Embed Size (px)
Citation preview

PHYSIQUE CELLULAIREFusion mitochondriale et autres curiosités
membranaires (8)
Jean-Pierre HENRY4 Juin 2009

QUELQUES RAPPELS: Physique de la fusion
• Nécessité d’un rapprochement des membranes• Nécessité d’une « exposition » des cœurs hydrophobes:
– Courbure « spontanée »: nature des phospholipides– Courbure induite: insertion partielle de protéines, effet de
protéines fusogènes– Tension superficielle
• Intermédiaires identifiables– Hémifusion: fusion des seules monocouches adjacentes– Pore de fusion: conductance mesurable

Fusion et déformations membranaires
• L’étude des fusions cellule-cellule suggère que lesdéformations membranaires pourraient jouer un rôleimportant dans certaines fusions
• Ces déformations sont induites de manièresdifférentes, souvent par des protéines en interactionavec les têtes polaires d’une monocouche
• Les fusions mitochondriales peuvent être un autreexemple de ce type
• Indépendamment des fusions, des déformationsmembranaires spectaculaires peuvent êtreobservées
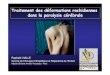
Fusion mitochondriale
• La microscopie électroniquea montré la mitochondriecomme un objet autonome(ballon de rugby)
• La microscopie optique(marquage GFP) montre unréseau dynamique lié aucytosquelette
• Ce réseau montre desfusions et fissions multiples
(Chen et al (2003) J Cell Biol, 160, 189)
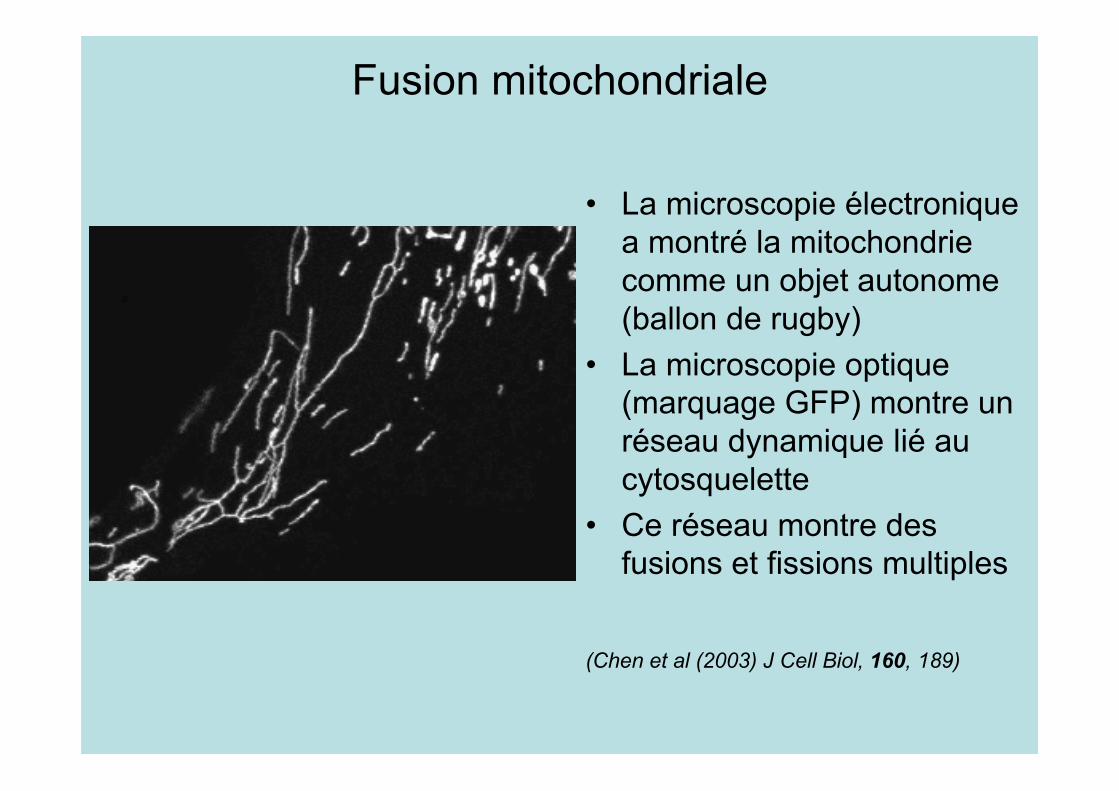
L’équilibre fusion-fission contrôle lamorphologie des mitochondries
• A gauche, mutant de la fusion, à droite de la fission• L’équilibre détermine la taille et le nombre de
mitochondries• Chez le sauvage, tube de 0,5 µ de diamètre et 1 à 10 µ
de longueur(Detmer and Chan (2007) Nature Review,8, 870)
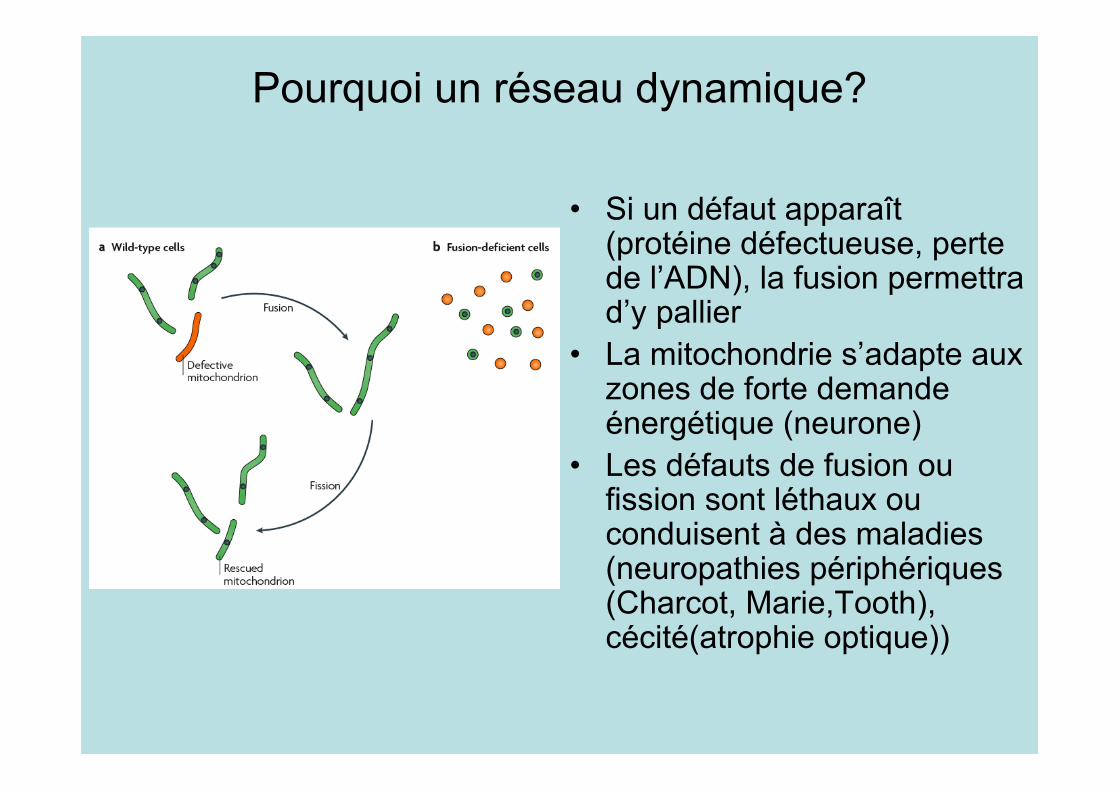
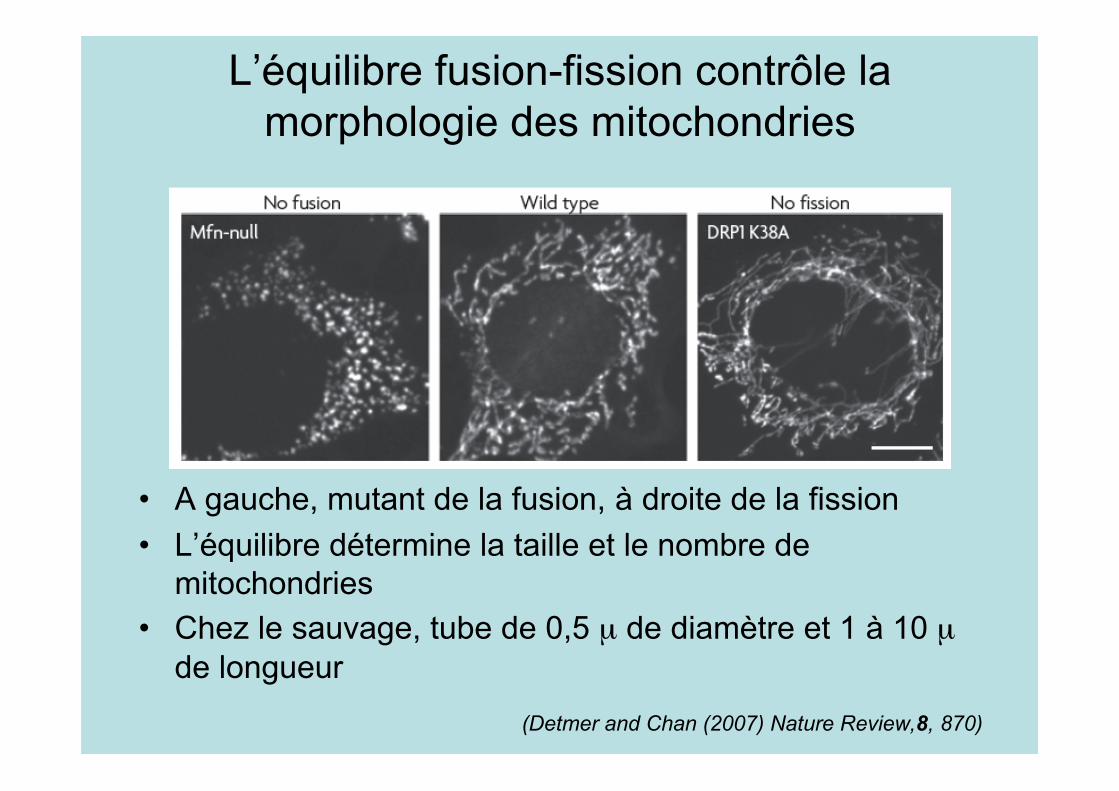
Pourquoi un réseau dynamique?
