Embed Size (px)
Citation preview

L3-BH01Cours n°12
Traduction (1)
Ce cours est présent sur le web à l’adresse suivante :http://www.univ-orleans.fr/sciences/BIOCHIMIE/L/ressources.htm
1

Introduction
I- Code génétique : comment l’ADN code-t-il l’information génétique?codons = triplets de basesdécryptage du codecaractéristiques
II- ARNt – aminoacylationstructures primaire & secondaire des ARNtstructure tertiaire des ARNtaminoacyl-ARNt synthétasesinteractions codon-anticodon
III- Ribosomesstructurefonction
IV- synthèse polypeptidiquevue d’ensembleinitiationélongationterminaisonprécisionantibiotiques
Récapitulatif
Plan
2

Synthèse des protéines : avancées historiques majeures
Début des années 1950 :
1- détermination du lieu de synthèse des protéinesau sein de particules ribonucléoprotéiques
2- mise en évidence de l’activation des acides aminésattachement des acides aminés à un ARN particulier
3- caractérisation d’une molécule « adaptatrice » ARNt « traduisent » la séquence nucléotidique du messager en
une séquence d’acides aminés
3

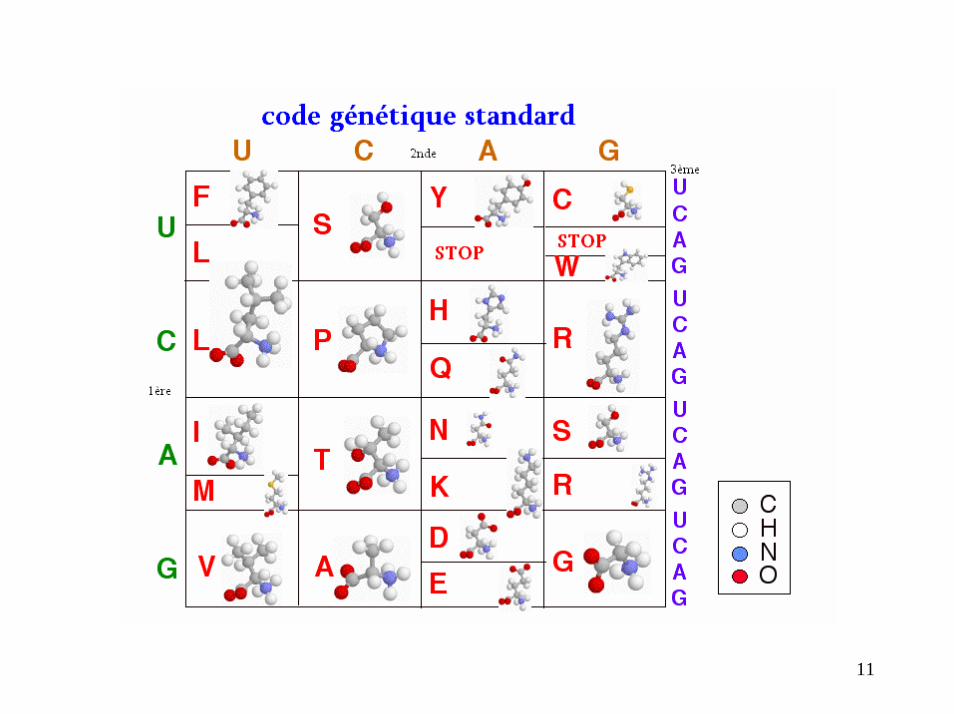
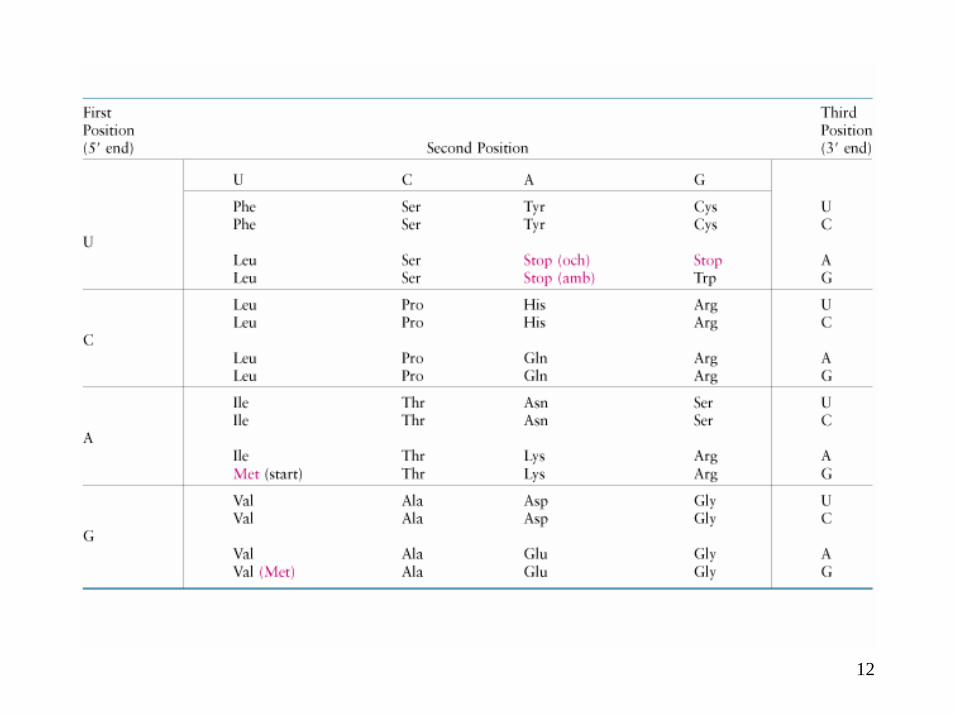
Code génétique
4

Les codons sont des triplets : démonstration expérimentale
Décryptage du code génétique
Caractéristiques principales du code génétique
ADN GAT
ARN CUA
ARNt GAU
aa Leu
5
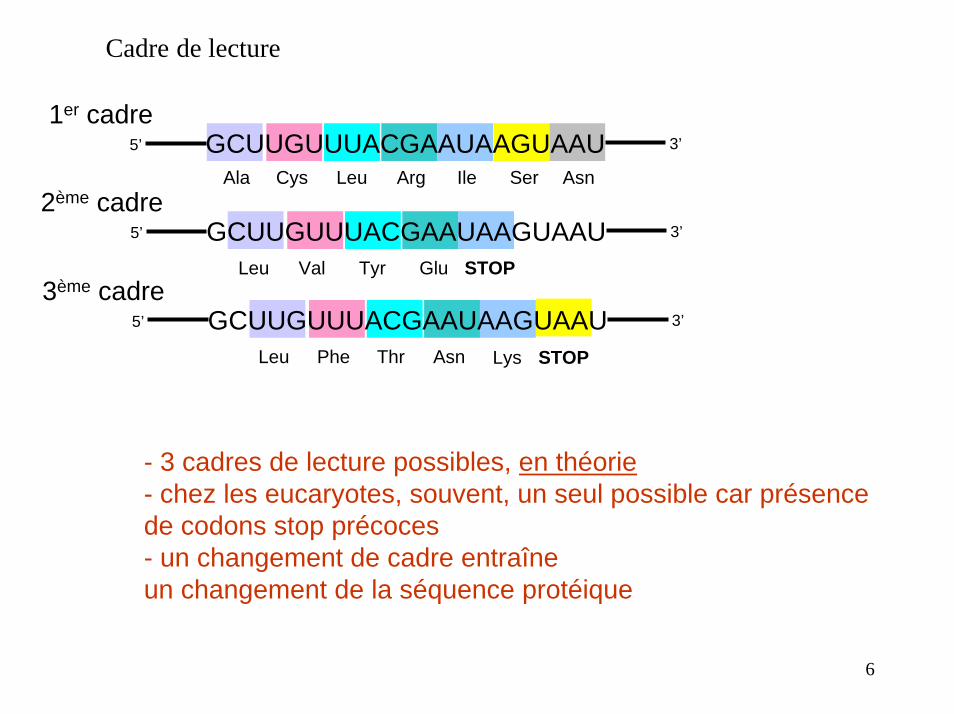
Cadre de lecture
1er cadreGCUUGUUUACGAAUAAGUAAU 3’5’
Ala Cys Leu Arg Ile Ser Asn2ème cadre
GCUUGUUUACGAAUAAGUAAU 3’5’
STOPLeu Val Tyr Glu3ème cadre
GCUUGUUUACGAAUAAGUAAU 3’5’
Leu Phe Thr Asn STOPLys
- 3 cadres de lecture possibles, en théorie- chez les eucaryotes, souvent, un seul possible car présencede codons stop précoces- un changement de cadre entraîne un changement de la séquence protéique
6

Incorporation des acides aminés dans des polypeptides en présence de polymères aléatoires d’ARN :
Expérience de Nirenberg & Matthaei (1961)
EXPERIENCEIncubation d’un polyribonucléotide synthétique polyurydilate (polyU) avec un extrait d’E. coli, du GTP et un mélange de 20 acides aminés (dans 20 tubes différents) : dans chaque tube un acide aminé différent est radioactif .
cet ARN doit engendrer la synthèse d’un polypeptide à partir d’un seul des 20 acides aminés codés par le triplet UUU
RESULTATFormation d’un polypeptide radioactif dans un seul des 20 tubes ;celui contenant la phénylalanine radioactive.
7

