Embed Size (px)
Citation preview

Chapitre 2
Le fonctionnement des organismes animaux en fonction du plan
d’organisation et du milieu de vie
Partie II - L’organisme vivant : un système physico-chimique en interaction avec son environnement
code des diapositives
✭ très important, à savoir avec précision❀ important pour comprendre
✄ pour approfondir, sinon à couper1

4 animaux observés en TPVertébrés Arthropodes
aérien
aquatique
❀
2
souris
criquet
maquereau
truite
langoustine
écrevisse

1. Les mêmes fonctionspour tous les animaux
3
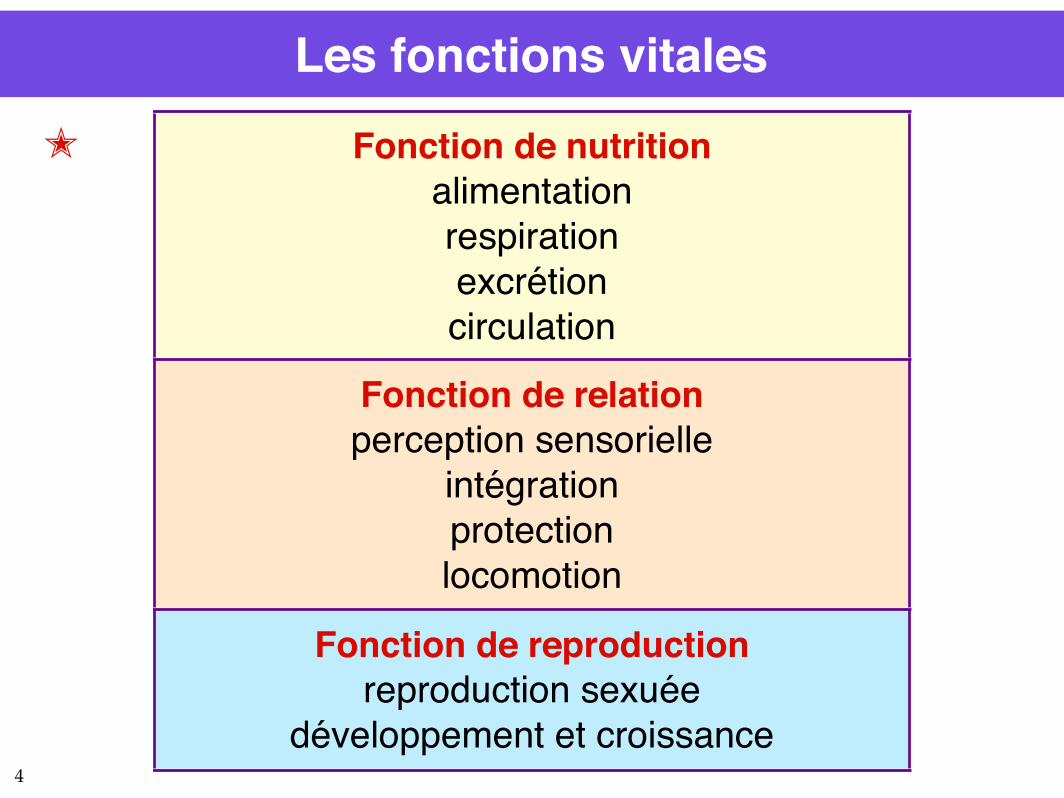
Les fonctions vitales
Fonction de nutritionalimentationrespirationexcrétioncirculation
Fonction de relationperception sensorielle
intégrationprotectionlocomotion
Fonction de reproductionreproduction sexuée
développement et croissance
✭
4
Excrétion, un mode universel... ou presque
16
Plans d’organisations et relation entre les organismes et leur milieu
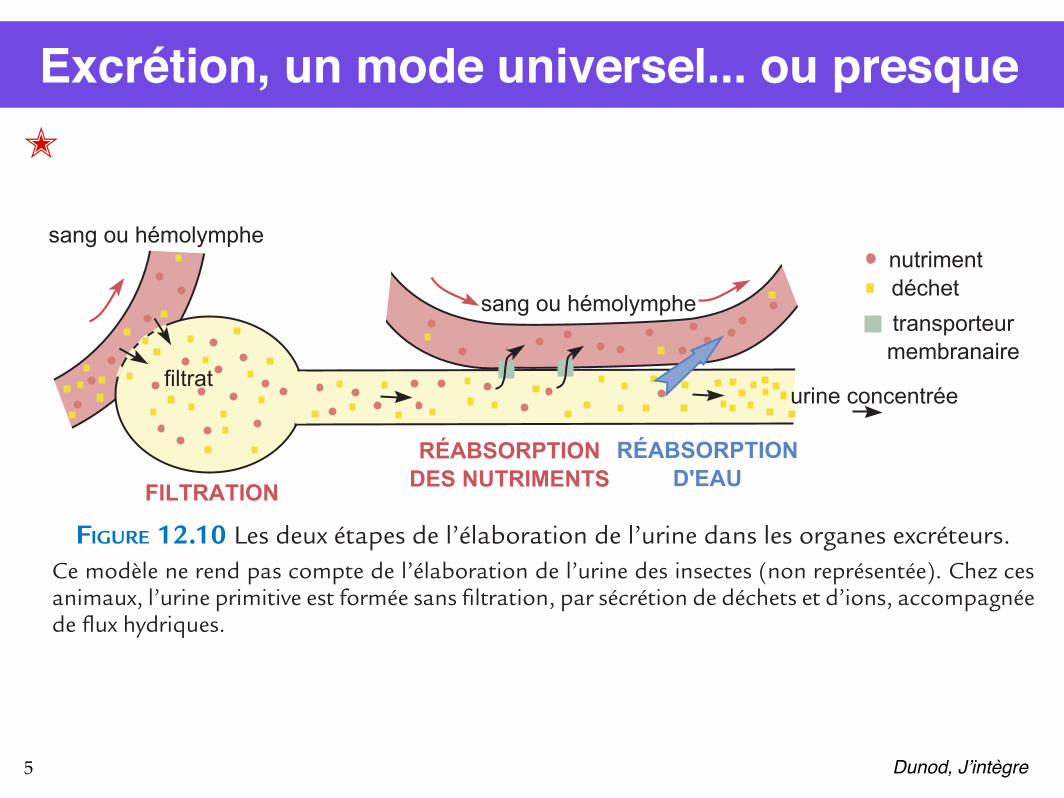
12.3.2 Excréter les déchets azotés dans différents milieuxLe catabolisme cellulaire aboutit par étapes successives à la formation de déchets comme CO2 (pour toutes les molécules organiques) ou NH3 pour les molécules azotées (protéines, acides nucléiques). Ces déchets sont d’abord libérés dans le sang ou l’hémolymphe. Le CO2 est éli-miné par les surfaces d’échanges respiratoires, lors de la respiration. C’est une autre fonction, l’excrétion, qui permet à l’organisme de se débarrasser des déchets azotés. Chez les vertébrés, deux reins élaborent l’urine dans des tubules, ou néphrons, fermés à une extrémité et ouverte à l’autre vers l’extérieur, où l’urine est évacuée par les voies urinaires. Le criquet et l’écrevisse ne possèdent pas de reins mais d’autres organes excréteurs. Chez l’écrevisse, il s’agit de deux glandes antennaires situées dans la tête de l’écrevisse et reliées à un canal dont l’orifice s’ouvre à la base du premier article de chaque antenne (a2). Les néphrons des vertébrés et les glandes antennaires de l’écrevisse élaborent l’urine suivant les deux mêmes étapes (figure 12.10). Le liquide circulant (plasma ou hémolymphe) est filtré à travers l’extrémité proximale du tubule, ce qui laisse passer toutes les petites molécules (qu’elles soient ou non des déchets) ; puis les nutriments et une partie de l’eau sont réabsorbés et seuls les déchets sont excrétés, avec une certaine quantité d’eau.
