Embed Size (px)
Citation preview

PAI,I ONTOLOGIE D E S VERTI BRI S INFI RIEURS - R E N O U V E A U
ET DI VELOPPEMENTS
CI~CILE POPLIN, ALAIN BLIECK, RICHARD CLOUTIER, CLAIRE DERYCKE, DANIEL GOUJET, PHILIPPE JANVIER, HERV]~ LELII~VRE, MONETTE VERAN • SYLVIE WENZ
POPLIN C., BLIECK A., CLOUTIER R., DERYCKE C., GOUJET D., JANVIER P., LELII~VRE H., VI~RAN M. & WENZ S. 1997. Pal6ontologie des vert6br6s inf6rieurs - Renouveau et d6veloppements. [Lower vertebrate palaeon- tology - Renewal and developments]. GEOBIOS, M.S. n ° 20: 437-446.
RI~SUMI~ - Pendant les 20 derni6res ann6es, grace ~ la mise en oeuvre des m6thodes cladistiques, la connaissance de la phylog6nie de nombreux groupes fossiles d'agnathes, de poissons et d'amphibiens primitifs a 6t6 pr6cis6e. Leur valeur de marqueurs biostratigraphiques, en particulier les microrestes, a 6t6 explor6e, surtout au Pal6ozoYque. I1 en est de m~me pour la reconstitution des pal6oenvironnements et le test des mod61es pal6og6ographiques propos6s par les g6ophysiciens. Les travaux interdisciplinaires et les coop6rations internationales ainsi que l 'ouverture de nombreux nouveaux chantiers de fouille ont 6t6 aussi d6terminants dans ce renouveau.
MOTS-CLES: VERTI~BRI~S INFERIEURS, PHYLOGI~NIE, BIOSTRATIGRAPHIE, PALI~OENVIRONNEMENT, PALt~OGI~OGRAPHIE.
ABSTRACT - During the last 20 years, thanks to the use of the cladistic methods, the knowledge of the phylogeny of many fossil groups of agnathans, fishes and primitive amphibians was specified. They have been used as bios- tratigraphic markers, especially in regard with Palaeozoic microremains. They also have been used for palaeoenvi- ronmental reconstructions and testing palaeogeographic models hypothetized by geophysicians. Interdisciplinary studies, international cooperations as well as exploration of many new fossiliferous localities also participated to this renewal.
KEYWORDS: LOWER VERTEBRATES, PHYLOGENY, BIOSTRATIGRAPHY, PALAEOENVIRONMENT, PALAEOGEO- GRAPHY.
INTRODUCTION: LES OUTILS DU PROGRES DES CONNAISSANCES
L'expression "vert6br6s inf6rieurs" a 6t6 utilis6e pa r des biologistes et des pal6ontologues pour grouper tout ce qui n ' appa r t i en t pas aux roam- miferes ou aux oiseaux. Dans ce travail , nous n'in- cluons sous ce vocable que les "agnathes" et les "poissons", tout en 6 tendant not re domaine de recherche aux tout p remiers amphibiens. Jalonn6e pa r une s6rie de congr6s in t e rna t ionaux consacr6s aux vert6br6s pal6ozoiques (Londres 1972; Paris 1973; Tal l inn 1976, 1989; P6kin 1987; Miguasha 1991; G~t t ingen 1993; Par is 1995) et aux poissons m6sozoiques (Eichst~t t 1993), la connaissance des vert6br6s inf6r ieurs a, au cours des vingt derni~res ann6es, cons id6rablement 6volu6. Ces r6unions ont 6t6 des 6tapes dans les progr6s de l '6tude de ces groupes qui sont au coeur des probl6mes impor- ran ts pos6s pa r les p remiers stades de l '6volution des vert6br6s. Elles ont permis ~ des m6thodolo-
gies jusqu 'a lors marginales de se d6velopper et d ' introduire, dans un domaine oh on ne l ' a t tenda i t pas, un renouveau 6pist6mologique g6n6ra teur de probl6matiques et de th6mat iques nouvelles. Au congr~s de 1973, les id6es qui domina ien t 6taient influenc6es par l 'autori t6 des mai t res d'alors. Ce congr6s fut marqu6 pa r l ' in t roduct ion en France de la Sys t6mat ique Phylog6n6tique de Hennig, ou cla- distique (Bonde in L e h m a n 1975; Miles in L e h m a n 1975). La cladistique avait p6n6tr~ d 'abord dans l 'univers des pal6oichthyologues des pays anglo- phones, puis, vers la fin des ann6es 70, en France. Son adoption ne se fit pas sans douleur (voir les pol6miques de N a tu r e en 1978-79) mais plus rapi- dement dans le domaine des vert6br~s inf6rieurs qu'ailleurs. Cette m6thodologie, qui impose une vision nouvelle de la ressemblance et doric des rela- tions de parent6, a contribu6 pour une pa r t impor- t an te h un regain d' int6r6t pour la pal6ontologie. Son efficacit6 s 'est accrue dans les ann6es 80 pa r la raise au point de logiciels d 'analyse cladistique.

438
La g~n6ralisation de la pr6paration par attaque l'acide et l'utilisation des r~sidus obtenus, dans la recherche des conodontes en particulier, conduisi- rent fi d'importants d6veloppements dans la d6couverte et l'~tude des microrestes et des micro- structures. Ces techniques b6n6fici~rent des inno- vations apport~es ~ la pr6paration des colles et des mati~res plastiques autorisant une consolida- tion durable des sp6cimens et des proc6d6s de transfert sur support permanent.
Les progr~s de la pal6ohistologie, qui utilise les microscopes min~ralogiques, sont surtout dus, pour la distinction des tissus, au microscope Nomarski fi interference diff6rentielle, au micro- scope fi cathodoluminescence (Derycke 1990) et au microscope ~lectronique fi balayage. De plus, l'interpr6tation des r~seaux observes ~ la surface des tissus hypermin6ralis~s, que ce soit sur des restes de chondrichthyens, d'actinopt6rygiens ou d'acanthodiens, permet de pr6ciser les m6ca- nismes de biomin6ralisation.
Les progr~s dans l'interpr6tation des conditions de fossilisation et la dgcouverte de fossiles de conservation exceptionnelle ont ~galement mar- qu~ cette p6riode. Ceci allait de pair avec l'ouver- ture de nombreux chantiers nouveaux au Moyen et Extreme Orient, en Australie, dans les zones arctiques et en Am~rique du Sud, impliquant des cooperations internationales (PICG 328 et 406 par exemple). Le d6veloppement de celles-ci avec des coll~gues ayant une exp6rience diff6rente et des terrains d'~tudes jusqu'alors ignores des cher- cheurs occidentaux (ouverture vers l'Est notam- ment) fur un 616ment important du regain d'int~r~t des vert6br6s inf6rieurs.
De fait, les avanc~es m6thodologiques et tech- niques jointes aux collaborations internationales ont donn~ aux vertebras inf6rieurs, au-delfi de leur dimension pal~obiologique, le r61e d'indica- teurs pr~cieux, t6moins des g6ographies pass6es permettant de tester les sch6mas pal~og6ogra- phiques, notamment pour le Pal6ozo~'que: leur localisation privil~gi~e fi la bordure des conti- nents est, ~ cet ~gard, d~terminante. Ainsi ces groupes ont-ils aussi acquis une signification pal~oenvironnementale, en particulier avec le d~veloppement de la recherche de vert6br6s dans les facies matins marginaux.
