Embed Size (px)
Citation preview

Le développement du système nerveux
2010
Pr Pierre-Simon Jouk - Département de Génétique et Procréation Il est impossible de retracer en quelques heures l’ensemble du développement du système
nerveux central (SNC). Pour donner un ordre de grandeur de l’étendue du problème, je
donnerais 2 chiffres : 104 et 1011 ; 104 c’est le nombre de cellules de l’ectoderme qui subit
l’induction neurale primaire, c’est donc le nombre initial des cellules neurales qui au gré des
multiplications et des organisations vont donner pour l’essentiel le système nerveux définitif ;
1011, c’est le nombre total de neurones de l’encéphale du cerveau adulte. Nous ne sommes
pas habitué à manipuler de tels nombres aussi faut’ il pour se représenter les choses, utiliser
quelques analogies ; 1011 c’est aussi l’ordre de grandeur du nombre d’arbres de la forêt
amazonienne. Si l’on s’intéresse maintenant au nombre de synapses présentes dans le
système nerveux, il est du même ordre de grandeur que le nombre total de feuilles de cette
même forêt amazonienne !
Ce vaste domaine s’est donc subdivisé en une multitude de domaines secondaires au sein
desquels se développe une recherche active. Devant un tel foisonnement, il faut bien cerner
le sujet et nous nous limiterons donc à l’étude de la mise en place du plan de base du
système nerveux, c'est-à-dire à la compartimentalisation du système nerveux qui aboutit à la
mise en place des agrégats de neurones, localisés (les noyaux du système nerveux central ;
les ganglions du système nerveux périphérique) ou diffus. Nous n’envisagerons que sous
une forme conceptuelle, schématique les liaisons qui se mettent en place entre ces
différentes structures.
Il faut aussi adopter un point de vue : il est clairement embryologique. Ce point de vue est
différent des autres points de vue : neuroanatomique, neurophysiologique, éthologique,
etc… auxquels il ne se substitue pas. Son intérêt est d’être très structurant. Nous décrirons
le système nerveux à des stades ou celui-ci est encore simple. Ces structures simples une
fois stabilisées vont être dotées de caractéristiques fonctionnelles propres ; ainsi la
connaissance de l’origine embryologique d’une structure du système nerveux procure des
renseignements précieux sur son fonctionnement quand bien même la morphogénèse
tardive a considérablement complexifié le schéma de base.
Il n’est pas encore très simple d’écrire un tel cours introductif, en effet l’embryologie du
système nerveux vient de connaître de grands bouleversements ainsi d’ailleurs que la
neuroanatomie comparée. Par exemple, des entités qui paraissaient pourtant établies, celles
statufiées dans le livre de neuroanatomie comparée d’Ariens Kappers (1ère édition 1936 ;
dernière édition 1960) d’archéo, paléo et néo encéphale sont maintenant déconsidérées.
C’est le processus normal de la science, un corpus se constitue il rend compte pendant un
1

temps des données puis se fissure sous l’apparition de nouvelles données dont il ne permet
plus de rendre compte. Non seulement cet ancien corpus n’a plus de valeurs explicative,
mais il pèse dans l’émergence des nouveaux concepts nécessaires, nous en sommes là.
Deuxième élément devant conduire à la prudence, la plupart des connaissances actuelles
ont nécessairement été obtenues à partir des données de l’expérimentation animale chez les
animaux modèles de biologie du développement : la drosophile, le nématode caenorhabditis
elegans, le poisson zèbre, le xénope, la souris et nombre d’autres petits rongeurs. Il faut
donc toujours garder une certaine prudence dans les extrapolations faites depuis ces
modèles animaux vers l’être humain.
Malgré toutes ces difficultés, nous sommes à un moment où ces données fondamentales
doivent être portées à la connaissance des étudiants et des médecins, afin de ne pas laisser
se creuser le fossé entre les disciplines scientifiques et médicales. C’est ce qu’essaye de
faire ce cours introductif qui ne se substitue pas aux excellents manuels sur l’embryologie du
système nerveux. Citons sans aucune exclusive ceux de Blechschmidt, de O’Rahilly et
Müller, d’Auroux et Haegel, de Catala, de Drews, de Langman, de Larsen, etc, etc… Ces
auteurs ont insisté chacun sur des aspects différents et surtout peut être, ils ont su
développer des collaborations avec des dessinateurs pour mettre au point des
représentations bi et tridimensionnelles qui constituent un véritable apport pédagogique.
C’est a chaque étudiant de déterminer quel est celui qui lui convient le mieux, selon qu’il
privilégie tel texte, tel graphisme et surtout tel type d’interaction entre les deux.
Ces manuels résultent d’un véritable travail d’édition qu’il faut soutenir pour permettre que
cette industrie continue à s’intéresser à l’édition de ce type d’ouvrage et à leur
réactualisation.
Quelques repères chronologiques seront donnés en se référant exclusivement au
développement humain. Ils doivent être pris comme des ordres de grandeur car les données
de l’observation humaines surtout pour les stades précoces sont extrêmement limitées et
l’extrapolation des données animales est difficile.
Nous envisagerons donc après un bref rappel du schéma d’organisation de l’embryon à 18
jours de développement embryonnaire (32 jours d’aménorrhée) successivement : 1)
l’induction neurale ; 2) le développement du tube neural jusqu’au stade phylotypique; 3) la
mise en place du système nerveux périphérique ; 4) Le développement du tube neural au
delà du stade phylotypique en insistant sur le positionnement du sulcus limitans ; 5) le
développement du cortex et des noyaux gris centraux.
Rappel organisation de l’embryon humain à J18 - L’embryon est triblastique, il a une
forme ovalaire plane et mesure entre 1 et 1,5 mm dans son grand axe (figure 1).
2

nœud de Hensen
ligne primitive membrane cloacale
mésoblaste latéral
Mésoblaste bourgeon caudal
allantoïde
vésicule vitelline
Processus chordal
Cavité amniotique
membrane cloacale
A B C D
Figure 1. Schémas de l’embryon à 18 jours de développement embryonnaire. A) vue supérieure du disque embryonnaire triblastique ; B) Coupe saggitale médiane ; C) Coupe transversale en avant du nœud de Hensen ; D) coupe transversale en arrière du nœud de Hensen, au niveau de la ligne primitive. Il est situé entre la cavité amniotique et la vésicule vitelline. L’amnios qui délimite la cavité
amniotique est en continuité au niveau du bord latéral de l’embryon avec l’ectoblaste
embryonnaire. L’entoblaste vitellin qui délimite la vésicule vitelline est en continuité avec
l’entoblaste embryonnaire. Au niveau du feuillet superficiel, l’ectoblaste on distingue la ligne
primitive qui a atteint sa longueur maximale d’environ 0,5 mm. C’est à son niveau que se
sont invaginées (on utilise aussi les termes d’enroulement ou d’involution), les cellules à
l’origine de la formation de l’entoblaste et du mésoblaste embryonnaire. Les cellules du
mésoblaste embryonnaire qui se sont invaginées au niveau des berges de la ligne primitive
atteignent les bords du disque embryonnaire ou elles font leur jonction avec le mésoblaste
extra-embryonnaire. Les cellules qui se sont invaginées au niveau du nœud de Hensen,
migrent en direction rostrale pour former la plaque prochordale (structure mésenchymateuse
immédiatement caudale à la membrane bucco-pharyngée) et le processus chordal qui
conserve une structure épithéliale en doigt de gant (le terme de processus chordal est
générique pour désigner ses 3 stades successifs : canal chordal, plaque chordale et
notochorde ou chorde définitive).
3

Du fait de l’existence du processus chordal, l’organisation du mésoblaste intra-embryonnaire
n’est pas la même en arrière et en avant du nœud de Hensen.
En arrière du nœud de Hensen, le mésoblaste invaginé au niveau des berges de la ligne
primitive occupe l’espace situé entre ecto et entoblaste, ou il forme le mésoblaste du
bourgeon caudal. A l’exception de la zone de la membrane cloacale, zone d’accolement
étroit de l’ecto et de l’entoblaste au pole caudal de l’embryon.
En avant du nœud de Hensen, et en arrière de la plaque prochordale le mésoblaste est
constitué du processus chordal axial et de 2 nappes de mésoblaste latéral. Ces 2 nappes
latérales, ont migrés vers l’avant depuis les berges de la ligne primitive, de part et d’autre du
processus chordal. Elles ne fusionnent sur la ligne médiane qu’en avant de la membrane
bucco-pharyngienne.
1) L’induction neurale C’est à 19 jours de développement embryonnaire au stade de l’embryon triblastique que
des interactions s’établissent à distance entre les cellules du mésoblaste axial (plaque
prochordale et plaque chordale), qui émettent une substance diffusible et les cellules de
l’ectoblaste sus-jacent, qui possèdent les récepteurs nécessaires et est donc compétent pour
répondre au signal diffusible. Ces cellules ectoblastiques axiales se différencient alors en
cellules neuroépithéliales de la plaque neurale. Ce processus de stimulation par un tissu
d’un autre tissu compétent qui se différencie est appelé induction, et dans ce cas particulier
d’interaction entre la plaque chordale et l’ectoblaste sus-jacent d’induction neurale primaire.
Les cellules à la jonction entre plaque neurale et ectoblaste constituent les cellules de la
crête neurale (figure 2).
4

Cellules neuroépithélialesDe la plaque neurale
Crêtes neurales
Plaque chordale
Vésicule vitelline
Entoblaste
Ectoblaste
Amnios
Figure 2. L’induction neurale primaire. Coupe transversale de l’embryon en avant du nœud de Hensen à 19 jours de développement embryonnaire.
Cette induction neurale primaire ne rend pas compte de la formation de la partie la plus
caudale du tube neural, en arrière du nœud de Hensen, qui provient d’un processus de
neurulation secondaire, par différenciation neurale de la partie médiane du mésoblaste du
bourgeon caudal (à ce niveau naîtra donc encore.4 paires de nerfs spinaux, 3 sacrées et une
coccygienne).
A 20 jours, la plaque neurale accentue sa plicature pour former la gouttière neurale. Dès ce
stade, on peut distinguer une portion crâniale à l’origine de l’encéphale (déjà divisé en
encéphale antérieur ou prosencéphale, moyen ou mésencéphale, et postérieur ou
rhombencéphale) et une portion caudale étroite, au niveau des somites en formation, à
l’origine de la moelle épinière.
S’il est logique d’insister sur cette interaction entre le mésoblaste axial et l’ectoblaste sus-
jacent, il ne faut pas non plus le schématiser outrancièrement. Les molécules morphogènes
qui diffusent depuis le processus chordal ont une concentration tissulaire qui décroit avec la
distance. D’autre part les autres régions embryonnaires : le mésoblaste latéral, l’entoblaste
émettent eux aussi des molécules morphogènes. Chaque territoire embryonnaire va donc
être soumis à une combinatoire propre de concentrations de morphogènes. C’est ce qui
permet d’expliquer la différenciation visible dès 19 jours entre la partie médiale de la plaque
neurale (forte concentration de morphogènes provenant du processus chordal) et les crêtes
neurales (concentration plus faible des morphogènes en provenance du processus chordal,
interactions plus fortes avec le mésoblaste latéral). Mais il ne faut pas perdre de vue que les
mécanismes inductifs à l’œuvre au niveau des régions latérales de la plaque neurale ne
5
sont pas strictement les mêmes qu’au niveau des régions médiales. Conséquences à garder
à l’esprit, la ligne médiane stricto sensu est constituée de toutes les structures dérivées des
régions médiales de la plaque neurale (futur plancher du tube neural), et non pas des
structures dérivées des régions latérales de la plaque neurale (futur toit du tube neural).
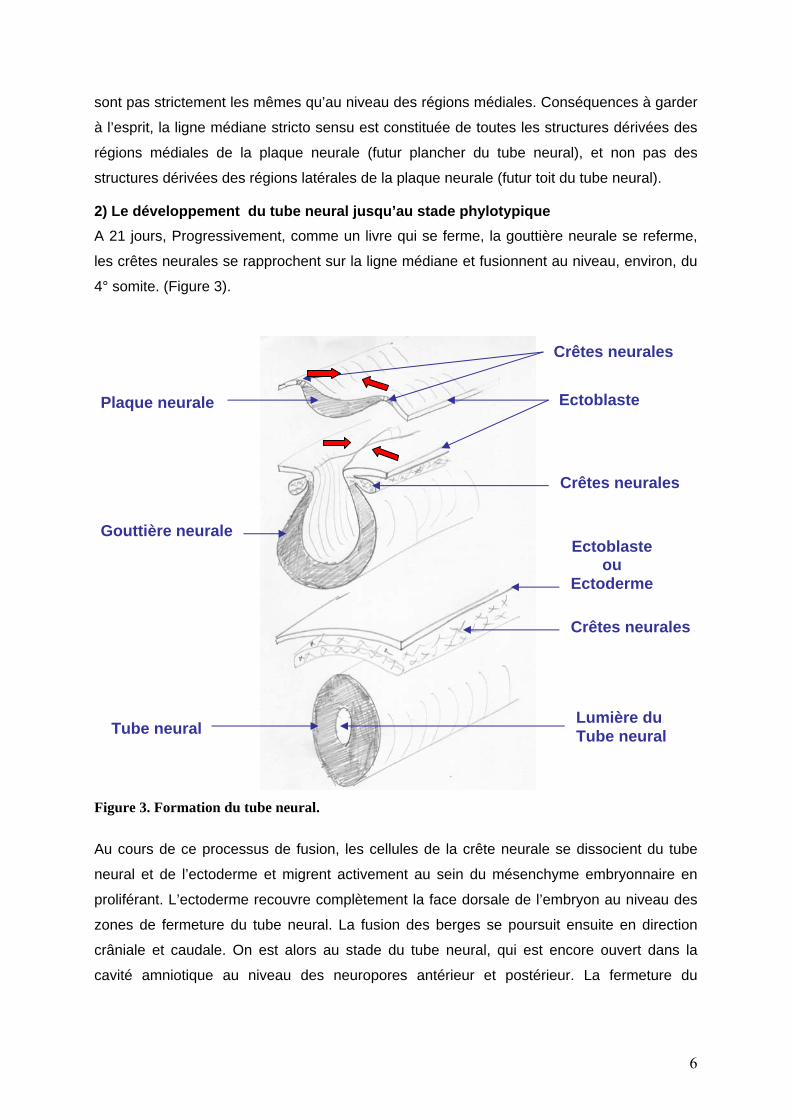
2) Le développement du tube neural jusqu’au stade phylotypique A 21 jours, Progressivement, comme un livre qui se ferme, la gouttière neurale se referme,
les crêtes neurales se rapprochent sur la ligne médiane et fusionnent au niveau, environ, du
4° somite. (Figure 3).
Plaque neurale
Gouttière neurale
Tube neural
Crêtes neurales
Ectoblaste
Crêtes neurales
Ectoblaste ou
Ectoderme
Crêtes neurales
Lumière du Tube neural
Figure 3. Formation du tube neural.
Au cours de ce processus de fusion, les cellules de la crête neurale se dissocient du tube
neural et de l’ectoderme et migrent activement au sein du mésenchyme embryonnaire en
proliférant. L’ectoderme recouvre complètement la face dorsale de l’embryon au niveau des
zones de fermeture du tube neural. La fusion des berges se poursuit ensuite en direction
crâniale et caudale. On est alors au stade du tube neural, qui est encore ouvert dans la
cavité amniotique au niveau des neuropores antérieur et postérieur. La fermeture du
6
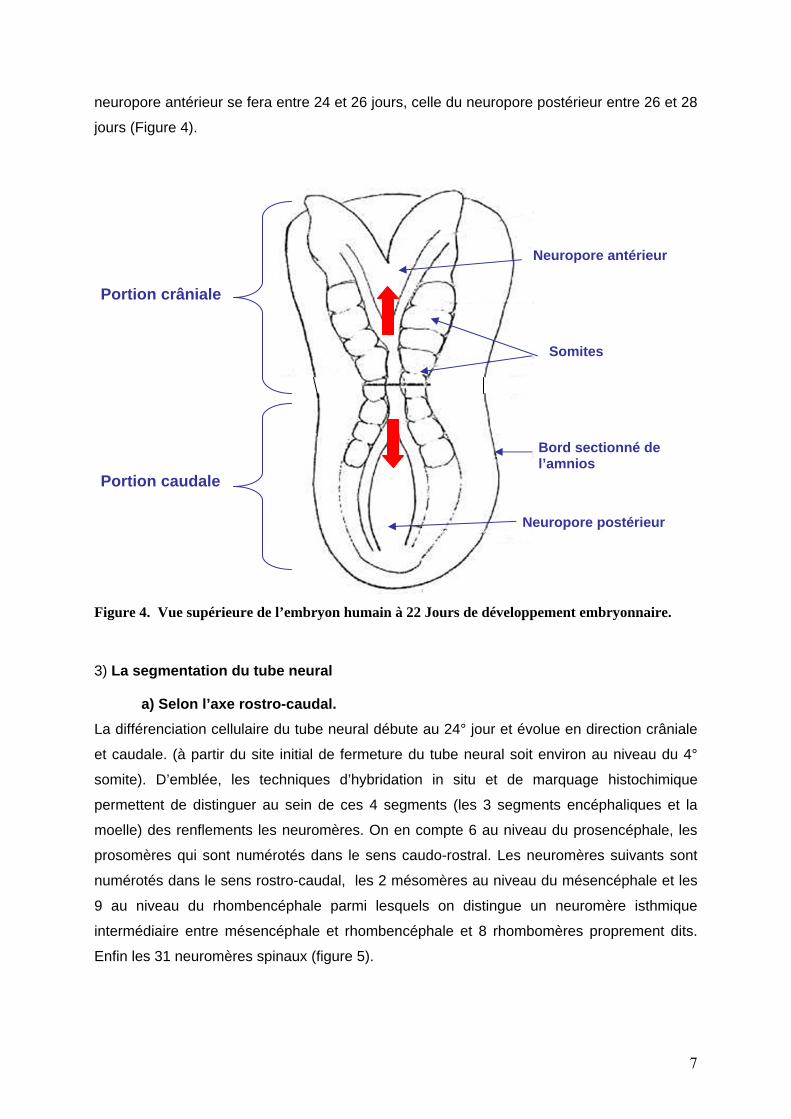
neuropore antérieur se fera entre 24 et 26 jours, celle du neuropore postérieur entre 26 et 28
jours (Figure 4).
Neuropore antérieur
Somites
Bord sectionné de l’amnios
Neuropore postérieur
Portion crâniale
Portion caudale
Figure 4. Vue supérieure de l’embryon humain à 22 Jours de développement embryonnaire.
3) La segmentation du tube neural
a) Selon l’axe rostro-caudal. La différenciation cellulaire du tube neural débute au 24° jour et évolue en direction crâniale
et caudale. (à partir du site initial de fermeture du tube neural soit environ au niveau du 4°
somite). D’emblée, les techniques d’hybridation in situ et de marquage histochimique
permettent de distinguer au sein de ces 4 segments (les 3 segments encéphaliques et la
moelle) des renflements les neuromères. On en compte 6 au niveau du prosencéphale, les
prosomères qui sont numérotés dans le sens caudo-rostral. Les neuromères suivants sont
numérotés dans le sens rostro-caudal, les 2 mésomères au niveau du mésencéphale et les
9 au niveau du rhombencéphale parmi lesquels on distingue un neuromère isthmique
intermédiaire entre mésencéphale et rhombencéphale et 8 rhombomères proprement dits.
Enfin les 31 neuromères spinaux (figure 5).
7

