Embed Size (px)
Citation preview

0
Hélène Deraison Master 2 Sciences Technologies Santé Parcours Ecologie Fonctionnelle, Comportementale et Evolutive Université de Rennes 1 Année 2009-2010
RAPPORT DE STAGE
Analyse de la structure de la communauté de Carabidae en système céréalier intensif
Organisme d'accueil : Centre d’Etudes Biologiques de Chizé (CNRS / INRA) Sous la direction de : Vincent Bretagnolle, Directeur

1
Sommaire
REMERCIEMENTS p.2 INTRODUCTION p.3 MATERIEL ET METHODES p.6
1. Site d’étude p.6
2. Présentation du jeu de données p.7
3. Méthode d’échantillonnage des Carabidae p.7
4. Validation de la méthode d’échantillonnage des coléoptères carabiques p.8
5. Description sommaire de la communauté des Carabidae de la Zone Atelier p.9
6. Analyse des traits biologiques des espèces p.10
7. Influence de la surface des parcelles sur les communautés de Carabidae p.11
RESULTATS p.11
1. Robustesse de la méthode d’échantillonnage des coléoptères carabiques p.11
2. Description de la communauté de Carabidae de la Zone Atelier « Plaine
& Val de Sèvres » p.13
3. Comparaison des communautés de coléoptères carabiques entre les cultures p.15
4. Influence de la surface des parcelles sur des communautés des Carabidae p.21
DISCUSSION p.22 CONCLUSION p.24
BIBLIOGRAPHIE p .26 Annexe 1. Tableau récapitulatif des espèces échantillonnées sur la Zone Atelier Plaine & Val de Sèvre p.28 Annexe 2. Tableau récapitulatif des traits biologiques p.29 Annexe 3. AFC sur différents jeux de données p.30

2
REMERCIEMENTS
Je remercie tout particulièrement Vincent Bretagnolle de m’avoir accueillie dans son
équipe et son laboratoire et de m’avoir offert cette opportunité de stage. Merci également de
m’avoir accompagnée, fait partager ses connaissances et son expérience, et de m’avoir
conseillée tout au long de la réalisation de ce rapport.
Merci à David Pinot pour ses formations R et ArcGIS.
Egalement un grand merci à Kevin pour sa patience et ses conseils avisés sur
l’utilisation du logiciel R.
Merci à Jérôme pour sa disponibilité. Nos nombreux échanges m’ont enrichie sur ce
groupe.
Je tiens également à remercier les étudiants du bureau M2 : ambiance et bonne humeur
à toute épreuve.
Merci enfin à Orianne pour son soutien dans les moments difficiles, une belle
complicité s’est construite au fil des mois.

3
INTRODUCTION
L’écologie des communautés est une discipline qui vise principalement à comprendre et
prédire les patterns de distribution de la biodiversité afin notamment de comprendre le rôle de
la biodiversité dans les écosystèmes (Holyoak et al., 2005). L’impact des activités
anthropiques sur l’environnement renforce notre volonté d’étudier les mécanismes régissant
les communautés et ce dans un but de conservation et de gestion des écosystèmes (Loreau et
al., 2001). En effet, les activités humaines ont bouleversé l’environnement global en altérant
profondément l’utilisation des terres et des eaux, les cycles biogéochimiques, la chimie
atmosphérique, la distribution et la dynamique de la biodiversité à l’échelle planétaire (Sala et
al., 2000). La transformation et la fragmentation des habitats naturels apparaissent clairement
comme les principales causes à l’origine de l’extinction et du changement d’abondance des
espèces et constituent une véritable menace pour la biodiversité (Schweiger et al., 2005).
Actuellement, le taux de disparition des espèces est 1000 fois supérieur au taux naturel et
nous entraîne vers la 6ème crise de la biodiversité.
Depuis 50 ans, de nombreuses théories ont été proposées pour expliquer et prédire la
structure et la dynamique des communautés. La première théorie est celle de la Biogéographie
Insulaire proposée par Mac Arthur et Wilson en 1963. Elle considère un réservoir d’espèces
correspondant à un continent. Autour de ce dernier se trouve un ensemble d’îles de taille
variable plus ou moins éloignées du continent. Le nombre d’espèces de chaque île va
dépendre de deux processus : le taux de colonisation à partir du continent et le taux
d’extinction des espèces une fois sur les îles. Le premier taux dépendant lui-même de la
distance entre l’île et le continent (plus l’île est éloignée du continent, plus le taux de
colonisation sera faible et inversement). Le second taux dépend de la taille de l’île, sous
l’hypothèse implicite qu’un milieu plus grand est plus hétérogène et diversifié (Tscharntke &
Brandl, 2004). Cette théorie considère les espèces de la communauté comme indépendantes
les unes des autres, et l’explication de leur distribution ne tient pas compte des interactions
éventuelles qui peuvent se produire entre elles.
Une autre théorie est celle des Métapopulations, proposée par Levins en 1969. Elle se
réfère, à l’inverse de la première théorie, à un ensemble de réservoirs d’espèces ou patches
d’habitat entourés d’une matrice hostile. Le nombre d’espèces sur chaque patch va dépendre
des mêmes paramètres que précédemment : le taux de colonisation et le taux d’extinction des
espèces. Les espèces sont toujours considérées comme indépendantes. En revanche, les
patches varient les uns par rapport aux autres en termes de qualité. Un patch de meilleure

4
qualité présentera une plus forte probabilité d’être colonisé qu’un patch de moins bonne
qualité.
La troisième théorie est celle des Métacommunautés (Holyoak, 2005), apparue
récemment. Elle considère un ensemble de communautés locales reliées par le phénomène de
dispersion. Cette théorie présente l’avantage de pouvoir expliquer des patterns de distribution
aussi bien à l’échelle locale que régionale. De plus, elle présente la caractéristique de prendre
en compte les processus intervenant dans les relations interspécifiques, telles que les relations
trophiques, susceptibles d’affecter le taux d’extinction et de colonisation des espèces au sein
des patches. Cette théorie reste cependant récente et encore très conceptuelle par rapport aux
autres.
Enfin la dernière théorie est la théorie neutraliste (Hubbell, 2001 in Alonso, 2006). Elle
prédit une distribution aléatoire des espèces dans leur environnement : par exemple, un milieu
riche présentera une richesse spécifique plus importante qu’un milieu pauvre. Les espèces
possèdent des capacités de compétition égale et leur distribution est toujours liée aux
processus de colonisation et d’extinction, mais le processus principal est ici stochastique.
Enfin, ces théories étant assez proches les unes des autres, il est difficile de les départager.
Les agro-écosystèmes apparaissent de ce fait comme des milieux parfaits pour ces études
sur les communautés. En effet, les paysages agricoles peuvent être considérés comme une
mosaïque hétérogène de « patches » ou d’îles, car ce sont des systèmes spatialement
complexes composés d'éléments divers plus ou moins proches, contrastés et de qualité
variable, chaque élément correspondant à une parcelle. Les contraintes spatiales y sont donc
claires et fortes. Par ailleurs, les agro-écosystèmes sont des milieux particulièrement perturbés
par l’action de l’homme (via l’agriculture), et dans ces systèmes, les populations animales
comme végétales sont soumises à des extinctions régulières et répétées. Les populations ne
peuvent donc se maintenir à l’échelle du paysage agricole que par la dispersion et la
colonisation de nouveaux habitats (Elzinga, 2007). Enfin, les conséquences des activités
anthropiques vont également dépendre des espèces et de leurs capacités à disperser (ainsi bien
sûr que de la structure du paysage agricole), et on peut donc s’attendre à ce que les différentes
espèces composant les communautés réagissent différemment au régime de perturbation.
Les agro-écosystèmes ont été soumis à une forte perte de biodiversité due à
l’intensification agricole qui agit à travers plusieurs aspects : une simplification du paysage,
une mécanisation des pratiques ainsi qu’une forte augmentation de l’usage des intrants
(Schweiger, 2005). Cette intensification a entrainé une uniformisation de l’occupation du sol,

5
la raréfaction de nombreuses espèces et une dégradation globale de la qualité des habitats
dans les agro-écosystèmes (Benton et al., 2003 ; Duelli et al., 2003). La mise en œuvre de la
PAC a de plus eu comme conséquence l’abandon de milieux naturels tels que les prairies bien
qu’important pour la biodiversité (Bàldi & Kisbenedek, 1997). Les communautés des agro-
écosystèmes sont donc numériquement plus faibles que ce qu’elles devraient être, mais elles
sont toutefois encore suffisamment riches pour des études pertinentes.
Les coléoptères carabiques représentent un modèle biologique original et pertinent pour
des études sur les communautés. D’une part, ils comportent une diversité spécifique élevée et
les espèces qui composent la communauté présentent des traits d’histoire de vie très
diversifiés (Dajoz, 2002). En effet, ils regroupent des taxa réagissant différemment aux
conditions biotiques et abiotiques de l'environnement et sont sensibles aux microclimats
(Gutierrez et al., 2004 ; Lambeets et al., 2008). Ces caractéristiques en font ainsi de bons bio-
indicateurs (Melnychuk et al., 2003). D’autre part, au sein des agro-écosystèmes de l’Europe
tempérée, les Carabidae constituent le groupe dominant d’Arthropodes épigés (Dajoz, 2002).
Ils forment un des maillons de base du réseau trophique dans ces milieux, en tant que proies
d’insectes, de reptiles, d’oiseaux et de mammifères et possèdent une position clef du point de
vue de l’agro-écologie en tant qu’auxiliaire des cultures (Melnychuk et al., 2003). Enfin, leur
forte abondance dans les zones agricoles permet de les utiliser pour des analyses statistiques
(Duelli, 1999).
Objectifs de cette étude
L’objectif de ce travail est d’étudier la communauté de Carabidae dans un paysage
agricole intensif, en particulier d’essayer de déterminer les règles d’assemblages d’espèces au
sein de la communauté et entre les cultures. En effet, de nombreuses études montrent que
d’une part les groupes d’espèces se distinguent selon le régime de perturbation agricole du
milieu considéré et d’autre part que les groupes d’espèces occupant ces milieux présentent des
traits d’histoire différents (Dufrêne, 1999 ; Niemela, 2001). Il est important de noter que les
Carabidae présentent un gradient de taille associé à leur capacité de dispersion. En effet, les
petites espèces sont macroptères, c'est-à-dire qu’elles possèdent des ailes fonctionnelles et
sont donc capables de disperser par le vol sur de plus grandes distances que les espèces de
grande taille, aptères, qui se déplacent uniquement par la marche (noter toutefois que quelle
que soit l’espèce, le mode de déplacement privilégié reste cependant la marche). Ainsi, au
sein de milieux plus faiblement perturbés, tels que les praires ou les luzernes, par rapport aux

6
cultures annuelles (colza, blé), on s’attend à trouver des espèces qui résistent difficilement aux
perturbations, c'est-à-dire des espèces de grande taille, spécialistes et brachyptères à faible
capacité de dispersion (Gobbi, 2008). A l’inverse, dans des milieux fortement perturbés
comme les céréales ou le colza, les espèces seraient des espèces généralistes de petite taille et
macroptères à forte capacité de dispersion. Cela leur permettant de se déplacer dans un
environnement favorable voisin pour s’alimenter.
Le deuxième enjeu de ce travail est d’analyser l’influence de la surface des parcelles sur la
richesse spécifique. Sous l’hypothèse que les parcelles de grande taille sont plus hétérogènes
et renferment des micro-habitats plus ou moins favorables on s’attend à ce que les espèces
spécialistes occupent des parcelles de grande taille et inversement pour les espèces
généralistes. Ces premières espèces présentent des exigences particulières puisqu’elles ont
une durée de vie plus longue et un plus faible taux de reproduction (Dufrêne, 1999). Ainsi,
une grande parcelle leur offre de meilleure condition de vie qu’une petite parcelle.
Le troisième enjeu de cette d’étude consiste à analyser la covariance entre les cultures et
les traits d’histoire de vie des espèces, ce qui permettra de confirmer ou non, que les cultures
telles que le colza et les céréales sont des habitats plus perturbés que les luzernes et les
prairies et ainsi que confirmer les hypothèses du première enjeu.
Au préalable, la qualité et la robustesse du protocole d’échantillonnage des Carabidae en
tant que moyen d’estimer la richesse spécifique à l’échelle de la zone d’étude et à l’échelle de
la parcelle sera étudiée.
MATERIEL ET METHODES
1. Site d’étude
Cette étude est réalisée au sud du département des Deux-Sèvres en région Poitou-
Charentes (France) dans la plaine céréalière de Niort Brioux (Fig1). Il s'agit d'une plaine
céréalière intensive, sur laquelle une Zone Atelier d’environ 500 km2 (et dont la moitié
environ appartient au réseau Natura 2000) a été labellisée. Elle comprend comme cultures
principales des céréales, du maïs, du tournesol, du pois et du colza ainsi que des élevages
bovins et caprins mais en diminution. Les plaines agricoles de la région Poitou-Charentes sont
des milieux abritant une très grande richesse biologique ayant une valeur patrimoniale comme
l'outarde canepetière (Centre d’études biologiques de Chizé : Zone atelier - Plaine & Val de
Sèvre [en ligne]. Disponible sur : <http://www. zaplainevaldesevre.fr /present.php> (consulté
le 28.03.2010)).

7
2. Présentation du jeu de données
Sur cette Zone Atelier, des piégeages de carabes sont réalisés depuis 1994, mais au fil des
années et des études spécifiques, 7 protocoles d’échantillonnage différents ont été mis en
place entre 1994 et 2009. Les données sont spatialement référencées et le type de culture
échantillonné est noté. Elles se présentent sous forme d’abondance ou de présence/absence et
ont été compilées dans une base de données avec le logiciel Access version 97. Cette base
regroupe un total de 94 espèces de coléoptères carabiques (Annexe 1) (ordre des coléoptères,
sous ordre des adephaga, famille des Carabidae ; Dajoz, 2002) et 1199 parcelles
échantillonnées. Seules les données comprises entre 2005 et 2009 sont analysées dans ce
travail afin d’éviter un biais inter-année ainsi qu’un biais d’identification (une seule personne
a en effet réalisé la totalité des identifications entre 2005 et 2009).
3. Méthode d’échantillonnage des Carabidae
Chaque protocole est réalisé avec des pièges de Barber, également appelés pots pièges, de
7cm de hauteur et de 8,5cm de diamètre. Ces pièges sont enfouis dans le sol de manière à ce
que le pot ne dépasse pas la surface du sol. Ils sont remplis avec de l’éthylène-glycol dilué à
50% avec de l’eau.
La méthode d’inventaire des Carabidae est spécifique à la zone d’étude en termes de
durée de piégeage utilisée et de nombre de pots par parcelle. En fait, il n’existe pas de
méthode standardisée (Pearsall, 2007). Ainsi, lors de cette étude la durée de piégeage est de 5
Fig1 : Localisation de la région Poitou Charentes sur la carte de la France. A droite : Localisation de la zone d’étude sur la carte de la région Poitou Charentes
Photographies de deux espèces de coléoptères carabiques. A gauche Poecilus cupreus, à droite Anchomenus dorsalis. Source : http://www.kerbtier.de

