Embed Size (px)
Citation preview

Projet P38 Etude de l’accès fluvial direct à Port 2000 Inventaire de la macrofaune benthique
Rapport de synthèse des deux campagnes 2016

PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
CELLULE DE SUIVI DU LITTORAL NORMAND
(Association régie par la Loi du 1ier juillet 1901)
53, rue de Prony
76600 LE HAVRE CEDEX
Tel. : 02.35.21.71.75 / Fax : 02.35.22.47.50
Rapport de synthèse des deux campagnes 2016
Version définitive
Par
Chloé DANCIE
Pour le
GRAND PORT MARITIME du HAVRE
Consultation n°15 du 19/11/15
Marché subséquent à l’accord cadre du 2 janvier 2014
Avec la collaboration de :
Céline CHAIGNON : Prélèvement, Tri, Détermination, Biomasse
Séverine DUBUT : Prélèvement, Tri, Détermination, Biomasse
Delphie Le THOER : Prélèvement, Tri, Détermination, Biomasse
Remerciements au personnel du navire « Le Marais » pour la préparation et la mise en œuvre du matériel de
prélèvements.
VD. 24.10.16

PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Table des matières
1. INTRODUCTION .............................................................................................................................................. 1
2. CONTEXTE GENERAL ....................................................................................................................................... 2
2.1. Compartiment benthique ...................................................................................................................... 2
2.2. Communautés et Habitats benthiques .................................................................................................. 3
2.3. Indicateurs de qualité écologique et « bon état écologique » .............................................................. 7
2.4. Effet des opérations de dragages .......................................................................................................... 8
3. MATERIELS ET METHODES ........................................................................................................................... 10
3.1. Déroulement des prélèvements et localisation des stations .............................................................. 10
3.2. Analyses au laboratoire ....................................................................................................................... 14
3.3. Analyses des données .......................................................................................................................... 16
4. CARACTERISTIQUES SEDIMENTAIRES ........................................................................................................... 23
5. COMMUNAUTES MACROBENTHIQUES ........................................................................................................ 29
5.1. Distribution et composition ................................................................................................................. 29
5.2. Variations saisonnières ........................................................................................................................ 42
5.3. Caractérisation des assemblages faunistiques et Habitats benthiques .............................................. 44
5.4. Sensibilité des peuplements, indices écologiques, et enjeux .............................................................. 48
6. CONCLUSION ................................................................................................................................................ 50
BIBLIOGRAPHIE ..................................................................................................................................................... 53
LISTE DES TABLEAUX ET FIGURES .......................................................................................................................... 58
ANNEXES ............................................................................................................................................................... 61
ANNEXE 1 : Analyses granulométriques ............................................................................................................... 62
ANNEXE 2 : Liste faunistique ................................................................................................................................. 70
ANNEXE 3 : Résultats faunistiques par station...................................................................................................... 74
ANNEXE 4 : Fiches descriptives des espèces dominantes ..................................................................................... 79

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 1
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
1. INTRODUCTION
L’objectif du projet P38 Etude de l’accès fluvial direct à Port 2000 est notamment de définir une solution permet-
tant un accès direct à Port 2000 pour tous types de bateaux fluviaux et par toutes conditions météo pour effec-
tuer des opérations commerciales.
Dans ce cadre, le Grand Port Maritime du Havre (GPMH) dresse un état initial des différentes alternatives et
souhaitait réaliser des inventaires de la macrofaune benthique afin de caractériser les communautés benthiques
subtidales présentes au sein des sédiments portuaires des bassins concernés par cet aménagement. En effet,
l’endofaune des bassins portuaires du Havre n’a été que très peu étudiée depuis une trentaine d’années (Pro-
niewsky, 1979 ; Breton, 2000, 2005 ; Vincent et al., 2006 ; Vincent, 2009 ; Dancie et al., 2007, 2010, 2011, 2012).
Or, une meilleure connaissance des peuplements benthiques ainsi que de leur habitat que sont les sédiments
superficiels est essentielle dans la mesure où d’une part, la faune benthique fournit une ressource alimentaire
pour certains poissons et d’autre part, la répartition, l’abondance et la persistance des peuplements benthiques
témoignent des diverses perturbations subies par les fonds. En effet, les espèces benthiques, et plus particuliè-
rement certains polychètes, sont des indicateurs des conditions du milieu dans les zones plus ou moins polluées
: ces espèces reflètent non seulement les conditions du milieu au moment de leur prélèvement, mais aussi celles
qui existaient depuis le moment où elles se sont installées dans leur biotope.
Le présent rapport, étude de la Cellule de Suivi du Littoral Normand sous le mandat de l’ONF, dresse l’état des
lieux des peuplements et habitats benthiques des fonds des avant-ports et du long de la digue Sud du port his-
torique et à l’ouest de la digue Nord de Port 2000 à partir des données bibliographiques et des données collectées
lors des campagnes de l’année 2016.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 2
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
2. CONTEXTE GENERAL
2.1. Compartiment benthique Au sein du benthos, le phytobenthos, regroupant les organismes végétaux, est distingué du zoobenthos qui re-
groupe les organismes animaux. Le zoobenthos constitué d’invertébrés (mollusques, annélides, crustacés, échi-
nodermes…) est découpé en six qualificatifs selon des critères de tailles (microfaune, méiofaune, macrofaune)
et de niveau par rapport au sédiment (endofaune, épifaune, suprabenthos). Au sein de cette macrofaune ben-
thique, les annélides polychètes vont dominer en termes de fréquence, abondance et diversité.
Au sein du réseau trophique, la macrofaune benthique constitue un élément indispensable de la chaine alimen-
taire estuarienne. Tout d’abord, la multiplicité de leurs régimes alimentaires les place à différents niveaux de la
chaine alimentaire. Ainsi, une large gamme de régimes alimentaires différents est distinguée avec :
- Les détritivores, qui se nourrissent de matière organique particulaire, détritique ou de débris animaux
(cadavres)
- Les déposivores, qui se nourrissent à partir de matière organique particulaire déposée sur le fond,
fraîche ou détritique
- Les suspensivores, qui se nourrissent à partir des particules nutritives (microphagie) en suspension dans
la colonne d’eau
- Les carnivores.
Ensuite, cette macrofaune benthique est la proie de nombreuses espèces de poissons comme les soles, les flets,
les bars, les plies… La composition des peuplements benthiques va donc régir étroitement la distribution des
peuplements ichtyologiques. De plus, en zone intertidale, les oiseaux côtiers vont également rechercher certains
invertébrés benthiques. Ainsi, en raison de leur rôle essentiel dans la chaîne alimentaire, le moindre changement
de composition faunistique au sein des peuplements benthiques peut induire un changement fonctionnel dans
l’écosystème.
Or, la distribution spatiale, la composition et l’état de ces peuplements benthiques vont être régis par de nom-
breux facteurs comme la nature sédimentaire, la profondeur qui influence l’éclairement, la température, la tur-
bidité… Néanmoins, la texture des sédiments est reconnue comme un des facteurs prépondérants de la distribu-
tion spatio-temporelle de la macrofaune benthique des substrats meubles que ce soit à échelle de temps ou
d’espace (Hermand, 2008).
Mais, ces organismes benthiques sont également exposés aux contaminants chimiques et aux stress hypoxiques
du milieu or leur sédentarité et leur durée de vie leur permettent de refléter les conditions environnementales
proches sur plusieurs années. En effet, le compartiment benthique va intégrer à la fois les variations et les per-
turbations de la colonne d'eau et de la colonne sédimentaire (Dauvin, 1993). Ainsi, chaque contrainte abiotique,
essentiellement les changements climatiques et l’anthropisation sous toutes ses formes, va provoquer au sein
des communautés benthiques un déséquilibre temporaire ou permanent modifiant les conditions physico-chi-
miques, y compris sédimentaires du milieu ce qui va se répercuter alors sur la composition qualitative et quanti-
tative des communautés (Hermand, 2008 ; Ware et al., 2010).
En conséquence, le macrozoobenthos étant considéré comme un bon indicateur des changements naturels et
anthropiques dans les écosystèmes marins (Dauvin, 1993), les instances européennes en 2000, ont adopté
comme outil de diagnostic de l'état des milieux estuariens et côtiers, l'utilisation d'indices biotiques basés sur les
communautés d'invertébrés benthiques (Borja et al., 2000 ; Simboura et Zenetos, 2002 ; Dauvin et Ruellet, 2007
; Blanchet et al., 2008).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 3
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
2.2. Communautés et Habitats benthiques
En termes de connaissance, gestion et conservation des milieux naturels et de la biodiversité, de nom-
breuses typologies existent et sont utilisées tant sur le plan international et européen que national, régional ou
local. Elles se basent sur la nature sédimentaire du substrat et sur les espèces présentes au sein de celui-ci mais
également sur l’étage occupé et les durées d’immersion.
Historiquement, les typologies utilisées dans les études sont :
- NATURA 2000,
- EUNIS (European Natura Information System) 2004, 2008,
- Corinne Biotope.
Ces typologies et les habitats associés sont décrits dans les documents de référence suivants :
- Directive européenne 92/43/CEE du Conseil du 21 mai 1992 concernant la conservation des habitats naturels
ainsi que la faune et la flore sauvages (liste des habitats génériques).
- Manuels d'interprétation des habitats de l'Union européenne. Ils constituent la référence européenne princi-
pale pour l’interprétation des Habitats génériques dans le milieu marin. La version originale a été produite en
octobre 1999 puis actualisée en 2003 et 2007.
- Les Cahiers d’habitats côtiers français (édition 2004), document réalisé au niveau français à partir du manuel
EUR 15, mais qui décline les Habitats génériques en Habitats élémentaires.
- La typologie EUNIS dans sa version 2004 et son dictionnaire de description des habitats établi dans la même
version. Une série de tables de correspondance permettant de jongler entre les différentes versions d’EUNIS
citées dans les divers documents utilisés.
- Divers documents, accompagnés d’une base de donnée, établis notamment par le JNCC (GB), décrivant de ma-
nière détaillée les habitats des îles britanniques, selon une classification propre à ces îles mais proche d’EUNIS.
- La table de correspondance entre EUNIS 2004, la typologie des îles britanniques, les Habitats génériques de la
directive Habitat et les Habitats prioritaires OSPAR établie en 2006 par D. Connor.
- Le document «Guidelines for the establishment of the NATURA 2000 network in the marine environment. Ap-
plication of the Habitats and Birds Directives» (mai 2007) qui propose de nouvelles définitions pour des habitats
dont la définition posait problème (notamment pour les Habitats marins 1110 et 1170), définitions par la suite
insérées dans le manuel - Description of habitats on the OSPAR list of threatened and/or declining species and
habitats (2008).
- Le document « Coastal and Marine Ecological Classification Standard » (CMECS Version III, Madden et al., 2008)
élaboré par NatureServe et NOAA, qui constitue le standard de classification pour les habitats côtiers et marins
des Etats-Unis.
- Le rapport de la deuxième version de la typologie des habitats marins benthiques de la Manche, de la Mer du
Nord et de l’Atlantique (Michez et al., 2015) expliquant de manière détaillée les changements opérés suite à la
mise à jour du référentiel national basé sur la proposition de typologie du REBENT (Guillaumont et al., 2009 et
Bajouk et al., 2011).
En ce qui concerne les habitats génériques et élémentaires de la Typologie Natura 2000, ils sont peu
nombreux. Ainsi, en estuaire de Seine, à proximité du site d’étude, 5 habitats génériques sont observés avec 3

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 4
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
habitats élémentaires pour la partie marine (1110-2 Sables moyens dunaires, 1110-3 Sables grossiers et graviers
et 1110-4 Sables mal triés) et 2 habitats élémentaires dans les fosses de flot (1140-1 Sables de hauts de plage à
Talitres et 1140-3 Estran de sable fin). Dans le cadre de notre étude, les deux stations de l’extérieur, au sud de la
digue, sont situées dans l’habitat 1110-4 Sables mal triés (Figure 1).
FIGURE 1 : HABITATS GENERIQUES ET ELEMENTAIRES DU RESEAU NATURA 2000 DU SITE D’ETUDE.
Le seul système de classification des habitats marins couvrant toutes les eaux européennes et ayant
pour vocation de devenir la référence en Europe fait partie du Système Européen d’Information sur la Nature
(EUNIS, EUropean Nature Information System), développé et géré par l’ETC/NTB (European Topic Center for Na-
ture Protection and Biodiversity) pour l’Agence Européenne pour l’environnement (EEA) et le Réseau d’Observa-
tion pour l’Information Environnementale Européenne (EIONET) (Davies et al., 2004). L’objectif du système EU-
NIS est d’avoir un système commun accepté pour définir l’ensemble des habitats et les nommer, sur l’ensemble
des territoires de l’Union Européenne. Il s’appuie donc sur les travaux britanniques du JNCC (Joint Nature Con-
servation Committee) (Connor et al., 2004), de l’OSPAR (Convention pour la protection du milieu marin de l'Atlan-
tique du Nord-Est) et du CIEM (Conseil International pour l’exploitation de la Mer ; ICES, 2002). Les données de
la classification EUNIS sont disponibles gratuitement sur internet (www.eunis.eea.europa.eu). Elles y sont com-
pilées, entretenues et complétées pour être utilisées dans des rapports environnementaux et soutenir le réseau
européen Natura 2000 (« Directive habitats »).
Jusqu’à présent, la plupart des cartes d’habitats disponibles étaient issues de la numérisation de cartes exis-
tantes, qui ont été converties en types d’habitats EUNIS. La carte présentée est issue de l’actualisation des habi-
tats à l’échelle de la baie de Seine à partir de la synthèse des travaux existants de la dernière décennie et des
dires d’experts et réalisée conjointement par le M2C, la CSLN et le GEMEL-Normandie dans le cadre de la mise à
jour des périmètres ZNIEFF pour la DREAL (Figure 2).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 5
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
L’estuaire de la Seine à proximité du site d’étude comprend 11 habitats traduisant la mosaïque d’habitats de cet
environnement. Les trois habitats principaux d’aval en amont sont les Vases sableuses infralittorales à Melinna
palmata, Magelonna spp. et Thyasira spp., les Sables envasés infralittoraux à Spisula subtruncata et Nephtys
hombergii et les Vases sableuses littorales à Nephtys hombergii, Macoma balthica et Streblospio shrubsolii (tra-
duction typologie EUNIS V2, MNHN, 2015). D’autres habitats sont extrêmement réduits comme les moulières
sur substrat anthropique qui sont situées sur les digues. Dans le cadre de notre étude, les deux stations de l’ex-
térieur, au sud des digues, sont situées dans l’habitat A5.244 Sables envasés infralittoraux à Spisula subtruncata
et Nephtys hombergii.
FIGURE 2 : COMMUNAUTES BENTHIQUES SELON LA CLASSIFICATION EUNIS DU SITE D’ETUDE.
Il existe également la typologie Corine Biotope mais elle est peu appropriée aux habitats marins et lit-
toraux et donc par là-même peu utilisée. Pour information, les habitats présents en zone d’étude sont néanmoins
listés ci-après. Ainsi, en ce qui concerne les Habitats littoraux et halophiles, les travaux de Lavrabre et Fisson
(2013), listent 15 habitats CORINE Biotopes (Rameau et al., 1997) au niveau de l’estuaire marin (salinité > 30 g/l ;
d’Honfleur à la proche baie de Seine) soit :
- 11 – Mers et Océans : 11.121 Eaux littorales et 11.31 Herbiers atlantiques à Zostères
- 13 – Estuaires et rivières tidales soumises à marée : 13.2 Estuaires
- 14 – Vasières et bancs de sables sans végétation
- 15 – Marais salés, prés salés (schorres), steppes salés et fourrés sur gypse : 15.1 Gazons pionniers salés
et 15.2 Prairies à spartines
- 16 – Dunes côtières et plages de sables : 16.11 Plages de sables sans végétation et 16.211 Dunes em-
bryonnaires
- 17 – Plages de galets : 17.1 Plages de galets sans végétation, 17.2 Végétation annuelle des laisses de
mer sur plages de galets et 17.3 Végétation vivace des bancs de galets à Crambe

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 6
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
- 18 – Côtes rocheuses et falaises maritimes : 18.1 Falaises maritime nues
- 19 – Ilot, Banc rocheux et Récifs.
Sur la zone d’étude, en prenant en compte le compartiment « Benthos », l’habitat en place à l’extérieur des
digues serait l’habitat 11.22 Zones benthiques sublittorales sur sédiments meubles.
Parallèlement, à ces systèmes de classification des habitats marins, les connaissances et l’expertise con-
jointe du M2C, de la CSLN et du GEMEL-Normandie ont permis de proposer de nouveaux périmètres ZNIEFF en
mer pour la Normandie et de mettre à jour le cas échéant les emprises géographiques des périmètres ZNIEFF
terrestres à l’interface terre-mer et leurs fiches en baie de Seine (Baffreau et Dauvin, 2015). A partir des données
agrégées depuis plusieurs dizaines d’années dans le cadre de divers programmes ainsi que des listes d’espèces
disponibles dans des publications et rapports scientifiques, il a été possible d’établir une liste d’espèces déter-
minantes ce qui a permis de proposer de nouveaux périmètres ZNIEFF mer sur les secteurs de la Manche centrale
et orientale haut-normande. Ces nouveaux périmètres sont visibles sur la Figure 3. En Baie de Seine, deux péri-
mètres de ZNIEFF marines sont visibles : ZNIEFF marine de type I – 23M000005 SABLES FINS ET VASEUX DE LA
BAIE DE SEINE ORIENTALE et ZNIEFF marine de type II - 23M000004 BAIE DE SEINE ORIENTALE.
Par définition, dans la circulaire n°91-71 du 14 mai 1991 à l’attention des préfets de région, la notion de ZNIEFF
de type I est définie comme suit : « Secteurs de superficie en général limitée, définis par la présence d’espèces,
d’associations d’espèces ou de milieux rares, remarquables ou caractéristiques du patrimoine naturel national
ou régional » et celle de ZNIEFF de type II comme cela : « Grands ensembles naturels riches ou peu modifiés, ou
offrant des potentialités biologiques importantes ».
Dans le cadre de notre étude, les deux stations de l’extérieur, au sud des digues, sont situées dans la ZNIEFF
marine de type II - 23M000004 BAIE DE SEINE ORIENTALE.
FIGURE 3 : ZONES NATURELLES D’INTERET ECOLOGIQUE FAUNISTIQUE ET FLORISTIQUE DU SITE D’ETUDE.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 7
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
2.3. Indicateurs de qualité écologique et « bon état écologique » Un grand nombre d’indices permettent d’évaluer l’état des environnements aquatiques, dont une partie a été
développée en vue de la mise en place de la Directive Cadre Eau (à partir de 2000) qui a imposé le rétablissement
du « bon état écologique » des milieux humides. Une partie d’entre eux est basée sur les communautés ben-
thiques. De nombreuses synthèses répertoriant ces listes d'indices existants sont disponibles (Diaz et al. 2004,
Grall et Coïc, 2005, Borja et al. 2009, Pinto et al. 2009, Martinez-Crego et al. 2010, Pedel et Fabri, 2012).
Au niveau de l’estuaire marin et moyen et de la partie orientale de la Baie de Seine, de nombreux indicateurs
sont utilisés pour décrire l’état écologique des peuplements benthiques. Ces indicateurs reposent sur l’étude des
groupes écologiques (AMBI, BOPA, I2EC, BENTIX), des groupes trophiques (ITI) ou de la diversité (BQI, H’). A partir
des relations entre les indices et les paramètres édaphiques, une synthèse point par point est possible donnant
ainsi une image de l’état moyen des communautés en place (Figure 4).
FIGURE 4 : QUALITE DES COMMUNAUTES BENTHIQUES DE LA PARTIE AVAL DE L’ESTUAIRE ET DE LA PARTIE ORIENTALE DE LA
BAIE DE SEINE ESTIMEE PAR LES INDICES BOPA, ITI, BQI ET SYNTHESE PAR POINT DONNANT UN ETAT MOYEN DES COMMU-
NAUTES (SOURCE : GIP SEINE-AVAL 2010, DAUVIN ET AL. 2009).
FIGURE 5 : ETAT DES PEUPLEMENTS BENTHIQUES SELON L’AMBI POUR LA PERIODE 1979-2006 (SOURCE : GIP SEINE-AVAL
2008, RUELLET ET DAUVIN, 2007).
Dans le cadre de notre étude, les communautés benthiques des deux stations de l’extérieur, au sud des digues,
apparaissent dans un état écologique moyen à l’issue de la synthèse des indices d’évaluation. Mais, selon les
indices, l’état écologique apparait plus ou moins dégradé. Ainsi, le ratio des Polychètes opportunistes et des
amphipodes donne l’image d’un très bon état écologique des communautés en place tandis que l’analyse des
abondances de chaque groupe trophique (Infaunal trophic Index, ITI) et des abondances des espèces sensibles
et tolérantes (valeur de tolérance de chaque espèce, Indice de qualité Benthique BQI) traduisent un état écolo-
gique moyen à médiocre.
Enfin, l’indice écologique basé sur l’étude des groupes écologiques de polluosensibilités (AZTI’s Marine Biotic
Index AMBI ; Figure 5) et la synthèse de celui-ci pour la période 1979-2006 traduit un état écologique bon à
moyen.
Ainsi, au vu des résultats, il apparait nécessaire d’utiliser plusieurs métriques en parallèle afin d’avoir une vue la
plus complète possible de la communauté benthique et de son état écologique.
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 8
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
2.4. Effet des opérations de dragages L’effet majeur des opérations de dragages sur la macrofaune benthique reste l’enlèvement du sédiment super-
ficiel et de la faune inféodée d’où la destruction partielle ou totale des habitats et donc des communautés ben-
thiques en place (réduction entre 30% et 90% du nombre d’espèces, de l’abondance et de la biomasse des inver-
tébrés benthiques ; Newell et al., 1998 ; Boyd et al., 2002 ; ICES, 2001 ; 2009). Le macrozoobenthos est concentré
essentiellement dans les premières dizaines de centimètres de sédiment ce qui correspond généralement à
l’épaisseur de sédiment draguée d’où une défaunation totale.
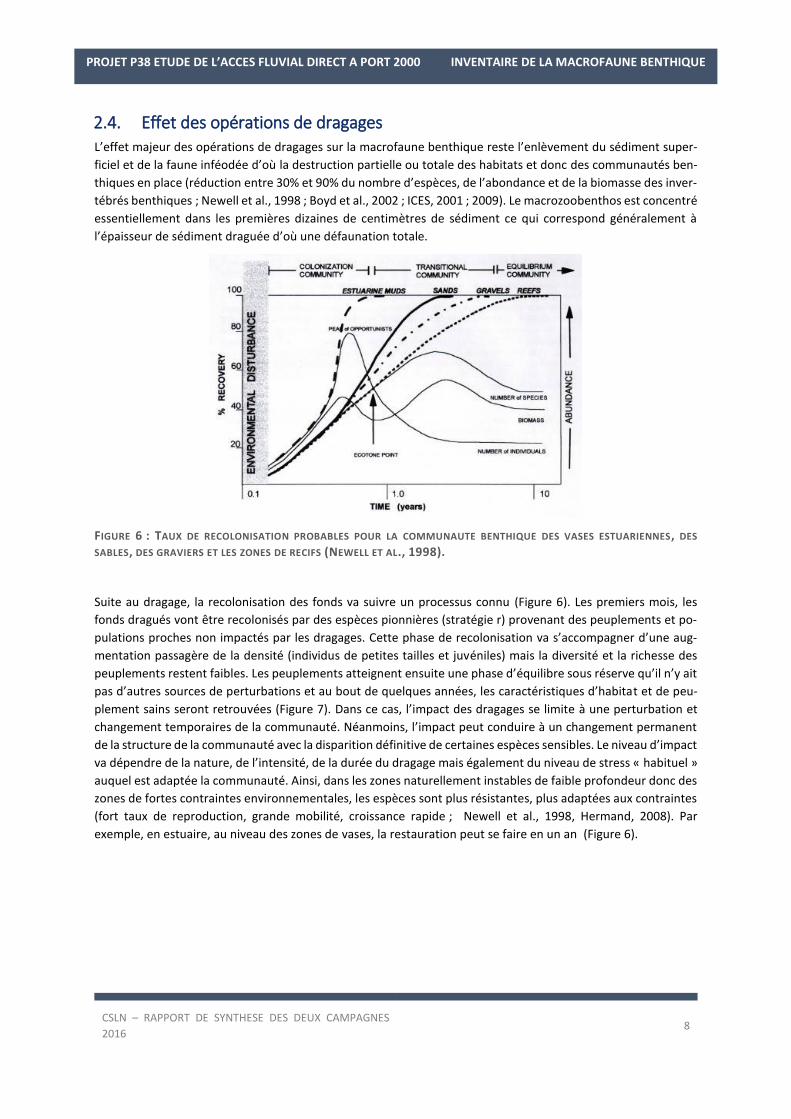
FIGURE 6 : TAUX DE RECOLONISATION PROBABLES POUR LA COMMUNAUTE BENTHIQUE DES VASES ESTUARIENNES, DES
SABLES, DES GRAVIERS ET LES ZONES DE RECIFS (NEWELL ET AL., 1998).
Suite au dragage, la recolonisation des fonds va suivre un processus connu (Figure 6). Les premiers mois, les
fonds dragués vont être recolonisés par des espèces pionnières (stratégie r) provenant des peuplements et po-
pulations proches non impactés par les dragages. Cette phase de recolonisation va s’accompagner d’une aug-
mentation passagère de la densité (individus de petites tailles et juvéniles) mais la diversité et la richesse des
peuplements restent faibles. Les peuplements atteignent ensuite une phase d’équilibre sous réserve qu’il n’y ait
pas d’autres sources de perturbations et au bout de quelques années, les caractéristiques d’habitat et de peu-
plement sains seront retrouvées (Figure 7). Dans ce cas, l’impact des dragages se limite à une perturbation et
changement temporaires de la communauté. Néanmoins, l’impact peut conduire à un changement permanent
de la structure de la communauté avec la disparition définitive de certaines espèces sensibles. Le niveau d’impact
va dépendre de la nature, de l’intensité, de la durée du dragage mais également du niveau de stress « habituel »
auquel est adaptée la communauté. Ainsi, dans les zones naturellement instables de faible profondeur donc des
zones de fortes contraintes environnementales, les espèces sont plus résistantes, plus adaptées aux contraintes
(fort taux de reproduction, grande mobilité, croissance rapide ; Newell et al., 1998, Hermand, 2008). Par
exemple, en estuaire, au niveau des zones de vases, la restauration peut se faire en un an (Figure 6).
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 9
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
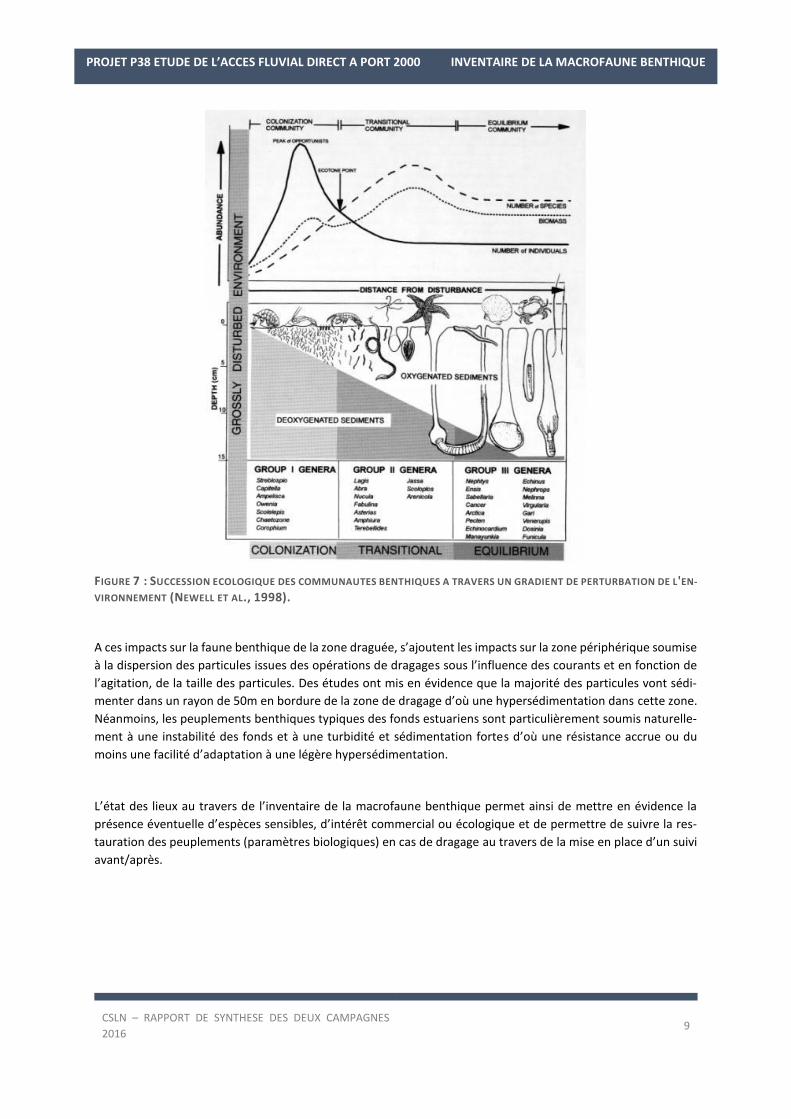
FIGURE 7 : SUCCESSION ECOLOGIQUE DES COMMUNAUTES BENTHIQUES A TRAVERS UN GRADIENT DE PERTURBATION DE L'EN-
VIRONNEMENT (NEWELL ET AL., 1998).
A ces impacts sur la faune benthique de la zone draguée, s’ajoutent les impacts sur la zone périphérique soumise
à la dispersion des particules issues des opérations de dragages sous l’influence des courants et en fonction de
l’agitation, de la taille des particules. Des études ont mis en évidence que la majorité des particules vont sédi-
menter dans un rayon de 50m en bordure de la zone de dragage d’où une hypersédimentation dans cette zone.
Néanmoins, les peuplements benthiques typiques des fonds estuariens sont particulièrement soumis naturelle-
ment à une instabilité des fonds et à une turbidité et sédimentation fortes d’où une résistance accrue ou du
moins une facilité d’adaptation à une légère hypersédimentation.
L’état des lieux au travers de l’inventaire de la macrofaune benthique permet ainsi de mettre en évidence la
présence éventuelle d’espèces sensibles, d’intérêt commercial ou écologique et de permettre de suivre la res-
tauration des peuplements (paramètres biologiques) en cas de dragage au travers de la mise en place d’un suivi
avant/après.
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 10
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
3. MATERIELS ET METHODES
Le prélèvement, le tri et la détermination de la macrofaune benthique de cet inventaire ont été réalisés confor-
mément à la norme ISO/FDIS 16665, 2005 (Qualité de l'eau — Lignes directrices pour l'échantillonnage quanti-
tatif et le traitement d'échantillons de la macrofaune marine des fonds meubles) et selon les recommandations
du "Guide pour l’étude du macrobenthos de l’estuaire de la Seine" (Dauvin, 2013, documents GIP-Seine Aval) ;
protocoles utilisés en routine par la Cellule de Suivi du Littoral Normand.
3.1. Déroulement des prélèvements et localisation des stations En amont, le Maître d’Ouvrage a identifié quatre stations de prélèvement pertinentes vis-à-vis d l’aménagement
projeté en termes de localisation et de gradient (Figure 1). Les stations situées dans l’avant-port historique, le
long de la digue Ouest et dans l’avant-port de Port 2000 seront sur le secteur impacté par les travaux envisagés
tandis que la station à l’angle de la digue Sud est hors du secteur impacté directement par les travaux envisagés.
Les prélèvements ont été positionnés au plus près des coordonnées théoriques en fonction de la hauteur d’eau
présente et la nature sédimentaire. Le navire se repositionnait pour chaque coup de benne.
Les opérations de prélèvements se sont déroulées à partir du moyen nautique du GPMH, le remorqueur « Le
Marais » basé au port du Havre le mardi 16 février 2016 pour la campagne de fin d’hiver et les 28 septembre et
04 octobre pour la campagne de fin d’été 2016. Les conditions météorologiques sont visibles dans le Tableau 1
et les conditions marégraphiques figurent dans le Tableau 2.
TABLEAU 1 : CONDITIONS METEOROLOGIQUES - FRANCE - LA HEVE (LE HAVRE).
TABLEAU 2 : CONDITIONS MAREGRAPHIQUES.
FIGURE 8 : HAUTEUR D’EAU (M) - BALISE A PENDANT LES PERIODES DE PRELEVEMENTS (DONNEES SHOM).
02h 05h 08h 11h 14h 17h 20h 23h 02h 05h 08h 11h 14h 17h 20h 23h 02h 05h 08h 11h 14h 17h 20h 23h 02h 05h 08h 11h 14h 17h 20h 23h 02h 05h 08h 11h 14h 17h 20h 23h 02h 05h 08h 11h 14h 17h 20h 23h 02h 05h 08h 11h 14h 17h 20h 23h
16.02.2016 4 3 2 1 1 1 2 3 6 7 9 9 9 10 12 12 6 6 5 5 6 6 6 5 8 10 44
28.09.2016 4 4 4 4 3 3 2 3 5 5 5 5 5 5 6 6 17 17 16 17 19 20 19 18 91 38 27 67 34 30 34 16
04.10.2016 4 4 4 4 4 4 5 5 4 6 6 6 6 4 5 5 14 13 13 14 18 19 17 16 15 31 65 60 100 98 92
Lat: 49.52, Lon: 0.07 (www.windguru.cz)
GFS 27 kmVitesse du vent (Bft) Direction du vent Direction des vagues Période des vagues (s) Température (°C) Pluie (mm/3h) Couverture nuageuse (%)
h. PM Hauteur Coeff h. PM Hauteur Coeff h. BM Hauteur h. BM Hauteur
Le Havre matin m soir m matin m soir m
16/02/2016 04h21 7,05 59 17h03 6,85 53 11h18 2,50 23h52 2,70
28/09/2016 10h01 7,40 70 22h17 7,50 76 04h39 2,10 17h07 2,15
04/10/2016 01h06 7,85 85 13h23 7,75 82 08h16 1,65 20h28 1,75

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 11
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
La localisation théorique et réelle des stations figurent sur la Figure 9.
FIGURE 9 : LOCALISATION DES STATIONS (COORDONNEES THEORIQUES ET REELLES - CAMPAGNES 2016).
Le faible tirant d’eau au niveau des stations AD_06 et AD_11 pendant la campagne de fin d’hiver a entraîné
respectivement un décalage vers l’ouest et le sud des stations de prélèvement par rapport aux stations théo-
riques.
La présence de graviers, galets et cailloutis a entraîné de nombreux incidents de prélèvements en empêchant la
fermeture de la benne au niveau de la station AD_12 lors des deux campagnes 2016 (jusqu’à 15 coups de benne
nécessaire en fin d’été pour valider les six réplicats).
L’échantillonnage a été réalisé à l’aide de la benne Smith McIntyre du GPMH afin de caractériser quantitative-
ment les peuplements subtidaux d’endofaune et d’épifaune de substrats meubles. D’environ 120 kg, cette benne
est composée de deux mâchoires et est montée dans un cadre métallique pour une meilleure pénétration dans
le sédiment (ajout de poids). Son déclenchement immédiat dès qu’elle touche le fond est très utile pour l’échan-
tillonnage de la faune vagile. Prélevant sur une surface de 0,1m², six prélèvements ont été effectués sur chaque
station pour l’échantillonnage du peuplement benthique (cinq réplicats, 0,5m²) et du sédiment (un réplicat,
0,1m²).
Les cinq réplicats ont été tamisés séparément à bord sur un tamis à maille ronde de 1 mm ne retenant de la
faune benthique que ce qui est qualifié comme macrofaune et laissant passer les sédiments les plus fins gênants
pour le tri ultérieur. Le refus de tamis est ensuite conditionné dans des seaux ou pots et fixé avec une solution
d’eau de mer et de formaldéhyde 35-40% p/v stabilisé avec méthanol à 4%. De la phloxine, colorant rouge du
matériel biologique, est rajoutée à cette solution afin de faciliter le tri de celui-ci au laboratoire.
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 12
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Le prélèvement supplémentaire « coup de benne » pour l’analyse granulométrique a été conditionné en sac
plastique et stocké en milieu réfrigéré et à l’abri de la lumière avant d’être transmis au laboratoire sous-traitant
dans les meilleurs délais.
La mesure de la température et de la salinité à chaque station au fond de la colonne d’eau a été réalisée avec
une sonde HANNA HI-9828. Les résultats sont consignés dans le Tableau 3.
A chaque station, une fiche de relevés a été renseignée.
TABLEAU 3 : DATES ET MESURES HYDROLOGIQUES EN FOND DE COLONNE D'EAU – CAMPAGNES 2016.
Les étapes de prélèvements sont résumées sur la Figure 10.
Station Heure Température Salinité
AD_01 (°C)
16/02/2016 15h45 8,09 32,83
28/09/2016 09h00 18,78 30,26
AD_06
16/02/2016 14h35 8,2 33,28
04/10/2016 15h07 17,79 30,9
AD_11
16/02/2016 13h55 8,16 32,83
28/09/2016 10h00 19,09 30,95
AD_12
16/02/2016 15h00 8,08 32,75
04/10/2016 13h52 17,85 32

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES 2016 13
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 10 : ETAPES DES PRELEVEMENTS.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 14
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
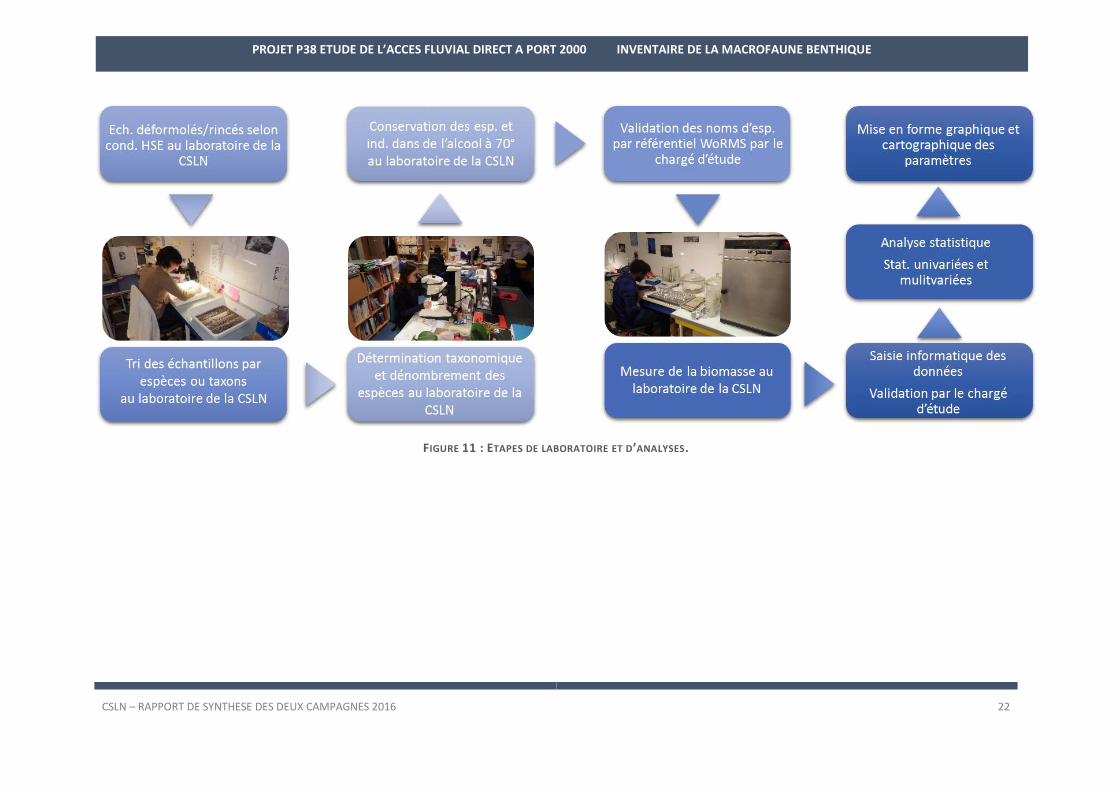
3.2. Analyses au laboratoire Analyses au laboratoire de la CSLN
Les échantillons de faune sont rincés en respectant les conditions d’hygiène et sécurité requises lors de
manipulation de produits cancérigènes puis rincés de nouveau sur une maille de 0,5 mm dans le but d’enlever le
maximum de sédiment fin.
Le tri de chacun des échantillons est effectué à l’œil nu. La totalité de la faune est extraite. La détermi-
nation est effectuée sous loupe binoculaire LEICA M165C, S8APO, MZ16. Dans la mesure du possible, chaque
taxon est identifié jusqu’à l’espèce à l’aide de clés de détermination. Toutefois certains individus abîmés lors des
différentes manipulations ou bien de taille très petite (< 1mm) ne peuvent faire l’objet d’une telle précision.
Certains individus ne sont identifiés qu’à des niveaux élevés de taxonomie (Anthozoaires, Sipunculiens, Né-
mertes, Nématodes, Bryozoaires, Tuniciers ou les genres problématiques). Une veille bibliographique est réalisée
afin de connaitre les dernières mises à jour des clés, mises à jour relativement fréquentes. Des référentiels taxo-
nomiques ont également été développés tels que l’ERMS (European Register of Marine Species,
http://www.marbef.org/data/erms.php) ou le WoRMS (World Registrer of Marine Species, http://www.marines-
pecies.org/). Ils correspondent aux listes taxonomiques des espèces récoltées dans les eaux européennes et
mondiales et sont mis à jour quotidiennement grâce à la veille d’experts internationaux en taxonomie. La liste
des espèces déterminées est validée par le référentiel WoRMS et la date de vérification précisée.
Les individus et espèces sont ensuite conservés dans de l’alcool à 70° en vue de traitements ultérieurs
comme la détermination de leur biomasse.
La biomasse est mesurée de façon à obtenir un poids total par station ainsi que les valeurs relatives aux
principales espèces. Ces mesures, effectuées par espèce, sont issues du cumul des réplicats par station. La faune
qui contribue faiblement à ce paramètre est regroupée par groupe faunistique. La mesure de la masse sèche
libre de cendre (MSLC) est quantifiée par la différence entre le poids étuvé des individus de chaque espèce,
obtenue après passage à l’étuve MEMMERT (48 heures à 80°C) et le poids calciné après passage au four CARBO-
LITE 1100 (2 heures à 550°C). Les pesées sont effectuées sur balance de précision.
Les étapes de laboratoire sont résumées sur la Figure 11.
Analyses granulométriques
Les prélèvements de sédiments destinés aux analyses granulométriques sont stockés en milieu réfrigéré
et à l’abri de la lumière avant d’être transmis au laboratoire sous-traitant.
Les analyses granulométriques des échantillons prélevés ont été réalisées par le Laboratoire de Morphologie
Continentale et Côtière de l’Université de Caen.
La technique utilisée est la granulométrie laser mesurant les particules comprises entre 0,04 µm et 2 mm. Les
différentes fractions granulométriques et leur désignation seront conformes à celles de la norme AFNOR (voir
tableau ci-après). Pour la station AD_12, une granulométrie pondérale est également réalisée selon la norme
AFNOR en raison d’une fraction grossière trop importante et donc inadaptée à une analyse laser.
Le Tableau 4 ci-après récapitule les gammes de taille correspondant à chacune des différentes fractions.
TABLEAU 4: CLASSES GRANULOMETRIQUES.
Taille min Type sédimentaire Taille max
Silts et Argiles ≤ 50 µm
50 µm < Sables fins ≤ 200 µm
200 µm < Sables moyens ≤ 500 µm
500 µm < Sables grossiers ≤ 2 mm
2 mm < Graviers ≤ 2 cm
2 cm < Galets et Cailloutis ≤ 12 cm

