Embed Size (px)
Citation preview

J. MoL Biol. (1968) 35, 311-331
Quelques Propri&~s des RNA ~ Marquage Rapide Extraits de Cellules Foliaires
CLAUDE GIGOT, GABRIEL PH,,.rppS ET L~O~ ~raTH
Inatitut de Botanique, Universit~ de Strasbourg, France et Centre de Rechervhes ~ud~aires, ApTlication~ biologiques
Stra~bourg-Cronenbourg, France
(Received 9 October 1967, and in revised form 22 March 1968)
82p incorporation was followed in Chinese cabbage leaves (Brassica chiner~is var. Po tsa'~') isolated from plants. After pulses of different lengths, the total RNA of leaves was extracted and run on a sucrose gradient.
I t has been possible to isolate different rapidly labelled RI~A's with sedimenta- tion coefficients increasing from 5 to 35 s while the pulse length increased from 7 to 25 minutes. These heterogeneous RNA's were divided into four major classes: 4 to 10 s; 15 s; 23 s; 35 s.
The following points were established. (1) Base composition of rapidly labelled RNA's ~s very different from those of
ribosomal and soluble RNA's. The base composition varies also with the size of the molecule.
(2) The participation of rapidly labelled RNA's in polysome building and their ability to stimulate ,,mlno acid incorporation into proteins within a cell-free system are in agreement with messenger function for these RNA's.
(3) The meaning of the increase in size of rapidly labelled RlqA with the length of 32p pulse is discussed.
(4) 18 hours incubation with high concentration of actinomycin D intfibits ~he synthesis both of ribosomal and soluble RNA's to the extent of 85 and 75%, respectively. After the same treatment, the decrease of a2p incorporation into rapidly labelled RNA is abou~ 60%. At the same time, protein synthesis is not affected by actinomycin D treatment. This result implies a very high stability of leaf mRNA.
(5) Prelhninary studies on the effect of light on rapidly labelled RNA syn- thesis illustrates the importance of this factor.
1. I n t r o d u c t i o n
Depuis la d6couverte et l '~laboration do la th6orie du R N A messagcr chez Esc/~r/eh/¢ coli (Jacob et Monod, 1961), un nombre impor tan t de t r avaux ont p o r ~ sur ce type de R N A dans les cellules bact6riennes et an~males. Les r6sultats obtenus ont permis de pr6ciser le m6tabolisme et diff6rents earact~res de ces RNA.
Chez les plantes, un certain nombre d 'observat ions ont conduit k penser que les m6canismes fondamentaux 6taient les m~mes que chez les bact~ries et les aulmaux. Cette affirmation s 'appuie sur des f r avaux effec~u6s sur de jeunes plan~ules (Loening, 1965; Ingle, K e y et Holm, 1965; Chroboczek et Cherry, 1966; Lonberg-Holm, 1967) ou sur des graines en voie de germination (Waters et Dure, 1966). P a r centre, aueune 6rude syst6matique n ' a port6 sur des organes diff6renci6s k haute ac~ivlt~ m6tabolique (notamment en ce qui concerne la synth&se des prot~ines), tels ClUe les feuflles.
311

~12 C. GIGOT, G. P H I L I P P S ET L. H I R T H
L'in%6r~t des feuilles r6side en plus dans le fair que les ceUules foliaires peuvent h6berger des virus et que l'influence de la multiplication de ceux-ci sur la syn~h6se des RNA des cellules-hStes n 'est pas eonnue. C'est le cas notamment de B r a ~ # ~ chi~s~s qui peut h6berger un virus ~ RNA tel que le virus de la mosaique jaune du navet. Cependant, avant d'entreprendre une telle ~tude, il ~tait n6cessaire de carac- t~riser, au sours d 'une premi6re 4tape, les diff6rents RNA des feuiUes saines de ~rcus~ca chinen~i~.
Co m~moire pr~sente la raise en ~vidence, la earaet~risation et l '6tude de eergaines propri~t~s des RlgA messagers dans co materiel, sans chercher ~ distinguer Ieur loca/isation eytoplasmique, nucl4aire ou chloroplastique.
Les experiences rappor~6es ont 6t6 r6alis6es ~ la lnmi6re dent le r61e ne sera en- visag~ que tr6s bri6vement.
2. Materiel et Techniques
Les planfes utflis6es sent des ohoux de Chine (Bramric~ chir~r~gs var. Po Tsal) cultiv6es en serre climatis6e ~ une %emp6rature moyenne de 20°C, un degr~ hygrom6trique de 60 ~. 70% et un 4clairement de 14 hr par jour avec une intensit~ ]umlneuse de 2000 lux. Lors- qu'elles sent employees, les plantes sent gg6es de 5 k 7 :semaines, les feuilles ayant entre 15 et 20 cm de long.
(a) Teohn~ & ~rqua4e Apr6s une exposition pr~a/able d'au moins 5 hr/~ la 1-mi6ro, les feuilles sent pr61ev~es en
sectdonnant la base du p~tlole. Ce p6tiole est plong4 dans une solution mln~ra/e de Knop dilute au 1/3 o5 le phosphate a 6t6 remplac6 par de l'acide phosphorique marqu4 sur le phosphore. La quantit~ de s2p est de 1 mc/ml. Entre le moment du d6tachement de la plante et le d~but du marquage, il s'~coule entre 7 et 10 min. Pendant la dur4e du marquage les feuilles sent toujours maintenues/~ la ]nmi6re. On a pu v~ri~er par l'utilisation de feuiUes sf~rilis~es par l'hypoehlorite d'une part, et par le d6nombrement de bact~ries ou de champignons d'autre part, que notre technique de marquage excluait route contamin- ation par du RNA d'origine bacb6rienne ou fongique. Aprgs le marquage, la partie des p6tioles ayant plong6 dans le 82p est couple et les RNA totaux des feuilles extraits.
(b) EzZr~£on des RNA Los feuilles sent broy6es au moyen d'un homog~n~iseur Vir~is, par trois broyages
successffs do 20 sec chacun, ~ vitesse ma.Timum dans ~ tampon acetate de sodium 0,01 x,- Na6~ 0,1 ~ k pH 5,0, en presence de bentonite k raison de 10 mg/g de maf~riel frais. Pendant la duroc du broyage, la fiole plonge dans la glace, afin d'6viter un 6chauffement trop important du au frottement des couteaux de l'homog6n~iseur. A l'homog4nat on ajoute du lauryl sulfate de sodium de mani6re ~ obtenir une concentration finale de 0,5/~ 1%, puis un re1. de phenol fralchement distill~ et satur~ d'eau. Le m~lange est agit4 au froid pendant 9.0 rnin~ puis centrifug6 k 3000 g pendant 20 mln. Le surnageant aqueux est reeuciUi et subit un deuxi6me traitement ph6nolique et une autre centrifugation. A l'inter- phase et k la phase ph~noliquo r~sultant de la premi6re centrifugation, on ajoute 1/2 vol. de f~anpon, on agite 15 mln et on centrifuge. Les 2 phases aqueuses r~unios sent a/ors trait~es 3 fois par 2 voL d'6ther pour ~limlner le phenol, puis l'6ther ost chass~ par un courant d'azote. La preparation est so-mi~e a/ors ~ une dialyse d'une nuit centre 21. du tampon d'extraction. Toutes ces manipulations so font au froid. Apr6s la dialyse, le RIgA es% pr~cipif~ par 2 vol. d'a/cool ~ 95 deg pendant 4hr/~ --20~C. Apr6s centrifugation, le pr~cipit6 est rem;- on suspension dans un tampon Tris-HC10,01 ,~ (pH 7,4) contenant du magn4sium ~ la concentration de 0,001 Met t rai~ par la Dlgase 2 ~g/mL pendant 30 min
0°C. Dans cer~ina cas, ce traif~ment a ~t4 supprim~ pour d~celer la pr6senee ~ventuelle dans nos preparations de DIgA. Apr6s repr~cipitation par l'a/cool et deux autres dissolu- tions, les preparations de RIgA on~ 4f~ ana/ys~s par deux techniques diff4rentes.

R N A DE C E L L U L E S F O L I A I R E S 313
(o) ~q$paration des d~ff~ren~s RiVA Fr&snt~ darts lss l~r~araz /~ Los deux techniques utflis~es sent la eentrifugation en gradient de densit~ de saceharose
et la chromatographie sur colonnes d'alburnlne m~thyl~e. Los gradients de densif~ de saccharose ont ~t~ prepares dans le m&me tampon clue celui
qui a servi R l 'extraction des RNA. Leurs centrifugation et fractionnement ont ~t~ r~al- isis dans les conditions eouramment utilis~es pour l '~nalyse et |a s~paration de R N A contenns darts un m~lange. La radioactivit~ de ehaque fraction a ~t~ d~termin~e apr~s precipitation par le TCAt 5% et filtration sur filtres Millipore.
La mSthylation de la s~rum-alb~mlne bovine a St~ faite d'apr~s la technique originale de Mandell et Hershey (1960). Les colonnes ont ~t~ prSpar~es solon la technique simplifi~e indiqu~e par Monier, Naono, l=[ayes, Hayes et Gros (1962). La colonne, charg~e de 5 ~. 10 mg de RNA, est $1u~e par un gradient lin~aire de concentration en NaC1 de 0,2 ~ 0,9 M. Des fractions de 5 xnl. sent recueillies, dent on d~termlne la densit~ optique ~ 260 mp et la radioactivitY. Avant l 'hydrolyse, los R N A prSeipit~s par l'alcool sent laves et purifi~s par une technique voisine de cello d 'Ogur et Rosen (1950).
Apr&s une hydrolyse alcaline, effeetu~e solon la technique classique, le m~lange de nucl~otides neutralis~ par l 'acide perehlorique est analys~ sur une colonne de Dowex 1 X 8 pr~par$ sous la forme formiate, et ~lu~ par un gradient de concentration d'acide formique. La radioactivit~ de chaclue nucl~otide permet de calculer Ia composition en bases du R N A StudiO. Chaque nucl$otide concentr~ a ~t~ analys~ par chromatographie bidimensionnelle sur touche mince de cellulose, solon une technique originale mise au point dans ce laboratoire (Kumar, sous presse). Les autoradiogrammes de chacun de cos nucl~otides ont d~montr~ leur puretg, saul dans lecas des RI~TA de faible masse (4 R 10 s), oh est apparu un contaminant radioactif ~lu~ avec la gu~nlne, et dent fl sera reparl~ plus loin.
