Embed Size (px)
Citation preview

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 90
Faculté des Sciences
Laboratoire de Zoologie
Structuration de l’espace de vol en fonction du vent chez Bombus terrestris (L.)
(Hymenoptera, Apidae) Promoteur: Prof. P. Rasmont
Mémoire de fin d’étude présenté par Benoît Namur en vue de l'obtention du diplôme de Licencié en Sciences Biologiques
Année académique 2004-2005

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 89
Namur B., 2005. Structuration de l’espace de vol en fonction du vent chez Bombus terrestris (L.) (Hymenoptera, Apidae). Mémoire de Licence, Université de Mons-Hainaut, XXpp. + X planche.
Résumé: La structuration de l’espace de vol a été souvent étudiée par divers auteurs sur le terrain ou par le biais de modèles. Cependant les influences de certains paramètres météorologiques sur la structuration de l’espace de vol des bourdons restent peu connues et en particulier les effets du vent. C’est pourtant un facteur qui influence fortement le vol, que ce soit celui des avions ou des animaux.
Cette étude s’est déroulé au Pla del Bac (France, Pyrénées-Orientales, Eyne) durant le mois de juillet 2004. Le Pla del Bac est caractérisé par des changements réguliers du sens du vent. En observant les changements éventuels de site de butinage lors des différents vents, on peut comprendre l’effet du vent sur la structuration de l’espace de vol chez les bourdons.
Des colonies d’élevages sont disposées sur un site exposé à des vents de directions et de vitesses variables. La zone d’étude est un cercle de 500 m de rayon centré sur les nids. Les ressources florales sont dispersées et réparties en abondance dans toutes les directions dans la zone d’étude. L’ensemble des spécimens des colonies est marqué à l’aide d’une étiquette colorée et numérotée pour pouvoir les reconnaître sur la zone d’étude. Les étendues florales de la zone d’étude sont ensuite toutes parcourues aléatoirement pour retrouver les bourdons. La vitesse et la direction du vent ainsi que les autres paramètres météorologiques sont enregistrés pour vérifier si les recaptures y sont corrélées.
Les ouvrières de cette espèce butinent sur des grandes étendues de florales proches du nid avec une grande fidélité. Vicia cracca L. est parmi les plantes les plus attractives pour cette espèce.
Les recaptures suivent en majorité l’axe principal du vent et sont faites sur deux grandes étendues de Vicia cracca L. disposées de part et d’autre des colonies.
La distance de butinage varie entre 1 et 331m. Elle ne semble pas être fonction du sens du vent, ni de sa vitesse.
Le sens du vent ne semble pas avoir une influence sur le choix du site de butinage des ouvrières et sa direction par rapport au nid.
Les ouvrières vont butiner sur les plantes attractives qui sont situé dans l’axe du vent. Les mêmes plantes attractives en dehors de cet axe ne sont que très peu visitées.
Mots-clés: Bombus terrestris (L.), distance de butinage, vent, Vicia cracca L.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 88
Remerciements
C’est par ces quelques lignes que j’aimerais exprimer ma reconnaissance envers toutes les personnes qui m’ont aidé, conseillé et soutenu tout au long de cette année.
Je pense tout particulièrement au Professeur Pierre Rasmont qui m’a accueilli dans son service et m’a permis de mener à bien ce mémoire.
J’adresse également mes sincères remerciements au Dr. P. Flamang pour la mise à ma disposition de son matériel et au Professeur P. Grosjean pour son aide pour les analyses statistiques.
Toute ma gratitude va également à M. Ir. Denis Michez pour ses conseils et son aide sur le terrain.
Je remercie le Dr. Michäel Terzo, Mlles Stéphanie Iserbyt et Olivia Ponchau pour leurs conseils et leurs aides, ainsi que leur patience.
J’aimerais encore exprimer ma gratitude envers toute l’équipe qui m’a aidé sur le terrain. Je pense au Dr. A. M. Aytekin et à M. M. Podrecca et A. Seché ainsi que Mlle K. Gras.
Je remercie M. A. Bousquet (Maire d’Eyne), M. Baracetti (Gestionnaire de la Réserve Naturelle d’Eyne) pour leur accueil chaleureux, et M. M. Baillot pour les possibilités de parcours de ses parcelles pour la réalisation de la présente étude.
Mes remerciements vont aussi à M. F. Delmarquette, P. Postiau et P. Bailly pour leurs aides techniques.
Je remercie également A. Secret et les étudiants de ma promotion ainsi que toutes les autres personnes qui m’ont aidé et soutenu tout au long de l’année.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 86
Table des matières
1. INTRODUCTION............................................................................................................................................. 1
1.1. GENERALITES............................................................................................................................................... 1
1.2. HISTOIRE NATURELLE .................................................................................................................................. 1 1.2.1. Cadre spécifique.................................................................................................................................. 1 1.2.2. Optimalisation du butinage (« Optimal foraging ») ............................................................................ 2
1.2.2.1. Exploitation des ressources............................................................................................................................ 3 1.2.2.2. Fidélité........................................................................................................................................................... 3 1.2.2.3. Distance de recaptures ................................................................................................................................... 4
1.2.3. Contraintes aérodynamiques............................................................................................................... 6 1.2.3.1. Vent ............................................................................................................................................................... 6 1.2.3.2. Température................................................................................................................................................... 7
2. OBJECTIFS ...................................................................................................................................................... 8
3. MATERIELS ET METHODES ...................................................................................................................... 9
3.1. PRINCIPE GENERALE..................................................................................................................................... 9
3.2. BOURDONS UTILISES .................................................................................................................................... 9
3.3. ZONE D’ETUDE ............................................................................................................................................. 9 3.3.1. Présentation générale........................................................................................................................ 11 3.3.2. Relevés phytosociologiques et occupation du sol .............................................................................. 11 3.3.3. Conditions météorologiques .............................................................................................................. 14
3.4. MATERIELS UTILISES.................................................................................................................................. 15 3.4.1. Cartographie ..................................................................................................................................... 15 3.4.2. Relevés météorologiques ................................................................................................................... 15
3.5. DETAILS DES METHODES ............................................................................................................................ 15 3.5.1. Marquage .......................................................................................................................................... 15 3.5.2. Installation des colonies .................................................................................................................... 16 3.5.3. Mesure de la structuration de l’espace de vol ................................................................................... 16 3.5.4. Mesures des conditions environnementales....................................................................................... 17 3.5.5. Cartographie de la zone d’étude ....................................................................................................... 17 3.5.6. Analyse palynologique....................................................................................................................... 18 3.5.7. Analyse des données .......................................................................................................................... 20
4. RESULTATS................................................................................................................................................... 22
4.1. MODELE NUMERIQUE DE TERRAIN.............................................................................................................. 22
4.2. CHOIX FLORAUX......................................................................................................................................... 24 4.2.1. Plantes butinées................................................................................................................................. 24 4.2.2. Analyse palynologique....................................................................................................................... 24
4.3. DISPERSION THEORIQUE ............................................................................................................................. 25
4.4. DISTANCE DE BUTINAGE............................................................................................................................. 26

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 87
4.5. FIDELITE AUX RESSOURCES ........................................................................................................................ 27
4.6. INFLUENCE DU VENT SUR LA DISTANCE DE BUTINAGE................................................................................ 31
4.7. INFLUENCE DU VENT SUR L’ORIENTATION DE BUTINAGE............................................................................ 33
4.8. STRUCTURATION DE L’ESPACE DE VOL....................................................................................................... 37
5. DISCUSSION .................................................................................................................................................. 40
5.1. CHOIX FLORAUX ET FIDELITE ..................................................................................................................... 40
5.2. DISTANCE DE BUTINAGE............................................................................................................................. 41
5.3. INFLUENCE DU VENT .................................................................................................................................. 42
5.4. STRUCTURATION DE L’ESPACE DE VOL....................................................................................................... 43
6. CONCLUSION ............................................................................................................................................... 44
7. PERSPECTIVES ............................................................................................................................................ 45
8. BIBLIOGRAPHIE.......................................................................................................................................... 46
9. ANNEXE I: LES PARCELLES .................................................................................................................... 49
10. ANNEXE II: LES RELEVES PHYTOSOCIOLOGIQUES DES PARCELLES.................................... 50
11. ANNEXE III: DONNEES DE RECAPTURES .......................................................................................... 73

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 1
1. Introduction
1.1. Généralités
Les bourdons (Hymenoptera, Apidae, Bombini) sont des insectes eusociaux qui possèdent une thermorégulation endotherme élaborée (Heinrich, 1979a). Cette particularité leur permet d’avoir une activité de butinage peu variable en fonction de la température au contraire des abeilles domestiques (Delbrassinne & Rasmont, 1988). Ils occupent de ce fait une place importante dans les milieux froids et tempérés, ce qui comprend les écosystèmes montagnards (Rasmont, 1988).
Ils sont pratiquement les seuls pollinisateurs d’un très grand nombre de plantes (Heinrich, 1979a; Rasmont, 1988) et notamment d’un grand nombre de plantes cultivées (Ruszkowsky, 1971; Heinrich, 1979a).
La structuration de l’espace de vol a été souvent étudiée par divers auteurs sur le terrain ou par le biais de modèles (Heinrich, 1979a; Pyke, 1984; Dukas & Edelstein-Keshet, 1998; Walter-Hellwig & Frankl, 1999; Durieux, 2000; Osborne et al., 2000; Goulson, 2003).
Cependant les influences de certains paramètres météorologiques sur la structuration de l’espace de vol des bourdons restent peu connues et en particulier les effets du vent. C’est pourtant un facteur qui influence fortement le vol, que ce soit celui des avions ou des animaux.
Dans l’optique de comprendre les effets du vent sur l’orientation et la distance de butinage des bourdons, nous avons étudié le comportement de 2 colonies de Bombus terrestris (L.) déposées sur un site exposé quotidiennement à des vents de directions différentes, le site du Pla del Bac à Eyne (France, Pyrénées-Orientales)
1.2. Histoire naturelle
1.2.1. Cadre spécifique
Bombus terrestris (L.) est une espèce très polylectique et ubiquiste (Ruszkowsky, 1971; Alford, 1975; Rasmont, 1988; Durieux, 2000; Gosselin, 2004). Cette espèce a pourtant une préférence pour les Fabaceae et les Asteraceae (Rasmont, 1988). Ruszkowski (1971) ajoute aussi les Lamiaceae, les Liliaceae, les Ranunculaceae, les Rosaceae et les Scrophulariaceae dans les préférences de cette espèce.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 2
Cette espèce est également capable de vol de nectar. Le bourdon perce un orifice à la base de la corolle pour accéder directement aux nectaires (Alford, 1975).
La vitesse de butinage de Bombus terrestris (L.) est de 32,2 fleurs de colza par minute et ne varie pas significativement entre 13 et 16°C (Delbrassinne & Rasmont, 1988). Celle d’Apis mellifera varie selon la température (13 – 16°C) et est de 10,6 à 14 fleurs par minute seulement (Delbrassinne & Rasmont, 1988).
1.2.2. Optimalisation du butinage (« Optimal foraging »)
Le butinage peut être influencé par les choix floraux, la dispersion des ressources, ainsi que les facteurs climatiques tels que la température et le vent. La contribution individuelle pour la génération suivante, l’aptitude relative à la survie et à la reproduction, dépend ainsi du comportement de butinage (Heinrich, 1979a). Selon Heinrich (1979a), les bourdons optimisent en permanence leur butinage de manière à maximiser leur aptitude. Cette théorie du butinage optimisé (« Optimal foraging ») est donc une extension de la théorie de l’évolution.
Pour Andersson (1978, cité par Pyke, 1984), l’aptitude relative à la reproduction augmente linéairement avec la nourriture récoltée durant un temps fixé, ou diminue avec le temps nécessaire pour récolter une quantité de nourriture fixée.
Dans le comportement de butinage le temps dépensé pour butiner est très important pour l’accumulation de profit (Heinrich, 1979a). Le profit peut être définit comme ceci:
Profit Temps
dépenséeénergieingéréetotalenourriture −=
Le bourdon peut augmenter le profit en visitant plus de fleurs pour un laps temps donné. Cela augmente la nourriture totale ingérée mais aussi l’énergie dépensée. Le profit augmente néanmoins, car l’énergie obtenue par la nourriture totale ingérée augmente plus rapidement que la dépense d’énergie qui est limitée physiologiquement (Heinrich, 1979a).
Le choix de l’espèce butinée est très important (Heinrich, 1979a; Pyke, 1984). Pour un même résultat, les butineuses se dirigent vers la fleur la plus proche (Pyke, 1984). Le temps de vol entre les inflorescences diminue lorsque la densité florale augmente (Ranta, 1983). Une forte densité florale permet donc de limiter la distance et le temps de vol ainsi que le coût énergétique (Heinrich, 1979a).

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 3
1.2.2.1. Exploitation des ressources
Les bourdons utilisent le pollen et le nectar comme nourriture pour les adultes et les larves (Alford, 1975). Le nectar est une source de sucre alors que le pollen est une source d’acides aminés, de sels minéraux et de vitamines (dont les stérols) (Alford, 1975). Au cours de l’évolution, certains groupes d’angiospermes ont été favorisés parce que les insectes les pollinisaient et recevaient en retour une récompense (Leppik, 1957, cité par Durieux, 2000).
Rasmont (1988) distingue quatre niveaux de spécialisation dans les choix floraux. Les espèces polylectiques qui peuvent butiner un grand nombre d’espèces différentes. Les espèces oligolectiques qui n’en butinent qu’un faible nombre. Les espèces monolectiques qui se nourrissent exclusivement d’une ou quelques espèces apparentées, même si d’autres plantes leurs sont accessibles. Les espèces mésolectiques qui butinent un grand nombre de plantes tout en exerçant un choix clairement discernable.
Les dimensions d’une aire de butinage dépendent de la densité des fleurs, de leur teneur en nectar et/ou en, et de la compétition spécifique (Heinrich, 1979a).
1.2.2.2. Fidélité
Pouvreau (2004) différencie la constance et la fidélité. La constance est un caractère inné et spécifique, qui lie l’insecte à un type de fleurs. La fidélité est un caractère temporel en rapport avec les avantages offerts par une espèce de fleur à un moment. L’apprentissage se fait sur un même type de fleurs pendant un certain temps (Pouvreau, 2004).
En terme de butinage optimisé, la fidélité à une espèce florale est très intéressante car cela permet d’augmenter la vitesse de butinage et donc la performance de la butineuse (Heinrich, 1976; Pyke, 1984).
Pour Laverty (1994), le butinage des fleurs simples est programmé chez les bourdons. Par contre, l’utilisation des fleurs à morphologie complexe demande un long apprentissage. La morphologie florale, la couleur, les parfums floraux sont des stimuli importants. L’ouvrière naïve se dirige vers la plante intéressante grâce à des stimuli visuels. Une ouvrière naïve qui sort du nid teste différentes espèces de plantes et se fidélise à celle qui lui fournit nectar et pollen de qualité supérieure et/ou en quantité supérieure. Ce sont les plantes « majeures » (Heinrich, 1979b). Les bourdons gardent cependant la capacité de basculer leur choix vers des plantes moins attrayantes en cas d’épuisement des ressources principales et de modification des conditions du milieu (Heinrich, 1979b).

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 4
Heinrich (1979b) s’est intéressé aux phénomènes d’apprentissage dans les choix floraux des bourdons. Pour lui, il n’existe pas de communication ni de trophallaxie. Il pense que l’apprentissage est individuel.
Par contre, Dornhaus & Chittka (1999, 2001) ont démontré que de retour au nid, les ouvrières de Bombus terrestris (L.) qui ont trouvé de la nourriture stimulent les autres ouvrières à aller butiner. Cette stimulation se fait par l’odeur de la source de nourriture qu’elles ont localisée. L’ouvrière de retour parcourt la surface du nid en manifestant une grande excitation, elle bouscule les autres ouvrières en faisant vibrer ses ailes. Les ouvrières stimulées quittent le nid et recherchent la source de l’odeur.
De plus, les expériences en laboratoire de Cameron (1981) et Pouvreau (1990) mettent en évidence une phéromone odorante que les bourdons déposent sur les fleurs qu’ils viennent de butiner. Celles-ci permettent aux autres ouvrières de distinguer les plantes intéressantes des autres plantes. Les bourdons n’utilisent leur mémoire visuelle que pour la reconnaissance à longue distance et ces phéromones les guideraient pour la reconnaissance à courte distance. Néanmoins, l’importance de ce phénomène dans le milieu naturel n’est pas connue. Goulson et al. (2000) ont montré que ces phéromones, qui proviennent des glandes tarsales, sont de même composition chez trois espèces différentes de bourdons. Ils ont également remarqué que ces phéromones à différentes dilutions ont un effet répulsif chez les butineuses.
1.2.2.3. Distance de recaptures
La plupart des études ont été menées sur la distance entre deux recaptures mais beaucoup moins entre le nid et la capture. Cette dernière a été étudiée chez les bourdons par Osborne et al. (1999) et Walther-Hellwig & Frankl (2000).
Walther-Hellwig & Frankl (2000) ont étudié notamment Bombus terrestris (L.) et les ont recapturés sur des sources de nourritures très abondantes à proximité du nid (fig.1).
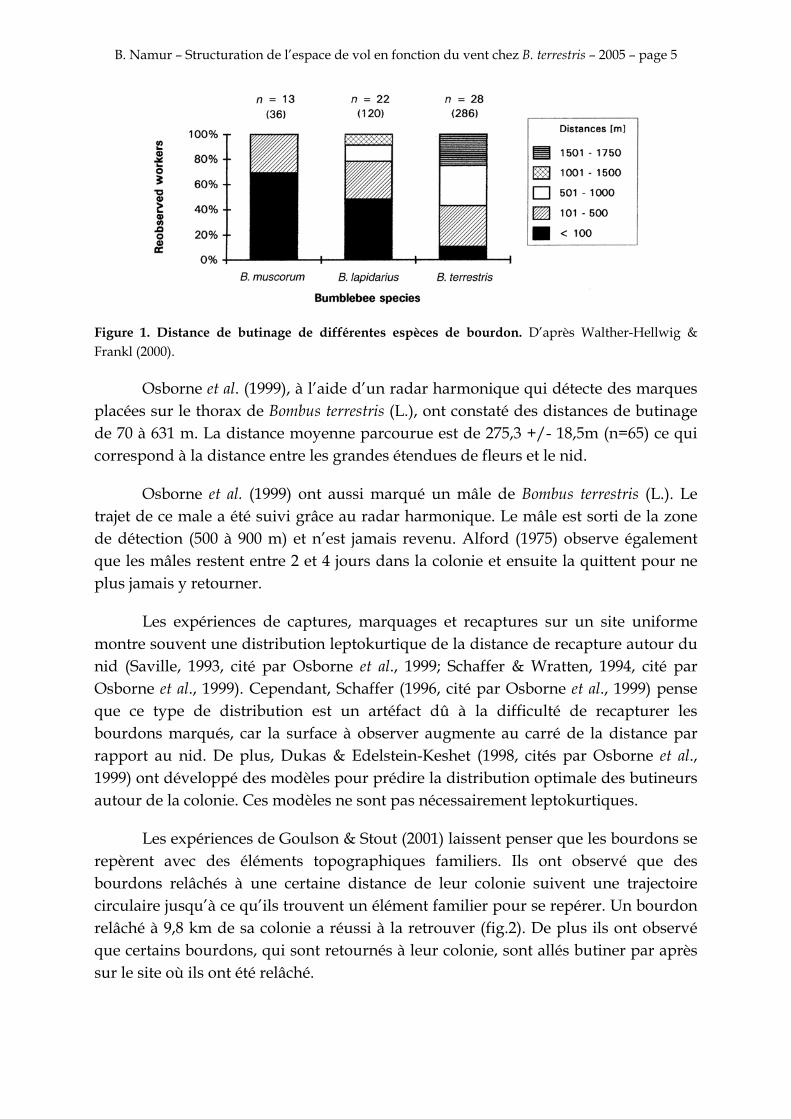
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 5
Figure 1. Distance de butinage de différentes espèces de bourdon. D’après Walther-Hellwig & Frankl (2000).
Osborne et al. (1999), à l’aide d’un radar harmonique qui détecte des marques placées sur le thorax de Bombus terrestris (L.), ont constaté des distances de butinage de 70 à 631 m. La distance moyenne parcourue est de 275,3 +/- 18,5m (n=65) ce qui correspond à la distance entre les grandes étendues de fleurs et le nid.
Osborne et al. (1999) ont aussi marqué un mâle de Bombus terrestris (L.). Le trajet de ce male a été suivi grâce au radar harmonique. Le mâle est sorti de la zone de détection (500 à 900 m) et n’est jamais revenu. Alford (1975) observe également que les mâles restent entre 2 et 4 jours dans la colonie et ensuite la quittent pour ne plus jamais y retourner.
Les expériences de captures, marquages et recaptures sur un site uniforme montre souvent une distribution leptokurtique de la distance de recapture autour du nid (Saville, 1993, cité par Osborne et al., 1999; Schaffer & Wratten, 1994, cité par Osborne et al., 1999). Cependant, Schaffer (1996, cité par Osborne et al., 1999) pense que ce type de distribution est un artéfact dû à la difficulté de recapturer les bourdons marqués, car la surface à observer augmente au carré de la distance par rapport au nid. De plus, Dukas & Edelstein-Keshet (1998, cités par Osborne et al., 1999) ont développé des modèles pour prédire la distribution optimale des butineurs autour de la colonie. Ces modèles ne sont pas nécessairement leptokurtiques.
Les expériences de Goulson & Stout (2001) laissent penser que les bourdons se repèrent avec des éléments topographiques familiers. Ils ont observé que des bourdons relâchés à une certaine distance de leur colonie suivent une trajectoire circulaire jusqu’à ce qu’ils trouvent un élément familier pour se repérer. Un bourdon relâché à 9,8 km de sa colonie a réussi à la retrouver (fig.2). De plus ils ont observé que certains bourdons, qui sont retournés à leur colonie, sont allés butiner par après sur le site où ils ont été relâché.
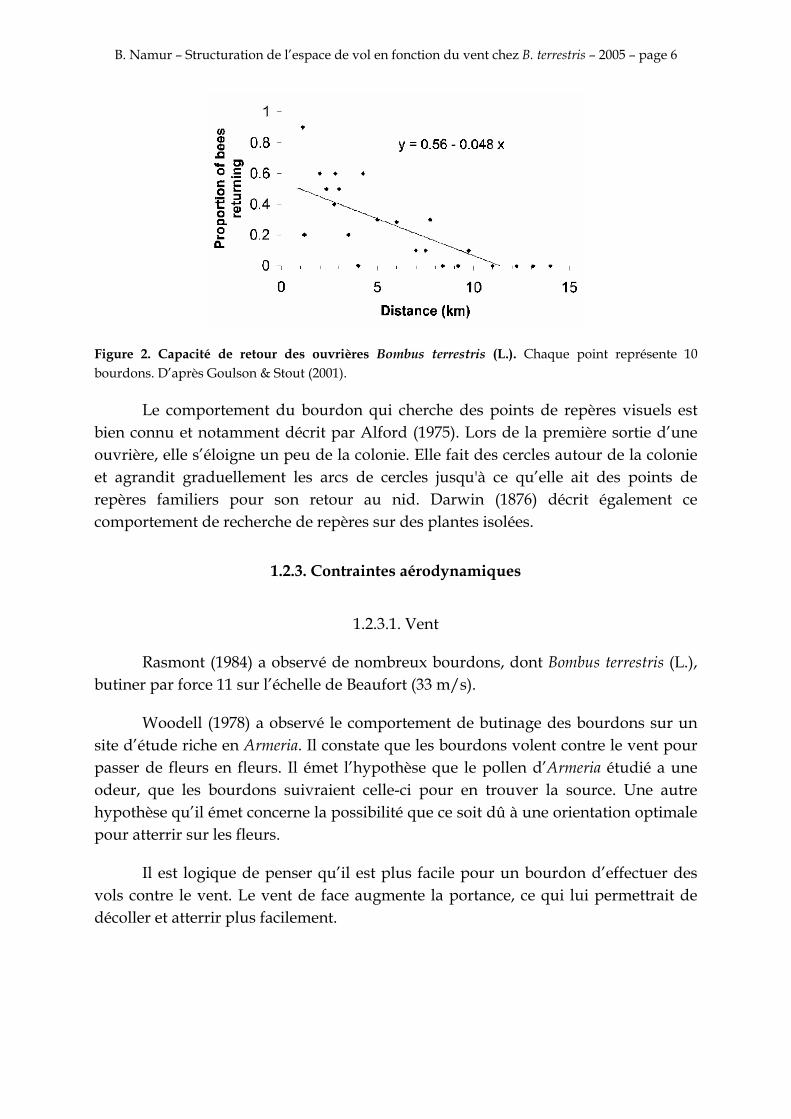
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 6
Figure 2. Capacité de retour des ouvrières Bombus terrestris (L.). Chaque point représente 10 bourdons. D’après Goulson & Stout (2001).
Le comportement du bourdon qui cherche des points de repères visuels est bien connu et notamment décrit par Alford (1975). Lors de la première sortie d’une ouvrière, elle s’éloigne un peu de la colonie. Elle fait des cercles autour de la colonie et agrandit graduellement les arcs de cercles jusqu'à ce qu’elle ait des points de repères familiers pour son retour au nid. Darwin (1876) décrit également ce comportement de recherche de repères sur des plantes isolées.
1.2.3. Contraintes aérodynamiques
1.2.3.1. Vent
Rasmont (1984) a observé de nombreux bourdons, dont Bombus terrestris (L.), butiner par force 11 sur l’échelle de Beaufort (33 m/s).
Woodell (1978) a observé le comportement de butinage des bourdons sur un site d’étude riche en Armeria. Il constate que les bourdons volent contre le vent pour passer de fleurs en fleurs. Il émet l’hypothèse que le pollen d’Armeria étudié a une odeur, que les bourdons suivraient celle-ci pour en trouver la source. Une autre hypothèse qu’il émet concerne la possibilité que ce soit dû à une orientation optimale pour atterrir sur les fleurs.
Il est logique de penser qu’il est plus facile pour un bourdon d’effectuer des vols contre le vent. Le vent de face augmente la portance, ce qui lui permettrait de décoller et atterrir plus facilement.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 7
1.2.3.2. Température
L’endothermie leur permet d’être actif dans une gamme large de température. Un bourdon en train de voler a été vu alors que la température était de -3,6°C (Bruggemann, 1958). La plupart des bourdons sont capable de butiner à 16°C (Heinrich, 1979a).
La vitesse de butinage n’est pas significativement différente chez les bourdons entre 13 et 16°C ce qui n’est pas le cas chez les abeilles domestiques (Delbrassinne & Rasmont, 1988). Ils commencent à butiner très tôt, 5h30 (heures solaires), et jusque 18-19h avec un ralentissement durant le heures les plus chaudes de la journée (Delbrassinne & Rasmont, 1988). Ce ralentissement durant la mi-journée est également décrit par Løken (1949, cité par Alford, 1975).
Les températures élevées peuvent provoquer la surchauffe des muscles thoraciques. À 35°C, les vols sont encore observés mais la température thoracique atteint 45°C, ce qui est proche de la température létale (Heinrich, 1979a).
D’après Bruggemann (1958), même une tempête de neige, le vent et la pluie n’empêchent pas les bourdons de voler.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 8
2. Objectifs
De nombreuses questions restent en suspens dans l’étude de la structuration de l’espace de vol des bourdons.
- Le vent a-t-il une influence sur la dispersion de butinage ?
- La direction et le sens du vent ont-ils une influence sur l’orientation de butinage ?
- Comment les bourdons tiennent-ils compte du vent par rapport à la disposition des ressources florales ?

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 9
3. Matériels et méthodes
3.1. Principe générale
Des colonies d’élevages sont disposées sur un site exposé à des vents de directions et de vitesses variables. La zone d’étude est un cercle de 500 m de rayon centré sur les nids. Les ressources florales sont dispersées et réparties en abondance dans toutes les directions dans la zone d’étude. L’ensemble des spécimens des colonies est marqué à l’aide d’une étiquette colorée et numérotée pour pouvoir les reconnaître sur la zone d’étude. Les étendues florales de la zone d’étude sont ensuite toutes parcourues aléatoirement pour retrouver les bourdons. La vitesse et la direction du vent ainsi que les autres paramètres météorologiques sont enregistrés pour vérifier si les recaptures y sont corrélées.
3.2. Bourdons utilisés
Deux colonies de stades différents ont été utilisées pour pallier aux problèmes les risques liés à l’acclimatation et à l’adaptation. Ce sont des colonies d’élevage fournies par la société Biobest (photo 1, planche I).
L’espèce étudiée dans la présente étude est Bombus terrestris (L.). Bombus terrestris (L.) est une espèce considérée comme très polylectique et ubiquiste (Ruszkowsky, 1971; Alford, 1975; Rasmont, 1988; Durieux, 2000; Gosselin, 2004).
3.3. Zone d’étude
Le zone d’étude se situe au lieu-dit le Pla del Bac, sur la commune d’Eyne (France, Pyrénées-Orientales) (fig.3 et 4).
Les colonies sont situées aux coordonnées géographiques 42°28’23,5’’N 02°04’25,3’’E (WGS84), 31TDH2395002935 (UTM) à 1615 m.
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 10
Figure 3. Département des Pyrénées-Orientales. En gris clair la Cerdagne et en gris foncé la commune d’Eyne, d’après S. Iserbyt (2000).
Figure 4. Photo aérienne d’Eyne. Mission FD 66 2000 1/25000 de l’IGN français.
500m
Eyne Eyne 2600
Pla del Bac
La Cabanasse
♦
Col de la Perche

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 11
3.3.1. Présentation générale
Eyne se trouve au sud du département des Pyrénées-Orientales, dans la région de la Cerdagne (fig.3 et photo 2, planche I). Il appartient au système hydrographique de l’Ebre. Il se situe au sud-est du plateau Cerdan, plateau de 900 à 1600 m d’altitude, entourés de hautes montagnes (fig.3).
Le Pla del Bac est recouvert de prés de fauche et de quelques surfaces emblavées (fig.4 et photo 3, planche I).
3.3.2. Relevés phytosociologiques et occupation du sol
Les prés de fauche d’Eyne présentent une flore très riche et diversifiée (Association Charles Flahaut, 1984). La zone a été divisée en parcelle caractérisée par une végétation différente (Annexe I). Cette année, le Pla del Bac a fait l’objet d’un inventaire floristique par D. Michez, M. Podrecca, P. Rasmont, A. Seché et moi même (tab.1) pour chacune des parcelles délimitées (Annexe II). Les plantes ont été déterminées avec différentes flores (Coste, 1927; Saule, 1991; Kerguélen, 1993; Grey-Wilson & Blamey, 1995).