Portion crânialeEncéphale
Portion caudaleMoelle
[ Pro- ][ més- ][ Rhomb- ]
Plaque prochordale Chorde
Figure 5. Représentation schématique latérale du tube neural, région encéphalique au stade 3 vésicules et région spinale. Ce schéma permet d’insister sur les interactions étroites entre d’une part la plaque prochordale et le prosencéphale, et d’autre part la chorde et le reste du tube neural. Nous étudierons la spécialisation de ces neuromères en fonction du niveau après avoir
étudié les grands principes de la différenciation dorso-ventrale.
Au 23° jour, l’extrémité céphalique accentue son inflexion en direction ventrale. Cette
inflexion : la courbure céphalique, se produit au niveau du mésencéphale. Il devient alors
plus facile de distinguer sur des critères morphologiques externes : les différents segments
du SNC : prosencéphale, le mésencéphale et rhombencéphale suivi de la future moelle
épinière (figure 6). Par ailleurs les vésicules optiques et otiques sont visibles. Le mécanisme
de formation de chacune de ces vésicules est fondamentalement différent, la vésicule
optique est une expansion directe du tube neural, la vésicule otique résulte d’une
invagination de l’ectoderme de la placode otique.
8
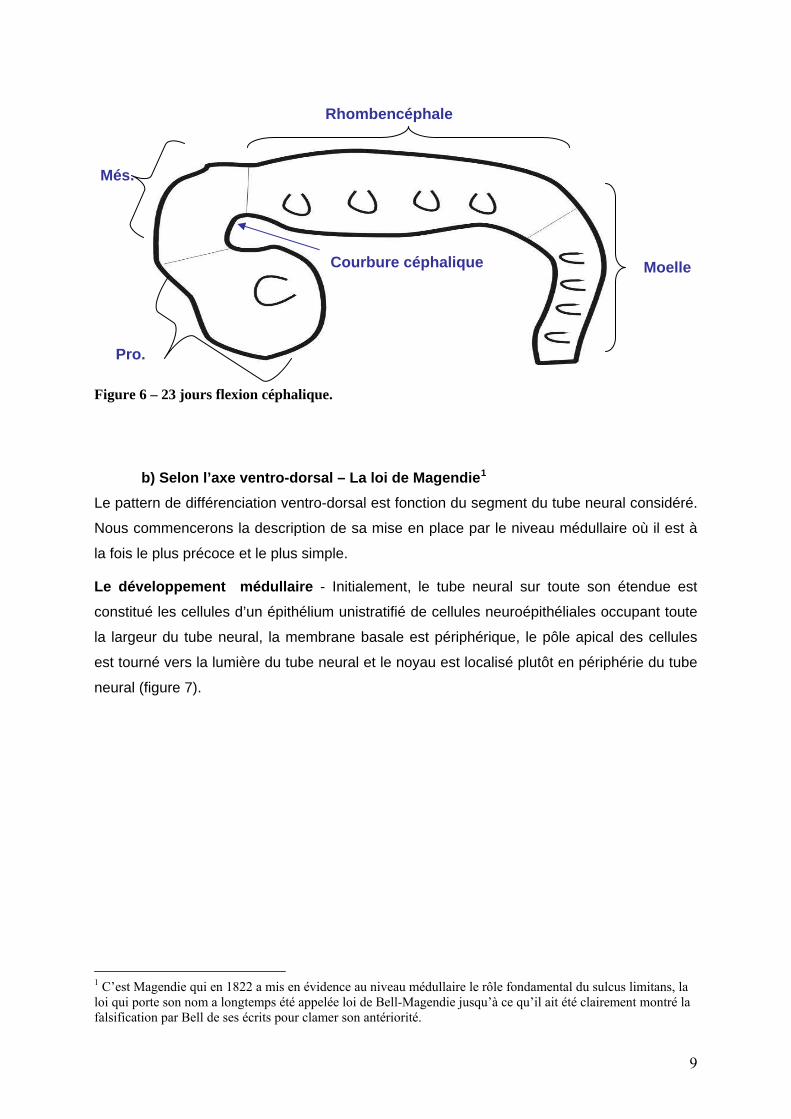
Figure 6 – 23 jours flexion céphalique.
Pro.
Més.
Rhombencéphale
Courbure céphalique Moelle
b) Selon l’axe ventro-dorsal – La loi de Magendie1 Le pattern de différenciation ventro-dorsal est fonction du segment du tube neural considéré.
Nous commencerons la description de sa mise en place par le niveau médullaire où il est à
la fois le plus précoce et le plus simple.
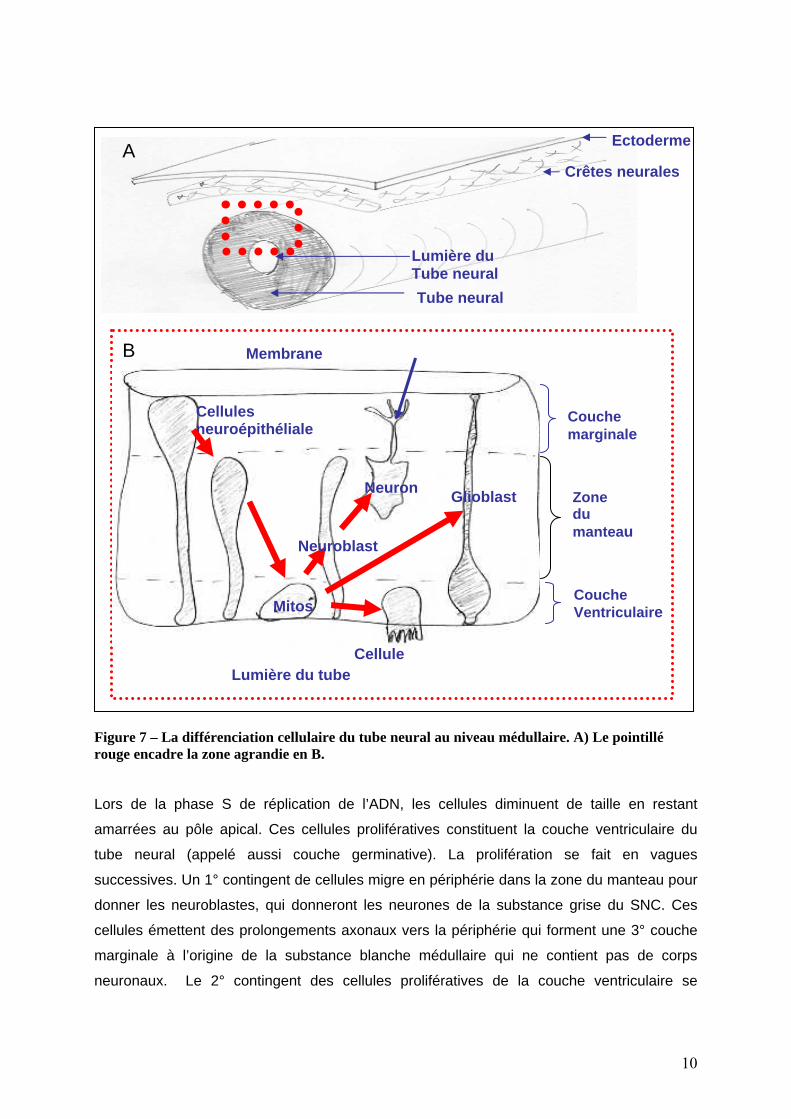
Le développement médullaire - Initialement, le tube neural sur toute son étendue est
constitué les cellules d’un épithélium unistratifié de cellules neuroépithéliales occupant toute
la largeur du tube neural, la membrane basale est périphérique, le pôle apical des cellules
est tourné vers la lumière du tube neural et le noyau est localisé plutôt en périphérie du tube
neural (figure 7).
1 C’est Magendie qui en 1822 a mis en évidence au niveau médullaire le rôle fondamental du sulcus limitans, la loi qui porte son nom a longtemps été appelée loi de Bell-Magendie jusqu’à ce qu’il ait été clairement montré la falsification par Bell de ses écrits pour clamer son antériorité.
9
Tube neural
Ectoderme
Crêtes neurales
Lumière du Tube neural
Lumière du tube
Cellules neuroépithéliale
Membrane
MitosCouche Ventriculaire
Zone du manteau
Couche marginale
Neuroblast
Neuron Glioblast
Cellule
A B
Figure 7 – La différenciation cellulaire du tube neural au niveau médullaire. A) Le pointillé rouge encadre la zone agrandie en B.
Lors de la phase S de réplication de l’ADN, les cellules diminuent de taille en restant
amarrées au pôle apical. Ces cellules prolifératives constituent la couche ventriculaire du
tube neural (appelé aussi couche germinative). La prolifération se fait en vagues
successives. Un 1° contingent de cellules migre en périphérie dans la zone du manteau pour
donner les neuroblastes, qui donneront les neurones de la substance grise du SNC. Ces
cellules émettent des prolongements axonaux vers la périphérie qui forment une 3° couche
marginale à l’origine de la substance blanche médullaire qui ne contient pas de corps
neuronaux. Le 2° contingent des cellules prolifératives de la couche ventriculaire se
10

