Embed Size (px)
Citation preview

262
BBA 52013
Biochimtcu et Btophystco Actcr X36 (1985) 262- 2h6
ElSZ\Wr
Metabolism of plastid terpenoids: lycopene cyclization by Capsicum chromoplast
membranes
Bilal Camara ‘, Odette Dogbo ‘, Alain d’Harlingue ‘, Hans Kleinig ’ and Renk Monkger ’
‘I Laboratoire de RPgulattons MPtaboliques et Diff&-enciution des Plastes, lJrtiuer.~ttP Pierre et Mane Curie. 4 P&e Jussreu,
75230 Paris Cddex 05 (France), und h Institut ftir Biologic II, Zellbtologie, Albert-Ludwigs Urticersitiit, Schiinzlestrusse I,
D-7800 Freihurg IF. R. G.)
(Received March 27th. 1985)
(Revised manuscript received June 14th, 1985)
Key words: Terpenoid: Carotenoid biosynthesis; Lycopene cyclase; Chromoplast membrane: (C‘. clnnuum)
The cyclization of lycopene to /3,/3-carotene (p-carotene) was studied using chromoplast membranes prepared from Cupsicum fruits. The enzymic activity is tightly bound to the membranes and, under optimal conditions, lycopene is cyclized to p-carotene without accumulation of the intermediate /3,+carotene (y-carotene). In this system, no formation of (6,R)-P,r-carotene (a-carotene) occurs. The optimal pH for lycopene cyclization is 6.8. Nucleotide cofactors are not required for activity. The enzymic activity is stimulated in the presence of Tween 80 in contrast to the inhibitory effect of Triton X-100. Thiol reagents such as mersalyl, N-ethyl- maleimide and p-chloromercuribenzoate strongly inhibit the cyclization of lycopene to p-carotene. Oc- tyldiethylamine and dodecyl diethylamine are potent inhibitors of lycopene cyclase activity from Cupsicum chromoplast membranes.
Introduction
It has recently been shown that plastidial mem- branes from higher plants perform the later reac-
tions (desaturation, isomerization, cyclization.. ) of carotenoid biosynthesis [1,2]. The cyclization
reaction, i.e., the conversion of neurosporene or
lycopene to P-carotene, represents a key step in that neither neurosporene nor lycopene is in- tegrated in photosynthetic membranes. Further-
Abbreviations: AMO-1618, 2’-isopropyl-4’-(trimethylam-
monium chloride)-5’-methylphenylpiperidine-l-carboxylate;
pCMB: p-chloromercuribenzoate; p-carotene, p,p-carotene:
u-carotene, (6’R )-/3, c-carotene; y-carotene, p,+carotene:
canthaxanthin, /3,B-carotene-4,4’-dione; lycopene. $,+caro- tene; neurosporene, 7,8-dihydro-$,$-carotene.
more, their accumulation in plastids leads to pre- cipitation as lycopene crystals [3].
Using Cupsicum chromoplasts, we have shown
that neurosporene as well as lycopene can be used as a substrate for cyclization but, at a definite
concentration, there was a preference for lycopene cyclization [4]. In pursuing these studies, we now present detailed experiments which were aimed at further defining some properties of lycopene cyclase activity from Cupsicum chromoplast mem- branes.
Materials and Methods
Preparation of chromoplast membranes. The chromoplast membranes obtained from isolated Capsicum chromoplasts as described previously [5]
0005-2760/85/$03.30 (3 1985 Elsevier Science Publishers B.V. (Biomedical Division)
263
were subjected to osmotic shock in a medium
containing 1 mM MgCl,, 10 mM dithiothreitol,
and 50 mM Tris-HCl buffer (pH 7.6). The result- ing homogenate was frozen at -20°C overnight. The thawed membrane suspension was pelleted at
100000 x g for 2 h and finally suspended in 50
mM Tris-maleate buffer (pH 6.8). The protein content was determined according to a previously
described procedure [6]. Reaction conditions and analytical procedures.