Incorporation des acides aminés dans des polypeptides en présence de polymères aléatoires d’ARN
8
Le triplet AAAAAA code la lysine
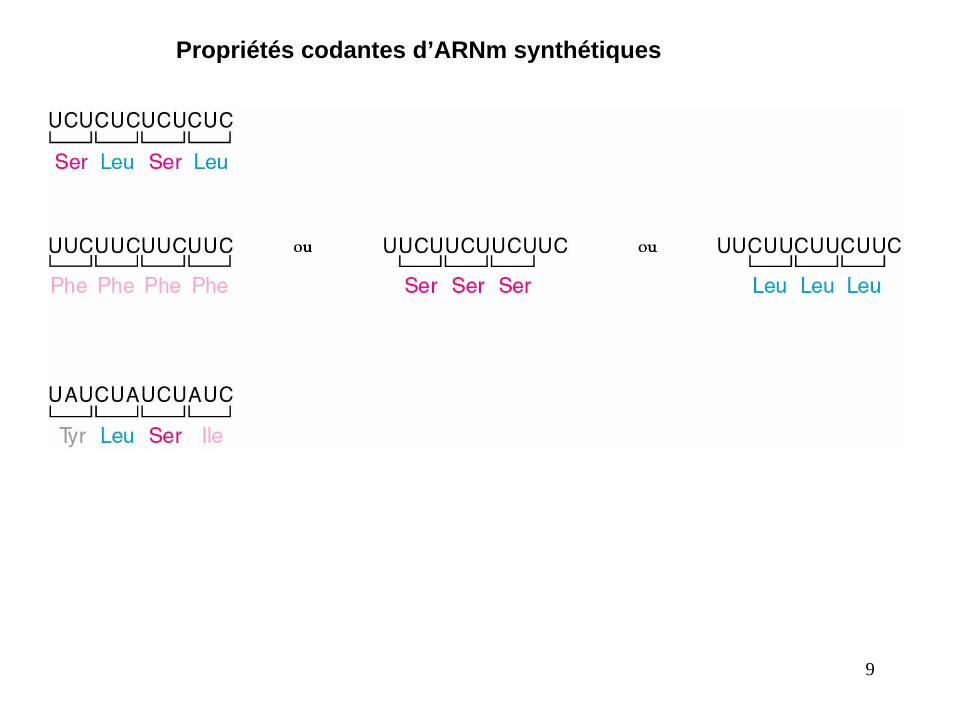
Propriétés codantes d’ARNm synthétiques
9
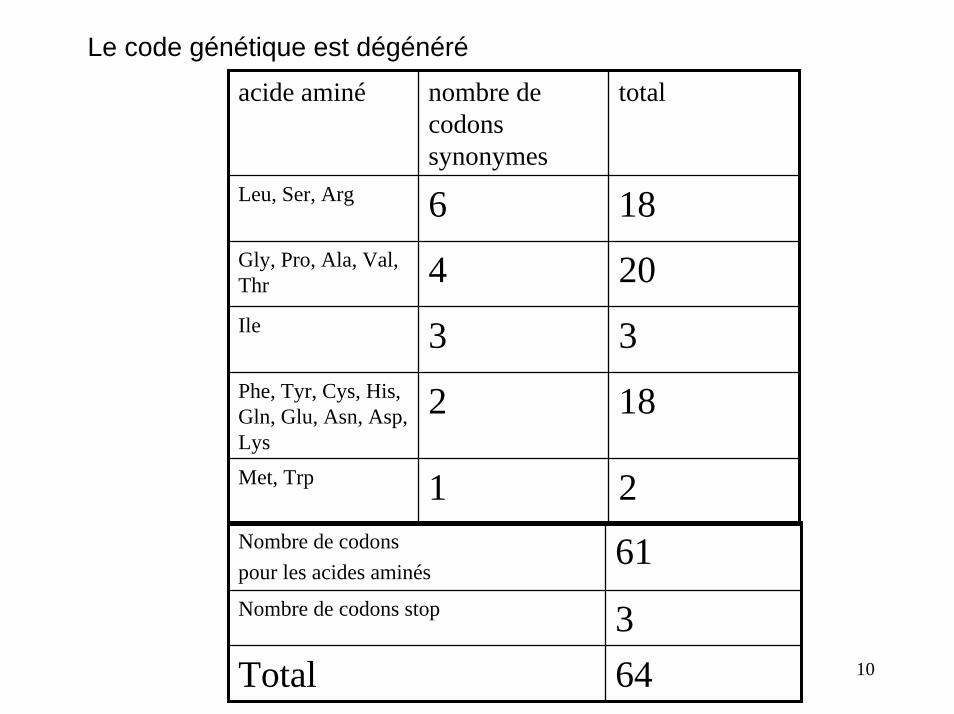
Le code génétique est dégénéré
acide aminé nombre de codons synonymes
total
Leu, Ser, Arg 6 18Gly, Pro, Ala, Val, Thr 4 20Ile 3 3Phe, Tyr, Cys, His, Gln, Glu, Asn, Asp, Lys
2 18
Met, Trp 1 2Nombre de codonspour les acides aminés
61Nombre de codons stop 3Total 64 10
11
12
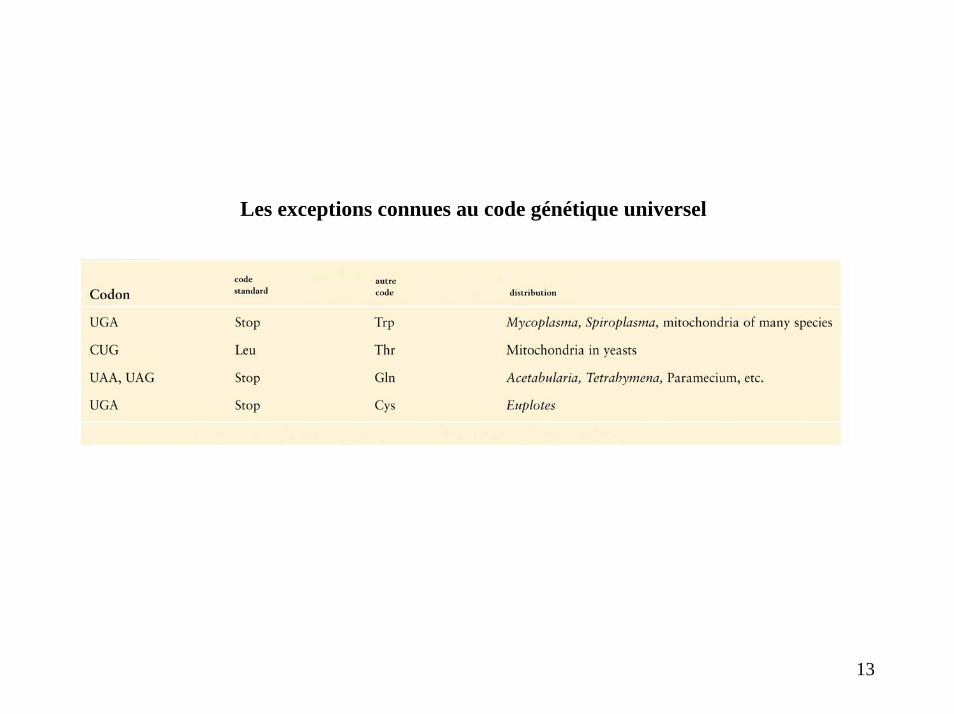
Les exceptions connues au code génétique universel
13
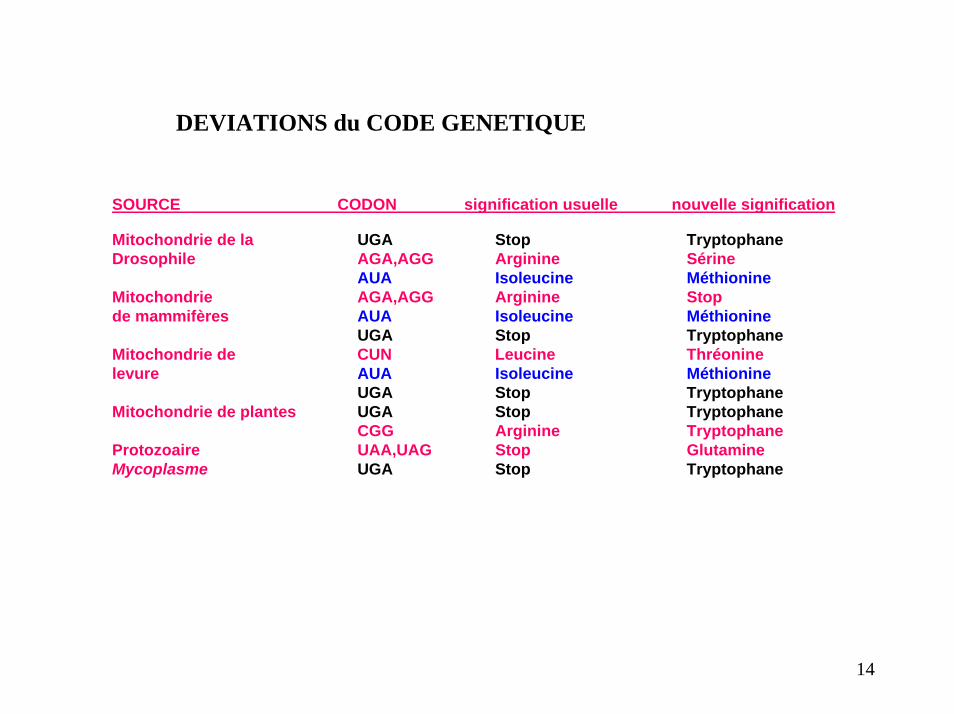
DEVIATIONS du CODE GENETIQUE
SOURCE CODON signification usuelle nouvelle signification
Mitochondrie de la UGA Stop TryptophaneDrosophile AGA,AGG Arginine Sérine
AUA Isoleucine MéthionineMitochondrie AGA,AGG Arginine Stopde mammifères AUA Isoleucine Méthionine
UGA Stop TryptophaneMitochondrie de CUN Leucine Thréoninelevure AUA Isoleucine Méthionine
UGA Stop TryptophaneMitochondrie de plantes UGA Stop Tryptophane
CGG Arginine TryptophaneProtozoaire UAA,UAG Stop GlutamineMycoplasme UGA Stop Tryptophane
14

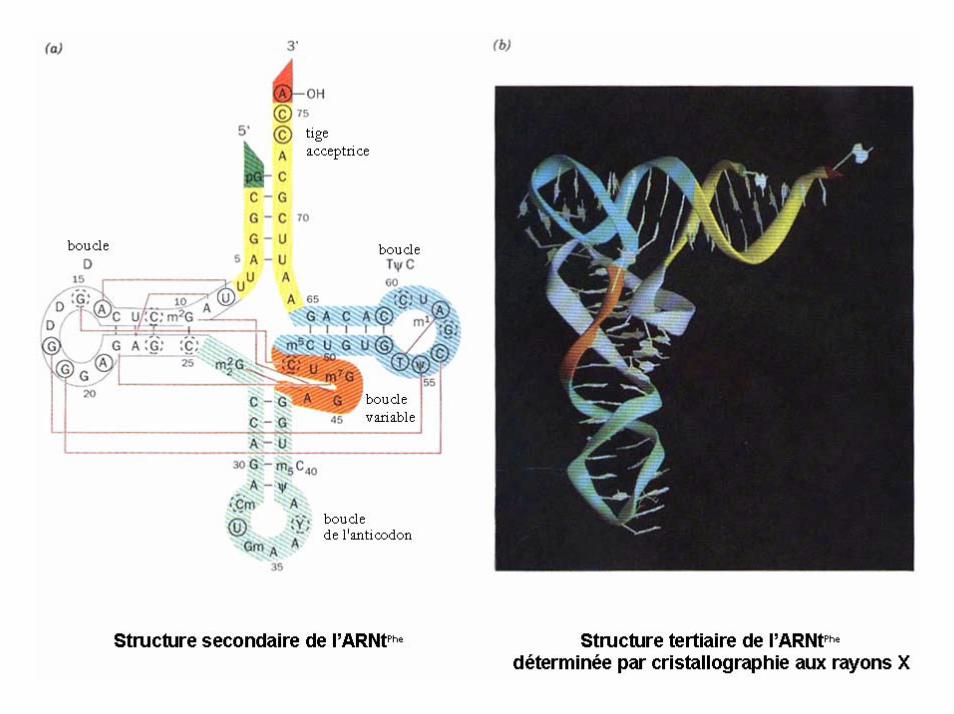
ARN de transfert
15
16
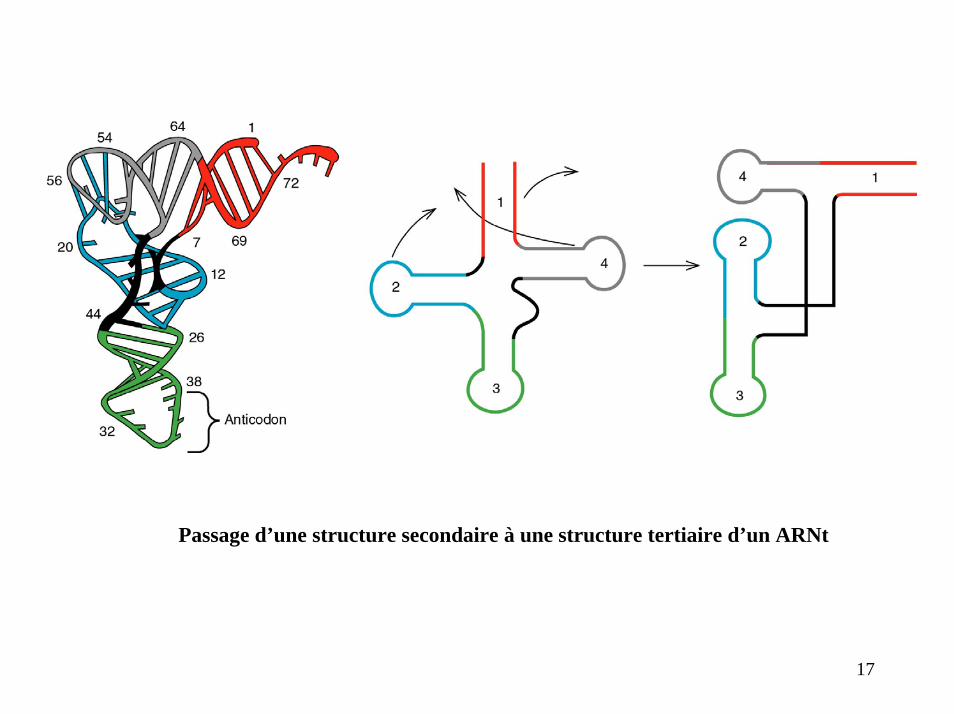
Passage d’une structure secondaire à une structure tertiaire d’un ARNt
17
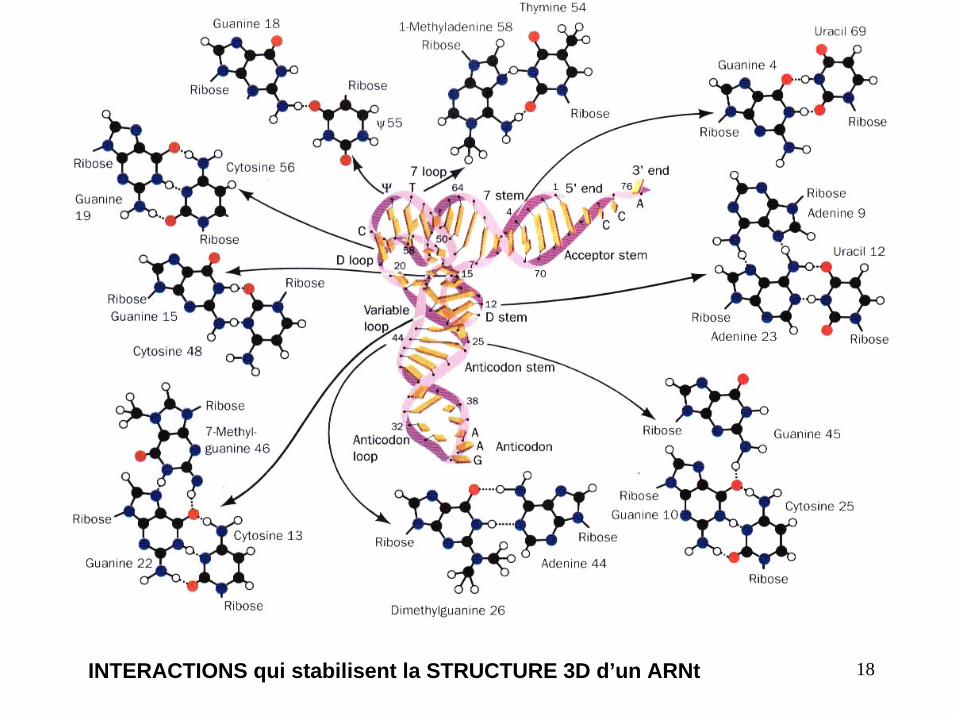
18INTERACTIONS qui stabilisent la STRUCTURE 3D d’un ARNt
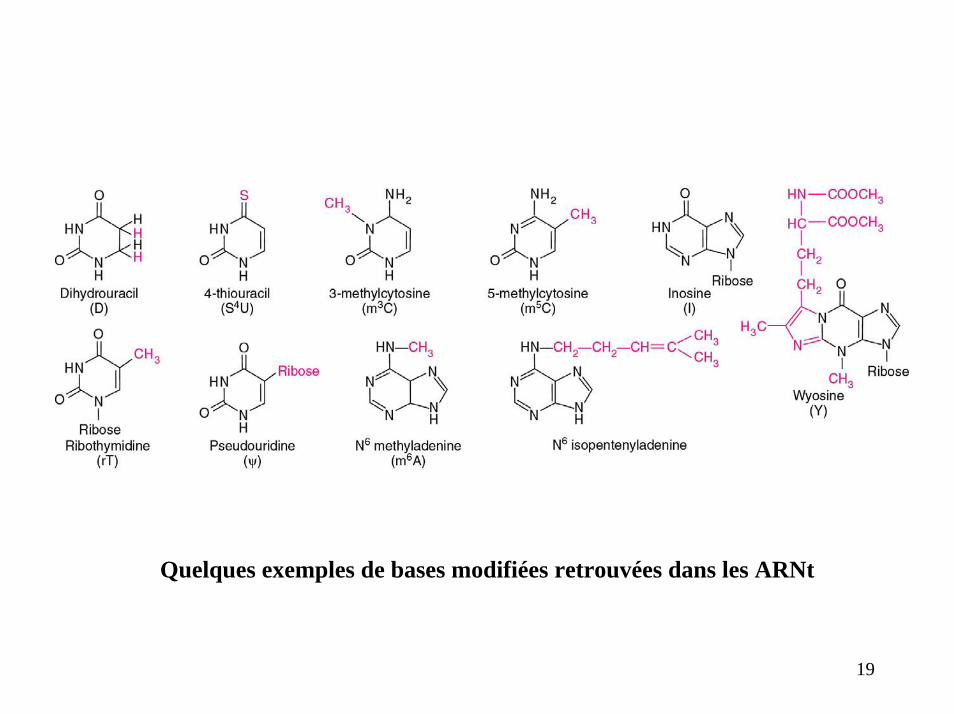
Quelques exemples de bases modifiées retrouvées dans les ARNt
19
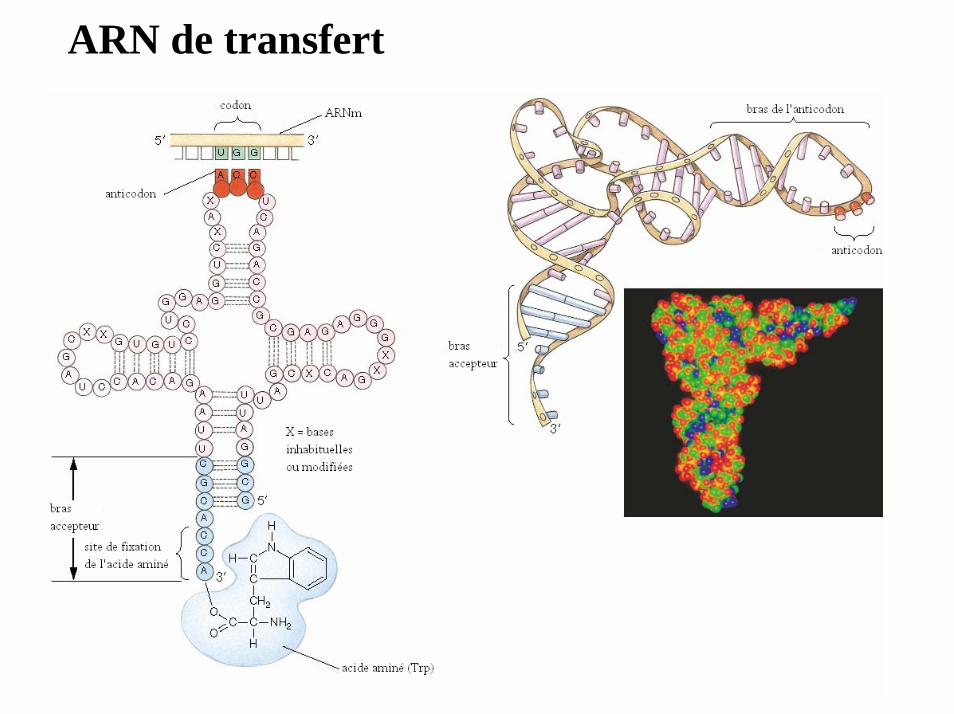
ARN de transfert
20