différencie en glioblastes (cellule souche gliale : oligodendrocytes, astrocytes). Enfin, le 3°
contingent se différencie en cellules épendymaires.
Le réarrangement de la zone du manteau et de la couche marginale au niveau de la moelle
épinière débute dès la fin de la 4ème semaine de développement embryonnaire. (figure 8).
Lumière du tube neural
Couche Ventriculaire
Zone du manteau
Couche marginale
Colonnes ventrales(ou fondamentales)
Plancher
Colonnes dorsales (ou alaires)
Toit
Sulcus limitans
Colonnes Intermédio-latérales
Figure 8. Différenciation ventro-dorsale du tube neural au niveau médullaire.
A la fin de la 4° semaine, la zone du manteau forme 4 colonnes : 2 ventrales (ou
fondamentales) où se situent les corps cellulaires des motoneurones somatiques assurant
l’innervation des muscles volontaires striés ; 2 colonnes dorsales (ou alaires) où se situent
les neurones d’association entre terminaison axonale des neurones des ganglions rachidiens
dorsaux et les motoneurones. Enfin, à partir de la partie la plus dorsale des colonnes
ventrales, s’individualise dès 28 jours : du 1er neuromère thoracique au 3ème neuromère
lombaire (T1-L3), les colonnes intermédio-latérales où se situent les 1° neurones du système
sympathique ; et aux niveaux des neuromères sacrés 2 à 4 (S2-S4) les 1° neurones du
système parasympathique sacral. Les colonnes ventrales et dorsales sont séparées par le
sillon limitant ou « sulcus limitans » qui séparent donc les versants moteurs et sensitifs du
tube neural. La zone marginale ventrale forme une fine lame le plancher. La zone marginale
dorsale forme une fine lame le toit.
11
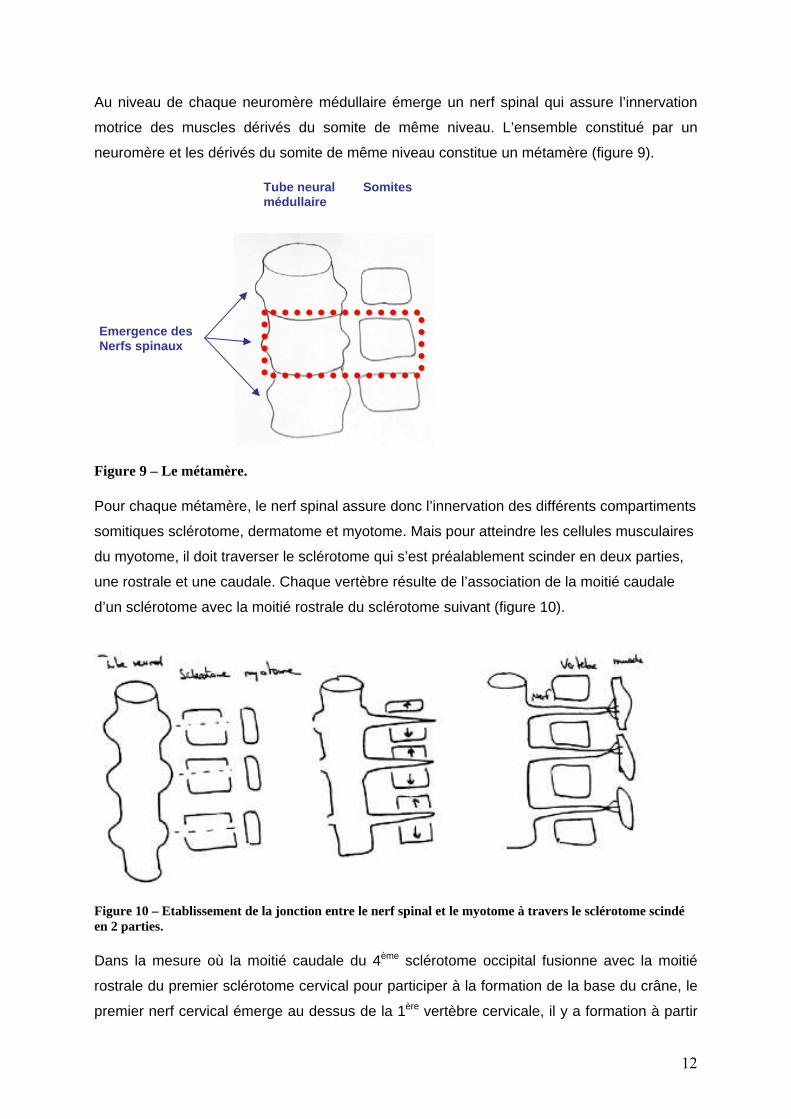
Au niveau de chaque neuromère médullaire émerge un nerf spinal qui assure l’innervation
motrice des muscles dérivés du somite de même niveau. L’ensemble constitué par un
neuromère et les dérivés du somite de même niveau constitue un métamère (figure 9).
Tube neural Somitesmédullaire
Emergence des Nerfs spinaux
Figure 9 – Le métamère. Pour chaque métamère, le nerf spinal assure donc l’innervation des différents compartiments
somitiques sclérotome, dermatome et myotome. Mais pour atteindre les cellules musculaires
du myotome, il doit traverser le sclérotome qui s’est préalablement scinder en deux parties,
une rostrale et une caudale. Chaque vertèbre résulte de l’association de la moitié caudale
d’un sclérotome avec la moitié rostrale du sclérotome suivant (figure 10).
Figure 10 – Etablissement de la jonction entre le nerf spinal et le myotome à travers le sclérotome scindé en 2 parties. Dans la mesure où la moitié caudale du 4ème sclérotome occipital fusionne avec la moitié
rostrale du premier sclérotome cervical pour participer à la formation de la base du crâne, le
premier nerf cervical émerge au dessus de la 1ère vertèbre cervicale, il y a formation à partir
12

de 8 somites cervicaux de 7 vertèbres cervicales et le 8ème nerf cervical émerge sous la 7ème
vertèbre cervicale (figure 11).
Figure 11 – Rapport entre les nerfs crâniens et les vertèbres Finalement, l’innervation des dérivés somitiques cervico-thoraco-lombo-sacro-coccygiens est
assuré par les 31 paires de nerfs spinaux (figure 12).
13

Tube neural embryonnaire Tube neural avant fermeture des neuropores adulte
Neurulation primaire
Neurulation secondaire
5 paires de nerfs spinaux lombaires
12 paires de nerfs spinaux thoraciques
8 paires de nerfs spinaux cervicaux
2 paires de nerfs Spinaux sacrés 3 paires de nerfs Spinaux sacrés 1 paire de nerfs Spinaux coccygiens
4 paires Somites occipitaux 8 paires Somites cervicaux
12 paires Somites thoraciques
5 paires Somites lombaires
5 paires Somites sacrées
3 ou 4 paires Somites coccygiennes
Somites embryons
Figure 12 – Innervation des dérivés somitiques par les nerfs spinaux.
Digression 1 : Le stade Phylotypique
A ce stade 28 jours de développement embryonnaire, il est particulièrement intéressant de
replacer le système nerveux central au sein de l’ensemble de l’embryon, nous sommes en
effet au stade dit phylotypique. L’embryon est délimité, relié au placenta per le cordon
ombilical, le système cardiovasculaire est fonctionnel dans sa forme embryonnaire, les
bourgeons des membres sont apparus ainsi que les arcs branchiaux qui modèlent la région
cervico-faciale. L’extrémité cervico-céphalique est au début de la 4ème semaine constitué par
2 feuillets épithéliaux ecto- et entoblastique entre lesquels se situe un tissu mésenchymateux
contenant des cellules dérivées des cellules de crêtes neurales et du mésoblaste. Ces
épithéliums constitue un patron (au sens du patron de couture nécessaire à la confection
d’un vêtement, d’une robe…) qui modèle l’expansion du mésenchyme sous jacent. Au cours
de la 4° semaine, on note l’apparition successive dans le sens crânio-caudal des arcs
branchiaux séparés par des poches branchiales ecto- et entoblastique, l’ébauche optique et
la vésicule otique sont apparus (figure 13).
14

Arcs Branchiaux
Figure 13 – Stade phylotypique – 28 Jours –à droite coupe horizontale du pôle céphalique de l »’embryon selon le trait hachuré rouge permettant de visualiser les arcs branchiaux ainsi que les poches ecto-et entobranchiales.
L’embryon à ce stade a atteint le stade dit phylotypique, car très conservé au cours de
l’évolution (ou phylogénèse). Quels que soient les mécanismes initiaux de l’embryogenèse
qui sont très divers chez les vertébrés, ils convergent vers ce stade qui constitue un véritable
goulot d’étranglement. Ce n’est qu’après ce stade que la morphogenèse ultérieure permettra
de distinguer les différentes espèces animales (figure 14).
Renflement cardiaque
Bourgeonsdes
membres 28 Jours – 3,5 mm
entoblaste ectoblaste
I II III IV
I II III IV
Poches branchiales
15

Figure 14 – Le stade phylotypique. A ce stade le développement du système nerveux central est très semblable pour toutes les espèces du phylum des vertébrés quelque soit le mode de développement préalable et ultérieur. L’étude du devenir des différents arcs branchiaux est au-delà du cadre de ce cours consacré
au développement du SNC. Nous rappellerons simplement que l’origine embryologique du
mésenchyme des arcs branchiaux est double (mésoblastique et ectomésenchyme dérivé
des crêtes neurales mésoblastiques et rhombencéphaliques). Le mésenchyme de chaque
arc va donner des dérivés vasculaires, cartilagineux et osseux. Chaque arc reçoit un
contingent de cellules musculaires du mésoderme para axial innervé par un nerf crânien
propre à chaque arc. Chaque arc est parcouru par une paire d’arc aortique (un droit et un
gauche) reliant l’aorte ventrale et les aortes dorsales encore paires à ce stade du
développement. L’innervation sensitive et motrice de chaque arc est assurée par une paire
de nerfs crâniens. Le premier arc mandibulaire est innervé par la 5ème paire des nerfs
crâniens, le trijumeau dont l’action prédominante est la commande des muscles de la
mastication. Le 2ème arc Hyoïdien est innervé par la 7ème paire des nerfs crâniens le facial
dont l’action prédominante est la commande des muscles de la mimique. Le 3ème arc
16

branchial est innervé par la 9ème paire des nerfs crâniens, le glosso-pharyngien qui innerve le
muscle stylo-pharyngien qui soulève le pharynx lors de la phonation et la déglutition. Enfin
les muscles des 4ème et 6ème arcs branchiaux qui interviennent dans la phonation et la
déglutition sont innervés par la 10ème paire des nerfs crâniens le nerf vague ou
pneumogastrique.
Alors qu’au niveau du tronc et des membres il a été relativement facile d’introduire la notion
de métamère permettant d’établir une correspondance entre les dérivés du tube neural et du
mésoderme d’un même niveau ; au niveau cervico-facial l’adjonction des arcs branchiaux
rends cette description plus difficile en effet la segmentation des arcs branchiaux n’est pas
identique à la segmentation neurale et mésodermique. Cette mise en correspondance est
illustrée dans la figure 15.
Tube neural Mésoblaste para-axial Arcs branchiaux
Rhombomère 1
Rhombomère 2
Rhombomère 3
Rhombomère 4
Rhombomère 5
Rhombomère 6
Rhombomère 7
Rhombomère 8
Somitomères
Somites
4 5 6 7
1 2 3 4
I II III V VI
Figure 15 – Mise en correspondance schématique des arcs branchiaux (à gauche, avec la vésicule otique représentée entre le 2ème et le 3ème arc branchial), les rhombomères et les dérivés mésodermiques (à droite).
Le développement du Système nerveux périphérique (SNP). Les cellules du SNP dérivent essentiellement des cellules des crêtes neurales. Nous
n’évoquerons pas dans ce cours introductif la contribution des placodes ectodermiques
17

cervico-faciales dont dérive un contingent minoritaire de cellules du SNP. Pour étudier cette
mise en place du SNP, il faut décrire successivement la migration des cellules des crêtes
neurales puis la mise en place des connexions axonales et dendritiques entre l’ensemble
des cellules du SNP. Cette description sera faite au niveau médullaire puis au niveau
céphalique.
La migration des cellules des crêtes neurales au niveau médullaire.
Les cellules de la crête neurale prolifèrent intensément et migrent de façon active selon 2
voies : Superficielle dans le mésenchyme sous ectodermique avant d’atteindre l’épiderme où
ils se différencieront en mélanocytes ; Profonde entre le tube neural et les dérivés somitiques
(figure 16).
Voie de migration superficielle
Figure 16 – Les cellules des crêtes neurales migrent en proliférant dès le stade de la gouttière neurale. Elles peuvent emprunter 2 voies de migration, une superficielle, l’autre profonde. Leur devenir est différent selon le niveau spinal ou crânien. Au niveau spinal, la migration de
ces cellules peut s’interrompre a 4 niveaux distincts (figure 17).
18

1- Ganglions de la racine dorsale
2- Chaîne ganglionnaire Sympathique paravertébrale
3- Ganglions préaortiques
Figure 17 – Représentation schématique des 4 niveaux distincts d’arrêt de migration des cellules des crêtes neurales. Attention c’est exclusivement au niveau cervical que ces 4 arrêts peuvent être observés sur une même coupe Le premier arrêt se fait latéralement au tube neural pour former les ganglions de la racine
dorsale. Ces cellules dérivées des crêtes neurales se différencient en neurones (de forme
pseudo -unipolaire) qui donneront naissances aux fibres nerveuses sensitives qui conduisent
l’influx nerveux depuis les terminaisons nerveuses périphériques vers la moelle épinière et
en cellules gliales. La distribution de ces ganglions de la racine dorsale est segmentaire à
l’exception du 1° neuromère cervical et des 2° et 3° neuromères coccygiens. Ceci aboutit à
la formation de : 7 paires de ganglions cervicaux, 12 thoraciques ; 5 lombaires ; 5 sacrés et 1
coccygien (figure 13).
Les cellules qui migreront au delà de ce premier arrêt donneront essentiellement les corps
cellulaires des neurones des ganglions du SNA. Cette migration se fait différemment selon le
niveau spinal.
Le 2° arrêt se fait latéralement et ventralement au tube neural. Ces cellules des CN nées des
crêtes neurales médullaire cervicales, thoraciques, lombaires, sacrées et coccygiennes,
formeront les neurones et les cellules gliales de la chaîne ganglionnaire sympathique
paravertébrale. Ses neurones constitueront les fibres post-ganglionnaires sympathiques qui
assurent le contrôle moteur des fonctions involontaires (rythme cardiaque, secrétions
glandulaires, mouvements intestinaux) au cours des situations d’alerte (stress). Alors qu’au
19

niveau thoracique, lombaire et sacral, il y a formation d’une paire de ganglion au niveau de
chaque neuromère ; au niveau de la CN cervicale il y a formation simplement de 3 paires de
volumineux ganglions sympathiques cervicaux supérieurs, moyens et inférieurs. Enfin, les
cellules de la CN coccygienne forment une seule paire de ganglions coccygiens (figure 18).
La plupart des axones de la chaine sympathique rejoignent le nerf spinal par le rameau
communiquant gris. Il y a quelques exceptions à ce pattern général. Certaines fibres post-
ganglionnaires issues du ganglion cervical supérieur suivent les vaisseaux jusqu’au
structures cervico-céphaliques qu’elles innervent (muqueuses orales et nasales, glandes
lacrymales, muscle dilatateur pupillaire de l’iris). Certaines fibres post-ganglionnaires issues
des ganglions cervicaux et thoraciques T1 à T4 forment les nerfs des plexus cardiaques et
pulmonaires.
Rx communicants blancs
Tronc sympathique
Chaîne sympathique Crêtes neurales
12 paires de ggls Σ T
5 paires de ggls Σ L
5 paires de ggls Σ S
3 paires de ggls Σ CSup, moy et inf.
Figure 18 - Représentation schématique de la chaîne ganglionnaire sympathique paravertébrale.
20

