Embed Size (px)
Citation preview

É
Q
i
MNNLADI
gtlipcsssa
itplps2dd
dlllsddtn
a
1h
Revue du rhumatisme 80 (2013) 102–105
Disponible en ligne sur
www.sciencedirect.com
ditorial
uand les neutrophiles jettent leurs filets�
n f o a r t i c l e
ots clés :eutrophileétoseupusuto-immunitééfense anti-infectieuse
mmunité innée
Les polynucléaires neutrophiles (PN) sont les leucocytes san-uins les plus nombreux chez l’homme. Leur rôle premier a long-emps été considéré comme étant celui de protéger l’hôte contrees micro-organismes. Ils sont donc essentiels dans la réponse anti-nfectieuse. Cependant, depuis quelques années, de nombreusesreuves expérimentales ont documenté leur implication commeellules effectrices et régulatrices des réponses immunitaires noneulement innées, mais aussi adaptatives. C’est à ce titre que l’onait maintenant que les PN jouent un rôle fondamental dans la phy-iopathologie de maladies inflammatoires chroniques, notammentuto-immunes, ainsi que dans le cancer [1].
Les PN sont les premières cellules qui migrent vers un sitenflammatoire, infectieux ou non. Ils sont doués de capacités fonc-ionnelles nombreuses, grâce à un arsenal d’outils originaux. Oneut citer, en particulier, la synthèse de protéases intragranu-
aires préformées (élastase, myéloperoxidase, lactoferrine. . .), laroduction de formes réactives de l’oxygène et de l’azote, et laynthèse d’un grand nombre de cytokines (IL-8, TNF, IL-1. . .). En004, une nouvelle propriété fonctionnelle a été mise en évi-ence, la nétose, qui a été étudiée de manière intensive depuis saécouverte [2].
La nétose est une nouvelle forme de mort cellulaire, différentee l’apoptose et de la nécrose. Au cours de ce phénomène, les PN
ibèrent des filaments d’ADN extracellulaires (neutrophil extracellu-ar traps [NETs]) dont la propriété principale semble être de piégeres micro-organismes. Des phénomènes morphologiques originauxont visibles au cours de ce phénomène. En premier, la décon-ensation de la chromatine nucléaire donne lieu à l’observation
e noyaux unilobés, avec perte de l’aspect caractéristique mul-ilobé de ces cellules. Puis, la désintégration des membranesucléaires, mitochondriales et granulaires, permet le mélange deDOI de l’article original : http://dx.doi.org/10.1016/j.jbspin.2013.01.006.� Ne pas utiliser pour citation, la référence franc aise de cet article, mais la référencenglaise de Joint Bone Spine avec le DOI ci-dessus.
169-8330/$ – see front matter © 2012 Société Française de Rhumatologie. Publié par Elsttp://dx.doi.org/10.1016/j.rhum.2012.11.007
leurs nombreux constituants. La phase finale de la nétose consisteen l’expulsion extracellulaire de filaments de chromatine parse-més de protéines issues des différents compartiments cellulaires(noyau, cytoplasme, granulations. . .).
La nétose dépend de l’explosion oxydative ; en effet, la NADPH-oxydase-2 (NOX2) et les petites protéines, rho GTPase Rac2,semblent nécessaires à son induction, même si certains travauxrécents suggèrent la possibilité d’induire une nétose en l’absencede NOX2. La citrullination des histones par la peptidylargininedeiminase (PAD4) est la signature biologique de la nétose et estindispensable au désenroulement de la chromatine [3,4]. La trans-location nucléaire de myéloperoxydase (MPO) et d’élastase (ELA),provenant des granulations, est également essentielle afin de per-mettre les modifications enzymatiques à l’origine de l’extrusiond’ADN hors de la cellule [5]. Récemment, des observations dyna-miques in vivo suggèrent que les PN porteurs de NETs conserventleurs capacités migratoires, et qu’ils peuvent engager certainsrécepteurs pour les fragments Fc des immunoglobulines (Fc�RIIA)afin d’internaliser les complexe immuns [6,7]. Ce phénomène denétose n’est pas l’apanage des neutrophiles. En effet, d’autres typescellulaires peuvent également produire des NETs, comme les éosi-nophiles ou les basophiles et mastocytes.
