Embed Size (px)
Citation preview

Plate-forme de Kaw
(Aquarelle T. Deville)
D. Guiral
IRD Cayenne
Mars Avril 2003
'1>., : ....<.
'-:".-;..:>., ';':' .• ,".

1. Introduction
Une plate-fonne a été déposée par l'IRD dans le marais de Kaw par hélicoptère le 4 décembre2001. Cette opération a fait l'objet d'un rapport transmis le 17/1212001 conformément à la clause 3 del'arrêté 1183 IDI1B/ENV du 20 juillet 2001 autorisant l'IRD à installer une plate-forme de recherchedans la Réserve Naturelle de Kaw-Roura. Cette première mission avait été consacrée:
à l'installation et au montage de la plate-fonneà l'implantation d'une station météorologique et d'un limnigrapheà une première estimation de la densité de caïmans réalisée par B. de Thoisy (AssociationKwata)
Au cours de l'année 2002, au total 9 missions ont été réaliséesDu 16/02 au 18/02
Caractérisation de la végétation par JJ de GranviJIe, JM Denhin et F. Crozier (lRDCayenne)
Etude physico-chimique des eaux par D. Guiral (IRD Cayenne)Prélèvements pour l'étude des communautés phyto- et zoo-planctoniques (échantillons
expédiés à l'université Montpellier 2)Du 30/3 au 3/4
Etude de l'avifaune par O. Tostain (Conservateur Réserve du Grand Connétable)Etude physico-chimique des eaux par D. Guiral (!RD Cayenne)Prélèvements pour l'étude des communautés phyto et zoo-planctoniques (échantillons
expédiés à l'université Montpellier 2).Du 12/05 au 15/05
Etude de l'avifaune par P. Reynaud (IRD Dakar) et T. Deville (Ecobios)Etude physico-chimique des eaux par D. Guiral (IRD Cayenne)Prélèvements pour l'étude des communautés phyto- et zoo-planctoniques (échantillons
expédiés à l'université Montpellier 2).Du 1 au 6/06
Etude de l'avifaune par T. Deville (Ecobios)Du 8 au14/06
Etude de l'ichtyofaune par F. Meunier et P. Keith (MNHN)Etude physico-chimique des eaux par D. Guiral (IRD Cayenne)Prélèvements pour l'étude des communautés phyto- et zoo-planctoniques (échantillons
expédiés à l'université Montpellier 2).Le 26/06
Cartographie de la mare et oxygène dissous par D. Guiral (IRD Cayenne)Du 4 au 7/10
Etude de l'ichtyofaune par F. Meunier (MNHN)Du 13 au 18110
Etude physico-chimique des eaux par D. Guiral (IRD Cayenne)Prélèvements pour l'étude des communautés phyto- et zoo-planctoniques (échantillons
expédiés à l'université Montpellier 2).Du 24 au 26/11
Etude physico-chimique des eaux par D. Guiral (lRD Cayenne)Prélèvements pour l'étude des communautés phyto- et zoo-planctoniques (échantillons
expédiés à l'université Montpellier 2).
Au cours de ces diverses missions scientifiques une équipe pour la réalisation d'un film en coproduction entre France 2, Mona Lisa et l'IRD est venue à 4 reprises entre décembre 2001 etnovembre 2002. Cette équipe était constituée pour toutes les missions de L. Riolon (réalisateur, IRDMona Lisa) et de B. Oses (ingénieur du son, IRD, 2 missions), G. Sergent (caméraman Mona Lisa, 3missions), de R. Le Guen (caméraman, Mona Lisa, 1 mission) et de M. Quendal (caméraman, MonaLisa, 1 mission). M. Blanc a participé à 2 misions en tant qu'expert faune aquatique. Ont aussi étéinvités à effectuer un séjour sur la plate-forme A. Joly et B. Goguillon en tant que conservateurs de la

Réserve de Km,v-Roura ainsi que pour rendre compte des modalités de travail au cours des missionsscientifiques
de deux gardes de la Réserveet de deux personnes de la DIREN (S. Lochon et I. Delafosse).
La DIREN (G. Tiegoet S. Lochon) et un représentant de l'ONCFS ont en outre été embarqués lorsde rotation pour un survol de la zone où est déposée la plate forme. Enfm, trois photographesjournalistes M. Dukhan (IRD Montpellier), Le Guen (reportage pour VSD, 2 missions) et M. Soutif(reportage pour Terre sauvage, 1 mission) ont réalisé au cours de leur séjour sur la plate-fonne desimages qui sont actuellement toutes en dépôt à la base indigo de l'IRD.
En dehors du film actuellement en cours de montage et des articles sous presse (VSD, TerreSauvage, Espaces naturels) cette opération a déjà fait l'objet d'une forte médiatisation avec lapublication de deux annonces par AFP (7/12/2001 et 28/1/2003), d'articles courts dans Le Point (4janvier 2002) Sciences et Avenir (No février 2002), National Géographic (No de juin 2002), d'articlesplus développés Libération (12 mars 2002), France Guyane (9-10 décembre 2001, 9-10 novembre2002), Midi libre (9 février 2003) et dans la revue de l'IRD Sciences au Sud (N°14, mars-avril 2002,N°16, septembre-octobre 2002).
2. Les données environnementales2.1. La climatologie
2.1.1. La mare
A
29,028,0 __1IIIFi__- ---I_----
27,0 r--=--11111..---l ----::F--},..----___
26,0 a:-:Ac--__~....._::=----r--....._==--
25,0 +-----'~fJrI-+_-----:Jr-'.,.....-__:~
24,0 +------=--""":"O'l<=-------'lII~'-=-
23,0 -+--,---,.-----r-r--------,----,---,17/5 21/5 25/5 29/5 216 6/6 10/6
B
29,0
28,0 ~~iI!~~~~~~~~~27,0 ,
26,0 -+-----~lb.---___.:-
25,0 ....e.tl.--}"\..,'I"'*'.-+--lF.III.....CL.-~~
24,0 +--~--.....-----
23,0 +--,------r_---,----,----,------,
6/10 10/10 14110 18/10 22110 26110 30/10
Fig 1 A et B. Variations journalières des températures moyennesde l'air (courbe rouge) et de l'eau (courbe bleu)
A, en saison des pluies (17/5 -10/6) - B, en saison sèche (6110 -30/10)
Les températures moyennes journalières de l'air et de l'eau présentent sensiblement les mêmesévolutions au cours du temps. Les températures moyennes de l'eau sont peu différentes en fonctiondes saisons (saison des pluies: 25,09°C - saison des sèche 25,07). Les eaux en saison de pluies sont enmoyenne 2°C plus chaudes que l'air. Cet écart est plus important au cours de la saison sèche (2,5 OC).
Les coefficients de variations des températures journalières pour l'eau et l'air sont au cours de lasaison des pluies plus élevées (CVair 3,3 - CVeau 2,8) qu'en saison sèche (CVair 1,8 - CVeau 0,9).