FILTRATION
RÉABSORPTION
DES NUTRIMENTS
RÉABSORPTION
D'EAU
urine concentréefiltrat
sang ou hémolymphe
sang ou hémolymphe
nutriment
déchet
transporteur
membranaire
FIGURE 12.10 Les deux étapes de l’élaboration de l’urine dans les organes excréteurs.Ce modèle ne rend pas compte de l’élaboration de l’urine des insectes (non représentée). Chez ces animaux, l’urine primitive est formée sans filtration, par sécrétion de déchets et d’ions, accompagnée de flux hydriques.
Les animaux excrètent l’une des trois molécules azotées (ammoniaques, urée ou acide urique) comme indiqué sur le tableau 12.5.
TABLEAU 12.5 LES DÉCHETS AZOTÉS DES ANIMAUX ÉTUDIÉS EN TRAVAUX PRATIQUES.
Écrevisse Criquet Téléostéens Souris
Ammoniac
Solubilité : 52,4 mM
Acide urique
Solubilité : 0,0015 mM
Ammoniac
Solubilité : 52,4 mM
Urée
Solubilité : 39,8 mM
L’ammoniac est excrété par les téléostéens et l’écrevisse, mais ni par le criquet ni par la souris. Ce fait peut être généralisé. En effet, l’ammoniac est très hydrosoluble mais aussi très toxique. Il peut être excrété par tout échangeur en contact avec l’eau (comme la peau des téléostéens ou les branchies). Ainsi les animaux aquatiques, traversés par un fort flux d’eau, excrètent leur azote sous forme d’ammoniac qu’ils peuvent éliminer facilement, ce qui n’est pas le cas des animaux terrestres. L’urée est aussi très hydrosoluble. Elle est synthétisée à partir de CO2 et de NH3, par une succession de réactions catalysées par des enzymes spécifiques. Sa synthèse représente donc
✭
Dunod, J’intègre5
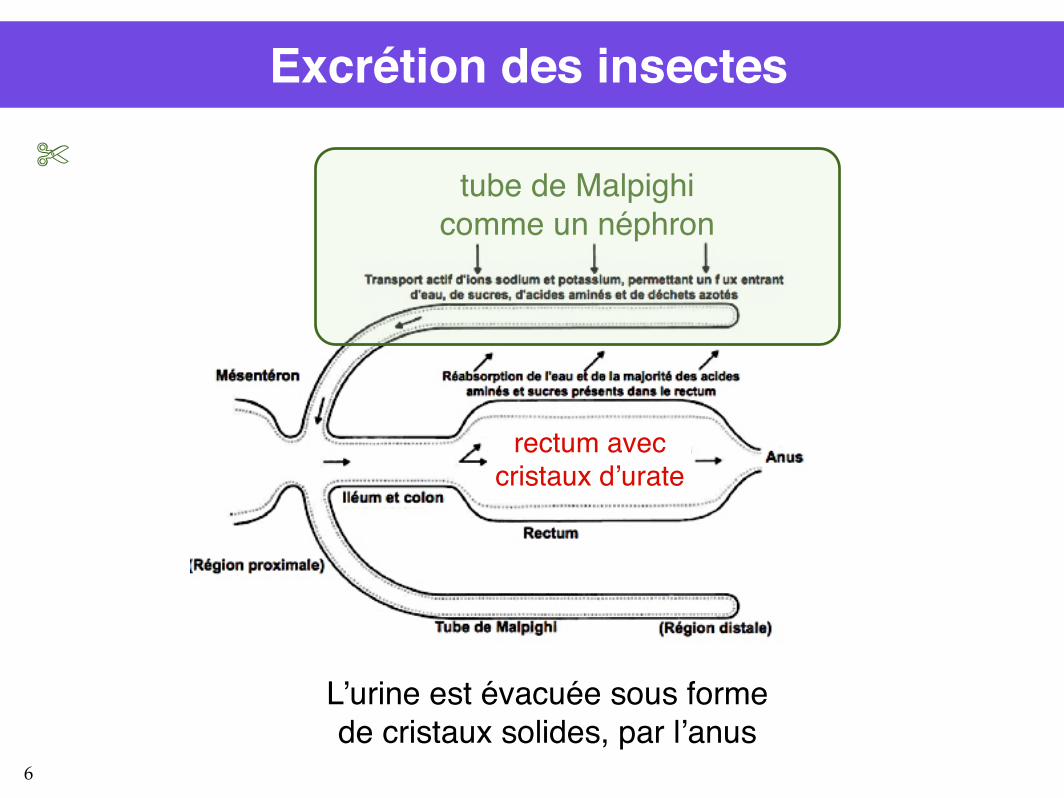
Excrétion des insectes
tube de Malpighicomme un néphron
rectum avec cristaux d’urate
L’urine est évacuée sous forme de cristaux solides, par l’anus
✄
6
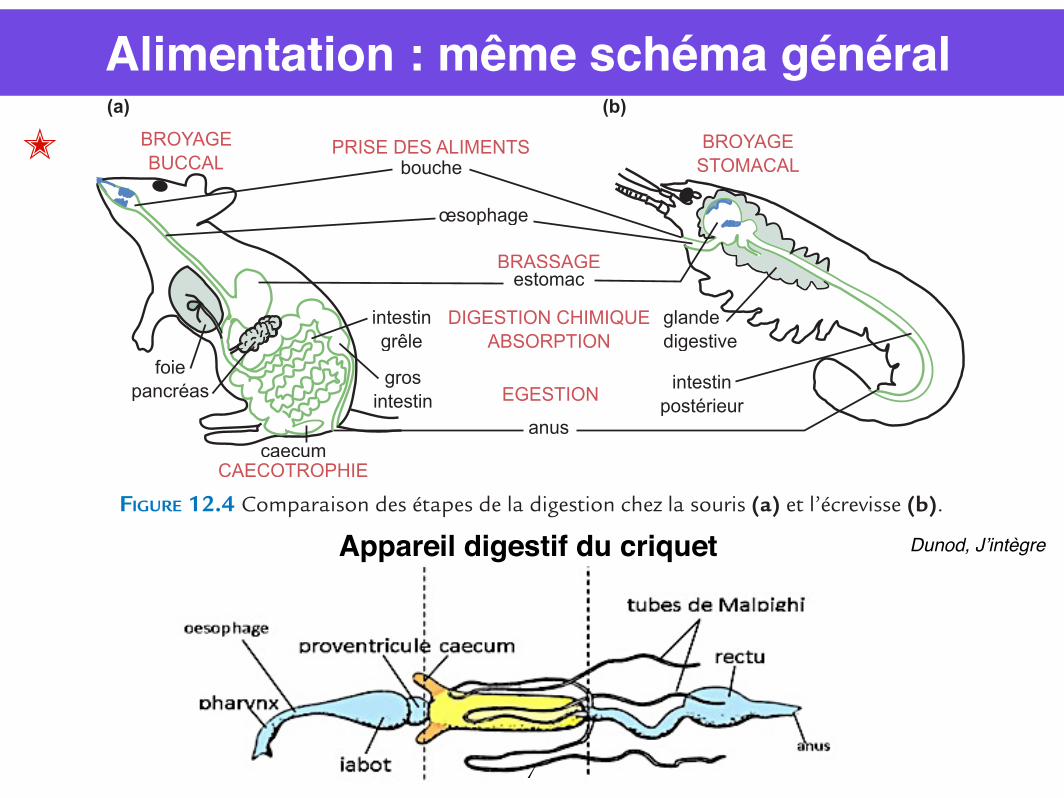
Alimentation : même schéma général
bouche
PRISE DES ALIMENTSBROYAGE
BUCCAL
BROYAGE
STOMACAL
œsophage
estomac
BRASSAGE
anus
gros
intestin
intestin
postérieur
DIGESTION CHIMIQUE
ABSORPTION
intestin
grêle
glande
digestive
caecum
CAECOTROPHIE
foie
pancréas EGESTION
(a) (b)
FIGURE 12.4 Comparaison des étapes de la digestion chez la souris (a) et l’écrevisse (b).