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 12
Tableau 1. Inventaire des plantes observées dans la zone d’étude par D. Michez,
B. Namur, M. Podrecca, P. Rasmont et A. Seché
APIACEAE Anthriscus sylvestris (L.) Hoffmann Apiaceae sp. Astrantia cf. major L. Cf. Daucus sp. Eryngium bourgati Gouan. Peucedanum austriacum (Jacq.) Koch.
ASTERACEAE Achillea millefolium L. Arctium lappa L. Artemisia vulgaris L. Asteraceae indéterminée Carduus spp. Centaurea cyanus L. Centaurea jacea L. Centaurea scabiosa L. Cirsium ssp. Chamomilla recutita (L.) Rauschert Crepis spp. Hieracium pilosella L. Leucanthemum vulgare Lam. Onopordum sp. Senecio adonidifolius Loisel. Tanacetum vulgare L. Taraxacum spp. Tragopogon pratensis L.
BETULACEAE Alnus glutinosa (L.) Gaertn. Betula pendula Roth
BORAGINACEAE Echium vulgare L. Myosotis arvensis (L.) Hill
BRASSICACEAE Arabis glabra (L.) Bernh. Biscutella sp. Brassicaceae indéterminée Thlaspi arvense L.
CAMPANULACEAECampanula latifolia L. Campanula rotundifolia L. Campanula sp. Jasione montana L. Phyteuma orbiculare L.
CARYOPHYLLACEAE Agrostemma githago L. Cerastium brachypetalum Desportes ex
Pers. Dianthus carthusianorum L. Dianthus deltoides L. Dianthus hyssopifolius L.
Silene latifolia (P. Mill.) Britten & Rendle, non Poir.
Silene vulgaris (Moench) Garcke Stellaria graminea L.
CHENOPODIACEAEChenopodium album L.
CISTACEAEHelianthemum nummularium (L.) Miller
CONVOLVULACEAEConvolvulus arvensis L.
CRASSULACEAESedum acre L. Sedum monregalense Balbis Sedum rupestre L. Sedum sexangulare L. Sempervivum montanum L. Sempervirum arachnoideum L.
CUPRESSACEAEJuniperus communis nana Syme
CYPERACEAECarex sp.
DIPSACACEAEKnautia arvensis (L.) Coult.
ERICACEAECalluna vulgaris (L.) Hull
EUPHORBIACEAEEuphorbia cyparissias L.
FABACEAEChamaespartium sagittale (L.) P. Gibbs Genista anglica L. Genista tinctoria L. Lathyrus pratensis L. Lotus corniculatus L. Medicago sativa L. Onobrychis viciifolia Scop. Trifolium arvense L. Trifolium dubium Sibthorp Trifolium ochroleucum Hudson Trifolium pratense L. Trifolium repens L. Vicia cracca L. Vicia hirsuta (L.) S.F. Gray Vicia onobrychioides L. Vicia pannonica Crantz Vicia sp.
GENTIANACEAEGentiana lutea L. Gentiana sp.
GERANIACEAEErodium cicutarium (L.) L'Hér. ex Ait.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 13
Geranium pratense L. Geranium sp.
HYPERICACEAE Hypericum perforatum L.
JUNCACEAE Juncus sp.
LAMIACEAE Galeopsis tetrahit L. Prunella grandiflora (L.) Scholler Stachys officinalis (L.) Trev. Thymus cf. serpyllum L.
LILIACEAE Veratrum album L.
MALVACEAE Malva moschata L.
OLEACEAE Fraxinus excelsior L.
ORCHIDACEAE Dactylorhiza maculata (L.) Soo Gymnadenia conopsea (L.) R. Br. ex Ait. f. Orchis sp.
OROBANCHACEAEOrobanche gracilis Sm.
PINACEAE Pinus sylvestris L. Pinus uncinata Miller ex Mirbel
PLANTAGINACEAEPlantago lanceolata L. Plantago major L. Plantago media L.
PLUMBAGINACEAEArmeria maritima Willd.
POACEAE Alopecurus pratensis L. Avenula sp. Briza media L. Briza minor L. Dactylis glomerata L. Festuca cf. rubra L. Lolium sp. Phleum sp. Poa sp.
POLYGALACEAE Polygala alpestris Reich.
POLYGONACEAE Polygonum bistorta L. Rumex acetosa L. Rumex longifolius DC.
RANUNCULACEAE
Aconitum napellus L. Caltha palustris L. Ranunculus arvensis L. Ranunculus bulbosus L. Ranunculus montanus Willd. R. Ranunculus sp. Trollius europaeus L.
ROSACEAEAmelanchier obovalis (Michx.) Ashe Cotoneaster integerrimus Medik. Filipendula ulmaria (L.) Maxim. Malus sp. Prunus sp. Prunus spinosa L. Rosa canina L. Rosa pimpinellifolia L. Rosa sp. Rubus sp. Sanguisorba minor Scop. Sanguisorba officinalis L. Sorbus aucuparia L.
RUBIACEAEGalium palustre L. Galium pumilum Murray Galium verum L.
SALICACEAEPopulus tremula L. Salix spp. Salix caprea L.
SCROPHULARIACEAE Euphrasia Cf. nemorosa (Pers.) Wallr. Linaria repens (L.) Miller Pedicularis comosa L. Rhinanthus pumilus (Sterneck) Soldano Verbascum lychnitis L.
SOLANACEAESolanum tuberosum L.
THYMELEACEAEDaphne mezereum L.
URTICACEAEUrtica dioica L. Urtica pilulifera L.
VIOLACEAEViola arvensis Murray Viola sp. Viola tricolor L.
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 14
3.3.3. Conditions météorologiques
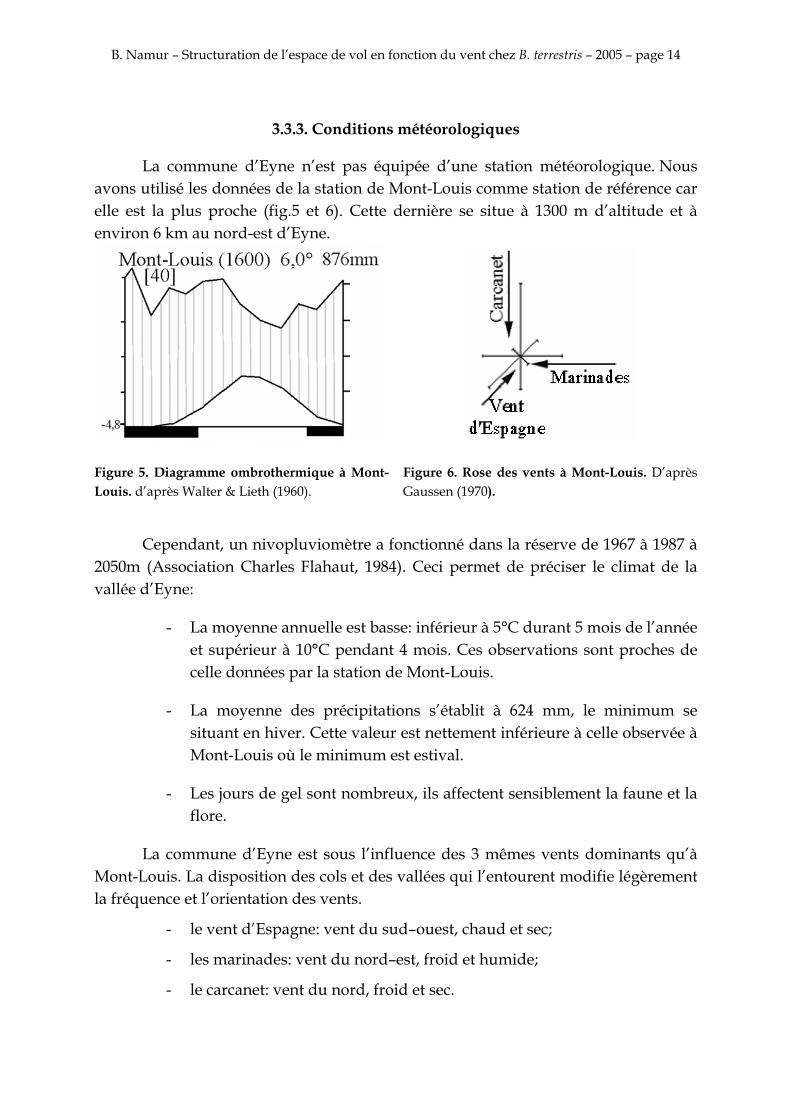
La commune d’Eyne n’est pas équipée d’une station météorologique. Nous avons utilisé les données de la station de Mont-Louis comme station de référence car elle est la plus proche (fig.5 et 6). Cette dernière se situe à 1300 m d’altitude et à environ 6 km au nord-est d’Eyne.
Figure 5. Diagramme ombrothermique à Mont-Louis. d’après Walter & Lieth (1960).
Figure 6. Rose des vents à Mont-Louis. D’après Gaussen (1970).
Cependant, un nivopluviomètre a fonctionné dans la réserve de 1967 à 1987 à 2050m (Association Charles Flahaut, 1984). Ceci permet de préciser le climat de la vallée d’Eyne:
- La moyenne annuelle est basse: inférieur à 5°C durant 5 mois de l’année et supérieur à 10°C pendant 4 mois. Ces observations sont proches de celle données par la station de Mont-Louis.
- La moyenne des précipitations s’établit à 624 mm, le minimum se situant en hiver. Cette valeur est nettement inférieure à celle observée à Mont-Louis où le minimum est estival.
- Les jours de gel sont nombreux, ils affectent sensiblement la faune et la flore.
La commune d’Eyne est sous l’influence des 3 mêmes vents dominants qu’à Mont-Louis. La disposition des cols et des vallées qui l’entourent modifie légèrement la fréquence et l’orientation des vents.
- le vent d’Espagne: vent du sud–ouest, chaud et sec;
- les marinades: vent du nord–est, froid et humide;
- le carcanet: vent du nord, froid et sec.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 15
Le carcanet y est beaucoup moins fréquent qu’à Mont-Louis à cause du Col de la Perche situé au nord-est du Pla del Bac.
3.4. Matériels utilisés
3.4.1. Cartographie
Les GPS utilisés sont des Magellan Sport Track Pro. Dans les conditions d’emploi la précision de cet instrument est de 1 à 10 m.
Les points enregistrés et le trajet effectué sont téléchargés chaque soir à partir du GPS vers un ordinateur portable grâce au logiciel MapSend Street Europe version 4.10d.
L’ensemble des points récoltés durant l’étude est utilisé pour établir un modèle numérique de terrain, une carte topographique détaillée de la zone d’étude. Le logiciel MapInfo Professional 5.5 permet de faire cette carte ainsi qu’une représentation topographique en trois dimensions de la zone d’étude (MapInfo Professionnel version 5.5, 1999).
3.4.2. Relevés météorologiques
Le vent a été mesuré avec un anémomètre de la marque Kaindl Windmaster 2. Ce modèle permet de mesurer la vitesse du vent en m/s et en Beaufort quelque soit la direction du vent. L’affichage numérique donne la vitesse instantanée, la vitesse maximale et la vitesse moyenne du vent en m/s sur le temps où l’appareil est allumé. La vitesse en Beaufort est également indiquée. La précision est de +/- 4%.
La température et l’hygrométrie absolue ont été mesurées avec un psychromètre de la marque Lufft C210. L’affichage numérique donne la température et l’hygrométrie instantanées en gramme d’eau par mètre cube ou l’hygrométrie relative en %. La précision est de +/- 0,3°C et de +/- 2% pour l’humidité relative.
3.5. Détails des méthodes
3.5.1. Marquage
L’ensemble des spécimens des deux colonies est marqué avec des étiquettes colorées et numérotées. Cela permet de distinguer sur le terrain chaque spécimen et sa colonie d’origine. Lors du marquage, on différencie les ouvrières des mâles.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 16
Pour effectuer ce marquage, on isole un spécimen dans un tube de contention (photo 4, planche I). Un piston permet d’immobiliser le spécimen. L’étiquette colorée et numérotée est fixée sur le thorax à l’aide de colle cyanoacrylique (photo 5, planche I).
Après le marquage, les spécimens sont réintroduits dans leur colonie.
Les bourdons qui viennent d’émerger sont capturés à la sortie des colonies et marqués sur le terrain.
3.5.2. Installation des colonies
Les colonies sont placées dans des ruches en bois. Pour améliorer l’isolation thermique, des plaques de polystyrène extrudé et de la paille sont disposées entre les planches de la ruche et les boites contenant les colonies. L’entrée de la ruche est raccordée à la sortie via un tuyau. Les colonies sont ensuite placées au centre de la zone d’étude à l’abri d’un arbre.
3.5.3. Mesure de la structuration de l’espace de vol
Un cercle de 500 m de rayon autour des colonies a été choisi comme zone d’étude. La dimension de la zone d’étude a été choisie en fonction de la littérature sur les distances de recaptures (Osborne et al., 1999; Walther-Hellwig & Frankl, 2000) et des possibilités d’accès pour les observateurs. Dans ce rayon de 500 m, on a parcouru toutes les étendues florales de manière aléatoire afin de retrouver les spécimens marqués. Lorsqu’un bourdon marqué est observé, on note:
- la date et l’heure;
- la couleur et le numéro de l’étiquette du bourdon;
- la localisation précise de l’observation à l’aide du GPS;
- la plante butinée.
Les colonies sont installées au centre de la zone d’étude le 9 juillet 2004.
La période d’observation va du 11 au 27 juillet 2004, avec un jour d’interruption pour des raisons logistiques le 17 (tab.2). Au total, les observations ont duré 16 jours. Les observations ont commencé deux jours après l’installation des colonies pour laissez le temps aux bourdons de s’acclimater. Le 27 juillet au soir, les colonies sont retirées et sacrifiées par congélation.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 17
Tableau 2. Calendrier des prospections. Sur fond noir les jours sans observations et sur fond blanc les jours d’observation
Lundi Mardi Mercredi Jeudi Vendredi Samedi Dimanche
9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
3.5.4. Mesures des conditions environnementales
Les paramètres météorologiques ont été mesurés à l’aide d’un anémomètre et d’un psychromètre.
Les relevés météorologiques sont faits toutes les demi-heures pendant une minute à l’emplacement des colonies à 2 m du sol.
Lors des relevés météorologiques, nous avons noté:
- la date et l’heure;
- la direction du vent;
- la vitesse moyenne du vent en m/s;
- la vitesse maximale du vent en m/s;
- la vitesse moyenne du vent en Beaufort;
- la température;
- l’hygrométrie absolue en g/m³;
- une estimation de la couverture nuageuse en %.
Par la suite chaque recapture est associée à un relevé météorologique qui lui correspond.
3.5.5. Cartographie de la zone d’étude
En plus d’une cartographie classique de la zone d’étude, on a réalisé un modèle numérique de terrain. Le modèle numérique de terrain est une représentation simple d’une surface continue dans l’espace à partir de points irrégulièrement espacés mais représentatifs du relief (Wolf, 2001). Il repose sur le principe de l’interpolation spatiale et sur le fait que deux observations proches dans l’espace se ressemblent plus que deux observations éloignées (auto-corrélation, Burrough & McDonnell, 2000).

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 18
Le modèle numérique de terrain global se base sur la connaissance de X stations à l’échelle de la zone d’étude. L’interpolation s’effectue sur l’ensemble de cette zone.
La technique utilisée est celle de la moyenne mobile. L’altitude d’un pixel est calculée (centroïde) sur la base de l’altitude moyenne d’une fenêtre d’au moins 30 points d’altitude connue (DeMers, 1997).
303021 ptduAltitudeptduAltitudeptduAltitudeXptduAltitude +++
=Κ
La fenêtre de 30 points se déplace et permet le calcul de l’altitude du pixel suivant. Cette technique a un inconvénient majeur: elle laisse subsister des flous sur les bords de la zone d’étude. Pour cette raison, on a étendu la zone topographiée à un rayon de 750 m autour des colonies, alors que les observations se limitent à un rayon de 500 m.
Le modèle numérique de terrain est réalisé à l’aide du logiciel MapInfo Professional 5.5 (MapInfo Professionnel version 5.5, 1999).
3.5.6. Analyse palynologique
Pour vérifier que les ouvrières n’allaient pas sur d’autres plantes que celles obtenues par les recaptures, une analyse des pollens présents dans les colonies est effectuée a posteriori.
A la fin de l’étude, les colonies sont sacrifiées par congélation et conservées au congélateur à -30°C jusqu’à leur dissection. Le couvain est dégagé en retirant la canopée construite par les ouvrières et l’ouate isolante. Une photo du couvain complètement dégagé est prise. Cette photo permet de numéroter toutes les cellules pour les identifier en vue de l’analyse palynologique.
On étudie séparément les pollens contenus dans la cire de 33 cellules d’une des colonies ainsi que les pollens mélangés au nectar retrouvé dans cette colonie. Le choix des 33 cellules est obtenu par tirage de nombre aléatoire.
Les pollens sont ensuite comparés à 18 échantillons de pollens témoins prélevés sur les plantes majoritairement présentes dans la zone d’étude. Ces échantillons de pollens ont été prélevés sur: Achillea millefolium L., Anthriscus sylvestris (L.) Hoffmann, Centaurea jacea L., Centaurea scabiosa L., Echium vulgare L., Geranium pratense L., Helianthemum nummularium (L.) Miller, Hypericum perforatum L., Knautia arvensis (L.) Coult., Lathyrus pratensis L., Malva moschata L., Medicago sativa L.,

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 19
Rhinanthus pumilus (Sterneck) Soldano, Rosa canina L., Senecio adonidifolius Loisel., Trifolium pratense L., Trifolium repens L. et Vicia cracca L.
L’analyse palynologique requiert un traitement chimique acétolytique (9 parts d’anhydride acétique pour 1 part d’acide sulfurique) qui permet d’enlever tout l’exine qui entoure les grains de pollens (Delcourt et al., 1959; Erdtman, 1960; Wodehouse, 1965).
En ce qui concerne les pelotes de pollen et les échantillons frais, ils sont mis pendant 30 minutes au minimum dans un tube Ependorf rempli d’acide acétique glacial. Une centrifugation à 2400 tours par minute est réalisée pendant 5 minutes pour enlever ensuite le surnageant. Au culot qui contient les pollens, on ajoute la solution acétolytique et on secoue pour le dissoudre. La solution acétolytique est une solution de 9 volumes d’anhydride acétique pur dans laquelle on verse goutte à goutte 1 volume d’acide sulfurique concentré. Juste après l’addition de la mixture acétolytique, le tube Ependorf est placé dans un bain-marie. On monte la température jusqu’au point d’ébullition. Le tube doit être remué occasionnellement. Quand le point d’ébullition est atteint, on arrête de chauffer et on laisse les tubes refroidir dans le bain pendant 15 minutes. Grâce à une seconde centrifugation de 5 minutes à 2400 tours par minute, la mixture acétolytique est éliminée. Le culot est nettoyé avec une solution d’eau et de alcool 95% (3: 1) dans laquelle il est dissout. Une dernière centrifugation de 5 minutes à 2400 tours par minute permet d’éliminer le surnageant. Le culot est séché pendant une nuit au minimum.
Le montage sur lame microscopique se fait avec de la gélatine glycérinée. Pour 30 cm³ de cette gélatine, il faut ramollir 7 g de gélatine dans 25 ml d’eau distillée pendant 2 heures. Ensuite, on ajoute 20 ml de glycérine pure et, pour éviter la moisissure, 1 ml de phénol. On chauffe légèrement en agitant pendant 1 minute. Après on verse dans une boite de pétri et on laisse refroidir.
Pour le montage sur lame microscopique, on prélève 1 mm³ de gélatine glycérinée à l’aide d’une pince et on la frotte au culot séché pour ramasser les grains de pollen. On dépose le petit morceau de gélatine glycérinée au milieu de la lame microscopique. On chauffe doucement la lame microscopique jusqu'à la fusion de la gélatine et la formation d’une goutte de 5 mm de diamètre maximum. Ensuite tant que la gélatine est liquide, on la couvre avec une lamelle couvre objet. On coule de la paraffine sur le bord de la lamelle pour pouvoir la conserver. La lamelle est tournée vers le bas le temps qu’elle sèche.
Pour les pollens contenus dans la cire des pains des bourdons, on prélève un fragment de cire et on la met dans un Ependorf. On rempli le tube Ependorf avec de l’acétone. On fragmente la cire et on agite fortement, cela permet de dissoudre la cire et libérer les grains de pollen. Une centrifugation pendant 5 minutes à 2400 tours par

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 20
minute permet d’éliminer le surnageant. Le culot est ensuite traité comme une pelote de pollen ou un échantillon frais.
Pour les pollens mélangés au nectar, on prélève le nectar avec une pipette pasteur, en le diluant légèrement avec de l’eau si nécessaire. On met le nectar dans un tube Ependorf et on rajoute de l’eau. On homogénéise la solution avant de la centrifuger pendant 5 minutes à 2400 tours par minute. On élimine le surnageant et on traite le culot comme une pelote de pollen ou un échantillon frais.
3.5.7. Analyse des données
Pour cette étude, deux méthodes de prospection étaient possibles: la prospection en transects et la prospection aléatoire de toutes les étendues florales.
La prospection selon un quadrillage centré sur les colonies couvre très peu de surface par rapport à la surface total de la zone d’étude. Le nombre de recaptures est donc toujours très faible et peu représentatif de ce qui se passe sur l’ensemble de la zone d’étude. Ce manque de représentativité est causé par les pics de distribution qui se resserrent aux niveaux des grandes étendues florales. L’avantage est que l’on peut plus facilement utiliser des tests statistiques. Cependant le très faible nombre de recaptures et le manque de représentativité de ce qui se passe sur l’ensemble de la zone d’étude diminue fortement la robustesse de ces tests.
La prospection aléatoire de toutes étendues florales est la méthode utilisée dans cette étude. Cette technique permet de maximiser le nombre de recaptures et de couvrir une surface hautement représentative de l’ensemble de la zone d’étude. Le problème est que les analyses statistiques sont rendues très difficiles ou nécessitent une simplification à l’extrême. L’analyse de ce très grand nombre de recaptures, qui est très représentatif de ce qui se passe sur l’ensemble de la zone d’étude, peut se faire de manière descriptive.
Un test de χ2 qui compare le nombre de recaptures à un nombre de recaptures théoriques permet de dire si les bourdons se distribuent de façon homogène sur les parcelles recouvertes de plantes attractives. Les parcelles considérées comme attractives sont celles qui ont un coefficient d’abondance, reprit en annexe 2, de minimum 2 pour l’une de ces plantes considérées comme attractives par la littérature (Ruszkowsky, 1971; Alford, 1975; Rasmont, 1988; Durieux, 2000; Gosselin, 2004) et les résultats des choix floraux de la présente étude. Pour calculer le nombre de recaptures théoriques d’une parcelle, le nombre de recaptures totales sur les parcelles attractives est multiplié par la fraction de la surface de la parcelle sur la surface totale des parcelles attractives. L’hypothèse H0 est que les valeurs observées sont égales aux

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 21
valeurs théoriques. L’hypothèse H0 est rejetée si χ2 observé est supérieur au χ2 critique au seuil α.
L’analyse de l’effet de la direction du vent sur le site de butinage des bourdons est une analyse essentiellement descriptive. L’interprétation des données passe par les pointages des observations sur la représentation topographique du modèle numérique de terrain, par des graphiques simples et par des éthogrammes.
Trois catégories de vent sont différenciées: le vent considéré comme nul (≤ 2 Beaufort = 11,9 m/s), le vent du sud-ouest (> 2 Beaufort) et le vent du nord-est (> 2 Beaufort).
Pour étudier l’effet du vent sur la distance de butinage, un test de Kolmogorov-Smirnov est utilisé. Les classes de distance de butinage sont de 10 m. Le test de Kolmogorov-Smirnov permet de mettre en évidence si les nombres de recaptures dans les différentes classes de distances sont significativement différents pour les trois catégories de vent. Le test de Kolmogorov-Smirnov ne fait pas d’hypothèse sur la distribution des données. C’est un test cumulatif non-paramétrique. L’hypothèse H0 est rejetée si l’écart maximum, Dmax, entre les données cumulées (fréquences relatives) et la distribution normale cumulée dépasse une valeur critique, définie en fonction du degré de signification choisi.
L’estimation de la fidélité se fait à l’aide de la technique des flow-diagrams. Sur les schémas, la taille des cercles est proportionnelle au nombre d’observations réalisées sur la parcelle qu’ils représentent. Les cercles sont disposés de manière analogue à la zone d’étude. Les flèches sont proportionnelles à la probabilité qu’a une ouvrière qui butine dans cette parcelle d’être recapturée sur dans une autres parcelle désignée par la flèche lors de la recapture suivante. Les données utilisées pour cette représentation ne concernent que les bourdons recapturés au moins deux fois.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 22
4. Résultats
Le tableau 3 résume les observations de marquages – recaptures. L’Annexe III figure l’ensemble des recaptures associées aux relevés météorologiques.
Tableau 3. Résultats des marquages – recaptures
Nombre d’ouvrières marquées 262 Nombre de mâles marqués 15 Nombre de recaptures d’ouvrières 357 Nombre de recaptures de mâles 0 Nombre de relevés météorologiques 228 Distance de recaptures 1 à 330 m Nombre de points GPS utilisés pour le modèle topographique 18.501
4.1. Modèle numérique de terrain
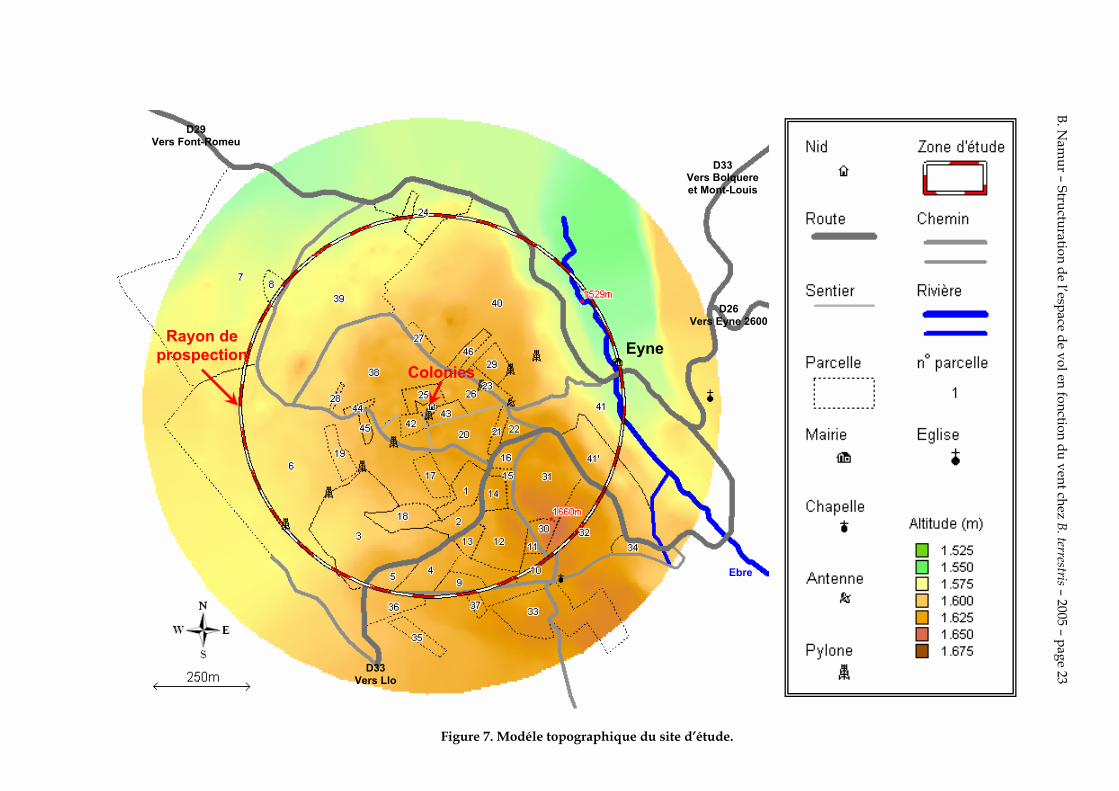
La zone d’étude d’un rayon de 500 m est subdivisée en 47 parcelles (Annexe I) caractérisées par un relevé phytosociologique en Annexe II. L’altitude de la zone d’étude varie entre 1529 et 1660 m, soit un dénivelé de 131 m
La figure 7 est une représentation topographique de la zone d’étude.
La grande partie de la zone d’étude est recouverte de prés de fauche avec de petites emblavures de seigle et de pomme de terre. Le nord et le nord-est de la zone d’étude sont légèrement boisés avec des Pinus uncinata Miller ex Mirbel, Pinus sylvestris L., Populus tremula L. et des buissons de Juniperus communis nana Syme
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 23
D33 Vers Bolquere et Mont-Louis
D26 Vers Eyne 2600
Ebre
Eyne Colonies
Rayon de prospection
D29 Vers Font-Romeu
D33 Vers Llo
Figure 7. Modéle topographique du site d’étude.
B. Nam
ur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 23

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 24
4.2. Choix floraux
4.2.1. Plantes butinées
La figure 8 montrent que l’essentiel des recaptures se sont faites sur Vicia cracca L. (photo 6, planche I)
82
5
3
3
2
1
1
1
1
1
1
0
0
0
0 10 20 30 40 50 60 70 80 90 100
Vicia cracca
Knautia arvensis
Echium vulgare
Rhinanthus pumilus
Anthriscus sylvestris
Medicago sativa
Trifolium pratense
Helianthemum nummularium
Rosa canina
Rubus sp.
Trifolium repens
Achillea millefolium
Lathyrus pratensis
Asteraceae indéterminée
Pourcentage de recapture (n = 336)
Figure 8. Choix floraux de Bombus terrestris (L.) dans la zone d’étude.
4.2.2. Analyse palynologique
La figure 9 concerne l’analyse palynologique de la somme des pollens retrouvés dans la cire et dans le nectar dans une des colonies. La différenciation des différents types de pollen montre que le pollen le plus abondant est le pollen utilisé comme ressource alimentaire par la société Biobest.
Le pollen d’élevage est constitué de nombreuses espèces florales de différentes familles. Les principaux pollens utilisés proviennent d’Apiaceae; d’Asteraceae du genre Bellis, Helianthus, Centaurea, cf. Chrysanthemum, Taraxacum; de Boraginaceae du genre Echium; de Brassicaceae du genre Brassica; de Campanulaceae du genre Campanula; de Chenopodiaceae; de Cistaceae; d’Ericaceae du genre Rhododendron, Calluna; de Fabaceae du genre Trifolium; de Lamiaceae; d’Oleaceae; de Plantaginaceae du genre Plantago; de Poaceae; de Ranunculaceae; de Rhamnaceae; de Rosaceae du genre Rubus et de Tiliaceae du genre Tilia.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 25
46
33
7
6
5
2
1
1
0
0
0
0
0
0
0
0
0
0 5 10 15 20 25 30 35 40 45 50
Pollens Biobest (PB)
Pollens indéterminables
Lathyrus pratensis + Vicia cracca
Helianthemum nummularium + PB
Echium vulgare + PB
Hypericum perforatum + Medicago sativa + Trifolium repens + PB
Rhinanthus pumilus
Achillea millefolium
Rosa canina
Centaurea jacea
Trifolium pratense
Centaurea scabiosa
Anthriscus sylvestris
Geranium pratense
Knautia arvensis
Malva moschata
Senecio adonidifolius
Pourcentage de pollens (n = 4285)
Figure 9. Analyse palynologique. Le noir représente les pollens d’espèces qui sont surestimées car le pollen des plantes de la zone d’étude se confondent avec celui utilisé pour l’élevage par Biobest.
Les pollens indéterminables sont les pollens qui sont déformés, détruits suite au traitement acétolytique ou masqués par les résidus de cire.
Certain pollens de plantes présentent sur la zone d’étude se confondent avec le pollen utilisé pour l’élevage. Les pollens qui sont confondus sont ceux d’Hypericum perforatum L., de Trifolium repens L., de Medicago sativa L., d’Echium vulgare L. et d’Helianthemum nummularium (L.) Miller qui sont donc surestimés. Les pollens les plus abondants et qui proviennent des plantes présentent dans la zone d’étude sont ceux de Vicia cracca L. et de Lathyrus pratensis L. puisqu’ils se confondent.
4.3. Dispersion théorique
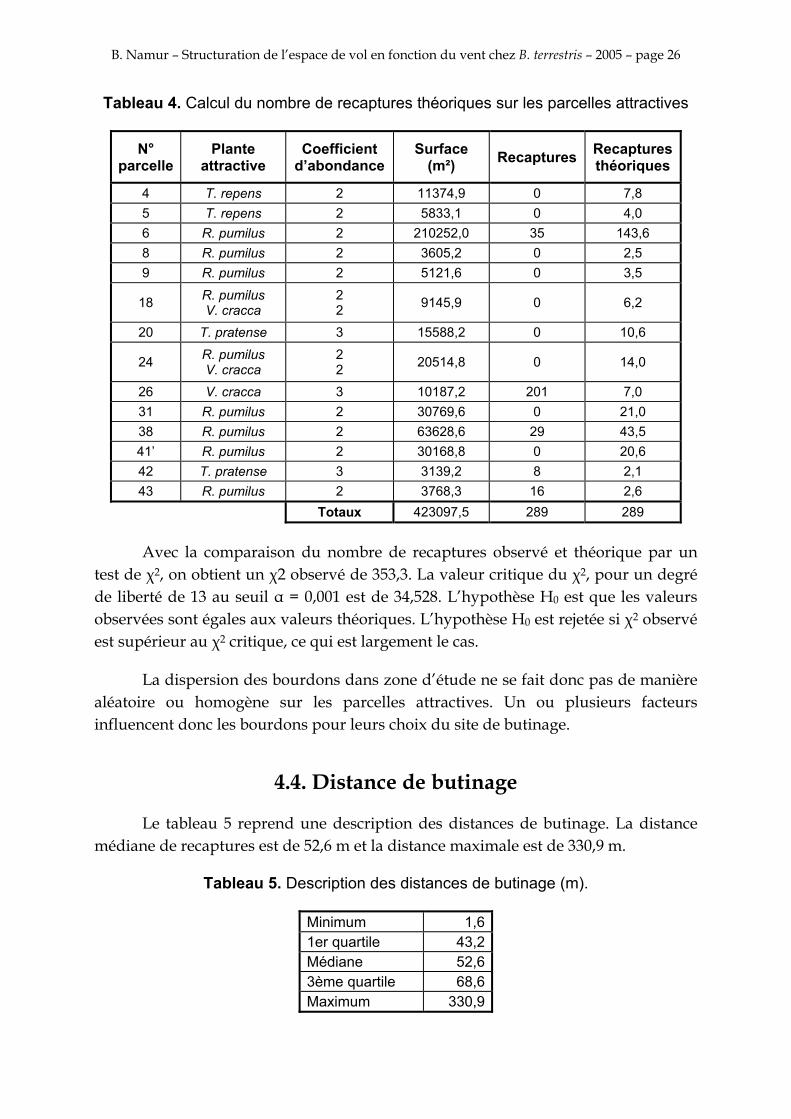
Les parcelles considérées comme attractives sont celles qui ont un coefficient d’abondance, reprit en Annexe II, de minimum 2 pour l’une de ces plantes considérées comme attractives: Vivia cracca L., Knautia arvensis (L.) Coult., Echium vulgare L., Rhinanthus pumilus (Sterneck) Soldano, Medicago sativa L., Trifolium pratense L., Trifolium repens L. et Lathyrus pratensis L.. Le nombre de recaptures théoriques sur les parcelles attractives est calculé dans le tableau 4.
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 26
Tableau 4. Calcul du nombre de recaptures théoriques sur les parcelles attractives
N° parcelle
Plante attractive
Coefficient d’abondance
Surface (m²) Recaptures Recaptures
théoriques
4 T. repens 2 11374,9 0 7,8 5 T. repens 2 5833,1 0 4,0 6 R. pumilus 2 210252,0 35 143,6 8 R. pumilus 2 3605,2 0 2,5 9 R. pumilus 2 5121,6 0 3,5
18 R. pumilus V. cracca
2 2 9145,9 0 6,2
20 T. pratense 3 15588,2 0 10,6
24 R. pumilus V. cracca
2 2 20514,8 0 14,0
26 V. cracca 3 10187,2 201 7,0 31 R. pumilus 2 30769,6 0 21,0 38 R. pumilus 2 63628,6 29 43,5 41’ R. pumilus 2 30168,8 0 20,6 42 T. pratense 3 3139,2 8 2,1 43 R. pumilus 2 3768,3 16 2,6
Totaux 423097,5 289 289
Avec la comparaison du nombre de recaptures observé et théorique par un test de χ2, on obtient un χ2 observé de 353,3. La valeur critique du χ2, pour un degré de liberté de 13 au seuil α = 0,001 est de 34,528. L’hypothèse H0 est que les valeurs observées sont égales aux valeurs théoriques. L’hypothèse H0 est rejetée si χ2 observé est supérieur au χ2 critique, ce qui est largement le cas.
La dispersion des bourdons dans zone d’étude ne se fait donc pas de manière aléatoire ou homogène sur les parcelles attractives. Un ou plusieurs facteurs influencent donc les bourdons pour leurs choix du site de butinage.
4.4. Distance de butinage
Le tableau 5 reprend une description des distances de butinage. La distance médiane de recaptures est de 52,6 m et la distance maximale est de 330,9 m.
Tableau 5. Description des distances de butinage (m).
Minimum 1,61er quartile 43,2Médiane 52,63ème quartile 68,6Maximum 330,9

