Embed Size (px)
Citation preview

BlOC~t/M/E, 1985, 67, 309-315 Revue
Immunological detection of lesions in DNA.
Marc LENG.
Centre de Biophysique Moldculaire, C.N.R.S. 1A, avenue de la Recherche Scientifique, 45045 Orldans Cedex, France.
(Refue le 9-1-1985, accept~e le 6-3-1985).
R6sum6 - - Le but de cet article est de montrer l'utilitd des anticorps anti-acides nucldiques, des anticorps anti-nucldosides et anti-DNA modifids par un agent physique ou chimique pour l'dtude de ddfauts dans le DNA et leur rdparation. Des rdsultats obtenus avec des anticorps anti-nu- cldosides, des anticorps anti-nucldosides et anti-DNA modifids par des cancdrogdnes chimiques sont des exemples du potentiel des mdthodes immunologiques dans trois domaines a) la sensibilitd de la ddtection et la quantification des adduits b) la visualisation des adduits dans les tissus, dans les cellules isoldes et le long de la double hdlice du DNA c) l'dtude des changements conformation- nels induits dans le DNA par les adduits.
Mots-cl~s : anticorps anti-nuel~osides / antieorps anti-DNA / eane~rog~nes ehimiques.
S u m m a r y - - The purpose of this paper is to show that the antibodies to nucleic acids, to nucleosides or to DNA damaged by a physical or a chemical agent, are usefid tools in the study of DNA damage and repair. The results obtained with antibodies to nucleosides, antibodies to nucleosides and DNA modified by chemical carcinogens emphasize the potential of hmnunological methods in three main areas, a) the sensitive detection and quantitation of adducts; b) the visualization of adducts in tissues, individual cells, and along the DNA double helix; c) the study of conformational changes of DNA induced by adducts.
Key-words : antibodies to nucleosides / antibodies to DNA / chemical carcinogens.
Numerous physical and chemical agents can damage DNA and these damaging agents induce mutations. The induction of mammary carcino- mas in rats by a single dose of nitroso-methylurea involves malignant activation of H-ras-I locus by single point mutations [1]. Generally, all chemical carcinogens bind covalently to DNA and it is assumed that this covalent binding is an impor- tant event in the initiation of multistage process of malignant transformation. The amount of bound carcinogen is quite low, of the order of one carcinogen molecule per 104-106 nucleotide resi- dues. One ultraviolet-induced pyrimidine cyclo- butane dimer in DNA can be a lethal lesion in E. coli. Thus a major problem encountered in the study of DNA damage and repair is to determine
the nature and amount of DNA lesions and to localize them along the DNA molecule.
The immunological methods appear efficient to bring an answer to this problem. The antigenic functions of nucleic acids were recognized much later than those of proteins or polysaccharides. It is now well-established that the synthesis of antibodies specifically recognizing bases, nucleo- sides, nucleotides, single stranded and multi- stranded polynucleotides, can be induced in animals immunized with the right antigen (gene- ral reviews), [2-6]. These antibodies are largely used in many laboratories, for example to titrate nucleosides or to detect Z-DNA. It is impoqant to note that the large expansion of immunological methods in several fields (biochemistry, pharma-

310 AL Leng
cology and cancer) is in part due to the develop- ment of the technique of radioimmunoassay [7].
The purpose of this paper is to give some hints on the possible use of specific antibodies to detect lesions in DNA. It is beyond the scope of this short review to summarize all the work done on the antibodies to nucleic acids (see general re- views [2-6, 59]).
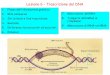
other biological functions. A schematic represen- tation of the basic immunoglobulin structure is shown in Figure I.
1. Antigen-antibody complex
Immunoglobulins
The ability of vertebrate species to cope with foreign substances depends in part on their capacity to synthesize antibodies which recognize specifically and bind to the foreign substances called antigens. An antibody is a serum protein belonging to the family called immunoglobulins. The immunoglobulins are biologically active glycoproteins capable of carrying out several different functions. They can bind specifically to antigens, activate the complement system, me- diate cytotropic reactions and act as antigen receptors on lymphocyte membranes.
