Embed Size (px)
Citation preview

Article original
Les principaux types de sapinières (Abies alba Mill)dans le massif du Jura (France et Suisse).
Étude phytoécologique
GD Bert
INRA, laboratoire de phytoécologie forestière, centre de recherches de Nancy,54280 Champenoux, France
(Reçu le 18 octobre 1991; accepté le 3 décembre 1991)
Résumé — Les premiers résultats d’une étude dendrochronologique ont montré que les peuple-ments de sapin (Abies alba Mill) dans le Jura ont traversé une période de crise entre 1973 et 1982,due en grande partie à des sécheresses prononcées. Considérées dans leur ensemble, ces sapi-nières bénéficient depuis d’un net rétablissement de leur croissance radiale. Cependant, certainesd’entre elles sont aujourd’hui encore dépérissantes. L’hypothèse d’une plus grande sensibilité decertaines stations vis-à-vis de stress climatiques a impliqué l’étude de l’interaction entre manifesta-tions du dépérissement et conditions écologiques locales, préalablement mises en évidence pourchaque type de station. L’échantillonnage de 208 sites de sapinières a permis de réaliser une typolo-gie des principales unités stationnelles sur toute l’aire de répartition du sapin dans le Jura. Cet ar-ticle présente les caractéristiques écologiques et floristiques des 11 principales unités rencontréesau sein de 6 associations phyto-sociologiques : hêtraie à laîche, hêtraie à tilleul, hêtraie à dentaire,hêtraie-sapinière, hêtraie à érable et pessière à doradille. La composition floristique des peuple-ments est essentiellement conditionnée par l’altitude et le bilan hydrique du site. Les influences cli-matiques, la biogéographie de certaines espèces et la dissymétrie de la chaîne montagneuse expli-quent la disjonction géographique entre certaines unités stationnelles. Certains types de stationsfloristiquement proches présentent d’importantes différences du point de vue de l’état sanitaire dessapins, apprécié par leur degré de défoliation et de jaunissement. Cette composante stationnelle dudépérissement sera précisée par une étude dendroécologique de la croissance radiale des sapins.
typologie des stations / phytoécologie / sapinière / Abies alba = sapin / Jura
Summary — Main types of silver fir (Abies alba Mill) stands in the Jura mountains (Franceand Switzerland). A phytoecological study. The initial results of a dendrochronological studyshowed that fir stands underwent a crisis period from 1973 to 1982, mainly due to severe droughts.These stands, considered as a whole, have recovered a normal radial growth level. Nevertheless,some of them are still declining, which suggests that some sites are more sensitive than others to cli-matic stress. In order to study the relationships between ecological characteristics of sites and die-back symptoms, we determined a typology for silver fir stands. Two hundred and eight sample plotswere selected that were as representative as possible of the natural range of silver fir in the Juramountains (eastern France). This paper describes 11 main types of sites belonging to 6 phytosocio-logic associations: Carici-Fagetum, Tilio-Fagetum, Dentario-Fagetum, Abieti-Fagetum, Aceri-Fagetum and Asplenio-Piceetum. The ground vegetation depends mainly upon altitude, soil watersupply, but also on geographic location due to the distribution of some species and site characteris-

tics. The soils of the Jura, which is a calcareous massif, consist of rendzina, humic cambisol, calciccambisol, chromic cambisol, orthic luvisol and chromic luvisols (FAO). A subsequent dendroecologicalstudy will deal with differences in state of health between site types.
site typology / phytoecology / fir stand /Abies alba = silver fir / Jura mountains
INTRODUCTION
Le sapin pectiné, essence indigène du Jura,de grande importance économique, montredes symptômes de dépérissement danscette région. Pour rechercher les causes dece dépérissement et en comprendre le mé-canisme, des recherches écologiques et
dendrochronologiques ont été entreprisesdans le contexte phytogéographique juras-sien, aux sols calcaires, calciques ou
acides. Ces travaux ont permis la comparai-son avec les études sur le dépérissementdu sapin dans les Vosges, sur des sols enmoyenne très acides. Les premiers résultatsd’une étude dendrochronologique ont mon-tré que, comme dans les Vosges (Becker,1987; Becker et Lévy, 1988), les sapinièresjurassiennes, considérées dans leur en-
semble, bénéficient d’un net rétablissementdu point de vue de la croissance radiale,après une période de crise entre 1973 et1982 due en grande partie à des séche-resses prononcées (Bert et Becker, 1990).
Cependant, certains peuplements sontaujourd’hui encore dépérissants. Au coursde travaux ultérieurs, nous chercheronsdonc à tester l’hypothèse d’une plusgrande sensibilité de certains arbres ou decertaines stations vis-à-vis de stress clima-
tiques, éventuellement aggravée par demauvaises conditions sylvicoles ou l’inter-vention de polluants. L’étude de l’interac-tion possible entre conditions écologiqueslocales et manifestation du dépérissementpasse par l’utilisation d’une typologie desstations préalablement construite. En effet,la réalisation d’une classification de la vé-
gétation met en évidence les principauxfacteurs du milieu qui structurent la flore
(Pfister, 1989) et les caractéristiques éco-logiques de chaque type de station. Cettetypologie était encore fragmentaire du pointde vue géographique et hétérogène pourune utilisation sur l’ensemble du massif.Moor (1952) a étudié les associations du
Faglon dans le Jura suisse, Richard (1961)a traité des forêts acidophiles du Jurasuisse et de la moitié nord du jura français,Guinochet (1973) a cartographié très fine-ment une petite région au sud de Pontarlier,Simmeray (1976) a interprété les groupe-ments de la région de Saint-Claude, Gaiffeet Schmitt (1980) ont analysé la végétationde Mouthe à Pontarlier, Gillet et al (1984)ont réalisé une étude cartographique de larégion sud-est du massif et Rameau (1988)a décrit les hêtraies mésoneutrophiles etacidiclines de la partie ouest du Jura.
Le dispositif initial de 87 placettes, misen place en 1987, dans les départementsdu Doubs et du Jura, a été étendu à l’en-semble du massif, tant dans sa partie fran-çaise que suisse. Ainsi, notre étude de ter-rain a permis de rassembler les élémentsnécessaires à un examen général de la va-riabilité stationnelle à l’échelle du massif.
L’analyse a conduit dans un premier tempsà identifier les associations végétales fo-restières déjà définies sur le Jura. Puisnous avons précisé la variabilité station-nelle de chacune d’elles afin de définir des
unités stationnelles plus homogènes. Cesunités stationnelles ne correspondent pastoujours à des «types de stations» (unitésparfaitement homogènes du point de vuestationnel) car l’échantillonnage a été réali-sé dans un but dendroécologique.
Cet article présente les résultats phytoé-cologiques issus de l’analyse de la végéta-

tion et les caractéristiques écologiques etgéographiques des unités stationnelles.
AIRE D’ÉTUDE, MÉTHODES
Limites géographiques
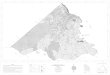
La surface étudiée couvre l’aire de répartition dusapin dans le massif du Jura. La limite des sapi-nières correspond au rebord du second plateauà 650 m d’altitude. Les précipitations moyennesannuelles dépassent 1 200 mm. L’ensemble des208 peuplements échantillonnés s’étend entre
Delémont (en Suisse, 30 km au SE de Bâle) etle lac du Bourget (35 km au Nord de Chambé-ry), soit une surface de 230 x 50 km (fig 1). Lapartie française du massif compte 144 sites : 77dans le département du Doubs, 41 dans le Juraet 26 dans l’Ain; les 64 sites suisses se parta-gent entre les cantons du Jura (11 placettes), deBerne (13), de Neuchâtel (19) et de Vaud (21 ).
Échantillonnage,observations phytoécologiques
L’échantillonnage n’a concerné que les stationssur lesquelles les peuplements forestiers com-

