Embed Size (px)
Citation preview

Rapport de stage 1ère année de Master Environnement
Spécialité Ecologie, Biodiversité, Evolution (EBE)
Valérie GUITTET
Soutenance : 7-8 juin 2010
Rôle de la PTOX dans la tolérance au stress lumineux
chez les plantes alpines
Etude d’écotypes alpins chez Arabidopsis thaliana
Laboratoire Ecologie Systématique Evolution (Université Paris-XI)
Ecophysiologie Végétale
Durée du stage : 15 février - 28 mars 2010 Responsable de stage : Peter Streb
Année universitaire : 2009-2010

2
Sommaire
Remerciements p3 Introduction p4 I. Matériels et méthodes p6 1.1. Mesure de fluorescence chlorophyllienne p6
1.1. a. Description de la technique de mesure de la fluorescence et des paramètres associés
1.1. b. Description du matériel utilisé
1.1. c. Description du protocole expérimental
1.2. Calcul de l’ETR (electron transport rate) p13 1.3. Extraction de la protéine PTOX par Western Blot p13 II. Résultats p15 2.1. Mesure de fluorescence chlorophyllienne : calculs des qP et NPQ p15
2.1. a. Quenching photochimique qP
2.1. b. Quenching non-photochimique NPQ
2.2. Calcul de l’ETR (electron transport rate) p19 2.3. Extraction de la protéine PTOX par Western Blot p21 III. Discussion p22 Conclusion p29 Bibliographie p30 Annexes p32 Abstract / Résumé p36

3
Remerciements
Je remercie tout d’abord mon responsable de stage Mr Peter Streb, pour m’avoir intégré à son
équipe, pour toute l’aide et le temps qu’il m’a consacré au cours du stage.
Je remercie également Constance Laureau, thésarde à l’ESE, pour toutes les réponses qu’elle m’a
apportée et sans qui je n’aurai pu réaliser les Western Blots.
Enfin je remercie toutes les personnes qui ont pu contribuer de près ou de loin au bon déroulement
de ce stage.

4
Introduction
Que se soit en conditions naturelles ou en culture, les plantes sont fréquemment exposées à
des conditions environnementales défavorables à l’origine d’un stress. Le stress est communément
défini comme un facteur environnemental ne permettant pas le développement optimal de la plante,
comme par exemple une forte lumière (stress abiotique). Les conditions environnementales
stressantes jouent un rôle majeur en déterminant comment le sol et le climat limitent la distribution
des espèces.
Les plantes alpines poussent à de hautes altitudes, elles sont ainsi exposées à des conditions
environnementales extrêmes, notamment de fortes intensités lumineuses et des températures
variables (faibles ou fortes). De plus, la saison de croissance des plantes herbacées alpines est très
courte (deux ou trois mois), elles ont donc besoin d’un système d’assimilation du carbone très
efficace (Streb et al, 1997 ; Streb et al, 2005). De nombreuses études ont montré que la
photosynthèse des plantes alpines était bien adaptée à ces conditions climatiques de haute altitude :
large optimum de température permettant l’assimilation efficace du carbone et maintien de la
photosynthèse sous de fortes radiations lumineuses.
Les plantes sont acclimatées à utiliser l’énergie absorbée si le flux quantique reçu est
similaire à celui de leur condition de croissance. A la lumière, selon le flux quantique absorbé, la
protéine D1 du photosystème II (PSII) est dégradée en permanence et resynthétisée à la même
vitesse. Si la lumière reçue est trop forte, c’est-à-dire si le flux quantique reçu excède la capacité
photosynthétique de la plante, l’inactivation peut excéder la réparation et la plante risque la
photoinhibition du PSII (diminution du rendement quantique induit par la lumière). L’exposition à
de fortes radiations lumineuses, associée à d’autres conditions de stress (une faible température par
exemple), induit la formation d’oxygène réactif (ROS : reactive oxygen species) à l’origine du
stress oxydant. L’oxygène réactif est formé en permanence dans les membranes, mais en grande
quantité, il peut être dangereux pour la plante. Les ROS sont généralement formés en conditions
réductrices (accumulation des électrons dans la chaine de transport), par réduction de O2 (transfert
d’un électron) en ion superoxyde O2-, à l’origine de destructions membranaires. De plus, O2
- peut
réagir (réaction de Haber Weiss) pour former des ions hydroxyles HO- très dangereux pour la plante
(destructions cellulaires). L’oxygène singulet (réaction de la chlorophylle avec l’oxygène) est
également très réactif. La production de ROS peut affecter la synthèse de protéines, et notamment la
protéine D1 du PSII. Une faible température associée au stress oxydant ralentit les réactions
enzymatiques et par conséquent le renouvellement de la protéine D1. Ainsi, les conditions
défavorables de haute altitude (forte lumière, faible température) peuvent induire la photoinhibition
du PSII chez des plantes non-acclimatées (Streb et al, 1997). De plus, une forte température peut
altérer la fluidité des membranes thylakoïdiennes et inhiber le transport d’électrons lors de la

5
photosynthèse (Yordanov et al, 1986 ; Schreiber & Berry, 1977). Les plantes ont développé de
nombreux systèmes de protection leur permettant d’éliminer cet oxygène réactif ou d’empêcher sa
production. L’enzyme superoxyde dismutase (SOD), associée aux enzymes ascorbate peroxydase
(APX) et glutathion peroxydase (GPX), est impliquée dans la détoxification des ions superoxydes
O2- par production d’H2O dans les chloroplastes (système antioxydant) (Foyer et al, 1994). La
quantité d’antioxydants dans les plantes alpines augmente avec l’altitude (Wildi & Lütz, 1996). Le
cycle des xanthophylles permet de dissiper l’énergie en excès sous forme de chaleur (Demming-
Adam & Adam, 1993). La photorespiration peut consommer jusqu’à 30% des électrons résultant de
la réaction primaire de la photosynthèse. Elle permet ainsi d’éviter une sur-réduction de la chaine de
transport photosynthétique à l’origine de la production de ROS (Heber et al, 1996). La quantité de
caroténoïdes dans les thylakoïdes augmente dans des plantes acclimatées, suggérant que les
caroténoïdes sont impliqués dans les mécanismes de photo-protection des plantes (Tallon & Quiles,
2007). Enfin, la protéine PTOX (plastid terminal oxidase), aussi connu sous le nom d’IMMUTANS,
serait impliquée dans la protection des plantes contre le stress oxydant, en prévenant une sur-
réduction du pool de plastoquinones dans les chloroplastes. Elle serait capable de transférer les
électrons de la plastoquinone à l’oxygène, immédiatement transformé en H2O dans le stroma, sans
générer d’espèces oxygènes réactives (Streb et al, 2005). Située du côté stroma de la membrane
thylakoïdienne, elle jouerait ainsi le rôle d’accepteur alternatif d’électrons. Cette enzyme, associée à
la NADH DH (NADH Déshydrogénase) est impliquée dans la chlororespiration (Quiles, 2006).
Bennoun (1982) définit la chlororespiration comme une chaine de transport d’électrons située dans
la membrane du thylakoïde, en interaction avec la chaine de transport photosynthétique (circulation
des électrons entre PSII et PSI) et impliquant l’oxydation non-photochimique des plastoquinones.
Cependant, la PTOX est une protéine mineure dans de nombreuses espèces de plantes déjà étudiées
et sa capacité à consommer les électrons en excès parait être faible (Peltier & Cournac, 2002). Cette
enzyme est pourtant très abondante chez Ranunculus glacialis, une plante alpine de haute altitude,
dont la quantité excède celles de nombreuses autres plantes, notamment des feuilles de tomates
transgéniques sur-exprimant la PTOX. De plus, la quantité de PTOX chez R. glacialis diminue
fortement lors d’une désacclimatation (22°C à faible lumière pendant trois semaines), suggérant
qu’elle tient un rôle important dans la protection contre la lumière forte. Enfin, l’abondance de la
PTOX chez Geum montanum, une autre plante alpine, est corrélée avec l’altitude (Streb et al,
2005). La PTOX serait de ce fait une protéine importante dans l’adaptation des plantes alpines aux
conditions environnementales difficiles induisant un stress oxydant. Cependant, aucun test n’est
actuellement disponible pour quantifier le rôle de la PTOX comme accepteur alternatif d’électrons.
Le but du stage est de développer une méthode, basée sur la fluorescence chlorophyllienne, afin de
mettre en évidence le rôle de la PTOX chez les plantes alpines.

6
I. Matériels et méthodes
1.1. Mesure de fluorescence chlorophyllienne
1.1. a. Description de la technique de mesure de la fluorescence et des paramètres associés
L’énergie lumineuse absorbée par la chlorophylle d’une feuille peut être utilisée de trois
façons différentes. Elle peut être utilisée pour réaliser la photosynthèse (photochimie), l’énergie en
excès peut être dissipée sous forme de chaleur ou réémise sous forme de lumière (fluorescence
chlorophyllienne). Cependant, la quantité de lumière absorbée ré-émise sous forme de fluorescence
est très faible : seulement 1 à 3% de la lumière totale absorbée. Cela représente donc un rendement
de fluorescence (énergie émise par fluorescence / énergie absorbée par le PSII) très faible.
Ces trois processus sont en compétition, du fait que si l’efficacité de l’un d’eux augmente,
celle des deux autres diminue. Ainsi, en mesurant le rendement de la fluorescence chlorophyllienne,
on peut obtenir des informations concernant la photochimie et la libération de chaleur (Maxwell &
Johnson, 2000).
Figure 1 : Les trois voies d’utilisation de l’énergie
La lumière bleue excite la chlorophylle vers un niveau énergétique S2, et la lumière rouge vers un
niveau S1. Lorsqu’un électron retombe du niveau S1, cela entraine une libération d’énergie qui peut
être utilisée par trois processus : photosynthèse, fluorescence et émission de chaleur.
Technique de mesure de l’émission de fluorescence de la chlorophylle d’une feuille : (Cornic, 2007)
Figure 2 : schéma du dispositif

7
Trois sources lumineuses sont produites :
- une lumière modulée ou lumière analytique (lambda < 680nm) émise par une LED. Il lui
correspond une fluorescence modulée qui est amplifiée. Cette lumière analytique de flux
quantique constant et très faible permet de mesurer la fluorescence Fo.
- une lumière actinique (lumière solaire ou toute autre source lumineuse susceptible d’activer
la photosynthèse) produisant une émission de fluorescence par la feuille, non amplifiée. La
lumière actinique modifie l’état redox de QA (et donc l’état d’ouverture des centres PSII),
qui résulte d’une part de l’afflux d’électrons par les antennes (réactions amonts) et d’autre
part de l’utilisation de ces électrons pour l’assimilation du CO2 et de l’O2, etc. (réactions
avales).
- une source de lumière sursaturante (flash lumineux) : environ 8000 – 10.000 µmol
photons/m2/s, qui provoque la fermeture des centres PSII. Quand tous les centres sont
fermés, la fluorescence est maximale (Fm).
Variation de la fluorescence en fonction du temps sur une feuille intacte (Cornic, 2007) :
La variabilité de la fluorescence est due principalement au PSII. En effet, au niveau du PSI,
le quenching photochimique (qP) est toujours maximal. La fluorescence est donc faible et ne varie
pas. La raison de cette faible fluorescence est que le PSI trouve toujours un accepteur d’électrons :
même si NADP+ n’est pas libre ou que le transport cyclique est bloqué, le PSI peut transférer les
électrons à l’oxygène. Au niveau du PSII, la fluorescence dépend de l’état redox du pool de
plastoquinones et en particulier de QA. Elle dépend également du gradient de protons.
La feuille est maintenue à l’obscurité 30 minutes environ avant d’être éclairée par une
lumière modulée (LM) de très faible intensité. La fluorescence atteint un niveau minimal Fo (les QA
sont oxydées au maximum et tous les centres PSII sont ouverts). La feuille reçoit ensuite un flash de
lumière sursaturante (LSS), qui provoque la fermeture des centres PSII (QA réduites brutalement au
maximum) qui ne peuvent plus faire de photochimie. La fluorescence atteint un niveau maximal
Fm. Puis QA donne ses électrons à QB et l’émission de fluorescence diminue (oxydation de QA).
La lumière actinique (LA < LSS) provoque la fermeture de quelques centres PSII (réduction
partielle des QA) ainsi qu’une augmentation de l’émission de fluorescence jusqu’à un niveau Fp
puis Fs (valeur stationnaire correspondant à la réoxydation des QA). En effet, la photosynthèse est
induite à la lumière et donc les électrons sont consommés. Lorsqu’un flash de lumière sursaturante
est donnée, on atteint le niveau de fluorescence Fm’ (fluorescence maximale à la lumière).
L’augmentation de la fluorescence lors du flash est proportionnelle au nombre de QA oxydées