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 27
La figure 10 montre clairement que la plupart des recaptures se sont faites entre 40 et 70m. Il y a aussi deux plus petits pics un entre 130 et 150m et l’autre entre 210 et 230m.
0
5
10
15
20
25
0 10 20 30 40 50 60 70 80 90 100
110
120
130
140
150
160
170
180
190
200
210
220
230
240
250
260
270
280
290
300
310
320
330
340
Distance de butinage (m)
Pour
cent
age
de re
capt
ure
(n=3
36)
Figure 10. Distance de butinage des ouvrières de Bombus terrestris (L.) par rapport à l’emplacement des colonies.
Aucun mâle sur les 15 marqués n’a été recapturé.
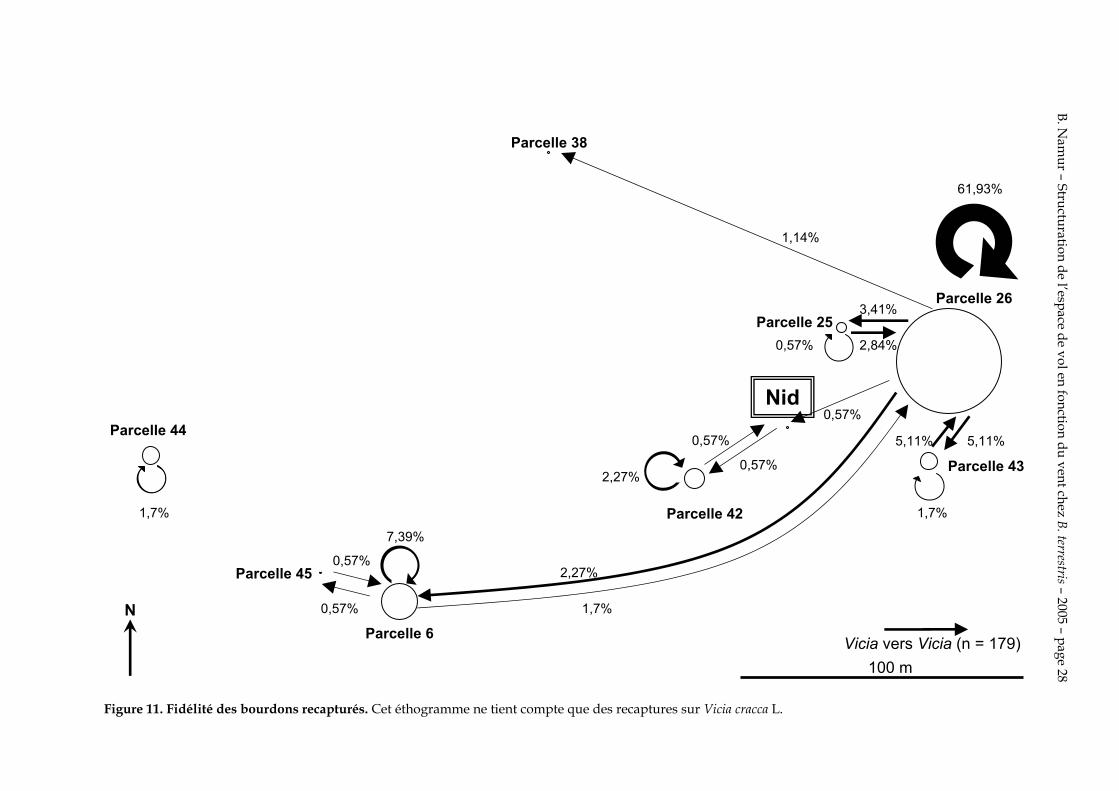
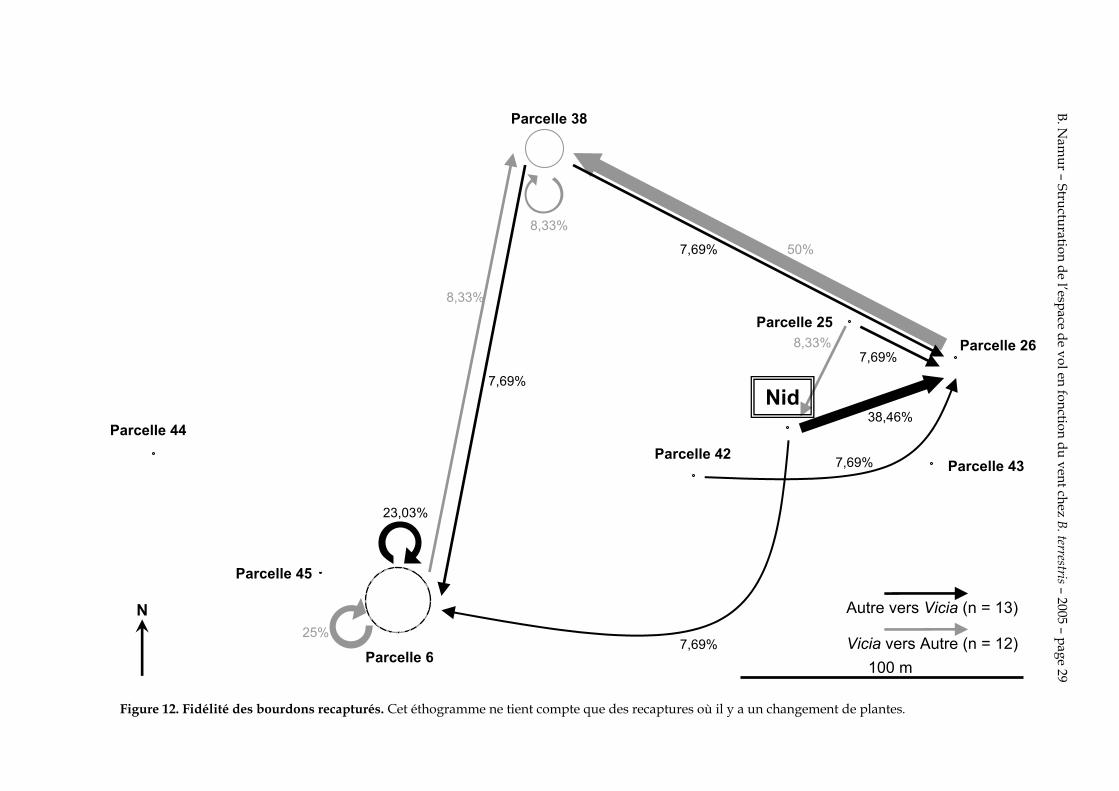
4.5. Fidélité aux ressources
Les figures 11, 12 et 13 expriment la fidélité des bourdons recapturés sur chaque parcelle.
La figure 11 montre l’attraction énorme des Vicia cracca L. de la parcelle 26 et de la parcelle 6. Elle montre également que peu de bourdons passent d’une parcelle à une autre.
La figure 12 montre que les bourdons qui changent de plantes ont tendance également à changer de site de butinage.
La figure 13 montre, comme la figure 11, que peu de bourdon change de site de butinage. Les bourdons recapturés sur une autre plante que Vicia cracca (L.) restent assez fidèles à cette autre plante.
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 28
Parcelle 38
Parcelle 26 Parcelle 25
Parcelle 44
Parcelle 45
Parcelle 43
Parcelle 42
Parcelle 6
7,39%
0,57%
61,93%
2,27%
1,7% 1,7%
2,27%0,57%
3,41%
1,7%
5,11%
1,14%
0,57%
0,57%
0,57%
0,57%
2,84%
5,11%
100 m Vicia vers Vicia (n = 179)
Figure 11. Fidélité des bourdons recapturés. Cet éthogramme ne tient compte que des recaptures sur Vicia cracca L.
Nid
N
B. Nam
ur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 28
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 29
100 m
Parcelle 38
Parcelle 26 Parcelle 25
Parcelle 44
Parcelle 45
Parcelle 43 Parcelle 42
Parcelle 6
23,03%
38,46%
8,33%
Autre vers Vicia (n = 13)
8,33%
8,33%
25%
50%7,69%
7,69%
7,69%
7,69%
7,69%
Vicia vers Autre (n = 12)
Figure 12. Fidélité des bourdons recapturés. Cet éthogramme ne tient compte que des recaptures où il y a un changement de plantes.
Nid
N
B. Nam
ur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 29
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 30
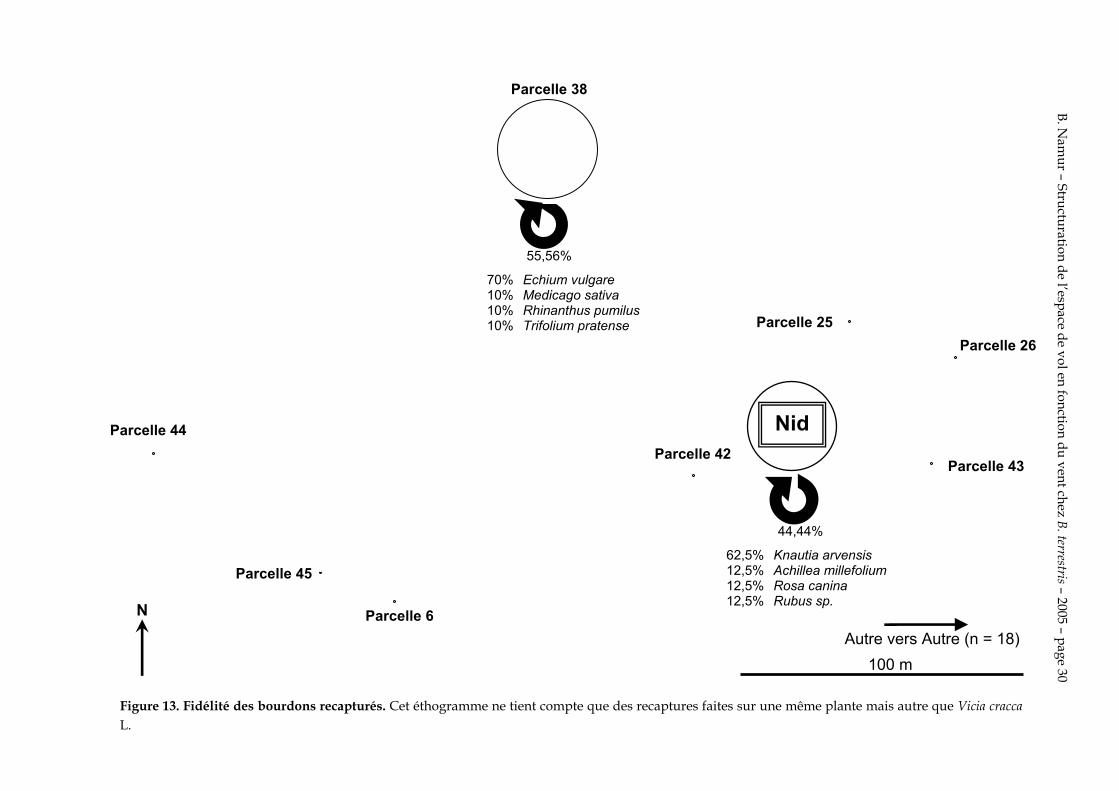
Figure 13. Fidélité des bourdons recapturés. Cet éthogramme ne tient compte que des recaptures faites sur une même plante mais autre que Vicia cracca L.
100 m
Parcelle 38
Parcelle 26 Parcelle 25
Parcelle 44
Parcelle 45
Parcelle 43 Parcelle 42
Parcelle 6Autre vers Autre (n = 18)
55,56%
44,44%
70% Echium vulgare 10% Medicago sativa 10% Rhinanthus pumilus 10% Trifolium pratense
62,5% Knautia arvensis 12,5% Achillea millefolium12,5% Rosa canina 12,5% Rubus sp.
Nid
N
B. Nam
ur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 30
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 31
On constate de plus une grande fidélité au Vicia cracca L. de la parcelle 26. Cependant certaines ouvrières changent leurs sites de butinage. Il est notable que certaines passent de la parcelle 26 à la 6, éloignée de près de 220 m et qui se trouve à l’opposée l’une de l’autre.
On remarque que c’est sur la figure 12 que les changements de site de butinage sont les plus fréquents que sur les figures 11 et 13. Les changements de site de butinage sont donc plus fréquents quand ils sont associés à un changement de plante. La fidélisation se fait donc à la fois à un site de butinage et à un type de fleur. Les bourdons qui changent de site de butinage ne recherchent donc pas systématiquement la même espèce de plante qu’ils ont butinée précédemment.
4.6. Influence du vent sur la distance de butinage
Le tableau 6 montre que le nombre d’observation lors des vents d’ouest, d’est et d’est-nord-est n’est pas suffisamment élevé pour les considérer dans le présent travail. Il y a donc un axe principal du vent nord-est sud-ouest.
Tableau 6. Nombre de recapture pendant les différents vents
Nom du vent Direction du vent Nombre de recaptures
Nombre de relevés météorologiques
SW 123 100 Vent d’Espagne W 5 1 NE 221 123 Les marinades
ENE 7 1 Carcanet E 1 2
Pour monter que les bourdons ne sortent pas plus quand le vent à un certain sens un coefficient de corrélation entre le nombre de recaptures et le nombre de relevé météorologique est fait. Le coefficient de corrélation de Pearson est de 0,973. Les nombres de recaptures pendant les différents vents sont donc fortement corrélés aux nombres de relevés météorologiques et donc à la durée proportionnelle des différents vents.
Les vitesses maximales du vent observées lors de cette étude sont des rafales à 9 m/s qui ne gênaient pas le butinage des bourdons.
Le tableau 7 reprend le nombre de recaptures selon la distance de butinage lorsque le vent est considéré comme nul (≤ 2 Beaufort = 11,9 m/s), le vent est du nord-est (> 2 Beaufort) et le vent est du sud-ouest (> 2 Beaufort).
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 32
Tableau 7. Nombre de recaptures pendant le vent nul (≤ 2 Beaufort), le vent du SW (> 2 Beaufort) et le vent du NE (> 2 Beaufort)
Distance NUL NE SW
0 - 10 9 2 1 10 - 20 8 3 0 20 - 30 2 0 1 30 - 40 6 8 0 40 - 50 37 21 26 50 - 60 29 27 20 60 - 70 15 14 13 70 - 80 7 4 1 80 - 90 0 2 2
90 - 100 2 3 0 100 - 110 3 1 3 110 - 120 2 0 1 120 - 130 1 0 0 130 - 140 3 4 1 140 - 150 7 5 4 150 - 160 1 0 1 160 - 170 2 1 1 170 - 180 5 0 1 180 - 190 1 0 0 190 - 200 0 0 0 200 - 210 0 0 0 210 - 220 7 0 0 220 - 230 5 3 0 230 - 240 1 0 0 240 - 250 0 0 0 250 - 260 0 0 0 260 - 270 0 0 0 270 - 280 0 0 0 280 - 290 0 0 0 290 - 300 0 0 0 300 - 310 0 0 0 310 - 320 0 0 0 320 - 330 0 0 0 330 - 340 0 1 0
Total 153 99 76
Le test de Kolmogorov-Smirnov permet de mettre en évidence si les nombres de recaptures dans les différentes classes de distances sont significativement différents pour les trois catégories de vent. Les résultats de ce test sont repris dans le tableau 8.
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 33
Tableau 8. Résultat du test de Kolmogorov-Smirnov
Nul vs NE Nul vs SW NE vs SW Dmax 0,097 0,111 0,105
D critique pour α = 0,05 0,175 0,191 0,207
Si Dmax > D critique test significatif NS NS NS
D critique pour α = 0,10 0,157 0,171 0,186
Si Dmax > D critique test significatif NS NS NS
Comme l’indique le tableau 8, la valeur critique est supérieur à la valeur observée, Dmax, au seuil de signification α = 0,05 ou même α = 0,10. On ne peut donc pas rejeter l'hypothèse nulle selon laquelle les échantillons ne sont pas différents. Autrement dit, la différence entre les trois vents n'est pas significative. Le nombre de recaptures dans les différentes classes de distance de butinage n’est pas différent pour les trois catégories de vent.
Les bourdons ne vont donc pas significativement plus loin ou moins loin qu’il y ait du vent ou pas et quelque soit son sens.
4.7. Influence du vent sur l’orientation de butinage
L’analyse descriptive de l’influence du vent sur l’orientation de butinage est faite sur les 3 catégories de vent: nul, nord-est et sud-ouest.
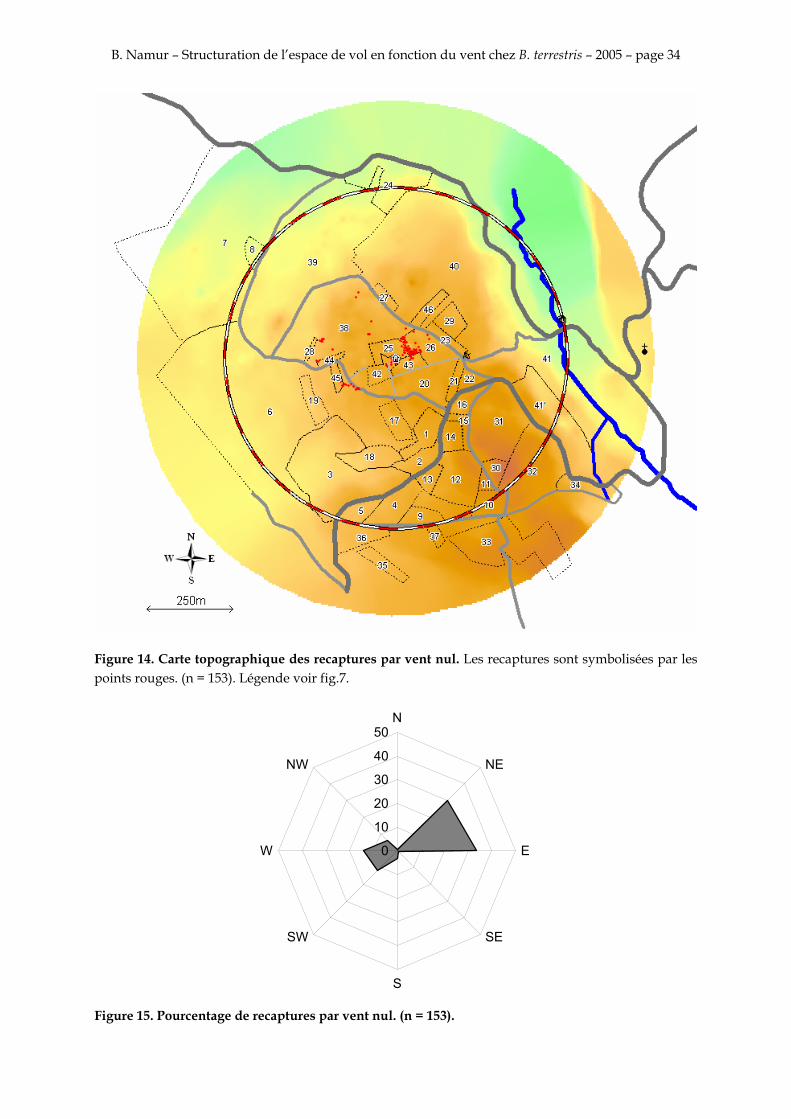
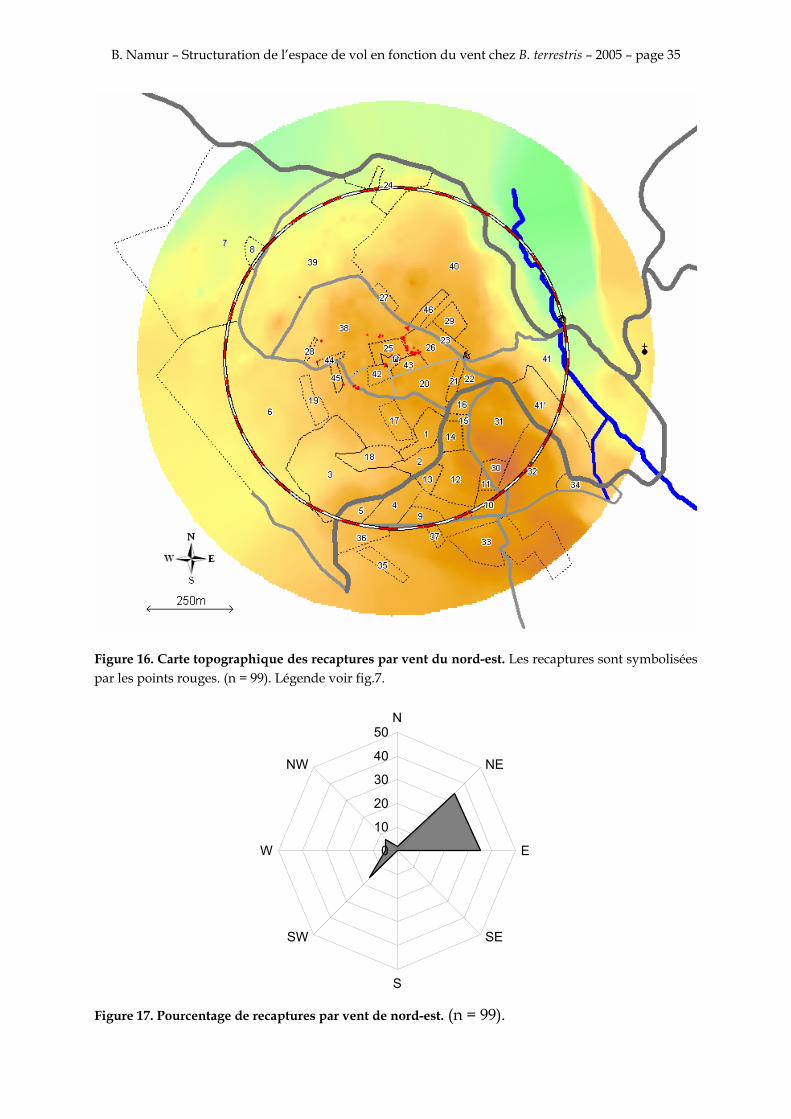
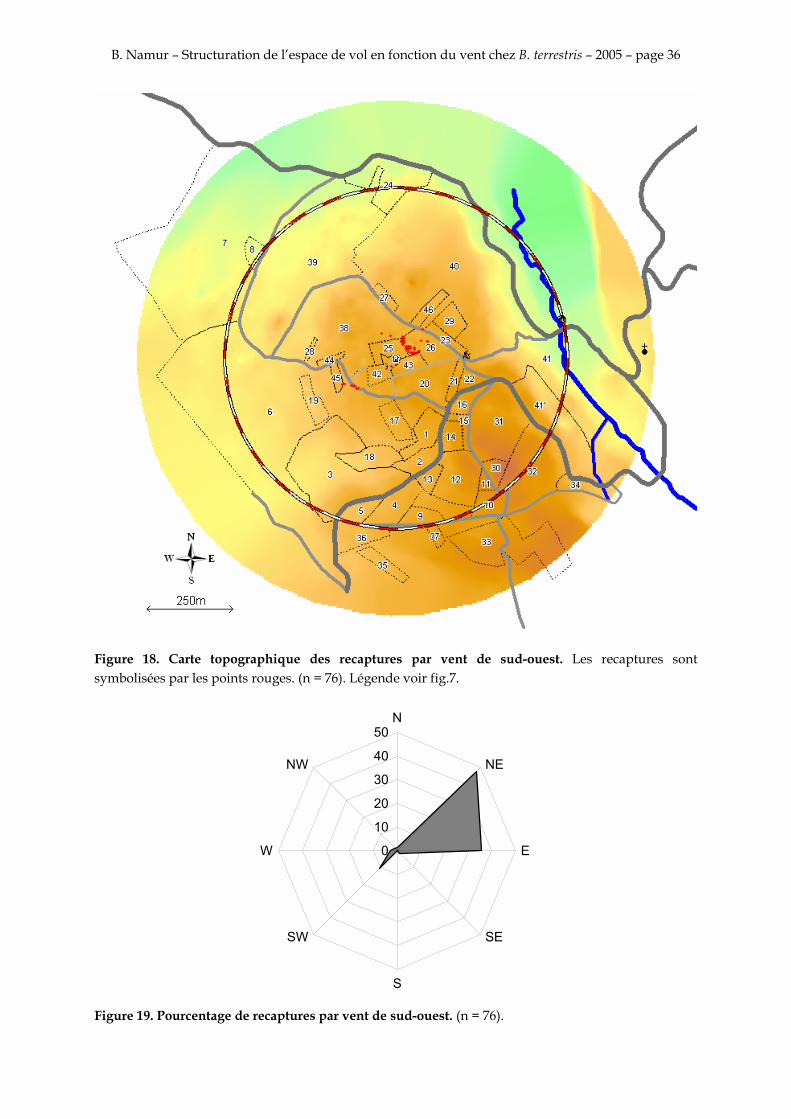
Les figures 14, 15, 16, 17, 18 et 19 montrent que la majorité des recapture sont faites sur au nord-est sur la parcelle 26 entre 40 et 70 m. Il y a aussi une deuxième concentration plus petite celle là au sud-ouest sur la parcelle 6 entre 130 et 150 m
La majorité des recaptures suit donc l’axe principal du vent. L’orientation des recaptures ne semble pas différente que le vent soit nul, de nord-est ou du sud-ouest.
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 34
Figure 14. Carte topographique des recaptures par vent nul. Les recaptures sont symbolisées par les points rouges. (n = 153). Légende voir fig.7.
0
10
20
30
40
50N
NE
E
SE
S
SW
W
NW
Figure 15. Pourcentage de recaptures par vent nul. (n = 153).
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 35
Figure 16. Carte topographique des recaptures par vent du nord-est. Les recaptures sont symbolisées par les points rouges. (n = 99). Légende voir fig.7.
0
10
20
30
40
50N
NE
E
SE
S
SW
W
NW
Figure 17. Pourcentage de recaptures par vent de nord-est. (n = 99).
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 36
Figure 18. Carte topographique des recaptures par vent de sud-ouest. Les recaptures sont symbolisées par les points rouges. (n = 76). Légende voir fig.7.
0
10
20
30
40
50N
NE
E
SE
S
SW
W
NW
Figure 19. Pourcentage de recaptures par vent de sud-ouest. (n = 76).

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 37
4.8. Structuration de l’espace de vol
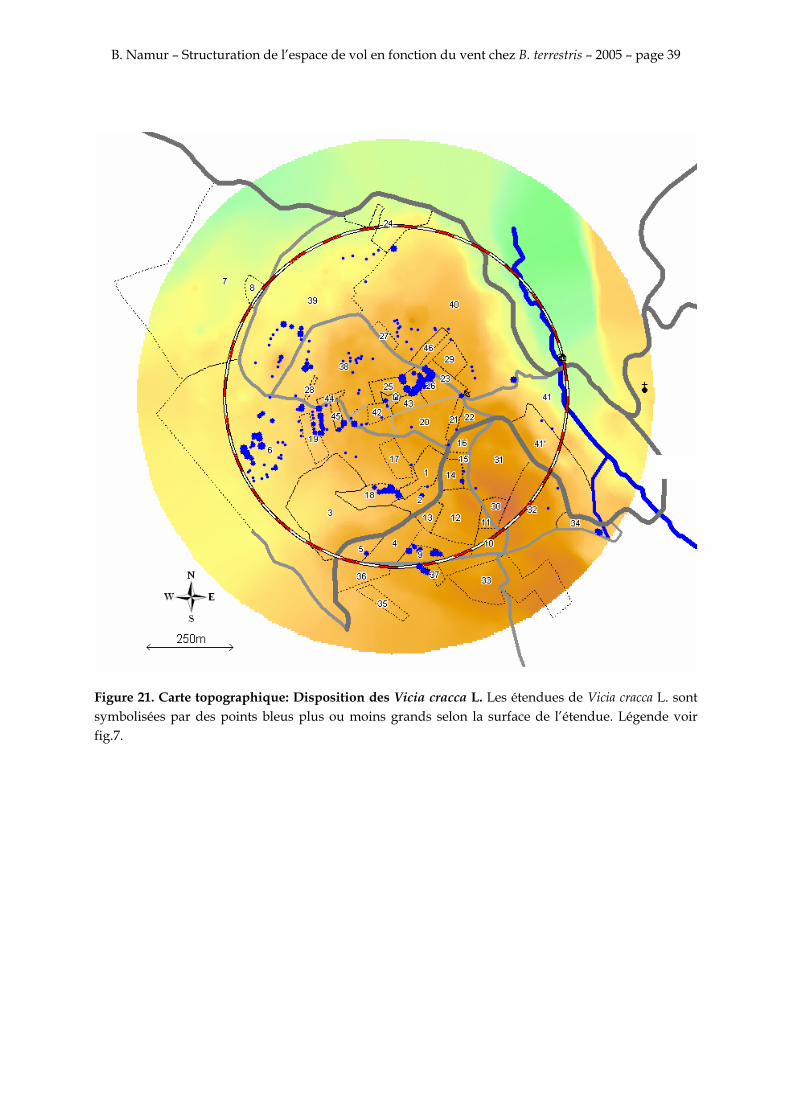
Les différents résultats s’accordent tous pour dire que la majorité des recaptures est faite sur une étendue de Vicia cracca L. au nord-est entre 40 et 70 m des colonies sur la parcelle 26. Une autre étendue de Vicia cracca L., plus petite cependant, est également attractive et se situe au sud-ouest sur la parcelle 6 entre 130 et 150 m des nids.
La figure 20 montre que même sur des parcelles pauvre en Vicia cracca L. la proportion de recaptures sur cette plante est très élevée. On constate que la parcelle 20 est très riche en Trifolium pratense L. et pourtant aucune recapture ne s’y est faite. Cependant cette parcelle n’est pas dans l’axe principal du vent et ne contient pas de Vicia cracca L..
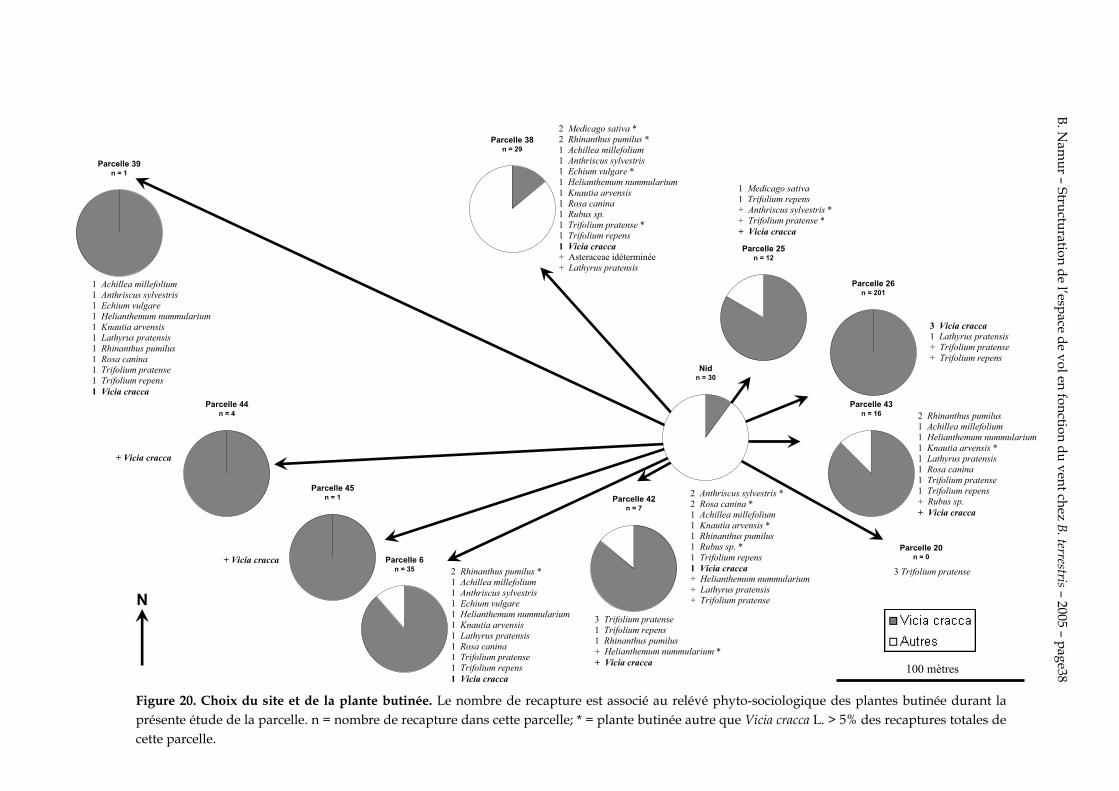
L’essentiel des recaptures se sont faites sur l’axe principal du vent. Il peut donc y avoir deux raisons possibles. Les Vicia cracca L. se concentrent sur l’axe principal du vent ou les bourdons suivent l’axe du vent quelque soit le sens.
Sur la figure 21, on peut observer que les Vicia cracca L. ne se situent pas uniquement dans les parcelles 6 et 26, qui sont dans l’axe du vent. Les parcelles 18 et 38 sont aussi recouvertes de grandes étendues de Vicia cracca L. et ne sont pas butinées. Cependant ces deux étendues ne se disposent pas dans l’axe principal du vent.
Les figures 20 et 21 permettent de dire clairement que les bourdons ne se distribuent pas au hasard sur les parcelles riches en plantes attractives. L’axe du vent a donc un effet très important sur le choix de l’orientation du site de butinage.
Tous ces résultats tendent tous vers la même conclusion. Les bourdons recherchent des plantes attractives dans l’axe du vent et délaissent la même plante en dehors de cet axe.
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 38
Parcelle 20n = 0Parcelle 6
n = 35
Parcelle 25n = 12
Parcelle 26n = 201
Parcelle 38n = 29
Parcelle 39n = 1
Parcelle 42n = 7
Parcelle 43n = 16
Parcelle 44n = 4
Parcelle 45n = 1
Figure 20. Choix du site et de la plante butinée. Le nombre de recapture est associé au relévé phyto-sociologique des plantes butinée durant la présente étude de la parcelle. n = nombre de recapture dans cette parcelle; * = plante butinée autre que Vicia cracca L. > 5% des recaptures totales de cette parcelle.
3 Trifolium pratense 2 Rhinanthus pumilus * 1 Achillea millefolium 1 Anthriscus sylvestris 1 Echium vulgare 1 Helianthemum nummularium 1 Knautia arvensis 1 Lathyrus pratensis 1 Rosa canina 1 Trifolium pratense 1 Trifolium repens 1 Vicia cracca
2 Anthriscus sylvestris * 2 Rosa canina * 1 Achillea millefolium 1 Knautia arvensis * 1 Rhinanthus pumilus 1 Rubus sp. * 1 Trifolium repens 1 Vicia cracca + Helianthemum nummularium + Lathyrus pratensis + Trifolium pratense
1 Medicago sativa 1 Trifolium repens + Anthriscus sylvestris * + Trifolium pratense * + Vicia cracca
3 Vicia cracca 1 Lathyrus pratensis + Trifolium pratense + Trifolium repens
2 Medicago sativa * 2 Rhinanthus pumilus * 1 Achillea millefolium 1 Anthriscus sylvestris 1 Echium vulgare * 1 Helianthemum nummularium 1 Knautia arvensis 1 Rosa canina 1 Rubus sp. 1 Trifolium pratense * 1 Trifolium repens 1 Vicia cracca + Asteraceae idéterminée + Lathyrus pratensis
3 Trifolium pratense 1 Trifolium repens 1 Rhinanthus pumilus + Helianthemum nummularium * + Vicia cracca
2 Rhinanthus pumilus 1 Achillea millefolium 1 Helianthemum nummularium 1 Knautia arvensis * 1 Lathyrus pratensis 1 Rosa canina 1 Trifolium pratense 1 Trifolium repens + Rubus sp. + Vicia cracca
+ Vicia cracca
+ Vicia cracca
100 mètres
1 Achillea millefolium1 Anthriscus sylvestris 1 Echium vulgare 1 Helianthemum nummularium 1 Knautia arvensis 1 Lathyrus pratensis 1 Rhinanthus pumilus 1 Rosa canina 1 Trifolium pratense 1 Trifolium repens 1 Vicia cracca
N
B. Nam
ur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page38
Nidn = 30
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 39
Figure 21. Carte topographique: Disposition des Vicia cracca L. Les étendues de Vicia cracca L. sont symbolisées par des points bleus plus ou moins grands selon la surface de l’étendue. Légende voir fig.7.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 40
5. Discussion
5.1. Choix floraux et fidélité
Ruszkowski (1971) observe Bombus terrestris (L.) et Bombus lucorum (L.), qu’il ne différencie pas sur le terrain, sur 570 espèces de plantes dont les principales sont Corydalis solida (L.) Clairv., Ribes nigrum L., Ribes uva-crispa L., Vaccinium myrtillus L., Trifolium pratense L., Medicago x-varia T. Martyn, Vicia villosa Roth, Vicia sepium L., Echium vulgare L., Melampyrum pratense L., Calluna vulgaris (L.) Hull., Helianthus annuus L., Carduus acanthoides L. et Dahlia variabilis (Willd.) Desf.. Les familles préférées sont 16% Asteraceae, 9% Fabaceae, 9% Lamiaceae, 8% Rosaceae, 5% Scrophulariaceae, 4% Ranunculaceae et 4% Liliaceae.
Gosselin (2003) étudie les choix floraux des bourdons dans la vallée de Nohèdes (France, Pyrénées-Orientales). Il constate que Bombus terrestris (L.) butine de nombreuses plantes: Fabaceae (Trifolium), Rosaceae (Rubus), Asteraceae (Cirsium), Ericaceae (Calluna et Rhododendron) et Lamiaceae (Teucrium) principalement.
Cette espèce, étudiée à Eyne par Durieux (2000), butine plus particulièrement les Fabaceae (Vicia, Genista, Trifolium) mais d’autres genres comme Rhinanthus, Centaurea, Epilobium, Rhododendron, Knautia,…
Rasmont (1988), dans son inventaire des plantes butinées par les bourdons du Languedoc-Rousillion, note le nombre de captures de Bombus terrestris (L.) sur un grand nombre de plantes, voici les principales: 395 sur Vicia cracca, 125 sur Lathyrus pratensis, 68 sur Ribes sanguineum, 67 sur Ononis spinosa, 59 sur Trifolium incarnatum, 45 sur Vicia pannonica, 28 sur Bituminaria bituminosa.
Les Fabaceae (Vicia, Trifolium), les Knautia, les Echium et les Rhinanthus forment l’essentiel des recaptures de cette présente étude ce qui est en accord avec les résultats obtenus par Ruszkowski (1971), Rasmont (1988), Durieux (2000) et Gosselin (2003).
La très grande attraction de Bombus terrestris (L.) pour Vicia cracca L., mise en évidence dans la présente étude, est également constatée par Rasmont (1988).
Dans la présente étude, on constate une grande fidélité à Vicia cracca L. sur la parcelle 26. Cependant quelques rares ouvrières changent leurs sites de butinage. Il est notable que certaines ouvrières passent de la parcelle 26 à la 6, éloignée de près de 220 m. Ces deux parcelles se situent dans des directions opposées par rapport au nid mais situées sur l’axe du vent. Les changements de site de butinage sont plus

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 41
fréquents quand ils sont associés à un changement de plante. La fidélisation se fait donc à la fois à un site de butinage et à un type de fleur. Les bourdons qui changent de site de butinage ne recherchent donc pas systématiquement la plante qu’ils ont butinée auparavant.
Le vol de nectar décrit par Alford (1975) n’a pas été observé. L’abondance de fleurs à corolles courtes dans zone d’étude ne nécessite probablement pas l’utilisation de vol de nectar. Le vol de nectar requiert de percer le bas de la corolle, ce qui demande un coût d’énergie et de temps. Cette technique est donc appliquée que si les conditions environnementales l’exigent.
Un recrutement comme celui décrit par Dornhaus & Chittka (1999, 2001) peut expliquer la très haute fidélité observée sur les Vicia cracca L. de la parcelle 26. Cependant l’attraction des Fabaceae décrit par Ruszkowski (1971), Rasmont (1988), Durieux (2000), Gosselin (2003) et leur proximité immédiate suffisent peut-être à fidéliser les ouvrières sans l’intervention de recrutement, décrit par Dornhaus & Chittka (1999, 2001), ou de marquage chimique, décrit par Cameron (1981) et Pouvreau (1990).
5.2. Distance de butinage
Walther-Hellwig & Frankl (2000) ont observé qu’une grande proportion des ouvrières de Bombus terrestris (L.) vont butiner à des distances qui vont à plus de 1,5 km. Les présentes observations indiquent que les distances de butinage sont nettement inférieures. Ceci peut s’expliquer par la proximité immédiate de grandes étendues florales très attractives.
Osborne et al. (1999) expriment leurs distances en moyenne plus ou moins écart-type et en amplitude. La moyenne ne peut être utilisée que dans le cas d’une distribution normale. Ils ne précisent pas si les distances de butinage suivent une distribution symétrique. A priori les distances de butinage ne peuvent pas être exprimées sous forme de moyenne car elles ne suivent pas une distribution normale. Ceci complique singuliérement les comparaisons avec la présente étude. L’utilisation de la médiane et des déciles pour décrire des distances de recaptures semblent mieux adaptées.
Les distances de butinage citées par Osborne et al. (1999) sont supérieures à celles observées pendant la présente étude. Une nouvelle fois la grande richesse florale de la présente zone d’étude en est probablement la cause.
Goulson & Stout (2001) ont étudié la capacité de retour des ouvrières de Bombus terrestris (L.). Ils capturent une ouvrière au niveau du nid et la relâchent à