The cyclization of lycopene was determined by
measuring the incorporation of lycopene into /3-
carotene. The standard incubation medium con-
tained, in 1 ml final volume, chromoplast mem- branes equivalent to 2 mg protein, 0.25 M sorbitol, 5 mM MgC12, 2 mM MnCl,, 5 mM dithiothreitol,
2 PM FAD, 1 mM NADP, 5 mg Tween 80,
480000 dpm 15,15’-[3H]lycopene (160 Ci/mol), 50 mM Tris-maleate buffer (pH 6.8). At the end of the incubation time, the reaction was stopped with
4 ml of chloroform/methanol (2: 1, v/v). p- Carotene purification and determination were as
described elsewhere [7]. As an alternative test for purity, the labelled p-carotene was converted to
canthaxanthin according to a procedure com- municated by Britton and Goswami. In this proce- dure, p-carotene was treated with sodium chlorate
in the presence of a catalytic amount of iodine. Canthaxanthin was separated from unreacted p- carotene by thin-layer chromatography on silica gel using petroleum ether/diethyl ether (98 : 2,
v/v). Chemical modifications of chromoplast mem-
branes with reagents. Membranes obtained from the above freeze-thaw procedure and those derived
from freshly prepared chromoplast suspensions
were kept on ice for 30 min in a medium contain-
ing 1-5 mM p-chloromercuribenzoate (pCMB), mersalyl, and N-ethylmaleimide, dissolved in 100 mM Tris-HCl buffer (pH 8). Treated membranes were then washed three times by centrifugation at 100000 X g for 15 min in the presence of cold Tris-HCl buffer.
Inhibition studies. Octyldiethylamine and dode- cyl diethylarnine prepared by reductive amination of the parent aldehydes [8], nicotine and AMO- 1618 (Calbiochem), were tested as lycopene cyclase inhibitors using the incubation medium described above.
20
f L Y
E
op
1
PROTEIN C mg )
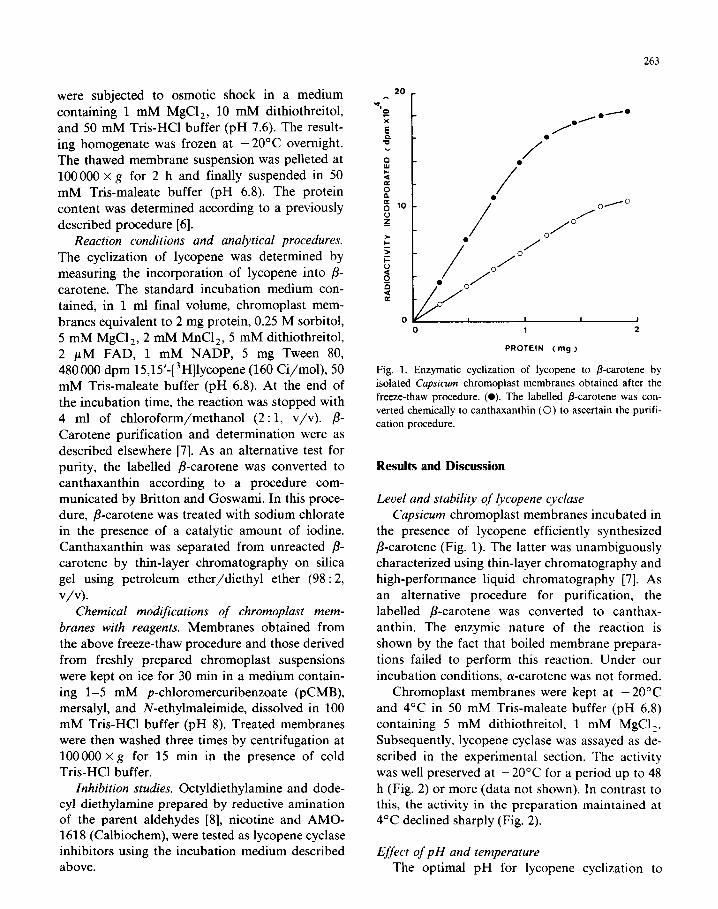
Fig. 1. Enzymatic cyclization of lycopene to p-carotene by
isolated Capsicum chromoplast membranes obtained after the
freeze-thaw procedure. (0). The labelled p-carotene was con-
verted chemically to canthaxanthin (0) to ascertain the purifi-
cation procedure.