• Si un défaut apparaît(protéine défectueuse, pertede l’ADN), la fusion permettrad’y pallier
• La mitochondrie s’adapte auxzones de forte demandeénergétique (neurone)
• Les défauts de fusion oufission sont léthaux ouconduisent à des maladies(neuropathies périphériques(Charcot, Marie,Tooth),cécité(atrophie optique))

Mécanisme de la fusion mitochondriale
• La mitochondrie possède deux membranes:– Membrane externe: rôle d’enveloppe, avec des pores
(porines, à structure en tonneau de feuillets β)– Membrane interne: très développée, avec des repliements
(crêtes), étanche aux protons
• La fusion doit s’effectuer sur les deux membranes enconservant à chacune ses propriétés
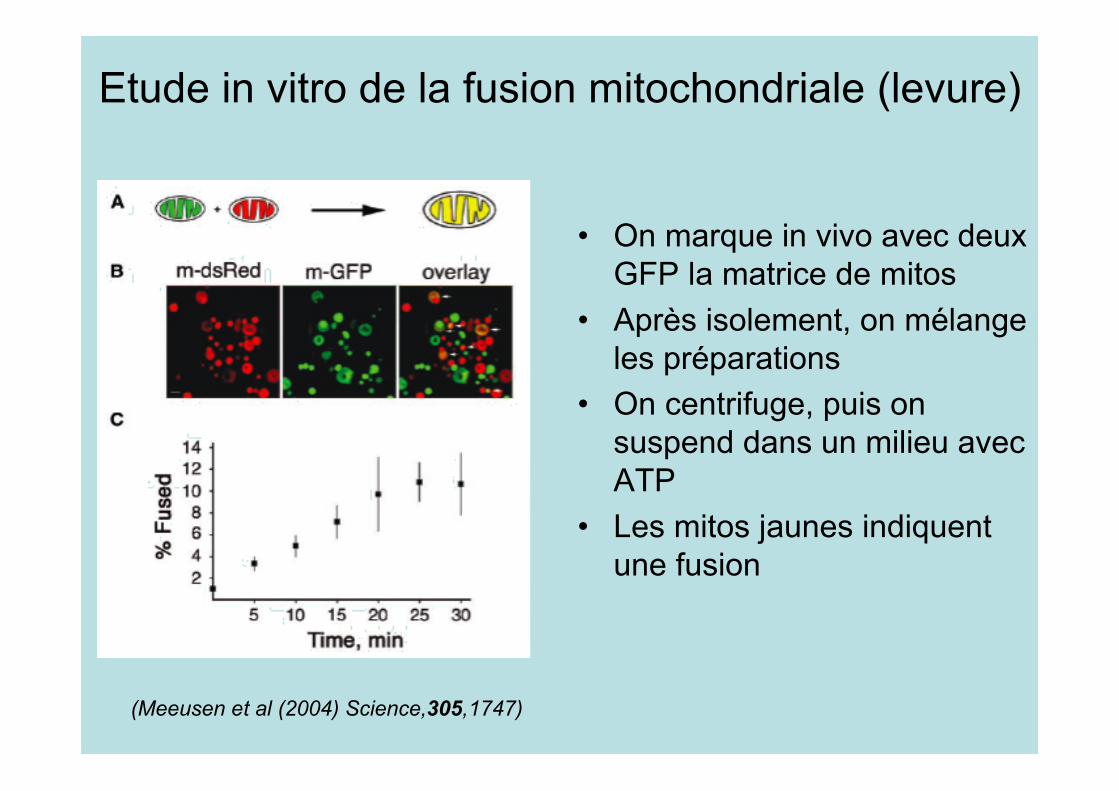
Etude in vitro de la fusion mitochondriale (levure)
• On marque in vivo avec deuxGFP la matrice de mitos
• Après isolement, on mélangeles préparations
• On centrifuge, puis onsuspend dans un milieu avecATP
• Les mitos jaunes indiquentune fusion
(Meeusen et al (2004) Science,305,1747)
Images de fusion mitochondriale
Images à 2 Hz
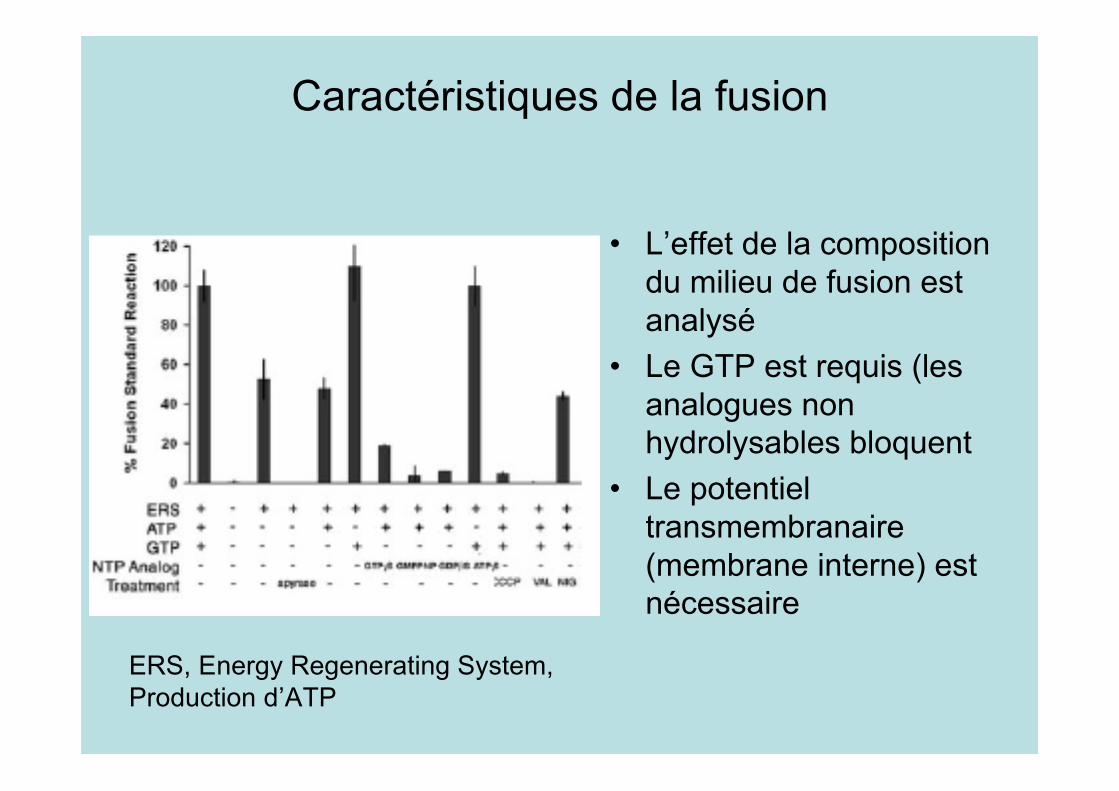
Caractéristiques de la fusion
• L’effet de la compositiondu milieu de fusion estanalysé
• Le GTP est requis (lesanalogues nonhydrolysables bloquent
• Le potentieltransmembranaire(membrane interne) estnécessaire
ERS, Energy Regenerating System,Production d’ATP

Fusion membranes externesmembranes internes
• Dans l’expérience précédente, l’index de la fusion estle mélange de protéines solubles de la matrice(GFP/DsRed)
• Ce protocole révèle la fusion des deux membranes(interne et externe)
• Le même travail décrit aussi une fusion limitée auxmembranes externes, par un processus différent
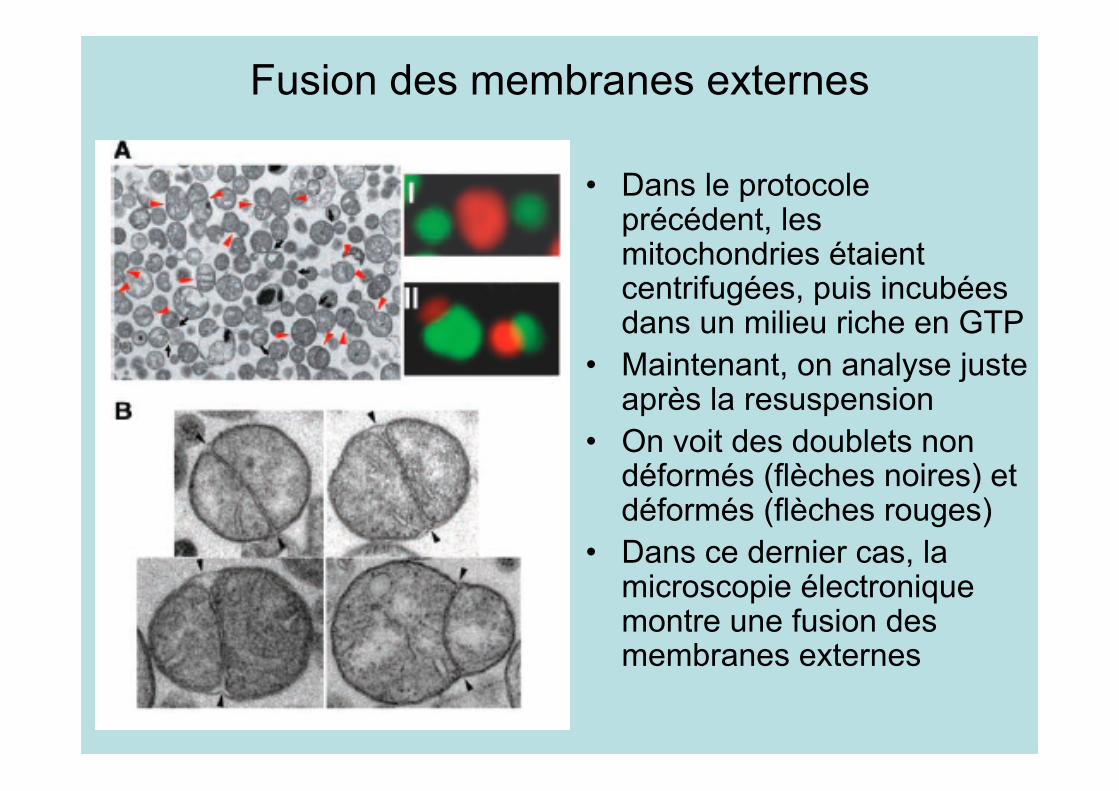
Fusion des membranes externes
• Dans le protocoleprécédent, lesmitochondries étaientcentrifugées, puis incubéesdans un milieu riche en GTP
• Maintenant, on analyse justeaprès la resuspension
• On voit des doublets nondéformés (flèches noires) etdéformés (flèches rouges)
• Dans ce dernier cas, lamicroscopie électroniquemontre une fusion desmembranes externes
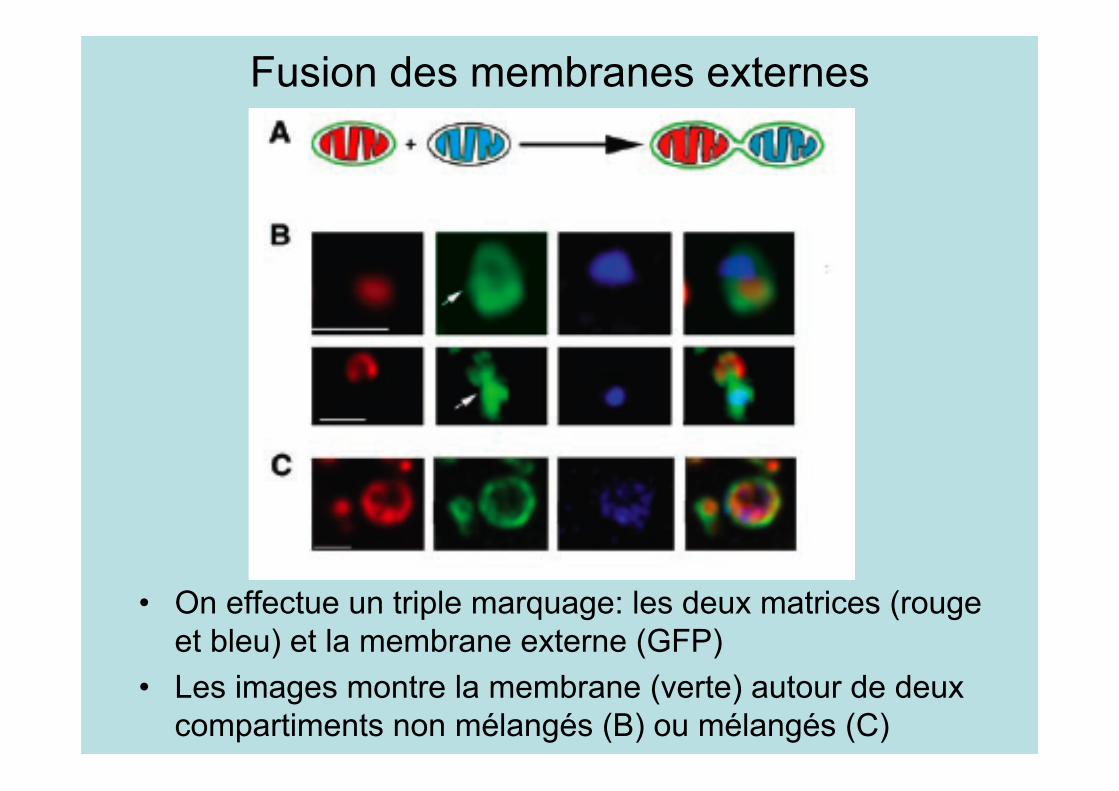
Fusion des membranes externes
