme : B/b d/d, génotype de la femme : B/b D/d, l’homme est la femme ne peuvent pas êtres homozygotes pour les deux caractères, car ils sont létaux sous cet état (homozygote). b) pour avoir un enfant [b] la probabilité est de 1/3 et pour avoir [d] c’est 1/2. La probabilité d’avoir un enfant [bd] est 1/3x1/2= 1/6, soit 0,166. c) pour avoir un enfant [B] la probabilité est de 2/3 et pour avoir [D] c’est 1/2. La probabilité d’avoir un enfant [BD] est 2/3x1/2= 2/6 ou 1/3, soit 0,666 ________________________________________________________________

Document réalisé par H.Belhocine 2011/2012
9
Exceptions au comportement Mendélien
Résumé du cours I- Le dihybridisme, gènes liés : I-1 Définition : Deux gènes liés sont deux gènes portés sur le même
chromosome.
I-2 Les types de positions : Les gènes sont dits liés en couplage (cis) si les deux allèles
dominants sont sur un chromosome et les allèles récessifs sont sur
l'autre chromosome homologue. Ils sont dits liés en répulsion (trans)
si sur chaque chromosome homologue on trouve simultanément un
allèle dominant et un allèle récessif. Voir figure ci-dessous.
I-3 Les types de liaisons :
a- liaison partielle : lorsque les deux gènes liés peuvent se
séparer avec une fréquence de recombinaison comprise
entre 0 et 1.
b- liaison absolue : les gènes ne se séparent jamais, la
fréquence de recombinaison est nulle (=0).
c- Synténie : les gènes liés sont très éloignés au point que
la fréquence de recombinaison atteint 0,5 les gènes se
comportent comme s'ils étaient indépendants.

Document réalisé par H.Belhocine 2011/2012
10
C'est pendant la méiose que les chromosomes ainsi que les allèles
forment de nouvelles combinaisons.
Conséquences génétiques de la méiose :
• Brassage intrachromosomique
• Brassage interchromosomique
• Maintient de l'état diploïde de l'espèce.
• Diversité génétique.
I-4 Distance génétique : la distance génétique est proportionnelle
à la fréquence de recombinaison entre deux gènes liés.
D (A-B) = (fréquence des gamètes recombinants pour les gènes A et B) X 100 cM
La fréquence des gamètes recombinants pour A et B s'obtient soit :
a- par l'étude du test cross, car dans un tes cross la fréquence des
gamètes recombinants est égale à la fréquence des individus
recombinants dans la descendance.
b- Par l’étude du croisement de deux hétérozygotes, dans ce cas on
se base sur le calcul de la fréquence du gamète double récessif
(ab).
Fr. gamètes (ab) = √ Fr. Individus [ab]
Fr. gamètes (ab) = Fr. gamètes (AB)
Fr. gamètes (Ab) = Fr. gamètes (aB)
Fr. gamètes (ab) +Fr. gamètes (AB) + Fr. gamètes (Ab) + Fr. gamètes (aB) =1

Document réalisé par H.Belhocine 2011/2012
11
-Si la fréquence des gamètes (ab) < 0,25 → d = (2x√ Fr. Individus [ab]) x 100
-Si la fréquence des gamètes (ab) > 0,25 → d = (1-2x√ Fr. Individus [ab]) x 100
Exercices Exercice N°1 : dihybridisme, gènes liés
Calculer les fréquences des gamètes recombinants Ab et Ab dans les
croisements suivants :
a) AB/ab * AB/ab: 30(AB); 9 (ab); 20 (Ab); 15 (aB)
b) AB/ab * ab/ab : 38 (AB); 37 (ab); 13 (Ab); 12 (aB)
Solution 1:
a)Croisement de deux hétérozygotes 1- Freq. gam. (ab) = √9/ (20+15+30+9) = √0,12 = 0,34 2- Freq. gam. (AB) = 0,34 3- Freq. gam. (Ab) = 0,50-0,34 =0,16 4- Freq. gam. (aB) = 0,50- 0,34 = 0,16 b) Croisement test-cross 1- Freq. gam. (ab) = 37/ (37+38+13+12) = 0,37 2- Freq. gam. (AB) = 38/ (37+38+13+12) = 0,38 3- Freq. gam. (Ab) = 13/ (37+38+13+12) = 0,13 4- Freq. gam. (aB) = 12/ ( 37+38+13+12) = 0,12
_________________________________
Exercice N°2 : dihybridisme, gènes liés
Un croisement entre deux homozygotes [AB] et [ab] produit en F1 le
phénotype [AB]; en F2 on obtient les résultats suivants :

Document réalisé par H.Belhocine 2011/2012
12
[AB]= 110 ; [Ab]= 16; [aB]=19; [ab]= 15
a) Les gènes des loci A et B sont-ils liés ou indépendants ?
b) Quelles proportions seraient attendues dans l'autre cas en F2 ?
Solution 2:
a) le croisement concerne deux hétérozygotes, pour savoir si les gènes sont liés il faut calculer la fréquence des gamètes (ab), si cette fréquence est de 0,25 on parlera de gènes indépendants, dans le cas contraire on parlera de gènes liés.
Freq. Gam. (ab) = √ Fr. Individus [ab] = √15/ (110 + 19 + 16 + 15) = 0,30 Les gènes sont donc liés. b) dans l’autre cas on obtient les proportions suivantes : [AB]= 9/16 ; [Ab]= 3/16; [aB]=3/16; [ab]= 1/16
__________________________________
Exercice N°3 : dihybridisme, gènes liés
Deux gènes a et b sont lés à 10%. Quels seraient les phénotypes et la
probabilité d'obtention de chaque type de descendant du croisement
suivant : Ab/aB*ab/ab ?
Solution 3:
Il faut remarquer que le croisement est un tes-cross d’un double hétérozygote. La probabilité d’obtention des individus de la

Document réalisé par H.Belhocine 2011/2012
13
descendance est égale à la probabilité d’obtention des gamètes de l’hétérozygote. Les gènes sont liés en répulsion (trans), comme le montre le génotype, les gamètes recombinants sont donc AB et ab et les parentaux sont Ab et aB. Pour trouver la proportion des individus de la descendance, il suffit de calculer la fréquence des gamètes donnés par l’hétérozygote. d= freq. Gam. (AB + ab) x100= 10 freq. Gam. (AB + ab)= 10/100= 0,1 et freq. Gam. (AB)= freq. Gam. (ab)= 0,05 freq. Gam. (Ab+ aB)= 1-0,1 = 0,9 et freq. Gam. (Ab)= freq. Gam (aB) = 0,45 Donc [AB] = 0,05 [ab] = 0,05 [Ab]= 0,45 [aB] = 0,45

Document réalisé par H.Belhocine 2011/2012
14
Résumé du cours II- codominance et absence de dominance: On parle de codominance lorsque deux allèles ont la même force
d'expression, ce ci ce traduit par l'expression des deux allèles dans le
phénotype. On peut citer pour cela l'exemple des groupes sanguins
ABO où il y a codominance entre l'allèle IA et l'allèle IB, un individu
de génotype IAIB aura un phénotype AB exprimant les deux
antigènes (Ag. A et Ag. B) dans le même globule rouge.
On parle d'absence de dominance lorsque aucun allèle ne domine
l'autre et aucun ne s'exprime dans le génotype hétérozygote, ceci se
traduit par l'apparition d'un phénotype intermédiaire. L'exemple de
la fleur mirabilis Jalapa illustre bien ce cas ; le croisement d'une
fleure rouge avec une fleure blanche donne des fleurs roses, la couleur
rose est intermédiaire entre le rouge et le blanc.
Codominance : hématie avec antigène A et antigène B chez un
individu de génotype IA/IB
Ccl : les deux allèles s’expriment complètement.
+
Absence de dominance : La fleure (mirabilis Jalapa) obtenue dans
ce croisement montre un phénotype rose, intermédiaire du rouge et
du blanc. Aucun des deux allèles ne s’est exprimé complètement.
B
B
A
A
H .Belhocine