Results and Discussion
Level and stability of lycopene cyclase Capsicum chromoplast membranes incubated in
the presence of lycopene efficiently synthesized
P-carotene (Fig. 1). The latter was unambiguously characterized using thin-layer chromatography and high-performance liquid chromatography [7]. As
an alternative procedure for purification, the labelled p-carotene was converted to canthax- anthin. The enzymic nature of the reaction is
shown by the fact that boiled membrane prepara-
tions failed to perform this reaction. Under our incubation conditions, a-carotene was not formed.
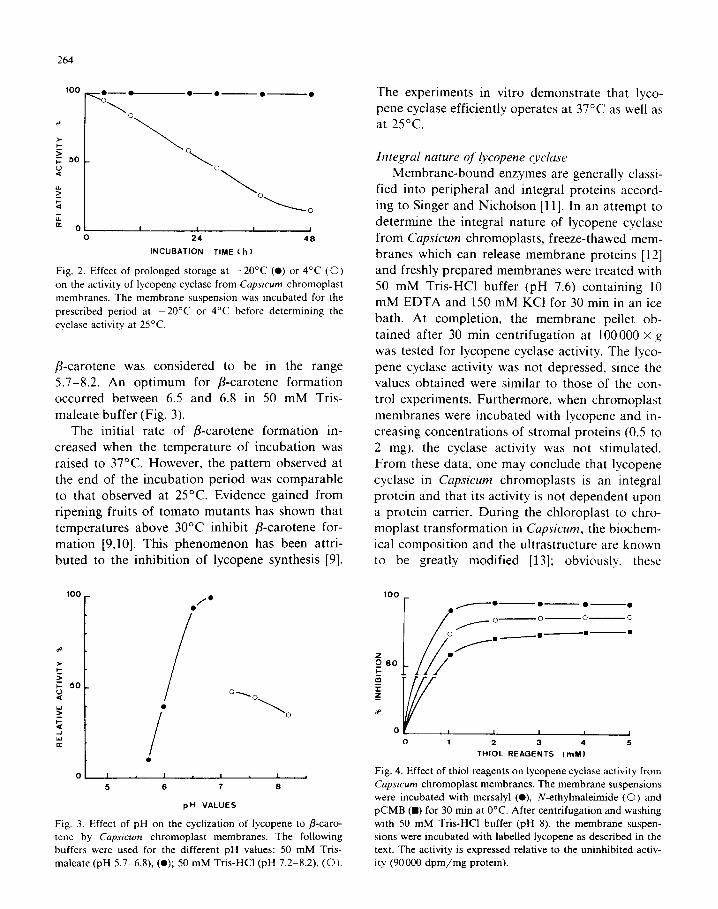
Chromoplast membranes were kept at -20°C and 4°C in 50 mM Tris-maleate buffer (pH 6.8) containing 5 mM dithiothreitol, 1 mM MgCl?.
Subsequently, lycopene cyclase was assayed as de- scribed in the experimental section. The activity
was well preserved at - 20°C for a period up to 48
h (Fig. 2) or more (data not shown). In contrast to this, the activity in the preparation maintained at 4’C declined sharply (Fig. 2).
Effect of pH and temperature The optimal pH for lycopene cyclization to
264
100
I‘
.-A.-.-.-.
O\_ 0
\ o\, \ Oio
INCUBATION TIME (h 1
Fig. 2. Effect of prolonged storage at - 20°C (0) or 4°C (0)
on the activity of lycopene cyclase from Cqsicum chromoplast
membranes. The membrane suspension was incubated for the
prescribed period at -20°C or 4°C before determining the
cyclase activity at 25°C.
P-carotene was considered to be in the range
5.7-8.2. An optimum for p-carotene formation
occurred between 6.5 and 6.8 in 50 mM Tris-
maleate buffer (Fig. 3). The initial rate of P-carotene formation in-
creased when the temperature of incubation was
raised to 37°C. However, the pattern observed at the end of the incubation period was comparable to that observed at 25°C. Evidence gained from
ripening fruits of tomato mutants has shown that temperatures above 30°C inhibit P-carotene for-
mation [9,10]. This phenomenon has been attri- buted to the inhibition of lycopene synthesis [9].
