Embed Size (px)
Citation preview

No d’ordre ………
UNIVERSITE DE OUAGADOUGOU ---------------
École Doctorale Sciences et Technologies
--------------- Laboratoire de Biologie et de
génétique moléculaires (Labiogene)
Thèse Présentée Par Cyrille BISSEYE
Pour obtenir le grade de
Docteur de l’Université de Ouagadougou
Option Sciences Biologiques Appliquées
Réponses immunes humorales et cellulaires dirigées contre les antigènes de Plasmodium falciparum candidats vaccins dans une zone d’endémie palustre à transmission saisonnière (Gambie)
Soutenue le 14 Décembre 2011 devant le jury composé de :
Président : David MODIANO, Professeur Titulaire, Université de Rome La Sapienza (Italie) Membres :
Yves TRAORE, Maître de Conférences, Université de Ouagadougou (Rapporteur)
Jacques SIMPORE, Professeur Titulaire, Université de Ouagadougou (Directeur de thèse)
Virginio Pietra, Chargé de Recherches, Université de Brescia (Italie)

2
DEDICACES Je dédie cette thèse de Doctorat :
A Feu Mon Père ANGOGHE NDONG ROBERT GUY pour l’amour, la patience et l’amitié.
A ma grand-mère AGNEGHE MARIE pour avoir fait de moi un homme.
A ma mère NZE ANTOINETTE (La rose), pour l’amour, la lumière et les prières.
A CYRIANE QUESSIA MARY pour être la raison de croire même quand les nuages s’amoncèlent.
A DZIMEZE MBA JEAN, EMANE, BISSOUE Bi Ekoh ALBERT, ESSINGONE ANGOGHE THIERRY pour l’aide précieuse et les encouragements dans les moments de doute.
A mes Frères, Sœurs, Parents et Amis, pour votre soutien moral, pour être ma famille et pour tous vos encouragements tout au long de ces années. A Yâ Nse et A Yâ Katoue A BETTY, VIVIE et LUCILE.
A mes Oncles OYE MENGOME, NKOUBA, NTSAME, WAGHA, ANGWE, NTOUTOUME, EKOMI, NDONG, BITEGHE OBAME pour l’aide et le soutien pendant mes études.
A mon frère et ami BATOUME CELESTIN pour l’amitié et le soutien dans nos différentes aventures.
A tous mes amis et collegues de FANGUINOVENY, MELEN et de l’EDR (Franceville).
Je n’oublie pas tous les frères et les sœurs d’ADOUMA (Lambaréné), mes compagnons de toujours.

3
REMERCIEMENTS
Ce travail a été réalisé en partie au Laboratoire dirigé par le Docteur Jamila Ismaili au Medical Research
Council (MRC) en Gambie, mais aussi au Laboratoire de Biologie Moléculaire et de Génétique (LABIOGENE de l’UFR-
SVT, Université de Ouagadougou), du Centre de Recherche Biomoléculaire Pietro Annigoni (CERBA) dirigé par le
Professeur Jacques SIMPORE.
Je tiens à exprimer ma profonde gratitude au Professeur Jacques SIMPORE, Directeur de cette thèse, membre du
jury et Directeur de plusieurs institutions : Laboratoire de Biologie Moléculaire et de Génétique (LABIOGENE de
l’UFR/SVT, Université de Ouagadougou) ; Centre de Recherche Biomoléculaire Pietro Annigoni (CERBA) ; Laboratoire du
Centre Médical Saint Camille de Ouagadougou et Recteur de l’Université Saint Thomas d’Aquin (USTA). Le
Professeur SIMPORE est le principal promoteur de la présente thèse, merci pour votre soutien moral et financier,
pour la qualité de l’encadrement scientifique, pour vos précieux conseils et votre disponibilité en dépit de vos
multiples fonctions.
Je remercie également le Docteur Jamila Ismaili pour la fructueuse collaboration au MRC en Gambie sans
laquelle cette thèse n’aurait peut-être pas aboutie.
Je remercie également le Professeur David Modiano (Université de Rome La Sapienza) qui a bien voulu
accepter d’évaluer et de présider le jury de cette thèse.
Je remercie Le Professeur Elie Mavoungou, Directeur de Recherches qui a bien voulu lire et juger la présente
thèse.
Mes remerciements vont aussi au Professeur Yves Traoré (Université de Ouagadougou) qui a accepté de juger la
présente thèse.
Je remercie Le Docteur Virginio Pietra, Médecin épidémiologiste, chargé de recherches (Université de Brescia,
Italie), qui a bien voulu participer au jury de la présente thèse.