Document réalisé par H.Belhocine 2011/2012
15
III- Polygénie : C’est le contrôle d'un caractère par plusieurs gènes. Il y a donc
plusieurs couples d'allèles qui occupent un ou plusieurs
chromosomes. On cite pour cela l'exemple du groupe sanguin rhésus
qui sont contrôlés par 3 gènes (C, D, E), avec comme gène majeur, le
gène "D". Les gènes C et E sont des gènes mineurs et pour l'écriture
du génotype, seul les allèles du gène "D" sont représentés.
Remarque : Les gènes C, D et E sont liés.
Diagramme : polygénie
Gène (n)
Gène 3
Gène2
Gène 1
Caractère
Document réalisé par H.Belhocine 2011/2012
16
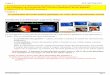
IV Polyallélisme : On parle de polyallélisme lorsqu’un caractère est
contrôlé par un gène représenté par plusieurs allèles situés au même
locus.
Caractère
Allèle 1
Ou Allèle 2
Ou Allèle n)
Allèle 1
Ou Allèle 2
Ou Allèle n)
Diagramme : polyallélisme
Chromosome paternel Chromosome maternel

Document réalisé par H.Belhocine 2011/2012
17
V- Pléiotropie : Il s'agit du contrôle de plusieurs caractères par un seul gène. La
pléiotropie se produit lorsque le produit du gène agit à différents
niveaux d'un organisme (protéine de structure) ou au début d'une
chaîne métabolique (enzyme).
Ex; Syndrome de MARFAN et ANEMIE FALCIFORME.
Hématie falciforme
Caractère(n)
Caractère 2
Caractère 1
1gène
Diagramme : pléiotropie

Document réalisé par H.Belhocine 2011/2012
18
Exercices
Exercice N°1 : codominance et absence de dominance, polyallélisme.
Un homme de groupe sanguin AB+ et son épouse A- ont eu 3
enfants.
Walid: B+; Karim O+; Malika: A+
a) Vous est-il possible de déterminer le génotype de la mère et du
père ?
b) Les parents craignent qu'il n'y ait eu, à la maternité, échange
pour l'un de leurs enfants; ces craintes vous semblent-elles
justifiées ?
Dans l'affirmative quel est l'enfant concerné ?
c) Est-il possible de connaître avec précision le groupe sanguin du
véritable père de cet enfant, si la mère présumée est de
groupe identique à celle sus-citée ?
Solution 1 : ☺
a) Le père est IA/IB D/d ou IA/IB D/D, la mère est forcément IA/io d/d (elle ne peut pas être IA/IA car elle a un fils, Walid qui est de groupe B).
b) Oui, l’enfant concerné est Karim. (le père de groupe AB ne peut
pas avoir d’enfants de groupe O).
c) Non, car il y a plusieurs possibilités pour le père de Karim, parmi lesquelles ; IA/io D/d, IB/io D/d, IA/io D/D etc...
______________________________

Document réalisé par H.Belhocine 2011/2012
19
Exercice N°2 : codominance et absence de dominance
a)Une femme de groupe sanguin B, N a un enfant de groupe sanguin
O, MN. Elle affirme qu'un homme de groupe sanguin A, M est le
père de l'enfant. Cette éventualité peut-elle être exclue, connaissant
ses groupes sanguins ?
b) Si l'homme et la femme étaient tous les deux Rh positif et l'enfant
Rh négatif,
Cette éventualité pourrait-elle être exclue ?
c)Si ses groupes sanguins étaient A, N Rh négatif, quelle serait votre
conclusion ?
Solution2 : ☺
a) Non, car l’enfant O, MN peut avoir un père AM (génotype IA/io M/M) est une mère BN (génotype IB/io N/N).
b) Non, car les parents peuvent êtres tout les deux D/d et l’enfant
d/d.
c) On peut conclure que l’homme ne peut pas être le père de
l’enfant, car un homme de groupe ne peut pas avoir d’enfants de groupe N.
_______________________________
Exercice N°3 : codominance et dominance/récessivité.
La recherche des groupes sanguins des membres d'une famille a
donné l'arbre généalogique ci- dessous :

Document réalisé par H.Belhocine 2011/2012
20
Dans cette famille sévit une maladie héréditaire représentée en
noir. Cette maladie se caractérise par des hémorragies nasales
fréquentes à partir de 8 à 10 ans.
1- Préciser les génotypes des individus : I1, I2, II2, III2.
2- Y a-t-il un enfant intrus dans cette famille ? Lequel et
pourquoi ?
- Les filles III7 et III8 de cette famille sont-elles de vraies ou
fausses jumelles ? Pourquoi ?
.
Solution 3:
1) I1 et I2 sont IA/iO et IB/iO. d’après l’arbre généalogique la maladie est récessive, puisqu’il y a saut de génération en II4 et II5, I1 est donc IA/iO M/m et I2 : IB/io m/m.II2 est IB/iO m/m, III2 est IB/IB m/m ou IB/io m/m.
2) L’enfant intrus est le III9 car il est de groupe B et
aucun des deux parents ne porte l’allèle B.

Document réalisé par H.Belhocine 2011/2012
21
3) Les filles sont de fausse jumelles d’abord par rapport au symbole qui est celui de fausses jumelles, puis par rapport à la maladie pour la quelle une qui est malade et l’autre non
________________________________
Exercice N°4 : polygénie
Une femme qui a été atteinte dans son enfance de la maladie
hémolytique du nouveau né, se marie avec un homme qui n'a pas eu
la maladie hémolytique du nouveau né, mais qui a plusieurs
germains plus âgés qui en on était atteints.
a) Donner les génotypes probables du couple et de leurs parents.
b) S'ils ont une famille nombreuse, quel risque ont-ils d'avoir un
enfant atteint de la maladie hémolytique ?
Solution 4 :
a) La femme atteinte est de génotype D/d, puisqu’ elle a été atteinte dans son enfance de la MHNN. L’homme est de génotype d/d puisque il n’a pas été atteint alors que ses frères et sœurs étaient atteints. La mère de la femme est d/d. Le père de la femme est D/D ou D/d.
La mère de l’homme est d/d et son père D/d.
b) Le risque est de 0, puisque la femme est rhésus positif, elle ne produit pas d’anticorps anti-rhésus contre les antigènes rhésus d’un fœtus éventuel « rhésus positif ».
____________________________

Document réalisé par H.Belhocine 2011/2012
22
Résumé du cours
VI- Hérédité liée au sexe :
VI-1) Définition : caractères contrôlés par des gènes situés sur les
gonosomes.
VI-2) chromosomes X et Y :
Le chromosome X comporte des gènes qui jouent un rôle vital. La
présence d'au moins un chromosome X est indispensable pour la
viabilité de l'individu.
Le chromosome Y ne comporte pas de gènes vitaux, il comporte
essentiellement des gènes du déterminisme du sexe mâle (ex. sry et
zfy qui permettent le développement des gonades mâles) et les gènes
de la spermatogenèse.
Même si le X et Y ne sont pas homologues, il existe une petite région
homologue entre X et Y dite région pseudo autosomique à chaque
extrémité.
Remarque : Les caractères dont les gènes sont portés par le
chromosome Y sont dits des caractères HOLANDRIQUES. Pour ce
type de gènes l'homme ne peut être ni homozygote ni hétérozygote,
mais HEMIZYGOTE, car les chromosomes X et Y n'ont pas les mêmes
gènes (ils ne sont pas homologues).
VI-3) Phénomène de Lyonisation : Le phénomène de lyonisation
permet d'éviter une « surproduction » des protéines codées par le
chromosome X chez la femme par rapport à l'homme. Ce phénomène
correspond à l'inactivation de l'un des chromosomes X chez la femme
par condensation sous forme d'un corpuscule de Barr ; ce dernier est
visible après coloration au Giemsa, il est localisé contre la face
interne de l'enveloppe nucléaire (lamina). Cette condensation débute
très tôt au cours de la vie embryonnaire, le chromosome X inactivé est
pris au hasard de sorte que dans le cas normal 50 % des corpuscules
de Barr sont d'origine maternelle et 50% d'origine paternelle. Pour
cette raison la femme est dite une mosaïque génétique. Cette
inactivation est transmise d'une génération cellulaire à une autre,
même si le corpuscule de Barr se décondense au cours de la mitose.

