Embed Size (px)
Citation preview

BIOCHIMIE, 1975, 57, 77-83.
Influence des
de l'6tat thyroidien sur le profil 61ectrophordtiquc protdines mitochondriales de tbie de rat
Miche l BAUDRY'), J e a n - P i e r r e CLOT et R a y m o n d MICHEL.
Endocrinologie, UER des Sciences Pharmaceutiques et Biologiques de Paris Luxembourg,
Universit~ Ren~ Descartes, 4, avenue de l'Observatoire, 75270 Paris Cedex 06. Biochimie Cellulaire, Coll~ge de France,
11 place Marcelin-Berthelot, 75231 Paris Cedex 05. (4-7-1974).
Summary. - - Liver mitoehondria were isolated f rom normal and thyroidectomized rats and their protein components analyzed' by polyaerylamide gel eleetrophoresis.
In whole mitoehondria 35 protein fractions wi th M,W ranging f rom 10,000 to 135,000 were characterized. In the absence of thyroid hormone secretion, the amount of a MW 54,000 fraction was always decreased. Injection of small doses of 3,5,3'-triiodo-L-thyronine to the thyroidectomized animal restored the quant i ty of that protein fraction to normal.
Isolated outer mitoehondrial membranes showed the presence of 20 protein fractions. These fractions revealed no change af ter thyroideetomy. The mitoplast, which contained 35 fractions, exhibited a decrease of the MW 54,000 component in thyroidectomized rats.
The mitoplast was separated into several fractions. Water soluble matr ix proteins pre- sent.ed molecular weights ranging between 40,{)00 and 55,000. Proteins, which were slightly bound to the inner mi'tochondrial membrane and could be extracted by KCI, presented molecular weights between 25,000 and 45,#00. Structural proteins showed a principal specific component of MW = 23,000. Electrophoretic patterns obtained with these submito- chondrial fractions were s imilar in normal and thyroidectomized animals. The mitoplast fraction which contained the insoluble cytochromes (a, a3, b, c~) was isolated ; its prin- cipal constituent, of MW 5~,000, was significantly decreased after thyroidectomy.
Thus, the lack of thyroid hormone secretion lowered the level of a protein constituent bound to the inner membrane of l iver mitochondria. The synthesis of this consti tuent could, be controlled by mitochondrial nucleic acids.
L ' o r g a n i s a t i o n g6n6ra le de la m i t o c h o n d r i e est ca rac t6 r i s6e p a r la p r 6 s e n c e de p lu s i eu r s c o m p a r - t iments , c o m p r e n a n t un sys t6me h doub le m e m - b r a n e et une ma t r i ce . Les m e m b r a n e s i n t e r n e et e x t e r n e d i f f6 ren t p a r l eu r s t r u c t u r e [1] et l eurs f o n c t i o n s [2]. La p a r o i i n t e r n e r e n f e r m e un 6qui- p e m e n t m a c r o m o l 6 c u l a i r e i m p l i q u 6 dans les r6ac- t ions de t r a n s f e r t des 61ectrons et de syn thbse de I 'ATP, l esque l les n6cess i t en t des a r r a n g e m e n t s st6- r 6osp6c i f i ques en t r e l es d ive r s composa:n ts de la c h a l n e r e s p i r a t o i r e . Hatef i et al. on t m o n t r 6 que la c h a i n e de t r a n s f e r t des 61ectrons isol6e de mi to - c h o n d r i e s de occur de b ceuf p e u t 6tre f r a c t i o n n 6 e en q u a t r e c o m p l e x e s p ro t6 iques f o n c t i o n n e l s [3].
Les t e c h n i q u e s d '61ec t rophor6se sur gel de po ly - a e r y l a m i d e on t p e r m i s u n e 6 tude p lus d6tail16e des p r o t 6 i n e s m i t o c h o n d r i a l e s . Ces m 6 t h o d e s on t 6t6 app l i qu6es aux o rgan i t e s en t i e r s et aux m e m - b r a n e s isol~es de fo ie de ra t [4, 5], a ins i q u ' a u x
(*) Ce travai l a ~t6 r6alis6 en partie grace ~ une sub- vention du C.N.R.S. et a v e c l a collaboration technique de Mademoiselle B. Souillat.
A qui route correspondance doit ~tre adress6e.
c o m p l e x e s r e s p i r a t o i r e s p r6pa r6s h p a r t i r de cceur de boeuf [6]. Ces t r a v a u x on t m o n t r 6 l 'h6t6- rog6n6i t6 des p r o t 6 i n e s m i t o c h o n d r i a l e s . Les qua- t re c o m p l e x e s r e s p i r a t o i r e s son t eux -mSmes cons- t i tu~s d ' u n g r a n d n o m b r e de pe t i t e s sous-uni t6s c o n t e n a n t des p r o t 6 i n e s aux g r o u p e m e n t s p ros th6- t iques connus , ma i s 6ga lemen t d ' au t r e s qu i n ' o n t pas e n c o r e 6t6 iden t i f i6es . La c a r a c t 6 r i s a t i o n d i r e c t e des p r o t 6 i n e s aprbs s6pa ra t i on 61ect ropho- r6 t ique 6 tan t trbs dif f ic i le , i l nous a p a r u int6- r e s s an t de su iv re les v a r i a t i o n s q u a n t i t a t i v e s de ces c o m p o s a n t s en r e l a t i o n avec les m o d i f i c a t i o n s fonc t i onne l l e s . Les h o r m o n e s t h y r o i d i e n n e s in- f l u e n c a n t p r o f o n d 6 m e n t les p rop r i6 t6 s des mi to - c h o n d r i e s , nous avons e n t r e p r i s une 6rude s t ruc- tu ra l e des p a r t i c u l e s i sol6es d ' a n i m a u x n o r m a u x et h y p o t h y r o i d i e n s r e c e v a n t ou non de la 3,5,3'.- t r i o d o - L - t h y r o n i n e . Nous avons m o n t r 6 p r 6 c 6 d e m - m e n t que ces h o r m o n e s ag issen t au n i v e a u de ]a m e m b r a n e i n t e r n e m i t o c h o n d r i a l e en s t i m u l a n t d ' u n e p a r t la v i tesse de r e n o u v e l l e m e n t de ses p r o - t6 ines [7~ et d ' a u t r e p a r t l ' a c t iv i t6 de c e r t a i n s sys- t6mes m u l t i e n z y m a t i q u e s i m p l i q u 6 s dans les m6- c a n i s m e s des o x y d o p h o s p h o r y l a t i o n s [8].