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 15
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Les échantillons traités sont caractérisés par :
- une courbe cumulative ainsi qu’une courbe différentielle des pourcentages volumiques,
- le mode de l’échantillon, c’est à dire la taille des particules correspondant au pic de fréquence de ces
dernières sur la courbe différentielle, soit la population la mieux représentée,
- le pourcentage des différentes classes granulométriques.
La définition des faciès sédimentaires permet de synthétiser l’information. Plusieurs classifications existent,
parmi lesquelles celle de Larsonneur modifiée (Lesourd, 2000), largement utilisée dans la caractérisation des
sédiments superficiels en baie de Seine Orientale et à l’embouchure. Actuellement, le choix des classifications
sédimentaires fait l’objet d’un débat au sein de la communauté. Dans le cadre de cette synthèse, il a été décidé
d’appliquer la classification de Larsonneur modifiée afin de permettre une meilleure comparaison avec les études
et travaux de la Baie de Seine antérieurs (Alizier, 2011 ; Lesourd 2000, Programme GIP-Seine-Aval Colmatage,
DEFHFIS). Il faut néanmoins préciser que cette classification permettant de mieux caractériser la teneur en vases
dans les sédiments, les mélanges vaseux apparaissent plus largement répartis que sur l’image donnée par la
classification de Folk. Plus précisément, les faciès sédimentaires ont été définis grâce à la proportion des grains
pour chaque taille de particules selon la classification de Larsonneur modifiée (Tableau 5).
TABLEAU 5 : CLASSIFICATION DE LARSONNEUR MODIFIEE (LESOURD, 2000) (CHIFFRES EXPRIMES EN % DE LA FRACTION TOTALE - G :
GALETS > 2CM - GR : GRAVIERS 2 CM A 2 MM - SG : SABLES GROSSIERS 500 ΜM A 2 MM - SM : SABLES MOYENS 200 A 500 ΜM- SF SABLES FINS 50
A 200 ΜM - V : VASES<50ΜM).
Résultats biologiques
Les résultats faunistiques sont exprimés à l'aide de descripteurs simples couramment utilisés pour ce type de
suivi :
- la richesse spécifique (nombre d’espèces identifiées par station),
- la densité des individus de chaque taxon (nombre d’individus/m²),
- la biomasse (g/m² de matière sèche libre de cendres).
Les résultats bruts sont donc exprimés pour chaque station, en nombre d’espèces, en nombre d’individus par
m2, espèce par espèce, et en grammes de matière sèche libre de cendres pour chaque espèce (ou groupe d’es-
pèces) ou groupe faunistique.
Ces résultats sont présentés sous forme de tableaux, graphiques ou cartes.
Silts et Argiles ≤ 50 µm Silts et Argiles ≤ 50 µm + Sables ]50-200µm] Médiane Faciès sédimentaire Code< 5% < 50% > 2 mm > 50% Galets (≥ 2 cm) > 70% Cailloutis Code
50% < Galets (≥ 2 cm) < 70% Cailloutis graveleux CGr
15% < Galets (≥ 2 cm) < 50% Gravier caillouteux GrC
Galets (≥ 2 cm) < 15% Gravier Gr
> 50% < 2 mm 15% < Gr + G < 50% % Graviers < % Galets Sable caillouteux SC
% Graviers > % Galets Sable graveleux Sgr
< 15% Sables grossiers majoritaires Sable grossier SG
Sables moyens majoritaires Sable moyen SM
Sables fins majoritaires Sable fin SF
> 5% V < 25% Sable vaseux SV
25% < V < 75% Vase sableuse VS
V > 75% Vase V
Graviers et Galets (>2mm)

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 16
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
3.3. Analyses des données
La description des communautés en place est réalisée à travers une série d'analyses.
Les étapes d’analyses sont résumées sur la Figure 11.
Indices classiques
L’Occurrence est le nombre de fois qu’un taxon donné est présent dans l'ensemble des stations d’échan-
tillonnage.
La Constance (Dajoz, 1985) est le rapport du nombre de fois où un taxon est enregistré sur le nombre
de stations prospectées à partir d’un même engin. La constance If peut alors se décliner selon les quatre catégo-
ries suivantes :
If < 12 % espèce rare
13 < If < 25 % espèce peu commune
26 < If < 50 % espèce commune
51 < If < 100 % espèce constante
L’indice de diversité de Shannon (H’) et l’Équitabilité de Pielou (J’) sont calculés pour chaque station :
H’= -Σs1pi log2 pi et J’=H’/H’max
Avec :
S : nombre total de taxons de la station,
pi : abondance relative de l’espèce i (pi= ni/N avec N : nombre total d’individus de la station et ni : nombre
d’individus de l’espèce i),
H’max : diversité maximale de la station comportant S taxons (H’max=log2 S).
L’indice de diversité de Shannon (Shannon, C.E. & Weaver W. 1949) prend non seulement en compte le
nombre d’espèces mais également la distribution des individus de ces espèces. Une station dominée par une
seule espèce présente un indice de diversité de Shannon moindre qu’une station dont les espèces sont codomi-
nantes. Sa valeur varie de 0 (une seule espèce ou très large domination d’une seule espèce) à log2S (toutes les
espèces ont la même abondance). Un indice de Shannon élevé indique donc la maturité de la communauté ben-
thique de la station considérée mais aussi un dynamisme faible. Un indice de Shannon faible ou en baisse traduit
un déséquilibre de la communauté du fait de la prolifération de quelques espèces. Vincent et al. (2002) ont défini
les équivalences entre les valeurs de l'indice de Shannon et les différents états écologiques définis par la Directive
Cadre sur l'Eau.
L’équitabilité permet de mesurer la répartition des individus des espèces indépendamment de la ri-
chesse spécifique. Il correspond au rapport de l’indice H’ sur un indice théorique H’max du même échantillon où
toutes les espèces auraient la même densité (H’max= Log2 S où S est la richesse spécifique). Plus le rapport est
élevé, plus la communauté de l’échantillon est homogène en densité et bien diversifiée ; à l’inverse, un faible
rapport indique la dominance d’une ou de quelques espèces. Sa valeur varie de 0 (dominance d’une espèce) à 1
(équirépartition des effectifs des espèces).
Ces deux indices permettent de suivre les modifications de structure des communautés dans le temps comme
dans l’espace, mais l’équitabilité est particulièrement adaptée à la comparaison des stations entre elles. La com-
binaison de ces deux indices fournit une indication sur la répartition des effectifs des taxons et ainsi les équilibres
ou déséquilibres pouvant exister.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 17
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
L’Abondance relative des espèces en % et dominance sont décrites au travers de Diagrammes rang /
fréquence soit le classement sur les densités de chaque espèce illustré sous forme de graphique bidimensionnel
par ordre d’abondance relative (%) décroissante des espèces. Les espèces occupant les premiers rangs caracté-
risent le peuplement en place avec leur cortège d’espèces associées.
Enfin, une attention particulière est portée à la présence (présence et abondance) d’espèces clés assu-
rant l’essentiel de la production de l’écosystème benthique.
Pour les communautés benthiques, les assemblages de stations présentant des similarités dans leur
composition faunistique sont identifiés grâce à une Classification Ascendante Hiérarchique (CAH) complétée
d’une Analyse Multidimensionelle non-métrique (MDS). Celles-ci sont réalisées à partir d’une matrice utilisant
les indices de similarité de Bray-Curtis obtenus à partir des données de densités transformées (√√) selon la tech-
nique de regroupement agglomératif moyen, recommandée par Legendre & Legendre (1998 ; dans Grall & Glé-
marec, 2001) et largement citée dans la littérature dans le cadre d’évaluations de la qualité du macrozoobenthos
en milieu côtier (Grall & Coïc, 2005). Le schéma d’arborescence (dendrogramme) résultant de la CAH relie les
données selon leur niveau de similarité et permet de déterminer des groupes de stations, la longueur des
branches représentant la distance moyenne (ou le pourcentage de similarité moyen) entre les stations ou
groupes de stations. Pour l’analyse prenant en compte les données des autres études réalisées au niveau des
bassins portuaires, seules les espèces présentes au niveau de 5% des observations ont été prises en compte.
L’analyse multidimensionnelle (MDS) permet de compléter l’analyse précédente en faisant abstraction des dis-
continuités mises en évidence par la CAH. Cette méthode d’ordination utilise les similitudes entre stations pour
élaborer un modèle de répartition des stations entres elles dans un repère non-métrique. En suivant un proces-
sus itératif, l’analyse va aboutir à la meilleure adéquation entre le modèle et les données. La représentation est
évaluée par le « Stress de Kruskal » dont la valeur traduit la représentativité du modèle (Grall & Coïc, 2005) :
> 0,5 : la représentation est probablement aléatoire,
Entre 0,5 et 0,25 : la qualité de la représentation est médiocre,
Entre 0,1 et 0,25 : la représentation est satisfaisante,
<0,1 : la représentation est excellente.
La méthode IndVal (Dufrêne et Legendre, 1997) est utilisée pour indiquer les espèces caractérisant les différents
assemblages de stations établis par la CAH en utilisant la méthode des valeurs indicatrices (logiciel IndVal, Du-
frêne et Legendre, 1997). La valeur indicatrice de chaque espèce est calculée au sein de chaque assemblage grâce
à la formule : IndVal = Aij x Bij
Avec :
- Aij = Densité moyenne de l’espèce i sur les stations de l’assemblage j / Somme des densités moyennes
de l’espèce i sur tous les assemblages.
- Bij = Nombre de stations de l’assemblage j où l’espèce i est présente / Nombre total de stations de
l’assemblage j.
Ainsi, une espèce i est indicatrice d’un assemblage j quand sa valeur indicatrice est la plus élevée de celles des
espèces de l’assemblage (IndVal espèce j = max[IndValk,j]. La significativité de cet indice est testée par combi-
naisons aléatoires des sites au sein des groupes (p < 0,05 ; n = 999). Plus la valeur indicatrice d’une espèce se
rapproche de 1, plus elle est « caractéristique » du groupe de stations.
Ces analyses, réalisées à l’aide du logiciel XLSTAT mettent en évidence des assemblages de stations traduisant
les différentes structures faunistiques rencontrées sur la zone d’étude.
D’autres tests statistiques sont réalisés avec le logiciel XLstat®. Ce sont des tests et des analyses classiquement
appliqués dans ce type d’études (Legendre et Legendre, 1998).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 18
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Le Box-plot est un système de représentation d’un jeu de données dont la légende est expliquée ci-
dessous :
Le test non paramétrique de Kruskall-Wallis (KW) permet la comparaison statistique entre plusieurs
échantillons. La p-value (donnée entre parenthèse) permet d’évaluer la significativité (%) des différences poten-
tielles mises en évidence. Une comparaison multiple par paires post hoc suivant la procédure de Conover-Iman
est réalisée quand p < 0,1 avec une correction de Bonferroni.
L’Analyse en Composantes Principales (ACP) est une méthode pour l’analyse de données quantitatives
(continues ou discrètes) se présentant sous forme de tableau à M lignes (stations) / N colonnes (variables de
pression). Elle permet de visualiser et analyser les corrélations entre N variables de pression et de visualiser et
analyser les M stations initialement décrites par N variables de pression sur un graphique à deux ou trois dimen-
sions, construit de manière à ce que la dispersion entre les données soit aussi bien préservée que possible.
Indices basés sur les groupes écologiques
Ces indices classiques sont être complétés par des indices biotiques et de qualité écologique.
En effet, l’état écologique du peuplement (EQS) est un des critères définis par la DCE pour comparer la
qualité des milieux entre les états membres. La qualification de l’état écologique du milieu (EQS) passe par l’ap-
plication d’indices appelés Indices Biotiques. La détermination de l’état de l’EQS passe par l’utilisation d’une grille
à 5 niveaux qui permet de qualifier l’état du peuplement de mauvais à bon. L’EQS peut être évalué à partir de
l’indice de Shannon (H’) considéré comme l’un des meilleurs indicateurs des perturbations importantes de la
communauté et l’indice biotique AMBI recommandé par la DCE.
L’indice AMBI pour AZTI’s Marine Biotic Index concernant le benthos de substrats meubles a été mis au
point pour les milieux côtiers européens (Borja et al., 2000 ; Borja et Muxika, 2005). Cet indice est fondé sur les
densités individuelles des espèces de cinq groupes écologiques définis en fonction de leur tolérance et/ou de
leur sensibilité à un gradient de stress environnemental (Hily, 1984 ; Grall et Glémarec, 1997). L'indice biotique
constitue une approche de la structure des peuplements benthiques en fonction du niveau de perturbation.
Il repose sur l'utilisation de 5 groupes écologiques de polluosensibilités différentes. Selon le gradient de pollution,
les 5 groupes écologiques vont se succéder selon 4 grandes étapes entre lesquelles s'intercalent des phases de
transition. Ces groupes ont été résumés par Grall et Glémarec (1997) :
- Groupe I : Espèces très sensibles à l’enrichissement en matière organique et présentes en milieu non
pollué (état initial),
- Groupe II : Espèces indifférentes à l’enrichissement, toujours présentes en faibles densités et sans
variations saisonnières prononcées,
- Groupe III : Espèces tolérantes à l’excès de matière organique. Ces espèces peuvent être présentes
dans les conditions normales, mais leurs populations sont stimulées par l’enrichissement en matière organique,
- Groupe IV : espèces opportunistes de second ordre : polychètes, déposivores de subsurface tels que
les cirratulidés,
- Groupe V : espèces opportunistes de premier ordre : déposivores proliférant dans les sédiments ré-
duits.
25% 75% 95%5% me
d.
mo
y.
min
max