Ces ganglions sympathiques reçoivent les axones des premiers neurones moteurs
sympathiques centraux (Rameaux communicants blancs) provenant des colonnes
intermédio-latérales des 12 neuromères thoraciques et des 3 premiers lombaires. Le
système sympathique est donc Thoraco -lombaire du point de vue de sa commande centrale
(T1-T4 : tête, cœur, poumons ; T5-L3 : tube digestif et dérivés). Ces fibres pré-
ganglionnaires sympathiques peuvent se poursuivre en direction crâniale et caudale avant
de faire synapse. L’ensemble des fibres pré-ganglionnaires qui relient les ganglions
sympathiques constitue le tronc sympathique qui assure l’innervation des ganglions
sympathiques. Du fait du décalage entre l’origine des 1° neurones sympathiques au niveau
du SNC (T1-L3) et l’origine des 2° neurones à partir des cellules des crêtes neurales (C, T,
L, S et C) les ganglions cervicaux, L4, L5, Sacrés et coccygiens ne reçoivent pas de
rameaux communicants blancs.
Tous les neurones sympathiques périphériques ne sont pas localisés au niveau de la chaîne
ganglionnaire sympathique. Certaines cellules de la crête neurale poursuivent leur migration
jusqu’au niveau de l’aorte dorsale ou elles forment les ganglions pré aortiques (3° arrêt) d’ou
sont issues les fibres sympathiques post ganglionnaires destinées à l’innervation de
l’intestin. Les cellules des crêtes neurales cervicales formeront une paire de ganglions à la
base du tronc cœliaque. Les cellules des crêtes neurales thoraciques formeront des
ganglions plus diffus associés à l’artère mésentérique supérieure. Les cellules des crêtes
neurales lombaires formeront des ganglions plus diffus associés à l’artère mésentérique
inférieure. L’innervation pré ganglionnaire de ces ganglions pré-aortique est assurée par les
nerfs splanchniques qui naissent aux niveaux T5-L3. Les axones à destinée de ces nerfs
splanchniques entrent dans les ganglions correspondants de la chaîne sympathique et en
sortent immédiatement sans y faire synapse pour former les nerfs splanchniques qui se
distribuent aux différents ganglions préaortiques (figure 19).
21
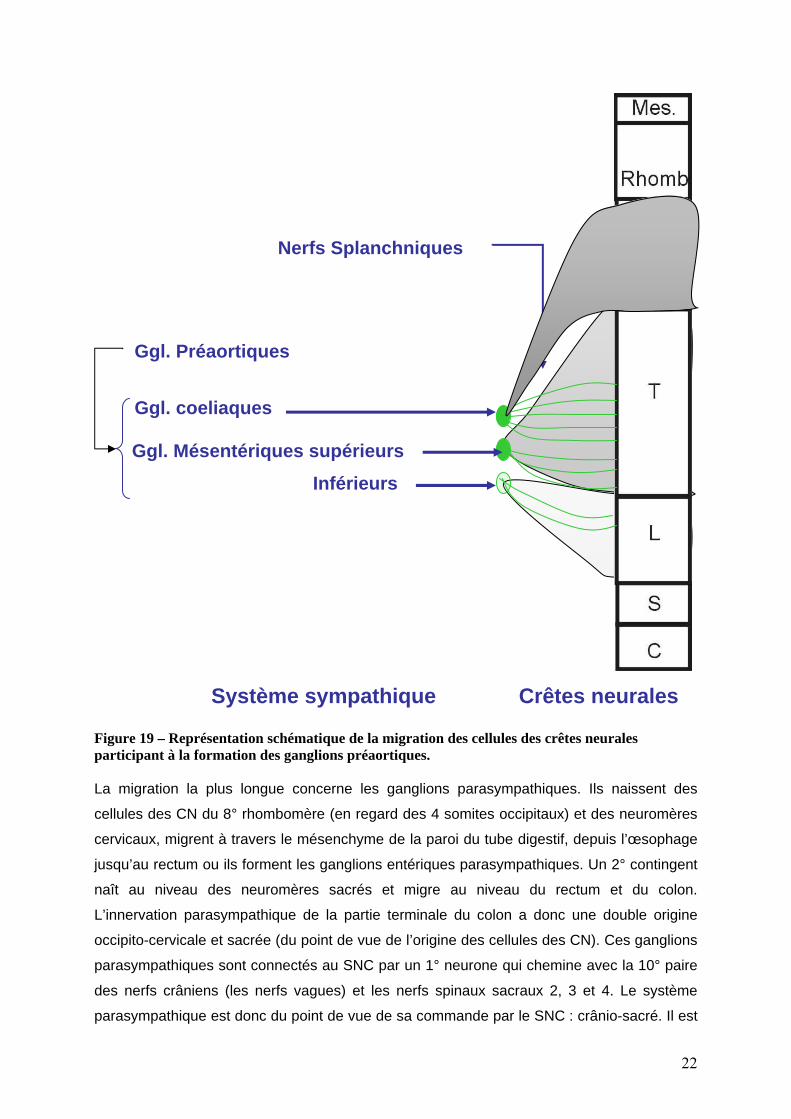
Nerfs Splanchniques
Ggl. Préaortiques
Système sympathique Crêtes neurales
Ggl. coeliaques
Ggl. Mésentériques supérieurs
Inférieurs
Figure 19 – Représentation schématique de la migration des cellules des crêtes neurales participant à la formation des ganglions préaortiques. La migration la plus longue concerne les ganglions parasympathiques. Ils naissent des
cellules des CN du 8° rhombomère (en regard des 4 somites occipitaux) et des neuromères
cervicaux, migrent à travers le mésenchyme de la paroi du tube digestif, depuis l’œsophage
jusqu’au rectum ou ils forment les ganglions entériques parasympathiques. Un 2° contingent
naît au niveau des neuromères sacrés et migre au niveau du rectum et du colon.
L’innervation parasympathique de la partie terminale du colon a donc une double origine
occipito-cervicale et sacrée (du point de vue de l’origine des cellules des CN). Ces ganglions
parasympathiques sont connectés au SNC par un 1° neurone qui chemine avec la 10° paire
des nerfs crâniens (les nerfs vagues) et les nerfs spinaux sacraux 2, 3 et 4. Le système
parasympathique est donc du point de vue de sa commande par le SNC : crânio-sacré. Il est
22
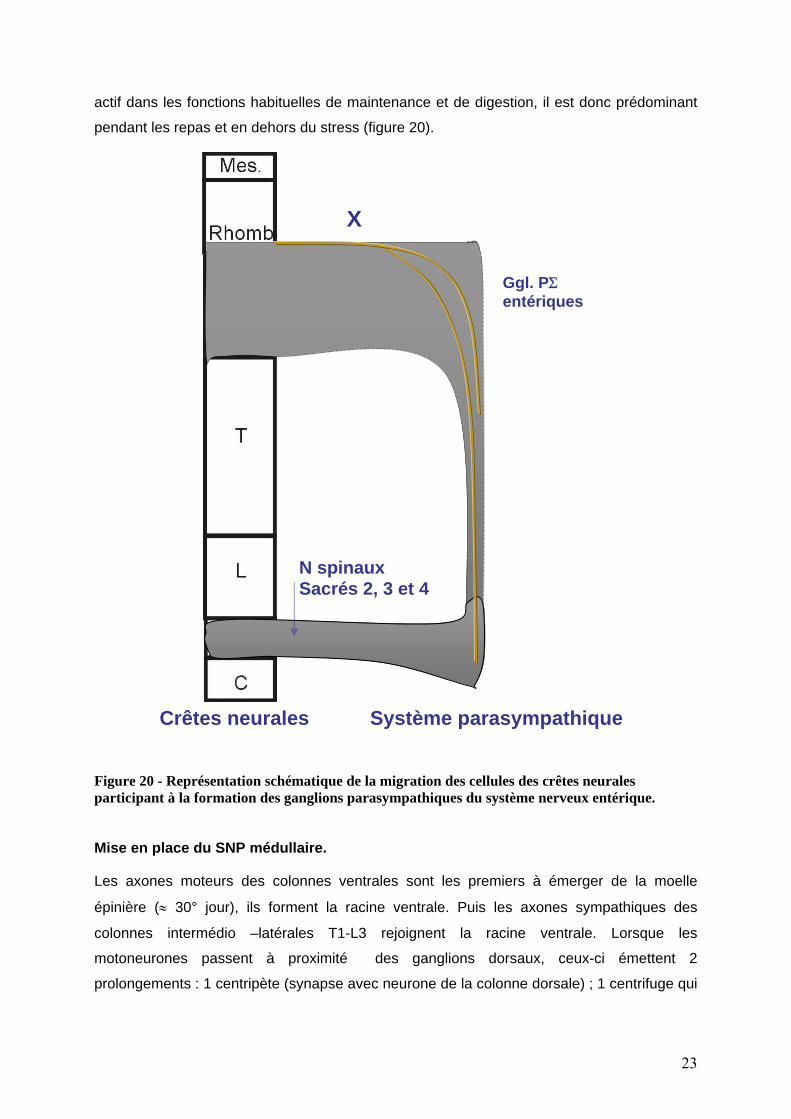
actif dans les fonctions habituelles de maintenance et de digestion, il est donc prédominant
pendant les repas et en dehors du stress (figure 20).
X
Ggl. PΣ entériques
Crêtes neurales Système parasympathique
N spinaux Sacrés 2, 3 et 4
Figure 20 - Représentation schématique de la migration des cellules des crêtes neurales participant à la formation des ganglions parasympathiques du système nerveux entériqu
e.
ise en place du SNP médullaire.
ventrales sont les premiers à émerger de la moelle
M
Les axones moteurs des colonnes
épinière (≈ 30° jour), ils forment la racine ventrale. Puis les axones sympathiques des
colonnes intermédio –latérales T1-L3 rejoignent la racine ventrale. Lorsque les
motoneurones passent à proximité des ganglions dorsaux, ceux-ci émettent 2
prolongements : 1 centripète (synapse avec neurone de la colonne dorsale) ; 1 centrifuge qui
23
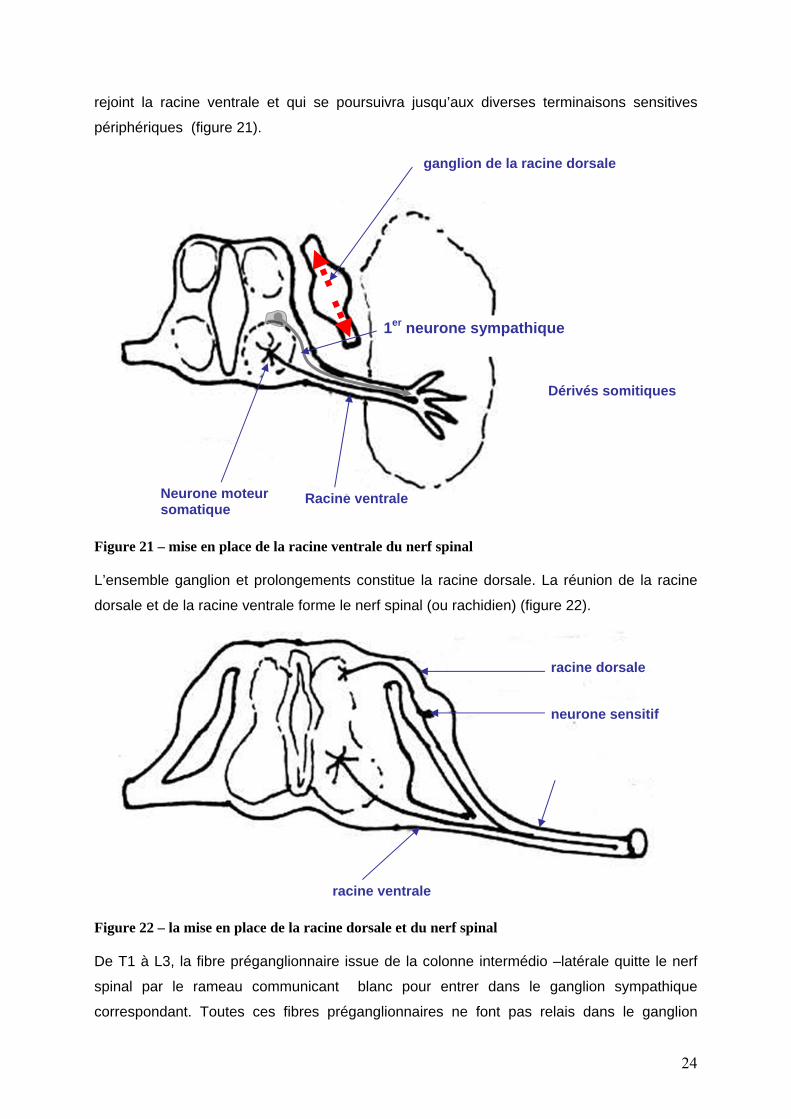
rejoint la racine ventrale et qui se poursuivra jusqu’aux diverses terminaisons sensitives
périphériques (figure 21).
Neurone moteur somatique
Racine ventrale
Dérivés somitiques
ganglion de la racine dorsale
1er neurone sympathique
Figure 21 – mise en place de la racine ventrale du nerf spinal L’ensemble ganglion et prolongements constitue la racine dorsale. La réunion de la racine
dorsale et de la racine ventrale forme le nerf spinal (ou rachidien) (figure 22).
racine ventrale
racine dorsale
neurone sensitif
Figure 22 – la mise en place de la racine dorsale et du nerf spinal De T1 à L3, la fibre préganglionnaire issue de la colonne intermédio –latérale quitte le nerf
spinal par le rameau communicant blanc pour entrer dans le ganglion sympathique
correspondant. Toutes ces fibres préganglionnaires ne font pas relais dans le ganglion
24
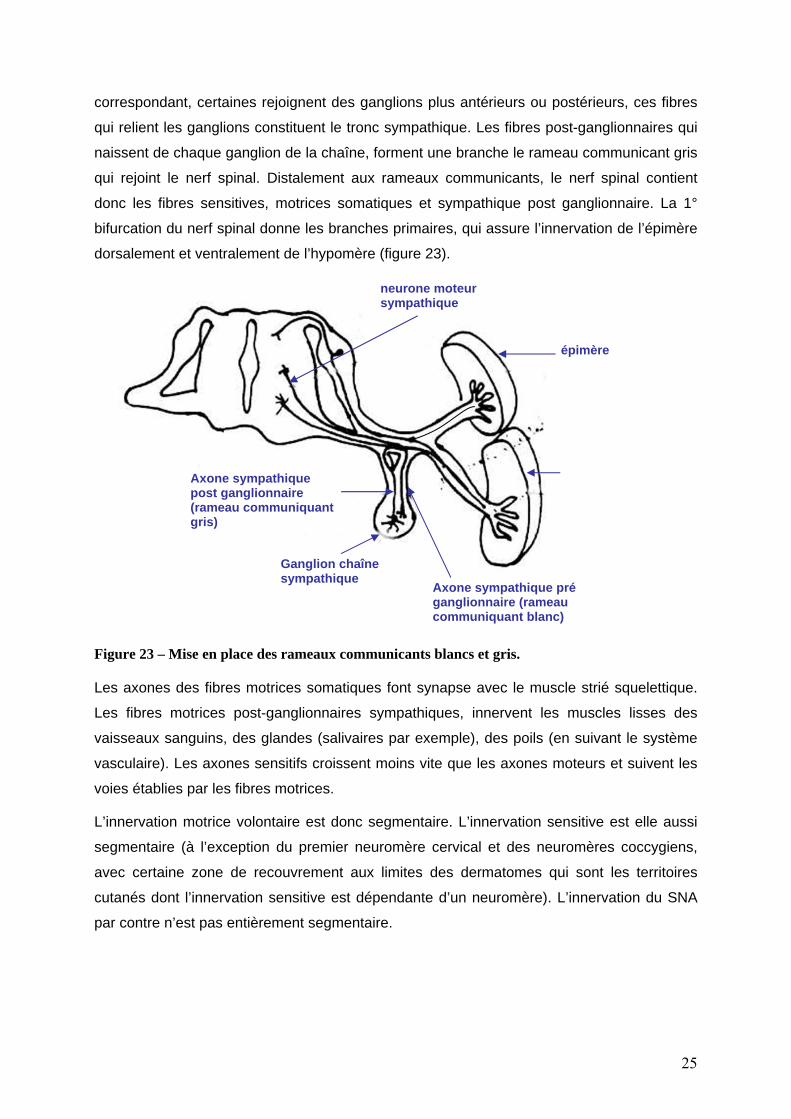
correspondant, certaines rejoignent des ganglions plus antérieurs ou postérieurs, ces fibres
qui relient les ganglions constituent le tronc sympathique. Les fibres post-ganglionnaires qui
naissent de chaque ganglion de la chaîne, forment une branche le rameau communicant gris
qui rejoint le nerf spinal. Distalement aux rameaux communicants, le nerf spinal contient
donc les fibres sensitives, motrices somatiques et sympathique post ganglionnaire. La 1°
bifurcation du nerf spinal donne les branches primaires, qui assure l’innervation de l’épimère
dorsalement et ventralement de l’hypomère (figure 23).
neurone moteur sympathique
Axone sympathique pré ganglionnaire (rameau communiquant blanc)
Ganglion chaîne sympathique
Axone sympathique post ganglionnaire (rameau communiquant gris)
épimère
Figure 23 – Mise en place des rameaux communicants blancs et gris. Les axones des fibres motrices somatiques font synapse avec le muscle strié squelettique.
Les fibres motrices post-ganglionnaires sympathiques, innervent les muscles lisses des
vaisseaux sanguins, des glandes (salivaires par exemple), des poils (en suivant le système
vasculaire). Les axones sensitifs croissent moins vite que les axones moteurs et suivent les
voies établies par les fibres motrices.
L’innervation motrice volontaire est donc segmentaire. L’innervation sensitive est elle aussi
segmentaire (à l’exception du premier neuromère cervical et des neuromères coccygiens,
avec certaine zone de recouvrement aux limites des dermatomes qui sont les territoires
cutanés dont l’innervation sensitive est dépendante d’un neuromère). L’innervation du SNA
par contre n’est pas entièrement segmentaire.
25