portaient plus de 50% de sapin; toutes les asso-ciations végétales ne pouvaient donc pas êtrerencontrées. Le choix des placettes a été fait enrespectant la variabilité des conditions station-nelles au sein d’une même petite région. Lespeuplements de sapins échantillonnés couvrentune large gamme d’âges, afin de pouvoir réali-ser une étude dendrochronologique. Chaqueplacette d’une surface de 400 m2 comportait 6sapins dominants sur lesquels diverses me-sures dendrométriques et observations sani-taires ont été réalisées. La défoliation (pourcen-tage d’aiguilles absentes par rapport à un sapinsain de même morphologie) et le jaunissement(pourcentage d’aiguilles jaunes dans le houp-pier) ont été appréciés par le même notateursur les 1 248 sapins. L’état sanitaire des sapinsn’a pas conditionné leur choix pour conserverune bonne représentativité de l’échantillon vis-à-vis du degré de dépérissement de l’ensembledes sapinières.
Un relevé phytoécologique a été réalisé surchaque placette. La composition floristique aété notée en nommant toutes les espèces selonla nomenclature de Flora Europaea (Halliday etBeadle, 1983) et en notant leur abondance-dominance selon la méthode «Braun-
Blanquet». Le peuplement a été brièvement ca-ractérisé par son type de traitement en «futaierégulière» ou «futaie jardinée». Le site a été dé-crit par : les coordonnées géographiques, l’alti-tude (mesurée avec un altimètre, précision =10 m), la pente (en degrés), l’azimut magnéti-que du haut de la pente, le masque (pente de ladroite joignant la placette au sommet du versantopposé); ces 3 dernières valeurs ont été inté-grées en un «indice de climat radiatif» (Becker,1982) qui varie dans l’échantillon entre 0,25(stations «froides») à 1,25 (stations«chaudes»). La position topographique est
notée comme «sommet», «haut de pente», «mi-pente», «plateau», «bas de pente» ou
«dépression». Le sol a été décrit grâce à unefosse pédologique ou un prélèvement à la ta-rière, selon sa charge en éléments grossiers;les descripteurs principaux sont : la profondeurde décarbonatation, la profondeur de sol, le
type de substrat classé en «marne», «banc cal-caire compact», «banc diaclasé», «banc
concassé», «éboulis fins», «éboulis grossiers»et «moraines». Ces éléments permettent de rat-tacher le sol aux types décrits par Gaiffe et
Schmitt (1980), Bruckert et Gaiffe (1980) en sui-vant la nomenclature de la classification des
sols CPCS (1967). Le rattachement a été par-fois compliqué par la disposition des sols en mo-saïque : le type de sol varie sur une courte dis-tance selon la microtopographie et la structuredu substrat sous-jacent. De même, l’abondancede cailloux dans les profils ont rendu délicate lamesure de profondeur de sol. Afin d’harmoniserles dénominations, nous avons également expri-mé les noms de sols selon la nomenclature duRéférentiel pédologique français (Baize et Gi-rard, 1990). La fosse pédologique la plus carac-téristique de chaque type de sol a fait l’objet deprélèvements et d’analyses; les résultats de
l’analyse granulométrique ont permis de calculerla réserve en eau moyenne.
Méthodes d’analyse
Les relevés floristiques en abondance-dominance ont été soumis aux analyses multiva-riées : analyse factorielle des correspondances(AFC) et classification ascendante hiérarchique(CAH) du logiciel SPAD N (Lebart et al, 1988).
L’analyse factorielle des correspondances estessentiellement un mode de représentation gra-phique du tableau de contingence constitué parles 208 relevés et les 199 espèces caractériséespar leur coefficient d’abondance-dominance (Le-bart et al, 1977). Elle utilise 2 types de variables.Les variables actives (ou principales) déterminentles axes; ce sont les 199 espèces de cette étude.Les variables supplémentaires ne participent pasau calcul des valeurs propres mais elles peuventêtre représentées sur les plans factoriels commele barycentre d’un groupe de relevés. Dans le casde notre étude, les relevés ont été regroupésselon 9 tranches d’altitude, puis le point moyen dechaque groupe a été projeté sur l’AFC. La mêmeprocédure a été employée pour les différentes va-riables stationnelles : type de sol, profondeur desol, indice de climat radiatif. L’agencement des npoints moyens correspondant aux n classes d’unevariable au long d’un axe permet de conforter soninterprétation écologique.
L’analyse factorielle permet d’établir la cor-
respondance entre le nuage de points des rele-vés et celui des espèces. Dans un premiertemps, l’interprétation de l’agencement des es-pèces les unes par rapport aux autres révèle lesfacteurs écologiques essentiels qui structurentla flore. Ceux-ci sont mis en évidence en obser-vant le mode de regroupement des espèces,

dont on connaît le caractère indicateur, quicontribuent le plus à la création d’un axe. Le ca-ractère indicateur de chaque espèce est révélépar l’observation de ses profils écologiques enfonction des différentes variables stationnelles.Les variables supplémentaires confortent l’inter-prétation des axes réalisée à partir des es-
pèces. Dans un second temps, le regroupementdes placettes est interprété à l’aide de cesmêmes facteurs.
Si les relevés ont été faits dans des milieuxnettement différents et distants les uns desautres, les groupes de relevés se séparent aisé-ment sur les plans factoriels. Au contraire, si lesrelevés ont été réalisés au sein de gradientsécologiques continus, au long d’un transect parexemple, les plans factoriels à 2 dimensionsmontrent un nuage de points homogène : les re-levés ne sont pas individualisés en groupes.Dans ce cas, il est nécessaire de réaliser uneclassification automatique des relevés.
La classification ascendante hiérarchiquefournit un ensemble de classes de moins enmoins fines obtenues par regroupement suc-cessif de parties. Le dendrogramme, ou arbrede classification, est obtenu de manière ascen-dante : on regroupe d’abord les 2 relevés floris-
tiquement les plus «proches» qui forment un«sommet», il ne reste alors plus que (n-1) ob-jets et le processus est réitéré jusqu’à regrou-pement complet (Saporta, 1990). L’algorithmede SPAD N fonctionne avec le critère d’agréga-tion de Ward qui utilise la distance euclidienne.Les calculs peuvent être effectués sur les rele-vés floristiques, ce qui est objectif mais en-globe les variations aléatoires de leur composi-tion, ou sur les premiers axes de l’AFC. Dansce cas, la classification ne prend en compteque l’information de ces axes et ignore les fac-teurs non interprétables ou de faible poids.Nous avons effectué la CAH sur les 10 pre-miers axes de l’AFC.
La qualité de l’appartenance d’un relevé à ungroupe issu de la classification hiérarchique aété chiffrée à l’aide d’une nouvelle méthode : laclassification par «ensembles flous». Le module
Fuzzy contenu dans le logiciel Modulad (Du-pouey, 1989) fonctionne selon ce principe. Laclassification par «ensembles flous» affecte à
chaque relevé une valeur d’appartenance (va-riant entre 0 et 1) à chacun des groupes floristi-ques. Avant cette analyse, les relevés sont re-groupés selon les résultats de la classificationhiérarchique; après calculs, les relevés
«typiques» de chacun des groupes peuvent êtredistingués des relevés «hybrides», situés entreles groupes (Dupouey, 1985; Equihua, 1990).
Pour situer notre typologie par rapport auxtravaux antérieurs et rattacher ses unités sta-tionnelles à des descriptions antérieures, 36 re-levés issu de la littérature ont été inclus dans
l’analyse factorielle des correspondances en
tant que relevés supplémentaires (Richard,1961; Simmeray, 1976; Gillet et al, 1984). Cesindividus supplémentaires ne peuvent pas modi-fier l’analyse; leur projection sur le plan formépar les 2 premiers axes de l’AFC est réalisée enprenant pour référence l’ensemble des relevéspropres à la présente étude.
RÉSULTATS
Interprétation de l’analyse factorielledes correspondances
Les 3 premiers axes expliquent 14,4% dela variance du nuage de points. Ils ont puêtre interprétés en termes de facteurs éco-logiques qui influent sur la végétation.
Axe 1 (6,2% de la variance du nuage)
Sur cet axe, les espèces dont le profil éco-logique indique une préférence pour lesfaibles altitudes se localisent du côté des
plus faibles abscisses : Carpinus betulus,Cornus sanguinea, Crataegus laevigata,C monogyna, Fraxinus exelsior, Ligustrumvulgare, Quercus petraea. Au contraire, lesespèces à caractère indicateur monta-
gnard correspondent aux plus fortes ab-scisses : Adenostyles alpina, Asplenium vi-ride, Lonicera alpigena, Orthilia secunda,Ranunculus aconitifolius, Rubus saxatilis,Vaccinium myrtillus. Par ailleurs, la projec-tion du point moyen des 9 classes d’alti-
tude montre un agencement très net aulong de l’axe 1 (fig 2). La corrélation entrela coordonnée sur l’axe 1 d’une placette etl’altitude est significative à 0,1% (r = 0,80).

