Embed Size (px)
Citation preview

Annales d’Endocrinologie 73 (2012) S9-S16
© 2012 Elsevier Masson SAS. Tous droits réservés.
* Auteur correspondant.Adresse e- mail : emmanuelle.kuhn@u- psud.fr (E. Kuhn).
Résumé
Cette revue fait le point sur un certain nombre de données récentes sur la biologie et la physiopathologie du syndrome métabolique (MetS) et l’implication des récepteurs nucléaires qui ont été présentées en juin 2012 lors du dernier congrès de l’Endocrine Society, à Houston. Plusieurs études ont rapporté les effets bénéfi ques de différents récepteurs nucléaires dits « orphelins », notamment SHP (Small Heterodimeric Partner, NR0B2) et LXR (Liver X Receptor, NR1H3 et NR1H2), sur diverses composantes du MetS. En utilisant un modèle d’invalidation de SHP chez la souris, l’équipe de David Moore a démontré que SHP avait des effets « anti- diabétiques », ceci aux dépens d’une stéatose hépatique. Par ailleurs, cette équipe a également montré que le DLPC (dilauroyl phosphatidylcholine), un phospholipide non classique, avait une action anti- diabétique via LRH- 1 (Liver Receptor Homolog- 1, NR5A2), un partenaire moléculaire de SHP. De manière très intéressante, Carolyn Cummins a présenté des données sur les mécanismes impliqués dans les effets métaboliques délétères des glu-cocorticoïdes en se focalisant notamment sur LXR, à l’aide de modèles murins d’invalidation des isoformes α et β de LXR. Ces données nouvelles et originales permettent d’envisager, à plus long terme, l’utilisation de modulateurs sélectifs de ces récepteurs dans la prise en charge des troubles métaboliques chez l’Homme.
© 2012 Elsevier Masson SAS. Tous droits réservés.
Abstract
This review focuses on a number of new data on biology and pathophysiology of the metabolic syndrome (MetS) and the involvement of nuclear receptors that have been presented during the last Endocrine Society meeting, held in Houston in June 2012. Several studies have reported benefi cial effects of various orphan nuclear receptors, including SHP (Small Heterodimeric Partner, NR0B2) and LXR (Liver X Receptor, NR1H3 and NR1H2), on various components of MetS. By using an inactivation model of SHP, David Moore has shown that SHP exerts “antidiabetic” effects but associated with hepatic steatosis development. He also showed that DLPC (dilauroyl phosphatidylcholine), an unconventional phospholipid, exhibited anti- diabetic properties through its binding to LRH- 1 (Liver Receptor Homolog- 1, NR5A2), a molecular partner of SHP. Interestingly, Carolyn Cummins investigated LXR α and β isoforms knock- out mice and provided experimental evidence for the detailed mechanisms involved in the deleterious metabolic effects of glucocorticoids, pointing out to the functional interac-tion between LXRβ, and the glucocorticoid receptor. These new and original studies open new therapeutic opportunities for the management of metabolic disorders in humans by selective modulators of these receptors.
© 2012 Elsevier Masson SAS. All rights reserved.
Mots clés : Récepteur nucléaire ; Syndrome métabolique ; SHP ; LXR ; LRH- 1
Keywords: Nuclear receptor; Metabolic syndrome; SHP; LXR; LRH- 1
Nouveaux mécanismes physiopathologiques du Syndrome Métabolique : implication des récepteurs nucléaires orphelins ?
New pathophysiological mechanisms of metabolic syndrome: implication of orphan nuclear receptors ?
E. Kuhna,b*, B. Fèvec, M. Lombèsa,b,d
aInserm U693, 94276 le Kremlin Bicêtre, FrancebFaculté de Médecine Paris- Sud, Université Paris Sud- 11, 94276 Le Kremlin Bicêtre, France
cService d’Endocrinologie, Hôpital St Antoine, Assistance Publique Hôpitaux de Paris, 75012 Paris, FrancedService d’Endocrinologie et des Maladies de la Reproduction, Hôpital de Bicêtre, Assistance Publique Hôpitaux de Paris, 94275 Le Kremlin Bicêtre,
France
9

S10 E. Kuhn et al. / Annales d’Endocrinologie 73 (2012) S9-S16
1. Introduction
L’augmentation de l’incidence de l’obésité dans nos socié-tés est en particulier due aux changements d’habitudes de vie, notamment nutritionnelles, survenus au cours des dernières décennies. Il est bien établi que l’obésité est associée à une surmortalité et à des facteurs de risques cardiovasculaires tels que l’hypertension artérielle (HTA), l’insulino- résistance, le diabète et la dyslipidémie. Essayer de comprendre les diffé-rents mécanismes physiopathologiques de ces maladies permet de mieux les prendre en charge. Des études récentes, utili-sant des modèles de souris invalidées pour certains récepteurs nucléaires indiquent qu’ils pourraient participer au dévelop-pement de ces troubles métaboliques.