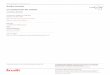
t~VOLUTION ET P H Y L O G E N I E (Fig. 1)
LES "AGNATHES"
Les vert6br~s furent longtemps consid6r6s comme comprenant deux sous-phyla en g~n~ral pr6sum~s monophyl~tiques: agnathes (sans m~choires) et
gnathostomes (avec m~choires). Depuis 20 ans, ce schema a 6volu~ pour les agnathes par l'am61iora- tion des connaissances de l'anatomie et de la diversit6 des fossiles (dont les Galeaspida, Wang N.Z., in Chang et al. 1991) et la d6couverte de nou- veaux groupes (Arandaspida ordoviciens, Gagnier 1993; Pituriaspida d~voniens, Young in Chang et al. 1991). I1 a d'abord ~t6 d6montr~ que les actuels cyclostomes sont paraphyl~tiques, les myxines 6rant le groupe-fr~re de tous les autres vert6br~s dont les lamproies. Ceci ~tant admis, les d~bats portent depuis sur les places respectives des formes fossiles par rapport aux groupes actuels. Selon une premiere analyse, les c6phalaspi- domorphes, clade rassemblant lamproies, anas- pides et ost~ostrac~s, auraient ~t~ les plus proches parents des gnathostomes. Cependant, les ost~os- trac~s partageant une majorit6 de synapomor- phies avec les gnathostomes, les c~phalaspi- domorphes apparaissent d~sormais comme para- ou polyphyl6tiques (Forey & Janvier 1993). Enfin l'analyse phylog~n6tique de Gagnier (1993), sui- vie de celle de Janvier (1996), sugg~re que tousles agnathes siluro-d6voniens ~ squelette min6ralis~ ("ostracodermes") sont plus 6troitement appa- rent,s aux gnathostomes qu'aux lamproies et, a fortiori, qu'aux myxines.
Les principaux d~bats actuels portent sur la monophylie ou la paraphylie des th61odontes (Turner 1991 in Chang et al. 1991; Janvier 1996), la position phylog~n6tique des "anaspides aty- piques" (Jamoytius, Euphanerops), les relations entre les pt~raspidomorphes (arandaspides, astraspides, eriptychiides et heterostraci) ordovi- ciens ~ d~voniens et la position syst~matique des conodontes dont l'anatomie sugg~re des affinit6s avec les Vertebras (Aldridge et al. 1993).
LES PREMIERS GNATHOSTOMES
L'apparition de la m~choire, issue des premiers arcs visc6raux (mandibulaire et hyo~'dien) est la principale synapomorphie soutenant la monophy- lie des gnathostomes. Le d6veloppement des recherches sur les acanthodiens et les placo- dermes a permis de pr~ciser la monophylie et la position respective de ces deux groupes par rap- port aux chondrichthyens et aux ost~ichthyens. Les acanthodiens qui furent longtemps associ6s aux chondrichthyens, sont aujourd'hui consid~r~s comme le groupe-fr~re des ost~ichthyens (actino- pt~rygiens et sarcopt~rygiens). L'anatomie cr~- nienne des premiers acanthodiens demeure 6nig- matique mais l'6tude de leurs 6cailles et la d~cou- verte d'individus en connexion (Valiukevicius in Mark-Kurik 1992) ~clairent certains aspects de leurs liens phylog6n~tiques et de la mise en place de la m~choire: il corroborent ainsi la th6orie d'Orvig (1973) selon laquelle les dents et les

439
T K J Tr P I Iyperotret i (Myxine~ ~,
| HyDerc~rti i (Lamproies) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Theladont i
- " L ' F ~ ....... , , A r a n d a s p i d a . . J II ~ Astrosp]da . _
H I I - - ~ Er iptychi lda . . . . . L - - Heterostraci . . . . . . . . . . . . . . . . . . .
q Anasp ida . . . . . . . . | I , Ga leasp id . . . . . . . . . . . . '
J l I . . . . . . . . . . O ~ e o ~ / r a d . . . . . . . . . . - - " 1 J ~ Pttur~aspldo . . . . . . . . . . . .
[ AcanthctJnoraci . . . . L . ~ J F - " - - - - - - - Rh . . . . id . . . . . . . . . . . . . . . . . . .
~ . 4 Ant la raha . . . . . . . . L ~ r - " Peta l i ch ihy ida
[ I - - ' - l _ _ P l yc todon t i da L ~ Ac t ino lop ido ido i
t t - - Ph lyc taen i ida . . . . . . . . . . ~ rachy ihorac i . . . .
. _ ~ Neoselochi l (~equins, Raies)
Hybodont i f . . . . . s [ _ : i I C t e n a c a n l h i c a e i i " - - . . . . . . . •
} In lopte~/g la " " "~ ~ 1 --~ t-.I-- He lodon f idae ........ ~.
] Chond renche t y i cae _ _~ . . . . M e n a s p o i d e a I .... ' COC hlJodontid Oe . . . . ~ I f E ch inoch i rnaer [doe . . . . . . . ~ . . . . . . . . . . Chi rnaero ide i (Chim6res] M o n g o l e p i d a . . . . . . . . . . . . .
~ ClJmat j ida . D ip lacan th ido . I schnacan th ida . . . . . . . . . A c a n t h o d i d a . . . . . . . . . . . . ~ . . . . . A n d r e o l a p ~ s . i "
D i ~ l ; p i n a . . . . . . . . . . . . . i . . . . . .
Che i ro lep id idae _ ~ ._ . Cladi~t ia (Pol,~pl&res) i . . . . . . . . i=~ Ac t inop ted basaux OU
I Pal@oniscofdes . . . . . . . . . . . . . . Chondroste i (Estcrgeons) i Sour ich thy idae . . . . Neoptery~ i i basQu× . . . . . . . . . . . . . . . . . . . . . . . . . . . - i G ing lymod i (L6p i ;os t6os) i I I Sorniono~idae . . . . . . . . . . . . . . . . . . . . . i l i l i i m Pycnodon f idae . . . . i i Para~emior;oJidQe Ca tu r idae .. A m i i d o e (Amias) . • I Teleostei ,¢l@l@o#¢er,~) ,., Onychodon t i f o rmes ~ Actinistia (Coelacanthes) • Porolepiform~s ................. P o w i c h t h y s . . . . . . . . . . . . . .
' r ' o u n g o l e p i ~ . _
O i a b o t e p i ~ . . . . . . .
Dipnoi (Dlpneustes) . • RhizodontiFo~mes . . . . . . . . . . . . . . Osteolepi forrr es Dpist os~egalia . . . . . . . . . . . . . O b r u c h e v i c h t h y s
g l g l n e r p e t o n . . . . . . . . . . . M e t a x y g n a t h u s . . . .
V e t t ~ t e ~ q a .
A c a n t h c s t e g a . . . . . . . . . . .
I c h l h y o s t e ~ a
T u f e r p e t o n . . . . . . . . . . . . . . . . . . . . . . . . .