Expérience montrant que des trinucléotides suffisent pour induire la liaison spécifique des aminoacyl-ARNt aux ribosomes
Liaison du LYSYL-ARNt aux RIBOSOMES selon les CODONS
21

Amino-acyl ARNt synthétases
22
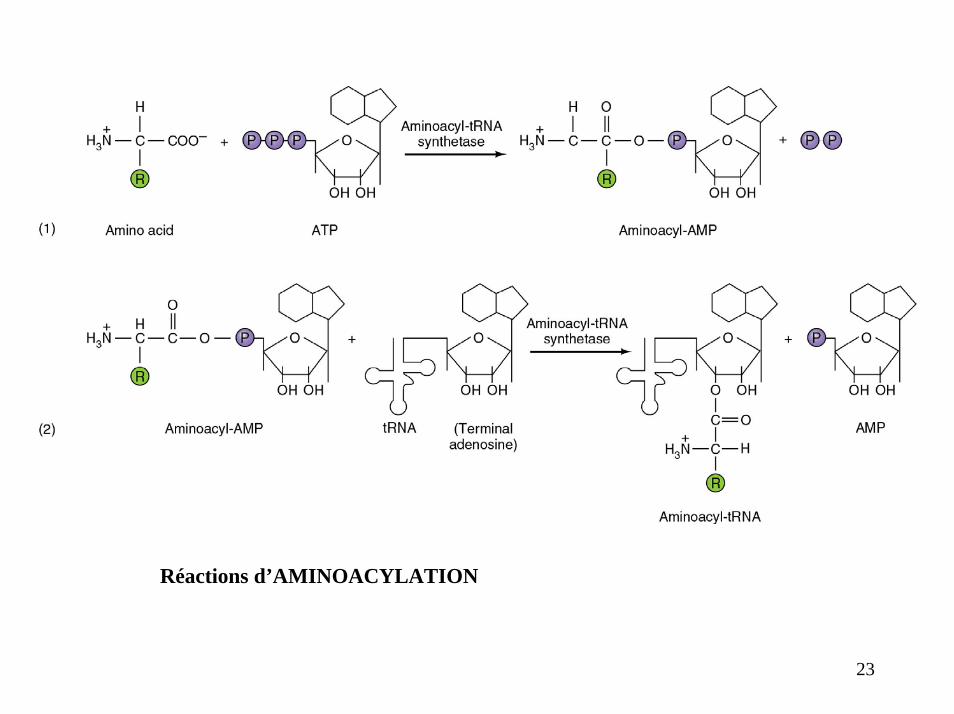
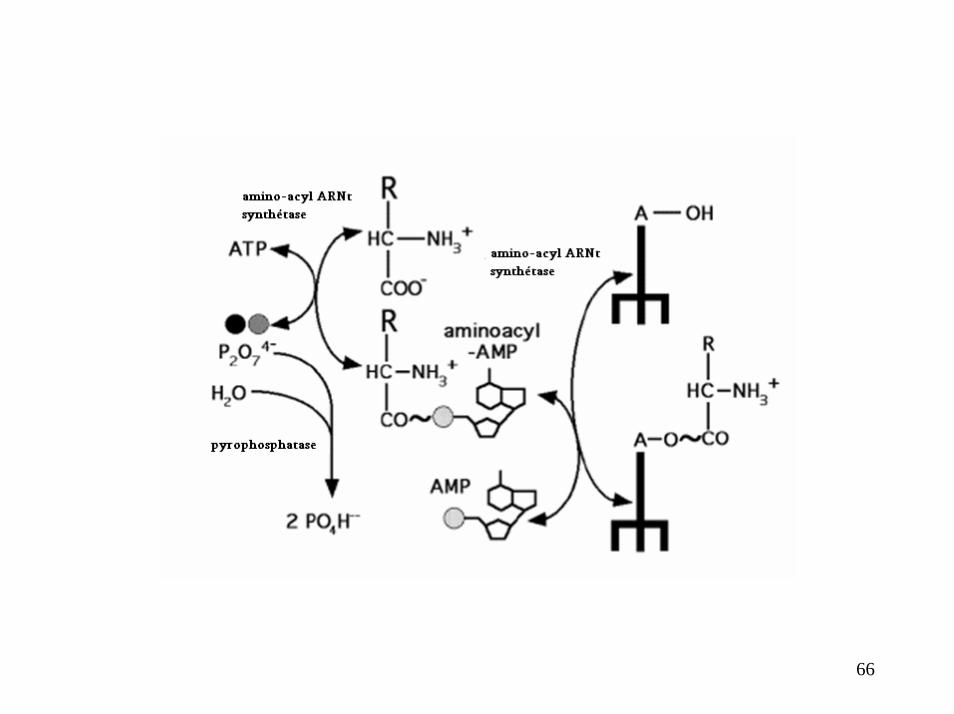
Réactions d’AMINOACYLATION
23
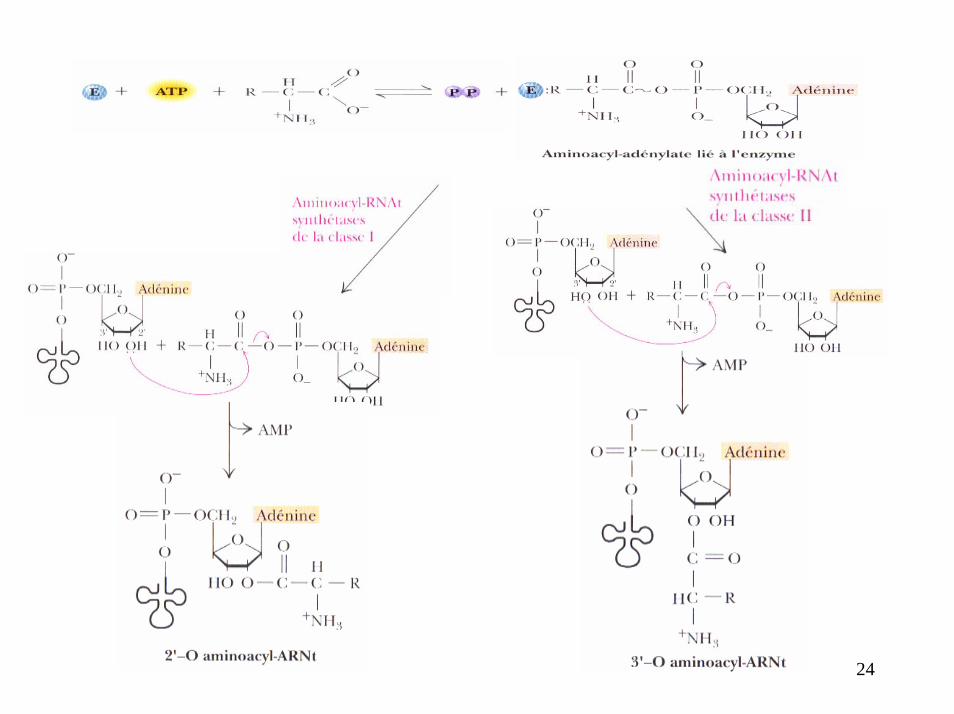
24
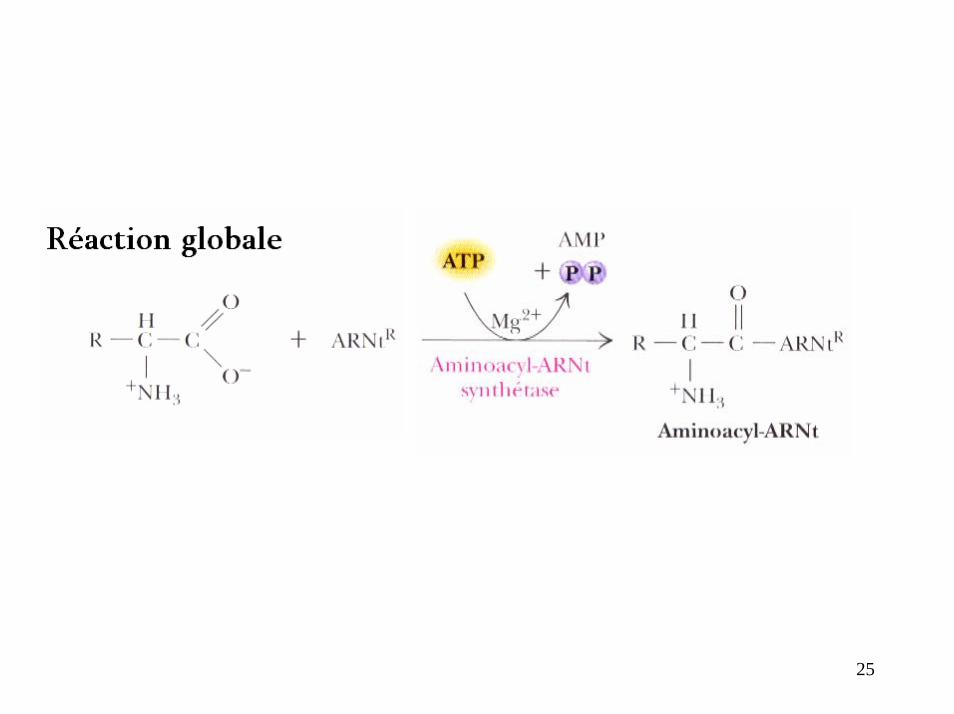
25
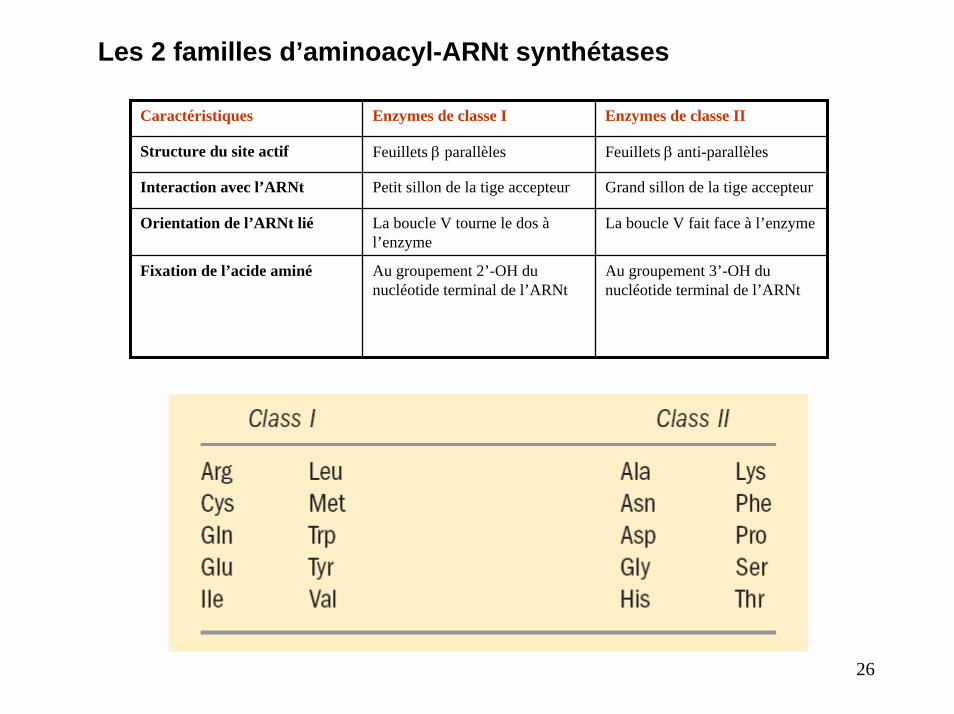
Les 2 familles d’aminoacyl-ARNt synthétases
Caractéristiques Enzymes de classe I Enzymes de classe II
Structure du site actif Feuillets β parallèles Feuillets β anti-parallèles
Interaction avec l’ARNt Petit sillon de la tige accepteur Grand sillon de la tige accepteur
Orientation de l’ARNt lié La boucle V tourne le dos à l’enzyme
La boucle V fait face à l’enzyme
Fixation de l’acide aminé Au groupement 2’-OH du nucléotide terminal de l’ARNt
Au groupement 3’-OH du nucléotide terminal de l’ARNt
26
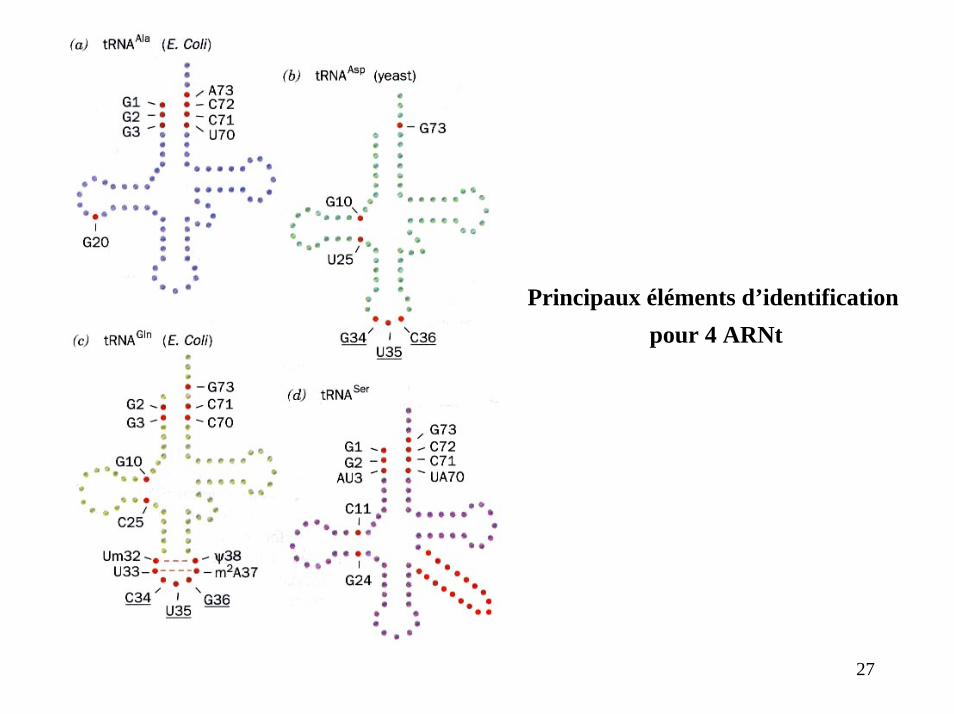
Principaux éléments d’identificationpour 4 ARNt
27

