Embed Size (px)
Citation preview

338 BIOCHIMICA ET BIOPHYSICA ACTA
BBA 95319
LE SYST]kME DES DNA-POLYMI~RASES DE FOIE DE RAT ET LA
TRANSFORMATION BIOLOGIQUE DU DNA NATIF EN DNA " P R I M E R "
ANNE-MARIE DE RECONDO
Centre de Recherches sur la CeUule Normale et Cancdreuse, Villejui], Seine (France)
(Re~u le 25 janvier, 1965) (Manuscrit r6vis6 re~u le 21 mai, 1965)
SUMMARY
Biological /actors affecting the primer /or DNA polymerase o/ regenerating rat liver Under standard assay conditions, the DNA polymerase (deoxynucleoside-
triphosphate:DNA deoxynucleotidyltransferase, EC 2.7.7.7) from regenerating rat liver has a specific requirement for heat-denatured DNA (single-stranded DNA) like calf-thymus polymerase. Native DNA (double-stranded DNA) preincubated in different biological media is, however, active. Native DNA incubated with regen- erating rat liver or hepatoma supernatant is even more active than the heat-de- naturated DNA.
The deoxyribonucleases I (EC 3.1.4.5) and II (EC 3.1.4.6) from these super- natants are not involved since: (I) the increase of priming effect is not blocked by specific inhibitor from deoxyribonuclease I and (2) the deoxyribonuclease II de- creases the priming capacity of native DNA.
The results may be explained by the action of an unidentified enzyme which would liberate active sites on the primer DNA molecule. These sites would facilitate the action of the terminal nucleotidyl transferase and the replicative DNA poly- merase. This enzyme appears to have some properties in common with the E. coli DNA phosphatase exonuclease described by RICHARDSON et al.
INTRODUCTION
Parmi les nombreux travaux concernant les DNA-polym6rases (d6soxy- ribonucl6osidetriphosphate:DNA d6soxyribonucl6otidyltransf~rase, EC 2.7.7.7) pu- blids ces derni~res ann~es, certains ont plus particuli~rement port6 sur le DNA n6cessaire au fonctionnement de ces enzymes et l '6tat physico-chimique dans lequel il dolt se trouver pour servir de "primer ''1-4. Certaines divergences peuvent ~tre relev6es k cet 6gard suivant les syst~mes enzymatiques 6tudids:
L'enzyme purifi6e ~ partir de thymus de veau par BOLLUM et son 6quipe 5,* utilise pr6f6rentiellement du DNA ~ simple chaine: DNA d6natur6 par la chaleur ou DNA de qsXI74.
Biochim. Biophys. Acta. 114 (1966) 338-348

DNA "PRIMER" POUR LA DNA-POLYMt~RASE 339
L'enzyme dtudide chez Escherichia coli par KORNBERG et ses collaborateurs ~ utilise aussi bien le DNA natif que le DNA d6naturd par la chaleur alors que la DNA-polymdrase induite chez cette bactdrie par le bactdriophage T 2 synthdtise du DNA ~ u n taux beaucoup plus dlevd lorsqu'on lui fournit comme "primer" un DNA /t simple chaine.
L'enzyme prdsente dans le surnageant de foie en hypertrophie compensatrice, et dont la purification est moins poussde, a donn6 lieu ~ des publications contra- dictoires: certains auteurs utilisent comme "primer" du DNA natif s-l°, d'autres obtiennent de meilleurs rdsultats avec du DNA ddnaturd par la chaleurn, 12. Pour d'autres enfin, la possibilitd d'utiliser le DNA natif comme "primer" varie suivant la fraction tissulaire consid6rde in ou suivant l'~ge de l'animal: cette propridtd disparal- trait chez les rats adultes et rdapparaitralt apr6s hdpatectomie partielle x4.
Cependant, malgr6 ces divergences dans l'utilisation du DNA, il a 6td signal6 pour ces diffdrents syst6mes enzymatiques, qu'une attaque mdnagde par la ddsoxyribonucldase I (EC 3.1.4.5) augmentait les qualitds de "primer" du DNA natif3,15-17.
Dans le cadre d'une 6tude d'ensemble du mdtabolisme du DNA dans le tissu hdpatique en voie de croissance, nous nous sommes propos6s: (a) de voir si les DNA- polym6rases du foie en hypertrophie compensatrice se comportaient comme l'enzyme purifide de thymus de veau en ce qui concerne le DNA "primer"; (b) de rechercher les mdcanismes qui physiologiquement pouvaient jouer le r61e de la ddnaturation tbermique.
MATI~RIEL ET METHODES
Des rats males de souche WAG, de 250-300 g, ont dtd hdpatectomisds selon la technique d'HIGGINS ET ANDERSON 18 et sacrifids 36 h apr6s, le sacrifice ayant toujours lieu le matin.
Le foie rapidement pr6levd est aussit6t homogdndisd au froid dans 3 vol. d'un milieu contenant 250 ml de saccharose 0.25 M, 15 ml de Tris-HC1 0.2 M (pH 8) et IO ml de KC1 o.15 M. L'bomogdnat est ensuite centrifugal I ou 2 h ~ lO5 ooo xg (40 ooo tours/min, rotor 40). Le surnageant prdlev6 par aspiration est conserv6 ~ --40° et l 'aetivitd de la DNA-polymdrase est dtudide dans ce surnageant suivant la mdthode III de BOLLUM ET POTTER 19.