2. Le syndrome métabolique
Le syndrome métabolique (MetS) est défi ni par l’asso-ciation d’une obésité abdominale, d’une insulino- résistance, d’une dyslipidémie, d’une élévation de la pression artérielle et d’une hyperglycémie [1]. D’autres co- morbidités sont égale-ment associées incluant un état pro- thrombotique, un état pro- infl ammatoire et une stéato- hépatite non alcoolique (NASH). Le MetS est associé à un risque accru de survenue d’un dia-bète de type 2 (DT2) et de complications cardiovasculaires. La surnutrition et les habitudes de vie sédentaire de nos socié-tés actuelles exposent à un excès d’adiposité qui concourt à la survenue du MetS.
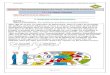
Le MetS résulte de l’association de 3 types d’altéra-tions : (i) l’accumulation ectopique de tissu adipeux (TA) notamment au niveau du muscle et du foie, conduisant à une lipo- toxicité des acides gras libres (AGL) et de leurs dérivés (diacylglycérols, acyl- coAs, céramides) au niveau de ces organes insulino- sensibles ; (ii) l’altération de la fonction endocrine du TA avec modifi cation de la produc-tion des adipokines (diminution de l’adiponectine insulino- sensibilisatrice, augmentation des adipokines impliquées dans l’insulino- résistance) ; et enfi n (iii) une infi ltration macro-phagique avec comme conséquence le développement d’un état pro- infl ammatoire (sécrétion inappropriée de TNF- α (Tumor Necrosis Factor- α) et d’IL- 1 (Interleukin- 1)) condui-sant à une insulino- résistance et à un hyperinsulinisme [2] (Fig. 1). Autrement dit, l’insulino- résistance induit un cercle vicieux (dit de McGarry) dans lequel l’hyper- insulinémie augmente la lipogenèse hépatique et donc l’accumulation ectopique de lipides au niveau du foie, et contribue à exa-cerber l’insulino- résistance elle- même.
Parmi les acteurs intervenant dans le développement du MetS, le tissu adipeux (TA) joue un rôle prépondérant. Il est non seulement constitué d’adipocytes mais également d’autres cellules (telles que macrophages, fi broblastes, cellules endo-théliales, préadipocytes). Au- delà de sa fonction classique de stockage énergétique sous forme de triglycérides (TG) et de lipo- mobilisation (lipolyse), le tissu adipeux est capable de synthétiser et de sécréter de nombreuses hormones, les adi-pokines, qui ont des rôles aussi divers que la régulation de
la prise alimentaire (leptine), la régulation du métabolisme glucidique et lipidique (adiponectine, résistine), la survenue de l’infl ammation (TNF- α, IL- 1), la régulation de la pres-sion artérielle (angiotensine II, angiotensinogène), ou la coa-gulation (PAI- 1 (Plasminogen Activator Inhibitor- 1)) [3]. Il est actuellement bien connu que ces différentes adipokines infl uencent le métabolisme et la fonction de divers organes ou tissus tels que le foie, le muscle, les vaisseaux. Leur taux plasmatique augmente en général avec la masse adipeuse et la taille des adipocytes, sauf pour l’adiponectine circulante qui est diminuée dans l’obésité.
3. Les récepteurs nucléaires
Les récepteurs nucléaires sont des facteurs de transcrip-tion, dont le rôle s’exerce au niveau de divers organes et types cellulaires, impliqués dans de nombreuses fonctions biolo-giques nécessaires au maintien et/ou à la survie de l’espèce (développement, différenciation cellulaire, homéostasie…). Ainsi, au cours de l’évolution, ils sont hautement conser-vés dans leur structure et leur fonction [4]. Ces récepteurs agissent au niveau du noyau, après liaison de leur ligand au niveau cytoplasmique ou nucléaire, où ils régulent l’ex-pression de nombreux gènes en activant ou réprimant leur expression, via leur interaction directe ou indirecte avec la machinerie transcriptionnelle et les séquences régulatrices de gènes- cibles sur l’ADN. Ils peuvent former des homo ou hétérodimères ou être simplement un monomère pour se lier à ADN. Ils recrutent des partenaires moléculaires, les co- régulateurs (co- activateurs ou corépresseurs) qui partici-pent à la stabilisation des complexes transcriptionnels et au remodelage chromatinien [5]. Les hormones stéroïdiennes, thyroïdiennes, les acides rétinoïques et certains dérivés des acides gras, petites molécules lipophiles, sont des ligands de ces récepteurs ; cependant il existe un grand nombre de récepteurs dits « orphelins » car leurs ligands naturels n’ont pas été identifi és à ce jour [4].
Fig. 1. Physiopathologie du syndrome métabolique.Fig. 1. Pathophysiology of the Metabolic Syndrome.TG : Triglycérides, HDLc : cholestérol HDL ; TNF- α : Tumor necrosis Factor- α ; IL- 6 : Interleukine- 6 ; HTA : Hypertension artérielle.