• On effectue un triple marquage: les deux matrices (rougeet bleu) et la membrane externe (GFP)
• Les images montre la membrane (verte) autour de deuxcompartiments non mélangés (B) ou mélangés (C)

Les deux membranes fusionnentindépendamment
• Fusions indépendantes, mais co-ordonnées: dans lesconditions cellulaires, on ne voit pas de fusionslimitées à la membrane externe
• Le test permet de déterminer les composantsnécessaires à la fusion des membranes externes
• Ces composants ne sont pas ceux nécessaires à lafusion des membranes internes: différence dans lesconcentrations de GTP, ΔpH plus important que ΔΨ
• Les systèmes de fusion sont différents
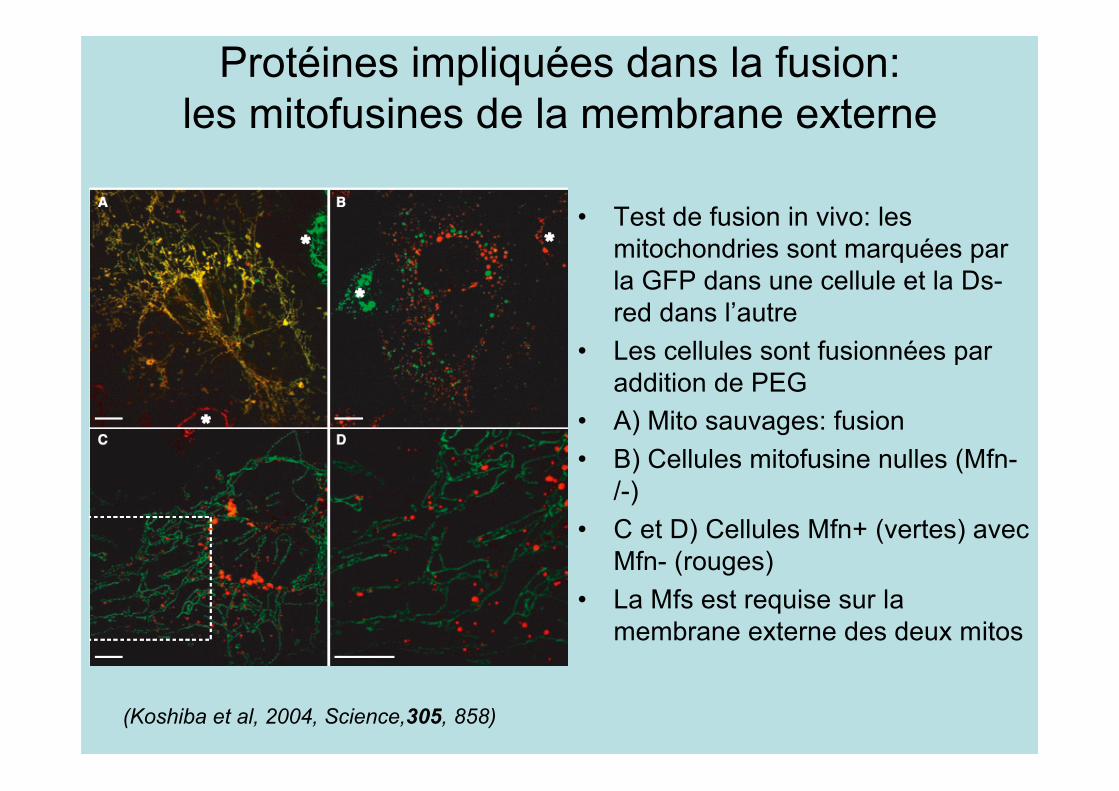
Protéines impliquées dans la fusion:les mitofusines de la membrane externe
• Test de fusion in vivo: lesmitochondries sont marquées parla GFP dans une cellule et la Ds-red dans l’autre
• Les cellules sont fusionnées paraddition de PEG
• A) Mito sauvages: fusion• B) Cellules mitofusine nulles (Mfn-
/-)• C et D) Cellules Mfn+ (vertes) avec
Mfn- (rouges)• La Mfs est requise sur la
membrane externe des deux mitos
(Koshiba et al, 2004, Science,305, 858)
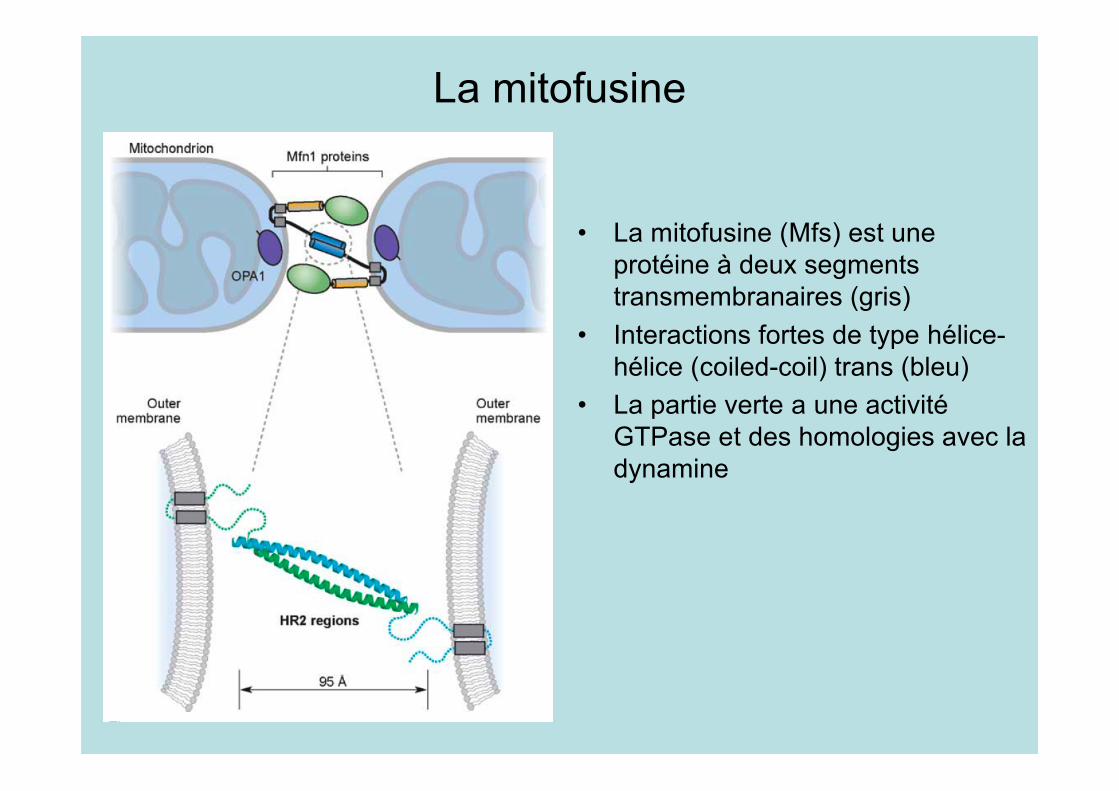
La mitofusine
• La mitofusine (Mfs) est uneprotéine à deux segmentstransmembranaires (gris)
• Interactions fortes de type hélice-hélice (coiled-coil) trans (bleu)
• La partie verte a une activitéGTPase et des homologies avec ladynamine
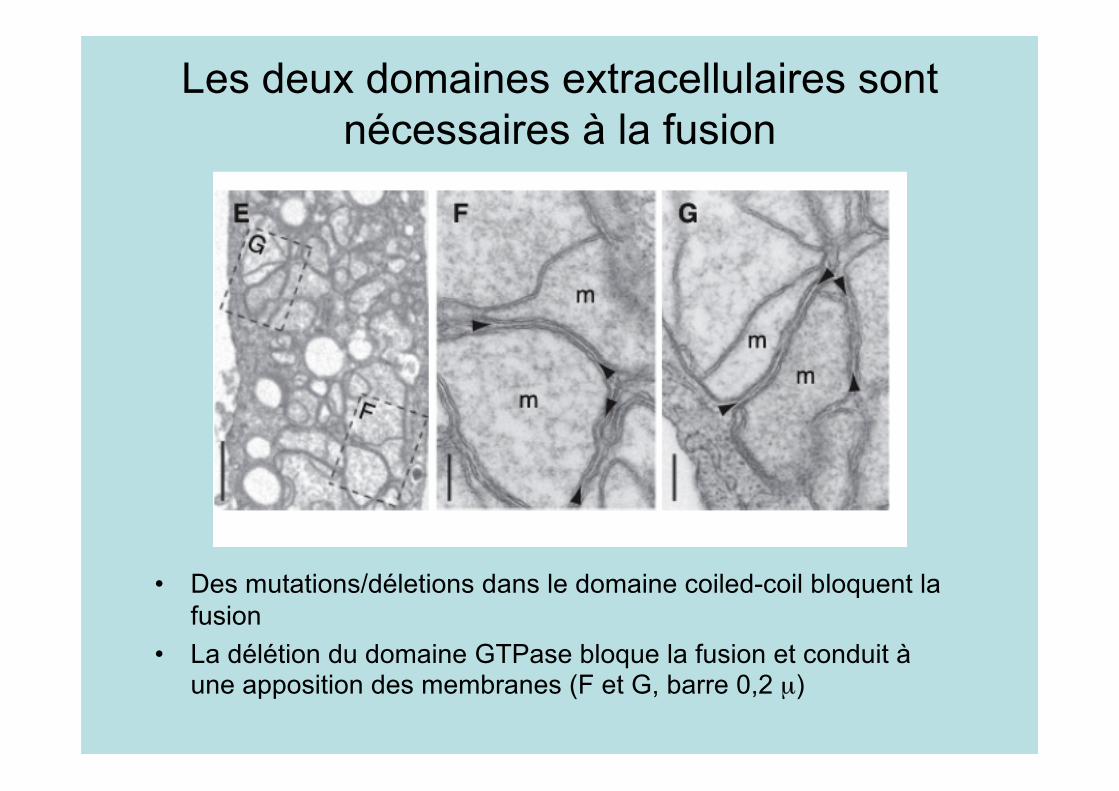
Les deux domaines extracellulaires sontnécessaires à la fusion
• Des mutations/déletions dans le domaine coiled-coil bloquent lafusion
• La délétion du domaine GTPase bloque la fusion et conduit àune apposition des membranes (F et G, barre 0,2 µ)
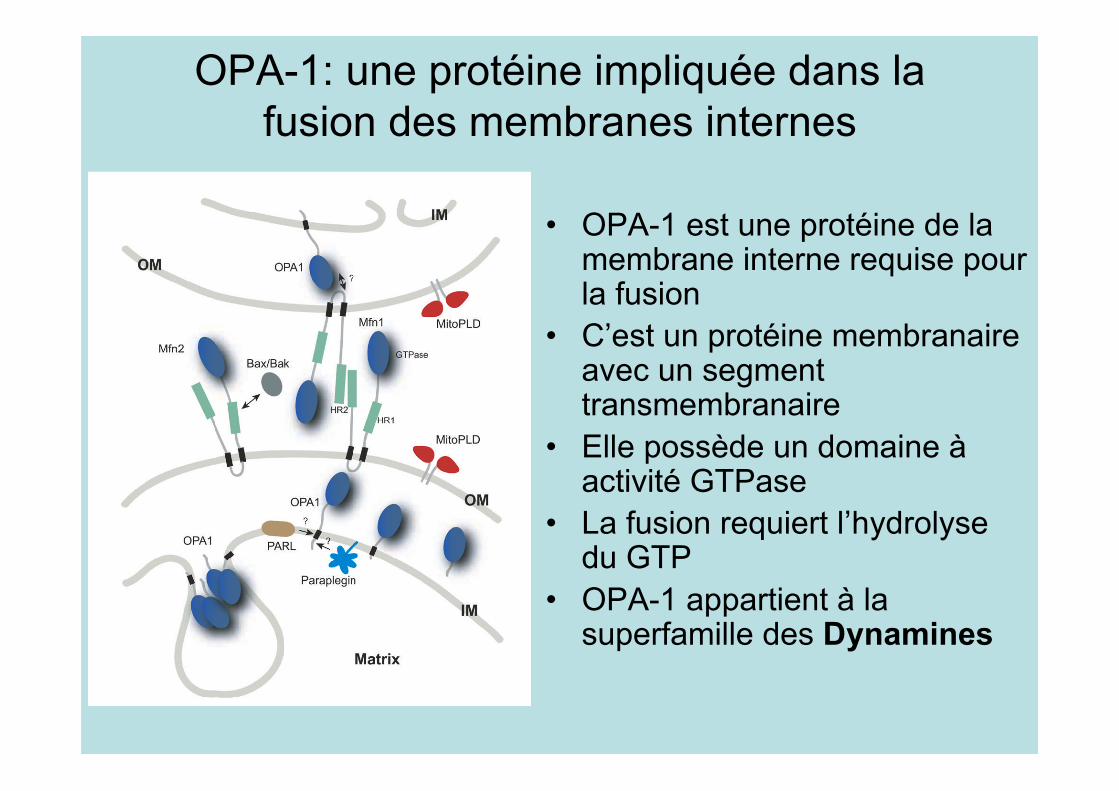
OPA-1: une protéine impliquée dans lafusion des membranes internes
• OPA-1 est une protéine de lamembrane interne requise pourla fusion
• C’est un protéine membranaireavec un segmenttransmembranaire
• Elle possède un domaine àactivité GTPase
• La fusion requiert l’hydrolysedu GTP
• OPA-1 appartient à lasuperfamille des Dynamines
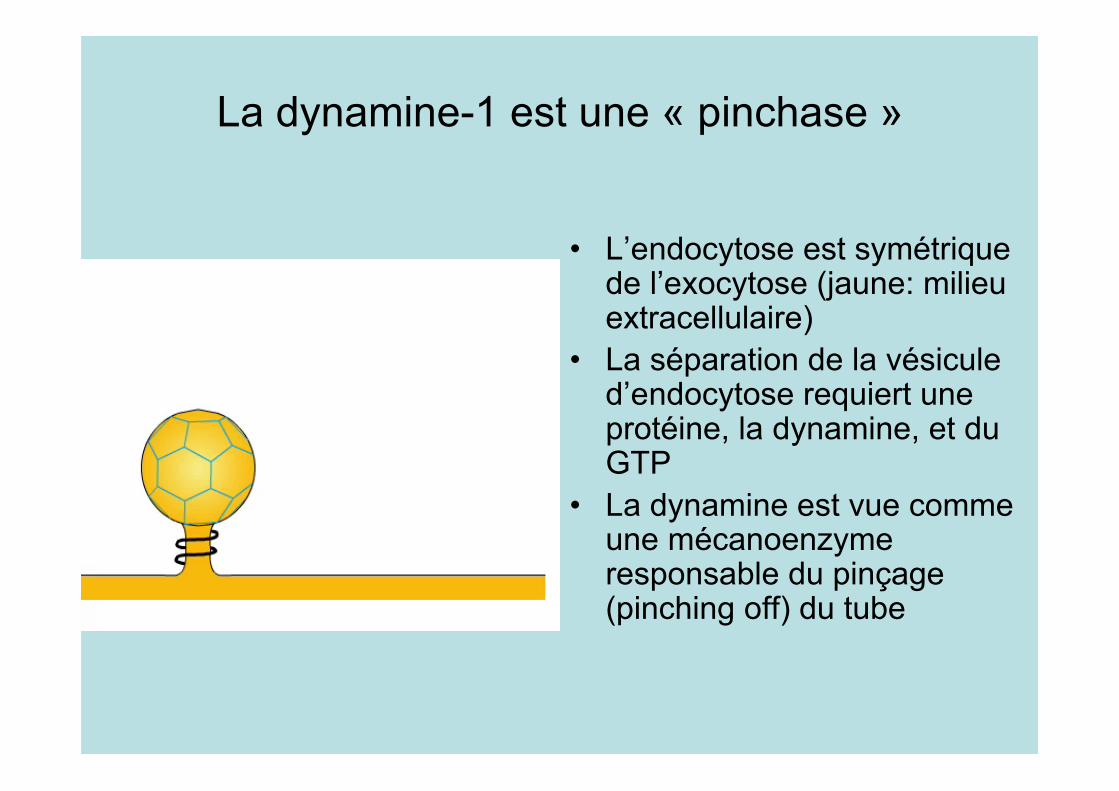
La dynamine-1 est une « pinchase »
• L’endocytose est symétriquede l’exocytose (jaune: milieuextracellulaire)
• La séparation de la vésiculed’endocytose requiert uneprotéine, la dynamine, et duGTP
• La dynamine est vue commeune mécanoenzymeresponsable du pinçage(pinching off) du tube
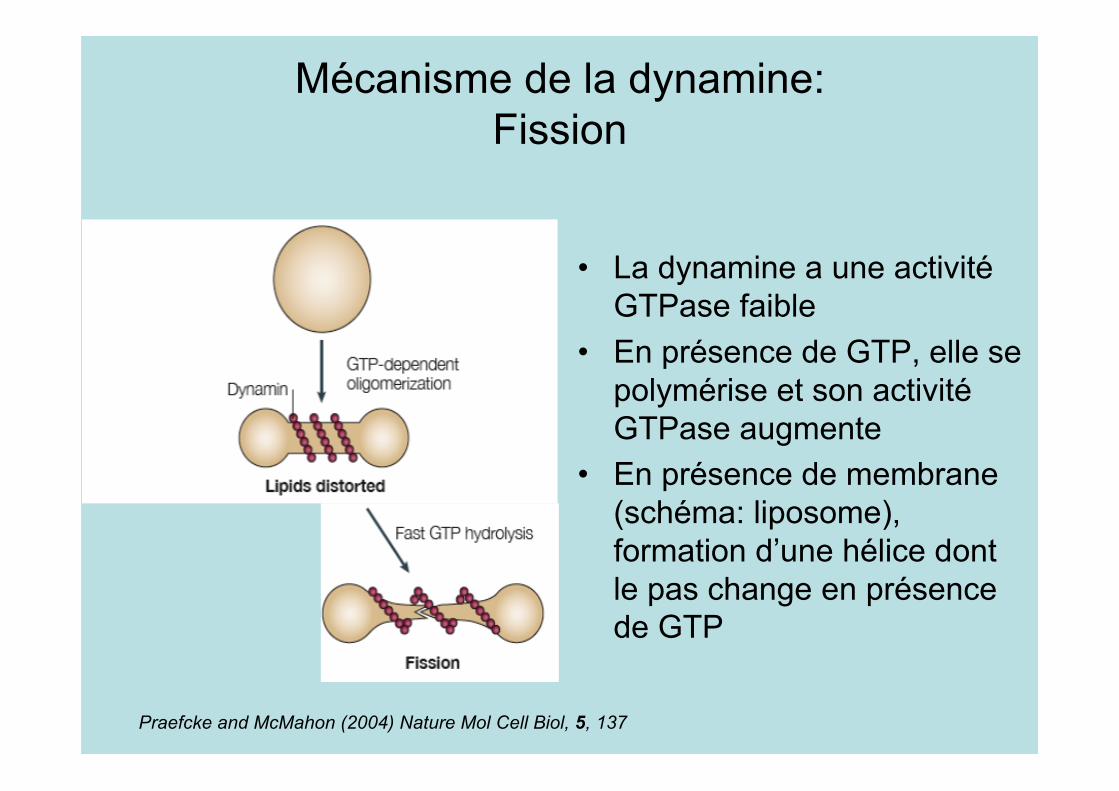
Mécanisme de la dynamine:Fission
• La dynamine a une activitéGTPase faible
• En présence de GTP, elle sepolymérise et son activitéGTPase augmente
• En présence de membrane(schéma: liposome),formation d’une hélice dontle pas change en présencede GTP
Praefcke and McMahon (2004) Nature Mol Cell Biol, 5, 137
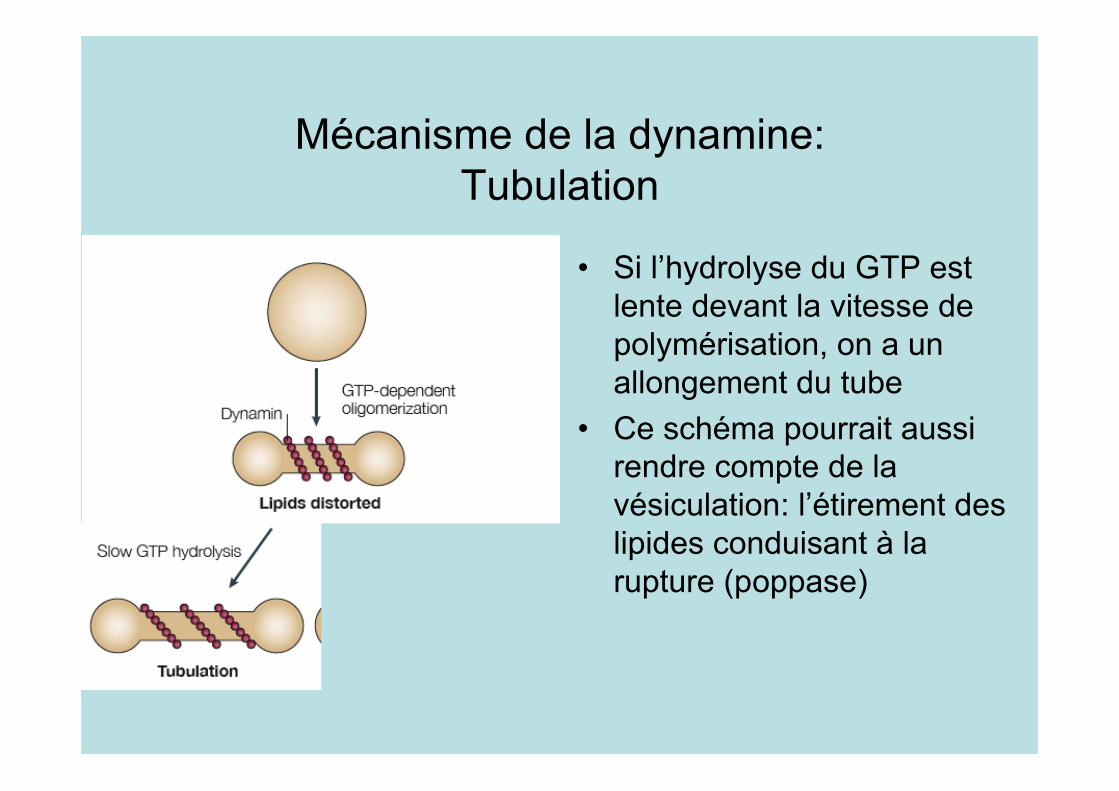
Mécanisme de la dynamine:Tubulation
• Si l’hydrolyse du GTP estlente devant la vitesse depolymérisation, on a unallongement du tube
• Ce schéma pourrait aussirendre compte de lavésiculation: l’étirement deslipides conduisant à larupture (poppase)
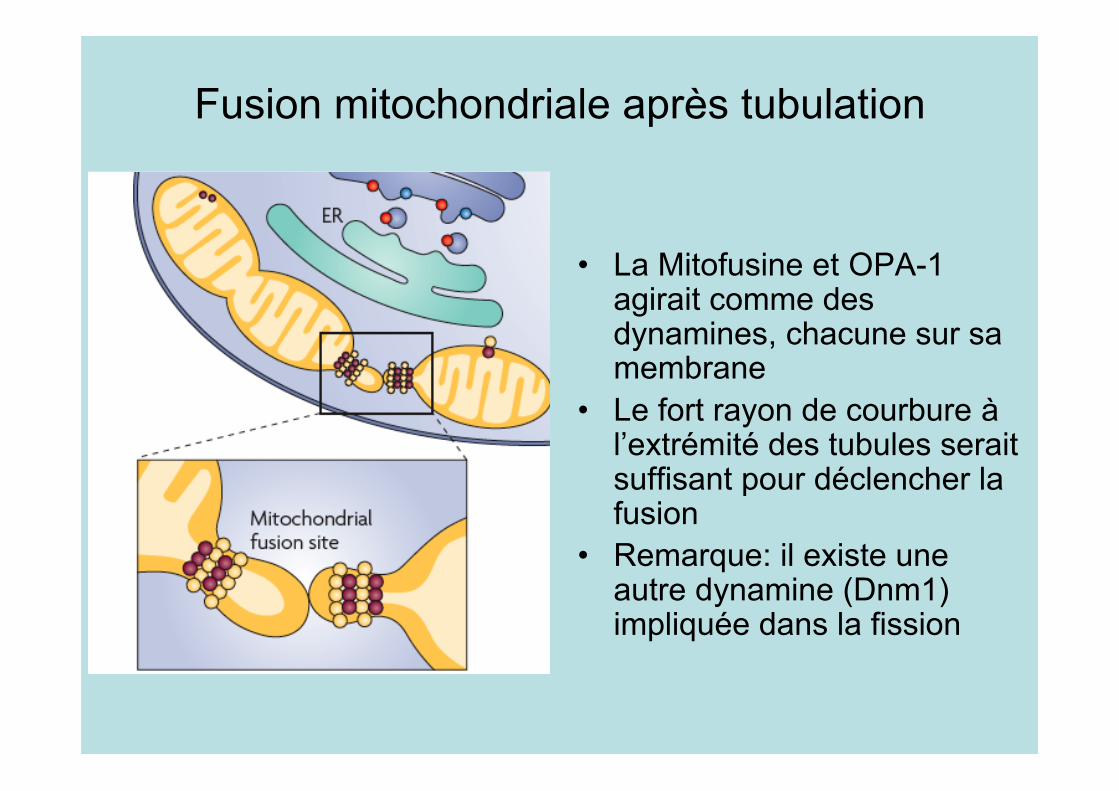
Fusion mitochondriale après tubulation
• La Mitofusine et OPA-1agirait comme desdynamines, chacune sur samembrane
• Le fort rayon de courbure àl’extrémité des tubules seraitsuffisant pour déclencher lafusion
• Remarque: il existe uneautre dynamine (Dnm1)impliquée dans la fission

Conclusions
• Les fusions mitochondriales sont intéressantes etcomplexes: deux systèmes indépendants, maiscoordonnés
• Les membranes possèdent un système dereconnaissance homotypique : la mitofusine
• La GTPase de la mitofusine pourrait soit induire unchangement de conformation de type fusion virale,soit être une » dynamine » mécanosensible
• Sur la membrane interne, OPA-1 est une dynamine;elle pourrait induire une tubulation et la fusion par lesextrémités des tubules

Conclusions (2)
• L’étude des fusions cellule-cellule et mitochondrialesmontrent que, dans la majorité des cas, on ne metpas en évidence de « protéines fusogènes »,analogues aux protéines virales ou aux SNARE
• Ces protéines pourraient être à découvrir• Une autre hypothèse est que la fusion impliquerait
des protéines de reconnaissance (non fusogènes) etune déformation de la membrane (ostéoclastes,myoblastes de mouche, mitochondries)
• Que sait-on des déformations de membrane?
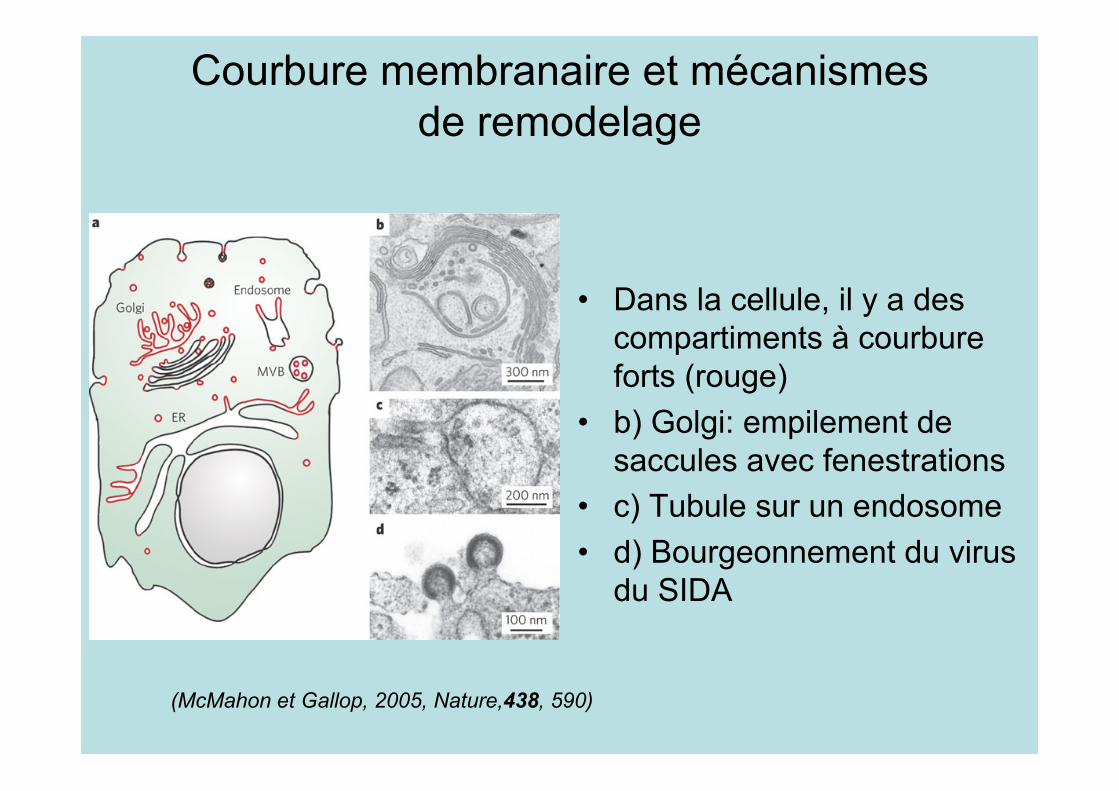
Courbure membranaire et mécanismesde remodelage
• Dans la cellule, il y a descompartiments à courbureforts (rouge)
• b) Golgi: empilement desaccules avec fenestrations
• c) Tubule sur un endosome• d) Bourgeonnement du virus
du SIDA
(McMahon et Gallop, 2005, Nature,438, 590)
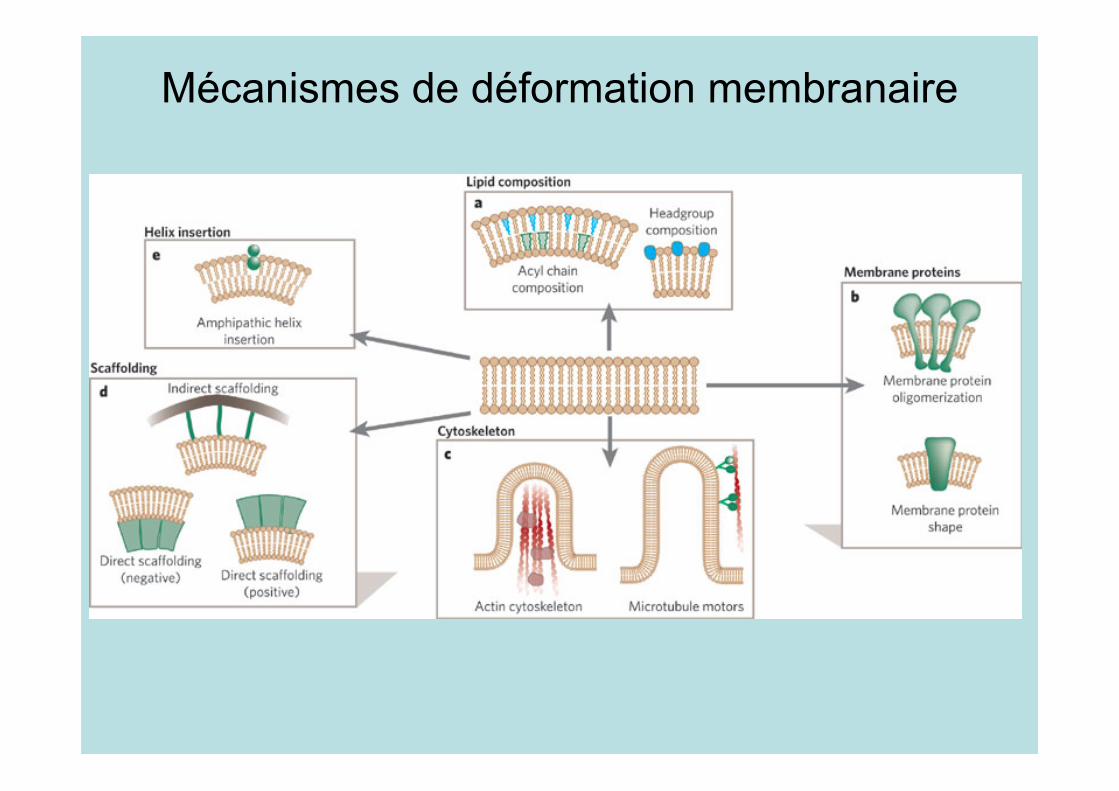
Mécanismes de déformation membranaire
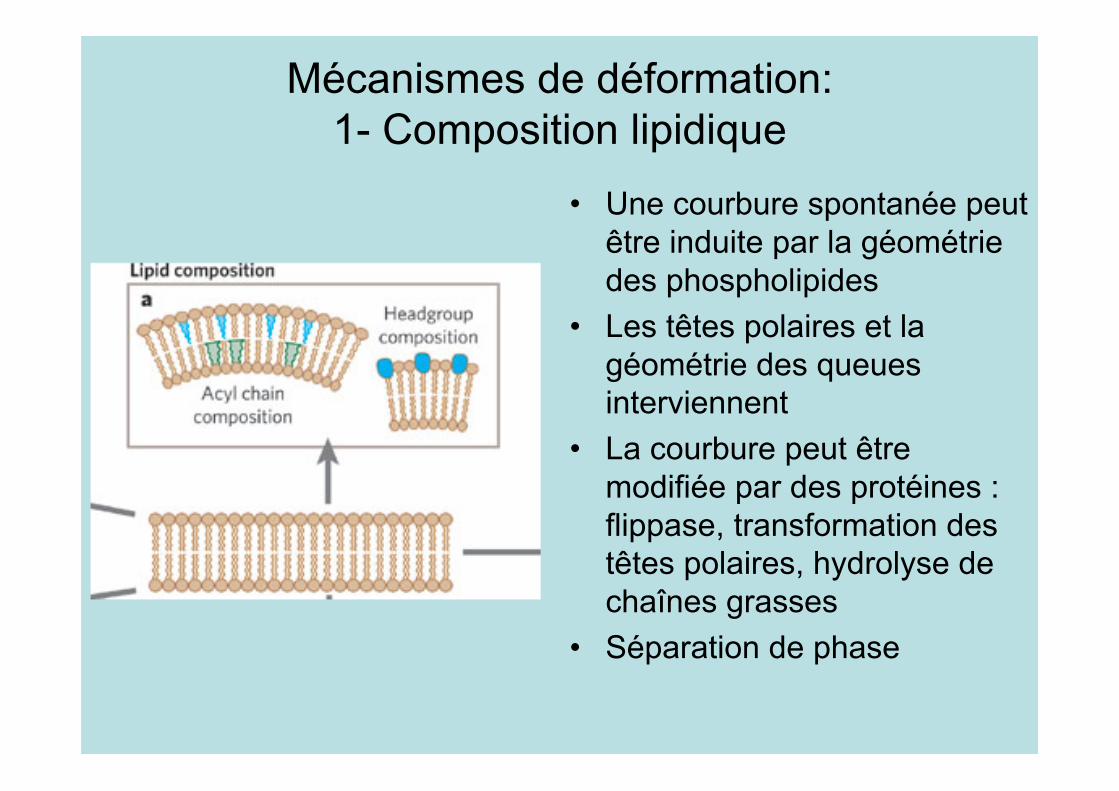
Mécanismes de déformation:1- Composition lipidique
• Une courbure spontanée peutêtre induite par la géométriedes phospholipides
• Les têtes polaires et lagéométrie des queuesinterviennent
• La courbure peut êtremodifiée par des protéines :flippase, transformation destêtes polaires, hydrolyse dechaînes grasses
• Séparation de phase
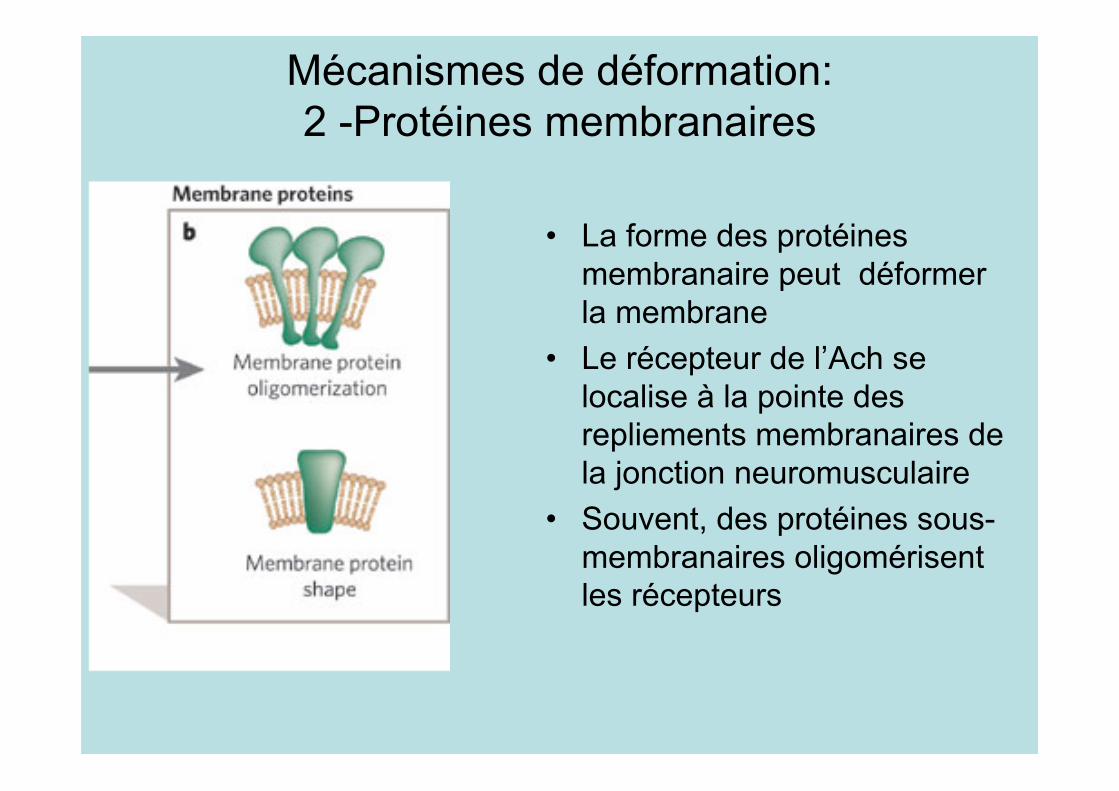
Mécanismes de déformation:2 -Protéines membranaires
• La forme des protéinesmembranaire peut déformerla membrane
• Le récepteur de l’Ach selocalise à la pointe desrepliements membranaires dela jonction neuromusculaire
• Souvent, des protéines sous-membranaires oligomérisentles récepteurs
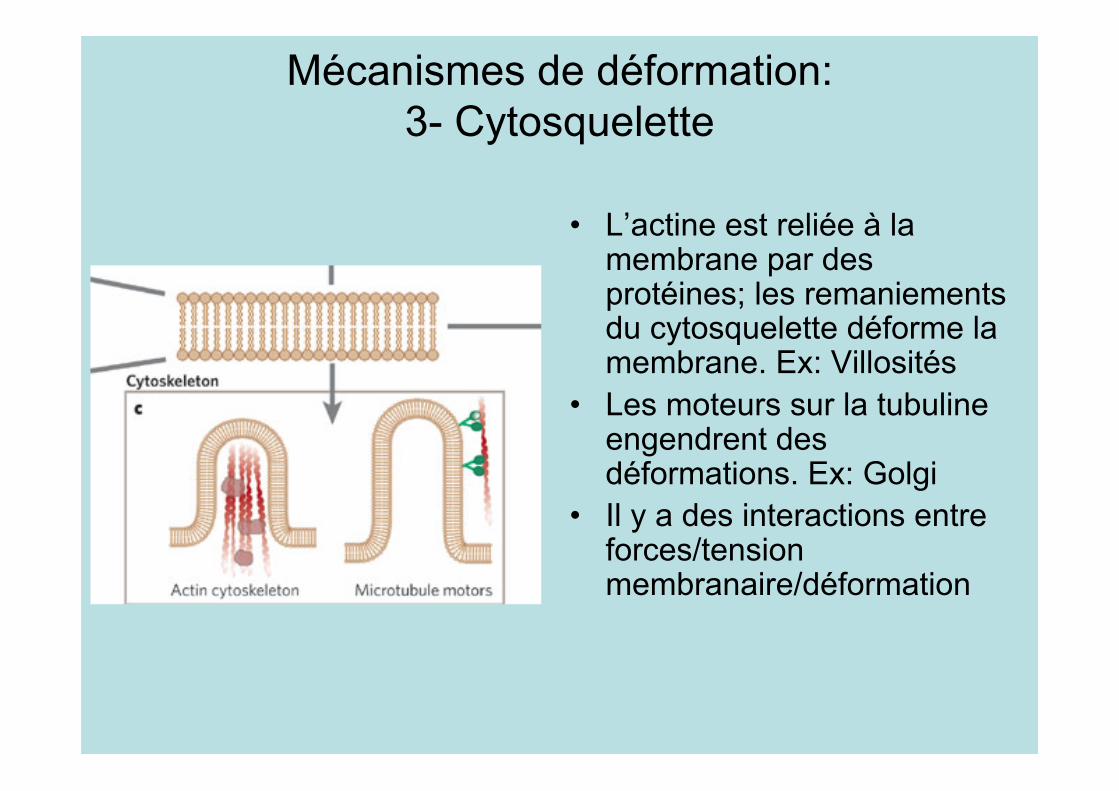
Mécanismes de déformation:3- Cytosquelette
• L’actine est reliée à lamembrane par desprotéines; les remaniementsdu cytosquelette déforme lamembrane. Ex: Villosités
• Les moteurs sur la tubulineengendrent desdéformations. Ex: Golgi
• Il y a des interactions entreforces/tensionmembranaire/déformation
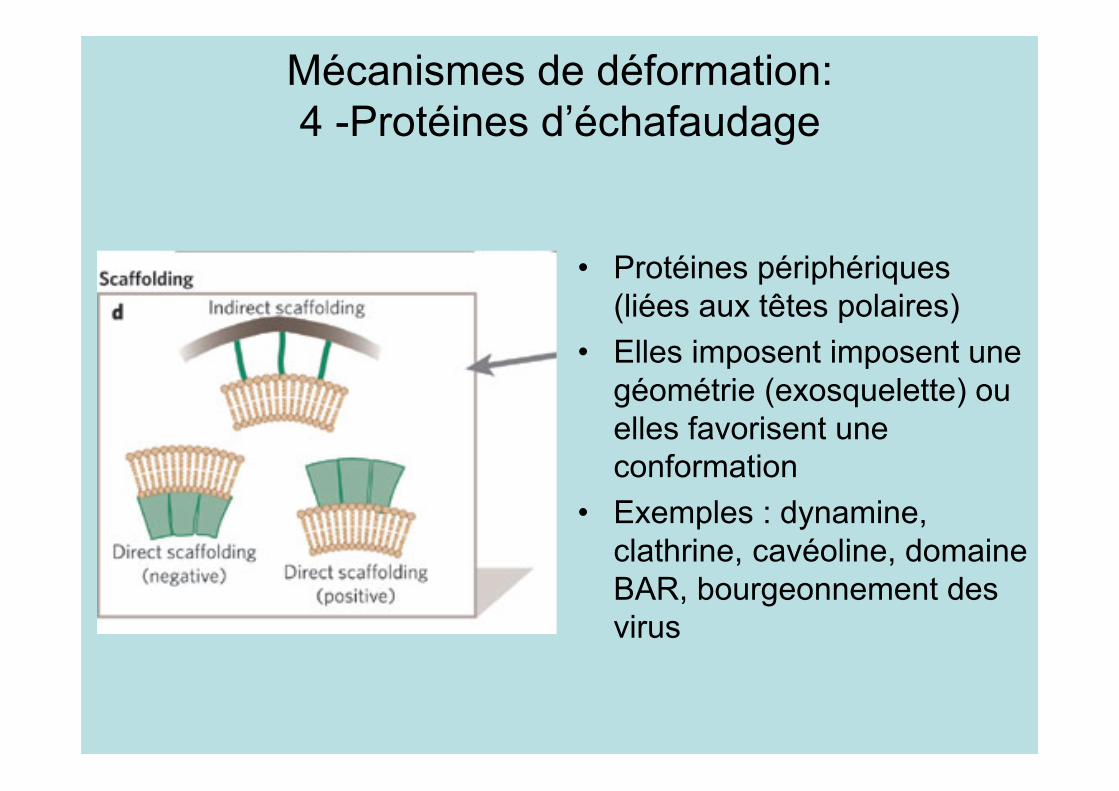
Mécanismes de déformation:4 -Protéines d’échafaudage
• Protéines périphériques(liées aux têtes polaires)
• Elles imposent imposent unegéométrie (exosquelette) ouelles favorisent uneconformation
• Exemples : dynamine,clathrine, cavéoline, domaineBAR, bourgeonnement desvirus
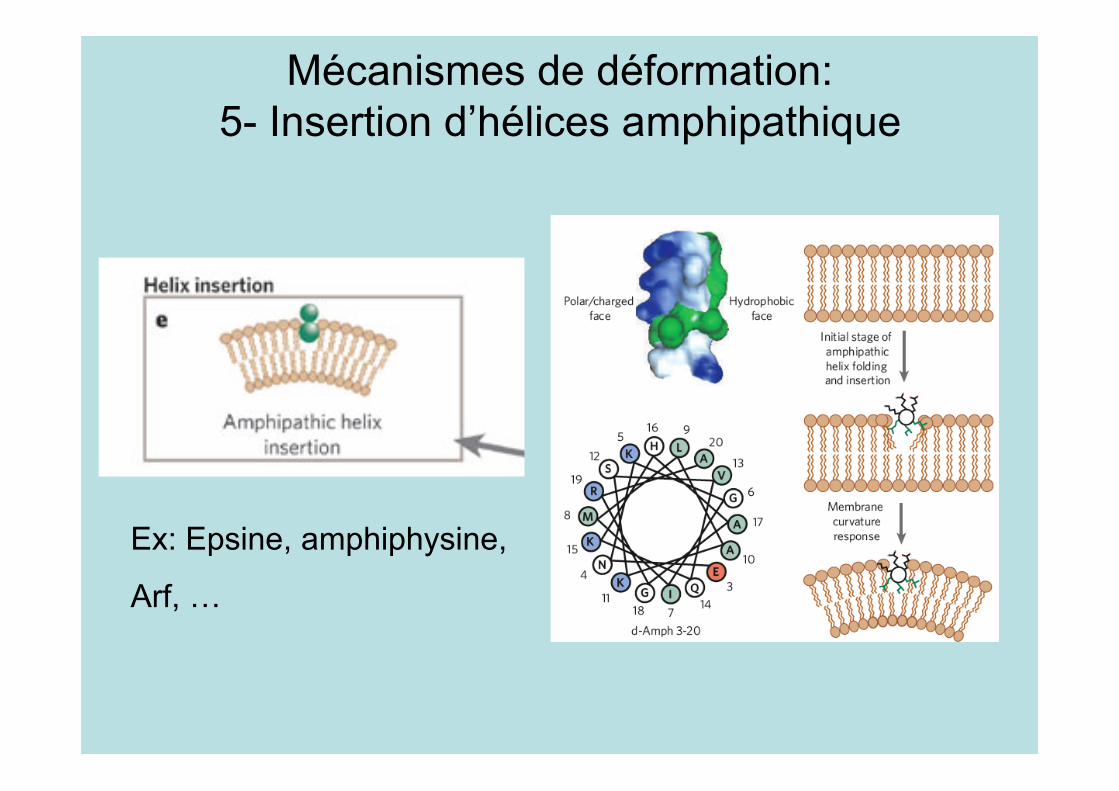
Mécanismes de déformation:5- Insertion d’hélices amphipathique
Ex: Epsine, amphiphysine,
Arf, …
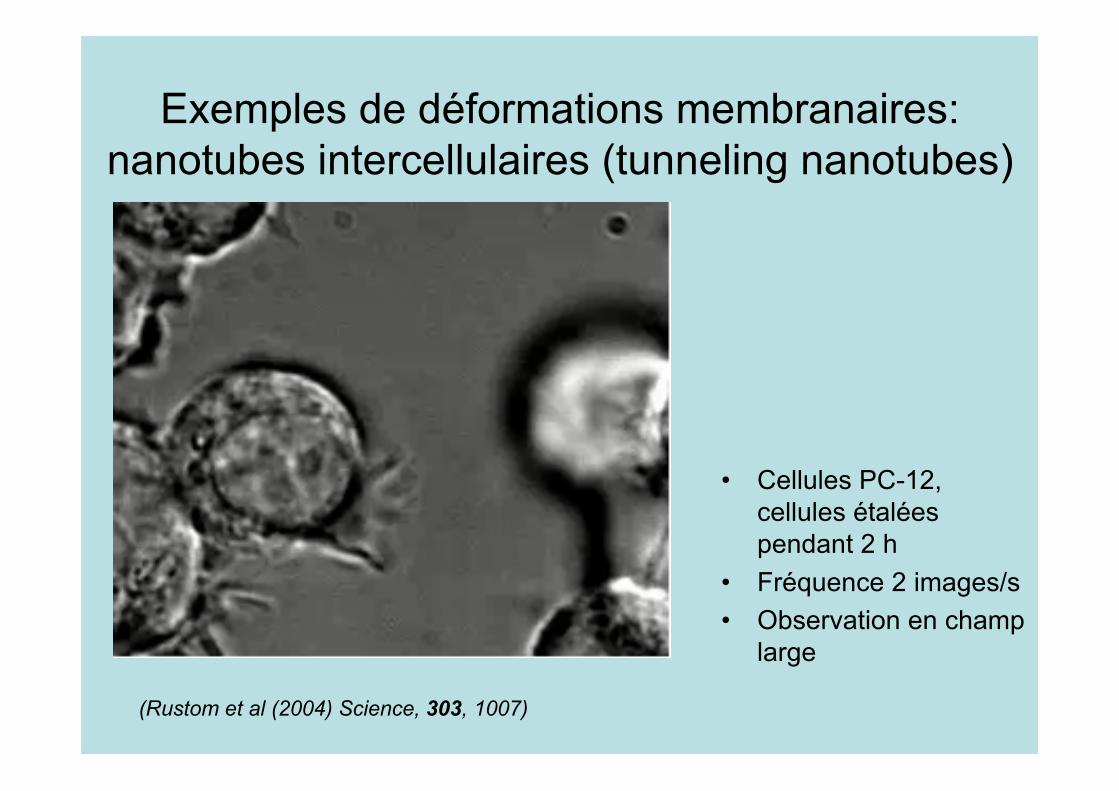
Exemples de déformations membranaires:nanotubes intercellulaires (tunneling nanotubes)
• Cellules PC-12,cellules étaléespendant 2 h
• Fréquence 2 images/s• Observation en champ
large
(Rustom et al (2004) Science, 303, 1007)
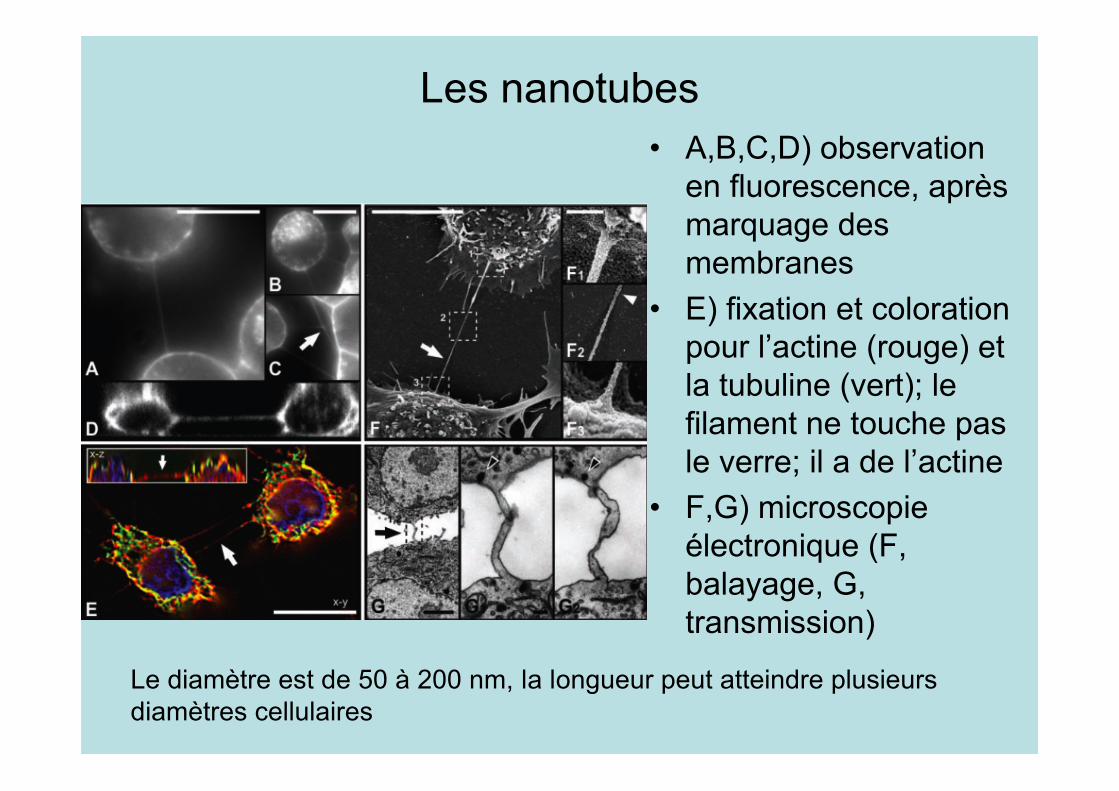
Les nanotubes• A,B,C,D) observation
en fluorescence, aprèsmarquage desmembranes
• E) fixation et colorationpour l’actine (rouge) etla tubuline (vert); lefilament ne touche pasle verre; il a de l’actine
• F,G) microscopieélectronique (F,balayage, G,transmission)
Le diamètre est de 50 à 200 nm, la longueur peut atteindre plusieursdiamètres cellulaires