Au eours d'une premi6re incubation les quatre nucldosides sont phosphorylds en de[3H~TTP, deCTP, deATP, et deGTP par les phosphokinases d'un surnageant de foie en hypertrophie compensatfice dans un milieu contenant: o.I2/*mole de [3H]thymidine marqude sur le cycle (Amersham) soit 5/*C de radioactivitd totale, o.Io/*mole de ddsoxyaddnosine, ddsoxyeytidine et ddsoxyguanosine, Io/*moles d'ATP, 12/*moles de 3-phosphoglyc6rate, IO/*moles de MgC12, 80/,moles de Tris- HC1 (pH 8.0) et 0.5 ml de surnageant de foie en hypertrophie compensatrice pour I ml de volume total. Au bout de I h d'incubation ~ 38°, les protdines sont prdcipitdes par dbullition et le surnageant est recueilli apr6s centrifugation.
L'activitd de la DNA-polymdrase est ensuite testde au cours d'une deuxi6me incubation dans un milieu contenant 0. 5 ml du surnageant prdcddent, 0. 4 ml de surnageant de foie en hypertrophie compensatfice (contenant environ 7 mg de
Biochim. Biophys. Acta, 114 (t966) 338-348

34 ° A. M. DE RECONDO
prot6ines) et 5o ~g de DNA "primer" dans un volume total de i ml. Nous avons utilis~ comme DNA "pr imer" soit du DNA natif, soit du DNA d6natur6 par la chaleur, soit du DNA natif pr6incubd avec du surnageant de foie en hypertrophie compensatrice ou d 'autres milieux biologiques d6pourvus d'activit~ polym6rasique, ceci avant l 'addition des triphosphates.
Apr~s une incubation de I h ~ 38°, on stoppe la r6action en plongeant les tubes dans de la glace et en y ajoutant un volume 6gal d'acide perchlorique ~ IO %. On ajoute 6galement ~ chaque essai 15 o/~g de DNA pour augmenter le volume du culot acido-insoluble et on sdpare syst6matiquement le DNA du RNA suivant la m6thode de SCHMIDT ET TANNHAUSER ~0 en prat iquant une hydrolyse alcaline douce (NaOH o.2 N, 20 h ~ 37°). La radioactivit6 des extraits perchloriques du DNA et du RNA est ensuite d6termin6e en scintillation liquide avee une TriCarb Packard. Les r6sultats sont finalement exprim6s en radioactivit6 incorpor6e en I h par I mg de prot6ines du surnageant, c'est-~-dire en d6sint~grations/min par mg de prot6ines.
Les activit6s des d6soxyribonucldases I e t I I (EC 3.1.4.5 et EC 3.1.4.6 ) ont 6t6 mesur6es selon la technique de KOWLESSAR e~ al. ~1, le ddsoxypentose lib6r6 6tant dos6 par la m6thode de BURTON z2.
L'6tat de d6naturation du DNA a 6t6 suivi par 6tude viscosim~trique/~ l'aide d'un viscosim~tre rotatif Epprecht Rh6omat 15.
La teneur en prot6ines du surnageant a 6t6 6valu6e par dosage de l 'azote l 'autoanalyseur Technicon (m~thode au ph6nol alcalin et ~ l 'hypochlorite de LOGS- DON~).
Le DNA pr~par~ selon la m~thode de KIRBY ~4 ~ partir de thymus de veau nous a 6td fourni par la maison Choay ainsi que la d6soxyribonucl6ase I cristallis6e (d6soxyribonucl6ase de pancr~as de boeuf).
Nous avons utilis6 de la d6soxyribonucl6ase I I purifi6e (Seravac Laboratories, Colnbrook, Angleterre) et l ' inhibiteur spdcifique de la d6soxyribonucl6ase I pr6par6 dans le laboratoire du Professeur LE BRETON par BERGER ET MAY z5 ~ part ir de s6rum de rat.
La [aH]thymidine, marqu6e sur le cycle, nous a fit6 fournie par Amersham et nous l 'avons amen6e dans nos exp6riences ~ une radioactivit6 sp6cifique de 4 ° mC/ mmole.
RESULTATS
Activitd de "primer" du DNA nati] et du DNA ddnaturd par la chaleur Dans ces conditions exp6rimentales, le surnageant de foie en hypertrophie
compensatrice incorpore effectivement beaucoup plus de IaH]thymidine lorsqu'on lui fournit comme module du DNA soumis pr6alablement /t une d6naturation thermique (Tableau I). Cette diff6rence est particuli~rement accentu6e lorsque les surnageants ont 6t6 centrifug6s 2 h ~. lO5 ooo×g. Sur dix-neuf homog6nats expdriment6s (chaque homog6nat correspondant ~ un lot de six rats au minimum), quinze ont r6agi de cette fa~on et quatre seulement ont incorpor6 de la EaHlthymidine aussi bien en pr6sence de DNA natif que de DNA d6natur6.