(d) Pr~Ioar~on des :polysome~ Los polysomes ont ~t6 extraits selon une ~ l m i q u e voisine de cello d~crite par BayIey
(1964). Apr~s marquage, los feuilles d~eoup~es en fines lamelles ont ~t~ broy~es grossi&rement
au moyen d 'un Ultraturax, puis ~ l 'aide d 'un homog~n~iseur Potter, en presence d 'un tampon Tris-HCl 0,01 M (pH 7,4) contenant du saccharose 0,25 ~a, du KC10,015 ~ et du MgCI~ 0,02 ~. L'homog~nat, filtr~ sur gaze, est centrifug~ deux lois 10 mln ~ 1000 g e t 7000g. Los culots sent je t& et le surnageant subit encore une eentrifugatlon de 105.000g pendant MgCI~ 2 hr. Le cuIot est remis en suspension dans un tampon Tris-HC~ 0,05 ~ (pH 7,4) et eontenant du KCI 0,015 Met du M g ~ 0,005 M. C'est une telle suspension qui sera een- trifug~e dans un gradient de densit~ de saecharose de 5 ~ 20% pendant 2 hr 30 ~ 24.000 tours/rain.
(e) Evalua~on de la lrrot~.osynth~se Apr~s marquage des feuilles pendant 30 mln par la proline [14C], eelles-ci sent eongel~es --20°C, puis broy~es au merrier daus t in,peti t volume de tampon phosphate 0,01 M
eentrifugatlon lento, Iv oulot est jet~ et le surnageant recentrifug~ (pH 7,4). Apr~s une " ~ " pendant 30 rnln ~ 17.000 g. On pr~cipite ce surnageant par 3 vol. d'ac~tene ou d'alcool. Los pr~eipit~s obtenus sent laves deux lois par l 'ac~tone 75%, puis dissous dans 2,5 ml. d 'acide formique 98%. De l 'eau est ajout~e jusqu'/~ dissolution compl&te du pr~cipit~, puis une fraction de 0,3 ml. de la solution est d~pos~e sur une planchette et la radioactivit~ d~termln~e. 2 ml. sent mln~ralis~s en presence d'acide sulfurique concentr6 et d 'un m~lange de sulfate de potassium et d'ac~tate mercurique, puis le contenu en azote est dos6 par la microm~thode de KjeldahL A partir de cos donn6es, on calcule la radioactivit~ sp~eifique des prot~ines extraites. La m~thode d~crite est simple. On a pu en effet montrer clue l 'activit~ sp~cifique des prot~ines d~termin~e eomme il vient d'Stre dit, est analogue /~ cello estim~e apr~s hydrolyse acide (24 hr / t 110°C dans HC161~) du pr~oipit~ et isolement sur Permuti te 50 des acides amines.
t Abbr6viations: VMJ~, virus de la mos~que jaune du hayer; TCA, acide trichlorac6tique.
314 C. GIGOT, G. P H I L I P P S ET L. H I R T H
(f) Extrui~ ace~lu~aire d'Escherichia coli L'apti~ude des diffdren~es fractions de 1%1~A k s~imuler la pro~osyn~h~se/~ vitro,
~ test~e par la ~echnlque de Nirenberg e~ Matthaei (1961), modifi~e par Willson et Gros (1964).
3 . R~stfltats
(a) Extraction des RI~A
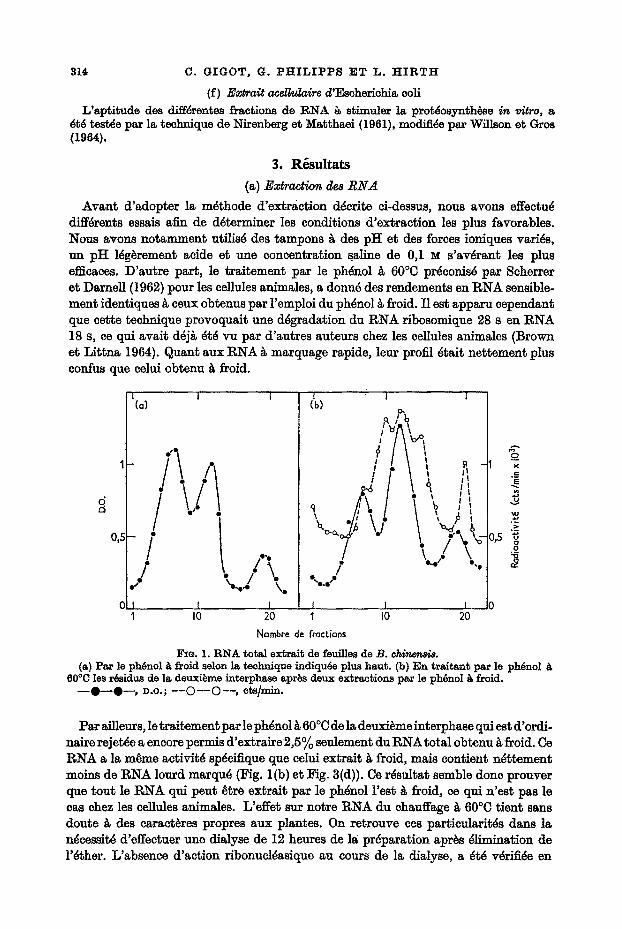
Avant d 'adopter la m~thode d'extr~ction ddcrite ci-dessus, nous avons effectu~ diff4rents essais ~fin de d4terminer les conditions d 'extraction les plus favorables. Nous avons notamment utilis4 des tampons ~ des p H et des forces ioniques vari4s, un p H 14g~rement acide et une concentration saline de 0,1 ~ s 'av~rant les plus efficaces. D'autre part , le t rai tement par le phenol ~ 60°0 pr~conis~ par Scherrer et Darnell (1962) pour les cellules aulmales, a donn4 des rendements en RNA sensible- ment identiques k ceux obtenus par remploi du ph4nol k froid. I1 est apparu cependant que cette technique provoquait une d4gradation du RNA ribosomique 28 s e n RNA 18 s, ce qui avait d~j~ 4td vu par d 'autres au~eurs chez les ceUules animales (Brown et Li t tna 1964). Quant aux RNA k marquage rapide, leur profil 4tait net tement plus confus que celui obtenu k froid.
d c~
I Ca)
'1-
0 ,5- f
/ Ol
1
I t
/\ :
~r I I I (b)
, i ~ o
o
, . , L \ , , a :
. / k., \, ",,.o." \ .
I 20
I I I I I0 I I0 20
x ._=
),5 .o
Nombre de fractions
FIG. 1. RNA total ex~rait de feuilles de B. c /~ne~. (a) Par le ph&nol ~ froid selon la technique indiqu6e plus haut. (b) En traitan~ par ]e ph6nol
60°C les r6sidus de la deuxi~me interphase apr~s deux extractions par le ph6nol ~ froid. --O--O--, p.o.; --0--0--, o~s/mm.
Par ailleurs, le t rai tement par le phenol ~ 60°C de 18, deuxi~me interphase qui est d ' ordi- naire rejet~e a encore permis d'extraire 2 , 5 ~ seulement du RNAtota l obtenu ~ froid. Ce RNA a la m~me activit~ sp~cifique que celui extrait ~ froid, mais contient n~t~ement moins de RNA lourd marqu~ (Fig. l(b) et Fig. 3(d)). Co r~sultat semble donc prouver que tou t le RNA qui peut ~tre extrai t par le phenol r e s t k froid, co qui n 'est pas le cas chez les cellules animales. L'effet sur notre RNA du chauffage ~ 60°0 t ient sans doute ~ des caract~res propres aux plantes. On retrouve cos partlcularit~s dans la n~cessit~ d'effectuer une dialyse de 12 heures de la preparation apr~s ~limination de l'~ther. L'absenee d'action ribonucl~asique au cours de la dialyse, a ~t~ v~rifi~e en
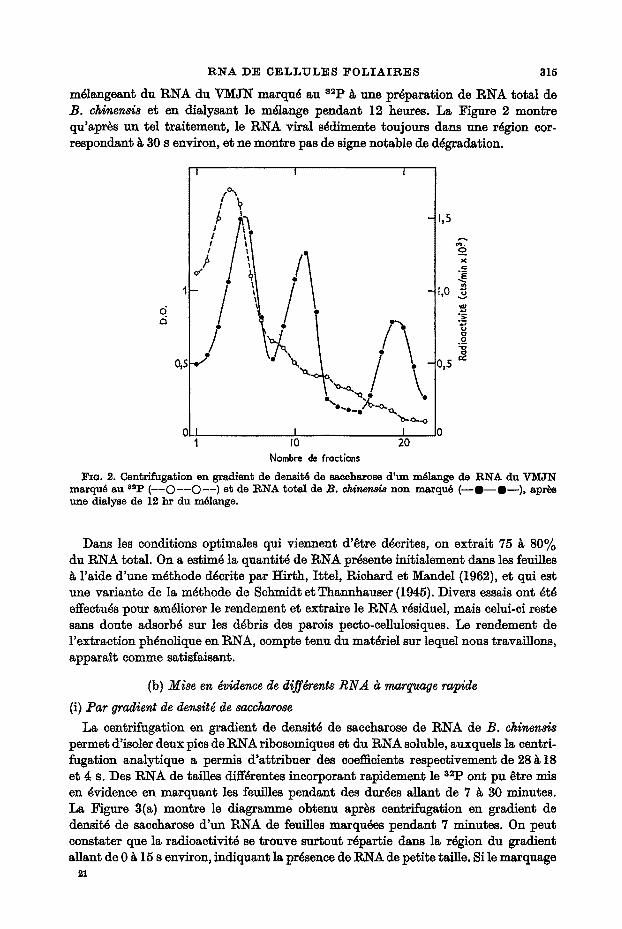
RNA DE CELLULES I~0LIAIRES 3 1 5
m~langeant du RNA du ~ J N marqu~ au 82P ~ une preparation de RNA total de B. ~ h ~ et en dialysant le m~lange pendant 12 heures. La Figure 2 montre qu'apr~s un tel traitement, le RNA viral s~dimente toujours dans une r~gion eor- respondant ~ 30 s environ, et ne montre pas de signe notable de d~gradation.
o"
I I I
!
0,51 /
[ . ,,
1 , J ......................... lO
Nombre de fractions
I 2o
- I , S
1,0
O
x
° ~
O
l~a . 2. Cen~rifuga~ion en gradlent de d e n s i ~ de saceharose d ' u n m61ange de R N A du Y M J N marqu6 au 82p ( - - O - - O - - ) e t de R N A ~otal de B. c ~ n ~ non marqu6 ( - - O - - S ~ ) , apr~s une dialyse de 12 h r du m61ange.
Dans les conditions optima]es qui viennent d'etre d~crites, on extrait 75 ~ 80% du RNA total. On a estim6 la quantit~ de RNA pr~sente initialement dans les feuiUes
l'aide d'une m~thode d6erite par Hirth, Ittel, Richard et Mandel (1962), et qui est une variante de la m~thode de Schmidt et Thannhauser (1945). Divers essais ont ~t~ effectu~s pour am61iorer le rendement et extraire le RNA r~siduel, mais celui-ci reste sans doute adsorb~ sur les d6bris des parois pecto-eellulosiques. Le rendement de l'extraction ph6nolique en RNA, eompte tenu du materiel sur lequel nous travaillons, apparalt comme satisfaisant.