78
Nous avons analys6 par ~leetrophor~se sur gel de polyacrylamide ]es const i tuants prot6iques des compar t iments de la i i tochondr ie et de certaines de leurs fract ions et, compte tenu de la sp6cificit6 des hormones thyro id iennes sur les propri6t6s et la prot6inog6n6se de cette part icule, nous avons ent repr is ces recherches dans divers Oats thy- roidiens.
MATERIEL ET TECHNIQUES.
ANIMAUX.
Les exp6riences sont r~alis6es avec des rats Wistar m~les normaux et thyroidectomis6s h 60 g par une m6thode chirurgicale. L 'hypothyro id ie est v6rifi6e par la mesure du m6tabolisme de base et en suivant l '6volution pond6rale des an imaux [9]. Des an imaux thyr6oprives sont trait6s par la 3,5,3 '- tr i iodo-L-thyronine (T 3) en admin is t ra t ion sous-cutan6e.
~SOLEMENT DES MITOCHONDRIES.
Les mi tochondr ies sont s6par6es par centr ifuga- t ions diff6rentieHes dans une solution de saccha- rose 0,25 M alnen6e h pH 7,4 h l 'a ide de Tris 2 mM [10]. La puret6 des pr6para t ions est contr616e en d6 te rminant l 'aetivit6 des marqueurs enzymati- ques sp6cifiques [7].
ISOLEMENT DES MEMBRANE:S¢ MITOCHONDRIALES.
Les membranes in i tochondr ia les sont isol6es en ut i l i sant la m6thode de Parsons et al. [11]. Apr6s t ra i tement des par t icules avec une solut ion hypo- tonique, la membrane externe et l ' ensemble mem- brane i n t e r n e - m a t r i c e (mitoplaste) sont s6parSs par cenlr i fugat ions diff6rentielles et purif i6s sur gradient de saccharose. La puret6 des pr6para- t ions est contr616e par la mesure de l 'activit6 des enzymes sp6cifiques, la monoamine oxydase [12] et la cytochrome oxydase [13], au moyen d 'une technique polarographique. Les prot6ines par t icu- laires sont dos6es par les m6thodes de Gornall et al. [14] et de Lowry et al. [15].
FRACTIONNEMENT DU MITOPLASTE.
La technique d6crile par Beattie et al. [16] a 6t6 adapt6e au f rac t ionnement du mitoplaste. Un choc osnmtique p rovoquan t la rup ture des s tructures membrana i r e s ent ra lne une raise en solut ion des prot6ines hydrosolubles essent iel lement matr i - clerics. Les const i tuants insolubles sont ensuite trait6s successivement avec des solutions de KC1
0,15 M e t 0,60 M pour extraire les prot6ines fai- b lement li~es h la membrane interne. Les prot6ines
BIOCHIMIE, 1975, 57, n ° 1.
M. Baudry , J.-P. Clot et R. Michel .
insolubles sont dispers6es ~ l 'a ide de d6tergents, cholate de sodium, d6soxycholate de sodium et dod6cylsulfate de sodium, puis fract ionn~es par addi t ion d 'une solution satur6e et neutral is6e de (NH,)2SO ,, Les prot6ines s tructurales pr6cip i tent
13 p. cent de saturat ion, et ~ 50 p. cent on obtient une f ract ion qui renferme ]es cytochromes structur6s a, a~, b et c v
ELECTROPHO RINSE.
Les prot6ines sont dissoutes dans le m61ange : acide ac6tiqne (20 p. cent), ur6e (24 p. cent), phe- nol (40 p. cent) et mercapto6thanol (5 p. cent) (p/v) [17]. Les gels utilis6s sont aux concentra- t ions finales de 6 p. cent en aerylamide, 0,16 p. cent en m6thyl6nebisacrylamide, 34 p. cent en ur6e, 28 p. cent en aeide ac6tique, 0,4 p. cent en persulTate d ' ammon ium et 0,5 p. cent en t6tram6- thyl6thyl6nediamine. On in t rodui t 13,5 ml du m6- lange dans des colonnes de 1 em de diambtre et la polym6risa t ion s'effectue h la temp6rature am- biante. Des 6chantiI lons con tenan t 1 mg de pro- t6ines par t ieula i res dispers6es dans 0,2,5 ml du m61ange dissolvant sont d6pos6s sur les gels, puts les colonnes et les euves de l ' appare i l (Buehler- 3-1071) sont rernplies avec une solution d 'aeide ae6tique glacial h 10 p. cent. L'61eetrophorbse est r6alis6e h une temp6rature de 4°C pendan t 22 heures sous une tens ion de 320 V. Les prot6ines sont ensuite fix6es e~ color6es dans une solut ion d 'acide t r iehlorac6t ique h 12,5 p. cent r en fe rman t 0,0'5 p. cent de bleu de Comassie. Les trac6s den- si tom6triques sont r6alis6s h 580 nm h l 'a ide d 'un speetrophotom6tre Gilford 6quip6 d 'un suppor t d6plaeement l in6aire.