Insuline eTG
HDLc TG
Hyperglycémie
Acides Gras Libres TNF-α, IL-6 Adiponectine e
HTA
Insulino-résistance

S11E. Kuhn et al. / Annales d’Endocrinologie 73 (2012) S9-S16
Tous ces récepteurs ont une même organisation structu-relle, comportant différents domaines ayant des fonctions spécifi ques (Fig. 2). Un domaine de liaison à ADN central (DBD : DNA- Binding Domain ou domaine C), composé de deux doigts de zinc hautement conservés caractérisant les récepteurs nucléaires, permet la fi xation du récepteur à des séquences spécifi ques sur l’ADN, appelées éléments de réponse à l’hormone (HRE : Hormone Responsive Ele-ment). Le domaine N- terminal (NTD : N- Terminal Domain ou domaine A/B) est très variable selon les différents récep-teurs nucléaires mais aussi au sein d’une même famille et contient un domaine de transactivation constitutivement actif (AF- 1) et plusieurs domaines de transactivation indé-pendants. Enfi n le domaine de liaison au ligand (LBD : Ligand Binding Domain ou domaine E), situé en partie C- terminale, permet une liaison spécifi que de l’hormone, ce qui entraîne un changement conformationnel du récep-teur et son activation. Un signal de localisation nucléaire, une interface de dimérisation, une interface d’interaction protéine- protéine ainsi qu’une fonction de transactivation ligand- dépendante (AF- 2) sont également présents à la par-tie C- terminale [6]. Par ailleurs, il existe une région char-nière (Hinge Region ou domaine D) située entre le DBD et le LBD et contenant un signal de localisation nucléaire. Elle possède un site de phosphorylation qui, une fois phos-phorylé, accroît les capacités de transactivation du récep-teur, mais toutes ses propriétés fonctionnelles ne sont pas encore totalement élucidées [6,7].
Les récepteurs nucléaires sont classés en 6 sous- familles, qui sont elles- mêmes subdivisées en sous- groupes (A à K), selon une nomenclature adoptée en 1999 par un consortium international [8]. Elle est basée sur l’évolution phylogéné-tique des 2 domaines les plus conservés : le DBD et le LBD.
4. Rôle des récepteurs nucléaires dans le développement du MetS ?
Différents modèles d’invalidation ou de surexpression de certains récepteurs nucléaires, mais aussi des modèles utili-sant des agents pharmacologiques (agonistes ou antagonistes), ont été développés afi n de déterminer quel était le rôle de ces récepteurs dans la physiopathologie du diabète, de l’obésité ou encore de la stéatose hépatique. Un nombre surprenant de récepteurs nucléaires (comme TRβ) ont des effets protecteurs contre le développement du MetS en inhibant notamment le facteur de transcription SREBP- 1c (Sterol Regulatory Element- Binding Protein- 1c), ce qui permet d’inverser le cercle vicieux de McGarry décrit plus haut (Fig. 3, Tableau 1) [9].
Fig. 3. Cercle vicieux de McGarry.Fig. 3. McGarry Vicious Cycle.L’obésité réduit la sensibilité à l’insuline, ce qui augmente les taux plasma-tiques d’insuline, et favorise la lipogenèse hépatique, puis la stéatose hépatique, qui serait à son tour délétère pour la sensibilité à l’insuline, permettant la mise en place du cercle vicieux (fl èches rouges). Plusieurs récepteurs nucléaires, en interférant avec la voie lipogénique, pourraient rompre ce cercle vicieux (fl èches vertes). SREBP- 1c : Sterol Regulatory Element- Binding Protein- 1c.
Insulinémie
Stéatose
Stéatose
Obésité
Insulinémie
Insulino-sensibilité
Insulino-sensibilité
SREBP-1c
Lipogenèse
SREBP-1cLipogenèse
Récepteurs
Nucléaires
Fig. 2. Structure classique des récepteurs nucléaires.Fig. 2. Nuclear receptor structure.NTD : domaine N- terminal ; DBD : domaine de liaison à ADN ; LBD : domaine de liaison au ligand.
NTD DBD LBD COOHNH2
Tableau 1Divers effets des récepteurs nucléaires sur l’insulino- résistanceTable 1Impact of several nuclear receptors on insulin resistance
Modèle animal Homme
GR - -
LXRα/β +/- ?
FXR +/- +?
PPARα/γ/δ +/+/+ +/+/?
ERα/β + +
CAR + +
HNF- 4α + +
VDR + ?
TRβ + ?
LRH- 1 + ?
NR4A1- 3 + ?
+ : évidence pour des effets anti- diabétiques ; - : évidence pour des effets délétères ; GR : récepteur des glucocorticoïdes ; LXRα/β : Liver X Receptor α/β ; FXR : récepteur X des farnésoïdes ; PPARα/γ/δ : Peroxisome Proliferator- Activated Receptor α/γ/δ ; ERα/β : récepteurs des estrogènes ; CAR : récepteur constitutif des androstanes ; HNF- 4α : Hepatocyte Nuclear Factor 4α ; VDR : récepteur de la vitamine D ; TRβ : récepteur des hormones thyroïdiennes β; LRH- 1 : Liver receptor homolog- 1 ; NR4A1- 3 : Growth factor- inducible immediate early gene nur/77- like receptor; Nur- related protein 1, Neuron- derived orphan receptor 1.