8
jours, alors que d’autres études en Europe sont basées sur une durée de piégeage pouvant aller
de 7 à 10 jours.
4. Validation de la méthode d’échantillonnage des coléoptères carabiques
La validation de la méthode d’inventaire des carabidae utilisée dans cette étude a consisté
à déterminer si le nombre de parcelles totales échantillonnées, le nombre de pièges placés par
parcelle ainsi que la durée de piégeage, permettent d’estimer correctement la richesse
spécifique présente dans une parcelle ou sur une culture donnée. La richesse spécifique est un
indice de diversité simple et facilement analysable (Gotelli & Colwell, 2001). Il sera alors
utilisé afin de déterminer si l’effort d’échantillonnage appliqué estime correctement cet
indice.
En ce qui concerne la validation de l’effort d’échantillonnage en termes de nombre de
parcelles échantillonnées et de nombre de pots placés par parcelle, elle fait classiquement
référence aux courbes de raréfaction (Colwell et al., 1994 ; Gotelli & Colwell, 2001 ; Colwell
et al., 2004). Cette méthode consiste à effectuer un tirage aléatoire sans remise de N parcelles
et à calculer la richesse spécifique cumulée obtenue en fonction du nombre de parcelles tirées.
Pour de nombreux taxa plus l’effort d’échantillonnage augmente plus on collecte de nouvelles
espèces. Les espèces généralistes sont capturées en premier puis lorsque l’effort
d’échantillonnage devient très grand on n’ajoute plus que des espèces rares. Cependant il peut
être difficile d’utiliser un effort d’échantillonnage approprié permettant d’obtenir la richesse
spécifique maximale de la zone étudiée. Ainsi, les courbes de raréfaction permettent à partir
de données observées, d’obtenir une estimation de la richesse spécifique cumulée en fonction
de l’effort d’échantillonnage fourni. Cette méthode présente également l’avantage de pouvoir
comparer différents jeux de données récoltés avec un effort d’échantillonnage différent. Cette
analyse est réalisée sur l’ensemble du jeu de données de 2005 à 2009 puis sur quatre sous jeux
de données qui séparent les parcelles selon le nombre de pots-pièges qu’elles contiennent.
Afin d’estimer la richesse spécifique maximale du jeu de données considéré la formule de
Chao est utilisé. Les analyses sont effectuées sous le logiciel R avec le package vegan
(Oksanen et al., 2009).
En ce qui concerne la durée de piégeage, elle est évaluée à partir d’un protocole spécifique
réalisé sur une unique parcelle (jachère sous contrat d’agriculture durable) en 2005. Dans
cette parcelle 150 pots ont été placés le même jour, puis tous les jours pendant 10 jours, 15
pots sont récoltés aléatoirement parmi les pots restant. Le nombre moyen d’espèces est
calculé par durée de pose de piège. Le test de Kruskall Wallis suivi d’un test post hoc avec

9
une correction de holm ont été utilisés afin de déterminer si le nombre moyen d’espèces
différait selon le nombre de jours de pose des pièges.
5. Description sommaire de la communauté des Carabidae de la Zone Atelier
Les 94 espèces identifiées depuis 1994 ne sont pas nécessairement présentes tous les ans
ou tous les mois, car les coléoptères carabiques sont des espèces sensibles aux conditions
abiotiques et présentent des variations dans leur période de reproduction. Ainsi, la richesse
spécifique et l’abondance des espèces sont comparées entre les années et les différents mois
échantillonnés. La description de la communauté se réfère également à l’étude de la
répartition des espèces sur la zone d’étude à partir d’une étude de la fréquence d’occurrence
des espèces et d’une analyse de la distribution de la richesse spécifique entre les parcelles.
Afin d’éviter tout biais relié à l’effort d’échantillonnage, seules les cultures présentant un
nombre suffisant de parcelles échantillonnées ont été analysées, et au final deux grandes
catégories de cultures ont pu être étudiées. Le premier type concerne les céréales et le colza,
qui sont des milieux considérés d’une part comme très perturbés d’un point de vue chimique
de par l’application des pesticides et d’autre part comme des milieux instables car les terres
sont labourées tous les ans. Le second type de culture à l’inverse se réfère aux prairies et aux
luzernes qui sont des habitats plus stables dans le temps car le labour n’est pas appliqué
chaque année et ce sont des milieux moins perturbés chimiquement. Les perturbations
physiques (fauche) y sont en revanche plus présentes.
Différents indices de diversité sont calculés afin de comparer la distribution des espèces
entre les cultures. Les indices sont les indices de Shannon, d’équitabilité et de biomasse.
L’indice de Shannon a pour formule : H’=-∑(Ni/N) x log(Ni/N) où Ni est le nombre
d’individus d’une espèce donnée (i allant de 1 à S (nombre total d’espèces)) et N est le
nombre total d’individus. Cet indice est égal à zéro lorsque la population est représentée par
une seule espèce et devient maximal lorsque la population est répartie de manière homogène
entre toutes les espèces. L’Indice d’Evenness ou Indice d’équitabilité est basé sur l’indice de
Shannon. La formule est : H=H’/log(S) où S est le nombre total d’espèces. Cet indice varie
entre 0 et 1, il est maximal lorsque la répartition de la population est homogène entre toutes
les espèces et minimal lorsque une espèce domine (Graham, 2009). Ces deux indices sont
calculés avec le logiciel PAST. En ce qui concerne la biomasse présente dans chaque pot-
piège, elle est calculée à partir de 5 classes de tailles, en multipliant le nombre d'individus de
chaque classe par le poids moyen des individus déterminé avec les équations de Hodar
(1996).