Document réalisé par H.Belhocine 2011/2012
23
Notons en fin que malgré cette inactivation, 25% des gènes d'un
corpuscule de Barr peuvent rester actifs. Parfois il y a un déséquilibre
dans la proportion des corpuscules d'origines maternelle et
paternelle, si pour une femme conductrice le chromosome X portant
l'allèle normal est préférentiellement condensé le caractère récessif
s'exprimera chez cette femme et on parle de LYONISATION
DEFAVORABLE.
Lyonisation
(●)= corpuscule de Barr
XH et Xh : respectivement allèle normal et allèle de l ‘hémophilie
La femme est conductrice « XH/Xh » et son phénotype est normal [H]
XH XH
Xh Xh ●
●
●
●
XH
Xh
Xh
Xh
●
●
●
●

Document réalisé par H.Belhocine 2011/2012
24
Lyonisation défavorable (●)= corpuscule de Barr XH et Xh : respectivement allèle normal et allèle de l ‘hémophilie
La femme est conductrice « XH/Xh » et son phénotype est
« hémophile » [h], car 75% des chromosomes Xh sont actifs et 25%
seulement pour XH
Exercices
Exercice N°1 : Hérédité liée au sexe.
Un homme (a) soufrant d'une maladie, se marie avec une femme
normale. Toutes les filles sont malades, les garçons sont normaux, les
fils de l'homme (a) se marient à des femmes normales, alors tous
leurs enfants sont normaux. Les filles de l'homme (a) se marient à des
hommes normaux alors 50% de leurs enfants sont malades.
Expliquez ces faits.
Solution 1 :
Il s’agit d’une maladie dominante liée à X. l’homme transmet le caractère uniquement aux filles, car il ne peut transmet le chromosome X aux garçons. Les fils de l’homme (a) qui sont tous normaux, ne portent que l’allèle normal dominant qu’ils transmettent à toutes les filles qui sont par conséquent toutes normales. Les filles de l’homme (a) sont hétérozygotes elles auront des filles et des garçons normaux, des garçons et filles malades, car le X portant l’allèle de la maladie est transmit aussi bien aux filles qu’aux garçons.
__________________________________

Document réalisé par H.Belhocine 2011/2012
25
Exercice N°2 : Hérédité liée au sexe.
Parmi les petits fils d'un homme hémophile, quel est le pourcentage
de ceux qui seront hémophiles ? (En admettant que sa femme ainsi
que les conjoints de ses enfants soient génotypiquement normaux.).
Solution 2 :
D’après le diagramme ci-dessus un garçon sur 4 sera malade parmi les petits fils, c'est-à-dire, 25%.
_______________________________
Exercice N°3 : Hérédité liée au sexe.
Chez l'homme on connaît un gène holandrique responsable de
l'apparition de poils dans les oreilles externes; si les hommes à
oreilles poilues se marient avec des femmes normales
a) Donner le pourcentage des hommes qui auront les oreilles poilues
parmi la descendance.
b) Même question pour les femmes.

Document réalisé par H.Belhocine 2011/2012
26
Solution3 : ☺
a)Un gène holandrique est porté par le chromosome Y, il est donc transmit à tout les garçons, mais jamais aux filles. 100% des hommes de la descendance auront des oreilles poilues. b) Aucune femme n’aura les oreilles poilues, car elles ne reçoivent pas le chromosome Y du père.
Exercice N°4 : Hérédité liée au sexe.
Un gène récessif "h" lié au sexe est responsable de l’hémophilie chez
l’être humain. Une femme non hémophile dont le père était
hémophile épouse un hémophile.
a) Quel génotype la mère de l’époux hémophile peut-elle avoir?
b) Quelle est la probabilité que le premier enfant de ce mariage soit
hémophile?
c) A quel pourcentage de filles hémophiles issues de cette union peut-
on s’attendre?
d) Quelle proportion des enfants issus de ce mariage ne sera pas
hémophile?
Solution4 : ☺
a) La mère de l’époux hémophile est XH/Xh b) la probabilité que le premier enfant de ce mariage soit hémophile est de 1 /3. c) aucune fille hémophile, 0% d) 2/3.

Document réalisé par H.Belhocine 2011/2012
27
Résumé du cours VII- Hérédité influencée par le sexe : on parle d’hérédité
influencée par le sexe lorsqu’ un caractère est contrôlé par des gènes
autosomiques, qui se comportent de façon différente chez l’homme est
la femme.
Exemple ; la calvitie dont l’allèle est dominant chez les garçons et
récessif chez les filles. Cette dominance du gène de la calvitie chez
l’homme est due à la présence de testostérone chez l’homme.
Ainsi si ‘C’ représente l’allèle de la calvitie et ‘c’ L’allèle de chevelure
normale, la relation entre phénotype et génotype sera différente chez
les deux sexes, à savoir :
C/c donne [C] chez l’home et [c] chez a femme.
C c
C/C C /c
C ♀ [C]
♂ [C]
♀ [c]
♂ [C]
C/c c/c
c ♀ [c]
♂ [C]
♀ [c]
♂ [c]
VIII- Hérédité limitée à un sexe : dans l’hérédité limitée à un sexe
le caractère est autosomique mais ne s’exprime que chez l’un des
sexes. Exemple la puberté précoce qui ne s’exprime que chez les
garçons.

Document réalisé par H.Belhocine 2011/2012
28
Résumé du cours
IX-Hérédité mitochondriale
IX.1) Définition : L’hérédité mitochondriale dite également hérédité
cytoplasmique, suit un mode de transmission non Mendélien.
IX.2) ADN mitochondrial (ADNmit) : La mitochondrie possède son
propre ADN appelé ADNmt, circulaire, L’ADNmt est répliqué
indépendamment de l’ADN nucléaire (ADNn), mais présente 10 fois
plus de mutions que ce dernier (à cause de l’absence du système de
correction lors de la réplication). Dans la même cellule c’est ADN
peut être de même type (sauvage ou muté) dans ce cas on parle
d’homoplasmie, ou bien de types différents et on parle alors
d’hétéroplasmie.
Les gènes mitochondriaux ne possèdent pas d’introns
IX.3) Mode de transmission : Le mode de transmission est ici exclusivement maternel, c'est-à-dire
que les caractères sont transmis uniquement de la mère aux
enfants.
Remarque : Certaines fonctions mitochondriales comme la
réplication, la stabilité de l’ADN mitochondrial et la fission des
mitochondries sont contrôlées par des gènes nucléaires. Une mutation
au niveau de l’un de ces gènes peut être à l’origine d’une maladie
mitochondriale, mais à transmission Mendélienne, pouvant être
autosomique dominante ou récessive.
IX.4) les maladies mitochondriales : Les maladies
mitochondriales varient dans leur sévérité d’un état clinique non
décelable vers un état fatal, l’expressivité de la maladie dépend de la
proportion des mitochondries anormales et du tissu touché.
Les organes les plus touchés sont ceux qui ont le plus besoin
d’énergie, à savoir ; le foie, les muscles, le cerveau, les reins et les
cellules de l’appareil respiratoire.
Les cytopathies mitochondriales sont dues principalement à un
déficit de la chaine respiratoire, et se manifestent souvent avant l’âge
de 1 an.

Document réalisé par H.Belhocine 2011/2012
29
Résumé du cours
X- LA GENEALOGIE X.1 Définition : en génétique; c'est la représentation schématique
de l'histoire génétique d'une famille.
X.2 Objectifs de la généalogie : L'étude de l'arbre généalogique
permet d'une part de déterminer le mode de transmission d'un
caractère et d'autre part d'établir un conseil génétique en
déterminant le risque de récurrence d'une maladie.
X.3 Les symboles : (voir schémas)
X.4 Détermination des modes de transmissions :
X.4.1 Autosomique dominant (AD) :
- le caractère est transmis du père et de la mère de la même façon
aux garçons et aux filles.
- il n’y a pas saut de génération (un enfant malade a au moins
l'un de ses parents malade).
X.4 .2 Autosomique récessif (AR):
- le caractère est transmis du père et de la mère de la même façon
aux garçons et aux filles.
- il y a saut de génération (un enfant malade peut avoir ses deux
parents sains).
X.4 .3 Gonosomique dominant (DLX) :
- le père transmet le caractère aux filles, mais jamais aux
garçons.
- il n’y a pas saut de génération (un enfant malade a au moins
l'un de ses parents malade).
X.4. 4 Gonosomique récessif (RLX) :
- la mère transmet le caractère aux garçons, alors que les filles
sont conductrices.

Document réalisé par H.Belhocine 2011/2012
30
- il y a saut de génération (un garçon malade peut avoir ses deux
parents sains).
X.4.5 Gonosomique lié à Y (HOLANDRIQUE) :
- Le père transmet à tous les garçons, mais jamais aux filles

Document réalisé par H.Belhocine 2011/2012
31
Exercices
Exercice N°1 : généalogie
Donner le mode de transmission du caractère représenté pour chacun des arbres
représenté ci-dessous, justifiez votre réponse.
Solution 1 : Arbre A : -Dominant, car un enfant malade à au moins l’un de ses parents malade - Autosomique car le père transmet le caractère aux garçons et aux filles Arbre B : - Récessif, car le garçon malade de la 3eme génération a ces deux parents sains.