I~ACTIFS.
L 'acrylamide, le m6thylbnebisacry lamide et le t6 t ram6thyl6thyl6nediamine sont fournis par East- man Kodak Company, Rochester. La 3,5,3'-triiodo- L-thyronine prov ien t de Smith, Kline et F r e n c h Laboratories, Phi ladelphie . Les autres r6actifs sont des produi ts Merck.
RESULTATS.
1. MITOCHONDRIES.
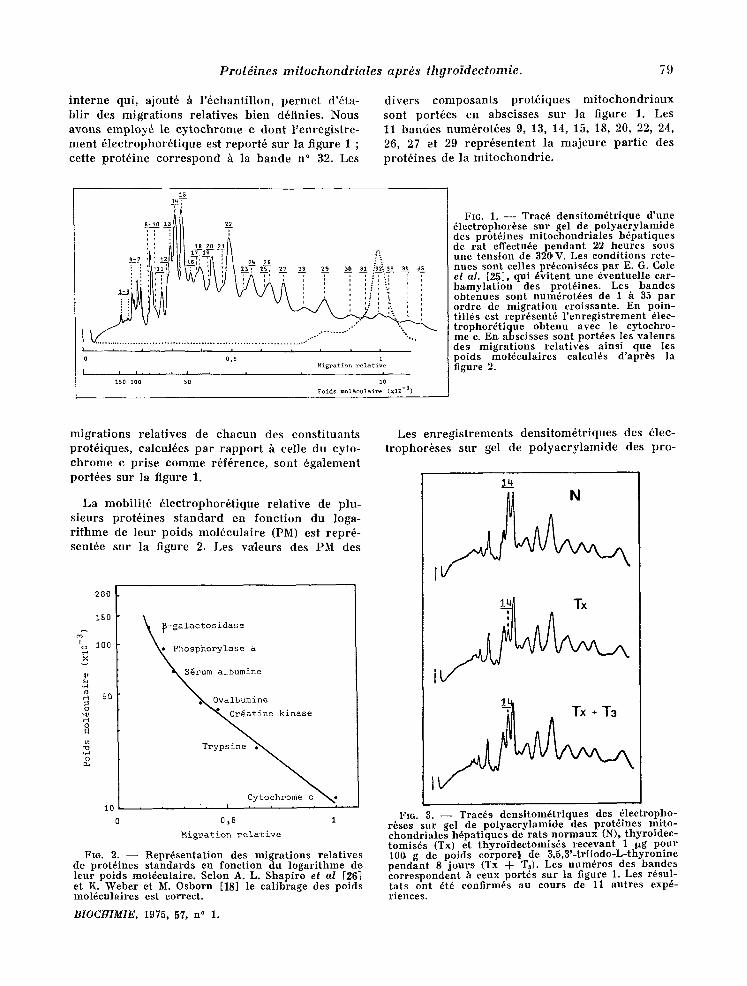
La technique d'61ectrophor6se employee permet la s6parat ion des prot6ines In i tochondr ia les en 35 const i tuants c o l m e le I ontre l ' enregis t rement densi tom6tr ique pr6sent6 sur la figure 1 ; les diff~rentes bandes sont num~rot6es par ordre de migra t ion croissante. Les distances de migra t ion peuvent var ier l~g6rement d 'une exp6rience h l 'autre, aussi est-il pr6f6rable d 'ut i l iser un t~moin
Protdines mitochondriales apr~s thgroidectomie. 79
i n t e r n e qui , a jou t6 h l ' 6 c h a n t i l l o n , p e r m e t d'&ta- b l i r des m i g r a t i o n s r e l a t i v e s b i e n d6f in ies . Nous a v o n s e m p l o y 6 le c y t o c h r o m e c d o n t l ' e n r e g i s t r e - m e n t 6 1 e c t r o p h o r 6 t i q u e est r e p o r t 6 s u r la f igu re 1 ; ce t t e p r o t 6 i n e c o r r e s p o n d & la b a n d e n ° 32. Les
d i v e r s c o m p o s a n t s p r o t 6 i q u e s m i t o c h o n d r i a u x s o n t p o r t 6 e s e n a b s c i s s e s s u r la f igure 1. Les 11 b a n d e s n u m ~ r o t ~ e s 9, 13, 14, 15, 18, 20, 22, 24, 26, 27 et 29 r e p r 6 s e n t e n t la m a j e u r e p a r t i e des p r o t 6 i n e s de la m i t o c h o n d r i e .
15
n-,
8 - 1 0 13 22
', ', ' ~ a 2o 2 1 • , t ~ - - l " ~ - - ' r [ -
~-7 , 12 - q " - - r '
I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . - ' " ' " . . . . . . . . . . . : " " " - . . . . '
(] 0 , 5 Migration relativle
[ i i , , , , I
1 5 0 l O 0 5 0 a0 Poids mol6~ulai~e (xlD -3)
Fro. 1. - - Trac6 dens i tom6t r ique d 'une ~lectrophor6se sur gel de po lyaery lamide des prot~ines mi tochondr ia les h6pat iques de r a t effectu4e pendan t 22 heures sous une tens ion de 320,V. Les condi t ions rete- nues sont celles pr6conis6es pa r E. G. Cole el al. [25], qui 6vi tent une 6ventuel le ear- b a m y l a t i o n des prot6ines. Les bandes obtenues sont num4rot4es de 1 h 35 pa r ordre de migra t ion croissante. En poin- t i l l6s est repr6sent6 l ' en reg i s t r ement 61ec- t rophor4 t iquc ob tenu avec le eytochro- me c. En~ abscisses sont port4es les va leurs des migra t ions re la t ives a ins i que les poids mol'~cnlaires caleul4s d 'apr6s la figure 2.
m i g r a t i o n s r e l a t i v e s de c h a c u n des c o n s t i t u a n t s p r o t 6 i q u e s , e a l cu l6es p a r r a p p o r t h ce l le d u cy to - c h r o m e c p r i s e e o m m e r 6 f 6 r e n c e , s o n t 6 g a l e m e n t p o r t 6 e s s u r la f igu re 1.