Ce premier axe est donc interprété commeun gradient climatique lié à l’altitude du site.
Axe 2 (4,6% de la variance du nuage)
Les espèces indicatrices de sols riches ensquelette calcaire (sols humo-calcaires,humo-calciques) ou d’une faible décarbo-natation possèdent les plus fortes coor-données sur l’axe 2, et inversement pourles espèces les plus acidiphiles, ce quisemblerait indiquer un gradient trophique.Cependant, l’étude de la répartition desespèces sensibles au régime hydrique dusol a révélé le véritable facteur écologiqueexprimé par cet axe : un gradient d’humidi-té, ou de bilan hydrique de la station. Lebilan hydrique a été estimé grâce à la pro-fondeur de sol, la position topographique,la pente de la station ou la perméabilité dela roche mère. Les 2 hypothèses sont ce-pendant compatibles car les sols calcairessont également les moins épais (30-50cm environ) et les plus secs, tandis queles sols bruns plus acides sont épais (60 àplus de 110 cm) et présents sur des sta-tions où le drainage est inférieur aux ap-ports d’eau. La projection des pointsmoyens des variables stationnelles met enévidence les mêmes résultats, avec moinsd’efficacité (fig 2). D’autres résultats, por-tant notamment sur l’analyse de la crois-sance en hauteur des sapins, ont confirménettement l’interprétation de l’axe 2 grâceau bilan hydrique (Bert, 1992).
Axe 3 (3,6% de la variance du nuage)
Les variables stationnelles ne permettentpas une interprétation satisfaisante del’axe 3. On a cherché à voir si la positionrelative des points relevés le long de cetaxe ne mettait pas en évidence une struc-
turation d’ordre géographique à laquellecorrespondrait un déterminisme écologi-que. Les placettes dont la coordonnée sur
l’axe 3 est négative se localisent géogra-phiquement sur la bordure ouest du Jura;les placettes avec des coordonnées posi-tives se localisent à l’est du massif. Par
ailleurs, la répartition géographique desespèces au sein du massif permet de lesrattacher à 5 principaux types :- espèces collinéennes sur le pourtour duJura : Acer campestre, Crataegus laeviga-ta, Ligustrum vulgare, Quercus petraea;- espèces montagnardes sur la Haute-Chaîne : Lonicera alpigena, Moehringiamuscosa, Ranunculus aconitifolius, R lanu-ginosus, Valeriana montana, Veronica urti-cifolia;- espèces xérophiles au sud : Carex alba,Coronilla emerus, Laburnum anagyroides,Teucrium scorodonia;- espèces continentales situées préféren-tiellement au nord-est : Lilium martagon,Maianthemum bifolium, Prunus avium,Sorbus mougeoti, Stachys officinalis;- espèces mésophiles ou hygroclines, sub-océaniques, plus abondantes dans le quartnord-ouest : Carex remota, Impatiens noli-tangere, Glechoma hederacea, Senecio
fuchsii, Veronica montana...
L’axe 3 révèle donc un gradient decontinentalité. Il apparaît au travers du dis-positif car le Jura est soumis aux 3 grandstypes d’influences climatiques et de cou-rants de migrations végétales : méditerra-néenne, atlantique et continentale.
Structuration des relevés
L’AFC a montré que la composition floristi-que des groupements végétaux dépendprincipalement de l’action conjuguée del’altitude, de l’humidité du sol et de leur po-sition géographique. La classification as-cendante hiérarchique a débord séparé lesrelevés d’altitude des stations de basse al-titude. Au sein de ces dernières, les sta-

tions sèches ont été séparées des stationshygrosciaphiles; enfin, ces dernières ontété subdivisées en hêtraies à dentaire et
hêtraies-sapinières (fig 3). Finalement, 11 unités stationnelles ont été identifiées et
ont pu être rattachées à des unités dé-crites par Moor (1968), Gaiffe et Schmitt
(1980), Rameau et al (1980) et Rameau(1987).
La projection des groupes de relevéssur le plan formé par les 2 premiers axesde l’AFC (en 2 dimensions) montre des en-sembles assez enchevêtrés. Pour en épu-rer la représentation, nous avons transfor-

mé ce plan en un diagramme qui combineles gradients altitudinal et hydrique; ilmontre l’amplitude de répartition écologi-que des diverses associations (fig 4).
Les facteurs essentiels qui structurentla végétation ont été utilisés pour présen-ter les unités stationnelles forestières sousforme d’un tableau floristique (tableau I).Les relevés ont été ordonnés en fonctionde l’altitude, exprimée par leur coordonnéesur l’axe 1. Les associations les plus colli-néennes sont sur la gauche du tableau etles plus montagnardes sont à l’autre extré-mité. Le découpage présenté dans le ta-bleau a été conditionné par la taille del’échantillon réalisé; les unités pourraientêtre plus finement décomposées en aug-mentant le nombre de points de relevés.
Les 4 groupes d’espèces individualiséspar une CAH correspondent globalementaux étages bioclimatiques submonta-
gnard, montagnard inférieur, montagnardmoyen, montagnard supérieur/subalpin.Plus précisément, certaines espèces sonteffectivement bien liées à une variationd’altitude tandis que le classementd’autres espèces dans un «étage» est dûà l’échantillon de relevés disponibles pourcette étude. Ce gradient a cependant étéconservé pour classer les espèces dans letableau pour mettre en évidence les affini-tés entre espèces aux diverses altitudes.Au sein de chaque étage, les espèces ontété ordonnées par groupes écologiques,en fonction d’un gradient hydrique (pré-pondérant) et trophique, comme l’indiquela légende au bas du tableau. Les es-
pèces différentielles de chaque groupesont encadrées.
Associations végétaleset unités stationnelles
La description des différents groupes com-porte leurs principales différentielles floris-
tiques, écologiques et pédologiques. Lesassociations végétales sont nomméesselon la nomenclature européenne reprisepar Gillet et al (1984). Elles sont présen-tées selon le gradient principal d’altitudecroissante. Le tableau II donne le résultatdes analyses effectuées sur les 10 sols lesplus typiques de l’échantillon.
Hêtraie à laîches
Cette association appartient au Carici-
Fagetum Moor 1952. Elle s’individualise
floristiquement grâce à la présence d’es-pèces de «faible» altitude (étages submon-tagnard et montagnard inférieur), calcari-
coles, calcicoles, neutrocalcicoles et
calciclines : Carex alba, Mercurialis peren-nis (caractérisent le groupement), Acer
campestre, Carex flacca, Clematis vitalba,Cornus sanguinea, Crataegus monogyna,Daphne laureola, Ligustrum vulgare, Loni-cera xylosteum, Melica mutans, Sesleria
albicans, Teucrium scorodonia. Les pla-cettes qui représentent la hêtraie à laîchess’étagent principalement entre 580 et 950m d’altitude (fig 5), de préférence sur lesversants chauds. Elles sont situées sur
des éboulis grossiers, en position de mi-pente sur des pentes moyennes à fortes(10-25°).
La hêtraie à laîches se rencontre surdes sols humifères carbonatés ou peu dé-carbonatés : humo-calcaire ou humo-
calcique. Les sapins de ce milieu montrentun important manque d’aiguilles dans leurhouppier, en moyenne 21%, et les plusforts jaunissements du feuillage.
Hêtraie à laîches neutrophile
Richard (1961) a distingué 2 sous-
associations de hêtraie à laîches : le Cari-
ci-Fagetum caricetosum albae (Moor,1952), caractérisé par Carex alba et que