Document réalisé par H.Belhocine 2011/2012
32
- Gonosomique car la mère de la première génération transmet uniquement aux filles. Arbre C : - Dominant, car un enfant malade à au moins l’un de ses parents malade - gonosomique, car le père transmet le caractère uniquement aux filles Arbre D : -Récessif, car un garçon et une fille ont deux parents sains -Autosomique, car la fille de la deuxième génération est atteinte alors que son père est sain. Arbre E : Holandrique (porté par Y), car le père transmet le caractère à tout les garçons sans exception, mais à aucune fille.
_______________________________ Exercice N°2 : généalogie
Un homme à vision normale atteint de puberté précoce et de calvitie
épouse une femme daltonienne, atteinte de calvitie. Si la mère de
l'homme était elle-même atteinte de calvitie alors que son père ne
l'était pas.
Schématisez l'arbre généalogique de cette famille.
Solution 2 :
____________________________

Document réalisé par H.Belhocine 2011/2012
33
Exercice N°3 : généalogie
Si pour un caractère donné on ne voit jamais de transmission père
fils et jamais de filles indemnes (saines) nées d'un père malade. Quel
est le mode de transmission ? Justifier.
Solution 3 :
C’est un mode gonosomique (lié à X) dominant. Justification : Jamais de transmission père fils, car le père ne transmet pas le chromosome X aux garçons. Jamais de filles indemne car le seul X que possède le père et qui est transmis à toutes les filles véhicule un gène malade dominant.
_____________________________
Exercice N°4 : généalogie
Un homme hémophile épouse une femme conductrice de l'hémophilie.
Le couple a 4 enfants
Ali (hémophile), Rabah (non hémophile), Samira (non hémophile),
Sara non hémophile. Dans cette famille il y a eu un avortement.
Samira épouse un homme non hémophile, elle a un enfant hémophile.
a- Dresser l'arbre généalogique de cette famille
b- Définir les génotypes de Samira, Rabah et Ali
c- Donner les fréquences phénotypiques des enfants de Samira
d- Expliquer pourquoi il y a eu un avortement dans la famille?
e- Si Samira et son mari avaient la calvitie, on supposant que le
mari est hétérozygote, donner les fréquences phénotypiques des
enfants de Samira pour l'hémophilie et la calvitie.
Document réalisé par H.Belhocine 2011/2012
34
Solution 4 :
b-Samira : XHXh ; Rabah XHY ; XhY
c-1 /2 filles [H] ; 1/4 Garçons [H] ; 1/4 Garçons [h]
d-L’avortement correspond à un embryon de sexe femelle de génotype XhXh
e- 1/4♀[HC] ; 1/4♀[HC] ; 1/4♂[HC] ; 1/4♂[hC]
______________________________

Document réalisé par H.Belhocine 2011/2012
35
Résumé du cours
XI- HEREDITE MULTIFACTORIELLE
XI.1-Définition : on parle d’hérédité multifactorielle lorsque
l’expression d’un caractère dépend de l’action d’un ensemble de gènes
et de l’effet de l’environnement.
Phénotype= Gènes + effets de l’environnement
XI.2- Notion d’héritabilité : l’héritabilité désigne la part des gènes
par rapport à l’effet de l’environnement dans l’expression d’un
caractère donné, dans une population donnée.
XI.3- Notion d’anticipation : dans l’hérédité multifactorielle une
maladie où un syndrome peut avoir une manifestation clinique plus
précoce ou plus sévère chez les enfants que chez leurs parents. Ce
phénomène est appelé phénomène d’anticipation.
XI.4- Notion d’allèles de susceptibilité : c’est un ensemble
d’allèles de plusieurs gènes différents, dont la combinaison constitue
un terrain génétique de prédisposition.
XI.5 Notion de gène majeur de susceptibilité : c’est un gène qui
détermine la susceptibilité vis-à-vis d’une maladie, mais dont l’effet
est modulé par un ensemble de gènes mineurs et de l’environnement. XI.6- Types d’hérédités multifactorielles :
XI.6-1 Hérédité multifactorielle à seuil : la maladie est contrôlée
par des allèles de susceptibilité et l’environnement.
Exemples : fente labio-palatine, sténose du pylore, Spina-bifida,
luxation de la hanche, pied bot.
XI.6-2 Hérédité multifactorielle mixte : dans ce cas il y a
intervention d’un gène majeur de susceptibilité, un ensemble de
gènes mineurs et des facteurs de l’environnement.
Exemples : HTA, asthme, DNID, certains cancers
XI.7- Estimation du risque de récurrence : le risque de
récurrence dans le cas d’un caractère multifactoriel et dit risque

Document réalisé par H.Belhocine 2011/2012
36
empirique. Le calcul de ce risque est basé sur des observations
effectuées dans la population générale et non pas sur un model
mathématique précis.
XI.7-1- Risque relatif : c’est le rapport du risque pour l’apparenté
d’un malade d’être atteint sur le risque dans la population générale.
Risque relatif = Risque pour l’apparenté/Risque dans la population générale
Exemple :
Parenté
% atteints
Risque relatif
Fratrie
4,1
X40
Enfants
3,5
X35
Oncles/tantes
0,7
X7
Neveux
0,8
X8
Risque relatif pour la fente labio-palatine en fonction de la
proximité génétique, par rapport à la population générale
(1/10000)
La proximité génétique représente le pourcentage de gènes en
commun entre deux individus.
Proximité génétique = (1/2) degré de parenté ou p= (1/2) d
Document réalisé par H.Belhocine 2011/2012
37
Exemple :
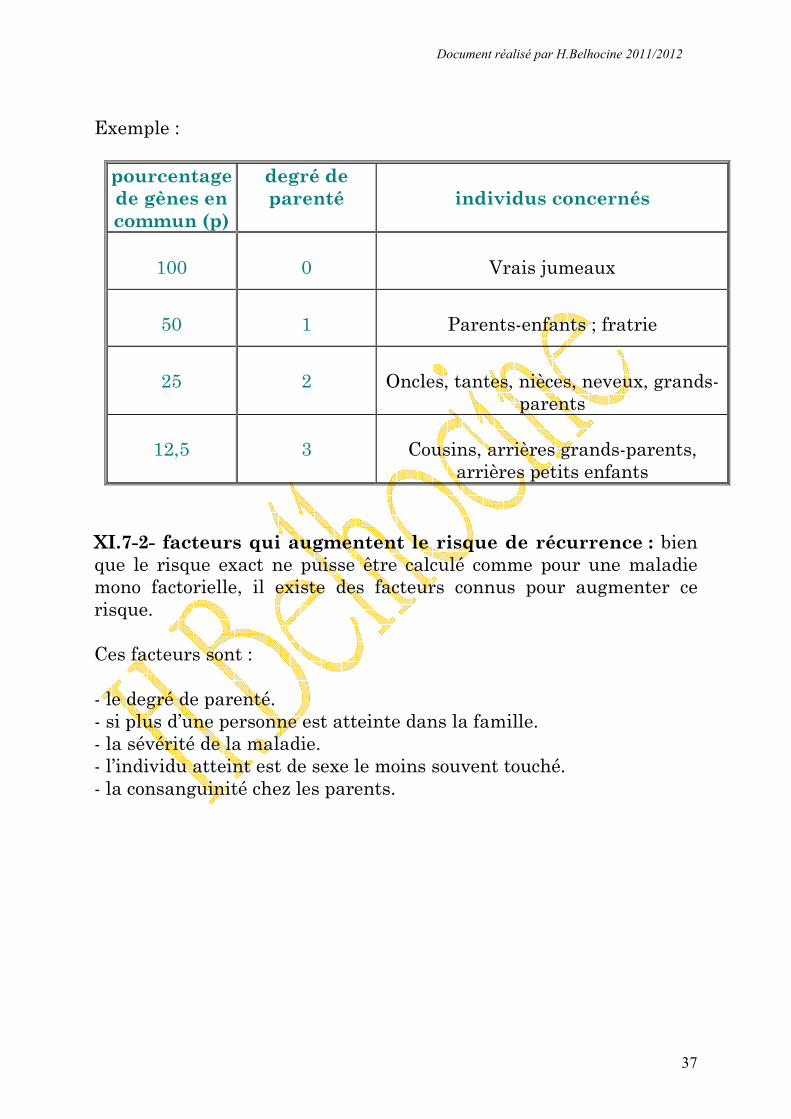
pourcentage de gènes en commun (p)
degré de parenté
individus concernés
100
0
Vrais jumeaux
50
1
Parents-enfants ; fratrie
25
2
Oncles, tantes, nièces, neveux, grands-
parents
12,5
3
Cousins, arrières grands-parents,
arrières petits enfants
XI.7-2- facteurs qui augmentent le risque de récurrence : bien
que le risque exact ne puisse être calculé comme pour une maladie
mono factorielle, il existe des facteurs connus pour augmenter ce
risque.
Ces facteurs sont :
- le degré de parenté.
- si plus d’une personne est atteinte dans la famille.
- la sévérité de la maladie.
- l’individu atteint est de sexe le moins souvent touché.
- la consanguinité chez les parents.