Au cours de cette 6tude, nous nous sommes aper~us que la fraction RNA, s~par~e selon la m6thode de SCHMIDT ET TANNHAUSER, contenait une certaine
Biochim. Biophys, Acta, 114 (1966) 338-348
DNA "PRIMER" POUR LA DNA-POLYM]~RASE
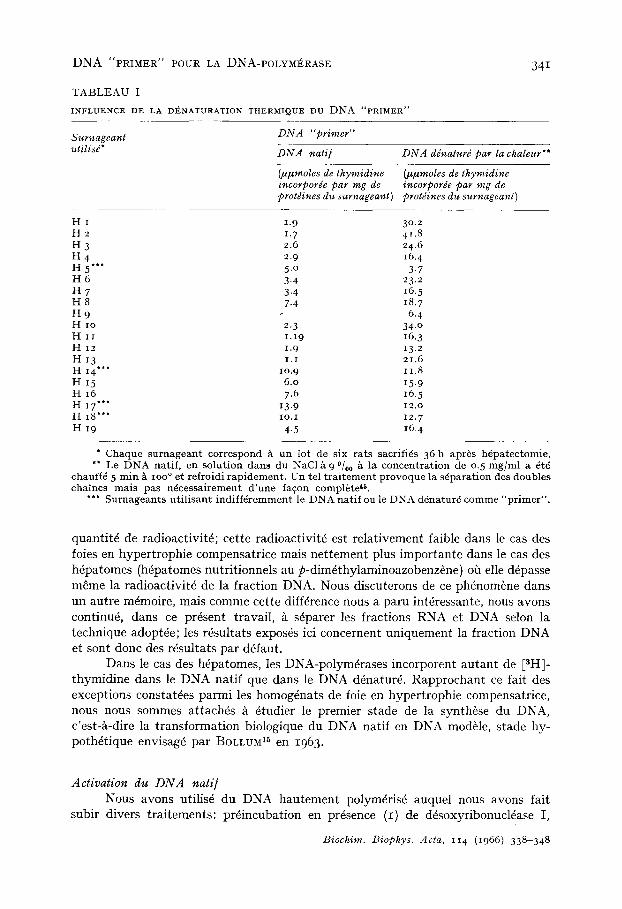
T A B L E A U I
INFLUENCE DE LA DENATURATION THERMIQUE DU D N A "PRIMER"
341
Surnageant utilisd*
DNA "primer"
DNA nati] DNA ddnaturd par la chaleur**
(t~ttmoles de thymidine incorporde par mg de protdines du surnageant)
(IHzmoles de thymidine incorporde par mg de protdines du surnageant)
H I 1.9 30.2 H 2 1. 7 41.8 H 3 2.6 24.6 H 4 2.9 I6. 4 H 5"** 5 .0 3-7 H 6 3.4 23.2 H 7 3.4 16.5 H 8 7.4 I8.7 H 9 -- 6.4 H io 2. 3 34.o H I r 1.19 16.3 H 12 1.9 13.2 H 13 i . i 21.6 H I 4 " " lO.9 11.8 H 15 6.o 15. 9 H I6 7.6 16. 5 H 17"** 13.9 12.o H i8 '*" IO.I 12.7 H 19 4.5 16.4
" Chaque s u r n a g e a n t cor respond ~, un lot de s ix r a t s sacrifiGs 36 h apr~s hdpa tec tomie . ** Le D N A nat i f , cn s o l u t i o n dans du NaC1 A 9 °/00 ~, la c o n c e n t r a t i o n de 0.5 m g / m l a 6td
chauff6 5 m i n ~, ioo ° e t re f ro id i r a p i d e m e n t . Un te l t r a i t e m e n t p rovoque la s6pa ra t ion des doubles cha ines m a i s pas ndces sa i r emen t d ' u n e fa~on compl6 te 4~.
**" S u r n a g e a n t s u t i l i s a n t i n d i f f 6 r e m m e n t le D N A n a t i f ou le DNA dGnatur6 comme " p r i m e r " .
quantit6 de radioactivitG cette radioactivit6 est relativement faible dans le cas des foies en hypertrophie compensatrice mais net tement plus importante dans le cas des h6patomes (h6patomes nutritionnels au p-dim6thylaminoazobenz6ne) oh elle d6passe m~me la radioactivit6 de la fraction DNA. Nous discuterons de ce phdnom6ne dans un autre m6moire, mais comme cette diff6rence nous a paru int6ressante, nous avons cont inue dans ce pr6sent travail, /~ s6parer les fractions RNA et DNA selon la technique adopt6e; les r6sultats expos6s ici concernent uniquement la fraction DNA et sont donc des r6sultats par d6faut.
Dans le cas des h6patomes, les DNA-polym6rases incorporent autant de E~H] - thymidine dans le DNA natif que dans le DNA d6natur6. Rapprochant ce fait des exceptions constat6es parmi les homog6nats de foie en hypertrophie compensatrice, nous nous sommes attach6s A 6tudier le premier stade de la synth6se du DNA, c'est-~-dire la transformation biologique du DNA natif en DNA mod61e, stade hy- poth6tique envisag6 par BOLLUM 15 en 1963.