L a m o b i l i t 6 6 1 e e t r o p h o r 6 1 i q u e r e l a t i v e de p lu - s i e u r s p r o t 6 i n e s s t a n d a r d e n f o n c t i o n d u loga- r i t h m e de l e u r p o i d s m o l & c u l a i r e (PM) es t r e p r 6 - s en t6e s u r la f igure 2. Les v a l e u r s des P M des
200
~-galactosidase
to 100 k- Phosphorylase a v~ \ ~ u m albumine
~o ~valbumine x~ ~ine kinase Trypsine
o
Cytochrome c ~, 10 , , , ", I , , , , i
0,5 1
Migration ~elative
FIG. 2. -- Repr6senta t ion des migra t ions re la t ives de prot4ines s t andards cn fonet ion du loga r i thme de leur poids mol6culaire. Selon A. L. Shapi ro et al [26] et K. Weber et M. Osborn [18] le eal ibrage des poids mol6culaires est correct.
BIOCHIMIE, 1975, 57, n ° 1.
Les e n r e g i s t r e m e n t s d e n s i t o m 6 t r i q u e s des 61ec- t r o p h o r ~ s e s s u r gel de p o l y a c r y l a m i d e des p r o -
Tx
Fro. 3. - - Trac~s dens i tom6t r iques des 61eetropho- r~ses sur gel de po lyacry lamide des prot~ines mi to- chondr ia les h6pa t iques de r a t s n o r m a u x (N), thyro idee- tomis6s (Tx) et thyro idec tomis6s recevalLt 1 ~g pour 10(~ g de poids corpore[ de 3,5,3 ' - t r i iodo-L-thyronine pendan t 8 j ou r s (Tx + Ts). Les num6ros des bandes cor respondent h ceux p o r t , s sur la figure 1. Les r6sul- t a t s ont ~td conflrmfis au eonrs de 11 aut res exp$- riences.
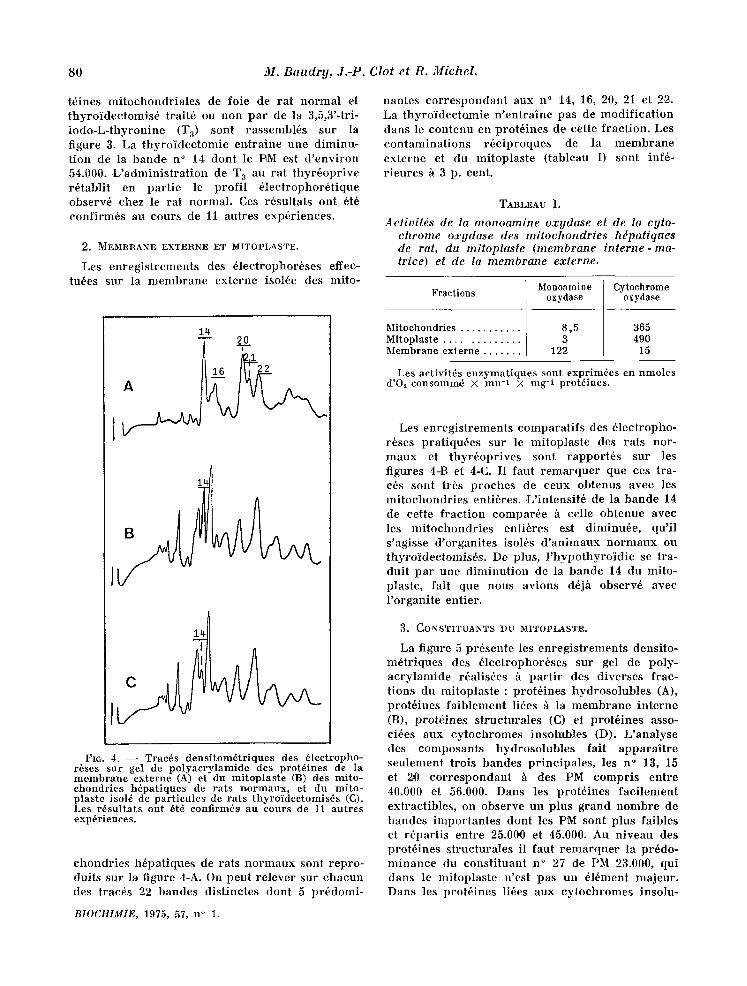
80 M. Baudrg, J.-P. Clot et R. Michel.
t6ines mi tochondr ia les de foie de rat normal et thyro idec tomis6 trait6 ou non par de la 3,5,3'-tri- iodo-L- thyroniue (T 8) sont rassembl6s sur la figure 3. La thyro ' idectomie ent ra ine uue diminu- t ion de la bande n ° 14 dont le PM est d ' env i ron 54.0,00. L ' admin i s t r a t i on de T 3 all rat thyr6opr ive r6tablit en par t ie le prof i l 61ectrophor6tique observ6 chez le rat normal. Ces r6sultats ont 6t6 confirm6s au cours de 11 autres exp6riences.