Document réalisé par H.Belhocine 2011/2012
38
Résumé du cours XII-Le conseil génétique
XII.1) BUT
Le but du conseil génétique est une meilleure prise en charge du
malade et d’aider la famille à y faire face.
XII.2) LES INDICATIONS DU CONSEIL GENETIQUE
Les indications du conseil génétique sont :
- antécédents de retard mental dans la famille
- antécédents d’enfants malformés dans la famille
- avortements répétés
- antécédents d’aberrations chromosomiques dans la famille
- couple consanguin
- femme âgée de plus de 35 ans
XII.3) CALCUL DU RISQUE DE RECURRENCE :
a- Pour un caractère autosomique récessif : Si les deux parents sont
phénotypiquement sains, le risque est de ¼.
R = Pht X mht x 1/4
R : risque de récurrence
(Pht) = probabilité que le père soit hétérozygote
(mht) = probabilité que la mère soit hétérozygote
1/4= Ehm = probabilité que l’enfant soit homozygote
La récurrence des maladies récessive est favorisée par la
consanguinité, qui est définie par le coefficient de consanguinité d’un
individu. Ce coefficient indique la probabilité pour qu’un individu soit

Document réalisé par H.Belhocine 2011/2012
39
autozygote (individu ayant reçu deux allèles identiques par
descendance d’un même ancêtre) en un locus donné.
En pratique le coefficient de consanguinité d’un individu est égal au
coefficient de parenté de ses parents.
Le coefficient de parenté représente la probabilité pour que deux
individus apparentés possèdent deux allèles identiques hérités d’un
ancêtre commun.
Le coefficient de parenté entre deux individus est calculé par la
relation :
ex, y = ∑ (FA+1) (1/2) p+m+1
Où FA = coefficient de consanguinité de l’ancêtre commun à « p »
et « m ».
P = nombre de générations entre le père et l’ancêtre commun
m= nombre de générations entre la mère et l’ancêtre commun
ex, y = coefficient de parenté entre l’individu x et l’individu y
∑ = somme des relations pour un ensemble d’ancêtres communs
b- pour un caractère autosomique dominant :
-En pratique des parents sains ont une probabilité nulle (0) d’avoir
des enfants atteints, sauf dans le cas d’une nouvelle mutation ou de
pénétrance incomplète du caractère chez l’un des parents.
-Si l’un des parents est atteint (généralement hétérozygote) le risque
de récurrence est de 0,5.
-Si les deux parents sont atteints le risque de récurrence est de 0,75
c- Pour un caractère gonosomique(X) dominant :
Le risque de récurrence dépend du sexe du parent atteint :

Document réalisé par H.Belhocine 2011/2012
40
- Si c’est le père qui est atteint, le risque de récurrence pour les
garçons est de « 0 », car le père ne transmet pas le « X » à ses garçons.
Par contre le risque de récurrence pour les filles est de (1), car le X du
père est transmis à toutes les filles.
- Si c’est la mère qui est atteinte, la récurrence pour les filles et les
garçons est la même, elle est de 0,5 pour les deux sexes.
d- Pour un caractère gonosomique(X) récessif :
- si c’est le père qui est atteint, le risque de récurrence est nul pour les
deux sexes.
- si c’est la mère qui est atteinte, le risque de récurrence est de 1 pour
les garçons (car hémizygote) et nul pour les filles (conductrices).
- si les deux parents sont sains, on recherche au niveau de l’arbre
généalogique la probabilité que la mère soit vectrice. Le risque de
récurrence dépend de cette probabilité.
e- pour un caractère holandrique:
- dans ce cas le risque est de 1 pour les garçons mais nul pour les
filles.
f- pour un caractère multifactoriel :
Pour un caractère multifactoriel, il est très difficile de faire un calcul
de risque exact.

Document réalisé par H.Belhocine 2011/2012
41
Exercices
Exercice N°1 : hérédité multifactorielle/ conseil génétique
Calculer le coefficient de consanguinité d'un enfant issu d'un mariage
entre un cousin et sa cousine sur 4 générations.
Solution 1 :
coefficient de consanguinité, calcul direct La probabilité qu'un allèle (a1, a2, a3 ou a4) présent à la génération I, soit transmis à l'enfant IV1 est 1/8 (1/2 à chaque génération) si l'on considère indépendamment la voie passant par le père ou la mère. Par conséquent, la probabilité pour que cet allèle soit transmis par les deux parents est 1/64. Comme il y a quatre allèles à la génération I qui peuvent êtres transmis, la probabilité pour que l'enfant IV1 reçoive deux fois le même allèle est 4 X (1/64) = 1/16. Le coefficient de consanguinité est 1/16. Dans ce cas, pour l’équation : p=m=2
(1/2) 5 + (1/2) 5 = 1/16
a 1 a 2 a 3 a 4
I
II
III
IV
1 2
1 2 3 4
1 2
1

Document réalisé par H.Belhocine 2011/2012
42
Exercice N°2 : hérédité multifactorielle/ conseil génétique
Calculer le risque de récurrence de la mucoviscidose (autosomique
récessif)
Pour un couple sain, non apparenté dans une population qui compte
1/25 d’hétérozygotes.
Solution 2 :
Chacun dans le couple a 1/25 de risque d’être hétérozygote, si tout les deux sont hétérozygotes ils on une chance sue 4 d’avoir un enfant homozygote, donc malade soit : 1/25 X 1/25 X 1/4 = 1/2500
Exercice N°3 : hérédité multifactorielle/ conseil génétique
Calculer le risque de récurrence de la mucoviscidose (autosomique
récessif) pour un couple sain, non apparenté, avec un premier enfant
atteint dans une population qui compte 1/25 d’hétérozygotes.
Solution 3 :
Le couple a un premier enfant atteint, les deux parents sont donc tout les deux hétérozygotes (la probabilité d’être hétérozygote = 1)
Risque de récurrence = 1X1X1/4= 1 /4
Exercice N°4 : hérédité multifactorielle/ conseil génétique
Calculer le risque de récurrence de la mucoviscidose (autosomique
récessif) pour un couple sain, non apparenté, avec un membre du
couple qui a un neveu atteint dans une population qui compte
1/25d’hétérozygotes

Document réalisé par H.Belhocine 2011/2012
43
Solution 4 :
Le membre du couple qui a un neveu atteint a 1/2 de chance d’être hétérozygote (puisque son frère qui a un garçon atteint est forcément hétérozygote), l’autre membre du couple a 1/25 de chance d’être hétérozygote. Le risque de récurrence = 1/25 X 1/2 X 1/4 = 1/200 Exercice N°5 : hérédité multifactorielle/ conseil génétique
Calculer le risque de récurrence de la mucoviscidose (autosomique
récessif) pour un couple sain, non apparenté, avec un membre du
couple qui a un neveu atteint et l’autre membre hétérozygote, dans
une population qui compte 1/25d’hétérozygotes Solution 5 : Même raisonnement que pour l’exercice précédent avec un risque de 1 pour l’hétérozygote, soit : Le risque de récurrence = 1 X 1/2 X 1/4 = 1/8
?
1 1/2 1/25