nous avons nommé «hêtraie à laîches», etune sous-association légèrement acido-cline de la hêtraie à laîches : le Carici-
Fagetum caricetosum montanae, caractéri-sé par Carex montana. L’analyse a distin-gué ce second groupement grâce à la pré-sence d’espèces neutrocalcicoles, calci-
clines, neutroclines ou acidiclines : Athy-rium filix-femina, Carex sylvatica, Dryopte-ris dilatata, D filix-mas, Hylocomium splen-dens, Hordelymus europaeus, Rubus sp,Rhytidiadelphus loreus, Tilia platyphyllos,Thuidium tamariscinum. Cette sous-
association se rencontre à l’étage submon-

tagnard/montagnard inférieur entre 700 et860 m. Le substrat est constitué par desbancs calcaires diaclasés ou concassés,ou des éboulis grossiers, en positions to-pographiques variées (srutout en mi-
pente) sur des pentes faibles de 3-16°.
Cette dernière caractéristique expliquel’absence de colluvionnement des sols
humo-calciques ou humo-calcaires. La
perméabilité du substrat confère un carac-tère relativement xérique aux stations dehêtraie à laîches : précipitations de 1 300 à

1 500 mm, réserve estimée en eau utile de70 mm. Les sapins dominants sont sou-vent parasités par du gui (Viscum album)et jaunissants. Les 24 sites qui représen-tent la hêtraie à laîches n’ont pas permisde distinguer les 15 sous-associations dé-crites plus récemment par Moor (1972).
Hêtraie à seslérie
L’association Seslerio-Fagetum a été dé-crite par Moor (1952); elle est caractériséepar Sesleria albicans. Elle se rencontre surles stations les plus sèches, sur des solscarbonatés superficiels. Un seul site la re-présente dans notre échantillon et nous
l’avons regroupé avec ceux de la hêtraie àlaîches.
Hêtraie à tilleul
Le Tilio-Fagetum Moor 1952 se rencontresur pente forte (12-35°) et peu stabilisée,en position de haut de pente ou de mi-pente, entre 800 et 1 050 m. La flore est
marquée par la présence d’espèces calci-coles : Acer campestre, Daphne laureola,Helleborus foetidus, Melittis melissophyl-lum, Melica nutans, Tilia platyphyllos.L’abondance d’espèces neutroclines et
neutronitroclines la distingue de la hêtraieà laîches neutrophile : Acer pseudoplata-nus, Euphorbia amygdaloides, Fissidens
taxifolius, Lilium martagon, Phyteuma spi-catum, Prenanthes purpurea, Solidago vir-gaurea. Les stations sont exposées aunord-ouest ou au nord-est et plutôt au nordsi l’altitude est faible.
Les substrats sont drainants : banc cal-caire concassé ou éboulis. Les sols sont le
plus souvent du type humo-calcaire collu-vial (Calcosol humique caillouteux sur
éboulis, selon le Référentiel pédologique)ou humocalcique; ils renferment peu deterre fine entre les éléments du squelettecalcaire, ce qui les distingue des sols de la
hêtraie à dentaire. Ce type de sol estformé de 2 horizons très caillouteux (plusde 65% en poids) riches en matière organi-que, aux pH élevés (pH eau = 7,7 dansl’horizon A1 et 8 dans le A1C); la terre finefait effervescence sous l’action de l’acideet contient 50 à 100 g de CaCO3 total parkg. Les sols humo-calcaires ont une épais-seur de 30 à 55 cm et leur réserve en eauest très faible : environ 60 mm.
Hêtraie à dentaire
Les stations de hêtraie à dentaire font par-tie du Dentario-Fagetum Moor, 1952; ThMüll 1966. L’association est caractériséepar Fagus sylvatica, Cardamine heptaphyl-la et Actaea spicata. Les sols à réserveassez importante expliquent le caractère
mésophile de la flore de ces stations : Acerpseudoplatanus, Bromus ramosus, Carexsylvatica, Circaea lutetiana, Fraxinus exel-sior, Geranium robertianum, Paris quadri-folia, Prenanthes purpurea, Sambuscus
nigra, Sanicula europaea, Plagiomniumundulatum. Ce groupement est très répan-du à l’étage montagnard moyen, entre 800et 1 050 m, sur des pentes moyennes de6-22°. Il se rencontre principalement surdes substrats calcaires drainants (bancdiaclasé ou concassé, éboulis) qui permet-tent l’existence de sols bruns calciques(Calcisol argileux, selon le Référentiel pé-dologique). La flore traduit le caractère cal-cicole ou neutrocalcicole du milieu : Carda-mine heptaphylla, Mercurialis perennis,Mycelis muralis, Fissidens taxifolius. Les
précipitations, comprises entre 1 400 et
1 600 mm/an, et réparties régulièrement,fournissent l’humidité nécessaire à la crois-sance du hêtre. La moyenne annuelle des
températures oscille entre 7 et 8 °C (Sim-meray, 1976).
Le sol brun calcique est décarbonatésur la majeure partie de son profil. L’hori-zon A1, de 15 cm, est limoneux-argileux et

contient 15% de cailloux calcaires, son pHest 6,6; l’horizon (B), de 15 cm, est plus ar-gileux et plus caillouteux (45% en poids).La base du profil est un horizon de 10 cmen farine calcaire argileuse. La réserve eneau estimée est meilleure que celle dessols humifères : 75 mm d’eau utile.
L’analyse factorielle des correspon-dance a regroupé avec les relevés de hê-
traie à dentaire quelques relevés localiséssur des moraines au pied du Jura suisse.Ces sites montrent une flore plus acidi-
cline : Galeopsis tetrahit, Galium rotundifo-lium, Milium effusum, Maianthemum bifo-lium. Les résidus granitiques ou
gneissiques des moraines d’origine alpinepermettent l’existence de sols bruns acides(Brunisol désaturé sur moraines acides).

La végétation qui recouvre ce type de solest à rapprocher de la hêtraie à millet étaléou Milio-Fagetum Frehner, 1963. Elle peutse rencontrer sur des placages de limonsrecouvrant le substrat calcaire, limons à
l’origine de sols bruns lessivés (Rameau,1988). De même, quelques relevés effec-tués sur des moraines alpines présententdes ressemblances avec la hêtraie à lu-
zules (Luzulo-Fagetum Meus, 1937), quise distingue des hêtraies calcicoles par Lu-zula luzuloides, Luzula nivea, Veronica of-ficinalis et Galium rotundifolium (Richard etFavarger, 1960). Leur faible nombre nenous a pas permis d’individualiser ce
groupe.Le sol brun acide est épais (75 à plus
de 110 cm), acide (pH eau A1 : 4,8) car il

est constitué par un horizon (B) de 70 cmissu de dépôts morainiques acides defaible taille granulométrique : 45% desables et 37% de limons dans la terre fine.Cette composition lui confère une bonneréserve en eau, environ 100 mm d’eauutile.
Nous avons vu au paragraphe Axe 2 del’Interprétation de l’analyse factorielle descorrespondances que l’axe 2 a été inter-prété comme un gradient de bilan hydri-que croissant allant globalement de pairavec un gradient de pH décroissant. Dansle détail, ce parallélisme n’est pas toujoursvérifié : les sols bruns calciques, peu acidi-fés, côtoient les sols bruns acides au
centre du nuage de points projeté sur leplan 1 x 2 de l’AFC. Cependant ce regrou-pement de sites calcicoles avec des sites
acidiphiles peut se comprendre en consi-dérant la réserve en eau assez semblablede ces sols, proche de 75 à 100 mm.
Hêtraie à adénostyle
Quelques relevés, très proches floristique-ment de la hêtraie à tilleul, ont pu être rat-tachés à la hêtraie à adénostyle. L’Adenos-tylo-Fagetum Moor 1971, s’individualise
grâce à quelques espèces neutrocalci-coles ou ne craignant pas les variationsd’humidité du sol : Adenostyles alpina,Carex flacca, Centaurea montana, Hyperi-cum hirsutum, Lathyrus vernus, Stachysofficinalis... Le peuplement est habituelle-ment dominé par le hêtre associé à l’érable
sycomore. Le sapin et l’épicéa peuvent do-miner aux altitudes moyennes et supé-