A
32,030,0 +------------
28,0 ~œ~D~~~~~~~~ 26,0 ~24,022,0 -+----------20,0 ~------,---___r_--r_--
00:00 06:00 12:00 18:00
Heures
B
32,0
30,0 +-----~-~~~~~~28,0 ~~~~itt.!~:::::~
~ 26,0 +:24,0 +----l'J--------""22 0 20:0 -I-----r------r--.---
00:00 06:00 12:00 18:00
Heures
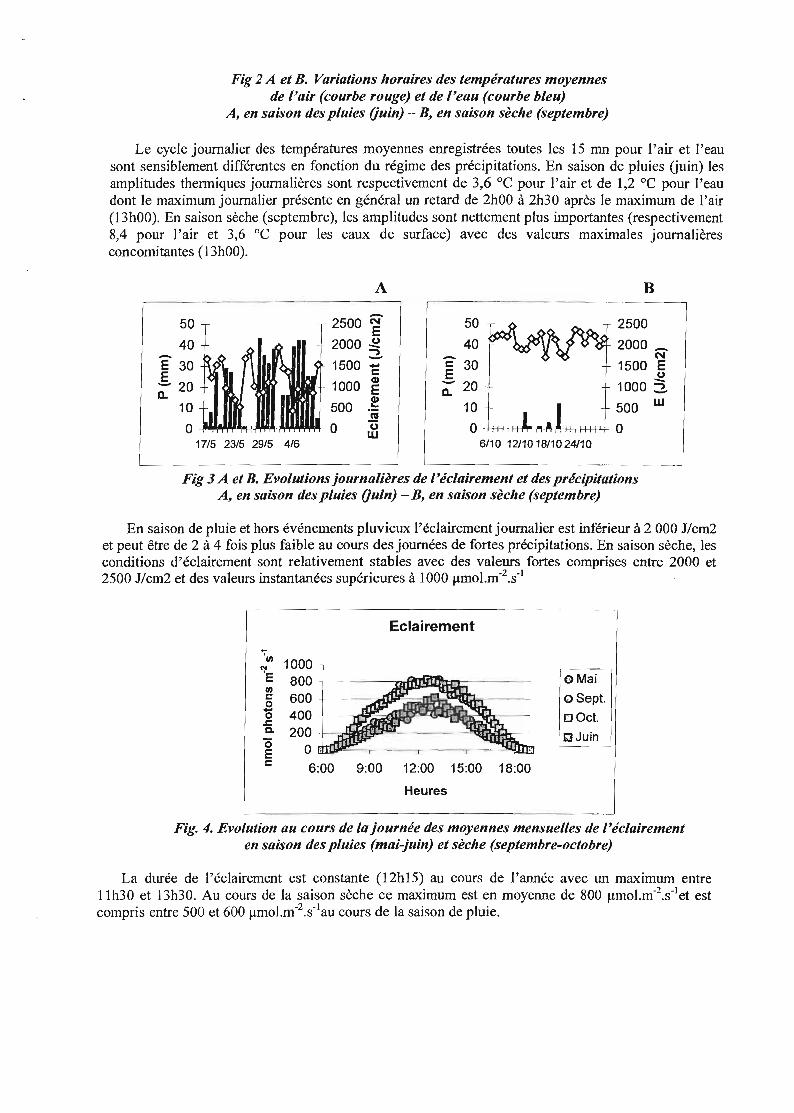
Fig 2 A et B. Variations horaires des températures moyennesde l'air (courbe rouge) et de l'eau (courbe bleu)
A, en saison des pluies Ouin) - B, en saison sèche (septembre)
Le cycle journalier des températures moyennes enregistrées toutes les 15 mn pour l'air et l'eausont sensiblement différentes en fonction du régime des précipitations. En saison de pluies (juin) lesamplitudes thermiques journalières sont respectivement de 3,6 oc pour l'air et de 1,2 oC pour l'eaudont le maximum journalier présente en général un retard de 2hOO à 2h30 après le maximum de l'air(13h00). En saison sèche (septembre), les amplitudes sont nettement plus importantes (respectivement8,4 pour l'air et 3,6 oC pour les eaux de surface) avec des valeurs maximales journalièresconcomitantes (13h00).
50
40-E 30E- 20a.
10
o17/5 23/5 29/5 416
A
2500 ~
- 2000 ~
1500 ~
1000 ~G)
500 .=cuo üw
B
50 2500
40 2000 _N
ê 30 1500 EE 0
0:- 20 1000 2-10 500 wo +t++++1-/'1"1--~~+t+1+t+- 0
6/10 121101811024110
Fig 3 A et B. Evolutions journalières de l'éclairement et des précipitationsA, en saison des pluies Ouin) - B, en saison sèche (septembre)
En saison de pluie et hors événements pluvieux l'éc1airementjournalier est inférieur à 2000 J/cm2et peut être de 2 à 4 fois plus faible au cours des journées de fortes précipitations. En saison sèche, lesconditions d'éclairement sont relativement stables avec des valeurs fortes comprises entre 2000 et2500 J/cm2 et des valeurs instantanées supérieures à 1000 J.I.mol.m'2.s·1 .
Eclairement
-'Ill 1000
1: 800 +------------:lmiJmuk~---IIIc 600 +---~~~nCI~~l-.--.so 400 +----.n~
.J:c. 200'0 0Ec 6:00 9:00 12:00 15:00 18:00
Heures
oMai
Q Sept.
oOct.
e Juin
Fig. 4. Evolution au cours de la journée des moyennes mensuelles de l'éclairementen saison des pluies (mai-juin) et sèche (septembre-octobre)
La durée de l'éclairement est constante (l2h15) au cours de l'année avec un maximum entre11h30 et 13h30. Au cours de la saison sèche ce maximum est en moyenne de 800 J.LIl1ol.m'2.s·Iet estcompris entre 500 et 600 flmoJ.m,2.s·Iau cours de la saison de pluie.
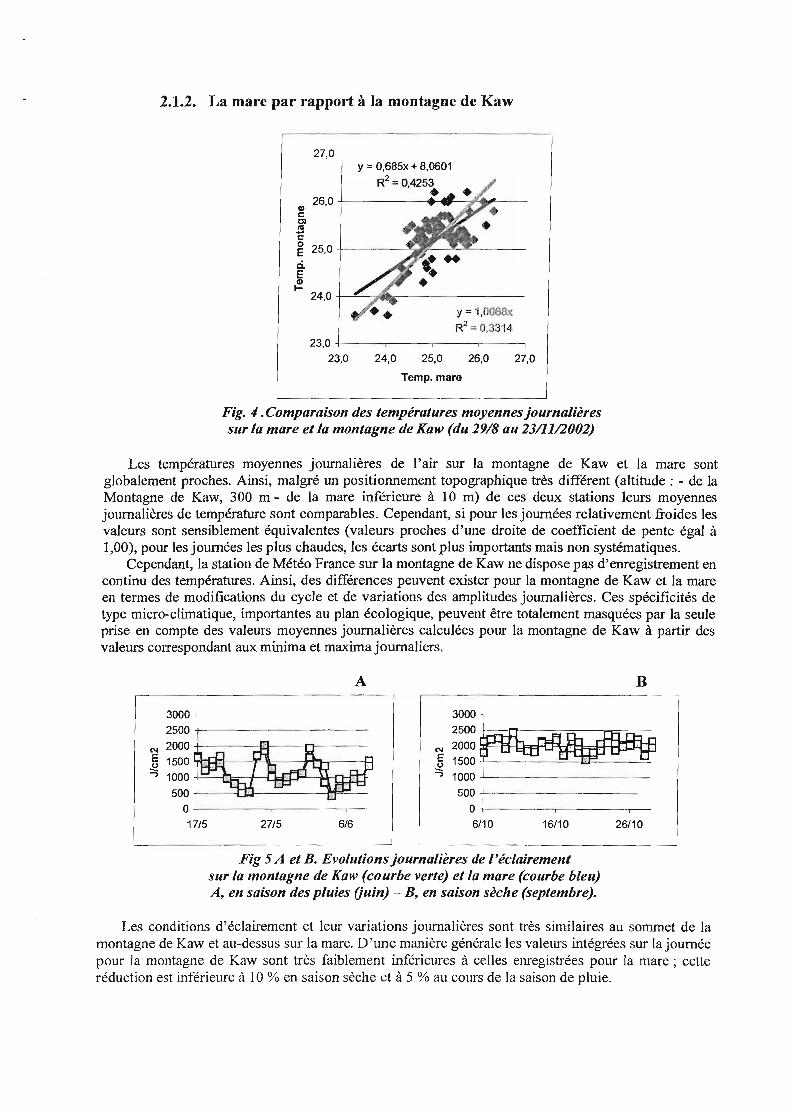
2.1.2. La mare par rapport à la montagne de Kaw
27,0y = O,685x + 8,0601
R2 = 0,4253•Cl) 26,0 +------.......r-~._..--
cClcuc~ 25,0 +----~-I'7---'<--'IL----
ci.EcDt-
24,0 +-~~~--------
y =-I,0063x
R2 =0,33'14
27,026,025,024,0
23,0 +---.-----,....---.-----,23,0
Temp. mare
Fig. " .Comparaison des températures moyennes journalièressur la mare et la montagne de Kaw (du 29/8 au 23/11/2002)
Les températures moyennes journalières de l'air sur la montagne de Kaw et la mare sontglobalement proches. Ainsi, malgré un positionnement topographique très différent (altitude; - de laMontagne de Kaw, 300 m - de la mare inférieure à 10 m) de ces deux stations leurs moyennesjournalières de température sont comparables. Cependant, si pour les journées relativement froides lesvaleurs sont sensiblement équivalentes (valeurs proches d'une droite de coefficient de pente égal à1,00), pour les journées les plus chaudes, les écarts sont plus importants mais non systématiques.
Cependant, la station de Météo France sur la montagne de Kaw ne dispose pas d'enregistrement encontinu des températures. Ainsi, des différences peuvent exister pour la montagne de Kaw et la mareen termes de modifications du cycle et de variations des amplitudes journalières. Ces spécificités detype micro-climatique, importantes au plan écologique, peuvent être totalement masquées par la seuleprise en compte des valeurs moyennes journalières calculées pour la montagne de Kaw à partir desvaleurs correspondant aux minima et maxima journaliers.
A B
3000 3000
2500 2500
N 2000 N 2000
5 1500 E 1500
:::; 1000 ~ 1000
500 500
0 O'
17/5 27/5 6/6 6/10 16/10 26/10
Fig 5 A et B. Evolutionsjournalières de l'éclairementsur la montagne de Kaw (courbe verte) et la mare (courbe bleu)A, en saison des pluies (juin) - B, en saison sèche (septembre).
Les conditions d'éclairement et leur variations journalières sont très similaires au sommet de lamontagne de Kaw et au-dessus sur la mare. D'une manière générale les valeurs intégrées sur lajouméepour la montagne de Kaw sont très faiblement inférieures à celles enregistrées pour la mare; cetteréduction est inférieure à 10 % en saison sèche et à 5 % au cours de la saison de pluie.

500400 300200 100
o~.....
A
250200 - __
150 -------:--100 -_
50 --_oo.....<D
B
Fig 6 A et B. Précipitations journalières cumuléesen saison des pluies (A) et saison sèche (B)
sur la montagne de Kaw (en vert) (Données Météo France) et sur la mare (en bleu).
En saison de pluies les précipitations journalières cwnulées sont très similaires sur la montagne deKaw et la mare. En effet, si toutes les précipitations reçues sur la montagne ne concernent pas la marela réciproque est aussi vraie et le nombre d'évènements pluvieux est comparable pour les eux sites demême que les quantités de pluie par précipitation. Par contre, en fin de saison sèche, il est possible denoter une reprise des précipitations plus précose sur la montagne qui ne semble pas se compenserultérieurement. Ce décalage devrait se traduire par un bilan annuel des précipitations sensiblementplus faible sur la mare que sur la montagne.

2.2. L'hydrologie
700
E 650Ec(1)
L- 600::s(1)-::s 550"'::I:
500\:) '!\,.....<::> .....<::> ~.....<::> .......... .......... .......... ..........
<-J." .....'11 ~ rvl{) r» ~ ~ rv~" "
50 E
40 Ec(1)
30 ~o
20 ~-.ë..10 ë3
>(1)L-o Q.
Fig 7. Variations de la hauteur d'eau dans la mare et des précipitations(en vert, montagne de Kaw et en bleu, mare)
au cours de la période de transition fin de saison sèche début de saison de pluie
Le niveau de l'eau dans la mare est remarquablement stable au cours de la saison sèche. Il nesemble donc pas exister d'influence notable de l'évaporation. Cette situation traduit uneévapotranspiration faible en accord avec les valeurs observées pour la station climatologique ausommet de la montagne. En effet en relation avec des valeurs moyennes annuèlles pour l'humidité del'air de 90 % et des vitesses du vent moyennes inférieures à 2 m.s') (1,74 m.s·1
) l'ErP ne représenteque moins de 30 % de la somme des précipitations annuelles En outre, l'importance de la couverturevégétale à l'échelle du marais ne laisse que très peu de zones d'eau libre susceptibles de conduire àune forte évaporation.
Des évènements pluvieux significatifs principalement localisés au sommet de la montagne deKaw (les 0l et 02 /11/2002 respectivement 10 et 19 mm pour la montagne et 3 et 4 mm pour la mare)ne se traduisent pas par des variations notables du niveau du plan d'eau. Une première variationsignificative du niveau de l'eau dans la mare est notée le 7/11/2002 pour des précipitationsimportantes tant pour la montagne que pour la mare (respectivement 47 mm et 28 mm). Enfin à partirdu 15/11, l'augmentation la plus importante est consécutive à plusieurs jours de fortes précipitationsau niveau de la mare et sur la montagne de Kaw.
Date Hauteurs Précipitations Précipitationsd'eau mare Mare Montagne de Kaw
(mm) (mm) (mm)15/11/02 550 1 116/11/02 580 14 2317/11/02 600 24 1818111/02 650 29 24
Tab. 1 Pluie hauteur d'eau
Ces résultats suggèrent un rôle essentiel des précipitations directes sur les variations desniveaux du plan d'eau en fin de saison sèche. Il est en outre possible qu'à cette période de transitionl'impact des premières précipitations principalement reçues sur la montagne de Kaw soit masqué parl'existence d'un phénomène de seuil lié, dans un premier temps, au remplissage de la zone de maraissituée plus en amont gue la mare. EnfIn, une augmentation de niveau de 10 cm de la mare correspondà des précipitations de l'ordre de 6,5 cm.

12
10 EE
8 cCIlc
6 0..S
4 ï5.u
2 'CD...e.
0
l~~
1"-.-
~
[] 1~C1 ]- --
~ • ].-... ..... ..
~. l;J ç ÇJ t l 7 ~
660E
640EcCD 620:::Jl'll
.CD 600-c...
580:::J.5:::J 560l'llJ:
540
16/11 16/11 17/11 17/11 18/11 18/11
Fig. 8. Variations limnigraphiques et précipitations hi horaires sur la mare.
Le suivi simultané des précipitations et des hauteurs d'eau confinne l'importance desprécipitations locales dans le fonctionnement hydrologique de la mare. Chaque événement pluvieuxengendre des variations de niveau d'eau quasi instantané.
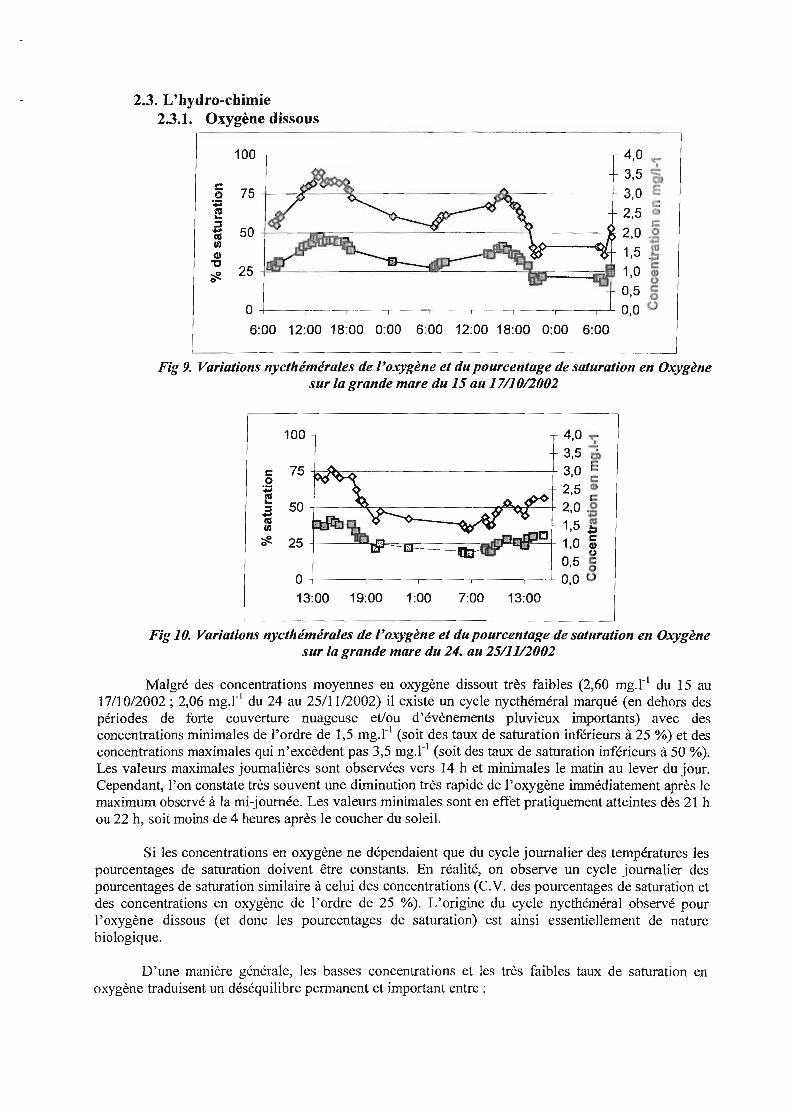
2.3. L'hydro-cbimie2.3.1. Oxygène dissous
4,0 ....J
315 =::
75 4-------;;~-~---------.R.,.___----_+ 3 0 E, c:2,5
c50 1~2iIiiï~---~-----:;;;:-t==~ 2,0 ~
1,5 E25 -j!I~-----~~:::::~~--_----=:!~:F::::::==~ 1,0 8
0,5 So -j---,--,-----,---,--,-----,--,----,---'-- 0 0 0,6:00 12:00 18:00 0:00 6:00 12:00 18:00 0:00 6:00
100
Fig 9. Variations nycthémérales de l'oxygène et du pourcentage de saturation en Oxygènesur la grande mare du 15 au 17/10/2002
100 4,0 ....-l
3,5 ~
c 75 3,0 E0 c
:0:; 2,5 CD
~ C::::1 50 2,0 :8-ca 1,5 ~III
?fe. 25 1,0 cCDCJ
0,5 c0
a 0,0 (,)
13:00 19:00 1:00 7:00 13:00
Fig 10. Variations nycthémérales de l'oxygène et du pourcentage de saturation en Oxygènesur la grande mare du 24. au 25/11/2002
Malgré des concentrations moyennes en oxygène dissout très faibles (2,60 mg.r l du 15 au17/10/2002; 2,06 mg.r l du 24 au 25/1112002) il existe un cycle nycthéméral marqué (en dehors despériodes de forte couverture nuageuse et/ou d'évènements pluvieux importants) avec desconcentrations minimales de l'ordre de 1,5 mg.r l (soit des taux de saturation inférieurs à 25 %) et desconcentrations maximales qui n'excèdent pas 3,5 mg.r l (soit des taux de saturation inférieurs à 50 %).Les valeurs maximales journalières sont observées vers 14 h et minimales le matin au lever du jour.Cependant, l'on constate très souvent une diminution très rapide de J'oxygène immédiatement après lemaximum observé à la mi-journée. Les valeurs minimales sont en effet pratiquement atteintes dès 21 hou 22 h, soit moins de 4 heures après le coucher du soleil.
Si les concentrations en oxygène ne dépendaient que du cycle journalier des températures lespourcentages de saturation doivent être constants. En réalité, on observe un cycle journalier despourcentages de saturation similaire à celui des concentrations (C.V. des pourcentages de saturation etdes concentrations en oxygène de l'ordre de 25 %). L'origine du cycle nycthéméral observé pourl'oxygène dissous (et donc les pourcentages de saturation) est ainsi essentiellement de naturebiologique.
D'une manière générale, les basses concentrations et les très faibles taux de saturation enoxygène traduisent un déséquilibre permanent et important entre:

- les apports: - production photosynthétique et flux diffusif à l'interface eau-atmosphère- les pertes: - consommations résultant des activités respiratoires et cataboliques au sein
de la colonne d'eau et des dépôts organiques accumulés au fond de la mare.En particulier, la minéralisation de la matière organique des macrophytes doit représenter un
pourcentage important de la consommation diurne et nocturne de l'oxygène dissous.Dans un tel contexte, la rapide diminution observée après 14 h peut résulter d'une stimulation
des activités respiratoires et minéralisatrices qui sont en permanence contrôlée par les faiblesconcentrations en oxygène. Sous cette hypothèse, l'augmentation des concentrations en oxygène parphotosynthèse au cours de la première partie de la journée contribuerait à une intensification desprocessus de minéralisation aérobie au cours de la seconde partie de la journée. Progressivement avecle coucher du soleil, la baisse de l'énergie incidente entraîne une diminution de la productionphotosynthétique qui devient alors inférieure à la consommation. Rapidement, l'oxygène produit par laphotosynthèse des communautés phytoplanctoniques est consommé par les activités respiratoires del'ensemble de la biocénose et surtout par les processus de minéralisation aérobie. Ainsi, dès le débutde la nuit, les concentrations sont, à nouveau, basses et engendrent un ralentissement des activitésminéralisatrices qui se traduit par une quasi-stabilisation des concentrations en oxygène. Au cours dela nuit, la consommation d'oxygène par les activités cataboliques sédimentaires et respiratoires descommunautés vivantes au sein de la colonne d'eau est ainsi essentiellement contrôlée par le fluxdiffusif d'oxygène à l'interface eau-atmosphère dont l'intensité augmente en fonction de la baisseprogressive des pourcentages de saturation en oxygène dans l'eau.
026/06 (11h15 à 13h45)
014110 (15h35 à 17h20)
016110 (8h45 à 14h10)
024/11 (11h30 à 13h20)
œ25111 (6h45 à 7h55)
Fig 11. Distribution spatiale des concentrations en oxygène
Au plan spatial et pour les 3 périodes étudiées (saison de pluie: juin - saison sèche: octobre fin de saison sèche: novembre) les valeurs en oxygène n'excédent jamais 5,0 mg.1-1 et correspondentainsi à des eaux plus ou moins fortement dessaturées. On constate en outre une très importantevariabilité nycthémérale qui masque d'éventuelles variations en fonction de la localisation des stationsétudiées.

2.3.2. Conductivité et pH
6,00 140
120 ......-5,80 100 E
u::I:
80 vic. 60
_.-
5,60 - -040 r:
0
20
5,40 0M. Hoat. M.Aga. M.Aig. F.Sav Gr. mare
Fig. 12. Valeurs moyennes du pH et de la conductivité des eaux des diverses mares.Hoatzin (M. Hoat.), Agami (M. Aga.), Aigrette (M. Aig.), Forêt Savacou. (F. Sav.) et Grande mare
(Gr. mare)
Les eaux des mares sont globalement très peu minéralisées et acides. Les mares situées àproximité des aires de nidification des oiseaux d'eau sont significativement plus minéralisées.
2.3.3. Sels nutritifs
5,004035 +-------=---u----/--\----I ~
30 +----L..I==--------",/--\------I 4,00 ~CO')
25 -+--------f-~~~'-'---_=I 3,00 ~
20 +-------r------\----j (!;15 -+----W----€:!"""'------.-,---\---1 2, 00 ~
10 +-------------y<---~-._+r-! "'lt1,00 i
5 +-----------/----~--Io -I---- -..,..-=lI===;-__.-==r==lIl==;==O--!- 0,00
M. Hoat M. Aga. M. Aig. F. Say. Gr. mare
Fig. 13. Valeurs moyennes des concentrations en sels nutritifs (en pM.rJ; des eaux des maresHoatzin (M. Hoat.), Agami (M. Aga.), Aigrette (M. Aig.), Forêt Savacou. (F. Sav.) et Grande mare
(Gr. mare)
Les concentrations en nitrites et nitrates sont excessivement basses (valeur maximale 0,1 /lM.r1). L'azote minéral est ainsi essentiellement sous forme réduite. Ces résultats sont probablement uneconséquence des faibles niveaux d'oxygénation des eaux (faible activité nitrifiante) et d'une très forteproductivité des macrophytes flottants (forte immobilisation biologique des nitrates).
Les concentrations en phosphore minéral sont inférieures à 0,5 /lM.rl à l'exception des eauxsituées sous les arbres utilisés par les savacous pour leur nidification. C'est aussi à cette station quel'on observe les plus fortes concentrations en azote ammoniacal. En absence de végétation aquatiqueflottante (très fort ombrage par les arbres) la fertilisation des eaux par les déjections des oiseaux estainsi mise en évidence.
Les concentrations en silice soluble sont comparativement aux autres sels nutritifs nettementplus importantes(de l'ordre de 30 /lM.r l
). Cependant ces valeurs sont faibles comparativement aux
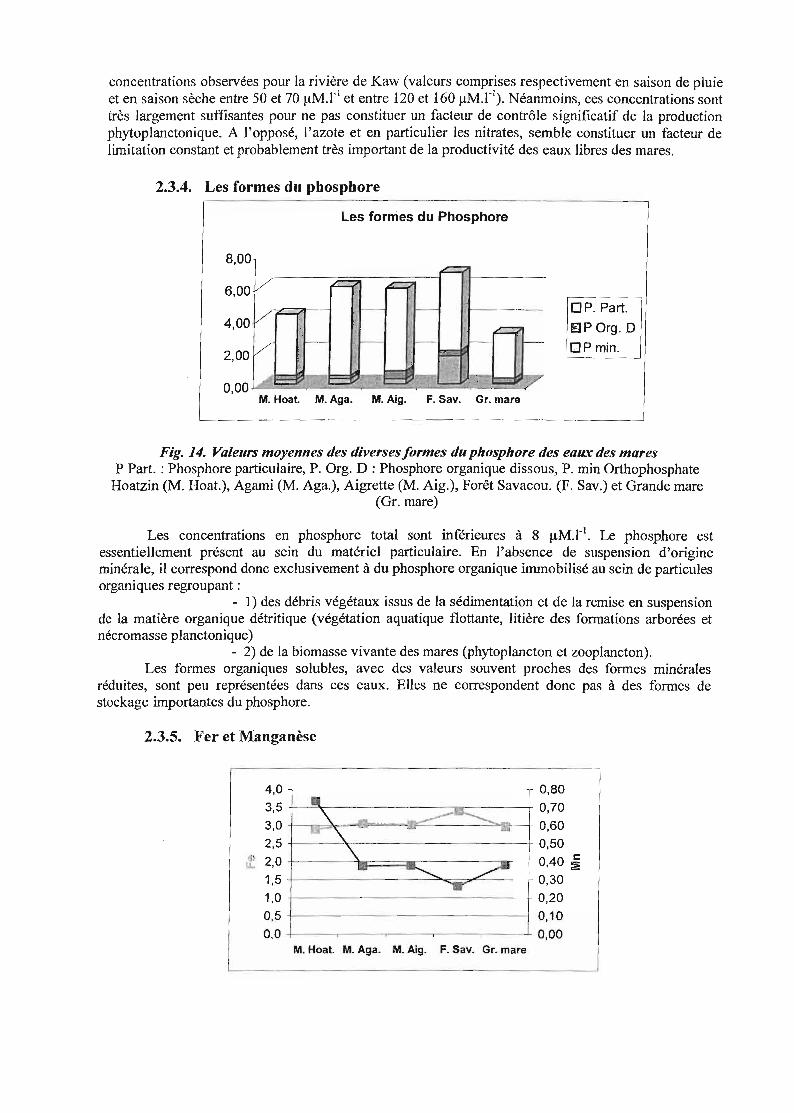
concentrations observées pour la rivière de Kaw (valeurs comprises respectivement en saison de pluieet en saison sèche entre 50 et 70 /lM. rI et entre 120 et 160 /lM. rI). Néanmoins, ces concentrations sonttrès largement suffisantes pour ne pas constituer un facteur de contrôle significatif de la productionphytoplanctonique. A l'opposé, l'azote et en particulier les nitrates, semble constituer un facteur delimitation constant et probablement très important de la productivité des eaux libres des mares.
2.3.4. Les formes du phosphore
Les formes du Phosphore
8,00
4,00
2,00
M. Hoat. M. Aga. M. Aig. F. Say. Gr. mare
OP. Part.
If]P Org. D
OP min.
Fig. 14. Valeurs moyennes des diverses formes du phosphore des eaux des maresP Part. : Phosphore particulaire, P. Org. D : Phosphore organique dissous, P. min Orthophosphate
Hoatzin (M. Hoat.), Agami (M. Aga.), Aigrette (M. Aig.), Forêt Savacou. (F. Sav.) et Grande mare(Gr. mare)
Les concentrations en phosphore total sont inférieures à 8 IlM.l-I. Le phosphore estessentiellement présent au sein du matériel particulaire. En l'absence de suspension d'origineminérale, il correspond donc exclusivement à du phosphore organique immobilisé au sein de particulesorganiques regroupant:
- 1) des débris végétaux issus de la sédimentation et de la remise en suspensionde la matière organique détritique (végétation aquatique flottante, litière des formations arborées etnécromasse planctonique)
- 2) de la biomasse vivante des mares (phytoplancton et zooplancton).Les formes organiques solubles, avec des valeurs souvent proches des formes minérales
réduites, sont peu représentées dans ces eaux. Elles ne correspondent donc pas à des formes destockage importantes du phosphore.
2.3.5. Fer et Manganèse
4,0 O,SO
3,5 +--=.c---------rv----_t_ 0,70
3,0 0,60
2,5 0,50
~. 2,0 0,40 i1,5 . 0,30
1,0 0,20
0,5 0,10
0,0 0,00M. Hoat. M. Aga. M. Aig. F. Say. Gr. mare
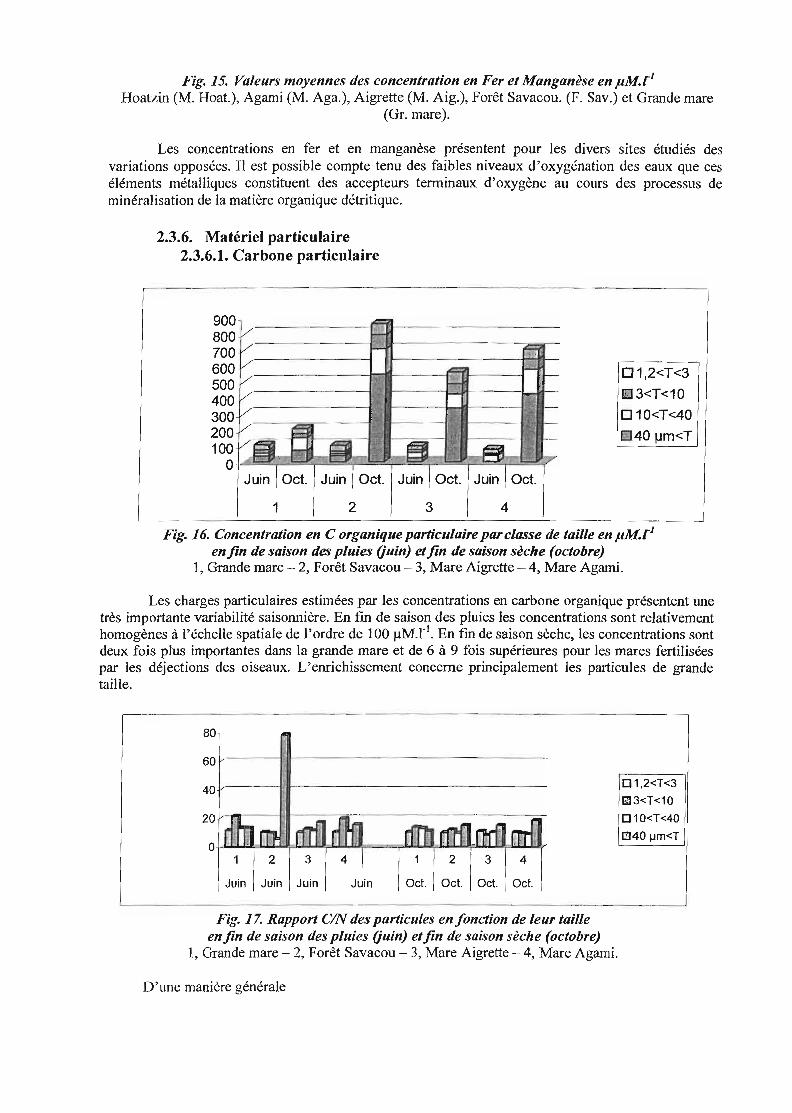
Fig. 15. Valeurs moyennes des concentration en Fer et Manganèse en pM.riHoatzin (M. Hoat.), Agami (M. Aga.), Aigrette (M. Aig.), Forêt Savacou. (p. Sav.) et Grande mare
(Gr. mare).
Les concentrations en fer et en manganèse présentent pour les divers sites étudiés desvariations opposées. Il est possible compte tenu des faibles niveaux d'oxygénation des eaux que ceséléments métalliques constituent des accepteurs terminaux d'oxygène au cours des processus deminéralisation de la matière organique détritique.
2.3.6. Matériel particulaire2.3.6.1. Carbone particulaire
-
900 11=V ' ..
800 ~
700 V .-.~
600 /" e::;. - D1,2<T<3/--_.~r
~
500 H
400 /" ~ r;- El 3<T<10
300 V ~
e=:lD10<T<40
200 /' ~ J; '04O l-lm<T100 /~ h:: ~ -~ ~
0 .LI '
Juin 1 Oct. Juin 1Oct. Juin' Oct. Juin 1 Oct.
1 2 3 4
Fig. 16. Concentration en C organique particulaire parc/asse de taille en pM.r]enfin de saison des pluies Ouin) etfin de saison sèche (octobre)
1, Grande mare - 2, Forêt Savacou - 3, Mare Aigrette - 4, Mare Agami.
Les charges particulaires estimées par les concentrations en carbone organique présentent unetrès importante variabilité saisonnière. En fm de saison des pluies les concentrations sont relativementhomogènes à l'échelle spatiale de l'ordre de 100 ~M.rl. En fin de saison sèche, les concentrations sontdeux fois plus importantes dans la grande mare et de 6 à 9 fois supérieures pour les mares fertiliséespar les déjections des oiseaux. L'enrichissement concerne principalement les particules de grandetaille.
Oct. Oct. Oct. Oct.
80
60
40 ---
20
02 3 4
Juin Juin Juin Juin
2 3 4
01,2<T<3
EJ3<T<10
010<T<40
[J40 IJm<T
Fig. 17. Rapport C/N des particules enfonction de leur tailleenfin de saison des pluies Ouin) etfin de saison sèche (octobre)
1, Grande mare - 2, Forêt Savacou - 3, Mare Aigrette - 4, Mare Agami.
D'une manière générale
1) Les rapports C/N des particules inférieures à 40 j.lm sont voisines de 10. Ellesdoivent ainsi correspondre essentiellement à des particules actives ou peu dégradées.
2) Les particules de grande taille (T> 40 j.lm) présentent des rapports C/Nsupérieurs à ceux des particules de taille inférieure. Cette augmentation comparativement aux cellulesde petite taille traduit la présence d'une matière organique particulaire plus évoluée ou héritée desformations végétales terrestres.
3) Les faibles concentration de particules en fin de saison des pluies présentent desrapports C/N généralement supérieurs à ceux observés au cours de la [m de saison sèche. Au cours dela saison des pluies les cellules phyto-planctoniques ne représentent doivent représentées qu'unebiomasse et une proportion relative faible des particules en suspension dans les eaux de la mare.
2.3.6.2.Les biomasses chlorophylliennesA B
r------------------
3,00 ,-----L.JI2,50 l ,------1r-n-----[llI2,00 ,--4-1f---H§1,50 /O&:::l.---t1----Ill~....1,000,500,00 CD 0 CD 0
::ë al ::ë al
() cf () cf2
DO,7<T<3
D3<T<10
Eil10<T<40
IiIT>40
30,0 c25,01/ .u------
20,0 [/~~-z::or--15,0100 v I~5:0 v -"':;"F,Il~-lIiiiII-
0,0..1::111 g .!!! gr() cf fj cf
2
DO,7<T<3
o 3<T<1 0
g10<T<40
BT>40
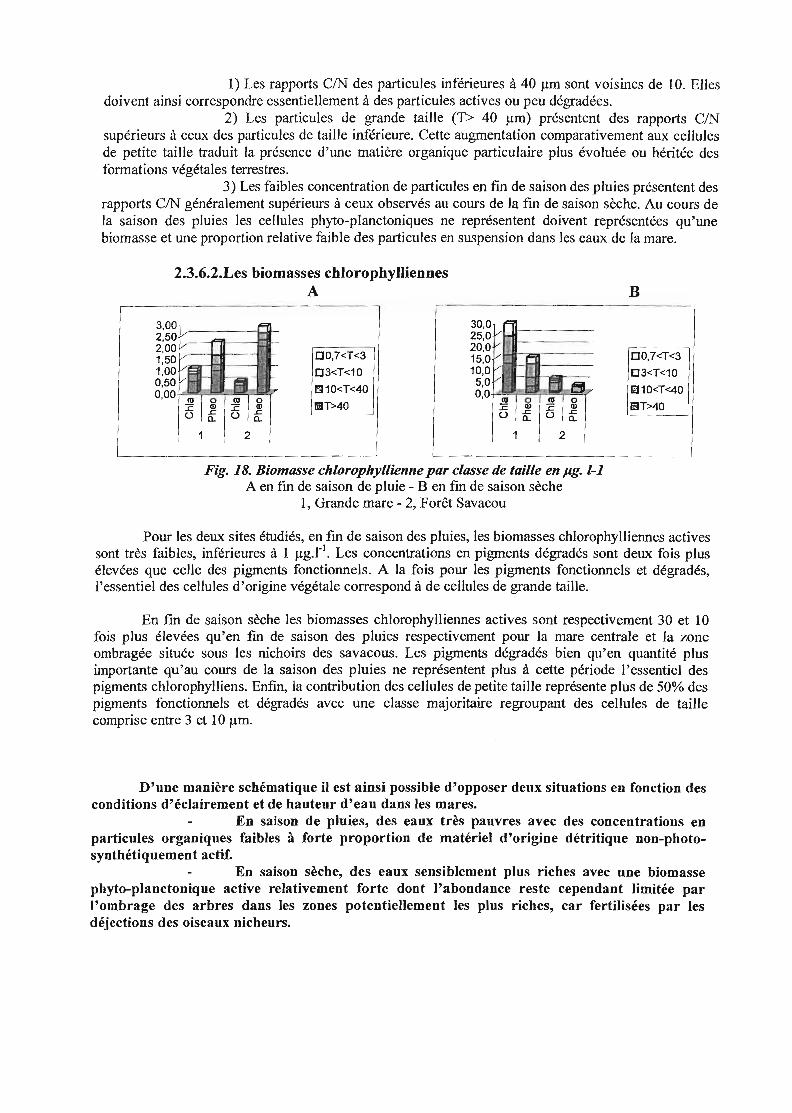
Fig. 18. Biomasse chlorophyllienne par classe de taille en pg. 1-1A en [m de saison de pluie - Ben [m de saison sèche
1, Grande mare - 2, Forêt Savacou
Pour les deux sites étudiés, en fm de saison des pluies, les biomasses chlorophylliennes activessont très faibles, inférieures à 1 j.lg.r1
• Les concentrations en pigments dégradés sont deux fois plusélevées que celle des pigments fonctionnels. A la fois pour les pigments fonctionnels et dégradés,l'essentiel des cellules d'origine végétale correspond à de cellules de grande taille.
En [m de saison sèche les biomasses chlorophylliennes actives sont respectivement 30 et 10fois plus élevées qu'en fm de saison des pluies respectivement pour la mare centrale et la zoneombragée située sous les nichoirs des savacous. Les pigments dégradés bien qu'en quantité plusimportante qu'au cours de la saison des pluies ne représentent plus à cette période l'essentiel despigments chlorophylliens. Enfin, la contribution des cellules de petite taille représente plus de 50% despigments fonctionnels et dégradés avec une classe majoritaire regroupant des cellules de taillecomprise entre 3 et 10 j.lm.
D'une manière schématique il est ainsi possible d'opposer deux situations en fonction desconditions d'éclairement et de hauteur d'eau dans les mares.
En saison de pluies, des eaux très pauvres avec des concentrations enparticules organiques faibles à forte proportion de matériel d'origine détritique non-photosynthétiquement actif.
En saison sèche, des eaux sensiblement plus riches avec une biomassephyto-planctonique active relativement forte dont l'abondance reste cependant limitée parl'ombrage des arbres dans les zones potentiellement les plus riches, car fertilisées par lesdéjections des oiseaux nicheurs.

3. Les données biologiques3.1. La végétation (JJ de Granville)
La zone centrale d'eau libre est occupée par une végétation aquatique à hydrophytesimmergés et flottants enracinés au non au fond de la mare. Cette zone regroupe diverses unités.
1a le groupement à Nympheae rudgeana, Cabomba aquatica et Sa/viniaauricu/tata. Sa biodiversité floristique est faible
1b le peuplement monospécifique à E/eocharis interstincta1c les petits îlot de pégase flottant sont essentiellement colonisés par des Cypéracées
(Fuirena umbellata et Cyperus haspan parfois des Poaceae (Panicum parvifolium, Sacciolepis striata).On peut en outre y observer Xyris laxifolia et l'orchidée Habenaria laugicauda.
Le marais d'eau douce à Cypéracées et fougères sur pégasse constitue la zone la plusétendue et la plus riche en terme de diversité floristique. A ce groupement se mêlent des espèces desmarais à Montrichardia arborescens faciès à Crinum erubescens. C'est une flore intermédiaire entreces deux groupements que l'on observe au nord et au sud de la mare. La végétation y est dense etbasse, dominée par Rhynchopora gigantea, Sagittaria lancifolia et les fougères Thelypteris interruptaet Blechun serrulatum. Deux espèces herbacées plus ou moins ligneuses à fleurs jaunes dépassant lavégétation herbacée Ludwigia nervosa et L. turulosa sont abondantes. Dans les zones moins denses enbordure de l'eau libre fleurit Crinum erubescens. Montrichardia arborescens ne forme pas depeuplement dense. L'espèce la plus remarquable est un arbuste du genre [lex à fleurs blanches. Cetteespèce très rare en Guyane n'est représentée dans l'herbier que par deux spécimens provenant de lamême localité (Savane Angélique). Des spécimens sont actuellement à l'étude de l'herbier de Genève.
Les marécages boisés de hauteur et de densité variable limitent la mare à l'est et à l'ouèst. Ilssont principalement constitués par Ptérocarpus officinalis et Chrysobalanus icaco auxquels se mêlentd'autres espèces ligneuses en faible densité Palicourea croceoides, Annoma ga/bra, Topcocaguianenesis ....
Dans la partie à l'ouest de la mare on voit en sous bois de nombreux pieds de [lex quis étiolent sans fleurir et la présence d'une aracée peu commune DiejJènbachia paludicola.
Dans la zone située à l'est poussent également quelques pabniers Euterpe o/eracea etBactris campestris. Les arbres bas et peu denses laissent passer une grande quantité de lumièrefavorisant la prolifération des épiphytes bas en particulier Codonanthe crassifolia, Peperomia sp. pl.,des orchidées Epidendrum nocturnum et surtout Catasetum cf saccatum, des Araceae et des fougèresen grande densité notamment Polypodium attenuatum, Capyloneurum phyllitis, Vittaria /ineata. Lalisère avec l'eau libre ainsi que les chenaux qui s'enfoncent dans la zone boisée sont marqués par depetits îlots de végétation flottante ou l'on remarque en autres Sacciolepsis striata, Bacopa aquatica,Hydrocotyle umbellata et quelques rares pieds d'une espèce nouvelle pour la Guyane Boehmeriaramiflora (Urticaceae).
3.2. Les poissons (F. Meunier)Les deux campagnes d'échantillonnage en fm de saison de pluie et sèche ont permis la
capture, tous engins de pêche confondus, de 9 familles regroupant un total de 24 espèces. Malgré uneffort important de prospection cette richesse spécifique est significativement plus faible que lenombre initialement envisagé (30 à 35) compte tenu des caractéristiques de la zone. En particulier, onnote une très grande pauvreté des Silurifonnes puisque deux espèces seulement ont été capturées(Parauchenipterus galeatus et Rhamdia quelen). De même, un seul Gymnotiforme est présent(Electrophorus e/ectricus). Ces deux groupes sont représentés par 33 (Ariidae non compris) et 12espèces dans les zones inondables de la rivière de Kaw. De même, dans le Pri-pri Yiyi, un marais àpriori similaire à celui de Kaw, les siluriformes et les Gymnotiformes sont respectivement riches de 21et de 3 espèces (Electrophorus electricus étant lui absent). Il faut aussi noter l'absence totale decapture d'Atipas tant de Hoplosternum littorale que de Megalechis thorocatum.

Au plan spatial il est possible d'identifier deux types de communautés les grandes zones d'eaulibre colonisées par la végétation aquatique avec une très grande abondance de l'espèce herbivoreMetynnis cf lippincottianus. Les zones sous couvert forestier avec la présence de poisons benthiquesParauchenipterus ga/eatus et Rhamdia que/en.
D'une manière générale les densité sont très élevées avec un peuplement dominé pour les deuxzones au plan structurel par des espèces prédatrices ichtyophages: Hop/erythrinus unitaeniatuis,Hoplias ma/abrica, Acestrorhynchius fa/acatus et diverses espèces de Cichlidae. Certains individus dece groupe (Hoplias ma/abrica) ont des tailles, dans ce milieu, supérieures au plus grande connue pources espèces (en juin individus compris entre 315 et 390 mm, en octobre entre 285 et 415 mm.L'abondance des prédateurs de grande taille est en outre associée à une assez grande pauvretéd'espèces de taille moyenne comme les Astyanx ou les Moenkhausia. A la fm de la saison de pluiel'hypothèse d'une répartition de ces espèces au sein de l'ensemble de la zone humide avait étéproposée pour interpréter leur absence dans les captures et expliquer cette structuration dimensionnelletrès déséquilibrée du peuplement. Cependant, en saison sèche si les juvéniles et les jeunes poissonsétaient très abondants les seuls poissons de taille intermédiaire entre les très petites espèces(Hemigrammnus, Pristella et Nannostomus) et les poissons de la taille des Heros, Hoplia,Acestrorynchus étaient des juvéniles et/ou des sub-adultes d'espèces comme les Cich/asoma, Krobiaou encore Methynnis.
Bnfm, un Cichlidae qui n'était connu que de la rive droite de l'Oyapock Heros efasciatus a étécapturé en juin et en octobre. Sa présence dans le marais de Kaw élargit ainsi l'aire de répartitionconnue pour cette espèce avec d'intéressants développements possibles au plan biogéographique. Eneffet, en l'état actuel des connaissances, cette espèce est apparemment absente dans les marais situésentre l'Oyapock et l'Approuague et à l'est du marais de Kaw. Eofm, son observation au voisinage deKourou interprétée comme une introduction accidentelle par des aquariophiles mérite peut être d'êtremaintenant réexaminée.
Au plan éco-physiologique il est important de noter que ces populations de poissons semblenttrès adaptées à une vie dans des eaux à très faible concentration en oxygène. Cette adaptation àl 'hypoxie se traduit pas une totale incapacité de survivre dans des eaux présentant des tauxd'oxygénation proches de la saturation.
3.3. Les oiseaux (0. Tostain, T. Deville et P. Reynaud)87 espèces d'oiseaux ont été recensées dans ce site au cours de 3 missions de courte durée
entre les mois de mars et juin 2002. Outre cette richesse spécifique remarquable une espèce demarouette nouvelle pour la Guyane (Latera/lus me/anophaius) a été observée dans la végétationherbacée en limite de la mare. Cependant, ce site est surtout exceptionnel pour sa fonction de zone dereproduction pour divers oiseaux et en particulier pour de très rares hérons sud-américains. Au totalsept espèces ont successivement utilisé les arbres limitrophes de la mare comme site de nidification.Par ordre d'arrivée au cours de l'année ces espèces sont: Nycticorax nycticorax, Ardea cocoï, Agamiaagamia, Egretta a/ba, Coch/earius coch/earius, Pha/acrocorax brasilianus et Anhinga anihngaConcernant la population nicheuse d'A. agamia avec plus de 1000 couples cette mare constitueactuellement la zone la plus importante au monde identifiée comme aire de reproduction pour cetteespèce. La seule zone précédemment connue au Costa Rica ne comptait que 12 couples.En outre en fm de saison sèche, des parades nuptiales très exubérantes de Cairina moschata se sontdéroulées sur la mare et, au début de la saison de pluie, ses abords immédiats ont été régulièrementutilisés comme dortoir pour une colonie très importante (3000 individus) de Hirundo rustica. Bofm, lavégétation arborée de la mare est utilisée pratiquement toute l'année pour la reproduction etl'alimentation de familles d'hoatzins regroupées à la limite des eaux libres en trois colonies quisemblent présenter une assez forte cohésion sociale.
3.4. Les caimans (B. de Thoisy)Rapport non parvenu.

Premières conclusions
Le choix de cette mare sur des critères d'hétérogénéité structurelle a permis la mise enévidence d'un site écologique remarque pour la Guyane, mais aussi, d'une très grandeimportance pour diverses espèces de plantes, de poissons, de caüuans et d'oiseaux sudaméricains naturellement très rares et/ou en voie de disparition.
Les connaissances acquises sur le plan physico-chimique ont permis, en outre, demettre en évidence un fonctionnement écologique tout à fait original de cette mare dont laproductivité biologique repose sur sa fonction d'aire de reproduction pour diverses espècesd'oiseaux d'eau dont certaines populations, dans ce milieu, sont les plus importantesactuellement connues au monde. Les oiseaux par la prédation qu'il exerce hors de la mare etl'alimentation de leur poussin au nid assurent un retour des sels nutritifs exportés via lesmigrations saisonnières des poissons en fonction des variations de niveau des eaux dans lemarais et donc des saisons hydro-climatiques. Dans ce mode de fonctionnement les caÜllansoccupent une place écologique importante en tant que super-prédateur. Ils interviennent eneffet à un double niveau 1) en occupant seul la niche trophique des prédateurs terminaux degrande taille et des nécrophages 2) en protégeant la mare de la présence de prédateurspotentiels d'œuf (iguanes et singes). Par leur pression de prédation ils contribuent ainsi àassurer la pérennité de cette aire de reproduction d'importance mondiale pour pratiquementl'ensemble des espèces oiseaux d'eau colonisant les zones humides sud américaines.
Au cours de l'année 2003 les études seront poursuivies selon quatre axes principaux :- poursuivre les inventaires biologiques et en particulier ceux relatifs à
l'entomofaune et à la végétation aquatique immergée,- étudier les relations fonctionnelles qui existent entre la mare et l'ensemble de
la zone inondable du marais de Kaw à partir d'une poursuite et d'une intensification desétudes reliant la climatologie et l'hydrologie comparée de la mare et de la rivière de Kaw,
- confirmer et préciser l'impact en termes de fonctionnement et de productivitédes eaux de la fertilisation par les déjections des oiseaux,
- réaliser des carottages pour étudier les modes de comblement de la mare enrelation avec les informations acquises lors de l'étude des 3 carottages réalisés en 2002 sur lesberges de la rivière de Kaw.
L'avifaune, élément déterminant de la valeur écologique et patrimoniale du site, nefera l'objet en 2003 que de suivis à caractère descriptif (nombre de nid et estimation du succèsreproducteur) avec comme premier objectif l'évaluation du niveau de nuisance occasionnéepar une présence humaine temporaire sur cette mare. Ces études seront menées enconcertation avec l'IRD par le conservateur de la réserve et l'association du Gepog. Selon lesrésultats de cette étude de transition, il sera recherché des moyens et un contexte scientifiquepour la réalisation d'études plus importantes et durables afm de déterminer pour les deuxespèces de hérons (agami et savacou) les plus emblématiques de la mare:
1) la fréquence et la durée de leur déplacement journalier et leurs sites degagnage lors de leur période d'occupation de la mare,
2) leur fidélité à la mare,3) leur éthologie et leur régime alimentaire en période de reproduction.
Des études similaires sur l'éthologie des caüuans noirs pourraient aussi être envisagés.
D'une manière plus globale ce site de très haute valeur biologique et son infrastructurescientifique (la plate-forme, les stations météorologique et limnigraphique) peuvent constituer

une base d'étude très importante pour la connaissance des zones humides de Guyane dans lecadre de programmes d'intérêt régional voire international du type par exemple: Rôle deszones humides dans la production de gaz à effet de serre.
Compte tenu des observations déjà acquises et des potentialités que cette plate-formeoffre en tenues d'études multidisciplinaires et de coopération multi-organisme pour laréalisation de recherches appliquées (contribution à la connaissance, la sauvegarde et à laprotection d'espèces rares) et fondamentales (rôle fonctionnel des zones d'eau libre au seind'un marais d'hydrophytes flottants - rôle des zones humides littorales dans la régulation desgrands flux biogéochimiques) il semble important de tout mettre en œuvre pour assurer lapérennité à moyen terme de cette plate-forme sur ce site. Cependant, dès à présent, elle apermis de notablement accroître la connaissance scientifique du marais de Kaw et, aposteriori, elle a confirmé le bien fondé des nombreuses décisions conservatoires dont cemilieu a fait l'objet depuis 1993 (Inscription sur la liste des sites Ramsar).





