Les nanotubes peuvent fusionner avec lacellule cible
• Images en microscopieélectronique àtransmission
• A) fusion avec la cible• C1) extrémité fermée• Hypothèse: la courbure
très forte de l’extrémitédu tube favorise lafusion
(Gerdes and Carvalho, 2008, Curr Op Cell Biol, 20, 470)
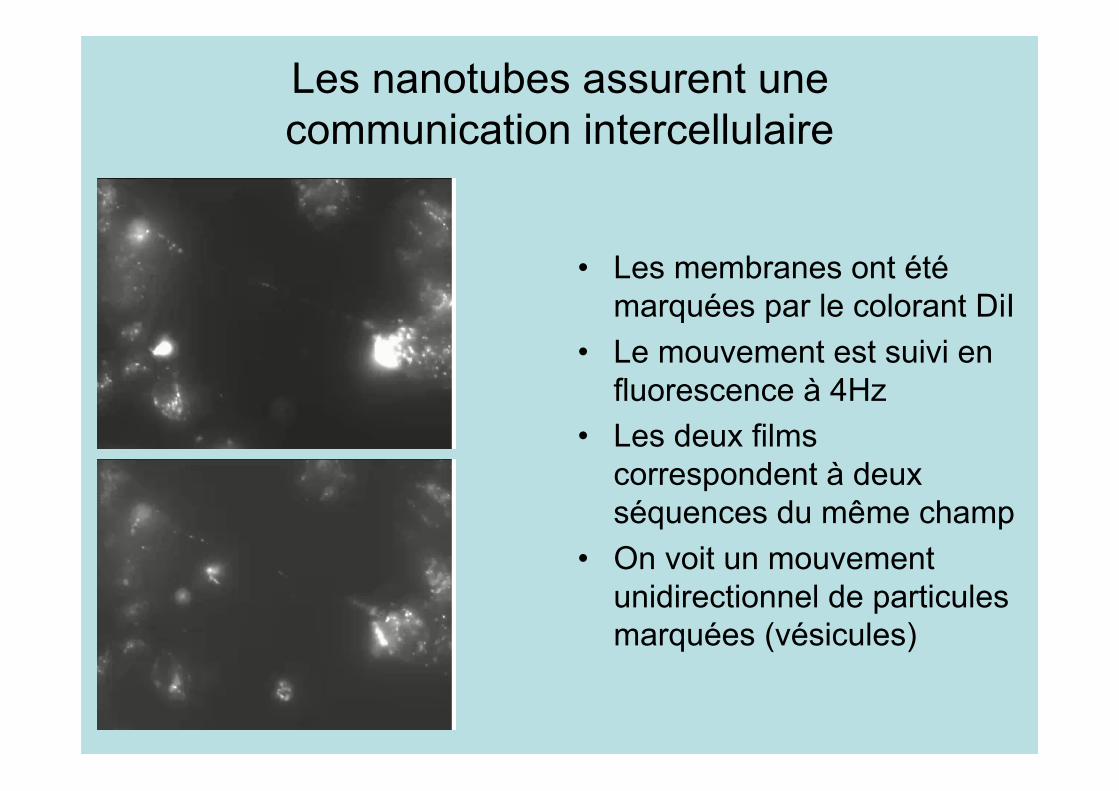
Les nanotubes assurent unecommunication intercellulaire
• Les membranes ont étémarquées par le colorant DiI
• Le mouvement est suivi enfluorescence à 4Hz
• Les deux filmscorrespondent à deuxséquences du même champ
• On voit un mouvementunidirectionnel de particulesmarquées (vésicules)
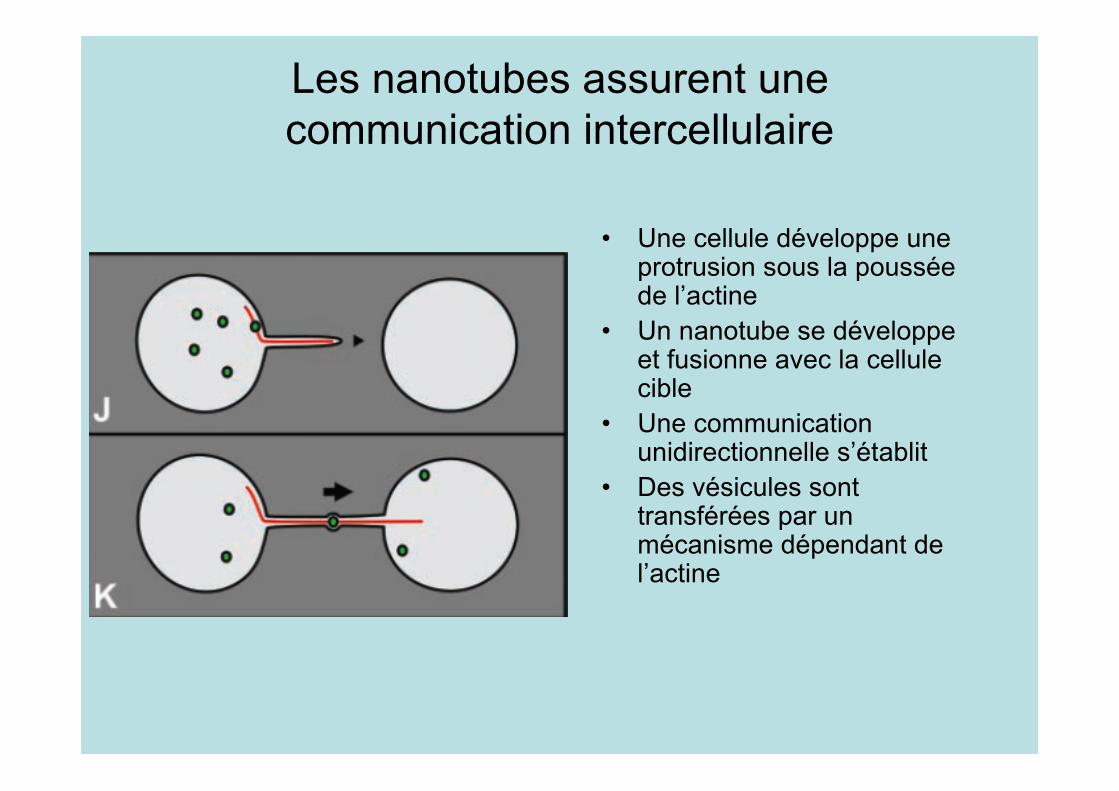
Les nanotubes assurent unecommunication intercellulaire
• Une cellule développe uneprotrusion sous la pousséede l’actine
• Un nanotube se développeet fusionne avec la cellulecible
• Une communicationunidirectionnelle s’établit
• Des vésicules sonttransférées par unmécanisme dépendant del’actine
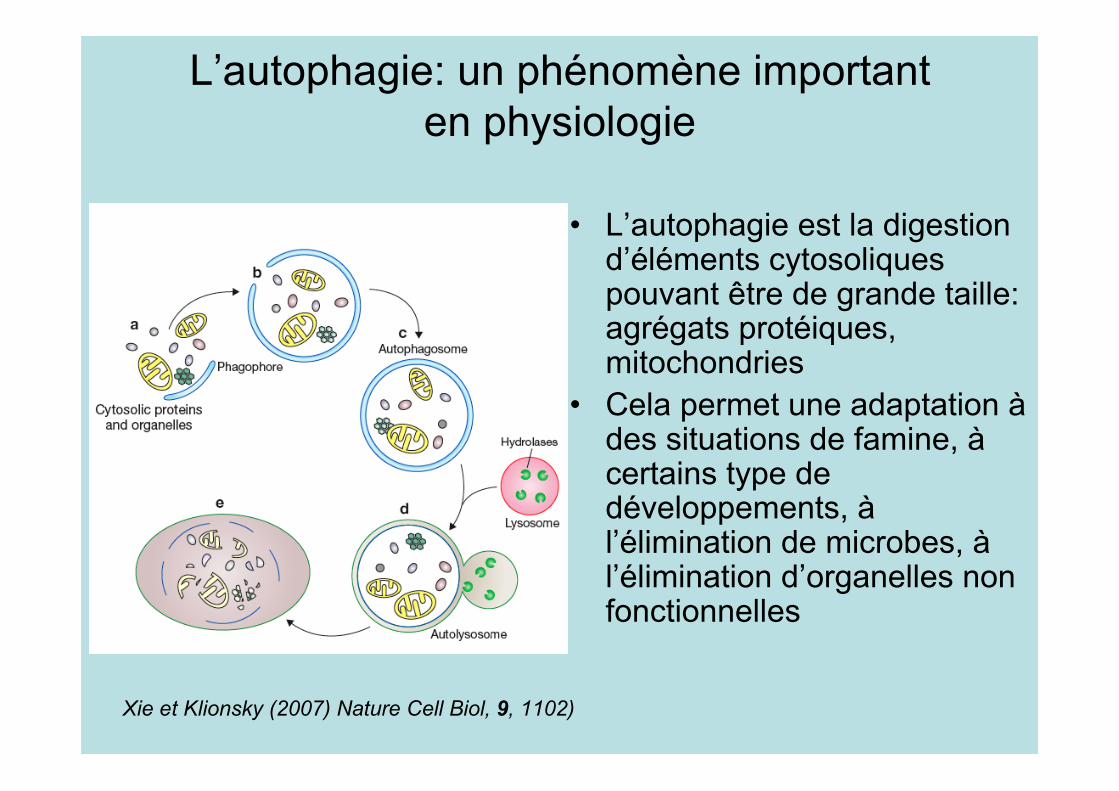
L’autophagie: un phénomène importanten physiologie
• L’autophagie est la digestiond’éléments cytosoliquespouvant être de grande taille:agrégats protéiques,mitochondries
• Cela permet une adaptation àdes situations de famine, àcertains type dedéveloppements, àl’élimination de microbes, àl’élimination d’organelles nonfonctionnelles