Activation du DNA nati/ Nous avons utilis6 du DNA hautement polym6ris6 auquel nous avons fait
subir divers traitements: prdincubation en pr6sence (I) de d6soxyribonucl6ase I,
Biochim. Biophys. Acta, 114 (1966) 338-348
~42 A. M. DE RECONDO
(2) de s4rum de rat h@atectomisd (3) de surnageant d'homog6nat de foie en hyper- trophie compensatrice ou (4) de surnageant d'homog6nat d 'h@atome. Nous avons testd ensure les qualitds de "pr imer" de ce DNA, sans le r6isoler au pr~alable de peur de d6truire un 6tat physiologiquement actif et particuli&rement instable. De cette fa£on, nous avons pu comparer l 'activit6 de "primer" du DNA natif ainsi trait~ & celle du m6me DNA d~natur~ par la chaleur. Voici l'essentiel de nos r6sultats:
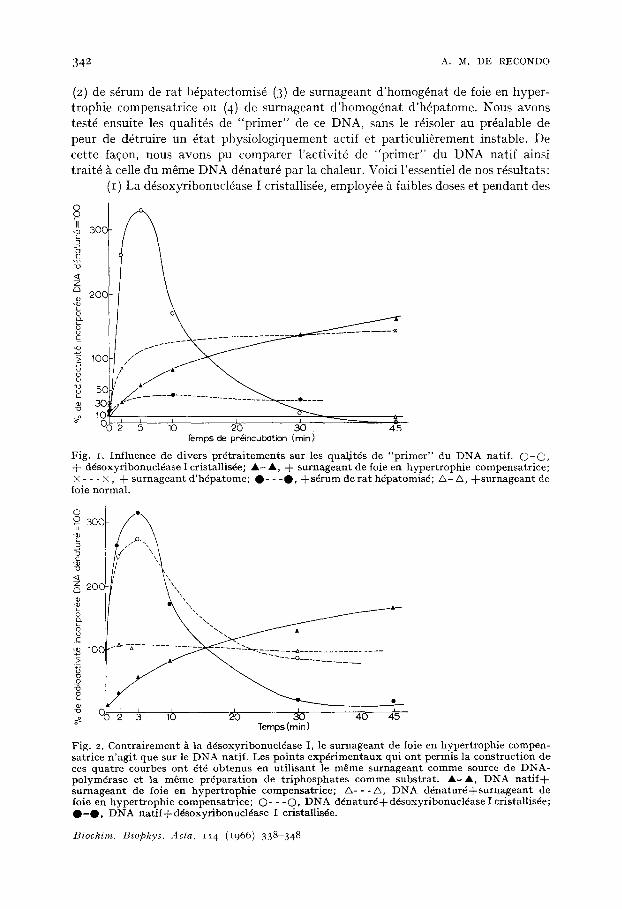
(I) La d6soxyribonucldase I cristallis6e, employee h faibles doses et pendant des
(5
OO . . . . . . . . . . -X C . . . . . . . . . . . . . . . . . . . - - - -
10 // 0
Temps de pr~incubetion (rain)
Fig. I, In f luence de d ive rs p r 6 t r a i t e m e n t s sur les qua t i t6s de " p r i m e r " du D N A nat i f . 0 - 0 , + ddsoxyr ibonuc l~ase I cristall is~e; A - A, + s u r n a g e a n t de foie en hype r t roph i e compensa t r i ce ; × - - - × , + s u r n a g e a n t d ' h 6 p a t o m e ; O - - - O , + s 6 r u m de r a t h6pa tomisd ; & - A, + s u r n a g e a n t de
foie normal .
0.04/\ "~ / / .~-. \
// ' L ' , g 2oo-! \ " , ,
°/ 2 100 - ' ~ -Z . . . . . . . . . . ~ _ _ " - ~ _ _ ~ . . . . . . . . . . . . . . .
Temps (rain)
Fig. 2. C o n t r a i r e m e n t & la d6soxyr ibonuc l6ase I, le s u r n a g e a n t de foie en hype r t roph i e c o m p e n - sa t r ice n ' a g i t que su r le D N A na t i f . Les p o i n t s e x p 6 r i m e n t a u x qu i o n t pe rmis la c o n s t r u c t i o n de ces q u a t r e courbes o n t 6t6 o b t e n u s en u t i l i s an t le m ~ m e s u r n a g e a n t comrne source de D N A - po lym6ra se et la m 6 m e p r6pa ra t i on de t r i p h o s p h a t e s c o m m e s u b s t r a t . A - A , D N A n a t i f + s u r n a g e a n t de foie en h y p e r t r o p h i e compensa t r i ce ; & - - - A , D N A d d n a t u r d + s u r n a g e a n t de foie en h y p e r t r o p h i e compensa t r i c e ; O - - - O , D N A d d n a t u r d + d d s o x y r i b o n u c l d a s e I cr is tal l is6e; O - O , D N A n a t i f + d d s o x y r i b o n u c l d a s e I cristallisde.
Biochim. Biophys. Acta, 114 (1966) 338-348

D N A "PRIMER" POUR LA DNA-POLYMI~RASE 343
temps tr~s courts augmente consid6rablement la radioactivitd incorpor6e dans du DNA natif: mais, d~s qu'on prolonge le traitement, on voit d~croitre les qualit6s de "pr imer" des produits form6s. Ces r6sultats sont en accord avec ceux publi~s par GOUTIER ET BOLOGNA 9, MANTSAVINOS ET CANELLAKIS 17, SMELLIE e t a / . 2,2e, SAR- KAR 2v et BOLLUM 15,16. (Fig. I).
TABLEAU II
I N F L U E N C E DE L ' I N H I B I T E U R DE LA D~SOXYRIBONUCL~ASE I SUR LA TRANSFORMATION DU DNA NATIF EN D N A " P R I M E R "
Les r6sultats sont exprim~s en d6sint6grat ions/min par mg de protdines du surnageant.
D N A prdincubd D N A prdincubd D N A prdincubd 2 rain avec de 45 rain avec un 45 rain avec du la ddsoxyribonu- surnageant de surnageant cldase I /oie en hypertro- d'hdpatome
phie compensatrice
E n absence d ' inh ib i teur 363 °
En pr6sence d ' inh ib i teur i i i
Inhib i teur agissant apr~s le pr~trai- 3225 t emen t du DNA "pr imer" pendan t la polymdrisat ion du DNA ndoformd
1285 128o I675 1855 175o 153 ° 144 ° 133o 1738
(2) Une pr6incubation du DNA natif avec du s6rum de rat h6patectomis6, dont nous avons constat6 l 'hyperactivit6 d6soxyribonucldasique I (r6f. 28) et dans lequel de nombreux auteurs ont recherch6 un facteur stimulant les multiplications cellulaires 29-~3, augmente l~g~rement les qualit6s de module d'un DNA hautement polym6ris6. (Fig. I). Par contre le s6rum de rat normal n'a pratiquement pas d'effet.