8
pouvant être réduites. La différence entre Fm et Fm’ (avec Fm > Fm’) est proportionnelle à
l’énergie émise sous la forme de chaleur, qui dépend du gradient de protons. Lorsque la lumière
actinique est éteinte, on atteint un niveau Fo’. Le niveau Fo est récupéré si la feuille est maintenue à
l’obscurité. Le schéma ci-dessous représente les variations de fluorescence d’une feuille selon les
traitements lumineux décrits précédemment.
Figure 3 : Variation de la fluorescence en fonction du temps sur une feuille intacte
La fluorescence sera supprimée par deux phénomènes : le quenching non-photochimique
(émission d’énergie sous forme de chaleur) et le quenching photochimique (lié au nombre de QA
oxydées pouvant être réduites en réalisant la photosynthèse).
- le quenching photochimique qP :
Il correspond à une estimation de la capacité du PSII à atténuer la fluorescence en réalisant la
photochimie. Cette capacité est liée à la concentration de centres ouverts susceptibles d’accepter les
électrons, donc liée à l’état redox de QA. Si la photosynthèse est maximale, la fluorescence ainsi que
l’émission de chaleur sont minimales. On appelle cette suppression quenching photochimique.
Le quenching photochimique se calcule selon la formule suivante : qP = Fq’ / Fv’
Fq’ correspond à la différence entre Fm’ (fluorescence maximale d’une feuille adaptée à la lumière)
et F’ (fluorescence émise par une feuille recevant une lumière actinique). Fv’ correspond à la
différence entre Fm’ et Fo’ (fluorescence minimale d’une feuille adaptée à la lumière).
- le quenching non-photochimique qN ou NPQ :
NPQ estime la capacité du PSII à supprimer la fluorescence en diminuant l’énergie d’excitation par
perte de chaleur. Tous changements dans les mesures de NPQ indiquent un changement dans
l’efficacité de la dissipation thermique. En général, une augmentation de NPQ est le résultat de la
mise en place de mécanismes de protection de la feuille contre des dommages pouvant être induits
par une forte lumière par exemple.

9
NPQ peut être décomposé en trois paramètres :
- qE (quenching énergétique) : est associé au transport de protons vers le lumen sous lumière
forte et donc régule le niveau d’excitation du PSII
- qI (quenching par la photoinhibition) : résulte de la photoinhibition du PSII
- qT (quenching associé aux états de transition) : résulte de la phosphorylation des complexes
recevant la lumière associés au PSII
Généralement, sur des plantes non-stressées exposées à une lumière modérée à saturante, qE est le
paramètre principal.
Afin de diminuer l’énergie lumineuse absorbée en excès, la plante est capable de dissiper
cette énergie sous forme de chaleur. Ce mécanisme met en jeu le cycle des xanthophylles, la
protéine PsbS associée au PSII ainsi qu’une enzyme présente dans le lumen du thylakoïde : la
violaxanthine dé-époxidase (VDE) (cf Fig. 4).
Une forte intensité lumineuse induit un flux électronique important entre PSII et PSI, ce qui
provoque l’acidification du lumen (accumulation de protons H+ transportés depuis le stroma vers le
lumen par le pool de PQ). Il en résulte l’activation de la VDE qui se fixe à la membrane du
thylakoïde et convertit la violaxanthine en zéaxanthine, induisant la protonation de PsbS (cf Fig. 4
et 5). Ceci provoque un changement conformationnel des antennes liées au PSII, la quantité de
chaleur émise par le PsbS augmente, et qE augmente. Lorsque l’intensité lumineuse diminue, la
déprotonation de PsbS et la reconversion de la zéaxanthine en violaxanthine provoque une
diminution de qE.
C’est ce que l’on appelle le quenching non-photochimique (la photochimie n’intervient pas
dans cette extinction puisque la lumière sursaturante ferme tous les centres). La diminution de Fm’
à la lumière (Fm’< Fm) est due à l’augmentation de la dissipation de chaleur.
Figure 4 : organisation du PSII et son rôle dans la dissipation de l’énergie en excès par émission de
chaleur

10
Figure 5 : cycle des xanthophylles
Le quenching non-photochimique se calcule de la façon suivante : NPQ = (Fm-Fm’) / Fm’.
1.1. b. Description du matériel utilisé
- Arabidopsis thaliana : un écotype contrôle Col0 et deux écotypes alpins Jo et J3E
- DBMIB (2,5-dibromo-3-methyl-6-isopropyl-p-benzoquinone) et DCMU (3-(3,4-
dichlorophenyl)-1,1-dimethylurea) : inhibiteurs du transport des électrons au niveau de la
chaine photosynthétique (voir principe des inhibiteurs en page 12)
- Mini-PAM (pulse amplitude modulated fluorometre) relié à une source lumineuse
- Un enregistreur (LINSEIS) L70 25 11
Figure 6 : Matériel utilisé pour les mesures de fluorescence chlorophyllienne
Mini -PAM
Source lumineuse
Enregistreur

11
1.1. c. Description du protocole expérimental
Après acclimatation à l’obscurité pendant 30 minutes, une feuille d’Arabidopsis thaliana est placée
sur la trajectoire du faisceau lumineux, associée à une bande de papier filtre imbibée d’eau distillée
pour éviter son desséchement.
Figure 7 : Détails du dispositif
La feuille est soumise à différentes intensités lumineuses successives (10, 50, 100, 250
µmol/m2/s), chacune des séries lumineuses étant séparée par une minute d’obscurité. Un flash de
lumière sursaturante est également programmé pour émettre toutes les 5minutes. Le mini-PAM
enregistre les variations de fluorescence émises par la feuille, retransmises ensuite à l’enregistreur
qui fournit un enregistrement papier de ces variations. A partir de cet enregistrement papier seront
relevées les différentes valeurs nécessaires aux calculs du qP et du NPQ.
La feuille subit ensuite un traitement au DBMIB (immersion dans une solution de DBMIB
25 µM et passage sous vide). Les séries lumineuses sont répétées de la même façon que
précédemment.
La feuille subit pour finir un traitement au DCMU (immersion dans une solution de DCMU
50 µM et passage sous vide, puis incubation 10 min (écotype Col0) ou 20 min (écotypes alpins Jo et
J3E)) et les mêmes séries lumineuses sont répétées.
L’objectif étant de montrer l’activité de la protéine PTOX dans des feuilles d’A. thaliana, les
différentes intensités lumineuses ainsi que les concentrations en inhibiteurs ont été choisi de façon à
ne pas entrainer la destruction du PSII (lumière trop forte, concentration trop élevée en inhibiteurs).
En effet, si le PSII est détruit, les flux électroniques circulant dans la chaine de transport seront
inexistants, et aucune observation concernant la protéine PTOX ne pourra être faite sur les
différents écotypes. Le but est de moduler l’activité photosynthétique sans entrainer la destruction
du PSII.

12
Principe des inhibiteurs DBMIB et DCMU :
Le DBMIB et le DCMU sont des inhibiteurs qui agissent au niveau de la chaine de transport
photosynthétique des électrons. Ils sont utilisés dans le même objectif : bloquer le transport des
électrons du PSII vers le PSI, et par conséquent induire la réduction des quinones du PSII. Leur site
d’action est cependant différent : le DBMIB se fixe sur le Cytochrome b6f à la place de la
plastoquinone ; le DCMU (herbicide, nom commercial : diuron) se fixe par contre directement sur
le PSII à la place de QB, ce qui induit la réduction de toutes les QA (accumulation des électrons qui
ne sont plus utilisés dans la chaine de transport). La fluorescence est alors maximale. La
photosynthèse en présence de DCMU est nulle, l’énergie est donc entièrement émise sous forme de
chaleur et de fluorescence.
L’utilisation de ces deux inhibiteurs permettra d’apporter des réponses d’une part concernant
la fonctionnalité de la protéine PTOX et d’autre part sur son rôle dans le maintien du flux
électronique en PSII et PSI permettant la réalisation de la photosynthèse.
Figure 8 : Chaine de transport des électrons entre PSII et PSI
Localisation de la PTOX et des cibles du DBMIB et du DCMU
Les différents résultats obtenus par fluorescence chlorophyllienne (qP et NPQ pour les trois
écotypes et pour chaque traitement : témoin, DBMIB, DCMU) seront analysés par des tests
statistiques de comparaison de moyennes (critère de test t : Student).
DCMU
DBMIB

13
1.2. Calcul de l’ETR (electron transport rate)
Comme nous l’avons vu précédemment, les inhibiteurs utilisés ont pour objectif de bloquer le
transport des électrons au sein de la chaine de transport. Nous allons donc quantifier ce transport
électronique au sein de feuilles non traitées (témoins) et de feuilles traitées au DBMIB et au
DCMU.
L’ETR se calcule de la manière suivante : ETR = ФPSII * PFD * a * (0,5) avec :
- ФPSII : mesure la proportion de lumière absorbée par la chlorophylle associée au PSII et qui
sera utilisée dans la photochimie, et plus particulièrement dans la réduction de QA (=
efficacité du PSII) ;
- PFD * a : lumière absorbée (µmol photon/m2/s) sachant que PFD (Photon Flux Density)
correspond à l’intensité lumineuse reçue par la feuille (10, 50, 100 et 250 µmol/m2/s) et que
a est calculé de la manière suivante : (100% - % T - % R)/100 (avec %T : % lumière reçue
par la feuille et %R : % lumière réfléchie par la feuille) ;
- 0,5 : facteur tenant compte de la répartition de l’énergie entre le PSII et le PSI.
Mesure de ФPSII :
Les résultats obtenus par fluorescence chlorophyllienne (cf chapitre 1.1. Mesure de fluorescence
chlorophyllienne) sont réutilisés ici puisque ФPSII correspond au ratio Fm’ / Fq’ (paramètres
décrits précédemment).
Mesure de a :
Les paramètres T et R s’obtiennent par le biais de la spectrophotométrie. Une feuille par écotype est
soumise à un rayon lumineux. La quantité de lumière transmise et réfléchie par la feuille est
calculée automatiquement pour chaque longueur d’onde entre 400 et 700 nm (300 valeurs par
prise). Il suffit par la suite de calculer la moyenne des %T et %R entre 400 et 700 nm pour chaque
écotype, puis de faire le calcul (100% - %T - %R)/100 pour obtenir les trois valeurs de a.
Les résultats obtenus seront également traités par des tests statistiques de comparaison de moyennes
(critère de test t : Student).
1.3. Extraction de la protéine PTOX par Western Blot
L’objectif est de quantifier la protéine PTOX présente dans les trois écotypes d’A. thaliana, et de
déterminer si des conditions de stress (faible ou forte température, forte lumière) peuvent entrainer
des modifications dans la quantité de cette protéine.