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 19
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Bien que l’AMBI soit désormais couramment utilisé, la pertinence du classement des espèces défini par Borja et
al. a fait l’objet de critiques. La participation de nombreuses équipes de recherche au classement des espèces a
permis l’édition d’une nouvelle liste en mars 2012.
La distribution de ces groupes écologiques en fonction de leur sensibilité aux perturbations environnementales,
permet l’élaboration d’un indice biotique allant de 0 à 7 (7 : azoïque).
Borja et al. (2000) ont défini les équivalences entre les valeurs de l'AMBI et les différents états écologiques définis
par la Directive cadre sur l'Eau (Tableau 3).
TABLEAU 6 : SIGNIFICATION DE L'AMBI POUR LA COMMUNAUTE A ABRA ALBA – LAGIS KORENI (BORJA ET AL., 2000).
Cette définition des statuts n'étant pas adaptée aux communautés estuariennes, cet indice n’est retenu en
France que pour qualifier les eaux côtières (mais pas les estuaires) dans le cadre de la DCE (communauté Abra
alba – Lagis koreni).
Son utilisation en eaux de transition et plus particulièrement dans les estuaires ou les bassins rencontre
donc quelques limites et nécessite quelques précautions. En effet, la qualité des sites dépendant des seuils choi-
sis, des études sont en cours afin d’établir des valeurs de seuils adaptés aux conditions estuariennes. Ruellet et
Dauvin (2007) ont proposé une harmonisation des seuils des différents indices existants mais la nécessité de
créer un indicateur unique ou un ensemble d'indicateurs, reste d’actualité pour évaluer l'état de qualité écolo-
gique des communautés benthiques dans les eaux de transition. Ces seuils seront appliqués aux biocénoses des
stations du bassin étudié.
TABLEAU 7 : SIGNIFICATION DE L'AMBI POUR LES COMMUNAUTES ESTUARIENNES (RUELLET ET DAUVIN, 2007).
L’indice d’évaluation de l’endofaune côtière I2EC s’avère également indispensable pour juger de la dé-
gradation du milieu portuaire (Grall et al., 2003). Basé également sur les proportions des groupes écologiques, il
permet de révéler l’état de santé du milieu (Tableau 5). Ces groupes écologiques feront également l’objet d’une
note. Afin de limiter la part de subjectivité, l’indice est attribué par le biais d’une clé (Grall et Glémarec, 2003).
La première étape consiste à comparer les richesses spécifiques et abondances du site étudié avec celles d’un
état de référence pour l’habitat considéré (Tableau 8).
Etat écologique AMBI Classification de la pollution
Mauvais ]5,5 - 7] Azoïque
Médiocre ]4,3 - 5,5] Gravement pollué
Moyen ]3,4 - 4,3] Modérement pollué
Bon ]1,2 - 3,3] Légèrement pollué, état transitoire
Très bon ]0 - 1,2] Normal
Etat écologique AMBI Classification de la pollution
Mauvais ]4,3 - 7] Azoïque
Médiocre ]3,4 - 4,3] Gravement pollué
Moyen ]2 - 3,4] Modérement pollué
Bon ]0,15 - 1,2] Légèrement pollué, état transitoire
Très bon ]0 - 0,15] Normal
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 20
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
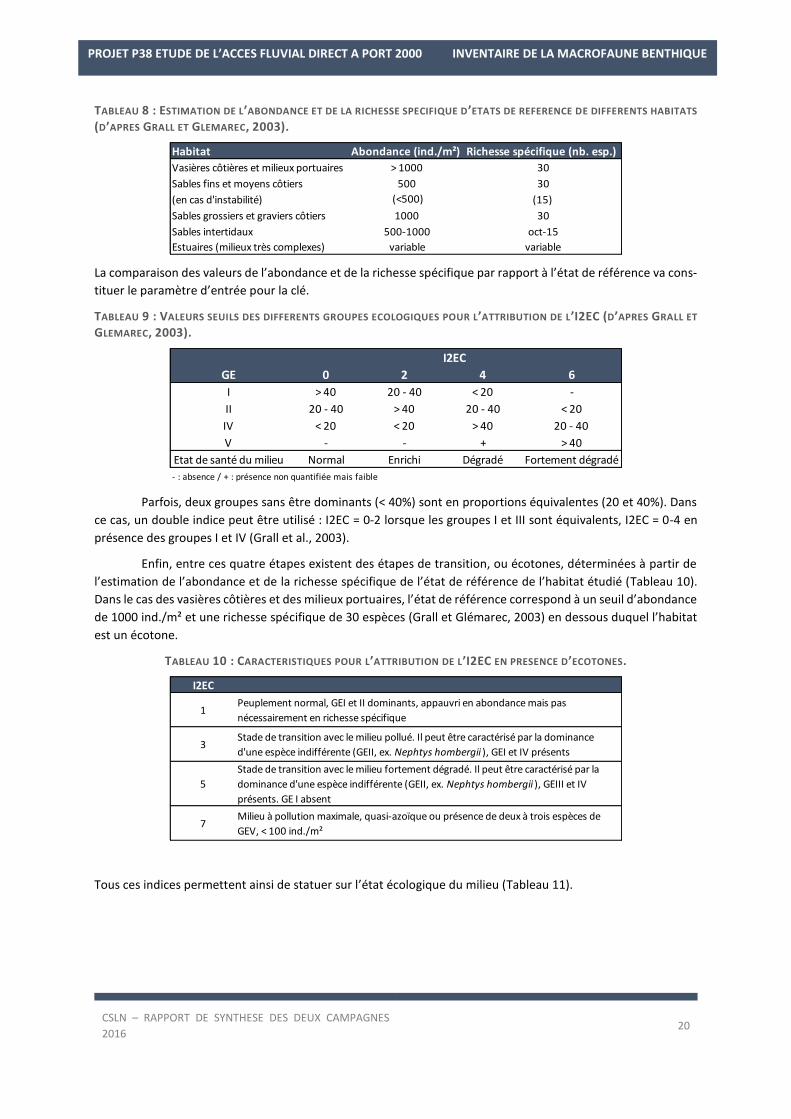
TABLEAU 8 : ESTIMATION DE L’ABONDANCE ET DE LA RICHESSE SPECIFIQUE D’ETATS DE REFERENCE DE DIFFERENTS HABITATS
(D’APRES GRALL ET GLEMAREC, 2003).
La comparaison des valeurs de l’abondance et de la richesse spécifique par rapport à l’état de référence va cons-
tituer le paramètre d’entrée pour la clé.
TABLEAU 9 : VALEURS SEUILS DES DIFFERENTS GROUPES ECOLOGIQUES POUR L’ATTRIBUTION DE L’I2EC (D’APRES GRALL ET
GLEMAREC, 2003).
Parfois, deux groupes sans être dominants (< 40%) sont en proportions équivalentes (20 et 40%). Dans
ce cas, un double indice peut être utilisé : I2EC = 0-2 lorsque les groupes I et III sont équivalents, I2EC = 0-4 en
présence des groupes I et IV (Grall et al., 2003).
Enfin, entre ces quatre étapes existent des étapes de transition, ou écotones, déterminées à partir de
l’estimation de l’abondance et de la richesse spécifique de l’état de référence de l’habitat étudié (Tableau 10).
Dans le cas des vasières côtières et des milieux portuaires, l’état de référence correspond à un seuil d’abondance
de 1000 ind./m² et une richesse spécifique de 30 espèces (Grall et Glémarec, 2003) en dessous duquel l’habitat
est un écotone.
TABLEAU 10 : CARACTERISTIQUES POUR L’ATTRIBUTION DE L’I2EC EN PRESENCE D’ECOTONES.
Tous ces indices permettent ainsi de statuer sur l’état écologique du milieu (Tableau 11).
Habitat Abondance (ind./m²) Richesse spécifique (nb. esp.)
Vasières côtières et milieux portuaires > 1000 30
Sables fins et moyens côtiers 500 30
(en cas d'instabilité) (<500) (15)
Sables grossiers et graviers côtiers 1000 30
Sables intertidaux 500-1000 oct-15
Estuaires (milieux très complexes) variable variable
GE 0 2 4 6
I > 40 20 - 40 < 20 -
II 20 - 40 > 40 20 - 40 < 20
IV < 20 < 20 > 40 20 - 40
V - - + > 40
Etat de santé du milieu Normal Enrichi Dégradé Fortement dégradé
- : absence / + : présence non quantifiée mais faible
I2EC
I2EC
1Peuplement normal, GEI et II dominants, appauvri en abondance mais pas
nécessairement en richesse spécifique
3Stade de transition avec le milieu pollué. Il peut être caractérisé par la dominance
d'une espèce indifférente (GEII, ex. Nephtys hombergii ), GEI et IV présents
5
Stade de transition avec le milieu fortement dégradé. Il peut être caractérisé par la
dominance d'une espèce indifférente (GEII, ex. Nephtys hombergii ), GEIII et IV
présents. GE I absent
7Milieu à pollution maximale, quasi-azoïque ou présence de deux à trois espèces de
GEV, < 100 ind./m²
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 21
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
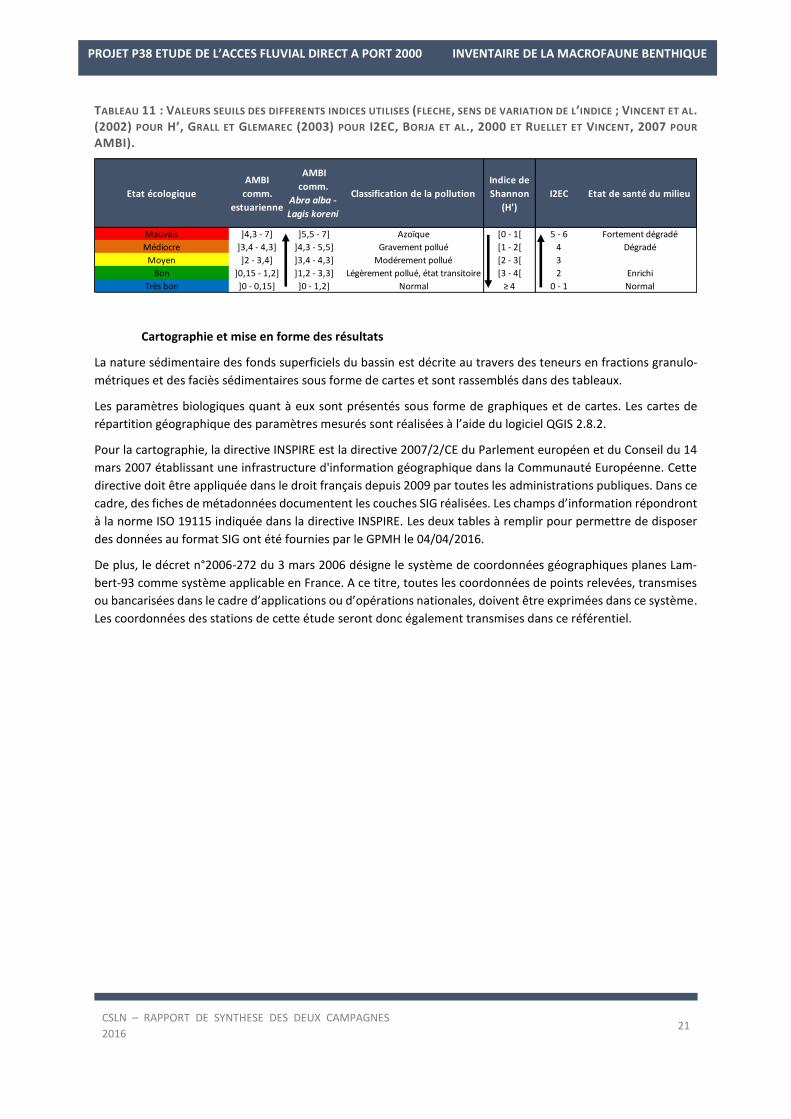
TABLEAU 11 : VALEURS SEUILS DES DIFFERENTS INDICES UTILISES (FLECHE, SENS DE VARIATION DE L’INDICE ; VINCENT ET AL.
(2002) POUR H’, GRALL ET GLEMAREC (2003) POUR I2EC, BORJA ET AL., 2000 ET RUELLET ET VINCENT, 2007 POUR
AMBI).
Cartographie et mise en forme des résultats
La nature sédimentaire des fonds superficiels du bassin est décrite au travers des teneurs en fractions granulo-
métriques et des faciès sédimentaires sous forme de cartes et sont rassemblés dans des tableaux.
Les paramètres biologiques quant à eux sont présentés sous forme de graphiques et de cartes. Les cartes de
répartition géographique des paramètres mesurés sont réalisées à l’aide du logiciel QGIS 2.8.2.
Pour la cartographie, la directive INSPIRE est la directive 2007/2/CE du Parlement européen et du Conseil du 14
mars 2007 établissant une infrastructure d'information géographique dans la Communauté Européenne. Cette
directive doit être appliquée dans le droit français depuis 2009 par toutes les administrations publiques. Dans ce
cadre, des fiches de métadonnées documentent les couches SIG réalisées. Les champs d’information répondront
à la norme ISO 19115 indiquée dans la directive INSPIRE. Les deux tables à remplir pour permettre de disposer
des données au format SIG ont été fournies par le GPMH le 04/04/2016.
De plus, le décret n°2006-272 du 3 mars 2006 désigne le système de coordonnées géographiques planes Lam-
bert-93 comme système applicable en France. A ce titre, toutes les coordonnées de points relevées, transmises
ou bancarisées dans le cadre d’applications ou d’opérations nationales, doivent être exprimées dans ce système.
Les coordonnées des stations de cette étude seront donc également transmises dans ce référentiel.
Etat écologique
AMBI
comm.
estuarienne
AMBI
comm.
Abra alba -
Lagis koreni
Classification de la pollution
Indice de
Shannon
(H')
I2EC Etat de santé du milieu
Mauvais ]4,3 - 7] ]5,5 - 7] Azoïque [0 - 1[ 5 - 6 Fortement dégradé
Médiocre ]3,4 - 4,3] ]4,3 - 5,5] Gravement pollué [1 - 2[ 4 Dégradé
Moyen ]2 - 3,4] ]3,4 - 4,3] Modérement pollué [2 - 3[ 3
Bon ]0,15 - 1,2] ]1,2 - 3,3] Légèrement pollué, état transitoire [3 - 4[ 2 Enrichi
Très bon ]0 - 0,15] ]0 - 1,2] Normal ≥ 4 0 - 1 Normal
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES 2016 22
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 11 : ETAPES DE LABORATOIRE ET D’ANALYSES.
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 23
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
4. CARACTERISTIQUES SEDIMENTAIRES
Les résultats par station et par campagne sont en Annexe 1 avec les spectres granulométriques (courbes
cumulatives et différentielle des pourcentages volumiques des fractions) et les tableaux regroupant les modes,
médiane, moyenne et statistiques de l’échantillon ainsi que les pourcentages des fractions sédimentaires.
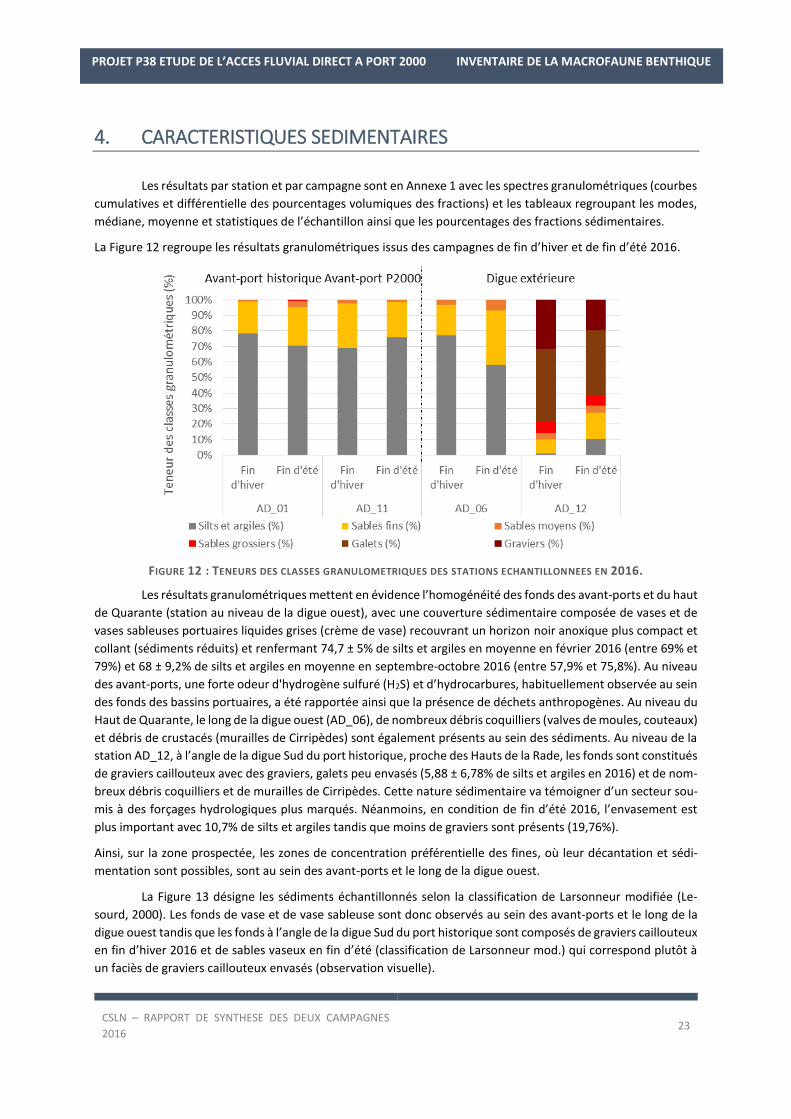
La Figure 12 regroupe les résultats granulométriques issus des campagnes de fin d’hiver et de fin d’été 2016.
FIGURE 12 : TENEURS DES CLASSES GRANULOMETRIQUES DES STATIONS ECHANTILLONNEES EN 2016.
Les résultats granulométriques mettent en évidence l’homogénéité des fonds des avant-ports et du haut
de Quarante (station au niveau de la digue ouest), avec une couverture sédimentaire composée de vases et de
vases sableuses portuaires liquides grises (crème de vase) recouvrant un horizon noir anoxique plus compact et
collant (sédiments réduits) et renfermant 74,7 ± 5% de silts et argiles en moyenne en février 2016 (entre 69% et
79%) et 68 ± 9,2% de silts et argiles en moyenne en septembre-octobre 2016 (entre 57,9% et 75,8%). Au niveau
des avant-ports, une forte odeur d'hydrogène sulfuré (H2S) et d’hydrocarbures, habituellement observée au sein
des fonds des bassins portuaires, a été rapportée ainsi que la présence de déchets anthropogènes. Au niveau du
Haut de Quarante, le long de la digue ouest (AD_06), de nombreux débris coquilliers (valves de moules, couteaux)
et débris de crustacés (murailles de Cirripèdes) sont également présents au sein des sédiments. Au niveau de la
station AD_12, à l’angle de la digue Sud du port historique, proche des Hauts de la Rade, les fonds sont constitués
de graviers caillouteux avec des graviers, galets peu envasés (5,88 ± 6,78% de silts et argiles en 2016) et de nom-
breux débris coquilliers et de murailles de Cirripèdes. Cette nature sédimentaire va témoigner d’un secteur sou-
mis à des forçages hydrologiques plus marqués. Néanmoins, en condition de fin d’été 2016, l’envasement est
plus important avec 10,7% de silts et argiles tandis que moins de graviers sont présents (19,76%).
Ainsi, sur la zone prospectée, les zones de concentration préférentielle des fines, où leur décantation et sédi-
mentation sont possibles, sont au sein des avant-ports et le long de la digue ouest.
La Figure 13 désigne les sédiments échantillonnés selon la classification de Larsonneur modifiée (Le-
sourd, 2000). Les fonds de vase et de vase sableuse sont donc observés au sein des avant-ports et le long de la
digue ouest tandis que les fonds à l’angle de la digue Sud du port historique sont composés de graviers caillouteux
en fin d’hiver 2016 et de sables vaseux en fin d’été (classification de Larsonneur mod.) qui correspond plutôt à
un faciès de graviers caillouteux envasés (observation visuelle).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 24
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 13 : REPARTITION SPATIALE DES FACIES SEDIMENTAIRES (CL. DE LARSONNEUR MODIFIEE ; LESOURD, 2000) AUX
STATIONS ECHANTILLONNEES EN 2016.
AD_12 AD_01
AD_06
AD_11
AD_12 AD_01
AD_06
AD_11
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 25
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
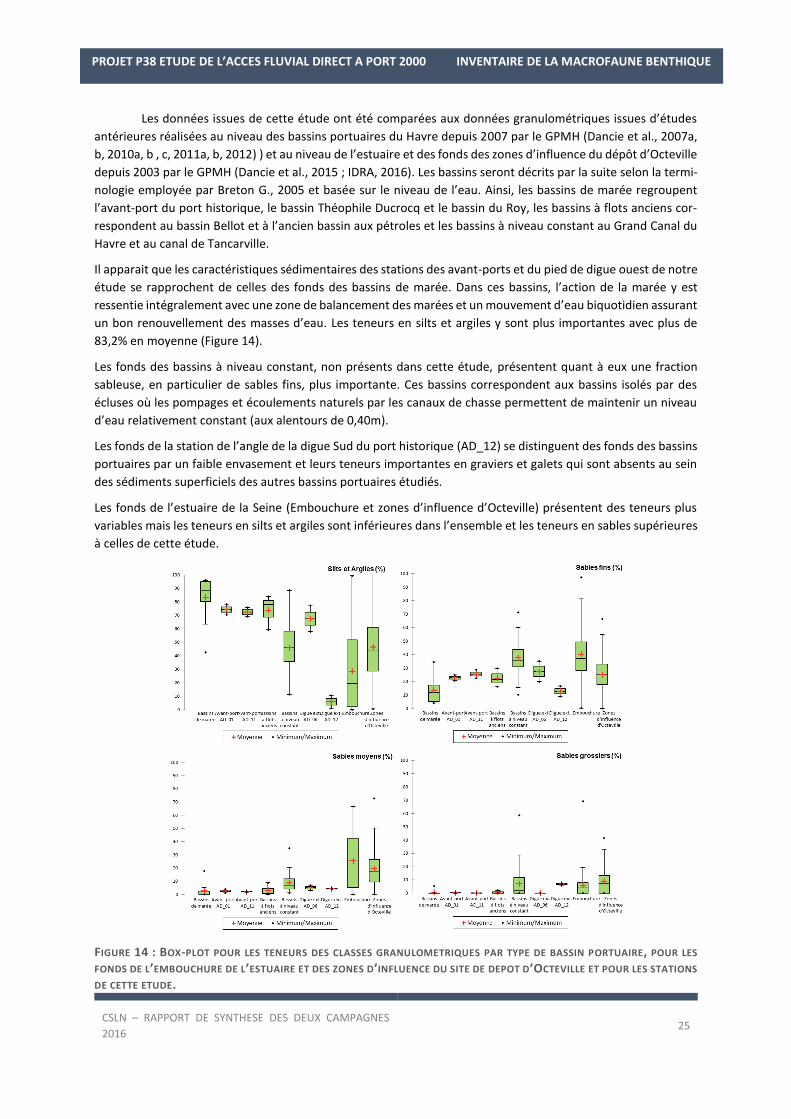
Les données issues de cette étude ont été comparées aux données granulométriques issues d’études
antérieures réalisées au niveau des bassins portuaires du Havre depuis 2007 par le GPMH (Dancie et al., 2007a,
b, 2010a, b , c, 2011a, b, 2012) ) et au niveau de l’estuaire et des fonds des zones d’influence du dépôt d’Octeville
depuis 2003 par le GPMH (Dancie et al., 2015 ; IDRA, 2016). Les bassins seront décrits par la suite selon la termi-
nologie employée par Breton G., 2005 et basée sur le niveau de l’eau. Ainsi, les bassins de marée regroupent
l’avant-port du port historique, le bassin Théophile Ducrocq et le bassin du Roy, les bassins à flots anciens cor-
respondent au bassin Bellot et à l’ancien bassin aux pétroles et les bassins à niveau constant au Grand Canal du
Havre et au canal de Tancarville.
Il apparait que les caractéristiques sédimentaires des stations des avant-ports et du pied de digue ouest de notre
étude se rapprochent de celles des fonds des bassins de marée. Dans ces bassins, l’action de la marée y est
ressentie intégralement avec une zone de balancement des marées et un mouvement d’eau biquotidien assurant
un bon renouvellement des masses d’eau. Les teneurs en silts et argiles y sont plus importantes avec plus de
83,2% en moyenne (Figure 14).
Les fonds des bassins à niveau constant, non présents dans cette étude, présentent quant à eux une fraction
sableuse, en particulier de sables fins, plus importante. Ces bassins correspondent aux bassins isolés par des
écluses où les pompages et écoulements naturels par les canaux de chasse permettent de maintenir un niveau
d’eau relativement constant (aux alentours de 0,40m).
Les fonds de la station de l’angle de la digue Sud du port historique (AD_12) se distinguent des fonds des bassins
portuaires par un faible envasement et leurs teneurs importantes en graviers et galets qui sont absents au sein
des sédiments superficiels des autres bassins portuaires étudiés.
Les fonds de l’estuaire de la Seine (Embouchure et zones d’influence d’Octeville) présentent des teneurs plus
variables mais les teneurs en silts et argiles sont inférieures dans l’ensemble et les teneurs en sables supérieures
à celles de cette étude.
FIGURE 14 : BOX-PLOT POUR LES TENEURS DES CLASSES GRANULOMETRIQUES PAR TYPE DE BASSIN PORTUAIRE, POUR LES
FONDS DE L’EMBOUCHURE DE L’ESTUAIRE ET DES ZONES D’INFLUENCE DU SITE DE DEPOT D’OCTEVILLE ET POUR LES STATIONS
DE CETTE ETUDE.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 26
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
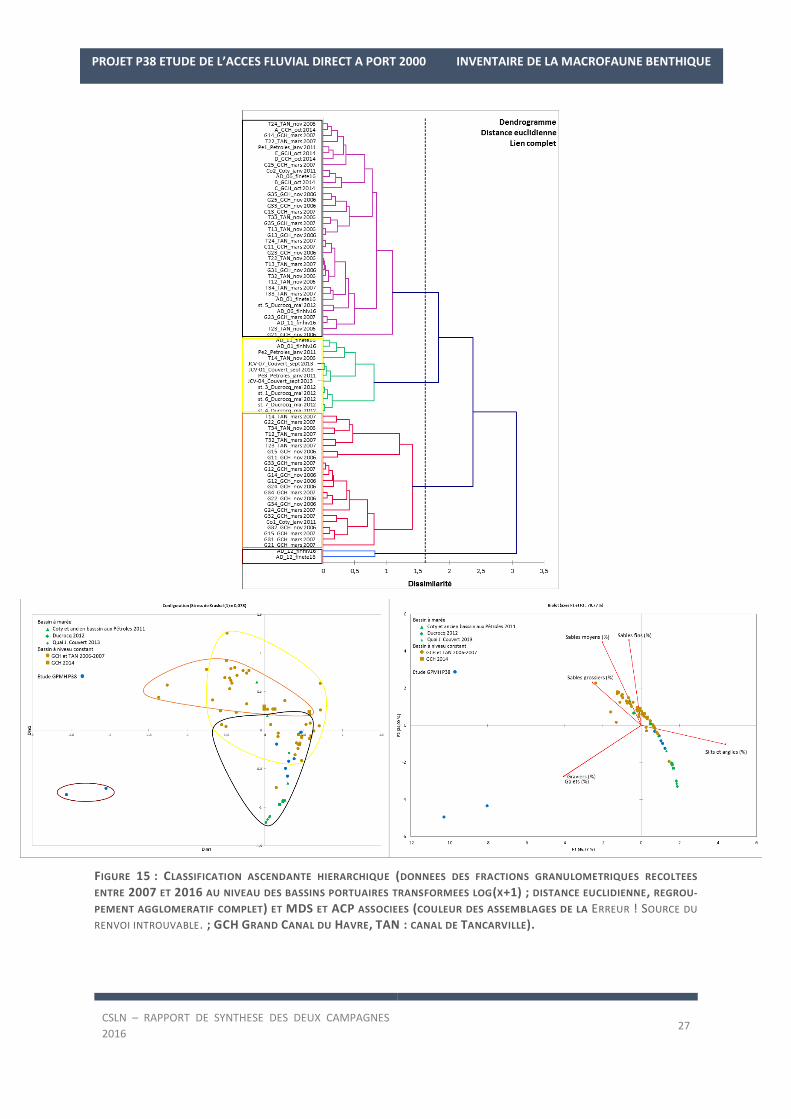
Ceci est confirmé par la Figure 15 qui illustre la CAH des données granulométriques transformées issues
de ces études et la MDS et ACP associées avec les caractéristiques sédimentaires de chaque assemblage.
En effet, les fonds de la station AD_12 sont distingués des autres fonds du complexe portuaires en raison de leurs
caractéristiques sédimentaires plus grossières (44,5 ± 3,2% de graviers et 25,7 ± 8,4% de galets, Assemblage 3).
Parallèlement, trois autres assemblages sont mis en évidence.
L’assemblage 1 regroupe les stations de la digue ouest et des avant-ports (station AD_01 en fin d’été et AD_11
en fin d’hiver) de notre étude avec deux stations de bassin de marée, une station située au niveau de l’ancien
bassin à Pétroles et une grande partie des stations situées au milieu du Grand Canal du Havre et du canal de
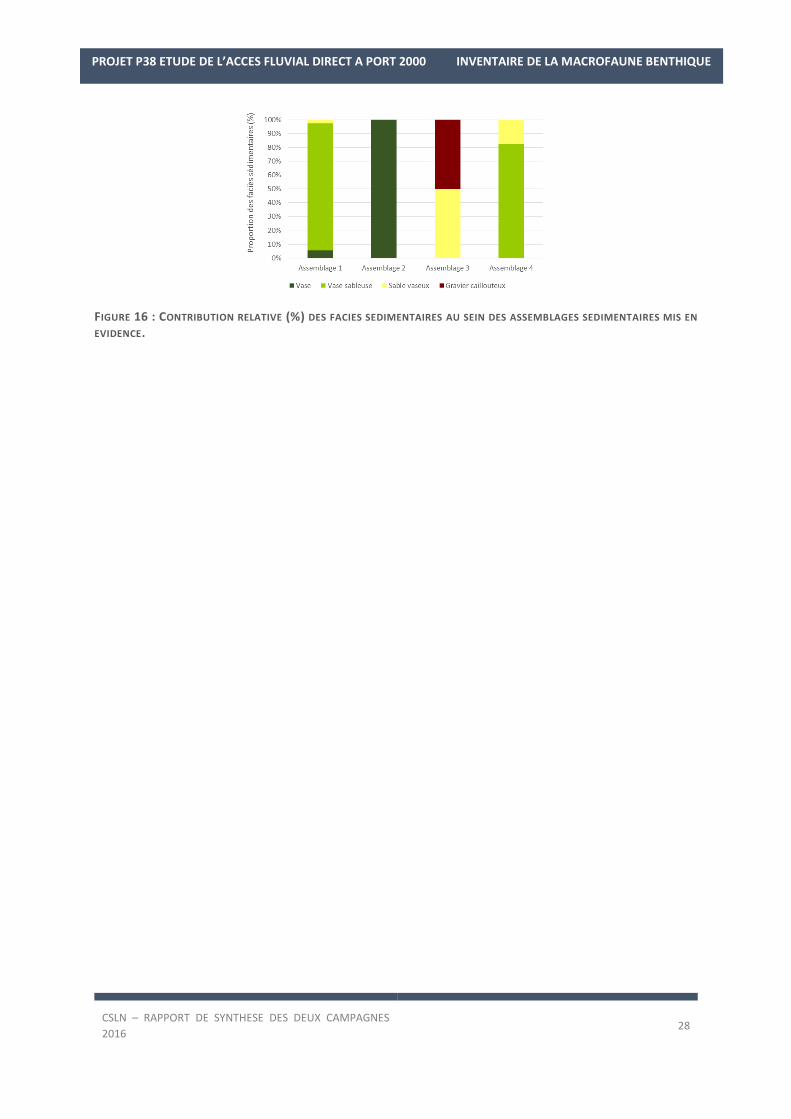
Tancarville. Les fonds superficiels de ces stations, de vases sableuses majoritairement (Figure 16), se caractéri-
sent par une dominance des fractions de silts et argiles et de sables fins (53,5 ± 13,5% et 39,2 ± 12,4 %).
L’assemblage 2 caractérise les fonds superficiels composés de vases portuaires avec une fraction importante de
particules fines (87,6 ± 7,3%). Il regroupe la majorité des stations des bassins portuaires à marée du Havre, des
avant-ports (station AD_01 en fin d’hiver et AD_11 en fin d’été) à l’enclave sur le quai nord du bassin René Coty.
Enfin, l’assemblage 4 concerne essentiellement les stations étudiées au niveau du Grand Canal du Havre et à l’est
du canal de Tancarvillle vers l’écluse. Les teneurs en sables moyens et en sables grossiers à ces niveaux sont plus
importants qu’au sein des autres bassins portuaires (12 ± 7,7% et 16,3 ± 13,6 %). Aucune station de notre étude
n’est rattachée à cet assemblage.
Ce gradient d’envasement est visible sur l’ACP avec une transition des sédiments des canaux portuaires caracté-
risés par les fractions sableuses vers les bassins portuaires à marée caractérisés par les fractions de silts et argiles
(bassin René Coty, Théophile Ducrocq, Ancien bassin à pétroles ; Figure 15). Les fonds étudiés lors de notre étude
se retrouvent à la jonction entre ces deux groupes, mis à part les fonds de la station AD_12 qui se distinguent et
sont isolés de par la fraction de graviers et galets (partie gauche de l’ACP).
Les résultats d’études menées dans d’autres ports comme à Dunkerque dans un bassin à niveau constant (écluse)
mettent en évidence des fonds de vases sableuses avec des caractéristiques sédimentaires également similaires
à celles de notre étude (56,5 ± 1,7% de silts et argiles en moyenne en mars 2014 et 70,9 ± 4,9% de silts et argiles
en moyenne en octobre 2014 ; Dancie et al., 2015).
Les tests de corrélations (Pearson, α=0,01) mettent en évidence une corrélation négative significative des te-
neurs en silts et argiles avec les teneurs en sables fins, moyens et grossiers (p-values<0,0001) et avec les teneurs
en graviers et galets (p-values=0,0001 et 0,002). Il existe également une corrélation négative significative entre
les teneurs en sables fins et en graviers (p-values=0,02). Enfin, une corrélation positive significative entre les
teneurs en sables fins et en sables moyens est également établie (p-values=0,014).
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 27
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 15 : CLASSIFICATION ASCENDANTE HIERARCHIQUE (DONNEES DES FRACTIONS GRANULOMETRIQUES RECOLTEES
ENTRE 2007 ET 2016 AU NIVEAU DES BASSINS PORTUAIRES TRANSFORMEES LOG(X+1) ; DISTANCE EUCLIDIENNE, REGROU-
PEMENT AGGLOMERATIF COMPLET) ET MDS ET ACP ASSOCIEES (COULEUR DES ASSEMBLAGES DE LA ERREUR ! SOURCE DU
RENVOI INTROUVABLE. ; GCH GRAND CANAL DU HAVRE, TAN : CANAL DE TANCARVILLE).
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 28
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 16 : CONTRIBUTION RELATIVE (%) DES FACIES SEDIMENTAIRES AU SEIN DES ASSEMBLAGES SEDIMENTAIRES MIS EN
EVIDENCE.
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 29
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
5. COMMUNAUTES MACROBENTHIQUES
5.1. Distribution et composition
La liste des espèces observées lors des deux campagnes de 2016 est en Annexe 2.
Les résultats sur les densités et les biomasses de la macrofaune benthique, par station, sont en Annexe 3.
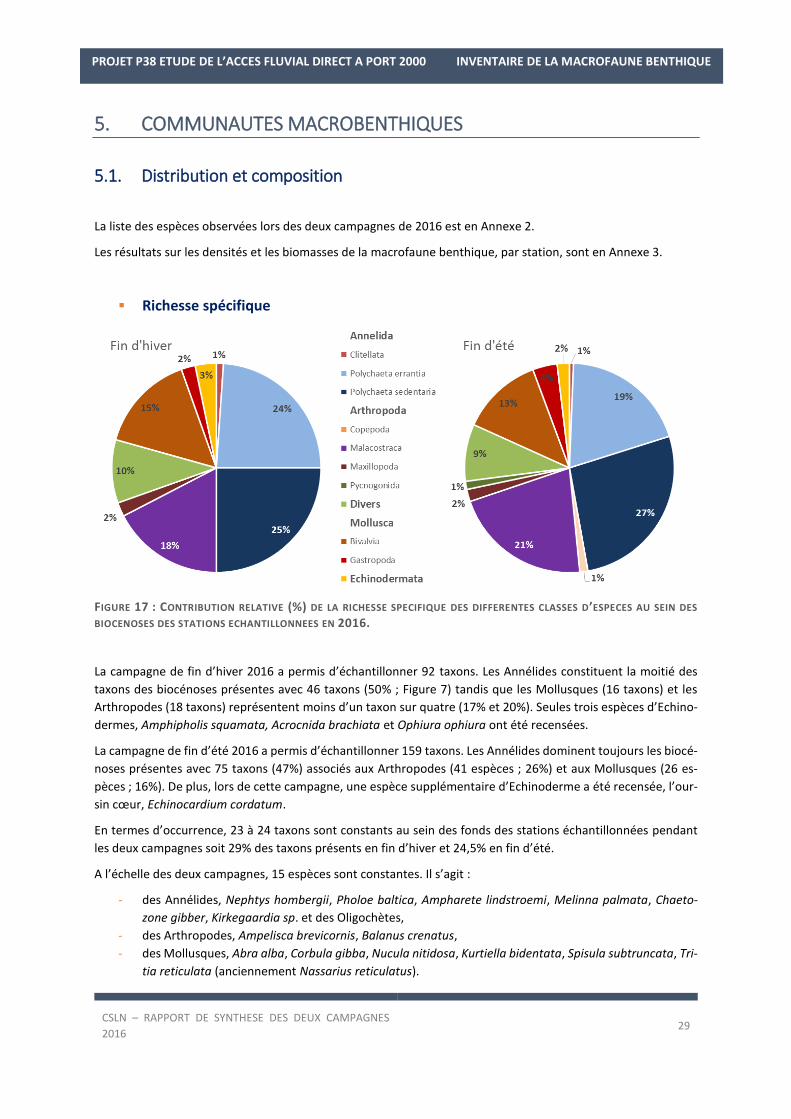
Richesse spécifique
FIGURE 17 : CONTRIBUTION RELATIVE (%) DE LA RICHESSE SPECIFIQUE DES DIFFERENTES CLASSES D’ESPECES AU SEIN DES
BIOCENOSES DES STATIONS ECHANTILLONNEES EN 2016.
La campagne de fin d’hiver 2016 a permis d’échantillonner 92 taxons. Les Annélides constituent la moitié des
taxons des biocénoses présentes avec 46 taxons (50% ; Figure 7) tandis que les Mollusques (16 taxons) et les
Arthropodes (18 taxons) représentent moins d’un taxon sur quatre (17% et 20%). Seules trois espèces d’Echino-
dermes, Amphipholis squamata, Acrocnida brachiata et Ophiura ophiura ont été recensées.
La campagne de fin d’été 2016 a permis d’échantillonner 159 taxons. Les Annélides dominent toujours les biocé-
noses présentes avec 75 taxons (47%) associés aux Arthropodes (41 espèces ; 26%) et aux Mollusques (26 es-
pèces ; 16%). De plus, lors de cette campagne, une espèce supplémentaire d’Echinoderme a été recensée, l’our-
sin cœur, Echinocardium cordatum.
En termes d’occurrence, 23 à 24 taxons sont constants au sein des fonds des stations échantillonnées pendant
les deux campagnes soit 29% des taxons présents en fin d’hiver et 24,5% en fin d’été.
A l’échelle des deux campagnes, 15 espèces sont constantes. Il s’agit :
- des Annélides, Nephtys hombergii, Pholoe baltica, Ampharete lindstroemi, Melinna palmata, Chaeto-
zone gibber, Kirkegaardia sp. et des Oligochètes,
- des Arthropodes, Ampelisca brevicornis, Balanus crenatus,
- des Mollusques, Abra alba, Corbula gibba, Nucula nitidosa, Kurtiella bidentata, Spisula subtruncata, Tri-
tia reticulata (anciennement Nassarius reticulatus).
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 30
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
De plus, 9 espèces introduites sont présentes au sein des biocénoses échantillonnées en 2016, à savoir, les An-
nélides Ficopotomatus enigmaticus (1ère observation dans les bassins portuaires du Havre dans les années 1990),
Hydroides ezoensis (1ère observation dans les bassins portuaires du Havre en 1997), Streblospio benedicti, la ba-
lane, Austrominus modestus (1ère observation dans les bassins portuaires du Havre, env. 1960) et les Mollusques
Ensis directus (1ère observation dans les bassins portuaires du Havre en 2003), Petricolaria pholadiformis (1ère
observation dans les bassins portuaires du Havre dans les années 1990) et Crepidula fornicata (1ère observation
dans les bassins portuaires du Havre entre 1950-1960) et l’ascidie Styela clava (1ère observation dans les bassins
portuaires du Havre en 1968). Une autre espèce d’Annélide Melinna palmata est observée depuis 2006 dans
l’estuaire de Seine et dans le cadre de cette étude mais il s’agit dans son cas, d’une extension de répartition
géographique et non d’une introduction.
Enfin, à l’issue de ces deux campagnes, 28 espèces sont sur la liste des espèces de macrozoobenthos subtidales
déterminantes pour l’inventaire ZNIEFF mer en Haute-Normandie (validée par le CSRPN HN le 10 décembre
2013). Comparativement, entre 2000 et 2014, à l’issue de 14 années de suivi, lors d’inventaires similaires, 40
espèces de la liste des espèces de macrozoobenthos subtidales déterminantes pour l’inventaire ZNIEFF mer en
Haute-Normandie étaient observées au niveau des fonds de l’embouchure, 35 espèces au niveau des fonds du
large de l’estuaire de la Seine et 50 espèces au niveau des fonds des zones d’influence du site d’immersion d’Oc-
teville.
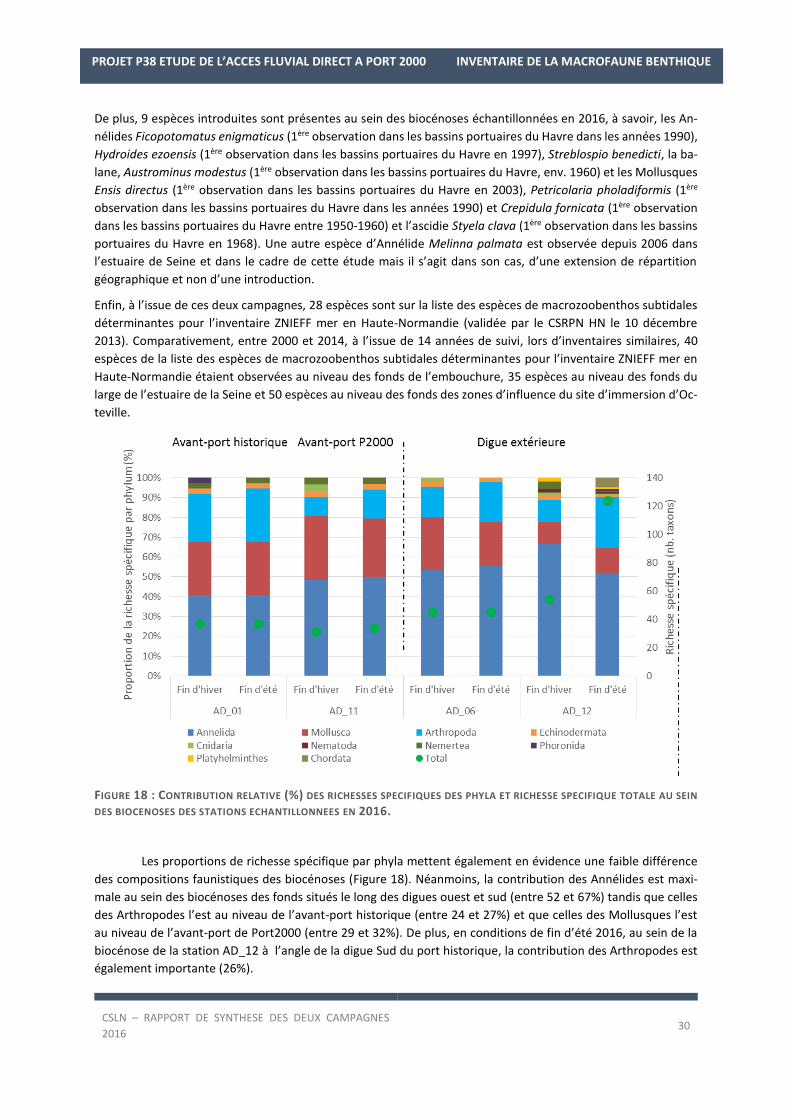
FIGURE 18 : CONTRIBUTION RELATIVE (%) DES RICHESSES SPECIFIQUES DES PHYLA ET RICHESSE SPECIFIQUE TOTALE AU SEIN
DES BIOCENOSES DES STATIONS ECHANTILLONNEES EN 2016.
Les proportions de richesse spécifique par phyla mettent également en évidence une faible différence
des compositions faunistiques des biocénoses (Figure 18). Néanmoins, la contribution des Annélides est maxi-
male au sein des biocénoses des fonds situés le long des digues ouest et sud (entre 52 et 67%) tandis que celles
des Arthropodes l’est au niveau de l’avant-port historique (entre 24 et 27%) et que celles des Mollusques l’est
au niveau de l’avant-port de Port2000 (entre 29 et 32%). De plus, en conditions de fin d’été 2016, au sein de la
biocénose de la station AD_12 à l’angle de la digue Sud du port historique, la contribution des Arthropodes est
également importante (26%).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 31
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Les richesses spécifiques mesurées au sein des biocénoses des stations en 2016 traduisent une diversité
homogène dans l’ensemble avec entre 37 et 54 taxons recensés en fin d’hiver et entre 37 et 45 taxons en fin
d’été observés (en moyenne, 40 ± 8 taxons), seule la richesse spécifique de la biocénose des fonds de l’angle de
la digue Sud du port historique se détachant avec 124 taxons échantillonnés en fin d’été 2016 (Figure 19).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 32
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 19 : REPARTITION SPATIALE DES RICHESSES SPECIFIQUES DES BIOCENOSES AUX STATIONS ECHANTILLONNEES EN
2016.
AD_12 AD_01
AD_06
AD_11
AD_12 AD_01
AD_06
AD_11
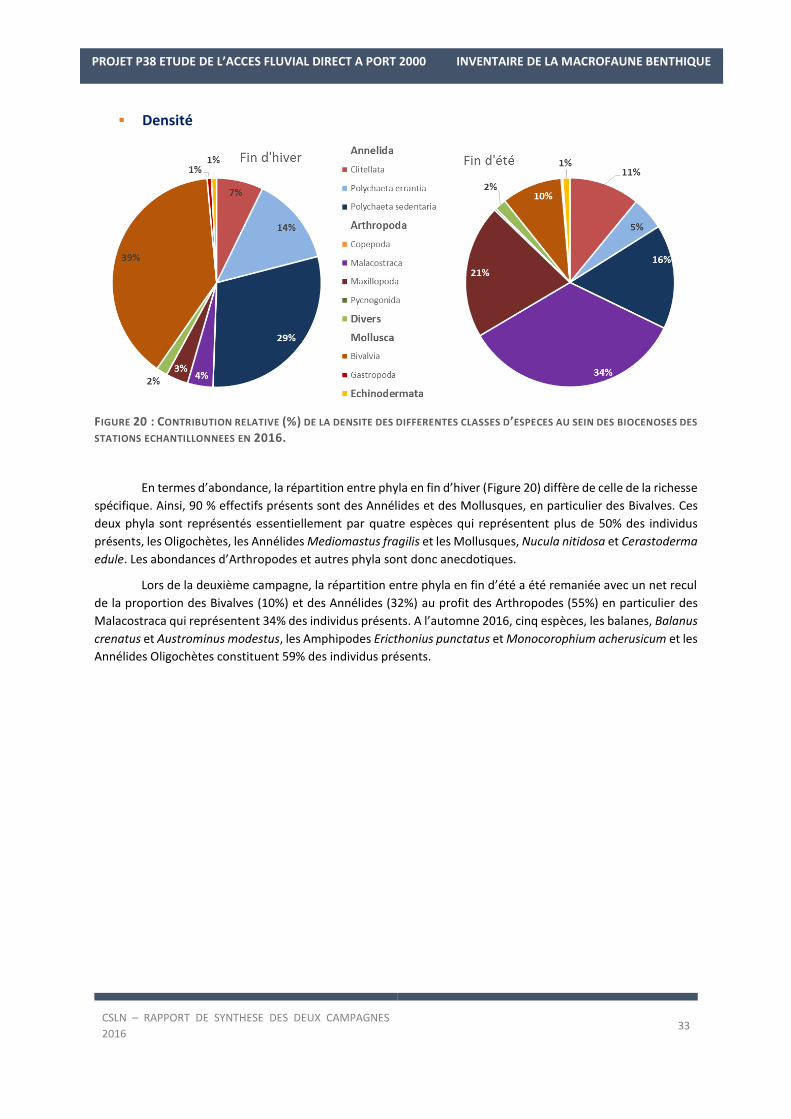
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 33
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Densité
FIGURE 20 : CONTRIBUTION RELATIVE (%) DE LA DENSITE DES DIFFERENTES CLASSES D’ESPECES AU SEIN DES BIOCENOSES DES
STATIONS ECHANTILLONNEES EN 2016.
En termes d’abondance, la répartition entre phyla en fin d’hiver (Figure 20) diffère de celle de la richesse
spécifique. Ainsi, 90 % effectifs présents sont des Annélides et des Mollusques, en particulier des Bivalves. Ces
deux phyla sont représentés essentiellement par quatre espèces qui représentent plus de 50% des individus
présents, les Oligochètes, les Annélides Mediomastus fragilis et les Mollusques, Nucula nitidosa et Cerastoderma
edule. Les abondances d’Arthropodes et autres phyla sont donc anecdotiques.
Lors de la deuxième campagne, la répartition entre phyla en fin d’été a été remaniée avec un net recul
de la proportion des Bivalves (10%) et des Annélides (32%) au profit des Arthropodes (55%) en particulier des
Malacostraca qui représentent 34% des individus présents. A l’automne 2016, cinq espèces, les balanes, Balanus
crenatus et Austrominus modestus, les Amphipodes Ericthonius punctatus et Monocorophium acherusicum et les
Annélides Oligochètes constituent 59% des individus présents.
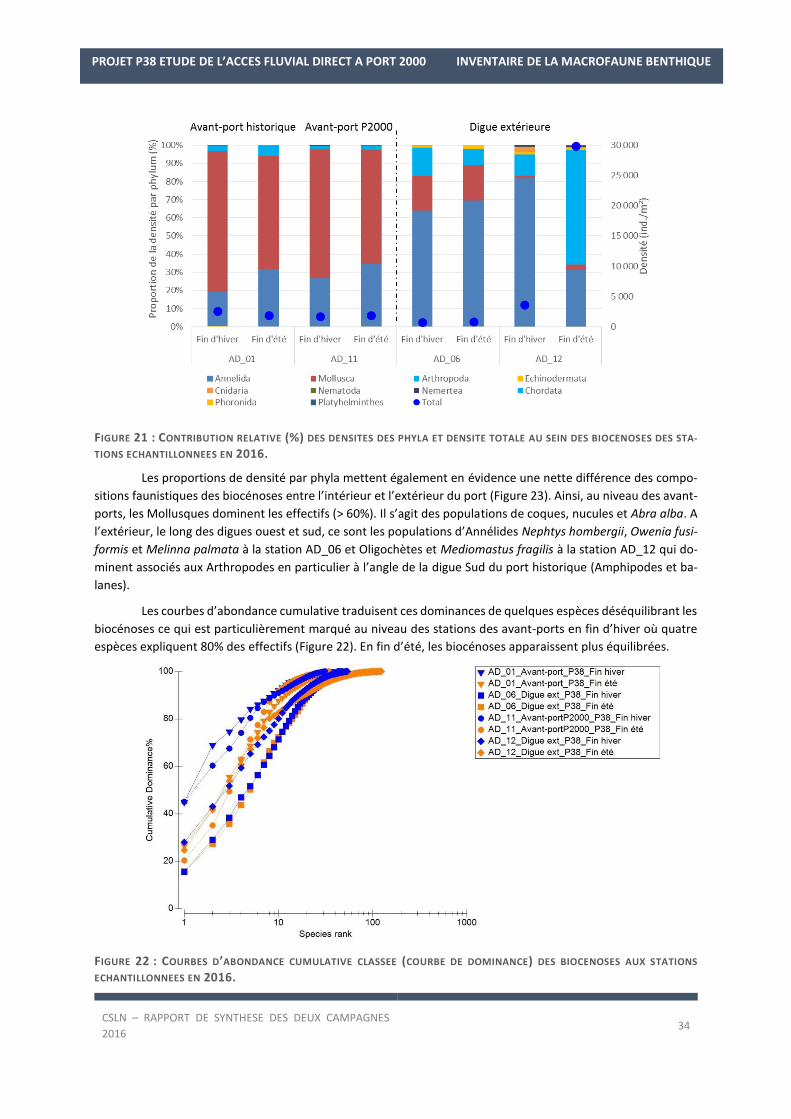
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 34
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 21 : CONTRIBUTION RELATIVE (%) DES DENSITES DES PHYLA ET DENSITE TOTALE AU SEIN DES BIOCENOSES DES STA-
TIONS ECHANTILLONNEES EN 2016.
Les proportions de densité par phyla mettent également en évidence une nette différence des compo-
sitions faunistiques des biocénoses entre l’intérieur et l’extérieur du port (Figure 23). Ainsi, au niveau des avant-
ports, les Mollusques dominent les effectifs (> 60%). Il s’agit des populations de coques, nucules et Abra alba. A
l’extérieur, le long des digues ouest et sud, ce sont les populations d’Annélides Nephtys hombergii, Owenia fusi-
formis et Melinna palmata à la station AD_06 et Oligochètes et Mediomastus fragilis à la station AD_12 qui do-
minent associés aux Arthropodes en particulier à l’angle de la digue Sud du port historique (Amphipodes et ba-
lanes).
Les courbes d’abondance cumulative traduisent ces dominances de quelques espèces déséquilibrant les
biocénoses ce qui est particulièrement marqué au niveau des stations des avant-ports en fin d’hiver où quatre
espèces expliquent 80% des effectifs (Figure 22). En fin d’été, les biocénoses apparaissent plus équilibrées.
FIGURE 22 : COURBES D’ABONDANCE CUMULATIVE CLASSEE (COURBE DE DOMINANCE) DES BIOCENOSES AUX STATIONS
ECHANTILLONNEES EN 2016.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 35
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 23 : REPARTITION SPATIALE DES DENSITES TOTALES DES BIOCENOSES AUX STATIONS ECHANTILLONNEES EN 2016.
AD_12 AD_01
AD_06
AD_11
AD_12
AD_01
AD_06
AD_11
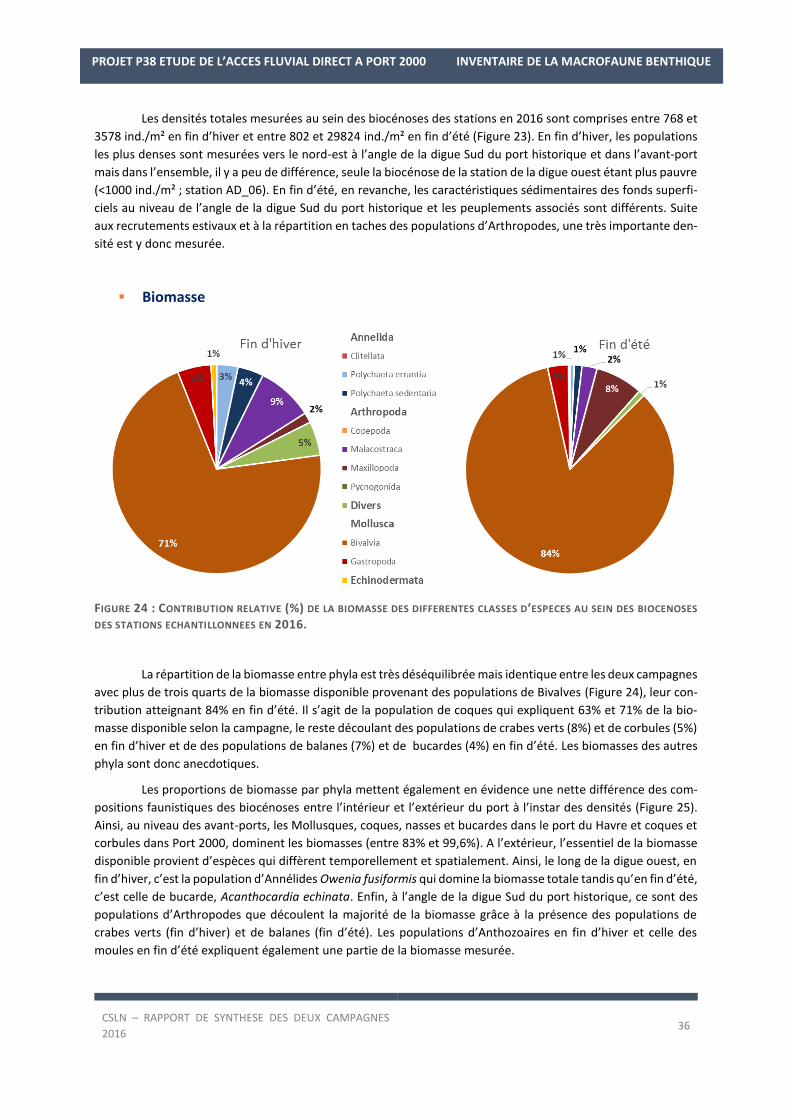
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 36
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Les densités totales mesurées au sein des biocénoses des stations en 2016 sont comprises entre 768 et
3578 ind./m² en fin d’hiver et entre 802 et 29824 ind./m² en fin d’été (Figure 23). En fin d’hiver, les populations
les plus denses sont mesurées vers le nord-est à l’angle de la digue Sud du port historique et dans l’avant-port
mais dans l’ensemble, il y a peu de différence, seule la biocénose de la station de la digue ouest étant plus pauvre
(<1000 ind./m² ; station AD_06). En fin d’été, en revanche, les caractéristiques sédimentaires des fonds superfi-
ciels au niveau de l’angle de la digue Sud du port historique et les peuplements associés sont différents. Suite
aux recrutements estivaux et à la répartition en taches des populations d’Arthropodes, une très importante den-
sité est y donc mesurée.
Biomasse
FIGURE 24 : CONTRIBUTION RELATIVE (%) DE LA BIOMASSE DES DIFFERENTES CLASSES D’ESPECES AU SEIN DES BIOCENOSES
DES STATIONS ECHANTILLONNEES EN 2016.
La répartition de la biomasse entre phyla est très déséquilibrée mais identique entre les deux campagnes
avec plus de trois quarts de la biomasse disponible provenant des populations de Bivalves (Figure 24), leur con-
tribution atteignant 84% en fin d’été. Il s’agit de la population de coques qui expliquent 63% et 71% de la bio-
masse disponible selon la campagne, le reste découlant des populations de crabes verts (8%) et de corbules (5%)
en fin d’hiver et de des populations de balanes (7%) et de bucardes (4%) en fin d’été. Les biomasses des autres
phyla sont donc anecdotiques.
Les proportions de biomasse par phyla mettent également en évidence une nette différence des com-
positions faunistiques des biocénoses entre l’intérieur et l’extérieur du port à l’instar des densités (Figure 25).
Ainsi, au niveau des avant-ports, les Mollusques, coques, nasses et bucardes dans le port du Havre et coques et
corbules dans Port 2000, dominent les biomasses (entre 83% et 99,6%). A l’extérieur, l’essentiel de la biomasse
disponible provient d’espèces qui diffèrent temporellement et spatialement. Ainsi, le long de la digue ouest, en
fin d’hiver, c’est la population d’Annélides Owenia fusiformis qui domine la biomasse totale tandis qu’en fin d’été,
c’est celle de bucarde, Acanthocardia echinata. Enfin, à l’angle de la digue Sud du port historique, ce sont des
populations d’Arthropodes que découlent la majorité de la biomasse grâce à la présence des populations de
crabes verts (fin d’hiver) et de balanes (fin d’été). Les populations d’Anthozoaires en fin d’hiver et celle des
moules en fin d’été expliquent également une partie de la biomasse mesurée.
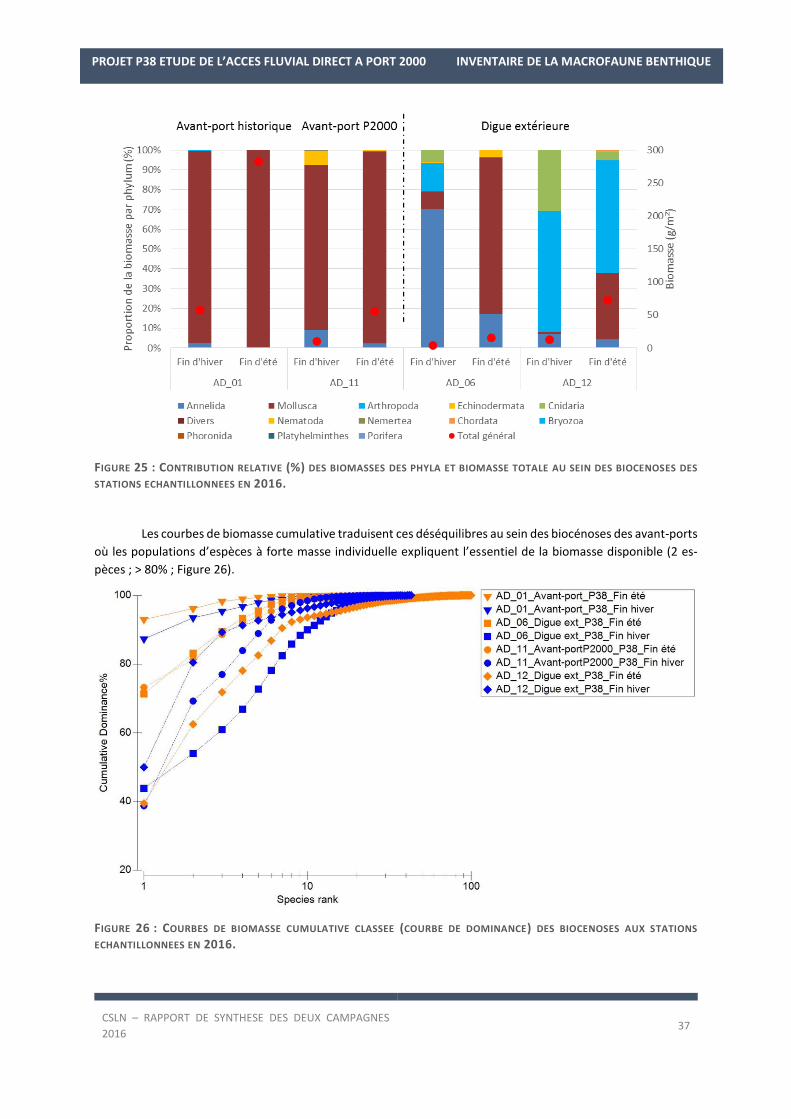
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 37
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 25 : CONTRIBUTION RELATIVE (%) DES BIOMASSES DES PHYLA ET BIOMASSE TOTALE AU SEIN DES BIOCENOSES DES
STATIONS ECHANTILLONNEES EN 2016.
Les courbes de biomasse cumulative traduisent ces déséquilibres au sein des biocénoses des avant-ports
où les populations d’espèces à forte masse individuelle expliquent l’essentiel de la biomasse disponible (2 es-
pèces ; > 80% ; Figure 26).
FIGURE 26 : COURBES DE BIOMASSE CUMULATIVE CLASSEE (COURBE DE DOMINANCE) DES BIOCENOSES AUX STATIONS
ECHANTILLONNEES EN 2016.
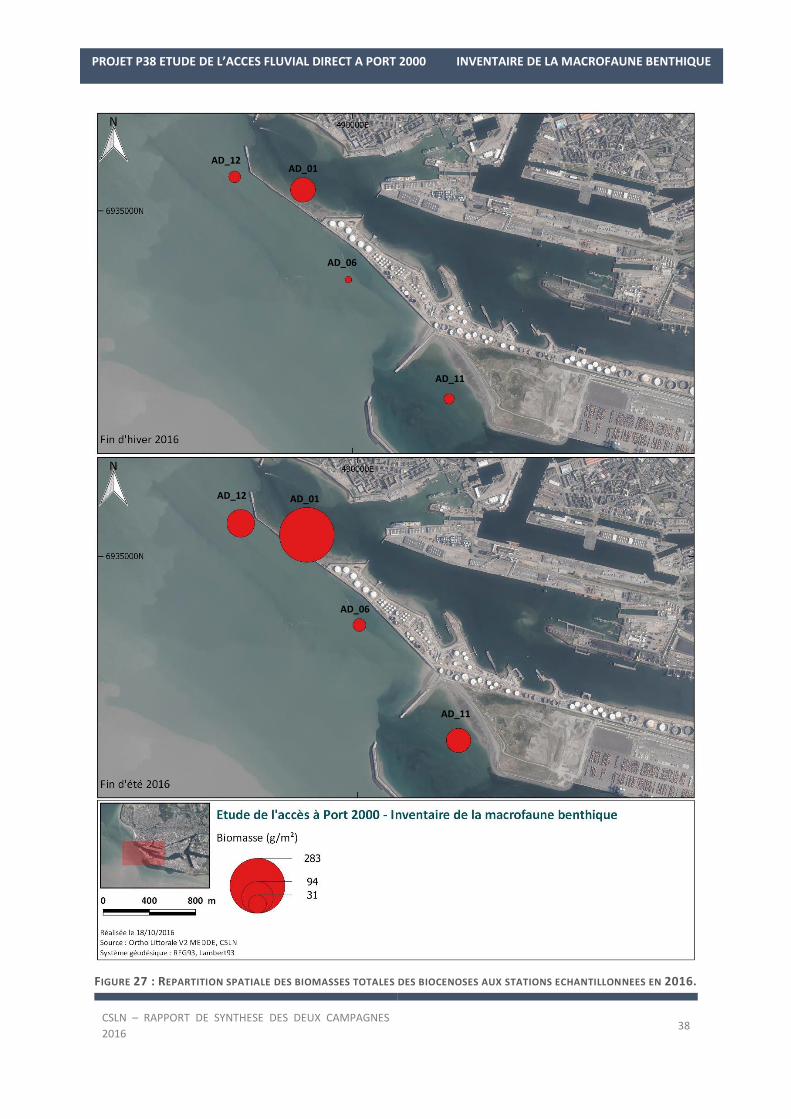
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 38
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 27 : REPARTITION SPATIALE DES BIOMASSES TOTALES DES BIOCENOSES AUX STATIONS ECHANTILLONNEES EN 2016.
AD_12 AD_01
AD_06
AD_11
AD_12 AD_01
AD_06
AD_11
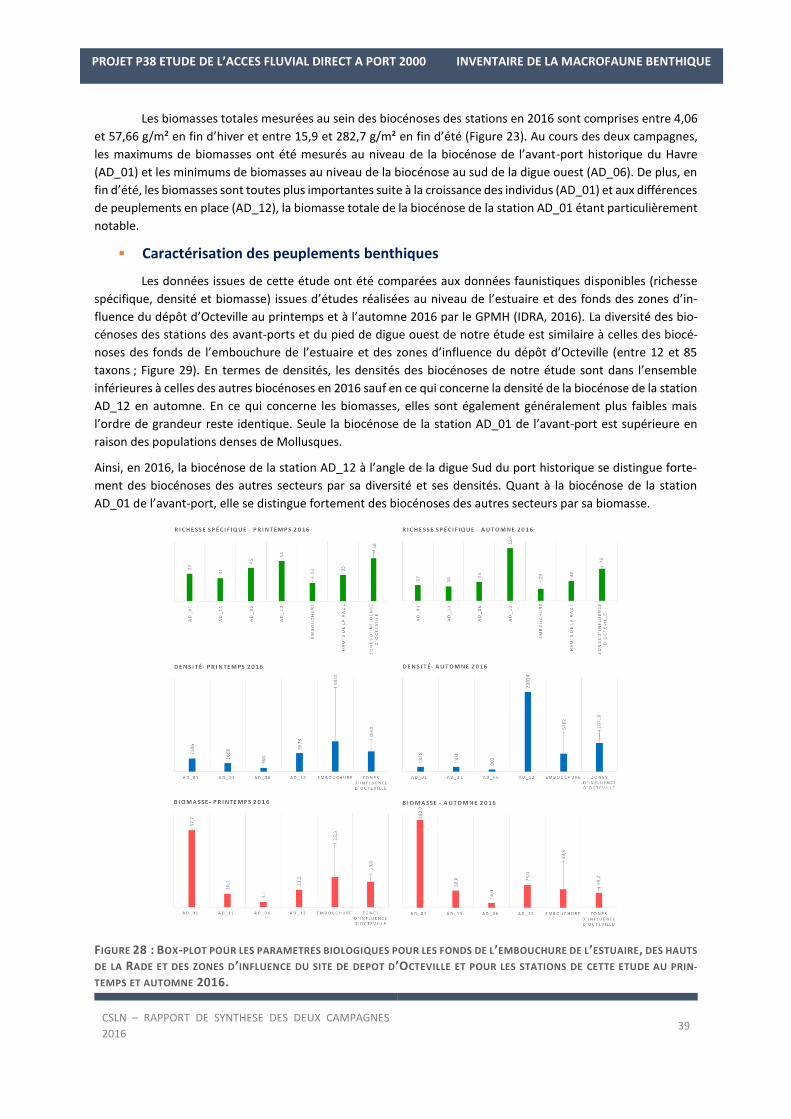
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 39
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Les biomasses totales mesurées au sein des biocénoses des stations en 2016 sont comprises entre 4,06
et 57,66 g/m² en fin d’hiver et entre 15,9 et 282,7 g/m² en fin d’été (Figure 23). Au cours des deux campagnes,
les maximums de biomasses ont été mesurés au niveau de la biocénose de l’avant-port historique du Havre
(AD_01) et les minimums de biomasses au niveau de la biocénose au sud de la digue ouest (AD_06). De plus, en
fin d’été, les biomasses sont toutes plus importantes suite à la croissance des individus (AD_01) et aux différences
de peuplements en place (AD_12), la biomasse totale de la biocénose de la station AD_01 étant particulièrement
notable.
Caractérisation des peuplements benthiques
Les données issues de cette étude ont été comparées aux données faunistiques disponibles (richesse
spécifique, densité et biomasse) issues d’études réalisées au niveau de l’estuaire et des fonds des zones d’in-
fluence du dépôt d’Octeville au printemps et à l’automne 2016 par le GPMH (IDRA, 2016). La diversité des bio-
cénoses des stations des avant-ports et du pied de digue ouest de notre étude est similaire à celles des biocé-
noses des fonds de l’embouchure de l’estuaire et des zones d’influence du dépôt d’Octeville (entre 12 et 85
taxons ; Figure 29). En termes de densités, les densités des biocénoses de notre étude sont dans l’ensemble
inférieures à celles des autres biocénoses en 2016 sauf en ce qui concerne la densité de la biocénose de la station
AD_12 en automne. En ce qui concerne les biomasses, elles sont également généralement plus faibles mais
l’ordre de grandeur reste identique. Seule la biocénose de la station AD_01 de l’avant-port est supérieure en
raison des populations denses de Mollusques.
Ainsi, en 2016, la biocénose de la station AD_12 à l’angle de la digue Sud du port historique se distingue forte-
ment des biocénoses des autres secteurs par sa diversité et ses densités. Quant à la biocénose de la station
AD_01 de l’avant-port, elle se distingue fortement des biocénoses des autres secteurs par sa biomasse.
FIGURE 28 : BOX-PLOT POUR LES PARAMETRES BIOLOGIQUES POUR LES FONDS DE L’EMBOUCHURE DE L’ESTUAIRE, DES HAUTS
DE LA RADE ET DES ZONES D’INFLUENCE DU SITE DE DEPOT D’OCTEVILLE ET POUR LES STATIONS DE CETTE ETUDE AU PRIN-
TEMPS ET AUTOMNE 2016.
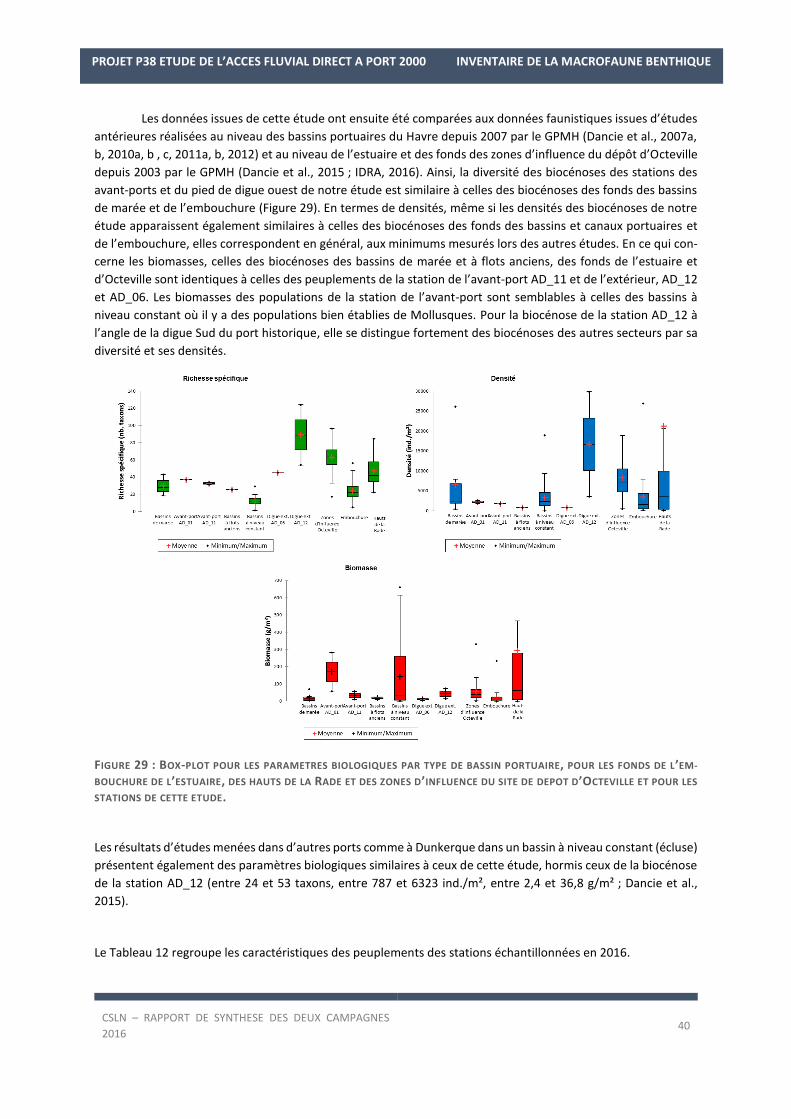
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 40
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Les données issues de cette étude ont ensuite été comparées aux données faunistiques issues d’études
antérieures réalisées au niveau des bassins portuaires du Havre depuis 2007 par le GPMH (Dancie et al., 2007a,
b, 2010a, b , c, 2011a, b, 2012) et au niveau de l’estuaire et des fonds des zones d’influence du dépôt d’Octeville
depuis 2003 par le GPMH (Dancie et al., 2015 ; IDRA, 2016). Ainsi, la diversité des biocénoses des stations des
avant-ports et du pied de digue ouest de notre étude est similaire à celles des biocénoses des fonds des bassins
de marée et de l’embouchure (Figure 29). En termes de densités, même si les densités des biocénoses de notre
étude apparaissent également similaires à celles des biocénoses des fonds des bassins et canaux portuaires et
de l’embouchure, elles correspondent en général, aux minimums mesurés lors des autres études. En ce qui con-
cerne les biomasses, celles des biocénoses des bassins de marée et à flots anciens, des fonds de l’estuaire et
d’Octeville sont identiques à celles des peuplements de la station de l’avant-port AD_11 et de l’extérieur, AD_12
et AD_06. Les biomasses des populations de la station de l’avant-port sont semblables à celles des bassins à
niveau constant où il y a des populations bien établies de Mollusques. Pour la biocénose de la station AD_12 à
l’angle de la digue Sud du port historique, elle se distingue fortement des biocénoses des autres secteurs par sa
diversité et ses densités.
FIGURE 29 : BOX-PLOT POUR LES PARAMETRES BIOLOGIQUES PAR TYPE DE BASSIN PORTUAIRE, POUR LES FONDS DE L’EM-
BOUCHURE DE L’ESTUAIRE, DES HAUTS DE LA RADE ET DES ZONES D’INFLUENCE DU SITE DE DEPOT D’OCTEVILLE ET POUR LES
STATIONS DE CETTE ETUDE.
Les résultats d’études menées dans d’autres ports comme à Dunkerque dans un bassin à niveau constant (écluse)
présentent également des paramètres biologiques similaires à ceux de cette étude, hormis ceux de la biocénose
de la station AD_12 (entre 24 et 53 taxons, entre 787 et 6323 ind./m², entre 2,4 et 36,8 g/m² ; Dancie et al.,
2015).
Le Tableau 12 regroupe les caractéristiques des peuplements des stations échantillonnées en 2016.
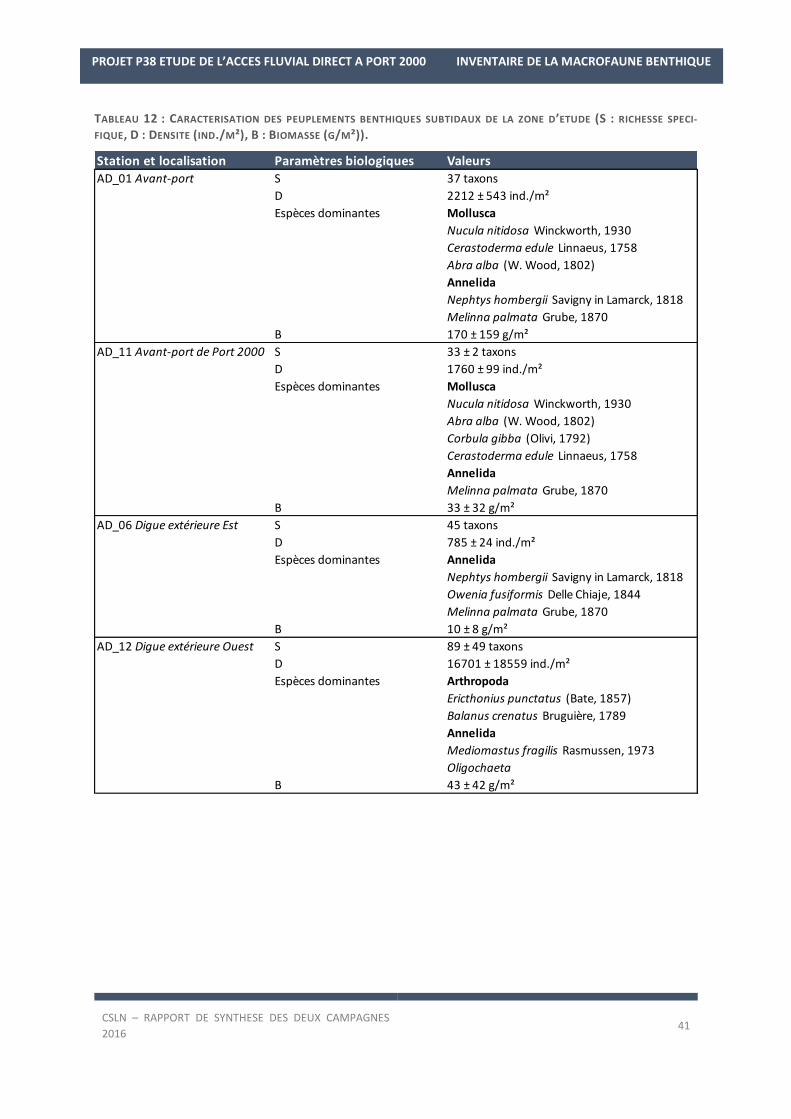
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 41
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
TABLEAU 12 : CARACTERISATION DES PEUPLEMENTS BENTHIQUES SUBTIDAUX DE LA ZONE D’ETUDE (S : RICHESSE SPECI-
FIQUE, D : DENSITE (IND./M²), B : BIOMASSE (G/M²)).
Station et localisation Paramètres biologiques ValeursAD_01 Avant-port S 37 taxons
D 2212 ± 543 ind./m²
Espèces dominantes Mollusca
Nucula nitidosa Winckworth, 1930
Cerastoderma edule Linnaeus, 1758
Abra alba (W. Wood, 1802)
Annelida
Nephtys hombergii Savigny in Lamarck, 1818
Melinna palmata Grube, 1870
B 170 ± 159 g/m²
AD_11 Avant-port de Port 2000 S 33 ± 2 taxons
D 1760 ± 99 ind./m²
Espèces dominantes Mollusca
Nucula nitidosa Winckworth, 1930
Abra alba (W. Wood, 1802)
Corbula gibba (Olivi, 1792)
Cerastoderma edule Linnaeus, 1758
Annelida
Melinna palmata Grube, 1870
B 33 ± 32 g/m²
AD_06 Digue extérieure Est S 45 taxons
D 785 ± 24 ind./m²
Espèces dominantes Annelida
Nephtys hombergii Savigny in Lamarck, 1818
Owenia fusiformis Delle Chiaje, 1844
Melinna palmata Grube, 1870
B 10 ± 8 g/m²
AD_12 Digue extérieure Ouest S 89 ± 49 taxons
D 16701 ± 18559 ind./m²
Espèces dominantes Arthropoda
Ericthonius punctatus (Bate, 1857)
Balanus crenatus Bruguière, 1789
Annelida
Mediomastus fragilis Rasmussen, 1973
Oligochaeta
B 43 ± 42 g/m²
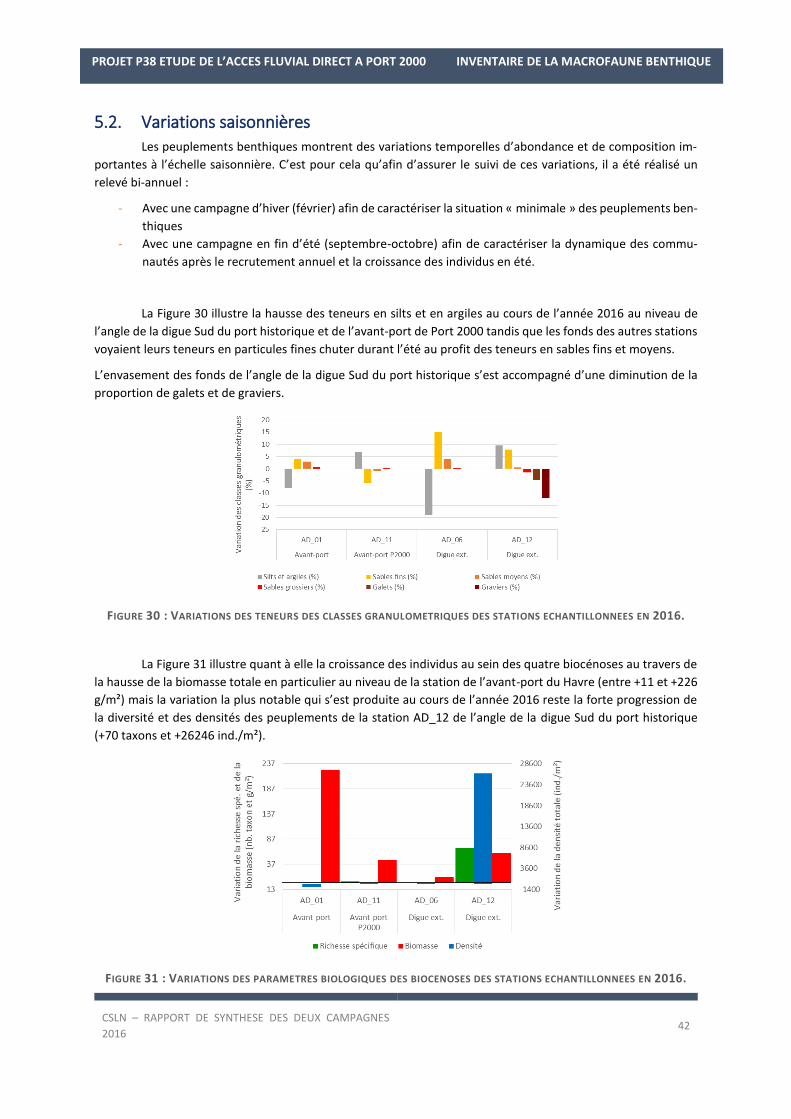
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 42
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
5.2. Variations saisonnières Les peuplements benthiques montrent des variations temporelles d’abondance et de composition im-
portantes à l’échelle saisonnière. C’est pour cela qu’afin d’assurer le suivi de ces variations, il a été réalisé un
relevé bi-annuel :
- Avec une campagne d’hiver (février) afin de caractériser la situation « minimale » des peuplements ben-
thiques
- Avec une campagne en fin d’été (septembre-octobre) afin de caractériser la dynamique des commu-
nautés après le recrutement annuel et la croissance des individus en été.
La Figure 30 illustre la hausse des teneurs en silts et en argiles au cours de l’année 2016 au niveau de
l’angle de la digue Sud du port historique et de l’avant-port de Port 2000 tandis que les fonds des autres stations
voyaient leurs teneurs en particules fines chuter durant l’été au profit des teneurs en sables fins et moyens.
L’envasement des fonds de l’angle de la digue Sud du port historique s’est accompagné d’une diminution de la
proportion de galets et de graviers.
FIGURE 30 : VARIATIONS DES TENEURS DES CLASSES GRANULOMETRIQUES DES STATIONS ECHANTILLONNEES EN 2016.
La Figure 31 illustre quant à elle la croissance des individus au sein des quatre biocénoses au travers de
la hausse de la biomasse totale en particulier au niveau de la station de l’avant-port du Havre (entre +11 et +226
g/m²) mais la variation la plus notable qui s’est produite au cours de l’année 2016 reste la forte progression de
la diversité et des densités des peuplements de la station AD_12 de l’angle de la digue Sud du port historique
(+70 taxons et +26246 ind./m²).
FIGURE 31 : VARIATIONS DES PARAMETRES BIOLOGIQUES DES BIOCENOSES DES STATIONS ECHANTILLONNEES EN 2016.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 43
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
A l’issue de la dernière campagne, les biocénoses des stations des avant-ports sont sensiblement iden-
tiques en termes de composition faunistique. Au niveau de l’avant-port historique du Havre, les populations de
Mollusques dominants, coques et Nucula nitidosa ont vu leur densité chuter mais la croissance des individus a
entrainé une hausse notable des biomasses spécifiques qui se sont répercutées sur la biomasse totale. La popu-
lation de nucules de l’avant-port de Port2000 s’est également appauvrie mais celles d’Abra alba et coques pré-
sentent des densités et des biomasses en hausse. Les populations de Melinna palmata et d’Annélides opportu-
nistes progressent également. Mais, les contributions en termes de richesse spécifique, densité et biomasse des
phyla se maintiennent entre les deux campagnes.
Le long de la digue ouest, la biocénose est également similaire entre les campagnes, une légère diversi-
fication des espèces opportunistes étant néanmoins observée. La hausse de la biomasse découle quant à elle de
l’échantillonnage en septembre 2016 de bucarde, espèce à forte biomasse individuelle ce qui explique la forte
hausse de la contribution des Mollusques à la biomasse totale.
Au niveau de l’angle de la digue Sud du port historique, les fonds se sont envasés entre les deux cam-
pagnes et une importante diversification des peuplements est mesurée avec la présence d’espèces d’Arthro-
podes, cirripèdes, amphipodes et décapodes, supplémentaires. La forte hausse de la densité totale s’explique
par l’échantillonnage de nombreux galets recouverts de balanes, Austrominius modestus et Balanus crenatus qui
peuvent présenter des très importantes abondances mais également par la présence de nombreux amphipodes
tubicoles comme Ericthonius punctatus qui présentent des répartitions en taches et donc de fortes densités. Ces
nouvelles populations très denses ont remanié les contributions des phyla avec une nette hausse de celle des
Arthropodes au déficit de celle des Annélides. En ce qui concerne les biomasses, la composition est également
remaniée avec une baisse de la contribution des Anthozoaires au profit de celle des Mollusques ce qui s’explique
par l’échantillonnage d’une population dense de moules, espèce à forte biomasse individuelle.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 44
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
5.3. Caractérisation des assemblages faunistiques et Habitats benthiques La classification ascendante hiérarchique (CAH) a été appliquée aux résultats de densité transformés
issues des stations échantillonnées en 2016 dans le cadre de cette étude (Fourth-Root ; dissimilarité de Bray-
Curtis). Le dendrogramme et l’analyse multidimensionelle non-métrique (MDS) obtenus illustrent les assem-
blages de stations (Figure 32 : Classification ascendante hiérarchique (données des fractions faunistiques trans-
formées fourth root ; distance de Bray-curtis, regroupement agglomératif moyen) et MDS associées.Figure 32).
La CAH, le test SIMPROF et l’analyse conjointe de la MDS ont permis d’isoler trois assemblages de stations. La
typologie d’habitats des habitats marins benthiques de la Manche, de la Mer du Nord et de l’Atlantique (Michez
et al., 2015) et celle EUNIS Habitat Classification (2004 et 2012 - a revision of the habitat classification descrip-
tions), MNHN (Michez et al., 2015) sont précisées.
FIGURE 32 : CLASSIFICATION ASCENDANTE HIERARCHIQUE (DONNEES DES FRACTIONS FAUNISTIQUES TRANSFORMEES
FOURTH ROOT ; DISTANCE DE BRAY-CURTIS, REGROUPEMENT AGGLOMERATIF MOYEN) ET MDS ASSOCIEES.
Ainsi, sont isolées et regroupées les biocénoses des fonds de vases portuaires des avant-ports (72,4 ± 4,4% de
silts et argiles en moyenne) caractérisées par les richesses spécifiques les plus faibles de l’étude (35 ± 3 taxons
en moyenne) et des densités moyennes (1986 ±412 ind./m²) mais des biomasses importantes (101 ± 123 g/m²).
Les espèces indicatrices de cet assemblage sont les coques, les nucules, les corbules, les nasses et Euchone lim-
nicola. Ces populations de Mollusques associées aux populations d’Annélides, Melinna palmata et Nephtys hom-
bergii dominent les effectifs. Il correspond à la communauté à Abra alba et Lagis koreni dans une variante ap-
pauvrie de cette communauté avec la présence de Corbicula gibba qui caractérise un faciès très particulier de la
communauté à Abra alba, espèce qu’il tend à remplacer. Il peut correspondre à deux habitats i) l’habitat A5.331
Nephtys hombergii and Macoma balthica in infralittoral sandy mud (M10.01.01.01) retrouvé dans les vases sa-
bleuses et vases, et parfois sédiments hétérogènes, riches en contenu organique, en eau peu profonde près du
rivage, pouvant être caractérisés par la présence du polychète Nephtys hombergii et du bivalve Macoma balthica,
Abra alba et Nucula nitidosa peuvent aussi être des espèces importantes ; ii) l’habitat A5.341 Cerastoderma edule
with Abra nitida in infralittoral mud (M10.02.01.01 Vases infralittorales à Cerastoderma edule avec Abra nitida),
rencontré au niveau des vases et vases graveleuses infralittorales des ports, et hébergeant des populations den-
ses de coques. Les autres taxons représentés peuvent comprendre le gastéropode Peringia ulvae (anciennement