S12 E. Kuhn et al. / Annales d’Endocrinologie 73 (2012) S9-S16
les mécanismes moléculaires sous- jacents à cette apparente divergence, l’équipe de David Moore a réévalué l’impact de l’invalidation de SHP, chez des souris sur ce même fond génétique (C57/Bl6), en caractérisant l’insulino- résistance et le développement de l’obésité induites par un régime riche en graisses [16]. Après 6 mois d’un régime riche en graisses (42 % des calories d’origine lipidique), les souris SHP- /- pré-sentent, comme attendu, une moindre prise de poids (poids à la fi n du régime de : 41 g ± 0,9 pour les souris sauvages et 29 g ± 0,7 pour les souris SHP- /- ; p < 0,002) en rapport avec une diminution du pourcentage de masse grasse mesuré par absorptiométrie (31 % ± 0,7 vs 15 % ± 0,7 ; p < 0,002). Cette résistance à l’obésité ne s’explique pas par une modifi cation de la prise alimentaire ni de l’activité locomotrice.
Le régime riche en graisses est connu pour induire une stéatose hépatique. De façon intéressante, les souris SHP- /- ne présentent pas de stéatose hépatique contrairement aux souris contrôles comme l’atteste la plus faible accumulation lipidique au niveau du foie. Cette diminution du stockage lipidique s’ex-plique par une augmentation de la β- oxydation hépatique des acides gras libres (AGL), comme en témoignent l’élévation du taux plasmatique des corps cétoniques chez les souris SHP- /- et la surexpression de gènes impliqués dans la β- oxydation tels que Cpt1a (Carnitine palmitoyltransferase 1a), HMGCoAS (3- hydroxy- 3- methyl- glutaryl- CoA reductase). A l’inverse, l’expression hépatique de gènes impliqués dans la lipogenèse (tels que la FAS (Fatty Acid Syntase), ACC1 (AcetylCoA Car-boxylase)) ainsi que celle de SREBP- 1c et PPAR γ est for-tement diminuée chez les souris SHP- /- . L’augmentation de la β- oxydation chez les souris SHP- /- a conduit les auteurs à émettre l’hypothèse d’une implication de PPAR- α, facteur de transcription régulateur clé de la β- oxydation. L’utilisation de technique de co- transfection dans un modèle cellulaire hépa-tocytaire et d’immunoprécipitation de la chromatine (CHIP) a permis de démontrer que SHP réprime la fonction transac-tivatrice de PPAR- α et donc de ses gènes cibles.
Enfi n, pour déterminer le statut de l’homéostasie gluci-dique, les auteurs ont soumis les souris à un test de tolérance au glucose (administration d’une solution glucosée par voie intra- péritonéale à 1 g/kg) et à un test de sensibilité à l’insu-line (injection intra- péritonéale d’insuline 1 U/kg). De façon surprenante, les souris SHP- /- , malgré une moindre prise de poids ainsi qu’une glycémie et une insulinémie basales simi-laires aux souris contrôles, présentent une intolérance glu-cidique marquée sans modifi cation de l’insulino- sensibilité globale. En revanche, elles développent une insulino- résistance hépatique, comme en témoigne la diminution de la phos-phorylation de la protéine kinase B (PKB/Akt), en réponse à l’insuline. Cette insulino- résistance hépatique est en partie compensée par une meilleure sensibilité musculaire à l’insu-line permettant le maintien d’un équilibre glycémique adapté (glycémie basale normale). Cette association « intolérance au glucose- sensibilité à l’insuline similaire » pourrait être expli-quée par un défaut d’insulino- sécrétion. Il a été montré, sur îlots pancréatiques isolés, qu’à la fois le contenu insulinique et la sécrétion d’insuline induite par le glucose sont forte-
Lors du congrès annuel de l’Endocrine Society, qui s’est déroulé du 23 au 26 juin 2012 à Houston, Texas, plusieurs équipes ont ainsi présenté les résultats d’études récentes qui démontrent l’implication potentielle de certains récepteurs nucléaires dans le développement d’une dysglycémie voire d’un diabète et de la stéatose hépatique, deux composantes importantes du syndrome métabolique. Ainsi David Moore (Baylor College of Medecine, Houston, TX) a démontré que SHP (Small Heterodimeric Partner) et LRH- 1 (Liver Recep-tor Homolog- 1) avaient des effets anti- diabétiques en utili-sant des souris invalidées pour ces 2 récepteurs.