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 42
une certaine distance de la colonie et observent si elle arrive à rejoindre sa colonie. Ces distances obtenues ne peuvent pas être comparée à des distances de butinage. Cependant leurs observations sur la manière dont les bourdons se repèrent sont intéressantes. Goulson & Stout (2001) observent que les ouvrières relâchées font des vols circulaires jusqu’à ce qu’elles trouvent un élément familier pour se repérer. Lors de la présente étude, les premières sorties de la journée d’une ouvrière, elle vole en arc de cercle autour de la colonie en s’éloignant à reculons. Cela permet de mémoriser des points de repères visuels pour le retour. Ce comportement de recherche d’éléments familiers est bien connu (Darwin, 1876; Alford, 1975).
En ce qui concerne la dispersion des sexués, la présente étude confirme les observations d’Osborne et al. (1999). Aucun des 15 mâles marqués n’est resté dans la zone d’étude. Les mâles sortent avant les femelles et s’éloignent de leur colonie, ce phénomène est en effet bien connu (Alford, 1975). Cela leur permet d’éviter de rencontrer des femelles issues de la même colonie.
5.3. Influence du vent
Woodell (1978) remarque que les bourdons se dirigent contre le vent pour aller de fleurs en fleurs. Il ne décrit pas si le vent à un effet sur l’orientation du site de butinage. Ces observations permettent de conclure que le vent est un paramètre tenu en compte par les ouvrières dans leurs comportements de butinage.
Dans la présente étude, le sens du vent ne semble pas avoir une influence sur l’orientation de butinage. La direction, par contre sembleavoir une influence sur l’orientation de butinage. La distance de butinage ne varie pas qu’il y ait du vent ou, pas et quelque soit son sens.
L’hypothèse qu’émet Woodell (1978) sur l’effet de la perception des odeurs florales par l’ouvrière, ne semble pas être confirmé dans la présente étude pour le trajet du nid au site de butinage. Cependant l’influence de la perception des odeurs florales lors des trajets de fleurs à fleurs n’est pas remise en cause.
Rasmont (1984) observe des bourdons par un vent de 33 m/s qui ne semble pas les gêner. Les vitesses les plus grandes observées lors de cette étude sont des rafales à 9 m/s qui ne gênait pas le butinage des ouvrières.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 43
5.4. Structuration de l’espace de vol
La structuration de l’espace de vol se fait donc en fonction du choix et de l’abondance des ressources alimentaires mais est aussi fonction de la direction du vent.
L’attractivité des Vicia cracca L., mise en évidence dans cette étude et celle de Rasmont (1988), et celle des Fabaceae en générale (Ruszkowsky, 1971; Alford, 1975; Rasmont, 1988; Durieux, 2000; Gosselin, 2003) sont très élevées. Cependant cette attractivité n’est pas suffisante pour que les bourdons les butinent en dehors de l’axe du vent, s’il y en a aussi sur cet axe. L’axe principal du vent a donc un effet supérieur à l’attraction des Vicia cracca L. qui se situent en dehors de cette axe.
L’effet de l’axe principal du vent est mis en évidence par la présente étude, cependant le sens du vent pourrait avoir une influence sans que la présente étude l’ait mise en évidence. Les bourdons ont une grande fidélité et donc l’auto-corrélation entre les recaptures n’est pas négligeable. Lors de changement de la direction du vent les bourdons n’auraient peut-être pas changé de site de butinage pour des raisons de fidélité au site de butinage.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 44
6. Conclusion
La fidélité des ouvrières de Bombus terrestris (L.) à un site de butinage est très élevée.
Les ouvrières de cette espèce butinent sur des grandes étendues de florales proches du nid. Vicia cracca L. est parmi les plantes les plus attractives pour cette espèce.
La distance de butinage varie entre 1 et 331m. Il semble qu’elle n’est pas fonction de l’orientation du vent, ni de sa vitesse.
Le sens du vent ne semble pas avoir une influence sur le choix du site de butinage des ouvrières et sa direction par rapport au nid.
Les ouvrières vont butiner sur les plantes attractives qui sont situées dans l’axe du vent. Les mêmes plantes attractives en dehors de cet axe ne sont que très peu visitées.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 45
7. Perspectives
Maintenant que l’étude de Woodell (1978) et la présente étude ont clairement établi que le vent à un effet sur le comportement de butinage, d’autres expériences seraient intéressantes pour notamment mettre en évidence si le sens a ou non un effet sur l’orientation du site de butinage.
Une expérience sur une zone d’étude à végétation homogène sous l’influence d’un vent à vitesse et sens constant pourrait être envisagée.
Le recrutement, décrit par Dornhaus & Chittka (1999, 2001) n’aurait pas d’influence sur un site homogène. Le bourdon qui recherche la même odeur que celle offerte par la recruteuse n’aurait pas d’influence sur l’orientation du site de butinage dans une zone d’étude à végétation homogène.
Le marquage chimique, décrit par Cameron (1981) et Pouvreau (1990), aurait quand à lui une possible influence, mais si sur la zone d’étude la compétition entre les butineuses est faible, son effet pourrait être minimisé.
La prospection d’une telle zone d’étude serait également facilitée. La prospection pourrait se faire à vitesse constante. Les trajets suivraient des cercles centrés sur les colonies. La comparaison du nombre de recaptures faites dans des classes d’orientation permettrait de dire simplement si le sens du vent à une influence sur l’orientation du site de butinage.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 46
8. Bibliographie
Alford D.V., 1975. Bumblebees. Davis, Poynter, London, XII + 352 pp., 16pls.
Andersson M., 1978. Optimal foraging area: Size and allocation of search effort. Theoretical Population Biology, 13: 397-409.
Association Charles Flahaut, (sous la direction de Amigo, J.J.), 1984. Réserve naturelle d’Eyne. Proposition de classement. Etude scientifique, Perpignan, DPN. Paris 17pp.
Bruggemann P.F., 1958. Insect environment of the hight Arctic. In Proceedings of the tenth international congress of entomology, vol.1, 695-702.
Burrough P.A. & McDonnell R.A., 2000. Principes of Geographical Information Systems. Spatial Information Systems and Geostatistics, Oxford, University Press, 333pp.
Cameron S. A., 1981. Chemical signals in bumble bee foraging. Behavioral Ecology and Sociobiology, 9: 257-260.
Coste abbé H., 1927. Flore descriptive et illustrée de la France, de la Corse et des contrées limitrophes. Librairie scientifique et technique Albert Blanchard, tome I: 416 pp., tom II: 627 pp., tome III: 807pp. Paris.
Darwin C., 1876. The effects of cross and self fertilisation in the vegetable kingdom. London.
DeMers M.N., 1997. Fondamentals of Geographic Information Systems. New Mexico State University, John Wiley & Sons, Inc., 486pp.
Delbrassinne S. & Rasmont P., 1988. Contribution à l’étude de la pollinisation du Colza Brassica napus L. var. oleifera (Moench) Delile) en Belgique. Bulletin des Recherches Agronomiques de Gembloux, 23 (2): 123-152.
Delcourt A., Mullenders W. & Piérart P., 1959. La préparation des spores et des grains de pollen, actuels et fossiles. Naturaliste Belge: 90-120.
Dornhaus A. & Chittka L., 1999. Insect behaviour – Evolutionary origins of bee dances. Nature, 401: 38.
Dornhaus A. & Chittka L., 2001. Food alert in bumblebees (Bombus terrestris): possible mechanisms and evolutionary implications. Behavioral Ecology and Sociobiology, 50: 570-576.
Dukas R. & Edelstein-Keshet L., 1998. The spatial distribution of colonial food provisioners. Journal of Theoretical Biology, 190: 121-134.
Durieux E.A., 2000. Etude des choix floraux des bourdons (Hymenoptera, Apidae) de la vallée d’Eyne (France, Pyrénées-Orientales). Mémoire de licence, Université de Mons-Hainaut, 101pp.
Erdtman G., 1960. The Acetolysis Method. A revisited description. Svensk Botanisk Tidskrift, 54: 561-564.
Gaussen H., 1970. Carte de végétation de la France. n°78, 2ème édition, Perpignan.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 47
Gosselin M., 2003. Ecologie des bourdons (Hymenoptera, Apidae) de la vallée de Nohèdes (France, Pyrénées-Orientales). Mémoire de licence, Université de Mons-Hainaut, 109pp.
Goulson D. & Stout J.C., 2001. Homing ability of the bumblebee Bombus terrestris (Hymenoptera: Apidae). Apidologie, 32: 105-111.
Goulson D., 2003. Bumblebees: their behaviour and ecology. Oxford University Press, New York. 235pp.
Goulson D., Stout J.C., Langley J. & Hughes W.O.H., 2000. Identity and function of scent marks deposited by foraging bumblebees. Journal of Chemical Ecology, 26 (12): 2897-2911.
Grey-Wilson C. & Blamey M., 1995. Le guide des fleurs de montagne. Delachaux et Niestlé, Paris, 384 pp.
Heinrich B., 1976. Ressouces partitioning among some eusocial insect: Bumblebees. Ecology, 57: 874-889.
Heinrich B., 1979a. Bumblebees economics. Harvard University press, Cambridge, 246pp.
Heinrich B., 1979b. "Majoring" and “Minoring" by foraging bumblebees, Bombus vagans: an experimental analysis. Ecology, 60: 245-255.
Iserbyt S., 2000. Ecologie des bourdons (Hymenoptera, Apidae) de la commune d’Eyne (France, Pyrénées-Orientales). Mémoires de licence, Université de Mons-Hainaut. 96pp.
Kerguélen M., 1993. Index synonymique de la flore de France. Secrétariat de faune et de la flore. Muséum national d’Histoire naturelle, Paris, 197pp.
Laverty T. M., 1994. Bumble bee learning and flower morphology. Animal behaviour, 47: 531-545.
Leppik E.E., 1957. Evolutionary relationship between entomophilars plants and anthophilars insects. Evolution, 11: 466-481.
Løken A., 1949. Bumble bees in relation to Aconitum septentrionale in Central Norway (Oeyer). Nytt Magasin for Naturvidensk, 87: 1-60.
MapInfo Professionnel version 5.5, 1999. User’s guide. MapInfo corporation Troy, New York, 589pp.
Osborne J.L., Clark S.J., Morris R.J., Williams I.H., Riley J.R., Smith A.D., Reynolds D.R. & Edwards A.S., 1999. A landscape-scale study of bumble bee foraging range and constancy, using harmonic radar. Journal of Applied Ecology, 36: 519-533.
Pouvreau A., 1990. Sur le marquage des sources de nourriture chez les bourdons. Actes colloq. Insectes sociaux, 6: 83-90.
Pouvreau A., 2004. Les insectes pollinisateurs. Delachaux et Niestlé, Paris, 190pp.
Pyke, G. 1984. Optimal foraging theory: a critical review. Annual Review of Ecology and Systematics, 15: 523-575.
Ranta E., 1983. Foraging differences in bumblebees. Annales Entomologici Fennici, 49: 17-22.
Rasmont P., 1984. Les bourdons du genre Bombus Latreille sensu stricto en Europe Occidentale et Centrale (Hymenoptera, Apidae). Spixiana, München, 7: 135-160.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 48
Rasmont P., 1988. Monographie écologique et zoogéographique des bourdons de France et de Belgique (Hymenoptera, Apidae, Bombinae). Dissertation de doctorat, Faculté des sciences agronimiques de Gembloux, Gembloux, 310 + LXIIpp.
Rasmont P., Verhaeghe J. C., Rasmont R. & Terzo M. (en préparation). West-Palaearctic bumblebees. Apollo, Svenstrup, 500pp + 38 pls, 300 figs.
Ruszkowski A., 1971. Rośliny pokarmowe i znaczenie gospodarcze trzmiela ziemnego – Bombus terrestris (L.) i trzmiela gajowego – Bombus lucorum (L.). Pamiętnik Puławski, 47: 215-250.
Saule M., 1991. La grande Flore illustrée des Pyrénées. Edition Milan, 730pp.
Saville N.M., 1993. Bumblebee ecology in woodlands and arable farmland. PhD Thesis. Cambridge University, Cambridge, UK.
Schaffer M. & Wratten S.D., 1994. Bumblebee (Bombus terrestris) movement in an intensive farm landscape. Proceeding of the 47th New Zealand Plant Protection Conference, (ed. A. J. Popay), 253-256. New Zealand Plant Protection Societry, Rotorua, New Zealand.
Schaffer M.J., 1996. Spatial aspects of bumble bee (Bombus spp: Apidae) foraging in farm landscape. Master of Applied Science Thesis. Lincoln University, Lincoln, New Zealand.
Walter H. & Lieth H., 1960. World atlas of climatic diagrams. Jena.
Walther-Hellwig K. & Frankl R., 2000. Foraging habitats and foraging distances of bumblebees, Bombus spp. (Hym., Apidae), in an agricultural landscape. Journal of Applied Ecology, 124: 299-306.
Wodehouse R.P., 1965. Pollen grains. New York and London: Hafner Publishing Compagny, 574pp.
Wolf E., 2001. Système d’information géographique. Notes de cours, Université Libre de Bruxelles, 247pp.
Woodell S.R.J., 1978. Directionality in bumblebees in relation to environmental factors. In: The pollination of floxers by insects (ed. Richards AJ), 31-39. Linnean Society Symposium Series, no.6. Academic Press, London, UK.
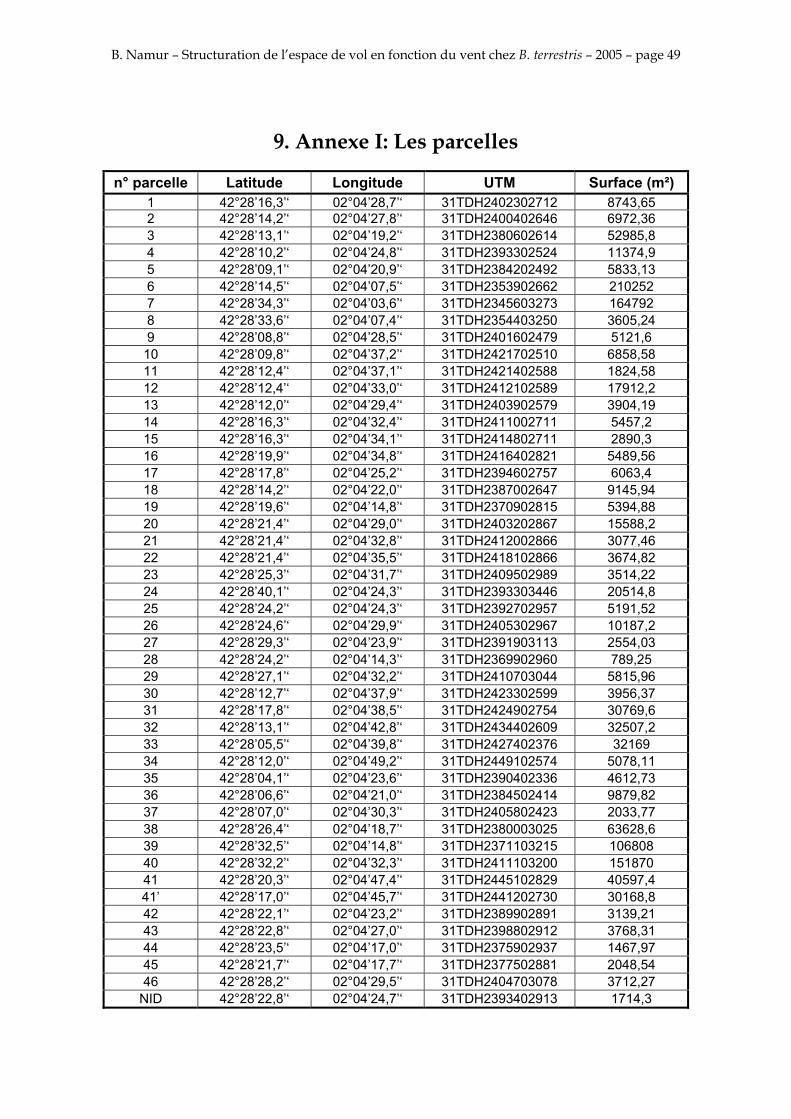
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 49
9. Annexe I: Les parcelles
n° parcelle Latitude Longitude UTM Surface (m²) 1 42°28’16,3’‘ 02°04’28,7’‘ 31TDH2402302712 8743,652 42°28’14,2’‘ 02°04’27,8’‘ 31TDH2400402646 6972,36 3 42°28’13,1’‘ 02°04’19,2’‘ 31TDH2380602614 52985,8 4 42°28’10,2’‘ 02°04’24,8’‘ 31TDH2393302524 11374,9 5 42°28’09,1’‘ 02°04’20,9’‘ 31TDH2384202492 5833,13 6 42°28’14,5’‘ 02°04’07,5’‘ 31TDH2353902662 210252 7 42°28’34,3’‘ 02°04’03,6’‘ 31TDH2345603273 164792 8 42°28’33,6’‘ 02°04’07,4’‘ 31TDH2354403250 3605,24 9 42°28’08,8’‘ 02°04’28,5’‘ 31TDH2401602479 5121,6 10 42°28’09,8’‘ 02°04’37,2’‘ 31TDH2421702510 6858,58 11 42°28’12,4’‘ 02°04’37,1’‘ 31TDH2421402588 1824,58 12 42°28’12,4’‘ 02°04’33,0’‘ 31TDH2412102589 17912,2 13 42°28’12,0’‘ 02°04’29,4’‘ 31TDH2403902579 3904,19 14 42°28’16,3’‘ 02°04’32,4’‘ 31TDH2411002711 5457,2 15 42°28’16,3’‘ 02°04’34,1’‘ 31TDH2414802711 2890,3 16 42°28’19,9’‘ 02°04’34,8’‘ 31TDH2416402821 5489,56 17 42°28’17,8’‘ 02°04’25,2’‘ 31TDH2394602757 6063,4 18 42°28’14,2’‘ 02°04’22,0’‘ 31TDH2387002647 9145,94 19 42°28’19,6’‘ 02°04’14,8’‘ 31TDH2370902815 5394,88 20 42°28’21,4’‘ 02°04’29,0’‘ 31TDH2403202867 15588,2 21 42°28’21,4’‘ 02°04’32,8’‘ 31TDH2412002866 3077,46 22 42°28’21,4’‘ 02°04’35,5’‘ 31TDH2418102866 3674,82 23 42°28’25,3’‘ 02°04’31,7’‘ 31TDH2409502989 3514,22 24 42°28’40,1’‘ 02°04’24,3’‘ 31TDH2393303446 20514,8 25 42°28’24,2’‘ 02°04’24,3’‘ 31TDH2392702957 5191,52 26 42°28’24,6’‘ 02°04’29,9’‘ 31TDH2405302967 10187,2 27 42°28’29,3’‘ 02°04’23,9’‘ 31TDH2391903113 2554,03 28 42°28’24,2’‘ 02°04’14,3’‘ 31TDH2369902960 789,25 29 42°28’27,1’‘ 02°04’32,2’‘ 31TDH2410703044 5815,96 30 42°28’12,7’‘ 02°04’37,9’‘ 31TDH2423302599 3956,37 31 42°28’17,8’‘ 02°04’38,5’‘ 31TDH2424902754 30769,6 32 42°28’13,1’‘ 02°04’42,8’‘ 31TDH2434402609 32507,2 33 42°28’05,5’‘ 02°04’39,8’‘ 31TDH2427402376 32169 34 42°28’12,0’‘ 02°04’49,2’‘ 31TDH2449102574 5078,11 35 42°28’04,1’‘ 02°04’23,6’‘ 31TDH2390402336 4612,73 36 42°28’06,6’‘ 02°04’21,0’‘ 31TDH2384502414 9879,82 37 42°28’07,0’‘ 02°04’30,3’‘ 31TDH2405802423 2033,77 38 42°28’26,4’‘ 02°04’18,7’‘ 31TDH2380003025 63628,6 39 42°28’32,5’‘ 02°04’14,8’‘ 31TDH2371103215 106808 40 42°28’32,2’‘ 02°04’32,3’‘ 31TDH2411103200 151870 41 42°28’20,3’‘ 02°04’47,4’‘ 31TDH2445102829 40597,4 41’ 42°28’17,0’‘ 02°04’45,7’‘ 31TDH2441202730 30168,8 42 42°28’22,1’‘ 02°04’23,2’‘ 31TDH2389902891 3139,21 43 42°28’22,8’‘ 02°04’27,0’‘ 31TDH2398802912 3768,31 44 42°28’23,5’‘ 02°04’17,0’‘ 31TDH2375902937 1467,97 45 42°28’21,7’‘ 02°04’17,7’‘ 31TDH2377502881 2048,54 46 42°28’28,2’‘ 02°04’29,5’‘ 31TDH2404703078 3712,27
NID 42°28’22,8’‘ 02°04’24,7’‘ 31TDH2393402913 1714,3
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 50
10. Annexe II: Les relevés phytosociologiques des parcelles
Parcelle 1 Prairie fauchée.
Parcelle 2 Prairie non fauchée haute (fauchée le 14/07/2004).
Espèces Coefficients d’abondance
Achillea millefolium L. 2 Anthriscus sylvestris (L.) Hoffmann 2 Galium verum L. 2 Geranium pratense L. 2 Tanacetum vulgare L. 2 Urtica dioica L. 2 Carduus sp. 1 Centaurea jacea L. 1 Convolvulus arvensis L. 1 Crepis sp. 1 Dactylis glomerata L. 1 Geranium sp. 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Lotus corniculatus L. 1 Plantago lanceolata L. 1 Ranunculus sp. 1 Rhinanthus pumilus (Sterneck) Soldano 1 Rosa canina L. 1 Rosa pimpinellifolia L. 1 Rumex longifolius DC. 1 Stellaria graminea L. 1 Taraxacum sp. 1 Tragopogon pratensis L. 1 Trifolium pratense L. 1 Trollius europaeus L. 1 Vicia cracca L. 1 Vicia onobrychioides L. 1 Viola tricolor L. 1 Centaurea scabiosa L. +
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 51
Parcelle 3 Prairie pâturée par des chevaux.
Espèces Coefficients d’abondance
Achillea millefolium L. 1 Brassicaceae indéterminée 1 Caltha palustris L. 1 Carduus sp. 1 Centaurea jacea L. 1 Dianthus carthusianorum L. 1 Galium verum L. 1 Genista tinctoria L. 1 Geranium pratense L. 1 Juncus sp. 1 Myosotis arvensis (L.) Hill 1 Rosa canina L. 1 Rosa pimpinellifolia L. 1 Tanacetum vulgare L. 1 Thymus serpyllum L. 1 Anthriscus sylvestris (L.) Hoffmann + Carex sp. + Cerastium brachypetalum Desportes ex Pers. + Convolvulus arvensis L. + Crepis sp. 1 + Crepis sp. 2 + Dactylis glomerata L. + Echium vulgare L. + Geranium sp. + Lathyrus pratensis L. + Lotus corniculatus L. + Plantago lanceolata L. + Plantago media L. + Polygonum bistorta L. + Ranunculus montanus Willd. R. + Ranunculus sp. + Rhinanthus pumilus (Sterneck) Soldano + Rumex longifolius DC. + Sedum rupestre L. + Tragopogon pratensis L. + Trifolium dubium Sibthorp + Trifolium ochroleucum Hudson + Trifolium pratense L. + Trifolium repens L. + Trollius europaeus L. + Veratrum album L. + Verbascum lychnitis L. + Vicia cracca L. + Vicia hirsuta (L.) S.F. Gray + Vicia onobrychioides L. + Vicia pannonica Crantz + Viola tricolor L. +
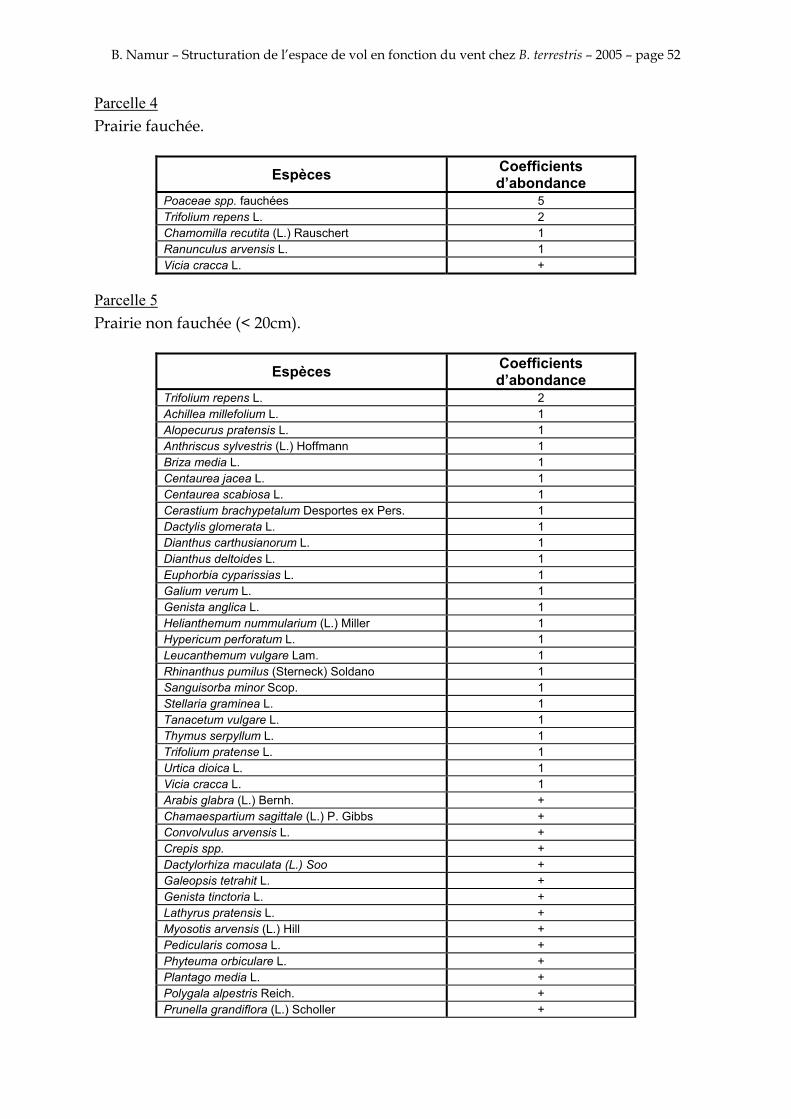
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 52
Parcelle 4 Prairie fauchée.
Espèces Coefficients d’abondance
Poaceae spp. fauchées 5 Trifolium repens L. 2 Chamomilla recutita (L.) Rauschert 1 Ranunculus arvensis L. 1 Vicia cracca L. +
Parcelle 5 Prairie non fauchée (< 20cm).
Espèces Coefficients d’abondance
Trifolium repens L. 2 Achillea millefolium L. 1 Alopecurus pratensis L. 1 Anthriscus sylvestris (L.) Hoffmann 1 Briza media L. 1 Centaurea jacea L. 1 Centaurea scabiosa L. 1 Cerastium brachypetalum Desportes ex Pers. 1 Dactylis glomerata L. 1 Dianthus carthusianorum L. 1 Dianthus deltoides L. 1 Euphorbia cyparissias L. 1 Galium verum L. 1 Genista anglica L. 1 Helianthemum nummularium (L.) Miller 1 Hypericum perforatum L. 1 Leucanthemum vulgare Lam. 1 Rhinanthus pumilus (Sterneck) Soldano 1 Sanguisorba minor Scop. 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1 Thymus serpyllum L. 1 Trifolium pratense L. 1 Urtica dioica L. 1 Vicia cracca L. 1 Arabis glabra (L.) Bernh. + Chamaespartium sagittale (L.) P. Gibbs + Convolvulus arvensis L. + Crepis spp. + Dactylorhiza maculata (L.) Soo + Galeopsis tetrahit L. + Genista tinctoria L. + Lathyrus pratensis L. + Myosotis arvensis (L.) Hill + Pedicularis comosa L. + Phyteuma orbiculare L. + Plantago media L. + Polygala alpestris Reich. + Prunella grandiflora (L.) Scholler +
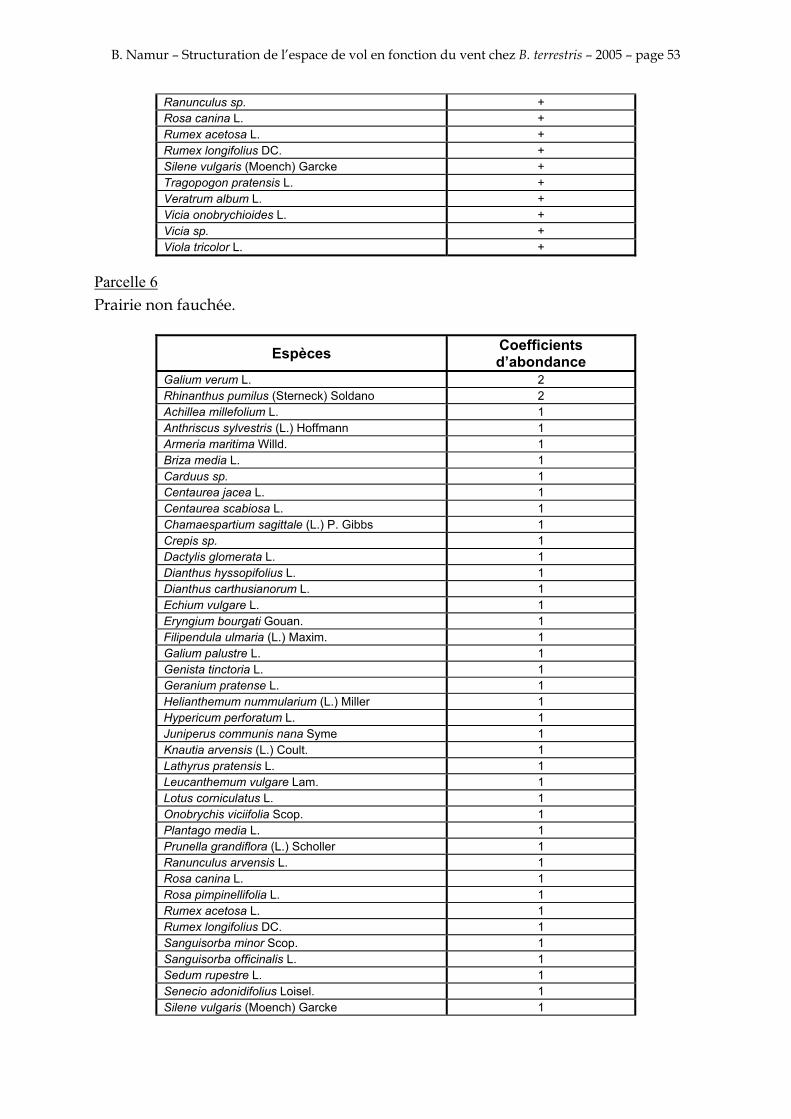
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 53
Ranunculus sp. + Rosa canina L. + Rumex acetosa L. + Rumex longifolius DC. + Silene vulgaris (Moench) Garcke + Tragopogon pratensis L. + Veratrum album L. + Vicia onobrychioides L. + Vicia sp. + Viola tricolor L. +
Parcelle 6 Prairie non fauchée.
Espèces Coefficients d’abondance
Galium verum L. 2 Rhinanthus pumilus (Sterneck) Soldano 2 Achillea millefolium L. 1 Anthriscus sylvestris (L.) Hoffmann 1 Armeria maritima Willd. 1 Briza media L. 1 Carduus sp. 1 Centaurea jacea L. 1 Centaurea scabiosa L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Crepis sp. 1 Dactylis glomerata L. 1 Dianthus hyssopifolius L. 1 Dianthus carthusianorum L. 1 Echium vulgare L. 1 Eryngium bourgati Gouan. 1 Filipendula ulmaria (L.) Maxim. 1 Galium palustre L. 1 Genista tinctoria L. 1 Geranium pratense L. 1 Helianthemum nummularium (L.) Miller 1 Hypericum perforatum L. 1 Juniperus communis nana Syme 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1 Lotus corniculatus L. 1 Onobrychis viciifolia Scop. 1 Plantago media L. 1 Prunella grandiflora (L.) Scholler 1 Ranunculus arvensis L. 1 Rosa canina L. 1 Rosa pimpinellifolia L. 1 Rumex acetosa L. 1 Rumex longifolius DC. 1 Sanguisorba minor Scop. 1 Sanguisorba officinalis L. 1 Sedum rupestre L. 1 Senecio adonidifolius Loisel. 1 Silene vulgaris (Moench) Garcke 1
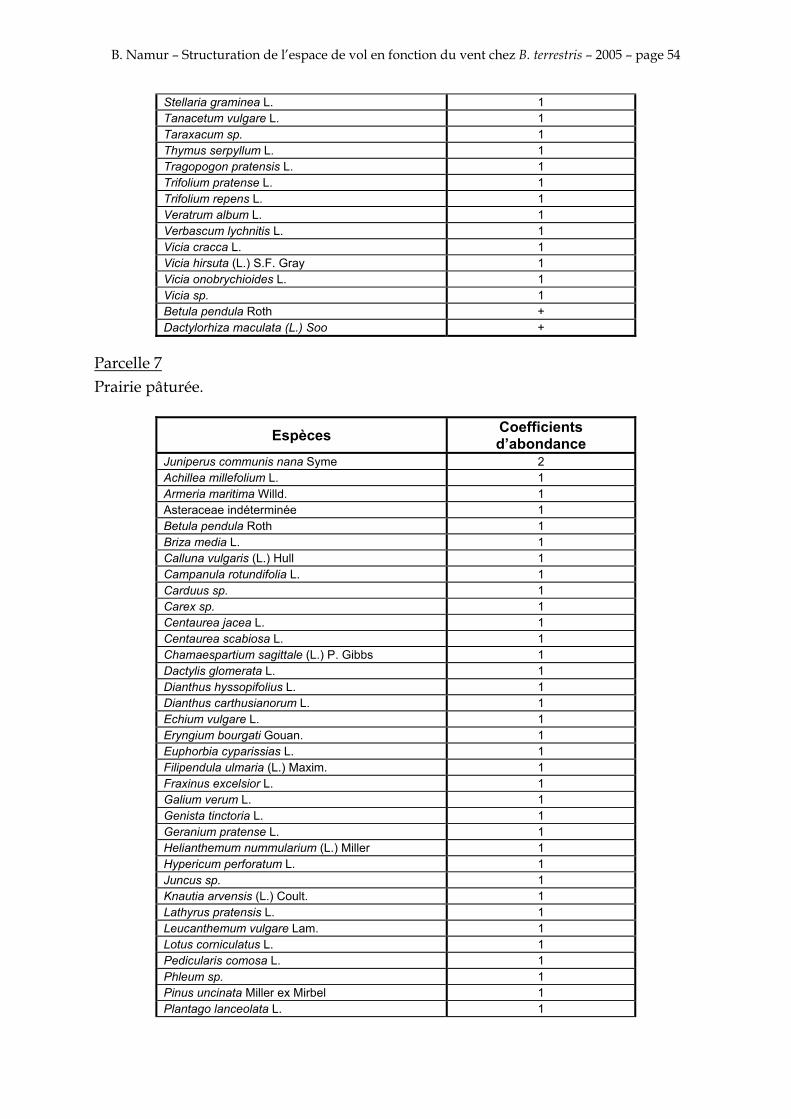
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 54
Stellaria graminea L. 1 Tanacetum vulgare L. 1 Taraxacum sp. 1 Thymus serpyllum L. 1 Tragopogon pratensis L. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Veratrum album L. 1 Verbascum lychnitis L. 1 Vicia cracca L. 1 Vicia hirsuta (L.) S.F. Gray 1 Vicia onobrychioides L. 1 Vicia sp. 1 Betula pendula Roth + Dactylorhiza maculata (L.) Soo +
Parcelle 7 Prairie pâturée.
Espèces Coefficients d’abondance
Juniperus communis nana Syme 2 Achillea millefolium L. 1 Armeria maritima Willd. 1 Asteraceae indéterminée 1 Betula pendula Roth 1 Briza media L. 1 Calluna vulgaris (L.) Hull 1 Campanula rotundifolia L. 1 Carduus sp. 1 Carex sp. 1 Centaurea jacea L. 1 Centaurea scabiosa L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Dactylis glomerata L. 1 Dianthus hyssopifolius L. 1 Dianthus carthusianorum L. 1 Echium vulgare L. 1 Eryngium bourgati Gouan. 1 Euphorbia cyparissias L. 1 Filipendula ulmaria (L.) Maxim. 1 Fraxinus excelsior L. 1 Galium verum L. 1 Genista tinctoria L. 1 Geranium pratense L. 1 Helianthemum nummularium (L.) Miller 1 Hypericum perforatum L. 1 Juncus sp. 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1 Lotus corniculatus L. 1 Pedicularis comosa L. 1 Phleum sp. 1 Pinus uncinata Miller ex Mirbel 1 Plantago lanceolata L. 1
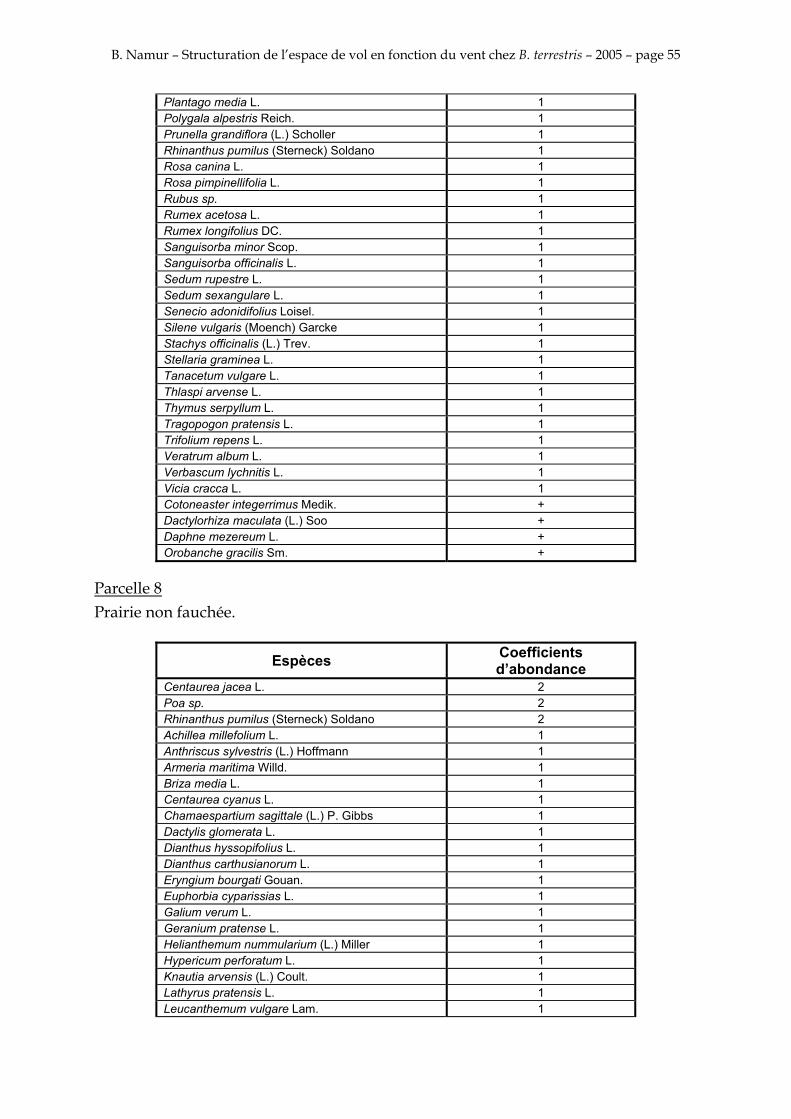
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 55
Plantago media L. 1 Polygala alpestris Reich. 1 Prunella grandiflora (L.) Scholler 1 Rhinanthus pumilus (Sterneck) Soldano 1 Rosa canina L. 1 Rosa pimpinellifolia L. 1 Rubus sp. 1 Rumex acetosa L. 1 Rumex longifolius DC. 1 Sanguisorba minor Scop. 1 Sanguisorba officinalis L. 1 Sedum rupestre L. 1 Sedum sexangulare L. 1 Senecio adonidifolius Loisel. 1 Silene vulgaris (Moench) Garcke 1 Stachys officinalis (L.) Trev. 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1 Thlaspi arvense L. 1 Thymus serpyllum L. 1 Tragopogon pratensis L. 1 Trifolium repens L. 1 Veratrum album L. 1 Verbascum lychnitis L. 1 Vicia cracca L. 1 Cotoneaster integerrimus Medik. + Dactylorhiza maculata (L.) Soo + Daphne mezereum L. + Orobanche gracilis Sm. +
Parcelle 8 Prairie non fauchée.
Espèces Coefficients d’abondance
Centaurea jacea L. 2 Poa sp. 2 Rhinanthus pumilus (Sterneck) Soldano 2 Achillea millefolium L. 1 Anthriscus sylvestris (L.) Hoffmann 1 Armeria maritima Willd. 1 Briza media L. 1 Centaurea cyanus L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Dactylis glomerata L. 1 Dianthus hyssopifolius L. 1 Dianthus carthusianorum L. 1 Eryngium bourgati Gouan. 1 Euphorbia cyparissias L. 1 Galium verum L. 1 Geranium pratense L. 1 Helianthemum nummularium (L.) Miller 1 Hypericum perforatum L. 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1
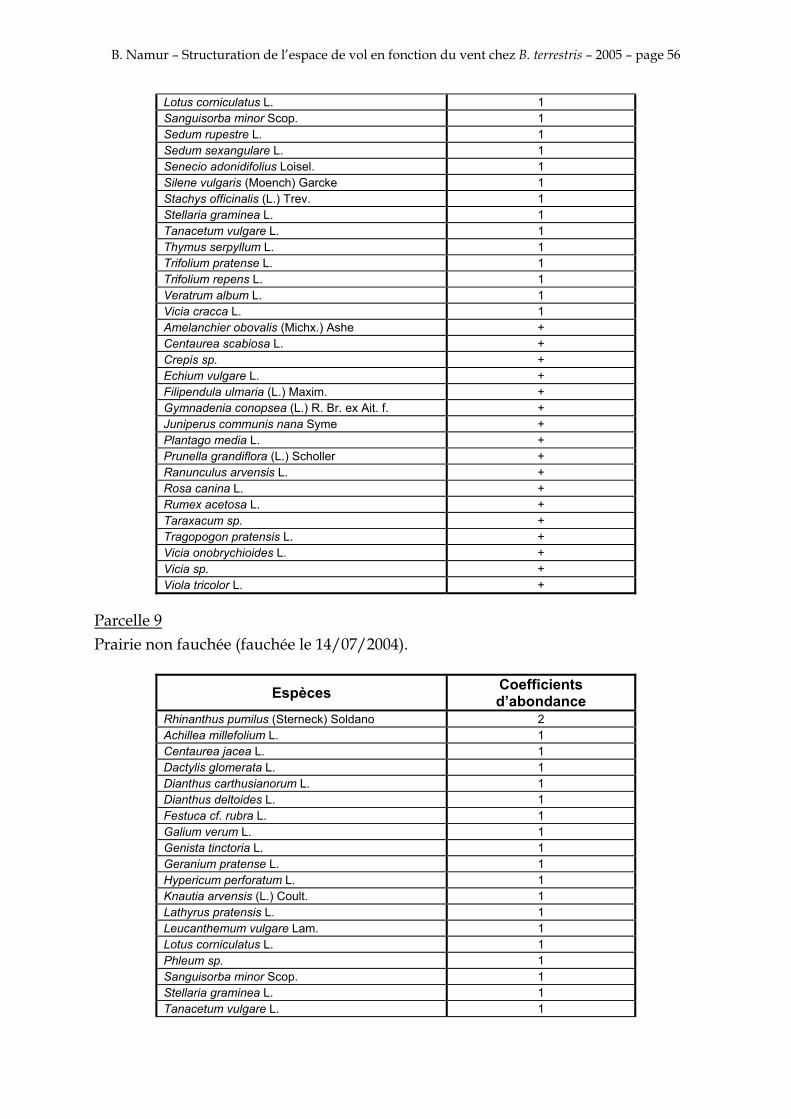
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 56
Lotus corniculatus L. 1 Sanguisorba minor Scop. 1 Sedum rupestre L. 1 Sedum sexangulare L. 1 Senecio adonidifolius Loisel. 1 Silene vulgaris (Moench) Garcke 1 Stachys officinalis (L.) Trev. 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1 Thymus serpyllum L. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Veratrum album L. 1 Vicia cracca L. 1 Amelanchier obovalis (Michx.) Ashe + Centaurea scabiosa L. + Crepis sp. + Echium vulgare L. + Filipendula ulmaria (L.) Maxim. + Gymnadenia conopsea (L.) R. Br. ex Ait. f. + Juniperus communis nana Syme + Plantago media L. + Prunella grandiflora (L.) Scholler + Ranunculus arvensis L. + Rosa canina L. + Rumex acetosa L. + Taraxacum sp. + Tragopogon pratensis L. + Vicia onobrychioides L. + Vicia sp. + Viola tricolor L. +
Parcelle 9 Prairie non fauchée (fauchée le 14/07/2004).
Espèces Coefficients d’abondance
Rhinanthus pumilus (Sterneck) Soldano 2 Achillea millefolium L. 1 Centaurea jacea L. 1 Dactylis glomerata L. 1 Dianthus carthusianorum L. 1 Dianthus deltoides L. 1 Festuca cf. rubra L. 1 Galium verum L. 1 Genista tinctoria L. 1 Geranium pratense L. 1 Hypericum perforatum L. 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1 Lotus corniculatus L. 1 Phleum sp. 1 Sanguisorba minor Scop. 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 57
Thymus serpyllum L. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Vicia cracca L. 1 Vicia onobrychioides L. 1 Armeria maritima Willd. + Carduus pp. + Chamaespartium sagittale (L.) P. Gibbs + Crepis sp. + Dactylorhiza maculata (L.) Soo + Plantago lanceolata L. + Ranunculus arvensis L. + Rosa canina L. + Rosa pimpinellifolia L. + Rumex acetosa L. + Senecio adonidifolius Loisel. + Stachys officinalis (L.) Trev. + Tragopogon pratensis L. + Verbascum lychnitis L. + Vicia hirsuta (L.) S.F. Gray + Vicia sp. +
Parcelle 10 Culture de seigle (Secale cereale L.).
Parcelle 11 Culture fauchée.
Espèces Coefficients d’abondance
Poaceae spp. fauchées 5 Trifolium repens L. +
Parcelle 12 Culture fauchée.
Espèces Coefficients d’abondance
Poaceae spp. fauchées 5 Trifolium repens L. +
Parcelle 13 Champ labouré.
Parcelle 14 Culture de pomme de terre (Solanum tuberosum L.).
Parcelle 15 Prairie basse.
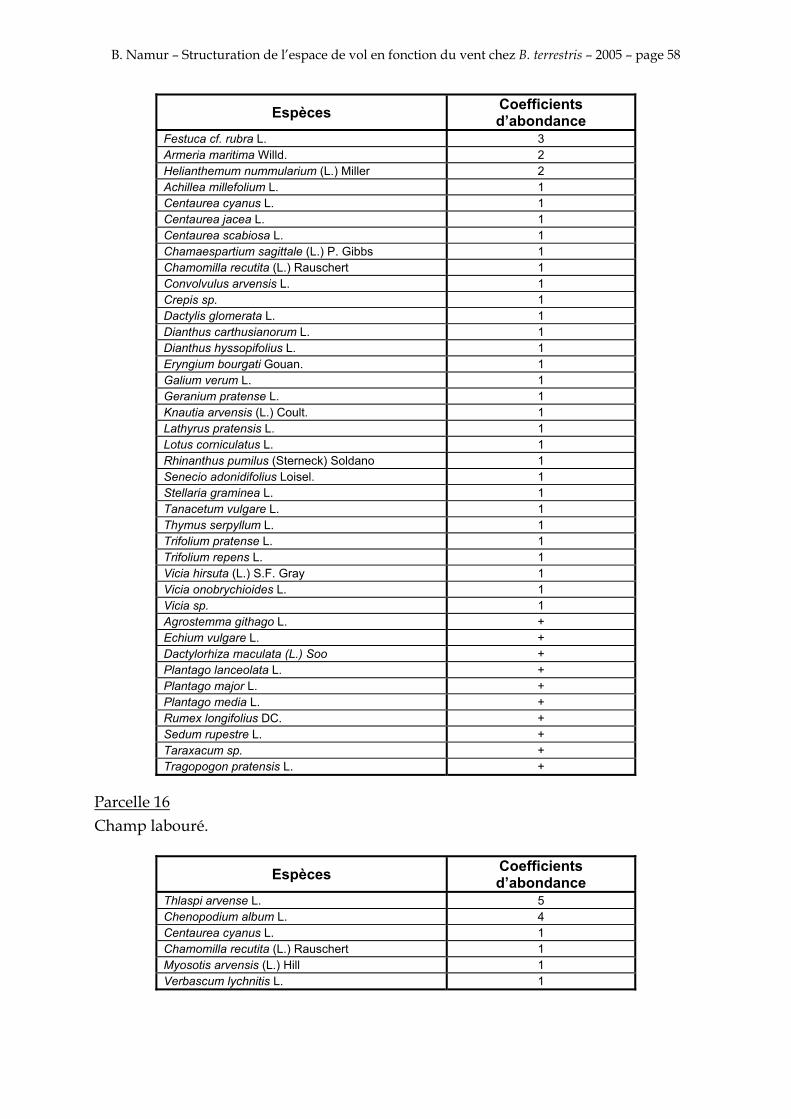
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 58
Espèces Coefficients d’abondance
Festuca cf. rubra L. 3 Armeria maritima Willd. 2 Helianthemum nummularium (L.) Miller 2 Achillea millefolium L. 1 Centaurea cyanus L. 1 Centaurea jacea L. 1 Centaurea scabiosa L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Chamomilla recutita (L.) Rauschert 1 Convolvulus arvensis L. 1 Crepis sp. 1 Dactylis glomerata L. 1 Dianthus carthusianorum L. 1 Dianthus hyssopifolius L. 1 Eryngium bourgati Gouan. 1 Galium verum L. 1 Geranium pratense L. 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Lotus corniculatus L. 1 Rhinanthus pumilus (Sterneck) Soldano 1 Senecio adonidifolius Loisel. 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1 Thymus serpyllum L. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Vicia hirsuta (L.) S.F. Gray 1 Vicia onobrychioides L. 1 Vicia sp. 1 Agrostemma githago L. + Echium vulgare L. + Dactylorhiza maculata (L.) Soo + Plantago lanceolata L. + Plantago major L. + Plantago media L. + Rumex longifolius DC. + Sedum rupestre L. + Taraxacum sp. + Tragopogon pratensis L. +
Parcelle 16 Champ labouré.
Espèces Coefficients d’abondance
Thlaspi arvense L. 5 Chenopodium album L. 4 Centaurea cyanus L. 1 Chamomilla recutita (L.) Rauschert 1 Myosotis arvensis (L.) Hill 1 Verbascum lychnitis L. 1
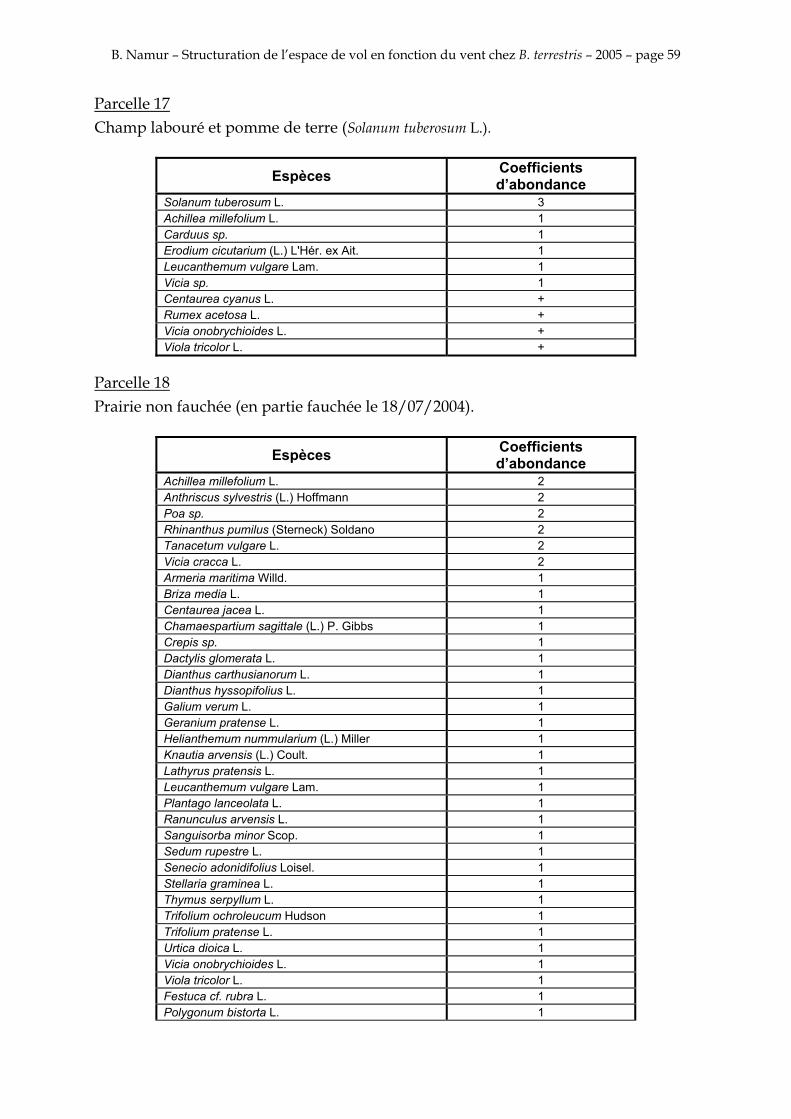
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 59
Parcelle 17 Champ labouré et pomme de terre (Solanum tuberosum L.).
Espèces Coefficients d’abondance
Solanum tuberosum L. 3 Achillea millefolium L. 1 Carduus sp. 1 Erodium cicutarium (L.) L'Hér. ex Ait. 1 Leucanthemum vulgare Lam. 1 Vicia sp. 1 Centaurea cyanus L. + Rumex acetosa L. + Vicia onobrychioides L. + Viola tricolor L. +
Parcelle 18 Prairie non fauchée (en partie fauchée le 18/07/2004).
Espèces Coefficients d’abondance
Achillea millefolium L. 2 Anthriscus sylvestris (L.) Hoffmann 2 Poa sp. 2 Rhinanthus pumilus (Sterneck) Soldano 2 Tanacetum vulgare L. 2 Vicia cracca L. 2 Armeria maritima Willd. 1 Briza media L. 1 Centaurea jacea L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Crepis sp. 1 Dactylis glomerata L. 1 Dianthus carthusianorum L. 1 Dianthus hyssopifolius L. 1 Galium verum L. 1 Geranium pratense L. 1 Helianthemum nummularium (L.) Miller 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1 Plantago lanceolata L. 1 Ranunculus arvensis L. 1 Sanguisorba minor Scop. 1 Sedum rupestre L. 1 Senecio adonidifolius Loisel. 1 Stellaria graminea L. 1 Thymus serpyllum L. 1 Trifolium ochroleucum Hudson 1 Trifolium pratense L. 1 Urtica dioica L. 1 Vicia onobrychioides L. 1 Viola tricolor L. 1 Festuca cf. rubra L. 1 Polygonum bistorta L. 1