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 45
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Hydrobia ulvae), des polychètes cirratulidés tels que Caulleriella spp., ainsi que d’autres polychètes, dont Hediste
diversicolor et Aphelochaeta marioni
La biocénose de la station AD_06 de la digue ouest est caractérisée par une richesse spécifique moyenne (45
taxons) et de faibles densités et biomasses (785 ± 24 ind./m² ; 10 ± 8 g/m²). Les espèces indicatrices sont les
amphipodes Ampelisca brevicornis et les amphiures, Acrocnida brachiata. Les populations d’Annélides Melinna
palmata, Owenia fusiformis et Nephtys hombergii dominent les effectifs.
Enfin, la biocénose de la station AD_12 de la digue Sud du port historique est isolée de par ses fractions impor-
tantes en graviers et galets au sein des sédiments superficiels qui abritent un grand nombre de peuplements très
denses (89 ± 49 taxons ; 16701 ± 18559 ind./m²). Les espèces indicatrices sont les Annélides Pholoe baltica et
Harmothoe impar. Les effectifs sont dominés par les Arthropodes, Ericthonius punctatus et les balanes ainsi que
par les Annélides Mediomastus fragilis et les Oligochètes. (Habitats A5.13 infralittoral coarse sediment et A5.14
Circalittoral coarse sediment dont A5.142 Mediomastus fragilis, Lumbrineris spp. and venerid bivalves in circalit-
toral coarse sand or gravel).
Les données issues de cette étude ont été comparées aux données faunistiques issues d’études anté-
rieures réalisées au niveau des bassins portuaires du Havre depuis 2007 par le GPMH (Dancie et al., 2007a, b,
2010a, b , c, 2011a, b, 2012). La Figure 15 présente le dendrogramme issu de cette analyse et la MDS et l’ACP
illustrent les caractéristiques faunistiques de chaque assemblage.
Cette CAH a permis d’identifier quatre assemblages dont un sera détaillé suivant les quatre sous–assemblages le
constituant. Cinq zones sont clairement distinguées : les canaux portuaires, les bassins de marée, l’ancien bassin
aux pétroles, le recoin dans le quai nord du bassin René Coty et des espaces plus restreints au niveau ou à proxi-
mité des digues.
Les cortèges faunistiques de ces assemblages se rattachent aux communautés et aux habitats habituellement
décrits dans l’estuaire de la Seine et dans les bassins portuaires et sont détaillées ci-après.
Ainsi, les biocénoses des avant-ports et de la digue ouest de la présente étude (AD_01, AD_11 et AD_06)
sont rattachées à celles des autres bassins de marée étudiés, le long du quai Joannès Couvert et du bassin Théo-
phile Ducrocq. Cet assemblage présente des paramètres biologiques moyens (34 ± 9 taxons, 2211 ± 2085 ind./m²
et 39 ± 79 g/m² en moyenne) et est caractérisé par Nephtys hombergii et Nucula nitidosa. Les Annélides Euchone
limnicola et les Oligochètes, les nucules et Abra alba dominent les effectifs.
Les peuplements présents au niveau de l’angle de la digue Sud du port historique (AD_12) sont rattachés
à ceux proches des digues du fond du bassin Théophile Ducrocq. Caractérisé par Pholoe inornata et les balanes,
Balanus crenatus, il est dominé par les espèces opportunistes, Oligochètes et Cirriformia tentaculata et les ba-
lanes ce qui explique les densités pouvant être élevées en raison de leur faculté à proliférer (en moyenne 8682
± 11902 ind./m²). La présence de populations d’Amphipodes en taches en est également une raison. Ce mode de
répartition explique également les écart-types élevés.
Les peuplements du bassin René Coty au niveau du renfoncement entre le quai de l’Atlantique et le quai
des Amériques sont isolés par la CAH avec l’assemblage à Phyllodoce mucosa et Capitella sp.. Dominée numéri-
quement par la population de Capitella sp., cette biocénose présente d’importantes densités, les plus impor-
tantes du complexe portuaire (22593 ± 5016 ind./m²), les richesses spécifiques et les biomasses étant relative-
ment faibles (22 ± 4 taxons et 16 ± 13 g/m² en moyenne).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 46
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Au niveau de l’ancien bassin aux pétroles, bassin à niveau constant, les biocénoses sont également dif-
férentes. Caractérisées par l’ascidie Ascidiella aspersa et l’amphipode, Ampelisca tenuicornis, les espèces oppor-
tunistes comme Cirriformia tentaculata dominent numériquement.
Les fonds des canaux portuaires, Grand Canal du Havre et Canal de Tancarville, composés de vases sa-
bleuses et de sables vaseux abritent des biocénoses pauvres (entre 13 ± 5 et 7 ± 1 taxons en moyenne) pouvant
présenter des densités notables découlant des populations de Mollusques, d’Annélides opportunistes et d’Am-
phipodes (entre 3427 ± 3204 et 3583 ± 4521 ind./m² en moyenne) et des biomasses importantes si des moulières
sont présentes (164 ± 202 g/m²). Ces biocénoses sont regroupées en deux assemblages caractérisés d’une part
par Alitta succinea et Polydora ciliata et d’autre part, par Gammarus locusta et Capitella sp.
Pour finir, un assemblage regroupe trois biocénoses situées au milieu du Grand Canal du Havre et ca-
ractérisées par un peuplement oligospécifique - seule espèce présente, Corbula gibba - peu dense et à faible
biomasse.
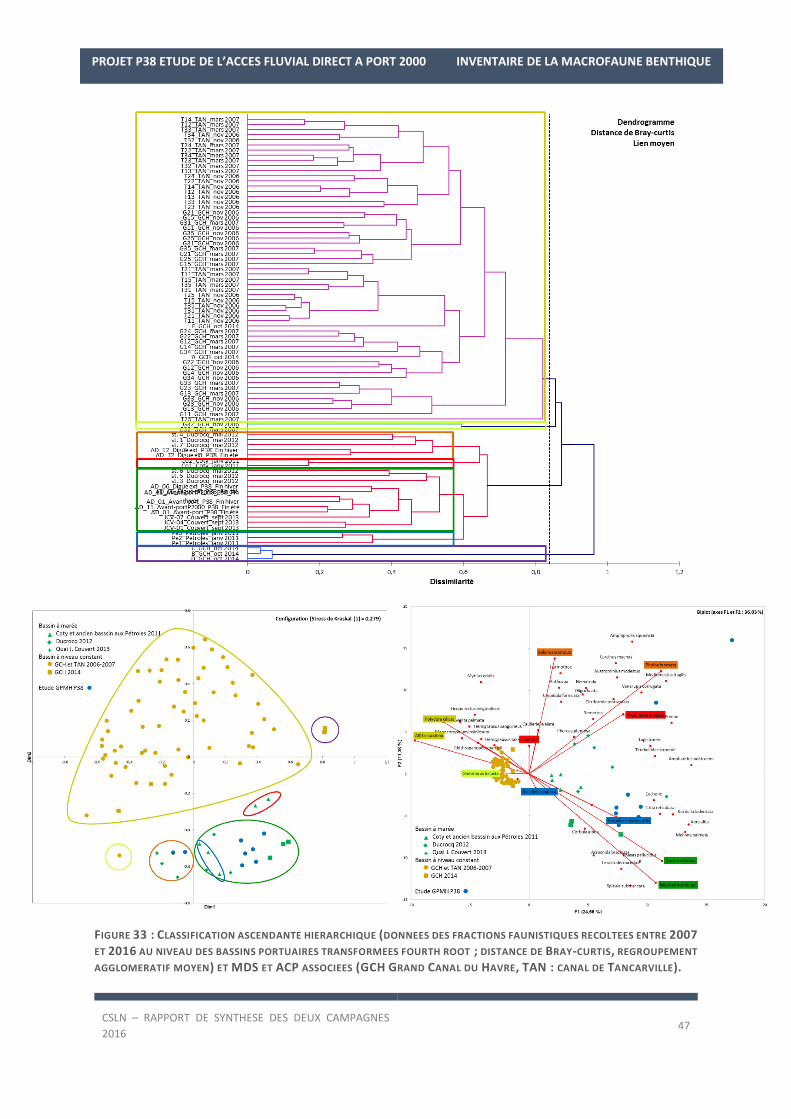
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 47
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
FIGURE 33 : CLASSIFICATION ASCENDANTE HIERARCHIQUE (DONNEES DES FRACTIONS FAUNISTIQUES RECOLTEES ENTRE 2007
ET 2016 AU NIVEAU DES BASSINS PORTUAIRES TRANSFORMEES FOURTH ROOT ; DISTANCE DE BRAY-CURTIS, REGROUPEMENT
AGGLOMERATIF MOYEN) ET MDS ET ACP ASSOCIEES (GCH GRAND CANAL DU HAVRE, TAN : CANAL DE TANCARVILLE).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 48
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
5.4. Sensibilité des peuplements, indices écologiques, et enjeux
FIGURE 34 : CONTRIBUTION RELATIVE (%) DES DENSITES DES GROUPES ECOLOGIQUES ET VALEUR DE L’INDICE AMBI AU SEIN
DES BIOCENOSES DES STATIONS ECHANTILLONNEES EN 2016.
Au sein des avant-ports, les espèces tolérantes stimulées par l’enrichissement en matière organique (GE
III), Abra alba, coques et Melinna palmata dominent les biocénoses associées aux espèces sensibles (GE I), Nucula
nitidosa en fin d’hiver et aux espèces opportunistes (GE IV et V), Corbula gibba, Oligochètes et Pholoe inornata
en fin d’été.
Au niveau de la digue ouest, ce sont les espèces indifférentes à l’enrichissement, toujours présentes en faibles
densités et sans variations saisonnières prononcées (GE II) Nephtys hombergii et Owenia fusiformis qui vont do-
miner numériquement la biocénose. Néanmoins, au cours de l’été 2016, les espèces tolérantes stimulées par
l’enrichissement en matière organique (GE III), Abra alba, Kurtiella bidentata et Melinna palmata ont progressé.
A l’angle de la digue Sud du port historique, en fin d’hiver, les espèces opportunistes dominent comme Cirrifor-
mia tentacula et Kirkegaardia sp. mais en fin d’été, la biocénose est différente et les espèces sensibles en parti-
culier Ericthonius punctatus et Pisidia longicornis présentent des populations très denses. Cette hausse s’est ré-
percutée sur la contribution des espèces tolérantes qui recule malgré des densités en hausse.
TABLEAU 13 : INDICES ECOLOGIQUES DES BIOCENOSES ET ETAT DE SANTE DU MILIEU AU NIVEAU DES STATIONS ECHANTIL-
LONNEES EN 2016.
Les indices de diversité en 2016 sont dans l’ensemble similaires entre les biocénoses avec un indice de
Shannon compris entre 2,76 et 4,405 et un indice de Piélou (équitabilité) entre 0,53 et 0,806 (Tableau 13). Ainsi,
d’après les seuils décrits dans la littérature, les communautés apparaissent globalement en état écologique bon
avec une légère pollution entrainant un état écologique transitoire, seules les biocénoses des avant-ports en fin
d’hiver ressortent en état écologique moyen, signe d’une pollution modérée.
Néanmoins, l’indice AMBI et l’indice IE2C en intégrant une notion de sensibilité des espèces permettent
de modérer les précédents résultats avec des indices témoignant de perturbations plus marquées. Ainsi, d’après
Fin d'hiver Fin d'été Fin d'hiver Fin d'été Fin d'hiver Fin d'été Fin d'hiver Fin d'été
Indice de diversité de Shannon (H') 2,760 3,620 2,970 3,515 4,405 4,427 3,901 3,974
Equitabilité de Piélou (J') 0,530 0,695 0,600 0,691 0,802 0,806 0,678 0,572
AMBI 1,458 2,537 1,761 2,775 1,653 2,353 3,454 2,182
Etat écologique (Ruellet et Dauvin, 2007) Moyen Moyen Moyen Moyen Moyen Moyen Médiocre Moyen
Classification de la pollution Modérement pollué Modérement pollué Modérement pollué Modérement pollué Modérement pollué Modérement pollué Gravement pollué Modérement pollué
I2EC 0-2 2 0-2 2 3 3 2 0
Etat de santé du milieu Normal à enrichi Enrichi Normal à enrichi Enrichi Enrichi NormalStade de transition avec le milieu pollué
AD_01 AD_11 AD_06 AD_12

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 49
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
les seuils décrits dans la littérature, les communautés apparaissent globalement toutes en état écologique moyen
et la biocénose de la station AD_12 ressortant en état écologique médiocre signe d’une pollution grave d’après
les seuils décrits par Ruellet et Dauvin (2007) pour les communautés estuariennes. L’indice I2EC confirme un état
écologique du milieu plus dégradé au niveau de la digue ouest en stade de transition avec le milieu pollué. Au
niveau des avant-ports, le milieu apparaît plus enrichi en conditions de fin d’été mais, cela ne traduit pas obliga-
toirement une pollution anthropique, toute source d’enrichissement en pouvant être à l’origine. Enfin, au niveau
de l’angle de la digue Sud du port historique, l’état du milieu, enrichi en fin d’hiver, s’améliore pendant l’été
comme en témoigne la hausse de la contribution des espèces sensibles.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 50
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
6. CONCLUSION
Les campagnes de fin d’hiver et de fin d’été 2016 s’inscrivaient dans un objectif de connaissance globale
des peuplements benthiques et des sédiments superficiels dans le cadre du projet de l’accès fluvial direct à Port
2000.
La composition sédimentaire des bassins portuaires comme ceux de l’étude n’a fait l’objet que de rares
études. Les fonds prospectés au niveau des avant-ports et à l’entrée de Port 2000 étaient caractérisés en 2016
par une fraction importante de silts et argiles (entre 57,9% et 79%) avec des teneurs similaires à celles des fonds
des bassins de marée du Havre (Théophile Ducrocq et Coty, études récentes menées par le GPMH) et par une
fraction importante de galets et graviers (>61%) au niveau de l’angle de la digue Sud du port historique. Ces
résultats recoupent les données historiques témoignant de la présence de vases molles ou vases compactes ré-
ductrices dans laquelle la phase sableuse à silteuse est pratiquement absente, au niveau des bassins de marée
(Breton et al., 2005 ; Breton, 2000 ; Dancie et al., 2011). Ainsi, à l’échelle du site d’étude, en 2016, les fonds au
sein des avant-ports et à l’entrée de Port2000 sont constitués d’un horizon inférieur de vases et vases sableuses
portuaires noires anoxiques, riches en matière organique, oxydée en surface et réduites suite à la colonisation
par les bactéries Beggiatoales expliquant la couche noire observée. Ces conditions diagénétiques anoxiques en-
traînent d’une part la coloration des fonds mais également une odeur prononcée de décomposition associée à
celle anthropique des hydrocarbures (Breton, 2005 ; Dancie et al., 2011 ; 2012). Cet horizon inférieur de vases
réductrices molles à compactes est recouvert d’un horizon de crème de vase et de vase sableuse. Des débris de
coquilles, essentiellement de moules et couteaux, et de murailles de Cirripèdes sont également présents au sein
des sédiments. A l’angle de la digue Sud du port historique, bien que les débris de coquilles, essentiellement de
moules et couteaux, et de murailles de Cirripèdes sont également présents, les fonds sont constitués de cailloutis
peu envasés, l’hydrodynamisme du secteur ne permettant pas la décantation et la sédimentation des particules
fines. Au cours de l’année 2016, les caractéristiques sédimentaires ont évolué différemment selon les stations,
les variations des teneurs en silts et en argiles pouvant s’expliquer d’une part, par les conditions naturelles de
débits, les crues ayant été exceptionnelles entre le 31 mai et le 23 juin (débit > 1000 m3/s) et d’autre part, par
les activités anthropiques (dragages d’entretien, carottage et déplacement de la plateforme en juillet pouvant
entrainer des dispersions de particules fines).
Or, ces caractéristiques sédimentaires, en particulier cette forte charge organique ainsi que les teneurs
importantes en particules fines, sont des facteurs de turbidité pouvant avoir comme conséquences, selon le
nombre et la nature chimique des matières en suspension, une diminution du taux de filtration des bivalves, un
accroissement du taux de mortalité des organismes ou de l’adsorption de polluants. Les données historiques
montrent que les milieux portuaires en particulier les vasières portuaires abritent des communautés dégradées
dominées par les espèces opportunistes.
Les campagnes de prélèvement se sont déroulées le 16 février 2016, en conditions post-hivernales avec
une température moyenne de l’eau de 8,1 ± 0,1 °C et une salinité moyenne de de 32,9 ± 0,2 et les 28 septembre
et 04 octobre 2016, en conditions post-estivale avec une température moyenne de l’eau de 18,4 ± 0,7 °C, l’aug-
mentation de la température de l’eau ayant permis le recrutement estival des populations (salinité moyenne de
de 31 ± 0,7).
D’un point de vue faunistique, la campagne de prospection 2016 a permis de recenser 176 taxons (92
taxons en fin d’hiver et 159 taxons en fin d’été) soit 79 taxons d’Annélides, 49 taxons d’Arthropodes, 28 taxons
de Mollusques et 4 espèces d’Echinodermes. Par rapport aux études réalisées en 2007 et 2012, les communautés
des avant-ports et à l’entrée de Port 2000 sont similaires à celles des bassins à marée déjà étudiées en termes
de richesse spécifique (maximums).