Appareil digestif du criquet
✭
Dunod, J’intègre
7
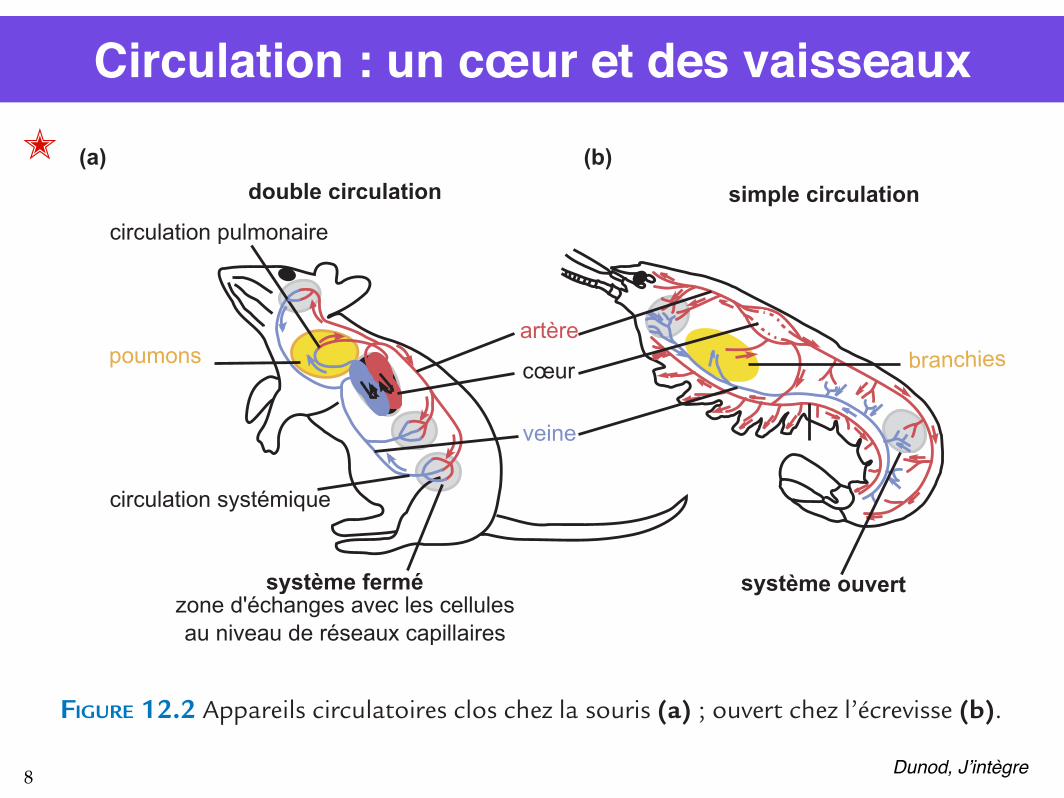
Circulation : un cœur et des vaisseaux
4
Plans d’organisations et relation entre les organismes et leur milieu
circulation pulmonaire
système fermé
artère
veine
zone d'échanges avec les cellules
au niveau de réseaux capillaires
poumons
circulation systémique
cœurbranchies
système ouvert
double circulation simple circulation
(a) (b)
FIGURE 12.2 Appareils circulatoires clos chez la souris (a) ; ouvert chez l’écrevisse (b).
c) Les fonctions de reproduction
Des animaux gonochoriquesChez la souris et l’écrevisse, comme chez la plupart des animaux, les sexes sont séparés ; ces animaux sont gonochoriques. La souris qui vit 3 ou 4 ans, peut se reproduire dès 3 mois, toute l’année. L’écrevisse, dont la longévité peut atteindre 20 ans, est à maturité sexuelle au cours de sa troisième année, et sa période de reproduction est limitée à l’automne.
Un accouplement favorisant la rencontre des gamètesComme la vache, la souris n’accepte l’accouplement que lors de sa période d’œstrus ; cependant le cycle œstrien de la souris est beaucoup plus court que celui de la vache (4 à 5 jours). Grâce à son pénis, le mâle introduit son sperme dans le vagin de la femelle. La fécondation est interne : elle a lieu dans les voies génitales femelles.Les écrevisses s’accouplent aussi, en automne, le mâle et la femelle étant alors orientés l’un vers l’autre par leurs faces ventrales. Les spermatozoïdes agglomérés en amas cylindriques, les spermatophores, s’écoulent dans un canal délimité par les deux premières paires d’appendices abdominaux jusqu’à aller s’accrocher sur l’abdomen de la femelle où ils restent fixés jusqu’à la ponte quelques jours à quelques semaines plus tard ; les spermatophores libèrent alors leurs spermatozoïdes qui fécondent les ovules. La fécondation est externe.Dans les deux cas, l’accouplement augmente la probabilité de rencontre des gamètes mâles et femelles ; la fécondation interne des mammifères permet en outre de protéger les gamètes du caractère déshydratant et non porteur du milieu terrestre.
Une attention différente portée aux descendants lors de leur développementChez la souris comme chez la vache, le développement embryonnaire se déroule dans l’uté-rus maternel. La souris est vivipare. La gestation dure 19 à 21 jours. La portée comprend en moyenne 9 souriceaux. Nés aveugles, ils restent au nid et sont allaités par leur mère pendant 2 à 3 semaines avant de prendre leur essor.L’écrevisse pond entre 100 et 150 œufs, qui, après la fécondation, restent fixés à son abdomen pendant les mois que dure le développement embryonnaire. L’écrevisse est ovipare. La femelle favorise régulièrement l’oxygénation de ses œufs en se plaçant dans un courant d’eau. Après l’éclosion, les jeunes écrevisses vivent encore quelques semaines à proximité de leur mère qui les protège mais ne les nourrit pas. Cependant, un fort pourcentage d’œufs n’éclôt jamais et moins de 15 % des écrevisses ayant éclos survivent au-delà de leur premier été.
✭
Dunod, J’intègre8

2. Influence du plan d’organisationsur la réalisation des fonctions
9
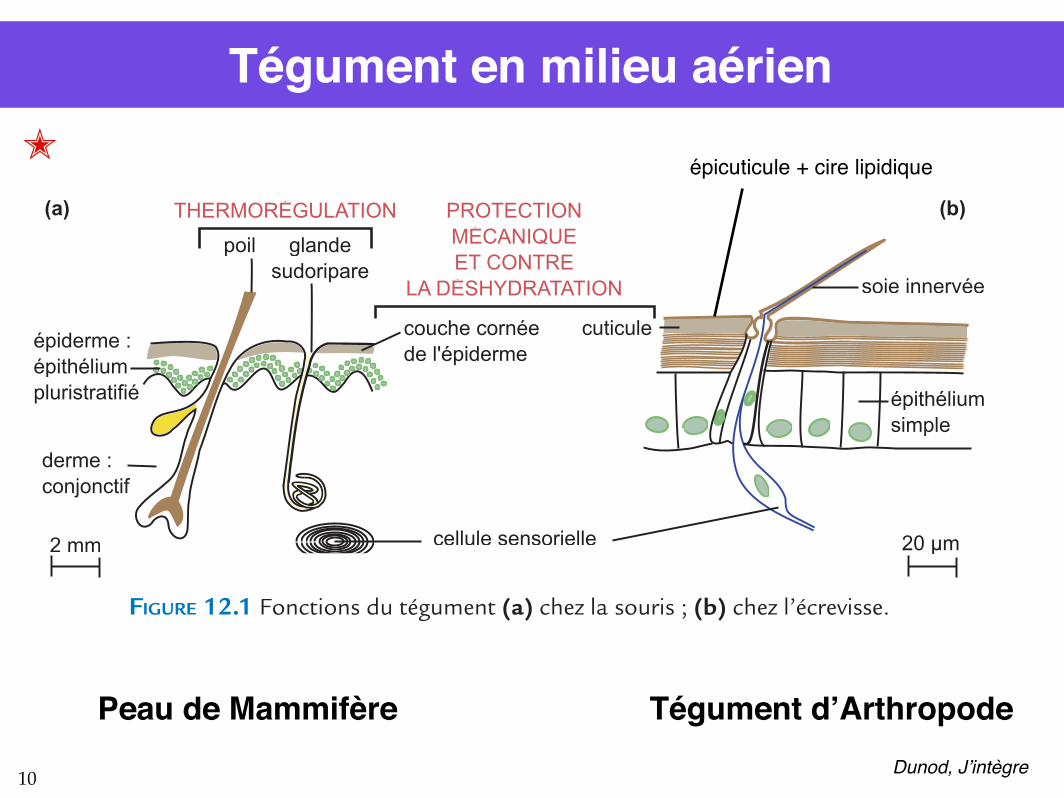
Tégument en milieu aérien
2
Plans d’organisations et relation entre les organismes et leur milieu
TABLEAU 12.1 RÉCEPTEURS SENSORIELS CHEZ LA SOURIS ET L’ÉCREVISSE.