rieures, ce qui indique que la hêtraie àadénostyle n’est pas une forêt pionnière.Cette association se rencontre entre 900et 1 100 m, sur des éboulis grossiers, desbancs calcaires concassés, en positionsommitale ou de mi-pente, sur des pentes8-27°. Elle constitue un groupement vica-riant altitudinal de la hêtraie à tilleul (Moor,1971).
Les sols humifères sont le plus souventdu type humo-calcique colluvial (Calcisolhumique caillouteux sur éboulis). L’horizonA1 de 20 cm est constitué de 75% decailloux calcaires de 2-20 cm, la terre fineest très organique : 20-30% de matière or-ganique; l’horizon brun A1C de 30 cmcontient 90% de cailloux calcaires demoins de 10 cm de diamètre. La terre finede l’horizon A1, totalement décarbonatée,est constituée d’un complexe argilo-humique à forte capacité d’échange catio-nique pouvant fixer plus de 70 meq de cal-cium pour 100 g de terre, ce qui est lemaximum pour les sols échantillonés. Par
contre, en raison de leur granulométrie,ces sols ont la plus faible réserve en eaupotentielle : 60 mm. Ce groupement serencontre également sur des sols humo-calcaires.
Hêtraie-sapinière
L’Abieti-Fagetum Oberd 1938 em 1957 estreprésenté dans notre échantillon par 53%des relevés. Elle est caractérisée par Fes-tuca altissima et Hordelymus europaeus.La CAH la subdivise en 2 groupes biendistincts : d’une part, les stations d’altitudemoyenne, qui rassemblent les sous-
associations «mésophile» et «hygrocline»;d’autre part un ensemble de stations surdes substrats plus drainants qui consti-tuent la sous-association d’altitude ou
«alticole». Les 2 premières sous-
associations sont constituées de peuple-
ments très sains; la défoliation moyenneest de 15% et le jaunissement est très peufréquent. Les plantations réalisées sur labordure ouest de massif appartiennent àces groupements, de même que les plusbelles forêts du massif : forêt de Levier,forêt de La Joux. La hêtraie-sapinière alti-cole est plus défoliée (19%) et plus jaunis-sante que la moyenne des peuplements.
Hêtraie-sapinière mésophileToutes ses espèces les plus caractéristi-ques appartiennent préférentiellement à
l’étage montagnard inférieur; elle partagece dernier avec la hêtraie à laîches neutro-
phile en se développant dans des milieuxnettement moins secs et moins calcaires,entre 710 et 880 m. C’est pourquoi les es-pèces calcicoles, peu représentées, lais-sent la place aux espèces neutroclines àacidiclines : Arum maculatum, Athyriumfilix-femina, Carex sylvatica, Circaea lute-tiana, Dryopteris dilatata, Glechoma hede-racea, Veronica montana. Cette forme de
hêtraie-sapinière se rencontre surtout surdes plateaux en bancs calcaires diaclaséset parfois sur des éboulis sur pente faible(4-19°). Les précipitations sont de 1 300-1 500 mm à ces altitudes.
Les sols sont des sols bruns de laizines
(dans les fentes des lapiaz), bruns limo-neux et surtout bruns à pellicule calcaire(Brunisol saturé, argilo-limoneux, mésosa-turé en surface). Ce type de sol est consti-tué d’un horizon A1 de 15 cm non calcaire,limono-argileux sans cailloux, à pH 5,6 etd’un horizon (B) de 25 cm argilo-limoneuxqui contient 45% de cailloux calcaires dé-composés par l’eau sur leur surface enune pellicule calcaire d’altération. Malgréce flux calcaire, le sol n’est pas saturé encalcium échangeable : 27 meq pour 100 g.La réserve en eau totale est assez bonne :90 mm. Cette sapinière est la plus riche ensapins de tous les groupes; elle est exclu-

sivement traitée en futaie régulière souslaquelle la végétation est moins abondanteque sous les autres sapinières.
Hêtraie-sapinière hygroclineElle occupe les stations sur lesquelles lebilan hydrique est le plus favorable. Laflore caractéristique comporte des es-
pèces neutroclines, neutronitrophiles etsurtout des espèces mésophiles ou hygro-clines : Anemone nemorosa, Atrichum un-dulatum, Cardamine pratensis, C flexuosa,Carex remota, C pendula, Circaea inter-media, Impatiens noli-tangere, Lysimachianemorum, Plagiomnium affine,P punctatum, P undulatum, Plagiochila as-plenioides. Cette hêtraie-sapinière est
étendue entre 790 et 1 030 m, sur dessubstrats assez imperméables : desmarnes, des bancs calcaires compacts oudiaclasés situés au fond de vastes dépres-sions, sur des plateaux, en position de basde pente ou sur pente faible (3-14°).
Les types de sols qui lui sont associéssont des sols bruns à pellicule calcaire oudes sols bruns eutrophes, les plus fré-
quents étant les sols bruns limoneux (Bru-nisol saturé, limoneux, pachique, mésosa-turé en surface) et les sols bruns lessivésargileux (Brunisol désaturé, luvique, argi-leux).
Les sols bruns limoneux formés à partirde limons éoliens post-glaciaires (Pochon,1978) sont les plus épais de tous, plus de80-110 cm, et ne contiennent pas de cal-caire. Ils sont constitués d’un horizon A1de 15 cm à pH 5 et d’un horizon (B) com-portant 60% de limons et 30% d’argile.Ces caractéristiques leur confèrent la
meilleure réserve en eau : 190 mm.
Les sols bruns argileux comportent unhorizon A1 limoneux brun foncé de 8 cmavec peu de matière organique (5,6%, pH4,5), un horizon A2 de 20 cm brun-jaunâtrelimono-argileux à pH 5,2 sans calcaire et
un horizon (B) de 40 à 80 cm argileux,sans cailloux. Ces sols profonds, situéssur des stations de bas-de-pente ou desdépressions, ont une réserve en eau esti-mée de l’ordre de 110 mm.
Hêtraie-sapinière alticole
Entre 900 et 1 100 m, la hêtraie-sapinières’enrichit en espèces de l’étage monta-
gnard supérieur et en espèces acidiclines :Asplenium viride, Dicranum scoparium,Festuca altissima, Hylocomium splendens,Moehringia muscosa, Orthilia secunda,Rhytidiadelphus triqueter, R loreus, Ribesalpinum, Rosa pendulina, Rubus saxatilis,Vaccinium myrtillus. Ce groupement se lo-calise sur les substrats perméables :bancs calcaires diaclasés ou concassés,éboulis fins ou grossiers, sur des pentesde 0 à 22°. Les précipitations s’élèvent à1 600-1 900 mm et la températuremoyenne est 6-7 °C.
Deux variantes se distinguent en fonctiondu type de sol : la majorité des sols sont hu-mifères (variante à sols humifères), particu-lièrement humo-calciques sur banc concas-sé (Calcisol humique, caillouteux, argileux,sur calcaire concassé); on peut égalementrencontrer des sols bruns à pellicule calcaireou bruns de laizines (variante à sols brunseutrophes superficiels). Les sols humo-
calciques sur banc concassé sont constituésd’un horizon LA0 de 5 cm, organique, fi-
breux, brun-rougeâtre qui comporte 90% decailloux calcaires anguleux; l’horizon A1 de20 cm gris très foncé est également riche encailloux de faibles dimensions, la terre fineest très organique (25-40% de matière or-ganique), son pH est de 7,5; l’horizon A1Cde 20 cm est un peu plus clair, plus argileuxque son homologue du type «colluvial» maiségalement décarbonaté et riche en calcium(60-70 meq). Ce type de sol, rencontré surdes bancs calcaires purs, durs et très fine-ment concassés, présente une mauvaise ré-serve en eau : 60 mm.