There are five classes of immunoglobulins (IgG, IgM, IgA, IgD and IgE) which are built of heavy and light chains. All the immunoglobulins can be regarded as being derived from a basic structure of two light polypeptide chains of molecular weight 22000-23000 (about 220 amino acid residues) and two heavy polypeptides chains of molecular weight 50000-70000, linked together by the disulphide bridges of cystine residues. The sequence analyses of numerous immunoglobulins have demonstrated the existence of distinct re- gions of sequence homology, called domains. Each light chain has two domains, one variable domain (V,) and one constant (C,). Each heavy chain has one variable domain (VH) and several constant domains. The constant domains are common to immunoglobulins of the same group. The variable domains vary from antibody to antibody. They have sections of variable amino- acid sequence that are involved in the antigen binding site (an IgG has two antigen binding sites) and determine the antigen binding specifi- city.
Under suitable conditions, limited proteolysis with papain results in three fragments of about equal molecular weight, two Fab fragments and one Fc fragment. The antigen binding site is localized on the Fab fragment while the Fc fragment is involved in complement fixation and
FIG. 1 . - Schematic representation of the basic immunoglo- bulin stntcture.
Antigen
An antigen is any substance which when injec- ted into an animal induces the formation of antibodies capable of reacting specifically with the antigen. The substance has to be of large molecular weight. Low molecular substances (ap- proximately less than 1500) do not induce the formation of antibodies. However, antibodies can be raised to small molecules by immunization with conjugates made up of low molecular weight sul~stances called haptens, covalently linked to a macromolecule (generally a protein).
Antigen-antibody complexes
The reaction of formation of a complex bet- ween an antibody and the corresponding antigen is reversible. Both products are not chemically modified. The specificity of the interaction is a function of the stability of the complex formed between the antigen and the antibody. The inter- molecular forces involved in the antibody-antigen complex are similar to those engaged in any type of molecular complex (electrostatic, hydrogen bonding, stacking and hydrophobic interactions). The thermodynamic parameters for numerous antibody-hapten complexes have been determi-

Immunological detection
ned. The values of the association constants vary in a large range ( - 104 - 10 u M-I). The thermo- dynamic parameters for the complexes between an antibody and an antigen bearing several antigenic determinants are more difficult to esti- mate. Depending upon their relative concentra- tions, the immunoglobulins (IgG have two antigen binding sites, IgM have ten...) can bind to the same macromolecule, or bind to several macromolecules leading to a large molecular weight soluble complex, or cross-link several macromolecules leading to the formation of a precipitate (general reviews [8-15]).
2. Immunochemical procedures
The three major steps involved in the produc- tion of antibodies to nucleic acids (or to nucleic acids modified by a damaging agent) are : a) the synthesis of the antigen; b) the immunization of animals; c) the characterization of the antibodies.
a) A n t i g e n
Low molecular weight compounds (haptens) are not antigenic and thus have to be covalently linked to a carrier [16]. Bovine serum albumin has often been used but some other proteins might be better carriers. The choice of the covalent linkage depends upon the nature of the hapten and several chemical reactions have been described [17]. When possible, the periodate oxidation reaction, as proposed by Erlanger and Beiser, is
o f lesions in DNA 311
one of the most convenient [18] and is summari- zed in Figure 2.
b) I m m u n i z a t i o n
There are many immunization protocols [3, 5, 6, 19]. In rabbits, large molecular weight nucleic acids are often mixed with positively charged methylated bovine serum albumin and form inso- luble complexes. These complexes or the hap- ten-protein conjugates are prepared for injection in the form of a stable emulsion by mixing with complete Freund's adjuvant. Several inoculations are done at various time intervals. Rabbits posses- sing antibodies are boosted with antigen emulsi- fied in incomplete Freund's adjuvant. The amount of antigen for each inoculation is of the order of 50-200 lag.