(3) Une pr~incubation du DNA natif avec du surnageant de foie en hyper- trophie compensatrice augmente en fonction du temps les qualit6s de "primer" du DNA qui peut devenir aussi actif ou plus actif que le DNA d~natur~ par la chaleur (rig. ~).
(4) De m~me, une prdincubation du DNA natif avec o.I ml de surnageant d'h6pa- tome nutritionnel ~ croissance lente augmente consid~rablement les qualit6s de mod~le du DNA utilis6, alors que la m~me quantit6 de surnageant de foie normal n'a aucun effet (Fig. I).
Ces r~sultats permettent de constater que certains facteurs biologiques sont capables de transformer le DNA natif en DNA "primer" et sont aussi efficaces ce point de rue que le proc~d6 artificiel de la d6naturation thermique.
Mdcanismes impliquds clans l'activalion du DNA nati/ par les surnageants de ]oie en hypertrophie compensatrice et d'hdpatome
Ddsoxyribonucldases I e t II . Etant donn~ l'action favorisante de la d6soxy- ribonucl6ase I sur la polym6risation du DNA in vitro, on pouvait envisager que les surnageants de foie en hypertrophie compensatrice et que les surnageants d'h6patomes agissaient sur le DNA natif par l'intermddiaire des d~soxyribonucldases. Nous avons pu v6rifier qu'il n'en 6tait rien.
Biochim. Biophys. Acta, 114 (1966) 338 348

344 A. M. DE RECONDO
(a) Le dosage syst6matique des activitds d6soxyribonucl6asiques I et I I de ces surnageants nous a permis de constater que l 'activit6 de la ddsoxyribonucl6ase I est en grande partie masqude par l ' inhibiteur tissulaire ~ et varie peu tandis que la d6soxyribonucl6ase II , qui n'est pas touch6e par l 'hypertrophie compensatrice, est cinq fois plus active dans le cas des h@atomes aS.
(b) L'allure des ph6nom~nes provoquds par un pr6traitement par la ddsoxy- ribonucl6ase I d'une part, et par un surnageant de foie en hypertrophie compen- satrice ou d 'h@atome d 'autre part, est tr6s diff6rente. La d6soxyribonucl6ase I a une action extr6mement rapide et brutale qui s'inverse d6s que l 'enzyme at taque trop profond6ment le DNA. Par contre les surnageants de foie en hypertrophie compensatrice et les surnageants d'h6patomes ont une action lente et progressive (Fig. I e t 2).
(c) En utilisant l ' inhibiteur sp6cifique de la d6soxyribonucl6ase I pr@ar6 dans le laboratoire du Professeur E. LE BRETON par G. BERGER Lr P. MAY, nous avons constat6 que cet inhibiteur bloquait compl6tement l 'action de la d6soxyribo- nucl6ase I alors qu'il ne touchait prat iquement pas Faction des surnageants de foies en hypertrophie compensatrice ni celle des surnageants de foies tumoraux (Tableau II) .
T A B L E A U I I I
IMPORTANCE D'UN pHI~NOMI'NE D'ADDITION EN BOUT DE CHA~NE
Les milieux a et b on t 6t6 pr6par6 simultalx6ment avec le m~me surnagean t de foie en hyper t rophie compensatr ice comme source de phosphokinases . Les r6sul tats sont exprim6s en d6sint~grations/ min par mg de prot6ines.
En prdsence des quatre En prdsence de de[3H]TTP nucldotides seul (a) (b)
Expdriences H1~ Hi7 H18 His H17 Hi8
DNA nat i f 555 934 1288 555 78o 9oo DNA nat i f pr6incub~ avec du surna- geant de foie ell hyper t roph ie compensat r ice 853 2523 2675 845 197 ° 194 ° D N A nat i f pr6incub6 avec du surna- geant d 'h~pa tome 1375 113 ° D N A d6natur6 par la chateur 147o 1176 lO95 563 830 820
(d) La d6soxyribonucl6ase I I tr~s active dans les surnageants d 'h@atomes 35 n 'a t taque prat iquement pas le DNA "pr imer" dans les conditions d'action de la DNA-polymfrase. Nous avons pu constater, en outre, comme SMELLIE 2~ et RICHARD- SON et al. 3 qu'un pr6traitement du DNA natif par une d6soxyribonucl6ase I I purifi6e diminuait l ' incorporation de thymidine dans ce DNA (Tableau IV). L'action de cette enzyme ne peut donc expliquer la transformation du DNA natif en DNA "pr imer" par les surnageants d'h6patomes.
Nucldotidyltrans[drase terminale. Plusieurs auteurs a~-~9 ont constat6 la prfsence dans les pr@arations de DNA-polym6rase d'une nucl6otidyltransffrase terminale
Biochim. Biophys. Acta, 114 (1966) 338-348
D N A "PRIMER" POUR LA DNA-POLYMt~RASE 345
capable de fixer l 'un des quatre nucl4otides au bout d'une des chalnes du DNA "primer". L'activit4 de cette enzyme est diminu4e par l 'addition des trois autres nucl4otides.