Document réalisé par H.Belhocine 2011/2012
44
XIII-CYTOGENETIQUE
I. LES ANOMALIES DE NOMBRE DES CHROMOSOMES : C’est
une variation du nombre de chromosomes, on distingue les
aneuploïdies et les polyploïdies.
I.1. Les aneuploïdies
Les aneuploïdies se traduisent par une modification du nombre total
de chromosomes ou. Les plus fréquentes sont les trisomies (2n+1) et
les monosomies (2n-1) dont l’origine est souvent, une non disjonction
méiotique. Cette non disjonction conduit à la formation de gamètes à
(n+1) ou à (n-1).
I.1.1 - Les trisomies
Le nombre total de chromosomes est de 47, car il existe un
chromosome en 3 exemplaires au lieu de deux. Ces anomalies peuvent
concerner les autosomes, dans ce cas on parle de trisomie autosomique, dont la majorité avorte. Il y a néanmoins une trisomie
autosomique fréquente et viable, c’est la trisomie 21 ou syndrome de
Down (47, XY, +21). Les autres trisomies comme la trisomie 13 ou
syndrome de Pateau et trisomie 18(47, XY, +13) ou syndrome
d’Edwards sont létales. (47, XY, +18)
La trisomie peut concerner les gonosomes, dans ce cas on parle de
trisomies gonosomiques, qui sont viables, par exemple : 47, XXY ;
47, XXX ; 47, XYY.
I.1.2 - Les monosomies
Le nombre total de chromosomes est de 45, car il existe un
chromosome en 1 exemplaire au lieu de deux. Les monosomies autosomiques ne sont jamais viables et provoquent un nombre
important d’avortements précoces. Parmi les monosomies
gonosomiques seule la monosomie X ou syndrome de Turner est
viable, avec un caryotype 45, X0 ou 45,X.
I.2. Les polyploïdies
Ce sont les cas dont le multiple de n est supérieur à 2. Elles sont
généralement provoquées par une dispérmie ou des gamètes à 2n. Le
cas le plus fréquent est la triploïdie (3n : 69, XXY ; 69, XXX ; 69,
XYY) qui est d’ailleurs non viable.

Document réalisé par H.Belhocine 2011/2012
45
II. LES ANOMALIES DE STRUCTURE DES CHROMOSOMES :
Les anomalies chromosomiques de structure peuvent être
équilibrées lorsqu’il n y a ni perte ni gain de matériel génétique
(donc pas d’effet sur l’individu).Dans le cas contraire elles sont non
équilibrées. On remarque que les anomalies chromosomiques de
structure les plus fréquentes sont les délétions et les translocations.
II.1 Les délétions : Elles peuvent être des
a- délétions terminales : Il y a perte d’un segment à l’extrémité du chromosome
b- délétions interstitielles Il y a perte d’un segment à l’intérieur du chromosome
Ces anomalies sont non équilibrées et l’expression phénotypique
dépends de la taille et du contenu du segment perdu.
-Exemples : 46, XY, del (13) (p32) : Délétion terminale du chromosome
13 avec un point de rupture situé au niveau de bande 2 de la région 3
du bras court.
46, XX, del (12) (q31 q21) : délétion interstitielle du chromosome 12
avec deux points de cassures bras long région 3 bande 1 et bras long
région 2 bande 1
En générale l’écriture obéit à la règle suivante :
Nombre de chromosomes, sexe de l’individu, abréviation de l’anomalie
(chromosome concerné) (bras région bande)
II.2 Les translocations : C’est le transfert d’un segment
chromosomique d’un chromosome vers un autre, qui ne lui est pas
homologue.
II.2.1 - translocations réciproques :
C’est un échange réciproque de segments chromosomiques (en dehors
des centromères) entre deux chromosomes non homologues. On
observe un déséquilibre dans 10% des cas seulement, lorsque le point
de cassure touche un gène.
-Exemple : 46, XY, t (9; 22) (q34; q11) : Translocation entre les
segments chromosomiques des bras longs des chromosomes 9 et 22
(retrouvées dans les leucémies myéloïdes chroniques).

Document réalisé par H.Belhocine 2011/2012
46
II.2.2-translocations robertsoniennes :
Elles sont également dites fusions centriques, car souvent la cassure
se produit au niveau du centromère. La conséquence est la fusion des
bras longs de deux chromosomes différents pour former un seul
chromosome, comme ce type de translocation touche les chromosomes
acrocentriques (13, 14, 15, 21,22), qui ont un bras court très petit
renfermant exclusivement des gènes des ARNr. Le chromosome formé
par les bras court est négligeable et on passe de 46 à 45
chromosomes, sans altérer le phénotype.
-exemple : 45, XX, t (14q 21q) : translocation robertsonienne entre
deux segments des bras longs des chromosomes 14 et 21
II.3- Les insertions (additions) : C’est un segment chromosomique
d’un chromosome qui est inséré dans un autre chromosome
II.4. Les inversions : Les inversions sont formées suite à une cassure
en deux points d’un segment chromosomique suivit d’une rotation de
180°, faisant intervenir un processus de recombinaison
intrachromosomique.
II.4.1- inversions péricentriques : le centromère est inclus dans le
segment inversé,
II.4.2- inversions paracentriques : le centromère n’est pas inclus dans
l’inversion.
Exemple : inv. (12) (q24q36).
II.5 Les duplications : elles sont provoquées par une recombinaison
inégale, est peuvent provoquer des trisomies partielles. Ces
anomalies sont rares.
Exemple : dup (11q).
II.6 Les isochromosomes : ils sont formés de deux bras identiques.
Exemple : i (Xq) isochromosome du bras long du chromosome X.
II.7-. Les chromosomes en anneau : ils sont formés par fusion des
extrémités d’un chromosome suite à la perte des segments distaux.
La formation d’un chromosome en anneau entraîne souvent une
monosomie partielle.

Document réalisé par H.Belhocine 2011/2012
47
II.8 Sites fragiles : la cassure de l’extrémité distale des bras longs du
chromosome X (site fragile de l’X) s’accompagne du syndrome de l’X
fragile.
Caryotype d’un Klinefelter Caryotype d’une femme normale

Document réalisé par H.Belhocine 2011/2012
48
Exercices
Exercice N°1 : cytogénétique
Une femme victime de plusieurs avortements spontanés fait réaliser
un caryotype pour elle et son mari. Les résultats montrent une
translocation entre les chromosomes 5 et 10. On a 5t (q23) et 10t
(q26).
a- Dites pourquoi l’anomalie dont cette femme est porteuse n’a pas été
pressentie avant les avortements ?
b- Quelle est la probabilité pour elle de mener à terme une nouvelle
grossesse ? En admettant que pour qu’un embryon soit viable il faut
que les gènes existent en deux exemplaires et deux seulement.
Solution :
a- La femme présente une translocation, mais il n ya ni perte ni gain de gènes. Le génome reste normal. b- Le père produit des gamètes normaux avec un chromosome 10 et un chromosome 5. La mère produit 4 types de gamètes : 5,10 ; 5,10t ; 5t, 10 ; 5t,10t Les zygotes possibles sont les suivants : 5,10 +5,10 = zygote 5,5,10,10 normal 5,10 + 5t, 10 + zygote 5,5t,10,10 : trisomie 10 partielle , non viable 5,10 + 5,10t = zygote 5,5,10,10t : trisomie 5 partielle , non viable 5,10 + 5t,10t = zygote 5,5t,10,10t : zygote équilibré viable La probabilité est donc de 1/2