La migration des cellules des crêtes neurales au niveau céphalique.
A la différence du niveau médullaire, les cellules des crêtes neurales céphaliques débutent
leur migration probablement avant même que le tube neural ne soit complètement fermé.
Elles contribuent à la formation de 2 structures ganglionnaires (et non pas de 4 comme au
niveau médullaire) : 1) les ganglions sensitifs des nerfs crâniens, et 2) les ganglions
parasympathiques des nerfs crâniens (figure 24). Enfin elles forment : 3) les cellules
mésenchymateuses du mésectoderme.
R 1
R 2
R 3
R 4
R 5
R 6
R 7
R 8
M1 M2
Ggls. Sensitifs N. crâniens Ggls. Para Σ des N crâniens
V Trijumeau
VII Facial
IX Glosso- pharyngien
X Vague
Ggls. Ciliaires m.Sphincter pupille
Ggls Ptérygo- palatins Gl. lacrymales
Ggls Submandibulaires Gl. Salivaires sous-max et sub linguales
Ggls. Otiques Gl. parotide
Ggls. entériques
III Oculomoteur
VII
IX
X
Nx Pupillaires
Nx lacrymaux
Nx salivaires sup.
Nx salivaires inf
Figure 24 – Les dérivés ganglionnaires des crêtes neurales céphaliques.
1) Les cellules des ganglions sensitifs des nerfs crâniens sont issues des crêtes neurales
rhombencéphaliques: au niveau du 2ème rhombomère (R2) elle forme les ganglions sensitifs
du nerf Trijumeau, nerf du 1er arc branchial ; au niveau R4, les ganglions sensitifs du nerf
facial, nerf du 2ème arc branchial ; au niveau R6 les ganglions sensitifs du nerf glosso-
pharyngien, nerf du 3ème arc branchial ; et au niveau R8 les ganglions sensitifs du nerf
Vague, nerf du 4ème arc branchial.
2) Les cellules des ganglions parasympathiques des nerfs crâniens proviennent des crêtes
neurales mésencéphaliques et rhombencéphaliques. Leur innervation est assurée par les
26

premiers neurones préganglionnaires parasympathiques issus des noyaux viscéro –moteurs
des nerfs crâniens.
Les cellules des crêtes neurales mésencéphaliques migrent pour former les : Ganglions
parasympathiques ciliaires (se projetant sur muscle sphincter de la pupille), et dont
l’innervation préganglionnaire se fait à partir des noyaux pupillaires (ou noyaux oculo –
moteur accessoire) de la III° Paire.
Les cellules des crêtes neurales rhombencéphaliques migrent pour former, Au niveau du 4°
rhombomère : les ganglions ptérygopalatins (ou sphénopalatins) se projetant sur les glandes
lacrymales et les muqueuses des fosses nasales, du voile et du pharynx, et dont
l’innervation préganglionnaire se fait à partir des noyaux lacrymaux de la VII° paire ; les
Ganglions submandibulaires (ou sous maxillaires) se projetant sur les glandes salivaires
sous maxillaires et sublinguales et dont l’innervation préganglionnaire se fait à partir des
noyaux salivaires supérieures de la VII° paire.
Au niveau du 6° rhombomère elles forment les ganglions parasympathiques otiques se
projetant sur la glande parotide et dont l’innervation préganglionnaire se fait à partir des
noyaux salivaires inférieurs de la IX° paire.
Au niveau du 8° rhombomère elles forment les ganglions parasympathiques se projetant sur
les viscères thoraco -abdominaux et dont l’innervation préganglionnaire se fait à partir des
noyaux dorsaux de la X° paire.
3) Alors qu’au niveau médullaire les dérivés non neuraux des crêtes neurales étaient
restreints aux mélanocytes, au niveau des crêtes neurales céphaliques les cellules des
crêtes neurales forment les cellules mésenchymateuses du mésectoderme qui contribueront
à la formation du derme et hypoderme de la face et du cou, du tissu conjonctif entourant l’œil
ainsi que les muscles pupillaires et ciliaires, les cartilages des arcs branchiaux, les
odontoblastes, le septum aortico-pulmonaire, l’arachnoïde et la pie-mère.
Le développement de l’encéphale au delà du stade phylotypique.
Mise en place des différents segments encéphaliques
A 28 jours, on atteint le stade dit 5 vésicules, du fait de la segmentation du prosencéphale en
3 segments distincts : le diencéphale et les cavités télencéphaliques.
Deux conceptions de subdivision du télencéphale coexistent actuellement. Une théorie
classique selon laquelle le sulcus limitans s’interrompt rostralement au niveau du
mésencéphale. Dans cette version, au niveau du prosencéphale apparaissent 2 expansions
27

antérolatérales, les 2 vésicules télencéphaliques ou hémisphères cérébraux primitifs au
niveau desquelles se développaient les bulbes olfactifs. Ces bulbes olfactifs étaient
considérés comme une expansion terminale du système nerveux. Le reste du prosencéphale
devenait dès lors le diencéphale ou cerveau intermédiaire au niveau duquel naissaient les
vésicules optiques ; l’épiphyse, au niveau du toit ; le thalamus ; l’hypothalamus ; la post
hypophyse, au niveau du plancher.
Dans la conception moderne, le prosencéphale se subdivise en diencéphale pour les
premiers prosomères (au niveau duquel se développent dans la partie alaire : l’épiphyse ou
corps pinéal au niveau du toit, l’épithalamus, le thalamus dorsal et dans la partie basale : le
thalamus ventral, l’hypothalamus. Les derniers prosomères sont à l’origine du prosencéphale
secondaire, au niveau duquel se développent dans la partie alaire les vésicules
télencéphaliques, les vésicules optiques et dans la partie basale la post hypophyse (figure
25). Dans cette nouvelle conception la partie terminale du système nerveux est très
logiquement la lame terminale, et les lobes olfactifs ne sont qu’un des territoires fonctionnels
des vésicules télencéphaliques. Cette nouvelle description, dans la mesure où elle permet
de mieux systématiser l’architecture fonctionnelle du système nerveux devient prédominante,
il persiste toutefois quelques points toujours en cours d’investigation : la situation du sulcus
limitans dans les régions les plus rostrales du prosencéphale, mais aussi la signification
fonctionnelle du sulcus limitans dans le prosencéphale.
28

Figure 25 – L’encéphale à 30 Jours de développement embryonnaire. Représentation schématique des 5 vésicules ou segments dans la conception moderne. De la région caudale vers la région rostrale on distingue le rhombencéphale, le mésencéphale, le diencéphale qui s’est différencié à partir des 4 premiers prosomères (on rappelle que les prosomères sont numérotés dans le sens caudo-rostral), et le prosencéphale secondaire. Au niveau de la région alaire du prosencéphale secondaire naissent les 2 vésicules télencéphaliques. La position du sulcus limitans est schématisée en tirets discontinus noirs, la position de la plaque prochordale et de la chorde en rouge. Deux courbures supplémentaires se sont constituées, les courbures rhombiques et cervicales. En pointillé cyan est représenté le plan de section de l’encéphale qui permet la représentation de la figure 26. A ce stade, pour représenter les cavités de l’ensemble des segments de l’encéphale il
devient nécessaire de procéder à une représentation aplatie de l’encéphale sectionné selon
une surface plane suivant le sulcus limitans pour le rhombencéphale, le mésencéphale, et le
diencéphale, mais qui doit s’en éloigner pour la partie la plus rostrale du prosencéphale
secondaire. Ce plan de coupe a été figuré, dans sa portion s’éloignant du sulcus limitans, en
pointillé cyan dans la figure 25, il passe par la vésicule optique et les cavités
télencéphaliques (figure 26).
Figure 26 – La lumière du tube neural à 30 j. La lumière médullaire est le canal de l’épendyme, la lumière du rhombencéphale est le 4ème ventricule, du mésencéphale est l’aqueduc de Sylvius, du diencéphale et du prosencéphale secondaire le 3ème ventricule qui communique par les foramen interventriculaires latéraux (ou trou de Monroe) avec les cavités télencéphaliques ou ventricules latéraux. L’ébauche optique est une évagination de la région alaire du prosencéphale secondaire, sa lumière est encore présente à ce stade du développement qui est celui de la cupule optique, elle est en continuité directe avec la lumière du 3ème ventricule. La partie rostrale et médiale du prosencéphale secondaire est la lame terminale, elle est limitée latéralement par les pédoncules olfactifs.
29