14
Des feuilles d’Arabidopsis thaliana (écotype Col0 et écotypes alpins Jo et J3E) ont subi au
préalable différents traitements de lumière ou de température : pour chaque écotype, quatre
traitements différents sont réalisés :
- aucun traitement : feuilles témoins (lumière reçue : 80 µmol/m2/s)
- exposition à une plus forte lumière (120 µmol/m2/s) pour Col0 uniquement
- traitement à faible température (4°C, lumière reçue : 100 µmol/m2/s) pendant 5 jours
- traitement à forte température (33°C, lumière reçue : 100 µmol/m2/s) pendant 4 jours
Les feuilles prélevées sont immédiatement congelées dans de l’azote liquide.
Méthode d’extraction protéique par Western Blot :
Les principales étapes sont les suivantes :
1. Préparation des échantillons : les feuilles préalablement congelées sont broyées dans l’azote
liquide, et le broyat est suspendu dans des solutions tampon. Les protéines membranaires
sont séparées des protéines solubles par centrifugations successives.
2. Electrophorèse : Les protéines migrent successivement dans un gel de concentration à 3%
de polyacrylamide (0,1% SDS), puis dans un gel de séparation à 13% (0,1% SDS). Dans le
champ électrique, les protéines sont donc séparées en fonction de leur masse et non en
fonction de leur charge électrique.
3. Transfert sur membrane de nitrocellulose : Les protéines sont électrotransférées sur une
membrane de nitrocellulose dans un appareil de transfert Biorad.
4. Immunodétection : Incubation en présence d’anticorps primaires anti-PTOX, puis
d’anticorps secondaires anti-Ig de lapin.
5. Révélation sur films photographiques : Dans une chambre noire, la membrane est mise à
incuber dans une solution de révélation (kit de chimioluminescence). Un film
photographique est ensuite placé au niveau de la membrane, mais le contact direct est évité à
l’aide d’un film plastique inséré entre les deux. Après un court instant, on réalise le
développement du film photographique.
Le détail des cinq étapes est disponible en Annexe 1.
15
II. Résultats :
2.1. Mesure de fluorescence chlorophyllienne : calculs des qP et NPQ
Pour chaque écotype d’A. thaliana, 4 à 6 répétitions sont effectuées. Les résultats obtenus sont
traités sous le logiciel Excel. La moyenne des valeurs de qP et NPQ obtenues pour chaque intensité
lumineuse (10, 50, 100, 250 µmol/m2/s) est calculée.
2.1. a. Quenching photochimique qP
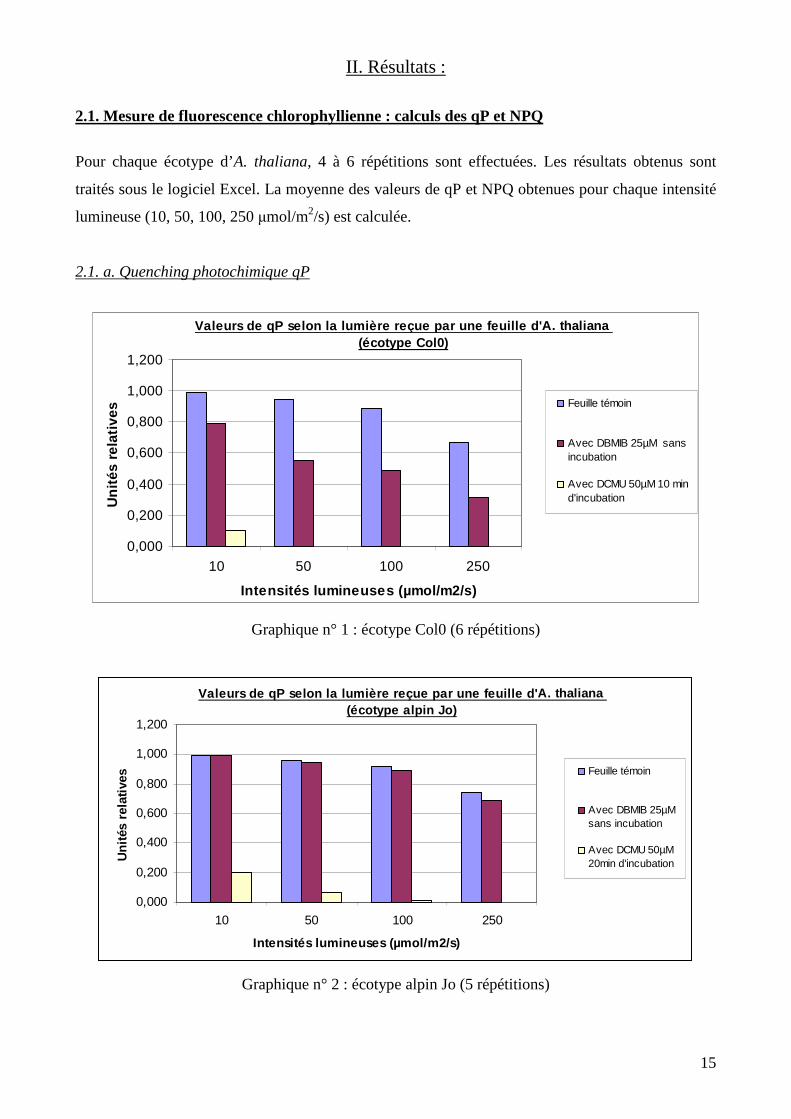
Graphique n° 1 : écotype Col0 (6 répétitions)
Graphique n° 2 : écotype alpin Jo (5 répétitions)
Valeurs de qP selon la lumière reçue par une feuill e d'A. thaliana (écotype alpin Jo)
0,000
0,200
0,400
0,600
0,800
1,000
1,200
10 50 100 250
Intensités lumineuses (µmol/m2/s)
Uni
tés
rela
tives
Feuille témoin
Avec DBMIB 25µMsans incubation
Avec DCMU 50µM20min d'incubation
Valeurs de qP selon la lumière reçue par une feuill e d'A. thaliana (écotype Col0)
0,000
0,200
0,400
0,600
0,800
1,000
1,200
10 50 100 250
Intensités lumineuses (µmol/m2/s)
Uni
tés
rela
tives
Feuille témoin
Avec DBMIB 25µM sansincubation
Avec DCMU 50µM 10 mind'incubation
16
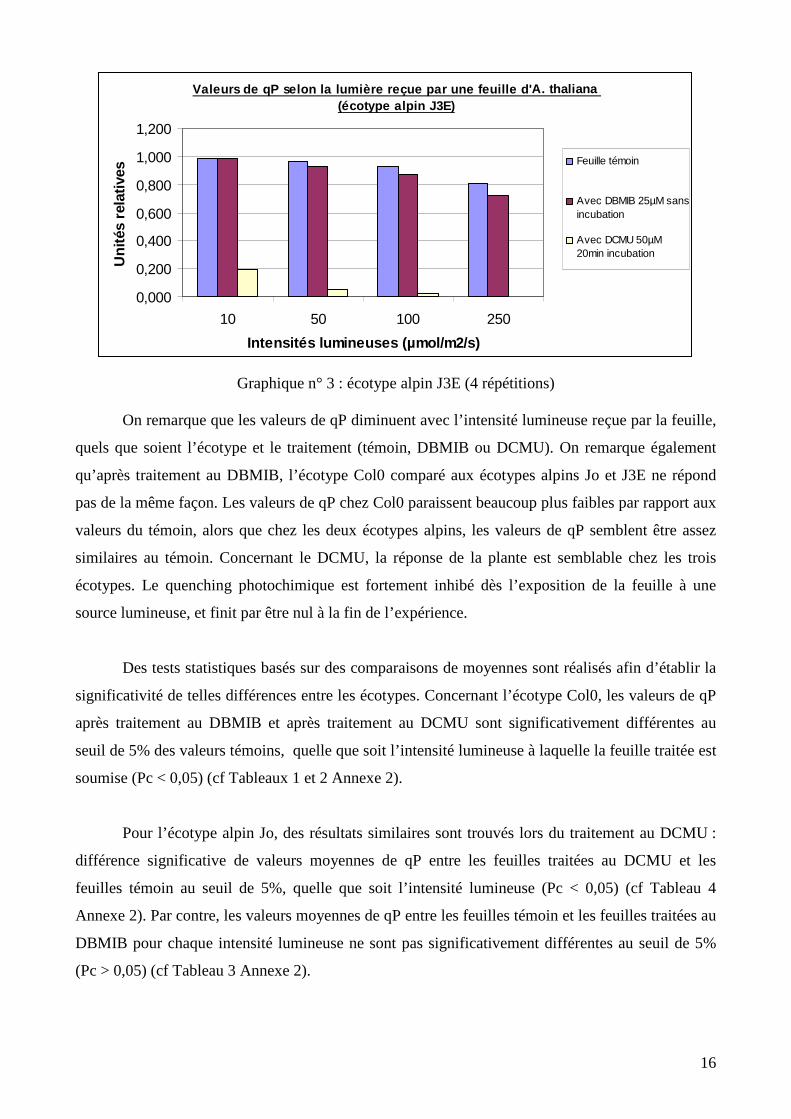
Graphique n° 3 : écotype alpin J3E (4 répétitions)
On remarque que les valeurs de qP diminuent avec l’intensité lumineuse reçue par la feuille,
quels que soient l’écotype et le traitement (témoin, DBMIB ou DCMU). On remarque également
qu’après traitement au DBMIB, l’écotype Col0 comparé aux écotypes alpins Jo et J3E ne répond
pas de la même façon. Les valeurs de qP chez Col0 paraissent beaucoup plus faibles par rapport aux
valeurs du témoin, alors que chez les deux écotypes alpins, les valeurs de qP semblent être assez
similaires au témoin. Concernant le DCMU, la réponse de la plante est semblable chez les trois
écotypes. Le quenching photochimique est fortement inhibé dès l’exposition de la feuille à une
source lumineuse, et finit par être nul à la fin de l’expérience.
Des tests statistiques basés sur des comparaisons de moyennes sont réalisés afin d’établir la
significativité de telles différences entre les écotypes. Concernant l’écotype Col0, les valeurs de qP
après traitement au DBMIB et après traitement au DCMU sont significativement différentes au
seuil de 5% des valeurs témoins, quelle que soit l’intensité lumineuse à laquelle la feuille traitée est
soumise (Pc < 0,05) (cf Tableaux 1 et 2 Annexe 2).
Pour l’écotype alpin Jo, des résultats similaires sont trouvés lors du traitement au DCMU :
différence significative de valeurs moyennes de qP entre les feuilles traitées au DCMU et les
feuilles témoin au seuil de 5%, quelle que soit l’intensité lumineuse (Pc < 0,05) (cf Tableau 4
Annexe 2). Par contre, les valeurs moyennes de qP entre les feuilles témoin et les feuilles traitées au
DBMIB pour chaque intensité lumineuse ne sont pas significativement différentes au seuil de 5%
(Pc > 0,05) (cf Tableau 3 Annexe 2).
Valeurs de qP selon la lumière reçue par une feuill e d'A. thaliana (écotype alpin J3E)
0,000
0,200
0,400
0,600
0,800
1,000
1,200
10 50 100 250
Intensités lumineuses (µmol/m2/s)
Uni
tés
rela
tives
Feuille témoin
Avec DBMIB 25µM sansincubation
Avec DCMU 50µM20min incubation
17
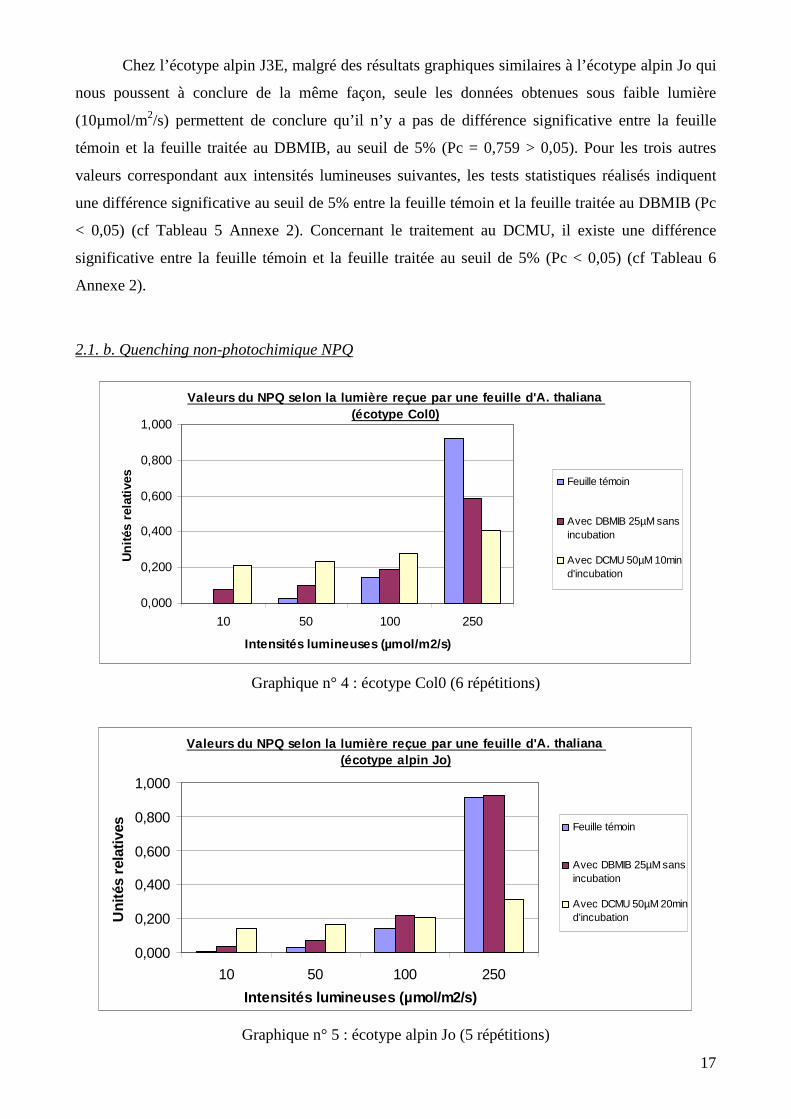
Chez l’écotype alpin J3E, malgré des résultats graphiques similaires à l’écotype alpin Jo qui
nous poussent à conclure de la même façon, seule les données obtenues sous faible lumière
(10µmol/m2/s) permettent de conclure qu’il n’y a pas de différence significative entre la feuille
témoin et la feuille traitée au DBMIB, au seuil de 5% (Pc = 0,759 > 0,05). Pour les trois autres
valeurs correspondant aux intensités lumineuses suivantes, les tests statistiques réalisés indiquent
une différence significative au seuil de 5% entre la feuille témoin et la feuille traitée au DBMIB (Pc
< 0,05) (cf Tableau 5 Annexe 2). Concernant le traitement au DCMU, il existe une différence
significative entre la feuille témoin et la feuille traitée au seuil de 5% (Pc < 0,05) (cf Tableau 6
Annexe 2).
2.1. b. Quenching non-photochimique NPQ
Graphique n° 4 : écotype Col0 (6 répétitions)
Graphique n° 5 : écotype alpin Jo (5 répétitions)
Valeurs du NPQ selon la lumière reçue par une feuil le d' A. thaliana (écotype alpin Jo)
0,000
0,200
0,400
0,600
0,800
1,000
10 50 100 250
Intensités lumineuses (µmol/m2/s)
Uni
tés
rela
tives
Feuille témoin
Avec DBMIB 25µM sansincubation
Avec DCMU 50µM 20mind'incubation
Valeurs du NPQ selon la lumière reçue par une feuil le d' A. thaliana (écotype Col0)
0,000
0,200
0,400
0,600
0,800
1,000
10 50 100 250
Intensités lumineuses (µmol/m2/s)
Uni
tés
rela
tives Feuille témoin
Avec DBMIB 25µM sansincubation
Avec DCMU 50µM 10mind'incubation
18
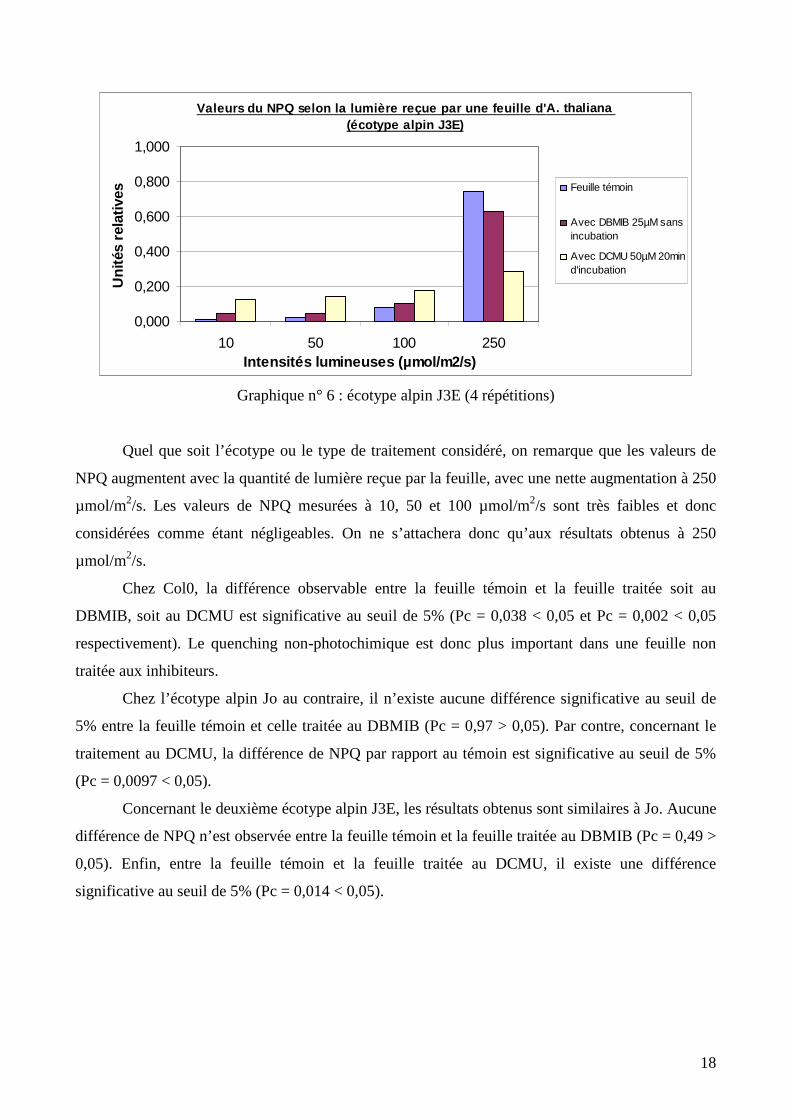
Graphique n° 6 : écotype alpin J3E (4 répétitions)
Quel que soit l’écotype ou le type de traitement considéré, on remarque que les valeurs de
NPQ augmentent avec la quantité de lumière reçue par la feuille, avec une nette augmentation à 250
µmol/m2/s. Les valeurs de NPQ mesurées à 10, 50 et 100 µmol/m2/s sont très faibles et donc
considérées comme étant négligeables. On ne s’attachera donc qu’aux résultats obtenus à 250
µmol/m2/s.
Chez Col0, la différence observable entre la feuille témoin et la feuille traitée soit au
DBMIB, soit au DCMU est significative au seuil de 5% (Pc = 0,038 < 0,05 et Pc = 0,002 < 0,05
respectivement). Le quenching non-photochimique est donc plus important dans une feuille non
traitée aux inhibiteurs.
Chez l’écotype alpin Jo au contraire, il n’existe aucune différence significative au seuil de
5% entre la feuille témoin et celle traitée au DBMIB (Pc = 0,97 > 0,05). Par contre, concernant le
traitement au DCMU, la différence de NPQ par rapport au témoin est significative au seuil de 5%
(Pc = 0,0097 < 0,05).
Concernant le deuxième écotype alpin J3E, les résultats obtenus sont similaires à Jo. Aucune
différence de NPQ n’est observée entre la feuille témoin et la feuille traitée au DBMIB (Pc = 0,49 >
0,05). Enfin, entre la feuille témoin et la feuille traitée au DCMU, il existe une différence
significative au seuil de 5% (Pc = 0,014 < 0,05).
Valeurs du NPQ selon la lumière reçue par une feuil le d' A. thaliana (écotype alpin J3E)
0,000
0,200
0,400
0,600
0,800
1,000
10 50 100 250Intensités lumineuses (µmol/m2/s)
Uni
tés
rela
tives
Feuille témoin
Avec DBMIB 25µM sansincubation
Avec DCMU 50µM 20mind'incubation
19
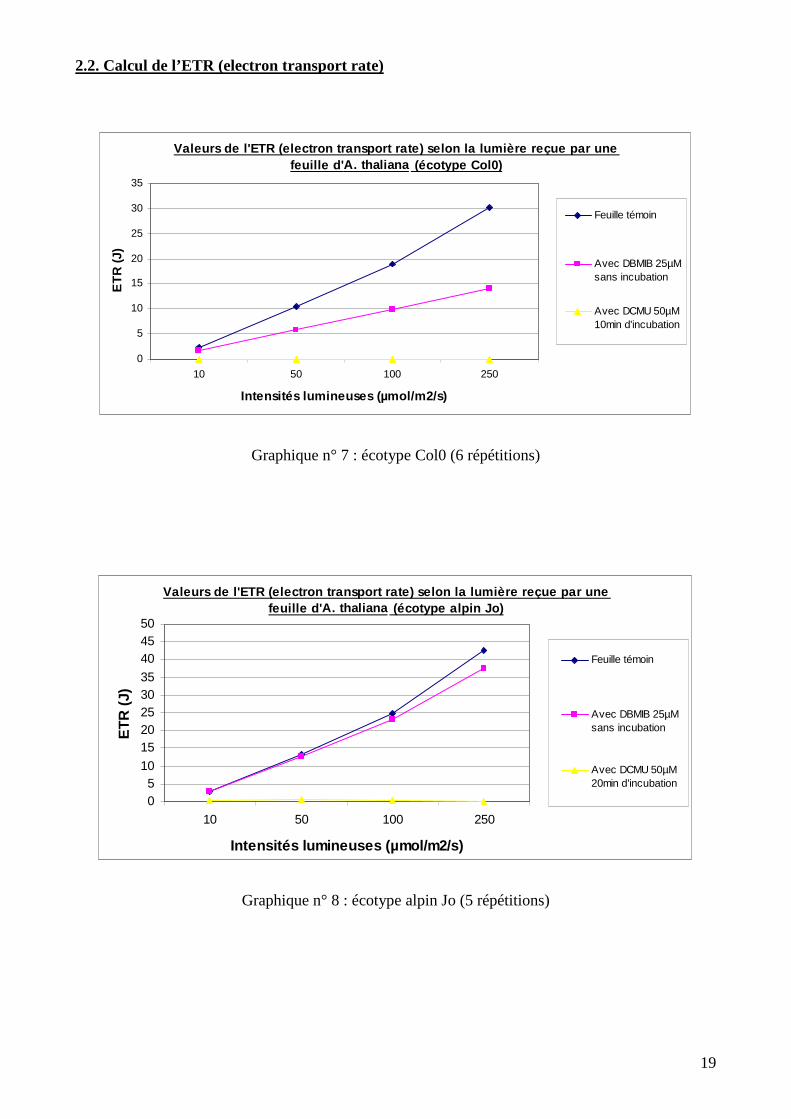
2.2. Calcul de l’ETR (electron transport rate)
Graphique n° 7 : écotype Col0 (6 répétitions)
Graphique n° 8 : écotype alpin Jo (5 répétitions)
Valeurs de l'ETR (electron transport rate) selon la lumière reçue par une feuille d' A. thaliana (écotype alpin Jo)
05
10
1520253035
404550
10 50 100 250
Intensités lumineuses (µmol/m2/s)
ET
R (
J)
Feuille témoin
Avec DBMIB 25µMsans incubation
Avec DCMU 50µM20min d'incubation
Valeurs de l'ETR (electron transport rate) selon la lumière reçue par une feuille d' A. thaliana (écotype Col0)
0
5
10
15
20
25
30
35
10 50 100 250
Intensités lumineuses (µmol/m2/s)
ET
R (
J)
Feuille témoin
Avec DBMIB 25µMsans incubation
Avec DCMU 50µM10min d'incubation
20
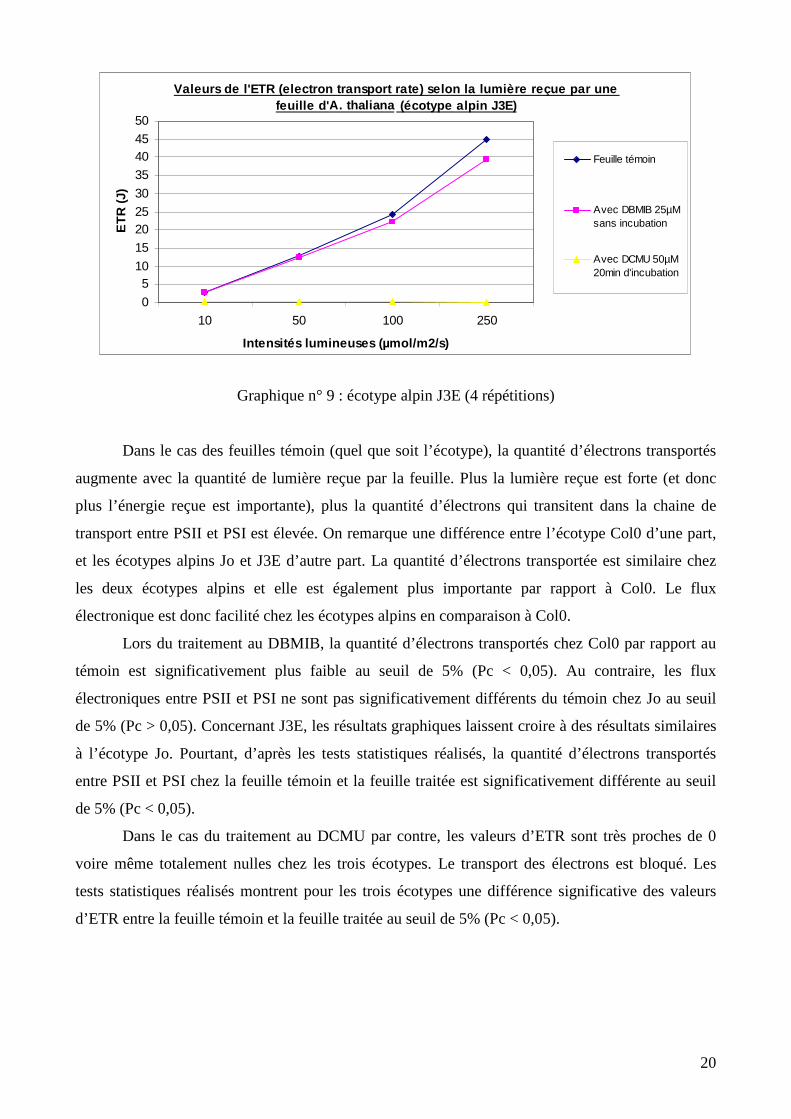
Graphique n° 9 : écotype alpin J3E (4 répétitions)
Dans le cas des feuilles témoin (quel que soit l’écotype), la quantité d’électrons transportés
augmente avec la quantité de lumière reçue par la feuille. Plus la lumière reçue est forte (et donc
plus l’énergie reçue est importante), plus la quantité d’électrons qui transitent dans la chaine de
transport entre PSII et PSI est élevée. On remarque une différence entre l’écotype Col0 d’une part,
et les écotypes alpins Jo et J3E d’autre part. La quantité d’électrons transportée est similaire chez
les deux écotypes alpins et elle est également plus importante par rapport à Col0. Le flux
électronique est donc facilité chez les écotypes alpins en comparaison à Col0.
Lors du traitement au DBMIB, la quantité d’électrons transportés chez Col0 par rapport au
témoin est significativement plus faible au seuil de 5% (Pc < 0,05). Au contraire, les flux
électroniques entre PSII et PSI ne sont pas significativement différents du témoin chez Jo au seuil
de 5% (Pc > 0,05). Concernant J3E, les résultats graphiques laissent croire à des résultats similaires
à l’écotype Jo. Pourtant, d’après les tests statistiques réalisés, la quantité d’électrons transportés
entre PSII et PSI chez la feuille témoin et la feuille traitée est significativement différente au seuil
de 5% (Pc < 0,05).
Dans le cas du traitement au DCMU par contre, les valeurs d’ETR sont très proches de 0
voire même totalement nulles chez les trois écotypes. Le transport des électrons est bloqué. Les
tests statistiques réalisés montrent pour les trois écotypes une différence significative des valeurs
d’ETR entre la feuille témoin et la feuille traitée au seuil de 5% (Pc < 0,05).
Valeurs de l'ETR (electron transport rate) selon la lumière reçue par une feuille d' A. thaliana (écotype alpin J3E)
0
510
15
2025
30
35
4045
50
10 50 100 250
Intensités lumineuses (µmol/m2/s)
ET
R (
J)
Feuille témoin
Avec DBMIB 25µMsans incubation
Avec DCMU 50µM20min d'incubation
21
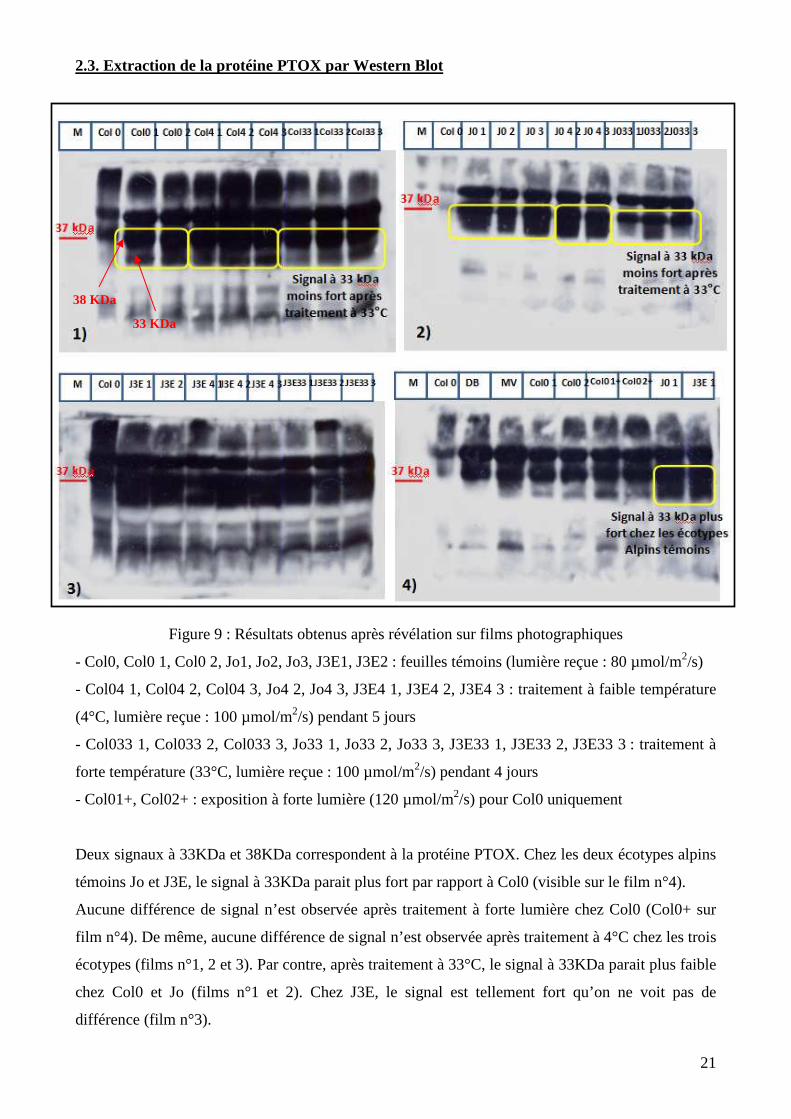
2.3. Extraction de la protéine PTOX par Western Blot
Figure 9 : Résultats obtenus après révélation sur films photographiques
- Col0, Col0 1, Col0 2, Jo1, Jo2, Jo3, J3E1, J3E2 : feuilles témoins (lumière reçue : 80 µmol/m2/s)
- Col04 1, Col04 2, Col04 3, Jo4 2, Jo4 3, J3E4 1, J3E4 2, J3E4 3 : traitement à faible température
(4°C, lumière reçue : 100 µmol/m2/s) pendant 5 jours
- Col033 1, Col033 2, Col033 3, Jo33 1, Jo33 2, Jo33 3, J3E33 1, J3E33 2, J3E33 3 : traitement à
forte température (33°C, lumière reçue : 100 µmol/m2/s) pendant 4 jours
- Col01+, Col02+ : exposition à forte lumière (120 µmol/m2/s) pour Col0 uniquement
Deux signaux à 33KDa et 38KDa correspondent à la protéine PTOX. Chez les deux écotypes alpins
témoins Jo et J3E, le signal à 33KDa parait plus fort par rapport à Col0 (visible sur le film n°4).
Aucune différence de signal n’est observée après traitement à forte lumière chez Col0 (Col0+ sur
film n°4). De même, aucune différence de signal n’est observée après traitement à 4°C chez les trois
écotypes (films n°1, 2 et 3). Par contre, après traitement à 33°C, le signal à 33KDa parait plus faible
chez Col0 et Jo (films n°1 et 2). Chez J3E, le signal est tellement fort qu’on ne voit pas de
différence (film n°3).
38 KDa
33 KDa