L’étage montagnard supérieur est le do-maine de 2 associations dans lesquelles lesapin laisse progressivement la place àl’épicéa : la hêtraie à érable et la pessièreà doradille. Les précipitations totales va-rient entre 1 800 et 2 600, et la tempéra-ture moyenne est inférieure à 5 °C.
Hêtraie à érable
Cette association dominée par le hêtre etl’érable sycomore se rencontre entre 1 100et 1 300 m, en situations froides. Les es-
pèces à tempérament montagnard carac-térisent ce groupement de l’Aceri-FagetumBartsch 1940 : Festuca altissima, Moehrin-gia muscosa, Polystichum aculeatum, Ra-nunculus aconitifolius, R lanuginosus,Rosa pendulina, Rubus saxatilis, Saxifragarotundifolia, Valeriana montana, Veronicaurticifolia. Deux sous-associations se dis-
tinguent selon le caractère plus ou moinsmésotrophe de la flore, en relation avec letype de sol.
Hêtraie à érable neutrophileSur les substrats drainants (bancs concas-sés, éboulis) à sols humifères sur pentesassez fortes (5-32°). Les espèces différen-tielles calcicoles, neutrocalcicoles ou neu-troclines peuvent s’accommoder de solsassez secs : Corylus avellana, Festuca al-tissima, Lonicera alpigena, Moehringiamuscosa, Sorbus aria, Valeriana montana,Veronica urticifolia. Cette sous-associationse rencontre sur des sols assez sem-
blables à ceux de la hêtraie-sapinière alti-cole mais, en plus de certaines différencesfloristiques, elle se localise sur des pentesplus prononcées et à des altitudes supé-rieures (200 m plus haut en moyenne).Cette dernière caractéristique est reflétéedans la flore par un enrichissement en es-
pèces montagnardes : Acer pseudoplata-nus, Adenostyles alpina, Lonicera alpige-na, Saxifraga rotundifolia, Senecio fuschii,
Valeriana montana, Veronica urticifolia.Les sapins de cette unité ont montré lesplus fortes défoliations (22,6% en
moyenne) et le jaunissement est juste unpeu inférieur à celui des hêtraies à laîches.
Hêtraie à érable mésotropheSur les substrats moins drainants couverts
par des sols habituellement plus désatu-rés : bruns à pellicule calcaire, bruns calci-ques ou bruns de laizines. Les espècessont neutrophiles à acidiclines et certainesd’entre elles indiquent un bilan hydriqueplus favorable que pour l’autre sous-
association : Athyrium filix-femina, Carexsylvatica, Fissidens taxifolius, Geum urba-num, Lysimachia nemorum, Plagiomniumundulatum, Paris quadrifolia, Plagiochilaasplenioides, Primula elatior, Ranunculuslanuginosus. Quelques plantes de méga-phorbiaies, telles que Cicerbita alpina oules renoncules peuvent s’y développerconsidérablement.
Pessière à doradille
Cette association de l’Asplenio-PiceetumKuoch 1954, se rencontre sur les plateauxde haute altitude (1 150 à 1 310 m) for-mant des lapiaz : bancs calcaires décou-pés en gros blocs. L’épicéa est davantageprésent qu’au sein des autres groupe-ments, ainsi que Asplenium viride (petitefougère : la doradille). Les espèces acidi-clines ou acidiphiles différencient la sta-
tion : Anthoxanthum odoratum, Dicranumscoparium, Maianthemum bifolium, Melam-pyrum sylvaticum, Polypodium vulgare,Polytrichum formosum, Rhytidiadelphus lo-reus, Vaccinium myrtillus.
Ce milieu constitue une mosaïque desols : sol lithocalcique humifère de quel-ques cm sur les blocs et sol brun de lai-zines (Organosol Folist mésosaturé) dansles fentes (laizines). Le sol brun de laizinesest constitué d’un humus de type mor sur

un horizon A11 de 8 cm brun foncé, richeen racines fines, myceliums et en matièreorganique (20%), à pH 5; l’horizon A12 de15 cm est un peu plus compact et contientégalement 75% de blocs calcaires. L’hori-zon (B) brun jaunâtre de 25 cm est plus ar-gileux et renferme plus de cailloux cal-caires empilés entre lesquels descendentles grosses racines jusqu’à 50 cm. La ré-serve en eau est estimée à 75 mm. Le
soutirage karstique intense, dû à des pré-cipitations supérieures à 2 000 mm, ap-pauvrit le complexe absorbant en calcium.En raison du faible effectif, les 2 variantesoligotrophe et mésotrophe qui peuventêtre distinguées selon l’épaisseur de sol,ont été regroupées.
Composition de la strate arborescente
Tous groupements végétaux confondus,l’épicéa et le hêtre se rencontrent dans lastrate arborescente de 75% des peuple-ments échantillonnés. L’érable sycomoreest présent sur 24% des sites. Les autresespèces arborescentes sont nettement
plus rares dans l’étage dominant : le frêne(fréquence : 11 %), le sorbier des oiseleurs(5,3%), l’alisier blanc (2,4%), le chêne ses-sile, le tilleul à grandes feuilles, l’érable
plane (moins de 2%)...En raison de l’autoécologie des es-
sences et des interventions sylvicoles, la
proportion de feuillus dans les peuple-ments de sapins change avec l’altitude;elle est maximale en dessous de 800 m etau-dessus de 1 100 m. Le caractère plusou moins montagnard des espèces arbo-rescentes, et sa prise en compte par desgestionnaires forestiers, conditionnent la
composition des peuplements au long dugradient d’altitude : Abies alba, Fraxinus
excelsior, Quercus petraea, Carpinus be-tulus, Acer campestre, A platanoides seraréfient ou disparaissent tandis que Picea
abies, Fagus sylvatica, Acer pseudoplata-nus, Sorbus aucuparia sont davantageprésents aux altitudes élevées.
Les conditions stationnelles et sylvi-coles modulent ces tendances généraleset favorisent localement certaines es-
pèces. Le sapin forme des peuplementspresque purs au sein des hêtraie-
sapinières mésophiles et hygrophiles etdes hêtraies à dentaire; ces groupes ca-ractérisent les milieux les plus mésophileset les altitudes moyennes (750-950 m). Leremplacement du sapin par l’épicéa quandl’altitude augmente est net dans la série
hêtraie-sapinière alticole, hêtraie à érable,pessière à doradille. Au sein de l’échan-tillon, la régénération du sapin est plusabondante dans les groupes formés de
peuplements clairs dans lesquels les sa-pins dominants sont en contact avec leursvoisins sur moins de 30% de la circonfé-rence du houppier; c’est le cas de la hê-traie à laîches et de la pessière à doradille.L’enrichissement en sapin de groupementsqui lui sont peu favorables créé des sylvo-faciès au sein desquels les sapins mon-trent des jaunissements importants; c’estle cas en hêtraie à laîches et hêtraie àérable neutrophile.
Le caractère xérotolérant du hêtre expli-que sa plus grande abondance sur les sta-tions sèches, particulièrement en hêtraie àtilleul et sa rareté en hêtraie-sapinière hy-grophile. L’érable sycomore se rencontre
essentiellement en hêtraie à érable et hê-traie à dentaire. On peut parfois trouver lefrêne en hêtraie à laîches et hêtraie à tilleul.
Phytogéographie
L’interprétation du troisième axe de l’AFC afait appel à la distribution des espèces ausein du massif jurassien, ce qui impliqueque la composition des groupements végé-taux dépend de leur position géographi-