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 60
Echium vulgare L. + Euphorbia cyparissias L. + Gymnadenia conopsea (L.) R. Br. ex Ait. f. + Hypericum perforatum L. + Orobanche gracilis Sm. + Rosa canina L. + Rumex acetosa L. + Salix sp. 1 + Salix sp. 2 + Trollius europaeus L. + Verbascum lychnitis L. +
Parcelle 19 Prairie pâturée par des chevaux.
Espèces Coefficients d’abondance
Tanacetum vulgare L. 2 Carduus sp. 1 Centaurea cyanus L. 1 Convolvulus arvensis L. 1 Erodium cicutarium (L.) L'Hér. ex Ait. 1 Leucanthemum vulgare Lam. 1 Plantago lanceolata L. 1 Rosa canina L. 1 Rosa sp. 1 Rubus sp. 1 Sedum rupestre L. 1 Stellaria graminea L. 1 Thymus serpyllum L. 1 Trifolium repens L. 1 Urtica dioica L. 1 Verbascum lychnitis L. 1 Vicia hirsuta (L.) S.F. Gray 1 Vicia sp. 1 Viola tricolor L. 1 Anthriscus sylvestris (L.) Hoffmann + Centaurea scabiosa L. + Chamaespartium sagittale (L.) P. Gibbs + Dianthus deltoides L. + Cotoneaster integerrimus Medik. + Euphorbia cyparissias L. + Galium verum L. + Genista tinctoria L. + Helianthemum nummularium (L.) Miller + Knautia arvensis (L.) Coult. + Lathyrus pratensis L. + Silene vulgaris (Moench) Garcke + Trifolium pratense L. +
Parcelle 20 Prairie à Trifolium pratense L. et Centaurea cyanus L. (fauchée le 15/07/2004).
Espèces Coefficients d’abondance

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 61
Trifolium pratense L. 3 Centaurea cyanus L. 2 Leucanthemum vulgare Lam. 1
Parcelle 21 Culture de seigle (Secale cereale L.).
Espèces Coefficients d’abondance
Secale cereale L. 5 Carduus sp. 1 Centaurea cyanus L. 1 Leucanthemum vulgare Lam. 1 Lolium sp. 1 Agrostemma githago L. +
Parcelle 22 Champ labouré.
Espèces Coefficients d’abondance
Chenopodium album L. 2 Carduus sp. 1 Chamomilla recutita (L.) Rauschert 1 Geranium pratense L. 1 Lolium sp. 1 Plantago major L. 1 Tanacetum vulgare L. 1
Parcelle 23 Culture de pomme de terre (Solanum tuberosum L.).
Espèces Coefficients d’abondance
Solanum tuberosum L. 5 Carduus sp. 1
Parcelle 24 Prairie non fauchée.
Espèces Coefficients d’abondance
Peucedanum austriacum (Jacq.) Koch. 2 Rhinanthus pumilus (Sterneck) Soldano 2 Tanacetum vulgare L. 2 Vicia cracca L. 2 Achillea millefolium L. 1 Anthriscus sylvestris (L.) Hoffmann 1 Armeria maritima Willd. 1 Betula pendula Roth 1 Briza media L. 1 Carduus sp. 1
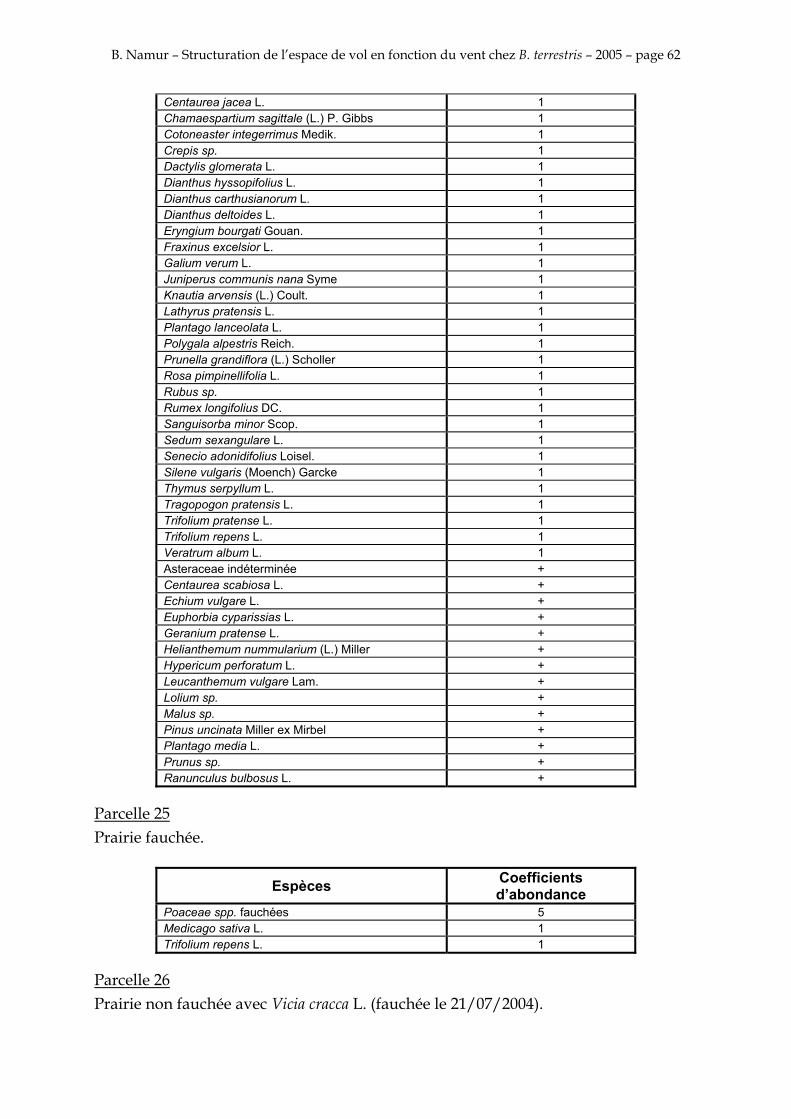
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 62
Centaurea jacea L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Cotoneaster integerrimus Medik. 1 Crepis sp. 1 Dactylis glomerata L. 1 Dianthus hyssopifolius L. 1 Dianthus carthusianorum L. 1 Dianthus deltoides L. 1 Eryngium bourgati Gouan. 1 Fraxinus excelsior L. 1 Galium verum L. 1 Juniperus communis nana Syme 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Plantago lanceolata L. 1 Polygala alpestris Reich. 1 Prunella grandiflora (L.) Scholler 1 Rosa pimpinellifolia L. 1 Rubus sp. 1 Rumex longifolius DC. 1 Sanguisorba minor Scop. 1 Sedum sexangulare L. 1 Senecio adonidifolius Loisel. 1 Silene vulgaris (Moench) Garcke 1 Thymus serpyllum L. 1 Tragopogon pratensis L. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Veratrum album L. 1 Asteraceae indéterminée + Centaurea scabiosa L. + Echium vulgare L. + Euphorbia cyparissias L. + Geranium pratense L. + Helianthemum nummularium (L.) Miller + Hypericum perforatum L. + Leucanthemum vulgare Lam. + Lolium sp. + Malus sp. + Pinus uncinata Miller ex Mirbel + Plantago media L. + Prunus sp. + Ranunculus bulbosus L. +
Parcelle 25 Prairie fauchée.
Espèces Coefficients d’abondance
Poaceae spp. fauchées 5 Medicago sativa L. 1 Trifolium repens L. 1
Parcelle 26 Prairie non fauchée avec Vicia cracca L. (fauchée le 21/07/2004).
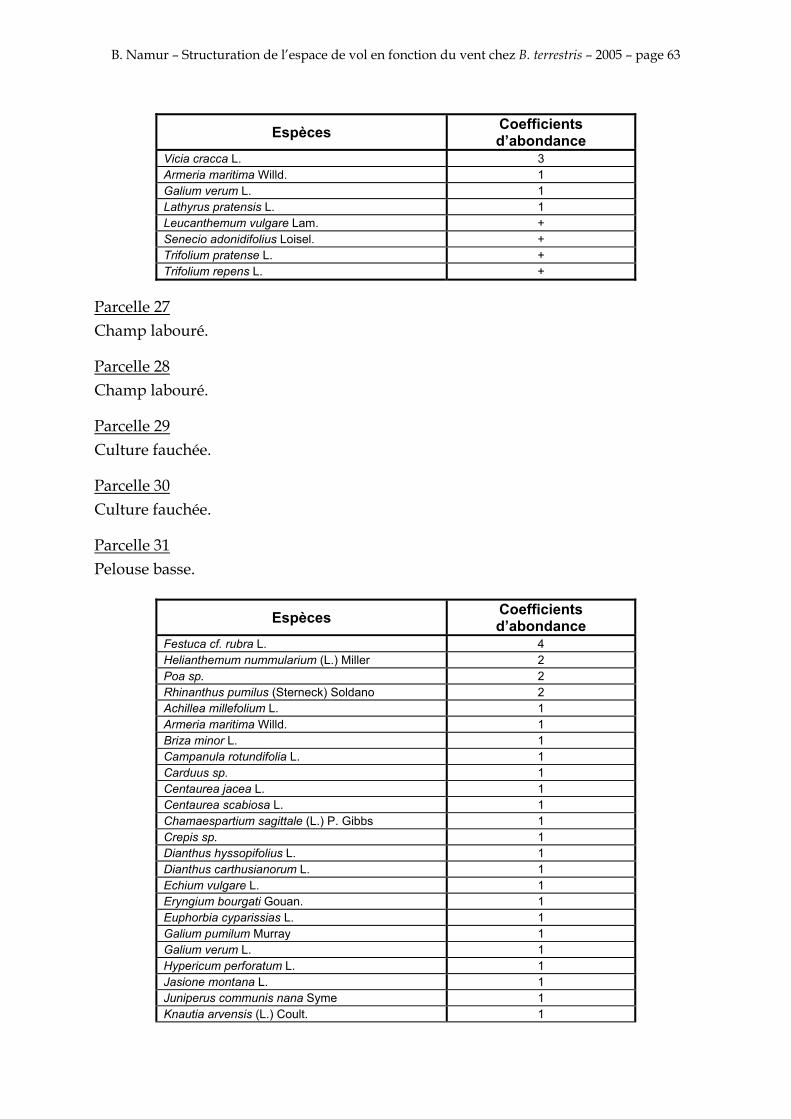
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 63
Espèces Coefficients d’abondance
Vicia cracca L. 3 Armeria maritima Willd. 1 Galium verum L. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. + Senecio adonidifolius Loisel. + Trifolium pratense L. + Trifolium repens L. +
Parcelle 27 Champ labouré.
Parcelle 28 Champ labouré.
Parcelle 29 Culture fauchée.
Parcelle 30 Culture fauchée.
Parcelle 31 Pelouse basse.
Espèces Coefficients d’abondance
Festuca cf. rubra L. 4 Helianthemum nummularium (L.) Miller 2 Poa sp. 2 Rhinanthus pumilus (Sterneck) Soldano 2 Achillea millefolium L. 1 Armeria maritima Willd. 1 Briza minor L. 1 Campanula rotundifolia L. 1 Carduus sp. 1 Centaurea jacea L. 1 Centaurea scabiosa L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Crepis sp. 1 Dianthus hyssopifolius L. 1 Dianthus carthusianorum L. 1 Echium vulgare L. 1 Eryngium bourgati Gouan. 1 Euphorbia cyparissias L. 1 Galium pumilum Murray 1 Galium verum L. 1 Hypericum perforatum L. 1 Jasione montana L. 1 Juniperus communis nana Syme 1 Knautia arvensis (L.) Coult. 1

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 64
Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1 Lotus corniculatus L. 1 Orobanche gracilis Sm. 1 Plantago media L. 1 Rosa canina L. 1 Rumex acetosa L. 1 Sanguisorba officinalis L. 1 Sedum acre L. 1 Sedum rupestre L. 1 Sempervivum montanum L. 1 Senecio adonidifolius Loisel. 1 Silene vulgaris (Moench) Garcke 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1 Taraxacum sp. 1 Thymus serpyllum L. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Verbascum lychnitis L. 1 Vicia cracca L. 1 Vicia onobrychioides L. 1 Viola tricolor L. 1 Arctium lappa L. + Biscutella sp. + Cf. Daucus sp. + Euphrasia Cf. nemorosa (Pers.) Wallr. + Onobrychis viciifolia Scop. + Pinus uncinata Miller ex Mirbel + Tragopogon pratensis L. +
Parcelle 32 Prairie non pâturée.
Espèces Coefficients d’abondance
Achillea millefolium L. 1 Dianthus carthusianorum L. 1 Galium verum L. 1 Knautia arvensis (L.) Coult. 1 Poa sp. 1 Rhinanthus pumilus (Sterneck) Soldano 1 Rosa canina L. 1 Trifolium pratense L. 1 Vicia cracca L. 1 Armeria maritima Willd. + Centaurea scabiosa L. + Cf. Daucus sp. + Crepis sp. + Dactylis glomerata L. + Dianthus deltoides L. + Echium vulgare L. + Gentiana lutea L. + Leucanthemum vulgare Lam. + Lotus corniculatus L. + Orobanche gracilis Sm. +
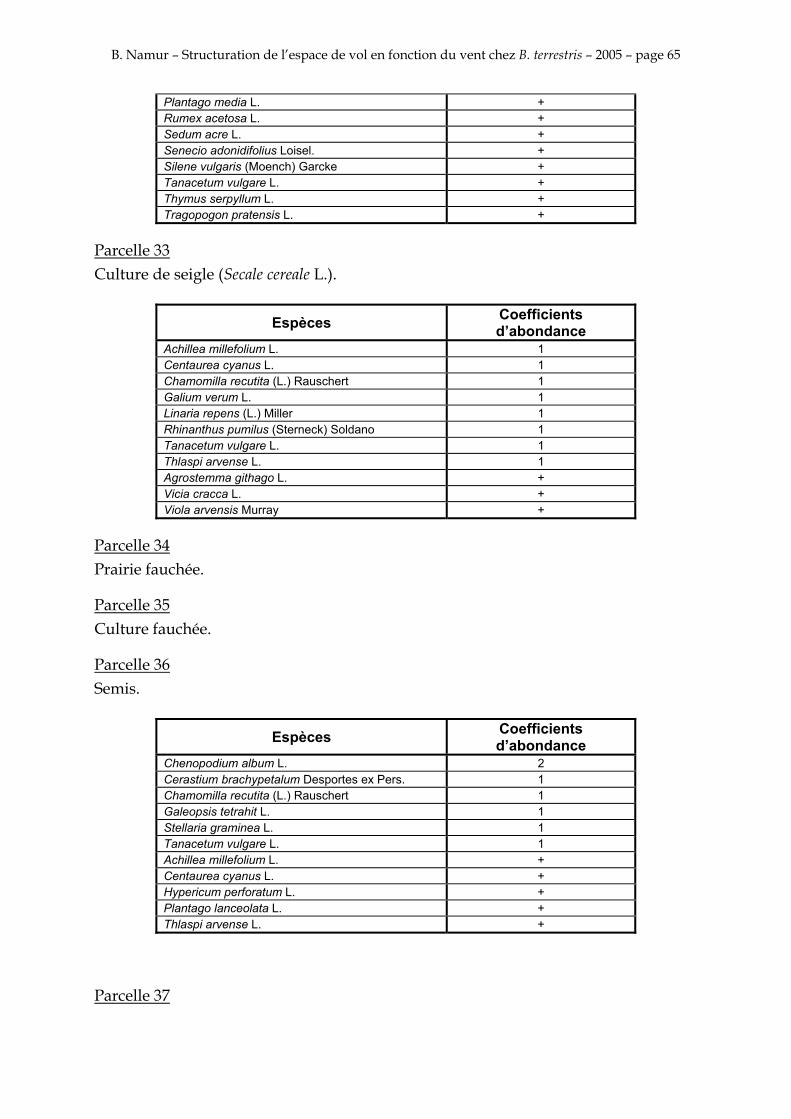
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 65
Plantago media L. + Rumex acetosa L. + Sedum acre L. + Senecio adonidifolius Loisel. + Silene vulgaris (Moench) Garcke + Tanacetum vulgare L. + Thymus serpyllum L. + Tragopogon pratensis L. +
Parcelle 33 Culture de seigle (Secale cereale L.).
Espèces Coefficients d’abondance
Achillea millefolium L. 1 Centaurea cyanus L. 1 Chamomilla recutita (L.) Rauschert 1 Galium verum L. 1 Linaria repens (L.) Miller 1 Rhinanthus pumilus (Sterneck) Soldano 1 Tanacetum vulgare L. 1 Thlaspi arvense L. 1 Agrostemma githago L. + Vicia cracca L. + Viola arvensis Murray +
Parcelle 34 Prairie fauchée.
Parcelle 35 Culture fauchée.
Parcelle 36 Semis.
Espèces Coefficients d’abondance
Chenopodium album L. 2 Cerastium brachypetalum Desportes ex Pers. 1 Chamomilla recutita (L.) Rauschert 1 Galeopsis tetrahit L. 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1 Achillea millefolium L. + Centaurea cyanus L. + Hypericum perforatum L. + Plantago lanceolata L. + Thlaspi arvense L. +
Parcelle 37
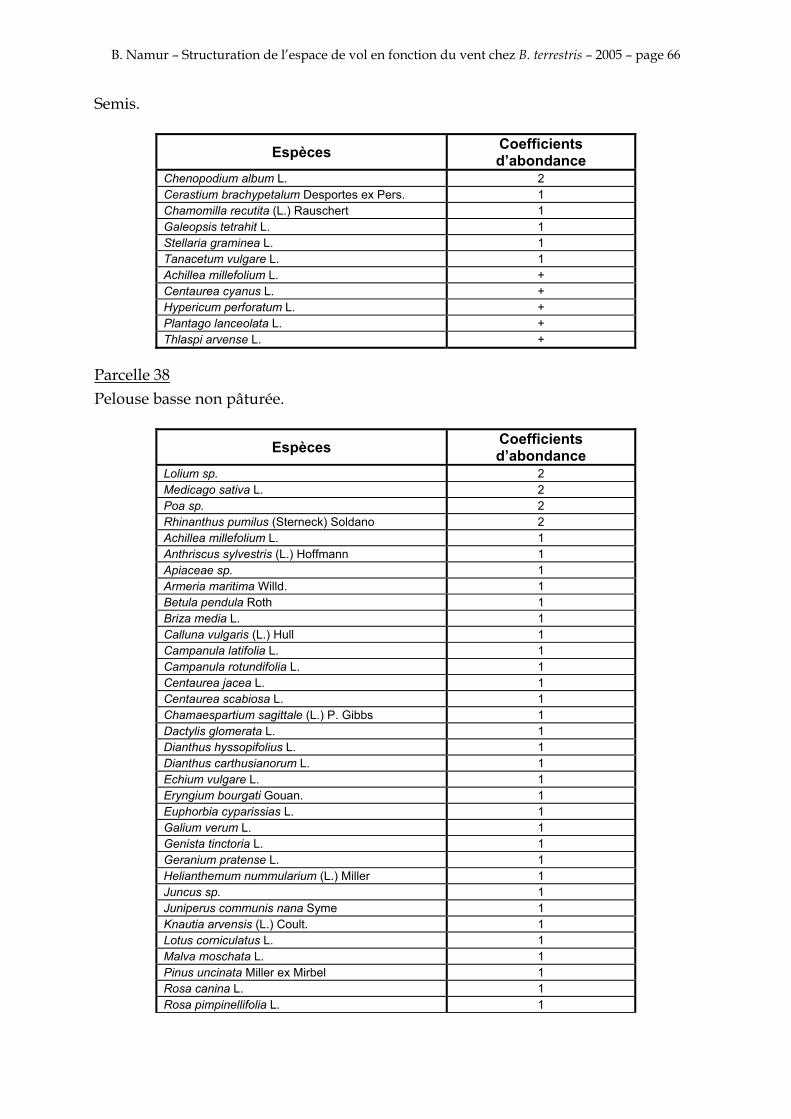
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 66
Semis.
Espèces Coefficients d’abondance
Chenopodium album L. 2 Cerastium brachypetalum Desportes ex Pers. 1 Chamomilla recutita (L.) Rauschert 1 Galeopsis tetrahit L. 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1 Achillea millefolium L. + Centaurea cyanus L. + Hypericum perforatum L. + Plantago lanceolata L. + Thlaspi arvense L. +
Parcelle 38 Pelouse basse non pâturée.
Espèces Coefficients d’abondance
Lolium sp. 2 Medicago sativa L. 2 Poa sp. 2 Rhinanthus pumilus (Sterneck) Soldano 2 Achillea millefolium L. 1 Anthriscus sylvestris (L.) Hoffmann 1 Apiaceae sp. 1 Armeria maritima Willd. 1 Betula pendula Roth 1 Briza media L. 1 Calluna vulgaris (L.) Hull 1 Campanula latifolia L. 1 Campanula rotundifolia L. 1 Centaurea jacea L. 1 Centaurea scabiosa L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Dactylis glomerata L. 1 Dianthus hyssopifolius L. 1 Dianthus carthusianorum L. 1 Echium vulgare L. 1 Eryngium bourgati Gouan. 1 Euphorbia cyparissias L. 1 Galium verum L. 1 Genista tinctoria L. 1 Geranium pratense L. 1 Helianthemum nummularium (L.) Miller 1 Juncus sp. 1 Juniperus communis nana Syme 1 Knautia arvensis (L.) Coult. 1 Lotus corniculatus L. 1 Malva moschata L. 1 Pinus uncinata Miller ex Mirbel 1 Rosa canina L. 1 Rosa pimpinellifolia L. 1
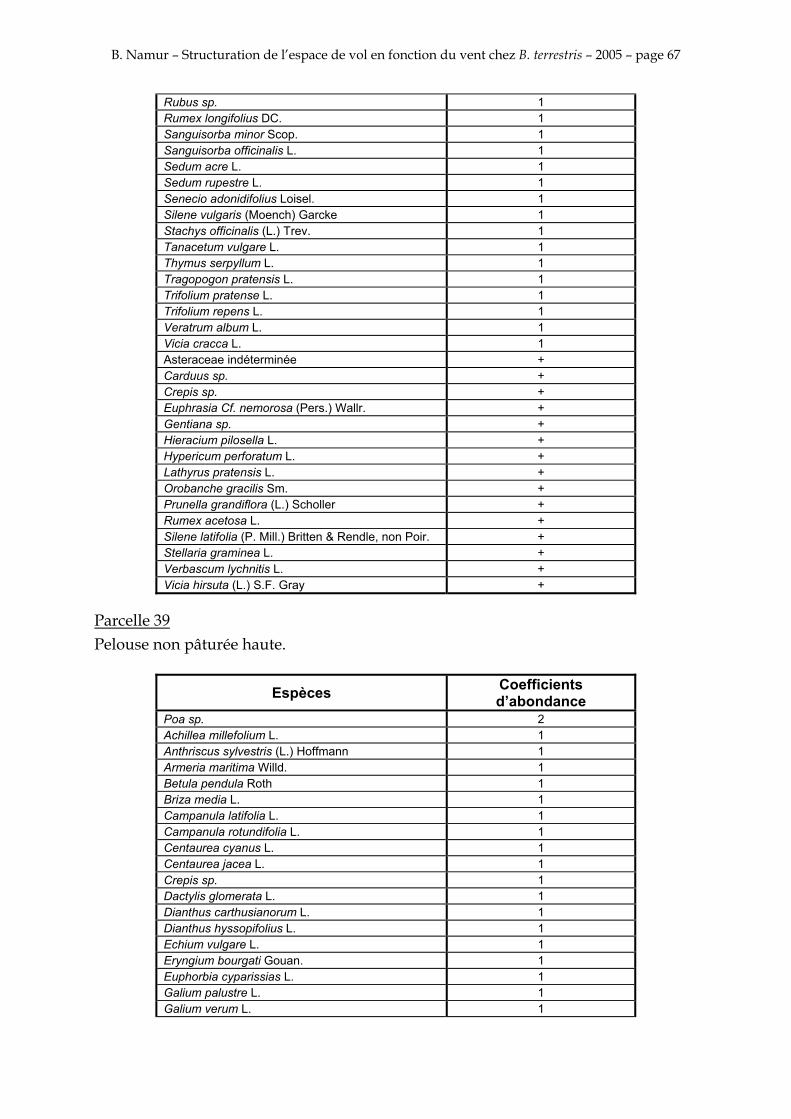
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 67
Rubus sp. 1 Rumex longifolius DC. 1 Sanguisorba minor Scop. 1 Sanguisorba officinalis L. 1 Sedum acre L. 1 Sedum rupestre L. 1 Senecio adonidifolius Loisel. 1 Silene vulgaris (Moench) Garcke 1 Stachys officinalis (L.) Trev. 1 Tanacetum vulgare L. 1 Thymus serpyllum L. 1 Tragopogon pratensis L. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Veratrum album L. 1 Vicia cracca L. 1 Asteraceae indéterminée + Carduus sp. + Crepis sp. + Euphrasia Cf. nemorosa (Pers.) Wallr. + Gentiana sp. + Hieracium pilosella L. + Hypericum perforatum L. + Lathyrus pratensis L. + Orobanche gracilis Sm. + Prunella grandiflora (L.) Scholler + Rumex acetosa L. + Silene latifolia (P. Mill.) Britten & Rendle, non Poir. + Stellaria graminea L. + Verbascum lychnitis L. + Vicia hirsuta (L.) S.F. Gray +
Parcelle 39 Pelouse non pâturée haute.
Espèces Coefficients d’abondance
Poa sp. 2 Achillea millefolium L. 1 Anthriscus sylvestris (L.) Hoffmann 1 Armeria maritima Willd. 1 Betula pendula Roth 1 Briza media L. 1 Campanula latifolia L. 1 Campanula rotundifolia L. 1 Centaurea cyanus L. 1 Centaurea jacea L. 1 Crepis sp. 1 Dactylis glomerata L. 1 Dianthus carthusianorum L. 1 Dianthus hyssopifolius L. 1 Echium vulgare L. 1 Eryngium bourgati Gouan. 1 Euphorbia cyparissias L. 1 Galium palustre L. 1 Galium verum L. 1
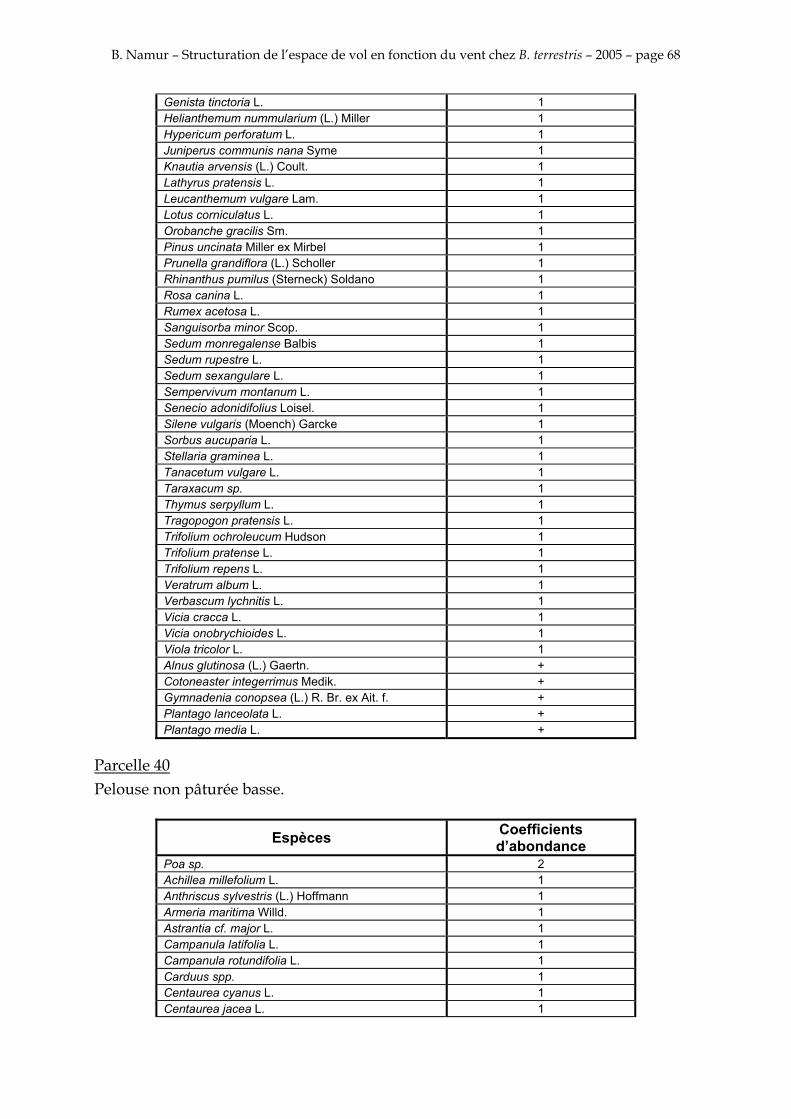
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 68
Genista tinctoria L. 1 Helianthemum nummularium (L.) Miller 1 Hypericum perforatum L. 1 Juniperus communis nana Syme 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1 Lotus corniculatus L. 1 Orobanche gracilis Sm. 1 Pinus uncinata Miller ex Mirbel 1 Prunella grandiflora (L.) Scholler 1 Rhinanthus pumilus (Sterneck) Soldano 1 Rosa canina L. 1 Rumex acetosa L. 1 Sanguisorba minor Scop. 1 Sedum monregalense Balbis 1 Sedum rupestre L. 1 Sedum sexangulare L. 1 Sempervivum montanum L. 1 Senecio adonidifolius Loisel. 1 Silene vulgaris (Moench) Garcke 1 Sorbus aucuparia L. 1 Stellaria graminea L. 1 Tanacetum vulgare L. 1 Taraxacum sp. 1 Thymus serpyllum L. 1 Tragopogon pratensis L. 1 Trifolium ochroleucum Hudson 1 Trifolium pratense L. 1 Trifolium repens L. 1 Veratrum album L. 1 Verbascum lychnitis L. 1 Vicia cracca L. 1 Vicia onobrychioides L. 1 Viola tricolor L. 1 Alnus glutinosa (L.) Gaertn. + Cotoneaster integerrimus Medik. + Gymnadenia conopsea (L.) R. Br. ex Ait. f. + Plantago lanceolata L. + Plantago media L. +
Parcelle 40 Pelouse non pâturée basse.
Espèces Coefficients d’abondance
Poa sp. 2 Achillea millefolium L. 1 Anthriscus sylvestris (L.) Hoffmann 1 Armeria maritima Willd. 1 Astrantia cf. major L. 1 Campanula latifolia L. 1 Campanula rotundifolia L. 1 Carduus spp. 1 Centaurea cyanus L. 1 Centaurea jacea L. 1