5. Les deux visages de SHP (Small Heterodimeric Partner)
SHP, récepteur nucléaire orphelin, est un membre aty-pique de cette super- famille (NR0B2). En effet, il ne possède pas le domaine classique de liaison à l’ADN mais contient probablement un LBD. Ce facteur de transcription exerce des fonctions biologiques majeures en particulier sur le métabo-lisme du cholestérol, des acides gras et des acides biliaires mais aussi sur la balance énergétique. SHP interagit avec plus de la moitié des récepteurs nucléaires et d’autres fac-teurs de transcription en formant un hétéro- dimère, son rôle principal étant d’inhiber la voie de transduction du signal. La recherche de ligands synthétiques agissant sur SHP consti-tue donc une voie de recherche importante dans la prise en charge des anomalies métaboliques.
Il est maintenant établi que SHP interagit avec HNF- 4α (Hepatocyte Nuclear Factor 4α, NR2A1), gène dont les muta-tions sont responsables du MODY 1 (Maturity- Onset Diabetes of the Young de type 1) [10,11]. Par ailleurs, des mutations hétérozygotes de SHP ont été retrouvées chez des sujets obèses [12], les mutations de SHP co- ségrégent avec l’obé-sité mais pas avec le diabète [13]. Enfi n, il a été montré que de nouveaux variants génétiques associés à une obésité, entraî-neraient une perte de l’inhibition de HNF- 4α [14].
C’est dans ce contexte que l’équipe de David Moore a déve-loppé des souris invalidées pour SHP (SHP- /- ) afi n d’évaluer l’implication de ce récepteur nucléaire dans le développement des composantes du MetS. Une première étude avec des sou-ris issues d’un fond génétique mixte montrait que les sou-ris SHP- /- étaient résistantes à l’obésité induite par le régime, avaient une amélioration de l’homéostasie énergétique ainsi qu’un moindre développement de la stéatose hépatique. Ce phénotype était associé à une surexpression de gènes impli-qués dans la dépense énergétique (comme PGC- 1α (PPAR γ Coactivator- 1α), UCP1 (Uncoupling Protein 1), DIO2 (Deio-dinase 2), LHS (Lipase Hormono- sensible) au niveau du tissu adipeux brun (BAT : Brown Adipose Tissue) [15]. Cepen-dant, de manière surprenante, les souris SHP- /- , établies sur un autre fond génétique (C57/Bl6), présentent, elles aussi, une résistance à l’obésité induite par le régime (DIO, Diet Induced- Obesity) mais sans modifi cation de l’expression des gènes PGC- 1α, UCP1, DIO2, ou LHS. Afi n de préciser

S13E. Kuhn et al. / Annales d’Endocrinologie 73 (2012) S9-S16
de la leptine), et donc génétiquement obèses et diabétiques, ou chez des souris insulino- résistantes suite à un régime riche en graisses, permet également de diminuer la stéatose hépatique et d’améliorer la tolérance au glucose. Ces 2 effets protecteurs (contre le diabète et la stéatose hépatique) sont abolis chez des souris invalidées pour LRH- 1 spécifi quement au niveau du foie. En conclusion, le DLPC en activant LRH- 1 semble améliorer l’homéostasie glucidique et des acides gras libres, et pourrait ainsi être un agent pharmacologique intéressant pour la prise en charge des troubles métaboliques chez l’Homme.
7. Vers une meilleure compréhension des effets métaboliques des glucocorticoïdes.
Dans le but de mieux comprendre les mécanismes phy-siopathologiques du MetS, Carolyn Cummins (Université de Toronto, Canada) a, quant à elle, présenté des résultats pro-metteurs sur les mécanismes aboutissant aux effets métabo-liques délétères des glucocorticoïdes (GC).
En effet, les GC sont largement utilisés en pratique courante pour leurs effets anti- infl ammatoires et immuno- suppresseurs. Malheureusement leur utilisation chronique et à fortes doses s’accompagne de nombreux effets indésirables notamment métaboliques tels qu’une insulino- résistance, une hypergly-cémie, voire un diabète, une stéatose hépatique, une obésité facio- tronculaire, une amyotrophie. Les mécanismes condui-sant à ces effets néfastes sont multifactoriels : (i) augmentation de la néoglucogenèse hépatique ; (ii) diminution du captage périphérique de glucose par le tissu adipeux et le muscle ; (iii) défaut d’apports de substrats énergétiques d’origine adipocy-
ment diminués chez les souris SHP- /- , confi rmant ainsi la dys-fonction β- pancréatique après inactivation génique de SHP.
L’ensemble de ces résultats démontre que les souris SHP- /- sont protégées du développement de l’obésité et de la stéatose hépatique mais au prix d’une insulino- résistance hépatique et d’une dysfonction β pancréatique conduisant au diabète (Fig. 4). L’utilisation de modulateurs sélectifs de SHP selon les tissus pourrait permettre de mieux prendre en charge les troubles métaboliques que sont l’obésité et la stéatose hépa-tique sans induire d’effets néfastes comme le diabète.