10
L’utilisation des courbes de raréfaction décrites précédemment permettront également de
comparer la diversité maximale entre les cultures.
Au-delà de la simple analyse quantitative des communautés (richesse spécifique), nous
avons également comparé ces communautés du point de vue de leur composition. Pour cette
analyse, les données étant des données de type « présence/absence », une analyse factorielle
des correspondances a été réalisée. Cependant les analyses de type AFC accordent un poids
relativement disproportionné aux espèces atypiques, c'est-à-dire rares ou particulières. Nous
avons donc dans un deuxième temps restreint le jeu de données, en retirant d’abord les
espèces rares, ou en ne conservant que les 10 espèces les plus communes, ou enfin les 10
parcelles (par culture) de plus forte richesse spécifique et en ne gardant que les espèces de
fréquence d’occurrence supérieure à 15 parcelles. Notre objectif était ici de séparer les
communautés par type de culture.
6. Analyse des traits biologiques des espèces
L’analyse des traits des espèces permettra d’expliquer les assemblages observés entre les
cultures. Chaque espèce recensée dans la base de données est caractérisée selon 6 catégories
de traits biologiques (Annexe 2) qui sont les suivantes : le régime alimentaire (généraliste,
spécialiste), le type de proie consommée (carnivore, phytophage, omnivore), la période de
reproduction (printemps, automne), le type d’habitat préférentiel (milieu sec, milieu humide,
milieu rivulaire, culture, milieu boisé), le mode de déplacement (macroptère, brachyptère,
dimorphique) et la taille séparée, en quatre classes (T1 :de 0 à 5 mm, T2 :de 5 à 10 mm, T3 :
de 10 à 15 mm, T4 :de 15 à 20 mm). Les informations ont été obtenues à partir d’articles
scientifiques (Purtauf et al., 2005 ; Desender et al., 2010), de livres de biologie des Carabidae
(Dajoz, 2002 ; Holland, 2002) ou encore de livres d’identification (Forel & Leplat, 2003a et
b).
Une analyse des correspondances multiples (ACM) est réalisée entre les espèces et leurs
traits biologiques. Seules les espèces renseignées pour chacune des variables sont gardées
pour réaliser cette analyse ce qui regroupe 55 espèces. Puis une analyse canonique (ACC) des
correspondances est réalisée entre la fréquence d’occurrence des espèces selon les cultures et
leurs traits d’histoire de vie afin de déterminer si les assemblages d’espèces entre les cultures
diffèrent selon les traits biologiques des espèces.
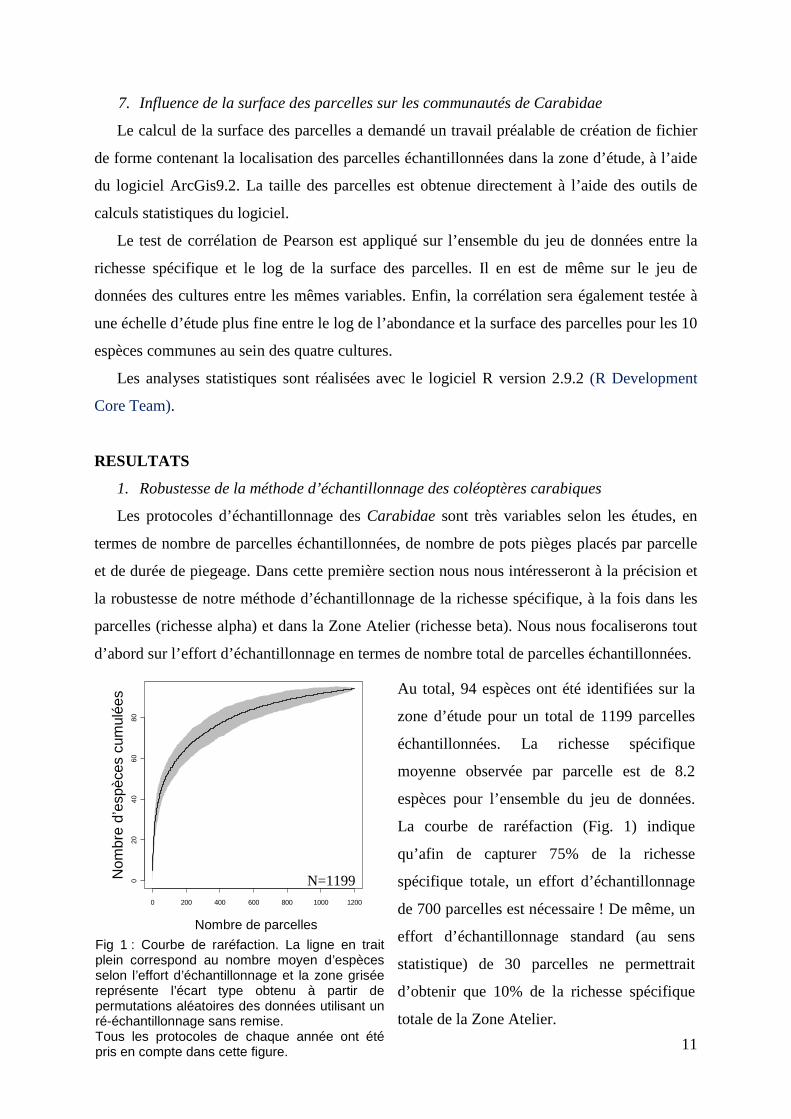
11
0 200 400 600 800 1000 1200
020
4060
80
7. Influence de la surface des parcelles sur les communautés de Carabidae
Le calcul de la surface des parcelles a demandé un travail préalable de création de fichier
de forme contenant la localisation des parcelles échantillonnées dans la zone d’étude, à l’aide
du logiciel ArcGis9.2. La taille des parcelles est obtenue directement à l’aide des outils de
calculs statistiques du logiciel.
Le test de corrélation de Pearson est appliqué sur l’ensemble du jeu de données entre la
richesse spécifique et le log de la surface des parcelles. Il en est de même sur le jeu de
données des cultures entre les mêmes variables. Enfin, la corrélation sera également testée à
une échelle d’étude plus fine entre le log de l’abondance et la surface des parcelles pour les 10
espèces communes au sein des quatre cultures.
Les analyses statistiques sont réalisées avec le logiciel R version 2.9.2 (R Development
Core Team).
RESULTATS
1. Robustesse de la méthode d’échantillonnage des coléoptères carabiques
Les protocoles d’échantillonnage des Carabidae sont très variables selon les études, en
termes de nombre de parcelles échantillonnées, de nombre de pots pièges placés par parcelle
et de durée de piegeage. Dans cette première section nous nous intéresseront à la précision et
la robustesse de notre méthode d’échantillonnage de la richesse spécifique, à la fois dans les
parcelles (richesse alpha) et dans la Zone Atelier (richesse beta). Nous nous focaliserons tout
d’abord sur l’effort d’échantillonnage en termes de nombre total de parcelles échantillonnées.
Fig 1 : Courbe de raréfaction. La ligne en trait plein correspond au nombre moyen d’espèces selon l’effort d’échantillonnage et la zone grisée représente l’écart type obtenu à partir de permutations aléatoires des données utilisant un ré-échantillonnage sans remise. Tous les protocoles de chaque année ont été pris en compte dans cette figure.
Nombre de parcelles
Nom
bre
d’es
pèce
s cu
mul
ées Au total, 94 espèces ont été identifiées sur la
zone d’étude pour un total de 1199 parcelles
échantillonnées. La richesse spécifique
moyenne observée par parcelle est de 8.2
espèces pour l’ensemble du jeu de données.
La courbe de raréfaction (Fig. 1) indique
qu’afin de capturer 75% de la richesse
spécifique totale, un effort d’échantillonnage
de 700 parcelles est nécessaire ! De même, un
effort d’échantillonnage standard (au sens
statistique) de 30 parcelles ne permettrait
d’obtenir que 10% de la richesse spécifique
totale de la Zone Atelier.
N=1199
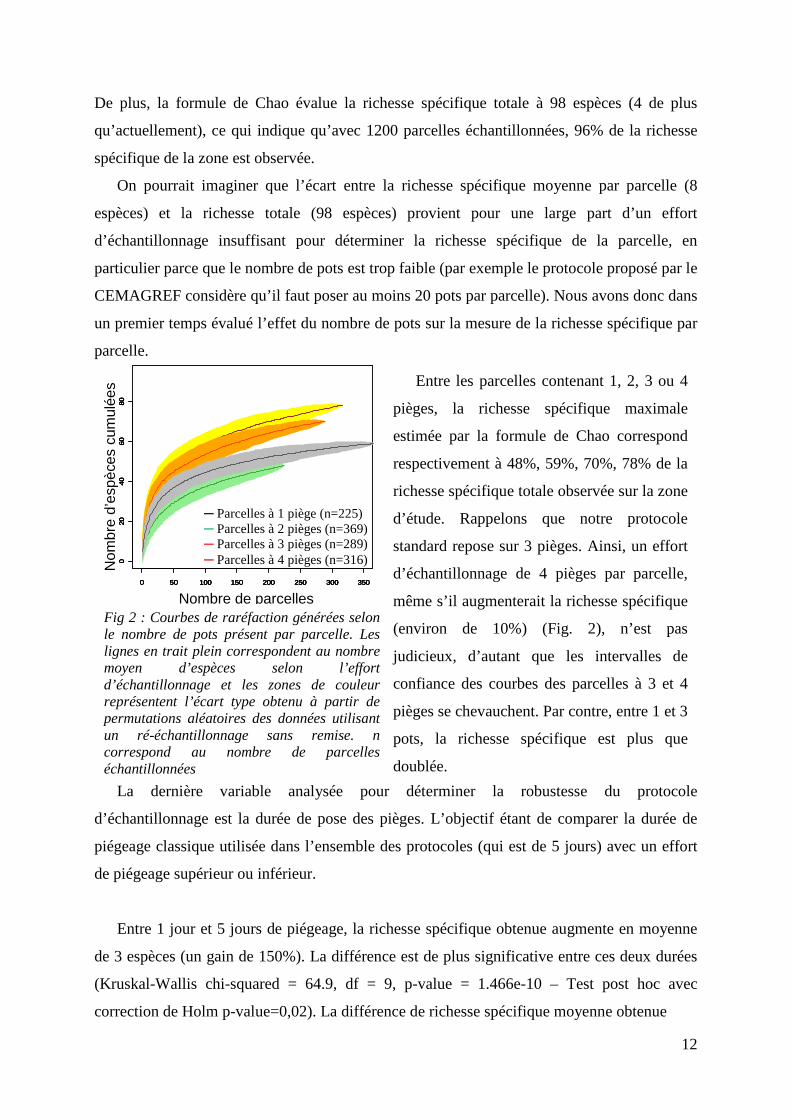
12
0 50 100 150 200 250 300 350
020
4060
80
0 50 100 150 200 250 300 350
020
4060
80
0 50 100 150 200 250 300 350
020
4060
80
0 50 100 150 200 250 300 350
020
4060
80
De plus, la formule de Chao évalue la richesse spécifique totale à 98 espèces (4 de plus
qu’actuellement), ce qui indique qu’avec 1200 parcelles échantillonnées, 96% de la richesse
spécifique de la zone est observée.
On pourrait imaginer que l’écart entre la richesse spécifique moyenne par parcelle (8
espèces) et la richesse totale (98 espèces) provient pour une large part d’un effort
d’échantillonnage insuffisant pour déterminer la richesse spécifique de la parcelle, en
particulier parce que le nombre de pots est trop faible (par exemple le protocole proposé par le
CEMAGREF considère qu’il faut poser au moins 20 pots par parcelle). Nous avons donc dans
un premier temps évalué l’effet du nombre de pots sur la mesure de la richesse spécifique par
parcelle.
La dernière variable analysée pour déterminer la robustesse du protocole
d’échantillonnage est la durée de pose des pièges. L’objectif étant de comparer la durée de
piégeage classique utilisée dans l’ensemble des protocoles (qui est de 5 jours) avec un effort
de piégeage supérieur ou inférieur.
Entre 1 jour et 5 jours de piégeage, la richesse spécifique obtenue augmente en moyenne
de 3 espèces (un gain de 150%). La différence est de plus significative entre ces deux durées
(Kruskal-Wallis chi-squared = 64.9, df = 9, p-value = 1.466e-10 – Test post hoc avec
correction de Holm p-value=0,02). La différence de richesse spécifique moyenne obtenue
Nombre de parcelles
Nom
bre
d’es
pèce
s cu
mul
ées
Fig 2 : Courbes de raréfaction générées selon le nombre de pots présent par parcelle. Les lignes en trait plein correspondent au nombre moyen d’espèces selon l’effort d’échantillonnage et les zones de couleur représentent l’écart type obtenu à partir de permutations aléatoires des données utilisant un ré-échantillonnage sans remise. n correspond au nombre de parcelles échantillonnées
─ Parcelles à 1 piège (n=225) ─ Parcelles à 2 pièges (n=369) ─ Parcelles à 3 pièges (n=289) ─ Parcelles à 4 pièges (n=316)
Entre les parcelles contenant 1, 2, 3 ou 4
pièges, la richesse spécifique maximale
estimée par la formule de Chao correspond
respectivement à 48%, 59%, 70%, 78% de la
richesse spécifique totale observée sur la zone
d’étude. Rappelons que notre protocole
standard repose sur 3 pièges. Ainsi, un effort
d’échantillonnage de 4 pièges par parcelle,
même s’il augmenterait la richesse spécifique
(environ de 10%) (Fig. 2), n’est pas
judicieux, d’autant que les intervalles de
confiance des courbes des parcelles à 3 et 4
pièges se chevauchent. Par contre, entre 1 et 3
pots, la richesse spécifique est plus que
doublée.
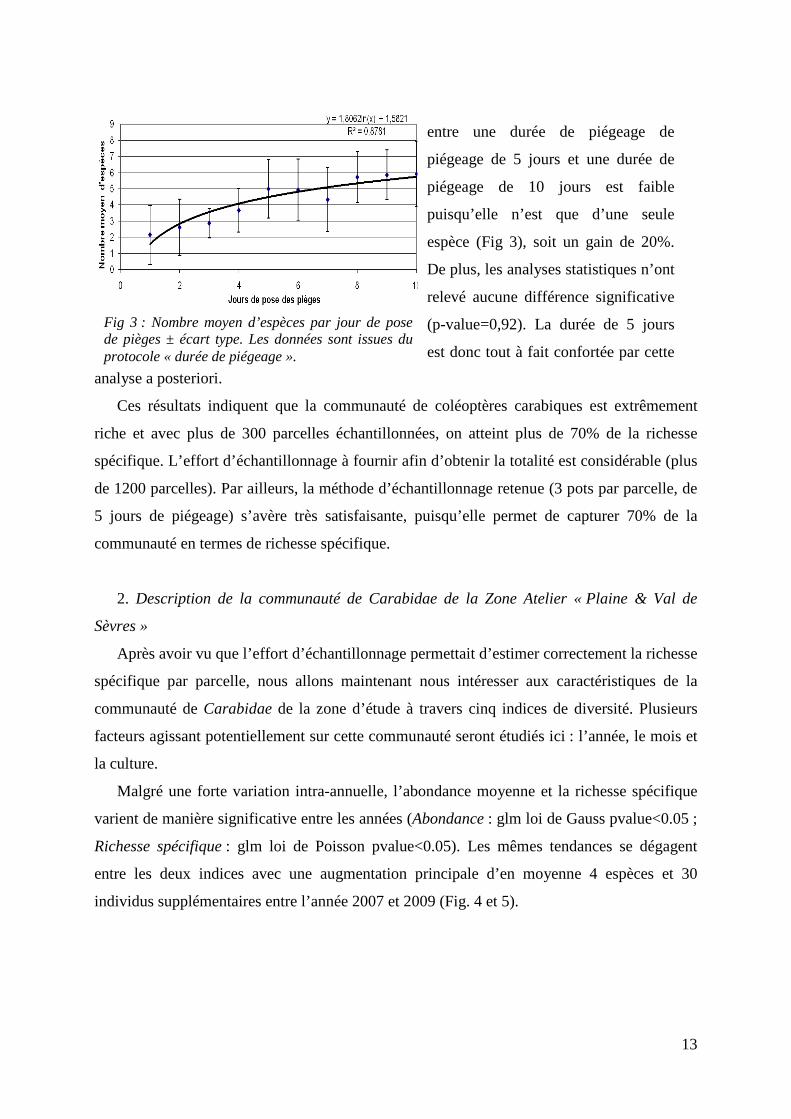
13
analyse a posteriori.
Ces résultats indiquent que la communauté de coléoptères carabiques est extrêmement
riche et avec plus de 300 parcelles échantillonnées, on atteint plus de 70% de la richesse
spécifique. L’effort d’échantillonnage à fournir afin d’obtenir la totalité est considérable (plus
de 1200 parcelles). Par ailleurs, la méthode d’échantillonnage retenue (3 pots par parcelle, de
5 jours de piégeage) s’avère très satisfaisante, puisqu’elle permet de capturer 70% de la
communauté en termes de richesse spécifique.
2. Description de la communauté de Carabidae de la Zone Atelier « Plaine & Val de
Sèvres »
Après avoir vu que l’effort d’échantillonnage permettait d’estimer correctement la richesse
spécifique par parcelle, nous allons maintenant nous intéresser aux caractéristiques de la
communauté de Carabidae de la zone d’étude à travers cinq indices de diversité. Plusieurs
facteurs agissant potentiellement sur cette communauté seront étudiés ici : l’année, le mois et
la culture.
Malgré une forte variation intra-annuelle, l’abondance moyenne et la richesse spécifique
varient de manière significative entre les années (Abondance : glm loi de Gauss pvalue<0.05 ;
Richesse spécifique : glm loi de Poisson pvalue<0.05). Les mêmes tendances se dégagent
entre les deux indices avec une augmentation principale d’en moyenne 4 espèces et 30
individus supplémentaires entre l’année 2007 et 2009 (Fig. 4 et 5).
Fig 3 : Nombre moyen d’espèces par jour de pose de pièges ± écart type. Les données sont issues du protocole « durée de piégeage ».
entre une durée de piégeage de
piégeage de 5 jours et une durée de
piégeage de 10 jours est faible
puisqu’elle n’est que d’une seule
espèce (Fig 3), soit un gain de 20%.
De plus, les analyses statistiques n’ont
relevé aucune différence significative
(p-value=0,92). La durée de 5 jours
est donc tout à fait confortée par cette
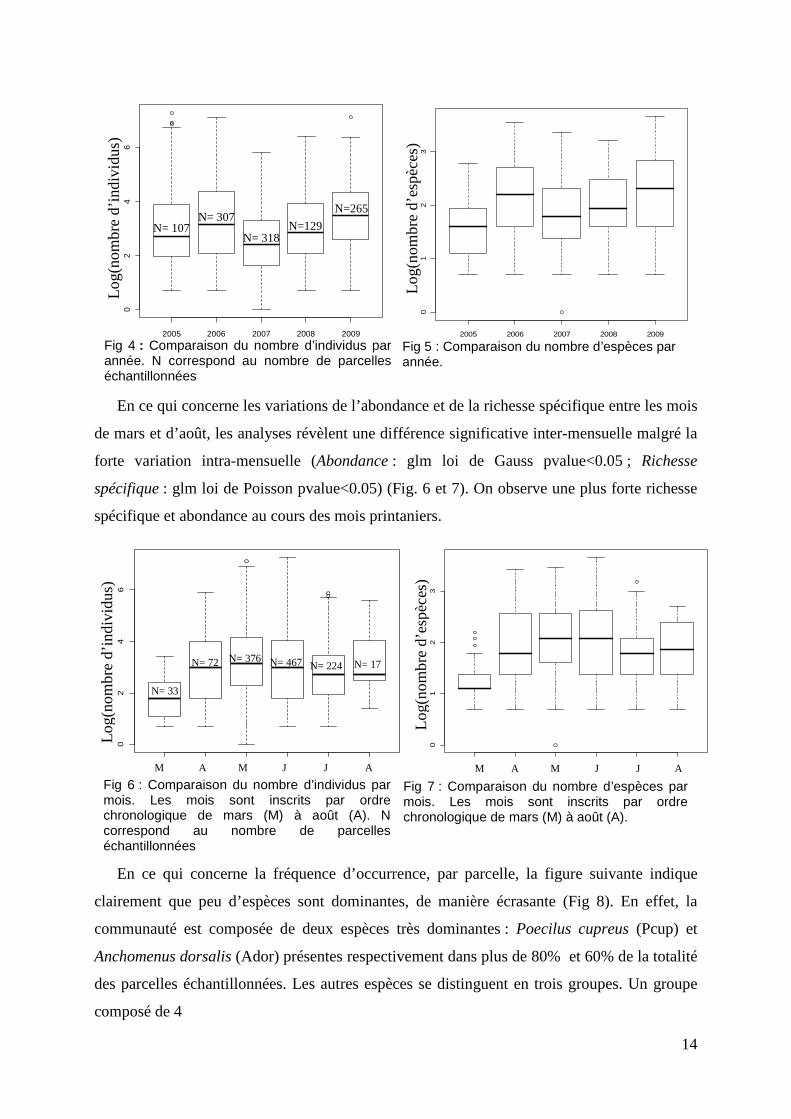
14
02
46
En ce qui concerne les variations de l’abondance et de la richesse spécifique entre les mois
de mars et d’août, les analyses révèlent une différence significative inter-mensuelle malgré la
forte variation intra-mensuelle (Abondance : glm loi de Gauss pvalue<0.05 ; Richesse
spécifique : glm loi de Poisson pvalue<0.05) (Fig. 6 et 7). On observe une plus forte richesse
spécifique et abondance au cours des mois printaniers.
En ce qui concerne la fréquence d’occurrence, par parcelle, la figure suivante indique
clairement que peu d’espèces sont dominantes, de manière écrasante (Fig 8). En effet, la
communauté est composée de deux espèces très dominantes : Poecilus cupreus (Pcup) et
Anchomenus dorsalis (Ador) présentes respectivement dans plus de 80% et 60% de la totalité
des parcelles échantillonnées. Les autres espèces se distinguent en trois groupes. Un groupe
composé de 4
Fig 6 : Comparaison du nombre d’individus par mois. Les mois sont inscrits par ordre chronologique de mars (M) à août (A). N correspond au nombre de parcelles échantillonnées
M A M J J A M A M J J A
Fig 7 : Comparaison du nombre d’espèces par mois. Les mois sont inscrits par ordre chronologique de mars (M) à août (A).
Fig 4 : Comparaison du nombre d’individus par année. N correspond au nombre de parcelles échantillonnées
Fig 5 : Comparaison du nombre d’espèces par année.
Log(
nom
bre
d’es
pèce
s)
Log(
nom
bre
d’in
divi
dus)
2005 2006 2007 2008 2009
02
46
N= 107 N= 318
N= 307 N=129
N=265
2005 2006 2007 2008 2009
01
23
N= 33
N= 72 N= 376 N= 467 N= 224 N= 17
Log(
nom
bre
d’in
divi
dus)
Log(
nom
bre
d’es
pèce
s) 0
12
3
15
0
10
20
30
40
50
60
70
80
90
100
Pcu
p
Ador
Bsc
l
Oru
f
Hdim
Tqua
Cfu
s
Bcr
e
Haf
f
Mlu
c
Aae
n
Nbre
Pvu
l
Pm
ad
Lqua
Cch
r
Oaz
u
Nqua
Lpil
Hsm
a
Afu
l
Am
on
Mm
in
Nru
f
Zten
Agl
a
Aeur
Tobt
Hdis
Pir
i
Ola
t
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
0.00
0.05
0.1
00.
15
d’occurrence inférieure à 10%.
D’après la figure 9, la majorité des parcelles possèdent une faible richesse spécifique. En
effet, plus de 80% d’entre elles possèdent moins de 7 espèces. La valeur moyenne étant de
6,67 espèces pour les parcelles contenant 3 ou 4 pots. Cette distribution suit une loi de
présentes dans moins de 5% des parcelles, et 51 espèces totalisent moins de 1% des captures).
Enfin, le nombre d’espèces par parcelle suit une loi de Poisson, distribution aléatoire attendue
pour ce type de variable, même si nous avons mis en évidence des effets de l’année et des
saisons.
3. Comparaison des communautés de coléoptères carabiques entre les cultures
Les coléoptères carabiques présentant des exigences alimentaires et des préférendum
d’habitat stricts pour la plupart des espèces, nous allons maintenant nous intéresser à la
distribution des coléoptères carabiques à une échelle d’analyse plus fine, celle de l’habitat et
Fig 8 : Fréquence d’occurrence par parcelle des 31 espèces les plus communes. Seules les parcelles contenant 3 ou 4 pièges sont utilisées.
Nom des espèces
Fré
quen
ce d
’occ
urre
nce
(%)
Nombre d’espèces
Nom
bre
de p
arce
lle
(fré
quen
ce)
espèces (soit 4,3% de la richesse
spécifique totale) qui présente une
fréquence d’occurrence comprise
entre 35 et 45%. Le deuxième
groupe se réfère à 12,8% des
espèces de fréquence d’occurrence
variant de 10% et 25%. Enfin, le
dernier groupe comprend plus de
74% des espèces de fréquence
Fig 9 : Distribution des fréquences d’occurrence des espèces en pourcentage de parcelles. La courbe bleue représente la distribution théorique de la loi de Poisson de moyenne 6,67. Seules les parcelles comprenant 3 ou 4 pièges sont utilisées.
Poisson. La richesse spécifique par
parcelle est donc aléatoire, autour d’une
valeur moyenne de 7-8 espèces. Ainsi,
cette communauté de Carabidae est
composée de deux espèces ultra-
dominantes, puis de 3 espèces dominantes.
A elles seules, ces 5 espèces totalisent
90% du total des captures. Viennent
ensuite un très grand nombre d’espèces
rares voire très rares (19 espèces sont
16
0
100
200
300
400
500
600
Cer Luz Prai LMT Col LP Jac RG
No
mb
reto
tal
de p
arc
elles
échan
tillo
nnées
Cultures
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
Col Luz Prai Cer LP LMT Jac RG
Tau
x d
'occ
up
atio
n d
es
cultu
res
Cultures
Col
JachCerLMT
Luz
PraiLP
RG
ColJach
Cer
LMTLuz
Prai
LP
RG
y = 1,415x + 0,4327R² = 0,7814
y = 0,9374x + 1,0193R² = 0,8785
0
0,5
1
1,5
2
2,5
0
0,5
1
1,5
2
2,5
0 0,2 0,4 0,6 0,8 1 1,2
log(
Biom
asse
moy
enne
)
log
(nom
bre
moy
en d
'indi
vidus
)
log(nombre moyen d'espèces)
Nombre moyen d'individusBiomasse moyenne
plus particulièrement à l’échelle des cultures. Nous nous intéresserons d’abord à une
description de la communauté des Carabidae parmi toutes les cultures échantillonnées, puis
nous focaliserons notre analyse sur les quatre cultures à la fois les plus riches et les mieux
échantillonnées : céréales, colza, luzernes et prairies.
Le nombre total de parcelles échantillonnées varie fortement selon les cultures (Fig 10).
En effet, l’effort d’échantillonnage est presque deux fois plus fort pour les céréales, avec 528
parcelles échantillonnées, que pour les luzernes et plus de cinq fois supérieure pour les autres
cultures. Cependant pour chaque culture, les coléoptères carabiques sont récoltés dans plus de
60% des parcelles (Fig. 11). Ainsi les coléoptères carabiques sont non seulement présents,
mais dominants, dans chacune des cultures échantillonnées.
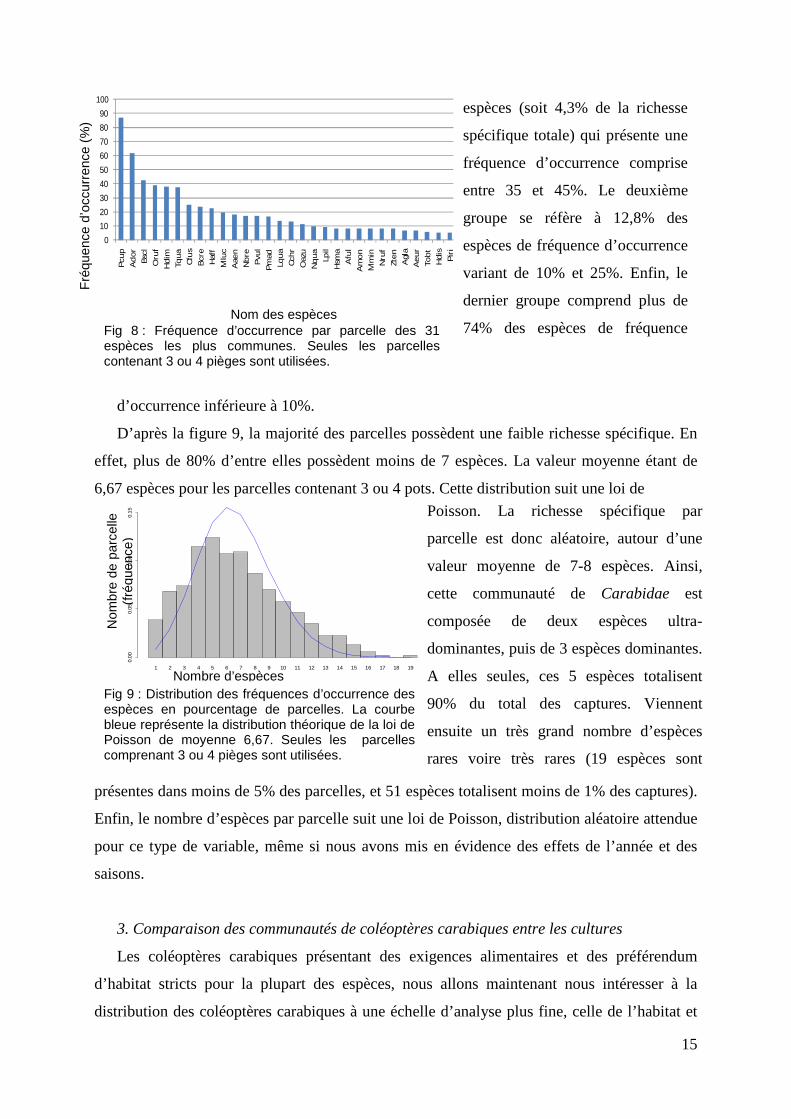
Fig 10 : A gauche - Nombre total de parcelles échantillonnées par culture. Pour des raisons de lisibilité le nom des cultures est abrégé de la manière suivante : Cer : céréales, Luz : luzernes, Prai : praries, LMT : labour-maïs-tournesol, Col : colza, LP : lin-pois, Jac : jachère et RG : ray-grass. Fig 11 : A droite - Taux de parcelles occupées par les coléoptères carabiques. Rapport entre le nombre de parcelles occupées sur le nombre total de parcelles par culture.
Nous nous sommes également
intéressés à la relation entre
richesse spécifique et abondance
(ou biomasse totale), par parcelle
et entre cultures. Le nombre
d’espèces et la biomasse sont
fortement corrélés (t = 58.448, df
= 1187, p-value < 2.2e-16). Il en
est de même avec le nombre
d’individus (t = 22.6101, df =
1187, p-value < 2.2e-16). Ce
pendant, la figure 12 nous indique
Fig12 - Corrélation entre le nombre moyen d’espèces par parcelle avec le nombre moyen d’individus ainsi qu’avec la biomasse moyenne par parcelle par culture. Pour des raisons de lisibilité le nom des cultures est abrégé de la manière suivante : Cer : céréales, Luz : luzerne, Prai : Praire, LMT : labour-maïs-tournesol, Col : colza, LP : lin-pois, Jach : jachère et RG : ray-grass
17
0
20
40
60
80
100
120
Cer Col Luz Prai
Nom
bre
moy
en d
'esp
èces
Cultures
Richesse spécifique moyenne observée Richesse spécifique maximale estimée par la formule Chao
Cer Col Jach LMT LP Luz Prai RG
0.0
0.5
1.0
1.5
2.0
Cultures
Indi
ce d
e S
hann
on
Cer Col Jach LMT LP Luz Prai RG
0.2
0.4
0.6
0.8
1.0
Cultures
Equ
itabi
lité
que les prairies abritent plutôt des espèces de grande taille, que Lin-Pois et Colza abritent
beaucoup d’individus plutôt de petite taille comparativement à leur richesse spécifique et
qu’enfin céréales et jachères sont plutôt riches en espèces. Colza abritent beaucoup
d’individus plutôt de petite taille comparativement à leur richesse spécifique et qu’enfin
céréales et jachères sont plutôt riches en espèces.
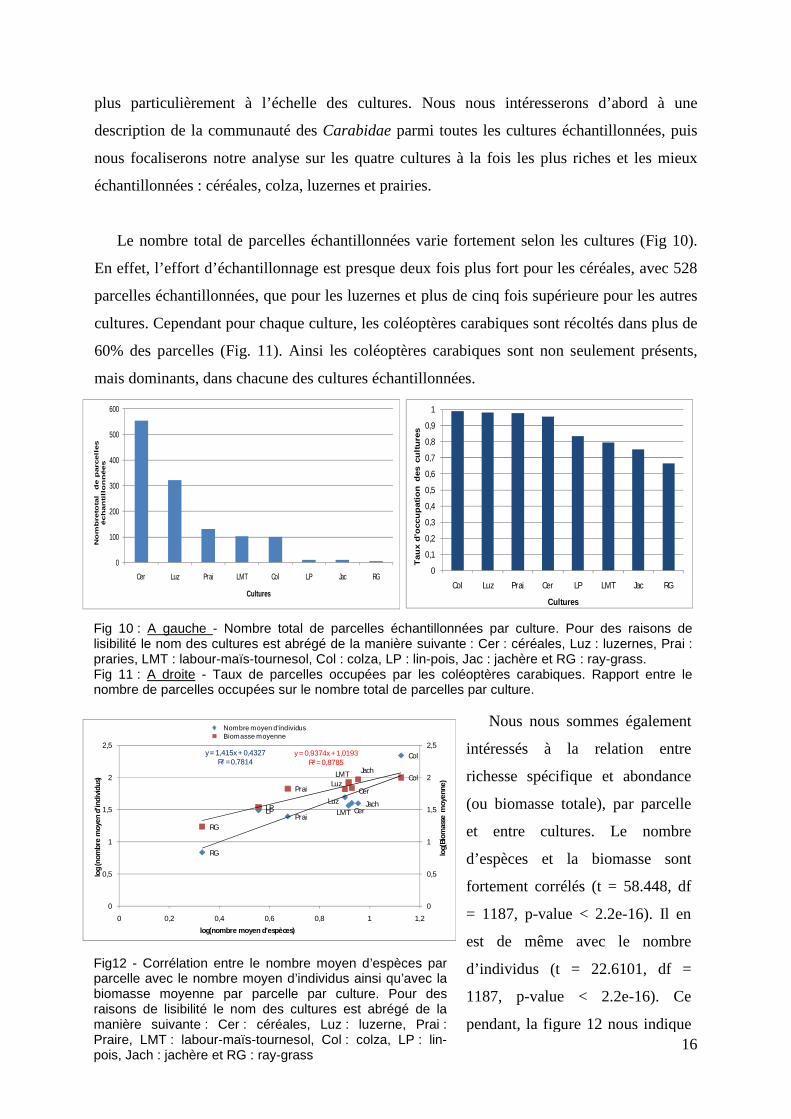
La richesse spécifique moyenne varie très peu entre les quatre cultures considérées et
correspond à une valeur moyenne de 7 espèces pour les parcelles contenant 3 ou 4 pots(Fig
13). De plus, cette valeur est très éloignée de la richesse spécifique totale de la communauté.
Les parcelles sont donc pauvres en espèces comparées au potentiel de la communauté totale.
Les analyses révèlent une différence significative de l’indice de Shannon entre culture
(glm loi de Gauss : F-statistic: 11.94 on 7 and 1181 DF, p-value: 9.028e-15). Il en est de
même pour l’équitabilité entre les cultures (glm loi de Gauss : F-statistic: 19.83 on 7 and 1181
DF, p-value: < 2.2e-16) (Fig 14 & 15). Les valeurs d’équitabilité, étant comprises entre 0,4 et
1, mettent en évidence que la répartition des espèces de Carabidae est plutôt homogène au
sein des cultures et entre les cultures.
Fig 13 : Comparaison entre la richesse spécifique moyenne observée et la richesse estimée par la formule de Chao. Pour des raisons de lisibilité le nom des cultures est abrégé de la manière suivante : Cer : céréales, Luz : luzernes, Prai : prairie, Col : colza
N=528
N=99
N=9
N=82
N=10
N=315
N=131
N=21
Fig 14 : Comparaison de l’indice de Shannon par culture. N correspond au nombre de parcelles échantillonnées
Fig 15 : Comparaison de l’équitabilité par culture
18
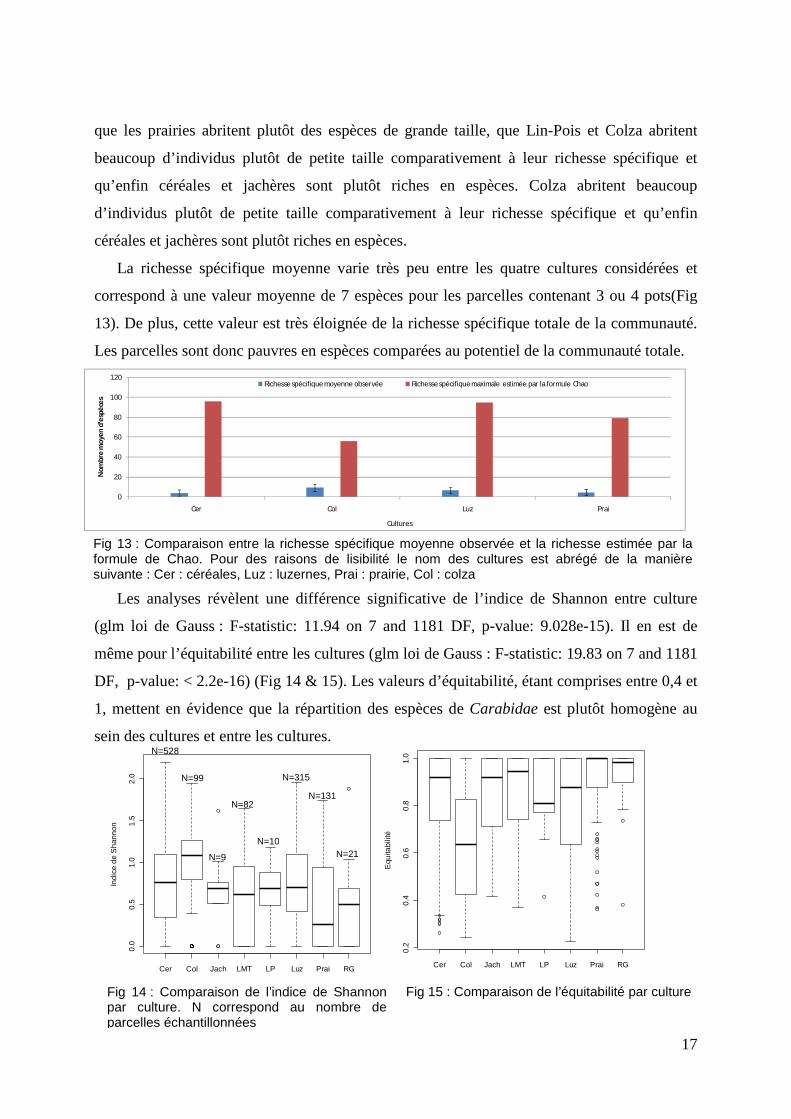
Les courbes de raréfaction permettent de comparer la richesse spécifique entre les
différentes cultures malgré la différence d’effort d’échantillonnage. De plus, la formule de
Chao donne une idée de la richesse spécifique maximale au sein de chacune de ces cultures.
La formule de Chao estime la richesse spécifique maximale à 96 espèces pour les céréales, 95
pour les luzernes, 79 pour les prairies et à 57 pour le colza. Ainsi la richesse spécifique
estimée varie fortement entre ces cultures. Cependant, l’effort d’échantillonnage effectué pour
chaque culture permet d’estimer plus de 70% de la richesse spécifique observée pour le milieu
considéré.
La richesse spécifique moyenne par parcelle entre les cultures est proche mais cela ne
présume en rien de la similarité entre les communautés de ces milieux. Ainsi, la suite des
analyses consiste à comparer les assemblages d’espèces entre les cultures à partir d’AFC. Sur
le jeu de données regroupant les dix espèces de plus forte fréquence d’occurrence ainsi que
sur les données rassemblant les 10 parcelles de plus forte richesse spécifique contenant
uniquement les espèces de fréquence d’occurrence supérieure à 15. Le même code couleur est
utilisé pour les représentations graphiques. Le vert correspondra aux luzernes, le rouge aux
céréales, le bleu au colza et le orange aux prairies.
Nombre de parcelles
Nom
bre
d’es
pèce
s cu
mul
ées
Fig 16 : Courbes de raréfaction générées par culture. Les lignes en trait plein correspondent au nombre moyen d’espèces selon l’effort d’échantillonnage et les zones de couleur représentent l’écart type obtenu à partir de permutations aléatoires des données utilisant un ré-échantillonnage sans remise. n correspond au nombre de parcelles total échantillonnées et s à la richesse spécifique observée.
Les luzernes et les céréales présentent une
richesse spécifique très proche et sont deux fois
plus riches que le colza et les prairies. Aucune
culture n’a atteint la richesse spécifique observée
sur la zone d’étude (Fig. 16). La richesse
spécifique estimée pour les céréales présente une
valeur supérieure à celle estimée pour l’ensemble
de la zone d’étude de la richesse spécifique (78
espèces). Les cultures présentent une richesse
spécifique très variable les unes par rapport aux
autres. Cependant la répartition des espèces au
sein des parcelles est assez similaire avec peu
d’espèces mais réparties équitablement. Enfin, la
richesse spécifique intra parcelle est très éloignée
de la richesse de la zone d’étude et de la culture
considérée.
─ Céréales (n=528, s=79) ─ Colza (n=99, s=44) ─ Luzerne (n=315, s=70) ─ Prairie (n=131, s=59)
0 100 200 300 400 500
020
4060
80
rand
om
0 100 200 300 400 500
020
4060
80
rand
om
0 100 200 300 400 500
020
4060
80
rand
om
0 100 200 300 400 500
020
4060
80
rand
om
19
d = 1
Cer Col Luz Prai
d = 1 d = 1
Ador Bcre
Bscl Cfus Haff
Hdim
Mluc
Oruf
Pcup Tqua
d = 0.5
Cer
Col Luz Prai
d = 1 d = 1
Aaen
Ador
Bcre
Bscl Cfus
Cchr
Haff
Hdim
Hsma
Lqua
Mluc
Oazu Oruf
Pcup
Pmad
Pvul Tqua
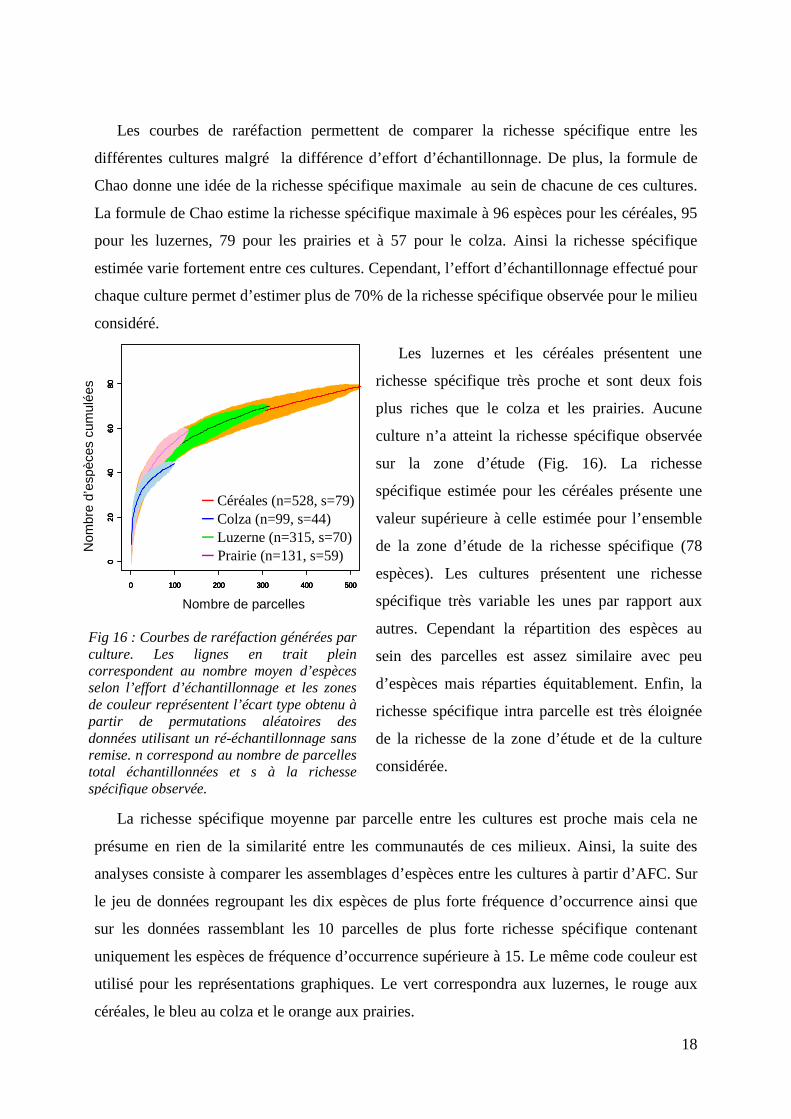
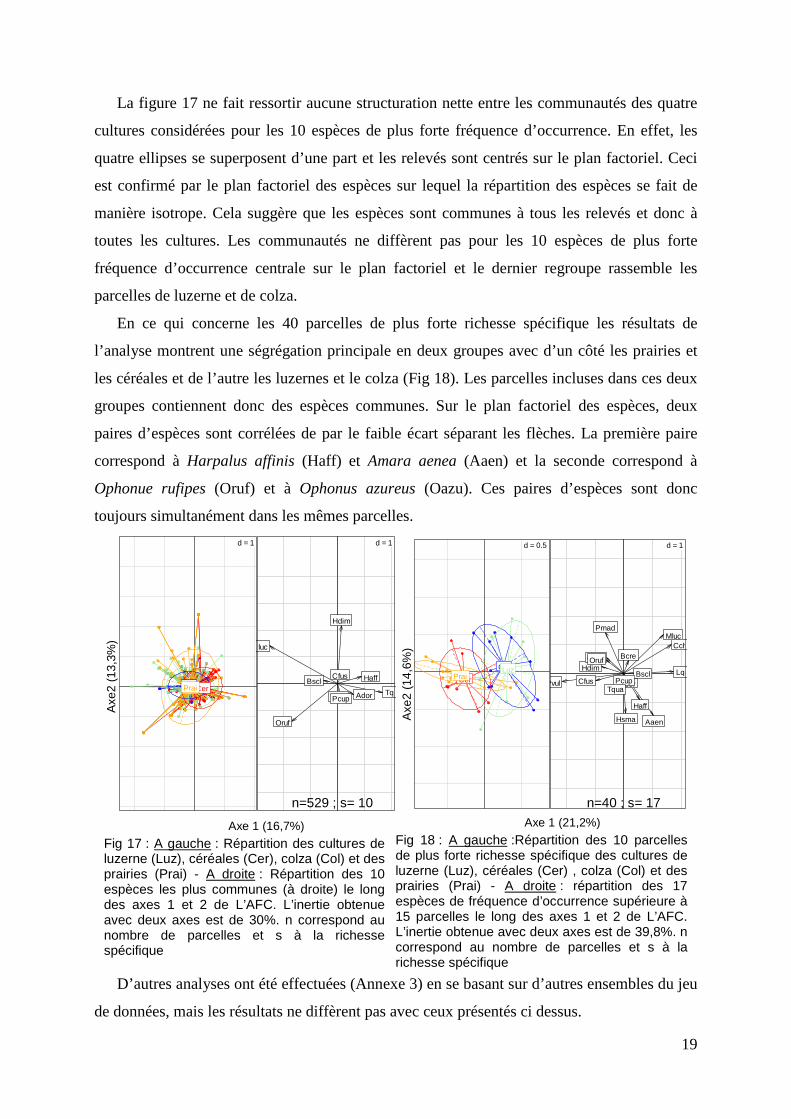
La figure 17 ne fait ressortir aucune structuration nette entre les communautés des quatre
cultures considérées pour les 10 espèces de plus forte fréquence d’occurrence. En effet, les
quatre ellipses se superposent d’une part et les relevés sont centrés sur le plan factoriel. Ceci
est confirmé par le plan factoriel des espèces sur lequel la répartition des espèces se fait de
manière isotrope. Cela suggère que les espèces sont communes à tous les relevés et donc à
toutes les cultures. Les communautés ne diffèrent pas pour les 10 espèces de plus forte
fréquence d’occurrence centrale sur le plan factoriel et le dernier regroupe rassemble les
parcelles de luzerne et de colza.
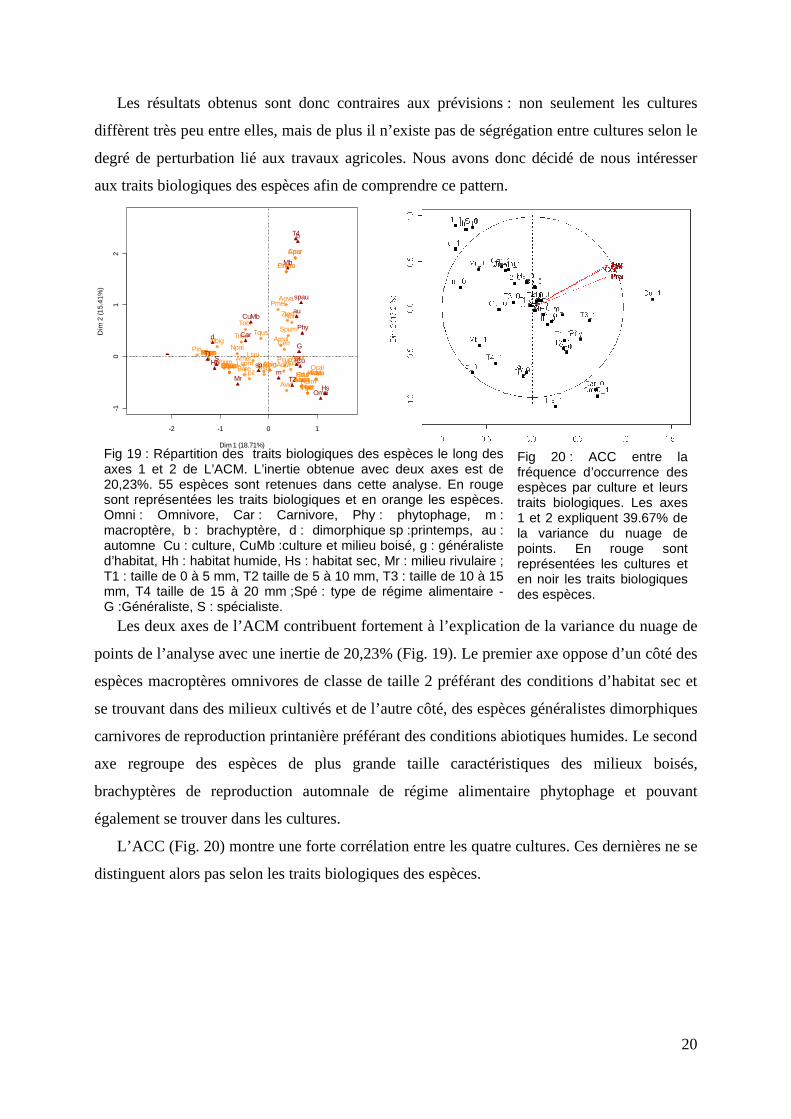
En ce qui concerne les 40 parcelles de plus forte richesse spécifique les résultats de
l’analyse montrent une ségrégation principale en deux groupes avec d’un côté les prairies et
les céréales et de l’autre les luzernes et le colza (Fig 18). Les parcelles incluses dans ces deux
groupes contiennent donc des espèces communes. Sur le plan factoriel des espèces, deux
paires d’espèces sont corrélées de par le faible écart séparant les flèches. La première paire
correspond à Harpalus affinis (Haff) et Amara aenea (Aaen) et la seconde correspond à
Ophonue rufipes (Oruf) et à Ophonus azureus (Oazu). Ces paires d’espèces sont donc
toujours simultanément dans les mêmes parcelles.
D’autres analyses ont été effectuées (Annexe 3) en se basant sur d’autres ensembles du jeu
de données, mais les résultats ne diffèrent pas avec ceux présentés ci dessus.
Fig 17 : A gauche : Répartition des cultures de luzerne (Luz), céréales (Cer), colza (Col) et des prairies (Prai) - A droite : Répartition des 10 espèces les plus communes (à droite) le long des axes 1 et 2 de L’AFC. L’inertie obtenue avec deux axes est de 30%. n correspond au nombre de parcelles et s à la richesse spécifique
Axe 1 (16,7%)
Axe
2 (1
3,3%
)
Axe
2 (1
4,6%
)
n=529 ; s= 10
Fig 18 : A gauche :Répartition des 10 parcelles de plus forte richesse spécifique des cultures de luzerne (Luz), céréales (Cer) , colza (Col) et des prairies (Prai) - A droite : répartition des 17 espèces de fréquence d’occurrence supérieure à 15 parcelles le long des axes 1 et 2 de L’AFC. L’inertie obtenue avec deux axes est de 39,8%. n correspond au nombre de parcelles et s à la richesse spécifique
Axe 1 (21,2%)
n=40 ; s= 17
20
Les résultats obtenus sont donc contraires aux prévisions : non seulement les cultures
diffèrent très peu entre elles, mais de plus il n’existe pas de ségrégation entre cultures selon le
degré de perturbation lié aux travaux agricoles. Nous avons donc décidé de nous intéresser
aux traits biologiques des espèces afin de comprendre ce pattern.
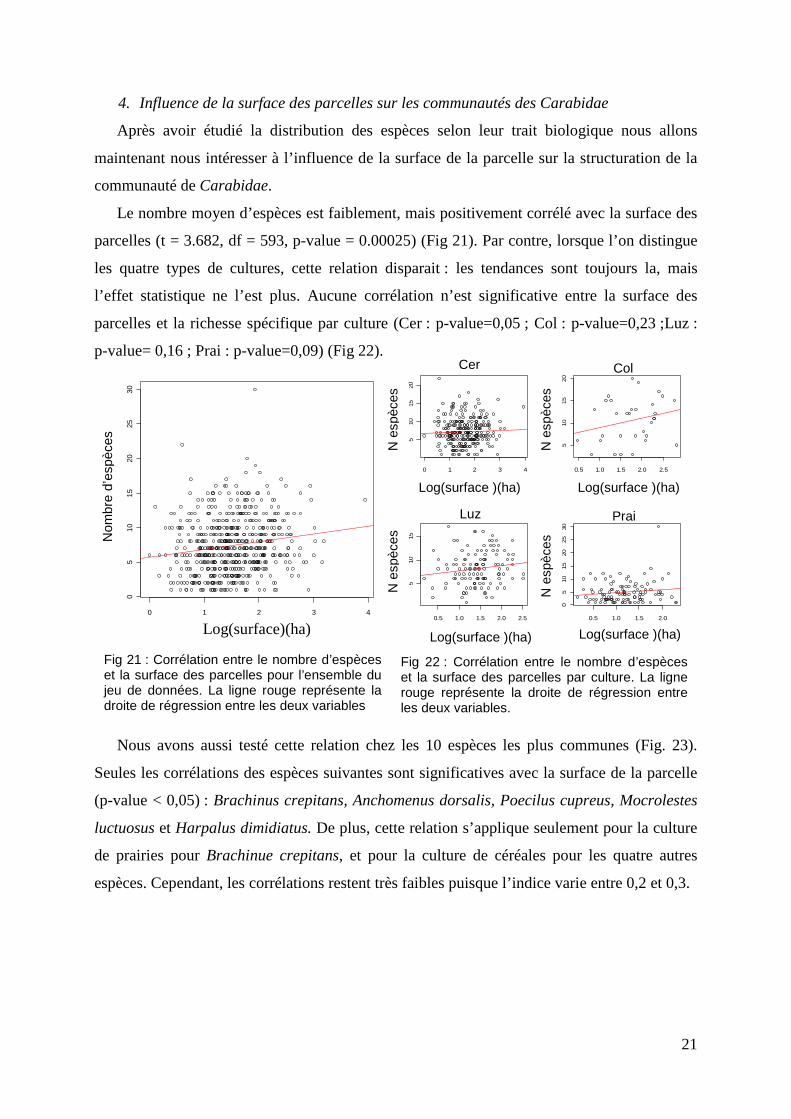
Les deux axes de l’ACM contribuent fortement à l’explication de la variance du nuage de
points de l’analyse avec une inertie de 20,23% (Fig. 19). Le premier axe oppose d’un côté des
espèces macroptères omnivores de classe de taille 2 préférant des conditions d’habitat sec et
se trouvant dans des milieux cultivés et de l’autre côté, des espèces généralistes dimorphiques
carnivores de reproduction printanière préférant des conditions abiotiques humides. Le second
axe regroupe des espèces de plus grande taille caractéristiques des milieux boisés,
brachyptères de reproduction automnale de régime alimentaire phytophage et pouvant
également se trouver dans les cultures.
L’ACC (Fig. 20) montre une forte corrélation entre les quatre cultures. Ces dernières ne se
distinguent alors pas selon les traits biologiques des espèces.
Fig 19 : Répartition des traits biologiques des espèces le long des axes 1 et 2 de L’ACM. L’inertie obtenue avec deux axes est de 20,23%. 55 espèces sont retenues dans cette analyse. En rouge sont représentées les traits biologiques et en orange les espèces. Omni : Omnivore, Car : Carnivore, Phy : phytophage, m : macroptère, b : brachyptère, d : dimorphique sp :printemps, au : automne Cu : culture, CuMb :culture et milieu boisé, g : généraliste d’habitat, Hh : habitat humide, Hs : habitat sec, Mr : milieu rivulaire ; T1 : taille de 0 à 5 mm, T2 taille de 5 à 10 mm, T3 : taille de 10 à 15 mm, T4 taille de 15 à 20 mm ;Spé : type de régime alimentaire - G :Généraliste, S : spécialiste.
Fig 20 : ACC entre la fréquence d’occurrence des espèces par culture et leurs traits biologiques. Les axes 1 et 2 expliquent 39.67% de la variance du nuage de points. En rouge sont représentées les cultures et en noir les traits biologiques des espèces.
-2 -1 0 1
-10
12
Dim 1 (18.71%)
Dim
2 (1
5.41
%)
Aaen
AbinAcon
AdorAeur
Aful
Amer
Amon
Amul
Aova
Apar
AsimAvid
BcreBhumBillBlunBqumBqup Bscl
Cerr
Cnem
Cnig
Cpur
Datr
Dger
Haff
HattHdim
HdisHsma
Htar
LcomLpil
Lqua
LspiMlucMmin
Nbig
Nbre
NpalNqua
OcalOruf
PbigPcup
Piri
Pmau
Pmel
SpumTnan
TobtTqua
Zten
Car
Omni
Phy
b
d
m
au
sp
spau
Cu
CuMb
Hh
Hs
Hsa
Mb
Mr
T1
T2
T3
T4
G
S
21
4. Influence de la surface des parcelles sur les communautés des Carabidae
Après avoir étudié la distribution des espèces selon leur trait biologique nous allons
maintenant nous intéresser à l’influence de la surface de la parcelle sur la structuration de la
communauté de Carabidae.
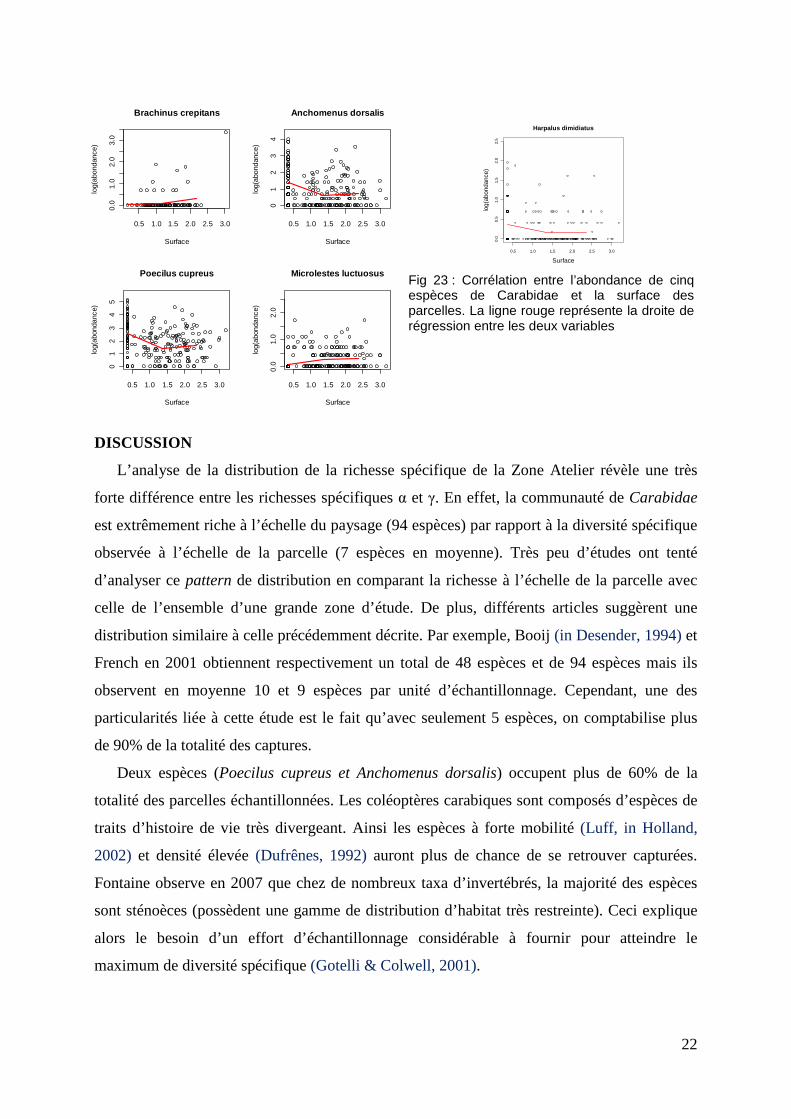
Le nombre moyen d’espèces est faiblement, mais positivement corrélé avec la surface des
parcelles (t = 3.682, df = 593, p-value = 0.00025) (Fig 21). Par contre, lorsque l’on distingue
les quatre types de cultures, cette relation disparait : les tendances sont toujours la, mais
l’effet statistique ne l’est plus. Aucune corrélation n’est significative entre la surface des
parcelles et la richesse spécifique par culture (Cer : p-value=0,05 ; Col : p-value=0,23 ;Luz :
p-value= 0,16 ; Prai : p-value=0,09) (Fig 22).
Nous avons aussi testé cette relation chez les 10 espèces les plus communes (Fig. 23).
Seules les corrélations des espèces suivantes sont significatives avec la surface de la parcelle
(p-value < 0,05) : Brachinus crepitans, Anchomenus dorsalis, Poecilus cupreus, Mocrolestes
luctuosus et Harpalus dimidiatus. De plus, cette relation s’applique seulement pour la culture
de prairies pour Brachinue crepitans, et pour la culture de céréales pour les quatre autres
espèces. Cependant, les corrélations restent très faibles puisque l’indice varie entre 0,2 et 0,3.
0 1 2 3 4
05
1015
2025
30
Log(surface)(ha)
Nom
bre
d’es
pèce
s
Fig 21 : Corrélation entre le nombre d’espèces et la surface des parcelles pour l’ensemble du jeu de données. La ligne rouge représente la droite de régression entre les deux variables
Fig 22 : Corrélation entre le nombre d’espèces et la surface des parcelles par culture. La ligne rouge représente la droite de régression entre les deux variables.
0 1 2 3 4
510
1520
MTailcer$a
MT
ailc
er$R
S
0.5 1.0 1.5 2.0 2.5
510
1520
MTailcol$a
MT
ailc
ol$R
S
0.5 1.0 1.5 2.0 2.5
510
15
MTailluz$a
MT
aillu
z$R
S
0.5 1.0 1.5 2.0
05
1015
2025
30
MTailprai$a
MTai
lpra
i$R
S
Log(surface )(ha)
N e
spèc
es
N e
spèc
es
N e
spèc
es
N e
spèc
es
Log(surface )(ha)
Log(surface )(ha) Log(surface )(ha)
Cer
Luz
Col
Prai
22
DISCUSSION
L’analyse de la distribution de la richesse spécifique de la Zone Atelier révèle une très
forte différence entre les richesses spécifiques α et γ. En effet, la communauté de Carabidae
est extrêmement riche à l’échelle du paysage (94 espèces) par rapport à la diversité spécifique
observée à l’échelle de la parcelle (7 espèces en moyenne). Très peu d’études ont tenté
d’analyser ce pattern de distribution en comparant la richesse à l’échelle de la parcelle avec
celle de l’ensemble d’une grande zone d’étude. De plus, différents articles suggèrent une
distribution similaire à celle précédemment décrite. Par exemple, Booij (in Desender, 1994) et
French en 2001 obtiennent respectivement un total de 48 espèces et de 94 espèces mais ils
observent en moyenne 10 et 9 espèces par unité d’échantillonnage. Cependant, une des
particularités liée à cette étude est le fait qu’avec seulement 5 espèces, on comptabilise plus
de 90% de la totalité des captures.
Deux espèces (Poecilus cupreus et Anchomenus dorsalis) occupent plus de 60% de la
totalité des parcelles échantillonnées. Les coléoptères carabiques sont composés d’espèces de
traits d’histoire de vie très divergeant. Ainsi les espèces à forte mobilité (Luff, in Holland,
2002) et densité élevée (Dufrênes, 1992) auront plus de chance de se retrouver capturées.
Fontaine observe en 2007 que chez de nombreux taxa d’invertébrés, la majorité des espèces
sont sténoèces (possèdent une gamme de distribution d’habitat très restreinte). Ceci explique
alors le besoin d’un effort d’échantillonnage considérable à fournir pour atteindre le
maximum de diversité spécifique (Gotelli & Colwell, 2001).
Fig 23 : Corrélation entre l’abondance de cinq espèces de Carabidae et la surface des parcelles. La ligne rouge représente la droite de régression entre les deux variables
0.5 1.0 1.5 2.0 2.5 3.0
0.0
1.0
2.0
3.0
Brachinus crepitans
Surface
log(
abon
danc
e)
0.5 1.0 1.5 2.0 2.5 3.0
01
23
4
Anchomenus dorsalis
Surface
log(
abon
danc
e)
0.5 1.0 1.5 2.0 2.5 3.0
01
23
45
Poecilus cupreus
Surface
log(
abon
danc
e)
0.5 1.0 1.5 2.0 2.5 3.0
0.0
1.0
2.0
Microlestes luctuosus
Surface
log(
abon
danc
e)0.5 1.0 1.5 2.0 2.5 3.0
0.0
0.5
1.0
1.5
2.0
2.5
Harpalus dimidiatus
Surface
log(
abon
danc
e)