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 51
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
En 2016, 23 à 24 taxons sont constants soit plus d’un quart des espèces recensées mais parallèlement,
96 espèces n’ont été observées qu’en une seule occasion (54,5% des espèces). Sur l’ensemble des deux cam-
pagnes, 15 espèces sont constantes et 9 espèces introduites sont présentes : les Annélides Ficopotomatus
enigmaticus (1ère observation dans les bassins portuaires du Havre dans les années 1990), Hydroides ezoensis
(1ère observation dans les bassins portuaires du Havre en 1997), Streblospio benedicti, la balane, Austrominus
modestus (1ère observation dans les bassins portuaires du Havre, env. 1960) et les Mollusques Ensis directus
(1ère observation dans les bassins portuaires du Havre en 2003), Petricolaria pholadiformis (1ère observation
dans les bassins portuaires du Havre dans les années 1990) et Crepidula fornicata (1ère observation dans les
bassins portuaires du Havre en sûrement 1950-1960) et l’ascidie Styela clava (1ère observation dans les bassins
portuaires du Havre en 1968). Une autre espèce d’Annélide Melinna palmata est observée depuis 2006 dans
l’estuaire de Seine et dans le cadre de cette étude mais il s’agit dans son cas, d’une extension de répartition
géographique et non d’une introduction. Enfin, 28 espèces sont sur la liste des espèces de macrozoobenthos
subtidales déterminantes pour l’inventaire ZNIEFF mer en Haute-Normandie (validée par le CSRPN HN le 10 dé-
cembre 2013).
En termes de richesses spécifiques stationnelles, les biocénoses présentent une répartition homogène
(entre 37 et 54 taxons dans l’ensemble), seule la richesse spécifique de la station AD_12 se détachant en fin d’été
suite à une importante diversification des peuplements (124 espèces).
En ce qui concerne les densités, elles sont dominées par les peuplements d’Annélides et de Mollusques
Bivalves en fin d’hiver (90% des effectifs) et par les peuplements d’Arthropodes et d’Annélides en fin d’été (87%
des effectifs). A l’échelle du site d’étude et de l’année 2016, la communauté est dominée numériquement par
les Amphipodes, Ericthonius punctatus et Monocorophium acherusicum, les balanes, Balanus crenatus et Austro-
minius modestus, les Annélides, Oligochètes et Mediomastus fragilis ; les Mollusques, Nucula nitidosa. Mais, au
cours de l’été 2016, les biocénoses ont été remaniées avec un recul de la contribution des Annelides, Mediomas-
tus fragilis, Mollusques, Nucula nitidosa et coques au profit de celle Amphipodes, Ericthonius punctatus, balanes,
Balanus crenatus et Oligochètes.
Au cours de l’année 2016, les densités des biocénoses échantillonnées au niveau des quatre stations de
prélèvement sont comprises entre 768 et 3578 ind./m² à la fin de l’hiver 2016 et entre 802 et 29824 ind./m² à la
fin de l’été. La répartition spatiale des densités est identique au cours de l’année avec les populations les plus
denses vers le nord-est à l’angle de la digue Sud du port historique et dans l’avant-port mais dans l’ensemble, il
y a peu de différence, seule la biocénose de la station de la digue ouest étant plus pauvre (station AD_06). Enfin,
en termes de recrutement, les densités se maintiennent dans l’ensemble, seule la densité de la station AD_12 au
niveau de l’angle de la digue Sud du port historique augmente fortement (+26246 ind./m²). Cela s’explique par
l’échantillonnage de peuplements différents suite à l’envasement des fonds et l’échantillonnage d’un nombre
plus important de galets encroutés qui a entrainé une hausse importante des densités des espèces sensibles,
Amphipodes, Ericthonius punctatus et balanes, entre autres. Au niveau des autres biocénoses, les espèces domi-
nantes sont dans l’ensemble identiques entre les deux campagnes.
Enfin, les biomasses de la communauté en place proviennent d’individus à forte biomasse individuelle
à savoir des Mollusques pour l’essentiel (>75%), essentiellement les coques (entre 62% et 71%) et à moindre
mesure, des populations de crabes verts (8%) et de corbules (5%) en fin d’hiver et de des populations de balanes
(7%) et de bucardes (4%) en fin d’été. Au cours de l’année 2016, les fortes biomasses sont donc retrouvées au
niveau des populations de Mollusques les plus denses, à savoir au niveau de la biocénose de l’avant-port du
Havre (AD_01) et les minimums de biomasses au niveau de la biocénose au sud de la digue ouest (AD_06). Au
cours de l’été, les biomasses ont toutes augmenté suite à la croissance des individus (AD_01) et aux différences

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 52
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
de peuplements en place (AD_12), la biomasse totale de la biocénose de la station AD_01 étant particulièrement
notable (entre 4,06 et 57,66 g/m² en fin d’hiver et entre 15,9 et 282,7 g/m² en fin d’été).
A l’issue de ces deux campagnes 2016, les peuplements benthiques identifiés dans les avant-ports et le
long de la digue ouest appartiennent à la communauté des sables fins envasés à Abra alba et Lagis koreni dans
une variante appauvrie de cette communauté et sont rattachés aux biocénoses précédemment étudiées des
bassins de marée du Havre. La présence de Corbicula gibba caractérise un faciès très particulier de la commu-
nauté à Abra alba, espèce qu’il tend à remplacer. La première source d’enrichissement de la communauté à Abra
alba va être la matière organique issues du confinement important des particules fines au niveau de ces stations
et ce, malgré les apports du milieu marin. Ce faciès a déjà été décrit à nombreuses reprises lors des études des
sédiments portuaires français, comme lors de la bioévaluation du triple ensemble portuaire Boulogne-Calais-
Dunkerque (Grall et al., 2003), ou étrangers (Reish, 1959 ; Bellan, 1967 ; Leung Tack Kit, 1971 ; Hily, 1985 ; Reb-
zani-Zahaf, 1992 ; Pruvot, 2000), et correspond donc à la biocénose des sédiments fins envasés à vases orga-
niques des milieux portuaires confinés. Caractéristique d’une zone subnormale (en transition vers un déséqui-
libre du milieu), ce peuplement se distingue donc de l’état normal des biocénoses par la réduction sensible du
nombre d’espèces et par la prolifération d’espèces les moins sensibles ou indicatrices d’un facteur particulier.
Cet état de fait est confirmé par l’indice I2EC dont les valeurs traduisent la présence d’un milieu qualifié d’enrichi
à transitoire en fin d’été. En dehors du secteur impacté directement par le projet, les fonds de la station AD_12
constituent un habitat de sédiments grossiers infralittoraux à circalittoraux. Modérément exposé, au niveau de
fonds de sable graveleux, galets, cailloutis et gravier, il est susceptible d’être perturbé par les courants de marée
et l’action des vagues mais reste caractérisé par d’importantes densités et une forte proportion d’espèces sen-
sibles essentiellement des Arthropodes.
Ainsi, l’état des lieux a permis de mettre en évidence la présence de populations d’espèces sensibles et
d’intérêt écologique en nombre notable, en particulier au niveau des fonds de l’angle de la digue Sud du port
historique, hors du secteur impacté directement par les travaux envisagés. Au niveau des fonds de vases et vases
sableuses portuaires étudiés lors de cette étude, les communautés dominées par des espèces tolérantes et op-
portunistes se rapprochent de celles jusqu’alors décrites dans les bassins à marée du Havre. Il conviendra d’en
tenir compte lors des travaux d’aménagements prévus afin de réduire au maximum les impacts sur la faune
benthique présente déjà soumise à de fortes pressions anthropiques. Un suivi d’impact des travaux pourrait en
soi constituer une solution d’appréciation des dérangements encourus au travers d’une étude de la recolonisa-
tion des fonds.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 53
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
BIBLIOGRAPHIE
ALIZIER S., 2011. Echelles spatio-temporelles d’observation des relations macrobenthos – sédiments : organisation
et changements à long terme (1988-2009) des communautés benthiques subtidales de la partie orientale de
la baie de Seine. Thèse de doctorat d’état de l’Université de Lille. 205 p.
BAFFREAU A., DAUVIN J.-C., 2015. Proposition de nouveaux périmètres ZNIEFF-Marines et mise à jour des ZNIEFF
existantes en Baie de Seine. Rapport final. Mission d’expertise. DREAL. 74 p.
BAJJOUK T., 2009 Cahier des charges pour la cartographie d’habitats des sites Natura 2000 littoraux Guide métho-
dologique RST/IFREMER/DYNECO/AG/09-01TB
BAJJOUK T., DERRIEN-COURTEL S., GENTIL F., HILY C., GRALL J., 2011. Typologie d'habitats marins benthiques : Analyse
de l’existant et propositions pour la cartographie. Habitats côtiers de la région Bretagne - Note de synthèse
n°2 - Habitats du circalittoral. Rebent et Natura Bretagne, Ref. RST/IFREMER/DYNECO/AG/11-03/TB, 39p.
http://archimer.ifremer.fr/doc/00063/17416/
BELLAN G., 1967. Pollution et peuplements benthiques sur substrats meubles dans la région de Marseille. Deu-
xième partie : l’ensemble portuaire marseillais. Revue int. Oceanogr. Med. 5 : 51-95.
BENSETTITI F., BIORET F., ROLAND G., LACOSTE J-PH, GEHU J-M., GLEMAREC M. ET BELLAN-SANTINI D., 2004. Connaissance et
gestion des habitats et des espèces d'intérêt communautaire. Tome 2 : Habitats côtiers - Cahiers d'habitats
Natura 2000. Ed. Paris la documentation Française. 399 p,
BLAIR T.C., MCPHERSON J.G., 1999. Grain-size and textural classification of coarse sedimentary particles. J. Sedim.
Res., 69 , 6 19.
BLANCHET, H., LAVESQUE, N., RUELLET, T., DAUVIN, J.C., SAURIAU, P.G., DESROY, N., DESCLAUX, C., LECONTE, M., BACHELET, G.,
JANSON, A.L., BESSINETON, C., DUHAMEL, S., JOURDE, J., MAYOT, S., SIMON, S., DE MONTAUDOUIN, X., 2008. Use of biotic
indices in semi-enclosed coastal ecosystems and transitional waters habitats - Implications for the implemen-
tation of the European Water Framework Directive. Ecological Indicators, 8, 360-372.
BORJA, A., FRANCO, J. AND PEREZ, V., 2000. A marine biotic index to establish the ecological quality of soft
bottom benthos within european estuarine and coastal environments. Marine Pollution Bulletin, 40 (12):
1100-1114.
BORJÁ, A., MUXIKA, I., 2005. Guidelines for the use of AMBI (AZTI’s marine biotic index) in the assessment of the
benthic ecological quality. Marine Pollution Bulletin, 50, 787–789.
BOYD S.E. et al., 2002. Guidelines for the conduct of benthic studies at aggregate dredging sites. DTLR/CEFAS, 117
pp.
BRETON G., 2000. Etude du benthos de sept sites du port du Havre (Seine-Maritime) : quai Joannès-Couvert-Est,
quai Mazeline, Môle Central et quai de l’Atlantique. Juin à Septembre 2000. Rapport Mu-séum d’Histoire
Naturelle du Havre / EDF – Service environnement ECOTOX. 5pp.
BRETON G., 2005. Le port du Havre (Manche orientale, France) et ses peuplements : un exemple de domaine pa-
ralique en climat tempéré. Bull. Soc. zool. Fr. 130 (4) : 381-423.
CONNOR D.W., ALLEN J.H., GOLDING N., HOWELL K., LIEBERKNECHT L.M., NORTHEN K.O. & REKER J.B., 2004. The marine
habitat classification for Britain and Ireland, Version 04.05 JNCC, Peterborough, ISBN 1 861 07561 8 (Internet
version http://www.jncc.gov.uk/page-1584)
CONSLEG 2004. Directive 92/43/CEE DU CONSEIL du 21 mai 1992 concernant la conservation des habitats naturels
ainsi que de la faune et de la flore sauvages. Réf. 1992L0043 - FR - 01.05.2004. 57p.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 54
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Commission Européenne, 1999. Manuel d'interprétation des habitats de l'Union Européenne. EUR 15/2. 132 p.
DAJOZ R., 1985. Dictionnaire d'entomologie : anatomie, systématique, biologie. Paris, Dunod, 549 p.
DANCIE C., 2015. Suivi des sédiments et des peuplements benthiques du site d’immersion d’Octeville, de l’estuaire
de la Seine et de la Réserve Naturelle de l’Estuaire de la Seine. Rapport Scientifique des trois sites - Automne
2014. Rapport CSLN / GPMH. pp. 65.
DANCIE C., ALLIGNY C., CHAIGNON C., DUBUT S., HANIN C., LANSHERE J., LE THOER D., 2010a. Étude biosédimentaire du port
d'Antifer. Campagne Printemps 2010. Rapport de synthèse. Rapport CSLN / SAFEGE / Gaz de Normandie. 53
p. + annexes.
DANCIE C., CHAIGNON C., CRAMET F., MANGANE A., 2011a. Etude biosédimentaire de l’ancien bassin aux pétroles –
Etat initial Hiver 2011. Rapport final de la campagne d’hiver 2011. Rapport CSLN / GPMH. 25 p. + annexes.
DANCIE C., CHAIGNON C., CRAMET F., MANGANE A., 2011b. Etude biosédimentaire du bassin René Coty, Quai de liaison
Atlantique/Amériques – Etat initial Hiver 2011. Rapport final de la campagne d’hiver 2011. Rapport CSLN /
GPMH. 26 p. + annexes.
DANCIE C., CHAIGNON C., DUBUT S., GAVALDA M., LE THOER D., 2012. Etude de l’endobenthos des bassins de marée du
port du Havre. Rapport de la campagne de printemps 2012. Rapport CSLN / Port Vivant / ECOTOX. 29 p. +
annexes.
DANCIE C., CHAIGNON C., DUBUT S., GESLAND R., HANIN C., LANSHERE J., LE THOER D., 2010b. Étude biosédimentaire du
port d'Antifer. Campagne Automne 2009. Rapport de synthèse. Rapport CSLN / SAFEGE / Gaz de Normandie.
43 p. + annexes.
DANCIE C., CHAIGNON C., DUBUT S., GESLAND R., HANIN C., LANSHERE J., LE THOER D., 2010c. Étude biosédimentaire du
port d'Antifer. Campagne Automne 2009. Rapport de synthèse. Rapport CSLN / Gaz de Normandie. 43 p. +
annexes.
DANCIE C., CHAIGNON C., DUBUT S., LE THOER D., MANGANE A., 2015. Etude de l’endofaune de trois stations de prélè-
vement dans une darse du port de Dunkerque - Année 2014. Rapport CSLN / ECOTOX. 32 p. + annexes.
DANCIE C., SIMON S., BERNO A., ECOLIVET E., HANIN C., JOURDE J., LEFRANCOIS T., FRIBOULET R., 2007a. Suivi des sédiments
et des peuplements du Grand Canal du Havre. Rapport de la campagne de mars 2007. Rapport CSLN / Port
Autonome du Havre. 17p. et annexes.
DANCIE C., SIMON S., BERNO A., ECOLIVET E., HANIN C., JOURDE J., LEFRANCOIS T., 2007b. Suivi des sédiments et des peu-
plements benthiques du canal de Tancarville. Rapport final des campagnes de novembre 2006 et mars 2007.
Rapport CSLN / Port Autonome du Havre. 19p. et annexes.
DAVIES C.E., MOSS D., HILL M.O., 2004. EUNIS habitat classification revised 2004. European Topic Centre on Nature
Protection and Biodiversity, European Environment Agency. 307 P.
DAVIES, C.E. & MOSS, D. 2004. EUNIS Habitat Classification. Marine Habitat Types: Proposals for Revised
Criteria, July 2004. Report to the European Topic Centre on Nature Protection and Biodiversity, European
Environment Agency. July 2004.
DAUVIN, J.C., 1993. Le benthos : témoin des variations de l'environnement. Oceanis, 19, 25-53.
DAUVIN, J.C., RUELLET, T., 2007. Polychaete/amphipod ratio revisited. Marine Pollution Bulletin, 55, 215–224.
DAUVIN J.C., VASSET N., PARAIS F., CHAUVIN C., ALIZIER S., BRUNAUD C., DELHAY J.-B., 2013. Stratégie d’observation à long
terme. Guide pour l’étude du macrobenthos de l’estuaire de la Seine. Rapport GIP Seine-Aval. 31 p.
DEWARUMEZ J.M., GHILLEBAERT F., 2011. Suivi de la Biodiversité du Bassin Maritime au niveau du site de la Centrale
de DK6. Campagne d’échantillonnage 2009. Rapport ECOTOX/Université de Lille. 33 p.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 55
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
DIAZ R.J, SOLAN M., VALENTE R.M., 2004. A review of approaches for classifying benthic habitats and evaluating
habitat quality. Journal of Environmental Management. 73, 765-181.
DUFRÊNE M, LEGENDRE P., 1997. Species assemblages and indicator species : the need for a flexible asymmetrical
approach. Ecol. Monogr. 67, 345-366.
European Commission, 2007. Interpretation manual of European Union Habitats. EUR 27. 144p.
European Commission, 2007. Guidelines for the establishment of the Natura 2000 network in the marine envi-
ronment. Application of the Habitats and Birds Directives. 112 p.
European Environement Agency, 2004. European Nature Information System (EUNIS), Version 2004.
http://unis.eea.europa.eu/habitats.jsp.
FOLK R.L., 1959, Practical petrographic classification of limestones: American Association of Petroleum Geologists
Bulletin, v. 43, p. 1-38.
FOLK R.L., 1965, Petrology of Sedimentary Rocks, Hemphill Publishing Company, Austin, Texas. 159 p.
FRONTIER S., 1976. Utilisation des diagrammes rang-fréquences dans l’analyse des écosystèmes. Journal de la Re-
cherche Océanographique, 1 : 35-48.
GHILLEBAERT F., 2007. Suivi de la Biodiversité du Bassin Maritime au niveau du site de la Centrale de DK6. Rapport
ECOTOX. 35 p.
GEODE., 2012. Guide sur le suivi environnemental des opérations de dragage et d'immersion. 133 p.
GUILLAUMONT B., BAJJOUK T., ROLLET C., HILY C., GENTIL F., 2009. Typologie d’habitats marins benthiques : analyse
de l’existant et propositions pour la cartographie (Habitats côtiers de la région Bretagne) - Note de syn-
thèse, Projets REBENT-Bretagne et Natura-Bretagne. RST/IFREMER/DYNECO/AG/08-06/BG; 22 pp. +Tab-
leaux.
GRALL J., COIC N., 2005. Synthèse des méthodes d’évaluation de la qualité du benthos en milieu côtier. Dyneco/Vi-
gies/Rebent. (http://www.rebent.org).
GRALL J., GLEMAREC M., 1997. Using biotic indices to estimate macrobenthic community perturbation in the Bay of
Brest. Estuarine, Costal and Shelf Science. 44 (suppl. A) : 43-53.
GRALL J., QUINIOU F., GLÉMAREC M., 2003. Bioévaluation de la qualité environnementale des milieux portuaires :
programme PNETOX. In ALZIEU (C.)(coord.) : Bioévaluation de la qualité environnementale des sédiments
portuaires et des zones d’immersion. IFREMER éd., p. 89-117.
GRALL J., GLEMAREC M., 2003. L’indice d’évaluation de l’endofaune côtière I2EC, Bioévaluation de la qualité envi-
ronnementale des sédiments portuaires et des zones d’immersion. ALZIEU C. (coord.) 51-86 .Editions IFRE-
MER.
HERMAND R., 2008. Réponse d'une communauté macrobenthique à des apports sédimentaires allochtones,
naturels ou anthropiques. Thèse de Doctorat, Université Aix-Marseille II. 238pp.
HILY C., 1984. Variabilité de la macrofaune benthique dans les milieux hypertrophique de la rade de Brest. Thèse
Doct. Etat, Univ. Bretagne Occident. (UBO), Brest (France), 696 p.
Hrs-Brenko M., 2006. The basket shell, Corbula gibba Olivi, 1792 (Bivalve Mollusks) as a species resistant to en-
vironmental disturbances: Acta Adriat., 47 (Suppl 1): 49-64.
ICES Marine Habitat Committee, 2001. Report of the Working Group on Marine Habitat Mapping (WGMHM), 3-
6 April 2001, Galway, Ireland, ICES CM 2001/E:07, 47p.
ICES Marine Habitat Committee, 2006. Report of the Working Group on Marine Habitat Mapping (WGMHM), 4–
7 April, 2006, Galway, Ireland, ICES CM 2006/MHC:05, Ref. FTC, ACE 136p.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 56
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
ICES Marine Habitat Committee, 2009. Report of the Working Group on Marine Habitat Mapping (WGMHM), 31-
24 April 2009, The National Institute of Aquatic Resources, Charlottenlund Castle, Copenhagen, Demark. ICES
CM 2009/MHC: 07. 78 pp.
LAVRABRE J., FISSON C., 2013. Les habitats naturels de l’estuaire de la Seine. Typologie et fonctions écologiques
associées. GIP Seine-aval. 74 p.
LEGENDRE P., LEGENDRE L., 1998. Numerical ecology. 2nd English edition. Elsevier, Amsterdam.
LESOURD S., 2000. Processus d’envasement d’un estuaire macrotidal : zoom temporel du siècle à l’heure ; appli-
cation à l’estuaire de la Seine. Thèse de doctorat, Université de Caen. 280p.
LEUNG T. K., D. 1971. Étude du milieu pollué: le Vieux-Port de Marseille. Influence des conditions physico-chi-
miques sur la physionomie du peuplement de quai. Thèse de 3e cycle : Océanographie biologique : Université
Aix-Marseille 2. 767 p.
MARTÍNEZ-CREGO B., ALCOVERRO T. & ROMERO J., 2010. Biotic indices for coastal waters quality assessment: a review
of strengths and weaknesses. Journal of Environmental Pollution. 12: 1013-1028.
MADDEN GOODIN C.K., ALLEE B., FINKBEINER M. ET BAMFORD D., 2008. Coastal and marine ecological classification stand-
ard. Noaa & N. Serve), p. 77.
MICHEZ N., BAJJOUK T., AISH A., ANDERSEN A.C., AR GALL E., BAFFREAU A., BLANCHET H., CHAUVET P., DAUVIN J.-C., DE CASA-
MAJOR M.-N., DERRIEN-COURTEL S., DUBOIS S., FABRI M.-C., HOUBIN C., LEGALL L., MENOT L., ROLET C., SAURIAU P.-G.,
THIEBAUT E., TOUROLLE J., VAN DER BELD I., 2015. Typologie des habitats marins benthiques de la Manche, de la
Mer du Nord et de l’Atlantique Version 2. Rapport SPN 2015 – 45, MNHN, Paris, 61 p.
NEWELL R.C., SEIDERER L.J. AND HITCHCOCK D.R., 1998. The impact of dredging works in coastal waters: Areview
of the sensitivity to disturbance and subsequent recovery of biological resources on the sea bed. Oceanogra-
phy and Marine Biology: An Annual Review36, 127-178.
NEWELL R.C., ROBINSON J.E., PEARCE BN, SEIDERER L.J., WARWICK R.M., SOMERFIELD P.M., CLARKE K.R., JENKINS H., BEER N.,
BURLINSON F. & STACEY K., 2007. A predictive framework for the assessment of recoverability of marine benthic
munities following cessation of aggregate dredging. In: Newell R.C. & Garner D.J. (Ed.). Marine aggregate
extraction: Helping to determine good practice. Marine ALSF Conference Proceedings, 158-163.com.
Norme ISO/FDIS 16665 : « 2005 Qualité de l’eau – Lignes directrices pour l’échantillonnage quantitatif et le trai-
tement d’échantillons de la macrofaune marine des fonds meubles ».
OSPAR list of threatened and/or declining species and habitats. 2008. Référence number OSPAR 2008-07.
PEDEL L, FABRI M.C., 2012. Etat de l'art sur les indices existant concernant l'Etat Ecologique des habitats
benthiques du domaine profond. DCSMM Bon Etat Ecologique (Ministère de l'Écologie, du
Développement Durable, des Transports et du Logement), Ifremer. p:71
PINTO, R., PATRICIO, J., BAETA, A., FATH, B.D., NETO, J.M., MARQUES, J.C., 2009. Review and evaluation of estuarine
biotic indices to assess benthic condition. Ecological Indicators, 9, 1–25.
POISSON E., CHAIGNON C., LE THOER D., 2013. Surveillance de l’impact du rejet des eaux de l’entreprise SEDIBEX sur
l’environnement dans le Grand Canal du Havre. Rapport de la campagne de janvier 2013. Rapport CSLN /
SEDIBEX. 23p + annexes.
PRUVOT C., 1999. Contribution à l'évaluation de l'impact des dragages et des immersions sur les communautés
macrozoobenthiques du port de Dunkerque et de ses zones de vidage. These de doctorat Environnement et
Ecosystèmes marins et continentaux, Université de Lille. 315 p.
PRUVOT C., EMPIS A., DHAINAUT-COURTOIS N., 2000. Présence du mollusque bivalve Corbula gibba (Olivi, 1792) dans
les sédiments meubles du port est de Dunkerque (mer du Nord). Bull. Soc., Zool. Fr., 125, 75-82.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 57
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
RAMEAU J.-C., BISSARDON M., GUIBAL L., 1997. CORINE Biotopes, types d’habitats français. ENGREF.
RAUJOUAN P. CETMEF., 2012. Suivi environnemental des aménagements portuaires et littoraux - Guide de recom-
mandations. 150 p.
REBZANI-ZAHAF C., 1992. Le peuplement macrobenthique du port d'Alger: impact de la pollution. Hydroécol. Appl.
4, 2, 91-103.
REISH D.J., 1959. A new species of Sabellidae (Annelida, Polychaeta) from southern California. Annals and Maga-
zine of Natural History, (Series 13) 2(24): 717-719
RIVIERE A., 1977. Méthodes granulométriques, techniques et interprétation, Paris, Masson, 170 p.
RUELLET, T. & DAUVIN, J.-C., 2007a. Base de Données MABES – Macrobenthos de la Baie et de l’Estuaire de la Seine–
version 2. GIP Seine-Aval, Rouen.
RUELLET, T., DAUVIN, J.C., 2007b. Benthic Indicators: analysis of the threshold values of Ecological Quality classifi-
cations for transitional waters. Mar. Pol. Bull. 54, 1707-1714. Scherrer, B., 1984. Biostatistique. Gaëtan Morin,
Paris.
SHANNON C. E., WEAVER W., 1949. The mathematical theory of communication, Urbana, University of Illinois Press.
SIMBOURA, N., ZENETOS, A., 2002. Benthic indicators to use in ecological quality classification of Mediterranean soft
bottom marine ecosystems, including a new biotic index. Mediterranean Marine Science, 3/2, 77-111.
VINCENT C., HEINRICH H., EDWARDS A., NYGAARD K., HAYTHORNTHWAITE J., 2002. Guidance on typology, reference condi-
tions and classification systems for transitional and coastal waters. Commission Européenne, CIS WG 2.4
(COAST). 119 pp.
WARE, S., BOLAM S.G., RESS H.L., 2010. Impact and recovery associated with the deposition of capital dredgings at
UK disposal sites: Lessons for future licensing and monitoring. Marine Pollution Bulletin, 60, 79-90.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 58
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
LISTE DES TABLEAUX ET FIGURES
Figure 1 : Habitats génériques et élémentaires du réseau Natura 2000 du site d’étude. ...................................... 4
Figure 2 : Communautés benthiques selon la classification EUNIS du site d’étude. .............................................. 5
Figure 3 : Zones Naturelles d’Intérêt Ecologique Faunistique et Floristique du site d’étude. ................................ 6
Figure 4 : Qualité des communautés benthiques de la partie aval de l’estuaire et de la partie orientale de la baie
de Seine estimée par les indices BOPA, ITI, BQI et synthèse par point donnant un état moyen des communautés
(source : GIP Seine-Aval 2010, Dauvin et al. 2009). ................................................................................................ 7
Figure 5 : Etat des peuplements benthiques selon l’AMBI pour la période 1979-2006 (source : GIP Seine-Aval
2008, Ruellet et Dauvin, 2007). ............................................................................................................................... 7
Figure 6 : Taux de recolonisation probables pour la communauté benthique des vases estuariennes, des sables,
des graviers et les zones de récifs (Newell et al., 1998).......................................................................................... 8
Figure 7 : Succession écologique des communautés benthiques à travers un gradient de perturbation de
l'environnement (Newell et al., 1998). ................................................................................................................... 9
Figure 8 : Hauteur d’eau (m) - Balise A pendant les périodes de prélèvements (données SHOM). ..................... 10
Figure 9 : Localisation des stations (coordonnées théoriques et réelles - Campagnes 2016). ............................. 11
Figure 10 : Etapes des prélèvements. ................................................................................................................... 13
Figure 11 : Etapes de laboratoire et d’analyses. ................................................................................................... 22
Figure 12 : Teneurs des classes granulométriques des stations échantillonnées en 2016. .................................. 23
Figure 13 : Répartition spatiale des faciès sédimentaires (cl. de Larsonneur modifiée ; Lesourd, 2000) aux stations
échantillonnées en 2016. ...................................................................................................................................... 24
Figure 14 : Box-plot pour les teneurs des classes granulométriques par type de bassin portuaire, pour les fonds
de l’embouchure de l’estuaire et des zones d’influence du site de dépôt d’Octeville et pour les stations de cette
étude. .................................................................................................................................................................... 25
Figure 15 : Classification ascendante hiérarchique (données des fractions granulométriques récoltées entre 2007
et 2016 au niveau des bassins portuaires transformées log(x+1) ; distance euclidienne, regroupement
agglomératif complet) et MDS et ACP associées (couleur des assemblages de la Erreur ! Source du renvoi
introuvable. ; GCH Grand Canal du Havre, TAN : canal de Tancarville). ............................................................... 27
Figure 16 : Contribution relative (%) des faciès sédimentaires au sein des assemblages sédimentaires mis en
évidence. ............................................................................................................................................................... 28
Figure 17 : Contribution relative (%) de la richesse spécifique des différentes classes d’espèces au sein des
biocénoses des stations échantillonnées en 2016. ............................................................................................... 29
Figure 18 : Contribution relative (%) des richesses spécifiques des phyla et richesse spécifique totale au sein des
biocénoses des stations échantillonnées en 2016. ............................................................................................... 30
Figure 19 : Répartition spatiale des richesses spécifiques des biocénoses aux stations échantillonnées en 2016.
.............................................................................................................................................................................. 32
Figure 20 : Contribution relative (%) de la densité des différentes classes d’espèces au sein des biocénoses des
stations échantillonnées en 2016. ........................................................................................................................ 33

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 59
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Figure 21 : Contribution relative (%) des densités des phyla et densité totale au sein des biocénoses des stations
échantillonnées en 2016. ...................................................................................................................................... 34
Figure 22 : Courbes d’abondance cumulative classée (courbe de dominance) des biocénoses aux stations
échantillonnées en 2016. ...................................................................................................................................... 34
Figure 23 : Répartition spatiale des densités totales des biocénoses aux stations échantillonnées en 2016. ..... 35
Figure 24 : Contribution relative (%) de la biomassé des différentes classes d’espèces au sein des biocénoses des
stations échantillonnées en 2016. ........................................................................................................................ 36
Figure 25 : Contribution relative (%) des biomasses des phyla et biomasse totale au sein des biocénoses des
stations échantillonnées en 2016. ........................................................................................................................ 37
Figure 26 : Courbes de biomasse cumulative classée (courbe de dominance) des biocénoses aux stations
échantillonnées en 2016. ...................................................................................................................................... 37
Figure 27 : Répartition spatiale des biomasses totales des biocénoses aux stations échantillonnées en 2016. .. 38
Figure 28 : Box-plot pour les paramètres biologiques pour les fonds de l’embouchure de l’estuaire, des hauts de
la Rade et des zones d’influence du site de dépôt d’Octeville et pour les stations de cette étude au printemps et
automne 2016. ...................................................................................................................................................... 39
Figure 29 : Box-plot pour les paramètres biologiques par type de bassin portuaire, pour les fonds de
l’embouchure de l’estuaire, des hauts de la Rade et des zones d’influence du site de dépôt d’Octeville et pour les
stations de cette étude. ........................................................................................................................................ 40
Figure 30 : Variations des teneurs des classes granulométriques des stations échantillonnées en 2016. ........... 42
Figure 31 : Variations des paramètres biologiques des biocénoses des stations échantillonnées en 2016. ........ 42
Figure 32 : Classification ascendante hiérarchique (données des fractions faunistiques transformées fourth root ;
distance de Bray-curtis, regroupement agglomératif moyen) et MDS associées. ................................................ 44
Figure 33 : Classification ascendante hiérarchique (données des fractions faunistiques récoltées entre 2007 et
2016 au niveau des bassins portuaires transformées fourth root ; distance de Bray-curtis, regroupement
agglomératif moyen) et MDS et ACP associées (GCH Grand Canal du Havre, TAN : canal de Tancarville). ......... 47
Figure 34 : Contribution relative (%) des densités des groupes écologiques et valeur de l’indice AMBI au sein des
biocénoses des stations échantillonnées en 2016. ............................................................................................... 48
Tableau 1 : Conditions météorologiques - France - La Heve (Le Havre). .............................................................. 10
Tableau 2 : Conditions marégraphiques. .............................................................................................................. 10
Tableau 3 : Dates et mesures hydrologiques en fond de colonne d'eau – Campagnes 2016. .............................. 12
Tableau 4: Classes granulométriques. .................................................................................................................. 14
Tableau 5 : Classification de Larsonneur modifiée (Lesourd, 2000) (chiffres exprimés en % de la fraction totale -
G : galets > 2cm - Gr : graviers 2 cm à 2 mm - Sg : sables grossiers 500 μm à 2 mm - Sm : sables moyens 200 à 500
μm- Sf sables fins 50 à 200 μm - V : vases<50μm). ............................................................................................... 15
Tableau 6 : Signification de l'AMBI pour la communauté à Abra alba – Lagis koreni (Borja et al., 2000). ........... 19
Tableau 7 : Signification de l'AMBI pour les communautés estuariennes (Ruellet et Dauvin, 2007). .................. 19
Tableau 8 : Estimation de l’abondance et de la richesse spécifique d’états de référence de différents habitats
(d’après Grall et Glémarec, 2003). ........................................................................................................................ 20

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 60
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Tableau 9 : Valeurs seuils des différents groupes écologiques pour l’attribution de l’I2EC (d’après Grall et
Glémarec, 2003). ................................................................................................................................................... 20
Tableau 10 : Caractéristiques pour l’attribution de l’I2EC en présence d’écotones. ............................................ 20
Tableau 11 : Valeurs seuils des différents indices utilisés (flèche, sens de variation de l’indice ; Vincent et al. (2002)
pour H’, Grall et Glémarec (2003) pour I2EC, Borja et al., 2000 et Ruellet et Vincent, 2007 pour AMBI). ........... 21
Tableau 12 : Caractérisation des peuplements benthiques subtidaux de la zone d’étude (S : richesse spécifique,
D : Densité (ind./m²), B : Biomasse (g/m²)). .......................................................................................................... 41
Tableau 13 : Indices écologiques des biocénoses et Etat de santé du milieu au niveau des stations échantillonnées
en 2016. ................................................................................................................................................................ 48