Dans nos conditions exp4rimentales, une grande partie de la radioactivit4 incorpor4e est due ~ l 'addition de [3H]thymidine en bout de chalne (Tableau uI). Cette incorporation est cependant augment4e par l 'addition des autres nucl4otides non radioactifs. La pr4incubation du DNA natif avec du surnageant, soit de foie en hypertrophie compensatrice, soit d'h4patome, augmente ce ph4nom~ne mais augmen- te aussi parall~lement la polym4risation des quatre nucl4otides. Lorsqu'on emploie un DNA d4natur4 comme "primer", seule la r4action de polym4risation est augment4e. Notre pr4paration contient donc & la fois la nucl4otidyltransf4rase terminale et la DNA-polym4rase proprement dire (nucl6otidyltransf4rase replicative).
T A B L E A U IV
IMPORTANCE DES GROUPEMENTS 3'-HYDROXYLS TERMINAUX DU DNA "PRIMER"
La d6soxyribonucl4ase II a 4t4 utilis4e & la dose de 4.5 unit4s par essai. Les milieux a et b ont 4td pr4par6s s imul tan4ment avec le m6me surnageant de foie en hypertrophie compensatrice comme source de phosphokinuses. Les r4sultats sont exprim4s en d4sint4grations/min par mg de prot4ines.
DNA "primer" En prdsence des qualm En presence de de ?HI T T P nucldotides seul (a) (b)
DNA nat i f 1288 DNA pr4incub4 avec du surnageant de foie en hypertrophie compensatrice 2675 DNA pr4incub4 avec du surnageant de foie ell hyper t rophie compensatr ice plus d4soxyribonucMase lI apr~s pr4incubation 1215 DNA nat i f pr~incub4 avec de la d4soxyribonucldase [I 718
900
194 o
813
465
Enzyme libdrant des 3'-hydroxyls terminaux. Puisque la pr4incubation du DNA natif avec un surnageant de foie en hypertrophie compensatrice ou un surnageant d 'h4patome augmente & la fois le ph4nom+ne de polym4risation et celui d'addition en bout de chaine, on est conduit & envisager la pr4sence dans ces surnageants d'une enzyme non encore identifi4e lib4rant des sites actifs de la molecule de DNA "primer" . Ces sites permettraient l 'action ult4rieure des deux enzymes de synth+se.
Or, RICHARDSON et al.3,4,7,4°, 41 ont isol4, & part ir des preparations de DNA- polym4rase d 'E. coli, une DNA-phosphatase exonucMase qui augmente l 'activit4 de la DNA-polym4rase purifi4e. Cette enzyme aurait pour effet de catalyser une hydrolyse propag4e d'une des chalnes du DNA ~t part ir du 3'-phosphoryl terminal, lib4rant des 5'-mononucl4otides et laissant en bout de chalne un 3 '-hydroxyl terminal n4cessaire/t la resynth~se du DNA.
Elle at taque le DNA & double h41ice dont elle n'hydrolyse que 45-50 % laissant finalement deux moiti4s de simple chaine termin4es par un 3 '-hydroxyl et r4unies encore entre elles par une faible portion de double chaine. Pour RICHARDSON, la resynth+se du DNA d4marrerait & part ir de ces 3'-hydroxyls, l 'une des h41ices
Biochim. Biophys. Acta, 114 (1966) 338-348

34 6 A . M . DE RECONDO
servant d 'amorce ou de "pr imer" l 'autre servant de mod61e ou de " templa te" pour l 'ordre de succession des bases.
D'autres auteurs 26 ont signal6 l ' importance de ces groupements 3 '-hydroxyls terminaux dans l 'initiation de la replication, importance qui peut expliquer l 'action stimulante de la d6soxyribonucl6ase I puisque cette enzyme coupe la chaine de DNA en lib6rant des 3'-hydroxylsa, 2~.
Effectivement le surnageant de foie en hypertrophie compensatrice pr6sente deux points communs avec la DNA-phosphatase exonucl6ase d6crite par RICHARD- SON: (a) il n'agit que sur le DNA natif n 'ayant aucune influence sur le DNA d6j~ d6natur6 par la chaleur ainsi qu'on peut le voir sar la Fig. 2; (b) il semble lib6rer sur la chalne du DNA des 3'-hydroxyls n6cessaires ~ l 'initiation de la replication car l 'action post6rieure de la d6soxyribonucl6ase II , qui coupe la chaine en lib6rant des 5'-hydroxyls, supprime ses effets (Tableau IV).
CONCLUSION
Comme la DNA-polymdrase purifi6e de thymus de veau, la DNA-polym6rase de foie en hypertrophie compensatrice utilise pr6f6rentiellement comme mod61e du DNA d6natur6 par la chaleur. I1 existe cependant dans les surnageants de foie en hypertrophie compensatrice et dans les surnageants d 'h6patomes des facteurs capables de transformer le DNA hautement polym6ris6 en DNA "pr imer" d'une fa~on aussi efficace que le proc6d6 artificiel de la d6naturation thermique.