6. LRH- 1 et le DLPC : nouvel agent pharmacologique anti- diabétique ?
Le récepteur orphelin LRH- 1 (NR5A2) régule la biosyn-thèse des acides biliaires. Les phospholipides seraient des ligands potentiels pour ce récepteur mais leur rôle fonctionnel est indéterminé. La même équipe de David Moore a montré que l’activation de LRH- 1 par le DLPC (dilauroyl phos-phatidylcholine), un phospholipide non classique, a un effet anti- diabétique et pourrait être ainsi un agent thérapeutique potentiel dans la prise en charge des troubles métaboliques chez l’Homme. En effet, l’administration de DLPC pendant 3 semaines chez des souris C57Bl/6 permet non seulement d’induire l’expression d’enzymes intervenant dans la bio-synthèse des acides biliaires (tels que CYP7A1, CYP8B1), et ainsi augmenter le taux des acides biliaires, mais aussi de diminuer le contenu intra- hépatique en triglycérides, le taux plasmatique des AGL et enfi n la glycémie. Le traitement par DLPC chez des souris db/db (souris défi cientes en récepteur
SHP-/-
Tissu adipeux brun
PancréasFoie
Muscle
Dysfonction β pancréatique
Surexpression de PGC1-αInsulino-sensibilité
Insulino-résistance
Stéatose Hépatique
Fig. 4. Phénotype Métabolique des souris SHP- /- .Fig. 4. Metabolic Phenotype of SHP- /- mice.PGC- 1α : PPARγ Coactivator 1- alpha.

S14 E. Kuhn et al. / Annales d’Endocrinologie 73 (2012) S9-S16
téronémie, hyperplasie bilatérale des surrénales sans obésité ni insulino- résistance) [23]. Afi n de préciser les interactions fonc-tionnelles entre ces deux voies de signalisation LXR et GR, l’équipe de Carolyn Cummins [24] a émis l’hypothèse que le recrutement des LXRs était nécessaire à l’action métabolique des glucocorticoïdes. Des souris sauvages et invalidées pour les deux gènes LXR (LXRα/β- /- ) ont été traitées par la dexamé-thasone (DXM 2,5 mg/kg) pendant 5 à 7 jours. Il a été montré que les souris double- mutantes LXRα/β- /- avaient une meilleure tolérance au glucose lors des tests dynamiques et étaient pro-tégées de l’insulino- résistance induite par les GC. En accord avec ces résultats, le niveau de phosphorylation de PKB/Akt hépatique est nettement supérieur dans les souris LXRα/β- /- comparé à celui des souris sauvages. Ces résultats ne sont pas en rapport avec une modifi cation de l’expression ou de l’acti-vation de GR. Par contre, l’expression des gènes de la néoglu-cogenèse hépatique (PEPCK, PGC1- α et G6P) est fortement réduite chez les souris double- mutantes. Ainsi, les récepteurs LXRs semblent jouer un rôle modulateur majeur dans l’ex-pression de certains gènes cibles du GR.
Afi n de déterminer la contribution de chacun des sous- types de LXRs, LXRα ou LXRβ, des expériences similaires ont été réalisées chez des souris invalidées sélectivement pour
taire ou musculaire pour la production hépatique de glucose ; enfi n (iv) inhibition de la sécrétion d’insuline par les cellules β pancréatiques. À long terme se développent une insulino- résistance et un MetS. La fi xation des GC à leur récepteur, le récepteur des glucocorticoïdes (GR), entraine l’activation de la transcription de gènes impliqués dans la néoglucoge-nèse (phosphoenolpyruvate carboxykinase (PEPCK), glu-cose 6 phosphatase (G6P)) et la répression de gènes impliqués dans l’infl ammation (TNF- α, IL- 1, IL- 6).
Mieux comprendre les partenaires moléculaires à l’origine de ces effets néfastes pourrait permettre de les prévenir lors de l’utilisation chronique des glucocorticoïdes. Les récepteurs nucléaires LXRα et LXRβ (Liver X Receptor, respectivement NR1H3 et NR1H2) sont activés par les oxysterols (métabo-lites du cholestérol), et exercent des effets anti- infl ammatoires, comme le GR, en régulant l’expression d’un certain nombre de gènes communs avec le GR [17]. En revanche, l’activation de LXRα permet d’améliorer le contrôle glycémique de sou-ris diabétiques [18- 20] en inhibant l’expression hépatique de PEPCK [19,20] et G6P [20,21] et en augmentant celle de GLUT4 au niveau musculaire [20,22]. De façon intéressante, les souris invalidées à la fois pour LXRα et LXRβ (LXRα/β- /- ) présentent une résistance aux glucocorticoïdes (hypercorticos-
LXRβ
GR
GC
GR
TranscriptionGènes cibles
(TNF-α)
GRE
CoR RNA Pol II
TTTTTTTTTraGèGR
HyperglycémieStéatose Hépatique
Inflammation
TranscriptionGènes cibles
(PEPCK)
GRE
CoR RNA Pol II GR
WT
LXRβ
GR
GC
GR
HyperglycémieStéatose Hépatique
TranscriptionGènes cibles
(PEPCK)
GRE
CoR RNA Pol II
TTTTTTTTTraGèGR
Lxrβ-/-
LXRβ
Fig. 5. Activation sélective de la voie de signalisation des glucocorticoïdes dans le foie.Fig. 5. Selective activation of hepatic GC- response pathway.WT : souris sauvage, LXRβ- /- : souris invalidée pour LXRβ ; GR : récepteur des glucocorticoïdes, GC : glucocorticoïdes ; GRE : éléments de réponse aux glucocorticoïdes ; LXRβ : Liver X Receptor β, CoR : co- régulateurs ; RNA pol II : ARN polymérase II, TNF- α : Tumor Necrosis Factor α ; PEPCK : Phos-phoenolpyruvate carboxykinase.