23
Les analyses multivariées ne révèlent aucune structuration nette des communautés entre
les cultures pérennes et les cultures annuelles. Ainsi, l’hypothèse selon laquelle les espèces,
étant sensibles aux conditions abiotiques, se ségrégent selon leur préférence d’habitat (Dajoz,
2002) n’est pas confortée. La Zone Atelier Plaine & Val de Sèvre fait partie d’un paysage
agricole très intensif. Les cultures pérennes telles que les luzernes et les prairies seraient donc
gérées de manière tout aussi intensive que les cultures annuelles, au moins du point de vue
d’un coléoptère carabique. Ainsi, des milieux considérés comme relativement stables dans le
temps et moins perturbés chimiquement peuvent être perçus par la microfaune comme
particulièrement perturbés car ce sont des milieux fauchés régulièrement et retournés tous les
deux ou trois ans détruisant ainsi les larves se développant dans le sol (Gobbi, 2008).
L’impact des perturbations physiques et mécaniques sur la communauté de coléoptères
carabiques en milieu pérenne serait alors tout aussi fort que les perturbations chimiques
(herbicides, insecticides) dans les cultures annuelles. De plus, les milieux pérennes incluent
les luzernes qui peuvent être gérées très intensivement par d’importants traitements
phytopharmaceutiques. Dans ce cas, leur gestion se rapproche davantage de celle d’une
culture annuelle comme le colza que de la gestion d’une prairie de graminées. Or l’ACM met
en évidence que certaines espèces se rassemblent selon des traits biologiques communs. On
aurait alors pu s’attendre à ce que des groupes d’espèces se distinguent selon les cultures
selon leurs traits d’histoire de vie. Toutefois, les résultats de l’ACC ne permettent pas
d’observer ce pattern de distribution ce qui conforte l’idée que les pratiques agricoles des
cultures pérennes et annuelles ont un impact similaire sur la structure de la communauté de
Carabidae. Une seconde explication pourrait permettre de comprendre l’absence de
structuration des communautés. Au sien d’une parcelle de céréales, avant le labour des terres,
le sol est ouvert et sec puis après le labour, les graines sont semés avec de l’engrais organique
rendant ainsi les conditions du sol plus humide (Thomas et al., in Desender 1994). Au cours
d’une saison, les conditions abiotiques au sein de cette culture sont alors très variables. Les
espèces, selon leurs exigences biologiques, peuvent avoir besoin de se réfugier dans des
habitats adjacents lorsque l’environnement devient défavorables afin d’y trouver de la
ressource et à termes se maintenir dans le paysage (French et al., 2001). Ainsi, les espèces
telles que Pterostichus niger dispersant facilement, peuvent modifier rapidement leur
distribution dans le paysage. Les cultures pérennes ont plusieurs fois montré leur rôle en tant
qu’habitat complémentaire dans les milieux perturbés (Le Roux et al., 2008). Ainsi, la
similarité de richesse spécifique entre les cultures pérennes et annuelles s’expliquerait par une
dispersion fréquente des espèces entre ces deux types de milieux (Diekotter et al., 2010).

24
En ce qui concerne l’influence de la surface des patches d’habitat sur la richesse
spécifique, une corrélation positive modeste apparait, ce qui est conforme aux prédictions
attendues par les différentes théories des communautés. D’après Dajoz (2002), la théorie de la
biogéographie insulaire ne s’applique pas dans ce type de systèmes car les facteurs
prédominants sur la richesse spécifique sont la perturbation et l’hétérogénéité de l’habitat. A
l’inverse, à l’échelle de la culture, aucune corrélation significative n’est observée. Bien que
l’on soit sur une plaine céréalière intensive, la taille des parcelles est petite (en moyenne
5.2Ha pour les céréales, 5.7Ha pour le colza, 2.7Ha pour les prairies et 3.8Ha pour les
luzernes) alors que d’autres études présentent des surfaces moyennes beaucoup plus grandes
(Moreby, 1999 : >20Ha ; Clark, 2006 : 16Ha). Ainsi, la faible variabilité de surface au sein de
chaque culture ne permettrait pas de détecter d’effet de la surface des parcelles sur la richesse
spécifique. Pour ce qui est de l’abondance (densité locale) des espèces, les résultats indiquent
que la surface de la parcelle a un effet significatif sur la moitié de ces espèces testées, mais
seulement 2 d’entre elles présentent une corrélation positive avec l’augmentation de la surface
de la parcelle, alors que trois espèces présentent une relation négative dans les cultures de
céréales contre une positive dans les céréales et une seconde positive dans les prairies. En
comparant les traits biologiques de ces espèces, on constate qu’elles possèdent cependant des
traits d’histoire de vie similaires. Ce sont des espèces de petite taille à forte capacité de
dispersion, spécialistes ou généralistes. La fragmentation des habitats provoquant une baisse
de la qualité des milieux, on aurait pu s’attendre à ce que l’abondance des espèces spécialistes
décroisse dans les petits patches de moins bonne qualité que dans les grands patches et que
l’effet soit nul pour les espèces généralistes, étant capable de disperser dans l’environnement
voisin en cas de perturbation du milieu. Ces résultats sont en accord cette prédiction pour
Brachinus crepitans et pour Microlestes luctuosus qui sont deux espèces spécialistes. A
l’inverse les trois autres espèces généralistes, présentent une tendance inverse aux prédictions.
Ceci peut être rattaché à l’hypothèse de Dajoz qui affirme que la taille du patch peut ne pas
être le facteur dominant à l’origine de la distribution des espèces. Il serait intéressant alors
d’approfondir les études en prenant en considération la structure spatiale de l’environnement
voisin ainsi que les variables environnementales associées telles que l’humidité ou l’ouverture
du paysage pour tenter de confirmer ou non l’hypothèse de Dajoz.