22
III. Discussion
La variation des valeurs de qP (quenching photochimique), NPQ (quenching non-
photochimique) et ETR (electron transport rate) selon l’écotype considéré, selon l’intensité
lumineuse reçue par la feuille et enfin selon le traitement (témoin, DBMIB ou DCMU) sont à mettre
en relation puisque ces trois paramètres dépendent notamment du flux électronique circulant au
niveau de la chaine de transport photosynthétique. On observe d’une manière globale que les
valeurs de qP (quenching photochimique) diminuent avec l’augmentation de l’intensité lumineuse
reçue par la feuille, contrairement aux valeurs de NPQ et d’ETR qui augmentent (quels que soient
l’écotype et le type de traitement). Rappelons que ces valeurs sont obtenues lors de flash lumineux
saturant, et que l’augmentation de la fluorescence lors de ce flash nous renseigne sur le nombre de
QA oxydées pouvant être réduites. L’augmentation de l’intensité lumineuse reçue par la feuille est à
l’origine d’une quantité plus importante d’énergie arrivant au niveau du PSII. Une quantité plus
importante d’électrons devra donc transiter par le PSII via le pool de plastoquinones (réduction des
QA) afin de réaliser la photosynthèse.
Comment expliquer une diminution du quenching photochimique avec l’augmentation de la
quantité de lumière reçue par la feuille ? Une faible intensité lumineuse sera à l’origine d’un faible
flux d’électrons au niveau du PSII, entrainant la réduction et la réoxydation rapide de QA,
rapidement disponible à nouveau pour réaliser la photosynthèse. Lors d’un flash lumineux saturant,
les électrons en excès seront facilement pris en charge par les QA alors à l’état oxydé (PSII ouvert).
La valeur de qP est donc élevée puisque la disponibilité des QA pour les électrons a permis d’éviter
la fluorescence en réalisant la photosynthèse. De fortes intensités lumineuses seront au contraire à
l’origine d’un fort flux électronique au niveau du PSII. QA est donc rapidement réduite, ce qui
provoque la fermeture du PSII. Lors d’un flash lumineux saturant, le nombre de QA alors oxydées et
disponibles pour prendre en charge les électrons est faible. La photosynthèse au niveau du PSII
n’est plus réalisable, et l’énergie doit donc être consommée d’une autre façon. Par conséquent
l’émission de fluorescence augmente. Les valeurs de qP diminuent avec l’intensité lumineuse,
puisque de plus en plus d’électrons s’accumulent dans les membranes et donc de moins en moins de
QA sont à l’état oxydé. Le système perd progressivement sa capacité à éviter la fluorescence par
photochimie. Le risque pour la plante est l’accumulation d’électrons dans les membranes, à
l’origine de la formation d’oxygène réactif. Ces ROS (espèces d’oxygène réactif), lorsqu’ils
s’accumulent dans les membranes, peuvent provoquer un stress oxydant aux conséquences néfastes
pour la plante : dégradation des membranes, des pigments, destruction de la protéine D1 du PSII
ayant pour conséquence la photoinhibition (inactivation de la photosynthèse sous lumière forte).