S15E. Kuhn et al. / Annales d’Endocrinologie 73 (2012) S9-S16
Références[1] Cornier MA, Dabelea D, Hernandez TL, Lindstrom RC, Steig
AJ, Stob NR, et al. The metabolic syndrome. Endocr Rev 2008;29:777- 822.
[2] Despres JP and Lemieux I. Abdominal obesity and metabolic syndrome. Nature 2006;444:881- 7.
[3] Feve B and Bastard JP. The role of interleukins in insulin resistance and type 2 diabetes mellitus. Nat Rev Endocrinol 2009;5:305- 11.
[4] Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schutz G, Umesono K, et al. The nuclear receptor superfamily: the second decade. Cell 1995;83:835- 9.
[5] Lonard DM and O’Malley BW. The expanding cosmos of nuclear receptor coactivators. Cell 2006;125:411- 4.
[6] Bain DL, Heneghan AF, Connaghan- Jones KD, Miura MT. Nuclear receptor structure: implications for function. Annu Rev Physiol 2007;69:201- 20.
[7] Lee YK, Choi YH, Chua S, Park YJ, Moore DD. Phosphoryla-tion of the hinge domain of the nuclear hormone receptor LRH- 1 stimulates transactivation. J Biol Chem 2006;281:7850- 5.
[8] Committee NRN. A unifi ed nomenclature system for the nuclear receptor superfamily. Cell 1999;97:161- 3.
[9] Moore DD. Nuclear receptors reverse McGarry’s vicious cycle to insulin resistance. Cell Metab 2012;15:615- 22.
[10] Lee YK, Dell H, Dowhan DH, Hadzopoulou- Cladaras M, Moore DD. The orphan nuclear receptor SHP inhibits hepa-tocyte nuclear factor 4 and retinoid X receptor transactivation: two mechanisms for repression. Mol Cell Biol 2000;20:187- 95.
[11] Mitchell SM, Weedon MN, Owen KR, Shields B, Wilkins- Wall B, Walker M, et al. Genetic variation in the small hetero-dimer partner gene and young- onset type 2 diabetes, obesity, and birth weight in U.K. subjects. Diabetes 2003;52:1276- 9.
[12] Nishigori H, Tomura H, Tonooka N, Kanamori M, Yamada S, Sho K, et al. Mutations in the small heterodimer partner gene are associated with mild obesity in Japanese subjects. Proc Natl Acad Sci U S A 2001;98:575- 80.
[13] Hung ML, Farooqi IS, Ong K, Luan J, Keogh JM, Pembrey M, et al. Contribution of variants in the small heterodimer partner gene to birthweight, adiposity, and insulin levels: mutational analysis and association studies in multiple populations. Dia-betes 2003;52:1288- 91.
[14] Echwald SM, Andersen KL, Sorensen TI, Larsen LH, Andersen T, Tonooka N, et al. Mutation analysis of NR0B2 among 1545 Danish men identifi es a novel c.278G> A (p.G93D) variant with reduced functional activity. Hum Mutat 2004;24:381- 7.
[15] Wang L, Liu J, Saha P, Huang J, Chan L, Spiegelman B, et al. The orphan nuclear receptor SHP regulates PGC- 1alpha expression and energy production in brown adipocytes. Cell Metab 2005;2:227- 38.
[16] Park YJ, Kim SC, Kim J, Anakk S, Lee JM, Tseng HT, et al. Dissociation of diabetes and obesity in mice lacking orphan nuclear receptor small heterodimer partner. J Lipid Res 2011;52:2234- 44.
[17] Ogawa S, Lozach J, Benner C, Pascual G, Tangirala RK, Westin S, et al. Molecular determinants of crosstalk between nuclear receptors and toll- like receptors. Cell 2005;122:707- 21.
[18] Zelcer N, Tontonoz P. Liver X receptors as integrators of metabo-lic and infl ammatory signaling. J Clin Invest 2006;116:607- 14.
[19] Cao G, Liang Y, Broderick CL, Oldham BA, Beyer TP, Schmidt RJ, et al. Antidiabetic action of a liver x receptor agonist media-ted by inhibition of hepatic gluconeogenesis. J Biol Chem 2003;278:1131- 6.