Crossig~rinida . . . . . . . . 1 : " I [ ' 1 [ L o x o m m a t i d a e . . . . . . . . } [ . i Anthracosaur:3 id~a . . . . . . . . . A m n i n t n • I I ] Ais topoda
. . . . . . . . . i
i
N ec t r idea . . . . . . . m , /
Coloste idae • _ . [ ~ A rnph ib in
i
C b S 0 Cb
Fmuas 1 - A gauche: 6tat actuel de la phyIog~nie des vertebras (theories de relations phylog6n~tiques les plus largement admises en 1996; les conodontes ne sont pas inclus dans cette phylog6nie). Traits en pointill~s: taxons dont la monophylie est contro- vers~e; traits doubl6s: taxons notoirement paraphyl~tiques mais dont les relations ne sont pas encore r~solues. A droite, r6parti- tion strat igraphique des taxons; r~partition connue avant 1976 (en nob) et depuis (en blanc); en pointill~s, r@art i t ion des taxons dont l 'attribution syst~matique est controvers~e. Les "Neopterygii basaux" sont compris au sens de Patterson & Rosen (1977). Left- hand side: present state of the vertebrate phylogeny (theories of phylogenetical relationships most widely admitted in 1996; cono- donts not included). Dotted lines: taxa, the monophyly of which is controversial; double lines: notoriously paraphyletic taxa, the interrelationships of which are not yet resolved. Right-hand side: stratigraphic distribution of the terminal taxa; black blocks: dis- tribution known before 1976; white blocks: distribution extended since 1976; dotted blocks: distribution of taxa based on material of controversial systematic attribution. "Basal Neopterygii" sensu Patterson & Rosen (1977).

440
machoires des acanthodiens sont profond6ment diff6rents de ceux des t616ostomes et des 61asmo- branchiomorphes.
Lee placodermes ont fait l 'objet d'un regain d'int6r~t apr6s la publication de la synth6se de Denison (1978). Depuis, le nombre de taxons d6crits s'est accru de pr6s de 40%. Leur apparition a 6t6 recul6e au Silurien inf6rieur (Wang J. 1991) et leur anatomie a 6t6 pr6cis6e grace aux mat6riels d'Australie et du Spitsberg. La recherche des caract6res utiles ~ la construction phylog6n6tique a mis en 6vidence une r~duction pr6coce de l'arc hyoY- dien chez un certain hombre d'entre eux (Goujet 1984), trait coexistant avec des structures endocra- niennes tr~s primitives (Young 1986; Goujet & Young in Leli6vre et al. 1995). Uexploration de nou- velles localit6s (Leli6vre et al. 1994) ont r6v616 l'im- portance des actinol6pides et des rh6nanides. Les placodermes sont aujourd'hui consid6r~s comme le groupe-fr6re de ]'ensemble des autres gnatho- stomes.
Les relations phylog6n6tiques ont 6t6 6tudi6es surtout chez les placodermes, pour lesquels on dispose de donn6es assez completes (Carr in Arsenault et al.; Leli6vre in Arsenault et al. 1995). Les d~bats portent surtout sur la position relative des antiarches et des arthrodires, et sur les formes ~ placer ~ la racine du groupe qui montre une radiat ion adaptat ive remarquable d6s le D6vonien inf6rieur.
LES CHONDRICHTHYENS
Selon la nature des mat6riels 6tudi6s, la connais- sance de l'histoire des chondrichthyens a pro- gress6 dans deux voies. La collecte de nombreux 616ments isol6s dans le Silurien de Mongolie et de Sib~rie (Mongolepida) et dans le D6vonien de l'h6- misph6re sud (Kara ta ju te-Tal imaa in Mark- Kurik 1992; in Leli6vre et al. 1995) a permis la compr6hension des processus tr6s vari6s de crois- sance des 6cailles non placoides.
La d6couverte, notamment dans le Mississippien du Montana (Lurid 1990), et la r6vision d'individus complets ont conduit ~ de nouvelles interpr6tations anatomiquee et fi de nouvelles analyses des rela- tions phylog~n6tiques des groupes. Ainsi une r6vi- sion de la phylog6nie des x6nacanthes a r6v616 que leur 6nigmatique "seconde nageoire anale" ne serait en fait que ]e lobe ventral de la caudale (Poplin & Heyler 1989); la structure dentaire de ce groupe a fait l'objet de r6visions approfondies (Hampe 1993). La connaissance de la phylog6nie du groupe dans son ensemble comporte encore des zones d'ombre: si la monophylie des chondrichthyens est corrobor6e par des apomorphies (notamment le cartilage pris- matique calcifi6, exclusif au groupe), les rapports entre fossiles et actuels sont encore tr6s discut6s.
Certains groupes pal6ozo~ques (Cladoselaches) ne sont probablement ni des 61asmobranches ni des holoc6phales. D'autres (stethacanthides, symmo- riides) sont probablement plus proches des holoc6- phales. Les 61asmobranches ont fait l'objet de plu- sieurs travaux de synth6se (Maisey 1984). I1 en est de m~me pour les holoc6phales, en particulier ceux du Montana (Lund 1986).
LES ACTINOPTt~RYGIENS
D6s les ann6es 1970, l'analyse cladistique a 6t6 le moteur principal des progr~s de l'6tude de la phy- log6nie des actinopt6rygiens. Les fossiles fournis- sent l'essentiel des donn6es utilis6es pour les acti- nopt6rygiens basaux tandis que la phylog6nie des t616ost6ens est fond6e sur les formes actuelles, sus- ceptibles d'analyses ontog6n6tiques et mol6culaires, avec introduction des taxons fossiles a posteriori. L'6tude des relations des grands groupes a conduit l'61aboration des sch6mas encore tr~s utilis6s de Patterson (1973) et Patterson & Rosen (1977): les Cladistia (polypt~ree) sont le groupe-fr6re des Actinopteri (Chondroetei, Neopterygii et plusieurs ensembles paraphyl6tiques dont les pal6onis- coldee); chez les Neopterygii, les Ginglymodi (16pi- sost6es) sont le groupe-fr~re des Halecostomi [Halecomorpha (Amies) + teleostei]. La paraphylie de t616ost6ens m6sozo~ques tels "Leptolepis" ou "Clupavus" a 6t6 reconnue. Des aspects de ce sch6- ma ont 6t6 discut6s en raison d'une r6vision de nora- breux caract6res: toit cranien, suspensorium (V6ran 1988), dents, uroneuraux, etc. Lesprincipales ana- lyses sur les Actinopteri basaux ont 6t6 celles de Gardiner & Schaeffer (1989) et le cladogramme pro- visoire de Lund et al. (in Leli6vre et al. 1995). A par- tir de 1990, la g6n6ralisation des analyses phy- log6n6tiques informatis6es a conduit ~ une remise en question des premiers r6sultats: d'abord, remise en question des relations phylog6n6tiques des t616ost6ens jurassiques (Arratia in Chang et al. 1991); ensuite, restauration de la monophylie des holost6ens (Olsen & McCune 1991); enfin, nouvelles hypoth6ses sur le groupe-fr6re des Teleostei: soit s6mionotiformes seuls (= Semionotidae + ensemble Macrosemiidae-Lepisosteidae) ou s6mionotiformes + hal6comorphes (Olsen & McCune 1991), soit pyc- nodontiformes (Nursall 1996). Pour les t61~ost6ens, la pr6cision des r6sultats sur les Osteoglosso- morpha, les Salmonidae et les Otophysi doit 6tre g6n6ralis6e fi l'ensemble du groupe. Des probl~mes subsistent, telle la coexistence de ]a double articula- tion mandibulaire des Halecomorpha et des uroneu- raux des t616ost6ens chez Vinctifer (Brito 1995).