c) Speci f ic i ty
The specificity of the antibodies can be charac- terized by several physico-chemical methods. Radioimmunoassays (RIA) are now largely used because of their sensitivity and reliability. After determination of conditions giving 50 % binding of the radioactive hapten or polymer (tracer), competition experiments are performed with unlabeled inhibitors [7, 20]. This enables to prove the specificity of the antibodies, to titrate the amount of ufilabeled antigen in the samples to be analyzed, and to get some knowledge on groups or atoms of the antigen involved in the recogni- tion by the antibodies. Another technique, also largely used, is the enzyme-linked immunosorbent
R O H 2 C ~ b o s e
OH OH nucleoside nucleotide oligonucleotide
IoZ ROH2C r~ base ROH2C ~ base
"" H2N - Protein "'
ICI Cii OH /C C x O O XN/ OH
I Protein
S ROH2C O bose
H,,'C\NzC'-.H I
Protein
FXG. 2. -- Conjugation of a ribonucleoside, ribonucleotide or oligoribonucleotides to protein, according to Erlanger and Beiser[l$].

312 M. Leng
assay (ELISA) which does not need a radioactive tracer [21].
3. Detection of lesions in DNA
For sake of clarity, we will limit ourselves to a few examples and we will consider antibodies to nucleosides, and to nucleosides and DNA modified by chemical carcinogens.
a) Antibodies to nucleosides
The base plays a major role in the recognition of the nucleoside by the antibodies. For example, antibodies to adenosine have a higher affinity for adenosine than for adenine. They do not bind to guanosine, thymidine or cytidine, i.e. guanosine is at least 10000-fold less reactive than adenosine. Depending upon the nature of the covalent link between adenosine and the protein carrier, deoxyadenosine can be slightly better recognized than adenosine (general reviews [5, 6, 22]). At least in rabbits, it does seem possible to induce the synthesis of antibodies which bind to adenosine and not to deoxyadenosine. These antibodies bind to oligo(A), poly(A) and denatured DNA. They do not bind to native DNA. In native DNA, the atoms and groups interacting with the amino acid residues of the antibody binding sites are no longer accessible. Thus, these antibodies can be used to titrate the amount of deoxyadenosine (or adenine) after DNA hydrolysis. They can also detect adenine residues in partially denatured DNA. They combine with fixed metaphase chro- mosomes but only if the chromosomes are par- tially denatured [23, 24]. However, it is difficult to know precisely the minimal size of a denatured region which makes an adenine residue accessible to the antibodies [24]. From a study of the complexes between anti-adenosine Fab fragments and oligo(A), it has been found that three nu- cleotide residues can fit in the binding site of the Fab fragment but one nucleotide residue interacts with the aminoacid residues of the binding site [28]. An estimate of the maximal dimensions of the size of the binding site can be deduced.
Antibodies to nucleosides were used to discri- minate between local defects induced in DNA by N-aminofluorene derivatives [26]. The antibodies to cytidine interact strongly with DNA modified by acetylamino residues (AAF) and weakly with DNA modified by aminofluorene (AF). Antibo- dies to adenosine do not bind to these DNAs. This suggests that a local denaturation is induced
by AAF residues and not by AF residues, which is in agreement with physico-chemical studies of these DNAs (general reviews [27,28]). On the other hand, the antibodies to cytidine do not bind to poly(dG-dC)-AFF in the Z conformation. It can be concluded that the localization of the carcinogen in B- and Z-DNA is not the same [29], again in agreement with physicochemical studies [29-32].