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 61
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
ANNEXES
ANNEXE 1 : ANALYSES GRANULOMETRIQUES
ANNEXE 2 : LISTE FAUNISTIQUE
ANNEXE 3 : RESULTATS FAUNISTIQUES PAR STATION
ANNEXE 4 : FICHES DESCRIPTIVES DES ESPECES DOMINANTES
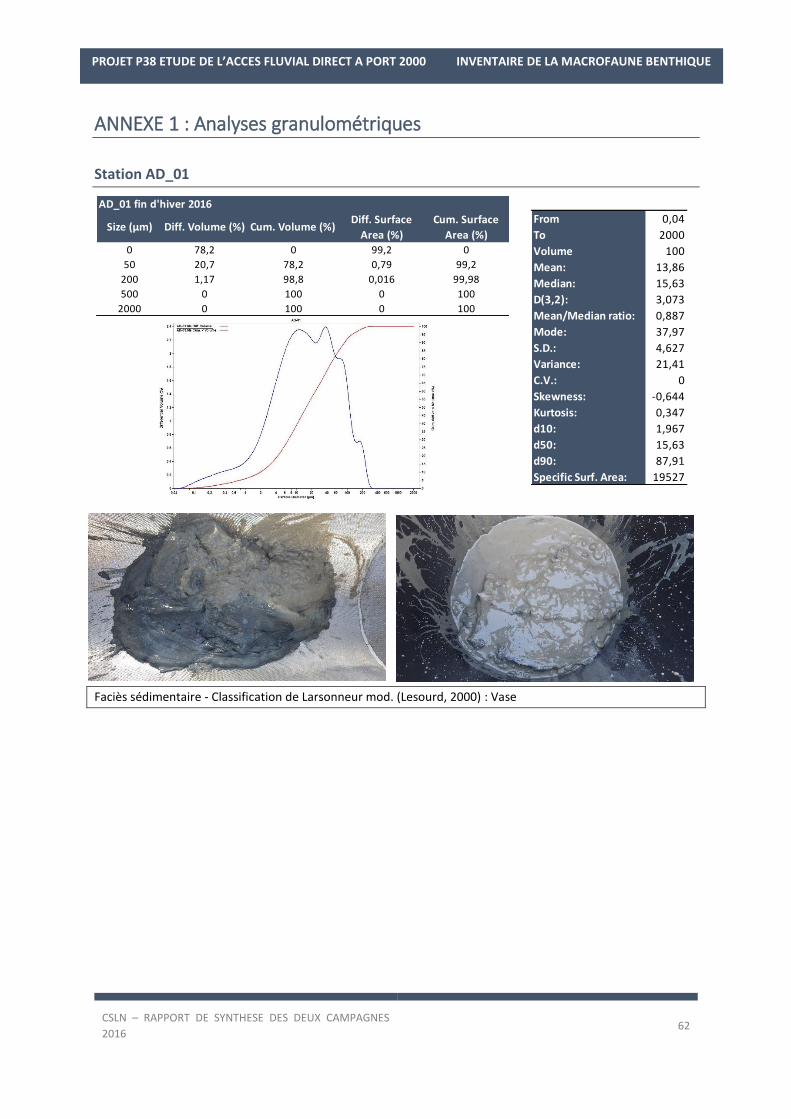
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 62
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
ANNEXE 1 : Analyses granulométriques
Station AD_01
Faciès sédimentaire - Classification de Larsonneur mod. (Lesourd, 2000) : Vase
AD_01 fin d'hiver 2016
Size (µm) Diff. Volume (%) Cum. Volume (%)Diff. Surface
Area (%)
Cum. Surface
Area (%)
0 78,2 0 99,2 0
50 20,7 78,2 0,79 99,2
200 1,17 98,8 0,016 99,98
500 0 100 0 100
2000 0 100 0 100
From 0,04
To 2000
Volume 100
Mean: 13,86
Median: 15,63
D(3,2): 3,073
Mean/Median ratio: 0,887
Mode: 37,97
S.D.: 4,627
Variance: 21,41
C.V.: 0
Skewness: -0,644
Kurtosis: 0,347
d10: 1,967
d50: 15,63
d90: 87,91
Specific Surf. Area: 19527
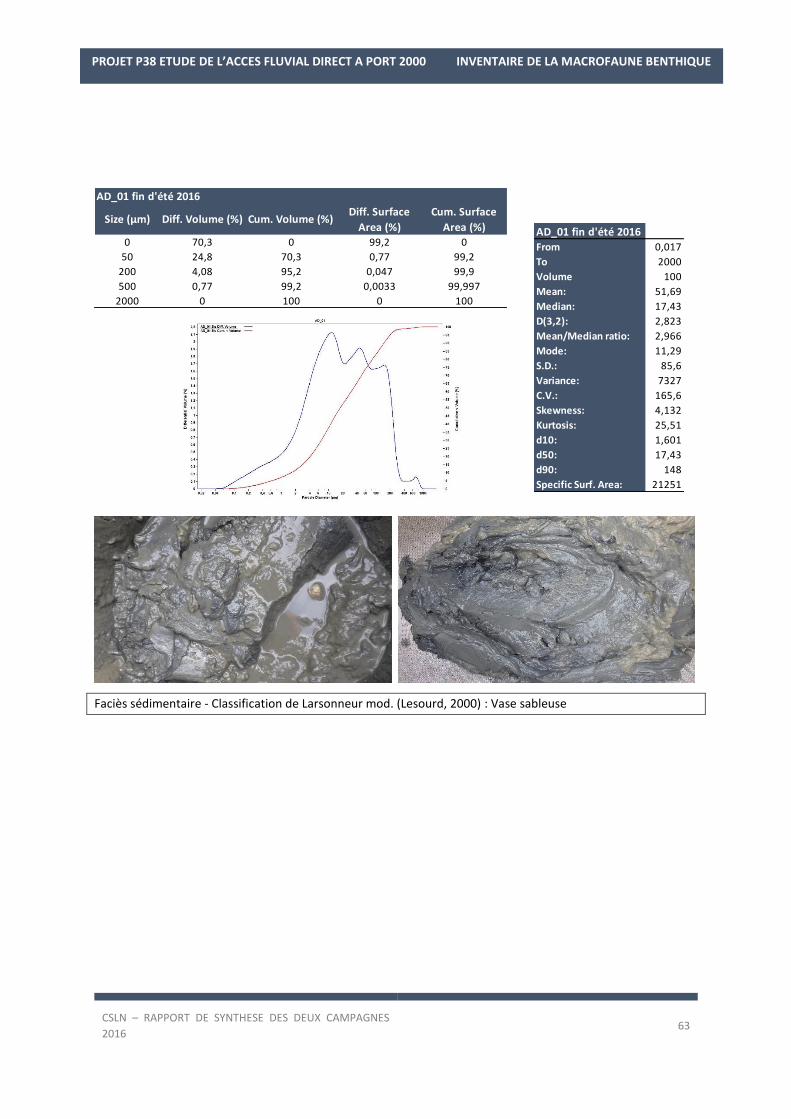
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 63
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Faciès sédimentaire - Classification de Larsonneur mod. (Lesourd, 2000) : Vase sableuse
AD_01 fin d'été 2016
Size (µm) Diff. Volume (%) Cum. Volume (%)Diff. Surface
Area (%)
Cum. Surface
Area (%)
0 70,3 0 99,2 0
50 24,8 70,3 0,77 99,2
200 4,08 95,2 0,047 99,9
500 0,77 99,2 0,0033 99,997
2000 0 100 0 100
AD_01 fin d'été 2016
From 0,017
To 2000
Volume 100
Mean: 51,69
Median: 17,43
D(3,2): 2,823
Mean/Median ratio: 2,966
Mode: 11,29
S.D.: 85,6
Variance: 7327
C.V.: 165,6
Skewness: 4,132
Kurtosis: 25,51
d10: 1,601
d50: 17,43
d90: 148
Specific Surf. Area: 21251
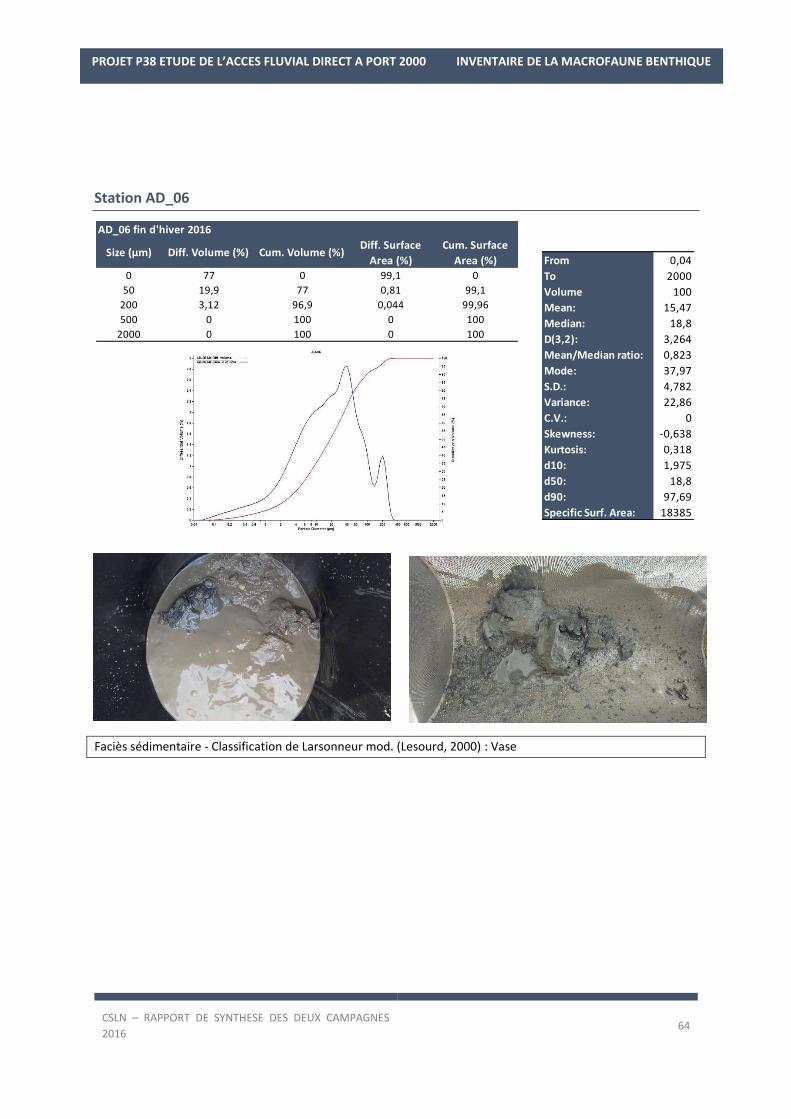
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 64
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Station AD_06
Faciès sédimentaire - Classification de Larsonneur mod. (Lesourd, 2000) : Vase
AD_06 fin d'hiver 2016
Size (µm) Diff. Volume (%) Cum. Volume (%)Diff. Surface
Area (%)
Cum. Surface
Area (%)
0 77 0 99,1 0
50 19,9 77 0,81 99,1
200 3,12 96,9 0,044 99,96
500 0 100 0 100
2000 0 100 0 100
From 0,04
To 2000
Volume 100
Mean: 15,47
Median: 18,8
D(3,2): 3,264
Mean/Median ratio: 0,823
Mode: 37,97
S.D.: 4,782
Variance: 22,86
C.V.: 0
Skewness: -0,638
Kurtosis: 0,318
d10: 1,975
d50: 18,8
d90: 97,69
Specific Surf. Area: 18385
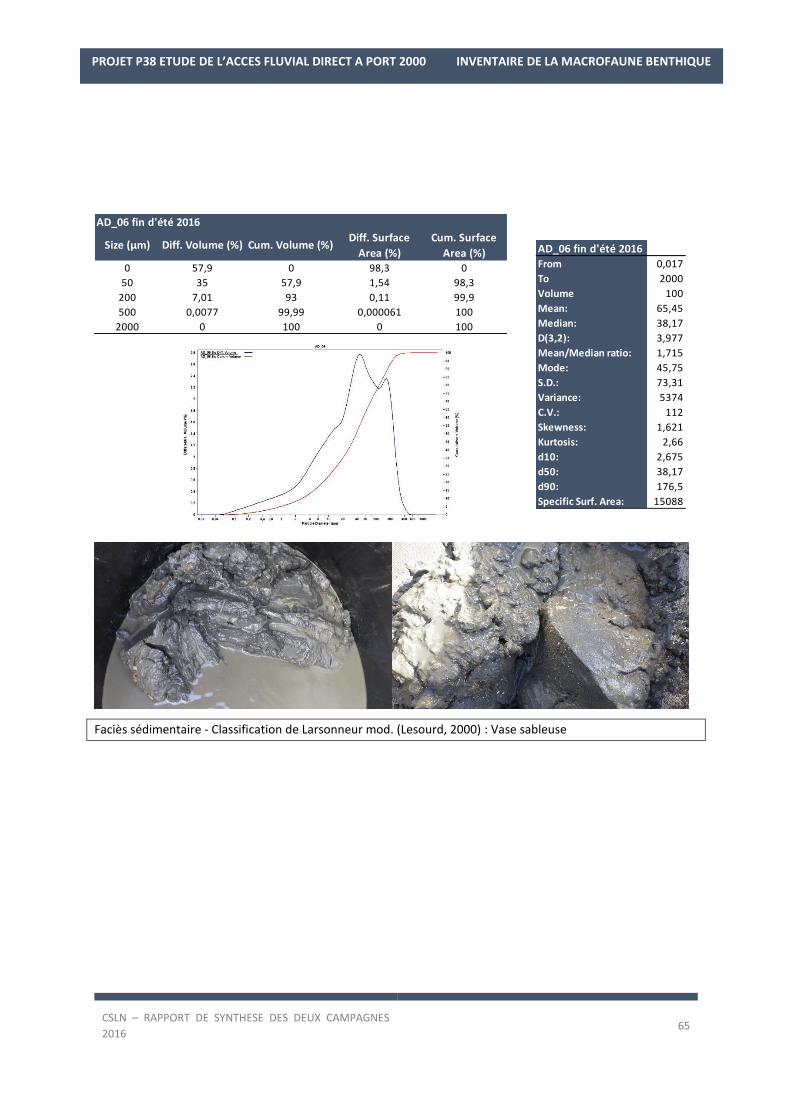
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 65
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Faciès sédimentaire - Classification de Larsonneur mod. (Lesourd, 2000) : Vase sableuse
AD_06 fin d'été 2016
Size (µm) Diff. Volume (%) Cum. Volume (%)Diff. Surface
Area (%)
Cum. Surface
Area (%)
0 57,9 0 98,3 0
50 35 57,9 1,54 98,3
200 7,01 93 0,11 99,9
500 0,0077 99,99 0,000061 100
2000 0 100 0 100
AD_06 fin d'été 2016
From 0,017
To 2000
Volume 100
Mean: 65,45
Median: 38,17
D(3,2): 3,977
Mean/Median ratio: 1,715
Mode: 45,75
S.D.: 73,31
Variance: 5374
C.V.: 112
Skewness: 1,621
Kurtosis: 2,66
d10: 2,675
d50: 38,17
d90: 176,5
Specific Surf. Area: 15088
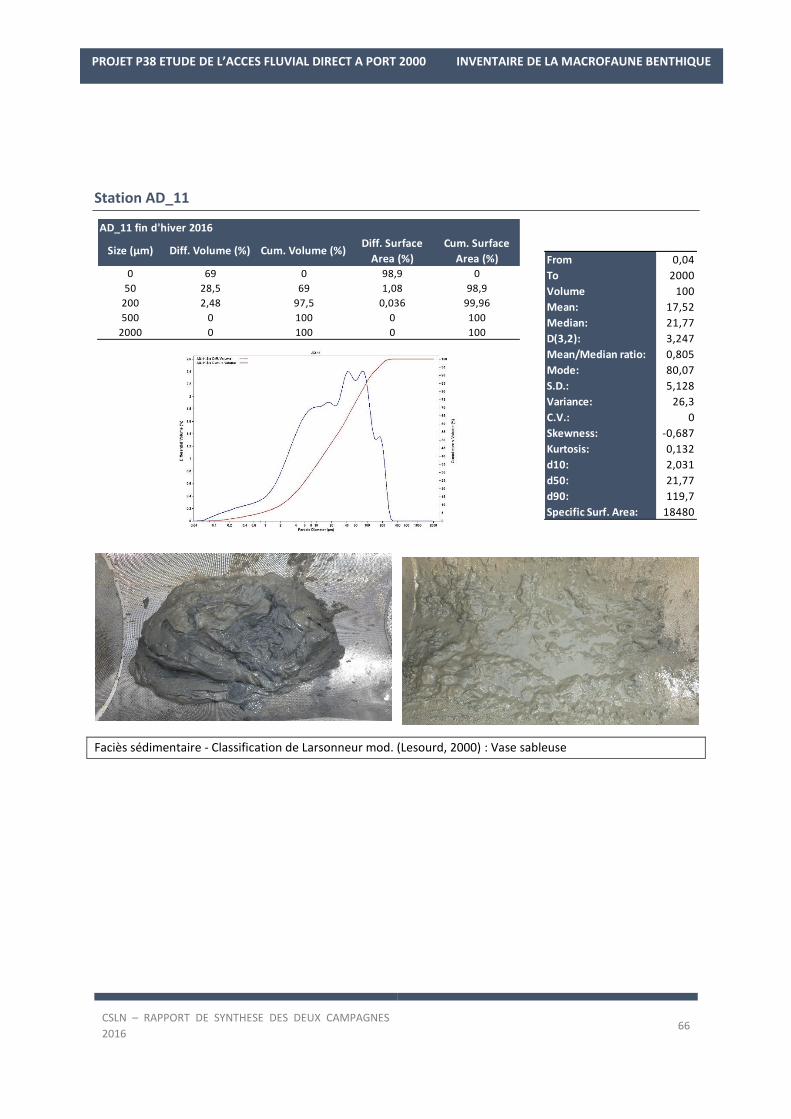
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 66
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Station AD_11
Faciès sédimentaire - Classification de Larsonneur mod. (Lesourd, 2000) : Vase sableuse
AD_11 fin d'hiver 2016
Size (µm) Diff. Volume (%) Cum. Volume (%)Diff. Surface
Area (%)
Cum. Surface
Area (%)
0 69 0 98,9 0
50 28,5 69 1,08 98,9
200 2,48 97,5 0,036 99,96
500 0 100 0 100
2000 0 100 0 100
From 0,04
To 2000
Volume 100
Mean: 17,52
Median: 21,77
D(3,2): 3,247
Mean/Median ratio: 0,805
Mode: 80,07
S.D.: 5,128
Variance: 26,3
C.V.: 0
Skewness: -0,687
Kurtosis: 0,132
d10: 2,031
d50: 21,77
d90: 119,7
Specific Surf. Area: 18480
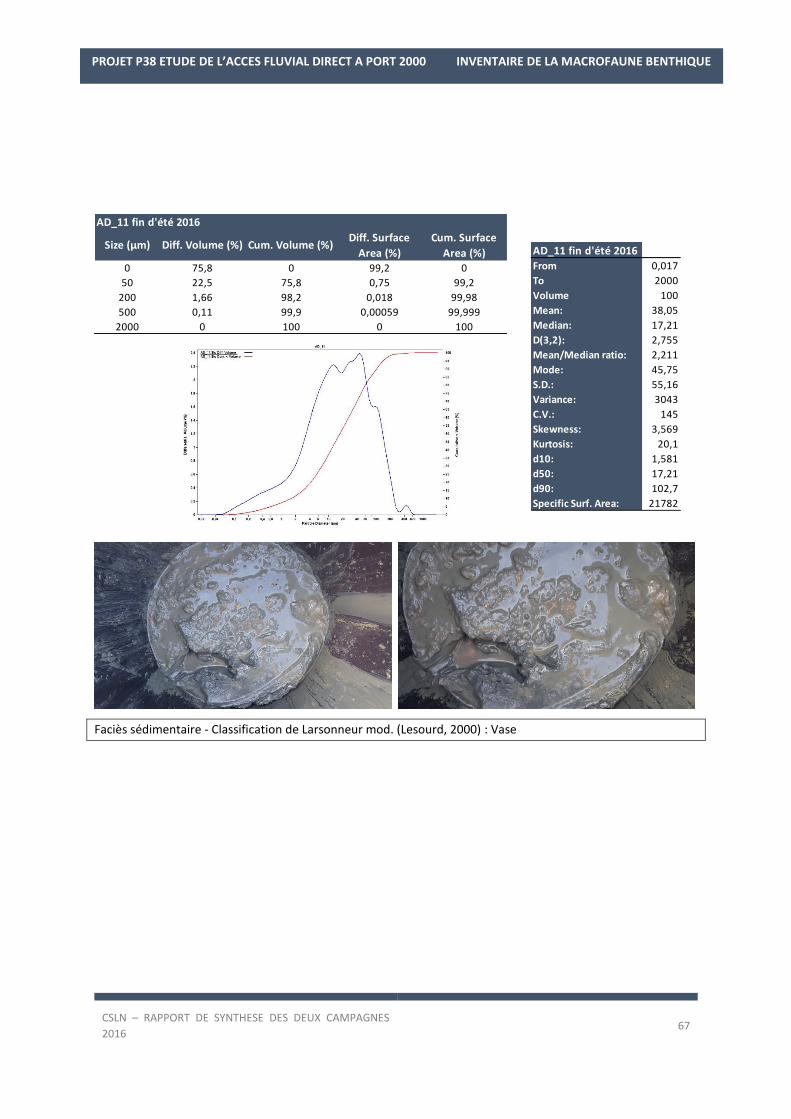
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 67
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Faciès sédimentaire - Classification de Larsonneur mod. (Lesourd, 2000) : Vase
AD_11 fin d'été 2016
Size (µm) Diff. Volume (%) Cum. Volume (%)Diff. Surface
Area (%)
Cum. Surface
Area (%)
0 75,8 0 99,2 0
50 22,5 75,8 0,75 99,2
200 1,66 98,2 0,018 99,98
500 0,11 99,9 0,00059 99,999
2000 0 100 0 100
AD_11 fin d'été 2016
From 0,017
To 2000
Volume 100
Mean: 38,05
Median: 17,21
D(3,2): 2,755
Mean/Median ratio: 2,211
Mode: 45,75
S.D.: 55,16
Variance: 3043
C.V.: 145
Skewness: 3,569
Kurtosis: 20,1
d10: 1,581
d50: 17,21
d90: 102,7
Specific Surf. Area: 21782
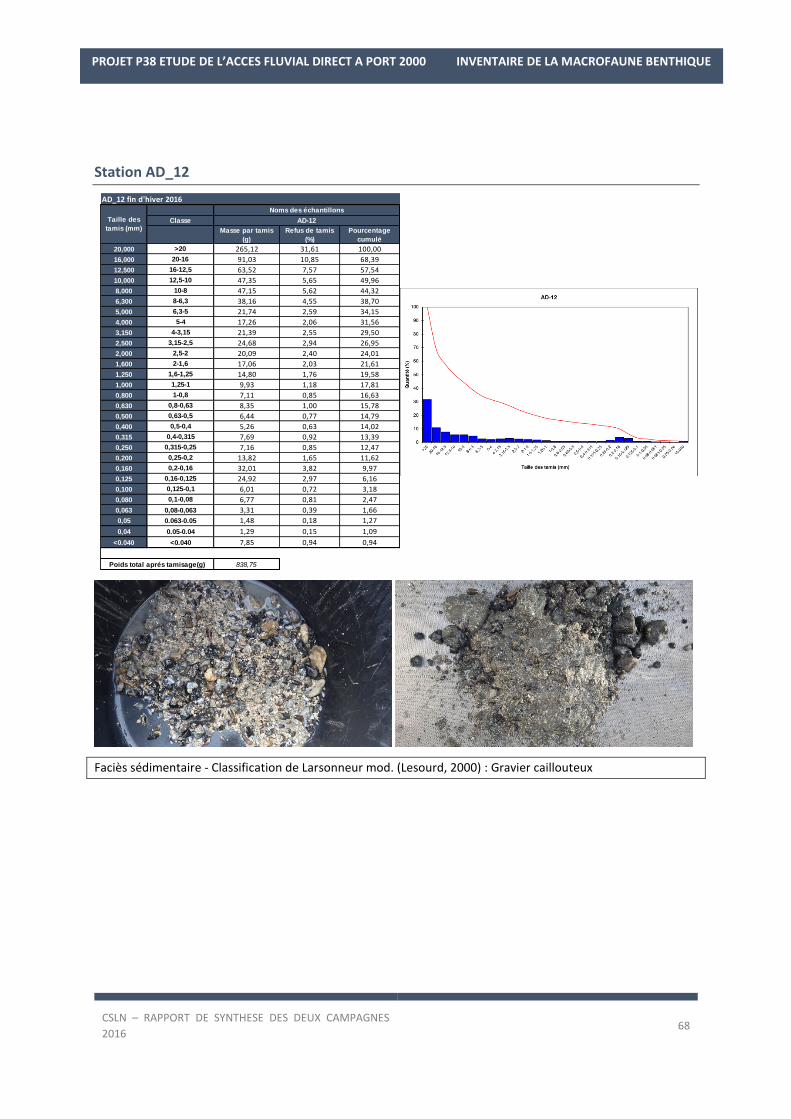
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 68
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Station AD_12
Faciès sédimentaire - Classification de Larsonneur mod. (Lesourd, 2000) : Gravier caillouteux
AD_12 fin d'hiver 2016
Classe
Masse par tamis
(g)
Refus de tamis
(%)
Pourcentage
cumulé
20,000 >20 265,12 31,61 100,00
16,000 20-16 91,03 10,85 68,39
12,500 16-12,5 63,52 7,57 57,54
10,000 12,5-10 47,35 5,65 49,96
8,000 10-8 47,15 5,62 44,32
6,300 8-6,3 38,16 4,55 38,70
5,000 6,3-5 21,74 2,59 34,15
4,000 5-4 17,26 2,06 31,56
3,150 4-3,15 21,39 2,55 29,50
2,500 3,15-2,5 24,68 2,94 26,95
2,000 2,5-2 20,09 2,40 24,01
1,600 2-1,6 17,06 2,03 21,61
1,250 1,6-1,25 14,80 1,76 19,58
1,000 1,25-1 9,93 1,18 17,81
0,800 1-0,8 7,11 0,85 16,63
0,630 0,8-0,63 8,35 1,00 15,78
0,500 0,63-0,5 6,44 0,77 14,79
0,400 0,5-0,4 5,26 0,63 14,02
0,315 0,4-0,315 7,69 0,92 13,39
0,250 0,315-0,25 7,16 0,85 12,47
0,200 0,25-0,2 13,82 1,65 11,62
0,160 0,2-0,16 32,01 3,82 9,97
0,125 0,16-0,125 24,92 2,97 6,16
0,100 0,125-0,1 6,01 0,72 3,18
0,080 0,1-0,08 6,77 0,81 2,47
0,063 0,08-0,063 3,31 0,39 1,66
0,05 0.063-0.05 1,48 0,18 1,27
0,04 0.05-0.04 1,29 0,15 1,09
<0.040 <0.040 7,85 0,94 0,94
838,75
Taille des
tamis (mm)
Noms des échantillons
AD-12
Poids total aprés tamisage(g)
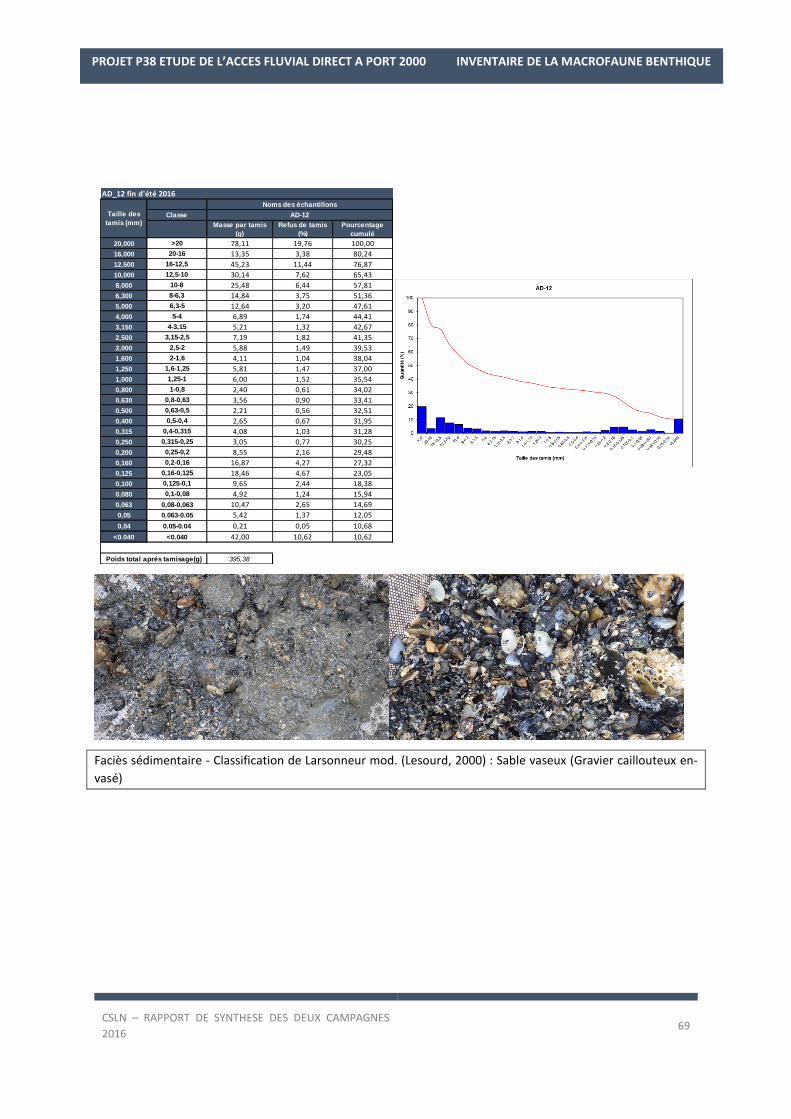
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 69
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Faciès sédimentaire - Classification de Larsonneur mod. (Lesourd, 2000) : Sable vaseux (Gravier caillouteux en-
vasé)
AD_12 fin d'été 2016
Classe
Masse par tamis
(g)
Refus de tamis
(%)
Pourcentage
cumulé
20,000 >20 78,11 19,76 100,00
16,000 20-16 13,35 3,38 80,24
12,500 16-12,5 45,23 11,44 76,87
10,000 12,5-10 30,14 7,62 65,43
8,000 10-8 25,48 6,44 57,81
6,300 8-6,3 14,84 3,75 51,36
5,000 6,3-5 12,64 3,20 47,61
4,000 5-4 6,89 1,74 44,41
3,150 4-3,15 5,21 1,32 42,67
2,500 3,15-2,5 7,19 1,82 41,35
2,000 2,5-2 5,88 1,49 39,53
1,600 2-1,6 4,11 1,04 38,04
1,250 1,6-1,25 5,81 1,47 37,00
1,000 1,25-1 6,00 1,52 35,54
0,800 1-0,8 2,40 0,61 34,02
0,630 0,8-0,63 3,56 0,90 33,41
0,500 0,63-0,5 2,21 0,56 32,51
0,400 0,5-0,4 2,65 0,67 31,95
0,315 0,4-0,315 4,08 1,03 31,28
0,250 0,315-0,25 3,05 0,77 30,25
0,200 0,25-0,2 8,55 2,16 29,48
0,160 0,2-0,16 16,87 4,27 27,32
0,125 0,16-0,125 18,46 4,67 23,05
0,100 0,125-0,1 9,65 2,44 18,38
0,080 0,1-0,08 4,92 1,24 15,94
0,063 0,08-0,063 10,47 2,65 14,69
0,05 0.063-0.05 5,42 1,37 12,05
0,04 0.05-0.04 0,21 0,05 10,68
<0.040 <0.040 42,00 10,62 10,62
395,38
Taille des
tamis (mm)
Noms des échantillons
AD-12
Poids total aprés tamisage(g)