(b) Mise en ~vidence de diffdrents R N A ~ marquage raTide
(i) Par gradient de densltd de sac~harose
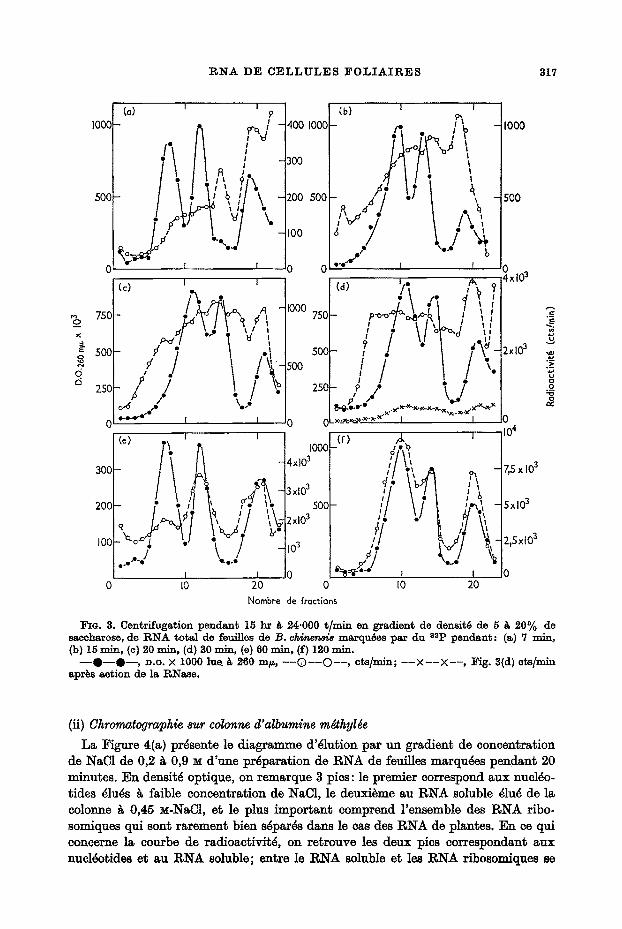
La centrifugation en gradient de densit4 de saccharose de RNA de B. c h i n ~ permet d'isoler deux pies de RNA ribosomiques et du RNA soluble, auxquels la centri- fugation analytique a permis d'attribuer des coefficients respectivement de 28~ 18 et 4 s. Des RNA de failles diff~rentes incorporant rapidement le 82p ont pu ~tre mis en ~vidence en marquant les feuilles pendant des duties allant de 7 ~ 30 minutes. La ~igure 3(a) montre le diagrs.mme obtenu apr~s centrifugation en gradient de densit~ de saccharose d'un RNA de feuilles marquees pendant 7 m~nutes. On peut constater que la radioactivit~ se trouve surtout r~partie dans la r~gion du gradient allant de 0 ~ 15 s environ, indiquant la presence de RNA de petite faille. Si le marquage
21

316 C. GIGOT, G. P H I L I P P S E T L. H I R T H
a lieu pendant 15 mlnutes, on volt un nouveau pic de radioaetivit6 plus lourd ap- paraitre. En effet, la Figure 3(b) montre qu'apr~s ce temps d'incorporation, un RNA radioactif s6ch'mente entre les deux RNA ribosomiques, avec un coefficient de 23 s environ. En6n, si le marquage se poursuit entre 20 et 30 minutes, on peut isoler comme le montrent les Figures 3(c) et 3(d) un troisi~me type de RNA ~ mar- quage rapide, qui s6dimente plus loin que le RNA ribosomique lourd. L'apparition en fonct~on du temps d'inc0rporation, de RNA k marquage rapide de plus en plus lourds est paffaitement reproduetible. D'autre part, le Tableau 1 montre que pour dix pr6parations diff6rentes provenant de feuilles marqu6es pendant 30 minutes, la rSpartition de la radioaetivit~ dans les trois types de RNA ~ marquage rapide est sensiblement la m~me. Toutefois il arrive que, suivant les exp6riences, le RNA 35 s se d6double en deux pics quelquefois tr~s bien sgpar6s (Fig. 5).
TABLEAU 1
_Rgpartltion de la radioactivitg dan~ les cliff grants types de R N A ~ ~narquage rapide, apr~z leur isolement .par centrifugation en gradient de densit$ de saccharose
Pourcentage de radioac~ivit6 dans diff~ren~s RNA 35s 23s 15s
40% 9.6% 34% 42 25,5 32,5 38,5 26 35,5 37,5 30,5 32 39 28 33 43 29 28 42,5 27,5 30 37 34 29 42,5 26 31,5 42,5 29 28,5 42,5 28 29,5
Si les temps de marquage deviennent longs (1 heure et plus) on assiste ~ l'incorpora- tion du 3~p daus les RNA ribosomiques, d'abord le 18 s puis le 28 s (Fig. 3(e) et (f)). Les diff6rents RNA ~ marquage rapide montrent une h6~rog6n~it6 remarquable. En effet, on trouve des fractions 16g~res de 8 ~ 15 s comme chez certaines bact~ries (Takai, Oota et Osawa, 1962), clans le eytoplasme des cellules de foie de rat (Munro et Korner, 1964) ou d'embryons de Xenopus laevis (Brown et Gurdon, 1966). La fraction qui s~climente entre les 2 RNA ribosomiques a la faille du RNA "DNA like" trouv6 par Ingle dans les hypocotyles de soja (1965). EnSn, on trouve des RNA de poids mol6culaire sup6rieur ~ celui du rRNA 28 s, semblables ~ ceux mis en ~vidence par diff~rents auteurs chez des cellules animales (Brown et Littna, 1964; Kempf et Mandel, 1966; Houssais et Attardi, 1966; Scherrer, lYlarcaud, Zajdela, London et Gros, 1966). En ce qui concerne le RNA lourd, l'id~e qu'il s'agit d'un art6fact prove- nant de la formation de complexes entre mRNA et RNA ribosomique peut ~tre ~cart~e. En effet, sous Faction de I'EDTA il reste parfaitement intact, prouvant que les cations divalents qui peuvent provoquer la formation de complexes, ne sont pour rien dans la pr6sence de ce type de RNA. I)'autre part, l'utilisation du ph6nol ~ froid 6rite la formation de complexes dissociables par I'EDTA ou la cha]eur (Ebel, com- munication personnelle).
RNA DE CELLULES FOLIAIRES 317
% m x
E
o; 0
IOOC
50C
75O
500
250
0
300
200
I00
(a) I , P
- ; I
_
r / . r t " ~o ,LI "...d
(c) I I ~ •
_ ! L,~L~ ~,
_,.,-o" I I
e) "'l I . . I
II 1 ~ - -
I I I0 20
Nombre
(b)
400 I000 - i I, / 3 I I .0
,t ~,o~ 300
200,00 ,4 Ji ! a o / . h
-1"7/ 3 .
p " / / \ d
~.~e.e "o X. X'.x..x..,~. ~..a~,,~ . . , ) f '
0 .x.._,~.x..x. I I
i0 3 ~,
0 I0 20
0
I (d)
I000 75C
5C 500
25
1 t
- IOO0
500
4xlO 3
2xlO 3
104
7,s x lo 3
5xlO ~
2,5×103
0
de fractions
Z
FIe. 3. Contrifugation pendant 15 hr k 24'000 t/rain en gradient do densit6 do 5 k 20% de saccharose, do R N A total de feuitles de B. c/viner~a marqu6es par du 82p pendant : (a) 7 mi,% (b) 15 rain, (c) 20 , . i . : (d) 30 mln, (e) 60 rain, (f) 120 rnlm
- - Q - - O - - , P.O. X 1000 lue_/~ 260 m/~, ----O----O----, cts /min; - - - X - - - X - - , ~ g . 3(d) ets/mln aprbs action de la RNaso.
(ii) Ohroma~ogrwlahie aur CO.he d" a~Sumine m~hyIde
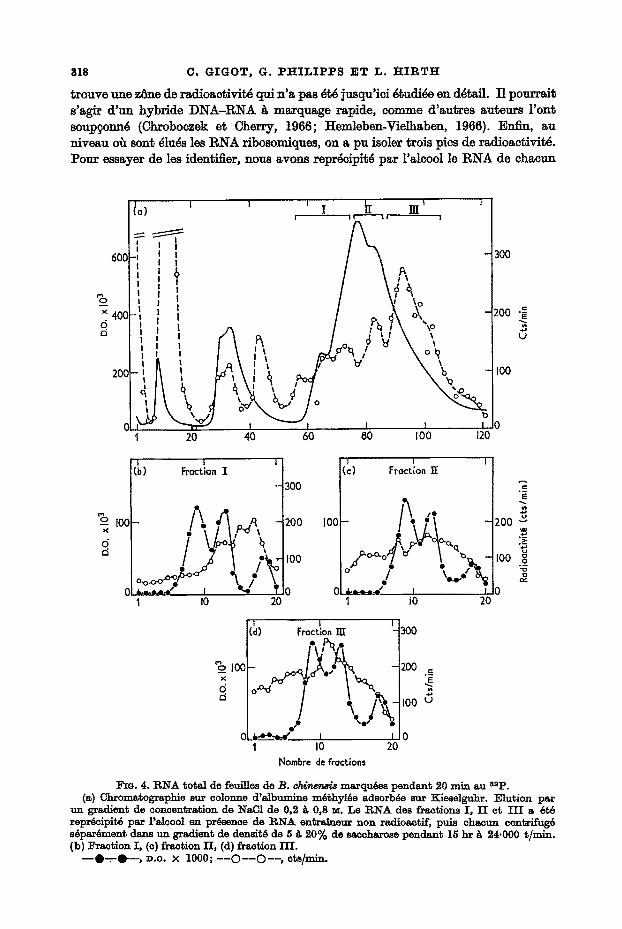
La Figure 4(a) pr4sente le diagramme d'61ution par un gradient de concentration de NaC1 de 0,2 g 0,9 • d 'une pr6paration de RNA de feuilles marquees pendant 20 minutes, En densit6 optique, on remarque 3 pics: le premier correspond aux nuel6o- tides 41u6s g faible concentration de NaC1, le deuxi~me au RNA soluble 61u6 de la colonne ~ 0,45 ~-NaCI, et le plus important comprend l'ensemble des RNA ribo- somiques qui sent raremen~ bien s6par6s dans le cas des RNA de plantes. En ee qui concerne la courbe de racUoactivit4, on retrouve les deux pies eorrespondant aux nucl6otides et au RNA soluble; entre le RNA soluble et les RNA ribosomiques se
318 C. G I G O T , G. P H I L I P P S E T L . H I R T H
trouve une zSne de radioactivit~ qui n'a pas ~t~ jusqu'ici ~tudi~e en d~tail. TEl pourrai~ s ' a ~ d'un hybride DNA-RNA ~ marquage r~pide, comme d'autres au~eurs l'on~ soup~onn~ (Chroboczek et Cherry, 1966; Hemleben-Vielhaben, 1966). Enfin; au niveau o~ son~ ~lu4s les RNA ribosomiques, on a pu isoler trois pics de radioactivitY. Pour essayer de les identifier, nous avons repr~cipit~ par l'aleool le RNA de chacun
~) I ' , ' ! , , k , , m . 1 , i
600t--I
11 ' ' ' t ° 06 . 4001-'I
~oo~ i / ~ \ ! " . ~ " ~ ' X \ -
I 20 40 60 80 I O0 120
300
2o0
u
I00
I00
d c~
Fraction I 3O0
:~_~7 '°°, 'V ',Io I0 20
IOO
I t 1"''} (c) Fraction ]I I
_ J,oo ' x:7 L> I'00
• ~.,:o" t ~10 I0 20
72_ IOC X
o ,.I
I I I (d) Fraction ]]I
fv~ o / ~ i~,~ / %.~ t I0 20
Nombre de fractions
300
2O0 ._=
I00 u
I~IG. 4. R N A %o~1 de feu~es de B. e A 4 ~ marquess pendant 20 rnin au s2p. (a) Chroma~ographie sur colonue d'albl~mi,~e m6thyl6e adsorb~e sur Kieselguhr. Elut ion par
un gradient de concentration de NaCI de 0,2 ~ 0,8 ~. Le R N A des fractions I , I I et I I I a 6t6 repr6cipit6 par l'alcool en pr6sence de R N A entra~ueur non radioactif, puis chacun centrifug6 s6par6ment dans un gradient de densif~ de 5 ~ 20% de saccharose pendant 15 hr ~ 24.000 ~/min. (b) Fraction I, (c) fraction II , (d) fraction HI .