25
CONCLUSION
Cette étude est la première étude réalisée sur la base de données de Coléoptères carabiques
de la Zone Atelier Plaine & Val de Sèvre. Elle reste ainsi très exploratoire et descriptive.
Cependant, des grandes tendances se dégagent et laissent ainsi place à de nombreuses
perspectives afin d’analyser plus précisément les résultats obtenus. En effet, la richesse
spécifique présente sur la zone d’étude est très inégalement répartie. Nos résultats indiquent
que les parcelles sont composées d’une ou deux espèces dominantes dans la communauté et
que sur les sept espèces présentes en moyenne au sein de ces patches d’habitats, les cinq
restantes sont des espèces de fréquence rare, qui différent selon les parcelles. Cela nous
suggère que la communauté de Carabidae fonctionnerait davantage selon un modèle
neutraliste, avec une répartition stochastique des espèces dont la dynamique serait gérée par le
taux de natalité, de mortalité et de dispersion des espèces (Alonso et al., 2006). Cependant,
cette théorie a été très controversée dans la littérature (Ricklefs, 2003 ; Adler, 2004). En effet,
ces auteurs affirment que certains processus, tels que les relations interspécifiques, sont
manquants dans cette théorie alors qu’ils sont nécessaires afin d’étudier des patterns de
distribution d’espèces. Ainsi, il serait intéressant de prendre en considération de tels
mécanismes pour de futures études de fonctionnement des communautés de coléoptères
carabiques. Apporter une dimension spatiale à cette étude permettrait de confirmer ou non la
non structuration des communautés observée sur la Zone Atelier en intégrant la densité des
éléments favorables ainsi que la distance entre les patches pouvant influencer le taux de
colonisation et d’extinction des espèces.