Document réalisé par H.Belhocine 2011/2012
49
XIV- STRUCTURE DE L'ADN
I- Structure de la molécule d'ADN
La structure de l'ADN est découverte par Crick et Watson en 1953.
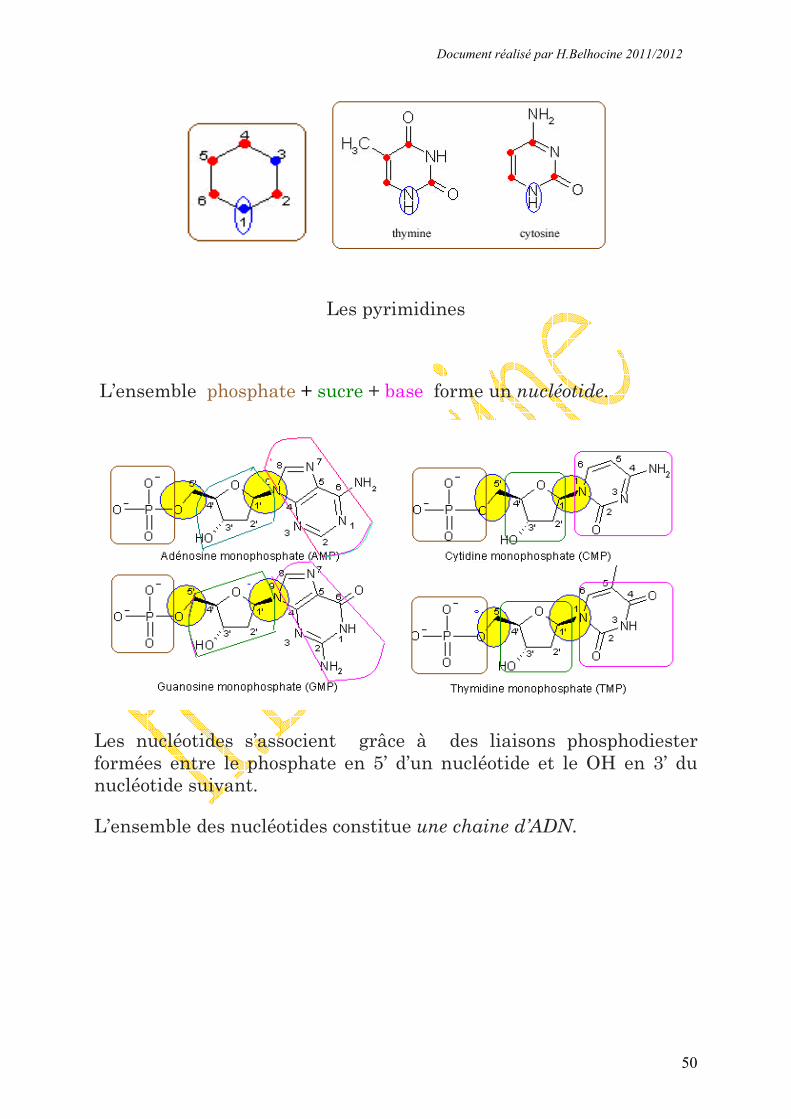
I.1- le nucléotide
Un nucléotide est composé d'une base, d'un sucre et d'un phosphate.
Le sucre présent dans l'ADN est le désoxyribose représenté ci-dessous
Les carbones sont numérotés à partir de la fonction aldéhyde de la
forme linéaire avec des numéros affectés d'un "prime". C’est au
niveau du Carbonne 1’ que sont liés les bases puriques (A et G) et
Les bases pyrimidiques (T et C) pour former une liaison N
glycosidique .Les bases puriques sont liés par l’azote (9) et les bases
pyrimidiques par l’azote (1). Voir schéma ci-dessous.
Les purines
Document réalisé par H.Belhocine 2011/2012
50
Les pyrimidines
L’ensemble phosphate + sucre + base forme un nucléotide.
Les nucléotides s’associent grâce à des liaisons phosphodiester
formées entre le phosphate en 5’ d’un nucléotide et le OH en 3’ du
nucléotide suivant.
L’ensemble des nucléotides constitue une chaine d’ADN.
Document réalisé par H.Belhocine 2011/2012
51
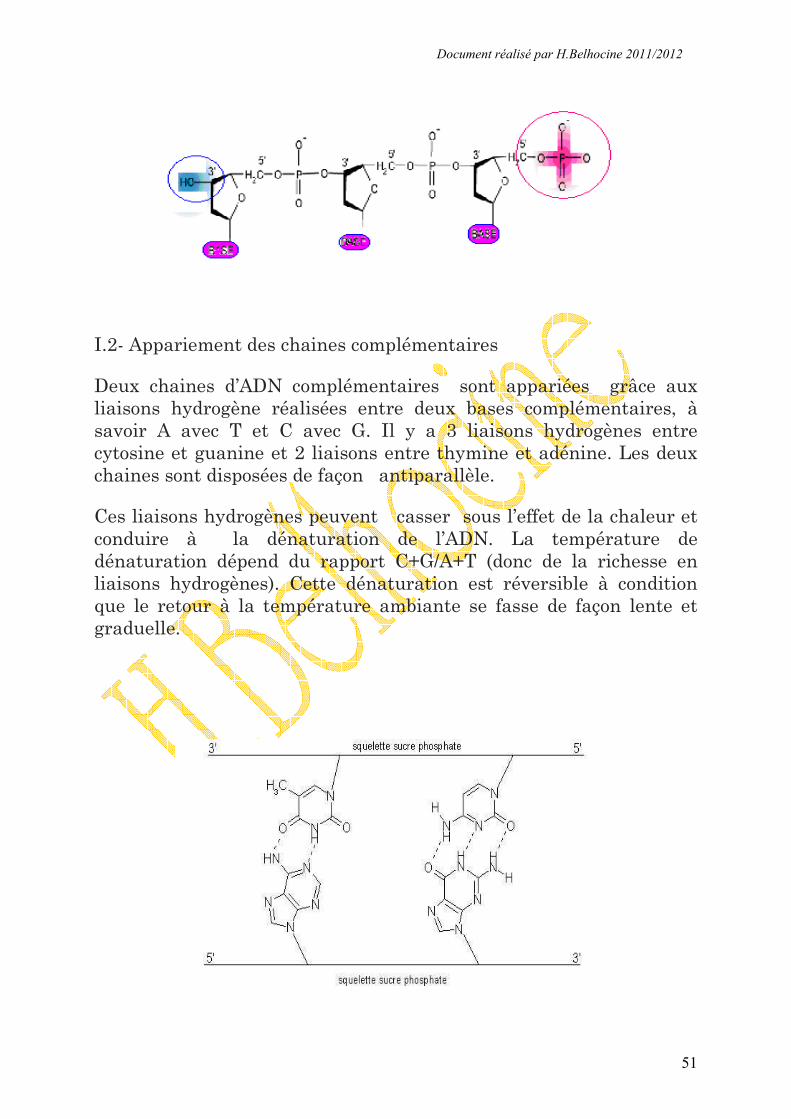
I.2- Appariement des chaines complémentaires
Deux chaines d’ADN complémentaires sont appariées grâce aux
liaisons hydrogène réalisées entre deux bases complémentaires, à
savoir A avec T et C avec G. Il y a 3 liaisons hydrogènes entre
cytosine et guanine et 2 liaisons entre thymine et adénine. Les deux
chaines sont disposées de façon antiparallèle.
Ces liaisons hydrogènes peuvent casser sous l’effet de la chaleur et
conduire à la dénaturation de l’ADN. La température de
dénaturation dépend du rapport C+G/A+T (donc de la richesse en
liaisons hydrogènes). Cette dénaturation est réversible à condition
que le retour à la température ambiante se fasse de façon lente et
graduelle.

Document réalisé par H.Belhocine 2011/2012
52
I.3- La double hélice
Les deux brins sont disposés de façons antiparallèles, c'est à dire que
le sens de lecture 5'-3' sur deux brins complémentaires est opposé. La
structure est stabilisée par les interactions entre bases : les liaisons
hydrogènes. La double hélice a un pas de 3,4 nm, chaque tour étant
constitué de 10 nucléotides (par brin) et présente un grand et un
petit sillon :
Il existe plusieurs structures différentes de la double hélice.
L'ADN B est la forme la plus commune : les paires de bases sont
perpendiculaires à l'axe de l'hélice, et globalement, ces paires ont leur
centre qui passe par l'axe. La répartition spatiale des paires de bases
est homogène.
Dans l'ADN A, l'axe de l'hélice passe par le grand sillon, qui est plus
profond que dans l'ADN B, tandis que le petit sillon est moins creusé.
Les bases sont décalées et inclinées par rapport à l'axe, laissant un
trou central dans l'hélice. Le taux d'hydratation est moins élevé que
dans l'ADN B, et la molécule est plus compacte. Le pas est de 2,8 nm
et il y a 11 résidus par tour.
Enfin, on trouve une structure assez différente d'hélice gauche dans
le cas très particulier des poly G-C (ADN Z).
I.3.1- Les interactions avec des protéines dans le grand sillon.
Les interactions qui ont lieu entre une protéine et une séquence
d'ADN spécifique (par exemple avec les enzymes de restriction ou les
protéines régulatrices agissant au niveau des promoteurs) se passent
dans le grand sillon.