2. MEMBRANE EXTERNE ~1' MITOPLASTE.
Les enregis t rements des 61ectrophor~ses effec- tu6es sur la membrane externe isol6e des ulito-
A
14 20
:t
Fro. 4. - - Trae6s densitom6triques des ~lectropho- r6ses sur gel de polyacrylamide des prot6ines de la membrane externe (A) et du mitoplaste (]3) des mito- chondries h6patiques de rats normaux, et du mito- plaste isold de particules de rats thyroidectomis~s (C). Les r~sultats ont 6t6 confirmds au cours de 11 autres experiences.
chondr ies h6pat iques de rats no rmaux sont repro- dnits sur la figure 4-A. On peut re lever sur chacun des trac6s 22 bandes dist iuctes dont 5 pr6domi-
nantes cor respondant aux n ° 14, 16, 20, 21 et 22. La thyr0idectoxnie n ' en t ra ine pas de modi f ica t ion dans le contenu en prot6ines de cette f ract ion. Les contamina t ions r6c iproques de la membrane externe et du mitoplas te (tableau I) sont inf6- r ieures h 3 p. cent.
TABLEAU ].
Activit~s de la monoamine oxgdase et de la cgto- chrome oxgdase des mitochondries hdpatiques de rat, du mitoplaste (membrane interne-ma- trice) et de la membrane externe.
Fractions
Mitochondries . . . . . . . . . . . Mitoplaste . . . . . . . . . . . . . Membrane externe . . . . . . .
Monoamine oxydase
8,5 3
122
Cytochrome oxydase
365 490
15
Les activitfis enzymatiques sont exprim~es en nmoles d'O2 consomm,6 × ran-1 X rag-1 prot~ines.
Les enregis t rements compara t i f s des 61ectropho- r6ses prat iqu6es sur le mitoplaste des rats nor- maux et thyr6opr ives sont rapport6s sur les figures 4~B et 4-C. n faut r emarque r que ces tra- c6s sont tr6s proches de ceux obtenus avec les mi tochondr ie s enti6res. L ' intensi t6 de la bande 14 de cette f rac t ion compar6e h celle obtenue avec les mi tochondr ie s enti6res est diminu6e, qu ' i l s 'agisse d 'organi tes isol6s d ' an imaux normaux ou thyroidectomis6s . De plus, l ' hypo thy ro id i e se tra- duit pa r une d iminut ion de la bandc 14 du mito- plaste, fait que nous avions d6jh observ6 avec l 'organi te entier.
3. CONSTITUANTS DU 3~IITOPLAST~.
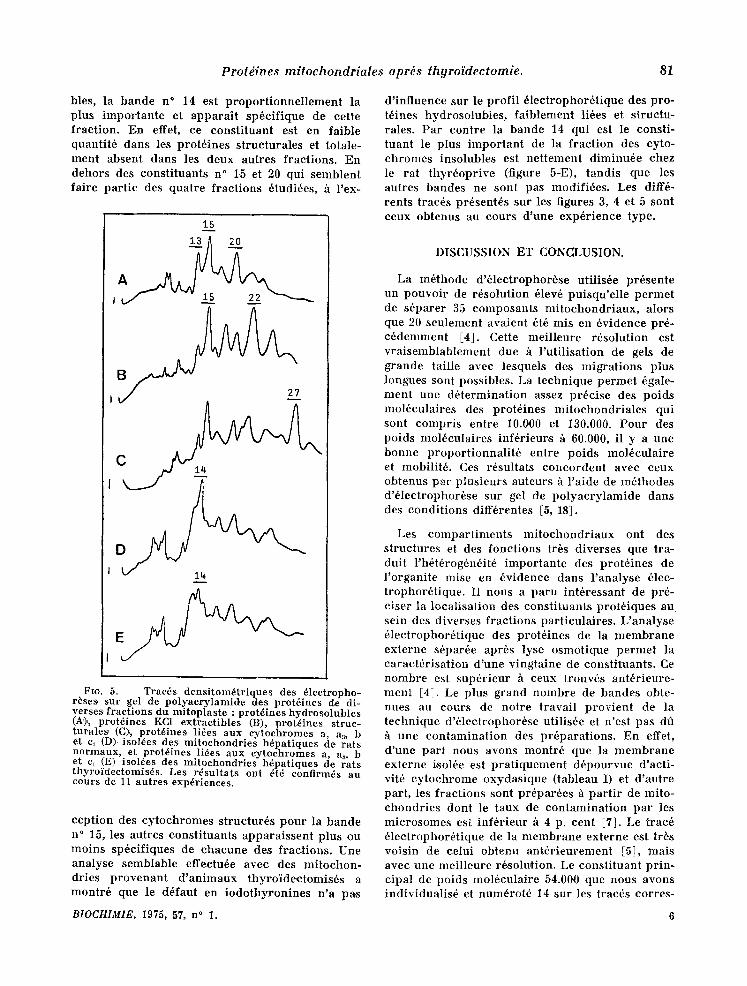
La figure 5 prdsente les enregis t rements densito- m6tr iques des 61ectrophor6ses sur gel de poly- ac ry lamide r6alis~es h pa r t i r des diverses frac- t ions du mitoplaste : prot6ines hydrosolubles (A), prot6ines faiblemen~t li6es h la membrane in te rne (B), prot6ines s tructura]es (C) et prot6ines asso- ci6es aux cy tochromes ins olubles (D). L 'analyse des composants hydrosolubles fait appara l t re seulement trois bandes pr inc ipa les , les n ° 13, 15 et 2~) co r respondan t h des PM eompr is entre 40.000 et 56.000. Dans les prot6ines fac i lement extract ibles, on observe un plus grand nombre de bandes impor tantes dont les PM sont plus faibles et r6part is entre 25.00,0 et 45.000. Au niveau des prot6ines s t ructurales il' faut r e m a r q u e r la pr6do- minance du const i tuant n ° 27 de PM 23.000, qui dans le mitoplaste n 'est pas un 616ment majeur . Darts les prot6ines li6es aux cy tochromes insolu-
BIOCHIMIE, 1975, 57, n ° 1.