26
BIBLIOGRAPHIE Adler, P. B. 2004. Neutral models fail to reproduce observed species-area and species-time
relationships in Kansas grasslands. Ecology 85:1265-1272. Alonso, D., R. S. Etienne, and A. J. McKane. 2006. The merits of neutral theory. Trends in Ecology
& Evolution 21:451-457. Baldi, A. and T. Kisbenedek. 1997. Orthopteran assemblages as indicators of grassland naturalness
in Hungary. Agriculture Ecosystems & Environment 66:121-129. Benton, T. G., D. M. Bryant, L. Cole, and H. Q. P. Crick. 2002. Linking agricultural practice to
insect and bird populations: a historical study over three decades. Journal of Applied Ecology 39:673-687.
Clark , S., K. Szlavecz, M. A. Cavigelli, and F. Purrington. 2006. Ground beetle (Coleoptera : Carabidae) assemblages in organic, no-till, and chisel-till cropping systems in Maryland. Environmental Entomology 35:1304-1312.
Colwell, R. K. and J. A. Coddington. 1994. ESTIMATING TERRESTRIAL BIODIVERSITY THROUGH EXTRAPOLATION. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 345:101-118.
Colwell, R. K., C. X. Mao, and J. Chang. 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85:2717-2727.
Dajoz R. Les Coléoptères Carabidés et Ténébrionidés. TEC & DOC, 2002 edition. Lavoisier, Paris. Desender K., D. M., Loreau M., Luff M.L. & Maelfait J.P . 1994. Carabid beetles ecology and
evolution. Kluwer Academic Publishers (Dordercht). Desender K., D. W., Dufrêne M. & Maes D. 2010. Changes in the distribution of carabid beetles in
Belgium revisited:Have we halted the diversity loss? Pages 1549-1557 Biological conservation. Diekotter, T., S. Wamser, V. Wolters, and K. Birkhofer. 2010. Landscape and management effects on
structure and function of soil arthropod communities in winter wheat. Agriculture Ecosystems & Environment 137:108-112.
Dufrêne M. 1992. Biogéographie et Écologie des Communautés de Carabidae en Wallonie. Thèse : Département de Biologie. Unité d'Écologie et de Biogéographie. Université Catholique de Louvain. 288p.
Elzinga, J. A., S. van Nouhuys, D. J. van Leeuwen, and A. Biere. 2007. Distribution and colonisation ability of three parasitoids and their herbivorous host in a fragmented landscape. Basic and Applied Ecology 8:75-88.
Fontaine et al. The European Union's 2010 target: Putting rare species in focus. Biological Conservation 139:167-185.
Forel J. & Leplat J.2003a. Faune des carabiqes de France - X. Systématique edition. Magellanes. Forel J. & Leplat J.2003b. Faune des carabiques de France - XI. Systématiques edition. Megallanes. French, B. W., N. C. Elliott, R. C. Berberet, and J. D. Burd. 2001. Effects of riparian and grassland
habitats on ground beetle (Coleoptera : Carabidae) assemblages in adjacent wheat fields. Environmental Entomology 30:225-234.
Gaublomme, E., F. Hendrickx, H. Dhuyvetter, and K. Desender. 2008. The effects of forest patch size and matrix type on changes in carabid beetle assemblages in an urbanized landscape. Biological Conservation 141:2585-2596.
Gobbi, M. and D. Fontaneto. 2008. Biodiversity of ground beetles (Coleoptera : Carabidae) in different habitats of the Italian Po lowland. Agriculture Ecosystems & Environment 127:273-276.
Gotelli, N. J. and R. K. Colwell. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4:379-391.
Graham, J. H. et al. 2009. Species richness, equitability, and abundance of ants in disturbed

27
landscapes. Ecological Indicators 9:866-877. Gutierrez, D., R. Menendez, and M. Mendez. 2004. Habitat-based conservation priorities for
carabid beetles within the Picos de Europa National Park, northern Spain. Biological Conservation 115:379-393.
Hanski, I. 1991. SINGLE-SPECIES METAPOPULATION DYNAMICS - CONCEPTS, MODELS AND OBSERVATIONS. Biological Journal of the Linnean Society 42:17-38.
Hodar, J. A. 1996. The use of regression equations for estimation of arthropod biomass in ecological studies. Acta Oecologica-International Journal of Ecology 17:421-433.
Holland, J. M. 2002. The agroecology of carabid beetles. 1 edition (January 9, 2004) edition. Intercept.
Holyoak M., L. M. A. H. R. D. 2005. Metacommunities : Spatial dynamics and ecological communities. New edition edition (Oct 1 2005) edition. University Of Chicago Press.
Lambeets, K., F. Hendrickx, S. Vanacker, K. Van Looy, J. P. Maelfait, and D. Bonte. 2008. Assemblage structure and conservation value of spiders and carabid beetles from restored lowland river banks. Biodiversity and Conservation 17:3133-3148.
Le Roux X. et al. 2008. Agriculture et biodiversité : valoriser les synergies une expertise scientifique collective conduite par l'INRA - Synthèse du rapport d'expertise.
Loreau, M. 2001. Biodiversity and ecosystem functioning: recent theoretical advances. Oikos 91:3-17.
Melnychuk, N. A., O. Olfert, B. Youngs, and C. Gillott. 2003. Abundance and diversity of Carabidae (Coleoptera) in different farming systems. Agriculture Ecosystems & Environment 95:69-72.
Moreby, S. J. and S. E. Southway. 1999. Influence of autumn applied herbicides on summer and autumn food available to birds in winter wheat fields in southern England. Agriculture Ecosystems & Environment 72:285-297.
Niemela, J. 2001. Carabid beetles (Coleoptera : Carabidae) and habitat fragmentation: a review. European Journal of Entomology 98:127-132.
Oksanen J. et al. 2009. Vegan: community ecology package, R. Pearsall I.A. 2007. Carabid beetles as ecological indicators. Paper presented at the "Monitoring the
effectiveness of biological conservation" conference, 2-4 november 2004, Richmond, BC. Purtauf, T., J. Dauber, and V. Wolters. 2005. The response of carabids to landscape simplification
differs between trophic groups. Oecologia 142:458-464. R Development Core Team. 2009. R: A language and environment for statistical computing. /R
Foundation forStatistical Computing, Vienna, Austria/. http://www.R-project.org <http://www.r-project.org/>.
Ricklefs R.E. 2003. A comment on Hubell's zero-sum ecological drift model. Pages 185-192 Oikos. Sala O.E., et al.. 2000. Biodiversity - Global biodiversity scenarios for the year 2100. Science
287:1770-1774. Schweiger, O. et al. 2005. Quantifying the impact of environmental factors on arthropod communities
in agricultural landscapes across organizational levels and spatial scales. Journal of Applied Ecology 42:1129-1139.
Tscharntke, T. and R. Brandl. 2004. Plant-insect interactions in fragmented landscapes. Annual Review of Entomology 49:405-430.
Volkov I., B. J. R., Hubbell S.P., Maritan A. 2003. Neutral theory and relative species abundance ecology. Pages 1-19 Nature.
Weibull, A. C., O. Ostman, and A. Granqvist. 2003. Species richness in agroecosystems: the effect of landscape, habitat and farm management. Biodiversity and Conservation 12:1335-1355.
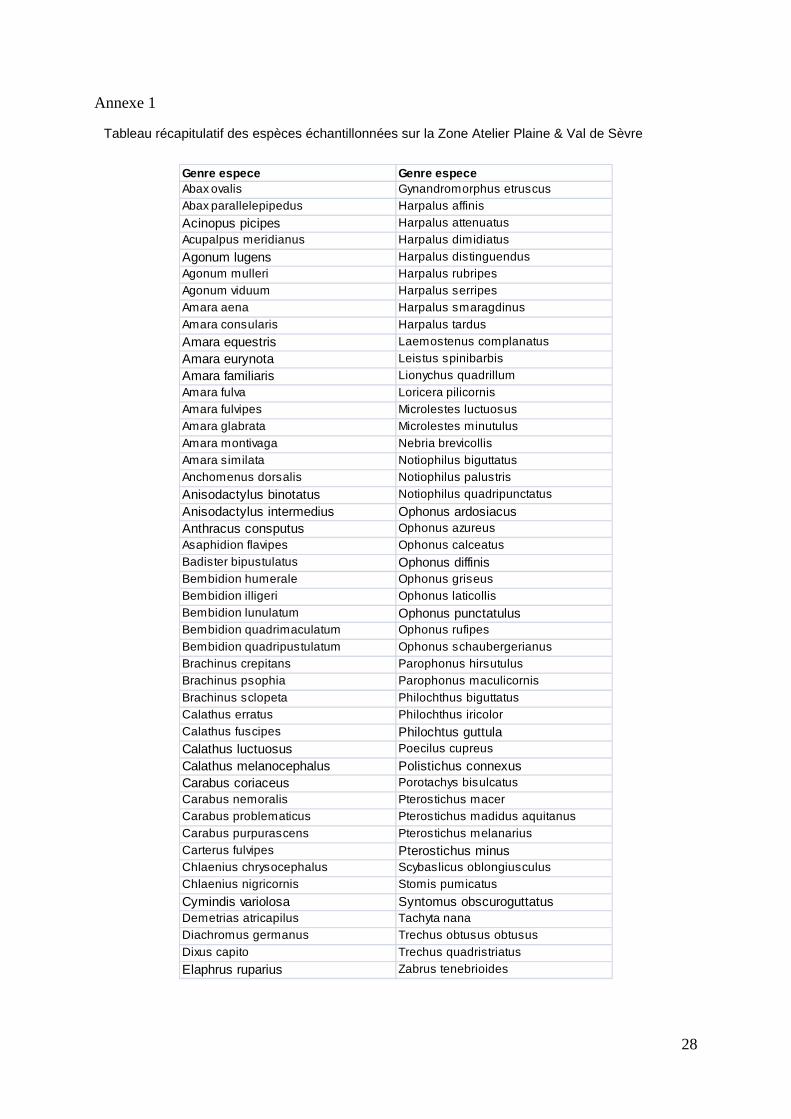
28
Genre espece Genre especeAbax ovalis Gynandromorphus etruscusAbax parallelepipedus Harpalus affinis
Acinopus picipes Harpalus attenuatusAcupalpus meridianus Harpalus dimidiatus
Agonum lugens Harpalus distinguendusAgonum mulleri Harpalus rubripesAgonum viduum Harpalus serripesAmara aena Harpalus smaragdinusAmara consularis Harpalus tardus
Amara equestris Laemostenus complanatus
Amara eurynota Leistus spinibarbis
Amara familiaris Lionychus quadrillumAmara fulva Loricera pilicornisAmara fulvipes Microlestes luctuosusAmara glabrata Microlestes minutulusAmara montivaga Nebria brevicollisAmara similata Notiophilus biguttatusAnchomenus dorsalis Notiophilus palustris
Anisodactylus binotatus Notiophilus quadripunctatus
Anisodactylus intermedius Ophonus ardosiacusAnthracus consputus Ophonus azureusAsaphidion flavipes Ophonus calceatusBadister bipustulatus Ophonus diffinisBembidion humerale Ophonus griseusBembidion illigeri Ophonus laticollisBembidion lunulatum Ophonus punctatulusBembidion quadrimaculatum Ophonus rufipesBembidion quadripustulatum Ophonus schaubergerianusBrachinus crepitans Parophonus hirsutulusBrachinus psophia Parophonus maculicornisBrachinus sclopeta Philochthus biguttatusCalathus erratus Philochthus iricolorCalathus fuscipes Philochtus guttulaCalathus luctuosus Poecilus cupreus
Calathus melanocephalus Polistichus connexusCarabus coriaceus Porotachys bisulcatusCarabus nemoralis Pterostichus macerCarabus problematicus Pterostichus madidus aquitanusCarabus purpurascens Pterostichus melanariusCarterus fulvipes Pterostichus minusChlaenius chrysocephalus Scybaslicus oblongiusculusChlaenius nigricornis Stomis pumicatus
Cymindis variolosa Syntomus obscuroguttatusDemetrias atricapilus Tachyta nanaDiachromus germanus Trechus obtusus obtususDixus capito Trechus quadristriatus
Elaphrus ruparius Zabrus tenebrioides
Annexe 1
Tableau récapitulatif des espèces échantillonnées sur la Zone Atelier Plaine & Val de Sèvre
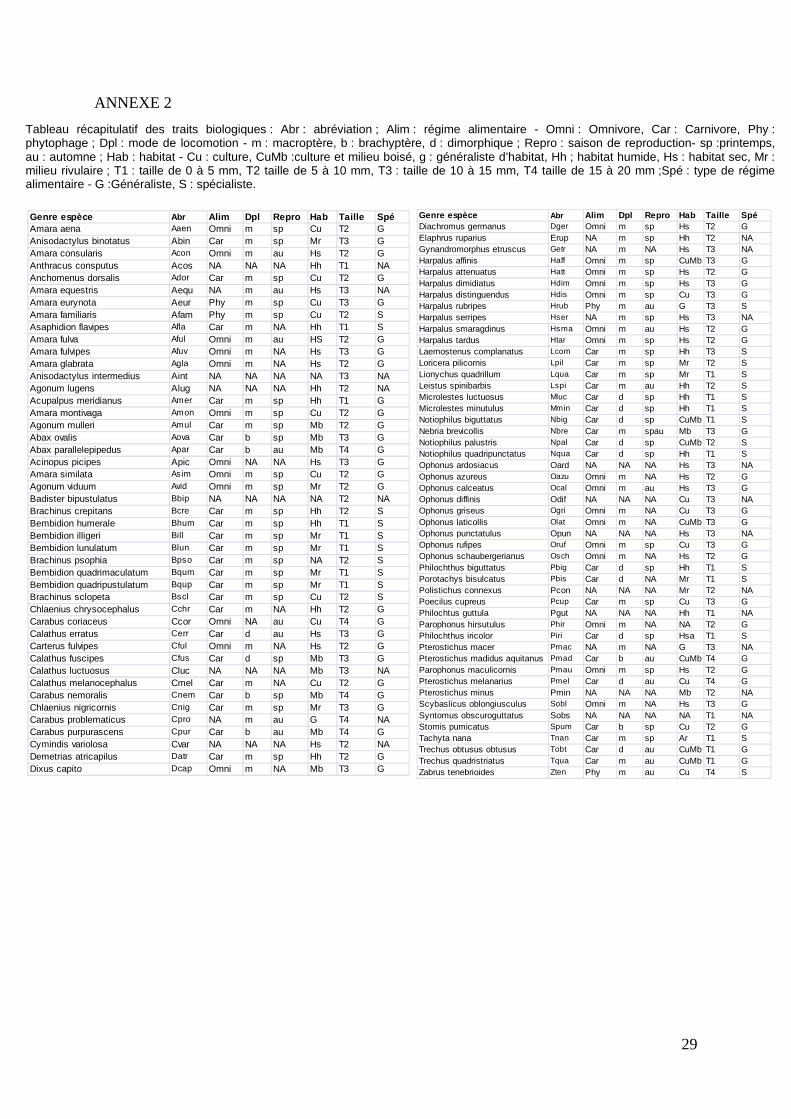
29
Genre espèce Abr Alim Dpl Repro Hab Taille SpéAmara aena Aaen Omni m sp Cu T2 GAnisodactylus binotatus Abin Car m sp Mr T3 GAmara consularis Acon Omni m au Hs T2 GAnthracus consputus Acos NA NA NA Hh T1 NAAnchomenus dorsalis Ador Car m sp Cu T2 GAmara equestris Aequ NA m au Hs T3 NAAmara eurynota Aeur Phy m sp Cu T3 GAmara familiaris Afam Phy m sp Cu T2 SAsaphidion flavipes Afla Car m NA Hh T1 SAmara fulva Aful Omni m au HS T2 GAmara fulvipes Afuv Omni m NA Hs T3 GAmara glabrata Agla Omni m NA Hs T2 GAnisodactylus intermedius Aint NA NA NA NA T3 NAAgonum lugens Alug NA NA NA Hh T2 NAAcupalpus meridianus Amer Car m sp Hh T1 GAmara montivaga Amon Omni m sp Cu T2 GAgonum mulleri Amul Car m sp Mb T2 GAbax ovalis Aova Car b sp Mb T3 GAbax parallelepipedus Apar Car b au Mb T4 GAcinopus picipes Apic Omni NA NA Hs T3 GAmara similata Asim Omni m sp Cu T2 GAgonum viduum Avid Omni m sp Mr T2 GBadister bipustulatus Bbip NA NA NA NA T2 NABrachinus crepitans Bcre Car m sp Hh T2 SBembidion humerale Bhum Car m sp Hh T1 SBembidion illigeri Bill Car m sp Mr T1 SBembidion lunulatum Blun Car m sp Mr T1 SBrachinus psophia Bpso Car m sp NA T2 SBembidion quadrimaculatum Bqum Car m sp Mr T1 SBembidion quadripustulatum Bqup Car m sp Mr T1 SBrachinus sclopeta Bscl Car m sp Cu T2 SChlaenius chrysocephalus Cchr Car m NA Hh T2 GCarabus coriaceus Ccor Omni NA au Cu T4 GCalathus erratus Cerr Car d au Hs T3 GCarterus fulvipes Cful Omni m NA Hs T2 GCalathus fuscipes Cfus Car d sp Mb T3 GCalathus luctuosus Cluc NA NA NA Mb T3 NACalathus melanocephalus Cmel Car m NA Cu T2 GCarabus nemoralis Cnem Car b sp Mb T4 GChlaenius nigricornis Cnig Car m sp Mr T3 GCarabus problematicus Cpro NA m au G T4 NACarabus purpurascens Cpur Car b au Mb T4 GCymindis variolosa Cvar NA NA NA Hs T2 NADemetrias atricapilus Datr Car m sp Hh T2 GDixus capito Dcap Omni m NA Mb T3 G
Genre espèce Abr Alim Dpl Repro Hab Taille SpéDiachromus germanus Dger Omni m sp Hs T2 GElaphrus ruparius Erup NA m sp Hh T2 NAGynandromorphus etruscus Getr NA m NA Hs T3 NAHarpalus affinis Haff Omni m sp CuMb T3 GHarpalus attenuatus Hatt Omni m sp Hs T2 GHarpalus dimidiatus Hdim Omni m sp Hs T3 GHarpalus distinguendus Hdis Omni m sp Cu T3 GHarpalus rubripes Hrub Phy m au G T3 SHarpalus serripes Hser NA m sp Hs T3 NAHarpalus smaragdinus Hsma Omni m au Hs T2 GHarpalus tardus Htar Omni m sp Hs T2 GLaemostenus complanatus Lcom Car m sp Hh T3 SLoricera pilicornis Lpil Car m sp Mr T2 SLionychus quadrillum Lqua Car m sp Mr T1 SLeistus spinibarbis Lspi Car m au Hh T2 SMicrolestes luctuosus Mluc Car d sp Hh T1 SMicrolestes minutulus Mmin Car d sp Hh T1 SNotiophilus biguttatus Nbig Car d sp CuMb T1 SNebria brevicollis Nbre Car m spau Mb T3 GNotiophilus palustris Npal Car d sp CuMb T2 SNotiophilus quadripunctatus Nqua Car d sp Hh T1 SOphonus ardosiacus Oard NA NA NA Hs T3 NAOphonus azureus Oazu Omni m NA Hs T2 GOphonus calceatus Ocal Omni m au Hs T3 GOphonus diffinis Odif NA NA NA Cu T3 NAOphonus griseus Ogri Omni m NA Cu T3 GOphonus laticollis Olat Omni m NA CuMb T3 GOphonus punctatulus Opun NA NA NA Hs T3 NAOphonus rufipes Oruf Omni m sp Cu T3 GOphonus schaubergerianus Osch Omni m NA Hs T2 GPhilochthus biguttatus Pbig Car d sp Hh T1 SPorotachys bisulcatus Pbis Car d NA Mr T1 SPolistichus connexus Pcon NA NA NA Mr T2 NAPoecilus cupreus Pcup Car m sp Cu T3 GPhilochtus guttula Pgut NA NA NA Hh T1 NAParophonus hirsutulus Phir Omni m NA NA T2 GPhilochthus iricolor Piri Car d sp Hsa T1 SPterostichus macer Pmac NA m NA G T3 NAPterostichus madidus aquitanus Pmad Car b au CuMb T4 GParophonus maculicornis Pmau Omni m sp Hs T2 GPterostichus melanarius Pmel Car d au Cu T4 GPterostichus minus Pmin NA NA NA Mb T2 NAScybaslicus oblongiusculus Sobl Omni m NA Hs T3 GSyntomus obscuroguttatus Sobs NA NA NA NA T1 NAStomis pumicatus Spum Car b sp Cu T2 GTachyta nana Tnan Car m sp Ar T1 STrechus obtusus obtusus Tobt Car d au CuMb T1 GTrechus quadristriatus Tqua Car m au CuMb T1 GZabrus tenebrioides Zten Phy m au Cu T4 S
ANNEXE 2
Tableau récapitulatif des traits biologiques : Abr : abréviation ; Alim : régime alimentaire - Omni : Omnivore, Car : Carnivore, Phy : phytophage ; Dpl : mode de locomotion - m : macroptère, b : brachyptère, d : dimorphique ; Repro : saison de reproduction- sp :printemps, au : automne ; Hab : habitat - Cu : culture, CuMb :culture et milieu boisé, g : généraliste d’habitat, Hh ; habitat humide, Hs : habitat sec, Mr : milieu rivulaire ; T1 : taille de 0 à 5 mm, T2 taille de 5 à 10 mm, T3 : taille de 10 à 15 mm, T4 taille de 15 à 20 mm ;Spé : type de régime alimentaire - G :Généraliste, S : spécialiste.
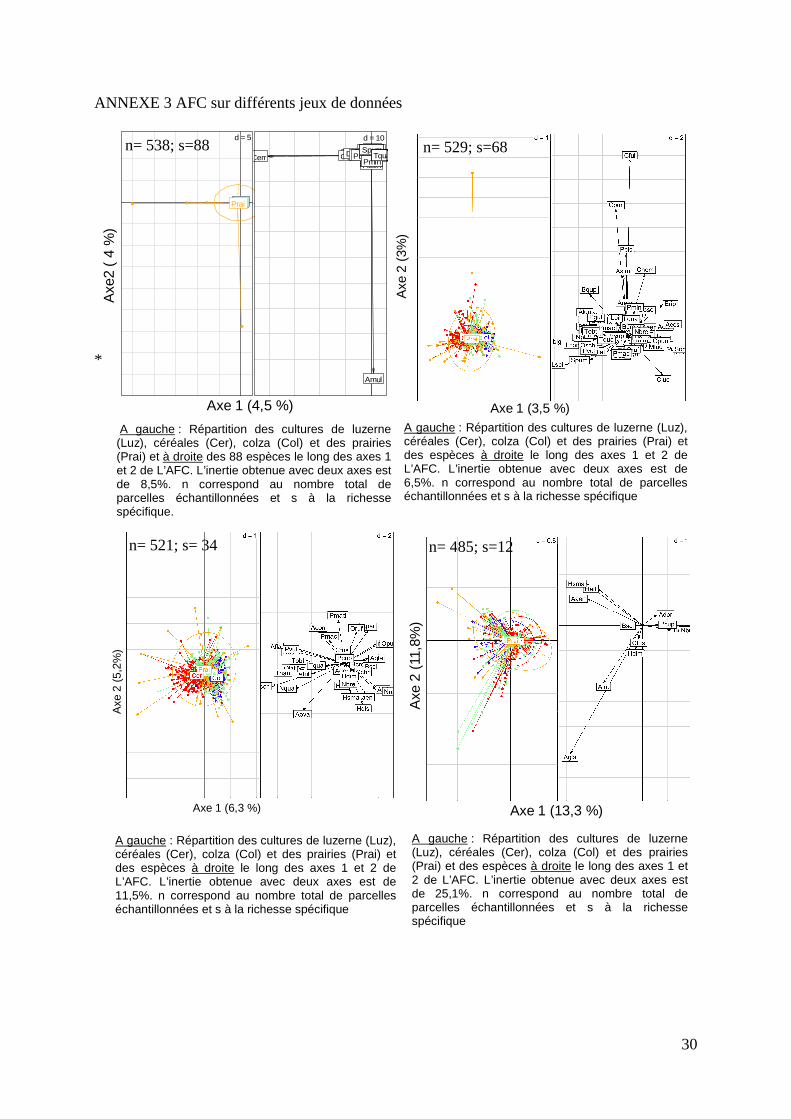
30
ANNEXE 3 AFC sur différents jeux de données *
Axe 1 (4,5 %)
Axe
2 (
4 %
)
d = 5
Cer Col Luz Prai
d = 10 d = 10
Aova Apar
Apic Amer Alug
Amul
Avid Aaen
Acon Aeur Afam
Aful Afuv
Agla Amon Asim
Ador Abin Aint Acon.1
Afla Bbip Bhum Bill Blun
Bqum Bqup Bcre Bpso
Bscl Cerr Cfus Cluc Ccor Cnem Cpur Cful Cchr Cnig
Datr Dger Dcap Erip
Getr Hatt Haff Hdim Hdis Hsma Htar Lcom Lspi Lqua Lpil Mluc Mmin Nbre Nbig Nqua
Npal Nruf Oard
Oazu Ocal Odif Ogri
Olat Opun Oruf Osch Pgut Phir
Piri Pcup Pcon Pbis Pbig Pmac Pmad Pmin Pvul Sobl
Sobs Spum
Tnan Tobt Tqua n= 538; s=88
A gauche : Répartition des cultures de luzerne (Luz), céréales (Cer), colza (Col) et des prairies (Prai) et des espèces à droite le long des axes 1 et 2 de L’AFC. L’inertie obtenue avec deux axes est de 6,5%. n correspond au nombre total de parcelles échantillonnées et s à la richesse spécifique
Axe 1 (3,5 %)A
xe 2
(3%
)
n= 529; s=68
A gauche : Répartition des cultures de luzerne (Luz), céréales (Cer), colza (Col) et des prairies (Prai) et à droite des 88 espèces le long des axes 1 et 2 de L’AFC. L’inertie obtenue avec deux axes est de 8,5%. n correspond au nombre total de parcelles échantillonnées et s à la richesse spécifique.
n= 485; s=12
Axe 1 (13,3 %)
Axe
2 (
11,8
%)
Axe 1 (6,3 %)
Axe
2 (
5,2%
)
n= 521; s= 34
A gauche : Répartition des cultures de luzerne (Luz), céréales (Cer), colza (Col) et des prairies (Prai) et des espèces à droite le long des axes 1 et 2 de L’AFC. L’inertie obtenue avec deux axes est de 25,1%. n correspond au nombre total de parcelles échantillonnées et s à la richesse spécifique
A gauche : Répartition des cultures de luzerne (Luz), céréales (Cer), colza (Col) et des prairies (Prai) et des espèces à droite le long des axes 1 et 2 de L’AFC. L’inertie obtenue avec deux axes est de 11,5%. n correspond au nombre total de parcelles échantillonnées et s à la richesse spécifique

31
Résumé
Depuis 50 ans, de nombreuses théories ont été proposées pour expliquer et prédire la structure et la dynamique des communautés. Ces théories présentent des caractéristiques communes liées au processus d’extinction et de colonisation des espèces dépendant de facteurs environnementaux tels que la taille ou la qualité du patch. Les agro-écosystèmes apparaissent comme des milieux parfaits pour ces études de communautés car ce sont des mosaïques hétérogènes de « patches » divers plus ou moins proches, contrastés et de qualité variable. Les coléoptères carabiques représentent un modèle biologique original et pertinent pour ce type d’étude de par leur diversité spécifique élevée et leur importance en termes d’agroécologie. L’objectif de ce travail est d’étudier la communauté de Carabidae dans un paysage agricole intensif, et en particulier les règles d’assemblages des espèces au sein de la communauté et entre les cultures. Le deuxième enjeu est d’étudier l’influence de la surface de la parcelle sur les communautés et enfin d’analyser la covariance entre les cultures et les traits d’histoire de vie des espèces. Les résultats montrent que la communauté est composée de quelques espèces dominantes et de beaucoup d’espèces rares. De plus la répartition des espèces semble suivre une distribution aléatoire entre les parcelles. Aucune structuration nette des communautés ni dominance des traits d’histoire de vie entre les cultures n’ont été mises en évidence. L’influence de la surface de la parcelle se fait ressentir seulement à une échelle globale mais pas à l’échelle de la culture. Les résultats suggèrent que l’effet des pratiques agricoles au sein cultures pérennes est tout aussi importance que celui des cultures annuelles et expliquerait l’absence de structuration de la communauté. De part la répartition des espèces dans l’environnement nous pouvons supposer un fonctionnement de la communauté selon un modèle neutraliste.
Summary
For 50 years, many theories have been proposed to explain and predict the structure and community dynamics. These theories have common features related to the process of extinction and colonization of species dependent on environmental factors such as size or quality of the patch. Agro-ecosystems are seen as ideal settings for these studies because the communities are heterogeneous mosaics of "patches" more or less similar, contrasting and varying quality. The carabid beetles are an original and relevant biological model for this type of study because of their high species diversity and their importance in terms of agroecology. The objective of this work is to study the community ofCarabidae in intensive agricultural landscapes, and especially the rules of assemblages of species within communities and between cultures. The second issue is to study the influence of the patch size on the communities and to analyze the covariance between cultures and life history traits of species. The results show that the community is composed of a few dominant species and many rare species. Moreover, the distribution of species seems to follow a random distribution between plots. No clear structure or community dominance of life history traits between cultures have been identified. The influence of the surface of the patch is felt only at a global scale but not at the culture level. The results suggest that the effect of agricultural practices in perennial crops is as important as that of annual crops and explain the lack of community structure. Due to the distribution of species in the environment we can assume that the community follows as a neutralist model.







![La bourdaine - Abeille sentinelle de... · 2017-05-16 · nourricières des chenilles d’un papillon actuellement en forte raréfaction, le citron (Gonepteryx rhamni) [fig. 5]. intérêt](https://img.pdfslide.fr/doc/110x75/5e74da4bf2f9011fb90cb7e7/la-bourdaine-abeille-sentinelle-de-2017-05-16-nourricires-des-chenilles.jpg)





