Fonctionsensorielle
Organes - Type de récepteur
Souris (§ 4.1.2) Écrevisse (figure TP6.1)
vision Yeux (× 2) - photorécepteurs Yeux (× 2) - photorécepteurs
audition Tympan dans l’oreille moyenne (× 2) - mécano-récepteurs
gustation Langue - chimiorécepteurs Antennes (× 2) et antennules (2) ; soies - mécanorécepteursolfaction Muqueuse nasale - chimiorécepteurs
équilibration Oreille interne (× 2) - mécanorécepteurs Statocystes (× 2, à la base de a1)
somesthésie Peau - divers Soies dispersées sur la cuticule
Protection
couche cornée
de l'épiderme
PROTECTION
MÉCANIQUE
ET CONTRE
LA DÉSHYDRATATION
cuticule
poil glande
sudoripare
THERMORÉGULATION
cellule sensorielle
SENSIBILITE TACTILE
soie innervée
épiderme :
épithélium
pluristratifié épithélium
simple
derme :
conjonctif
2 mm 20 µm
(a) (b)
FIGURE 12.1 Fonctions du tégument (a) chez la souris ; (b) chez l’écrevisse.
Le corps de la souris est protégé mécaniquement, thermiquement et physiologiquement par sa peau. Le tégument de l’écrevisse est un épithélium simple (figure 12.1) doublé d’une cuticule formée pour moitié de chitine associée à des protéines et pour moitié à des sels de calcium. Cette enveloppe dure offre une protection mécanique. Elle est recouverte d’une couche de cires qui empêchent la pénétration de l’eau dans l’organisme à travers le tégument. Elle est aussi un obstacle aux agents pathogènes. En revanche, elle ne joue aucun rôle dans la thermorégulation. En effet, contrairement aux mammifères, l’écrevisse ne régule pas sa température corporelle et, selon les espèces, supporte plus ou moins bien les eaux de températures supérieures à 25 °C. La cuticule limite aussi la sensibilité tactile et la croissance ne pourra se faire que lors des mues successives.
Activités et fonctions motricesLa souris et l’écrevisse se déplacent grâce à la contraction de leurs muscles qui, commandée par des neurones moteurs, met en mouvement des pièces squelettiques rigides et articulées. Chez la souris, comme chez la vache, le squelette est formé par des os, visibles seulement à la dis-section : c’est un endosquelette. Chez l’écrevisse, la cuticule constitue un exosquelette. Dans tous les cas, un muscle ne peut faire bouger un élément du squelette que dans une direction et un sens précis. En général, il existe un muscle antagoniste qui permet le retour à la position de repos. Les tensions développées par la contraction musculaire sont amplifiées par des bras de levier qui dépendent de la distance entre le point d’insertion du muscle et le centre de rotation des pièces articulées. La locomotion résulte de la succession de phases variables d’un mode à un
Voir TP4, § TP4.1.1
Voir chapitre 2, § 2.12b
Peau de Mammifère Tégument d’Arthropode
épicuticule + cire lipidique✭
Dunod, J’intègre10
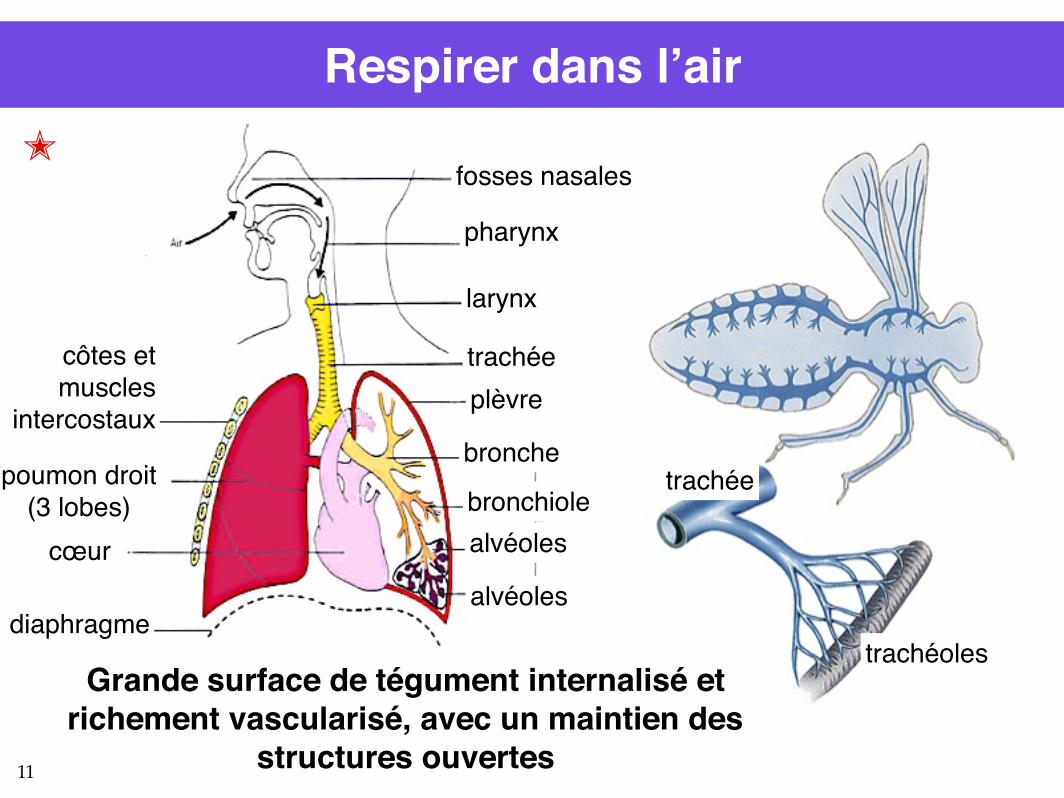
Respirer dans l’air✭
Grande surface de tégument internalisé etrichement vascularisé, avec un maintien des
structures ouvertes
fosses nasales
pharynx
larynx
trachéeplèvre
bronche
bronchiolealvéoles
alvéoles
côtes et muscles
intercostaux
poumon droit (3 lobes)
cœur
diaphragme
trachée
trachéoles
11
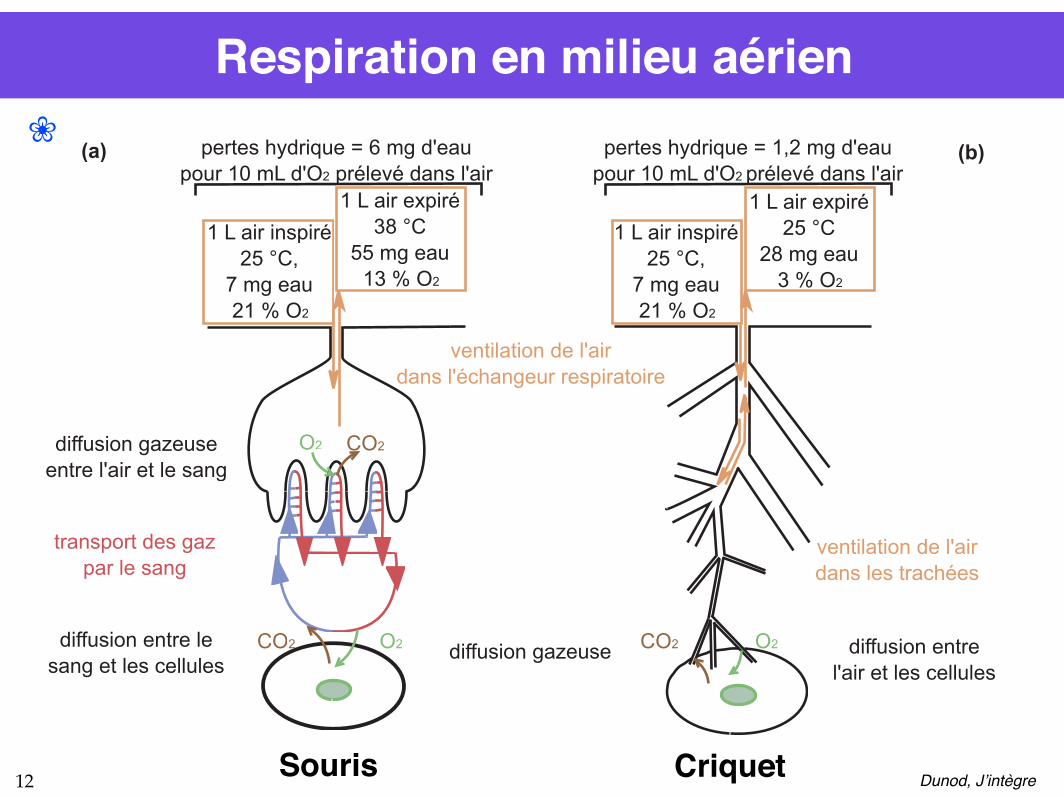
Respiration en milieu aérien
1 L air expiré
25 °C
28 mg eau
3 % O2
O2 CO2
O2CO2
(a)
1 L air inspiré
25 °C,
7 mg eau
21 % O2