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 69
Centaurea scabiosa L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Crepis sp. 1 Dianthus carthusianorum L. 1 Dianthus hyssopifolius L. 1 Echium vulgare L. 1 Eryngium bourgati Gouan. 1 Euphorbia cyparissias L. 1 Euphrasia Cf. nemorosa (Pers.) Wallr. 1 Fraxinus excelsior L. 1 Galium palustre L. 1 Galium verum L. 1 Helianthemum nummularium (L.) Miller 1 Hieracium pilosella L. 1 Hypericum perforatum L. 1 Jasione montana L. 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1 Linaria repens (L.) Miller 1 Lolium sp. 1 Lotus corniculatus L. 1 Onopordum sp. 1 Pinus uncinata Miller ex Mirbel 1 Prunus spinosa L. 1 Rhinanthus pumilus (Sterneck) Soldano 1 Rosa canina L. 1 Rosa pimpinellifolia L. 1 Sanguisorba officinalis L. 1 Sedum rupestre L. 1 Sedum sexangulare L. 1 Sempervivum montanum L. 1 Senecio adonidifolius Loisel. 1 Stachys officinalis (L.) Trev. 1 Tanacetum vulgare L. 1 Thymus serpyllum L. 1 Tragopogon pratensis L. 1 Trifolium arvense L. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Trollius europaeus L. 1 Veratrum album L. 1 Vicia cracca L. 1 Aconitum napellus L. + Amelanchier obovalis (Michx.) Ashe + Artemisia vulgaris L. + Cf. Daucus sp. + Chamomilla recutita (L.) Rauschert + Convolvulus arvensis L. + Dactylis glomerata L. + Galeopsis tetrahit L. + Orchis sp. + Pinus sylvestris L. + Populus tremula L. + Rumex acetosa L. + Salix caprea L. + Sanguisorba minor Scop. +
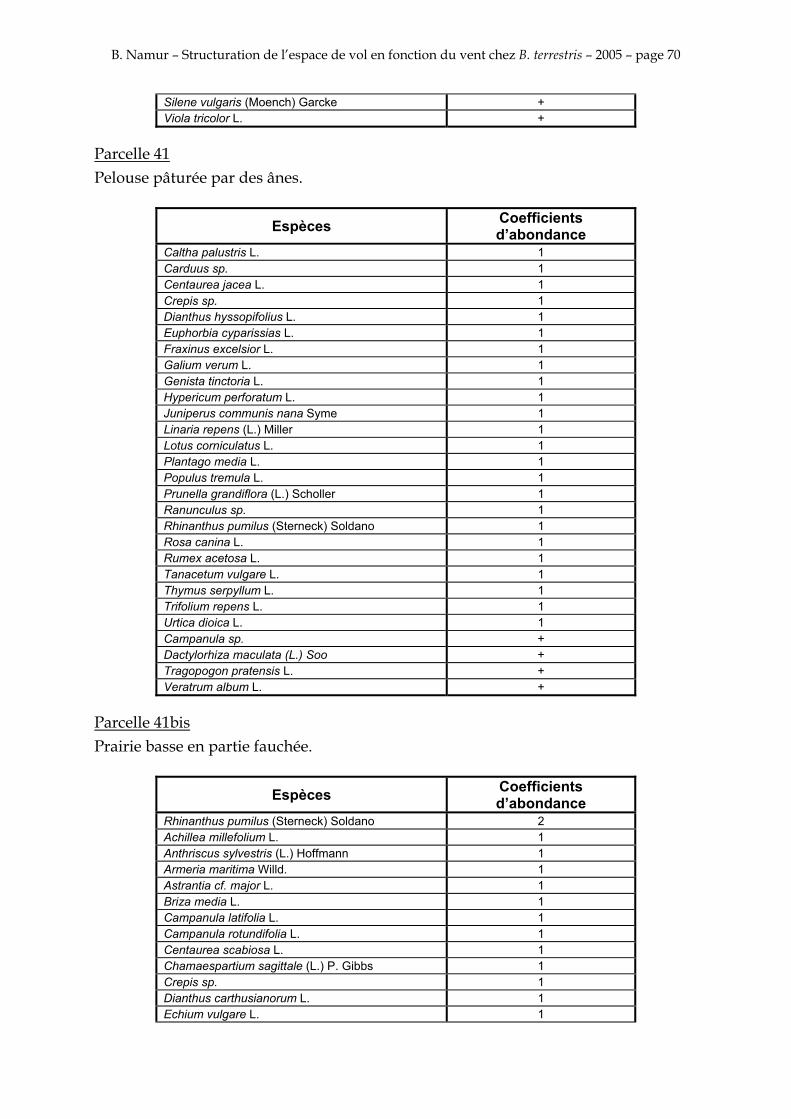
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 70
Silene vulgaris (Moench) Garcke + Viola tricolor L. +
Parcelle 41 Pelouse pâturée par des ânes.
Espèces Coefficients d’abondance
Caltha palustris L. 1 Carduus sp. 1 Centaurea jacea L. 1 Crepis sp. 1 Dianthus hyssopifolius L. 1 Euphorbia cyparissias L. 1 Fraxinus excelsior L. 1 Galium verum L. 1 Genista tinctoria L. 1 Hypericum perforatum L. 1 Juniperus communis nana Syme 1 Linaria repens (L.) Miller 1 Lotus corniculatus L. 1 Plantago media L. 1 Populus tremula L. 1 Prunella grandiflora (L.) Scholler 1 Ranunculus sp. 1 Rhinanthus pumilus (Sterneck) Soldano 1 Rosa canina L. 1 Rumex acetosa L. 1 Tanacetum vulgare L. 1 Thymus serpyllum L. 1 Trifolium repens L. 1 Urtica dioica L. 1 Campanula sp. + Dactylorhiza maculata (L.) Soo + Tragopogon pratensis L. + Veratrum album L. +
Parcelle 41bis Prairie basse en partie fauchée.
Espèces Coefficients d’abondance
Rhinanthus pumilus (Sterneck) Soldano 2 Achillea millefolium L. 1 Anthriscus sylvestris (L.) Hoffmann 1 Armeria maritima Willd. 1 Astrantia cf. major L. 1 Briza media L. 1 Campanula latifolia L. 1 Campanula rotundifolia L. 1 Centaurea scabiosa L. 1 Chamaespartium sagittale (L.) P. Gibbs 1 Crepis sp. 1 Dianthus carthusianorum L. 1 Echium vulgare L. 1
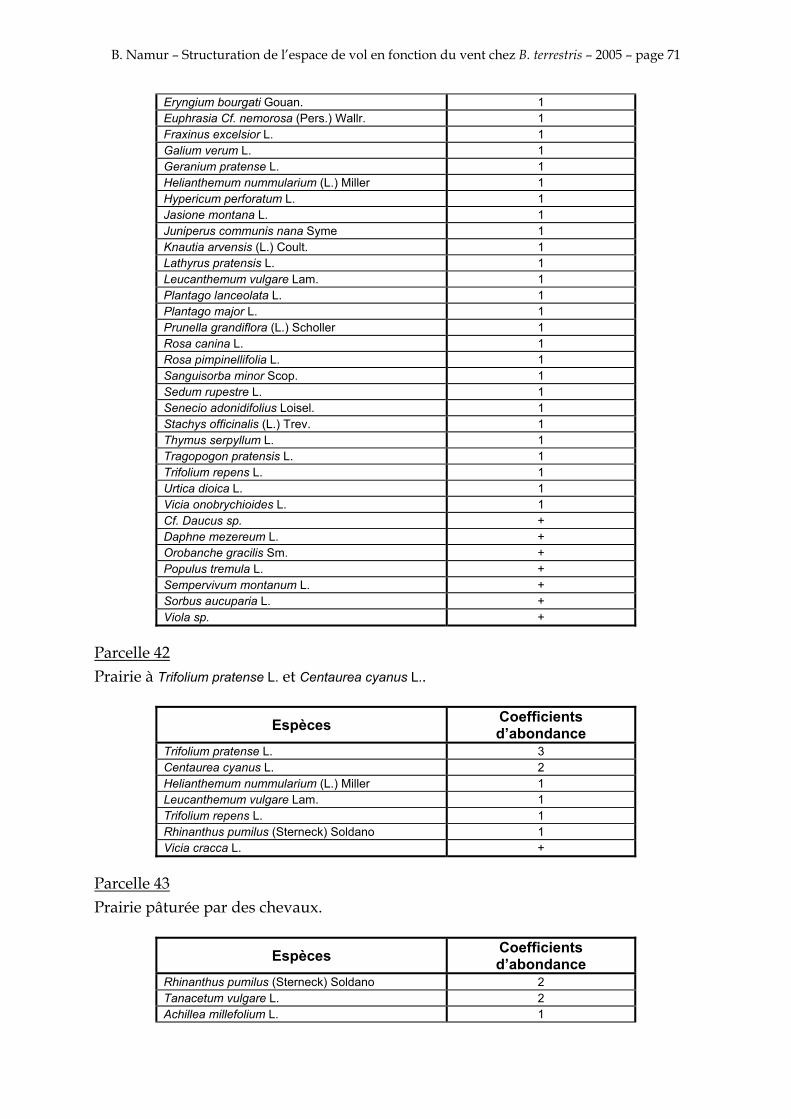
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 71
Eryngium bourgati Gouan. 1 Euphrasia Cf. nemorosa (Pers.) Wallr. 1 Fraxinus excelsior L. 1 Galium verum L. 1 Geranium pratense L. 1 Helianthemum nummularium (L.) Miller 1 Hypericum perforatum L. 1 Jasione montana L. 1 Juniperus communis nana Syme 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Leucanthemum vulgare Lam. 1 Plantago lanceolata L. 1 Plantago major L. 1 Prunella grandiflora (L.) Scholler 1 Rosa canina L. 1 Rosa pimpinellifolia L. 1 Sanguisorba minor Scop. 1 Sedum rupestre L. 1 Senecio adonidifolius Loisel. 1 Stachys officinalis (L.) Trev. 1 Thymus serpyllum L. 1 Tragopogon pratensis L. 1 Trifolium repens L. 1 Urtica dioica L. 1 Vicia onobrychioides L. 1 Cf. Daucus sp. + Daphne mezereum L. + Orobanche gracilis Sm. + Populus tremula L. + Sempervivum montanum L. + Sorbus aucuparia L. + Viola sp. +
Parcelle 42 Prairie à Trifolium pratense L. et Centaurea cyanus L..
Espèces Coefficients d’abondance
Trifolium pratense L. 3 Centaurea cyanus L. 2 Helianthemum nummularium (L.) Miller 1 Leucanthemum vulgare Lam. 1 Trifolium repens L. 1 Rhinanthus pumilus (Sterneck) Soldano 1 Vicia cracca L. +
Parcelle 43 Prairie pâturée par des chevaux.
Espèces Coefficients d’abondance
Rhinanthus pumilus (Sterneck) Soldano 2 Tanacetum vulgare L. 2 Achillea millefolium L. 1
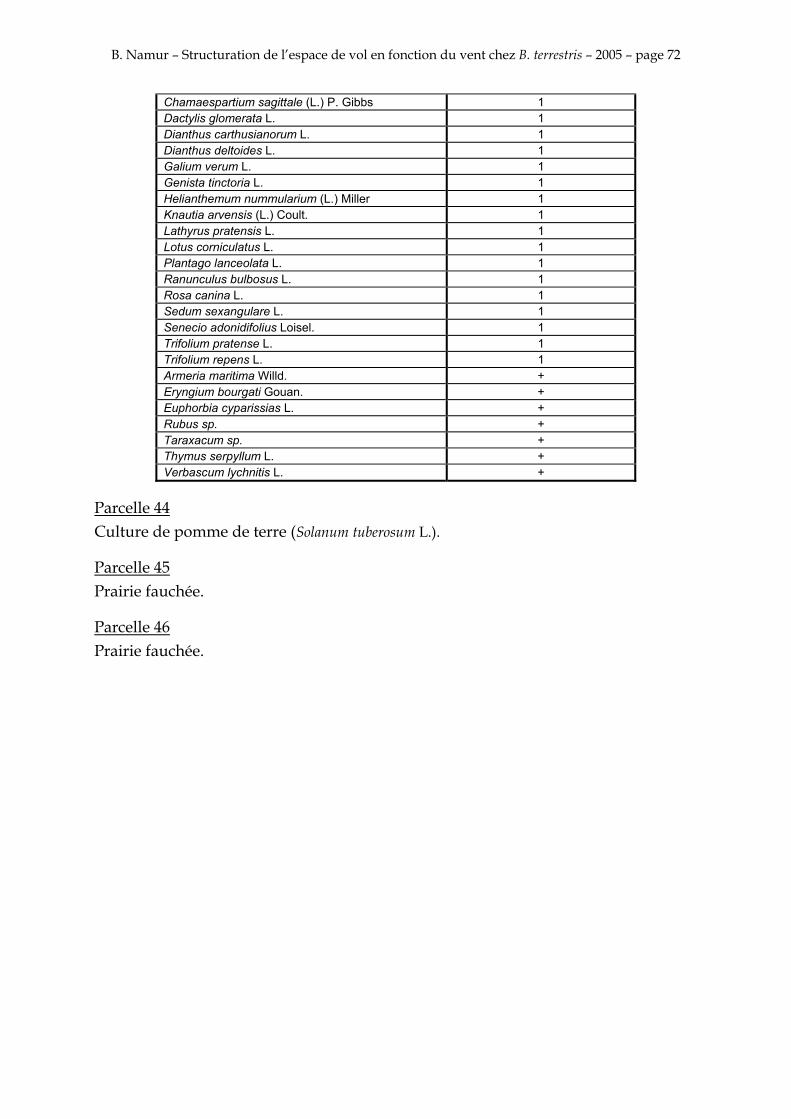
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 72
Chamaespartium sagittale (L.) P. Gibbs 1 Dactylis glomerata L. 1 Dianthus carthusianorum L. 1 Dianthus deltoides L. 1 Galium verum L. 1 Genista tinctoria L. 1 Helianthemum nummularium (L.) Miller 1 Knautia arvensis (L.) Coult. 1 Lathyrus pratensis L. 1 Lotus corniculatus L. 1 Plantago lanceolata L. 1 Ranunculus bulbosus L. 1 Rosa canina L. 1 Sedum sexangulare L. 1 Senecio adonidifolius Loisel. 1 Trifolium pratense L. 1 Trifolium repens L. 1 Armeria maritima Willd. + Eryngium bourgati Gouan. + Euphorbia cyparissias L. + Rubus sp. + Taraxacum sp. + Thymus serpyllum L. + Verbascum lychnitis L. +
Parcelle 44 Culture de pomme de terre (Solanum tuberosum L.).
Parcelle 45 Prairie fauchée.
Parcelle 46 Prairie fauchée.