Quels sont ces facteurs? Dans ce travail pr61iminaire, nous pensons avoir 61imin6 l ' intervention des d6soxyribonucl6ases I e t II . L 'augmentat ion parall61e du ph6nombne d'addition en bout de chaine et du ph6nom6ne de polym6risation implique l 'action d'une enzyme lib6rant les sites auxquels peuvent s'accrocher les d6soxyribonucl6otides: cette enzyme particuli6rement active dans les tissus en voie de croissance normale ou tumorale est-elle du type de la DNA-phosphatase exo- nucl6ase d6crite par RICHARDSON chez E. coli? Une telle hypoth6se serait particu- li6rement s6duisante car le DNA synth6tis6 en pr6sence d'un "pr imer" partiellement d6grad6 par la DNA-phosphatase exonucl6ase semble poss6der toutes les caract6ris- tiques du DNA naturel* alors que le produit form6 en pr6sence d'un DNA ~ double h61ice diff+re par ses propri6tds physico-chimiques et ultra-structurales du DNA normal42,43.
Nous essayons actuellement d'isoler la fraction capable d 'activer le DNA natif et de faire l '6tude physico-chimique du DNA synth6tis6 ~ partir d 'un tel "pr imer" .
Note: En cours de r6daction, nous avons eu connaissance d 'un article de MANTSAVINOS a4 concernant l 'enzyme partiellement purifi6e de foie en hypertrophie compensatrice. Ses conclusions diff6rent en plusieurs points des n6tres:
(I) D'apr6s lui, cette enzyme utiliserait aussi bien le DNA natif que le DNA d6natur6 et se rapprocherait en ce sens de la polym6rase d 'E. coli, il envisage ce- pendant la pr6sence de nucl6ases contaminantes.
(2) L'activit6 de sa pr6paration est net tement diminu6e par la suppression de l 'un des quatre nucl6osides triphosphates, la DNA-polym6rase replicative pr6domine donc dans ses pr6parations tandis que la nucl6otidyltransf6rase terminale est moins active que dans nos extraits.
Biochim. Biophys. dc/a, 114 (1966) 338-348

D N A "PRIMER" POUR LA DNA-POLYM]~RASE 347
Malgr4 ces divergences, un point int4ressant semble en accord avec nos propres r4sultats et les confirmer: MANTSAVINOS indique, en effet, que les d4soxyribonucl4o- sides 5'-triphosphates sont incorpor4s pr4f4rentiellement, ce qui concorde avec l ' importance des 3'-hydroxyls terminaux du DNA "primer".
REMERCIEMENTS
Ce travail a 4t4 r4alis4 sous la direction du Professeur E. LE BRETON, avec la collaboration technique de Mesdemoiselles M. HARDY et O. FICHOT et a b4n4fici6 de l 'aide d'une subvention de la Caisse des D4p6ts et Consignations.
Nous tenons ~ remercier G. BERGER qui nous a fourni l ' inhibiteur sp4cifique de la ddsoxyribonucl4ase I et les Docteurs Y. MOUL~, CH. FRAYSSINET et P. MAY qui nous ont aid4 dans l'41aboration et la r4daction de ce travail.
R~SUM~ Le syst~me des DNA-polym4rases (d4soxyribonucl4osidetriphosphate:DNA
d4soxyribonucl4otidyltransf4rase, EC 2.7.7.7) du foie en hypertrophie compensatrice est test4 quant h sa capacit4 ~ utiliser comme "pr imer" le DNA natif ou le DNA simple chaine.
Dans les conditions exp6rimentales adopt4es, ce syst~me enzymatique r4agit comme la DNA-polym4rase purifi4e de thymus de veau et utilise pr4f4rentiellement comme "pr imer" le DNA d4natur4 par la chaleur.
Ii peut cependant utiliser du DNA hautement polym6ris4 lorsque celui-ci a 4t4 soumis h divers pr4traitements: prdincubation en pr4sence de faibles doses de d4soxyribonucl4ase I cristallis~e, pr4incubation en pr4sence de sdrum de rat h4- patectomis4, de surnageant de foie en hypertrophie compensatrice ou de surnageant d 'h4patome.
An bout de 30-45 min, le DNA natif pr4incub4 en pr4sence de surnageant de foie en hypertrophie compensatrice ou de surnageant d 'h4patome peut ~tre plus actif comme "pr imer" que le m~me DNA d4natur4 par la chaleur.
Ce ph4nom~ne n'est pas dfi h l 'action des d4soxyribonucl4ases I (EC 3.1.4.5) et I I (EC 3.1.4.6) de ces surnageants: en effet, l ' inhibiteur sp4cifique de la d4soxy- ribonucl4ase I ne le bloque pas; quant h la d4soxyribonucl4ase I I tr~s active dans les surnageants d'h4patomes, elle n 'a t taque prat iquement pas le DNA primer dans les conditions d'action de la DNA-polym4rase, en outre une d4soxyribonucl4ase I I purifi4e diminue les qualitds de "pr imer" du DNA natif. On est conduit ~ envisager l 'action d'une enzyme lib4rant des sites actifs de la mol4cule de DNA "primer", ces sites permettraient l 'action ult4rieure de la nucl4otidyltransf4rase terminale et de la DNA-polym4rase proprement dire.
Cette enzyme pr4sente certaines analogies avec la DNA-phosphatase exo- nucl4ase d4crite par RICHARDSON et al. chez E. coll.
BIBLIOGRAPHIE
I F. J. BOLLUM, ./. Biol. Chem., 234 (1959) 2733 2 H. M. I~EIR, t3. BINNIE ET R. M. S. SMELLIE, Biochem. J., 82 (1962) 493.
Biochim. Biophys. Acta, 114 (1966) 338-348

348 A . M . DE RECONDO
3 C. C. RICHARDSON, C. L. SCHILDKRAUT ETA. KORNBERG, Cold Spring Harbor Syrup. Quant. Biol., 28 (1963) 9.
4 C. C. RICHARDSON, R. B. INMAN ETA. KORNBERG, J. iWlol. Biol., 9 (1964) 46. 5 F. J. BOLLUM, dr. Biol. Chem., 235 (196o) 2399. 6 F. J. BOLLUM, darts J. J. MITCHELL, The Cell Nucleus, But te rwor th , London, 196o, p. 60. 7 C. C. RICIIARDSON, C. L. SCHILDKRADT, ]7I. V. APOSHIAN ETA. •ORNBERG, J . Biol. Chem.,
239 (1964) 222. 8 R. MANTSAVINOS ET E. S. CANELLAKIS, dr. Biol. Chem., 234 (1958) 628. 9 R. GOUTIER ET [. BOLOGNA, Biochim. Biophys. Aeta, 72 (1963) 4 o.
io G. GIUDICE ET G. D. NOVELLI, Biochem. Biophys. Res. Commun., 12 (1963) 383 • I I E. D. GRAY, S. M. WEISSMAN, J. RICHARDS, D. BELL, H. M. I{EIR, R. M. S. SMELLIE ET
J. N. DAVlDSON, Biochim. Biophys. Acta, 45 (196o) I I I . I2 N. L. R. BUCHER, J. Cellular Comp. Physiol., 62 (1963) 69. 13 I{. M. BEHKI ET W'. C. SCHNEIDER, Biochim. Biophys. Acta, 68 (1963) 34. 14 -x{. n . MUKUNDAN, A. DEVI ET N. K. SARKAR, Biochem. Biophys. Res. Commun., i i (1963) 353. 15 F. J. BOLLUM, d r. Cellular Comp. Physiol., 62 (1963) 61. 16 b'. J. BOLLUM, darts J. N. DAVIDSON ET W. E. COHN, Progress in Nucleic Acid Research, Vol. i,
Academic Press, New York, 1963, p. I. 17 R. MANTSAVINOS E T E . S. CANELLAKIS, Cancer Res., 19 (1959) 1239. 18 G. M. HIGGINS ET R. M. ANDERSON, A . M . A . Arch. Palhol., 12 (1931) 186. 19 F. J. BOLLUM ET V. R. POTTER, J. Biol. Chem., 233 (1958) 478 . 20 G. SCHMIDT ET S. J. TANNItAUSER, d r. Biol. Chem., 161 (1945) 83. 21 O. D. KOWLESSAR, K. I. ALTMAN ET L. H. HEMPELMANN, Arch. Biochem. Biophys., 54 (1955)
355. 22 K. BURTON, Biochem. J., 62 (1956) 315 • 23 E. E. LOGSDON, Ann. N .Y . Acad. Sci., 87 (196o) 8Ol. 24 K. S. KIRBY, Biochem. J., 66 (1957) 495. 25 G. BERGER ET P. MAY, Compt. Rend., 258 (1964) lO93. 26 R. M. S. SMELLIE, Exptl. Cell Res., Suppl., 9 (1963) 245. 27 N. K. SARKAR, Arch. Biochem. Biophys., 93 (1961) 328. 28 A. M. DE RECONDO, CH. FRAYSSINET ET P. MAY, Compt. Rend., 255 (1962) 2667. 29 H. FRIEDRICH-FREKSA ET F. G. ZAKI, Z. Natur/orsch., 96 (1954) 394- 3 ° R. W. BEGG ET H. STICH, Abstr. 7th International Congress o/Cancer, London, 1958, p. 13o. 31 H. "~VRBA, H. RANZ ET M. RIPOLL-GOMEZ, Naturwissenschaften, 47 (196o) 306. 32 S. ADIBI, K. E. PASHRIS ET A. CANTAROW, Exptl. Cell Res., 18 (1959) 396. 33 R. KOHN, Exptl. Cell Res., 14 (1958) 228. 34 R. N. FEENSTEIN, J. Biol. Chem., 235 (196o) 733. 35 A. M. DE I~ECONDO ETCH. FRAYSSINET, J. Physiol. Paris, 56 (1964) 43 °. 36 J. S. KRAKOW, C. COUTSOGEORGOPOULOS E T E . S. CANELLAKIS, Biochim. Biophys. Acta, 55
(1962) 639. 37 H. M. KEIR ET M. J. SMITH, Biochim. Biophys. Acta, 68 (1963) 589 . 38 F. J. BOLLUM ET R. B. SETLOW, Biochim. Biophys. Acta, 68 (1963) 599. 39 F. J. BOLLUM, E. GROENIGER ET M. YONEDA, Proc. Natl. Acad. Sei. U.S., 51 (1964) 853. 4 ° C. C. RICHARDSON ETA. KORNBERG, J. Biol. Chem., 239 (1964) 242. 41 C. C. RICHARDSON, I. 1(. LEHMAN ET A. KORNBERG, J. Biol. Chem., 239 (1964) 251. 42 C. L. SCHILDKRAUT, C. C. RICHARDSON ET A. KORNBERG, J. Mol. Biol., 9 (1964) 24. 43 R. B. INMAN, C. L. SCHILDKRAUT ETA. KORNBERG, J. Mol. Biol., i i (1965) 285. 44 R. MANTSAVlNOS, J. Biol. Chem., 239 (1964) 3431. 45 M. R. CHAVALLIER ET G. BERNARDI, J. Mol. Biol., I I (1965) 658-
Biochim. Biophys. Acta, 114 (1966) 338-348





