Les protéines associées aux NETs sont très variées (Tableau 1).Une étude protéomique a identifié 24 protéines en 2009. Les his-tones, protéines nucléaires, correspondent à plus de 70 % desprotéines et sont associées à l’ADN. L’élastase, la calprotectine et lalactoferrine (LF) constituent environ 5 % des protéines totales [8].De plus, durant ces deux dernières années, notre groupe et d’autreséquipes ont identifié de nouveaux composants importants associésaux NETs comme la NOX2, la pentraxine 3, la �2 intégrine, la LL-37,l’annexine A1 ou la serpine B1 [9,10]. Il est cependant important denoter que le répertoire des protéines associées aux NETs n’est pasextensivement connu et que la découverte de nouvelles protéinesassociées à leur surface pourrait fournir de nouvelles pistes fonc-tionnelles pour ces NETs. De plus, la composition des NETs seraitdépendante de l’agent stimulant inducteur de nétose in vitro etprobablement in vivo [9,11]. Enfin, des travaux préliminaires denotre équipe (résultats non publiés) suggèrent une grande variabi-lité interindividuelle dans la composition des NETs, en réponse àun stimulus donné.
À ce jour, la fonction physiologique la plus importante des NETssemble être leur capacité à piéger et détruire les micro-organismes.
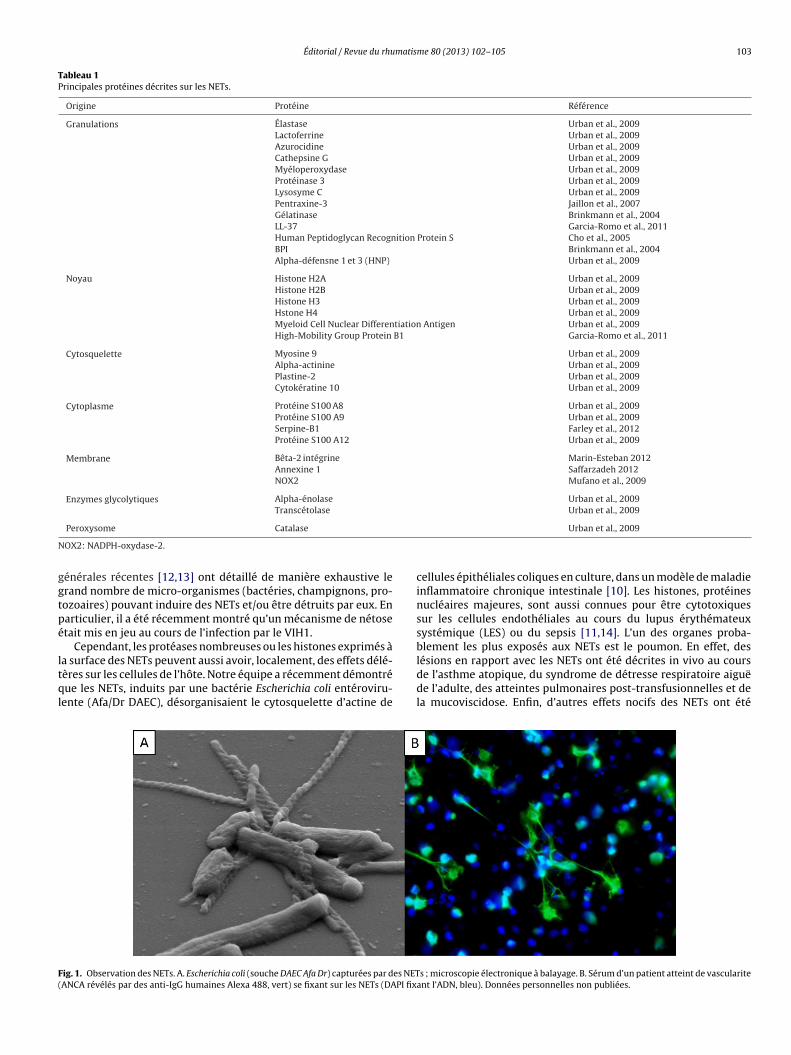
Les filaments de chromatine constituent un filet qui prévient ladissémination des agents infectieux (Fig. 1A), mais qui permetaussi de les tuer grâce à la forte concentration locale de compo-sants antimicrobiens, en particulier, des protéases. Deux revuesevier Masson SAS. Tous droits réservés.