L’aminoacylation doit être exacte : Fonction d’édition des aminoacyl-ARNt synthétases
Les ribosomes ne sélectionnent les aminoacyl-ARNt que grâce aux interactions codon-anticodon, et non en fonction de la nature de leurs groupes aminoacyls.
Une traduction fidèle nécessite donc, non seulement que chaque ARNt soit amino acylé par son aminoacyl ARNt synthétase spécifique mais aussi, qu’il ne soit pas aminoacylé par l’une ou l’autre de ses 19 aminoacyl-ARNt non spécifiques.
28

Fonction d’édition de l’Ile-ARNt synthétase
L’Ile-ARNt synthétase peut activer un certain nombre d’acides aminés de structure similaire mais ne peut attacher ces mauvais acides aminés à l’ARNt
29

Interaction codon-anticodon
30
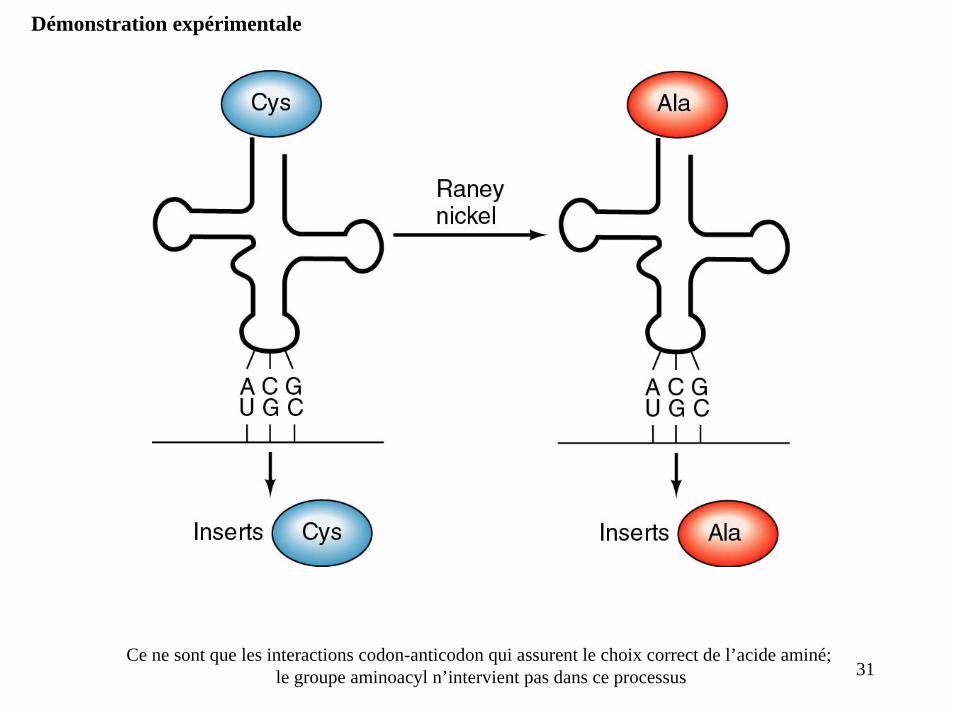
Démonstration expérimentale
Ce ne sont que les interactions codon-anticodon qui assurent le choix correct de l’acide aminé; le groupe aminoacyl n’intervient pas dans ce processus 31
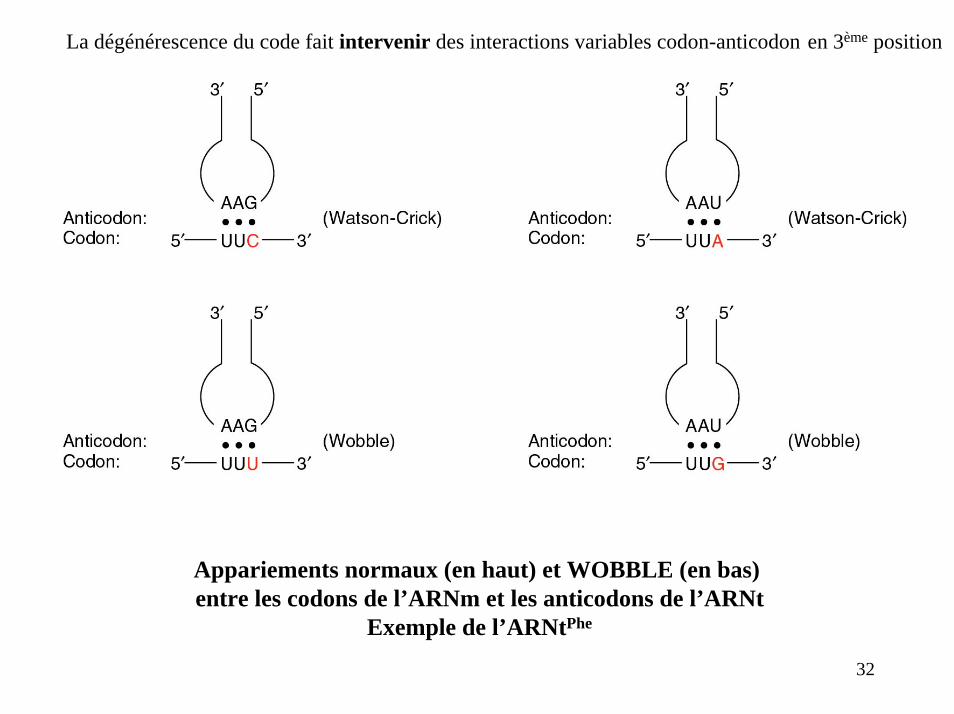
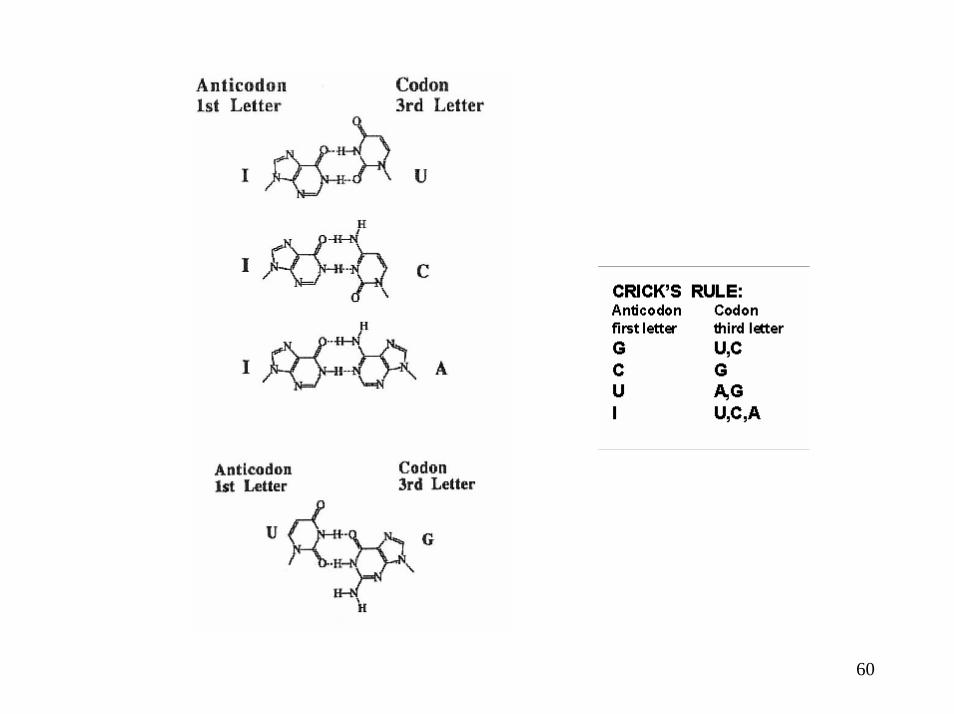
La dégénérescence du code fait intervenir des interactions variables codon-anticodon en 3ème position
Appariements normaux (en haut) et WOBBLE (en bas) entre les codons de l’ARNm et les anticodons de l’ARNt
Exemple de l’ARNtPhe
32
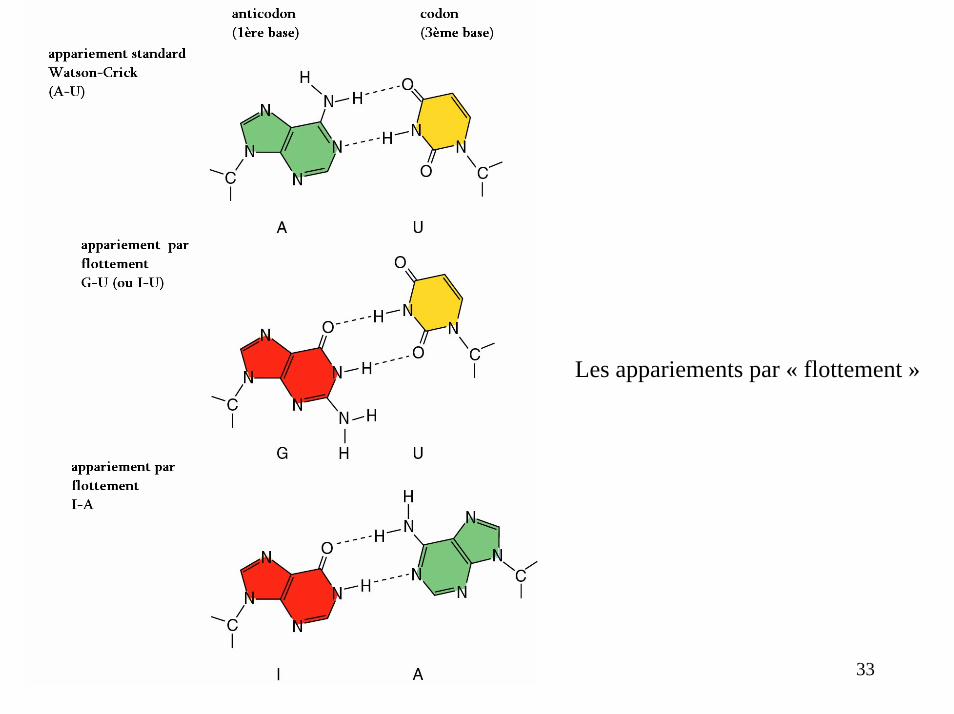
Les appariements par « flottement »
33
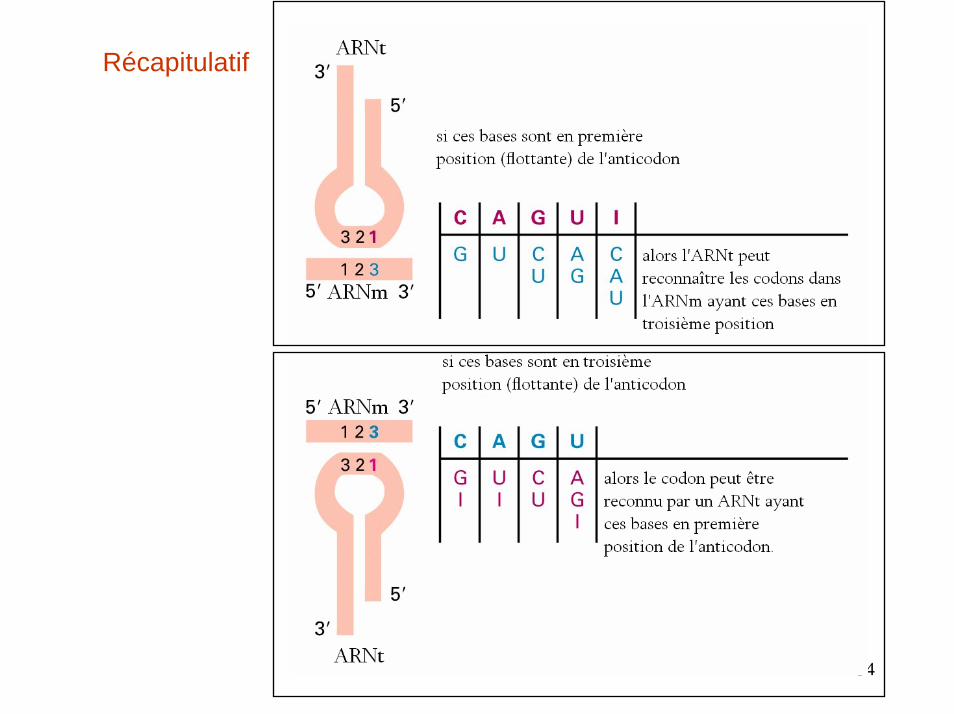
34
Récapitulatif
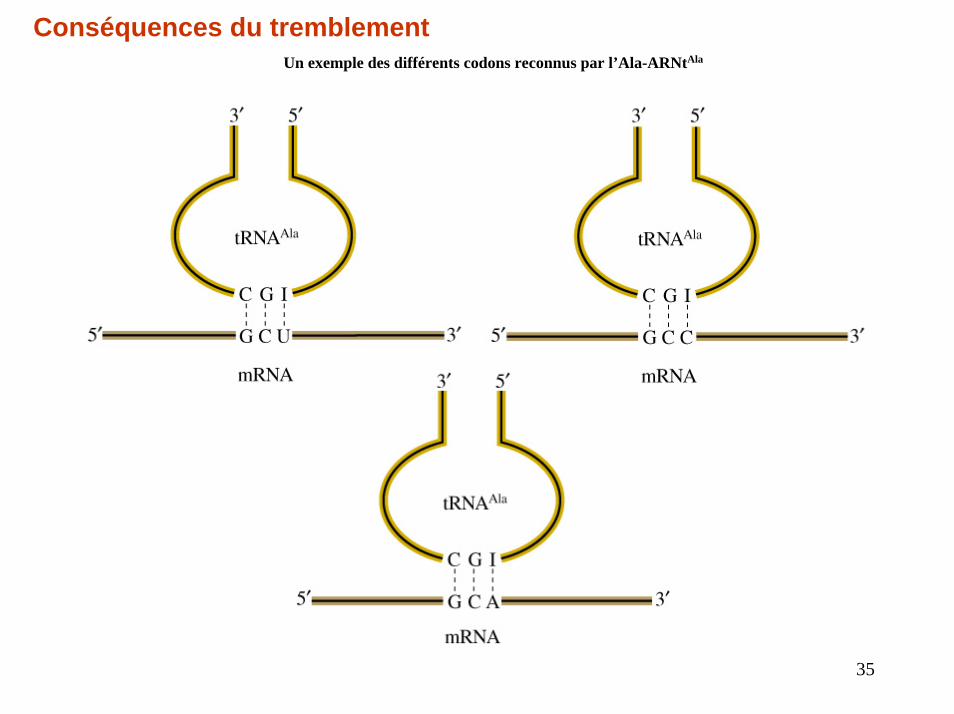
Conséquences du tremblementUn exemple des différents codons reconnus par l’Ala-ARNtAla
35