[20] Laffi tte BA, Chao LC, Li J, Walczak R, Hummasti S, Joseph SB, et al. Activation of liver X receptor improves glucose
LXRα (LXRα- /- ) ou pour LXRβ (LXRβ- /- ). Seules les souris LXRβ- /- présentent un phénotype métabolique similaire aux souris double- mutantes LXRα/β- /- (absence d’hyperglycémie et d’hyperinsulinémie induite par la DXM). Les souris LXRβ- /- n’ont pas d’accumulation hépatique de triglycérides et sont ainsi résistantes au développement de la stéatose hépatique induite par la corticothérapie. Par des techniques sophistiquées de ChIP (Chromatin Immnunoprecipitation) sur des cultures primaires d’hépatocytes issues de souris LXRβ- /- , combinées à l’étude de l’expression de PEPCK induite par les glucocor-ticoïdes, il a pu être formellement démontré que, dans le foie, le recrutement de LXRβ était indispensable à la liaison du GR sur le promoteur de PEPCK. Il est important de rappeler que chez ces souris LXRβ- /- , l’action anti- infl ammatoire des glu-cocorticoïdes est préservée, confi rmant la spécifi cité cellulaire de l’interférence fonctionnelle entre GR et LXRβ (Fig. 5). De plus, l’utilisation d’un antagoniste des LXRs, le composé 17, dans une culture primaire d’hépatocytes mime les effets de la perte de LXRβ sur le métabolisme hépatique.
En résumé, la caractérisation phénotypique de plusieurs modèles de souris invalidées pour LXR a permis de démon-trer le rôle clé de LXRβ, mais d’exclure celui de LXRα, dans la pathogenèse des conséquences métaboliques délétères des GC (hyperglycémie, stéatose hépatique).
Ces résultats originaux ouvrent à l’évidence sur des pers-pectives intéressantes mais posent aussi de nouvelles ques-tions. D’autres effets indésirables de la corticothérapie tels que l’ostéoporose ou l’amyotrophie, nécessitent- ils le recru-tement de LXRβ ? De nouvelles stratégies thérapeutiques utilisant des agonistes glucocorticoïdes ayant une action anti- infl ammatoire mais sans effet métabolique néfaste ou à l’in-verse des antagonistes spécifi ques de LXRβ pour prévenir les effets délétères des glucocorticoïdes pourraient voir le jour.
8. Conclusion
Les données récentes publiées et rapportées lors du congrès 2012 de l’Endocrine Society à Houston ont rappelé les liens étroits qui existent entre récepteurs nucléaires et régulation du métabolisme énergétique. Ces travaux ont aussi permis de préciser l’importance des interactions fonctionnelles entre les différents membres des récepteurs hormonaux classiques et les récepteurs orphelins. Bien sûr d’autres récepteurs nucléaires semblent également intervenir dans l’homéostasie énergétique tel que le récepteur des minéralocorticoïdes [25]. Ces études ouvrent sur des perspectives de compréhension physiopatho-logiques et à plus long terme thérapeutiques.
Déclarations d’intérêts
E. Kuhn, M. Lombès : aucun.B. Fève : conférences : invitations en qualité d’interve-
nant (Sanofi , Novo Nordisk, MSD) et en qualité d’auditeur (frais de déplacement et d’hébergement pris en charge par une entreprise) (Sanofi , Novo Nordisk, Lilly, MSD).

S16 E. Kuhn et al. / Annales d’Endocrinologie 73 (2012) S9-S16
[23] Cummins CL, Volle DH, Zhang Y, McDonald JG, Sion B, Lefrancois- Martinez AM, et al. Liver X receptors regulate adrenal cholesterol balance. J Clin Invest 2006;116:1902- 12.
[24] Patel R, Patel M, Tsai R, Lin V, Bookout AL, Zhang Y, et al. LXRbeta is required for glucocorticoid- induced hyperglycemia and hepatosteatosis in mice. J Clin Invest 2011;121:431- 41.
[25] Kuhn E, Le Menuet D, Keo V, Muscat A, Viengchareun S, Meduri G, et al. Mineralocorticoid Receptor overexpressing mice are resistant to high fat diet- induced obesity: implication for the pathophysiology of metabolic syndrome. 94th Annual Meeting of Endocrine Society, Houston, TX, Jun 23- 26 2012;Poster SUN- 91.
tolerance through coordinate regulation of glucose metabo-lism in liver and adipose tissue. Proc Natl Acad Sci U S A 2003;100:5419- 24.
[21] Grefhorst A, van Dijk TH, Hammer A, van der Sluijs FH, Havinga R, Havekes LM, et al. Differential effects of phar-macological liver X receptor activation on hepatic and peri-pheral insulin sensitivity in lean and ob/ob mice. Am J Phy-siol Endocrinol Metab 2005;289:E829- 38.
[22] Dalen KT, Ulven SM, Bamberg K, Gustafsson JA, Nebb HI. Expression of the insulin- responsive glucose transporter GLUT4 in adipocytes is dependent on liver X receptor alpha. J Biol Chem 2003;278:48283- 91.