A 35 jours de développement embryonnaire, le 1er rhombomère est à l’origine du
métencéphale et du cervelet. Et les 7 rhombomères suivants forment le myélencéphale
(figure 27).
Figure 27 – Vue schématique latérale de l’encéphale à 35 jours de développement embryonnaire. D’une façon très schématique on peut faire un parallèle entre la structuration ascendante du
SNC et l’évolution.
Au niveau médullaire sont retrouvées les structures nerveuses intervenant dans la motricité,
l’exemple le plus simple étant celui de l’arc réflexe. Toutefois, dès le niveau médullaire, il
faut introduire la notion de générateur central de pattern d’activation musculaire spatial et
temporel (CPG). L’exemple le plus simple est celui du réflexe de flexion de l’avant bras
mettant en jeu à la fois la contraction du biceps et le relâchement du triceps. Dans la mesure
où le relâchement ne peut pas provenir du largage d’un inhibiteur de la jonction neuro-
musculaire, la seule possibilité est un modèle à 4 populations de neurones : 1) un groupe de
neurones fléchisseurs ; 2) un groupe de neurones extenseurs ; 3) un groupe d’interneurones
excitateurs ; 4) un groupe d’interneurones inhibiteurs. Les 2 derniers appartiennent
proprement dit au CPG. Les données de l’examen neurologique et des explorations
neurophysiologiques montrent au niveau médullaire l’existence d’une hiérarchie de CPG
(exemple du réflexe bicipital, mais aussi du retrait de tout le bras après piqûre de l’extrémité,
30
et encore de la coordination des 4 membres pendant la marche). Il faut toutefois souligner
que la mise en évidence anatomique de ces réseaux de neurones reste très difficile et
fragmentaire.
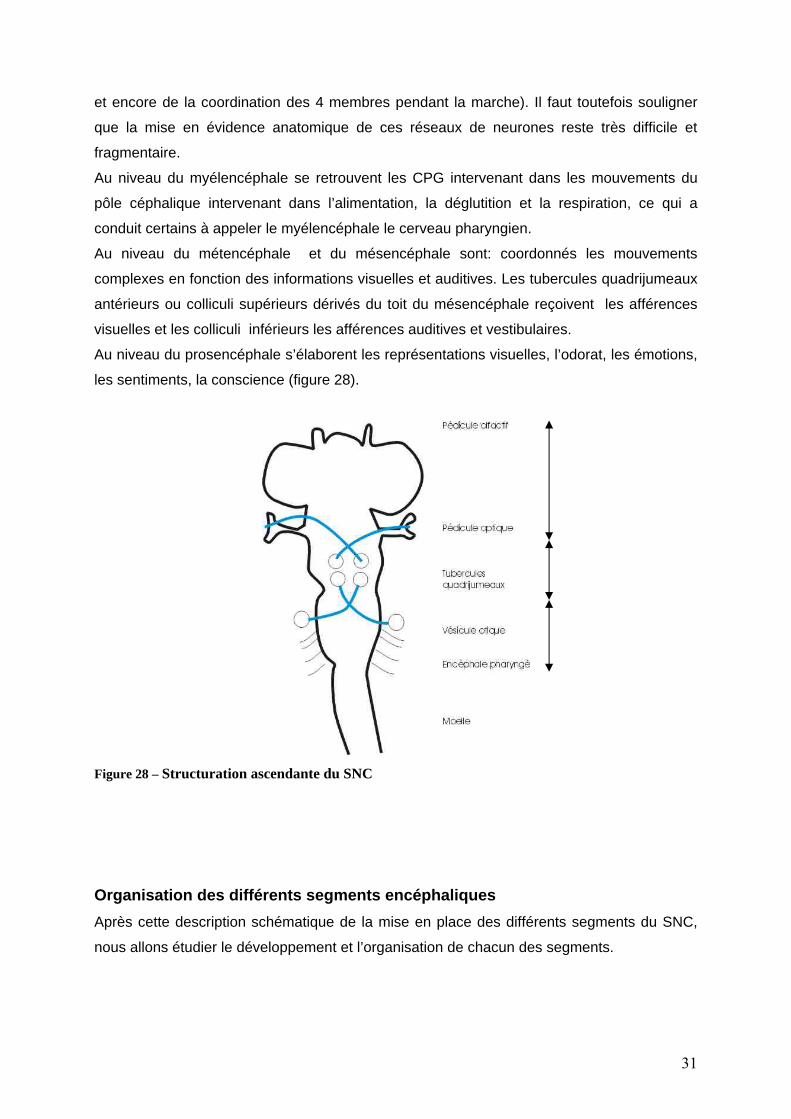
Au niveau du myélencéphale se retrouvent les CPG intervenant dans les mouvements du
nnés les mouvements
rat, les émotions,
igure 28 – St
rganisation des différents segments encéphaliques différents segments du SNC,
pôle céphalique intervenant dans l’alimentation, la déglutition et la respiration, ce qui a
conduit certains à appeler le myélencéphale le cerveau pharyngien.
Au niveau du métencéphale et du mésencéphale sont: coordo
complexes en fonction des informations visuelles et auditives. Les tubercules quadrijumeaux
antérieurs ou colliculi supérieurs dérivés du toit du mésencéphale reçoivent les afférences
visuelles et les colliculi inférieurs les afférences auditives et vestibulaires.
Au niveau du prosencéphale s’élaborent les représentations visuelles, l’odo
les sentiments, la conscience (figure 28).
F ructuration ascendante du SNC
OAprès cette description schématique de la mise en place des
nous allons étudier le développement et l’organisation de chacun des segments.
31

Figure 29 – Vue latérale schématique du tube neural à 35 J, sont représentés aussi les cavités ventriculaires du rhombencéphale et du mésencéphale, ainsi que les plans des coupes transverses 30, 31 et 32. A) myélencéphale ; B) métencéphale ; C) mésencéphale. Abréviations : IV, quatrième ventricule ; AS, aqueduc de Sylvius. Le rhombencéphale Il est dans la continuité de la moelle, et présente la même architecture générale avec le
sulcus limitans séparant les colonnes fondamentales et alaires. Toutefois, par rapport aux
nerfs spinaux, les fonctions des nerfs crâniens se diversifient, ils doivent en particulier
assurer la commande des structures dérivées des arcs branchiaux, d’où une certaine
complexification. Les parois du rhombencéphale s’écartent pour former un 4° ventricule
évasé, muni d’un toit mince constitué essentiellement par de l’épendyme recouvert par le
feuillet vascularisé de la méninge la pie-mère qui s’invagine pour donner le plexus choroïde
du 4° ventricule (figure 30).
Figure 30 – A. Schéma hémi-coupe transversale du rhombencéphale. Les colonnes des noyaux des paires crâniennes sont du plancher vers le sulcus limitans : la colonne efférente somatique (ES) ; la colonne efférente branchiale (EB) ou efférente viscérale spéciale (EVS) ; la colonne efférente viscérale (EV) ; du
32

sulcus limitans vers le toit ; la colonne afférente viscérale générale (AVG) ; la colonne afférente viscérale spéciale (AVS) appelé aussi afférente branchial spéciale (ABS) sensibilité gustative ; la colonne afférente somatique générale (ASG) ; la colonne afférente somatique spéciale (ASS). Le noyau olivaire a son origine au niveau des colonnes alaires et migre secondairement en position ventrale. Les noyaux des nerfs crâniens dans le rhombencéphale peuvent être schématiquement
réunis en 7 colonnes chacune d’elle ayant sous sa dépendance une fonction spécifique.
Depuis le plancher vers le toit on distingue : La colonne efférente somatique (muscle strié de
la langue et extrinsèque de l’œil) ; La colonne efférente branchiale (muscle strié branchial) ;
La colonne efférente viscérale parasympathique ; La colonne afférente ou sensitive viscérale
générale (sensibilité intéroceptive du vague) ; La colonne afférente branchiale spéciale
(bourgeons du goût) ; La colonne afférente somatique générale (sensibilité de la peau et
des muqueuses) ; La colonne afférente somatique spéciale (audition, équilibre, noyaux
vestibulo-cochléaire du VIII).
A partir de cette organisation commune à l’ensemble du rhombencéphale, des structures
spécifiques se différencient, par 2 mécanismes essentiels : 1) la migration de neurones des
colonnes alaires en direction ventrale ; et 2) le développement du bourrelet tectal qui est la
région qui constitue la jonction entre le toit (ou tectum) et la lame alaire.
Ainsi, au niveau du myélencéphale, on peut décrire la formation des noyaux olivaires dérivés
de la lame alaire qui interviennent dans le contrôle moteur et l’équilibre.
Et au niveau du métencéphale, la formation des noyaux pontins et du cervelet (figure 31).
Figure 31 – Schéma coupe transversale du métencéphale (coupe B du schéma 29).
33
Les noyaux pontins dérivent de la lame alaire, ils constitue un relais des signaux entre
moelle épinière, cortex cérébral et cérébelleux et sont localisés dans le pont ou
protubérance.
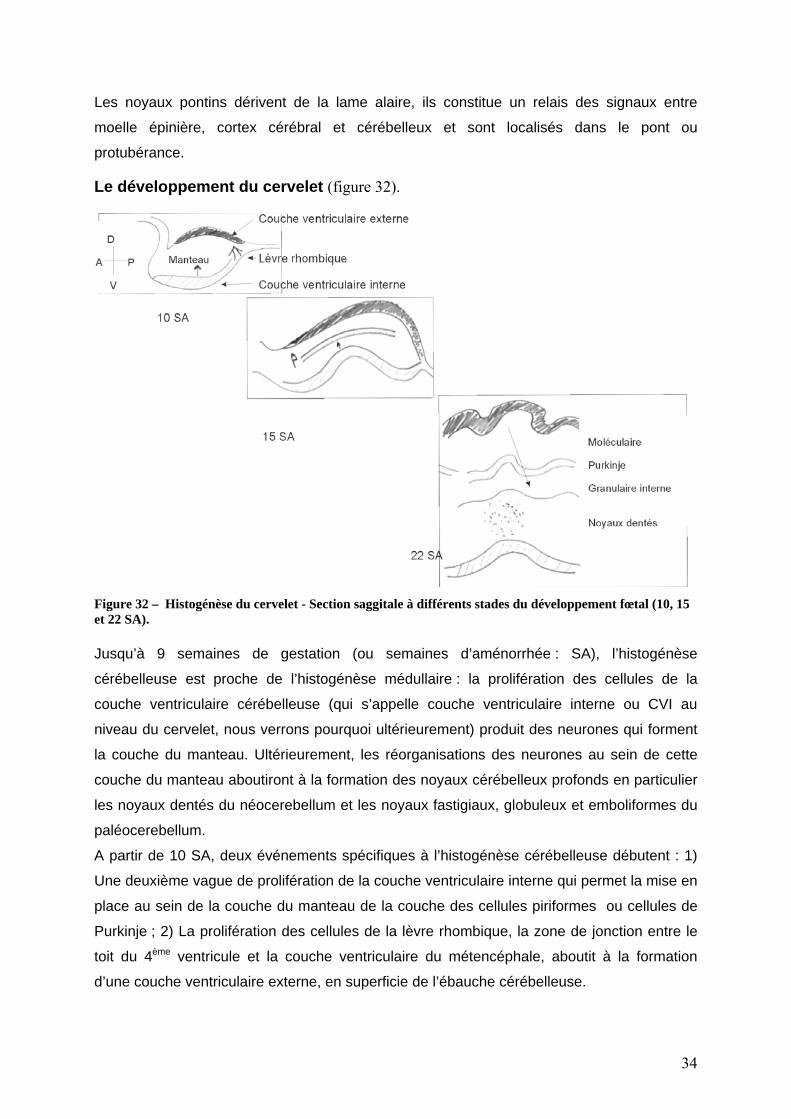
Le développement du cervelet (figure 32).
Figure 32 – Histogénèse du cervelet - Section saggitale à différents stades du développement fœtal (10, 15 et 22 SA). Jusqu’à 9 semaines de gestation (ou semaines d’aménorrhée : SA), l’histogénèse
cérébelleuse est proche de l’histogénèse médullaire : la prolifération des cellules de la
couche ventriculaire cérébelleuse (qui s’appelle couche ventriculaire interne ou CVI au
niveau du cervelet, nous verrons pourquoi ultérieurement) produit des neurones qui forment
la couche du manteau. Ultérieurement, les réorganisations des neurones au sein de cette
couche du manteau aboutiront à la formation des noyaux cérébelleux profonds en particulier
les noyaux dentés du néocerebellum et les noyaux fastigiaux, globuleux et emboliformes du
paléocerebellum.
A partir de 10 SA, deux événements spécifiques à l’histogénèse cérébelleuse débutent : 1)
Une deuxième vague de prolifération de la couche ventriculaire interne qui permet la mise en
place au sein de la couche du manteau de la couche des cellules piriformes ou cellules de
Purkinje ; 2) La prolifération des cellules de la lèvre rhombique, la zone de jonction entre le
toit du 4ème ventricule et la couche ventriculaire du métencéphale, aboutit à la formation
d’une couche ventriculaire externe, en superficie de l’ébauche cérébelleuse.
34

Enfin à partir de 16 SA, cette couche ventriculaire externe produit des cellules qui migrent
vers l’intérieur et forment la couche granulaire interne. On aboutit ainsi à la disposition finale
avec du centre vers la périphérie l’épendyme, la substance blanche avec en son sein les
noyaux profonds, puis la couche granulaire du cortex, la couche des cellules piriformes, la
couche moléculaire du cortex.
Cette zone matrice externe persistera jusqu’à la fin de la 2° année de vie post-natale (les
cellules qui persisteraient seraient à l’origine des médulloblastome).
Digression 2 – Cervelet, relation structure fonction. Des données expérimentales récentes ( R Thompson) nous aident à comprendre les liens
qui s’établissent entre la structure et la fonction du cervelet.
Les fibres de Purkinje sont en position idéale pour comparer les influx nerveux en
provenance des fibres moussues et grimpantes. Dans une expérience de conditionnement
positif, un stimulus inconditionnel un jet d’air sur la cornée est précédé de peu par l’envoi
d’un stimulus conditionnel sous la forme d’un « son de cloche », aboutissant par
apprentissage à un effet de protection par clignement préventif de la paupière. Les fibres
moussues transmettent le stimulus inconditionnel vers le noyau profond, et les fibres
grimpantes le stimulus conditionnel vers le noyau profond. Avant l’entraînement le signal des
fibres grimpantes (stimulus conditionnel, son de cloche) n’est pas suffisant pour déclencher
une réponse au niveau des noyaux, seul le stimulus inconditionnel permet au noyau profond
de déclencher une réponse qui sera véhiculée par les pédoncules cérébelleux aux
générateurs et initiateurs centraux de pattern du tronc cérébral et de la moelle ainsi qu’aux
hémisphères cérébraux (le système cognitif) via un relais thalamique. Puis au cours de
l’apprentissage les fibres de Purkinje interviennent pour renforcer la réponse jusqu’à ce que
le stimulus conditionnel déclenche la réponse (figure 33).
35

♪
Figure 33 – Diagramme du circuit élémentaire du cervelet.
Ces données associées aux données cliniques nous font progresser dans la compréhension
de la fonction du cervelet qui joue un rôle dans l’apprentissage et la coordination fine des
centaines de muscles impliqués dans les réponses orientées, la manipulation, la posture. Il
reçoit tous les types d’informations sensitives, directement de la moelle épinière et du tronc
cérébral ou indirectement du cortex cérébral, (par les fibres moussues de la substance grise
du pont). Le cervelet conserve essentiellement la trace mnésique des programmes moteurs
acquis.
Ceci explique aussi les données chirurgicales qui montrent que les agressions chirurgicales
du cortex cérébelleux sont assez bien tolérées cliniquement hormis les capacités
d’apprentissage moteur, alors que les noyaux cérébelleux eux sont très sensibles2.
Le mésencéphale Au niveau du mésencéphale, les dérivés de la colonne alaire sont les tubercules
quadrijumeaux (colliculi supérieurs et inférieurs) et la substance noire (migration ventrale).
Les dérivés de la colonne fondamentale, sont les noyaux viscéromoteurs des muscles
pupillaires et ciliaires ainsi que les noyaux rouges (figure 34).
2 La distinction des très nombreuses structures du vermis n’a que peu d’intérêt pratique, d’un point de vue clinique on distingue le vestibulo-cérebellum (afférences ces noyaux vestibulaires du tronc cérébral - hypertonie de décérébration), le spino-cérebellum vermien, le spino-cérebellum intermédiaire (afférences de la moelle - tremblement, dysmétrie, hypermétrie) et le cérébro-cérebellum (afférences du cortex cérébral - adiadococinésie).
36