LES SARCOPTt~RYGIENS
La d6couverte de nombreux taxons nouveaux a profond6ment modifi6 et pr6cis6 la phylog6nie des sarcopt6rygiens ~ trois niveaux. La relation 6troi-

441
te entre dipneustes et porol~piformes a ~t~ confirm~e par la description de Youngolepis et sur- tout Diabolepis (D4vonien inf@ieur, Chine): ce der- nier allie des caract~res de dipneustes (plaques dentaires, lignes de Westoll) et de porol~piformes (Rosen et al. 1981; Cloutier & Ahlberg in LeliSvre et al. 1995; Chang in Arsenault et al. 1995). La meilleure connaissance anatomique des Elpis- tostegalia (=Panderichthyida) montre que ces pois- sons ~ choanes ne sent pas des ost~ol@iformes mais le groupe-fr~re des Tetrapoda, avec lesquels ils partagent la perte des nageoires dorsales et anale, la presence de frontaux et un large contact jugal-quadratojugal (Vorobyeva & Schultze 1991). Enfin, pour les premiers Tetrapoda, de nouveaux genres du D~vonien sup@ieur (Obruchevichthys, Elginerpeton, Metaxygnathus, Ventastega ) assurent une transit ion morphologique entre les Elpis- tostegalia et les Tetrapoda pourvus de membres dactyliques (Ahlberg & Milner 1994). I1 a aussi ~t~ montr~ que, par sa polydactylie, son appareil bran- chial et sa nageoire caudale, Acanthostega est net- tement plus primitif qu'Ichthyostega, Tulerpeton et les t~trapodes post-d~voniens (Clack & Coates in Arsenault et al. 1995).
Les points controvers~s de la phylog~nie des sar- coptSrygiens res ten t la posit ion des onycho- dontiformes, la monophylie des ost~ol~piformes et les relations entre les premiers Tetrapoda post- dSvoniens. La principale ~nigme reste l'origine des phalanges: de r~centes recherches sur du mater ie l actuel sugg~rent que les phalanges seraient des n~oformations donc sans homologies avec les radiaux des nageoires paires.
BIOSTRATIGRAPHIE ET CORRI~LATIONS
Une tendance majeure de la pal~oichthyologie r@ente est la prise de conscience de l'utilit~ des restes de poissons pour la datation des terrains s~dimentaires. Ceci s'est t radui t par robtention d'un projet du Programme In te rna t iona l de Correlation G~ologique de I 'UNESCO (Projet n ° 328 "Palaeozoic microvertebrate biochronology and global marine/non-marine correlation" entre 1991, 1996; Turner 1993). On d~signe par "micro- vertSbr~s" les restes microscopiques ou de petite taille de vertebras (dents, ~cailles, os isol~s du cr~ne, ~pines, l~pidotriches, otolithes, vert~bres, etc). Souvent appel~s "ichthyolithes" ou "micro- restes", ils sent compos~s de t issus (dentine, ~mail, ~maillo~de, os) min@alis~s en phosphate de calcium d~s l'Ordovicien.
O r d o v i c i e n : en moins de 20 ans, l'aire de r~par- tition des vertebras ordoviciens s'est ~tendue de
l'Am@ique du Nord ~ l 'Australie et ~ l'Am~rique du Sud (Bolivie, Argentine) (Elliott et al. 1991). Les plus vieux vert~br6s connus sent de l'Ordo- vicien inf@ieur d'Australie (Young in Shergold et al. 1991). Cependant, les sites r~put~s ordoviciens de la vall~e du Rio Iglesiani (Bolivie), d'ofl 6tait cens~ provenir l 'Arandaspide Andinaspis suarezo- rum, se sent r~v~l~s d~voniens (Gagnier in Su4rez-Soruco 1991).
Silurien: l 'enregistrement en vertebras siluriens est relat ivement continu dans les s@ies de la r~gion balte, de Grande-Bretagne et de l'Arctique russe (Karatajute-Talimaa & Predtechenskyi in Arsenault et al 1995). Les s6ries de rArctique canadien, de Sib@ie, d'Am@ique du Sud et de Chine sent tr~s prometteuses (Blieck & Janvier 1991). Toutes ont permis l '~tablissement d'une 6chelle biostratigraphique fond~e sur les th~lo- dontes, les acanthodiens et les ost~ichthyens: les 12 biozones d~finies pour l 'ensemble du Silurien ont une valeur, sinon mondiale, du moins r~giona- le et sent corr~16es ~ d'autres @helles comme celles des conodontes et des graptolites (M~rss et al. in Leli~vre et al. 1995).
D6vonien: c'est le syst~me pal6ozo~que le plus riche en donn~es ichthyologiques. On distingue le D~vonien inf@ieur plus souvent siliciclastique (type Vieux Gr~s Rouges), du D~vonien moyen et sup@ieur plus souvent carbonate. Au D~vonien inf@ieur, on note un fort end~misme des ichthyo- faunes: ceci se t raduit par plusieurs ~chelles bios- tratigraphiques, ~ valeur r6gionale ou continenta- le, fond~es sur les pt@aspidomorphes, les th~lo- dontes, les gal~aspides ou les acanthodiens (Long 1993). Ily~s & Elliott (1994) ont propos~ de nou- velles datations pour la plupart des formations vertebras du D~vonien inf@ieur du centre-ouest des USA. Le D~vonien inf@ieur d'Arabie Saoudite a fourni plusieurs dizaines de niveaux fossiliFeres, riches en macro- et microrestes, en pat t ie inter- cal~s avec des niveaux ~ conodontes, brachiopodes et plantes (Leli~vre et al. 1994). Au D~vonien moyen et sup@ieur, trois ~chelles biostratigra- phiques compl~mentaires ~ vocation mondiale ont ~t6 ~labor~es: celles des acanthodiens, des chon- drichthyens Phoebodontidae (Ginter & Ivanov 1992) et des th~lodontes Turiniidae. En l 'absence de ces tax0ns "p~lagiques', d 'autres groupes sent utilis~s r~gionalement: les p lacodermes en Antarctique et en Chine, les sarcopt@ygiens dans le nord de la France et la Belgique, les actinopt~- rygiens en Russie (Chang et al. 1991; Long 1993; Leli~vre et al. 1995; Turner 1995). Au Canada, la Formation d'Escuminac, avec en parti- culier Eusthenopteron et Elpistostege, a ~t~ dat~e avec precision du Frasnien meyen ~ l'aide des miospores (Cloutier et al. sous presse). En Russie, la Formation Khovanshchina qui a fourni Tuler-

442
peton, un des plus anciens t@trapodes, est datEe du Famennien terminal sur la base des cono- dontes (Alekseev et al. 1994). D'autres dEcou- ver tes ont @t@ rEalis@es dans le D@vonien d'AmErique du Sud, du Maroc et d'Asie du Sud- Est (Long 1993). C'est la sErie des Pays Baltes, la plus complete, qui sert de rEfErence au moins pour le DEvonien moyen et sup6rieur du Continent des Vieux Gr~s Rouges, tandis que celle du Spitsberg est utilis6e pour le D@vonien inf6rieur circum-arc- tique. En Australie, les Echelles ichthyologiques ont ~t6 calEes sur celle des conodontes et par rap- port aux nouveaux figes radio-isotopiques (Young in Long 1993).
C a r b o n i f ~ r e : les chondrichthyens sont un outil performant pour la France-Belgique, l'Allemagne, la Pologne, la Russie et l'Angleterre. De re@me, aux USA, chondrichthyens et acanthodiens parti- cipent fi l 'Etablissement de corr@lations. Le passa- ge du D6vonien au Carbonifere a Et6 caract@risE dans son stratotype en Montagne Noire ainsi qu'en Turquie, en Pologne et en Russie. Le CarboniFere sup~rieur continental d'Europe cen- t ra le renferme des chondrichthyens Xena- canthida utiles pour des correlations au moins %gionales. En Australie les microrestes, entre autres de chondrichthyens, ont servi ~ des syn- theses biostratigraphiques (Turner 1990).