Protdines mitochondriales aprds thgro~'dectomie. 81
bles, la bande n ° 14 est p ropor t ionne l l emen t la plus impor tan te et appara i t sp6cifique de cette fraction. En effet, ce const i tuant est en faible quantit6 dans les prot6ines s tructurales et totale- ment absent dans les deux autres fractions. En dehors des const i tuants n ° 15 et 20 qui semblent faire par t ie des quatre fract ions 6tudi6es, ~ l 'ex-
15
D~ v~ ~ ~ ,-~,~.
FIG. 5. - - Trac~s densitom6triques des 61ectropho- r~ses sur gel de polyacrylamide des prot6ines de di- verses fractions du mitoplaste : prot~ines hydrosolubles (A:),, prot6ines KCI extractibles (B), prot~ines struc- turales (C)~ prot6ines liSes aux cytochromes a, as, b et c~ (D)isol~es des mitochondries htpatiques de rats normaux, et prot~ines li6es aux cytochromes a, aa, b et c~ (E) isol~es des mitoehondries h~patiques de rats thyrofdectomlsds. Les r~sultats ont ~td confirmds au cours de 11 autres experiences.
cept ion des cytochromes structur6s pour la bande n ° 15, les autres const i tuants apparaissent plus ou moins sp6cifiques de chacune des fractions. Une analyse semblable effectu6e avec des mitochon- dries p r o v e n a n t d ' an imaux thyroidectomis6s a montr6 que le d6faut en iodothyronines n 'a pas
BIOCHIMIE, 1975, 57, n ° I.
d ' inf luence sur le prof i l 61ectrophor6tique des pro- t6ines hydrosolubles , fa iblement li6es et s tructu- rales. Par contre la bande 14 qui est le consti- tuant le plus impor tan t de la fract ion des cyto- chromes insolubles est ne t tement diminu6e chez le rat thyr6oprive (figure 5-E), tandis que les autres bandes ne sont pas modifi6es. Lcs diff6- rents trac6s pr6sent6s sur les figures 3, 4 et 5 sont ceux obtenus au cours d 'une expdrience type.
DISCUSSION ET CONCLUSION.
La m6thode d'61ectrophor+se utilis6e pr6sente un pouvoir de r6solution 61ev6 puisqu'el le permet de s6parer 35 composants mi tochondr iaux , alors que 20 seulement avaient ~t6 mis en 6vidence pr6- c6demment [4]. Cette meil leure r6solution est vra isemblablement due h l 'u t i l i sa t ion de gels de grande taille avec lesquels des migrat ions plus longues sont possibles. La technique permet 6gale- ment une d6terminat ion assez pr6cise des poids mol6culaires des prot6ines mi tochondr ia les qui sont compris entre 10.0,00 et 130.000. Pour des poids mol6culaires inf6rieurs h 60.000, il y a une bonne propor t ionnal i t6 entre poids mol6culaire et mobilit6. Ces r6sultats concordent avec ceux obtenus par plusieurs auteurs ~t l 'a ide de m6thodes d'61ectrophor+se sur gel de po lyacry lamide dans des condi t ions diff6rentes ~5, 18].
Les compar t iments mi tochondr iaux ont des s t ructures et des fonct ions tr+s diverses que tra- duit l 'h6t6rog6n6it6 impor tante des prot6ines de l 'organi te nfise en 6vidence dans l 'analyse ~lec- trophor6tique. I1 nous a paru int6ressant de pr6- ciser la localisat ion des const i tuants prot~iques a u sein des diverses fract ions part iculaires . L 'analyse 61ectrophor6tique des prot6ines de la membrane externe s6par6e apr~s lyse osmotique permet la caract6risat ion d 'une vingta ine de const i tuants . Ce hombre est sup6rieur h ceux trouv6s ant~rieure- merit [4]. Le plus grand nombre de bandes obte- nues au cours de notre t ravai l provient de la technique d'61ectrophor~se utilis6e et n 'est pas dft h une contamina t ion des pr6parat ions. En offer, d 'une par t nous avons montr6 que la membrane externe isol6e est p ra t iquement d6pourvue d'acti- vit~ cytochrome oxydasique (tableau I) et d 'autre part, les fract ions sont pr6par6es h par t i r de mito- chondries dont le taux de con tamina t ion par les microsomes est inf6r ieur h 4 p. cent [7]. Le trac~ 61ectrophor6tique de la membrane externe est tr6s voisin de celui obtenu ant6r ieurement [5], mats avec une meil leure r6solution. Le const i tuant p r in- cipal de poids mol6culaire 54.000 que nous avons individual is6 et num~rot~ 14 sur les trac~s corres-

82 M. Baudry, J.-P. Clot et R. Michel.
pond ~ la f ract ion majeure de poids mol6culaire 50.000 envi ron mise 6galement en 6vidence par Melnick et al. [5].