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 70
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
ANNEXE 2 : Liste faunistique
Validité des noms et autorité des espèces : World Register Marine Species, 25/06/2014
GE : Species List v. Nov2014 (http://ambi.azti.es)/ espèce introduite
esp. constante > 50%
esp. commune ]25%-50%]
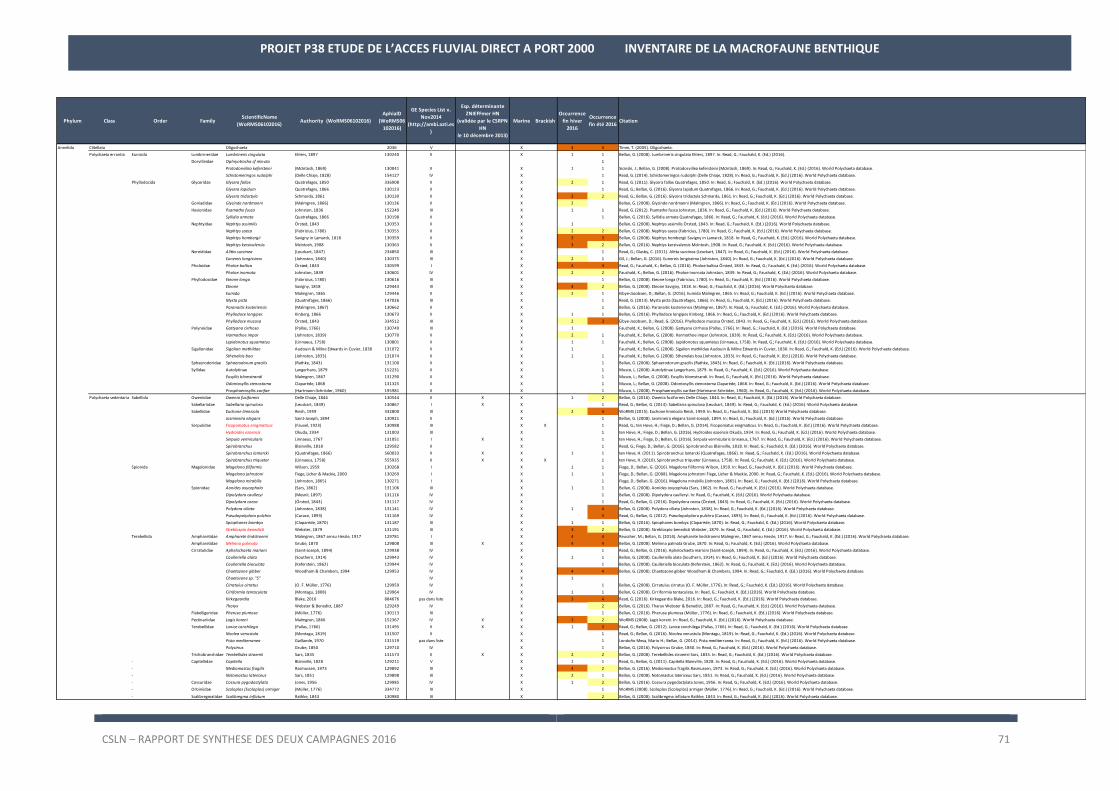
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES 2016 71
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Phylum Class Order FamilyScientificName
(WoRMS06102016)Authority (WoRMS06102016)
AphiaID
(WoRMS06
102016)
GE Species List v.
Nov2014
(http://ambi.azti.es
)
Esp. déterminante
ZNIEFFmer HN
(validée par le CSRPN
HN
le 10 décembre 2013)
Marine Brackish
Occurrence
fin hiver
2016
Occurrence
fin été 2016Citation
Annelida Clitellata Oligochaeta 2036 V X 4 4 Timm, T. (2005). Oligochaeta.
Polychaeta errantia Eunicida Lumbrineridae Lumbrineris cingulata Ehlers, 1897 130240 II X 1 1 Bellan, G. (2008). Lumbrineris cingulata Ehlers, 1897. In: Read, G.; Fauchald, K. (Ed.) (2016).
Dorvilleidae Ophryotrocha cf minuta 1
Protodorvillea kefersteini (McIntosh, 1869) 130041 II X 1 1 Sicinski, J.; Bellan, G. (2008). Protodorvillea kefersteini (McIntosh, 1869). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Schistomeringos rudolphi (Delle Chiaje, 1828) 154127 IV X 1 Read, G. (2014). Schistomeringos rudolphi (Delle Chiaje, 1828). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Phyllodocida Glyceridae Glycera fallax Quatrefages, 1850 336908 II X 2 1 Read, G. (2011). Glycera fallax Quatrefages, 1850. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Glycera lapidum Quatrefages, 1866 130123 II X 1 Read, G.; Bellan, G. (2016). Glycera lapidum Quatrefages, 1866. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Glycera tridactyla Schmarda, 1861 130130 II X 3 2 Read, G.; Bellan, G. (2016). Glycera tridactyla Schmarda, 1861. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Goniadidae Glycinde nordmanni (Malmgren, 1866) 130136 II X 2 Bellan, G. (2008). Glycinde nordmanni (Malmgren, 1866). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Hesionidae Psamathe fusca Johnston, 1836 152249 III X 1 1 Read, G. (2012). Psamathe fusca Johnston, 1836. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Syllidia armata Quatrefages, 1866 130198 II X 1 Bellan, G. (2016). Syllidia armata Quatrefages, 1866. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Nephtyidae Nephtys assimilis Örsted, 1843 130353 II X 1 Bellan, G. (2008). Nephtys assimilis Örsted, 1843. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Nephtys caeca (Fabricius, 1780) 130355 II X 2 2 Bellan, G. (2008). Nephtys caeca (Fabricius, 1780). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Nephtys hombergii Savigny in Lamarck, 1818 130359 II X 3 3 Bellan, G. (2008). Nephtys hombergii Savigny in Lamarck, 1818. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Nephtys kersivalensis McIntosh, 1908 130363 II X 3 2 Bellan, G. (2016). Nephtys kersivalensis McIntosh, 1908. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Nereididae Alitta succinea (Leuckart, 1847) 234850 III X 1 Read, G.; Glasby, C. (2011). Alitta succinea (Leuckart, 1847). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Eunereis longissima (Johnston, 1840) 130375 III X 2 1 Gil, J.; Bellan, G. (2016). Eunereis longissima (Johnston, 1840). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Pholoidae Pholoe baltica Örsted, 1843 130599 I X 4 4 Read, G.; Fauchald, K.; Bellan, G. (2016). Pholoe baltica Örsted, 1843. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Pholoe inornata Johnston, 1839 130601 IV X 2 2 Fauchald, K.; Bellan, G. (2016). Pholoe inornata Johnston, 1839. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Phyllodocidae Eteone longa (Fabricius, 1780) 130616 III X 1 Bellan, G. (2008). Eteone longa (Fabricius, 1780). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Eteone Savigny, 1818 129443 III X 4 2 Bellan, G. (2008). Eteone Savigny, 1818. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Eumida Malmgren, 1865 129446 II X 2 1 Eibye-Jacobsen, D.; Bellan, G. (2016). Eumida Malmgren, 1865. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Mysta picta (Quatrefages, 1866) 147026 III X 1 Read, G. (2013). Mysta picta (Quatrefages, 1866). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Paranaitis kosteriensis (Malmgren, 1867) 130662 II X 1 Bellan, G. (2016). Paranaitis kosteriensis (Malmgren, 1867). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Phyllodoce longipes Kinberg, 1866 130673 II X 1 1 Bellan, G. (2016). Phyllodoce longipes Kinberg, 1866. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Phyllodoce mucosa Örsted, 1843 334512 III X 2 3 Eibye-Jacobsen, D.; Read, G. (2016). Phyllodoce mucosa Örsted, 1843. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Polynoidae Gattyana cirrhosa (Pallas, 1766) 130749 III X 1 Fauchald, K.; Bellan, G. (2008). Gattyana cirrhosa (Pallas, 1766). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Harmothoe impar (Johnston, 1839) 130770 II X 2 1 Fauchald, K.; Bellan, G. (2008). Harmothoe impar (Johnston, 1839). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Lepidonotus squamatus (Linnaeus, 1758) 130801 II X 1 1 Fauchald, K.; Bellan, G. (2008). Lepidonotus squamatus (Linnaeus, 1758). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Sigalionidae Sigalion mathildae Audouin & Milne Edwards in Cuvier, 1830 131072 II X 1 Fauchald, K.; Bellan, G. (2008). Sigalion mathildae Audouin & Milne Edwards in Cuvier, 1830. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Sthenelais boa (Johnston, 1833) 131074 II X 1 1 Fauchald, K.; Bellan, G. (2008). Sthenelais boa (Johnston, 1833). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Sphaerodoridae Sphaerodorum gracilis (Rathke, 1843) 131100 II X 1 Bellan, G. (2008). Sphaerodorum gracilis (Rathke, 1843). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Syllidae Autolytinae Langerhans, 1879 152231 II X 1 Musco, L. (2008). Autolytinae Langerhans, 1879. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Eusyllis blomstrandi Malmgren, 1867 131290 II X 1 Musco, L.; Bellan, G. (2008). Eusyllis blomstrandi. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Odontosyllis ctenostoma Claparède, 1868 131325 II X 1 Musco, L.; Bellan, G. (2008). Odontosyllis ctenostoma Claparède, 1868. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Prosphaerosyllis xarifae (Hartmann-Schröder, 1960) 195981 II X 1 Musco, L. (2008). Prosphaerosyllis xarifae (Hartmann-Schröder, 1960). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Polychaeta sedentaria Sabellida Oweniidae Owenia fusiformis Delle Chiaje, 1844 130544 II X X 1 2 Bellan, G. (2016). Owenia fusiformis Delle Chiaje, 1844. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Sabellariidae Sabellaria spinulosa (Leuckart, 1849) 130867 I X X 1 Read, G.; Bellan, G. (2014). Sabellaria spinulosa (Leuckart, 1849). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Sabellidae Euchone limnicola Reish, 1959 332800 III X 2 4 WoRMS (2015). Euchone limnicola Reish, 1959. In: Read, G.; Fauchald, K. (Ed.) (2015) World Polychaeta database.
Jasmineira elegans Saint-Joseph, 1894 130921 II X 1 Bellan, G. (2008). Jasmineira elegans Saint-Joseph, 1894. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Serpulidae Ficopomatus enigmaticus (Fauvel, 1923) 130988 III X X 1 Read, G.; ten Hove, H.; Fiege, D.; Bellan, G. (2014). Ficopomatus enigmaticus. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Hydroides ezoensis Okuda, 1934 131003 III X 1 ten Hove, H.; Fiege, D.; Bellan, G. (2016). Hydroides ezoensis Okuda, 1934. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Serpula vermicularis Linnaeus, 1767 131051 I X X 1 ten Hove, H.; Fiege, D.; Bellan, G. (2016). Serpula vermicularis Linnaeus, 1767. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Spirobranchus Blainville, 1818 129582 II X 1 Read, G.; Fiege, D.; Bellan, G. (2016). Spirobranchus Blainville, 1818. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Spirobranchus lamarcki (Quatrefages, 1866) 560033 II X X 1 1 ten Hove, H. (2011). Spirobranchus lamarcki (Quatrefages, 1866). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Spirobranchus triqueter (Linnaeus, 1758) 555935 II X X X 1 ten Hove, H. (2010). Spirobranchus triqueter (Linnaeus, 1758). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Spionida Magelonidae Magelona filiformis Wilson, 1959 130268 I X 1 1 Fiege, D.; Bellan, G. (2016). Magelona filiformis Wilson, 1959. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Magelona johnstoni Fiege, Licher & Mackie, 2000 130269 I X 1 1 Fiege, D.; Bellan, G. (2008). Magelona johnstoni Fiege, Licher & Mackie, 2000. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Magelona mirabilis (Johnston, 1865) 130271 I X 1 Fiege, D.; Bellan, G. (2016). Magelona mirabilis (Johnston, 1865). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Spionidae Aonides oxycephala (Sars, 1862) 131106 III X 1 1 Bellan, G. (2008). Aonides oxycephala (Sars, 1862). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Dipolydora caulleryi (Mesnil, 1897) 131116 IV X 1 Bellan, G. (2008). Dipolydora caulleryi. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Dipolydora coeca (Örsted, 1843) 131117 IV X 1 Read, G.; Bellan, G. (2016). Dipolydora coeca (Örsted, 1843). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Polydora ciliata (Johnston, 1838) 131141 IV X 1 4 Bellan, G. (2008). Polydora ciliata (Johnston, 1838). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Pseudopolydora pulchra (Carazzi, 1893) 131169 IV X 4 Read, G.; Bellan, G. (2012). Pseudopolydora pulchra (Carazzi, 1893). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Spiophanes bombyx (Claparède, 1870) 131187 III X 1 1 Bellan, G. (2016). Spiophanes bombyx (Claparède, 1870). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Streblospio benedicti Webster, 1879 131191 III X 3 2 Bellan, G. (2008). Streblospio benedicti Webster, 1879. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Terebellida Ampharetidae Ampharete lindstroemi Malmgren, 1867 sensu Hessle, 1917 129781 I X 4 4 Reuscher, M.; Bellan, G. (2014). Ampharete lindstroemi Malmgren, 1867 sensu Hessle, 1917. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Ampharetidae Melinna palmata Grube, 1870 129808 III X X 4 4 Bellan, G. (2008). Melinna palmata Grube, 1870. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Cirratulidae Aphelochaeta marioni (Saint-Joseph, 1894) 129938 IV X 1 Read, G.; Bellan, G. (2016). Aphelochaeta marioni (Saint-Joseph, 1894). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Caulleriella alata (Southern, 1914) 129943 IV X 1 1 Bellan, G. (2008). Caulleriella alata (Southern, 1914). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Caulleriella bioculata (Keferstein, 1862) 129944 IV X 1 Bellan, G. (2008). Caulleriella bioculata (Keferstein, 1862). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Chaetozone gibber Woodham & Chambers, 1994 129953 IV X 4 4 Bellan, G. (2008). Chaetozone gibber Woodham & Chambers, 1994. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Chaetozone sp. "S" IV X 1
Cirratulus cirratus (O. F. Müller, 1776) 129959 IV X 1 Bellan, G. (2008). Cirratulus cirratus (O. F. Müller, 1776). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Cirriformia tentaculata (Montagu, 1808) 129964 IV X 1 1 Bellan, G. (2008). Cirriformia tentaculata. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Kirkegaardia Blake, 2016 884676 pas dans liste X 3 4 Read, G. (2016). Kirkegaardia Blake, 2016. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Tharyx Webster & Benedict, 1887 129249 IV X 2 Bellan, G. (2016). Tharyx Webster & Benedict, 1887. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Flabelligeridae Pherusa plumosa (Müller, 1776) 130113 III X 1 Bellan, G. (2016). Pherusa plumosa (Müller, 1776). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Pectinariidae Lagis koreni Malmgren, 1866 152367 IV X X 3 2 WoRMS (2008). Lagis koreni. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Terebellidae Lanice conchilega (Pallas, 1766) 131495 II X X 1 3 Read, G.; Bellan, G. (2012). Lanice conchilega (Pallas, 1766). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Nicolea venustula (Montagu, 1819) 131507 II X 1 Read, G.; Bellan, G. (2016). Nicolea venustula (Montagu, 1819). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Pista mediterranea Gaillande, 1970 131519 pas dans liste X 1 Londoño Mesa, Mario H.; Bellan, G. (2014). Pista mediterranea. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Polycirrus Grube, 1850 129710 IV X 1 Bellan, G. (2016). Polycirrus Grube, 1850. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
Trichobranchidae Terebellides stroemii Sars, 1835 131573 II X X 2 2 Bellan, G. (2008). Terebellides stroemii Sars, 1835. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
- Capitellidae Capitella Blainville, 1828 129211 V X 1 1 Read, G.; Bellan, G. (2011). Capitella Blainville, 1828. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
- Mediomastus fragilis Rasmussen, 1973 129892 III X 4 2 Bellan, G. (2016). Mediomastus fragilis Rasmussen, 1973. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
- Notomastus latericeus Sars, 1851 129898 III X 2 1 Bellan, G. (2008). Notomastus latericeus Sars, 1851. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
- Cossuridae Cossura pygodactylata Jones, 1956 129985 IV X 1 2 Bellan, G. (2016). Cossura pygodactylata Jones, 1956. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
- Orbiniidae Scoloplos (Scoloplos) armiger (Müller, 1776) 334772 III X 1 WoRMS (2008). Scoloplos (Scoloplos) armiger (Müller, 1776). In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.
- Scalibregmatidae Scalibregma inflatum Rathke, 1843 130980 III X 2 Bellan, G. (2008). Scalibregma inflatum Rathke, 1843. In: Read, G.; Fauchald, K. (Ed.) (2016). World Polychaeta database.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES 2016 72
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Arthropoda Malacostraca Amphipoda Ampeliscidae Ampelisca brevicornis (Costa, 1853) 101891 I X X 3 3 Costello, M.; Bellan-Santini, D. (2004). Ampelisca brevicornis (Costa, 1853).
Ampelisca diadema (Costa, 1853) 101896 II X X 1 Costello, M.; Bellan-Santini, D. (2016). Ampelisca diadema (Costa, 1853).
Ampelisca spinipes Boeck, 1861 101928 I X 1 Costello, M.; Bellan-Santini, D. (2016). Ampelisca spinipes Boeck, 1861.
Ampelisca tenuicornis Lilljeborg, 1855 101930 I X X 1 2 Costello, M.; Bellan-Santini, D. (2004). Ampelisca tenuicornis.
Amphilochidae Apolochus neapolitanus (Della Valle, 1893) 236495 II X 1 1 Kim, Y.; Costello, M.; Bellan-Santini, D. (2014). Amphilochus neapolitanus Della Valle, 1893.
Aoridae Aora gracilis (Bate, 1857) 102012 I X 1 Costello, M.; Bellan-Santini, D. (2016). Aora gracilis (Bate, 1857).
Atylidae Atylus vedlomensis (Bate & Westwood, 1862) 102132 I X 1 Costello, M.; Bellan-Santini, D. (2016). Atylus vedlomensis (Bate & Westwood, 1862).
Caprellidae Caprella erethizon Mayer, 1901 101831 pas dans liste X 1 Costello, M.; Bellan-Santini, D. (2004). Caprella erethizon Mayer, 1901.
Pariambus typicus (Krøyer, 1884) 101857 III X 1 Lowry, J.; Costello, M.; Bellan-Santini, D. (2009). Pariambus typicus (Krøyer, 1884).
Phtisica marina Slabber, 1769 101864 I X 1 Costello, M.; Bellan-Santini, D. (2010). Phtisica marina Slabber, 1769.
Cheirocratidae Cheirocratus intermedius G.O. Sars, 1894 102795 I X 1 Horton, T.; Lowry, J.; Costello, M.; Bellan-Santini, D. (2016). Cheirocratus intermedius G.O. Sars, 1894.
Corophiidae Monocorophium acherusicum (Costa, 1853) 225814 III X 3 d'Udekem d'Acoz, C.; Lowry, J. (2011). Monocorophium acherusicum (Costa, 1853).
Monocorophium insidiosum (Crawford, 1937) 148592 III X 1 Lowry, J. (2010). Monocorophium insidiosum (Crawford, 1937).
Iphimediidae Iphimedia minuta G. O. Sars, 1883 102345 I X 1 Costello, M.; Bellan-Santini, D. (2016). Iphimedia minuta G. O. Sars, 1883.
Ischyroceridae Ericthonius punctatus (Bate, 1857) 102408 I X 2 Costello, M.; Bellan-Santini, D. (2016). Ericthonius punctatus (Bate, 1857).
Jassa herdmani (Walker, 1893) 102432 pas dans liste X 1 Costello, M.; Bellan-Santini, D. (2004). Jassa herdmani (Walker, 1893).
Jassa marmorata Holmes, 1905 102433 V X 2 Lowry, J.; Costello, M.; Bellan-Santini, D. (2010). Jassa marmorata Holmes, 1905.
Leucothoidae Leucothoe incisa (Robertson, 1892) 102460 I X 1 1 Lowry, J.; Costello, M.; Bellan-Santini, D. (2010). Leucothoe incisa.
Melitidae Abludomelita obtusata (Montagu, 1813) 102788 III X 1 Costello, M.; Bellan-Santini, D. (2004). Abludomelita obtusata (Montagu, 1813).
Oedicerotidae Perioculodes longimanus (Bate & Westwood, 1868) 102915 II X 2 Costello, M.; Bellan-Santini, D. (2004). Perioculodes longimanus (Bate & Westwood, 1868).
Photidae Photis longicaudata (Bate & Westwood, 1862) 102383 I X 1 Costello, M.; Bellan-Santini, D. (2004). Photis longicaudata (Bate & Westwood, 1862).
Cumacea Diastylidae Diastylis bradyi Norman, 1879 110472 II X X 2 Watling, L. (2004). Diastylis bradyi Norman, 1879. In: Watling, L.; Gerken, S. (2016). World Cumacea database.
Leuconidae Eudorella truncatula (Bate, 1856) 110535 I X X 2 Watling, L. (2004). Eudorella truncatula (Bate, 1856). In: Watling, L.; Gerken, S. (2016). World Cumacea database.
Decapoda Carcinidae Carcinus maenas (Linnaeus, 1758) 107381 III X 1 1 Fransen, C.; Türkay, M. (2007). Carcinus maenas (Linnaeus, 1758).
Crangon crangon (Linnaeus, 1758) 107552 I X X 3 Fransen, C.; Türkay, M. (2007). Crangon crangon (Linnaeus, 1758).
Philocheras bispinosus bispinosus (Hailstone, 1835) 108207 I X 2 Fransen, C.; Türkay, M. (2011). Philocheras bispinosus bispinosus (Hailstone, 1835).
Diogenidae Diogenes pugilator (Roux, 1829) 107199 II X 1 Lemaitre, R.; Türkay, M. (2013). Diogenes pugilator (Roux, 1829). In: Lemaitre, R.; McLaughlin, P. (2016). World Paguroidea & Lomisoidea database.
Galatheidae Galathea intermedia Lilljeborg, 1851 107150 I X 1 Türkay, M. (2004). Galathea intermedia Lilljeborg, 1851.
Hippolytidae Hippolyte varians Leach, 1814 [in Leach, 1813-1814] 107518 I X 1 Fransen, C.; Türkay, M. (2011). Hippolyte varians Leach, 1814 [in Leach, 1813-1814].
Paguridae Pagurus bernhardus (Linnaeus, 1758) 107232 II X 2 Türkay, M. (2004). Pagurus bernhardus (Linnaeus, 1758). In: Lemaitre, R.; McLaughlin, P. (2016). World Paguroidea & Lomisoidea database.
Pandalidae Pandalina brevirostris (Rathke, 1843) 107647 II X X 1 Fransen, C.; Türkay, M. (2007). Pandalina brevirostris (Rathke, 1843).
Pilumnidae Pilumnus hirtellus (Linnaeus, 1761) 107418 I X 1 Fransen, C.; Türkay, M. (2007). Pilumnus hirtellus (Linnaeus, 1761).
Pinnotheridae Pinnotheres pisum (Linnaeus, 1767) 107473 III X 1 1 Fransen, C.; Türkay, M. (2007). Pinnotheres pisum (Linnaeus, 1767).
Polybiidae Liocarcinus depurator (Linnaeus, 1758) 107387 I X 1 Fransen, C.; Türkay, M. (2016). Liocarcinus depurator (Linnaeus, 1758).
Polybiidae Necora puber (Linnaeus, 1767) 107398 I X 1 Fransen, C.; Türkay, M. (2007). Necora puber (Linnaeus, 1767).
Porcellanidae Pisidia longicornis (Linnaeus, 1767) 107188 I X 1 2 Türkay, M. (2016). Pisidia longicornis (Linnaeus, 1767).
Processidae Processa edulis edulis (Risso, 1816) 108337 I X 1 Fransen, C.; Türkay, M. (2011). Processa edulis (Risso, 1816).
Thoridae Eualus cranchii (Leach, 1817 [in Leach, 1815-1875]) 156083 I X 1 Fransen, C.; De Grave, S. (2011). Eualus cranchii (Leach, 1817 [in Leach, 1815-1875]).
Isopoda Anthuridae Cyathura carinata (Krøyer, 1847) 118474 III X 1Poore, G.; Schotte, M. (2009). Cyathura carinata (Krøyer, 1847). In: Boyko, C.B; Bruce, N.L.; Merrin, K. L.; Ota, Y.; Poore, G.C.B.; Taiti, S.; Schotte, M. & Wilson, G.D.F. (Eds) (2008 onwards). World Marine, Freshwater and
Terrestrial Isopod Crustaceans database.
Bopyridae Pleurocrypta porcellanaelongicornis Hesse, 1876 593521 pas dans liste X 1Boyko, Christopher B.; Schotte, M.; Markham, J. (2016). Pleurocrypta porcellanae Hesse, 1876. In: Boyko, C.B; Bruce, N.L.; Merrin, K. L.; Ota, Y.; Poore, G.C.B.; Taiti, S.; Schotte, M. & Wilson, G.D.F. (Eds) (2008 onwards).
World Marine, Freshwater and Terrestrial Isopod Crustaceans database.
Mysida Mysidae Schistomysis ornata (G. O. Sars, 1864) 120204 II X 1 WoRMS (2004). Schistomysis ornata. In: Mees, J. & K. Meland (Eds) (2012 onwards). World List of Lophogastrida, Stygiomysida and Mysida.
Tanaidacea Tanaididae Zeuxo holdichi Bamber, 1990 416601 III X 2 Anderson, G. (2014). Zeuxo holdichi Bamber, 1990.
Pycnogonida Pantopoda Ammotheidae Achelia echinata Hodge, 1864 134599 I X 1 Bamber, R. (2007). Achelia echinata Hodge, 1864. In: Bamber, R.N., El Nagar, A. & Arango, C. (Eds) (2016). Pycnobase: World Pycnogonida Database.
Nymphonidae Nymphon brevirostre Hodge, 1863 150520 I X 1 Bamber, R. (2007). Nymphon brevirostre Hodge, 1863. In: Bamber, R.N., El Nagar, A. & Arango, C. (Eds) (2016). Pycnobase: World Pycnogonida Database.
- Sessilia Austrobalanidae Austrominius modestus (Darwin, 1854) 712167 II X 1 1 Van Syoc, R. (2016). Austrominius modestus (Darwin, 1854).
Balanidae Balanus crenatus Bruguière, 1789 106215 (Not assigned) X 3 3 Southward, A. (2004). Balanus crenatus Bruguière, 1789.
Perforatus perforatus (Bruguière, 1789) 535477 pas dans liste X 1 Chan, Benny K.K. (2010). Perforatus perforatus (Bruguière, 1789).
Siphonostomatoida Cancerillidae Cancerilla tubulata Dalyell, 1851 135789 pas dans liste X 1 Boxshall, G. (2004). Cancerilla tubulata Dalyell, 1851. In: Walter, T.C. & Boxshall, G. (2016). World of Copepods database.
- - - Copepoda Milne-Edwards, 1840 1080 pas dans liste X X 1 Boxshall, G. (2015). Copepoda.
- - - Larva 1
Echinodermata Echinoidea Spatangoida Loveniidae Echinocardium cordatum (Pennant, 1777) 124392 I X X 1 Kroh, A.; Hansson, H. (2013). Echinocardium cordatum (Pennant, 1777). In: Kroh, A. & Mooi, R. (2016). World Echinoidea Database.
Ophiuroidea Ophiurida Amphiuridae Acrocnida brachiata (Montagu, 1804) 236130 I X X 2 2 Stöhr, S. (2007). Acrocnida brachiata (Montagu, 1804). In: Stöhr, S.; O’Hara, T. & Thuy, B. (Eds) (2016). World Ophiuroidea database.
Amphipholis squamata (Delle Chiaje, 1828) 125064 I X X 1 1 Stöhr, S.; Hansson, H. (2016). Amphipholis squamata (Delle Chiaje, 1828). In: Stöhr, S.; O’Hara, T. & Thuy, B. (Eds) (2016). World Ophiuroidea database.
Ophiuridae Ophiura ophiura (Linnaeus, 1758) 124929 II X X 1 Stöhr, S.; Hansson, H. (2007). Ophiura ophiura (Linnaeus, 1758). In: Stöhr, S.; O’Hara, T. & Thuy, B. (Eds) (2016). World Ophiuroidea database.
Mollusca Bivalvia Adapedonta Pharidae Ensis directus (Conrad, 1843) 140732 I X X 1 1 von Cosel, R.; Gofas, S. (2009). Ensis directus (Conrad, 1843). In: MolluscaBase (2016).
Phaxas pellucidus (Pennant, 1777) 140737 I X X 3 1 Gofas, S. (2004). Phaxas pellucidus (Pennant, 1777). In: MolluscaBase (2016).
Solenidae Solen marginatus Pulteney, 1799 141546 I X 1 Citation: von Cosel, R.; Gofas, S. (2009). Solen marginatus Pulteney, 1799. In: MolluscaBase (2016).
Cardiida Cardiidae Acanthocardia echinata (Linnaeus, 1758) 138992 I X X 2 ter Poorten, J.; Gofas, S. (2012). Acanthocardia echinata (Linnaeus, 1758). In: MolluscaBase (2016).
Acanthocardia paucicostata (G. B. Sowerby II, 1834) 138993 I X 1 ter Poorten, J.; Gofas, S. (2014). Acanthocardia paucicostata (G. B. Sowerby II, 1834). In: MolluscaBase (2016).
Cerastoderma edule (Linnaeus, 1758) 138998 III X 2 2 ter Poorten, J.; Gofas, S. (2014). Cerastoderma edule (Linnaeus, 1758). In: MolluscaBase (2016).
Parvicardium scabrum (Philippi, 1844) 139012 I X 1 ter Poorten, J.; Gofas, S. (2012). Parvicardium scabrum (Philippi, 1844). In: MolluscaBase (2016).
Semelidae Abra alba (W. Wood, 1802) 141433 III X X 4 4 Huber, M.; Gofas, S. (2010). Abra alba (W. Wood, 1802). In: MolluscaBase (2016).
Tellinidae Fabulina fabula (Gmelin, 1791) 146907 I X 1 1 Sartori, André F.; Gofas, S.; Rosenberg, G. (2016). Fabulina fabula (Gmelin, 1791). In: MolluscaBase (2016).
Limecola balthica (Linnaeus, 1758) 880017 III X X 2 Sartori, André F. (2016). Limecola balthica (Linnaeus, 1758). In: MolluscaBase (2016).
Myida Corbulidae Corbula gibba (Olivi, 1792) 139410 IV X X 4 4 Gofas, S. (2008). Corbula gibba. In: MolluscaBase (2016).
Myidae Sphenia binghami Turton, 1822 140432 I X X 3 1 Gofas, S. (2016). Sphenia binghami Turton, 1822. In: MolluscaBase (2016).
Mytilida Mytilidae Mytilus edulis Linnaeus, 1758 140480 III X X 1 1 Gofas, S. (2004). Mytilus edulis Linnaeus, 1758. In: MolluscaBase (2016).
Nuculida Nuculidae Nucula nitidosa Winckworth, 1930 140589 I X X 3 3 Gofas, S. (2004). Nucula nitidosa. In: MolluscaBase (2016).
Pectinida Pectinidae Aequipecten opercularis (Linnaeus, 1758) 140687 I X X 1 Dijkstra, H.; Gofas, S. (2009). Aequipecten opercularis (Linnaeus, 1758). In: MolluscaBase (2016).
Venerida Veneridae Petricolaria pholadiformis (Lamarck, 1818) 156961 I X 1 Huber, M.; Gofas, S. (2010). Petricolaria pholadiformis (Lamarck, 1818). In: MolluscaBase (2016).
Venerupis corrugata (Gmelin, 1791) 181364 I X 2 3 Huber, M.; Gofas, S. (2010). Venerupis corrugata (Gmelin, 1791). In: MolluscaBase (2016).
- Montacutidae Kurtiella bidentata (Montagu, 1803) 345281 III X 4 4 Gofas, S. (2016). Kurtiella bidentata (Montagu, 1803). In: MolluscaBase (2016).
- Mactridae Mactra stultorum (Linnaeus, 1758) 140299 I X X 1 Gofas, S. (2004). Mactra stultorum (Linnaeus, 1758). In: MolluscaBase (2016).
- Pandoridae Pandora inaequivalvis (Linnaeus, 1758) 140674 I X 1 Gofas, S. (2004). Pandora inaequivalvis (Linnaeus, 1758). In: MolluscaBase (2016).
- Mactridae Spisula subtruncata (da Costa, 1778) 140302 I X X 3 3 Gofas, S. (2004). Spisula subtruncata (da Costa, 1778). In: MolluscaBase (2016).
- Thraciidae Thracia phaseolina (Lamarck, 1818) 152378 I X 1 Huber, M. (2016). Thracia phaseolina (Lamarck, 1818). In: MolluscaBase (2016).
Gastropoda [unassigned] Caenogastropoda Epitoniidae Epitonium clathrus (Linnaeus, 1758) 146905 I X 1 Bouchet, P.; Gofas, S. (2010). Epitonium clathrus (Linnaeus, 1758). In: MolluscaBase (2016).
Littorinimorpha Calyptraeidae Crepidula fornicata (Linnaeus, 1758) 138963 III X 1 Gofas, S. (2004). Crepidula fornicata. In: MolluscaBase (2016).
Neogastropoda Mangeliidae Mangelia costata (Pennant, 1777) 139272 II X 1 Gofas, S. (2009). Mangelia costata. In: MolluscaBase (2016).
Nassariidae Tritia incrassata (Strøm, 1768) 876825 II X 1 Gofas, S.; Marshall, B. (2016). Tritia incrassata (Strøm, 1768). In: MolluscaBase (2016).
Tritia reticulata (Linnaeus, 1758) 876821 II X 3 4 Gofas, S.; Marshall, B. (2016). Tritia reticulata (Linnaeus, 1758). In: MolluscaBase (2016).
- - Opisthobranchia 382226 pas dans liste X 1 1 Gofas, S. (2009). Opisthobranchia. In: MolluscaBase (2016).
In: Horton, T.; Lowry, J.; De
Broyer, C.; Bellan-Santini,
D.; Coleman, C. O.;
Daneliya, M.; Dauvin, J-C.;
Fišer, C.; Gasca, R.;
Grabowski, M.; Guerra-
García, J. M.; Hendrycks, E.;
Holsinger, J.; Hughes, L.;
Jaume, D.; Jazdzewski, K.;
Just, J.; Kamaltynov, R. M.;
Kim, Y.-H.; King, R.; Krapp-
Schickel, T.; LeCroy, S.; Lörz,
A.-N.; Senna, A. R.; Serejo,
C.; Sket, B.; Tandberg, A.H.;
Thomas, J.; Thurston, M.;
Vader, W.; Väinölä, R.;
Vonk, R.; White, K.; Zeidler,
W. (2016). World
Amphipoda Database.