Xie et Klionsky (2007) Nature Cell Biol, 9, 1102)
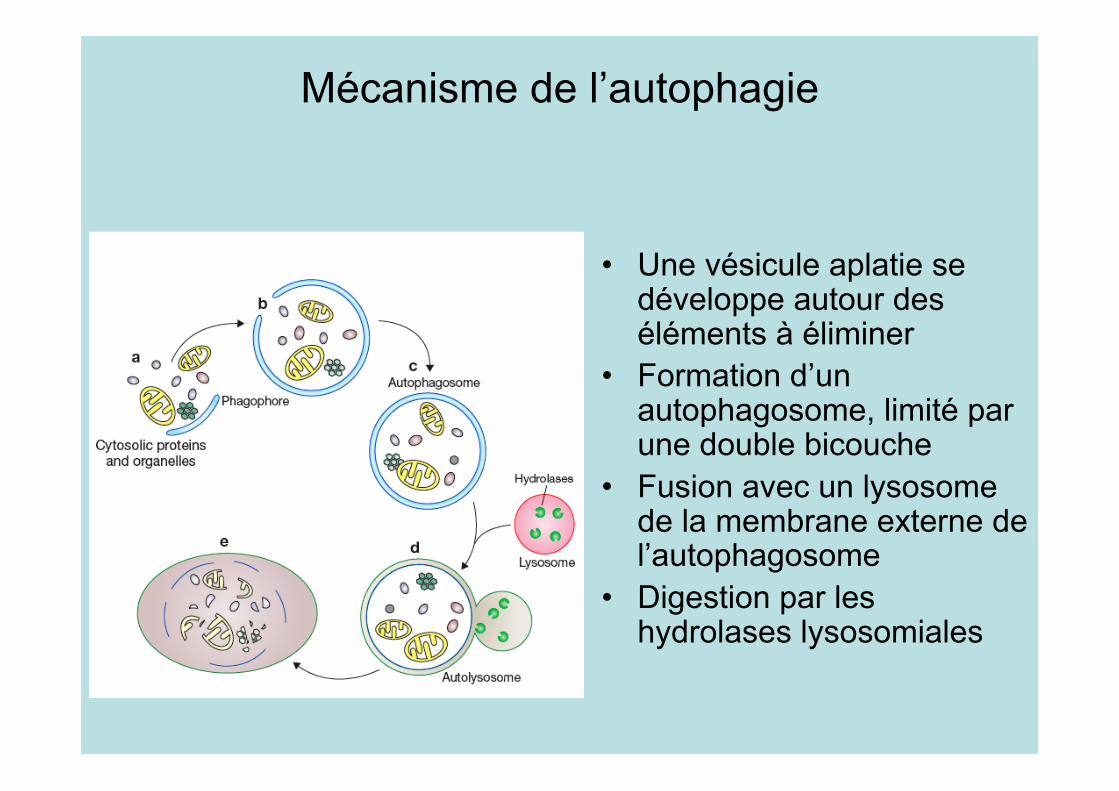
Mécanisme de l’autophagie
• Une vésicule aplatie sedéveloppe autour deséléments à éliminer
• Formation d’unautophagosome, limité parune double bicouche
• Fusion avec un lysosomede la membrane externe del’autophagosome
• Digestion par leshydrolases lysosomiales
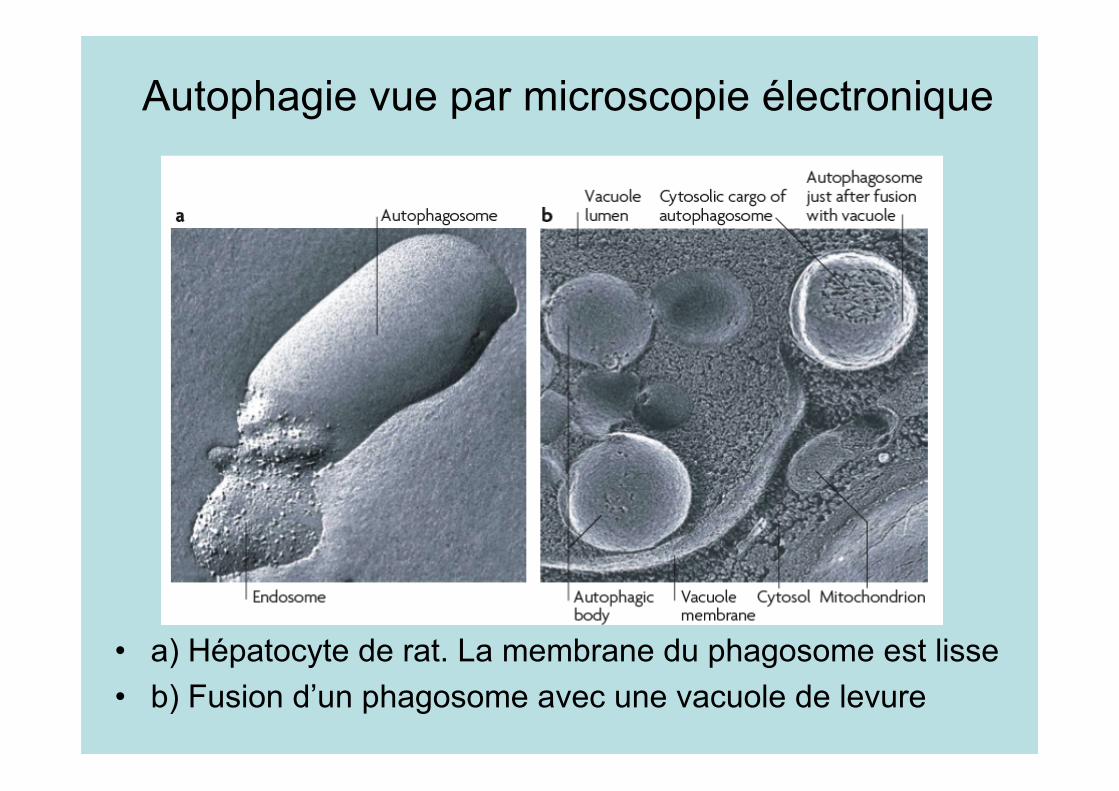
Autophagie vue par microscopie électronique
• a) Hépatocyte de rat. La membrane du phagosome est lisse• b) Fusion d’un phagosome avec une vacuole de levure
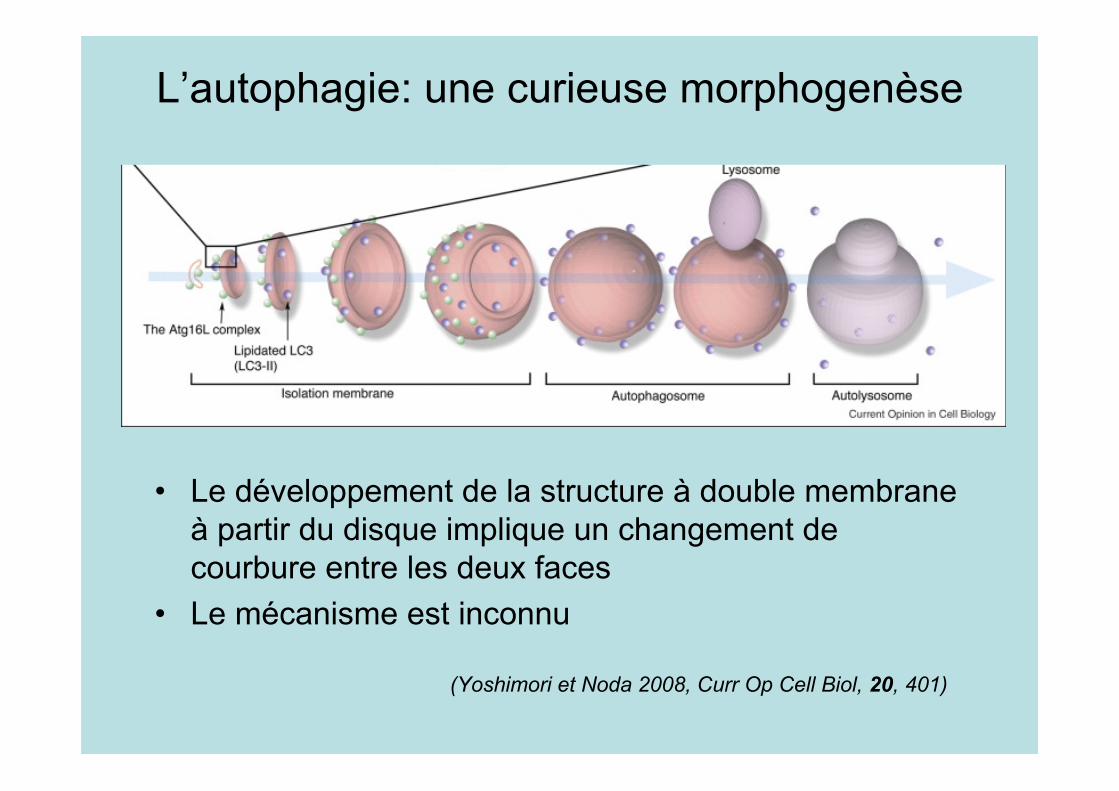
L’autophagie: une curieuse morphogenèse
• Le développement de la structure à double membraneà partir du disque implique un changement decourbure entre les deux faces
• Le mécanisme est inconnu
(Yoshimori et Noda 2008, Curr Op Cell Biol, 20, 401)

Conclusions
• Si la cellule comporte des compartiments quipermettent d’effectuer simultanément des opérationsdifférentes, cette compartimentation est dynamique
• Elle implique de nombreuses fusions membranaires,ainsi que des fissions; des protéines participent à cesétapes
• Mais, l’ensemble de la dynamique membranaire estloin d’être exploré
• Cette exploration laisse une large place à la physique