A quantitative analysis of these experiments is difficult because the antibodies to nucleosides can displace the equilibrium: double-stranded helix ~ single-stranded helix. Under normal salt conditions and at room temperature, there are some open regions in DNA [33] which can inte- ract with the antibodies. In other words, these antibodies have to be considered as melting proteins. Depending upon the concentration of DNA and antibodies, the value of the association constants, the equilibrium can be more or less shifted towards the right. It is expected that for example, high affinity antibodies to adenosine added in excess to a solution of DNA will completely open the double helix. Generally, any equilibrium (double-stranded helix ~ single- stranded helix, three-stranded helix --~ double- stranded helix, B-DNA ~ Z-DNA,...) can be shifted towards the left or the right in the pre- sence of antibodies having more affinity for one of the two conformations [34].
b) Antibodies to nucleosides modified by che- mical carcinogens
In the in vivo reaction between chemical carci- nogens and DNA, several adducts are formed. Some of them have been identified and synthe- sized chemically. These adducts covalently linked to a protein carrier are antigenic. Highly specific antibodies are now available (general reviews [35-36]). The best antibodies have affinity cons- tants for their respective haptens larger than 109M -I. Moreover, they show low degrees of cross-reactivity with the unmodified nucleosides. For example by RIA, antibodies to OLEtGuo detect O6-EtdGuo/dGuo molar ratio of 3.10 -7 in a sample of 100 lag of hydrolyzed DNA [37]. Antibodies to guanosine-acetylaminofluo- rene (Guo-AAF) detect by RIA 250 fmol of dGuo-AAF, by ELISA 40 fmol and by USERIA (ultra sensitive enzymatic radioimmuno assay) 4 fmol [38]. Thus, these antibodies are a powerful tool to titrate the adducts in in vivo modified DNA. However, it should be ascertained that there is no RNA. Most of the carcinogens bind

Immunological detection of lesions in DNA 313
to RNA and carcinogen-RNA is recognized by the antibodies. Moreover, since the affinity constants of these antibodies is larger for the free hapten than for modified DNA [39,40], the sensitivity of the titration is increased when performed with completely hydrolyzed DNA.
The antibodies to Guo-AAF bind more stron- gly to AAF-GpA than to ApG-AAF (K(Guo-AAF) > K(AAF-GpG) > K(ApG- AAF)). This shows that the accessibility of the hapten is hindered in the dinucleoside-monopho- sphate and that the interactions between the carcinogen residue and the bases are different in ApG and GpA, in agreement with circular di- chroism experiments [40]. In other words, from the determination of the relative affinities of the antibodies (or the Fab fragments) to nucleosides, oligonucleotides, and DNA modified by a chemi- cal carcinogen, one can obtain some knowledge on the localization of the carcinogen in the double helix.
The development of immunostaining techni- ques enables localization of bound carcinogens in tissues and individual cells. This seems to be appropriate to follow the binding of the carcino- gen and its repair in particular cell strata of whole tissues [41, 42]. At the DNA molecular level, the adducts can be visualized by electron microscopy observation of DNA-antibodies complexes. Using this technique, a random distribution of dGuo-AAF residues in in vitro modified ColEI DNA and a non-random distribution of O6-Etd - Guo in rat brain chromosomal DNA after in vivo administration of the N-nitroso carcinogen [43, 44] have been demonstrated.
c) Antibodies to DNA modified by chemical carcinogens
Despite numerous assays, it does not seem possible to induce the synthesis of antibodies specific for native B-DNA in rabbits or mice and it can be assumed that linear double-stranded B-DNA is not antigenic (denatured DNA is antigenic). On the other hand, double-stranded DNAs, chemically modified by several chemical carcinogens, are efficient antigens [45-51]. The antibodies are specific for damaged regions and do not cross-react with the unmodified native or denatured DNA. Immunization with the modified DNA as antigen has the advantage over immuni- zation with modified nucleoside-protein conju- gate that the antibodies recognize adducts in non-hydrolyzed DNA which makes the immuno- assays easier. Moreover, the chemical synthesis of
relatively large amounts of modified nucleoside can be very difficult and thus one has to use the modified DNA as antigen. However, since in the in vitro reaction between a carcinogen and DNA several types of adducts can be formed, the specificity of the antibodies has to be carefully characterized.