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES 2016 73
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Cnidaria Anthozoa Anthozoa Ehrenberg, 1834 1292 II X X 3 1 Hoeksema, B. (2013). Anthozoa.
Chordata Ascidiacea Ascidiacea Blainville, 1824 1839 III X 1 Gittenberger, A.; Shenkar, N.; Sanamyan, K. (2015). Ascidiacea. In: Shenkar, N.; Gittenberger, A.; Lambert, G.; Rius, M.; Moreira Da Rocha, R.; Swalla, B.J.; Turon, X. (2016). Ascidiacea World Database.
Ascidiacea sp1 Blainville, 1824 1839 - X 1 Gittenberger, A.; Shenkar, N.; Sanamyan, K. (2015). Ascidiacea. In: Shenkar, N.; Gittenberger, A.; Lambert, G.; Rius, M.; Moreira Da Rocha, R.; Swalla, B.J.; Turon, X. (2016). Ascidiacea World Database.
Ascidiacea sp2 Blainville, 1824 1839 - X 1 Gittenberger, A.; Shenkar, N.; Sanamyan, K. (2015). Ascidiacea. In: Shenkar, N.; Gittenberger, A.; Lambert, G.; Rius, M.; Moreira Da Rocha, R.; Swalla, B.J.; Turon, X. (2016). Ascidiacea World Database.
Ascidiacea sp3 Blainville, 1824 1839 - X 1 Gittenberger, A.; Shenkar, N.; Sanamyan, K. (2015). Ascidiacea. In: Shenkar, N.; Gittenberger, A.; Lambert, G.; Rius, M.; Moreira Da Rocha, R.; Swalla, B.J.; Turon, X. (2016). Ascidiacea World Database.
Ascidiacea sp4 Blainville, 1824 1839 - X 1 Gittenberger, A.; Shenkar, N.; Sanamyan, K. (2015). Ascidiacea. In: Shenkar, N.; Gittenberger, A.; Lambert, G.; Rius, M.; Moreira Da Rocha, R.; Swalla, B.J.; Turon, X. (2016). Ascidiacea World Database.
Bryozoa Bryozoa 146142 II X X Bock, P. (2014). Bryozoa.
Porifera Demospongiae Haplosclerida Chalinidae Haliclona (Haliclona) oculata (Linnaeus, 1759) 132833 pas dans liste Xvan Soest, R. (2013). Haliclona (Haliclona) oculata (Linnaeus, 1759). In: Van Soest, R.W.M; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez de Glasby, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schoenberg, C.;
Klautau, M.; Picton, B.; Kelly, M.; Vacelet, J.; Dohrmann, M.; Díaz, M.-C.; Cárdenas, P.; Carballo, J. L. (2016). World Porifera database.
Cnidaria Hydrozoa Hydrozoa Owen, 1843 1337 I X X Schuchert, P. (2009). Hydrozoa.
Nematoda Nematoda 799 III X 1 1WoRMS (2004). Nematoda. In: Guilini, K.; Bezerra, T.N.; Eisendle-Flöckner, U.; Deprez, T.; Fonseca, G.; Holovachov, O.; Leduc, D.; Miljutin, D.; Moens, T.; Sharma, J.; Smol, N.; Tchesunov, A.; Mokievsky, V.; Vanaverbeke, J.;
Vanreusel, A.; Venekey, V.; Vincx, M. (2016) NeMys: World Database of Free-Living Marine Nematodes.
Nemertea Nemertea spA 152391 III 3 2 Norenburg, J. (2010). Nemertea.
Nemertea spB 152391 III 1 1 Norenburg, J. (2010). Nemertea.
Phoronida Phoronidae Phoronis Wright, 1856 128545 II X 1 1 WoRMS (2016). Phoronis Wright, 1856.
Platyhelminthes Platyhelminthes Minot, 1876 793 II X X 1 1 Artois, T. (2015). Platyhelminthes.
Chordata Ascidiacea Stolidobranchia Styelidae Styela clava Herdman, 1881 103929 II X 1Gittenberger, A.; Moreira da Rocha, R.; Sanamyan, K.; Monniot, C. (2010). Styela clava Herdman, 1881. In: Shenkar, N.; Gittenberger, A.; Lambert, G.; Rius, M.; Moreira Da Rocha, R.; Swalla, B.J.; Turon, X. (2016). Ascidiacea
World Database.
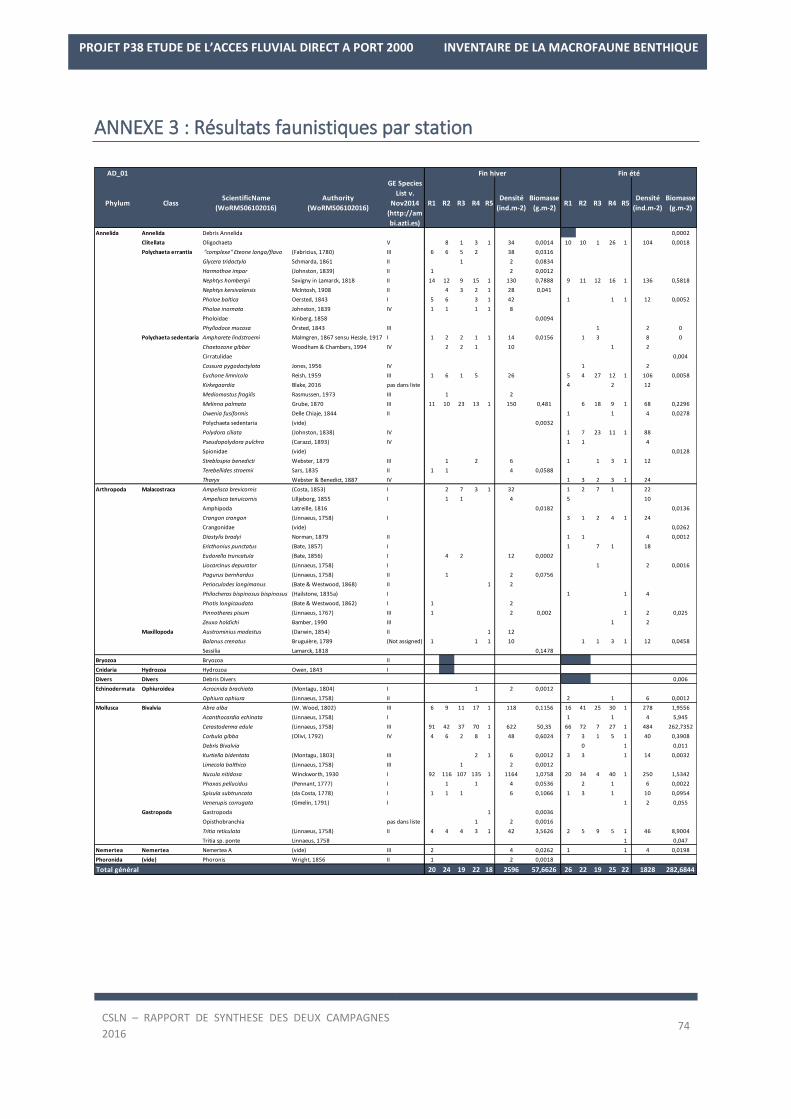
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 74
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
ANNEXE 3 : Résultats faunistiques par station
AD_01
Phylum ClassScientificName
(WoRMS06102016)
Authority
(WoRMS06102016)
GE Species
List v.
Nov2014
(http://am
bi.azti.es)
R1 R2 R3 R4 R5Densité
(ind.m-2)
Biomasse
(g.m-2)R1 R2 R3 R4 R5
Densité
(ind.m-2)
Biomasse
(g.m-2)
Annelida Annelida Debris Annelida 0,0002
Clitellata Oligochaeta V 8 1 3 1 34 0,0014 10 10 1 26 1 104 0,0018
Polychaeta errantia "complexe" Eteone longa/flava (Fabricius, 1780) III 6 6 5 2 38 0,0316
Glycera tridactyla Schmarda, 1861 II 1 2 0,0834
Harmothoe impar (Johnston, 1839) II 1 2 0,0012
Nephtys hombergii Savigny in Lamarck, 1818 II 14 12 9 15 1 130 0,7888 9 11 12 16 1 136 0,5818
Nephtys kersivalensis McIntosh, 1908 II 4 3 2 1 28 0,041
Pholoe baltica Oersted, 1843 I 5 6 3 1 42 1 1 1 12 0,0052
Pholoe inornata Johnston, 1839 IV 1 1 1 1 8
Pholoidae Kinberg, 1858 0,0094
Phyllodoce mucosa Örsted, 1843 III 1 2 0
Polychaeta sedentaria Ampharete lindstroemi Malmgren, 1867 sensu Hessle, 1917 I 1 2 2 1 1 14 0,0156 1 3 8 0
Chaetozone gibber Woodham & Chambers, 1994 IV 2 2 1 10 1 2
Cirratulidae 0,004
Cossura pygodactylata Jones, 1956 IV 1 2
Euchone limnicola Reish, 1959 III 1 6 1 5 26 5 4 27 12 1 106 0,0058
Kirkegaardia Blake, 2016 pas dans liste 4 2 12
Mediomastus fragilis Rasmussen, 1973 III 1 2
Melinna palmata Grube, 1870 III 11 10 23 13 1 150 0,481 6 18 9 1 68 0,2296
Owenia fusiformis Delle Chiaje, 1844 II 1 1 4 0,0278
Polychaeta sedentaria (vide) 0,0032
Polydora ciliata (Johnston, 1838) IV 1 7 23 11 1 88
Pseudopolydora pulchra (Carazzi, 1893) IV 1 1 4
Spionidae (vide) 0,0128
Streblospio benedicti Webster, 1879 III 1 2 6 1 1 3 1 12
Terebellides stroemii Sars, 1835 II 1 1 4 0,0588
Tharyx Webster & Benedict, 1887 IV 1 3 2 3 1 24
Arthropoda Malacostraca Ampelisca brevicornis (Costa, 1853) I 2 7 3 1 32 1 2 7 1 22
Ampelisca tenuicornis Lilljeborg, 1855 I 1 1 4 5 10
Amphipoda Latreille, 1816 0,0182 0,0136
Crangon crangon (Linnaeus, 1758) I 3 1 2 4 1 24
Crangonidae (vide) 0,0262
Diastylis bradyi Norman, 1879 II 1 1 4 0,0012
Ericthonius punctatus (Bate, 1857) I 1 7 1 18
Eudorella truncatula (Bate, 1856) I 4 2 12 0,0002
Liocarcinus depurator (Linnaeus, 1758) I 1 2 0,0016
Pagurus bernhardus (Linnaeus, 1758) II 1 2 0,0756
Perioculodes longimanus (Bate & Westwood, 1868) II 1 2
Philocheras bispinosus bispinosus (Hailstone, 1835a) I 1 1 4
Photis longicaudata (Bate & Westwood, 1862) I 1 2
Pinnotheres pisum (Linnaeus, 1767) III 1 2 0,002 1 2 0,025
Zeuxo holdichi Bamber, 1990 III 1 2
Maxillopoda Austrominius modestus (Darwin, 1854) II 1 12
Balanus crenatus Bruguière, 1789 (Not assigned) 1 1 1 10 1 1 3 1 12 0,0458
Sessilia Lamarck, 1818 0,1478
Bryozoa Bryozoa II
Cnidaria Hydrozoa Hydrozoa Owen, 1843 I
Divers Divers Debris Divers 0,006
Echinodermata Ophiuroidea Acrocnida brachiata (Montagu, 1804) I 1 2 0,0012
Ophiura ophiura (Linnaeus, 1758) II 2 1 6 0,0012
Mollusca Bivalvia Abra alba (W. Wood, 1802) III 6 9 11 17 1 118 0,1156 16 41 25 30 1 278 1,9556
Acanthocardia echinata (Linnaeus, 1758) I 1 1 4 5,945
Cerastoderma edule (Linnaeus, 1758) III 91 42 37 70 1 622 50,35 66 72 7 27 1 484 262,7352
Corbula gibba (Olivi, 1792) IV 4 6 2 8 1 48 0,6024 7 3 1 5 1 40 0,3908
Debris Bivalvia 0 1 0,011
Kurtiella bidentata (Montagu, 1803) III 2 1 6 0,0012 3 3 1 14 0,0032
Limecola balthica (Linnaeus, 1758) III 1 2 0,0012
Nucula nitidosa Winckworth, 1930 I 92 116 107 135 1 1164 1,0758 20 34 4 40 1 250 1,5342
Phaxas pellucidus (Pennant, 1777) I 1 1 4 0,0536 2 1 6 0,0022
Spisula subtruncata (da Costa, 1778) I 1 1 1 6 0,1066 1 3 1 10 0,0954
Venerupis corrugata (Gmelin, 1791) I 1 2 0,055
Gastropoda Gastropoda 1 0,0036
Opisthobranchia pas dans liste 1 2 0,0016
Tritia reticulata (Linnaeus, 1758) II 4 4 4 3 1 42 3,5626 2 5 9 5 1 46 8,9004
Tritia sp. ponte Linnaeus, 1758 1 0,047
Nemertea Nemertea Nemertea A (vide) III 2 4 0,0262 1 1 4 0,0198
Phoronida (vide) Phoronis Wright, 1856 II 1 2 0,0018
Total général 20 24 19 22 18 2596 57,6626 26 22 19 25 22 1828 282,6844
Fin étéFin hiver
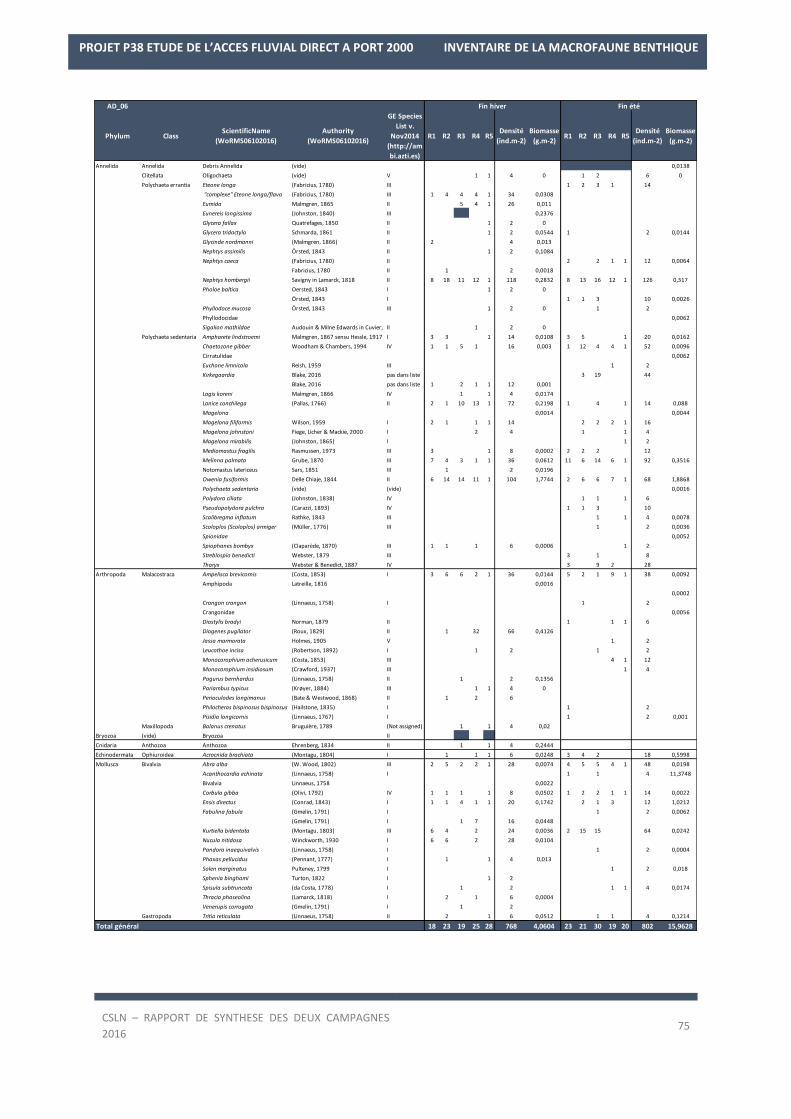
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 75
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
AD_06
Phylum ClassScientificName
(WoRMS06102016)
Authority
(WoRMS06102016)
GE Species
List v.
Nov2014
(http://am
bi.azti.es)
R1 R2 R3 R4 R5Densité
(ind.m-2)
Biomasse
(g.m-2)R1 R2 R3 R4 R5
Densité
(ind.m-2)
Biomasse
(g.m-2)
Annelida Annelida Debris Annelida (vide) 0,0138
Clitellata Oligochaeta (vide) V 1 1 4 0 1 2 6 0
Polychaeta errantia Eteone longa (Fabricius, 1780) III 1 2 3 1 14
"complexe" Eteone longa/flava (Fabricius, 1780) III 1 4 4 4 1 34 0,0308
Eumida Malmgren, 1865 II 5 4 1 26 0,011
Eunereis longissima (Johnston, 1840) III 0,2376
Glycera fallax Quatrefages, 1850 II 1 2 0
Glycera tridactyla Schmarda, 1861 II 1 2 0,0544 1 2 0,0144
Glycinde nordmanni (Malmgren, 1866) II 2 4 0,013
Nephtys assimilis Örsted, 1843 II 1 2 0,1084
Nephtys caeca (Fabricius, 1780) II 2 2 1 1 12 0,0064
Fabricius, 1780 II 1 2 0,0018
Nephtys hombergii Savigny in Lamarck, 1818 II 8 18 11 12 1 118 0,2832 8 13 16 12 1 126 0,317
Pholoe baltica Oersted, 1843 I 1 2 0
Örsted, 1843 I 1 1 3 10 0,0026
Phyllodoce mucosa Örsted, 1843 III 1 2 0 1 2
Phyllodocidae 0,0062
Sigalion mathildae Audouin & Milne Edwards in Cuvier, 1830II 1 2 0
Polychaeta sedentaria Ampharete lindstroemi Malmgren, 1867 sensu Hessle, 1917 I 3 3 1 14 0,0108 3 5 1 20 0,0162
Chaetozone gibber Woodham & Chambers, 1994 IV 1 1 5 1 16 0,003 1 12 4 4 1 52 0,0096
Cirratulidae 0,0062
Euchone limnicola Reish, 1959 III 1 2
Kirkegaardia Blake, 2016 pas dans liste 3 19 44
Blake, 2016 pas dans liste 1 2 1 1 12 0,001
Lagis koreni Malmgren, 1866 IV 1 1 4 0,0174
Lanice conchilega (Pallas, 1766) II 2 1 10 13 1 72 0,2198 1 4 1 14 0,088
Magelona 0,0014 0,0044
Magelona filiformis Wilson, 1959 I 2 1 1 1 14 2 2 2 1 16
Magelona johnstoni Fiege, Licher & Mackie, 2000 I 2 4 1 1 4
Magelona mirabilis (Johnston, 1865) I 1 2
Mediomastus fragilis Rasmussen, 1973 III 3 1 8 0,0002 2 2 2 12
Melinna palmata Grube, 1870 III 7 4 3 1 1 36 0,0612 11 6 14 6 1 92 0,3516
Notomastus latericeus Sars, 1851 III 1 2 0,0196
Owenia fusiformis Delle Chiaje, 1844 II 6 14 14 11 1 104 1,7744 2 6 6 7 1 68 1,8868
Polychaeta sedentaria (vide) (vide) 0,0016
Polydora ciliata (Johnston, 1838) IV 1 1 1 6
Pseudopolydora pulchra (Carazzi, 1893) IV 1 1 3 10
Scalibregma inflatum Rathke, 1843 III 1 1 4 0,0078
Scoloplos (Scoloplos) armiger (Müller, 1776) III 1 2 0,0036
Spionidae 0,0052
Spiophanes bombyx (Claparède, 1870) III 1 1 1 6 0,0006 1 2
Streblospio benedicti Webster, 1879 III 3 1 8
Tharyx Webster & Benedict, 1887 IV 3 9 2 28
Arthropoda Malacostraca Ampelisca brevicornis (Costa, 1853) I 3 6 6 2 1 36 0,0144 5 2 1 9 1 38 0,0092
Amphipoda Latreille, 1816 0,0016
0,0002
Crangon crangon (Linnaeus, 1758) I 1 2
Crangonidae 0,0056
Diastylis bradyi Norman, 1879 II 1 1 1 6
Diogenes pugilator (Roux, 1829) II 1 32 66 0,4126
Jassa marmorata Holmes, 1905 V 1 2
Leucothoe incisa (Robertson, 1892) I 1 2 1 2
Monocorophium acherusicum (Costa, 1853) III 4 1 12
Monocorophium insidiosum (Crawford, 1937) III 1 4
Pagurus bernhardus (Linnaeus, 1758) II 1 2 0,1356
Pariambus typicus (Krøyer, 1884) III 1 1 4 0
Perioculodes longimanus (Bate & Westwood, 1868) II 1 2 6
Philocheras bispinosus bispinosus (Hailstone, 1835) I 1 2
Pisidia longicornis (Linnaeus, 1767) I 1 2 0,001
Maxillopoda Balanus crenatus Bruguière, 1789 (Not assigned) 1 1 4 0,02
Bryozoa (vide) Bryozoa II
Cnidaria Anthozoa Anthozoa Ehrenberg, 1834 II 1 1 4 0,2444
Echinodermata Ophiuroidea Acrocnida brachiata (Montagu, 1804) I 1 1 1 6 0,0248 3 4 2 18 0,5998
Mollusca Bivalvia Abra alba (W. Wood, 1802) III 2 5 2 2 1 28 0,0074 4 5 5 4 1 48 0,0198
Acanthocardia echinata (Linnaeus, 1758) I 1 1 4 11,3748
Bivalvia Linnaeus, 1758 0,0022
Corbula gibba (Olivi, 1792) IV 1 1 1 1 8 0,0502 1 2 2 1 1 14 0,0022
Ensis directus (Conrad, 1843) I 1 1 4 1 1 20 0,1742 2 1 3 12 1,0212
Fabulina fabula (Gmelin, 1791) I 1 2 0,0062
(Gmelin, 1791) I 1 7 16 0,0448
Kurtiella bidentata (Montagu, 1803) III 6 4 2 24 0,0036 2 15 15 64 0,0242
Nucula nitidosa Winckworth, 1930 I 6 6 2 28 0,0104
Pandora inaequivalvis (Linnaeus, 1758) I 1 2 0,0004
Phaxas pellucidus (Pennant, 1777) I 1 1 4 0,013
Solen marginatus Pulteney, 1799 I 1 2 0,018
Sphenia binghami Turton, 1822 I 1 2
Spisula subtruncata (da Costa, 1778) I 1 2 1 1 4 0,0174
Thracia phaseolina (Lamarck, 1818) I 2 1 6 0,0004
Venerupis corrugata (Gmelin, 1791) I 1 2
Gastropoda Tritia reticulata (Linnaeus, 1758) II 2 1 6 0,0512 1 1 4 0,1214
Total général 18 23 19 25 28 768 4,0604 23 21 30 19 20 802 15,9628
Fin étéFin hiver
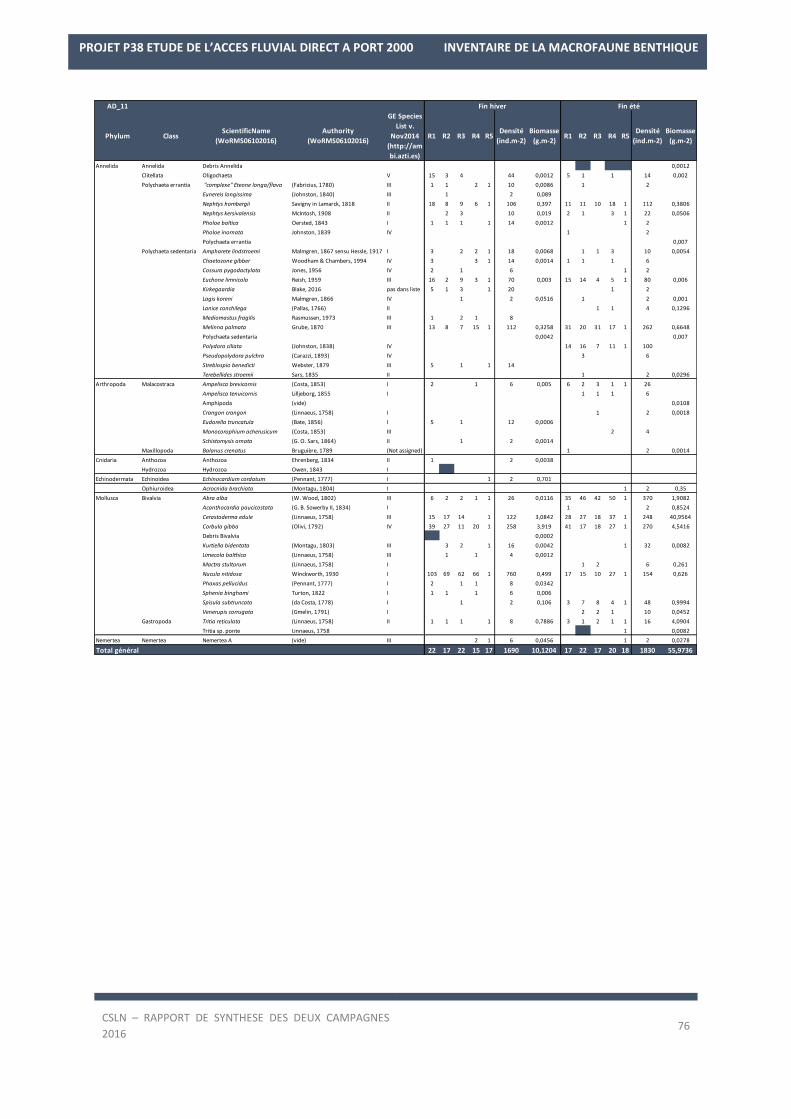
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 76
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
AD_11
Phylum ClassScientificName
(WoRMS06102016)
Authority
(WoRMS06102016)
GE Species
List v.
Nov2014
(http://am
bi.azti.es)
R1 R2 R3 R4 R5Densité
(ind.m-2)
Biomasse
(g.m-2)R1 R2 R3 R4 R5
Densité
(ind.m-2)
Biomasse
(g.m-2)
Annelida Annelida Debris Annelida 0,0012
Clitellata Oligochaeta V 15 3 4 44 0,0012 5 1 1 14 0,002
Polychaeta errantia "complexe" Eteone longa/flava (Fabricius, 1780) III 1 1 2 1 10 0,0086 1 2
Eunereis longissima (Johnston, 1840) III 1 2 0,089
Nephtys hombergii Savigny in Lamarck, 1818 II 18 8 9 6 1 106 0,397 11 11 10 18 1 112 0,3806
Nephtys kersivalensis McIntosh, 1908 II 2 3 10 0,019 2 1 3 1 22 0,0506
Pholoe baltica Oersted, 1843 I 1 1 1 1 14 0,0012 1 2
Pholoe inornata Johnston, 1839 IV 1 2
Polychaeta errantia 0,007
Polychaeta sedentaria Ampharete lindstroemi Malmgren, 1867 sensu Hessle, 1917 I 3 2 2 1 18 0,0068 1 1 3 10 0,0054
Chaetozone gibber Woodham & Chambers, 1994 IV 3 3 1 14 0,0014 1 1 1 6
Cossura pygodactylata Jones, 1956 IV 2 1 6 1 2
Euchone limnicola Reish, 1959 III 16 2 9 3 1 70 0,003 15 14 4 5 1 80 0,006
Kirkegaardia Blake, 2016 pas dans liste 5 1 3 1 20 1 2
Lagis koreni Malmgren, 1866 IV 1 2 0,0516 1 2 0,001
Lanice conchilega (Pallas, 1766) II 1 1 4 0,1296
Mediomastus fragilis Rasmussen, 1973 III 1 2 1 8
Melinna palmata Grube, 1870 III 13 8 7 15 1 112 0,3258 31 20 31 17 1 262 0,6648
Polychaeta sedentaria 0,0042 0,007
Polydora ciliata (Johnston, 1838) IV 14 16 7 11 1 100
Pseudopolydora pulchra (Carazzi, 1893) IV 3 6
Streblospio benedicti Webster, 1879 III 5 1 1 14
Terebellides stroemii Sars, 1835 II 1 2 0,0296
Arthropoda Malacostraca Ampelisca brevicornis (Costa, 1853) I 2 1 6 0,005 6 2 3 1 1 26
Ampelisca tenuicornis Lilljeborg, 1855 I 1 1 1 6
Amphipoda (vide) 0,0108
Crangon crangon (Linnaeus, 1758) I 1 2 0,0018
Eudorella truncatula (Bate, 1856) I 5 1 12 0,0006
Monocorophium acherusicum (Costa, 1853) III 2 4
Schistomysis ornata (G. O. Sars, 1864) II 1 2 0,0014
Maxillopoda Balanus crenatus Bruguière, 1789 (Not assigned) 1 2 0,0014
Cnidaria Anthozoa Anthozoa Ehrenberg, 1834 II 1 2 0,0038
Hydrozoa Hydrozoa Owen, 1843 I
Echinodermata Echinoidea Echinocardium cordatum (Pennant, 1777) I 1 2 0,701
Ophiuroidea Acrocnida brachiata (Montagu, 1804) I 1 2 0,35
Mollusca Bivalvia Abra alba (W. Wood, 1802) III 6 2 2 1 1 26 0,0116 35 46 42 50 1 370 1,9082
Acanthocardia paucicostata (G. B. Sowerby II, 1834) I 1 2 0,8524
Cerastoderma edule (Linnaeus, 1758) III 15 17 14 1 122 3,0842 28 27 18 37 1 248 40,9564
Corbula gibba (Olivi, 1792) IV 39 27 11 20 1 258 3,919 41 17 18 27 1 270 4,5416
Debris Bivalvia 0,0002
Kurtiella bidentata (Montagu, 1803) III 3 2 1 16 0,0042 1 32 0,0082
Limecola balthica (Linnaeus, 1758) III 1 1 4 0,0012
Mactra stultorum (Linnaeus, 1758) I 1 2 6 0,261
Nucula nitidosa Winckworth, 1930 I 103 69 62 66 1 760 0,499 17 15 10 27 1 154 0,626
Phaxas pellucidus (Pennant, 1777) I 2 1 1 8 0,0342
Sphenia binghami Turton, 1822 I 1 1 1 6 0,006
Spisula subtruncata (da Costa, 1778) I 1 2 0,106 3 7 8 4 1 48 0,9994
Venerupis corrugata (Gmelin, 1791) I 2 2 1 10 0,0452
Gastropoda Tritia reticulata (Linnaeus, 1758) II 1 1 1 1 8 0,7886 3 1 2 1 1 16 4,0904
Tritia sp. ponte Linnaeus, 1758 1 0,0082
Nemertea Nemertea Nemertea A (vide) III 2 1 6 0,0456 1 2 0,0278
Total général 22 17 22 15 17 1690 10,1204 17 22 17 20 18 1830 55,9736
Fin étéFin hiver
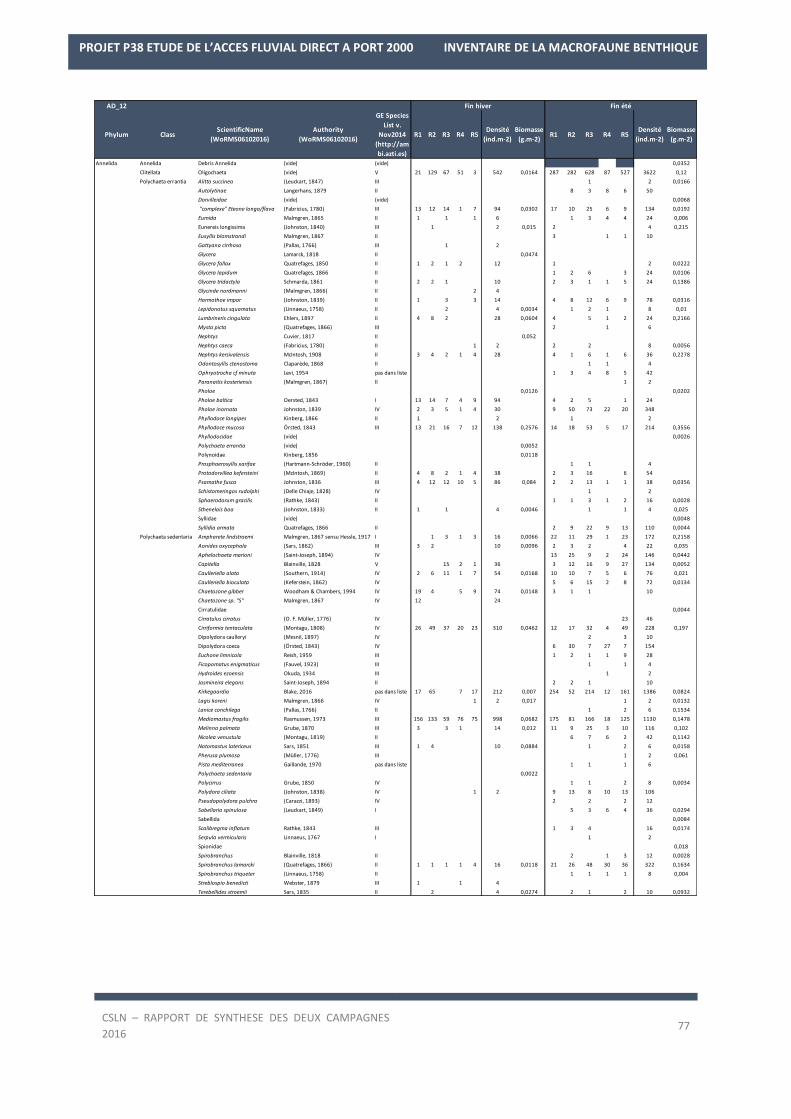
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 77
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
AD_12
Phylum ClassScientificName
(WoRMS06102016)
Authority
(WoRMS06102016)
GE Species
List v.
Nov2014
(http://am
bi.azti.es)
R1 R2 R3 R4 R5Densité
(ind.m-2)
Biomasse
(g.m-2)R1 R2 R3 R4 R5
Densité
(ind.m-2)
Biomasse
(g.m-2)
Annelida Annelida Debris Annelida (vide) (vide) 0,0352
Clitellata Oligochaeta (vide) V 21 129 67 51 3 542 0,0164 287 282 628 87 527 3622 0,12
Polychaeta errantia Alitta succinea (Leuckart, 1847) III 1 2 0,0166
Autolytinae Langerhans, 1879 II 8 3 8 6 50
Dorvilleidae (vide) (vide) 0,0068
"complexe" Eteone longa/flava (Fabricius, 1780) III 13 12 14 1 7 94 0,0302 17 10 25 6 9 134 0,0192
Eumida Malmgren, 1865 II 1 1 1 6 1 3 4 4 24 0,006
Eunereis longissima (Johnston, 1840) III 1 2 0,015 2 4 0,215
Eusyllis blomstrandi Malmgren, 1867 II 3 1 1 10
Gattyana cirrhosa (Pallas, 1766) III 1 2
Glycera Lamarck, 1818 II 0,0474
Glycera fallax Quatrefages, 1850 II 1 2 1 2 12 1 2 0,0222
Glycera lapidum Quatrefages, 1866 II 1 2 6 3 24 0,0106
Glycera tridactyla Schmarda, 1861 II 2 2 1 10 2 3 1 1 5 24 0,1386
Glycinde nordmanni (Malmgren, 1866) II 2 4
Harmothoe impar (Johnston, 1839) II 1 3 3 14 4 8 12 6 9 78 0,0316
Lepidonotus squamatus (Linnaeus, 1758) II 2 4 0,0034 1 2 1 8 0,01
Lumbrineris cingulata Ehlers, 1897 II 4 8 2 28 0,0604 4 5 1 2 24 0,2166
Mysta picta (Quatrefages, 1866) III 2 1 6
Nephtys Cuvier, 1817 II 0,052
Nephtys caeca (Fabricius, 1780) II 1 2 2 2 8 0,0056
Nephtys kersivalensis McIntosh, 1908 II 3 4 2 1 4 28 4 1 6 1 6 36 0,2278
Odontosyllis ctenostoma Claparède, 1868 II 1 1 4
Ophryotrocha cf minuta Levi, 1954 pas dans liste 1 3 4 8 5 42
Paranaitis kosteriensis (Malmgren, 1867) II 1 2
Pholoe 0,0126 0,0202
Pholoe baltica Oersted, 1843 I 13 14 7 4 9 94 4 2 5 1 24
Pholoe inornata Johnston, 1839 IV 2 3 5 1 4 30 9 50 73 22 20 348
Phyllodoce longipes Kinberg, 1866 II 1 2 1 2
Phyllodoce mucosa Örsted, 1843 III 13 21 16 7 12 138 0,2576 14 18 53 5 17 214 0,3556
Phyllodocidae (vide) 0,0026
Polychaeta errantia (vide) 0,0052
Polynoidae Kinberg, 1856 0,0118
Prosphaerosyllis xarifae (Hartmann-Schröder, 1960) II 1 1 4
Protodorvillea kefersteini (McIntosh, 1869) II 4 8 2 1 4 38 2 3 16 6 54
Psamathe fusca Johnston, 1836 III 4 12 12 10 5 86 0,084 2 2 13 1 1 38 0,0356
Schistomeringos rudolphi (Delle Chiaje, 1828) IV 1 2
Sphaerodorum gracilis (Rathke, 1843) II 1 1 3 1 2 16 0,0028
Sthenelais boa (Johnston, 1833) II 1 1 4 0,0046 1 1 4 0,025
Syllidae (vide) 0,0048
Syllidia armata Quatrefages, 1866 II 2 9 22 9 13 110 0,0044
Polychaeta sedentaria Ampharete lindstroemi Malmgren, 1867 sensu Hessle, 1917 I 1 3 1 3 16 0,0066 22 11 29 1 23 172 0,2158
Aonides oxycephala (Sars, 1862) III 3 2 10 0,0096 2 3 2 4 22 0,035
Aphelochaeta marioni (Saint-Joseph, 1894) IV 13 25 9 2 24 146 0,0442
Capitella Blainville, 1828 V 15 2 1 36 3 12 16 9 27 134 0,0052
Caulleriella alata (Southern, 1914) IV 2 6 11 1 7 54 0,0168 10 10 7 5 6 76 0,021
Caulleriella bioculata (Keferstein, 1862) IV 5 6 15 2 8 72 0,0134
Chaetozone gibber Woodham & Chambers, 1994 IV 19 4 5 9 74 0,0148 3 1 1 10
Chaetozone sp. "S" Malmgren, 1867 IV 12 24
Cirratulidae 0,0044
Cirratulus cirratus (O. F. Müller, 1776) IV 23 46
Cirriformia tentaculata (Montagu, 1808) IV 26 49 37 20 23 310 0,0462 12 17 32 4 49 228 0,197
Dipolydora caulleryi (Mesnil, 1897) IV 2 3 10
Dipolydora coeca (Örsted, 1843) IV 6 30 7 27 7 154
Euchone limnicola Reish, 1959 III 1 2 1 1 9 28
Ficopomatus enigmaticus (Fauvel, 1923) III 1 1 4
Hydroides ezoensis Okuda, 1934 III 1 2
Jasmineira elegans Saint-Joseph, 1894 II 2 2 1 10
Kirkegaardia Blake, 2016 pas dans liste 17 65 7 17 212 0,007 254 52 214 12 161 1386 0,0824
Lagis koreni Malmgren, 1866 IV 1 2 0,017 1 2 0,0132
Lanice conchilega (Pallas, 1766) II 1 2 6 0,1534
Mediomastus fragilis Rasmussen, 1973 III 156 133 59 76 75 998 0,0682 175 81 166 18 125 1130 0,1478
Melinna palmata Grube, 1870 III 3 3 1 14 0,012 11 9 25 3 10 116 0,102
Nicolea venustula (Montagu, 1819) II 6 7 6 2 42 0,1142
Notomastus latericeus Sars, 1851 III 1 4 10 0,0884 1 2 6 0,0158
Pherusa plumosa (Müller, 1776) III 1 2 0,061
Pista mediterranea Gaillande, 1970 pas dans liste 1 1 1 6
Polychaeta sedentaria 0,0022
Polycirrus Grube, 1850 IV 1 1 2 8 0,0034
Polydora ciliata (Johnston, 1838) IV 1 2 9 13 8 10 13 106
Pseudopolydora pulchra (Carazzi, 1893) IV 2 2 2 12
Sabellaria spinulosa (Leuckart, 1849) I 5 3 6 4 36 0,0294
Sabellida 0,0084
Scalibregma inflatum Rathke, 1843 III 1 3 4 16 0,0174
Serpula vermicularis Linnaeus, 1767 I 1 2
Spionidae 0,018
Spirobranchus Blainville, 1818 II 2 1 3 12 0,0028
Spirobranchus lamarcki (Quatrefages, 1866) II 1 1 1 1 4 16 0,0118 21 26 48 30 36 322 0,1634
Spirobranchus triqueter (Linnaeus, 1758) II 1 1 1 1 8 0,004
Streblospio benedicti Webster, 1879 III 1 1 4
Terebellides stroemii Sars, 1835 II 2 4 0,0274 2 1 2 10 0,0932
Fin hiver Fin été
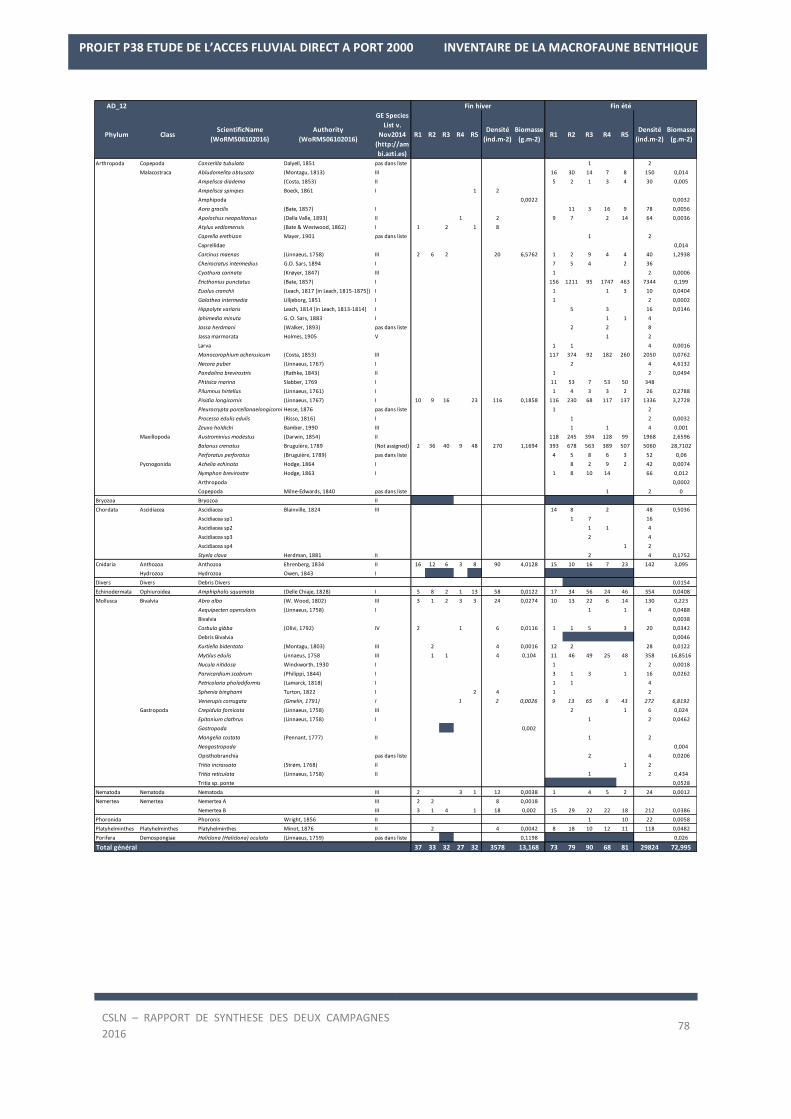
CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 78
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
AD_12
Phylum ClassScientificName
(WoRMS06102016)
Authority
(WoRMS06102016)
GE Species
List v.
Nov2014
(http://am
bi.azti.es)
R1 R2 R3 R4 R5Densité
(ind.m-2)
Biomasse
(g.m-2)R1 R2 R3 R4 R5
Densité
(ind.m-2)
Biomasse
(g.m-2)
Arthropoda Copepoda Cancerilla tubulata Dalyell, 1851 pas dans liste 1 2
Malacostraca Abludomelita obtusata (Montagu, 1813) III 16 30 14 7 8 150 0,014
Ampelisca diadema (Costa, 1853) II 5 2 1 3 4 30 0,005
Ampelisca spinipes Boeck, 1861 I 1 2
Amphipoda 0,0022 0,0032
Aora gracilis (Bate, 1857) I 11 3 16 9 78 0,0056
Apolochus neapolitanus (Della Valle, 1893) II 1 2 9 7 2 14 64 0,0036
Atylus vedlomensis (Bate & Westwood, 1862) I 1 2 1 8
Caprella erethizon Mayer, 1901 pas dans liste 1 2
Caprellidae 0,014
Carcinus maenas (Linnaeus, 1758) III 2 6 2 20 6,5762 1 2 9 4 4 40 1,2938
Cheirocratus intermedius G.O. Sars, 1894 I 7 5 4 2 36
Cyathura carinata (Krøyer, 1847) III 1 2 0,0006
Ericthonius punctatus (Bate, 1857) I 156 1211 95 1747 463 7344 0,199
Eualus cranchii (Leach, 1817 [in Leach, 1815-1875]) I 1 1 3 10 0,0404
Galathea intermedia Lilljeborg, 1851 I 1 2 0,0002
Hippolyte varians Leach, 1814 [in Leach, 1813-1814] I 5 3 16 0,0146
Iphimedia minuta G. O. Sars, 1883 I 1 1 4
Jassa herdmani (Walker, 1893) pas dans liste 2 2 8
Jassa marmorata Holmes, 1905 V 1 2
Larva 1 1 4 0,0016
Monocorophium acherusicum (Costa, 1853) III 117 374 92 182 260 2050 0,0762
Necora puber (Linnaeus, 1767) I 2 4 4,6132
Pandalina brevirostris (Rathke, 1843) II 1 2 0,0494
Phtisica marina Slabber, 1769 I 11 53 7 53 50 348
Pilumnus hirtellus (Linnaeus, 1761) I 1 4 3 3 2 26 0,2788
Pisidia longicornis (Linnaeus, 1767) I 10 9 16 23 116 0,1858 116 230 68 117 137 1336 3,2728
Pleurocrypta porcellanaelongicornisHesse, 1876 pas dans liste 1 2
Processa edulis edulis (Risso, 1816) I 1 2 0,0032
Zeuxo holdichi Bamber, 1990 III 1 1 4 0,001
Maxillopoda Austrominius modestus (Darwin, 1854) II 118 245 394 128 99 1968 2,6596
Balanus crenatus Bruguière, 1789 (Not assigned) 2 36 40 9 48 270 1,1694 393 678 563 389 507 5060 28,7102
Perforatus perforatus (Bruguière, 1789) pas dans liste 4 5 8 6 3 52 0,06
Pycnogonida Achelia echinata Hodge, 1864 I 8 2 9 2 42 0,0074
Nymphon brevirostre Hodge, 1863 I 1 8 10 14 66 0,012
Arthropoda 0,0002
Copepoda Milne-Edwards, 1840 pas dans liste 1 2 0
Bryozoa Bryozoa II
Chordata Ascidiacea Ascidiacea Blainville, 1824 III 14 8 2 48 0,5036
Ascidiacea sp1 1 7 16
Ascidiacea sp2 1 1 4
Ascidiacea sp3 2 4
Ascidiacea sp4 1 2
Styela clava Herdman, 1881 II 2 4 0,1752
Cnidaria Anthozoa Anthozoa Ehrenberg, 1834 II 16 12 6 3 8 90 4,0128 15 10 16 7 23 142 3,095
Hydrozoa Hydrozoa Owen, 1843 I
Divers Divers Debris Divers 0,0154
Echinodermata Ophiuroidea Amphipholis squamata (Delle Chiaje, 1828) I 5 8 2 1 13 58 0,0122 17 34 56 24 46 354 0,0408
Mollusca Bivalvia Abra alba (W. Wood, 1802) III 3 1 2 3 3 24 0,0274 10 13 22 6 14 130 0,223
Aequipecten opercularis (Linnaeus, 1758) I 1 1 4 0,0488
Bivalvia 0,0038
Corbula gibba (Olivi, 1792) IV 2 1 6 0,0116 1 1 5 3 20 0,0342
Debris Bivalvia 0,0046
Kurtiella bidentata (Montagu, 1803) III 2 4 0,0016 12 2 28 0,0122
Mytilus edulis Linnaeus, 1758 III 1 1 4 0,104 11 46 49 25 48 358 16,8516
Nucula nitidosa Winckworth, 1930 I 1 2 0,0018
Parvicardium scabrum (Philippi, 1844) I 3 1 3 1 16 0,0262
Petricolaria pholadiformis (Lamarck, 1818) I 1 1 4
Sphenia binghami Turton, 1822 I 2 4 1 2
Venerupis corrugata (Gmelin, 1791) I 1 2 0,0026 9 13 65 6 43 272 6,8192
Gastropoda Crepidula fornicata (Linnaeus, 1758) III 2 1 6 0,024
Epitonium clathrus (Linnaeus, 1758) I 1 2 0,0462
Gastropoda 0,002
Mangelia costata (Pennant, 1777) II 1 2
Neogastropoda 0,004
Opisthobranchia pas dans liste 2 4 0,0206
Tritia incrassata (Strøm, 1768) II 1 2
Tritia reticulata (Linnaeus, 1758) II 1 2 0,434
Tritia sp. ponte 0,0528
Nematoda Nematoda Nematoda III 2 3 1 12 0,0038 1 4 5 2 24 0,0012
Nemertea Nemertea Nemertea A III 2 2 8 0,0018
Nemertea B III 3 1 4 1 18 0,002 15 29 22 22 18 212 0,0386
Phoronida Phoronis Wright, 1856 II 1 10 22 0,0058
Platyhelminthes Platyhelminthes Platyhelminthes Minot, 1876 II 2 4 0,0042 8 18 10 12 11 118 0,0482
Porifera Demospongiae Haliclona (Haliclona) oculata (Linnaeus, 1759) pas dans liste 0,1198 0,026
Total général 37 33 32 27 32 3578 13,168 73 79 90 68 81 29824 72,995
Fin hiver Fin été

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 79
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
ANNEXE 4 : Fiches descriptives des espèces dominantes
Phylum
Classe
Ordre
Famille
—Développement
—Durée de vie
—Mobilité
—Position / substrat
—Habitat
—Bioturbation
—Groupe trophique
—Groupe écologique
—Affinité du substrat
—Taille maximale
—Résistance
Oligochaeta
Oligochètes
Systématique
Annelida
Direct (pas de phase planctonique)
De quelques semaines à plusieurs années
Fouisseur, peu mobile
Endofaune
-
-
Clitellata
Importance pour l'environnement
Traits d'histoire de vie
Les Oligochètes sont des vers aquatiques vivant dans les sédiments à l'intérieur d'un tube muqueux. Ces organismes
sédentaires et fouisseurs se caractérisent par leur capacité d’adaptation et de colonisation d’habitats très diversifiés.
Leur sensibilité ou leur résistance à la pollution varient selon l’espèce ce qui rend leur étude intéressante pour évaluer la
qualité des sédiments. Les oligochètes sont en nombre important en fonction de la richesse en matière organique des
sédiments dans lequel ils vivent. Ce sont des détritivores qui avalent le sédiment et les divers microorganismes présents
et en extraient les matières nutritives. Ils peuvent constituer des colonies très denses, en taches. En zone tempérée, ils
peuvent littéralement tapisser certaines parties des fonds anoxiques où ils sont pratiquement les seuls à supporter des
taux d'oxygène très bas, et un haut niveau de pollution organique. Ils jouent un rôle pionnier de dépollueur primaire
dans les sédiments très pollués en matière organique, mais ils peuvent aussi bioaccumuler certains polluants et
contribuer à les (re-)disperser (bioturbation). Ces vers peuvent toutefois constituer une source importante de nourriture
pour des invertébrés prédateurs et des poissons.
Espèce opportuniste de premier ordre (V)
Vases, vases sableuses
2 à 6 cm variable selon les espèces
Variable en fonction des espèces mais généralement forte résistance à l'anoxie et à la pollution
organique
Creuse des galeries plus ou moins permanentes dans les sédiments vaseux
Diffuseur à galerie, convoyeur vers le haut (selon l'espèce)
Dépositivores de sub-surface

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 80
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Phylum
Classe
Ordre
Famille
—Développement
—Durée de vie
—Mobilité
—Position / substrat
—Habitat
—Bioturbation
—Groupe trophique
—Groupe écologique
—Affinité du substrat
—Taille maximale
—Résistance
Savigny in Lamarck, 1818
Gravette blanche
Systématique
Nephtys hombergii
20 cm
Sensible à l’instabilité du sédiment, surtout lors du recrutement, forte résistance à l'anoxie,
faible tolérance à l'enrichissement en matière organique
Importance pour l'environnement
Nephtys hombergii est un annélide prédateur capable de coloniser une grande variété d'habitats tant en termes de
substrat (même s'il préfère les sédiments envasés) qu'en termes de salinité (préfère généralement les eaux légèrement
dessalées). Cette espèce est donc rencontrée pratiquement partout dans l'Estuaire aval de la Seine. Cependant, ses
densités restent généralement limitées. Il se nourrit principalement d'autres annélides, de petits bivalves et de crustacés,
et sert de proie à des invertébrés prédateurs (crabes, némertes) ainsi qu'aux poissons plats et aux oiseaux limicoles (dans
les zones intertidales où il est fréquent).
Annelida
Polychaeta errantia
Endofaune
Animal fouisseur ne formant pas de terrier
Biodiffuseur
Prédateur
Espèce indifférente à l’enrichissement (II)
Sables vaseux, vases sableuses
Phyllodocida
Nephtidae
Traits d'histoire de vie
Cycle bentho-pélagique (larves planctoniques)
5 à 7 ans
Fouisseur, mobile
Nephtys hombergii - CSLN

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 81
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Phylum
Classe
Ordre
Famille
—Développement
—Durée de vie
—Mobilité
—Position / substrat
—Habitat
—Bioturbation
—Groupe trophique
—Groupe écologique
—Affinité du substrat
—Taille maximale
—Résistance
Systématique
Annelida
Polychaeta sedentaria
Terebellida
Ampharetidae
Melinna palmataGrube, 1870
Convoyeur de surface
dépositivore de surface
Espèce tolérante à l’excès de matière organique (III)
Vases et vases sableuses
5 cm
pas de données
Traits d'histoire de vie
Cycle bentho-pélagique (larves planctoniques, mais phase planctonique courte)
5 ans
Immobile au stabe adulte
Endofaune
Animal tubicole
Importance pour l'environnement
Melinna palmata est un Annélide qui fabrique un tube, contitué d'un mucus durci sur lequel s'agglomèrent des
particules de vases, enfoncé verticalement dans le sédiment. Cette espèce se nourrit de particules déposées à l'interface
eau-sédiment, qu'elle ramène à sa bouche grâce à ses tentacules buccaux. Peu de données sont disponibles quant à sa
prédation, mais elle possède un mécanisme de défense, lui permettant d'accumuler de fortes concentrations de cuivre
dans ses branchies, la rendant inappétente pour certaines espèces de poisson.
Cette espèce était absente de l'estuaire de la Seine avant les années 2000. Depuis, suite à l'envasement progressif de
l'estuaire, elle l'a largement colonisé. Du fait de la courte phase planctonique des larves, la capacité de dispersion de cette
espèce est faible, et son arrivée dans l'estuaire de Seine est sans doute dû à un transport anthropique.
Melinna palmata - CSLN

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 82
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Phylum
Classe
Ordre
Famille
—Développement
—Durée de vie
—Mobilité
—Position / substrat
—Habitat
—Bioturbation
—Groupe trophique
—Groupe écologique
—Affinité du substrat
—Taille maximale
—Résistance
Cardiida
Semelidae
Traits d'histoire de vie
Cycle bentho-pélagique (larves planctoniques)
1 à 2 ans
Fouisseur, peu mobile, mais capable de migrations post-larvaires
Abra alba(W. Wood, 1802)
Telline blanche
Systématique
Mollusca
Bivalvia
2,5 cm
Très sensible à l'anoxie, tolérante aux perturbations chimiques, forte résilience, capacité de
recolonisation élevée
Importance pour l'environnement
Abra alba est un bivalve filtreur et dépositivore de surface, qui préfère les sédiments assez fortement envasés dans des
zones généralement peu dessalées. Elles se développent principalement près des côtes en zone subtidale. Abra alba est
sensible aux tempêtes et au perturbations physiques mais peut rapidement recoloniser les sites d’où elle a disparue.
Organisme mixte : déposivore de surface ou suspensivore ; il ne garde que les particules les plus fines, présumées plus
riches en micro-organismes. De ce fait, ses populations sont généralement stimulées par les apports de MES. Consommé
soit en entier soit partiellement (siphons) par les poissons plats tels que les soles, les limandes, les flets et les plies, par
des petits poissons tels que les gobies, par les polychètes prédateurs tels que les Nephtys et les Phyllodocidae, par les
ophiures et par les étoiles de mer.
Endofaune
Animal fouisseur
Biodiffuseur
Suspensivore actif, dépositivore de surface
Espèce tolérante à l’excès de matière organique (III)
Tous types de sédiments mais préférence pour les vases sableuses
Abra alba - CSLN

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 83
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Phylum
Classe
Ordre
Famille
—Développement
—Durée de vie
—Mobilité
—Position / substrat
—Habitat
—Bioturbation
—Groupe trophique
—Groupe écologique
—Affinité du substrat
—Taille maximale
—Résistance
Cerastoderma edule(Linnaeus, 1758)
Coque
Systématique
Mollusca
Bivalvia
Cardiida
Cardiidae
5 cm
Traits d'histoire de vie
Cycle bentho-pélagique (larves planctoniques)
2 à 4 ans (maximum 10 ans)
Fouisseur, peu mobile, mais capable de migrations
Importance pour l'environnement
La coque est un suspensivore actif qui se nourrit de phytoplancton, de zooplancton et de matière organique :
essentiellement du microphytobenthos pour le naissain et les juvéniles et surtout de la matière organique particulaire
pour les adultes. La coque est une source de nourriture pour de nombreux organismes, surtout lorsqu’elle est jeune. Elle
est consommée au stade de naissain par les poissons plats tels que le flet, puis par les crabes et les crevettes, et surtout
une fois adulte par les limicoles (huîtriers-pies, avocettes élégantes, courlis cendrés, bécasseaux variables, bécasseaux
maubèches, barges à queue noires, …) et les oiseaux marins tels que les goélands. Elle forme des gisements parfois
denses, mais dont la localisation et la densité peuvent être très variable d'une année sur l'autre, en grande majorité au
niveau de la partie inférieure de la zone intertidale.
Endofaune
Animal fouisseur
Biodiffuseur
Suspensivore actif
Espèce tolérante à l’excès de matière organique (III)
Vases sableuses à sables fins
Assez bonne résistance aux perturbations physiques et aux pollutions métalliques, apprécie
l'enrichissement en matière organique, très sensibles aux tensio-actifs
Cerastoderma edule - CSLN

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 84
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Phylum
Classe
Ordre
Famille
—Développement
—Durée de vie
—Mobilité
—Position / substrat
—Habitat
—Bioturbation
—Groupe trophique
—Groupe écologique
—Affinité du substrat
—Taille maximale
—Résistance
Corbula gibba
(Olivi, 1792)
Corbule
Systématique
2 ans
Fouisseur, très peu mobile une fois installé
Endofaune
Animal fouisseur
Biodiffuseur
Suspensivore actif
Mollusca
Bivalvia
Myida
Corbulidae
Traits d'histoire de vie
Cycle bentho-pélagique (larves planctoniques)
Espèce opportuniste de second ordre (IV)
Vases sableuses à graviers envasés
1,5 cm
Très forte résistance aux perturbations chimiques, à l'enrichissement en matière organique et à
l'anoxie, capable de recoloniser très rapidement des zones totalement défaunées.
Importance pour l'environnement
La corbule est un suspensivore actif qui se nourrit de phytoplancton, de zooplancton et de matière organique et est
capable de développer des populations très denses dans des zones très perturbées (notamment fortement hypoxiques).
Du fait de sa capacité de colonisation et de survie dans ces environnements hautement perturbés, la corbule compte peu
de prédateurs dans de tels environnements. Elle est toutefois consommées par divers prédateurs comme les étoiles de
mer, les crabes, les gastéropodes prédateurs, les poissons plats et les oiseaux dans les zones moins perturbées. Il semble
en général que se soit la compétition au moment des recrutements larvaires qui soit le principal facteur de régulation de
cette espèce.
Corbula gibba - CSLN

CSLN – RAPPORT DE SYNTHESE DES DEUX CAMPAGNES
2016 85
PROJET P38 ETUDE DE L’ACCES FLUVIAL DIRECT A PORT 2000 INVENTAIRE DE LA MACROFAUNE BENTHIQUE
Phylum
Classe
Ordre
Famille
—Développement
—Durée de vie
—Mobilité
—Position / substrat
—Habitat
—Bioturbation
—Groupe trophique
—Groupe écologique
—Affinité du substrat
—Taille maximale
—Résistance
Nucula nitidosa
Winckworth, 1930
Nucule
Systématique
Mollusca
Bivalvia
Endofaune
Animal fouisseur
Biodiffuseur
Dépositivore de surface, dépositivore de sub-surface
Espèce très sensible à l’enrichissement en matière organique (I)
Vases à sables fins
Nuculida
Nuculidae
Traits d'histoire de vie
Cycle bentho-pélagique (larves planctoniques, mais phase planctonique très courte)
7 à 10 ans
Fouisseur, peu mobile
1,3 cm
Très bonne résistance à l'anoxie, relativement tolérante aux perturbations physiques, peu de
données pour les perturbations chimiques
Importance pour l'environnement
La nucule se nourrit principalement en récoltant des microroganismes ou de la matière organique présente en surface
des sédiments ou dans ceux-ci. Elle est capable de filtrer l'eau mais ce mode d'alimentation reste très marginal. De petite
taille et généralement bien moins abondante que d'autres bivalves, elle ne représente pas une ressource alimentaire
majeure. Elle est toutefois consomée par des poissons plats comme la plie et la limande. La phase larvaire planctonnique
est très courte chez cette espèce. De ce fait, sa capacité de disperssion est relativement limitée. Du fait du grand nombres
d'oeufs émis, elle est donc capable de recoloniser rapidement des zones dépeuplées à petite échelle mais si une large
zone géographique est touchée par les perturbations, le temps de recolonisation pourra être très long, en fonction des
courants. Du fait de sa longévité, les populations sont généralement stables sur des périodes d'une dizaine d'années,
mais peuvent chuter de façon importante quand les vieux individus arrivent en fin de vie.
Nucula nitidosa - CSLN





