4
Je remercie aussi les Professeurs Bernard Vray et Sylvain Meuris de l’Université Libre de Bruxelles.
Je remercie le Professeur Bernard Vray pour m’avoir accueilli et initié à l’immunologie dans son laboratoire à
l’université Libre de Bruxelles. Le Professeur Vray me fait aussi la grande amitié de relire cette thèse et donc d’en
améliorer la qualité, Professeur je vous en remercie infiniment.
Je remercie également le Professeur Anthony Holder (National Institute of Medical Research, MRC, UK) pour la
précieuse collaboration dans la préparation de mes publications.
Je remercie également le Docteur Giorgio Sirugo (Université de Rome) pour notre collaboration en Gambie et
pour son aide précieuse.
Je voudrais aussi adresser mes remerciements au Docteur Mahamoudou Sanou Directeur du Centre National de
Transfusion sanguine pour notre fructueuse collaboration.
Je remercie Mr Bolni Marius Nagalo pour la sympathie, la franche collaboration mais aussi pour les bons repas
chez Tantine.
J’adresse mes remerciements à Dénis, Lucien, Robert, Georges et Patrice du Centre Médical Saint Camille
de Ouagadougou pour leur sympathie pendant toutes ces années.
Mes remerciements s’adressent aussi à l’ensemble de l’équipe du CERBA/LABIOGENE, Djeneba, Florencia, Tani,
Zoenabo, Zeba, Rémy, Bazié, Désiré, Stella Laure pour les moments d’échanges et de partage au
laboratoire.
Je remercie aussi mes amis de toujours, Guy Pierre Biteghe, Endama Mbang Anicet (Ziggy), Ango Billie
Alix qui malgré les années et la distance ont su garder le contact.
Je remercie enfin mon frère Engone Moure Engone Wilfried pour avoir été souvent le lien avec la famille.

5
TABLE DES MATIERES DEDICACES ........................................................................................................................................................... 2
REMERCIEMENTS................................................................................................................................................. 3
TABLE DES MATIERES .......................................................................................................................................... 5
LISTE DES FIGURES ............................................................................................................................................. 10
LISTE DES TABLEAUX ......................................................................................................................................... 11
SIGLES ET ABREVIATIONS .................................................................................................................................. 12
RESUME .............................................................................................................................................................. 15
INTRODUCTION ................................................................................................................................................. 18
Objectifs de la thèse .......................................................................................................................................... 19
I. GENERALITES ............................................................................................................................................. 21
I.1. Situation globale et historique ............................................................................................................... 21
I.1.1. Situation globale du paludisme ...................................................................................................... 21
I.1.2 Historique et pathologie .................................................................................................................. 21
I.2. Le parasite ............................................................................................................................................... 23
I.2.1. Cycle de vie de Plasmodium ............................................................................................................ 23
I.2.2. Pathogenèse du paludisme ............................................................................................................. 25
I.2.2.1. La Séquestration ....................................................................................................................... 25
I.2.2.2.Les Rosettes ............................................................................................................................... 26
I.2.2.3. La Variation antigénique .......................................................................................................... 26
I.2.3. Antigènes de surface de P. falciparum ........................................................................................... 27
I.2.3.1. Antigène de surface du mérozoïte 1 (MSP1) .......................................................................... 27
I.2.3.2. Apical membrane Antigen-1 (AMA1) ...................................................................................... 31
I.2.3.3. Protéine Circumsporozoite (CSP) ............................................................................................. 32
I.3. La maladie ................................................................................................................................................ 33
I.3.1. Caractéristiques cliniques ............................................................................................................... 33
I.3.1.1. Le paludisme simple ................................................................................................................. 34