- - O - r - O - - , p.o. X 1000; - - C ) - - O - - , cts/m~-.

RNA DE CELLULES FOLIAIRES 319
des trois pies en presence de RNA entra~meur non marque. Ces trois fractions ont ~t~ analys~es par centrifugation en gradient de densit~ de saccharose. Les Figures 4(b), (c) et (d) indiquent les diagrammes ob~enns.
La fraction I contient essentiellement du materiel l~ger, s~hnentant entre 4 et 18 s. La fraction II consiste en un m~lange de materiel l~ger et lourd, et en~n dans la fraction I I I on retrouve les RNA ~ marquage rapide 23 et 35 s. Le rendement des colonnes d'albumine m~thyl~e a ~t~ de 80% rant en densit~ optique qu'en radio° activit~ dans l'exp~rience dont le diagramme est donn~. Si, apr~s le gradient de NaC1 on 61ue la colonne par de l'ammoniaque 1 M selon la technique de Lingrel (1967), on parvient ~ d•orber les 20~/o restant encore sur la colonne. L'analyse de cette fraction a prouv~ qu'il s'agissait d'un m~lange des cliff, rents RNA marqu6s mis sur la co|onne.
La chromatographie sur colonne d'album~ne m~thyl~e a donc permis de s~parer les RNA lourds des l~gers. Elle a pcrmis 6galement de v~rifier qu'apr&s un temps de marquage de 20 minutes les RNA ribosomiques et soluble de la cellule ne sont prati- quement pas marquis, donc que leur presence n'est pas g~nante pour la d~tcrmlnation de la composition en nucl~otides radioactifs des d ~ r e n t s RNA ~ marquage rapide isol~s par eentrifugation en gradient de densit~ de saccharose. En effet, l'analyse de la composition en nucl~otides marquis des trois pics s6par~s sur colonne d'albumine m~thyl~e (Tableau 2) montre qu'fl s'agit bien de RNA riches en AMP et non de RNA ribosomiques.
(c) ~ompositlo~ e~ ba~e~ de8 R~A ~ ~r~ua~e raw,de
I1 convient de signaler au dSbut de ce paragraphe que la composition des fractions tr~s l~g~res comprises entre 4 eL 8 s, isol~es apr~s gradient de saccharose, apr~s un marquage tr~s bref, est tr~s sp~cia~e, comme le montre le Tableau 1. On pouvait s'attendre ~ ce que le marquage si rapide de cet~e fraction soit dfi ~ l'incorporation du 3~p dans le triplet terminal du sRNA pCpCpA, dont on a pu montrer lc renouvelle- ment rapide et ind~pendant de celui du reste de la chalne (Tamaok~ et Mueller, 1962; ~/Ierits, 1963). En fair, cette composition pr~sentait un fort exc~s en guanine (60 80~/o). En rue de d~celer des contaminations ~ventuelles dans les fractions s~par~es sur Dowex, chaque nucl~otide reconcentr~ a ~t~ chromatographi~ sur eouche m~nce de ce]lu]]ose (Kumar, sous presse). L'~tude de ces plaques par autoradiographie
montr~ clairement clue te pic de "guanine" provenant des RNA tr~s l~gers (4 ~ 8 s) contenait un contaminant radioactif. C'est pourquoi nous avons entrepris d'~tudier la composition de chacune des fractions par ~lectrophor~se (Click, 1966), technique qui nous a permis d'~l~m~uer ces contaminants. Si la composition des fractions sup~- rieures ~ 8 s a ~t~ trouv~e parfaitement indentique ~ celle d~term~n~e par chromato- graphie sur.colonne de Dowex, par contre pour la fraction l~g~re, le taux de guanine a baiss~, le pourcen~age se ma~ntenant ~ 40~/o environ.
En ce qui concerne les autres RNA, on peut constater sur le Tableau 2, ClUe comme chez tousles RNA de plantes ~tudi~s jusqu'~ present, les RNA ribosomiques de B. c h ~ n ~ sont riches en guanine, ce qui donne un rapport GMP]AMP nettement su- p~rieur ~ 1 (G]itz et Dekker, 1963; Hotta et Stern, 1963; Ingle et al., 1965). Par contre, ce qui caract~rise les RNA ~ marquage rapide, c'est leur forte teneur en adenine, donnant un rapport Gi~P]AMP inf~rieur ou voisin de 1, pour les RNA 23 et 35 s. En ce qui concerne le RNA 15 s, sa composition se rapproche davantage de celle des RNA ribosomiques par sa richesse en guanine. Dans le cas du RNA 35 s, ea teneur plus faible en AMP et plus forte en GMP que le RNA 23 s peu~ ~tre due
320 C. GIGOT, G. P H I L I P P S ET L. H I R T H
TABLEAU 2
Com~osiHon en nuddotides des diff~rente ~ N A des feuiUes de B. chinensis. Chaque valeur corresypond h la moyenne de 6 d~terminations au molns
RNA analys6s Composition pour 100 nucl6otides identifi6s GMP A + U Cyb Ad Gu Ur AMP G + C
RNA ribosomiquo 25,3~1,1 23,8+0,8 30,64-0,8 20,4±0,7 1,19 0,79 RNA soluble 28,8±0,7 22,8±0,4 29,54-1,2 18,6±0,8 1,28 0,71 RNA 35 s 20,4±1,0 31,6±0,7 28,34-1,4 19,6+0,9 0,89 1,05 RNA 23 s 20,0±0,6 34,2±1,2 24,5±1,4 21,4+0,5 0,75 1,25 RNA 12 s 18,4±0,8 27,4~1,0 31,8+1,2 22,6±0,6 1,16 1,00 RNA 4/~ 10 st 13,0±1,0 18,0±4,0 44,0+4,0 25,0~1,0 2,40 0,75
Fraction I:~ 21,6 27,8 28,0 22,6 1,04 1,01 Fraction LI~ 21,8 32,0 23,1 23,1 0,72 1,23 Fraction III~ 21,1 34,0 26,0 18,9 0,76 1,12
t Valeurs d6termin~s par 61ectrophor~se. :[ Composition en nucl6otides marqu6s des 3 fractions 61u~s de la colonno d'aJb-mlne
(volt Fig. 4). m6thyl6e
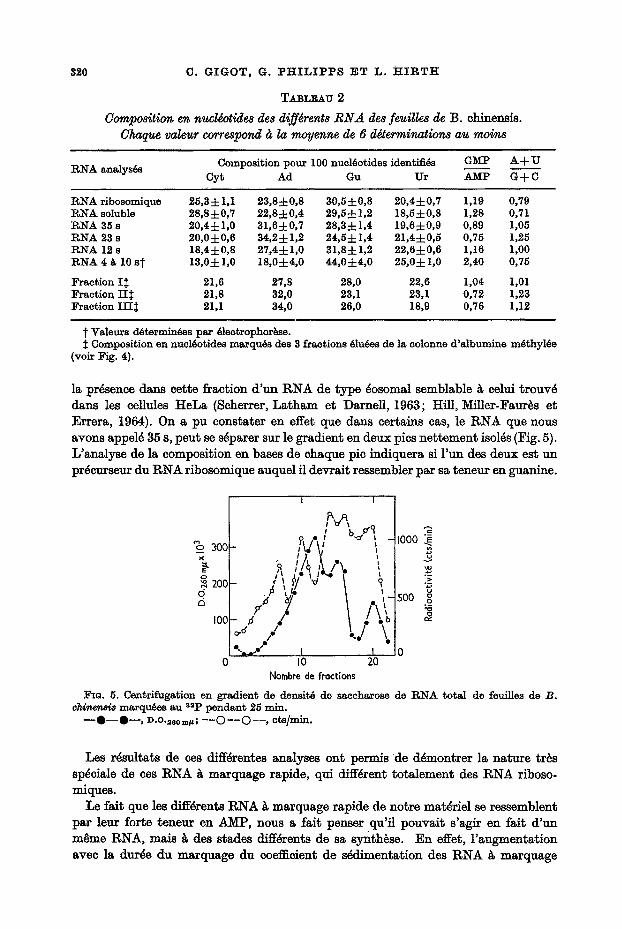
la prdsence dans cette fraction d 'un RNA de type dosomal semblable ~ celui trouv4 dans les cellu]es HeLa (Scherrer, La tham et Dame]l, 1963; Hill, MiUer-Faur~s et Errera, 1964). On a pu eonstater en effet que dans certains cas, le RNA que nous avons appeld 35 s, peut se sdparer sur le gradient en deux pics nettement isolds (Fig. 5). L'analyse de la composition en bases de chaque pic indiquera si Fun des deux est un prdcurseur du RNA ribosomique auquel il devrait ressembler par sa teneur en guanine.
% 3ooF ;V V
o 2oo[- / \ #" 'J"~ [ I I V \ . i
,oop, / t ; 0 I0 20
Nombre tie fractions
1000
500
~o . 5. Cen~rifugation en gradient de densit6 de saccharoso de RNA total de feuilles de B. chinen~ marquees au 8sp pendant 25 rnln.
- - i - - t - - , D.O.2eo m~ ; - - O - - O - - , cts/mln,
Les r ~ t a t s de ces diff~rentes analyses ont permis de d~montrer la nature tr~s sp~ciale de ces RNA ~ marquage rapide, qui diff4rent totalement des RNA riboso- miques.