pertes hydrique = 6 mg d'eau
pour 10 mL d'O2 prélevé dans l'air
1 L air expiré
38 °C
55 mg eau
13 % O2
1 L air inspiré
25 °C,
7 mg eau
21 % O2
pertes hydrique = 1,2 mg d'eau
pour 10 mL d'O2 prélevé dans l'air
ventilation de l'air
dans l'échangeur respiratoire
diffusion gazeuse
entre l'air et le sang
diffusion gazeuse diffusion entre le
sang et les cellulesdiffusion entre
l'air et les cellules
O2CO2
transport des gaz
par le sangventilation de l'air
dans les trachées
(b) ❀
Souris Criquet Dunod, J’intègre12

Les branchies de poisson et d’écrevisse
Grande surface de tégument flottant dans le milieu aquatiquerichement vascularisé, avec un courant d’eau régulé
Branchies à nombreux filaments disposant de centaines de lamelles
protection par un opercule
Branchies à nombreux filaments plumeux
protection par un repli du tégument (le branchiostégite)
✭
13

La locomotion dans l’air
6 pattespattes postérieures repliées en Z
2 paires d’ailes4 pattes
queue participant à l’équilibre
corps surélevé au-dessus du solmembres porteurs et articulés
musculature associée✭
14

La locomotion dans l’eau
palette natatoire d’écrevissenageoire caudale de gardon
Une surface large pour se propulser✭
15

3. Influence du milieu de viesur la réalisation des fonctions
16
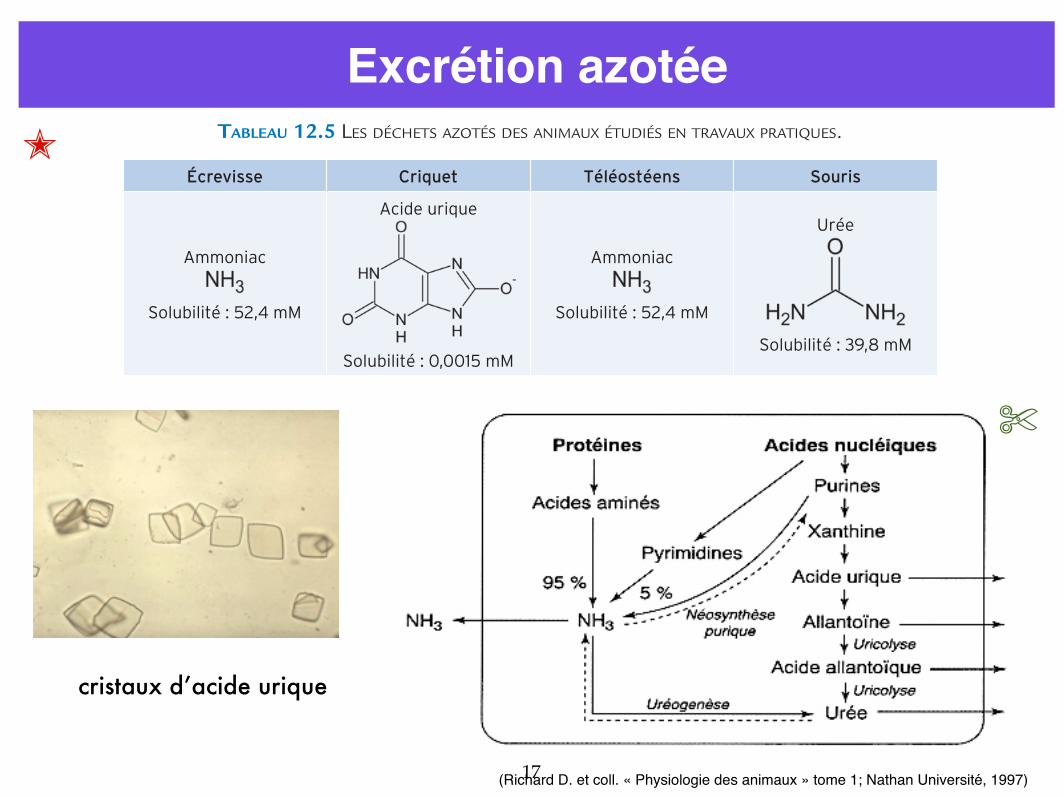
Excrétion azotée
16
Plans d’organisations et relation entre les organismes et leur milieu
12.3.2 Excréter les déchets azotés dans différents milieuxLe catabolisme cellulaire aboutit par étapes successives à la formation de déchets comme CO2 (pour toutes les molécules organiques) ou NH3 pour les molécules azotées (protéines, acides nucléiques). Ces déchets sont d’abord libérés dans le sang ou l’hémolymphe. Le CO2 est éli-miné par les surfaces d’échanges respiratoires, lors de la respiration. C’est une autre fonction, l’excrétion, qui permet à l’organisme de se débarrasser des déchets azotés. Chez les vertébrés, deux reins élaborent l’urine dans des tubules, ou néphrons, fermés à une extrémité et ouverte à l’autre vers l’extérieur, où l’urine est évacuée par les voies urinaires. Le criquet et l’écrevisse ne possèdent pas de reins mais d’autres organes excréteurs. Chez l’écrevisse, il s’agit de deux glandes antennaires situées dans la tête de l’écrevisse et reliées à un canal dont l’orifice s’ouvre à la base du premier article de chaque antenne (a2). Les néphrons des vertébrés et les glandes antennaires de l’écrevisse élaborent l’urine suivant les deux mêmes étapes (figure 12.10). Le liquide circulant (plasma ou hémolymphe) est filtré à travers l’extrémité proximale du tubule, ce qui laisse passer toutes les petites molécules (qu’elles soient ou non des déchets) ; puis les nutriments et une partie de l’eau sont réabsorbés et seuls les déchets sont excrétés, avec une certaine quantité d’eau.
FILTRATION
RÉABSORPTION
DES NUTRIMENTS
RÉABSORPTION
D'EAU
urine concentréefiltrat
sang ou hémolymphe
sang ou hémolymphe
nutriment
déchet
transporteur
membranaire
FIGURE 12.10 Les deux étapes de l’élaboration de l’urine dans les organes excréteurs.Ce modèle ne rend pas compte de l’élaboration de l’urine des insectes (non représentée). Chez ces animaux, l’urine primitive est formée sans filtration, par sécrétion de déchets et d’ions, accompagnée de flux hydriques.