Principales caractéristiques du code génétique
36
1- plusieurs codons ont une fonction particulièreAUG (ou codon d’initiation) : il signale le commencement d’une chaîne peptidique
(il code aussi pour les résidus Met en position interne des polypeptides)
UAG, UAA & UGA (ou codons de terminaison ou codons stop) : ils indiquent la fin de la synthèse d’une chaîne
Lorsque le cadre de lecture ne présente pas de codon stop pendant 50 codons environ (ou davantage), la région est appelée cadre ouvert de lecture (ORF en anglais : open reading frame) ; de longs cadres ouverts de lecture correspondent en général aux gènes codant des protéines.
2- le code est dégénéréun acide aminé peut être désigné par plus d’un codon
(la différence entre les codons pour un acide aminé donné se situe sur la 3ème base)le code génétique comprend 61 codons codants, on peut donc s’attendre à ce qu’il existe en tout 61 ARNt ;
En fait 32 ARNt suffisent pour « véhiculer » les 20 acides aminés, car un ARNt peut reconnaître plusieurs codons différents codant le même acide aminé.
les 2 premiers nucléotides d’un codon (sur l’ARNm) sont rigoureusement complémentaires de l’anticodon (de l’ARNt) selon la règle d’appariement classique ; il n’en est pas de même pour la 3ème base, un appariement moins classique est possible : il y a alors du jeu (Wobble ou flottement) dans la liaison codon-anticodon qui admet une certaine « liberté stérique ».
- Les liaisons wobble permettent à la cellule de faire des économies- La liaison plus faible entre la 1ère base de l’anticodon et la 3ème base du codon entraîne une dissociation
plus facile des ARNt fixés : la synthèse protéique est ainsi plus rapide.- Elle constitue un système de protection vis à vis des mutations qui peuvent se produire : un changement
de la 3ème base se révèlera le plus souvent sans conséquence puisque le codon muté positionnera le même ARNt
3- le code est non chevauchantla partie exprimée d’un ADN est transcrite puis traduite régulièrement de triplet en triplet.

Ribosomes
37
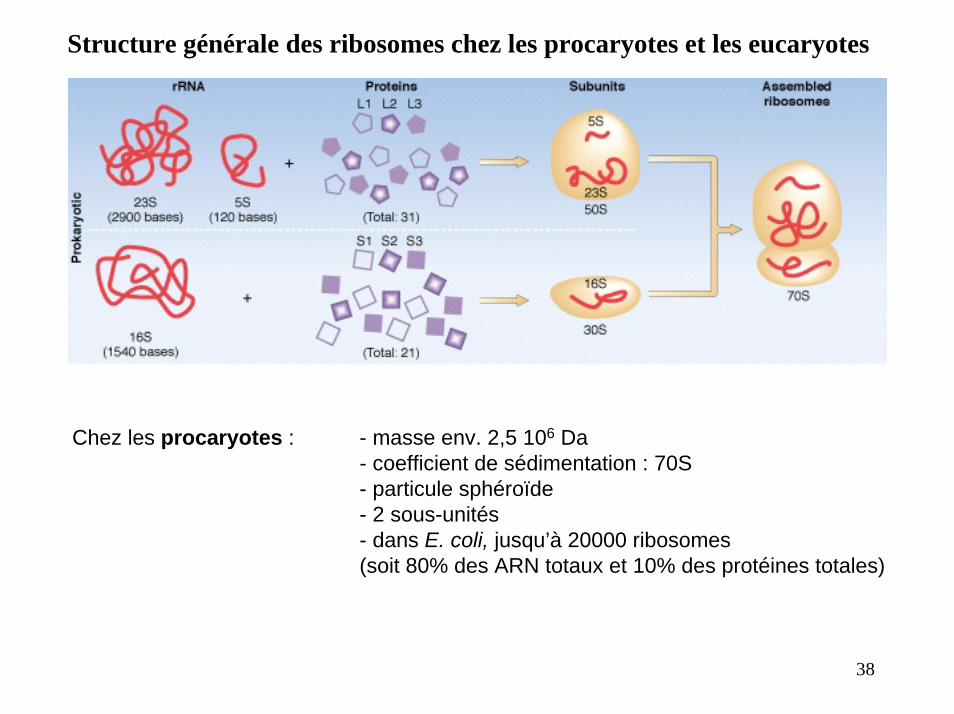
Structure générale des ribosomes chez les procaryotes et les eucaryotes
Chez les procaryotes : - masse env. 2,5 106 Da- coefficient de sédimentation : 70S- particule sphéroïde - 2 sous-unités- dans E. coli, jusqu’à 20000 ribosomes (soit 80% des ARN totaux et 10% des protéines totales)
38
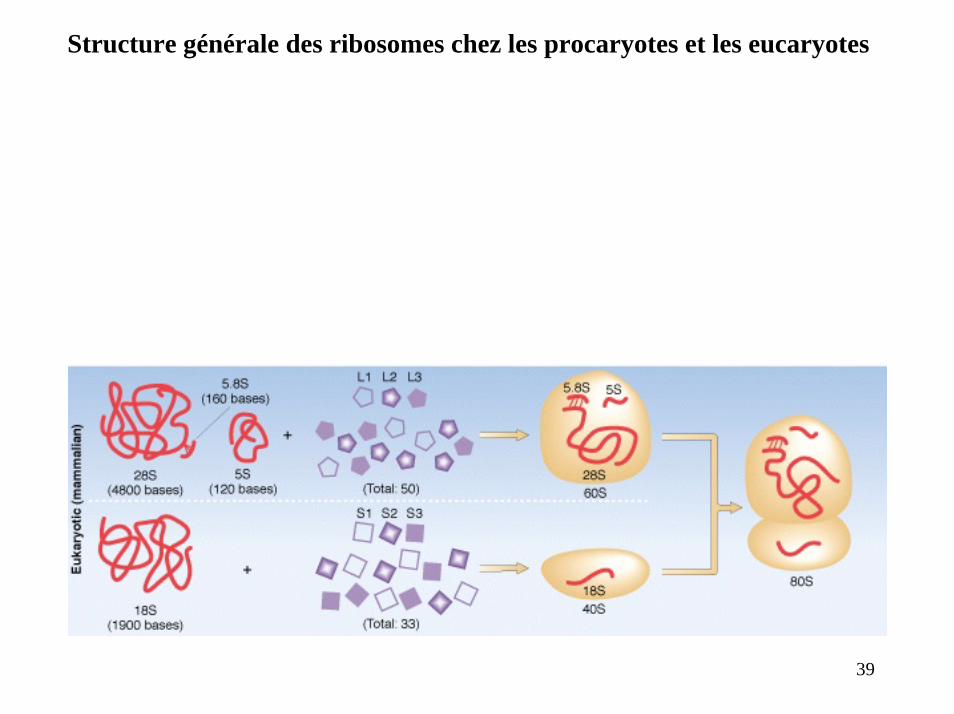
Structure générale des ribosomes chez les procaryotes et les eucaryotes
39
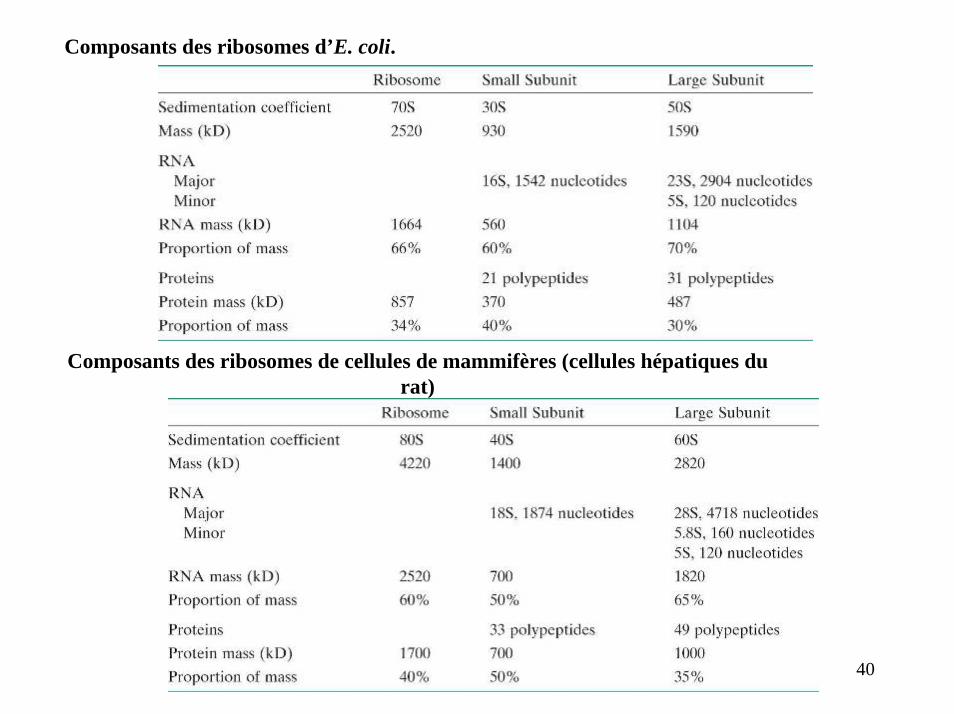
Composants des ribosomes d’E. coli.
Composants des ribosomes de cellules de mammifères (cellules hépatiques durat)
40

Composition des ribosomes
• 2 sous-unités• Petite sous-unité ribosomale
– 16S ARNr (bactéries), 18S ARNr (eucaryotes)– de 21 (bactéries) à 33 (eucaryotes) protéines
• Grande sous-unité ribosomale– 23S & 5S ARNr (bactéries), 28S, 5,8S & 5S ARNr
(eucaryotes)– de 31 (bactéries) à 49 (eucaryotes) protéines
41
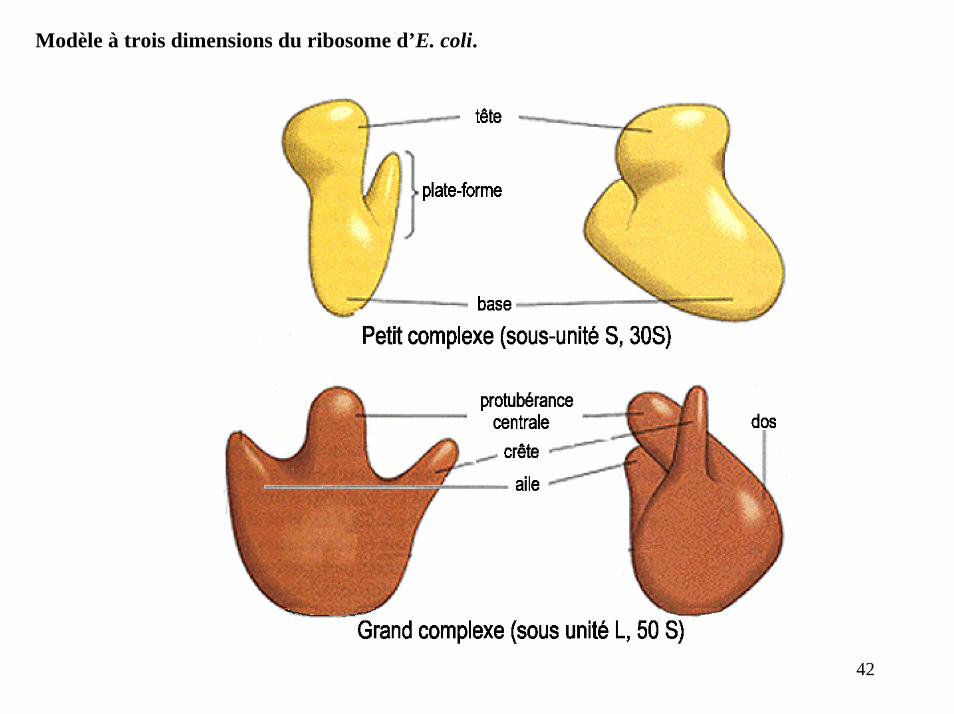
Modèle à trois dimensions du ribosome d’E. coli.
42
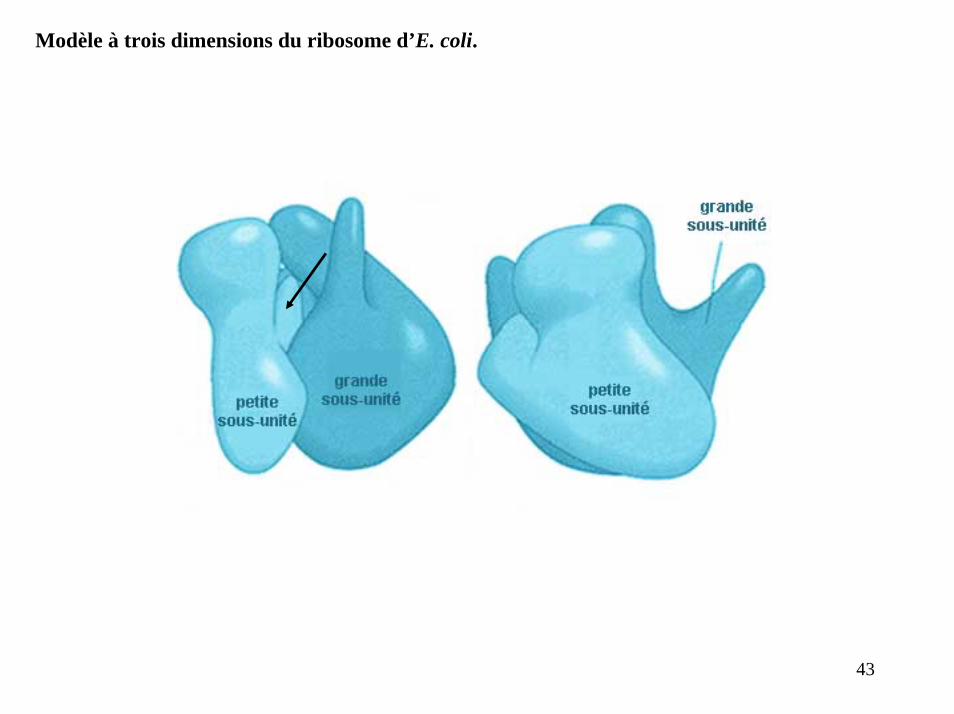
Modèle à trois dimensions du ribosome d’E. coli.
43

44
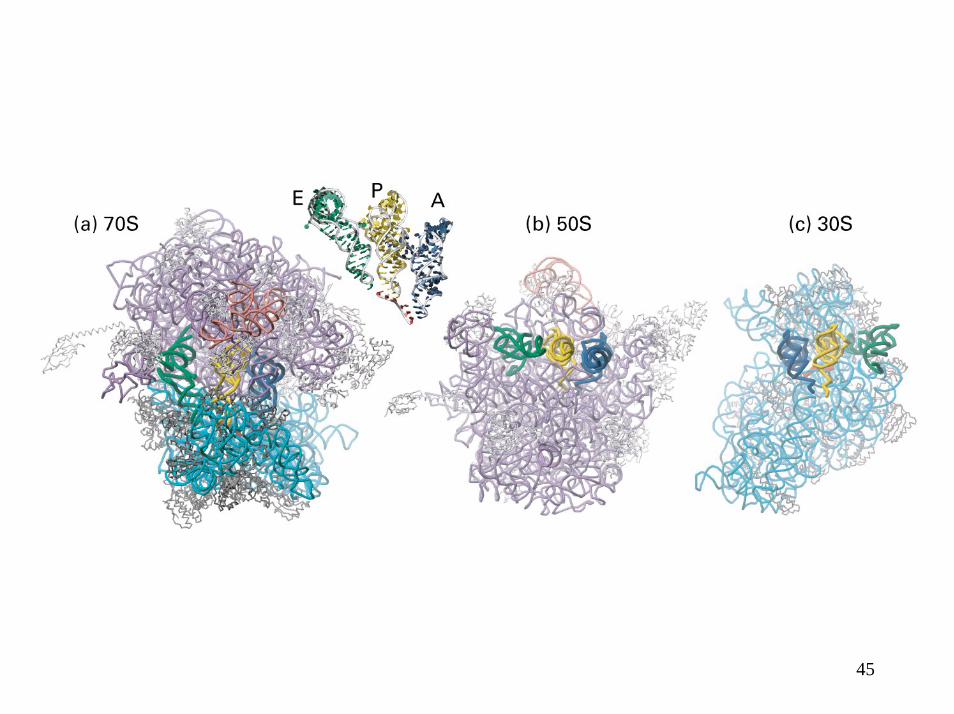
45
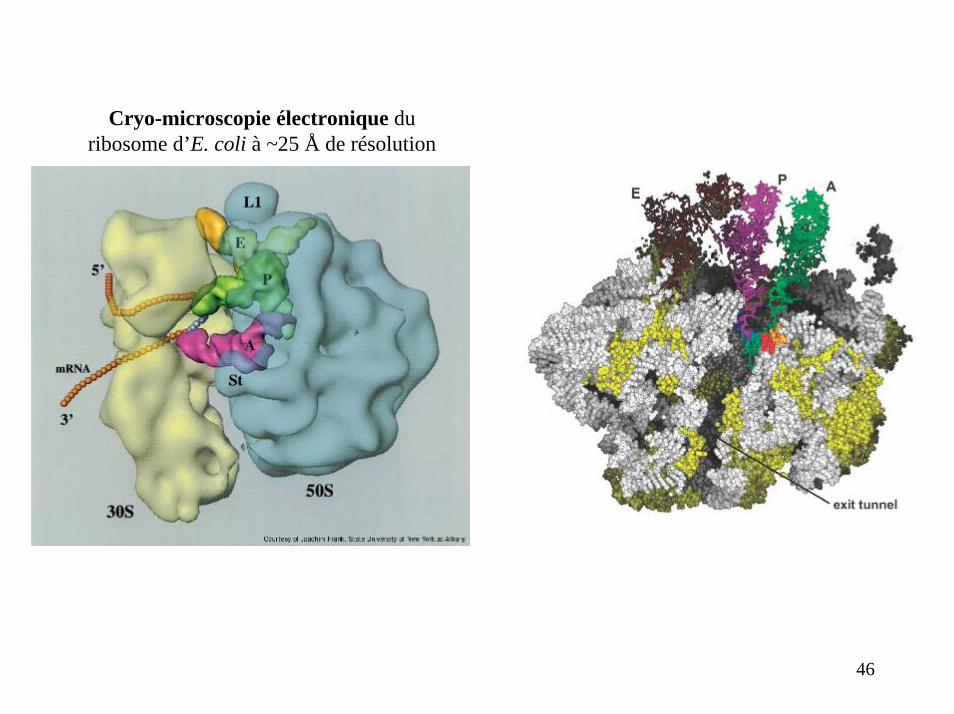
Cryo-microscopie électronique du ribosome d’E. coli à ~25 Å de résolution
46
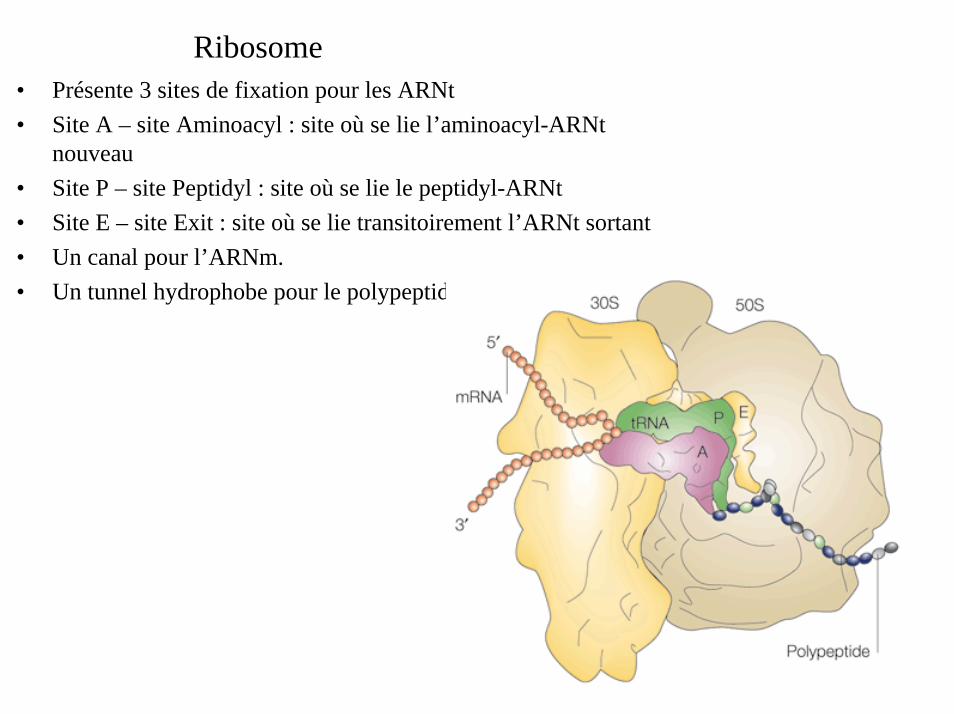
Ribosome• Présente 3 sites de fixation pour les ARNt• Site A – site Aminoacyl : site où se lie l’aminoacyl-ARNt
nouveau• Site P – site Peptidyl : site où se lie le peptidyl-ARNt• Site E – site Exit : site où se lie transitoirement l’ARNt sortant• Un canal pour l’ARNm.• Un tunnel hydrophobe pour le polypeptide.
47

Structure secondaire proposée de l’ARN 16S d’E. coli
48

Assemblage du ribosome
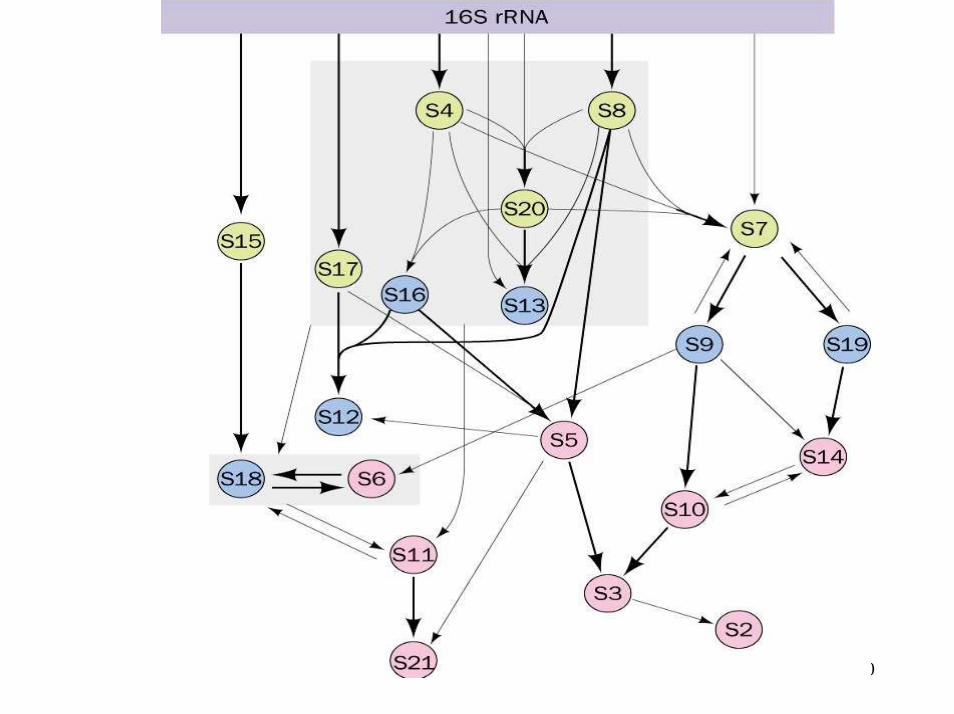
• Auto-assemblage – 1) avec repliement des ARNr– 2) assemblage des protéines (dans un ordre donné)
• Les cellules doivent coordonner l'expression de tous les composants du ribosome proportionnellement au taux de synthèse des protéines
• mécanismes de contrôle
49
50

Ribosomes : récapitulatif• Site de la synthèse des polypeptides
– Abondant : jusqu’à 20000 ribosomes par cellule d’E. coli• Structure énorme & complexe, 2520 kDa (E. coli) 4220 (homme)• De grands ARN et >50 protéines (E. coli), >80 protéines (mammifères)• Fonctions:
– Fixe les ARNm (lecture des codons)– 3 sites de fixation pour les ARNt– Fixe différentes protéines (non-ribosomal proteins) qui ont un rôle dans l’Initiation,
l’Elongation et la Termination– Catalyse la formation de la liaison peptidique– Parcours l’ARNm.
• Le Ribosome est un ribozyme.
51

Synthèse protéique : vue d’ensemble
52
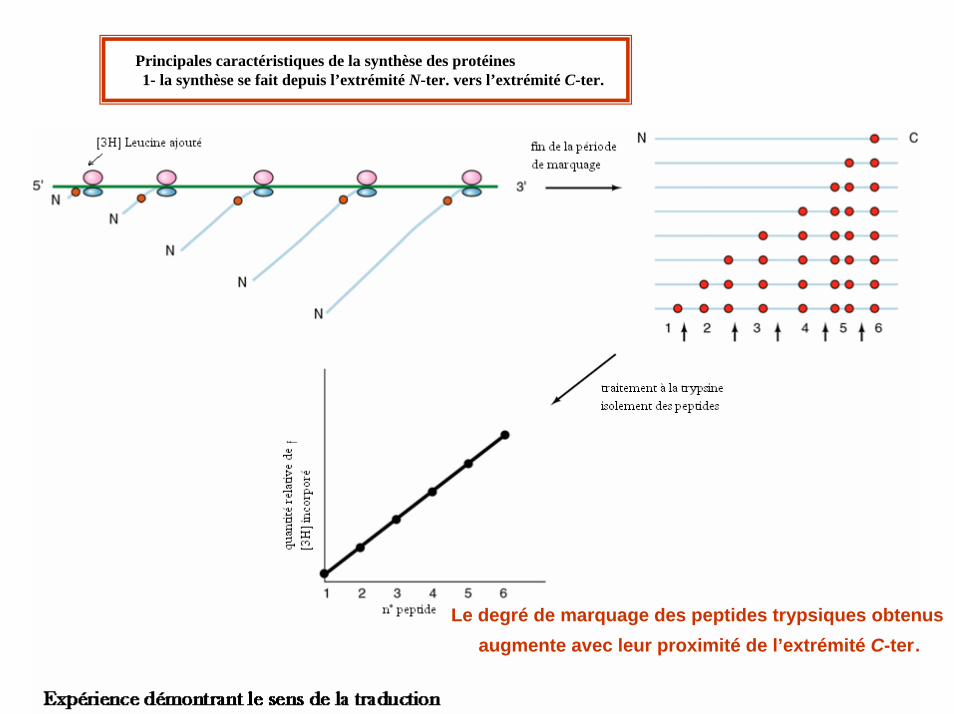
Principales caractéristiques de la synthèse des protéines1- la synthèse se fait depuis l’extrémité N-ter. vers l’extrémité C-ter.
53
Le degré de marquage des peptides trypsiques obtenus augmente avec leur proximité de l’extrémité C-ter.
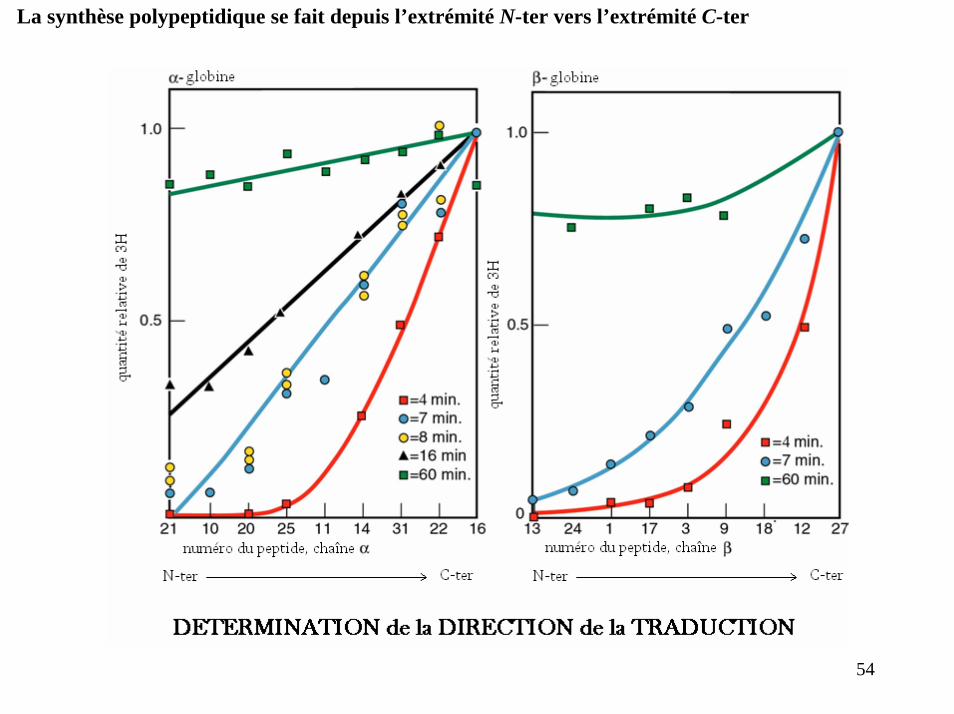
La synthèse polypeptidique se fait depuis l’extrémité N-ter vers l’extrémité C-ter
54

SCHEMA RECAPITULATIF
La synthèse peptidique se fait dans le sens N-ter. C-ter.
55

Principales caractéristiques de la synthèse des protéines2- les ribosomes lisent l’ARNm dans le sens 5’-3’.
Utilisation de systèmes de synthèse protéique acellulaires où l’ARNm est un poly(A) avec un C à son extrémité 3’ :
5’- A A A A A A … …A A A A C - 3’
Synthèse d’un poly(Lys) avec un Asn en C-ter.
Sachant que AAA et AAC codent pour Lys et Asn et que la synthèse se fait dans le sens N-ter C-ter, cela montre que le ribosome lit l’ARNm dans le sens 5’-3’.
56
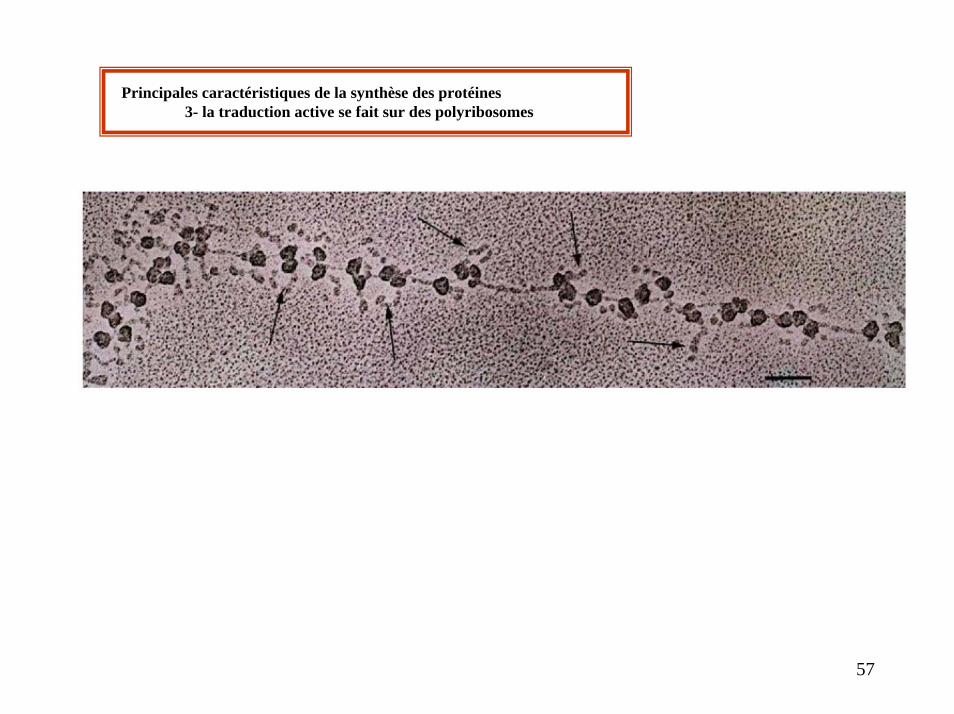
Principales caractéristiques de la synthèse des protéines3- la traduction active se fait sur des polyribosomes
57
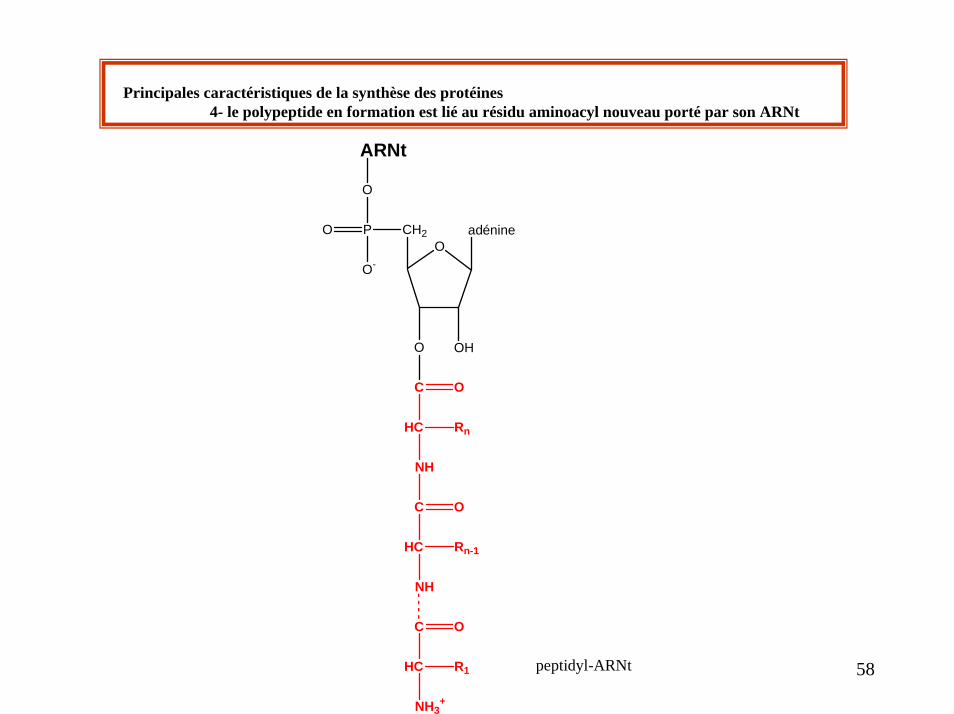
Principales caractéristiques de la synthèse des protéines4- le polypeptide en formation est lié au résidu aminoacyl nouveau porté par son ARNt
Oadénine
OHO
CH2P
O
O
O-
ARNt
C O
HC Rn
NH
C O
HC
NH
Rn-1
C
HC
O
R1
NH3+
peptidyl-ARNt 58

COMPLEMENTS
59
60
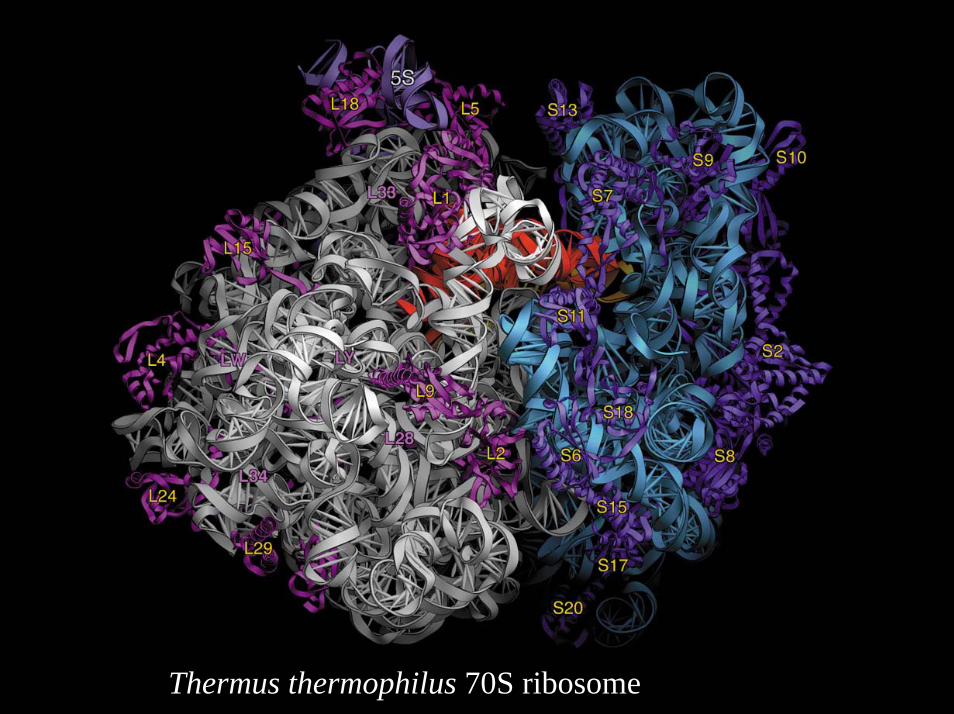
61Thermus thermophilus 70S ribosome
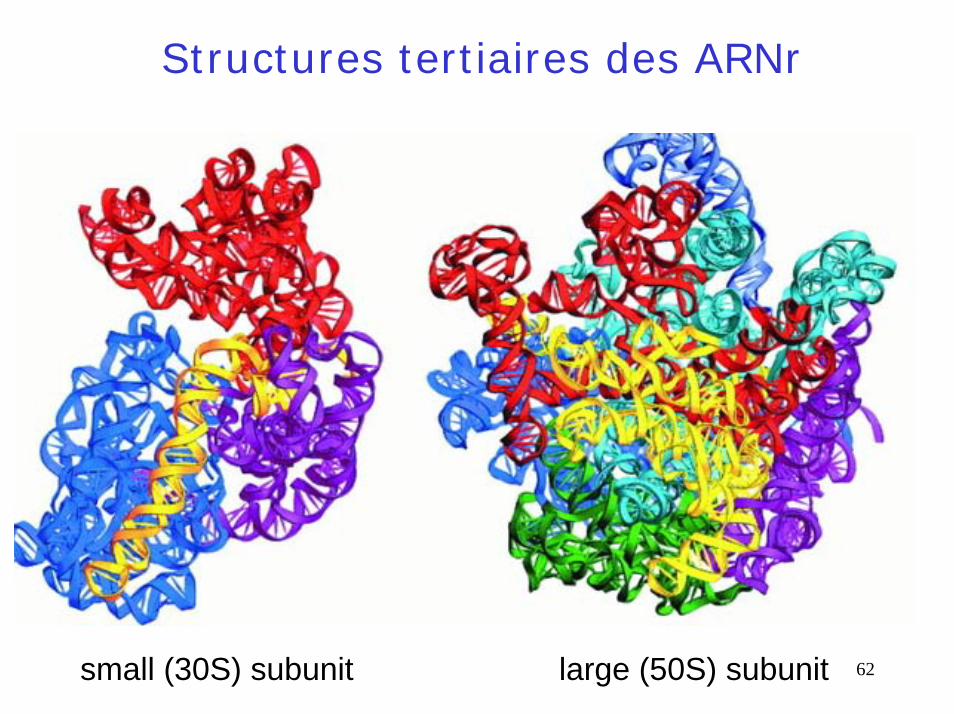
Structures tertiaires des ARNr
small (30S) subunit large (50S) subunit 62
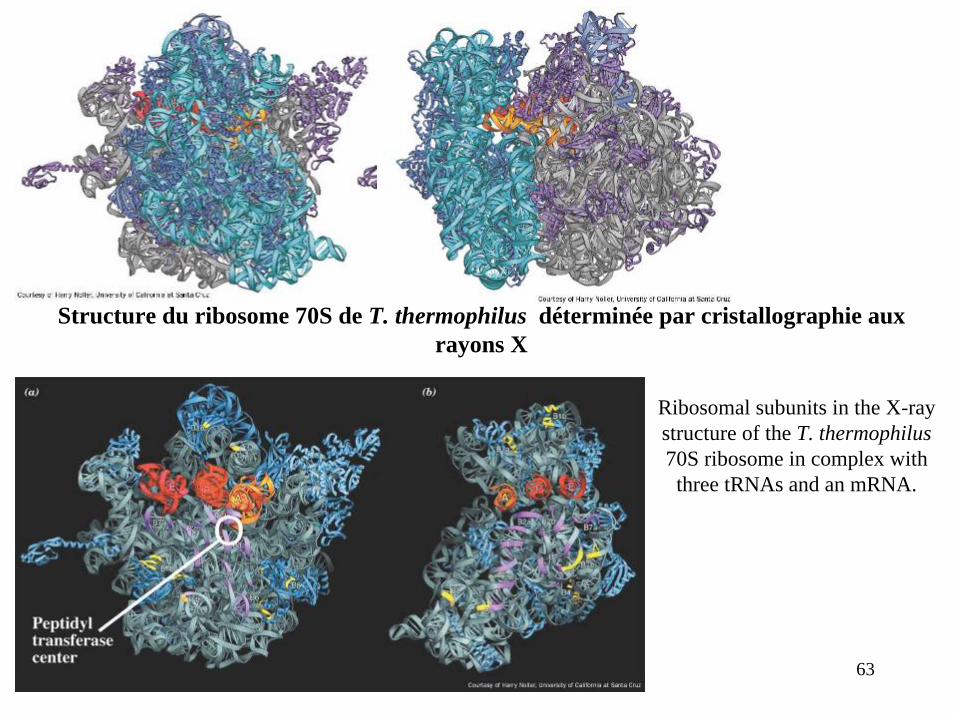
Structure du ribosome 70S de T. thermophilus déterminée par cristallographie aux rayons X
Page
131
5
63
Ribosomal subunits in the X-ray structure of the T. thermophilus70S ribosome in complex with three tRNAs and an mRNA.
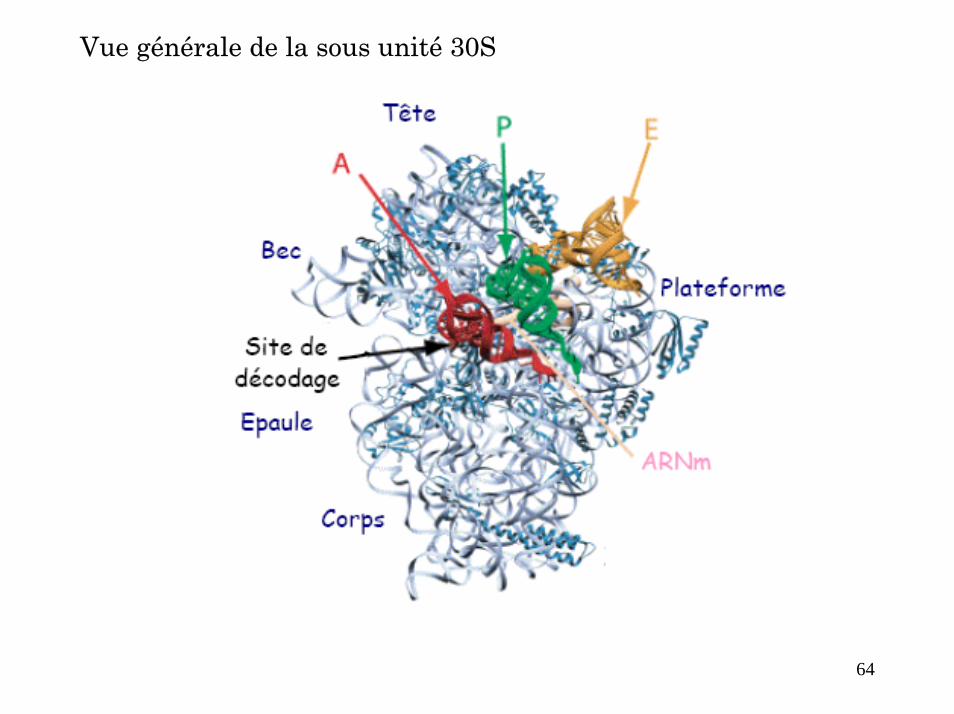
Vue générale de la sous unité 30S
64
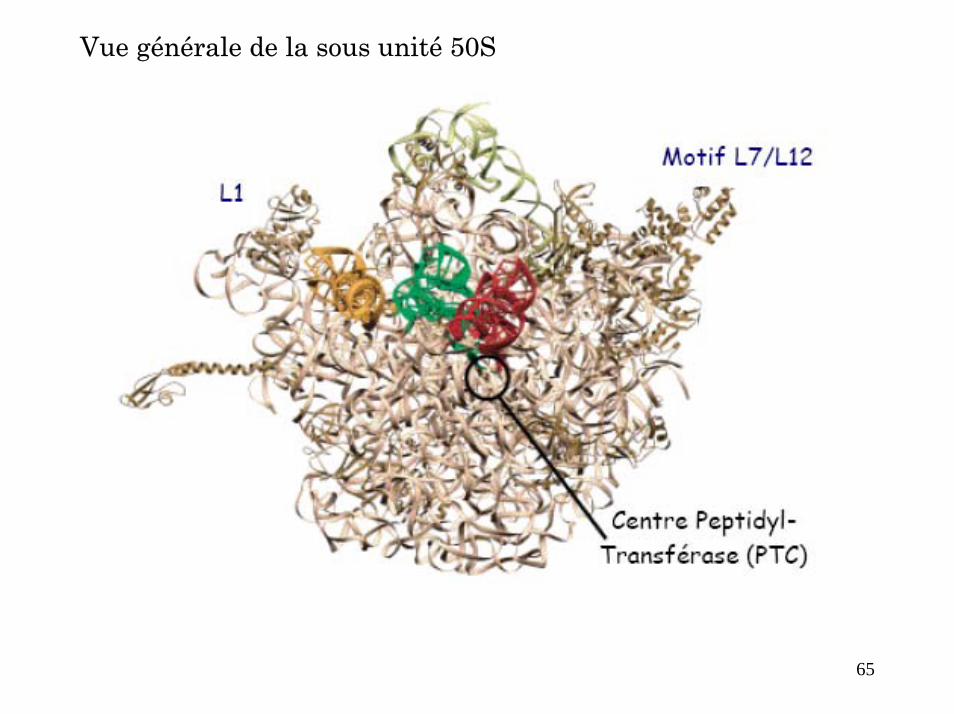
Vue générale de la sous unité 50S
65
66