P e r m i e n : des ~tudes biostrat igraphiques des chondr ichthyens du Carbonif~re terminal- Permien inf@rieur des USA et d'Europe, menEes par Johnson (1992) et Schneider et al. (1995), ont conduit h une synth~se stratigraphique en parti- culier pour l 'ensemble des grands bassins d'Europe, avec corr61ation aux bassins de Russie et calage partiel sur les donn@es radiochronolo- giques. Dans le PalEozo~'que alpin, c'est le Permien qui a fourni jusqu'h present le plus de restes de vert6br6s, ~ la lois en facies m a r i n e t en facies continental (Blieck et al. 1995).
P o s t - P e r m i e n : un apport majeur est la synth~se r@alisEe par Nolf (1985) sur les otolithes. Ces petites pi~ces min@ralisEes de l'oreille interne des poissons sont connues d~s le D6vonien, chez les acanthodiens, off elles sont trgs rares. Elles deviennent abondantes h partir du Jurassique. Elles sont beaucoup utilis@es en biostratigraphie du C@nozo~que d'Europe (bassins belge et aqui- tain).
PALEOENVIRONNEMENTS Avec la recherche des microrestes, l 'extension de l 'information h l'Echelle globale et la rE@valuation des donn~es, le r61e d'indicateurs pal@oenvironne- mentaux attribu6 ~ certains groupes de vertEbr@s
inf@rieurs a 6t6 remis en cause. On estime ainsi que toutes les localit6s h vert@br@s siluriens seraient marines, comme les localit@s ordovi- ciennes. De re@me au Pal@ozoique moyen, des formes pr@c@demment r6put@es dul~aquicoles (placodermes, chondrichthyens x6nacanthides, sarcopt@rygiens) ~taient pr@sentes dans des envi- ronnements marins. C'est ainsi que les faunes des Vieux Gr~s Rouges du Spitsberg sont maintenant consid~r~es comme marines (Goujet 1984; Blieck 1982, 1985). Cette interpr@tation a @t@ corrobor~e par l'analyse des relations entre les faunes du Boulonnais et celles de l'Orcadien d'Ecosse (Leli~vre et al. 1988). De m@me, l'@tude globale men@e sur le gisement de Miguasha (Schultze 1996), avec la g6ochimie (@tudes des proportions des isotopes stables de l'exyg~ne, du carbone et du bore), la s@dimentologie et les @tudes de palynofa- ci~s en plus des donn@es pal6obiologiques, permet de proposer plut6t un module d'estuaire pour la Formation d'Escuminac (Blieck et al. in Turner 1995; Cloutier et al. sous presse): ceci supposerait, en raison de la presence d'Elpistostege, que le pas- sage du milieu aquatique au milieu terrestre chez les vert@br6s aurait eu lieu en milieu estuarien et non lacustre.
A contrario, les donn6es sur certains gisements du Cr@tacE inf@rieur indiquent un environnement lacustre et non marin. C'est le cas des gisements espagnols de Las Hoyas (Serrania de Cuenca) et,
un moindre degrE de certitude, du Montsec (Province de Lleida). A Las Hoyas les arguments viennent des donnEes sEdimentologiques et pal~og@ographiques, de l'6tude de la flore et de la faune compl6tEes par les renseignements sur ]a taphonomie (Sanz et al. 1988; Mel@ndez 1995). L'6tude des compositions isotopiques du stron- tium relev6es sur les poissons et les calcaires encaissants (Talbot et al. 1995) confirme les donn@es pr~c6dentes.
Il faut souligner le nombre croissant des @tudes pluridisciplinaires qui ont coordonn6 les travaux de divers spEcialistes (pal@obotanistes, palEonto- logues invert@bristes et vertEbristes, g@ologues, s@dimentologues,...) sur un mSme bassin ou un m~me gisement pour aboutir h des reconstitutions palEoenvironnementales et pal@oEcologiques. C'est le cas, par exemple, pour plusieurs sites car- bonif~res des USA (Mazon Creek, Hami l ton Quarry, Kinney Brick Quarry) et d'Europe (East Kirkton et le bass in s tEphano-autunien de Montceau-les-Mines, Poplin & Heyler 1994). Le site du KimmEridgien sup@rieur de Cerin, exploit~ banc par banc depuis 20 ans, a fait l 'objet d'une synthgse similaire (Bernier & Gaillard 1994).

443
PALEOBIOGEOGRAPHIE
La g6odynamique globale du Pal6ozo~'que est celle d'une accr6tion des continents qui aboutit/~ une Pang6e permienne. Cette dynamique centrip6te est difficile ~ concilier avec un module d'analyse biog6ographique sur le mode dispersif/vicariant. On est donc amen6 fi comparer les provinces fau- niques fond6es sur des assemblages fossiles aux blocs continentaux propos6s par les g~odynami- ciens/pal6omagn6ticiens (Scotese & McKerrow in McKerrow & Scotese 1990; Li et al. in Long 1993). Plus on observe de taxons communs dans les assemblages de deux (ou plus) plates-formes continentales /~ une 6poque donn6e, plus on peut supposer qu'elles 6taient proches g6ographique- ment (Talent et al. 1987). I1 est 6galement justifi6 de t rai ter la plupart des assemblages de vert6br6s pal6ozo~ques comme ceux de brachiopodes ou de mollusques par exemple. En effet, comme pour l'Actuel, la majorit6 des poissons marins des pro- vinces n6ritiques pal6ozo~'ques 6taient 6troite- ment li6s aux plates-formes continentales au-des- sus desquelles ils vivaient. C'est ainsi qu'on obser- ve des end6mismes marqu i s fi l'Ordovicien (pt6- rasp idomorphes diff6rents sur Lauren t ia et Gondwana, Elliott et al. 1991), au Silurien (assemblages diff6rents sur Laurentia, Acadia [= Avalonia Ouest], Baltica, Siberia, Tuva et Chine du Sud, Blieck & Janvier 1991) et au D6vonien inf6rieur (d6finition de cinq provinces pal6oich- thyologiques: "Euramerica cephalaspid province", "Siberia amphiaspid province", "Tuva tannuaspid province", "South China galeaspid-yunnanolepid province" et "East Gondwana wuttagoonaspid- phyllolepid province", Young in Long 1993). Cependant la r6partition des taxons ordoviciens pose un probl6me majeur: la pr6sence de pt6ras- pidomorphes sur Laurent ia et Gondwana au moins d~s l'Arenig implique soit un contact entre ces deux blocs soit une aire oc6anique franchis- sable ~ l'Arenig ou plus tard (Caradoc), ce que ne montrent pas les reconsti tutions pal6og6ogra- phiques publi6es.