6
I.3.1.2. Paludisme grave ....................................................................................................................... 34
I.3.2. Paludisme et grossesse ................................................................................................................... 35
I.3.3. Epidémiologie du paludisme ........................................................................................................... 37
I.3.4. Résistance des parasites aux médicaments antipaludiques ......................................................... 40
I.3.4.1. Résistance à la chloroquine ..................................................................................................... 40
I.3.4.2. Résistance à la pyrimethamine-sulfadoxine ........................................................................... 41
I.3.4.3. Résistance à la quinine ............................................................................................................. 42
I.3.4.4. La résistance à la méfloquine .................................................................................................. 42
I.4. Paludisme à P. falciparum et vaccination .............................................................................................. 43
I.4.1. Vaccins pré-érythrocytaires ............................................................................................................ 44
I.4.2. Vaccins érythrocytaires ................................................................................................................... 45
I.5. Immunité antipalustre ............................................................................................................................ 46
I.5.1. Mécanismes effecteurs de protection contre le paludisme ......................................................... 46
I.5.1.1 Immunité au stade pré-érythrocytaire ..................................................................................... 46
I.5.1.2. Immunité au stade érythrocytaire ........................................................................................... 48
I.5.1.3. Immunité dirigée contre les antigènes variables de surface ................................................. 50
I.5.1.4. Mémoire immunologique ........................................................................................................ 51
I.5.2. Rôle des cytokines dans l’infection palustre .................................................................................. 52
I.5.2.1 Cytokines pro-inflammatoires et paludisme ............................................................................ 52
I.5.2.1.1. TNF- ................................................................................................................................. 52
I.5.2.1.2. Interféron-gamma (IFN-) ................................................................................................. 53
I.5.2.1.3. L’interleukine-12 (IL-12) ................................................................................................... 54
I.5.2.2. Cytokines anti-inflammatoires ................................................................................................ 55
I.5.2.2.1. Interleukine-4 .................................................................................................................... 56
I.5.2.2.2. Facteur de croissance transformant-beta (TGF-β).......................................................... 56
I.5.2.2.3. L’interleukine-10 (IL-10) ................................................................................................... 57
I.5.3. Les cellules présentatrices d’antigènes (CPA) ............................................................................... 58

7
I.5.3.1 Les cellules dendritiques (DC) ................................................................................................... 59
I.5.3.2. Les lymphocytes B..................................................................................................................... 59
I.5.3.3. Les macrophages/monocytes .................................................................................................. 60
I.5.4. Stratégies d’évasion immunitaire ................................................................................................... 60
I.5.4.1. Altération des ligands peptidiques .......................................................................................... 60
I.5.4.2. Altération des cellules présentatrices d’antigènes ................................................................. 61
I.5.5. Rôle des cellules T régulatrices naturelles dans l’infection palustre ........................................... 61
I.5.5.1. Rôle des Tregs naturelles dans le contrôle de l’infection palustre chez les rongeurs .......... 63
I.5.5.2. Rôle des cellules Tregs naturelles dans le contrôle du paludisme cérébral expérimental ... 64
I.5.5.3. Rôle des cellules T régulatrices dans le contrôle du paludisme chez l’homme..................... 65
II. Matériels et méthodes .................................................................................................................................. 68
II.1. Populations d’étude ............................................................................................................................... 68
II.2. Mesure des anticorps dirigés contre P. falciparum par Enzyme Linked Immunosorbent Assay (ELISA) ............................................................................................................................................................ 70
II.2.1. ELISA MSP119 et AMA1 ................................................................................................................... 70
II.2.2. ELISA pour mesurer les anticorps anti-R32LR (CSP) ..................................................................... 71
II.3. Culture in vitro de P. falciparum et préparation d’extraits de schizontes ......................................... 72
II.4. Séparation et analyse des cellules T CD4+CD25+.................................................................................. 73
II.5. Culture des cellules mononuclées du sang de cordon ........................................................................ 73
II.6. Détection des cellules T CD4+ productrices d’IFN-γ par culture ELISPOT (Enzyme Linked Immunoassay Spot) ....................................................................................................................................... 74
II.7. Extraction de l'ARN et transcription inverse de Foxp3 ........................................................................ 75
II.8. Génotypage de la G6PD et de l’hémoglobine S par PCR en temps réel ........................................... 75
III. Résultats ........................................................................................................................................................ 78
III.1. Article I. L’infection placentaire par Plasmodium falciparum interfère sur l’équilibre Th1/Th2 des cellules T néonatales à travers les cellules T régulatrices CD4+CD25+FoxP3+ et la production d’IL-10 ... 78
III.1.1. But de l’étude................................................................................................................................. 78
III.1.2. Population ...................................................................................................................................... 78
III.1.3. Résultats ....................................................................................................................................... 79
IV.1.3.1 Fréquence des cellules T CD4+CD25+ FoxP3+ parmi les CMSCs ............................................. 79