-\,
pii VALUES
Fig. 3. Effect of pH on the cyclization of lycopene to p-caro-
tene by Cqsicum chromoplast membranes. The following
buffers were used for the different pH values: 50 mM Tris-
maleate (pH 5.7-6.8), (0); 50 mM Tris-HC1 (pH 7.2-8.2) (0).
The experiments in vitro demonstrate that lyco- pene cyclase efficiently operates at 37°C as well as
at 25°C.
Integral nature of lycopene cyclase
Membrane-bound enzymes are generally classi-
fied into peripheral and integral proteins accord-
ing to Singer and Nicholson [ll]. In an attempt to
determine the integral nature of lycopene cyclase from Capsicum chromoplasts, freeze-thawed mem-
branes which can release membrane proteins [I 21 and freshly prepared membranes were treated with 50 mM Tris-HCl buffer (pH 7.6) containing 10 mM EDTA and 150 mM KC1 for 30 min in an ice
bath. At completion, the membrane pellet ob-
tained after 30 min centrifugation at 100000 x R
was tested for lycopene cyclase activity. The lyco- pene cyclase activity was not depressed, since the
values obtained were similar to those of the con-
trol experiments. Furthermore. when chromoplast membranes were incubated with lycopene and in-
creasing concentrations of stromal proteins (0.5 to 2 mg), the cyclase activity was not stimulated. From these data, one may conclude that lycopene
cyclase in Capsicum chromoplasts is an integral protein and that its activity is not dependent upon a protein carrier. During the chloroplast to chro-
moplast transformation in Capsicum, the biochem- ical composition and the ultrastructure are known
to be greatly modified [13]; obviously, these
0 1 2 3 4 5
THIOL REAGENTS rmM1
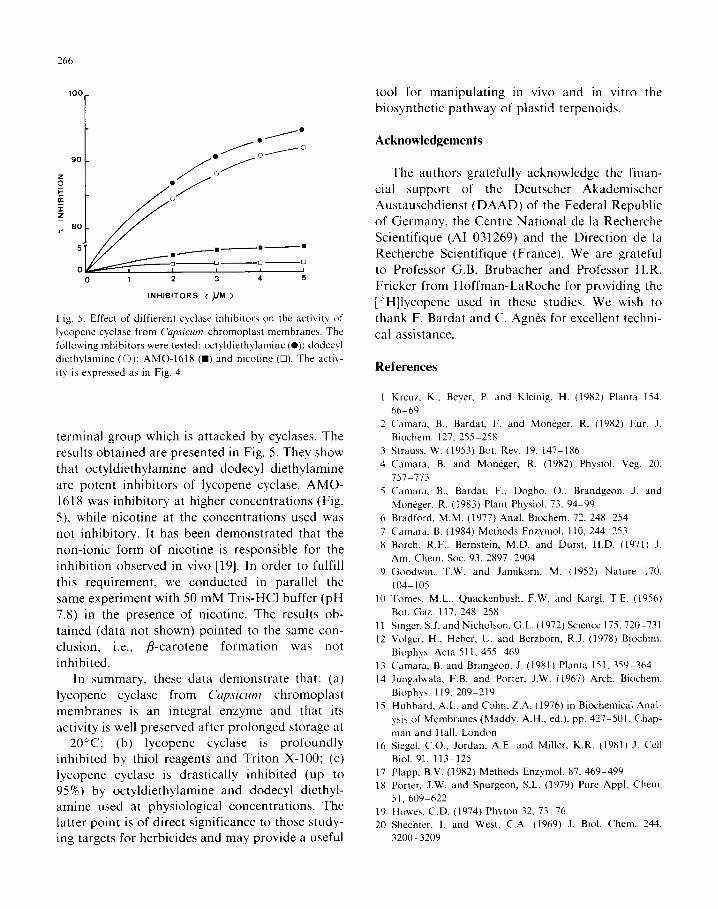
Fig. 4. Effect of thiol reagents on lycopene cyclase activity from
Cqwcum chromoplast membranes. The membrane suspensions
were incubated with mersalyl (o), N-ethylmaleimide (0) and
pCMB (B) for 30 min at 0°C. After centrifugation and washing
with 50 mM Tris-HCl buffer (pH 8), the membrane suspen- sions were incubated with labelled lycopene as described in the
text. The activity is expressed relative to the uninhibited activ-
ity (90000 dpm/mg protein).