Une 6rude comparat ive de l ' intensi t6 des bandes des diverses f ract ions dolt ten i r compte de l ' im- por tance relative de chaeun des compar t iments mi tochondr iaux . C'est ainsi que le profil 61ectro- phor6t ique du mitoplaste est trbs proche de celui des mi tochondr ies puisque la membrane externe et l 'espace i n t e rmembrana i r e ne repr6sentent que 10 p. cent envi ron des prot6ines totales [19, 20]. La seule diff6rence concerne la bande 14, Pun des const i tuants les plus impor tan ts de l ' enveloppe externe, qui est d iminu6e dans 1'ensemble mem- brane i n t e r n e - matrice. On ne peut pas cependant affirmer que les bandes num6rot6es 14 dans le mitoplaste et darts la membrane externe repr6- sentent soit le mSme const i tuant , soit deux pro- t6ines diff6rentes que la technique d'61ectropho- rbse ne permet pas de s bparer, leurs poids mol6- culaires 6taut tr6s voisins.
L 'analyse 61ectrophor6fique des fract ions pro- t6iques du mitoplaste a montr6 que les prot6ines hydrosolubles matriciell, es ont des poids mol6cu- laires compris entre 39.000 et 56.000. Par contre, les composants fa iblement li6s h la membrane in terne , KC,1 extractibles, out des poids mol6cu- h i r e s plus faibles compris entre 25.000 et 45.000. Darts la f ract ion contenant les prot6ines struc- turales, trois const i tuants para i ssen t impor tan t s dont le p r inc ipa l correspond h la prot6ine s t ruc- turale de poids mol6culaire 23.00,0 caract6ris6e par Criddle et al. [21]. Celle-ci est trbs sp6cifique de la fract ion s t ructurale et n ' appa ra i t pas sur le trac6 densi tom6tr ique des prot6ines li6es aux cytochromes insolubles dont le composant pr in- cipal a un poids mol6culaire d ' env i ron 54.000.
La r6par t i t ion des composants mi tochondr i aux dans les divers compar t iments et les fract ions du mitoplaste ayant 6t6 6tablie, nous avons compl6t6 ce t ravai l avec des pr6para t ions p rovenan t d 'ani- maux plac6s darts des 6tats m6taboliques et fonc- t ionnels vari6s. Compte tenu de la sp6cificit6 d 'act ion des hormones thyro id iennes sur tes pro- pri6t6s des mi tochondr ies , nous avons repris les analyses avec des pr6para t ions p rovenan t d 'ani- maux se t rouvant dans diff6rents 6tats thyroi- diens. La thyroidectomie ne semble pas affecter l'a teneur des const i tuants de la membrane externe et pour tan t les activit6s des enzymes de cette membrane sont net tement attgmen,t6es par le d6- taut d 'hormones thyro id iennes [81. Cependant , les modif icat ions d'activit6 n ' imp l iquen t pas n6ees- sa i rement des var ia t ions dans le contenu en pro-
t6ines enzymatiques. Nos r6sultats sont ~ rappro- cher de ceux obtenus par Gear au cours d'exp6- r iences r6alis6es avec le foie en r6g6n6ration apr6s h6patectomie part iel le [22]. Les activit6s de la monoamine oxydase et de la cynur6n ine hyi iroxylase 6taient alors abaiss6es, b ien qu ' i l n 'y ait pas de diff6renee dans les profils 61ectrophor6- t iques de la membrane externe. I1 est 6galement possible que les prot6iues enzymatiques ne repr6- sentent qu 'une faible par t ie des prot6ines mem- brana i res totales [23] et qu 'une modif ica t ion mi- neure dans leur teneur ne soit pas d6celable sur les trac6s 61eetrophor6tiques.
I1 ressort des r6sultats obtenus avec les orga- nites entiers que la bande n ° 14 d6pend de l'.~tat thyroidien. En effet elle est for tement d iminu6e chez les an imaux thyr6oprives, l ' in jec t ion de T z h ces derniers r amenan t la teneur de ce compo- sant presqu 'h la normale. Cette prot6ine fait par t ie des const i tuants for tement structur6s de la mem- b rane in te rne puisqu ' i l taut un t ra i tement 6ner- gique avec des d6tergents pour la solubiliser. Elle est associ6e ~t la f ract ion r en fe rman t les eyto- chromes a, a 3, b e t c I et il est possible qu'elle leur soit li6e. Ge const i tuant pour ra i t 6tre impli- qu6 dans les m6canismes d 'oxydat ions phospho- rylantes sans qu' i l s 'agisse n6cessairement d 'un coenzyme respiratoire. Beattie et al. [24~ ont mon- tr6 que l ' i neorpora t ion d 'acides amin6s marqu6s par les mi tochondr ies isol6es se fair h la fois dans les ¢ prot6ines s t ructurales >> et dans Ia f ract ion renfe rmant les cytochromes insolubles. II appa- rai t donc qu 'une ou plusieurs des prot6ines de eette dernibre fract ion que nous avons trouv6e 6tre modifi6e par la thyroidectomie sont sous la d6pendance direete des acides nuel6iques mito- chondr iaux . On peut envisager que les hormones thyro id iennes modif ient la teneur de ce consti- tuan t par l ' in term6dia i re du syst6me de synth6se prot6ique nfi tochondriale .
L 'act ion s61ective des iodothyronines sur les propri6t6s et la prot6inog6nbse de la membrane in te rne s 'accompagne ainsi de modif icat ions structurales. L 'ut i l isat ion ul t6rieure de pr6para- t ions mito.chondriales p rovenan t d ' an imaux se t rouvant dans diff6rents 6tats m6taboliques peut done fourn i r une m6thode indi rec te pour l '6tude des s t ructures de la mi tochondr ie .
P~suM~.