Au cours du D6vonien moyen et sup6rieur, on observe un cosmopolitisme de plus en plus mar- qu6 des ichthyofaunes fi mettre en relation avec l '6 tabl issement d'un contact entre Gondwana Ouest et Laurent ia + Baltica (fin des phases cal6- doniennes et d6but des phases varisques de l'oro- gen6se pal6ozo~'que). On peut rapprocher au D6vonien sup6rieur la Chine du Nord et la Chine du Sud du Gondwana Est sur la base de la r6par- tition des antiarches Sinolepidae (Long 1993). Dans les s6ries carbonat6es du D6vonien moyen et sup~rieur sont r6colt6s les assemblages taxons "p61agiques": chondrichthyens Phoebodon- tidae, th61odontes Turiniidae, associ6s aux cono-
dontes polygnathides et/ou palmatol6pides. On entend par "p61agiques" les taxons d6finis ainsi biologiquement (planctoniques ou nectoniques) et
vaste r6partition g6ographique, parfois mondia- le. Les autres tmxons p61agiques, ceux des pro- vinces marines n6ritiques, sont end6miques aux plates-formes continentales (au sens ggologique) et n 'autorisent pas de corr61ations mondiales; ils apportent le m~me type d'information que les taxons benthiques. Apr6s le D6vonien sup6rieur, la distinction entre assemblages aquatiques et terrestres est facilit6e par l 'abondance des t6tra- podes qui signent les pal6ocommunaut6s ter- restres. Au D6vonien sup6rieur une seule provin- ce fi t6trapodes a 6t6 identifi6e de la Laurent ia au Gondwana Est, mais avec un genre diff6rent dans chaque gisement (Long 1993; Ahlberg 1995). Cette r6part i t ion implique donc un contact entre Lauren t ia et Gondwana au Frasn ien ou au Famennien, 6bauche pr6coce d'une Pang6e que certains auteurs proposent m6me d~s le D6vonien inf6rieur (Kent & Van der Voo in McKerrow & Scotese 1990). Milner (in Long 1993) d6finit une seule province au Carbonif'ere (Am6rique du Nord + Europe de l'Ouest; probablement par manque de donnSes ailleurs), mais trois provinces au Permien inf6rieur ( "Edaphosaur id-Nect r idean province", "Mesosaurid province" et "Seymouria- morph province"). Les relations entre la phylog6- nie des t6trapodes basaux et l 'histoire pal6og6o- graphique du Pal6ozo~'que sup6rieur ne sont pour- rant pas r6solues. Sur la Pang6e du Permien sup6- rieur, les t6trapodes ont une r~partition quasi- mondiale qui serait le r6sultat de l 'extension de la province fi 6daphosaures-nectridiens aprSs la gla- ciation du Permien inf6rieur. N6anmoins quatre sous-provinces fond6es sur des taxons end6- miques sont reconnues. Les t6trapodes n'appor- tent pour l ' instant pas d 'arguments en ce qui concerne les relations pal6og6ographiques des nombreux blocs nord-gondwans en d6rive vers le nord ("Cimmeria").
CONCLUSION ET PERSPECTIVE S
Ce bilan de vingt ann6es d'6tudes sur les vert6- br6s inf6rieurs n'est 6videmment pas exhaustif. Nous avons sur tou t voulu souligner ici les grandes tendances et les faits principaux qui sont responsables de l'essor de ces recherches. Celui-ci est dfi d'abord ~ la diversit6 des th6mes et des approches et fi la fa~on novatrice avec laquelle ils ont 6t6 trait6s. I1 est dfi ensuite ~ la d6couverte de faunes dans des r6gions du monde jusqu'ici inex- plor6es (y compris celle de t6trapodes nouveaux dans des collections anciennes). Les domaines off les r6sultats sont encore incertains ont l 'avantage d'avoir 6t6 rep6r6s et identifi6s.

444
Le bilan de l 'analyse phylog6n6tique app]iqu6e aux vert6br6s inf6rieurs est d6monstra t i f Le r61e des fossiles dans les constructions phylog6n6- tiques, en d6pit de l ' information partielle qui les caract6rise, a 6t6 reconnu: en effet ils pr6sentent des combinaisons de caract6res inconnues chez les actuels, si bien qu'ils peuvent, sous certaines conditions de fossilisation et d'age, am61iorer l'es- t imation d'une hypoth6se, voire stabiliser les rela- tions de parent6 entre des taxons actuels. Cet aspect nouveau de l 'analyse phylog6n6tique a per- mis de sou]igner l ' importance des taxons basaux.
L'utilisation des vert6br6s pal6ozoiques en bios- t ra t igraphie s'est consid6rablement d6velopp6e, no tamment grace au rS]e f6d6rateur du Projet 328 du PICG. Les r6sultats synth6tiques de ce pro- gramme seront publi6s prochainement. Le Projet 406 du PICG sur les vert6br6s pal6ozoiques cir- cum-arctiques prendra le relais et exploitera le mat6riel abondant r6colt6 au cours de deux exp6- ditions dans l'Arctique canadien en 1994 et 1995. Certains groupes ou types de mat6riels pr6c6dem- ment d61aiss6s dans ce domaine ont 6t6 pris en compte, en part iculier les microrestes. Ainsi de nombreuses 6chelles biostratigraphiques ~ valeur r6gionale ou mondiale ont 6t6 61abor6es et leurs corr61ations discut6es; le niveau strat igraphique de certains sites a 6t6 pr6cis6.
La discussion du rSle d'indicateurs pal6oenviron- nementaux de nombreux taxons a provoqu6 la remise en cause de certaines id6es revues ou la mise en 6vidence de la na ture de certains pal6o- milieux.
Ces derni~res ann6es les 6tudes en pal6obiog6ogra- phie ont dispos6 de reconstitutions plus pr6cises des terres 6merg6es et des aires marines, fond6es sur diverses donn6es: c]imat, pal6omagn6tisme, analyse du niveau marin entre autres. Ces sch6mas sont confront6s aux hypoth6ses phylog6n6tiques concernant les vert6br6s inf6rieurs et r6sultant de l'application de la m6thode cladistique.
Ce tableau favorable des r6sultats de la pal6onto- logie des vert6br6s inf6rieurs ne lui est certes pas exclusif et les recherches sur bien d 'aut res groupes fossiles ont un bilan positif pour des rai- sons similaires. Mais quel sera l 'avenir des tra- vaux ainsi engag6s? Le renouvel lement des id6es amorc6 ~ la fin des ann6es soixante-dix a montr6 que les nouvelles m6thodes sont g6n6ratrices de th6ories f6condes et que le caract6re transitoire des hypoth6ses qu'elles g6n~rent est en soi por- teur de projets d'avenir. Uanalyse phylog6n6tique permet de pr6dire la d6couverte de certains taxons darts des niveaux beaucoup plus anciens et la logique de la pal6obiog6ographie cible les recherches sur certaines r6gions. Mais les avan- c6es de la recherche sont directement li6es aux
6quipes en place: si le train, actuel lement bien trop lent, du remplacement des sp6cialistes ne s'acc6l~re pas, combien serons-nous dans vingt ans ? Les fluctuations des 6quipes au gr6 des conjonctures causent d' irr6parables pertes dans le suivi de la connaissance, du savoir-faire et du mat6riel.
(Le pr6sent travail est en part ie une contribution au Projet 328 "Palaeozoic microvertebrate bio- chronology and global marine/non marine correla- tions" du PICG.)
RI{FI~RENCES AHLBERG P.E. & MILNER A.R. 1994 - The origin and early
diversification of tetrapods. Nature, 368: 507-514. AHLBERG P.E. 1995 - Elginerpeton paneheni and the ear-
liest tetrapod elade. Nature, 373 (6513): 420-425. ALDRIDGE R.J., BRINGS D.E.G., SMITH M.P., CLARKSON
E.N.K. & CLARK N.D.L. 1993 - The anatomy of Conodonts. Philosophical Transactions of the Royal Society of London, B, 340: 405-421.
ALEKSEEV A.A., LEBEDEV O.A., BARSKOV I.S., BARSKOVA M.I., KONONOVA L.I. & CHIZHOVA V.A. 1994 - On the stratigraphic position of the Famennian and Tournaisian fossil vertebrate beds in Andreyevka, Tula region, Central Russia. Proceedings of the Geologists Association, 105: 41-52.
ARSENAULT M., LELIl~VRE H. & JANVIER P. (eds) 1995 - ]~tudes sur les Vert6br6s inf6rieurs (VIIe Symposium International, Parc de Miguasha, Qu6bec, 9-22 Juin 1991). Bulletin du Musdum National d'Histoire Naturelle, Paris, 4 s6r., 17, C (1-4): 529 p.