8
III.1.3.2. Effet des cellules T régulatrices sur les cytokines Th1/Th2 produites par les CMSCs en réponse à la stimulation par la PHA .................................................................................................... 80
III.1.3.3. Effets des Tregs sur les cytokines Th1/Th2 produites par les CMSCs en réponses aux globules rouges intacts parasités et aux antigènes de P. falciparum ............................................... 82
III.1.3.5. Effet de l'IL-10 sur la production des cytokines Th1/Th2 par les CMSCs ............................ 84
III.1.4. Conclusion ...................................................................................................................................... 85
III.2. Article II. Un variant de la protéine de 19 kilodaltons de l’extrémité C-terminal du Merozoite Surface Protein 1 de Plasmodium falciparum candidat vaccin induit des taux élevés d’IFN-gamma associés à des réponses immunitaires cellulaires à des séquences peptidiques spécifiques chez les adultes gambiens naturellement exposés au paludisme ........................................................................... 86
Clinical and Experimental Immunology (2011), 166 (3):366-73 ............................................................ 86
III.2.1. But de l’étude................................................................................................................................. 86
III.2.2. Population ...................................................................................................................................... 87
III.2.3. Résultats ......................................................................................................................................... 87
III.2.3.1. Réponses des adultes gambiens aux protéines MSP119 par ELISPOT en culture ................ 87
III.2.3.2. Production des cytokines en réponses aux protéines MSP119 ............................................. 87
III.2.3.3. Réponses des adultes gambiens aux peptides MSP119 par ELISPOT en culture ................. 87
III.2.3.4. Production de cytokines en réponse aux peptides mutants et sauvages de MSP119 ......... 91
III.2.3.5.Typage HLA-DR et présentation des peptides MSP119 aux cellules T CD4+ ......................... 91
III.2.4. Conclusion ...................................................................................................................................... 94
III.3. Article III. Comparaison des réponses humorales dirigées contre CSP, AMA1 et MSP119 et la susceptibilité au paludisme chez les Peuls, Wolofs et Mandingues de Gambie ........................................... 95
Article soumis pour publication ................................................................................................................... 95
III.3.1. But de l’étude ..................................................................................................................................... 95
III.3.2. Population .......................................................................................................................................... 96
III.3.3. Résultats ............................................................................................................................................. 96
III.3.3.1. Caractéristiques des participants de l’étude .......................................................................... 96
III.3.3.2. Comparaisons interethniques de l'incidence du paludisme .................................................... 96
III.3.3.3. Comparaisons interethniques dans la réponse humorale à CSP, AMA1 et MSP119 ............... 98
III.3.3.4. Association anticorps et incidence du paludisme et de la parasitémie ................................101
III.3.4. Conclusion ........................................................................................................................................105

9
IV. