Embed Size (px)
DESCRIPTION
La rétine de poisson: un site de plasticité neuronale privilégié. Jean-Pierre Raynauld 6 Octobre 2006 Institut des Neurosciences de Bordeaux. Le héro de cette histoire: Le poisson rouge. Le cahier de charges pour le site de plasticité idéal: 1-situation physiologique. - PowerPoint PPT Presentation
Citation preview

La rétine de poisson: un site de plasticité neuronale privilégié.
Jean-Pierre Raynauld 6 Octobre 2006Institut des Neurosciences de Bordeaux

Le héro de cette histoire: Le poisson rouge

Le cahier de charges pour le site de plasticité idéal:
1-situation physiologique.
2-observation des plusieurs synapses ayant la même fonction.
3- une plasticité facilement observable dans ses aspects électrophysiologiques, anatomiques et biochimiques.
4-existence de contrôles.

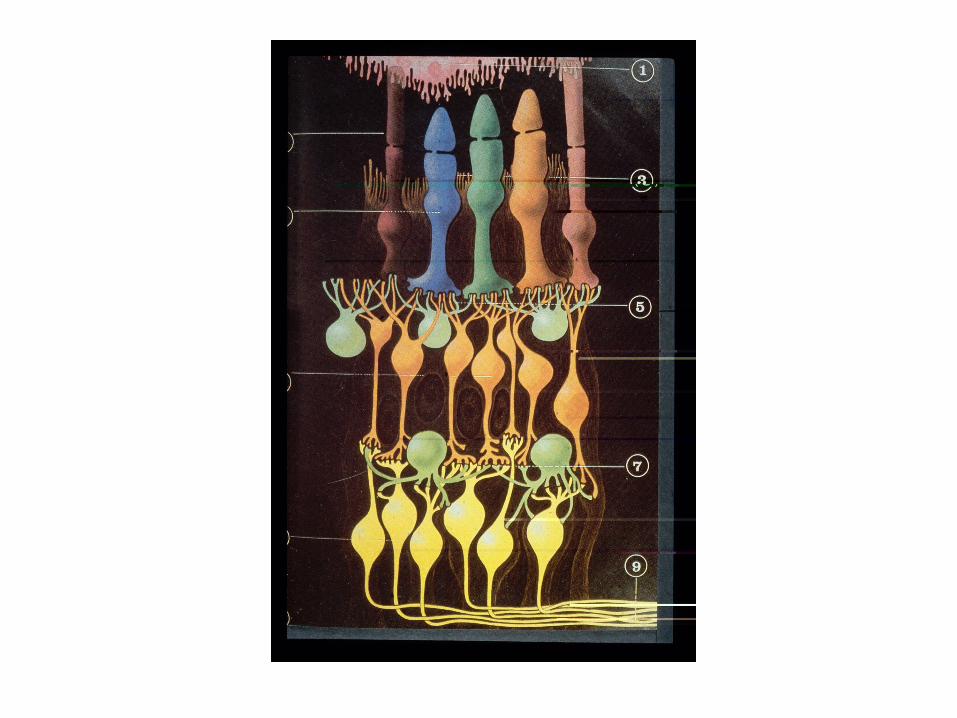
La rétine selon Cajal
Un important point à noter est la stratification des connexions synaptiques dans deux couches plexiformes. Le plexiforme externe étant plus mince que le plexiforme interne.

La rétine de poisson rouge: gracieuseté de Valentin Parthe
IVIC, Caracas, Venezuela
(mouvements mécaniques des segments externes lors de l’adaptation)
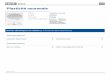
Une donnée spectaculaire dans l’électrophysiologie de la rétine de poisson.
Gunnar Svaetichin, IVIC, Caracas, Venezuela (1958)
Enrégistrements dans des cellules horizontales.
L’existence de cellules antagonistes-couleurs
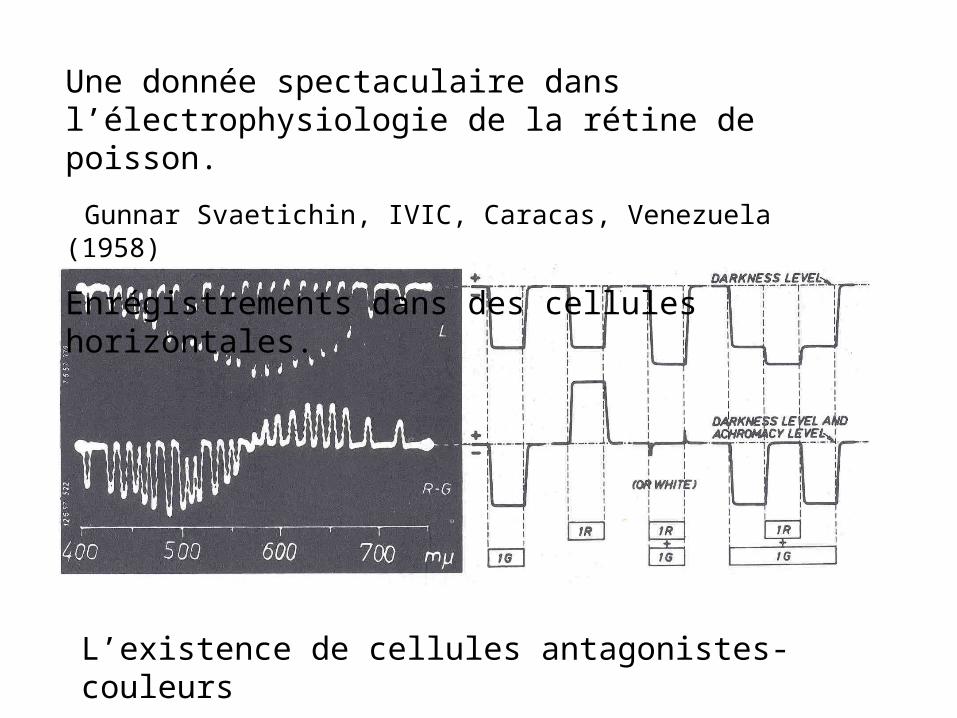
Cellule ganglionnaire: EF McNichol et al, (1960)
Cellule ganglionaire également antagoniste « couleurs »

Dans une rétine duplex, il n’y a pas de cellules ganglionnaires uniquement connectées aux bâtonnets.
Toutes ont des inputs des cônes et des bâtonnets.

Si une telle cellule doublement opposée aux couleurs possédait un input des bâtonnets, une série de questions se posaient:
1- l’input des bâtonnets avait-il le même effet que celui des cônes rouges?
2- l’input des bâtonnets avait-il le même effet que celui des cônes verts?
3-l’effet des bâtonnets était-il similaire d’une cellule ganglionnaire à l’autre?
NB- Ces questions attendaient une réponse depuis 10 ans!
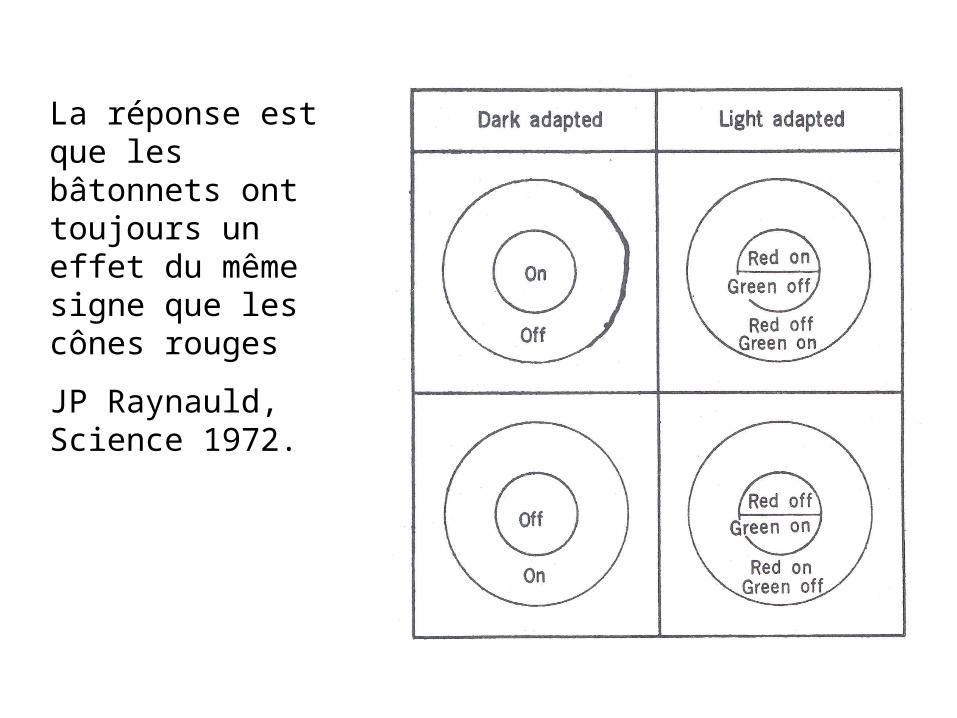
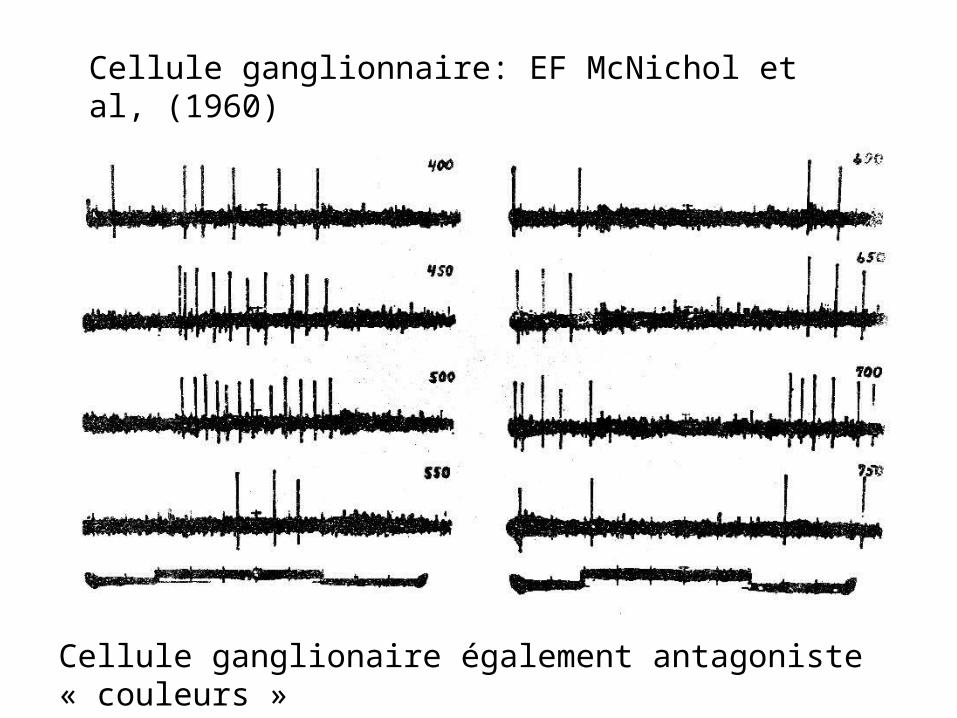
La réponse est que les bâtonnets ont toujours un effet du même signe que les cônes rouges
JP Raynauld, Science 1972.
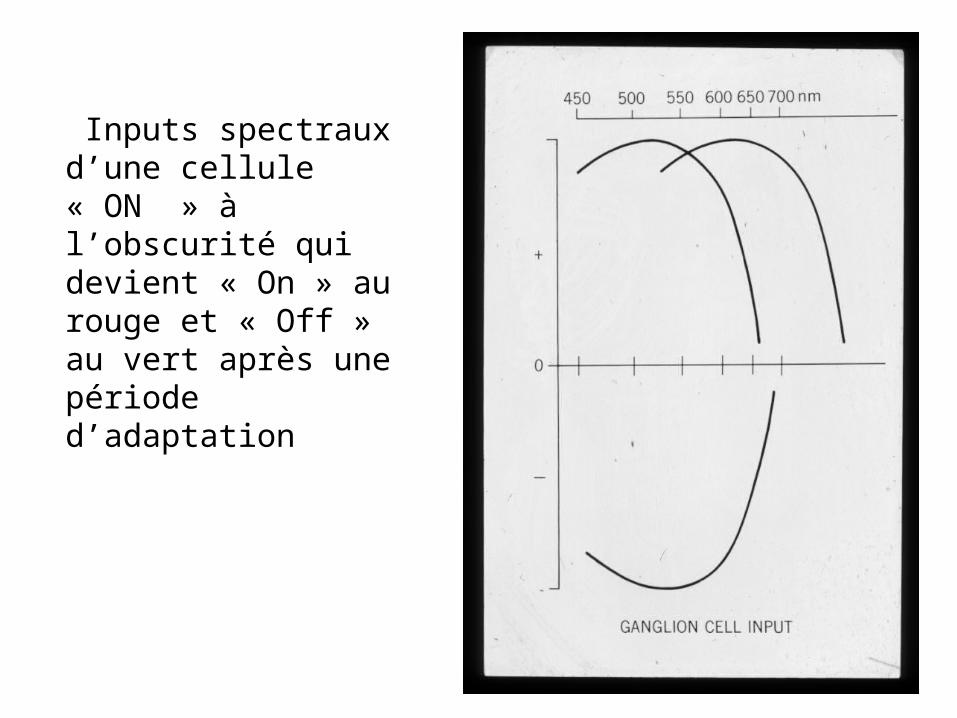
Inputs spectraux d’une cellule « ON » à l’obscurité qui devient « On » au rouge et « Off » au vert après une période d’adaptation
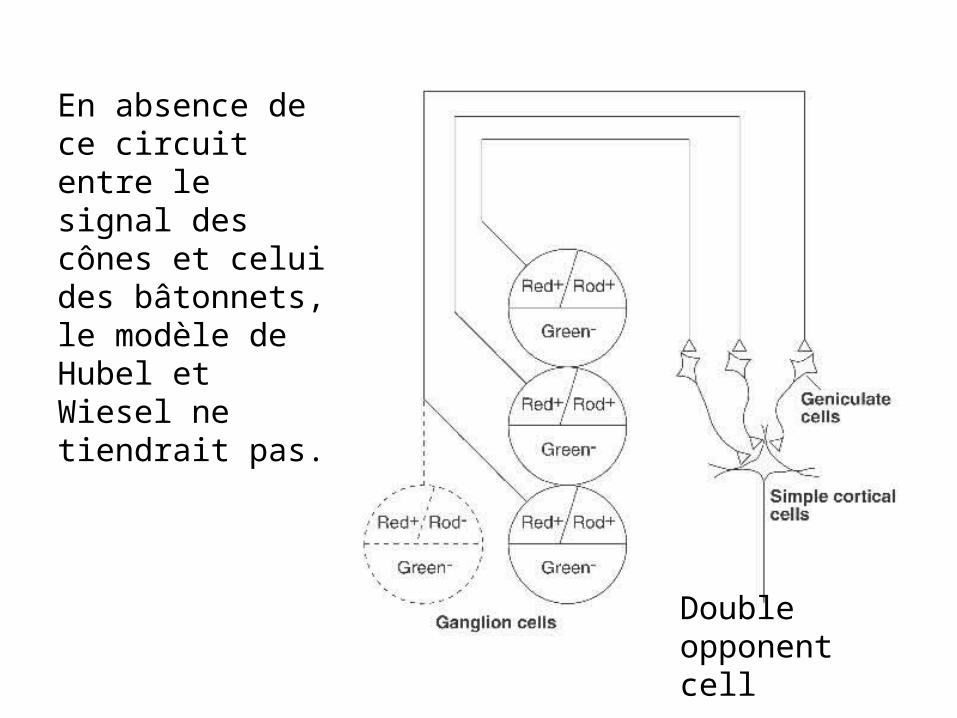
En absence de ce circuit entre le signal des cônes et celui des bâtonnets, le modèle de Hubel et Wiesel ne tiendrait pas.
Double opponent cell
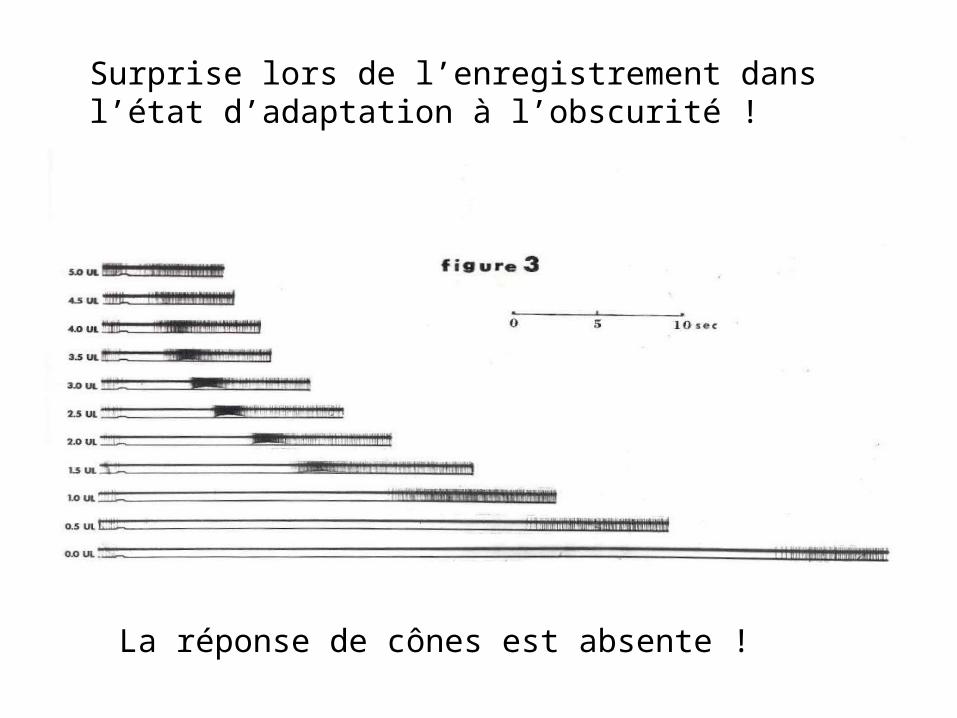
Surprise lors de l’enregistrement dans l’état d’adaptation à l’obscurité !
La réponse de cônes est absente !

En 1973 H.J. Wagner publiait dans la revue Nature une lettre faisant part de la disparition des rubans synaptiques la nuit dans les pédicules de cônes chez Nannacara Anomala.
Nature (London) New Biology 246,53 (1973).
Lors d’une collaboration initiale, il a semblé que le même phénomène était observable chez le poisson rouge.
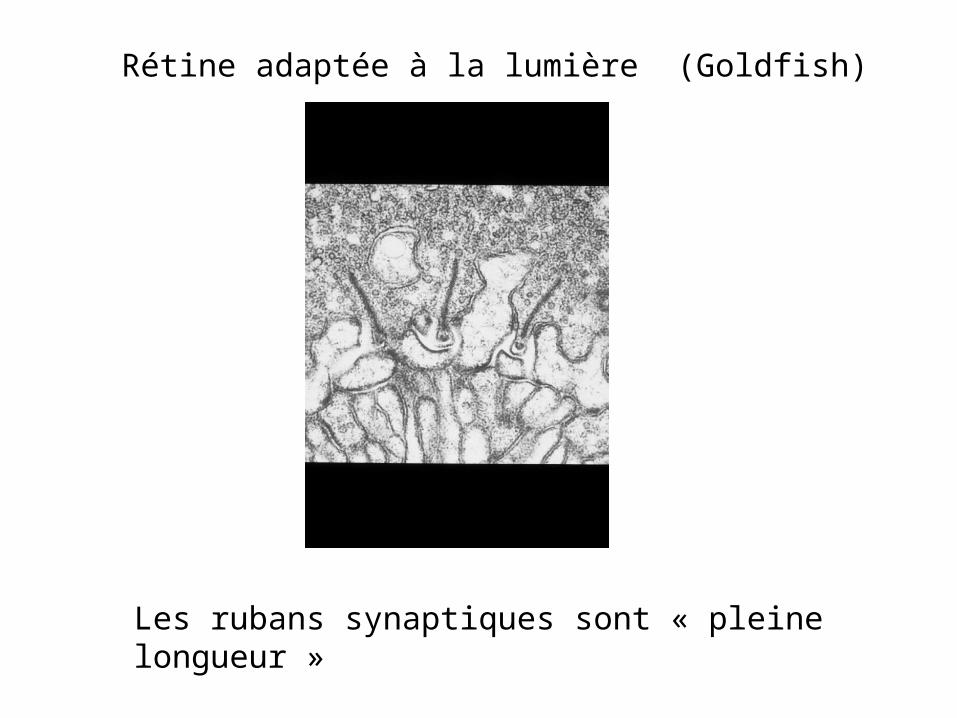
Rétine adaptée à la lumière (Goldfish)
Les rubans synaptiques sont « pleine longueur »
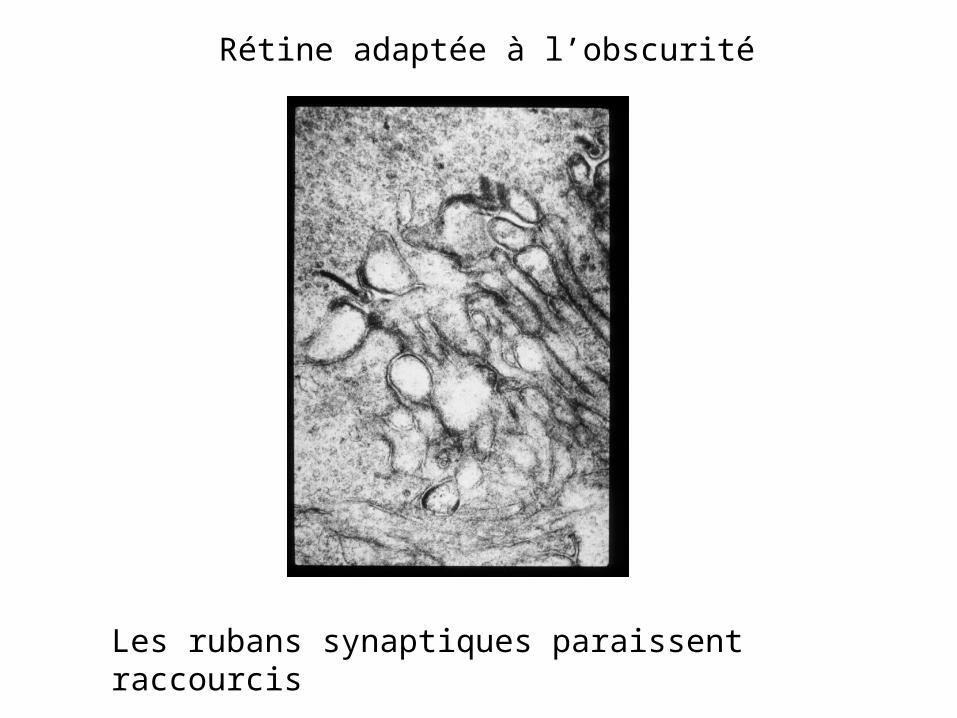
Rétine adaptée à l’obscurité
Les rubans synaptiques paraissent raccourcis

Il fut alors décider d’observer le comportement des cellules ganglionnaires lors de l’adaptation à l’obscurité et de tenter d’isoler les rétines dans l’état d’adaptation à la lumière.
A cause des mouvements mécaniques des cônes et des bâtonnets lors de l’adaptation, si les segments externes des bâtonnets étaient coupés et seuls les cônes seraient fonctionnels.
Les résultats furent surprenant.

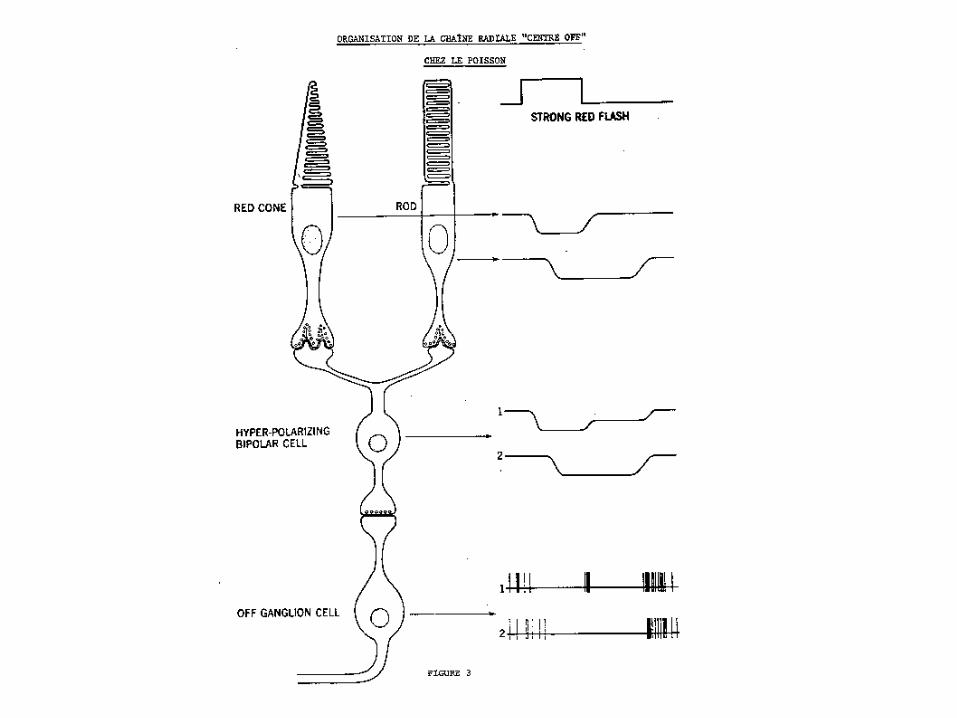
Cellule ganglionnaire « Off » au rouge et « On » au vert perdant son antagonisme couleur suite à une période d’une heure d’adaptation à l’obscurité. Cependant le signal provient toujours des cônes.

La longue latence de la réponse « Off » indique la présence d’un input des bâtonnets chez cette cellule.
Notez que la polarité « Off » est la même que celle produite par les cônes rouges.
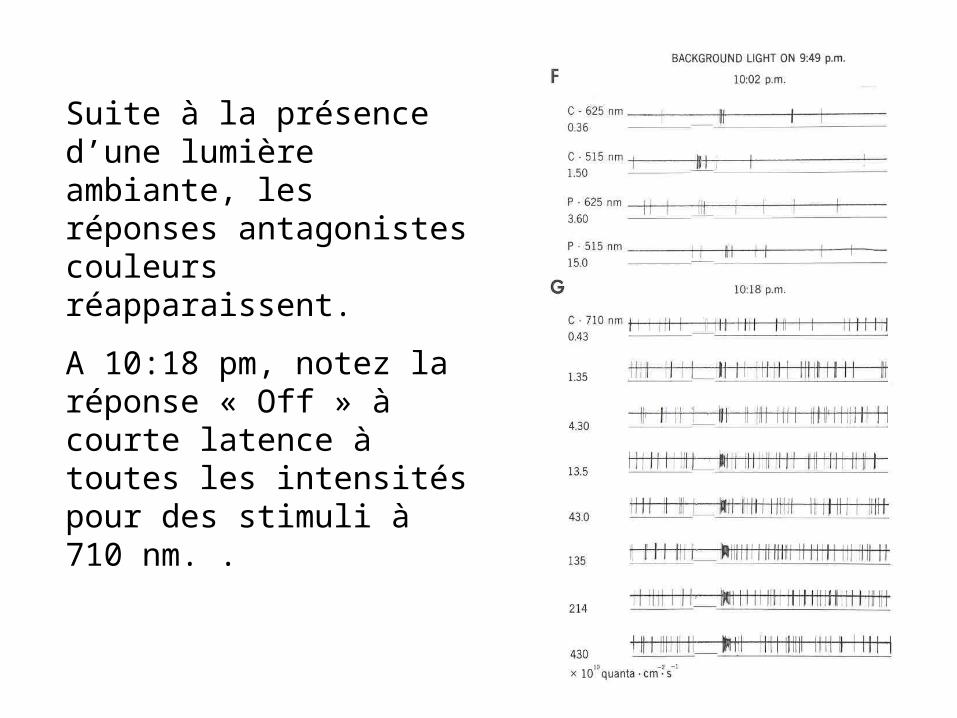
Suite à la présence d’une lumière ambiante, les réponses antagonistes couleurs réapparaissent.
A 10:18 pm, notez la réponse « Off » à courte latence à toutes les intensités pour des stimuli à 710 nm. .

Lors de ma formation en biophysique à l’Université Johns Hopkins, j’avais fait un court projet en microscopie électronique. J’ai donc tenté de reproduire les résultats de Wagner, i.e. la disparition des rubans synaptiques de cônes la nuit. Résultats négatifs.
Lors d’une collaboration avec Wagner et Laviollette, nous avons produit les résultats suivant:
Raynauld et al. Science 1979
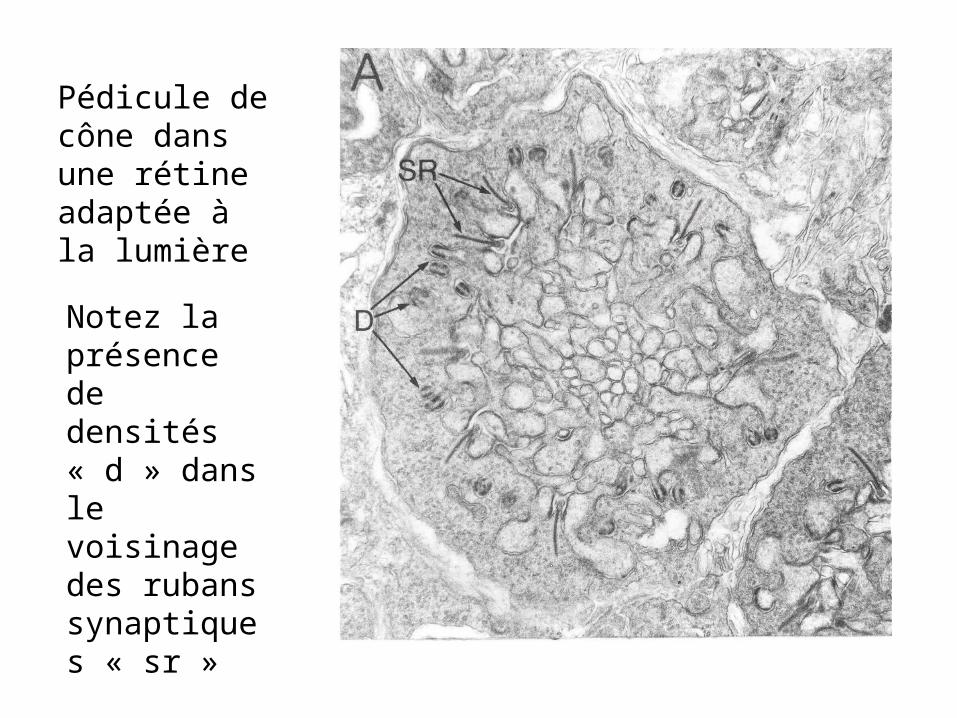
Pédicule de cône dans une rétine adaptée à la lumière
Notez la présence de densités « d » dans le voisinage des rubans synaptiques « sr »
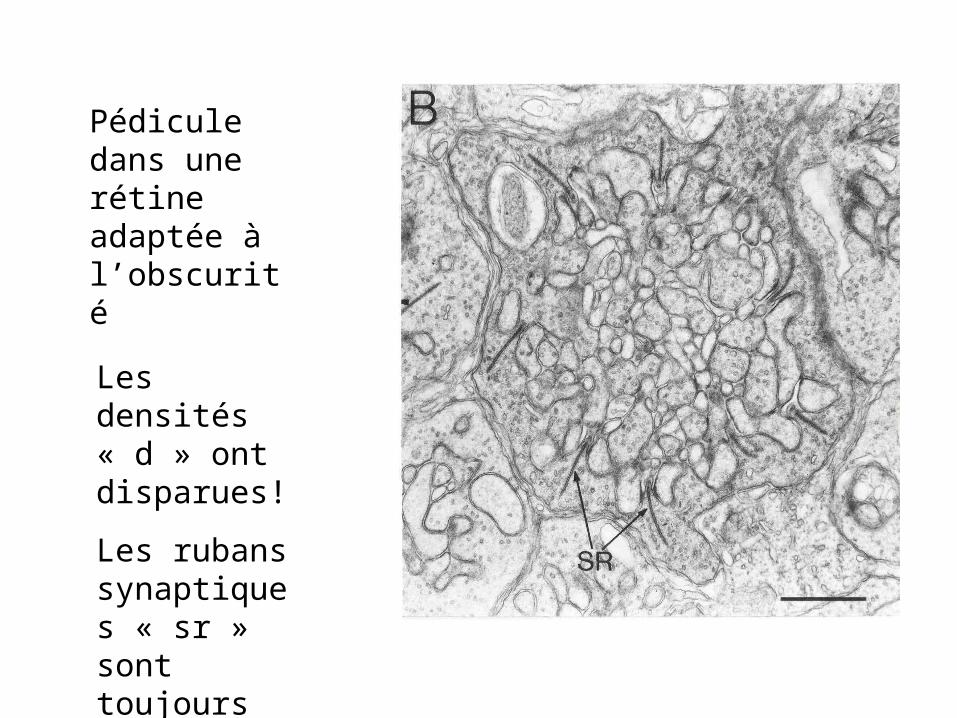
Pédicule dans une rétine adaptée à l’obscurité
Les densités « d » ont disparues!
Les rubans synaptiques « sr » sont toujours là.

Étant donné que les modifications anatomiques se produisaient dans les cellules horizontales, il fallait faire des enregistrements de longue durée dans ce type de cellules.
Lors d’une année sabbatique, je suis donc allé chez Gordon Fain au Jules Stein Eye Institute à UCLA pour faire ces enregistrements.
Dans cette préparation, la stabilité n’est pas très bonne et il est difficile d’enregistrer sur une période supérieure à 20 minutes.
Nous avons cependant obtenu un enregistrement exceptionnel:
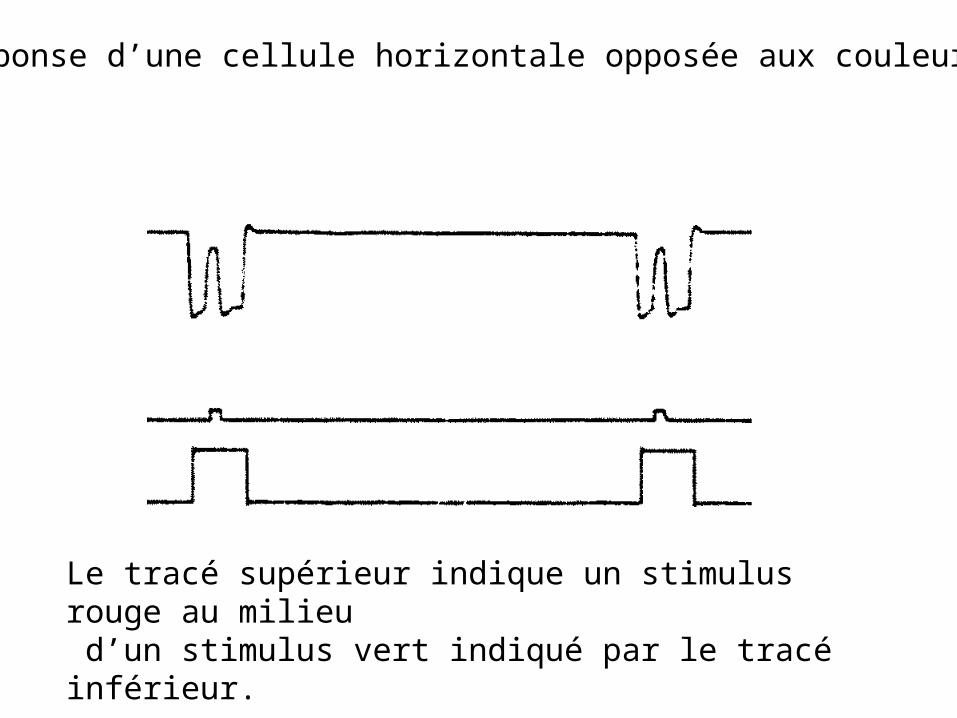
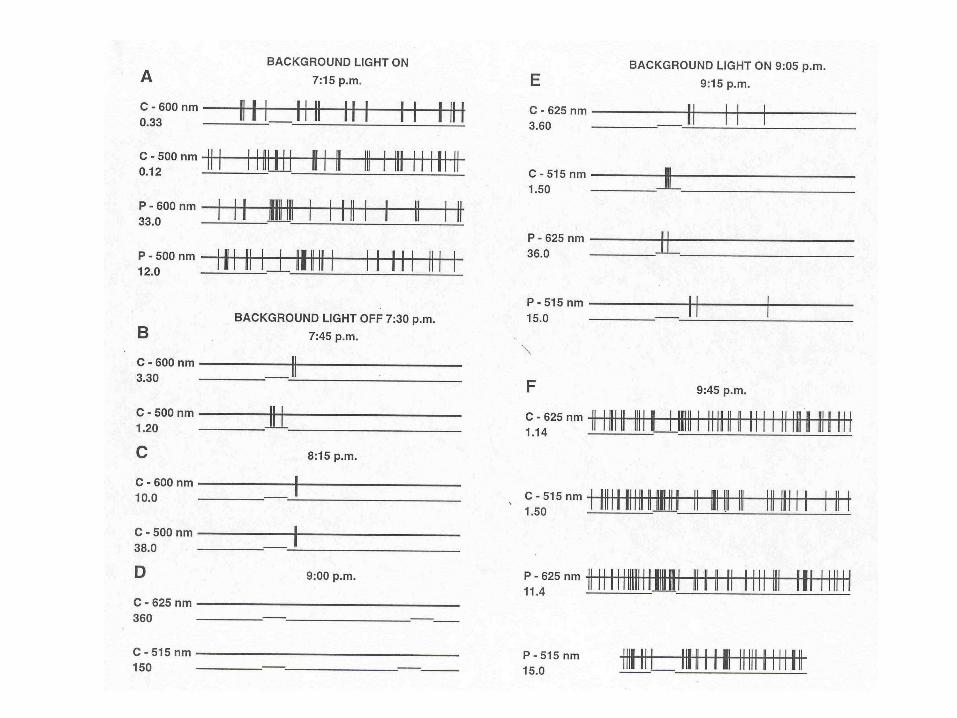
Réponse d’une cellule horizontale opposée aux couleurs.
Le tracé supérieur indique un stimulus rouge au milieu d’un stimulus vert indiqué par le tracé inférieur.
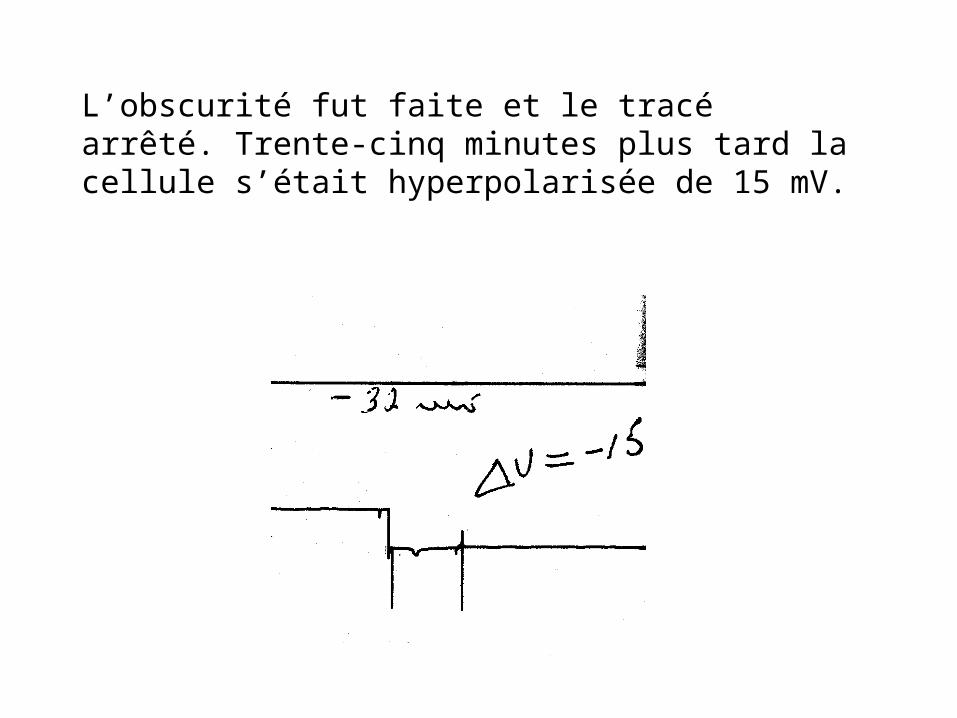
L’obscurité fut faite et le tracé arrêté. Trente-cinq minutes plus tard la cellule s’était hyperpolarisée de 15 mV.
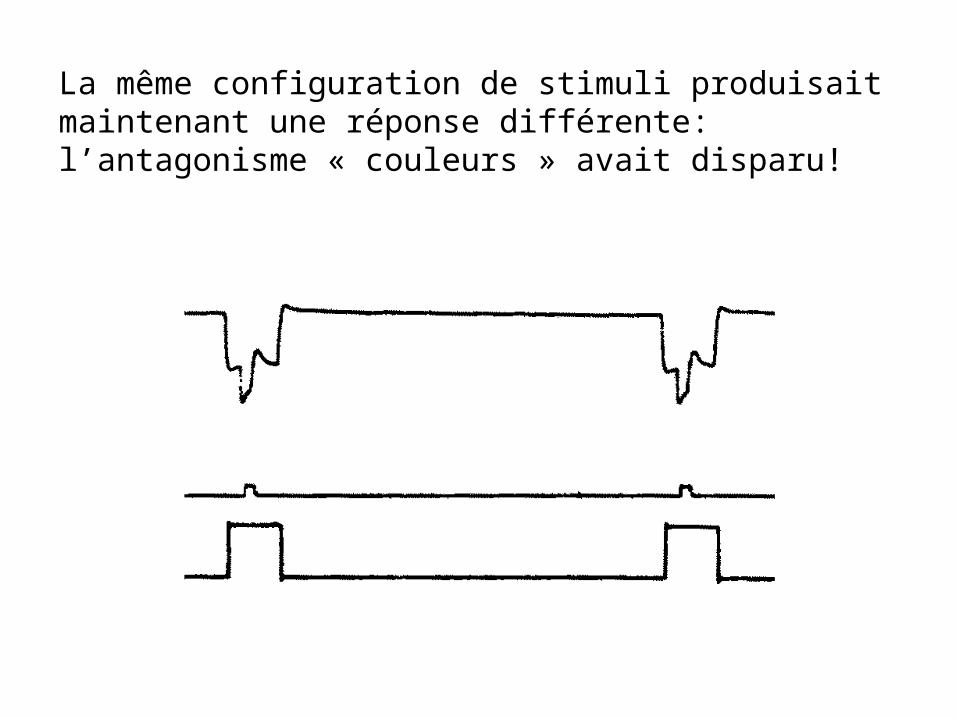
La même configuration de stimuli produisait maintenant une réponse différente: l’antagonisme « couleurs » avait disparu!
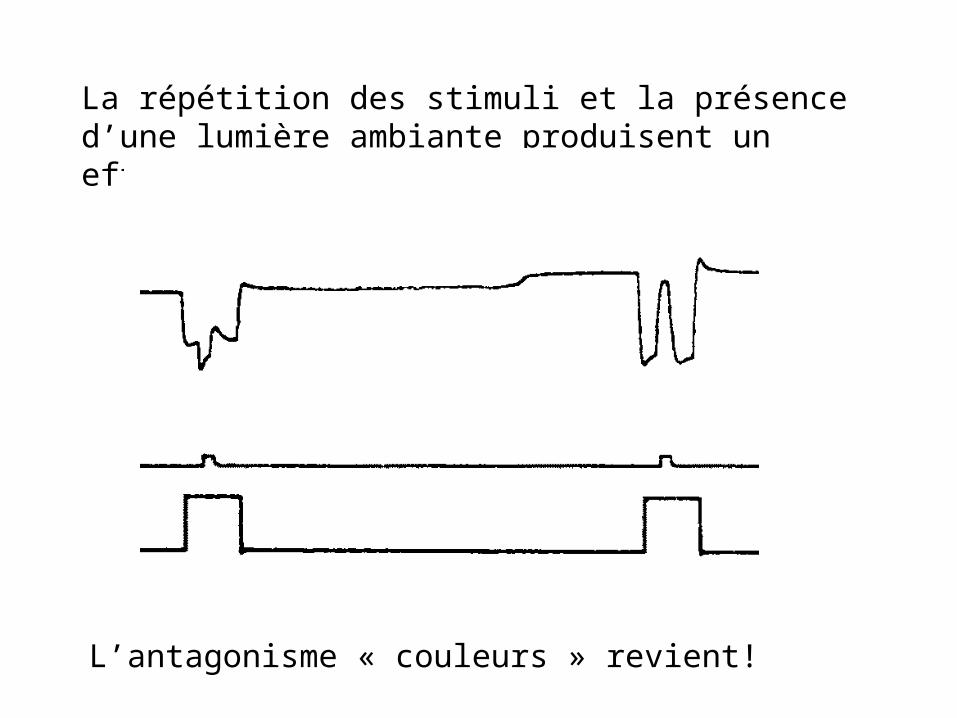
La répétition des stimuli et la présence d’une lumière ambiante produisent un effet dramatique
L’antagonisme « couleurs » revient!
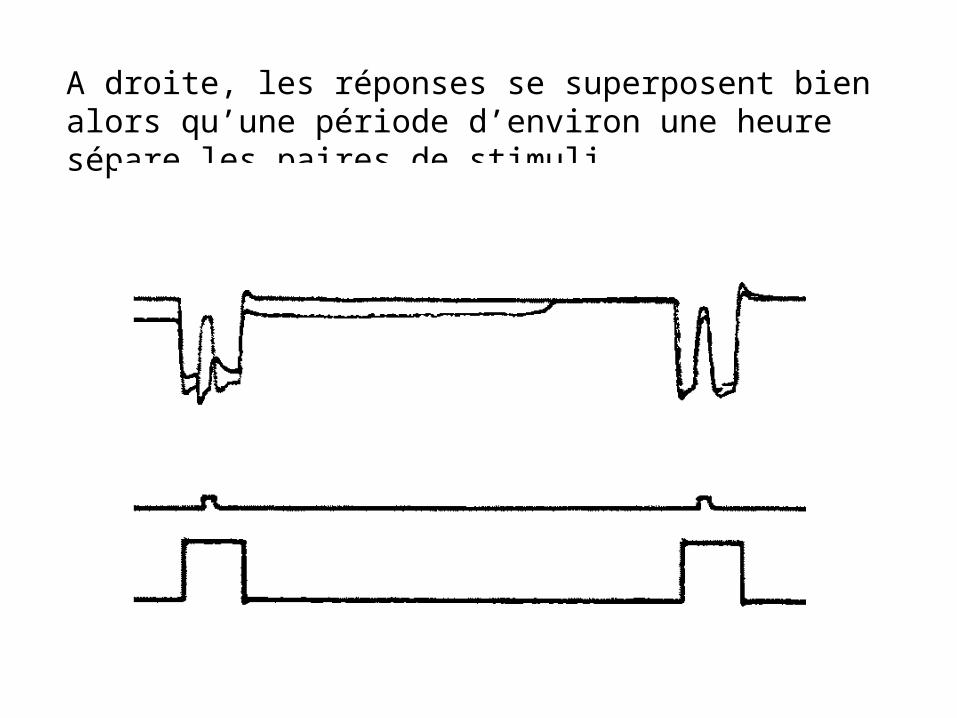
A droite, les réponses se superposent bien alors qu’une période d’environ une heure sépare les paires de stimuli.

Cet enregistrement montre un potentiel de membrane critique dans les cellules horizontales, potentiel au dessus du quel la réponse devient « antagoniste couleurs »
Des canaux différents opèrent-ils au dessus et au dessus de ce voltage critique?
Que représente la croissance des spinules? Si nous avons ici le substratum de la trace mémonique, est-ce le passage de la mémoire à court terme à la mémoire à long terme?
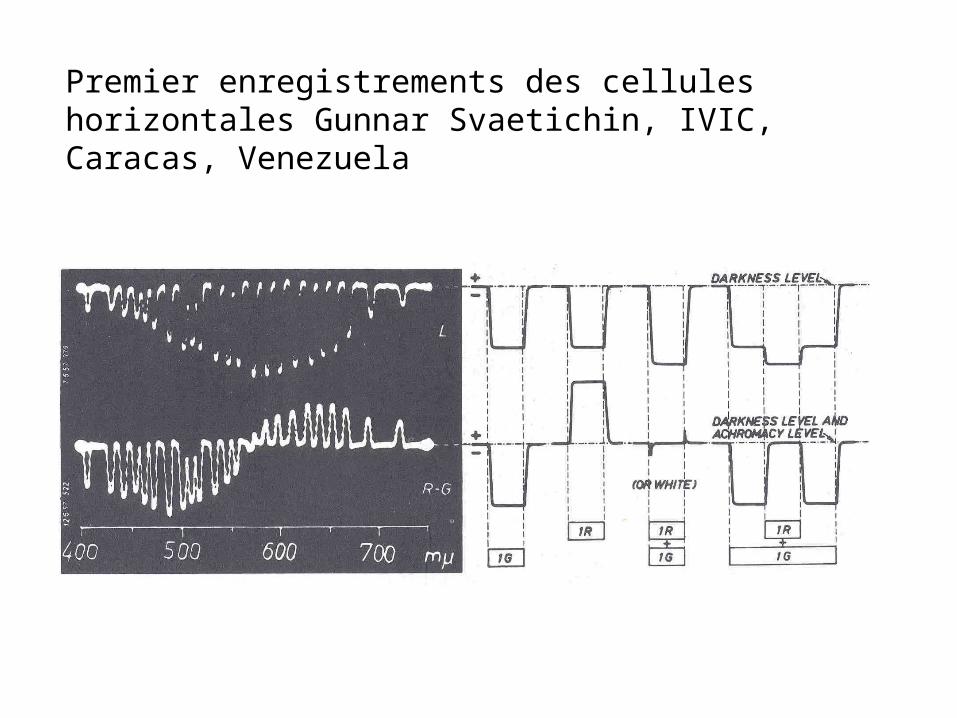
Premier enregistrements des cellules horizontales Gunnar Svaetichin, IVIC, Caracas, Venezuela

Il y a cependant un problème de synchronisation entre le phénomène électrique et les observations anatomiques. Les spinules prennent environ 30 minutes pour croitre à leur dimension maximum dans une rétine à 20 °C.

Quel est le futur de la rétine de poisson comme modèle de plasticité neuronale? Très bon
1- la situation est entièrement physiologique en opposition avec les tranches de cerveau,
2- l’input est la lumière facilement variable en intensité, en couleur et en géométrie,
3- le synapse impliqué secrète du glutamate,
4-c’est le premier synapse dans la rétine,
5-lors de section tangentielle, on peut observer des centaines de synapses ayant la même fonction,
6-l’œil opposé peut servir de contrôle lors de manipulation biochimique,
7-un contrôle existe dans une même rétine car les sphérules de bâtonnets ne montrent le phénomène.

Quelles sont les avenues prometteuses pour ce modèle de plasticité ?
1- l’identification moléculaire des spinules,
2-la mesure du Q10 de cette plasticité, le poisson tolère très bien des températures allant de 5°C à 30°C.
3-la mesure de la courbe d’apprentissage de la discrimination des couleurs en fonction de la température,
4-l’utilisation du Zebrafish.
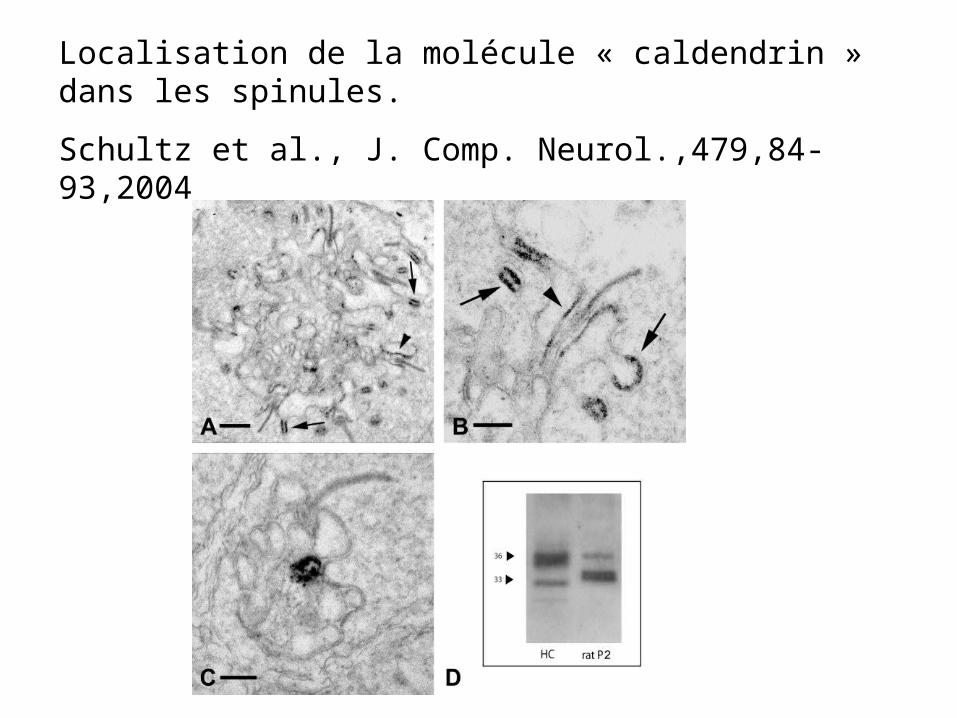
Localisation de la molécule « caldendrin » dans les spinules.
Schultz et al., J. Comp. Neurol.,479,84-93,2004
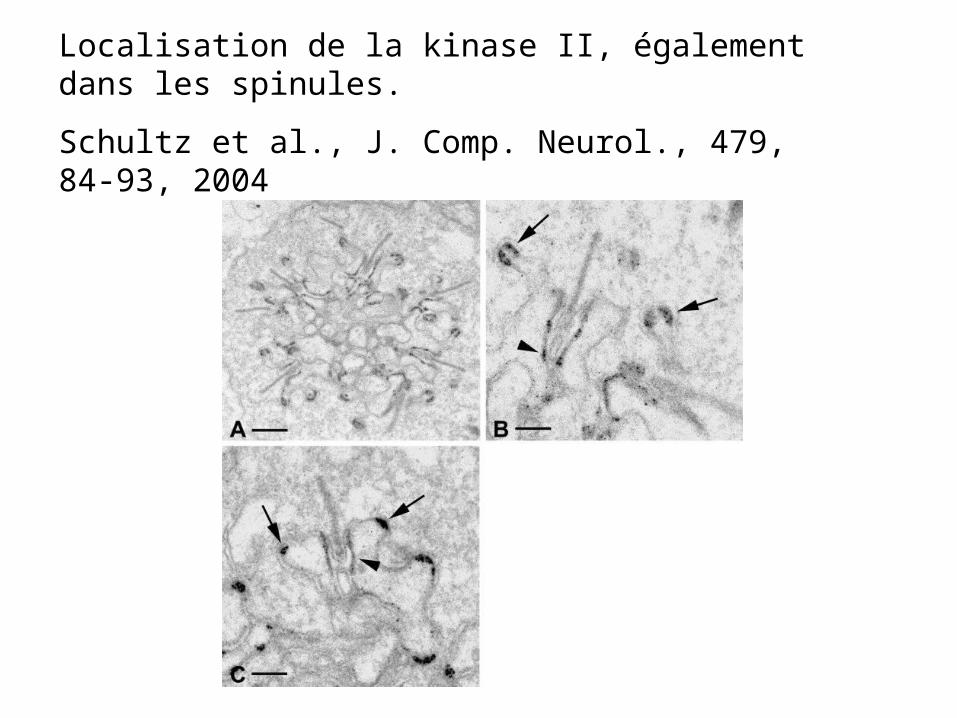
Localisation de la kinase II, également dans les spinules.
Schultz et al., J. Comp. Neurol., 479, 84-93, 2004

Données intéressantes sur « Zebrafish »
1- Zebrafish a des spinules dans les pédicules de cones.,
2-les spinules apparaissent à 5 dph (days post hatching),
3-Des molécules impliquées dans la structure des synapses telles que CaMKII et caldendrin marquées au GFP peuvent être exprimées chez ce poisson.
4-La possibilité de voir avec un microscope à deux photons la croissance des spinules.
5-Un « Zebrafish microarray » est disponible permettant d’autres expériences.
Pour plus de détails, visitez ma page Web où vous pouvez télécharger cette présentation.
http://www.mapageweb.umontreal.ca/raynauld