que. Cette influence s’explique par la ré-
partition hétérogène des conditions station-nelles en raison de la géomorphologie dela chaîne plissée calcaire. En effet, le reliefdu Jura est nettement dissymétrique : ducôté français, une succession de plateauxentre 600 et 900 m d’altitude permet d’at-teindre progressivement la Haute-Chaîne
(1 400-1 700 m), tandis que le rebordoriental s’abaisse brutalement sur la plainesuisse (500 m). La moitié nord du massifest essentiellement constituée par les pla-teaux, ceux-ci se réduisent vers le sud
pour laisser la place aux grands plis de laHaute-Chaîne. Cette double dissymétrieconditionne globalement la répartition destypes de sols, des mésoclimats, des es-pèces végétales et, ainsi explique la sépa-ration géographique de certains groupe-ments floristiques (fig 1).
La partie septentrionale peut être divi-sée en 2 zones parallèlement à son axe.La hêtraie-sapinière (climax climatique)constitue de belles forêts sur les grandsplateaux recouverts de limons de la moitiéouest; localement, selon le type de sol, ontrouve la sous-association mésophile ou lasous-association hygrophile. La hêtraie àdentaire ou la hêtraie à tilleul se rencon-trent sur le côté est, formé de plateaux cal-caires. Les pentes fortes et les sols humi-fères carbonatés sont favorables à lahêtraie à tilleul; la hêtraie à dentaire se dé-veloppe sur les sols bruns calciques despentes faibles. La hêtraie à adénostyle selocalise au sein de l’aire couverte par lahêtraie à dentaire, sur les stations froidesen altitude, à forte pente.
Dans la moitié sud, la hêtraie à Carexalba forme des taches sur le bord sud-ouest du massif sur les stations xérocalca-ricoles, où elle constitue un climax station-nel. Deux de nos sites représentent cegroupe sur le versant genevois, où il cons-titue un climax climatique grâce à l’effet defoehn du massif (Gillet et al, 1984). Deux
climax climatiques se rencontrent dans lapartie élevée du massif : la hêtraie-
sapinière d’altitude à l’étage montagnardmoyen, disjointe des autres hêtraie-
sapinières, et la hêtraie à érable à l’étagemontagnard supérieur. Les plateaux boi-sés des sommets de la Haute-Chaîne sontrecouverts par la pessière à doradille;cette association constitue un climax sta-tionnel sur certains sites à plus faibles alti-tude. Elle est principalement localisée surune petite partie du Jura : massif du Ri-
soux, Mont Tendre et Chasseron.
DISCUSSION
Les unités stationnellessont des «ensembles flous»
Bien que l’échantillonnage utilisé pourcette étude n’ait pas été réalisé dans une
optique purement typologique, les princi-paux groupes floristiques comportant dessapins en quantité assez importante ont puêtre identifiés. Le choix du site de relevésa été effectué sans tenir compte du typede station auquel il serait rattaché aprèsanalyse. Cette démarche a eu l’avantagede recueillir une image du milieu proche dela réalité de terrain, ce qui a permis de re-trouver le continuum écologique au seindes données floristiques et écologiques.Par contre, les limites entre les groupes derelevés n’apparaissent pas toujours trèsnettement lors des analyses. La typologieréalisée a donc été évaluée grâce à uneméthode de classification par ensemblesflous.
Avant l’analyse, les relevés ont été dis-tribués dans les 11 groupes décrits plushaut; après analyse, chaque groupecontient encore des relevés lui apparte-nant préférentiellement, ce qui confirme lavalidité de son existence. Les groupes les

plus solides ont conservé tous leurs rele-vés : hêtraie à laîche, hêtraie à tilleul, hê-traie à érable mésotrophe et pessière àdoradille. Par contre, certains relevés ontété réattribués à un autre groupe issu dela classification hiérarchique; ces sites
«hybrides» ont donc une composition flo-ristique intermédiaire entre 2 types de sta-tions. La quantité de relevés hybridesentre 2 groupes donne une indication surleur degré de ressemblance écologique.Généralement, 2 unités stationnelles avecde nombreux relevés hybrides sont géo-graphiquement voisines : une partie desrelevés sont intermédiaires entre la hê-
traie-sapinière mésophile et la hêtraie-
sapinière hygrophile, certains relevés de lahêtraie-sapinière alticole sont redistribuésdans la hêtraie à laîches neutrophile oudans la hêtraie à érable, qui la jouxtent àl’est et à l’ouest.
Les relevés les plus typiques d’un
groupe lui appartiennent presque exclusi-vement; sur le plan des 2 premiers axesde l’AFC, ils forment un noyau concentrédans la partie de l’aire de variabilité écolo-gique qui exprime le mieux le facteur res-ponsable de l’individualisation du groupe.Par exemple, la hêtraie-sapinière hygro-phile qui recouvre une large gamme d’alti-tudes (640-1 180 m) et d’humidité du sol,est plus caractéristique entre 750 et 970m, sur les sols les plus humides (bruns li-
moneux ou bruns lessivés argileux). Lamise en évidence des individus d’associa-tions les plus caractéristiques permet deles utiliser pour affiner la description del’ensemble floristique, du type de sol asso-cié, de la fertilité des peuplements.
La grande variabilité stationnelle deshêtraies à dentaire ou des hêtraie-
sapinières laisse penser que, avec un
échantillon plus important, certaines unitésstationnelles pourraient être subdivisées
jusqu’à définir de véritables «types de sta-tions forestières» plus homogènes (Del-pech et al, 1985).
Comparaisonsavec les études antérieures
Les relevés supplémentaires choisis pourreprésenter les divers groupements se
sont presque tous positionnés dans la par-tie du plan correspondant aux milieux rela-tivement secs et plutôt en altitude. Globa-lement, l’ordre des groupes floristiques aété conservé au long des gradients d’alti-tude et d’humidité du sol. Par exemple, les3 sous-associations de hêtraie-sapinière(melicetosum, typicum et adenostyleto-sum) décrites par Simmeray (1976) ont étébien classés des plus basses altitudesvers les plus hautes, mais leur position lesplace au sein de nos hêtraie-sapinières al-ticoles ou hêtraies à érable. En fait, cetteétude avait concerné la région de Saint-Claude et les relevés sont en majorité si-tués entre 1 000 m et 1 300 m, ce qui ex-plique l’absence des hêtraie-sapinière hy-grophile et hêtraie-sapinière mésophile.Cela laisse également penser que notrehêtraie-sapinière alticole pourrait se subdi-viser en une variante à sols humifères
(Abieti-Fagetum melicetosum selon Sim-
meray) et une variante à sols bruns (Abie-ti-Fagetum typicum). La projection des re-levés de hêtraie à laîches, de hêtraie àdentaire, de hêtraie à érable et de pessièreà doradille de Richard (1961) ou de Gilletet al (1984) corrobore notre analyse de lavégétation.
La classification ascendante hiérarchi-
que a individualisé un groupe de 4 relevés
qui ont été regroupés avec la hêtraie-
sapinière hygrophile en raison du faible ef-fectif et des affinités floristiques. Ces sitesont une flore acidiphile qui se développesur des sols bruns lessivés limoneux ou ar-
gileux; ces peuplements peuvent être rat-tachés à la sapinière à prêles (Equiseto-Abietetum, Moor, 1952) qui se développesur les stations les plus humides : fond dedoline, de vallon, dépressions. La projec-