Éditorial / Revue du rhumatisme 80 (2013) 102–105 103
Tableau 1Principales protéines décrites sur les NETs.
Origine Protéine Référence
Granulations Élastase Urban et al., 2009Lactoferrine Urban et al., 2009Azurocidine Urban et al., 2009Cathepsine G Urban et al., 2009Myéloperoxydase Urban et al., 2009Protéinase 3 Urban et al., 2009Lysosyme C Urban et al., 2009Pentraxine-3 Jaillon et al., 2007Gélatinase Brinkmann et al., 2004LL-37 Garcia-Romo et al., 2011Human Peptidoglycan Recognition Protein S Cho et al., 2005BPI Brinkmann et al., 2004Alpha-défensne 1 et 3 (HNP) Urban et al., 2009
Noyau Histone H2A Urban et al., 2009Histone H2B Urban et al., 2009Histone H3 Urban et al., 2009Hstone H4 Urban et al., 2009Myeloid Cell Nuclear Differentiation Antigen Urban et al., 2009High-Mobility Group Protein B1 Garcia-Romo et al., 2011
Cytosquelette Myosine 9 Urban et al., 2009Alpha-actinine Urban et al., 2009Plastine-2 Urban et al., 2009Cytokératine 10 Urban et al., 2009
Cytoplasme Protéine S100 A8 Urban et al., 2009Protéine S100 A9 Urban et al., 2009Serpine-B1 Farley et al., 2012Protéine S100 A12 Urban et al., 2009
Membrane Bêta-2 intégrine Marin-Esteban 2012Annexine 1 Saffarzadeh 2012NOX2 Mufano et al., 2009
Enzymes glycolytiques Alpha-énolase Urban et al., 2009Transcétolase Urban et al., 2009
N
ggtpé
ltql
F(
Peroxysome Catalase
OX2: NADPH-oxydase-2.
énérales récentes [12,13] ont détaillé de manière exhaustive lerand nombre de micro-organismes (bactéries, champignons, pro-ozoaires) pouvant induire des NETs et/ou être détruits par eux. Enarticulier, il a été récemment montré qu’un mécanisme de nétosetait mis en jeu au cours de l’infection par le VIH1.
Cependant, les protéases nombreuses ou les histones exprimés à
a surface des NETs peuvent aussi avoir, localement, des effets délé-ères sur les cellules de l’hôte. Notre équipe a récemment démontréue les NETs, induits par une bactérie Escherichia coli entéroviru-ente (Afa/Dr DAEC), désorganisaient le cytosquelette d’actine de
ig. 1. Observation des NETs. A. Escherichia coli (souche DAEC Afa Dr) capturées par des NETANCA révélés par des anti-IgG humaines Alexa 488, vert) se fixant sur les NETs (DAPI fix
Urban et al., 2009
cellules épithéliales coliques en culture, dans un modèle de maladieinflammatoire chronique intestinale [10]. Les histones, protéinesnucléaires majeures, sont aussi connues pour être cytotoxiquessur les cellules endothéliales au cours du lupus érythémateuxsystémique (LES) ou du sepsis [11,14]. L’un des organes proba-blement les plus exposés aux NETs est le poumon. En effet, des
lésions en rapport avec les NETs ont été décrites in vivo au coursde l’asthme atopique, du syndrome de détresse respiratoire aiguëde l’adulte, des atteintes pulmonaires post-transfusionnelles et dela mucoviscidose. Enfin, d’autres effets nocifs des NETs ont étés ; microscopie électronique à balayage. B. Sérum d’un patient atteint de vasculariteant l’ADN, bleu). Données personnelles non publiées.