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 73
11. Annexe III: Données de recaptures
n° = Référence de l’étiquette du bourdon
Alt. = Altitude
Dir. V. = Direction du vent
Vmax = Vitesse maximal du vent (m/s)
Vmoy = Vitesse moyenne du vent (m/s)
T = Température
H.A. = Humidité absolue (g/m³)
Nuag. = Couverture nuageuse (%)
Dist. = Distance de recapture (m)
Angle = Angle de recapture (par rapport au nord)
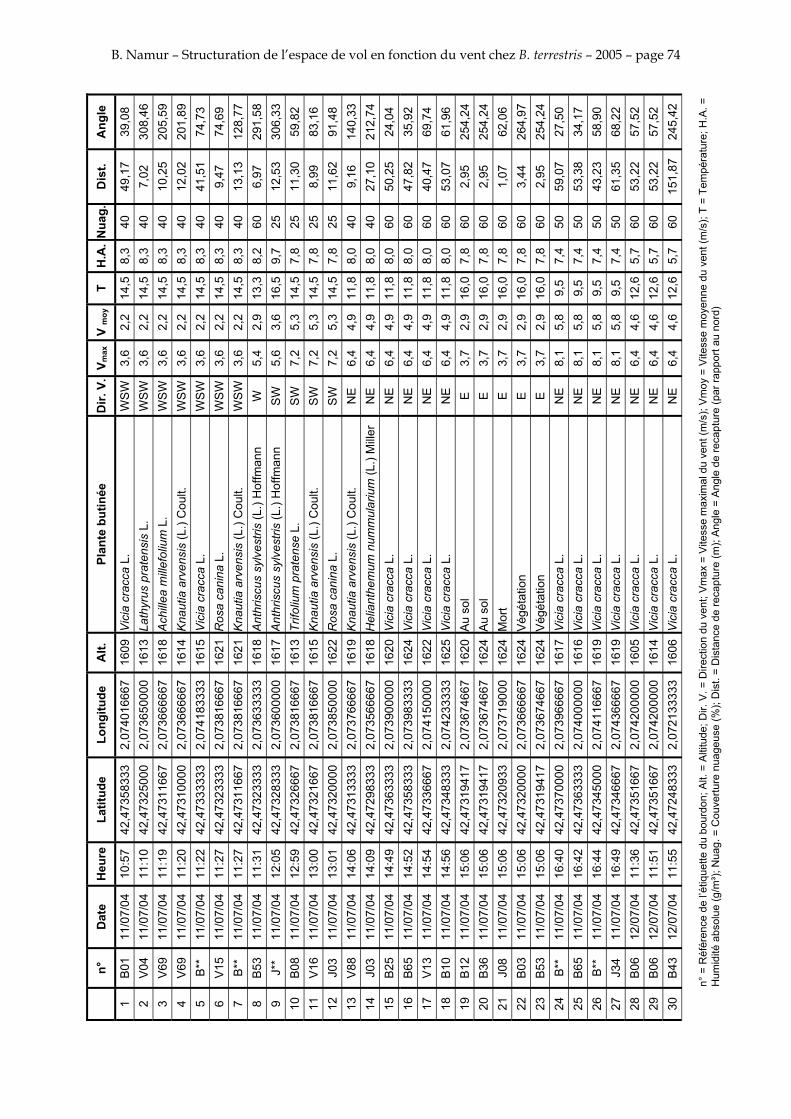
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 74
Ang
le
39,0
8 30
8,46
20
5,59
20
1,89
74
,73
74,6
9 12
8,77
29
1,58
30
6,33
59
,82
83,1
6 91
,48
140,
33
212,
74
24,0
4 35
,92
69,7
4 61
,96
254,
24
254,
24
62,0
6 26
4,97
25
4,24
27
,50
34,1
7 58
,90
68,2
2 57
,52
57,5
2 24
5,42
Dis
t.
49,1
7 7,
02
10,2
5 12
,02
41,5
1 9,
47
13,1
3 6,
97
12,5
3 11
,30
8,99
11
,62
9,16
27
,10
50,2
5 47
,82
40,4
7 53
,07
2,95
2,
95
1,07
3,
44
2,95
59
,07
53,3
8 43
,23
61,3
5 53
,22
53,2
2 15
1,87
Nua
g.
40
40
40
40
40
40
40
60
25
25
25
25
40
40
60
60
60
60
60
60
60
60
60
50
50
50
50
60
60
60
H.A
.
8,3
8,3
8,3
8,3
8,3
8,3
8,3
8,2
9,7
7,8
7,8
7,8
8,0
8,0
8,0
8,0
8,0
8,0
7,8
7,8
7,8
7,8
7,8
7,4
7,4
7,4
7,4
5,7
5,7
5,7
T
14,5
14
,5
14,5
14
,5
14,5
14
,5
14,5
13
,3
16,5
14
,5
14,5
14
,5
11,8
11
,8
11,8
11
,8
11,8
11
,8
16,0
16
,0
16,0
16
,0
16,0
9,
5 9,
5 9,
5 9,
5 12
,6
12,6
12
,6
V m
oy
2,2
2,2
2,2
2,2
2,2
2,2
2,2
2,9
3,6
5,3
5,3
5,3
4,9
4,9
4,9
4,9
4,9
4,9
2,9
2,9
2,9
2,9
2,9
5,8
5,8
5,8
5,8
4,6
4,6
4,6
V max
3,6
3,6
3,6
3,6
3,6
3,6
3,6
5,4
5,6
7,2
7,2
7,2
6,4
6,4
6,4
6,4
6,4
6,4
3,7
3,7
3,7
3,7
3,7
8,1
8,1
8,1
8,1
6,4
6,4
6,4
Dir.
V.
WS
W
WS
W
WS
W
WS
W
WS
W
WS
W
WS
W
W
SW
S
W
SW
S
W
NE
N
E
NE
N
E
NE
N
E
E
E
E
E
E
NE
N
E
NE
N
E
NE
N
E
NE
Plan
te b
utin
ée
Vic
ia c
racc
a L.
La
thyr
us p
rate
nsis
L.
Ach
illea
mill
efol
ium
L.
Kna
utia
arv
ensi
s (L
.) C
oult.
V
icia
cra
cca
L.
Ros
a ca
nina
L.
Kna
utia
arv
ensi
s (L
.) C
oult.
A
nthr
iscu
s sy
lves
tris
(L.)
Hof
fman
n A
nthr
iscu
s sy
lves
tris
(L.)
Hof
fman
n Tr
ifoliu
m p
rate
nse
L.
Kna
utia
arv
ensi
s (L
.) C
oult.
R
osa
cani
na L
. K
naut
ia a
rven
sis
(L.)
Cou
lt.
Hel
iant
hem
um n
umm
ular
ium
(L.)
Mille
r V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
A
u so
l A
u so
l M
ort
Végé
tatio
n Vé
géta
tion
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Alt.
1609
16
13
1618
16
14
1615
16
21
1621
16
18
1617
16
13
1615
16
22
1619
16
18
1620
16
24
1622
16
25
1620
16
24
1624
16
24
1624
16
17
1616
16
19
1619
16
05
1614
16
06
Long
itude
2,07
4016
667
2,07
3650
000
2,07
3666
667
2,07
3666
667
2,07
4183
333
2,07
3816
667
2,07
3816
667
2,07
3633
333
2,07
3600
000
2,07
3816
667
2,07
3816
667
2,07
3850
000
2,07
3766
667
2,07
3566
667
2,07
3900
000
2,07
3983
333
2,07
4150
000
2,07
4233
333
2,07
3674
667
2,07
3674
667
2,07
3719
000
2,07
3666
667
2,07
3674
667
2,07
3966
667
2,07
4000
000
2,07
4116
667
2,07
4366
667
2,07
4200
000
2,07
4200
000
2,07
2133
333
Latit
ude
42,4
7358
333
42,4
7325
000
42,4
7311
667
42,4
7310
000
42,4
7333
333
42,4
7323
333
42,4
7311
667
42,4
7323
333
42,4
7328
333
42,4
7326
667
42,4
7321
667
42,4
7320
000
42,4
7313
333
42,4
7298
333
42,4
7363
333
42,4
7358
333
42,4
7336
667
42,4
7348
333
42,4
7319
417
42,4
7319
417
42,4
7320
933
42,4
7320
000
42,4
7319
417
42,4
7370
000
42,4
7363
333
42,4
7345
000
42,4
7346
667
42,4
7351
667
42,4
7351
667
42,4
7248
333
Heu
re
10:5
7 11
:10
11:1
9 11
:20
11:2
2 11
:27
11:2
7 11
:31
12:0
5 12
:59
13:0
0 13
:01
14:0
6 14
:09
14:4
9 14
:52
14:5
4 14
:56
15:0
6 15
:06
15:0
6 15
:06
15:0
6 16
:40
16:4
2 16
:44
16:4
9 11
:36
11:5
1 11
:55
Dat
e
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
11
/07/
04
11/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
n°
B01
V04
V69
V69
B**
V15
B**
B53
J**
B08
V16
J03
V88
J03
B25
B65
V13
B10
B12
B36
J08
B03
B53
B**
B65
B**
J34
B06
B06
B43
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
1 2 3 4 5 6 7 8 9 10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
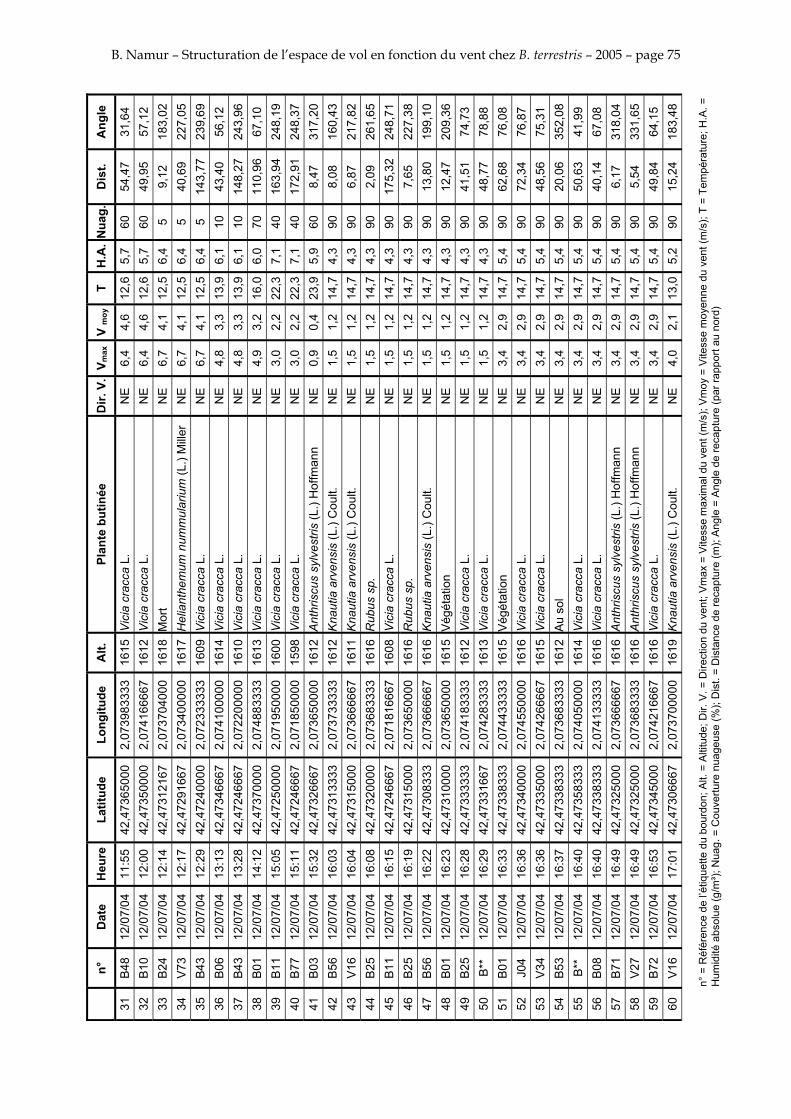
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 75
Ang
le
31,6
4 57
,12
183,
02
227,
05
239,
69
56,1
2 24
3,96
67
,10
248,
19
248,
37
317,
20
160,
43
217,
82
261,
65
248,
71
227,
38
199,
10
209,
36
74,7
3 78
,88
76,0
8 76
,87
75,3
1 35
2,08
41
,99
67,0
8 31
8,04
33
1,65
64
,15
183,
48
Dis
t.
54,4
7 49
,95
9,12
40
,69
143,
77
43,4
0 14
8,27
11
0,96
16
3,94
17
2,91
8,
47
8,08
6,
87
2,09
17
5,32
7,
65
13,8
0 12
,47
41,5
1 48
,77
62,6
8 72
,34
48,5
6 20
,06
50,6
3 40
,14
6,17
5,
54
49,8
4 15
,24
Nua
g.
60
60
5 5 5 10
10
70
40
40
60
90
90
90
90
90
90
90
90
90
90
90
90
90
90
90
90
90
90
90
H.A
.
5,7
5,7
6,4
6,4
6,4
6,1
6,1
6,0
7,1
7,1
5,9
4,3
4,3
4,3
4,3
4,3
4,3
4,3
4,3
4,3
5,4
5,4
5,4
5,4
5,4
5,4
5,4
5,4
5,4
5,2
T
12,6
12
,6
12,5
12
,5
12,5
13
,9
13,9
16
,0
22,3
22
,3
23,9
14
,7
14,7
14
,7
14,7
14
,7
14,7
14
,7
14,7
14
,7
14,7
14
,7
14,7
14
,7
14,7
14
,7
14,7
14
,7
14,7
13
,0
V m
oy
4,6
4,6
4,1
4,1
4,1
3,3
3,3
3,2
2,2
2,2
0,4
1,2
1,2
1,2
1,2
1,2
1,2
1,2
1,2
1,2
2,9
2,9
2,9
2,9
2,9
2,9
2,9
2,9
2,9
2,1
V max
6,4
6,4
6,7
6,7
6,7
4,8
4,8
4,9
3,0
3,0
0,9
1,5
1,5
1,5
1,5
1,5
1,5
1,5
1,5
1,5
3,4
3,4
3,4
3,4
3,4
3,4
3,4
3,4
3,4
4,0
Dir.
V.
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
Plan
te b
utin
ée
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Mor
t H
elia
nthe
mum
num
mul
ariu
m (L
.) M
iller
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Ant
hris
cus
sylv
estri
s (L
.) H
offm
ann
Kna
utia
arv
ensi
s (L
.) C
oult.
K
naut
ia a
rven
sis
(L.)
Cou
lt.
Rub
us s
p.
Vic
ia c
racc
a L.
R
ubus
sp.
K
naut
ia a
rven
sis
(L.)
Cou
lt.
Végé
tatio
n V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Vé
géta
tion
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Au
sol
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Ant
hris
cus
sylv
estri
s (L
.) H
offm
ann
Ant
hris
cus
sylv
estri
s (L
.) H
offm
ann
Vic
ia c
racc
a L.
K
naut
ia a
rven
sis
(L.)
Cou
lt.
Alt.
1615
16
12
1618
16
17
1609
16
14
1610
16
13
1600
15
98
1612
16
12
1611
16
16
1608
16
16
1616
16
15
1612
16
13
1615
16
16
1615
16
12
1614
16
16
1616
16
16
1616
16
19
Long
itude
2,07
3983
333
2,07
4166
667
2,07
3704
000
2,07
3400
000
2,07
2333
333
2,07
4100
000
2,07
2200
000
2,07
4883
333
2,07
1950
000
2,07
1850
000
2,07
3650
000
2,07
3733
333
2,07
3666
667
2,07
3683
333
2,07
1816
667
2,07
3650
000
2,07
3666
667
2,07
3650
000
2,07
4183
333
2,07
4283
333
2,07
4433
333
2,07
4550
000
2,07
4266
667
2,07
3683
333
2,07
4050
000
2,07
4133
333
2,07
3666
667
2,07
3683
333
2,07
4216
667
2,07
3700
000
Latit
ude
42,4
7365
000
42,4
7350
000
42,4
7312
167
42,4
7291
667
42,4
7240
000
42,4
7346
667
42,4
7246
667
42,4
7370
000
42,4
7250
000
42,4
7246
667
42,4
7326
667
42,4
7313
333
42,4
7315
000
42,4
7320
000
42,4
7246
667
42,4
7315
000
42,4
7308
333
42,4
7310
000
42,4
7333
333
42,4
7331
667
42,4
7338
333
42,4
7340
000
42,4
7335
000
42,4
7338
333
42,4
7358
333
42,4
7338
333
42,4
7325
000
42,4
7325
000
42,4
7345
000
42,4
7306
667
Heu
re
11:5
5 12
:00
12:1
4 12
:17
12:2
9 13
:13
13:2
8 14
:12
15:0
5 15
:11
15:3
2 16
:03
16:0
4 16
:08
16:1
5 16
:19
16:2
2 16
:23
16:2
8 16
:29
16:3
3 16
:36
16:3
6 16
:37
16:4
0 16
:40
16:4
9 16
:49
16:5
3 17
:01
Dat
e
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
n°
B48
B10
B24
V73
B43
B06
B43
B01
B11
B77
B03
B56
V16
B25
B11
B25
B56
B01
B25
B**
B01
J04
V34
B53
B**
B08
B71
V27
B72
V16
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
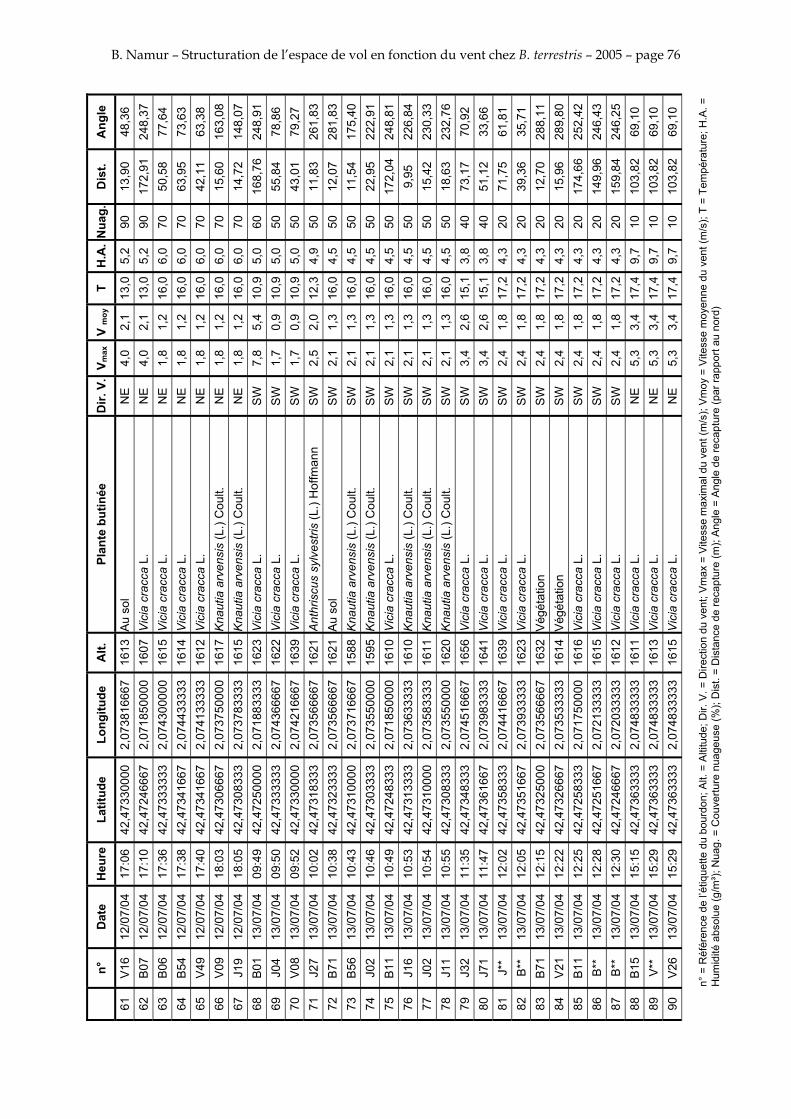
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 76
Ang
le
48,3
6 24
8,37
77
,64
73,6
3 63
,38
163,
08
148,
07
248,
91
78,8
6 79
,27
261,
83
281,
83
175,
40
222,
91
248,
81
226,
84
230,
33
232,
76
70,9
2 33
,66
61,8
1 35
,71
288,
11
289,
80
252,
42
246,
43
246,
25
69,1
0 69
,10
69,1
0
Dis
t.
13,9
0 17
2,91
50
,58
63,9
5 42
,11
15,6
0 14
,72
168,
76
55,8
4 43
,01
11,8
3 12
,07
11,5
4 22
,95
172,
04
9,95
15
,42
18,6
3 73
,17
51,1
2 71
,75
39,3
6 12
,70
15,9
6 17
4,66
14
9,96
15
9,84
10
3,82
10
3,82
10
3,82
Nua
g.
90
90
70
70
70
70
70
60
50
50
50
50
50
50
50
50
50
50
40
40
20
20
20
20
20
20
20
10
10
10
H.A
.
5,2
5,2
6,0
6,0
6,0
6,0
6,0
5,0
5,0
5,0
4,9
4,5
4,5
4,5
4,5
4,5
4,5
4,5
3,8
3,8
4,3
4,3
4,3
4,3
4,3
4,3
4,3
9,7
9,7
9,7
T
13,0
13
,0
16,0
16
,0
16,0
16
,0
16,0
10
,9
10,9
10
,9
12,3
16
,0
16,0
16
,0
16,0
16
,0
16,0
16
,0
15,1
15
,1
17,2
17
,2
17,2
17
,2
17,2
17
,2
17,2
17
,4
17,4
17
,4
V m
oy
2,1
2,1
1,2
1,2
1,2
1,2
1,2
5,4
0,9
0,9
2,0
1,3
1,3
1,3
1,3
1,3
1,3
1,3
2,6
2,6
1,8
1,8
1,8
1,8
1,8
1,8
1,8
3,4
3,4
3,4
V max
4,0
4,0
1,8
1,8
1,8
1,8
1,8
7,8
1,7
1,7
2,5
2,1
2,1
2,1
2,1
2,1
2,1
2,1
3,4
3,4
2,4
2,4
2,4
2,4
2,4
2,4
2,4
5,3
5,3
5,3
Dir.
V.
NE
N
E
NE
N
E
NE
N
E
NE
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
N
E
NE
N
E
Plan
te b
utin
ée
Au
sol
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Kna
utia
arv
ensi
s (L
.) C
oult.
K
naut
ia a
rven
sis
(L.)
Cou
lt.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
A
nthr
iscu
s sy
lves
tris
(L.)
Hof
fman
n A
u so
l K
naut
ia a
rven
sis
(L.)
Cou
lt.
Kna
utia
arv
ensi
s (L
.) C
oult.
V
icia
cra
cca
L.
Kna
utia
arv
ensi
s (L
.) C
oult.
K
naut
ia a
rven
sis
(L.)
Cou
lt.
Kna
utia
arv
ensi
s (L
.) C
oult.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Vé
géta
tion
Végé
tatio
n V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Alt.
1613
16
07
1615
16
14
1612
16
17
1615
16
23
1622
16
39
1621
16
21
1588
15
95
1610
16
10
1611
16
20
1656
16
41
1639
16
23
1632
16
14
1616
16
15
1612
16
11
1613
16
15
Long
itude
2,07
3816
667
2,07
1850
000
2,07
4300
000
2,07
4433
333
2,07
4133
333
2,07
3750
000
2,07
3783
333
2,07
1883
333
2,07
4366
667
2,07
4216
667
2,07
3566
667
2,07
3566
667
2,07
3716
667
2,07
3550
000
2,07
1850
000
2,07
3633
333
2,07
3583
333
2,07
3550
000
2,07
4516
667
2,07
3983
333
2,07
4416
667
2,07
3933
333
2,07
3566
667
2,07
3533
333
2,07
1750
000
2,07
2133
333
2,07
2033
333
2,07
4833
333
2,07
4833
333
2,07
4833
333
Latit
ude
42,4
7330
000
42,4
7246
667
42,4
7333
333
42,4
7341
667
42,4
7341
667
42,4
7306
667
42,4
7308
333
42,4
7250
000
42,4
7333
333
42,4
7330
000
42,4
7318
333
42,4
7323
333
42,4
7310
000
42,4
7303
333
42,4
7248
333
42,4
7313
333
42,4
7310
000
42,4
7308
333
42,4
7348
333
42,4
7361
667
42,4
7358
333
42,4
7351
667
42,4
7325
000
42,4
7326
667
42,4
7258
333
42,4
7251
667
42,4
7246
667
42,4
7363
333
42,4
7363
333
42,4
7363
333
Heu
re
17:0
6 17
:10
17:3
6 17
:38
17:4
0 18
:03
18:0
5 09
:49
09:5
0 09
:52
10:0
2 10
:38
10:4
3 10
:46
10:4
9 10
:53
10:5
4 10
:55
11:3
5 11
:47
12:0
2 12
:05
12:1
5 12
:22
12:2
5 12
:28
12:3
0 15
:15
15:2
9 15
:29
Dat
e
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
12
/07/
04
12/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
n°
V16
B07
B06
B54
V49
V09
J19
B01
J04
V08
J27
B71
B56
J02
B11
J16
J02
J11
J32
J71
J**
B**
B71
V21
B11
B**
B**
B15
V**
V26
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
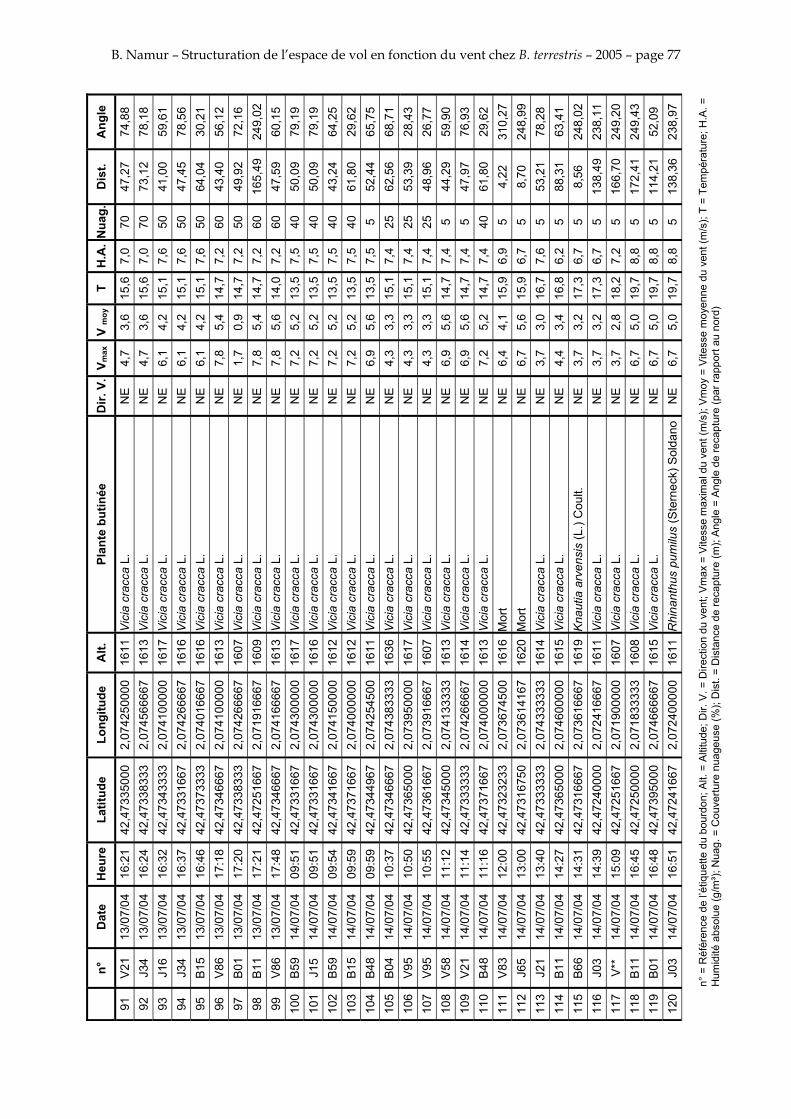
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 77
Ang
le
74,8
8 78
,18
59,6
1 78
,56
30,2
1 56
,12
72,1
6 24
9,02
60
,15
79,1
9 79
,19
64,2
5 29
,62
65,7
5 68
,71
28,4
3 26
,77
59,9
0 76
,93
29,6
2 31
0,27
24
8,99
78
,28
63,4
1 24
8,02
23
8,11
24
9,20
24
9,43
52
,09
238,
97
Dis
t.
47,2
7 73
,12
41,0
0 47
,45
64,0
4 43
,40
49,9
2 16
5,49
47
,59
50,0
9 50
,09
43,2
4 61
,80
52,4
4 62
,56
53,3
9 48
,96
44,2
9 47
,97
61,8
0 4,
22
8,70
53
,21
88,3
1 8,
56
138,
49
166,
70
172,
41
114,
21
138,
36
Nua
g.
70
70
50
50
50
60
50
60
60
40
40
40
40
5 25
25
25
5 5 40
5 5 5 5 5 5 5 5 5 5
H.A
.
7,0
7,0
7,6
7,6
7,6
7,2
7,2
7,2
7,2
7,5
7,5
7,5
7,5
7,5
7,4
7,4
7,4
7,4
7,4
7,4
6,9
6,7
7,6
6,2
6,7
6,7
7,2
8,8
8,8
8,8
T
15,6
15
,6
15,1
15
,1
15,1
14
,7
14,7
14
,7
14,0
13
,5
13,5
13
,5
13,5
13
,5
15,1
15
,1
15,1
14
,7
14,7
14
,7
15,9
15
,9
16,7
16
,8
17,3
17
,3
18,2
19
,7
19,7
19
,7
V m
oy
3,6
3,6
4,2
4,2
4,2
5,4
0,9
5,4
5,6
5,2
5,2
5,2
5,2
5,6
3,3
3,3
3,3
5,6
5,6
5,2
4,1
5,6
3,0
3,4
3,2
3,2
2,8
5,0
5,0
5,0
V max
4,7
4,7
6,1
6,1
6,1
7,8
1,7
7,8
7,8
7,2
7,2
7,2
7,2
6,9
4,3
4,3
4,3
6,9
6,9
7,2
6,4
6,7
3,7
4,4
3,7
3,7
3,7
6,7
6,7
6,7
Dir.
V.
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
Plan
te b
utin
ée
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Mor
t M
ort
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Kna
utia
arv
ensi
s (L
.) C
oult.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
R
hina
nthu
s pu
milu
s (S
tern
eck)
Sol
dano
Alt.
1611
16
13
1617
16
16
1616
16
13
1607
16
09
1613
16
17
1616
16
12
1612
16
11
1636
16
17
1607
16
13
1614
16
13
1616
16
20
1614
16
15
1619
16
11
1607
16
08
1615
16
11
Long
itude
2,07
4250
000
2,07
4566
667
2,07
4100
000
2,07
4266
667
2,07
4016
667
2,07
4100
000
2,07
4266
667
2,07
1916
667
2,07
4166
667
2,07
4300
000
2,07
4300
000
2,07
4150
000
2,07
4000
000
2,07
4254
500
2,07
4383
333
2,07
3950
000
2,07
3916
667
2,07
4133
333
2,07
4266
667
2,07
4000
000
2,07
3674
500
2,07
3614
167
2,07
4333
333
2,07
4600
000
2,07
3616
667
2,07
2416
667
2,07
1900
000
2,07
1833
333
2,07
4666
667
2,07
2400
000
Latit
ude
42,4
7335
000
42,4
7338
333
42,4
7343
333
42,4
7331
667
42,4
7373
333
42,4
7346
667
42,4
7338
333
42,4
7251
667
42,4
7346
667
42,4
7331
667
42,4
7331
667
42,4
7341
667
42,4
7371
667
42,4
7344
967
42,4
7346
667
42,4
7365
000
42,4
7361
667
42,4
7345
000
42,4
7333
333
42,4
7371
667
42,4
7323
233
42,4
7316
750
42,4
7333
333
42,4
7365
000
42,4
7316
667
42,4
7240
000
42,4
7251
667
42,4
7250
000
42,4
7395
000
42,4
7241
667
Heu
re
16:2
1 16
:24
16:3
2 16
:37
16:4
6 17
:18
17:2
0 17
:21
17:4
8 09
:51
09:5
1 09
:54
09:5
9 09
:59
10:3
7 10
:50
10:5
5 11
:12
11:1
4 11
:16
12:0
0 13
:00
13:4
0 14
:27
14:3
1 14
:39
15:0
9 16
:45
16:4
8 16
:51
Dat
e
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
13
/07/
04
13/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
n°
V21
J34
J16
J34
B15
V86
B01
B11
V86
B59
J15
B59
B15
B48
B04
V95
V95
V58
V21
B48
V83
J65
J21
B11
B66
J03
V**
B11
B01
J03
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
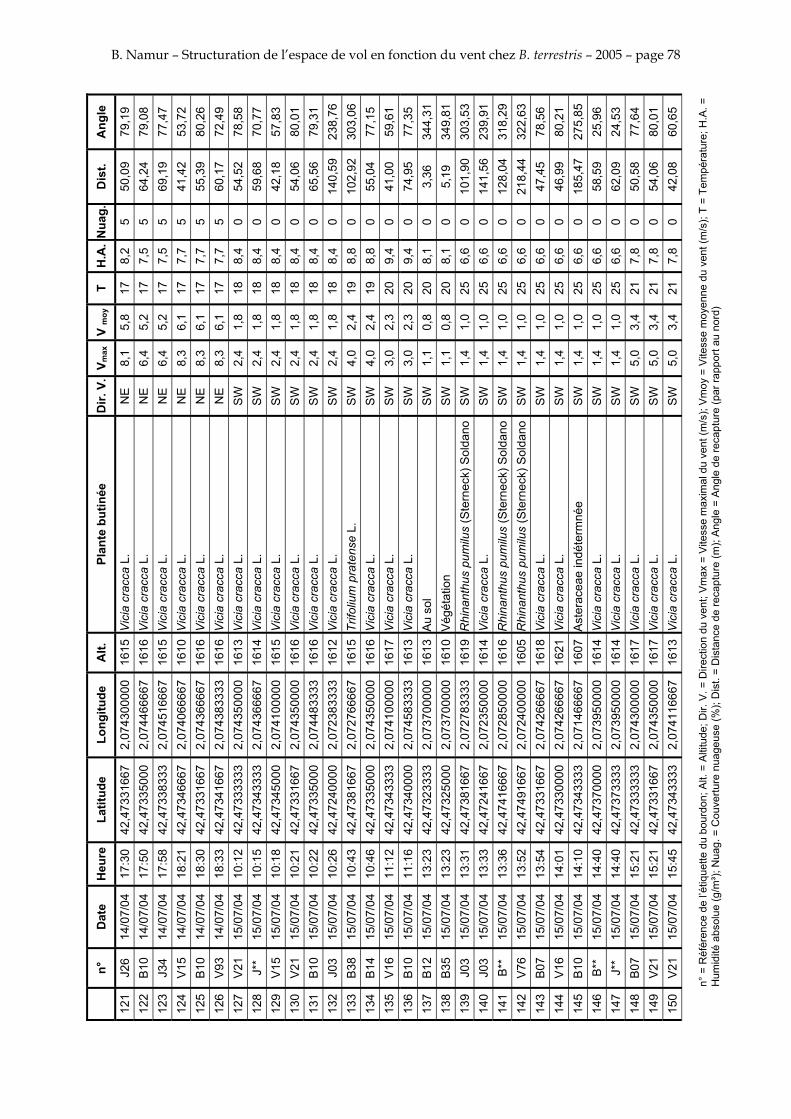
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 78
Ang
le
79,1
9 79
,08
77,4
7 53
,72
80,2
6 72
,49
78,5
8 70
,77
57,8
3 80
,01
79,3
1 23
8,76
30
3,06
77
,15
59,6
1 77
,35
344,
31
349,
81
303,
53
239,
91
318,
29
322,
63
78,5
6 80
,21
275,
85
25,9
6 24
,53
77,6
4 80
,01
60,6
5
Dis
t.
50,0
9 64
,24
69,1
9 41
,42
55,3
9 60
,17
54,5
2 59
,68
42,1
8 54
,06
65,5
6 14
0,59
10
2,92
55
,04
41,0
0 74
,95
3,36
5,
19
101,
90
141,
56
128,
04
218,
44
47,4
5 46
,99
185,
47
58,5
9 62
,09
50,5
8 54
,06
42,0
8
Nua
g.
5 5 5 5 5 5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
H.A
.
8,2
7,5
7,5
7,7
7,7
7,7
8,4
8,4
8,4
8,4
8,4
8,4
8,8
8,8
9,4
9,4
8,1
8,1
6,6
6,6
6,6
6,6
6,6
6,6
6,6
6,6
6,6
7,8
7,8
7,8
T 17
17
17
17
17
17
18
18
18
18
18
18
19
19
20
20
20
20
25
25
25
25
25
25
25
25
25
21
21
21
V m
oy
5,8
5,2
5,2
6,1
6,1
6,1
1,8
1,8
1,8
1,8
1,8
1,8
2,4
2,4
2,3
2,3
0,8
0,8
1,0
1,0
1,0
1,0
1,0
1,0
1,0
1,0
1,0
3,4
3,4
3,4
V max
8,1
6,4
6,4
8,3
8,3
8,3
2,4
2,4
2,4
2,4
2,4
2,4
4,0
4,0
3,0
3,0
1,1
1,1
1,4
1,4
1,4
1,4
1,4
1,4
1,4
1,4
1,4
5,0
5,0
5,0
Dir.
V.
NE
N
E
NE
N
E
NE
N
E
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
Plan
te b
utin
ée
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Trifo
lium
pra
tens
e L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Au
sol
Végé
tatio
n R
hina
nthu
s pu
milu
s (S
tern
eck)
Sol
dano
V
icia
cra
cca
L.
Rhi
nant
hus
pum
ilus
(Ste
rnec
k) S
olda
no
Rhi
nant
hus
pum
ilus
(Ste
rnec
k) S
olda
no
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Ast
erac
eae
indé
term
née
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Alt.
1615
16
16
1615
16
10
1616
16
16
1613
16
14
1615
16
16
1616
16
12
1615
16
16
1617
16
13
1613
16
10
1619
16
14
1616
16
05
1618
16
21
1607
16
14
1614
16
17
1617
16
13
Long
itude
2,07
4300
000
2,07
4466
667
2,07
4516
667
2,07
4066
667
2,07
4366
667
2,07
4383
333
2,07
4350
000
2,07
4366
667
2,07
4100
000
2,07
4350
000
2,07
4483
333
2,07
2383
333
2,07
2766
667
2,07
4350
000
2,07
4100
000
2,07
4583
333
2,07
3700
000
2,07
3700
000
2,07
2783
333
2,07
2350
000
2,07
2850
000
2,07
2400
000
2,07
4266
667
2,07
4266
667
2,07
1466
667
2,07
3950
000
2,07
3950
000
2,07
4300
000
2,07
4350
000
2,07
4116
667
Latit
ude
42,4
7331
667
42,4
7335
000
42,4
7338
333
42,4
7346
667
42,4
7331
667
42,4
7341
667
42,4
7333
333
42,4
7343
333
42,4
7345
000
42,4
7331
667
42,4
7335
000
42,4
7240
000
42,4
7381
667
42,4
7335
000
42,4
7343
333
42,4
7340
000
42,4
7323
333
42,4
7325
000
42,4
7381
667
42,4
7241
667
42,4
7416
667
42,4
7491
667
42,4
7331
667
42,4
7330
000
42,4
7343
333
42,4
7370
000
42,4
7373
333
42,4
7333
333
42,4
7331
667
42,4
7343
333
Heu
re
17:3
0 17
:50
17:5
8 18
:21
18:3
0 18
:33
10:1
2 10
:15
10:1
8 10
:21
10:2
2 10
:26
10:4
3 10
:46
11:1
2 11
:16
13:2
3 13
:23
13:3
1 13
:33
13:3
6 13
:52
13:5
4 14
:01
14:1
0 14
:40
14:4
0 15
:21
15:2
1 15
:45
Dat
e
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
14/0
7/04
14
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
n°
J26
B10
J34
V15
B10
V93
V21
J**
V15
V21
B10
J03
B38
B14
V16
B10
B12
B35
J03
J03
B**
V76
B07
V16
B10
B**
J**
B07
V21
V21
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
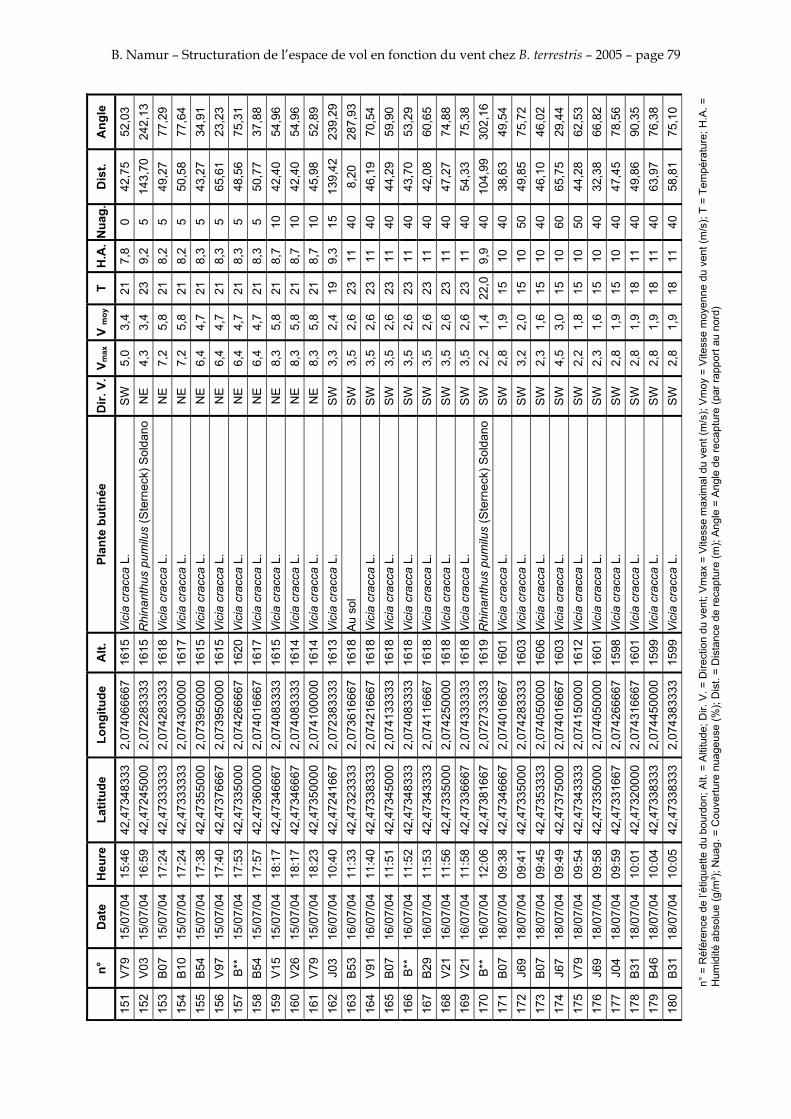
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 79
Ang
le
52,0
3 24
2,13
77
,29
77,6
4 34
,91
23,2
3 75
,31
37,8
8 54
,96
54,9
6 52
,89
239,
29
287,
93
70,5
4 59
,90
53,2
9 60
,65
74,8
8 75
,38
302,
16
49,5
4 75
,72
46,0
2 29
,44
62,5
3 66
,82
78,5
6 90
,35
76,3
8 75
,10
Dis
t.
42,7
5 14
3,70
49
,27
50,5
8 43
,27
65,6
1 48
,56
50,7
7 42
,40
42,4
0 45
,98
139,
42
8,20
46
,19
44,2
9 43
,70
42,0
8 47
,27
54,3
3 10
4,99
38
,63
49,8
5 46
,10
65,7
5 44
,28
32,3
8 47
,45
49,8
6 63
,97
58,8
1
Nua
g.
0 5 5 5 5 5 5 5 10
10
10
15
40
40
40
40
40
40
40
40
40
50
40
60
50
40
40
40
40
40
H.A
.
7,8
9,2
8,2
8,2
8,3
8,3
8,3
8,3
8,7
8,7
8,7
9,3
11
11
11
11
11
11
11
9,9
10
10
10
10
10
10
10
11
11
11
T 21
23
21
21
21
21
21
21
21
21
21
19
23
23
23
23
23
23
23
22,0
15
15
15
15
15
15
15
18
18
18
V m
oy
3,4
3,4
5,8
5,8
4,7
4,7
4,7
4,7
5,8
5,8
5,8
2,4
2,6
2,6
2,6
2,6
2,6
2,6
2,6
1,4
1,9
2,0
1,6
3,0
1,8
1,6
1,9
1,9
1,9
1,9
V max
5,0
4,3
7,2
7,2
6,4
6,4
6,4
6,4
8,3
8,3
8,3
3,3
3,5
3,5
3,5
3,5
3,5
3,5
3,5
2,2
2,8
3,2
2,3
4,5
2,2
2,3
2,8
2,8
2,8
2,8
Dir.
V.
SW
N
E
NE
N
E
NE
N
E
NE
N
E
NE
N
E
NE
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
Plan
te b
utin
ée
Vic
ia c
racc
a L.
R
hina
nthu
s pu
milu
s (S
tern
eck)
Sol
dano
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
A
u so
l V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
R
hina
nthu
s pu
milu
s (S
tern
eck)
Sol
dano
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Alt.
1615
16
15
1618
16
17
1615
16
15
1620
16
17
1615
16
14
1614
16
13
1618
16
18
1618
16
18
1618
16
18
1618
16
19
1601
16
03
1606
16
03
1612
16
01
1598
16
01
1599
15
99
Long
itude
2,07
4066
667
2,07
2283
333
2,07
4283
333
2,07
4300
000
2,07
3950
000
2,07
3950
000
2,07
4266
667
2,07
4016
667
2,07
4083
333
2,07
4083
333
2,07
4100
000
2,07
2383
333
2,07
3616
667
2,07
4216
667
2,07
4133
333
2,07
4083
333
2,07
4116
667
2,07
4250
000
2,07
4333
333
2,07
2733
333
2,07
4016
667
2,07
4283
333
2,07
4050
000
2,07
4016
667
2,07
4150
000
2,07
4050
000
2,07
4266
667
2,07
4316
667
2,07
4450
000
2,07
4383
333
Latit
ude
42,4
7348
333
42,4
7245
000
42,4
7333
333
42,4
7333
333
42,4
7355
000
42,4
7376
667
42,4
7335
000
42,4
7360
000
42,4
7346
667
42,4
7346
667
42,4
7350
000
42,4
7241
667
42,4
7323
333
42,4
7338
333
42,4
7345
000
42,4
7348
333
42,4
7343
333
42,4
7335
000
42,4
7336
667
42,4
7381
667
42,4
7346
667
42,4
7335
000
42,4
7353
333
42,4
7375
000
42,4
7343
333
42,4
7335
000
42,4
7331
667
42,4
7320
000
42,4
7338
333
42,4
7338
333