tion de relevés anciens (Richard, 1961) co-ïncide bien avec la localisation de ce
groupe sur le plan des 2 premiers axes del’AFC.
CONCLUSIONS
L’analyse de la végétation des sapinièresjurassiennes a mis en évidence les princi-paux facteurs écologiques qui condition-nent la composition floristique d’un site. Laflore spontanée intègre de nombreuses ca-ractéristiques du milieu, tant climatiquesqu’édaphiques; par le jeu de la composi-tion en espèces et de leur abondance, ellefournit des indications difficiles à obtenir
par des mesures ou des descriptions sta-tionnelles. L’importance de l’alimentationen eau a notamment été bien révélée parla flore alors que son évaluation pédologi-que nécessite des moyens plus lourds.
Les principales unités phytosociologi-ques, à partir du niveau «association»,s’individualisent par la combinaison de l’al-titude et du bilan hydrique du site, qui dé-pend de la position topographique, de la
pente et du drainage. La notion opération-nelle de «type de station» ne correspondpas toujours à celle d’association, surtoutdans le cadre des grandes associations cli-maciques au sein desquelles on peut dis-tinguer de nombreuses sous-unités en
fonction des nuances pédologiques et alti-tudinales.
Cette étude a distingué 11 principalesunités stationnelles qui appartiennent à 6associations : hêtraie à laîche, hêtraie à
tilleul, hêtraie à dentaire, hêtraie-sapinière,hêtraie à érable et pessière à doradille. Lagrande étendue de la surface étudiée dansle massif du Jura a permis d’apprécier lavariabilité écologique et de cerner la répar-tition géographique de chaque unité sta-tionnelle. À travers l’analyse des divers
types de groupements auxquels le sapinpectiné participe, et des conditions écologi-ques correspondantes, cette étude phytoé-cologique a précisé l’écologie du sapin surtoute l’étendue de son aire de répartitiondans le Jura.
Dans la suite de nos travaux, cette typo-logie servira de base pour étudier les rela-tions entre les conditions stationnelles et
l’expression de dépérissements forestiersdans le Jura. En particulier, les peuple-ments de sapins de hêtraie à tilleul mon-trent des signes de dépérissement tandisque les peuplements géographiquement etfloristiquement voisins de hêtraie à den-taire sont dans un état sanitaire plus satis-faisant. Les unités stationnelles seront
également étudiées du point de vue deleur potentialité de croissance en hauteuret de l’évolution de la croissance radialedes arbres.
REMERCIEMENTS
Le travail présenté a constitué un des aspectsde l’étude dendroécologique du dépérissementdu sapin dans le Jura. Nous remercions la Di-rection de l’espace rural et des forêts, le pro-gramme Deforpa et l’INRA, qui ont financé cesrecherches, R Schipfer pour sa collaborationtechnique pendant de la phase de terrain, MBecker et JL Dupouey pour leurs conseils lorsdes analyses, BS Tan pour son aide pédologi-que et 2 lecteurs anonymes pour leurs critiqueset suggestions. Nous sommes reconnaissantsenvers les Services forestiers suisses des 4cantons concernés par l’étude, envers l’Officenational des forêts de Franche-Comté et de l’Ain
pour leur autorisation de carottage et leurs indi-cations sur les peuplements de sapins.
RÉFÉRENCES
Baize D, Girard MC (1990) Référentiel pédologi-que français. Troisième proposition. Ed
AFES-INRA, Plaisir, 279 p

Becker M (1982) Influence relative du climat etdu sol sur les potentialités forestières enmoyenne montagne. Exemple des sapi-nières à Fétuque (Festuca altissima Vill)dans les Vosges alsaciennes. Ann Sci For39, 1-32
Becker M (1987) Bilan de santé actuel et rétros-pectif du sapin dans les Vosges. Étude éco-logique et dendrochronologique. Ann Sci For44, 379-402
Becker M, Lévy G (1988) A propos du dépéris-sement des forêts : climat, sylviculture et vi-talité de la sapinière vosgienne. Rev For Fr40, 345-358
Bert GD (1992) Potentialité du sapin pectiné(Abies alba Mill) et qualité du bois sur lesstations de sapinière dans le Jura (France etSuisse). Rev For Fr (sous presse)
Bert GD, Becker M (1990) Vitalité actuelle etpassée du sapin (Abies alba Mill) dans leJura. Étude dendroécologique. Ann Sci For47, 395-412
Bruckert S, Gaiffe M (1980) Analyse des fac-teurs de formation et de distribution des solsen pays calcaire ou karstique. Ann Sci UnivBesançon, 4e Sér Biol Vég, 19-67
CPCS (1967) Commission de pédologie et decartographie de sols : classification des sols.ENSA, Grignon, 96 p
Delpech R, Dumé G, Galmiche P (1985) Typo-logie des stations forestières. Vocabulaire.IDF, Paris, 243 p
Dupouey JL (1985) Intérêt de la notion d’en-semble flou en phytosociologie forestière.
Application à la classification des relevés devégétation. In: Colloques Phytosiologiques.14, «Phytosociologie et foresterie», 43-53
Dupouey JL (1989) Classification floue non hié-rarchique : le programme Fuzzy. Rev Modu-lad 4, 1-7
Equihua M (1990) Fuzzy clustering of ecologicaldata. J Ecol 78, 519-534
Gaiffe M, Schmitt A (1980) Sols et végétation àl’étage montagnard dans les forêts du JuraCentral. Science du sol. Bull AFES 4, 265-296
Gillet F, Lhote P, Trivaudey MJ (1984) Étudecartographique de la végétation et des mi-lieux naturels du Jura Gessien. Laboratoirede Taxonomie expérimentale et de Phytoso-
ciologie. Doc Univ Besançon, 82 p + an-nexes
Guinochet M (1973) Carte phytosociologiquePontarlier 5-6 au 1/20 000. In: Phytosociolo-gie. Masson, Paris, 227 p
Halliday G, Beadle M (1983) Flora Europaea. 5Vol. Cambridge Univ Press
Lebart L, Morineau A, Tabard N (1977) Techni-ques de la description statistique, méthodeset logiciels pour l’analyse des grands ta-bleaux. Dunod, Paris
Lebart L, Morineau A, Lambert T (1988) SPADN : système portable l’analyse des données.Manuel de référence. Version 1.2. CISIA,Sèvres
Moor M (1952) Die Fagion-Gesellschaften (Bu-chen, Tannen-Buchen und Ahornwälder) imSchweizer Jura. Beiträge zur geobotanischenLandesaufnahme der Schweiz Heft 31. Hans
Huber, Berne, 200 pMoor M (1968) Der Linden-Buchenwald. Vege-
tatio. Acta Geobot Vol 16, Fasc 1-4, 159-191
Moor M (1971) Adenostylo-Fagetum Höhenvika-riant des Linden-Buchenwaldes. Bauhinia. 4
Moor M (1972) Versuch einer soziologisch-systematischen Gliederung des Carici-
Fagetum. Vegetatio 24, 1-3, 31-69Pfister RD (1989) Basic concepts of using vege-
tation to build a site classification system. In:Proceedings-Land classifications based onvegetation: applications for resource mana-gement. Intermountain Research station,Odgen, UT, 22-29
Pochon M (1978) Origine et évolution des solsdu Haut-Jura suisse. Thèse de l’Universitéde Neuchâtel, 190 p
Rameau JC (1987) Contribution phytoécologi-que et dynamique à l’étude des écosys-tèmes forestiers : application aux forêts duNord-Est de la France. Thèse Université Be-
sançon. 340 p
Rameau JC (1988) Les Hêtraies mésoneutro-philes et acidiclines (Milio-Fagetum) du Nord-Est de la France. Doc Phytosoc. Nouvellesérie. Vol 11. Camerino. Univ degli Studi,205-220
Rameau JC, Schmitt A, Bidault M, Gaiffe M
(1980) Végétation et écologie des forêts

comtoises. In: Bull Soc Hist Nat Doubs AssocUNIVERS. N° spécial. 80-116
Richard JL (1961) Les forêts acidophiles du Jura.Étude phytosociologique et écologique. Maté-riaux pour le levé géobotanique de la Suisse.Fascicule 38, Hans Huber Berne. 164 p
Richard JL, Favarger C (1960) Les enclaves devégétation acidophile dans le Jura et le pro-
blème du climax. In : Rapports du sol et de lavégétation. Masson, Paris, 99-109
Saporta G (1990) Probabilités, analyses de don-nées et statistiques. Technip, Paris
Simmeray J (1976) Essai d’interprétation desgroupements végétaux de la région de Saint-Claude en vue d’une synthèse cartographi-que. Thèse Univ Besançon. 214 p