An important point is that DNA can adopt different conformations. B-DNA is not antigenic but A-DNA or Z-DNA are antigenic. Antibodies recognizing specifically A-DNA or Z-DNA have been produced in animals immunized with hy- brids in A conformation [5,52] or double-stran- ded polydeoxyribonucleotides in Z conformation [53-56], respectively. The conformational changes of DNA can be induced by numerous factors such as salt, proteins, folding of the double helix, damaging agents... The binding of some carcino- gens favors or prevents the B-Z transition [29-32, 57, 58]. All these factors can play a role in the conformation of the antigen after injection in animals and thus modify the antigenic characte- ristics. Nevertheless, the antibodies to modified nucleic acids like the antibodies to modified nucleosides can reveal the lesions in cells and along a DNA molecule.
Conclusion
Many physical and chemical agents can da- mage DNA. The nature and the extent of adducts (modified DNA constituents) even at low levels, the kinetic studies of their elimination (or per- sistence) from DNA can be studied with anti- bodies specificaJly recognizing these adducts. Another attractive possibility of the antibodies is the localization of the adducts in tissues or individual cells. At the molecular level, the mapping of adduct sites along a DNA molecule can be done by electron microscopy observation. Moreover, the immunological methods can reveal the accessibility of modified DNA constituents in DNA and conformational changes of the double helix. They are complementary to some physico* chemical methods.
These remarks underline the potential of im- munological methods in the understanding of DNA damage and repair. It seems certain that they will be widely used in numerous research fields. However, there are some difficulties. In addition to the synthesis of the antigens, many pitfalls can be encountered in the choice of the animals, in the choice of the hapten carrier, in the

314 M. Leng
choice of the immunization procedures, and in the characterization of the antibodies. The antibodies have to be highly specific. The sensi- tivity of the immunological methods depends upon the affinity of the antibody for the antigen. The affinity constants have to be high but some antibodies have a low affinity for the antigen. Also, it must be recalled that an unexpected cross-reaction can occur.
It is on purpose that we did not discuss whether it is better to use monoclonal or polyclo- nal antibodies. The choice between monoclonal and polyclonal depends upon a variety factors. In fact, for many purposes, they are complementary and enhance the potential of immunological methods.
Acknowledgments
This work was supported in part by La Fondation pour la Recherche M~dicale, La Ligue Nationale Franfaise contre le Cancer and l'Association pour la Recherche sur le Cancer.
REFERENCES
1. Sukumar, S., Notario, V., Martin-Zanca, D. & Barbacid, M. (1983) Nature, 396, 658-661.
2. Plescia, O.J. & Braun, W. (1967) Adranc. ImmunoL, 6, 231-252.
3. Lacour, F., Nahon-Merlin, E. & Michelson, A.M. (1973) Curr. Top. Microbiol. and ImmunoL, 62, 1-39.
4. Stollar, B.D. (1973) in : "The Antigens" (M. Sela ed.) Vol. 1, p. 1-85, Academic Press, New York.
5. Stollar, B.D. (1975) Crit. Rev. Biochem., ,45-69. 6. Munns, T. & Liszewski, K.T. (1980) Prog. Nucl. AcM
Res. Mol. BioL, 24, 109-165. 7. Yalow, R.G. (1978) Science, 200, 1236-1245. 8. Kabat, E.A. & Mayer, M.M. (1961) Experimental
lmmunochemistry, C.C. Thomas Publisher, Springfield.
9. Fougereau, M. (1975) Elements d'lmmunologie Fondamentale, Masson et Cie, Paris.
10. Day, E.D. (1972) Advanced Immunochemistry, The Williams and Wilkins Company, Baltimore.
1 I. Nisonoff, A., Hopper, J.E. & Spring, S.S. (1975) The Antibody Molecule (F.J. Dixon & Kunkel, H.G., ed.) Academic Press, New York.