Figure 34 – Schéma coupe transversale du mésencéphale
Le prosencéphale L’étude de la systématisation du prosencéphale est plus complexe, en effet les
remaniements tridimensionnels rendent plus difficilement perceptible la systématisation que
nous avions cherché à mettre en évidence au niveau du rhombencéphale et du
mésencéphale. Pour s’affranchir de ces difficultés de représentation, nous allons remonter
au tout début du développement su système nerveux, au stade de la plaque neurale. En
effet dès ce stade, il est possible de tracer une carte des territoires présomptifs3
embryonnaires (figure 35). Cette carte à ce stade est précieuse car elle permet de mettre en
évidence toutes les régions embryonnaires qui interviendront dans la formation du système
nerveux : 1) la plaque neurale proprement dite. A son niveau, il est dès ce stade possible de
reconnaitre les principales subdivisions que nous avons déjà décrites, la place du sulcus
3 La carte des territoires présomptifs d’un embryon indique le devenir des cellules lors du développement embryonnaire normal. Pour établir une carte des territoires présomptifs, il faut marquer au stade embryonnaire étudié les différentes régions avec un marqueur le plus stable possible, interférant le moins possible avec le développement normal, puis étudier la localisation de ces marques aux stades ultérieurs de l’embryon puis de l’adulte. La carte peut être établie lorsque l’on met en évidence de façon reproductible une relation entre le marquage embryonnaire et la localisation finale, ce qui implique qu’il n’y ait pas eu trop de brassage des cellules. Une analogie peut être faite entre ces cartes des territoires présomptifs et la carte des horaires de chemin de fer. La carte donne en fonction de leur localisation à l’instant t0, la destination des trains à l’instant t1. C’est ce qui est sensé ce passé si tout se passe bien. Dans le système nerveux comme dans le train, en cas d’avarie sur une voie ou sur une machine, de nouveaux itinéraires de détournement (de suppléance) vont pouvoir se mettre en place.
37
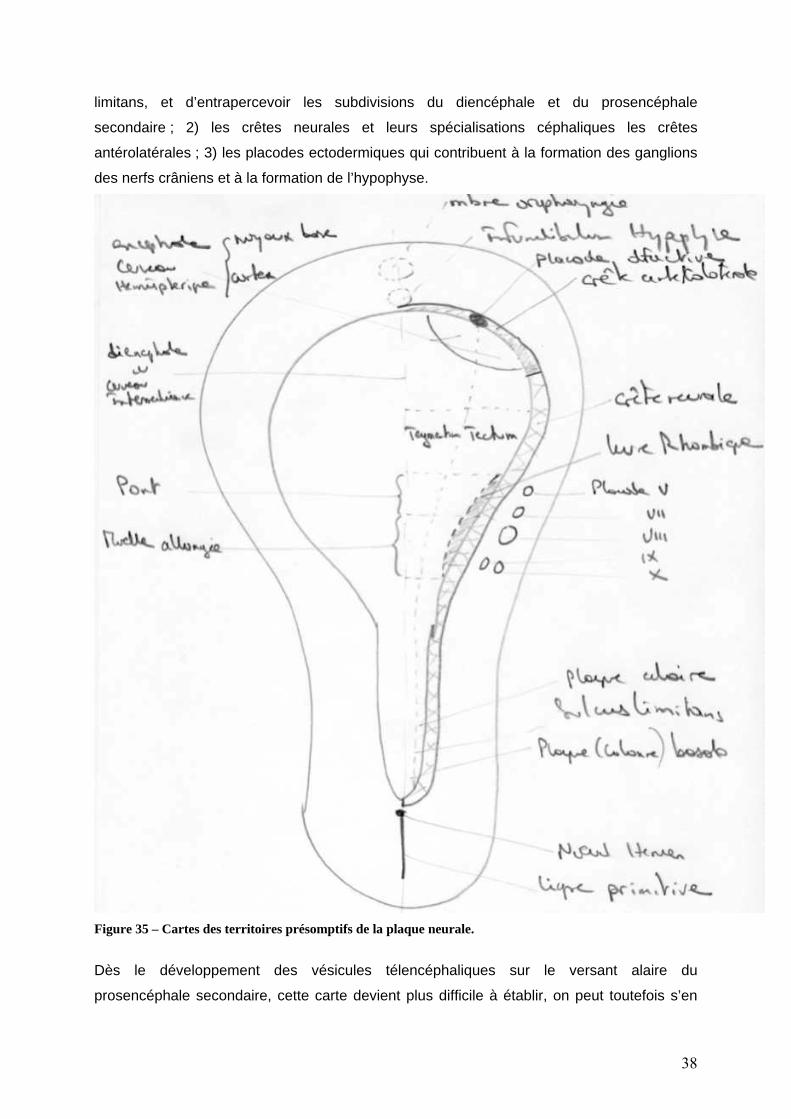
limitans, et d’entrapercevoir les subdivisions du diencéphale et du prosencéphale
secondaire ; 2) les crêtes neurales et leurs spécialisations céphaliques les crêtes
antérolatérales ; 3) les placodes ectodermiques qui contribuent à la formation des ganglions
des nerfs crâniens et à la formation de l’hypophyse.
Figure 35 – Cartes des territoires présomptifs de la plaque neurale.
Dès le développement des vésicules télencéphaliques sur le versant alaire du
prosencéphale secondaire, cette carte devient plus difficile à établir, on peut toutefois s’en
38

rapprocher en utilisant une représentation aplatie, établie en sectionnant médialement le toit
du tube neural et en projetant les moitiés latérales sur le plan (figure 36).
Figure 36 – Représentation aplatie du système nerveux au-delà du stade 5 vésicules.
Le diencéphale
Au niveau du diencéphale le sulcus limitans4 partage le diencéphale en sa partie alaire et
basale au niveau du thalamus, puis 2 sillons supplémentaires se constituent de part et
d’autre du sillon médian, d’abord le sillon hypothalamique, puis le sillon habénulaire pour
aboutir finalement à la disposition définitive en 4 régions longitudinales, de la région dorsale
vers la région ventrale, l’épithalamus, le thalamus dorsal, le thalamus ventral et
l’hypothalamus.
Digression 3 – Générateurs, initiateurs et contrôleurs de patterns. Nous avons déjà introduit, le concept de CPG (générateur central de pattern), en insistant
sur leur hiérarchisation. A ce stade de l’enseignement, il devient utile de mieux systématiser
cette notion (figure 37).
4 Signalons que certains auteurs préfèrent au niveau du diencéphale appelé sillon médian cette subdivision longitudinale, car ils émettent des doutes sur son analogie complète avec le sulcus limitans observé au niveau du mésencéphale et du rhombencéphale.
39

Figure 37 – Diagramme des générateurs, initiateurs et contrôleurs de pattern. Place du cervelet en
dérivation.
Au total, l’organisation du système moteur est hiérarchisée avec à la base des ensembles de
motoneurones bien caractérisés. Les circuits de contrôle restent eux mal définis et on en est
réduit à un modèle, dont l’idée est la suivante : un générateur primaire de pattern central
contrôle un groupe de motoneurones et produit un comportement spécifique en induisant
une réponse motrice (flexion de l’avant bras mettant en jeu contraction du biceps et
relâchement du triceps). Un générateur secondaire innerve un ensemble de générateurs
primaires pour produire un comportement coordonné plus vaste (exemple flexion de
l’ensemble du bras et de l’avant bras). Les CPG peuvent être activés autrement que par des
réflexes spinaux, ceci peut être mis en évidence par les expériences de stimulation de la
région locomotrice du mésencéphale qui entraîne l’activation des CPG d’aval sans
stimulation de l’arc réflexe spinal. Les régions stimulées responsables de l’activation des
CPG sont les initiateurs centraux de patterns (CPI). Ces initiateurs de pattern central sont
eux-mêmes sous le contrôle de contrôleurs centraux de pattern (CPC) qui sont capables de
conduire l’activité motrice en rapport avec certains comportements instinctuels. Les
contrôleurs sont eux mêmes hiérarchisés, avec au sommet un contrôleur au niveau de la
région motrice hypothalamique. On est souvent surpris par la complexité des comportements
sous le contrôle de ces CPC. Ainsi, les noyaux médians de l’hypothalamus sont des
contrôleurs pour 3 comportements motivés : alimentation (boire et manger), défense (attaque
ou fuite), reproduction (sexualité et parentalité). Ces derniers ont particulièrement été
explorés par les éthologistes (Nikolaas Timbergen, prix Nobel de médecine en 1973) chez
l’épinoche (Gasterosteus aculeatus). Il a ainsi distingué au sein de l’instinct (ou
40

comportement motivé) de reproduction 4 types de comportement stéréotypés : 1) combat ;
2) construction du nid; 3) fécondation ; 4) soins à la portée (figure 38).
Figure 38 – Instinct de reproduction de l’épinoche – les différents types de comportement stéréotypés.
Une expérience fondamentale souligne le rôle de l’hypothalamus dans ces comportements
stéréotypés complexes. On peut retirer tout le prosencéphale à l’exception de cette région et
on obtient toujours une activité spontanée ou intrinsèque.
Vous remarquerez enfin la place accordée au cervelet dans ce diagramme. La coordination
et l’apprentissage moteur sont modulés par le cervelet, qui apparaît être plus en parallèle
sur l’ensemble des structures précédemment décrites que hiérarchiquement situé.
Bien entendu ces instincts ou comportements motivés peuvent aussi être activés de façon
consciente, on rentre alors dans le royaume du prosencéphale secondaire.
Le prosencéphale secondaire – les hémisphères cérébraux- La pensée et le
contrôle volontaire de la motricité
Récapitulatif, vous avez probablement remarqué qu’entre le schéma de la figure 35 et celui
de la figure 36, qui sont si utiles pour comprendre le plan général du système nerveux en
développement, il y a eu un sérieux raccourci. C’est cette lacune que nous allons combler
maintenant en nous focalisant sur le développement de ce segment tout en restant fidèle à
notre parti pris de représentation schématique et plane. Pour cela il nous faut revenir une
représentation proche de celle de la figure 26 (figure 39).
41

Figure 39 – Carte des territoires présomptifs du prosencéphale secondaire au stade 5 vésicules (30 jours). Lame terminale (Lam) ; éminence médiane (m) ; éminence latérale (l) ; foramen interventriculaire ou trou de Monroe (IVF) ; Pallidum (P) ; striatum (S) ; Cortex (CTX). Les principales subdivisions du prosencéphale sont reconnaissables dès ce stade. Les
cavités des vésicules télencéphaliques, les ventricules latéraux communiquent avec le 3ème
ventricule au niveau des foramen interventriculaires de Monroe (FIV). Au niveau des parois
des vésicules télencéphaliques, il est possible de distinguer 2 renflements de la paroi : les
éminences médiane et latérale. L’éminence médiane, futur Pallidum est séparée
médialement des parois du 3ème ventricule par les berges du FIV et latéralement de
l’éminence latérale par le sillon pallido-striatal. L’éminence latérale, futur Striatum, est séparé
des parois latérales des vésicules télencéphaliques, futur cortex ou Pallium, par le sillon
cortico-basal (ou bien encore cortico-striatal). Ainsi, à ce stade les relations entre le cortex
cérébral et les noyaux gris centraux le pallidum et le striatum sont on ne peu plus simple.
A ce stade aussi, ou le cortex de chaque hémisphère est limité à une demi-sphère, il est
encore simple « en se plaçant dans le 3ème ventricule » d’observer les cartes des territoires
présomptifs télencéphaliques qui préfigurent les principales subdivisions corticales (figure
40). Ultérieurement les processus de plissement, l’apparition des sillons corticaux qui
séparent les circonvolutions cérébrales, rendront plus difficile la reconnaissance des rapports
entre ces différents territoires corticaux, alors même que les rapports topologiques sont les
mêmes.
42

a
b
Figure 40 – Vue schématique depuis le 3ème ventricule du prosencéphale d’un embryon humain au stade 5 vésicules. ab limite inférieure du trou de Monro ; Pal pallidum ; S Striatum ; Ol lobe olfactif ; PF lobe préfrontal ; C lobe cingulaire ; H Hippocampe; I Insula ; FPOT : frontal, pariétal, occipital, temporal.
L’histogénèse du cortex cérébral
Nous avons déjà étudié l’histogénèse du cortex cérébelleux, la formation du cortex cérébral
est très différente hormis pour les premiers stades (figure 41). A ce niveau, deux
mécanismes distincts de mise en place des cellules neuronales doivent être étudiés : la
migration radiaire et la migration latérale.
La migration radiaire. Au niveau du cortex cérébral, comme au niveau du tube neural spinal,
les neurones naissent dans la couche ventriculaire par mitose asymétrique. Cette
prolifération est remarquablement prolongée dans l’espèce humaine de 6 SA à 30 SA.
Le premier temps spécifique est la mise en place de la pré-plaque majoritairement à partir
des cellules de la couche ventriculaires (8 à 9 semaines de développement embryonnaire
chez l’homme, 10 à 11 SA). Elle est constituée des neurones de Cajal-Retzius et d’un plexus
de fibres nerveuses. Les mutations inactivatrices du gène de la Reeline sont responsables
43

de sa non mise en place. Ultérieurement (semaines 10 à 18) les neuroblastes traversent les
couches profondes de la pré-plaque, et la scinde en 2 parties : une superficielle dite aussi
moléculaire, futur couche 1 du cortex, et une profonde la sous-plaque, qui a un rôle essentiel
dans la mise en place des projections thalamo-corticales. Ces cellules neuronales qui ont
traversées la sous-plaque forment la plaque corticale qui donne naissance aux couches 2 à
6. La formation de ces couches se fait par « inside-out layering », les cellules qui arrivent
tardivement passent à travers la couche des précédentes (figure 41).
La migration latérale mise en évidence récemment par John Rubenstein en 2001. A partir
des éminences médiales et latérales naissent des neuroblastes gabaergiques qui migrent
dans la sous-plaque et dans la plaque corticale.
Figure 41 – Histogénèse du cortex cérébral. Sur ce schéma de base de cortex à 6 couches, il existe de larges variations régionales. Et, il
n’est pas possible en 2009 de ne pas célébrer le 100ème anniversaire de la description par
Broadman sur des critères cyto-architectonique de 50 aires corticales distinctes.
Actuellement, selon les critères retenus, on distingue de 20 à 400 aires distinctes.
D’un point de vue fonctionnel, il est plus simple de distinguer 3 super couches :
Une supercouche granulaire correspondant à la couche 4 caractérisée par l’importance des
entrées en provenance du thalamus et essentiellement des sorties vers la couche
supragranulaire.
44