BERNIER P. & GAILLARD C. (eds) 1994 - Les calcaires litho- graphiques. S6dimentologie, Pal6ontologie, Taphono- mie. Geobios, M6m. sp. 16: 3-70.
BLmCI< A. 1982 - Les grandes lignes de la biog6ographie des H6t6rostrac6s du Silurien Sup6rieur-D6vonien Inf6rieur dans le domaine Nord-Atlantique. Palaeo- geography, Palaeoclimatology, Palaeoecology, 38: 283- 316.
BLIECK A. 1985 - Pal6oenvironnements des H6t6ros- trac6s, Vert6br6s agnathes ordoviciens ~ d6voniens. In FISCHER J.-C. (organ.): Journ6es d'6tude sur les indicateurs pal6obiologiques de milieux (RCP 641, Paris, 1984). Bulletin du Musdum National d'Histoire Naturelle, 4 s6r, 7, C (2): 143-155.
BLIECK A., CONTI M.A., FL~JGEL H.W., GAND G., HUBMANN B.~ LELII~VRE H., MARIOTTI N., NIcosm U., POPLIN C., SCHNEIDER J.V¢ ~. • WERNEBURG R. 1995 - The Alps, a quasi-desert in Palaeozoic vertebrate database, and the Gondwana-Laureuropa palaeogeographic rela- tionships. Ichthyolith Issues, 15: 8-13.
BLIECK A. & JANVIER P. 1991 - Silurian vertebrates. In BASSETT M.G., LANE P.D. & EDWARDS D. (eds.): The Murchison Symposium. Palaeontology, Special Paper 44: 345-389.
BRITO P.M. 1995 - R6vision des Aspidorhynchidae (Pisces, Actinopterygii) du M6sozoique: ost6ologie et relations

445
phylog6n6tiques. Th6se de Doctorat du Mus6um nat ional d 'Histoire naturel le , Paris, 188 p. (in6dit).
CHANG MEE-MANN, LIU YU-HAI & ZHANG GUO-RUI (eds.) 1991 - E a r l y v e r t e b r a t e s and r e l a t ed p rob lems of e v o l u t i o n a r y biology ( I n t e r n a t i o n a l Sympos ium, Bei j ing , 12-26 Sept . 1987). Science Press, Beijing: 514 p.
CLOUTIER R., LOBOZIAK S., CANDILIER A.-M. & BLIECK A. 1996 - Biostratigraphy of the Upper Devonian Escuminae Formation, eastern Qu6bee, Canada: a comparative study based on miospores and fishes. In FAIRONT-DEMARET M. (ed.), Special Issue: Maurice Streel . Review of Palaeobotany and Palynology, 93 (1-4): 191-215.
DENISON R.H. 1978 - Placodermi, Gus tav Fischer, S tu t tga r t , 128 p.
DERYCKE C. 1.990 - Appl ica t ion de la cathodolumines- cence h la pal6ohistologie. Comptes Rendus de l'Aca- dgmie des Sciences, Paris, 310, 2: 1561-1565.
ELLIOTT D.K., BLIECK A.R.M. & GAGNIER P.-Y. 1991 - Ordovician ver tebra tes . In BARNES C.R. & WILLIAMS S.H. (ed.s): Advances in Ordovician geology. Geolo- gical Survey of Canada Papers 90 (9): 93-106.
FOREY P.L. & JANVIER P. 1993 - Agna thans and the origin of j awed ver tebra tes . Nature, 361: 129-134.
GAGNIER P.-Y. 1993 - Sacabambaspis janvieri, Vert6br6 ordovicien de Bolivie. 1, Ana lyse phylog6n6tique. Annales de Palgontologie, 79: 119-166.
GARDINER B.G. & SCHhEF~ER B. 1989 - In te r re la t ionsh ips of lower ac t inop te ryg ian fishes. Zoological Journal of the Linnean Society, 9 (7): 135-187.
GINTER M. & IVANOV A. 1992 - Devonian pheobodont teeth. Acta Palaeontologica Polonica, 37 (1): 55-75.
CxOUJET D. 1984 - Les Poissons placodermes du Spits- berg. Ar th rod i res Dolichothoraci de la Format ion de Wood-Bay (D6vonien Inf6rieur). t~ditions du CNRS: 284 p.
HAMPE O. 1993 - Var ia t ion of xenacan th id tee th in the Pe rmo-Carbon i f e rous depos i t s of the S a a r - N a h e Basin (SW-Germany). In HEIDTKE U. (Compiler): New Research on Permo-Carboniferous Faunas. Pollichia- Buch 29: 37-51, Bad Dtirkheim.
ILYI~S R.R. & ELLIOTT D.K. 1994 - New Ear ly Devonian p t e r a s p i d i d s (Agnatha , He te ros t rac i ) from Eas t - Cen t ra l Nevada . Journal of Paleontology, 68 (4): 878- 892.
JANVmR P. 1996 - The dawn of the ver tebra tes : charac- te rs ve r sus common ascent in cur ren t c ran ia te phy- logenies. Palaeontology, 39: 259-287.
JOHNSON G.D. 1992 - Chondr ich thyan b ios t r a t ig raphy of the Nor th Amer ican Pe rmian System. In Contri- butions to Eurasian Geology. Occasional Publica- tions ESRI, New Series, 8B: 41-50.
LEHMAN J.-P. (ed.) 1975 - Probl6mes ac tue ls de Pa l6on to log ie -Evo lu t ion des Vert6br6s. Par is , Colloque international C.N.R.S. N 218 :909 p.
LELII~VRE H., GOUJET D., BLIECK A. & JANVIER P. 1988 - Poissons du D6vonien du Boulonnais (France). In D. BRICE (ed.): Le Dgvonien de Ferques, Bas Boulonnais. Biostratigraphie du Paldozo~que, 7: 503-522.
LELII~VRE H., JANJOU D., HALAWANI M., JANVIER P., AL MUALLEM M.S., WYNNS R. & ROBELIN C. 1994 -
Nouveaux Vert6br6s (Placodermes, Acanthodiens , Chondr ichthyens et Sarcopt6rygiens) de la Forma- tion de J a u f (D6vonien inf6rieur, r6gion de A1 Huj, Arable Saoudite) . Comptes Rendus de l'Acaddmie des Sciences, Paris, 319, 2: 1247-1254.
LELII~VRE H., WENZ S., BLIECK A. & CLOUTIER R. (eds.) 1995 - Premiers Vert6br6s et Vert6br6s inf6rieurs (VIIIe Congr6s In te rna t iona l , Paris , 4-9 Sept . 1995). Geobios, M6m. sp. 19:409 p.
LONG J.A. (ed.) 1993 - Palaeozoic vertebrate biostrati- graphy and biogeography. Belhaven Press, London, 369 p.
LUND R. 1986 - The d ivers i ty and re la t ionsh ips of the Holocephal i . In T. UYENO, R. ARAI. T. TAINDI & MATSUURA K. (eds.), Indo-Pacific F i sh Biology. Procee- dings of the second International Conference on Indo- Pacific Fishes. Ichthyological Society of Japan: 97- 106.
LUND R. 1990 - Chondr ich thyan life h is tory s tyles as revealed by the 320 mil l ion years old Miss i s s ipp ian of Montana. Environmental Biology of Fishes, 27: 1- 19.
MAISEY J.G. 1984 - Higher e l a smobranch phylogeny and bios t ra t igraphy. Zoological Journal of the Linnean Society, 82: 33-54.