Document réalisé par H.Belhocine 2011/2012
53
1.3.2- Les liaisons dans la molécule d'ADN :
Deux nucléotides adjacents sont liés entre eux par des liaisons
phosphodiesters, qui mettent en jeux les carbones des oses.
Deux bases en vis avis sont liées entre elles par des liaisons
hydrogènes; les liaisons hydrogènes sont faibles et peuvent êtres
détruites à la chaleur, il y a deux liaisons entre A et T et trois entre
G et C. La destruction des liaisons hydrogènes provoque la séparation
des deux brins complémentaires ; c'est la dénaturation de l'ADN; Le
phénomène de dénaturation est réversible, les simples brins peuvent
se réassociés par complémentarité entre les bases pour reformer un
ADN bicentenaire, dans ce cas on parle de renaturation.
I.4- compaction de la molécule d’ADN
La molécule d'ADN avec tous ses constituants forme une double
hélice. La longueur totale de la molécule d'ADN dans une cellule est
en moyenne de 2 m (pour les 46 chromosomes), cet ADN est donc
fortement compacté dans le noyau, La compaction de l'ADN est
rendue possible grâce à 4 types de protéines histones qui sont : H2A/
H2B/H3/H4. L'ADN s'enroule au tour de ces protéines pour former un
nucléosome " 2x (H2A+H2B+H3+H4) + (ADN)= 1 nucléosome. Une
série d'enroulement et de surenroulement provoquent la compaction
de l'ADN pour aboutir à la forme finale d'un chromosome avec ses
deux chromatides.
I.5- caractéristiques de la double hélice :
- Antiparallèle : les deux brins sont parallèles mais orientés dans
deux sens différents.
- Complémentarité entre les bases : entre A et T et G et C.
- Formation de deux sillons : un grand et un petit sillon.
- Dénaturation et renaturation : L'ADN est dénaturé à la chaleur
à une température comprise entre 60 et 90° C, en fonction de sa
richesse en bases GC. La renaturation est possible s'il y a retour
à la température ambiante de façon lente et graduelle.
- Absorption des UV à 260 nm.

Document réalisé par H.Belhocine 2011/2012
54
II- Structure d'un gène :
II.1- Chez les eucaryotes :
Entre 50000 et 100000 gènes composent la banque d'information
génomique de l'homme. Ces gènes sont répartis dans 23 groupes de
liaisons ou chromosomes, certains sont des gènes de structure,
renfermant une information sur la séquence en acide aminés d'une
protéine de structure qui rentre dans la composition, d'un organite ou
d'un tissu ; d'autres renferment une information de la structure d'une
protéine enzymatique qui intervient dans le métabolisme de la
cellule. Certains gènes de régulations ne sont ni transcrits, ni
traduits, ce sont des sites de reconnaissances pour certains enzymes
de régulation de la fonction des gènes.
On distingue les gènes exprimés dans toutes les cellules dits
housekipping genes, qui sont essentiels aux fonctions générales de la
cellule et les gènes exprimés spécifiquement dans certaines cellules
ou certains tissus, dits tissus specific genes.
Le gène est composé d'une succession de séquences codantes
(EXONS) et des séquences non codantes (INTRONS). Le gène
commence toujours par un exon et se termine par un exon. Le
premier et le dernier exon renferment chacun une séquence non
traduite, mais transcrite dans l'ARNm ; ce sont les séquences UTR
(utranslated region) qui portent des séquences signales. L'UTR du
premier exon renferme la séquence signale de la "cap", qui est un
7methylguanosinephosphate ajouté à l'extrémité5' de l'ARNm pour le
protéger contre les exonucléases et qui sert aussi de site de
reconnaissance pour le ribosome du début de la traduction. L'UTR du
dernier Exon renferme le signal de Polyadénylation, qui permet
l'addition de 100 à 200 résidus Adénine à l'extrémité 3' est qui sert à
stabiliser l'ARNm.
La partie codante du premier exon commence par le génon ATG sur le
brin sens (ou informatif) et la partie codante du dernier exon se
termine par l'un des 3 génons suivants : TAA, TAG, TGA.
Au brin sens on oppose le brin antisens ou brin matrice qui sert de
modèle pour la polymérisation de l'ARNm. C'est le brin sens qui est
écrit quant il faut décrire un gène.
En amont du gène de structure se trouvent deux séquences
régulatrices qui sont :

Document réalisé par H.Belhocine 2011/2012
55
a- le promoteur : Il détermine le début et l'orientation de la
transcription du gène, c'est a son niveau que se fixe l'ARN polymérase
pour démarrer la transcription. Le promoteur comporte plusieurs
séquences régulatrices qui peuvent êtres spécifiques à un tissus ou
ubiquitaires, ces séquences sont reconnues par des facteurs de
transcriptions, on peut citer pour cela la TATA box dont la séquence
consensus 5'TATAAA3' est rencontrée dans toutes les cellules.
b- Le silencer : Situé entre le promoteur et le gène de structure,
permet de ralentir la transcription.
c- L'enhancer : C'est un activateur de la transcription
(transactivateur), il agit à distance et peut se trouver en amont ou en
aval du gène de structure.
Structure d'un gène eucaryote et relation avec les gènes de
régulations
Certains gènes dérivent les uns des autres par duplication d'un gène
ancestral commun ils peuvent êtres identiques ou semblables et
interviennent dans la même fonction; ils sont appelés famille
multigénique. .on peut citer pour cela la famille multigénique simple
comme celle des ARNr 5S ou la famille multigénique de la bêta

Document réalisé par H.Belhocine 2011/2012
56
globine composée des gènes epsilon, alpha, bêta intervenant tous
dans le formation de l'hémoglobine et à différents stades du
développement. A coté des gènes qui composent la famille
multigénique on trouve des gènes qui ne s'expriment pas, ce sont les
pseudogènes, ils résultent de la duplication de gènes fonctionnels
suivis de nombreuses mutations qui les rendent inactifs. A coté des
pseudogènes on rencontre des retropseudogènes qui dérivent de la
rétrotranscription d'un ARNm mature d'un gène, pour cette raison les
retropseudogènes n'ont pas d'introns.
II.2- Chez les procaryotes
Notion d'opéron ou unité de régulation :
Il existe, dans le génome des bactéries des groupements de cistrons,
qui sont des gènes de structures contigus sous le contrôle des mêmes
régulateurs.
Le groupement polycistronique comporte 3 cistrons contigus :
1- cistron "z" de la bétagalactosidase avec 3700pb.
2- Cistron "y" de la perméase, qui permet la pénétration du lactose
dans la cellule.
3- Cistron "a" de la thiogalactoside- acetylase
Le fonctionnement de ces 3 cistrons est régulé par deux gènes : le
gène "i", qui est le gène du répresseur, le gène promoteur "p" qui
permet la fixation de l'ARN polymérase et le gène "o" qui permet la
fixation du répresseur.
Le promoteur long de 80 Pb est lui-même subdivisé en deux
segments; le site CAP qui fixe la protéine CAP/AMPc active et le site
ARN pol. qui fixe l'ARN polymérase. La fixation de la protéine cap
active favorise la fixation de l'ARN polymérase.
Le gène "i" synthétise un répresseur représenté par un tétramère à
l'état actif, mais qui est inactivé lorsqu'il est associé à l’allolactose, ce
dernier est considéré comme l'inducteur du fonctionnement de
l'opéron lactose. L'allolactose dérive de la transformation du lactose
par la bétagalactosidase.
Le gène "o" ou opérateur permet la fixation du répresseur et empêche
ainsi la fixation de l'ARN pol.

Document réalisé par H.Belhocine 2011/2012
57
I-2) Mode de fonctionnement :
Si le lactose est absent du milieu, la molécule de répresseur fixée sur
l'opérateur bloque le fonctionnement du promoteur, donc l'action de
l'ARN polymérase. Il n'y a pas, pas d'ARNm et pas de synthèse
d'enzymes.
Si le lactose est présent dans le milieu la bétagalactosidase
transforme une partie de ce lactose en allolactose, sous l'effet de
l'allolactose le répresseur se dissocie et devient inactif, il se détache
de l'opérateur et permet à l'ARN polymérase de procédé à la
transcription de l'ARNm. Dès que le lactose a été entièrement
consommé, le répresseur reprend sa place et bloque l'opéron.
Si le glucose et le lactose sont présents en même temps dans le milieu
la bactérie utilise le glucose en priorité, qui est le sucre le plus simple;
la raison en est que, en présence de glucose, le taux d'AMPc diminue
dans le milieu, la protéine cap devient inactive. L'inactivation de la
cap diminue l'affinité du site promoteur à l'ARN pol ce qui arrête la
transcription de l'opéron lactose. En absence de glucose le phénomène
inverse se produit et l'opéron lactose devient actif en présence de
lactose.
L'opéron lactose et dit inductible car son fonctionnement est induit
par la présence de l'inducteur qui est l'allolactose.
La répression de l'opéron lactose par la présence du glucose est
appelée répression catabolique.

Document réalisé par H.Belhocine 2011/2012
58
STRUCTURE DE L'OPERON LACTOSE.
Légende :
R : Gène répresseur
P : Gène promoteur
O : Gène opérateur
Z : Gène de la bétagalactosidase
Y : Gène de la perméase
A : Gène de la trans-acétylasse