changes do not affect the physical state of lyco-
pene cyclase which remains tightly bound to the membranes.
Thiol reagents inhibit lycopene cyclase activity
Considering the importance of thiol groups in
phytoene synthesis [14], we treated the chro- moplast membranes with permeant (mersalyl and
N-ethylmaleimide) and slowly permeant (pCMB)
thiol reagents [15]. As shown in Fig. 4, lycopene
cyclase activity was readily inhibited in all cases.
Freshly prepared membranes gave similar results (unpublished results). Obviously, the different
probes affect a reactive group required for the enzymic activity. Indeed, when the chromoplast membranes were fortified with unlabelled lyco-
pene by the freeze-thaw method of Siegel et al. [16], subsequent addition of mersalyl (the more
active probe) resulted in a less inhibitory effect.
Whether or not lycopene afforded a protection to the active site is debatable. It must be stressed that
substrates can bind, in addition to the active site,
at a low affinity site of the same enzyme [17]. Thus, more detailed investigations are required in
order to determine the role of thiol groups in the lycopene cyclase activity.
Effect of nucleotides
The effects of NADP and FAD were tested. Under our incubation conditions addition of
NADP and FAD were not required for the conver-
sion of lycopene to P-carotene by Cap&urn chro- moplasts membranes. However, the reaction was slightly stimulated by NADP (5%). The same ef-
fect was also noted after solubilization of lycopene
TABLE I
265
cyclase activity (unpublished results). This situa-
tion is different from that described for tomato
fruits [18]. Whatever their effects, the mechanism of nucleotide action in this reaction is not estab-
lished.
Effect of detergents
The chromoplast membranes were treated with increasing concentrations of Tween 80 and Triton
X-100 directly added in the incubation medium.
The cyclase activity was severely inhibited in the
presence of Triton X-100 (Table I), while Tween 80 stimulated this activity. These observations ex-
tend previous data, showing that the incorporation
of isopentenyl pyrophosphate into carotenes be- yond phytoene was severely inhibited (up to 99%) by Triton X-100 [2,4]. Therefore, Triton X-100
inhibits membrane-bound desaturases and cyclases
from Capsicum chromoplasts.
Amphiphilic inhibitors of lycopene cyclase
Due to the lipophilic nature of lycopene, we have synthesized amphiphilic compounds having
the general formula CH,(CH,),,CH,N(CH,), or CH,(CH,),CH,N(C,H,), as possible lycopene
cyclase inhibitors. Two of them (octyldiethylamine and dodecyl diethylamine) were tested using the
incubation procedure described in the experimen-
tal section. Their effects were compared to those of nicotine, which induces in vivo lycopene accumu-
lation in higher plants [19], and of AMO-1618, which inhibits the cyclization of geranylgeranyl
pyrophosphate to copalyl pyrophosphate [20]. In
the latter case, it is worthy of note that geranyl-
geranyl pyrophosphate and lycopene have the same
EFFECT OF DETERGENTS (TWEEN 80 AND TRITON X-100) ON THE ACTIVITY OF LYCOPENE CYCLASE FROM
CAPSICUM CHROMOPLAST MEMBRANES
The incubation conditions were as described in Materials and Methods, except that Tween 80 or Triton X-100 was used to solubilize
the substrate. The different detergents were added to the incubation medium at different detergent-to-protein ratios. After incubation,
the labelled P-carotene was isolated and purified.
Detergents Radioactivity incorporated (dpm)
Ratio
(detergent(mg)/protein (mg)): 4.1: 1 8.2 : 1 12.3 : 1 16.4 : 1 20.5 : 1
Tween 80 90000 90 000 171000 197000 175 000 Triton X-100 36000 32400 18900 15300 14400
266
loo_
$5 l -----o
90 _ v---o
2 . 0
Fig
.-.- n -m
0 1 2 3 4 5
INHIBITORS ()/M 1
5. Effect of diffierent cyclase inhibitors on the activity of
lycopene cyclase from Cqvcum chromoplast membranes. The
following inhibitors were tested: octyldiethylamine (0); dodecyl
diethylamine (0); AMO-1618 (m) and nicotine (0). The acti\-
ity is expressed as in Fig. 4.
terminal group which is attacked by cyclases. The
results obtained are presented in Fig. 5. They show
that octyldiethylamine and dodecyl diethylamine are potent inhibitors of lycopene cyclase. AMO-
1618 was inhibitory at higher concentrations (Fig.