Les animaux excrètent l’une des trois molécules azotées (ammoniaques, urée ou acide urique) comme indiqué sur le tableau 12.5.
TABLEAU 12.5 LES DÉCHETS AZOTÉS DES ANIMAUX ÉTUDIÉS EN TRAVAUX PRATIQUES.
Écrevisse Criquet Téléostéens Souris
Ammoniac
Solubilité : 52,4 mM
Acide urique
Solubilité : 0,0015 mM
Ammoniac
Solubilité : 52,4 mM
Urée
Solubilité : 39,8 mM
L’ammoniac est excrété par les téléostéens et l’écrevisse, mais ni par le criquet ni par la souris. Ce fait peut être généralisé. En effet, l’ammoniac est très hydrosoluble mais aussi très toxique. Il peut être excrété par tout échangeur en contact avec l’eau (comme la peau des téléostéens ou les branchies). Ainsi les animaux aquatiques, traversés par un fort flux d’eau, excrètent leur azote sous forme d’ammoniac qu’ils peuvent éliminer facilement, ce qui n’est pas le cas des animaux terrestres. L’urée est aussi très hydrosoluble. Elle est synthétisée à partir de CO2 et de NH3, par une succession de réactions catalysées par des enzymes spécifiques. Sa synthèse représente donc
(Richard D. et coll. « Physiologie des animaux » tome 1; Nathan Université, 1997)
cristaux d’acide urique
✭
✄
17