104 Éditorial / Revue du rhumatisme 80 (2013) 102–105
, à la
rlpl
avedcpaapmto3cspdlutDdLlànaaicaCdP
Fig. 2. Synthèse des principales données et hypothèse sur le rôle des NETS
apportés via leur capacité à activer la coagulation, en raison de’expression de facteur tissulaire à leur surface, et par le fait que leslaquettes activent la nétose ; les NETs sont donc impliqués dans
’athérosclérose, les thromboses, mais également dans le sepsis.L’implication des NETs dans la physiopathologie de maladies
uto-immunes et la rupture de tolérance constituent un nou-eau sujet d’investigation, à l’origine de multiples hypothèses. Enffet, la nétose est caractérisée par l’exposition de composantsu Soi granulaires, cytoplasmiques et nucléaires, accessibles auxellules du système immunitaire. L’ADN et les histones, d’uneart, la myéloperoxydase ou la protéinase 3, d’autre part, sontutant d’auto-antigènes impliqués dans la genèse de maladiesuto-immunes bien caractérisées, dont les chefs de file sont res-ectivement le LES et les vascularites à ANCA. L’antigénicité de cesolécules est susceptible d’être altérée par des modifications post-
ranscriptionnelles comme l’oxydation ou la citrullination [4,5,15],u par des associations moléculaires. Par exemple, le peptide LL-7 permet à l’ADN des NETs d’être immunogène et d’activer lesellules dendritiques plasmacytoïdes via le TLR9, aboutissant à laécrétion d’interférons de type 1 [9]. Cette observation résulte delusieurs travaux réalisés ces deux dernières années, qui ont permise démontrer que les NETs étaient impliqués dans la physiopatho-
ogie du LES [9,16]. Les PN de patients atteints de LES possèdentne forte capacité à induire des NETs ex vivo ; ces NETs sont résis-ants à la dégradation, en raison de la présence d’inhibiteurs desNases, comme le C1q par exemple. La Fig. 2 résume l’implicatione la nétose au cours du LES. Si le nombre de travaux sur leES est croissant, la démonstration la plus claire de lien entrees NETs et l’auto-immunité a été faite au cours des vascularites
ANCA. En effet, le lien entre la myéloperoxidase et la protéi-ase 3 présents sur les NETs et le développement de la maladie
été établi comme l’illustre notamment la fixation spécifique desnticorps anti-PR3 des patients sur les NETs (Fig. 1B) [17]. Enfin,l semble licite d’émettre l’hypothèse selon laquelle les protéinesitrullinées à la surface des NETs peuvent constituer des néo-
ntigènes à l’origine de la production d’auto-anticorps spécifiques.ela est suggéré par des travaux récents au cours du syndromee Felty [18]. En revanche, l’invalidation du gène qui code pour laAD4 n’empêche pas le développement des lésions dans un modèlefois dans la défense anti-infectieuse et dans la physiopathologie du lupus.
murin de polyarthrite spontanée [19]. Enfin, au cours de la goutte,les neutrophiles sanguins et synoviaux produisent des NETs ex vivoen réponse aux cristaux d’urate de sodium, dans un mécanismedépendant de l’IL-1� et de l’autophagie [20].
Depuis leur première description en 2004, de nombreux scien-tifiques se sont laissés prendre dans les filets de ces NETs. Toutd’abord les microbiologistes, qui ont pu démontrer leur capacitéparticulière à immobiliser et tuer les micro-organismes, puis lesimmunologistes en décrivant les mécanismes intracellulaires dela nétose, mais aussi depuis peu les interactions des composantsdes NETs avec les cellules de l’immunité innée et de l’immunitéadaptative. L’implication de ce phénomène semble particulière-ment vraisemblable au cours du processus d’auto-immunité, maiselle fait actuellement l’objet d’investigations dans d’autres cadresnosologiques, aigus ou chroniques, au cours desquels le PN a déjàété identifié comme un acteur important, sinon majoritaire. Enperspective, la modulation pharmacologique de la nétose pourraitouvrir de nouveaux horizons thérapeutiques, dans le vaste spectredes maladies inflammatoires chroniques.
Déclaration d’intérêts
Les auteurs déclarent ne pas avoir de conflits d’intérêts en rela-tion avec cet article.
Références
[1] Mantovani A, Cassatella MA, Costantini C, et al. Neutrophils in the activationand regulation of innate and adaptive immunity. Nat Rev 2011;11:519–31.
[2] Brinkmann V, Reichard U, Goosmann C, et al. Neutrophil extracellular traps killbacteria. Science 2004;303:1532–5.
[3] Li P, Li M, Lindberg MR, et al. PAD4 is essential for antibacterial innate immunitymediated by neutrophil extracellular traps. J Exp Med 2010;207:1853–62.
[4] Neeli I, Radic M. Knotting the NETs: Analyzing histone modifications in neu-trophil extracellular traps. Arthritis Res Ther 2012;14:115.