Heu
re
15:4
6 16
:59
17:2
4 17
:24
17:3
8 17
:40
17:5
3 17
:57
18:1
7 18
:17
18:2
3 10
:40
11:3
3 11
:40
11:5
1 11
:52
11:5
3 11
:56
11:5
8 12
:06
09:3
8 09
:41
09:4
5 09
:49
09:5
4 09
:58
09:5
9 10
:01
10:0
4 10
:05
Dat
e
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
15
/07/
04
15/0
7/04
16
/07/
04
16/0
7/04
16
/07/
04
16/0
7/04
16
/07/
04
16/0
7/04
16
/07/
04
16/0
7/04
16
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
n°
V79
V03
B07
B10
B54
V97
B**
B54
V15
V26
V79
J03
B53
V91
B07
B**
B29
V21
V21
B**
B07
J69
B07
J67
V79
J69
J04
B31
B46
B31
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
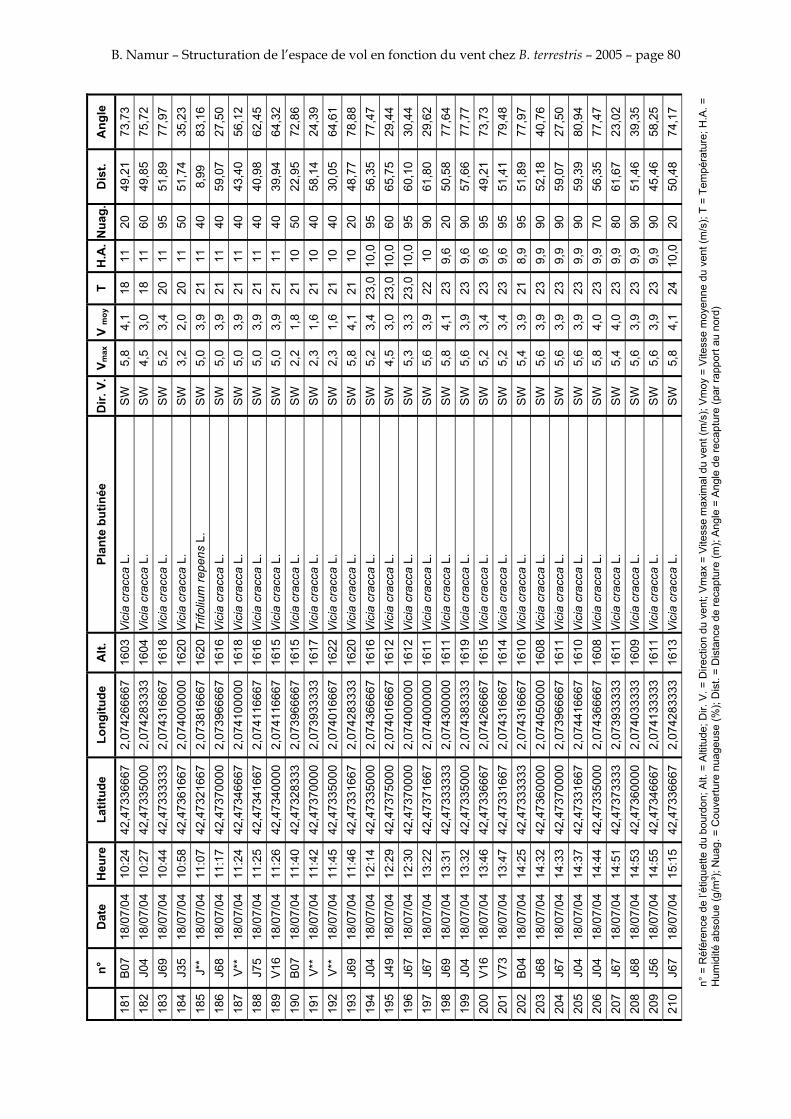
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 80
Ang
le
73,7
3 75
,72
77,9
7 35
,23
83,1
6 27
,50
56,1
2 62
,45
64,3
2 72
,86
24,3
9 64
,61
78,8
8 77
,47
29,4
4 30
,44
29,6
2 77
,64
77,7
7 73
,73
79,4
8 77
,97
40,7
6 27
,50
80,9
4 77
,47
23,0
2 39
,35
58,2
5 74
,17
Dis
t.
49,2
1 49
,85
51,8
9 51
,74
8,99
59
,07
43,4
0 40
,98
39,9
4 22
,95
58,1
4 30
,05
48,7
7 56
,35
65,7
5 60
,10
61,8
0 50
,58
57,6
6 49
,21
51,4
1 51
,89
52,1
8 59
,07
59,3
9 56
,35
61,6
7 51
,46
45,4
6 50
,48
Nua
g.
20
60
95
50
40
40
40
40
40
50
40
40
20
95
60
95
90
20
90
95
95
95
90
90
90
70
80
90
90
20
H.A
.
11
11
11
11
11
11
11
11
11
10
10
10
10
10,0
10
,0
10,0
10
9,
6 9,
6 9,
6 9,
6 8,
9 9,
9 9,
9 9,
9 9,
9 9,
9 9,
9 9,
9 10
,0
T 18
18
20
20
21
21
21
21
21
21
21
21
21
23,0
23
,0
23,0
22
23
23
23
23
21
23
23
23
23
23
23
23
24
V m
oy
4,1
3,0
3,4
2,0
3,9
3,9
3,9
3,9
3,9
1,8
1,6
1,6
4,1
3,4
3,0
3,3
3,9
4,1
3,9
3,4
3,4
3,9
3,9
3,9
3,9
4,0
4,0
3,9
3,9
4,1
V max
5,8
4,5
5,2
3,2
5,0
5,0
5,0
5,0
5,0
2,2
2,3
2,3
5,8
5,2
4,5
5,3
5,6
5,8
5,6
5,2
5,2
5,4
5,6
5,6
5,6
5,8
5,4
5,6
5,6
5,8
Dir.
V.
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
Plan
te b
utin
ée
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Trifo
lium
repe
ns L
. V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Alt.
1603
16
04
1618
16
20
1620
16
16
1618
16
16
1615
16
15
1617
16
22
1620
16
16
1612
16
12
1611
16
11
1619
16
15
1614
16
10
1608
16
11
1610
16
08
1611
16
09
1611
16
13
Long
itude
2,07
4266
667
2,07
4283
333
2,07
4316
667
2,07
4000
000
2,07
3816
667
2,07
3966
667
2,07
4100
000
2,07
4116
667
2,07
4116
667
2,07
3966
667
2,07
3933
333
2,07
4016
667
2,07
4283
333
2,07
4366
667
2,07
4016
667
2,07
4000
000
2,07
4000
000
2,07
4300
000
2,07
4383
333
2,07
4266
667
2,07
4316
667
2,07
4316
667
2,07
4050
000
2,07
3966
667
2,07
4416
667
2,07
4366
667
2,07
3933
333
2,07
4033
333
2,07
4133
333
2,07
4283
333
Latit
ude
42,4
7336
667
42,4
7335
000
42,4
7333
333
42,4
7361
667
42,4
7321
667
42,4
7370
000
42,4
7346
667
42,4
7341
667
42,4
7340
000
42,4
7328
333
42,4
7370
000
42,4
7335
000
42,4
7331
667
42,4
7335
000
42,4
7375
000
42,4
7370
000
42,4
7371
667
42,4
7333
333
42,4
7335
000
42,4
7336
667
42,4
7331
667
42,4
7333
333
42,4
7360
000
42,4
7370
000
42,4
7331
667
42,4
7335
000
42,4
7373
333
42,4
7360
000
42,4
7346
667
42,4
7336
667
Heu
re
10:2
4 10
:27
10:4
4 10
:58
11:0
7 11
:17
11:2
4 11
:25
11:2
6 11
:40
11:4
2 11
:45
11:4
6 12
:14
12:2
9 12
:30
13:2
2 13
:31
13:3
2 13
:46
13:4
7 14
:25
14:3
2 14
:33
14:3
7 14
:44
14:5
1 14
:53
14:5
5 15
:15
Dat
e
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
n°
B07
J04
J69
J35
J**
J68
V**
J75
V16
B07
V**
V**
J69
J04
J49
J67
J67
J69
J04
V16
V73
B04
J68
J67
J04
J04
J67
J68
J56
J67
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
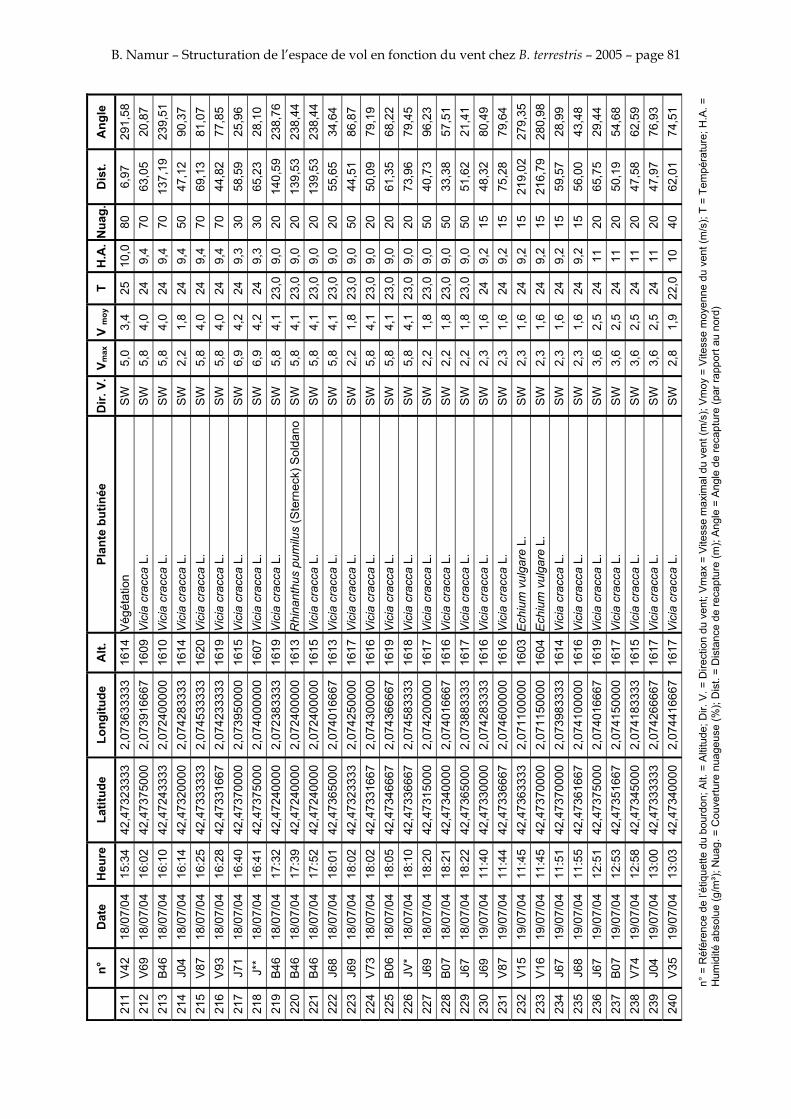
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 81
Ang
le
291,
58
20,8
7 23
9,51
90
,37
81,0
7 77
,85
25,9
6 28
,10
238,
76
238,
44
238,
44
34,6
4 86
,87
79,1
9 68
,22
79,4
5 96
,23
57,5
1 21
,41
80,4
9 79
,64
279,
35
280,
98
28,9
9 43
,48
29,4
4 54
,68
62,5
9 76
,93
74,5
1
Dis
t.
6,97
63
,05
137,
19
47,1
2 69
,13
44,8
2 58
,59
65,2
3 14
0,59
13
9,53
13
9,53
55
,65
44,5
1 50
,09
61,3
5 73
,96
40,7
3 33
,38
51,6
2 48
,32
75,2
8 21
9,02
21
6,79
59
,57
56,0
0 65
,75
50,1
9 47
,58
47,9
7 62
,01
Nua
g.
80
70
70
50
70
70
30
30
20
20
20
20
50
20
20
20
50
50
50
15
15
15
15
15
15
20
20
20
20
40
H.A
.
10,0
9,
4 9,
4 9,
4 9,
4 9,
4 9,
3 9,
3 9,
0 9,
0 9,
0 9,
0 9,
0 9,
0 9,
0 9,
0 9,
0 9,
0 9,
0 9,
2 9,
2 9,
2 9,
2 9,
2 9,
2 11
11
11
11
10
T 25
24
24
24
24
24
24
24
23,0
23
,0
23,0
23
,0
23,0
23
,0
23,0
23
,0
23,0
23
,0
23,0
24
24
24
24
24
24
24
24
24
24
22
,0
V m
oy
3,4
4,0
4,0
1,8
4,0
4,0
4,2
4,2
4,1
4,1
4,1
4,1
1,8
4,1
4,1
4,1
1,8
1,8
1,8
1,6
1,6
1,6
1,6
1,6
1,6
2,5
2,5
2,5
2,5
1,9
V max
5,0
5,8
5,8
2,2
5,8
5,8
6,9
6,9
5,8
5,8
5,8
5,8
2,2
5,8
5,8
5,8
2,2
2,2
2,2
2,3
2,3
2,3
2,3
2,3
2,3
3,6
3,6
3,6
3,6
2,8
Dir.
V.
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
Plan
te b
utin
ée
Végé
tatio
n V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
R
hina
nthu
s pu
milu
s (S
tern
eck)
Sol
dano
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Ech
ium
vul
gare
L.
Ech
ium
vul
gare
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Alt.
1614
16
09
1610
16
14
1620
16
19
1615
16
07
1619
16
13
1615
16
13
1617
16
16
1619
16
18
1617
16
16
1617
16
16
1616
16
03
1604
16
14
1616
16
19
1617
16
15
1617
16
17
Long
itude
2,07
3633
333
2,07
3916
667
2,07
2400
000
2,07
4283
333
2,07
4533
333
2,07
4233
333
2,07
3950
000
2,07
4000
000
2,07
2383
333
2,07
2400
000
2,07
2400
000
2,07
4016
667
2,07
4250
000
2,07
4300
000
2,07
4366
667
2,07
4583
333
2,07
4200
000
2,07
4016
667
2,07
3883
333
2,07
4283
333
2,07
4600
000
2,07
1100
000
2,07
1150
000
2,07
3983
333
2,07
4100
000
2,07
4016
667
2,07
4150
000
2,07
4183
333
2,07
4266
667
2,07
4416
667
Latit
ude
42,4
7323
333
42,4
7375
000
42,4
7243
333
42,4
7320
000
42,4
7333
333
42,4
7331
667
42,4
7370
000
42,4
7375
000
42,4
7240
000
42,4
7240
000
42,4
7240
000
42,4
7365
000
42,4
7323
333
42,4
7331
667
42,4
7346
667
42,4
7336
667
42,4
7315
000
42,4
7340
000
42,4
7365
000
42,4
7330
000
42,4
7336
667
42,4
7363
333
42,4
7370
000
42,4
7370
000
42,4
7361
667
42,4
7375
000
42,4
7351
667
42,4
7345
000
42,4
7333
333
42,4
7340
000
Heu
re
15:3
4 16
:02
16:1
0 16
:14
16:2
5 16
:28
16:4
0 16
:41
17:3
2 17
:39
17:5
2 18
:01
18:0
2 18
:02
18:0
5 18
:10
18:2
0 18
:21
18:2
2 11
:40
11:4
4 11
:45
11:4
5 11
:51
11:5
5 12
:51
12:5
3 12
:58
13:0
0 13
:03
Dat
e
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
18
/07/
04
18/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
n°
V42
V69
B46
J04
V87
V93
J71
J**
B46
B46
B46
J68
J69
V73
B06
JV*
J69
B07
J67
J69
V87
V15
V16
J67
J68
J67
B07
V74
J04
V35
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
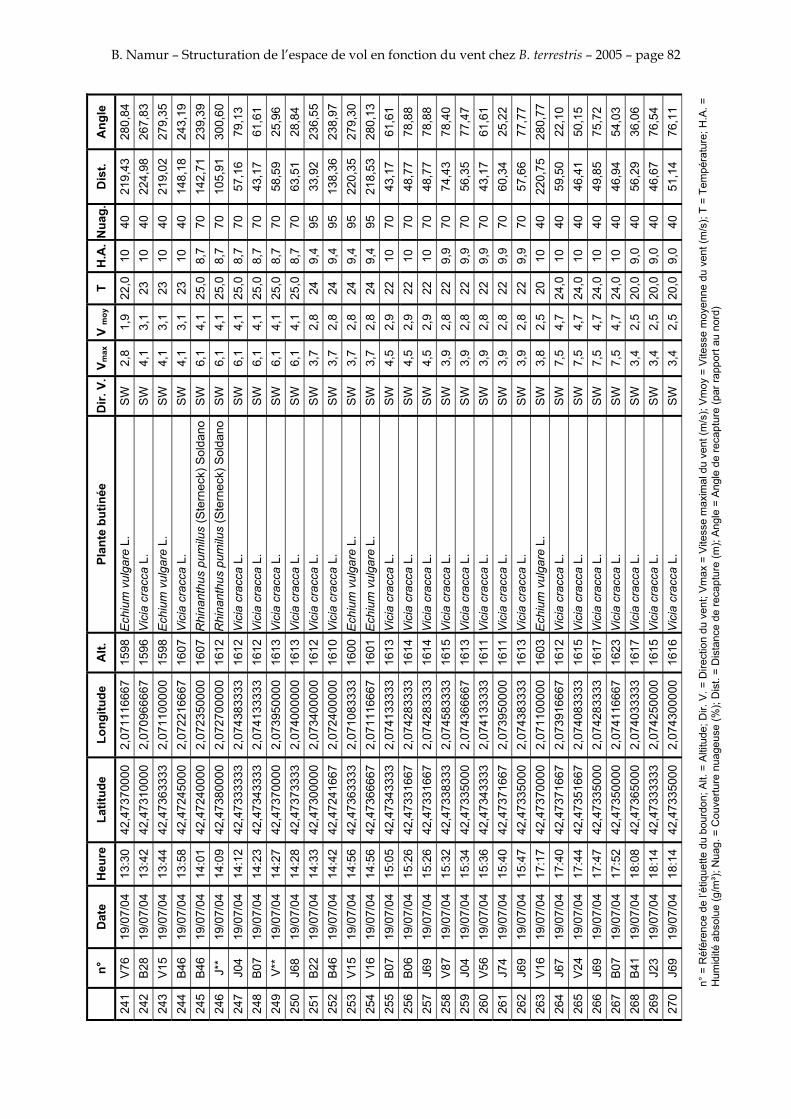
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 82
Ang
le
280,
84
267,
83
279,
35
243,
19
239,
39
300,
60
79,1
3 61
,61
25,9
6 28
,84
236,
55
238,
97
279,
30
280,
13
61,6
1 78
,88
78,8
8 78
,40
77,4
7 61
,61
25,2
2 77
,77
280,
77
22,1
0 50
,15
75,7
2 54
,03
36,0
6 76
,54
76,1
1
Dis
t.
219,
43
224,
98
219,
02
148,
18
142,
71
105,
91
57,1
6 43
,17
58,5
9 63
,51
33,9
2 13
8,36
22
0,35
21
8,53
43
,17
48,7
7 48
,77
74,4
3 56
,35
43,1
7 60
,34
57,6
6 22
0,75
59
,50
46,4
1 49
,85
46,9
4 56
,29
46,6
7 51
,14
Nua
g.
40
40
40
40
70
70
70
70
70
70
95
95
95
95
70
70
70
70
70
70
70
70
40
40
40
40
40
40
40
40
H.A
.
10
10
10
10
8,7
8,7
8,7
8,7
8,7
8,7
9,4
9,4
9,4
9,4
10
10
10
9,9
9,9
9,9
9,9
9,9
10
10
10
10
10
9,0
9,0
9,0
T
22,0
23
23
23
25
,0
25,0
25
,0
25,0
25
,0
25,0
24
24
24
24
22
22
22
22
22
22
22
22
20
24
,0
24,0
24
,0
24,0
20
,0
20,0
20
,0
V m
oy
1,9
3,1
3,1
3,1
4,1
4,1
4,1
4,1
4,1
4,1
2,8
2,8
2,8
2,8
2,9
2,9
2,9
2,8
2,8
2,8
2,8
2,8
2,5
4,7
4,7
4,7
4,7
2,5
2,5
2,5
V max
2,8
4,1
4,1
4,1
6,1
6,1
6,1
6,1
6,1
6,1
3,7
3,7
3,7
3,7
4,5
4,5
4,5
3,9
3,9
3,9
3,9
3,9
3,8
7,5
7,5
7,5
7,5
3,4
3,4
3,4
Dir.
V.
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
Plan
te b
utin
ée
Ech
ium
vul
gare
L.
Vic
ia c
racc
a L.
E
chiu
m v
ulga
re L
. V
icia
cra
cca
L.
Rhi
nant
hus
pum
ilus
(Ste
rnec
k) S
olda
no
Rhi
nant
hus
pum
ilus
(Ste
rnec
k) S
olda
no
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Ech
ium
vul
gare
L.
Ech
ium
vul
gare
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Ech
ium
vul
gare
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Alt.
1598
15
96
1598
16
07
1607
16
12
1612
16
12
1613
16
13
1612
16
10
1600
16
01
1613
16
14
1614
16
15
1613
16
11
1611
16
13
1603
16
12
1615
16
17
1623
16
17
1615
16
16
Long
itude
2,07
1116
667
2,07
0966
667
2,07
1100
000
2,07
2216
667
2,07
2350
000
2,07
2700
000
2,07
4383
333
2,07
4133
333
2,07
3950
000
2,07
4000
000
2,07
3400
000
2,07
2400
000
2,07
1083
333
2,07
1116
667
2,07
4133
333
2,07
4283
333
2,07
4283
333
2,07
4583
333
2,07
4366
667
2,07
4133
333
2,07
3950
000
2,07
4383
333
2,07
1100
000
2,07
3916
667
2,07
4083
333
2,07
4283
333
2,07
4116
667
2,07
4033
333
2,07
4250
000
2,07
4300
000
Latit
ude
42,4
7370
000
42,4
7310
000
42,4
7363
333
42,4
7245
000
42,4
7240
000
42,4
7380
000
42,4
7333
333
42,4
7343
333
42,4
7370
000
42,4
7373
333
42,4
7300
000
42,4
7241
667
42,4
7363
333
42,4
7366
667
42,4
7343
333
42,4
7331
667
42,4
7331
667
42,4
7338
333
42,4
7335
000
42,4
7343
333
42,4
7371
667
42,4
7335
000
42,4
7370
000
42,4
7371
667
42,4
7351
667
42,4
7335
000
42,4
7350
000
42,4
7365
000
42,4
7333
333
42,4
7335
000
Heu
re
13:3
0 13
:42
13:4
4 13
:58
14:0
1 14
:09
14:1
2 14
:23
14:2
7 14
:28
14:3
3 14
:42
14:5
6 14
:56
15:0
5 15
:26
15:2
6 15
:32
15:3
4 15
:36
15:4
0 15
:47
17:1
7 17
:40
17:4
4 17
:47
17:5
2 18
:08
18:1
4 18
:14
Dat
e
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
n°
V76
B28
V15
B46
B46
J**
J04
B07
V**
J68
B22
B46
V15
V16
B07
B06
J69
V87
J04
V56
J74
J69
V16
J67
V24
J69
B07
B41
J23
J69
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
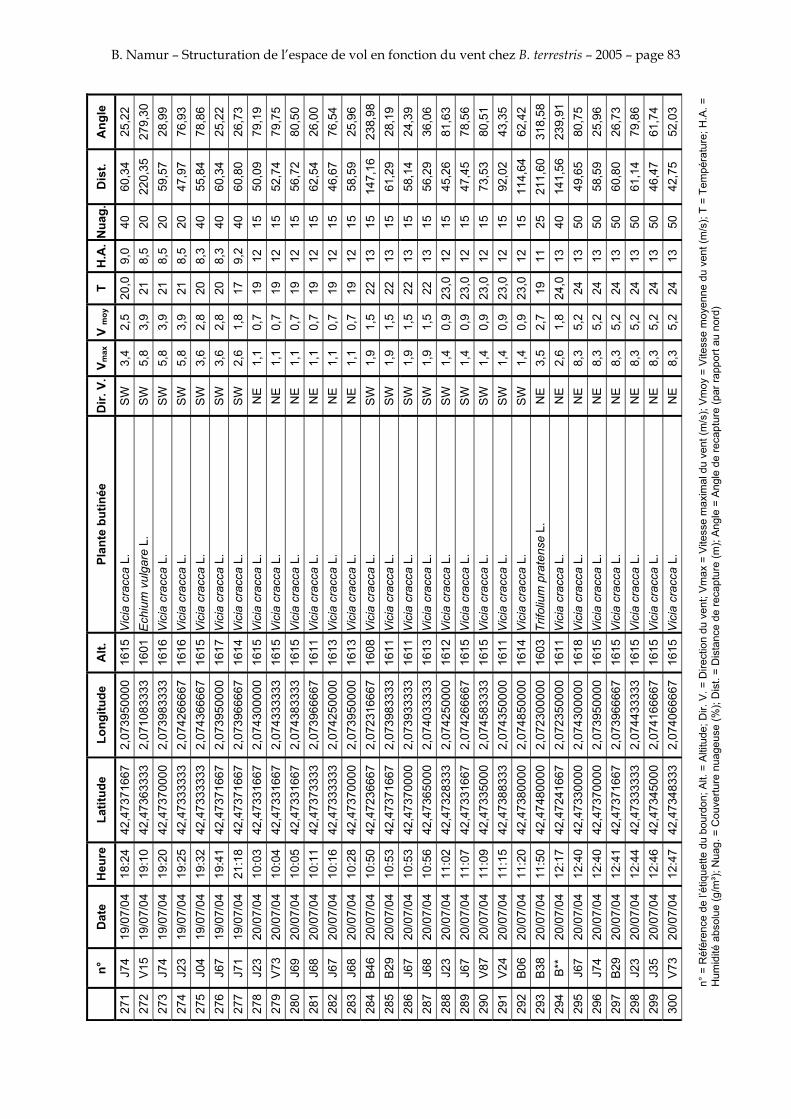
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 83
Ang
le
25,2
2 27
9,30
28
,99
76,9
3 78
,86
25,2
2 26
,73
79,1
9 79
,75
80,5
0 26
,00
76,5
4 25
,96
238,
98
28,1
9 24
,39
36,0
6 81
,63
78,5
6 80
,51
43,3
5 62
,42
318,
58
239,
91
80,7
5 25
,96
26,7
3 79
,86
61,7
4 52
,03
Dis
t.
60,3
4 22
0,35
59
,57
47,9
7 55
,84
60,3
4 60
,80
50,0
9 52
,74
56,7
2 62
,54
46,6
7 58
,59
147,
16
61,2
9 58
,14
56,2
9 45
,26
47,4
5 73
,53
92,0
2 11
4,64
21
1,60
14
1,56
49
,65
58,5
9 60
,80
61,1
4 46
,47
42,7
5
Nua
g.
40
20
20
20
40
40
40
15
15
15
15
15
15
15
15
15
15
15
15
15
15
15
25
40
50
50
50
50
50
50
H.A
.
9,0
8,5
8,5
8,5
8,3
8,3
9,2
12
12
12
12
12
12
13
13
13
13
12
12
12
12
12
11
13
13
13
13
13
13
13
T
20,0
21
21
21
20
20
17
19
19
19
19
19
19
22
22
22
22
23
,0
23,0
23
,0
23,0
23
,0
19
24,0
24
24
24
24
24
24
V m
oy
2,5
3,9
3,9
3,9
2,8
2,8
1,8
0,7
0,7
0,7
0,7
0,7
0,7
1,5
1,5
1,5
1,5
0,9
0,9
0,9
0,9
0,9
2,7
1,8
5,2
5,2
5,2
5,2
5,2
5,2
V max
3,4
5,8
5,8
5,8
3,6
3,6
2,6
1,1
1,1
1,1
1,1
1,1
1,1
1,9
1,9
1,9
1,9
1,4
1,4
1,4
1,4
1,4
3,5
2,6
8,3
8,3
8,3
8,3
8,3
8,3
Dir.
V.
SW
S
W
SW
S
W
SW
S
W
SW
N
E
NE
N
E
NE
N
E
NE
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
NE
N
E
NE
N
E
NE
N
E
NE
N
E
Plan
te b
utin
ée
Vic
ia c
racc
a L.
E
chiu
m v
ulga
re L
. V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Tr
ifoliu
m p
rate
nse
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Alt.
1615
16
01
1616
16
16
1615
16
17
1614
16
15
1615
16
15
1611
16
13
1613
16
08
1611
16
11
1613
16
12
1615
16
15
1611
16
14
1603
16
11
1618
16
15
1615
16
15
1615
16
15
Long
itude
2,07
3950
000
2,07
1083
333
2,07
3983
333
2,07
4266
667
2,07
4366
667
2,07
3950
000
2,07
3966
667
2,07
4300
000
2,07
4333
333
2,07
4383
333
2,07
3966
667
2,07
4250
000
2,07
3950
000
2,07
2316
667
2,07
3983
333
2,07
3933
333
2,07
4033
333
2,07
4250
000
2,07
4266
667
2,07
4583
333
2,07
4350
000
2,07
4850
000
2,07
2300
000
2,07
2350
000
2,07
4300
000
2,07
3950
000
2,07
3966
667
2,07
4433
333
2,07
4166
667
2,07
4066
667
Latit
ude
42,4
7371
667
42,4
7363
333
42,4
7370
000
42,4
7333
333
42,4
7333
333
42,4
7371
667
42,4
7371
667
42,4
7331
667
42,4
7331
667
42,4
7331
667
42,4
7373
333
42,4
7333
333
42,4
7370
000
42,4
7236
667
42,4
7371
667
42,4
7370
000
42,4
7365
000
42,4
7328
333
42,4
7331
667
42,4
7335
000
42,4
7388
333
42,4
7380
000
42,4
7480
000
42,4
7241
667
42,4
7330
000
42,4
7370
000
42,4
7371
667
42,4
7333
333
42,4
7345
000
42,4
7348
333
Heu
re
18:2
4 19
:10
19:2
0 19
:25
19:3
2 19
:41
21:1
8 10
:03
10:0
4 10
:05
10:1
1 10
:16
10:2
8 10
:50
10:5
3 10
:53
10:5
6 11
:02
11:0
7 11
:09
11:1
5 11
:20
11:5
0 12
:17
12:4
0 12
:40
12:4
1 12
:44
12:4
6 12
:47
Dat
e
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
19
/07/
04
19/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
n°
J74
V15
J74
J23
J04
J67
J71
J23
V73
J69
J68
J67
J68
B46
B29
J67
J68
J23
J67
V87
V24
B06
B38
B**
J67
J74
B29
J23
J35
V73
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
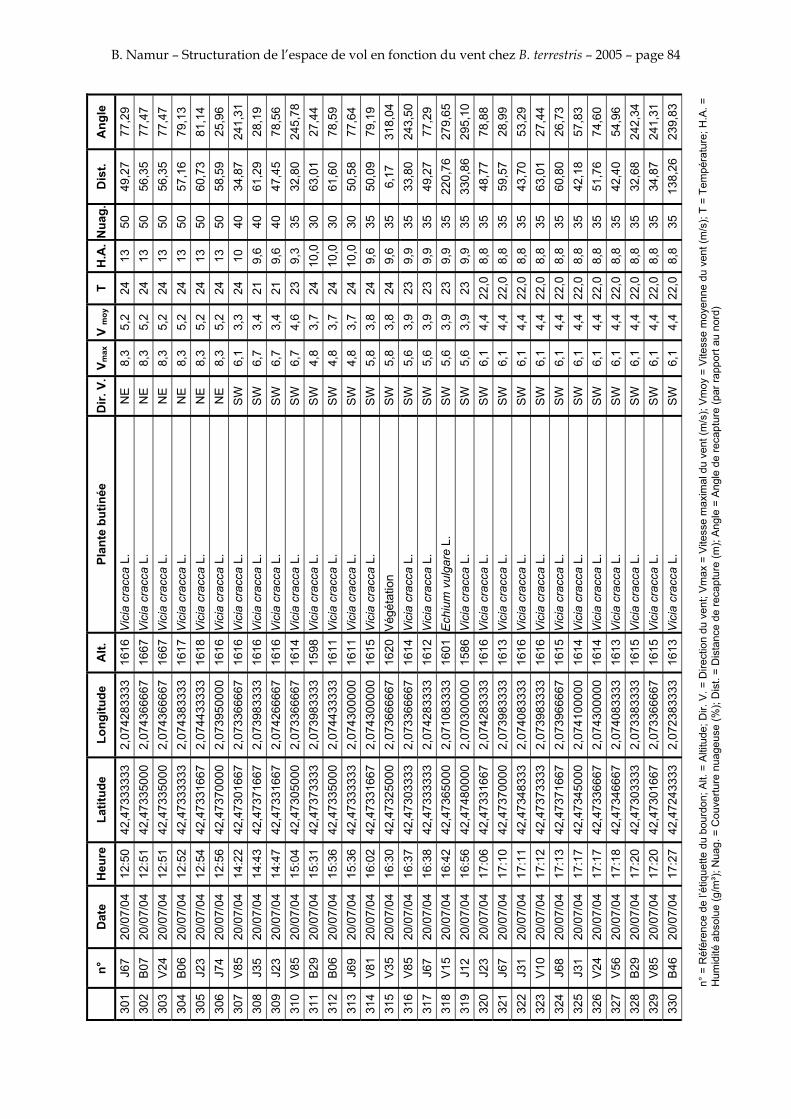
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 84
Ang
le
77,2
9 77
,47
77,4
7 79
,13
81,1
4 25
,96
241,
31
28,1
9 78
,56
245,
78
27,4
4 78
,59
77,6
4 79
,19
318,
04
243,
50
77,2
9 27
9,65
29
5,10
78
,88
28,9
9 53
,29
27,4
4 26
,73
57,8
3 74
,60
54,9
6 24
2,34
24
1,31
23
9,83
Dis
t.
49,2
7 56
,35
56,3
5 57
,16
60,7
3 58
,59
34,8
7 61
,29
47,4
5 32
,80
63,0
1 61
,60
50,5
8 50
,09
6,17
33
,80
49,2
7 22
0,76
33
0,86
48
,77
59,5
7 43
,70
63,0
1 60
,80
42,1
8 51
,76
42,4
0 32
,68
34,8
7 13
8,26
Nua
g.
50
50
50
50
50
50
40
40
40
35
30
30
30
35
35
35
35
35
35
35
35
35
35
35
35
35
35
35
35
35
H.A
.
13
13
13
13
13
13
10
9,6
9,6
9,3
10,0
10
,0
10,0
9,
6 9,
6 9,
9 9,
9 9,
9 9,
9 8,
8 8,
8 8,
8 8,
8 8,
8 8,
8 8,
8 8,
8 8,
8 8,
8 8,
8
T 24
24
24
24
24
24
24
21
21
23
24
24
24
24
24
23
23
23
23
22,0
22
,0
22,0
22
,0
22,0
22
,0
22,0
22
,0
22,0
22
,0
22,0
V m
oy
5,2
5,2
5,2
5,2
5,2
5,2
3,3
3,4
3,4
4,6
3,7
3,7
3,7
3,8
3,8
3,9
3,9
3,9
3,9
4,4
4,4
4,4
4,4
4,4
4,4
4,4
4,4
4,4
4,4
4,4
V max
8,3
8,3
8,3
8,3
8,3
8,3
6,1
6,7
6,7
6,7
4,8
4,8
4,8
5,8
5,8
5,6
5,6
5,6
5,6
6,1
6,1
6,1
6,1
6,1
6,1
6,1
6,1
6,1
6,1
6,1
Dir.
V.
NE
N
E
NE
N
E
NE
N
E
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
Plan
te b
utin
ée
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Végé
tatio
n V
icia
cra
cca
L.
Vic
ia c
racc
a L.
E
chiu
m v
ulga
re L
. V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Alt.
1616
16
67
1667
16
17
1618
16
16
1616
16
16
1616
16
14
1598
16
11
1611
16
15
1620
16
14
1612
16
01
1586
16
16
1613
16
16
1616
16
15
1614
16
14
1613
16
15
1615
16
13
Long
itude
2,07
4283
333
2,07
4366
667
2,07
4366
667
2,07
4383
333
2,07
4433
333
2,07
3950
000
2,07
3366
667
2,07
3983
333
2,07
4266
667
2,07
3366
667
2,07
3983
333
2,07
4433
333
2,07
4300
000
2,07
4300
000
2,07
3666
667
2,07
3366
667
2,07
4283
333
2,07
1083
333
2,07
0300
000
2,07
4283
333
2,07
3983
333
2,07
4083
333
2,07
3983
333
2,07
3966
667
2,07
4100
000
2,07
4300
000
2,07
4083
333
2,07
3383
333
2,07
3366
667
2,07
2383
333
Latit
ude
42,4
7333
333
42,4
7335
000
42,4
7335
000
42,4
7333
333
42,4
7331
667
42,4
7370
000
42,4
7301
667
42,4
7371
667
42,4
7331
667
42,4
7305
000
42,4
7373
333
42,4
7335
000
42,4
7333
333
42,4
7331
667
42,4
7325
000
42,4
7303
333
42,4
7333
333
42,4
7365
000
42,4
7480
000
42,4
7331
667
42,4
7370
000
42,4
7348
333
42,4
7373
333
42,4
7371
667
42,4
7345
000
42,4
7336
667
42,4
7346
667
42,4
7303
333
42,4
7301
667
42,4
7243
333
Heu
re
12:5
0 12
:51
12:5
1 12
:52
12:5
4 12
:56
14:2
2 14
:43
14:4
7 15
:04
15:3
1 15
:36
15:3
6 16
:02
16:3
0 16
:37
16:3
8 16
:42
16:5
6 17
:06
17:1
0 17
:11
17:1
2 17
:13
17:1
7 17
:17
17:1
8 17
:20
17:2
0 17
:27
Dat
e
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
20/0
7/04
20
/07/
04
n°
J67
B07
V24
B06
J23
J74
V85
J35
J23
V85
B29
B06
J69
V81
V35
V85
J67
V15
J12
J23
J67
J31
V10
J68
J31
V24
V56
B29
V85
B46
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
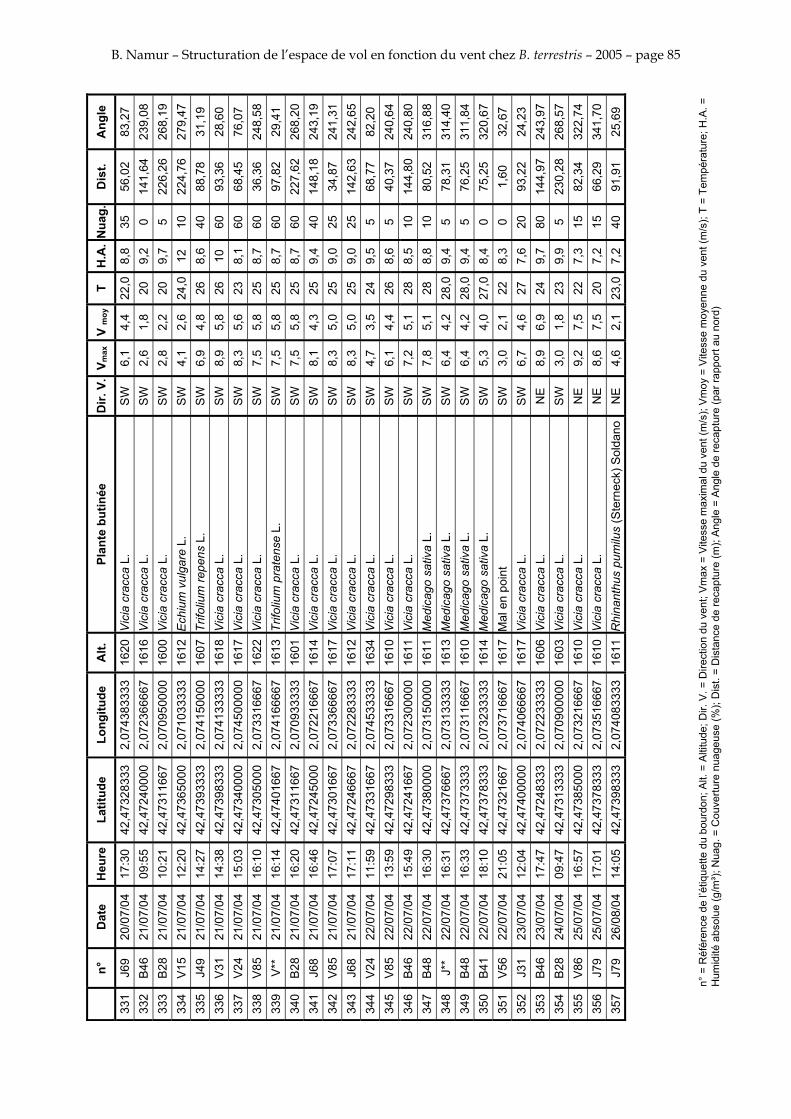
B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – page 85
Ang
le
83,2
7 23
9,08
26
8,19
27
9,47
31
,19
28,6
0 76
,07
248,
58
29,4
1 26
8,20
24
3,19
24
1,31
24
2,65
82
,20
240,
64
240,
80
316,
88
314,
40
311,
84
320,
67
32,6
7 24
,23
243,
97
268,
57
322,
74
341,
70
25,6
9
Dis
t.
56,0
2 14
1,64
22
6,26
22
4,76
88
,78
93,3
6 68
,45
36,3
6 97
,82
227,
62
148,
18
34,8
7 14
2,63
68
,77
40,3
7 14
4,80
80
,52
78,3
1 76
,25
75,2
5 1,
60
93,2
2 14
4,97
23
0,28
82
,34
66,2
9 91
,91
Nua
g.
35
0 5 10
40
60
60
60
60
60
40
25
25
5 5 10
10
5 5 0 0 20
80
5 15
15
40
H.A
.
8,8
9,2
9,7
12
8,6
10
8,1
8,7
8,7
8,7
9,4
9,0
9,0
9,5
8,6
8,5
8,8
9,4
9,4
8,4
8,3
7,6
9,7
9,9
7,3
7,2
7,2
T
22,0
20
20
24
,0
26
26
23
25
25
25
25
25
25
24
26
28
28
28,0
28
,0
27,0
22
27
24
23
22
20
23
,0
V m
oy
4,4
1,8
2,2
2,6
4,8
5,8
5,6
5,8
5,8
5,8
4,3
5,0
5,0
3,5
4,4
5,1
5,1
4,2
4,2
4,0
2,1
4,6
6,9
1,8
7,5
7,5
2,1
V max
6,1
2,6
2,8
4,1
6,9
8,9
8,3
7,5
7,5
7,5
8,1
8,3
8,3
4,7
6,1
7,2
7,8
6,4
6,4
5,3
3,0
6,7
8,9
3,0
9,2
8,6
4,6
Dir.
V.
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
SW
S
W
NE
S
W
NE
N
E
NE
Plan
te b
utin
ée
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
E
chiu
m v
ulga
re L
. Tr
ifoliu
m re
pens
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
Tr
ifoliu
m p
rate
nse
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
M
edic
ago
sativ
a L.
M
edic
ago
sativ
a L.
M
edic
ago
sativ
a L.
M
edic
ago
sativ
a L.
M
al e
n po
int
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
V
icia
cra
cca
L.
Vic
ia c
racc
a L.
R
hina
nthu
s pu
milu
s (S
tern
eck)
Sol
dano
Alt.
1620
16
16
1600
16
12
1607
16
18
1617
16
22
1613
16
01
1614
16
17
1612
16
34
1610
16
11
1611
16
13
1610
16
14
1617
16
17
1606
16
03
1610
16
10
1611
Long
itude
2,07
4383
333
2,07
2366
667
2,07
0950
000
2,07
1033
333
2,07
4150
000
2,07
4133
333
2,07
4500
000
2,07
3316
667
2,07
4166
667
2,07
0933
333
2,07
2216
667
2,07
3366
667
2,07
2283
333
2,07
4533
333
2,07
3316
667
2,07
2300
000
2,07
3150
000
2,07
3133
333
2,07
3116
667
2,07
3233
333
2,07
3716
667
2,07
4066
667
2,07
2233
333
2,07
0900
000
2,07
3216
667
2,07
3516
667
2,07
4083
333
Latit
ude
42,4
7328
333
42,4
7240
000
42,4
7311
667
42,4
7365
000
42,4
7393
333
42,4
7398
333
42,4
7340
000
42,4
7305
000
42,4
7401
667
42,4
7311
667
42,4
7245
000
42,4
7301
667
42,4
7246
667
42,4
7331
667
42,4
7298
333
42,4
7241
667
42,4
7380
000
42,4
7376
667
42,4
7373
333
42,4
7378
333
42,4
7321
667
42,4
7400
000
42,4
7248
333
42,4
7313
333
42,4
7385
000
42,4
7378
333
42,4
7398
333
Heu
re
17:3
0 09
:55
10:2
1 12
:20
14:2
7 14
:38
15:0
3 16
:10
16:1
4 16
:20
16:4
6 17
:07
17:1
1 11
:59
13:5
9 15
:49
16:3
0 16
:31
16:3
3 18
:10
21:0
5 12
:04
17:4
7 09
:47
16:5
7 17
:01
14:0
5
Dat
e
20/0
7/04
21
/07/
04
21/0
7/04
21
/07/
04
21/0
7/04
21
/07/
04
21/0
7/04
21
/07/
04
21/0
7/04
21
/07/
04
21/0
7/04
21
/07/
04
21/0
7/04
22
/07/
04
22/0
7/04
22
/07/
04
22/0
7/04
22
/07/
04
22/0
7/04
22
/07/
04
22/0
7/04
23
/07/
04
23/0
7/04
24
/07/
04
25/0
7/04
25
/07/
04
26/0
8/04
n°
J69
B46
B28
V15
J49
V31
V24
V85
V**
B28
J68
V85
J68
V24
V85
B46
B48
J**
B48
B41
V56
J31
B46
B28
V86
J79
J79
n° =
Réf
éren
ce d
e l’é
tique
tte d
u bo
urdo
n; A
lt. =
Alti
tude
; Dir.
V. =
Dire
ctio
n du
ven
t; V
max
= V
itess
e m
axim
al d
u ve
nt (m
/s);
Vm
oy =
Vite
sse
moy
enne
du
vent
(m/s
); T
= Te
mpé
ratu
re; H
.A. =
H
umid
ité a
bsol
ue (g
/m³)
; Nua
g. =
Cou
vertu
re n
uage
use
(%);
Dis
t. =
Dis
tanc
e de
reca
ptur
e (m
); A
ngle
= A
ngle
de
reca
ptur
e (p
ar ra
ppor
t au
nord
)
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357

B. Namur – Structuration de l’espace de vol en fonction du vent chez B. terrestris – 2005 – planche I
Photo 1. Colonie de Bombus terrestris (L.)
(Photo B. Namur)
Photo 3. Bourdon immobilisé et marqué
(Photo B. Namur)
Photo 5. Près de fauche du Pla del Bac
(Photo B. Namur)
Photo 2. Tube de contention (Photo B.
Namur)
Photo 4. Le village d’Eyne
(Photo B. Namur)
Photo 6. Bourdon marqué sur Vicia cracca
L. (Photo B. Namur)





