Le fait que les diff~rents RNA ~ marquage rapide de notre mat4riel se ressemblent par leur forte teneur en AMP, nous a fair penser qu'f l pouvait s'agir en fair d 'un m~me RNA, mais k des stades cliff, rents de sa synth~se. En effet, l 'augmentation a v e c l a dur4e du marquage du coefficient de s~dimentation des RNA ~ marquage

RNA DE CELLULES F O L I A I R E S 321
rapide fair penser A la eroissance de molecules de RNA passant de 10 ~ 35 s, ph@no- m&ne qui rend compte d 'une transcription s6quentielle du DNA cellul~ire. 11 aurait ~t~ int6ressant de voir si les RNA 15 et 23 s 6talent des chalnes de RNA 35 s in- completes, ou s'il s'agissait de RNA auxquels leur taille finale conf&re de tels coeffi- cients de s&dimentation. L'accumulation ou non de la radioactivit~ dans Ies RNA lourds, apr~s un marquage bref suivi d 'une incubation sur milieu non marque, aurait pu renseigner k ee sujet. Les experiences de ee type, r~alis~s avec notre materiel, n 'ont pas donn~ les r6sultats escompt~s. L'aecumulation d~s le d~but du marquage de 32p et d 'autres pr6curseurs marquis sur leur phosphore dans les vaisseaux conducteurs des p6tioles et dans les eellules, rend cos experiences de chassage impossibles A r~aliser.
(d) Ex?~riences de l~r~in~ubatlo~
Etan t donn~ que le 32p n'est pas incorpor$ apr~s un temps court dans les RNA lourds des feuilles couples, on pouvait s'interroger sur le rSle jou4 par le traumatisme ~nfllg~ A la feuille par ce dStachement. On salt en effet qu 'un tel t rai tement provoque des perturbations importantes dans le m~tabolisme des RNA et des prot@ines. On pouvait alors se demander ce qui se passerait si les feuilles ~taient, avant le marquage, pr~incubSes dans un milieu mineral sans s2p. A cet effet, on plonge pendant 10 minutes les p~tioles de feuilles qui viennent d 'etre prglev~es dans un m]lleu m~ngral non marque, puis on les transf~re sur un milieu contenant le s~P pendant 15 minutes. Dans ce cas, l 'activit~ sp~cifique des RNA ~ marquage rapide est de 30~/o plus impor- tante que celle des RNA marquSs 15 minutes ggalement mais sans prgincubation (Tableau 3).
TABLEAU 3
Augmentation de l' activit~ sp~cifique des different8 RNA & marquaqe rapide aTr~s 10 rain de prdincubation, 8uivie d'un marquage de 15 min dan~ les condition~
habituelles
RNA 6tudi6 Augmentation de l'activit~ spdcifique apr~s pr~incubation (%)
35 s 27 23 s 26 12 s 17
L'augmentation a 6t~ d6term~n6e par comparaison avec du RNA de feuilles marqu6es pendant 15 rain, sans pr6incubation.
A la lumi~re de cette experience, on pourrait penser que l 'obtention relativement tardive de RNA lourd est due ~ une reprise de la transcription du DNA, transcription qui serait interrompue au moment du d~tachement de la feuille. Ce point sera 6voqu~ dans la discussion.
Cette experience exclut aussi le fair, d'ailleurs peu vraisemblable, que l'absence de pic 35 s pour les marquages de courte dur~e puisse ~tre due ~ une augmentation transitoire de l 'activit6 ribonucl~asique des cellules apr~s le pr~l~vement. Cette hypoth~se est d'ailleurs contredite par la constance remarquable de la composition en bases des RNA 23 s, 15 s e t 5 s queue que soit la dur~e du marquage. Si, en effet,
$22 C. GIGOT, G. PHILIPPS ET L. I~IRTH
apr~s 7 rnlnutes de marquage, l'absence de RNA 35 se t 23 s ~tait due ~ une d~grada- tion de eeux-ci, la composition des RNA 15 s et,5 s devrait varlet avec le temps, ces quatre types de RNA ayant des compositions sensiblement diff~rentcs.
(e) Oaro.,e~re ~ e ~ g e r oT~ ~ N A ~ ~n, crg[~a, ge raIff, de L'activit~ messag~re d'un RNA peat ~tre d~montr~e par diff~rentes techniques,
fond~es sur les caraet~ristiques et les propri~t~s de ces molecules (Jacob et Monod, 1961). Par la rapidit~ avec laqueUe ils incorporent le ~ P et lear h~t~rog~n~it~, les RNA dent il a ~t~ question jusqu'ici r~pondent d~j~ ~ deux crit~res fondamentaux. D'autre part, l'aptitude ~ s'hybrider avec le DNA, la capacit~ de sthnuler la pro- t~osynth~se ~ v~ro et la contribution de ce RNA ~ Ia formation de polysomes (Risebrough, Tissi~res et Watson, 1962), constituent d'autres propri~t~s des RNA messagers.
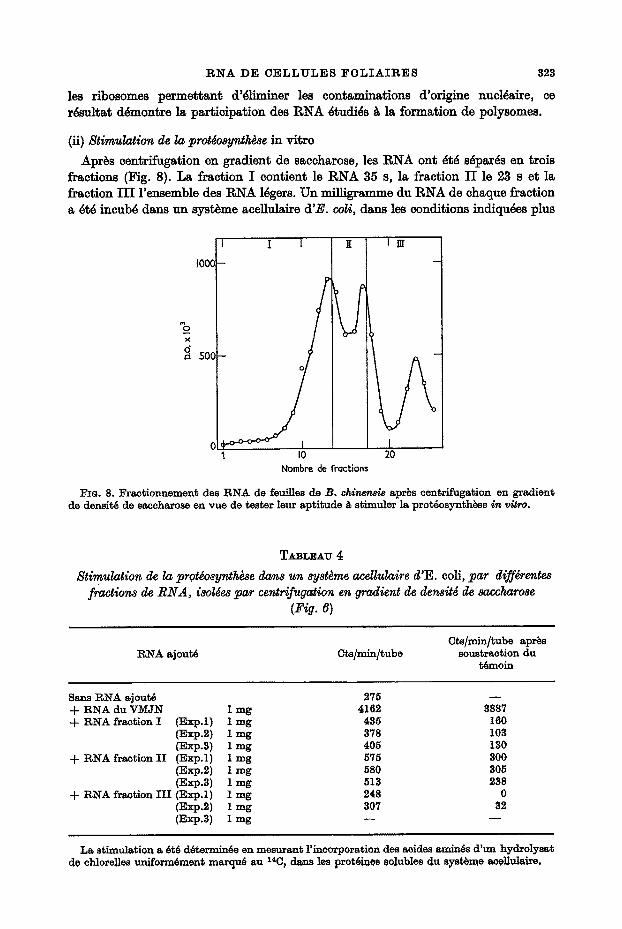
(i) Etude des ~olyr~bo~o~s La Figure 6 pr~sente le dJ&gramme obtenu apr~s centrifugation en gradient de
densit~ de saccharose d'une preparation de ribosomes provenant de feuilles marquees pendant 20 mlnut~s dans les conditions d$finles plus haat. La presence dans les rSgions denses du gradient d'un certain nombre de pic~ de radioactivitY, ainsi que leur
0
x. I0O o r~ I00
1 1
• i ~ 3x lO 3
; ~ 2xi03
~"J~U~'~ ~ r - "o~ I 0 ~
I0 20 o o
1
] i -
/ ?
to
400 c
,200 "I00 u
FIG. 6 Nombre de fractions FIG.7 Fro. 6. Fro. 7.
Fie. 6. Centrifugation en gradient de denslt6 de saccharose de 5 ~ 20% pendant 2 hr 30 rnln 24"000 t/min, d'une pr6paration de ribosomes extraits ~de feuilles de B. chi~e~r/e marqu6es au
s~p pendant 20 rain. ~ 0 - - 0 ~ , p.o. X 10s; - - O - - O - - , cts/mln.
~o. 7. RNA ex~rai~s d'une pr6paration de ribosomes de feuilles de B. c /~ne~ marqu6es pendant 20 min au e2p. Centrifugation en gradient de densit6 de 5 & 20% de saceharose pendant 15 hr ~ 24.000 t/rni~,
~ 0 ~ 0 ~ , p.o. X lOa; - - 0 - - 0 - - , ots/m~n,
disparition apr~s action de la RNase, prouve bien la presence de polysomes. Lear frag~t6 ne permettant pas une purification plus pous~e, c'est ~ partir d'une telle preparation de ribosomes que l'on a extrait le RNA. La Figure 7 indique le diagramme obtenu avec ce RNA. On peat eonstater que le profil de radioactivit~ dans les r~gions 23 et 35 s est absolument semblable ~ celui que donne un RNA extrait directement de feuilles marquees pendant 20 minutes. T,es fractions l~g~res sent eontamin~es rant au point de rue radioactivit~ ClUe densit6 optique par les impuret~s pr~sentes dans les preparations de ribosomes peu purifi~s. T,a technique employee pour preparer
R N A D E C E L L U L E S F O L I A I R E S 328
les r ibosomes p e r m e t t a n t d 'dliminer les con tsmina t ions d 'or igine nucldalre, ee rdsul ta t ddmont re Ia par t ic ipa t ion des R N A dtudids ~t la fo rmat ion de polysomes.
(ii) ~tlmulation de la larotdoxynth~e in vi t ro
Apr~s centr i fugat ion en gradient de saccharose, les R N A ont dtd sdpards en trois fract ions (Fig. 8). La f ract ion I cont ient le R N A 35 s, la f ract ion I I le 23 s e t la f ract ion I I I l 'ensemble des R N A ldgers. U n mil l lgramme du R N A de chaque f ract ion a dtd incubd dans u n systAme aceUulaire d 'E . coli, dans les condit ions indiqudes plus
% x
d r~
100(1
SOC
I0 Nombre de fractions
IDI
I 2O
FIG. 8. Fractionnement des RNA de feuilles de B. c h / ~ apr~s centrifugation en gradient de densit6 de sacoharose en rue de tester leur aptitude k stimuler la prot6osynthbse in vitro.
TABLEAU 4
~timuZation de Za protdosynth&e dans un ayst~me ace~lu~aire d'E. coli, Tar diffdrente~ fraction8 de RNA, isot~e~ par c~ntrifugation en gradient de densitd de 8accharose
( ~ . 6)
Cts/min/tube apr~s RNA ajout6 Cts/min/tube soustraction du
htmoin
Sans RNA ajout6 276 -{- RNA du VMJN I m g 4162 3887 -{- RNA fraction I (Exp.1) 1 mg 435 160
(Exp.2) 1 mg 378 103 (Exp.3) 1 mg 405 130
+ RNA fraction I I (Exp.1) 1 mg 675 300 (Exp.2) 1 mg 580 305 (Exp.3) 1 mg 513 238
+ RNA fraction H I (Exp.1) 1 mg 248 0 (Exp.2) 1 mg 307 32 (Exp.3) 1 mg - -
La stimulation a 6t6 d~termlu~e en mesurant l'inoorporation des aoides amln6s d'un hydrolysat de ehlorelles uniform6ment marqu6 au 14C, dans les prot6baes solubles du s ~ e aoollu~ire.

824 C. G I G O T , G. P H I L I P P S E T L . H I R T H
haut. I~ Tableau 4 rend compte de l'aptitude de chaque fraction ~t stimuler l'in- corporation des acides s.m~nds marquds dams les protdines.
De ces rdsultats, il ressort que les valeurs trouvdes sent significatives et repro- ductibles. Des trois fractions dtudides, c'est celle contenant le RNA 23 s qui est la plus active, la fraction lourde l'dtant sensiblement moins, tandis que les RNA de faible faille ne sent que peu ou pas actifs. Ces rdsultats peuvent ~tre rapprochds de ceux rapportds par Willson et Gros (1964) chez E. co//et par Di Girolamo, Henshaw et Hiatt (1964) ehez le foie de rat, organismes o~ ce sent dgalement les RNA 18 ~. 23 s qui sent les plus actifs, le matdriel lourd l'dtant sensiblement moins.
Le fair que ces RNA & marquage rapide stimulent la protdosynthAse, est une raison suppldmentaire pour admettre leur role de RNA messagers. I1 devenait alors int6ressant d'dtudier la stabilitd de ces RNA messagers, les rdsultats obtenus & ce propos par diffdrents auteurs avec d'autres organismes, dtant des plus variables.
(f) Stabilit~ des RNA ~ marquage raloide
Comme il a ~t6 dit plus haut, l'~tude de la stabilit$ de ces RNA ne pouvait ~tre entreprise par la technique des marquages rapides suivis d'un chassage, ~tant doung nos conditions exp~rimentales et le materiel employS. 1]a done fallu recourir ~ un moyen indirect: ~ savoir l'effet de l'actinomycine D sur la syath~se des RNA d'une part et la protgosynth~se de l'autre. En effet, fl est connu que l'action de cet anti- biotique sur le DNA (Goldberg et Rabinowitz, 1962) emp~che sa transcription par l'intermddiare de la RNA polymdrase DNA-ddpendante. C'est ainsi qu'un certain nombre d'auteurs ont pu ddmontrer cette action inhibitrice sur Ia synth~se des RNA de ceUules animales (Reich, 1961; Tamaoki et Mueller, 1962; Merits, 1963), ainsi que sur certains organes de plantes supdrieures (S~nger et Knight, 1963; Click et Hackett, 1963). La comparaison de l'inhibition de la synthAse des RNA messagers ~t celle de la protdosynthAse pouvait done permettre certaines conclusions quant & la stabilitd du RNA dans notre matdriel.
0
.E
~ so
v
I00 0
I I I I
I I I I I0 20 30 40 50
/~g cl' actlnomyclne
FIG. 9. Inhib i t ion de l ' incorporat ion du sap dans les R N A ribosomiques et soluble apr~s 18hr de t r a i t emen t pa r diff6rentes concentrat ions d 'act inomyeine D. Apr~s t ra i tement , les feuilles on t 6t6 marquees pendan t 4 h r par du 82p (100/~c/ral.). Apr~s isolement des cliff, rents R N A en gra- client de densit~ de saceharose, l ' inhibi t ion a 6t~ calculde pa r rappor t ~ du R N A t6moin p rovenan t de feuilles incubdes pendan t 18 h r 6galement meds sans act inomychte D. --@--@--, sRNA; --0--0~, rRl~A.

R N A D E C E L L U L E S F O L I A I R E S 855
(i) Inhibitio~ de la synth&e de8 RNA
La leigure 9 montre Faction de l'actinomycine D sur l'incorporation du 82p dans les RNA ribosomiques et soluble, en fonc~ion de la concentration en antibiotique. En utilisant une concentration de 50 ~g d'actinomycine D/ml., on peut obtenir une inhibition presque totale de la synth~se des RNA ribosomiques, puisqu'elle est rSduite de 95% par rapport ~ un lot t~moin non pr~trait~. Par contre, l'incorporation du pr~curseur dans le RNA soluble n'est dJmlnu~e que de 75%. I1 a ~t~ d~montr~ en effet (Merits, 1963) clue le RNA soluble continue £ incorporer le s2P dans son triplet terminal pCpCpA m~me en prSsence d'inhibiteur. CeaSe quantit~ de 50 ~g/ml. aurait tr~s bien convenu s'il n'~tait apparu au niveau des feuilles des phSnom~nes d'intoxiea- tion qui pouvaient rendre douteuse l'interpr~tation des rgsultats obtenus. C'est pour cette raison que dans les cas de traitement par l'actinomycine, on a utilis~ une concentration de 30 ~g]ml., permettant d'inhlber la synth~se des RNA ribosomiques de 80% environ (~g. 10).
300 %
E zoo
o. o 100
(a) I m"l i
_
l ,, i
, , ] (b) r" ~'~ ~ 'X -7,.5
/ . 2 ~ -
~o.o.o" I °'~"~ I b 0
o s ._=
2,5 U
0 10 20 0 10 20
Nombre de fract|ons
FTG. 10. Act ion de l 'act inomycine D sur l ' incorporat ion du a2p dans los RNAribosomiques et soluble de feuilles de B. c/dnsns/s. Apr~s u n t r a i t emen t de 18 h r pa r 90 Fg d 'ac t inomycine D/m]., les feuilles ont 6t6 marqu6es pendan t 4 h r pa r du 32p (100/~c/ml.).
(a) R N A de feui]les non trait6es. (b) R N A de feuilles apr~s act inomycins D.
~ i I i I 0
~a
50
I 0 3 6 9 12 15 IB Temps d'actipn de I'acUnomycine
(hr) a 30Fg/ml.
FIG. 11. Inhib i t ion de l ' incorporat ion du s2p dens los R N A ~ marquage rapids en fonetion du temps de t ra ibement pa r 80 Fg d 'act lnomycine D/ml. L ' inhlb i t ion a 6t6 d6termin6e apr~s isolement des R N A sur gradient de denslt6 de saoeharose, pa r r appor t ~ du R N A do feuilles non trait6es.

326 C. GIGOT, G. P H I L I P P S E T L. H I R T H
La Figure 11 indique l 'inhlbition de l ' incorporation du 82p dans los RNA ~ marquage rapide apr~s des duties variables de trai tement par 30 /zg d'actinomycine D/ml. Cette inhibition a ~td d~termin$e en compa~ant, apr~s isolement sur gradient de densit~ de saccharose, l 'activit~ sp~cifique des RNA ~ marquage rapide provenant de feuilles trait~es par l 'actinomycine ~ cello des m~mes RNA provenant de feuilles non trait~es, los deux lots ayant ~t~ marquis pendant 20 minutes.
On obtient ainsi apr~s 15 heures de pr~traitement une inhibition qui est settlement de 55 ~ 60~/o. Comte inhibition incomplete de la synth~se du RNA messager a d~jk ~t$ mise en ~vidence chez certaines cellules animales (Revel e~ Hiatt , 1964) et v~g~tales (Hotta et Stern, 1963; Chroboezek et Cherry, 1966).
(fi) Action sur Ia ~rot&synth&e
Pour d4termlner l 'inhibition de la protgosynth~se, on a ~tudi~ l ' incorporation de la prolinep~C] dans les prot6ines solubles apr~s des dut ies de prgtraitement allant jusqu'g 15 heures. Le Tableau 5 compare l 'inhibition de l ' incorporation de la proline dans los prot4ines calcul~e apr~s 3, 9 et 15 heures de traitement, par rapport un t4moin ayant subi los m~mes pr$incubations mais sans actinomycine. Avec un pr4traitement qui provoque une chute importante de la synth~se des RNA, on ne note aucune baisse clans l'intensitd de la prot4osynth~se. Au con~raire, on a pu constater une exaltation de cette synth~se apr~s 15 heures de traitement.
TABLEAU 5
Gomparaison de l'inhibition de la synthese du RIgA h mar~uage rapide et de la ~rotgosynth&e a~r~ des duties variables de traitement ~ar l'actinomycine D
(aO ~l~l.) Dur6e du trai~ement par
l'actinomycino D 3 hr 9 hr 15 hr
Variation de l'incorporation du 32p daus --27 --48 --56 los RNA g marquage rapide on % par rapport au t6moin
Variation de l'incorporation de ta proline [l~G] dans les prot6ines solubles on % par --0,6 -{-7 +35 rapport au t6moin
Apr~s ce traitemont, un lot do feuilles a 6t~ marqu6 par lo a2p pendant 20 rain pour 6tudier l'inhibition do la synth~se du mRNA, et un autre lot marqu6 pendant 30 min par la proline[14C] a servi ~ l'6tude dos variations de la prot~osynth~se. Los variations de l'incorporation do chacun des pr4murseurs ont ~tx~ d~termin~es par rapport & des fouiUes marqu6es dans los m~mes conditions mais non trait~es par l'actinomycine D.
II n 'apparalt pas d'explication claire g ce phSnom~ne pour l ' instant. Toutefois, le fair que ta prot~osynth~se ne soit pas inhib~e semble prouver la stabilit~ pendant au moins 15 heures du mRNA de notre materiel. Ce r~sultat le range parmi los mRNA stables dgjg misen ~vidence chez des cellules animales (Marks, Burka et Sehlessinger, 1962; Gross et Cousineau, 1963; Revel et Hiatt , 1964) et d'autres v~g~taux (Dure et Waters, 1965).
(g) Effet de la lumi&e sur Ia synth&e des RN A h marquage rapide
Clark, Matthews et Ralph (1964) ont pu eonstater l ' importance de la lumi~re clans la formation des polysomes dans les fouilles. Los Figures 12 montrent la difference de
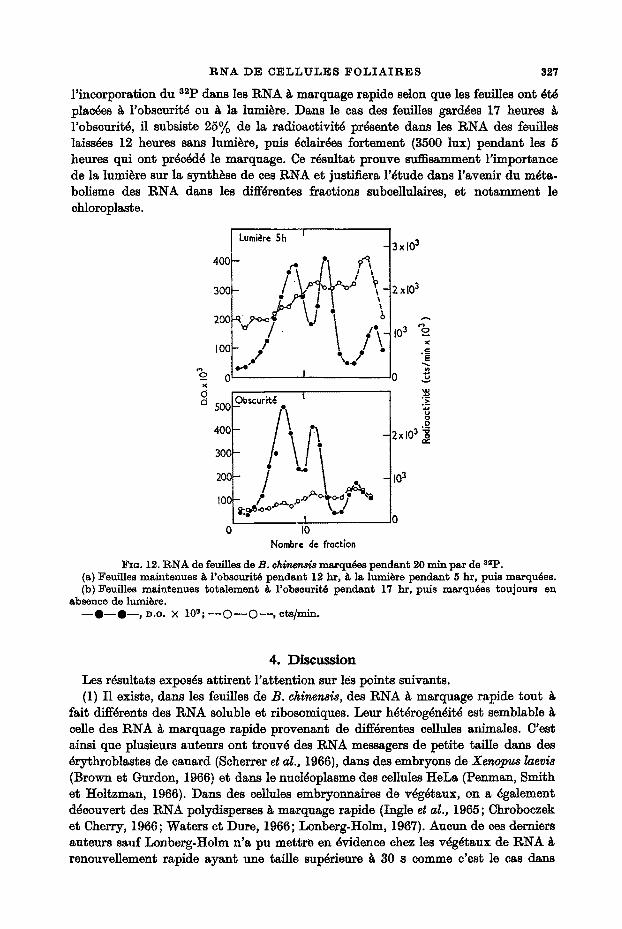
R N A D E C E L L U L E S I ~ O L I A I R E S 327
l ' incorporation du 32p dans les RNA ~ marquage rapide selon que les feuilles ont ~t~ plac~es ~ l'obseurit~ ou ~ la lumi~re. Dans le cas des feuilles gard~es 17 heures l'obscurit~, fl subsiste 25~/o de la radioactivit~ pr~sente dans les RNA des feuilles laiss~es 12 heures sans luml~re, puis ~elair~es for~emen~ (3500 lux) pendant les 5 heures qui ont prSe~cl~ le marquage. Ce rgsultat prouve suffisamment l ' importance de ]a lumi~re sur la synthfise de ces RNA et justifiera l '$tude dans l 'avenir du m~ta- bolisme des RNA dans les diff~rentes fractions subcellulaires, e~ notamment le
% >¢
o cJ
chloroplaste.
/Lumi~re 3 x 10 3 1 5 h
ooI- \ :.,o: I / ~ l o t I
:t;/. IO .~
I ......... ~
500 Obscurlt~ I .->
400 2xlO 3
'3OO
200 - I03
I o o IO
Nombre de fraction
FIo. 12. RNA de feuilles de B. ch i~n~ marqu6es pendant 20 mln par de 3=p. (a) Feuilles maintenues ~ l'obseurit~ pendant 12 hr, ~ la lumi~re pendant 5 hr, puls marqu6es. (b) Feuilles malntenues totalement ~, l'obscurit~ pendant 17 hr, puis marqu6es toujours en
absence de lumi&re. - - 0 - - 0 - - , D.O. × 10s; - - C ) - - O - - , ets/min.
4. D i s c u s s i o n
Les r~sultats exposes at t i rent l 'a t tention sur les points sulvants. (1) I1 existe, dans les feuilles de 27. chinensis, des RNA ~ marquage rapide tout
fait diffdrents des RNA soluble et ribosomiques. Leur h4tgrog4n$itg est semblable celle des RNA ~ marquage rapide provenant de diff4rentes cellules animales. C'est ainsi que plusieurs auteurs ont trouv4 des RNA messagers de petite taille dans des drythroblastes de canard (Seherrer et al., 1966), dans des embryons de Xeno]ous laevis (Brown et Gurdon, 1966) et dans le nuclgoplasme des cellules HeLa (Penman, Smith et Holtzman, 1966). Dans des cellules embryonnaires de v~g4taux, on a 6galement d~couvert des RNA polydisperses k marquage rapide (Ingle et al., 1965; Chroboczek et Cherry, 1966; Waters et Dure, 1966; Lonberg-Holm, 1967). Aucun de ces derniers auteurs sauf Lonberg-Holm n 'a pu mettre en 4videnee chez les v~g4taux de RNA renouve]lement rapide ayant une taiUe sup~rieure ~ 30 s comme c'est le cas dans

328 C. GIGOT, G. PHILIPPS ET L . H I R T H
notre materiel. Ce RNA, trouv~ dans les feuflles, n'est pas sans rappeler les RNA 32 s et 45 s isol~s ~ partir de nombreuses eellules anlmales: des ce]lules HeLa (Houssais et, Attardi, 1966; Warner, Soeiro, Birnboim, Girard et Dame]l, 1966), des ~rythro- blastes de canard (Scherrer et aL, 1966), des oeufs de X. ~evis (Brown et Littna, 1964) et du sarcome plasmocytaire de la souris (Kempf et Mandel, 1966).
Le comportement fie ces RNA lourds au cours de centrifugations en faible con- centration saline ou en presence d'EDTA permet d'exclure qu'fl s'agit d'agr~gats form,s au cours de l'extraction.
(2) Les m~thodes de fractionnement utilis~es ont permis d'&udier les propri&~s des divers pics de RNA ~ marquage rapide. Les RNA 18 ~ 23 s stimulant d'une mani~re significative et reproductible la prot~osynth~se dans un syst~me acellulaire d'E. toll, fl est logique de leur attribuer des propri&~s messag~res. Par contre, le RNA 35 s n'est que peu stimulant. Ayant d~montr6 que la possibilit~ d'un art~fact pouvait &re ~cart~e, il fallait envisager d'autres explications.
L'hypoth~se de la nature polycistronique de ces RNA lourds ~mise par Wfllson et Gros (1964) a ~t~ reprise depuis par diff~rentes autres auteurs. C'est ainsi que Scherrer et al. (1966) et Warner ~ al. (1966) ont suppos~ que ces molecules g~antes de RNA seraient d~coup~es selon un m~canisme analogu~ ~ celui qui preside ~ la formation des RNA ribosomiques 18 et 28 s ~ partir des pr~curseurs 45 et 32 s. D'autre part, Brown et Gurdon (1966) ont pu montrer dans les embryons anucl~ol~s de X. ~evis, la transformation au cours du d~veloppement embryonnaire du RNA de haut poids mol~culaire instable en RNA de faible poids mais stable. ]~n6n, un autre argument en faveur de cette hypoth~se est la faible faille (I0 s environ) du mRNA que l'on trouve associ6 aux polysomes dans le cytoptasme des ~rythroblastes de canard alors que l'on trouve dans le noyau des RNA s~dimentant jusqu'~ 70 s. Une telle structure poly- eistronique du RNA 35 s pourrait expliquer en partie sa faible activit~ clans un syst~me acellulaire. Notons que dans le cas des feuilles de B. chine~is, la composition en bases tr~s sp~ciale de ce RNA permet d'~carter la possibilit~ qu'fl s'agisse exclusive- ment d'un pr~curseur du RNA ribosomique du type 45 et 32 s.
Une autre hypoth~se consiste ~ l~enser que cette fraction correspondrait en fair un m~lange de RNA. Ce point de rue est conflrm~ par des experiences r~centes montrant qu'il est possible de s~parer dans certaines conditions ce RNA 35 s e n au moins deux fractions ayant chacune des propri~t~s stimulantes tr~s diff~rentes (Gigot, Phih'pps et Hirth, r~sultats non publi~s).
La capacit~ de stimulation du RNA lourd total pourrait alors &re ~m~nu~e par une fraction exer~ant une action ~nhlbitrice, comme c'est le eas pour le RNA du ~rMJN dont les propri~t~s stimulantes sont fortement inhib~es par la presence de RNA ribosomique (Gigot, Philipps et Hirth, 1965). Des experiences actuellement en cours devraient permettre de choisir entre ces diff~rentes hypotheses.
(3) En ce qui concerne la composition en bases de ces RNA ~ marquage rapide, fl est apparu un caract~re constant, e'est la richesse on AMP. Cette richesse a d'aflleurs pu &re raise en ~vidence chez tons les RNA ~ marquage rapide isol~s ~ partir de plantes, sans que l'on puisse ~donner ~ ce r~sultat une signification biologique (Hotta et Stern, 1963; Ingle e~ ~., 1965; Loening, 1965; Chroboczek et Cherry, 1966).
L'impossib~t~ dans laque]le nous nous sommes trouv~s malgr~ l'utilisation de diff~rentes techniques (Reddi, 1963; Ralph et Matthews, 1963) d'isoler un DNA de B. ch~ne~i~ suffisamment p u r e t en quantit~s convenables, ne nous a pas permis de comparer la composition des RNA messagers ~. ceUe du DNA. Des experiences en

R N A DE CELLULES FOLIAIRES 329
cours permettent de penser toutefois qu'iI est possible, par une technique appropri6e, d'extraire et de purifier du DNA; on pourra ainsi ob%enir sa composition en bases et ~tudier les capacit~s des cliff, rents types de RNA ~ marquage rapide ~ former des hybrides avec ce DNA.
(4) Un point tr~s particulier rapport~ dans ce m~moire est rineorporation du pr~curseur radioactif dans les RNA K marquage rapide de faille croissant avec |e temps d'incorporation. Jusqu'K present, selon les r~sultats obtenus avec les cellules animales, c'est au ph4nom~ne inverse clue ron assistait. En effet, on salt clue dans le cas des RNA ribosomiques, les pr~curseurs radioactifs sont d'abord incorpor4s dans des RNA de haut poids mol~culaire, ces RNA subissant par la suite un fractionnement en RNA 18 et 28 s (Penman, 1966). I)e m~me, dans le cas des embryons anucl~ol4s de X. b~evis (Brown et Gurdon, 1966), la radioactivit6 apparalt d'abord dans un RNA tr~s lourd et instable, puis par la suite, dans un RNA 16 s stable ayant des propri4t~s messag~res. Chez les cellules animales, le processus g4n6ral de synth%se des RNA semble donc passer par un pr~curseur de haut poids mo]~culaire sans doute poly- cistronique, qui sera ensuite fragment~ pour donner des RNA fonctionnels de masse plus faible. Pour expliquer les particularit~s des RNA de B. chinensis, plusieurs hypotheses peuvent ~tre formul~es.
Une hypoth~se vraisemblable est que certains compar~iments esllulaires sont plus rapidement accessibles au pr~curseur radioactif et que les premiers RNA marquis extraits proviennent par exemple des chloroplastes dont on sait qu'fls contiennent du DNA et sont le si~ge d'une synth%se active de RNA. Par la suite, le 82p atteignant Ie noyau serait incorpor~ dans les grosses molecules qui correspondraient donc au RNA
marquage rapide d'origine nucl~aire. Pour confinner cette explication, il serait indispensable de travailler sur des fractions subcellulaires pures, ce qui est pratique- ment incompatible avecla notion de marquage rapide: les ~chniques de purification des organites ceUulaires sont en effect tr~s longues sans toujours donner des fractions d'une purct~ parfaite.
I1 reste en~n une derni~re hypoth~se. On salt que leur d~tachement de la plante provoque chez les feuilles de tr~s importantes perturbations m4taboliques et notam- ment un arr~t de la synth~se des RNA(Hirth, 1958), et de eelle des pro~ines (Chibnall, 1954). Les r~sultats obtenus au cours des experiences de pr~incubation semblent indiquer que l'on assiste bien ~ un blocage de la synth~se des RNA apr~s le d~tache- ment de la feuiUe. Cet arr~t de synth~se provoquerait donc une synchronisation de la transcription des RNA dans l'ensemble des cellules au moment du marquage. Ce ph~nom~ne peut permettre d'e~xpliquer l'incorporation du ssP dans des RNA de plus en plus lourds. En effet, cctte croissance peut tout simplement correspondre K la visualisation de l~a transcription s4quentielle de certains fragments du I)NA en RNA.
L'4tat actuel des travaux portant sur les probl~mes de transcription chez les v~g~taux ne pcrmet pas de choisir entre ces diff~rentes interpretations.
(5) En ce qui concerns la vitesse de transcription du DNA en RNA, une estimation ne peut ~videmment ~tre faite qu'en formulant les m~mes restrictions clue clans le paragraphe precedent. Si l'on admet pour simplifier, que les diff4rents types de RNA mis en ~vidence proviennent tons du noyau, on peut arriver K une estimation grossi~re du temps n4cessaire pour la formation d'une cha~me de RNA 35 s. En effct, on peut estimer de 3 K 5 minutes le temps n~cessaire pour que la synth~se de RNA reprenne de mani~re normale dans les celiules. Or, comme il faut 12 ~ 15 minutes de marquage pour voir appara~tre le s2P dans les RNA 35 s, on peut 4valuer ~ 10 minutes environ

330 C. GIGOT, G. P H I L I P P S E T L. H I R T H
le temps n~cessaire pour construire uno relic mol6cule. Co temps pout 6videmment sembler eonsid6rable si on le compare/~ Ia vitesse de synth6se chez les bact~ries (Zimmermann et Levinthal, 1967) ou chez los animaux (Penman, 1966). Chez los bact6ries, Zjmmermaun et Levinthal ont pu d6montrer que pour transcrire une mol6cule de RNA ribosomique 16 s il fallait 18 secondes, ce qui repr6sente 0,3~/o du temps de g6n6ration des bact6ries utilis6es. D'autre part, Penman (1966) a prouv6 que le temps n6cessaire pour former un RNA 54 s chez des cellules HeLa est de 2,3 minutes environ, ce qui, compte tenu de la plus grande longueur do la chalne, repr6sente ]a m~me vitesse de synth6se que chez los bact6ries. 11 est 6vident que le temps estim6 dans notre cas pour la synth6se d 'un RNA 35 s est consid6rablement plus 61ev6. I1 n'est malheureusement pas possible de rattacher cette dur6e, d6termin6e tr6s grossi6rement, au temps do g6n6ration des cellules foliaires. En effet, la grande majorit6 de celles-ci, bien qu'6tant le si6ge d'une intense prot6osynth6se, no pr6sente plus aucune activit6 mitotique. Cette synth6se tr6s lento des RNA pout n'6tre, si elle est v6rifi6e, que le reflet de la grande stabilit6 des eellules 6tudi6es.
(6) Los exp6riences de chassage n'6tant pas r6alisables, c'es~ par l'utilisation de l'actinomycine D que l'on a 6tudi6 la stabilit6 des RNA messagers.
Los exp6riences rapporb6es sent critiquables/~ cause de l'inh~bition incompl6te do la synth6se des IRNA messagers. Elles permettent n6anmoins, en comparant l'action do l'actinomycine sur la prot6osynth6se (aucune inhibition apr6s 15 heures) et sur la synth6se des mRNA (inhlbition de 60~/o) d'envisager une stabilit6 de 15 heures au moins des RNA messagers de notre mat6riel. Une telle stabilit6 a d6j~ 6t6 trouv6e chez des oeufs d'oursins (Gross et Cousineau, 1963) et chez des graines de coton on germination (Waters et Dure, 1966).
On pout se demander toutefois si tousles mRNA ont le m6me degr6 de stabilit6. R6cemment, on offer, des auteurs ont montr6 clue chez un m6me mat6riel, la stabilit6 du mRNA varie avecla prot6ine dent il contrSle la synth6se (Marchis-Mouren et Cozzone, 1966; Appel, 1967). On dolt en effet so rappeler que l'actinomycine D n'inhibe dans nos conditions la synth6se des RNA messagers que de 60 ~/o, alors qu'eUe inhibe cello du RNA ribosomique de 85%.
D'autre part, nos recherches ont port6 sur la totalit6 des mRNA de la cellule. Or, on sait que los chloroplastes contiennent du DNA et peuvent synth6tiser des mRNA (Schweiger et Berger, 1964; Brachet et Ooffeau, 1964; Ooffeau et Brachet, 1965). On ignore actuellement si, in rive, le DNA chloroplastique est accessible & l'actinomycine D dans los m~mes conditions que le DNA nucl6aire. Cos remarques ainsi que cellos faites & propos des probl6mes pos6s par la transcription, montrent Fint6r~t qu'il y aura, dans de futures recherches, & d6termlner la part qui revient dans nos conditions exp6rimentales, aux RNA messagers d'origine nucl6aire et chloroplastique.
Nous tenons ~ remercier le Docteur Francois Gros pour los pr~cleux conseils qu'il a bien voulu nous prodiguer tout au long de ee travail. Nos remerciements vent 6galement Mine Mich/~le Meyer pour son assistance technique.
L'actinomycine D a 6t6 graciensement fournie par Merck, Sharp et Dohme, New York.
REFERENCES Appel, S. H. (1967). 2~a~urs, 218, 1253. Bayley, S. T. (1964). o r. ~o~. R~o~. 8, 231. Braehet, J. et Goffeau, A, (1964). C.R. ~lcad. ~c~. Par/a, 269, 2899. Brown, D. D. et Gurdon, J. B. (1966). or. fV~ot. B~ol. 19, 399. Brown, I), D. et Littna, E, (1964). or, .Mo~. ~o~, 8, 669.

RNA DE CELLULES I~OLIAIRES 331
Chibnall, A. C. (1954). New P h i l . 58, 31. Chroboezek, H. et Cherry, J . H. (1966). J. Mol. Biol. 19, 28. Clark, M. F., Matthews, R. E. F. et Ralph, R. K. (1964). Biochim. biophy~. Acta, 91, 281. Click, R. E. (1966). Biochim. biophya. Acta, 129, 426. Click, R. E. et Hackett, D. P. (1963). Proc. Nat. Acad. Sci., Wash. 5(), 243. Dure, L. et Waters, L. (1965). Scisncc, 147, 410. Di Girolamo, D., Henshaw, E. C. et Hiatt , H. H. (1964). J. Mol. Biol. 8, 479. Gigot, C., Philipps, G. et Hirth, L. (1965). Bull. Soc. Gh~m. Biol. 4"I, 1667. Glitz, D. G. et Dekker, C. A. (1963). Biochem. J. 2, 1185. Goffeau. A. et Brachet, J. (1965). Biochirm. biophys. Ast~, 95, 302. Goldberg, I. H. et Rabinowitz, M. (1962). Sc/~ncs, 136, 315. Gross, P. R. et Cousineau, G. H. (1963). Bioch~m. Biophys. Rea. Gomm. 1{}, 321. Hemleben-Vielhaben, V. (1966). Z. Naturf. 21b, 983. Hill, M., Miller-Faur~s, A. et Errera, M. (1964). Biochim. biophys. Acta, 80, 39. Hirth, L. (1958). Th~ss ds doctorat d'$tat, Paris. Hirth, L., Ittel, E., Richard, C. et Mandel, P. (1962). G.R. Acad. Sci. Paris, 255, 3217. Hotta, Y. et Stem, H. (1963). J. Gstl Biol. 19, 45. Houssais, J . F. et Attardi, G. (1966). Pros. Nat. Acad. Sci., Wash. 49, 240. Ingle, J., Key, 3. L. et Holm, R. E. (1965). J. MoL Biol. 11, 730. Jacob, F. et Monod, J . (1961). J. MoL Biol. 3, 318. Kempf, J. e~ Mandel, P. (1966). Bull. Soc. Ghim. Biol. 48, 211. Lingrel, J . B. (1967). Biochi,m. biophys. Ac~, 142, 75. Loeniug, U. E. (1965). Proc. Roy. Soc. B, 162, 121. Lonberg-Holm, K. K. (1967). Naturs, 218, 454. Mandell, J . D. et Hershey, A. D. (1960). Analyt. Bioche~. l , 66. Marchis-Mouren, G. et Cozzone, A. (1966). Biochemistry, 5, 3684. Marks, P., Burka, E. R. et Schlessinger, D. (1962). Prov. Nat. Acad. Sci., Wash. 48, 2163. Merits, I. (1963). Biochsm. Biophys. Rez. Gomm. 10, 254. Monier, R., l~aono, S., Hayes, D., Hayes, 1~. et Gros, F. (1962). J. Mol. Biol. 5, 311. Munro, A. J. , et Korner, A. (1964). Nature, 201, 1194. Nirenberg, M. W. et Matthaei, J. H. (1961). Proc. Na~. Acad. Svi., Wash. 47, 1580. Ogur, M. et Rosen, G. (1950). Arch. Biochsm. 25, 262. Penman, S. (1966). J. Mol. Biol. 17, 117. Penman, S., Smith, I. et Holtzman, E. (1966). ~cionce, 154, 786. Ralph, R. K. et Matthews, R. E. F. (1963). Bioch~m. Biophys. Res. Comm. 12, 287. Reddi, K. K. (1963}. Biochim. biophys. Acta, 72, 33. Reich, E. (1961). ScOnce, 134, 556. Revel, M. et Hiatb, H. H. (1964). Proc. Nat. Acad. Ssi., Wazh. 51, 810. Risebrough, R. W., Tissi~res, A. et Watson, J. D. (1962). Proc. Nat. Acad. Sci., Wash.
48, 430. S~nger, H. L. et Knight, C. A. (1963). Biochem. Biophys. Res. Gom~. 18, 455. Scherrer, K. ot Damell, J . E. (1962). Biochem. Biophys. Res. Gomm. 7, 486. Scherrer, K., Latham, H. ot D'~amell, J. E. (1963). Pros. Nat. Acad. Ssi., Wash. 49, 240. Seherrer, K., Marcaud, L., Zajdela, F., London, I. et Gros, le. (1966). Proc. Nat. Acad. Sci.,
Wash. 56, 1571. Sctmaidt, G. et Thannhauser, J . (1945). J. Biol. Ghsm., 161, 83. Schweigor, H. et Berger, S. (1964). Biochim. biophys. Acta, 87, 533. Takai, M., Oota, Y. et Osawa, S. (1962). Biochim. biophys. Acta, 55, 416. Tamaoki, T. ot i~Iueller, G. L. (1962). Biochem. Biophys. Rss. Gomm. 9, 451. Warner, J., Soeiro, 1%., Birnboim, H. C., Girard, M. et Damell, J. E. (1966). J. MoL Biol.
19, 349. Waters, L. C. ot Dure, L. S. (I966). J. Mol. Biol. 19, 1. Wfllson, C. et Gros, F. (1964). Biochim. biophys. Acta, 80, 478. Zimmermann, R. A. et Levinthal, C. (1967). J. MoZ. Biol. 36, 371.
22





