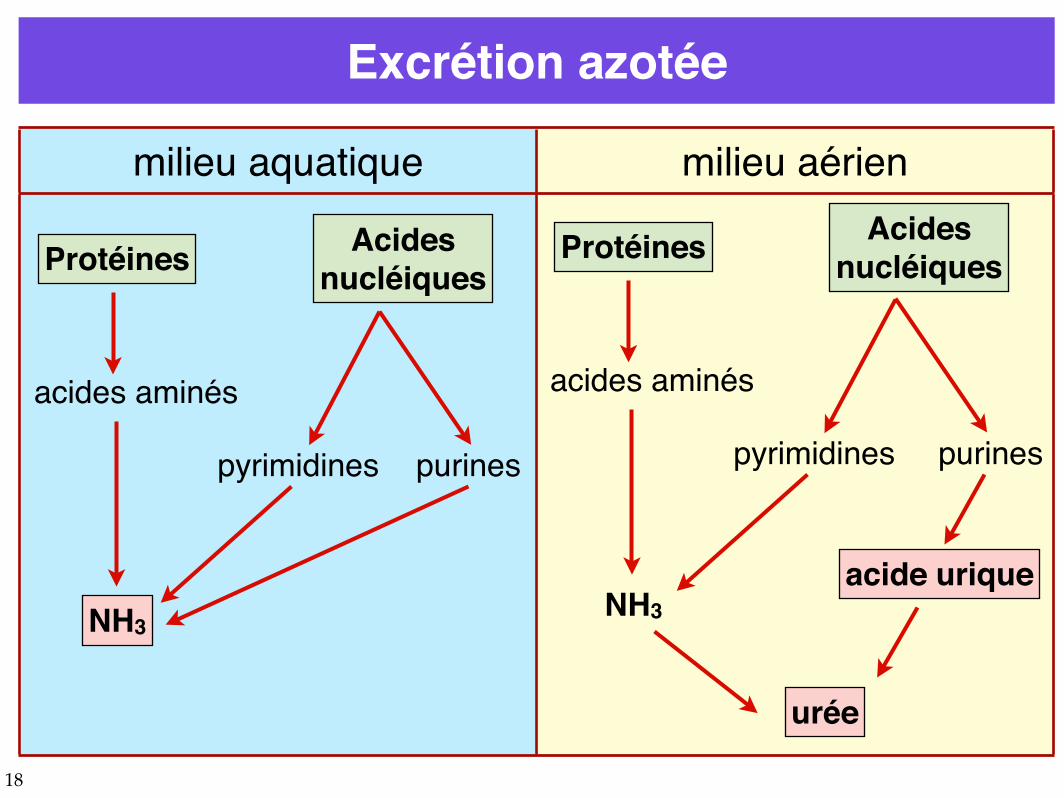
Excrétion azotée
milieu aquatique milieu aérien
Protéines Acidesnucléiques
acides aminés
pyrimidines purines
NH3
Protéines Acidesnucléiques
acides aminés
pyrimidines
NH3
purines
acide urique
urée
18
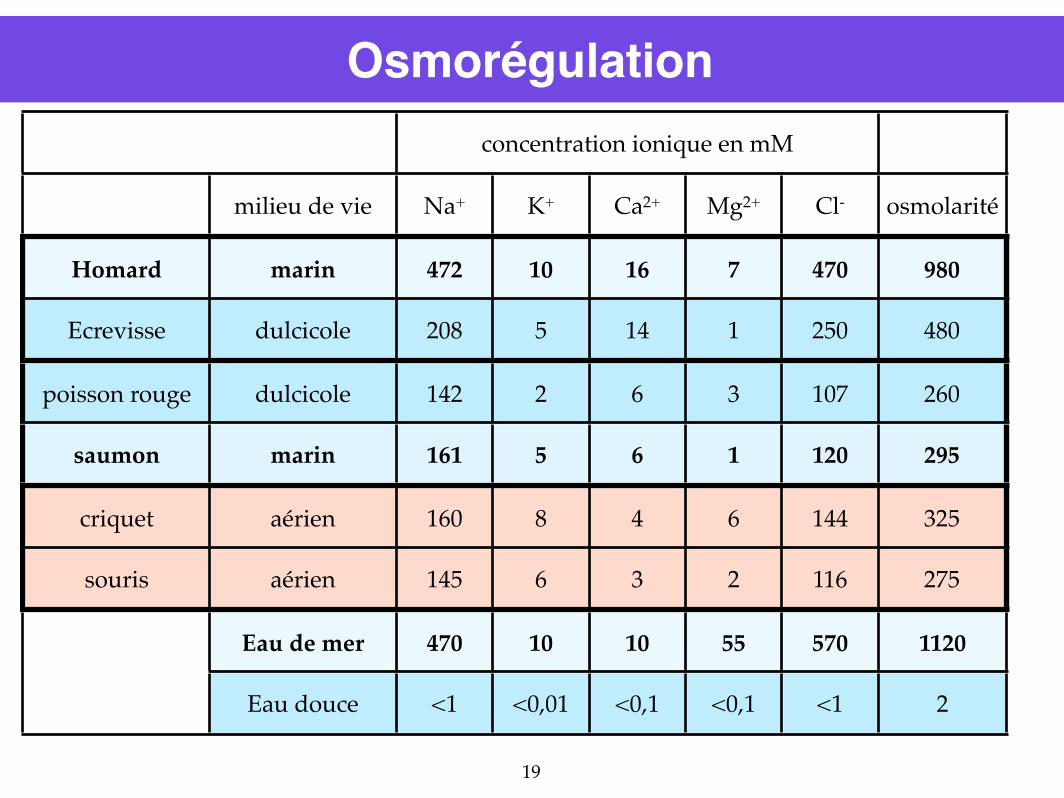
Osmorégulationconcentration ionique en mMconcentration ionique en mMconcentration ionique en mMconcentration ionique en mMconcentration ionique en mM
milieu de vie Na+ K+ Ca2+ Mg2+ Cl- osmolarité
Homard marin 472 10 16 7 470 980
Ecrevisse dulcicole 208 5 14 1 250 480
poisson rouge dulcicole 142 2 6 3 107 260
saumon marin 161 5 6 1 120 295
criquet aérien 160 8 4 6 144 325
souris aérien 145 6 3 2 116 275
Eau de mer 470 10 10 55 570 1120
Eau douce <1 <0,01 <0,1 <0,1 <1 2
19
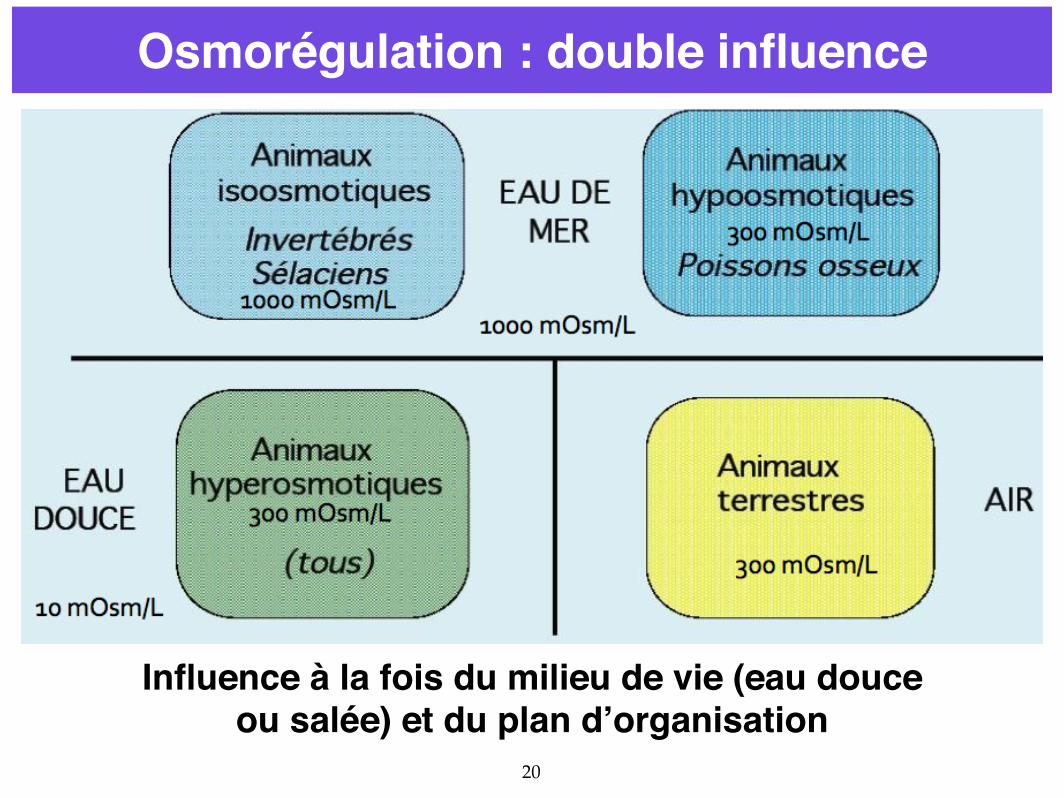
20
Osmorégulation : double influence
Influence à la fois du milieu de vie (eau douce ou salée) et du plan d’organisation
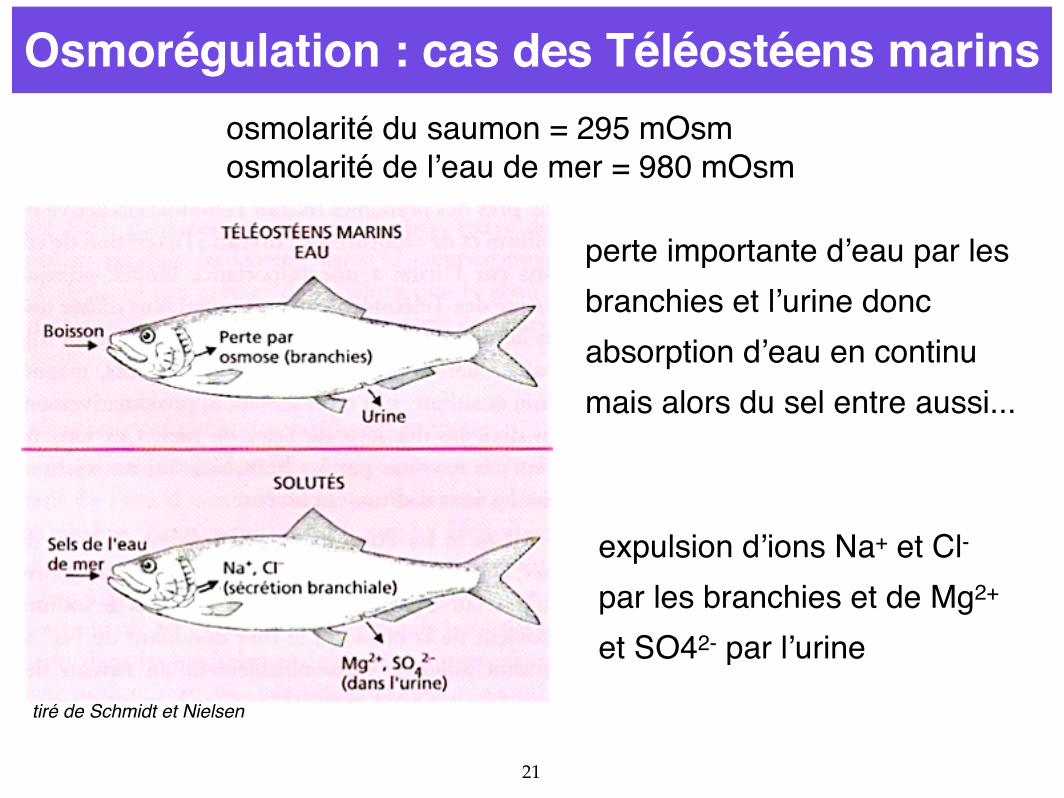
21
Osmorégulation : cas des Téléostéens marinsosmolarité du saumon = 295 mOsmosmolarité de l’eau de mer = 980 mOsm
perte importante d’eau par les branchies et l’urine donc absorption d’eau en continu mais alors du sel entre aussi...