23
La diminution de qP avec l’intensité lumineuse est observable chez les trois écotypes
témoins d’A. thaliana étudiés. Cependant, chez les deux écotypes alpins témoins, cette diminution
des valeurs de qP avec l’augmentation de la lumière reçue par la feuille semble être moins
importante que chez l’écotype Col0 (environ deux fois moins entre 10 et 250 µmol/m2/s). Les
écotypes alpins semblent subir moins fortement l’arrivée en excès d’électrons au niveau du PSII.
Les flux électroniques sont probablement plus importants chez ces écotypes.
Chez les feuilles témoins, les valeurs d’ETR (et donc le nombre d’électrons qui transitent
dans la chaine de transport) augmentent avec la quantité de lumière reçue par la feuille. En effet, la
lumière reçue est transmise à la feuille sous forme d’énergie, se traduisant par un flux électronique.
Plus la quantité de lumière reçue sera importante, plus la quantité d’électrons circulant dans les
membranes sera élevée. Ces résultats sont à mettre en relation avec le quenching photochimique
décrit précédemment. Contrairement aux valeurs d’ETR, les valeurs moyennes de qP chutaient avec
l’augmentation de l’intensité lumineuse reçue par la feuille. Plus le flux électronique est important
(ETR élevé), plus la quantité de QA oxydées diminue et donc plus les valeurs de qP diminuent.
On observe également que les valeurs d’ETR calculées dans les feuilles témoins sont plus élevées
chez les deux écotypes alpins Jo et J3E par rapport à Col0 (d’un facteur 1,5 environ), et qu’elles
sont comparables entre elles. Ces écotypes seraient capables de transporter un flux électronique plus
important lors de la photosynthèse, qui serait alors plus efficace dans des conditions
environnementales difficiles.
D’après les mesures de NPQ effectuées sur les trois écotypes, la quantité d’énergie libérée
sous forme de chaleur est très faible sous lumière faible à moyenne et donc considérée comme
négligeable. En effet, la quantité d’électrons transférés est faible, et par conséquent le gradient de
proton reste faible. L’énergie est utilisée pour la photosynthèse, et l’émission de chaleur (et de
fluorescence) reste faible. Avec l’augmentation de la quantité de lumière reçue par la feuille (250
µmol/m2/s), la quantité d’énergie libérée sous forme de chaleur augmente. En effet, plus la quantité
d’électrons qui transitent dans la chaine de transport est importante, plus la quantité de protons
transportés dans le lumen via le pool de plastoquinones sera importante. Rappelons que
l’acidification du lumen provoque la connexion de la violaxanthine dé-époxidase (VDE) à la
membrane du thylakoïde, aboutissant à une libération de chaleur par la protéine PsbS. Ainsi, des
valeurs élevées de NPQ indiquent que l’énergie est émise en grande partie sous forme de chaleur (et
plus faiblement sous forme de fluorescence, car rappelons que seuls 1 à 3% de l’énergie peuvent
être émis sous cette forme). Le gradient des protons est important. Ainsi, la libération d’énergie
sous forme de chaleur permet à la feuille de supporter de fortes lumières sans subir de dommages
physiologiques.