[5] Papayannopoulos V, Metzler KD, Hakkim A, et al. Neutrophil elastase and mye-
loperoxidase regulate the formation of neutrophil extracellular traps. J Cell Biol2010;191:677–91.[6] Chen K, Nishi H, Travers R, et al. Endocytosis of soluble immune complexesleads to their clearance by FcgammaRIIIB but induces neutrophil extracellulartraps via FcgammaRIIA in vivo. Blood 2012.

matis
[
[
[
[
[
[
[
[
[
[
[
Éditorial / Revue du rhu
[7] Yipp BG, Petri B, Salina D, et al. Infection-induced NETosis is a dynamic processinvolving neutrophil multitasking in vivo. Nat Med 2012.
[8] Urban CF, Ermert D, Schmid M, et al. Neutrophil extracellular traps contain cal-protectin, a cytosolic protein complex involved in host defense against Candidaalbicans. PLoS Pathog 2009;5:e1000639.
[9] Garcia-Romo GS, Caielli S, Vega B, et al. Netting neutrophils are major inducersof type I IFN production in pediatric systemic lupus erythematosus. Sci TranslMed 2011;3:73ra20.
10] Marin-Esteban V, Turbica I, Dufour G, et al. Afa/Dr diffusely adhering Escheri-chia coli strain C1845 induces neutrophil extracellular traps that kill bacteriaand damage human enterocyte-like cells. Infect Immun 2012;80:1891–9.
11] Villanueva E, Yalavarthi S, Berthier CC, et al. Netting neutrophils induce endo-thelial damage, infiltrate tissues, and expose immunostimulatory molecules insystemic lupus erythematosus. J Immunol 2012;187:538–52.
12] Brinkmann V, Zychlinsky A. Neutrophil extracellular traps: Is immunity thesecond function of chromatin ? J Cell Biol 2012;198:773–83.
13] Kaplan MJ, Radic M. Neutrophil extracellular traps: double-edged swords ofinnate immunity. J Immunol 2012;189:2689–95.
14] Xu J, Zhang X, Pelayo R, et al. Extracellular histones are major mediators ofdeath in sepsis. Nat Med 2009;15:1318–21.
15] Liu CL, Tangsombatvisit S, Rosenberg JM, et al. Specific post-translationalhistone modifications of neutrophil extracellular traps as immunogens andpotential targets of lupus autoantibodies. Arthritis Res Ther 2012;14:R25.
16] Lande R, Ganguly D, Facchinetti V, et al. Neutrophils activate plasmacytoiddendritic cells by releasing self-DNA-peptide complexes in systemic lupus ery-thematosus. Sci Transl Med 2011;3:ra19.
17] Sangaletti S, Tripodo C, Chiodoni C, et al. Neutrophil extracellular traps mediate
transfer of cytoplasmic neutrophil antigens to myeloid dendritic cells towardsANCA induction and associated autoimmunity. Blood 2012.18] Dwivedi N, Upadhyay J, Neeli I, et al. Felty’s syndrome autoantibodies bindto deiminated histones and neutrophil extracellular chromatin traps. ArthritisRheum 2012;64:982–92.
me 80 (2013) 102–105 105
19] Rohrbach AS, Hemmers S, Arandjelovic S, et al. PAD4 is not essential for diseasein the K/BxN murine autoantibody-mediated model of arthritis. Arthritis ResTher 2012;14:R104.
20] Mitroulis I, Kambas K, Chrysanthopoulou A, et al. Neutrophil extracellular trapformation is associated with IL-1� and autophagy-related signaling in gout.PLoS One 2011;6:e29318.
Luc de Chaisemartin a,b
Gilles Hayem c
Sylvie Chollet-Martin a,∗,b
a Inserm, UMR-S 996, faculté de pharmacie,universud, 5, rue J.-B.-Clément, 92296
Chatenay-Malabry cedex, Franceb Unité d’immunologie « auto-immunité et
hypersensibilités », hôpital Bichat, groupe hospitalierParis-Nord–Val-de-Seine, AP–HP, 75018 Paris, France
c Service de rhumatologie, hôpital Bichat, groupehospitalier Paris-Nord–Val-de-Seine, AP–HP, 75018
Paris, France
∗ Auteur correspondant.Adresse e-mail : [email protected]
(S. Chollet-Martin)
Accepté le 21 novembre 2012
Disponible sur Internet le 23 janvier 2013