Une supercouche supra granulaire de petits neurones pyramidaux générant l’essentiellement
des réseaux de connections entre les aires corticales et des sorties vers la supercouche
infragranulaire. C’est à ce niveau que s’élabore essentiellement les processus cognitifs
(penser, apprendre, se souvenir).
Une supercouche infragranulaire de grands neurones pyramidaux se projetant vers les
noyaux cérébraux (au sens large, incluant le thalamus), le tronc cérébral et la moelle spinale,
en rapport donc avec la commande volontaire motrice. Si l’on s’intéresse aux détails des
projections de cette supercouche infra granulaire on peut noter que la couche 6 se projette
essentiellement vers le thalamus, et la couche 5 vers le striatum et la moelle spinale.
Le développement des noyaux gris centraux
L’étude du développement des noyaux gris centraux à de longue date été source de
difficultés et de confusion. Je n’ai pas de baguette magique pour éviter ce long travail qu’il
faudra pourtant mener à bien pour ceux d’entre vous qui souhaitent se diriger vers les
disciplines neurologiques et cognitives, qu’elles soient anatomiques, cliniques, radiologiques
ou biologiques. Fidèle à notre parti-pris nous soulignerons simplement dans ce cours
introductif, quelques avancées récentes qui vont dans le sens d’une simplification de la
compréhension du développement des noyaux gris centraux et de leur rapport avec le cortex
(figure 42).
Figure 42 – Croissance différentielle des noyaux gris centraux et du cortex cérébral. De gauche à droite représentation schématique d’un stade précoce (12 SA), intermédiaire et tardif. En a position du sillon cortico-striatal. Le pointillé sépare la couche ventriculaire (v) de la couche du manteau (m). A titre illustratif ont été représenté 2 faisceaux reliant les noyaux gris centraux et le cortex, le fornix (Fx) et le faisceau médian frontal (medial forebrain bundle – mfb). L’élément essentiel à prendre en compte est la croissance relative initiale plus rapide des
noyaux gris centraux par rapport au cortex, ce qui les amènent à faire protrusion dans la
45

cavité ventriculaire et à s’accoler au cortex. Signalons qu’il n’existe pas de consensus actuel
pour la dénomination de ces noyaux gris centraux, ceci est source de beaucoup de
confusions. Le point de vue simple de la biologie du développement est de distinguer entre
cortex et noyaux gris centraux, et parmi ces derniers entre striatum et pallidum. Les
structures dérivées du striatum et du pallidum sont distinctes selon leur localisation dorsale,
ventrale, médiale, et selon l’axe rostro-caudal. En figure 43 sont donnés les origines
embryologiques des différentes structures du striatum et du pallidum.
Dorsal Ventral Médial Caudorostral Striatum Putamen Noyau accumbens
Fundus striatal
Tubercule olfactif
Complexe septal latéral Noyaux amygdaliens
Pallidum Globus pallidus
Interne et externe
Substance innominée
Noyau magnocellulaire préoptique
Noyaux de la Bandelette diagonale
Noyaux de la strie terminale
Figure 43 –Différentes composantes du Striatum et du Pallidum en fonction de leur topographie. Ce point de vue embryologique est soutenu par les données histochimiques. Il est devenu en
effet clair que les cellules pyramidales qui assurent les projections du cortex utilisent le
neurotransmetteur excitateur glutamate, alors que les projections descendantes des noyaux
gris centraux utilisent le neurotransmetteur inhibiteur GABA. Au sein du cortex il existe par
ailleurs des circuits locaux gabaergiques, les cellules qui les composent proviennent des
éminences médiales et latérales par migration latérale.
A ce stade, il est utile de faire un nouveau point sur les relations entre structures et fonctions.
Digression 4 – relation structure fonction entre cortex – Noyaux gris centraux et segments d’aval (figure 44).
Figure 44 – Diagramme des circuits élémentaires entre cortex- Noyaux gris centraux et structures d’aval.
46

Projection descendante triple du cerveau.
Tout le cortex se projette, depuis la couche 5 essentiellement, sur le striatum. Ces neurones
pyramidaux glutamatergique excitateurs de la couche 5 ont des axones qui se projettent
dans le tronc cérébral et la moelle spinale, et donnent des collatérales au striatum.
Il existe aussi une projection de l’ensemble du striatum sur le pallidum. Les neurones du
striatum gabaergique inhibiteurs ont des axones qui se projettent sur la substance noire
mésencéphalique compacte et réticulée, en donnant des collatérales au niveau du Pallidum
(souvenons nous que la substance noire réticulée appartient à la colonne de contrôle
comportemental des mouvements d’orientation des yeux et de la tête).
Les neurones gabaergiques inhibiteurs du pallidum ont des axones qui se projettent dans le
tronc cérébral et la moelle spinale, en donnant des collatérales au niveau du Thalamus.
Dans la mesure où le pallidum qui émet des axones inhibiteurs reçoit des afférences
inhibitrices du striatum, on peut considérer qu’il à un effet désinhibiteur au niveau du
système moteur.
Notons aussi que ce triple système descendant à une boucle de rétrocontrôle thalamo-
corticale.
Nous n’irons pas plus loin, dans le cadre de cet exposé introductif, dans l’étude des
interactions entre toutes les composantes du prosencéphale. Ce domaine fait l’objet de
recherches médicales et scientifiques très actives, qui posent d’ailleurs d’intéressantes
questions philosophiques et éthiques (Bernard Baertschi, 2009). Sans aller plus dans le
détail, il est utile de prendre un peu de recul et de considérer le chemin parcouru (ou qui
devrait avoir été parcouru) entre notre épinoche de la page 40 et l’être humain. Chez
l’épinoche, les comportements restent essentiellement innés et commandés par les
contrôleurs de patterns centraux hypothalamiques ; chez l’être humain le développement
post-natal du SNC, dans le cadre d’interaction culturelles fortes, abouti à des
comportements acquis essentiellement conscients commandés par les dérivés
télencéphaliques. Si vous étiez une épinoche et qu’un congénère vous barre la route en
rentrant au nid, votre comportement serait assez stéréotypé et conduirait à l’attaque et la
morsure ; lorsque l’on vous fait une queue de poisson sur l’autoroute vous savez bien en
général vous raisonnez, ne pas foncez immédiatement sur ce crétin dangereux, ce qui ne
vous empêche pas d’élaborer dans votre esprit toutes sortes de pensées diverses émaillées
de différents noms d’oiseaux. C’est ça l’hominisation, enfin un bref aperçu ! C’est comment
la conscience prend progressivement le contrôle des CPC sous jacents, en inhibant dans
notre exemple ceux qui conduirait à l’attaque et à la morsure.
47

Le développement ultérieur des vésicules télencéphaliques La croissance ultérieure du télencéphale est marquée par la forte croissance relative des
vésicules télencéphaliques qui viennent recouvrir le diencéphale puis le mésencéphale et le
cervelet (figure 45).
Figure 45 – Développement des vésicules télencéphaliques. La double flèche représente le plan de coupe représenté sur la figure 46. Ce mouvement concerne aussi bien le cortex que les noyaux gris centraux et les faisceaux
qui assurent les liaisons entre les différentes structures dont nous avons étudiées la mise en
place. Ainsi, à ce stade une coupe transversale selon la double flèche figurée dans le
schéma 45 permet d’illustrer les relations entre le diencéphale et le prosencéphale
secondaire.
48

Figure 46 – Coupe frontale du prosencéphale avant la formation du corps calleux à 10 SA. La double flèche figure la zone de rapprochement des parois médiales des vésicules télencéphaliques au niveau de la formation du corps calleux. Les 2 doubles flèches pointillées les foramen interventriculaires de Monroe faisant communiquer le 3ème ventricule (III) et les ventricules latéraux (VL). A partir de 12 SA, la principale commissure inter hémisphérique, le corps calleux commence
à se mettre en place. L’espace délimité, par le toit du 3ème ventricule, les parois médiales des
ventricules latéraux (qui deviendront très fines, transparentes ont dit aussi pellucide) au sein
desquelles s’individualise les piliers du fornix, et le corps calleux, forme la cavité du septum
pellucidum. Ultérieurement, le striatum se divisera en ses 2 principaux constituants le Noyau
caudé et le putamen, les principaux sillons corticaux apparaissent (figure 47). Surtout la
croissance du télencéphale et son repliement aboutissent à des remaniements
tridimensionnels complexifiant la description des structures cérébrales, c’est le domaine de
la neuroanatomie. Nous ne prendrons qu’un exemple pour illustrer la différence des points
de vue. Les anatomistes réunissent le globus pallidus et le putamen au sein d’une structure
le noyau lenticulaire. C’est effectivement une structure clairement reconnaissable sur une
coupe frontale, bien distincte du noyau caudé. La connaissance de l’hétérogénéité de son
origine embryologique pallidum pour le globus pallidus et striatum pour le putamen permet
de bien saisir la structure des interactions qui se jouent entre ces 2 composants du noyau
lenticulaire (CF digression 4).
49
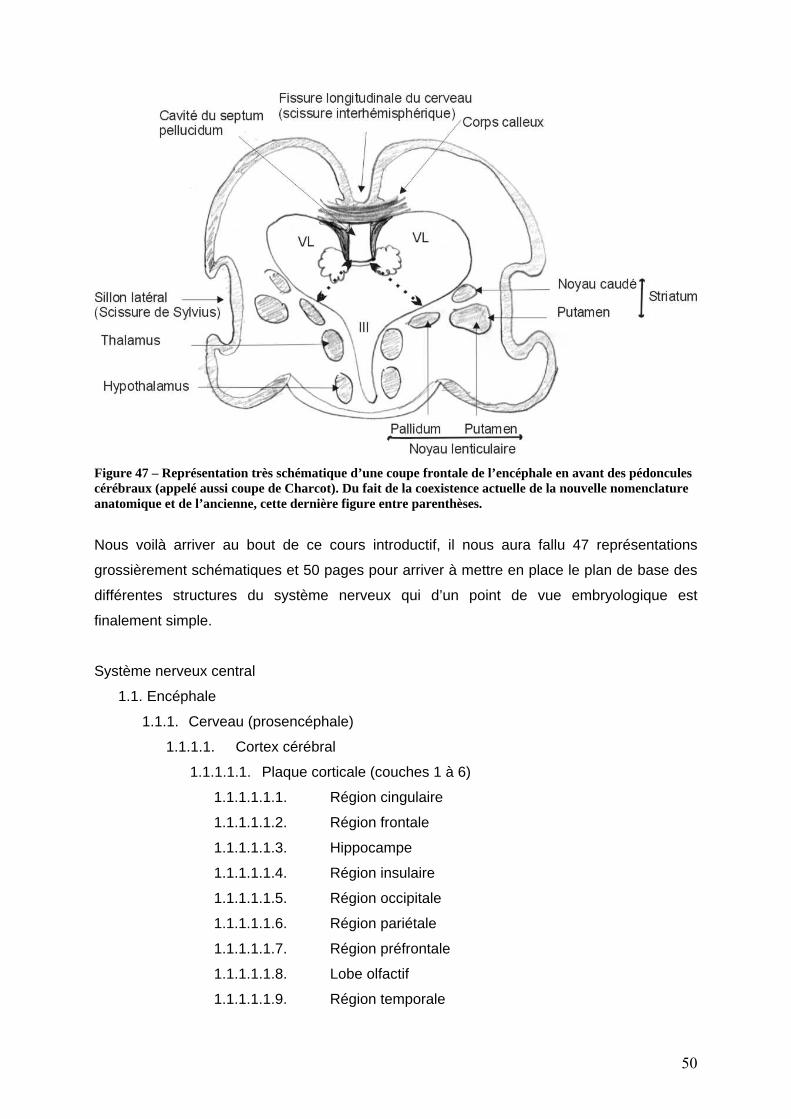
Figure 47 – Représentation très schématique d’une coupe frontale de l’encéphale en avant des pédoncules cérébraux (appelé aussi coupe de Charcot). Du fait de la coexistence actuelle de la nouvelle nomenclature anatomique et de l’ancienne, cette dernière figure entre parenthèses.
Nous voilà arriver au bout de ce cours introductif, il nous aura fallu 47 représentations
grossièrement schématiques et 50 pages pour arriver à mettre en place le plan de base des
différentes structures du système nerveux qui d’un point de vue embryologique est
finalement simple.
Système nerveux central
1.1. Encéphale
1.1.1. Cerveau (prosencéphale)
1.1.1.1. Cortex cérébral
1.1.1.1.1. Plaque corticale (couches 1 à 6)
1.1.1.1.1.1. Région cingulaire
1.1.1.1.1.2. Région frontale
1.1.1.1.1.3. Hippocampe
1.1.1.1.1.4. Région insulaire
1.1.1.1.1.5. Région occipitale
1.1.1.1.1.6. Région pariétale
1.1.1.1.1.7. Région préfrontale
1.1.1.1.1.8. Lobe olfactif
1.1.1.1.1.9. Région temporale
50

1.1.1.1.2. Sous-plaque (couche 7)
1.1.1.2. Noyaux gris centraux
1.1.1.2.1. Striatum
1.1.1.2.2. Pallidum
1.1.1.3. Cervelet
1.1.1.3.1. Cortex cérébelleux
1.1.1.3.1.1. Lobe antérieur
1.1.1.3.1.2. Lobe postérieur
1.1.1.3.1.3. Lobe floculo-nodulaire
1.1.1.3.2. Noyaux cérébelleux
1.1.1.4. Tronc cérébral
1.1.1.4.1. Diencéphale
1.1.1.4.1.1. Epithalamus
1.1.1.4.1.2. Thalamus dorsal
1.1.1.4.1.3. Thalamus ventral
1.1.1.4.1.4. Hypothalamus
1.1.1.4.2. Mésencéphale
1.1.1.4.2.1. Tectum
1.1.1.4.2.2. Tegmentum
1.1.1.4.2.3. Région prétectale
1.1.1.4.3. Rhombencéphale
1.1.1.4.3.1. Pont (protubérance)
1.1.1.4.3.2. Moelle allongée (bulbe)
1.2. Moelle spinale
1.2.1. Cervicale
1.2.2. Thoracique
1.2.3. Lombaire
1.2.4. Sacrée
1.2.5. Coccygienne
Figure 48 6 Classification embryologique des différentes structures du système nerveux central (d’après Swanson LW (2003).
51

Pour en savoir plus :
1. Allman J. Evolving Brains. Scientific American Library. New York: Freeman, 2000
2. Baertschi B. Ce que les neurosciences font à nos conceptions morales. La découverte, 2009
3. Bayer SA and Altman J. Atlas of the human central nervous system development, volumes 1 à 5. CRC Press, 2002
4. Held LI. Quirks of human anatomy. An evo-devo look at the human body. Cambridge university press, 2009
5. Jeannerod M, Le cerveau intime. Odile Jacob Poche, 2005
6. Swanson LW. Brain architecture. Oxford University press, 2003
52