24
Après traitement au DBMIB, la réponse des plantes varie selon l’écotype considéré. Chez
Col0, après traitement au DBMIB, les valeurs de qP obtenues aux différentes intensités lumineuses
sont significativement plus faibles que celles de la feuille témoin (d’un facteur 2 voire 3).
Rappelons que le DBMIB est un inhibiteur agissant au niveau de la chaine de transport
photosynthétique des électrons. Il bloque le transport des électrons du PSII vers le PSI en se fixant
sur le Cytochrome b6f à la place de la plastoquinone, entrainant la réduction des QA. De ce fait, la
photosynthèse n’est plus réalisable. Les valeurs de qP sont faibles (QA réduites) et la fluorescence
augmente afin de libérer l’énergie en excès. Ces valeurs de qP sont d’autant plus faibles que la
quantité de lumière (et donc d’énergie) absorbée par la feuille est importante. En effet, le nombre de
QA oxydées diminuent progressivement puisque la quantité d’électrons à prendre en charge
augmente, réduisant les QA encore disponibles. Cependant, le quenching photochimique n’est pas
nul, indiquant que des QA sont encore à l’état oxydé. Le but de l’expérience étant de moduler l’état
redox des QA en évitant d’induire la destruction du PSII, la concentration en DBMIB a été calculée
de façon à permettre un faible flux électronique au niveau de la chaine de transport. Ainsi, on peut
supposer que tous les sites accepteurs au niveau des Cytochromes b6f contenus dans les membranes
ne sont pas occupés par l’inhibiteur. Un faible flux d’électrons pourrait circuler, expliquant des
valeurs de qP faibles en présence de l’inhibiteur, mais pas nulles.
Chez l’écotype alpin Jo, contrairement à Col0, après traitement au DBMIB les valeurs de qP
obtenues sous différentes intensités lumineuses ne sont pas significativement différentes de celles
de la feuille témoin. Cela signifie que les électrons ne se sont pas accumulés au niveau du pool de
plastoquinones, malgré le blocage du site accepteur au niveau du Cytochrome b6f. Le flux
électronique est donc maintenu chez Jo, contrairement à Col0. La réoxydation des QA est alors
permise, d’où des valeurs de qP élevées (QA oxydées) et similaires à celles du témoin. Pourtant, ces
électrons n’ont pas pu être consommés par voie photochimique puisque la photosynthèse est
bloquée au niveau du Cytochrome b6f. Une autre voie de consommation des électrons doit donc
exister, et se situer avant le Cytochrome b6f dans la chaine de transport des électrons.
En ce qui concerne l’écotype alpin J3E, les résultats graphiques obtenus sont similaires à Jo,
avec apparemment très peu de différence de qP entre les feuilles témoins et les feuilles traitées au
DBMIB, suggérant que les flux électroniques entre PSII et PSI sont maintenus. Tout comme Jo,
chez J3E un accepteur alternatif permettrait le maintien du flux électronique au niveau de la chaine
de transport, évitant l’accumulation d’électrons et la formation de ROS. Pourtant, chez J3E les tests
statistiques montrent une différence significative entre les valeurs moyennes de qP des feuilles
témoins et traitées, remettant en question l’hypothèse précédente. En raison du faible nombre de
répétitions disponibles, les résultats statistiques sont cependant à interpréter avec précaution. Il
aurait été intéressant de faire d’avantage de répétitions afin d’avoir à traiter un plus grand nombre
25
de données, pouvant ainsi réduire les écarts dus aux aléas des expérimentations (âge de la plante,
pénétration de l’inhibiteur, etc.).
Concernant les valeurs d’ETR, après ajout de DBMIB, de même que lors du calcul de qP, la
réponse des feuilles varie selon l’écotype considéré. Chez Col0, l’ajout de DBMIB provoque une
réduction significative du flux d’électrons dans les membranes. Au contraire, chez l’écotype alpin
Jo, les valeurs d’ETR témoin et après ajout de DBMIB ne sont pas significativement différentes au
seuil de 5%, indiquant que le flux électronique chez les feuilles traitées est très similaire à celui des
feuilles témoins. Dans le cas de J3E, on pourrait conclure de la même façon que Jo au vue des
résultats graphiques. Les tests statistiques indiquent pourtant une différence significative des flux
entre les deux feuilles (témoin et traitée). Cependant, la faible robustesse des tests statistiques,
comme il a été expliqué précédemment, pourrait expliquer de tels résultats statistiques.
Chez l’écotype alpin Jo, le flux électronique entre PSII et PSI est donc maintenu malgré le
blocage du transport électronique au niveau du Cytochrome b6f (pas de diminution des valeurs de
qP et d’ETR), contrairement à l’écotype contrôle Col0. L’hypothèse est qu’un système alternatif
jouant le rôle d’accepteur d’électrons serait présent au niveau de la chaine de transport
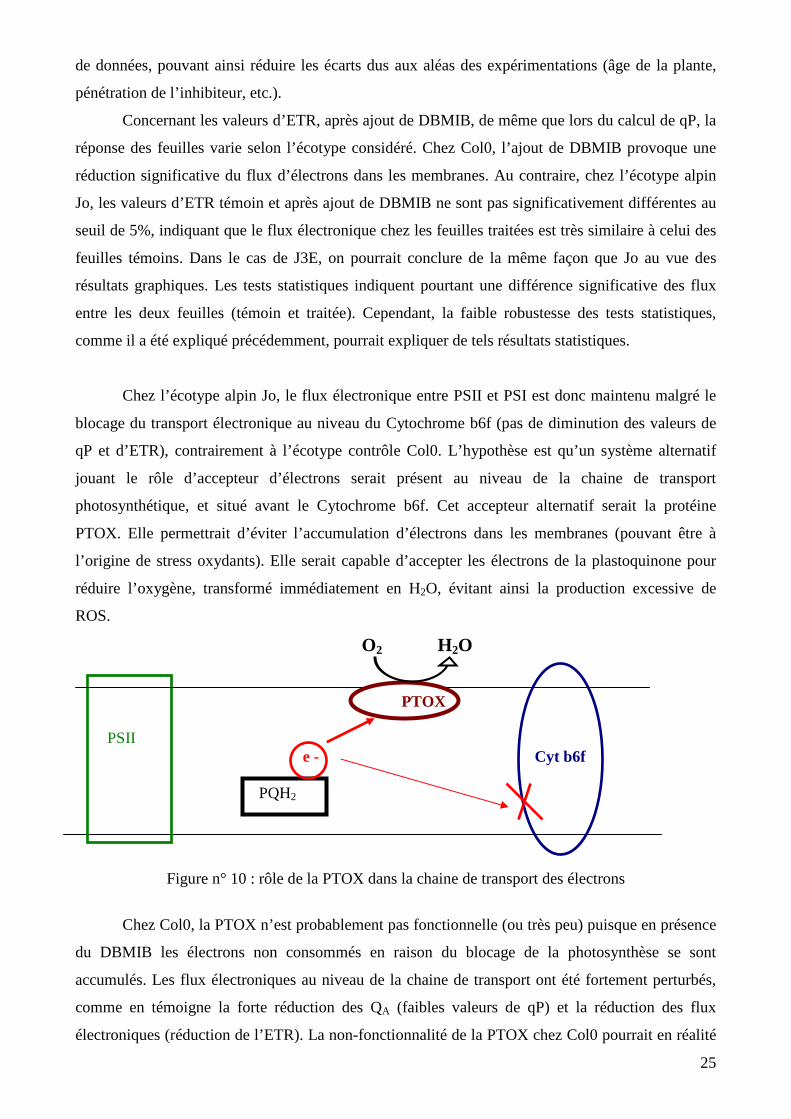
photosynthétique, et situé avant le Cytochrome b6f. Cet accepteur alternatif serait la protéine
PTOX. Elle permettrait d’éviter l’accumulation d’électrons dans les membranes (pouvant être à
l’origine de stress oxydants). Elle serait capable d’accepter les électrons de la plastoquinone pour
réduire l’oxygène, transformé immédiatement en H2O, évitant ainsi la production excessive de
ROS.
O2 H2O
PTOX PSII e - Cyt b6f PQH2
Figure n° 10 : rôle de la PTOX dans la chaine de transport des électrons
Chez Col0, la PTOX n’est probablement pas fonctionnelle (ou très peu) puisque en présence
du DBMIB les électrons non consommés en raison du blocage de la photosynthèse se sont
accumulés. Les flux électroniques au niveau de la chaine de transport ont été fortement perturbés,
comme en témoigne la forte réduction des QA (faibles valeurs de qP) et la réduction des flux
électroniques (réduction de l’ETR). La non-fonctionnalité de la PTOX chez Col0 pourrait en réalité

26
s’expliquer par le fait que cette protéine est présente en plus faible quantité par rapport aux écotypes
alpins (d’après les résultats du Western Blot). Il s’agirait dans ce cas non plus d’une question de
fonctionnalité mais de quantité.
On ne peut pas conclure à la fonctionnalité de cette protéine chez J3E malgré des résultats
graphiques qui, à première vue, nous amenaient à valider cette hypothèse (non confirmée
statistiquement). De plus nombreuses répétitions seraient nécessaires afin de confirmer ou
d’infirmer un éventuel rôle de la PTOX chez cet écotype alpin.
Chez Col 0, après ajout de DBMIB, la quantité de chaleur libérée par la feuille (NPQ)
diminue de façon très significative à 250 µmol/m2/s (environ de moitié). Ce résultat n’est cependant
pas surprenant. En effet, le DBMIB empêche le transfert des électrons du PSII vers le Cytochrome
b6f. Par conséquent, le gradient de protons diminue, l’acidification du lumen est de ce fait moins
importante et la libération d’énergie diminue. Chez les écotypes alpins Jo et J3E par contre, aucune
différence significative n’est observée entre les feuilles témoins et celles traitées au DBMIB. Le
gradient de protons serait ainsi maintenu à un niveau équivalent au témoin. Ceci implique que le
flux d’électrons soit maintenu, non pas par le biais de la chaine de transport (qui est bloquée) mais
par l’intermédiaire d’un accepteur alternatif d’électrons tel que la protéine PTOX. Ces résultats sont
en accord avec les précédentes conclusions concernant les écotypes alpins, et plus particulièrement
Jo. En effet, jusqu’à présent, les tests statistiques effectués sur J3E étaient contraires aux résultats
attendus. Les résultats obtenus pour le quenching non-photochimique confirment donc la possible
implication de la protéine PTOX comme accepteur alternatif d’électrons chez Jo, et remettent en
question la non-significativité des résultats obtenus pour J3E jusqu’à présent et qui ne nous
permettaient pas de conclure de façon similaire.
Les résultats obtenus avec le DCMU sont similaires pour les trois écotypes. Dans les trois
cas, les différences de valeurs de qP entre la feuille témoin et la feuille traitée sont significatives.
Rappelons que le DCMU, en se fixant directement sur le PSII à la place de QB, induit la réduction
de toutes les QA (accumulation des électrons qui ne sont plus transmis dans la chaine de transport).
La photosynthèse en présence de DCMU est donc nulle, l’énergie est alors entièrement émise sous
forme de chaleur et de fluorescence qui est maximale. Les valeurs de qP sont donc très faibles,
voire même nulles sous les plus fortes intensités lumineuses (QA réduites en quasi-totalité voire en
totalité). Dans ce cas là, la PTOX est inefficace puisque l’action de l’inhibiteur se situe en amont.
Après traitement au DCMU, les flux électroniques au sein de la chaine de transport sont
considérés comme étant nuls (ETR très proche ou égale à 0) chez les trois écotypes. Les électrons
sont bloqués au niveau du PSII et ne peuvent plus circuler jusqu’au PSI. La PTOX étant située en

27
aval du PSII, elle ne peut agir sur les flux électroniques. Ceci est en accord avec les précédentes
observations concernant le quenching photochimique.
L’ajout de DCMU, bloquant le transfert électronique au niveau du PSII, aurait dû provoquer
un arrêt de l’émission de chaleur chez les trois écotypes, par blocage de la circulation des protons
entre le stroma et le lumen. Pourtant, les valeurs de NPQ mesurées à 250 µmol/m2/s ne sont pas
nulles (bien que peu élevées). La PTOX étant ici inefficace, l’hypothèse envisageable est que les
sites de fixation du DCMU n’ont pas tous été occupés, permettant le passage d’une petite quantité
d’électrons et expliquant le faible gradient de protons (ce qui est très probable puisque la
concentration en DCMU a été choisi afin de ne pas provoquer la destruction du PSII).
Ces résultats nous permettent de vérifier deux éléments : d’une part la capacité de
pénétration des inhibiteurs au sein des structures membranaires et leur fixation sur les sites
spécifiques. En effet, le fait qu’il n’y ait visiblement pas d’impact du DBMIB sur le flux
électronique chez Jo et J3E (non vérifié statistiquement chez J3E) aurait pu s’expliquer par une
absence de pénétration de l’inhibiteur dans la feuille. Ceci n’est très probablement pas le cas
puisque dans des conditions similaires (mise sous vide et incubation quelques dizaines de minutes)
le DCMU a par contre un effet sur le flux électronique, en bloquant le flux au niveau du PSII chez
les trois écotypes. Ceci apporte la preuve d’une bonne pénétration des inhibiteurs et de leur fixation
sur les sites spécifiques. De plus, des expériences complémentaires effectuées en parallèle dans le
laboratoire ont montré qu’en présence de DBMIB, la réoxydation du PSI est moins rapide comparée
au témoin mais plus rapide qu’en présence de DCMU (résultats non communiqués). Cela indique
que non seulement le DBMIB a bien bloqué le transport d’électrons au niveau réducteur du
Cytochrome b6f, mais aussi que l'hypothèse d'un flux cyclique d'électrons entre le Cytochrome b6f
et le PSI n'est pas à exclure. Ce flux cyclique est probablement mis en place autour du PSI afin de
permettre l’élimination des électrons par formation de NADPH.
Et d’autre part ces résultats renforcent l’hypothèse selon laquelle la protéine PTOX serait
impliquée en tant qu’accepteur alternatif chez l’écotype alpin Jo (et probablement chez J3E). En
effet, le fait de bloquer le flux électronique d’abord en aval (Cyt b6f, DBMIB) puis en amont (PSII,
DCMU) de la PTOX permet d’établir une preuve que la PTOX agit effectivement comme accepteur
d’électrons. Les flux sont maintenus dans le premier cas, alors qu’ils sont inhibés dans le second
cas. Le DCMU a ici un rôle de contrôle. Un autre élément en faveur de cette hypothèse est que, en
présence d’une concentration deux fois plus élevée de DBMIB (50µM au lieu de 25µM) et d’un
temps d’incubation trois fois plus long (60min au lieu de 20min), les résultats obtenus chez
l’écotype alpin Jo sont similaires à ceux obtenus précédemment (résultats non communiqués).

28
Ainsi, les différentes expériences basées sur la fluorescence chlorophyllienne permettent de
mettre en évidence que le flux électronique entre PSII et PSI est favorisé chez les écotypes alpins
contrairement à l’écotype contrôle Col0. Ceci serait lié à la présence d’un accepteur d’électrons
localisé entre le PSII et le Cytochrome b6f, capable de prendre en charge l’excès d’électrons et
d’éviter le stress oxydant (formation de ROS en excès). Cet accepteur serait la protéine PTOX.
Les résultats du Western Blot nous montrent deux signaux correspondant à la protéine
PTOX, à 33KDa et 38KDa. Ces deux formes correspondraient respectivement à la forme mature et
à la forme non-mature de la protéine. La forme non mature (38KDa) contiendrait un peptide de
transfert permettant son adressage au chloroplaste. Une fois passé l’enveloppe chloroplastique, ce
peptide, d’une taille de 5KDa environ, serait éliminé, libérant la forme mature de la protéine (33
KDa). De plus, on remarque que la différence de taille entre la forme non-mature et la forme mature
(5KDa) correspond à la taille de ce peptide. Ceci n’est encore qu’une hypothèse. Des
expérimentations sont actuellement en cours au laboratoire d’Ecophysiologie Végétale (Orsay) afin
de vérifier cette hypothèse. Il s’agit d’isoler des chloroplastes et de les soumettre à l’action d’une
protéinase, afin de voir si la suppression du signal à 38KDa est possible, car on suppose que la
protéine non-mature reste à l’extérieur du chloroplaste. En éliminant l’extérieur du chloroplaste, on
élimine de ce fait la forme non-mature, mais pas la forme mature à l’intérieur du chloroplaste.
Les résultats obtenus chez les deux écotypes alpins Jo et J3E (signal plus fort à 33KDa,
forme mature) montrent que la quantité de protéine PTOX est plus importante dans les écotypes
alpins comparés à Col0. Ces résultats soutiennent l’hypothèse que la PTOX jouerait un rôle chez
ces écotypes en permettant un flux d’électrons plus important au niveau de la chaine de transport
photosynthétique. La non-significativité des résultats de qP et d’ETR chez J3E, ne permettant pas
de conclure à la présence d’une protéine PTOX fonctionnelle, est ainsi fortement remise en question
(tests probablement peu robustes). Les plantes sont soumises à différents traitements de température
et de lumière, l’objectif étant d’observer leur réponse face à un stress. Après traitement à 33°C,
seule la forme mature est apparemment dégradée (signal moins fort à 33KDa par rapport au témoin)
chez Col0 et Jo. Ceci reste pour l’instant inexpliqué, à savoir pourquoi seule la forme mature aurait
été dénaturée à forte température. Après traitement à 4°C, aucune différence avec les feuilles
témoins n’a été observée chez les trois écotypes. Chez les deux écotypes alpins, on pourrait faire
l’hypothèse suivante : qu’ils soient soumis à une faible température dès le début de leur
développement ou seulement à partir d’un stade de développement avancé, cela ne change pas le
fait qu’ils soient aptes à résister à ce type de stress (grande plasticité). D’où un signal à 33KDa de
même intensité que le témoin. Cependant, le fait que l’on trouve des résultats similaires chez
l’écotype Col0 n’est pas en accord avec cette hypothèse, puisque Col0 n’est en principe pas capable
de s’acclimater pour résister à de faibles températures. Les résultats obtenus après traitement à forte

29
lumière ne nous permettent pas non plus de conclure, puisque aucune différence avec le témoin n’a
été observée chez Col0. Ainsi, les différents traitements appliqués sur les trois écotypes (forte et
faible température, forte lumière) ne nous permettent pas de conclure à un éventuel rôle de la PTOX
dans leur capacité d’adaptation à des conditions environnementales contraignantes.
Conclusion
Ainsi, la PTOX jouerait le rôle d’accepteur alternatif d’électrons au sein de la chaine de
transport entre les photosystèmes II et I. Elle permettrait de maintenir un flux électronique entre
PSII et PSI, afin que la photosynthèse ne soit pas saturée à forte lumière, et également d’éviter un
stress oxydant à l’origine de destructions membranaires pouvant conduire à la mort de la plante. La
PTOX n’est cependant fonctionnelle que chez certaines plantes, les plantes alpines notamment
comme on a pu le montrer ici avec les écotypes alpins d’A. thaliana Jo et J3E (par comparaison
avec l’écotype contrôle Col0). Ceci n’a cependant pas été totalement confirmé statistiquement chez
J3E. Il aurait été utile de faire un plus grand nombre de répétitions sur chaque écotype afin de
pouvoir obtenir des résultats statistiques plus robustes. On peut néanmoins faire l’hypothèse que les
plantes alpines poussant dans des environnements de hautes altitudes très contraignants ont
développé des systèmes de protection très efficaces, avec notamment la présence d’une protéine
PTOX fonctionnelle.
Cette protéine n’est pas présente en même quantité chez toutes les plantes alpines. On sait
par exemple qu’elle est abondante chez Ranunculus glacialis, et qu’elle joue un rôle majeur par
rapport aux autres systèmes antioxydants (Streb et al, 2005). Il serait intéressant de faire cette étude
chez les écotypes alpins d’A. thaliana afin de comparer l’efficacité de la PTOX aux autres systèmes
de protection. Enfin, il pourrait être intéressant d’appliquer la méthode utilisée lors du stage sur des
feuilles de R. glacialis afin de confirmer la présence d’une protéine PTOX fonctionnelle chez cette
espèce alpine, malgré de nombreuses propriétés uniques le suggérant : concentration en PTOX bien
plus importante que dans d’autres espèces alpines, d’autres espèces de Ranunculaceae ou encore
dans des feuilles de tomates transgéniques sur-exprimant la PTOX ; découverte d’un ARNm dans
des feuilles matures de R. glacialis ; enfin forte concentration en NADH DH, une enzyme associée
à la PTOX dans la chlororespiration (Streb et al, 2005). Cette méthode pourrait être appliquée à
d’autres espèces de plantes alpines, avec probablement quelques ajustements (retrait de l’épiderme
coriace chez S. alpina par exemple pour favoriser la pénétration des inhibiteurs).

30
Bibliographie
� Baker (2008) : Chlorophyll fluorescence : a probe of photosynthesis in vivo – Plant Biology
59 : 89-113
� Bennoun (1982) : Evidence for a respiratory chain in the chloroplast – Proc. Natl. Acad.
Sci. USA 79 : 4352-4356
� Bukhov et al (2003) : Interaction of exogenous quinones with membranes of higher plant
chloroplasts : modulation of quinone capacities as photochemical and non-photochemical
quenchers of energy in Photosystem II during light-dark transitions – Biochimica et
Biophysica Acta 1604 : 115-123
� Cornic (2007) : L’émission de fluorescence chlorophyllienne. Régulation de l’activité du
PSII et estimation du flux d’électrons dans les thylakoïdes sur des feuilles intactes
� Demming-Adams & Adams (1993) : The Xanthophyll cycle – CRC Press : 91-110
� Foyer et al (1994) : Photooxidative stress in plants – Physiologia Plantarum 92 : 696-717
� Heber et al (1996) : Photorespiration is essential for the protection of the photosynthetic
apparatus of C3 plants against photoinactivation under sunlight – Botanica Acta 109 : 307-
315
� Maxwell & Johnson (2000) : Chlorophyll fluorescence : a practical guide – Journal of
Experimental Botany Vol 51, No 345 : 659-668
� Peltier & Cournac (2002) : Chlororespiration – Annual Review of Plant Biology 53 : 523-
550
� Quiles (2006) : Stimulation of chlororespiration by heat and high light intensity in oat plants
– Plant, Cell and Environment 29 : 1463-1470

31
� Schreiber & Berry (1977) : Heat-induced changes of chlorophyll fluorescence in intact
leaves correlated with damage of the photosynthetic apparatus - Planta 136 : 233–238
� Streb et al (1997) : Resistance to photoinhibition of photosystem II and catalase and
antioxidative protection in high mountain plants - Plant, Cell and Environment 20 : 1030-
1040
� Streb et al (2005) : Evidence for alternative electron sinks to photosynthetic carbon in th
high mountain plant species Ranunculus glacialis - Plant, Cell and Environment 28 : 1123-
1135
� Tallón & Quiles (2007) : Acclimatation to heat and high light intensity during the
development of oat leaves increases the NADH DH complex and PTOX levels in
chloroplasts – Plant Science 173 : 438-445
� Wildi & Lutz (1996) : Antioxidant composition of selected high alpine plant species from
different altitudes – Plant, Cell and Environment 19 : 138-146
� Yordanov et al (1986) : Mechanisms of the temperature damage and acclimation of the
photosynthetic apparatus - Photobiochemistry and Photobiophysics 12 : 147–155

32
Annexes
Annexe 1 Méthode d’extraction protéique par Western Blot :
1. Préparation des échantillons :
Les feuilles préalablement congelées sont broyées dans l’azote liquide, et le broyat est suspendu
dans 400 µl de solution tampon 1.
Composition de la solution tampon 1 : 27ml de Tris-HCl pH8 100mM, 3g de sucrose, 3ml NaCl
1,5M, 60µl βmercaptoéthanol 0,2% et 30 µl Pefablock 0,5M.
Les échantillons sont placés sous faible agitation à 4°C pendant 15 minutes, puis centrifugés à 4°C
et 15.000 rpm pendant 20 minutes. Le surnageant contient les protéines solubles. Le culot est
solubilisé dans 400µl de solution tampon 2.
Composition de la solution tampon 2 : 27ml de Tris-HCl pH8 100mM, 3g de sucrose, 60µl
βmercaptoéthanol 0,2%, 3ml de SDS 20% et 300µl de Triton.
Les échantillons sont placés sous centrifugation à 4°C et 15.000 rpm pendant 20 minutes.
Le culot est à nouveau solubilisé dans la solution tampon 2, mis sous agitation puis centrifugation
dans les mêmes conditions que précédemment. Le surnageant contient les protéines membranaires.
2. Electrophorèse :
Les protéines migrent successivement dans un gel de concentration à 3% de polyacrylamide (0,1%
SDS), puis dans un gel de séparation à 13% (0,1% SDS). Dans le champ électrique, les protéines
sont donc séparées en fonction de leur masse et non en fonction de leur charge électrique.
Dans un premier temps, les plaques en verre sont lavées à l’éthanol, et l’étanchéité du système est
vérifiée. Puis le gel de séparation est coulé jusqu’à quelques mm en dessous de la limite du peigne,
suivi de quelques mm d’eau additionnée de SDS (0,5%), ce qui aplanit le gel et l’isole de l’oxygène
de l’air. La prise du gel demande 45 minutes. L’eau de surface est retirée, puis le gel de
concentration est coulé jusqu’aux rebords, et les peignes sont placés immédiatement. La prise dure
également 45 minutes. Les peignes sont ensuite retirés, les puits sont repérés à l’aide d’une marque
sur la plaque en verre, et le système est placé dans la cuve d’électrophorèse remplie de tampon.
Les puits sont chargés avec 20-40µl de protéines additionnées de 50% de tampon de charge
dénaturant SBLU ainsi que de βmercaptoéthanol. Un des puits est chargé avec 3µl de marqueur
protéique Biorad 161-0373. Les protéines migrent sous électrophorèse pendant 1h à 150V dans un
tampon de migration Tris-Glycine-SDS pH 8,5. La migration est terminée quand le colorant est au
bas de la plaque. Les gels sont alors utilisés immédiatement pour le transfert.

33
3. Transfert sur membrane de nitrocellulose :
Les protéines sont électrotransférées sur une membrane de nitrocellulose dans un appareil de
transfert Biorad. Le gel est placé coté anode, et la membrane coté cathode. Les protéines, entrainées
par le courant électrique, vont sortir du gel et se fixer sur la membrane. Après 1h30 de transfert, les
membranes sont placées dans une solution de saturation contenant du lait écrémé (20mM Tris-HCl
pH7,4 ; 0,9% NaCl ; 5% Lait écrémé ; 0,1% Tween) pendant 45 minutes à température ambiante.
Ainsi, les protéines du lait vont saturer les sites de fixation libres, étape indispensable pour réaliser
par la suite une immunodétection.
4. Immunodétection :
Les membranes sont rincées 3*5min dans du tampon de rinçage (20mM Tris-HCl pH7,4 ; 0,9%
NaCl ; 0,1% Lait écrémé ; 0,1% Tween), puis incubées 1h30 à 2h en présence de l’anticorps
primaire anti-PTOX dilué dans du tampon de rinçage (4ml), sous faible agitation, dans un sachet en
plastique soudé (les bulles d’air sont enlevées régulièrement).
Les membranes sont à nouveau rincées 5*10min dans un tampon de rinçage, puis incubées 45min
en présence de l’anticorps secondaire (2µl par sachet de anti Ig de lapin couplé à la peroxydase
HRP) dilué dans du tampon de rinçage (4ml par sachet). Les membranes sont encore rincées
3*5min dans la solution de rinçage, puis 2*5 min dans une solution de rinçage finale (sans lait et
sans Tween), puis 2*5min dans de l’eau distillée.
5. Révélation sur films photographiques :
Dans une chambre noire, la membrane est mise à incuber dans une solution de révélation (kit de
chimioluminescence). Un film photographique est ensuite placé au niveau de la membrane, mais le
contact direct est évité à l’aide d’un film plastique inséré entre les deux. Après un court instant, on
réalise le développement du film photographique.
34
Annexe 2 Hypothèse nulle Ho : pas de différences significatives entre la feuille témoin et la feuille traitée au seuil de 5%.
Tableau n° 1 : résultats des tests statistiques concernant l’écotype Col0 : comparaison des valeurs moyennes de qP entre la feuille témoin et la feuille traitée au DBMIB
Tableau n° 2 : résultats des tests statistiques concernant l’écotype Col0 : comparaison des valeurs moyennes de qP entre la feuille témoin et la feuille traitée au DCMU
Intensités lumineuses (µmol/m2/s)
Valeur du critère de test : to
Probabilité critique : Pc
Comparaison au seuil de 5%
Conclusion sur Ho
10 53,998 1,150E-13 Pc < 0,05 rejetée 50 166,246 1,524E-18 Pc < 0,05 rejetée 100 61,734 3,024E-14 Pc < 0,05 rejetée 250 17,322 8,709E-09 Pc < 0,05 rejetée
Tableau n° 3 : résultats des tests statistiques concernant l’écotype alpin Jo : comparaison des valeurs
moyennes de qP entre la feuille témoin et la feuille traitée au DBMIB
Intensités lumineuses (µmol/m2/s)
Valeur du critère de test : to
Probabilité critique : Pc
Comparaison au seuil de 5%
Conclusion sur Ho
10 0,263 0,799 Pc > 0,05 acceptée 50 1,360 0,211 Pc > 0,05 acceptée 100 1,432 0,190 Pc > 0,05 acceptée 250 0,921 0,384 Pc > 0,05 acceptée
Tableau n° 4 : résultats des tests statistiques concernant l’écotype alpin Jo : comparaison des valeurs moyennes de qP entre la feuille témoin et la feuille traitée au DCMU
Intensités lumineuses (µmol/m2/s)
Valeur du critère de test : to
Probabilité critique : Pc
Comparaison au seuil de 5%
Conclusion sur Ho
10 2,434 0,035 Pc < 0,05 rejetée
50 3,569 0,005 Pc < 0,05 rejetée
100 3,853 0,003 Pc < 0,05 rejetée
250 4,096 0,002 Pc < 0,05 rejetée
Intensités lumineuses (µmol/m2/s)
Valeur du critère de test : to
Probabilité critique : Pc
Comparaison au seuil de 5%
Conclusion sur Ho
10 14,255 5,716E-07 Pc < 0,05 rejetée 50 24,630 7,889E-09 Pc < 0,05 rejetée 100 45,809 5,698E-11 Pc < 0,05 rejetée 250 17,095 1,393E-07 Pc < 0,05 rejetée
35
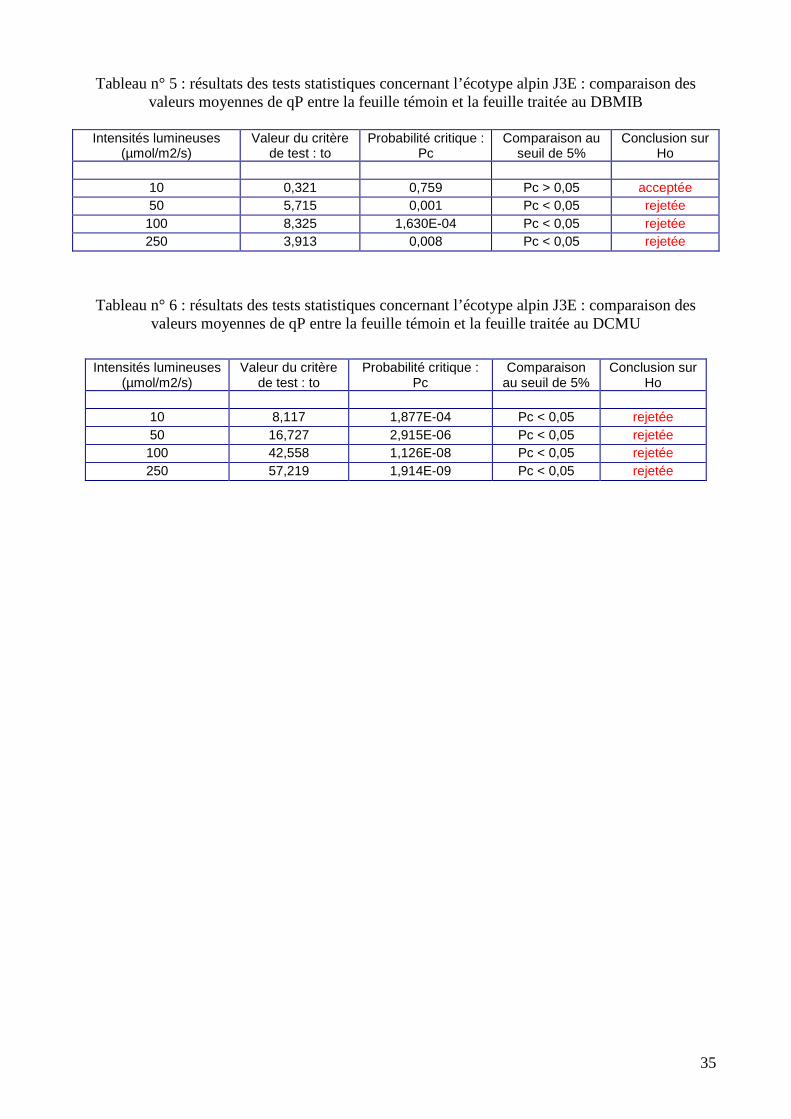
Tableau n° 5 : résultats des tests statistiques concernant l’écotype alpin J3E : comparaison des
valeurs moyennes de qP entre la feuille témoin et la feuille traitée au DBMIB
Intensités lumineuses (µmol/m2/s)
Valeur du critère de test : to
Probabilité critique : Pc
Comparaison au seuil de 5%
Conclusion sur Ho
10 0,321 0,759 Pc > 0,05 acceptée 50 5,715 0,001 Pc < 0,05 rejetée 100 8,325 1,630E-04 Pc < 0,05 rejetée 250 3,913 0,008 Pc < 0,05 rejetée
Tableau n° 6 : résultats des tests statistiques concernant l’écotype alpin J3E : comparaison des valeurs moyennes de qP entre la feuille témoin et la feuille traitée au DCMU
Intensités lumineuses (µmol/m2/s)
Valeur du critère de test : to
Probabilité critique : Pc
Comparaison au seuil de 5%
Conclusion sur Ho
10 8,117 1,877E-04 Pc < 0,05 rejetée 50 16,727 2,915E-06 Pc < 0,05 rejetée 100 42,558 1,126E-08 Pc < 0,05 rejetée 250 57,219 1,914E-09 Pc < 0,05 rejetée

36
Abstract
The PTOX protein is localized in the photosynthetic electron transport chain between the
PSII complex and the Cytochrome b6/f complex and can transfer electrons from plastoquinone to
molecular oxygen. In plants exposed to stress conditions like high irradiation and low temperature
as e.g. in alpine plants, the PTOX protein could alleviate photooxidative damage by reducing the
redox state of the photosynthetic electron transport chain. In order to demonstrate the PTOX
activity in vivo in Arabidopsis thaliana ecotypes (Col 0 as control and two alpine ecotypes Jo and
J3E) the reduction state of the PSII electron acceptor QA, the heat emission (NPQ) and the electron
transport rate (ETR) were characterized in leaves before and after blocking the photosynthetic
electron transport at the level of Cytochrome b6/f and at the level of PSII. The results show that
blocking the electron transport at the level of Cytochrome b6/f increase the reduction state of QA,
decrease the NPQ and the ETR at PSII in Col 0 but have only minor effects in the two alpine
varieties. In contrast, blocking the electron transport at the level of PSII blocks the electron
transport in all ecotypes at the same extent. These results suggest the presence of an alternative
electron acceptor between PSII and Cytochrome b6/f in the alpine ecotypes but not in Col 0.
Furthermore, Western Blot analysis shows that the amount of PTOX protein is more important in
two alpine ecotypes than in Col0. In conclusion, the PTOX protein will be functional only in alpine
ecotypes. This protein will be able to maintain an electron flux in photosynthetic transport chain
between PSII and PSI, to realise photosynthesis and to avoid accumulation of electrons originally
oxidative stress.

37
Résumé
La protéine PTOX est localisée au niveau de la chaine photosynthétique de transport
d’électrons entre le photosystème II et le Cytochrome b6f. Elle peut transférer des électrons de la
plastoquinone à l’oxygène moléculaire. Dans les plantes exposées à des conditions de stress telles
qu’une forte lumière ou une faible température, par exemple les plantes alpines, la protéine PTOX
pourrait réduire les dommages oxydants en diminuant l’état redox de la chaine photosynthétique de
transport. Afin de démontrer l’activité de la PTOX in vivo dans des écotypes d’Arabidopsis
thaliana (un écotype contrôle Col0 et deux écotypes alpins Jo et J3E), l’état de réduction de QA (site
accepteur d’électrons au niveau du PSII), l’émission de chaleur (NPQ) et la quantité d’électrons
transportés (ETR) sont mesurés dans les feuilles avant et après le blocage du transport électronique
au niveau du Cytochrome b6f et au niveau du PSII. Les résultats montrent que bloquer le transport
électronique au niveau du Cytochrome b6f provoque une réduction de QA, ainsi qu’une diminution
de NPQ et ETR chez Col0, mais a peu de conséquences sur les écotypes alpins. Au contraire, un
blocage au niveau du PSII empêche le transport électronique entre PSII et PSI chez les trois
écotypes, de façon similaire. Ces résultats suggèrent la présence d’un accepteur alternatif
d’électrons entre le PSII et le Cytochrome b6f dans les écotypes alpins, et pas chez Col0. De plus,
les extractions protéiques par Western Blot montrent que la quantité de protéine PTOX est plus
importante dans les écotypes alpins comparés à Col0. Pour conclure, la protéine PTOX serait
fonctionnelle uniquement chez les écotypes alpins. Elle permettrait de maintenir un flux
électronique au niveau de la chaine de transport entre les photosystèmes II et I, nécessaire à la
réalisation de la photosynthèse et évitant l’accumulation d’électrons à l’origine du stress oxydant.






















![[MétroPole] Principe du Block Automatique Lumineux](https://img.pdfslide.fr/doc/110x75/55cf9d52550346d033ad1e23/metropole-principe-du-block-automatique-lumineux.jpg)