expulsion d’ions Na+ et Cl- par les branchies et de Mg2+ et SO42- par l’urine
tiré de Schmidt et Nielsen

22
Osmorégulation : cas des Téléostéens dulcicolesosmolarité du gardon = 260 mOsmosmolarité de l’eau douce = 2 mOsm
entrée importante d’eau par les branchies donc urine abondante
prélèvement actif d’ions par les branchies et peu de perte d’ions par les reins
tiré de Schmidt et Nielsen

Audition des Vertébrés et milieu de vie
23
ligne latérale sensible à la pressionpas de tympan
ligne latérale
Poisson téléostéen Grenouille
ligne latérale chez la larve
tympan chez l’adultepas de pavillon externe
tympan
Souris
tympan + pavillon externe
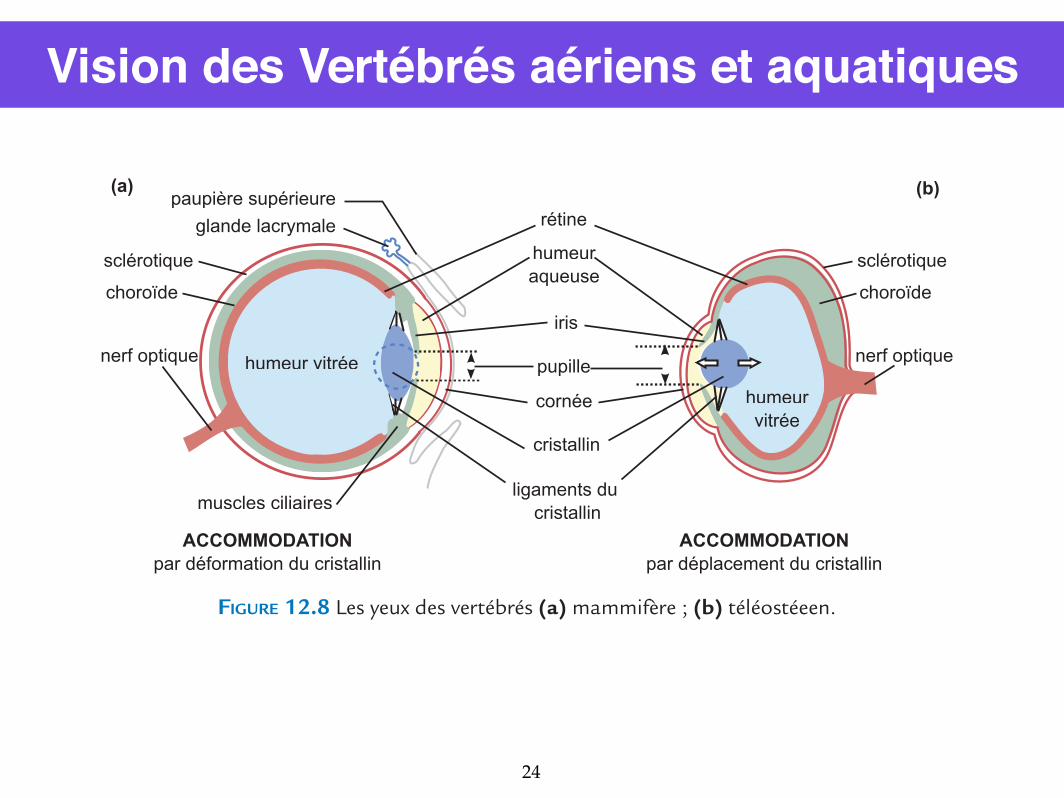
Vision des Vertébrés aériens et aquatiques
14
Plans d’organisations et relation entre les organismes et leur milieu
d’un orifice, la pupille, par lequel la lumière pénètre dans l’œil. Si la luminosité est forte, le diamètre de la pupille rétrécit, ajustant la quantité de lumière qui va frapper la rétine, à la manière du diaphragme d’un appareil photographique. À l’inverse, le diamètre augmente si la luminosité baisse. L’œil des mammifères est sphérique ; le cristallin biconvexe réfracte les rayons lumineux issus d’un objet situé à l’infini de façon à ce que l’image se forme sur la rétine. Pour les objets plus proches, le cristallin se déforme grâce à la contraction de muscles attachés aux ligaments du cristallin, ce qui le rend plus convergent et permet ainsi aux images des objets proches d’être formées sur la rétine : c’est l’accommodation. Les rayons lumineux qui frappent les photorécepteurs de la rétine, en un point variable selon l’objet émetteur, déclenchent une réaction photochimique qui, par une cascade de réactions est convertie en variation du potentiel membranaire des cellules photoréceptrices, elle-même à l’origine d’un message nerveux. Le traitement par les centres nerveux de cette information permet alors à l’objet d’être vu de façon nette.Chez les téléostéens, les yeux sont dépourvus de paupières ; c’est l’eau de l’environnement qui maintient la cornée hydratée. L’œil est aplati selon un axe antéro-postérieur, la cornée est plate, et le cristallin sphérique. Un tel œil permet au poisson de voir nettement les objets immergés ; mais, compte tenu de la différence d’indice entre l’eau et l’air, si cet œil fonc-tionnait dans l’air, l’image se formerait en avant de la rétine et le poisson serait myope. L’accommodation se fait d’une façon différente de celle des mammifères, par le déplacement du cristallin dans l’axe antéropostérieur de l’œil (un peu comme la lentille d’un appareil photographique lors de la mise au point).
glande lacrymale
pupille
cristallin
iris
rétine
nerf optique nerf optique
muscles ciliaires
sclérotique
choroïde
(a) (b)
sclérotique
choroïde
humeur vitrée
humeur
vitrée
humeur
aqueuse
cornée
ACCOMMODATION
par déformation du cristallin
ACCOMMODATION
par déplacement du cristallin
paupière supérieure
ligaments du
cristallin
FIGURE 12.8 Les yeux des vertébrés (a) mammifère ; (b) téléostéeen.
b) Les yeux composés des arthropodes : apposition ou superpositionUn œil composé d’arthropode est formé de la juxtaposition d’unités photoréceptrices, les ommatidies, constituées chacune comme un œil élémentaire : une cornée faite de cuticule transparente, un cristallin transparent et conique, 8 cellules rétiniennes. Ces dernières possè-dent des pigments photorécepteurs dans des microvillosités membranaires qui, en s’emboîtant les unes dans les autres, forment un bâtonnet photorécepteur. Le cristallin ne se déforme pas : il n’y a pas d’accommodation, mais la convergence des lentilles que forment cornée et cristal-lin, et la longueur du bâtonnet photorécepteur permettent de recevoir sur celui-ci les rayons lumineux émis à différentes distances. Comme chez les vertébrés, un stimulus lumineux, à la suite d’une réaction photochimique du pigment photorécepteur, génère un message nerveux qui, propagé jusqu’aux centres nerveux, est perçu comme une image élémentaire. Il existe en outre des pigments non photorécepteurs formant un écran autour de chaque ommatidie. Leur disposition varie en fonction de la luminosité du milieu de vie de l’animal.
Voir chapitre 5, § 5.2
24
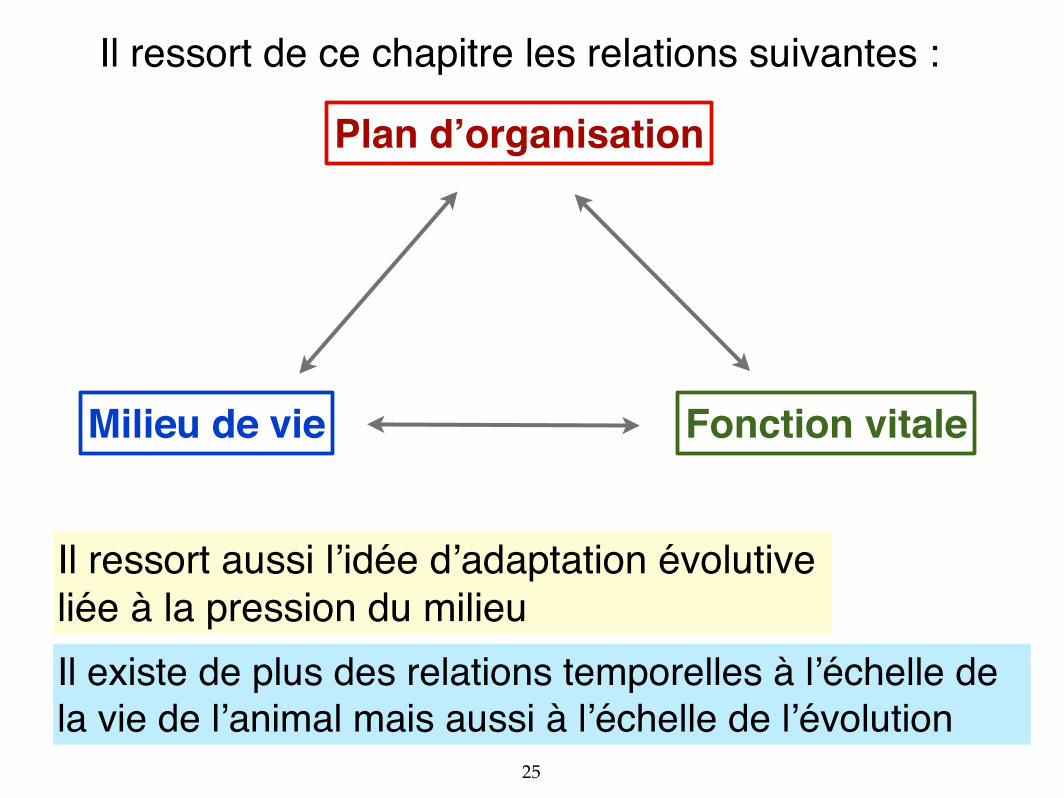
Plan d’organisation
Milieu de vie Fonction vitale
Il existe de plus des relations temporelles à l’échelle de la vie de l’animal mais aussi à l’échelle de l’évolution
Il ressort de ce chapitre les relations suivantes :
Il ressort aussi l’idée d’adaptation évolutiveliée à la pression du milieu
25