5). while nicotine at the concentrations used was
not inhibitory. It has been demonstrated that the non-ionic form of nicotine is responsible for the
inhibition observed in vivo [19]. In order to fulfill
this requirement, we conducted in parallel the same experiment with 50 mM Tris-HCI buffer (pH
7.8) in the presence of nicotine. The results ob- tained (data not shown) pointed to the same con-
clusion, i.e., p-carotene formation was not
inhibited. In summary, these data demonstrate that: (a)
lycopene cyclase from Cupsicum chromoplast membranes is an integral enzyme and that its
activity is well preserved after prolonged storage at -20°C; (b) lycopene cyclase is profoundly inhibited by thiol reagents and Triton X-100: (c) lycopene cyclase is drastically inhibited (up to 95%) by octyldiethylamine and dodecyl diethyl- amine used at physiological concentrations. The latter point is of direct significance to those study- ing targets for herbicides and may provide a useful
tool for manipulating in vivo and in vitro the
biosynthetic pathway of plastid terpenoids.
Acknowledgements
The authors gratefully acknowledge the finan- cial support of the Deutscher Akademischer
Austauschdienst (DAAD) of the Federal Republic
of Germany, the Centre National de la Recherche Scientifique (AI 031269) and the Direction de la
Recherche Scientifique (France). We are grateful to Professor G.B. Brubacher and Professor H.R.
Fricker from Hoffman-LaRoche for providing the
[“Hllycopene used in these studies. We wish to thank F. Bardat and C. Agnes for excellent techni-
cal assistance.
References
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
Kreut. K.. Beyer. P. and Kleinig, H. (1982) Planta 154.
66-69
Camara. B.. Bardat, F. and MonQer, R. (1982) Eur. J.
Biochem. 127. 255-258
Strauss. W. (1953) Bot. Rev. 19, 147-186
Camara. B. and MonCger, R. (1982) Physiol. Vtg. 20.
757-773
Camara. B.. Bardat. F.. Dogbo. 0.. Brandgeon, J. and
Moneger. R. (1983) Plant Physiol. 73. 94-99
Bradford. M.M. (1977) Anal. Biochem. 72. 248-254
Camara. B. (1984) Methods Enzymol. 110. 244-253
Borch. R.F.. Bernstein, M.D. and Dust. H.D. (1971) J.
Am. Chem. Sot. 93. 2897-2904
Goodwin. T.W. and Jamikorn. M. (1952) Nature 170.
104&105
Tomes. M.L.. Quackenbush. F.W. and Kargl. T.E. (1956)
Rot. Gaz. 117, 248-258
Singer. S.J. and Nicholson. G.L. (1972) Science 175.720-731
Volger. H.. Heber, U. and Berzborn. R.J. (1978) Bjochlm.
BiophSs. Acta 511. 455-469
Camara, B. and Brangeon. J. (1981) Planta 151. 359-364
Jungalwala. F.B. and Porter, J.W. (1967) Arch. Biochem.
Biophy\. 119, 209-219
Hubbard, A.L. and Cohn. Z.A. (1976) in Biochemical Anal-
ysis of Membranes (Maddy, A.H., ed.). pp. 427-501. Chap-
man and Hall. London
Siegel, (‘.O., Jordan, A.E. and Miller, K.R. ( 1981) J. C‘ell
Biol. 91. 113-125
Plapp. B.V. (1982) Methods Enzymol. X7. 469-499
Porter, J.W. and Spurgeon. S.L. (1979) Pure Appl. Chem.
51. 609-622
Howes. C.D. (1974) Phyton 32. 73-76 Shechter, I. and West, C.A. (1969) J. Blol. Chem. 244.
3200-3209





