Les mitochondries de foie de rats sont analysdes par 61eetrophor6se sur gel de polyacrylamide. Les parti- cules provenant d'animaux normanx et thyroidecto- mis6s renferment 35 constituants dont les poids mol6- culaires sont eompris entre 10 000 et 135 000. Le d6faut
BIOCHIMIE, 1975, 57, n ° 1.

Prot~ines mitochondriales apr~s thyroidectomie. 83
en iodothyronines se t radui t par une nette dimimation dans la teneur d 'un const i tuant de poids moldeulaire 54 000, l ' injeet ion de 3,5,3'-triiodo-L-thyronine h l 'ani- real thyrdoprive la rdtabl issant h une valeur proehe de la normale.
La membrane externe et le mitoplaste sont sdpards. La membrane externe est constitude de 20 fract ions prot~iques, l 'ablat ion de la thyroide n ' en t ra inan t pas de modification strueturale. On retrouve au niveau du mitoplaste la diffdrence observde avee les mitochon- dries entidres eoncernant le const i tuant de poids mo- 16eulaire 54 900 qui est diminu~ par rhypothyro id ie .
Le mitoplaste est fractionnd en protdines matr i - cielles, protdines fa ih lement lides aux membranes , pro- t~ines s t rueturales et prot~ines des cytochromes inso- luhles a, as, b e t el. Les protdines matrieiel les hydro- solubles ont des poids moldeulaires compris pr ineipa- lement entre 40000 et 55 000, les eomposants KC1- extraetibles entre 25 0O0 et 45 000. Les protdines struc- rurales renfe rment un eonst i tuant trds spdeifique de poids mol~culaire 23090. Les traeds densi tom~triques obtenus avee des f ract ions sont les m~mes quel que soit F~tat thyroidien tandis que le const i tuant prin- cipal des protdines lides aux cytoehromes insolubles dont le poids moldeulaire est de 54 000, est diminu6 chez / ' animal thyrdoprive.
La thyroideetomie provoque done la d iminut ion de la teneur d 'un const i tuant prot~ique for tement lid h la membrane interne mitochondria le et assoei~ aux eytoehromes a, a~, b e t c~. Cette fract ion fait part ie des eomposants dont certains sont sous le eontrdle des aeides nuel~iques mitochondriaux.
BIBLIOGRA.PHIE.
1. Parsons, D. F. & Yano, Y. (19~67) Biochim. Biophys . Acta, 135, 362-364.
2. Ashwell, M. & Work, T. S. (197'0) Ann. Rev. Bio- chem., 39, 251-290.
3. Hatefi, Y., Haavik, A. G., Fowler, L. R. & Griffiths, D. E. (1962). J. Biol. Chem., 237, 2661-2669.
4. Schnaitman, C. (1969) Proc. Nat. Acad. Sci., 63, 412-419.
5. Melniek, R. L., Tinberg, H. M., Maguire, J. & Packer, L. (1973) Biochim. Biophys. Acta, 311, 230-241.
6. Takayama, K., Mac Lennan, D. H., Tzagoloff, A. & Stoner, C. D. (1.966) Arch. Biochem. Biophys., 114, 223-230.
7. Baudry, M., Bouhnik, J., Michel, O. & Michel, R. (1972) Biochimie, 54, 219-227.
8. Clot, J.-P., Baudry, M. & Michel, R. (1973) Bio- chimie, 55, 451-456.
9. Bargeton, D. & Krumm-Heller, C. (1948) Compt. Rend. Soc. Biol., 142, 899-903.
10. Beattie, D. S. (1968) Biochem. Biophys. Res. Com- mun., 31, 901-907.
11. Parsons, D. F., Will iams, G. R. ~ Chance, B. (1966) Ann. N. Y. Acad. Sci., 137, 643-666.
12. Michel, R., Truchot, R., Autissier, N. & Rosner, B. (1966) Biochem. Pharmac., 15, 112r7-1138.
13. Sehnai tman, C., Erwin, V. G. & Greenawalt, J. W. (1967) J. of Cell Biol., 32, 71.9-735.
14. Gornall, A. G., Bardawill , C. J. & David, M. M. (1949) J. Biol. Chem., 177, 751-766.
15. Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall , R. J. (1951) J. Biol. Chem., 193, 265-275.
16. Beattie, D. S., Basford, R. E. & Koritz, S. B. (1966) Biochemis try , 5, 926-930.
17. Ray, T. K. & Marinetti, G. V. (1.9'71) Biochim. Bio- phys. Acta, 233, 787-791.
18. Weber, K. & Osborn, M. (1969) J. Biol. Chem., 244, 44'06-4412.
19. Sehnai tman, C. & Greenawalt, J. W. (1,968) J. of Cell Biol., 38, 158-175.
20. Brunner, G. & Bueeher, T. (1970) FEBS Left., 6, 1ff5-108.
21. Criddle, R. S., Boek, R. M., Green, D. E. & Tisdale, H. (1.962) Biochemis try , 1, 827-842.
22. Gear, A. R. L. (1970) Biochem. J., 120, 577-587. 23. Green,, D. E., Haard, N. E., Lenaz, G. & Silman, H. I.
(1968~ Proc. Nat. Acad. Sci., 60, 277-284. 24. Beattie, D. S., Basford, R. E. & Koritz, S. B. (19~7)
Biochemis try , 6, 3099-31,06. 25. Cole, E. G. ~ Mecham, D. K. (1966) Anal. Biochem.,
1~, 215-222. 26. Shapiro, A. L., Vinuela, E. & Maizel, J. V., Jr. (1967)
Biochem. Biophys. Res. Commnn., 28, 815-820.
BIOCHIMIE, 1975, 57, n o 1.