12. Davies, D.R., Padlan, E.A. & Segal, D.M. (1975) Ann. Rev. Biochem., 44, 639-667.
13. Poijak, R.J. (1973) Contemp. Top. Mol ImmunoL, 2, 1-26.
14. Beale, D. & Feinstein, A. (1976) Quat. Rev. Bio- phys., 9, 135-180.
15. Yelton, D.E. & Scharff, M.D. (1981) Ann. Rev. Biochem., 50, 657-680.
16. Landsteiner, K. (1945) The specificity o f serological reactions, Harvard Univ. Press, Cambridge.
17. Erlanger, B.F. (1980) In :'Methods in Enzymology" (Van Vunakis and J.J. Langone eds) vol. 70, part A, Academic Press, New York, pp. 85-104.
18. Erlanger, B.F. & Beiser, S.M. (1964) Proc. NatL Acad. ScL USA, 52, 68-74.
19. Hum, B.A.L. & Chantler, S.M. (1980) in : "Methods in Enzymology'" (H. Van Vunakis and J.J. Lan- gone eds) vol. 70, part A, Academic Press, New York, pp. 104-142.
20. Van Vunakis, H. (1980) in : "'Methods in Enzymo- logy" (H. Van Vunakis and J.J. Langone eds) vol. 70, part A, Academic Press, New York, pp. 201-209.
21. Enguall, E. (1980) in : "'Methods in Enzymology"(H. Van Vunakis and J.J. Langone eds) vol. 70, part A, Academic Press, New York, pp. 419-439.
22. Stollar, B.D. (1980) in : "Methods in Enzymology'" (H. Van Vunakis and J.J. Langone eds) vol. 70, part A, Academic Press, New York, pp. 70-85.
23. Schreck, R.R., Warburton, D., Miller, O.J., Beiser, S.M. & Erlanger, B.F. (1973) Proc. Natl. Acad. Sci. USA, 70, 804-807.
24. Schreck, R.R., Erlanger, B.F. & Miller, O.J. (1977) Exp. Cell. Res., 108, 403-411.
25. Lavayre, J. & Leng, M. (1977) Biochimie, 59, 33-42. 26. Sage, E., Spodheim-Maurizot, M., Rio, P., Leng, M.
& Fuchs, R.P.P. (1979) FEBS Letters, 108, 66-68. 27. Daune, M., Fuchs, R.P.P. & Leng, M. (1981)J. Natl.
Cancer Inst., 58, 201-210. 28. Grunberger, D. & Weinstein, I.B. (1979) in:
"C.R.C. Press, Chemical Carcinogens and DNA" (P. Grover, ed.) Voi. 2, Baca Raton, Florida, pp. 59-93.
29. Santella, R., Grunberger, D., Broyde, S. & Hin- gerty, B. (1981) Nucleic Acids Res., 9, 5459-5467.
30. Santeila, R., Grunberger, D., Weistein, B.I. & Rich, A. (1981) Proc. Natl. Acad. Sci., USA. 78, 1451-1455.
31. Sage, E. & Leng, M. (1980) Proc. Natl. Acad. Sci., USA, 77, 4597-4601.
32. Sage, E. & Leng, M. (1981) Nucleic Acids Res., 9, 1241-1250.
33. Englander, W.S. & Kallenbach, N.R., (1983) Quart. Rev. Biophys., 16, 521-655.
34. Guigues, M. & Leng, M. (1976) Eur. J. Biochem. 69, 615-624.
35. Polder, M.C. (1981) J. Natl. Cancer Inst., 67, 515-519.
36. Montesano, R. (1982) Cancer Res., 42, 5236-5239. 37. Rajewsky, M.F., MQller, R., Adamkiewicz, J. &
Drosdziok, W. (1980) in : Carcinogenesis, Funda- mental Mechanisms and Environmental Effects (B. Pullman, P.O.P. Ts'O and H. Gelboin eds) D. Reidel Publishing Company, Dordrecht, pp. 207-218.