MARK-KURIK E. (ed.) 1992 - Fossil f ishes as l iving ani- mals. Academia. 1 , Academy of Sciences of Estonia, Tallinn: 299 p.
McKERROW W.S. & SCOTESE C.R. (eds.) 1990 - Palaeozoic P a l a e o g e o g r a p h y and Biogeography. Geological Society of London, M. 12: 1-21.
MELI~NDEZ N. 1995 - Las Hoyas: a l acus t r ine Konservat - Lagerst~itte Cuenca, Spain. Editorial Universidad Complutense Madrid: V + 89 pp.
NOLF D. 1985 - Otoli thi Piscium. In SCHULTZE H.-P. (ed.): Handbook of Paleoichthyology, vol. 10, G. F ischer Publicat ions, S tu t tga r t : 145 p.
NURSALL J.R. 1996 - The phylogeny of the Pycnodont fishes. In G. ARRATIA & G. VIoHm (eds.): Mesozoic fishes - Systematies and Palaeoecology: 125-152.
OLSEN P.E. & MCCUNE A.R. 1991 - Morphology of the Semionotus elegans species group from the Ear]y Jurass ic Pa r t of the N e w a r k Supergroup of Eas t e rn N o r t h A m e r i c a wi th commen t s on the F a m i l y Semionot idae (Neopterygii) . Journal of Vertebrate Paleontology, 11 (3): 269-292.
ORVIG T. 1973 - Acanthodian dent i t ion and its bea r ing on the re la t ionships of the group. Palaeontographica, 143 (A): 119-150.
PATTERSON C. 1973 - In t e r re l a t ionsh ips of holosteans. In GREENWOOD P.H., MILES R.S. & C. PATTERSON (eds.) I n t e r r e l a t i o n s h i p s of f ishes. London, A c a d e m i c Press: 233-305.
PATTERSON C. & ROSEN D.E. 1977 - Review of ichthyo- dectiform and o ther Mesozoic te leos t fishes and the theory and pract ice of classifying fossils. Bulletin of the American Museum of Natural History, 158 (2): 82-172.
POPLIN C. & HEYLER D. 1989 - Evolut ion et phylog6nie des x6nacanthi formes (=pleuracanth i formes) (Pisces Chondrichthyes) . Annales de Paldontologie, 75 (4): 1- 34.

446
POPLIN C. 8z HEYLER D. 1994 - Quand le Mass i f Cent ra l ~tai t sous l%quateur . Un ~cosystNme carbonif'ere h Montceau-les-Mines. Mdmoires de la Section des Sciences 12, 331 p. sous la direct ion de C. Poplin & D.Heyler. Edi t ions du CTHS, Paris.
ROSEN D.E., FOREY P.L., GARDINER B.G. & PATTERSON C. 1981 - Lungfishes, te t rapods , paleontology and ple- siomorphy. Bulletin of the American Museum of Natural History, 167: 159-276.
SANZ J.L., WENZ S., YEBENES A., ESTES R., MARTINEZ- DELCLOS X., JIMENEZ-FUENTES E., DIEGUEZ C., BUSCALONI A.D., BARBADILLO L.J. & VIA L. 1988. - An E a r l y Cre taceous f auna l and f loral con t inen ta l assemblage: Las Hoyas fossil site (Cuenca, Spain). Geobios, 21 (5): 611-635.
SCHNEIDER J.W., R(3BLER R. & GAITZCH B.G. 1995 - XIII In t e rna t iona l Congress on Carboniferous-Permian (XIII ICC-P, Aug. 28 - Sept. 2, 1995, Krak6w, Poland). Guide to excursion A5: S t r a t i g r aphy and facies of the Middle European cont inen ta l Carboniferous and Pe rmian (Aug. 21-27, 1995). Panstwowy Instytut Geologiczny publ., Warszawa, 31 p.
SCHULTZE H.P. & CLOUTIER R. (eds.) 1996 - Devonian Fishes and Plants of Miguasha, Quebec, Canada. Verlag Dr. Fr iedr ich Pfeil. Munchen: 374 p.
SHERGOLD J.H. coord., wi th contr ibut ions by ELPHINS- TONE R., LAURIE J.R., NICOLL R.S., SHERGOLD J.H., WALTER M.R., YOUNG G.C. & ZANG W. 1991 - Late Proterozoic and Ear ly Palaeozoic palaeontology and b io s t r a t i g r aphy of the Amadeus Basin. In KORSAB D.J. & KENNARD J.M. (eds.): Geological and geophysi- cal studies in the Amadeus Basin, Central Australia. Bureau of Mineral Resources, Bulletin 236: 97-111.
SU~REz-SoRuco R. (ed.) 1991 - Fosiles y facies de Bolivia. VoI. I: Vertebrados. Revista Tgcnica de Yacimientos Petrolfferos Fiscales Bolivianos, 12 (3-4): 371-379.
TALBOT M.R., MEL]~NDEZ N. & M.A. FREGENAL-MARTINEZ 1995 - The waters of the Las Hoyas lake: sources and limnological characteristics. In M. N. MEL]~NDEZ (ed.): Las Hoyas, a lacustrine Konservat-Lagerstiitte Cuenca, Spain. Universidad Complutense de Ma- drid: 11-16.
TALENT J.A., GRATSIANOVA R.T. • YOLKIN E.A. 1987 - Prototethys: fact or phan tom? Palaeobiogeography in re la t ion to the crus ta l mosaic for the As ia -Aus t r a l i a hemisphere in Devonian-Ear ly Carboniferous t imes. In McKENzIE K.G. (ed.): Shallow Tethys 2. Balkema Publ., Rot terdam: 87-111.
TURNER S. 1990 - Ear ly Carboniferous Remains from the Rockhampton Distr ict , Queensland. Memoirs of the Queensland Museum, 28 (1): 65-73.
TURNER S. - 1993 - IGCP 328: Palaeozoic microver tebra- te workshop in 1992. Episodes, 16, 3: 398-401.
TURNER S. (ed.) 1995 -Moscow-94 Workshop and IGCP 328: Palaeozoic M i c r o v e r t e b r a t e s 1995 Report. Ichthyolith Issues, Special Publication 1:71 p.
VERAN M. 1988 - Les ~lNments accessoires de l 'arc hyo~- dien des poissons t~l~ostomes (Acan thod iens + Oste ich thyens) fossiles et actuels . Mdmoires du Musdum National d'Histoire Naturelle, Paris, Sciences de la Terre, 54: 1-98.
VOROBYEVA E.I. & SCHULTZE H.P. 1991 - Descr ipt ion and sys temat ics of pander ich thy id fishes wi th comments on the i r re la t ionships to te t rapods . In SCHULTZE H.P. & TRUEB L. (eds.): Origins of the higher groups of tetrapods. Controversy and consensus: 68-109, I thaca.
WANG J. 1991 - The Ant ia rch i from Ear ly S i lu r ian of Hunan. Vertebrata Palasiatica, 29, 3, 24 (3): 240-244.
YOUNO G.C. 1986 - The re la t ionships of p lacoderm fishes. Zoological Journal of the Linnean Society, 88: 1-57.
C. POPLIN, D. GOUJET, P. JANVIER, H. LELIEVRE, M. VERAN & S.WENZ
Museum national d'Histoire naturelle Laboratoire de Pal~ontologie - URA 12 du CNRS
8 rue Buffon, 75005 Paris France
A. BLIECK, R. CLOUTIER & C. DERYCKE Universit~ des Sciences et Technologies de Lille
Sciences de ]a Terre URA 1365 du CNRS
59655 Vitleneuve d'Ascq cedex