38. Hsu, I.C., Poirier, M.C., Yuspa, S.H., Yolken, R.H. & Harris, C.C. (1980) Carcinogenesis, 1, 455-458.

hnmunological detection o f lesions in DNA 315
39. Poirier, M.C., Yuspa, S.H., Weinstein, I.B. & Blobstein, S. (1977) Nature. 270, 186-188.
40. Guigues, M. & Leng, M. (1979) Nucleic Acids Res. 6, 733-744.
41. Sage, E., Gabelman, N., Mendez, F. & Bases, R. (198I) Cancer Letters, 14, 193-204.
42. Adamkiewicz, J., Drosdziok, W., Eberhardt, W., Langenberg, U. & Rajewsky, M.F. (1982) in : Indicators o f Genotoxic Exposure, Banbury Report 13, (Bridges, B.A., Butterworth, I.B. and Weins- tein, I.B. eds) Cold Spring Harbor Laboratory Press, New York, pp. 265-276.
43. De Murcia, G., Lang, M.C., Freund, A.M., Fuchs, R.P.P., Daune, M., Sage, E. & Leng, M. (1979) Proc. Natl. Acad. Sci., USA, 76, 6076-6080.
44. Nehls, P., Rajewsky, M.F., Spiess, E. & Werner, D. (1984) The Embo J., 3, 327-332.
45. Sage, E., Fuchs, R.P.P. & Leng, M. (1979) Bioche- mistry, 18, 1328-1332.
46. Spodheim-Maurizot, M., Saint-Ruf, G., & Leng, M. (1980) Carcinogeneis, 807-812.
47. Poirier, R., M.C., Santella, R., Weinstein, I.B., Grunberger, D. & Yuspa, S.T. (1980) Cancer Res., 40, 412-416.
48. Slor, H., Mizusawa, H., Neihart, N., Kakefuda, T., Day, R.S. & Bustin, M. (1981) Cancer Res., , 3111-3117.
49. Hertzog, P.J., Shaw, A., Lindsay Smith, J.R. & Gamier, R.C. (1983) J. lmmunol. Methods, 62, 49-58.
50. Haugen, A., Groopman, J.D., Hsu, I.C., Goodrich, G.R., Wogan, G.N. & Harris, C.C. (1981) Proc. Natl. Acad. Sci., USA, 7, 4124-4127.
51. Strickland, D.T. & Boyle, J.M. (1981) J. lmmunol. Methods, 41, 115-122.
52. Kitagawa, Y. & Stollar, B.D. (1982) Mol. lmmunol., 19, 413-420.
53. Lafer, E.M., M611er, A., Nordheim, A., Stollar, B.D. & Rich, A. (1981) Proc. Natl. Acad. Sci., USA, 78, 3546-3550.
54. Malfoy, B. & Leng, M. (1981) FEBS Letters, 132, 45-48.
55. Hanau, L.H., SanteIla, R.M., Grunberger, D. & Erlanger, B.F. (1984) J. Biol. Chem., 259, 173-178.
56. Zarling, D.A., Arndt-Jovin, D., Robert-Nicoud, M., McIntosh, L.P., Thomae, R. & Jovin, T.M. (1984) J. Mol. Biol.. 176, 369-415.
57. Nordheim, A., Hao, W.H., Wogan, G.N. & Rich, A. (1983) Science, , 1434-1436.
58. Wells, R.D., Miglietta, J.J., Klysik, J., Larson, J.E., Stirdivant, S.M. and Zacharias, W. (1982) J. Biol. Chem., 257, 10166-10171.
59. Strickland, P.T. and Boyle, J.M. (1984) Prog. Nucl. Acid Res. Mol. Biol.. 31, 1-58.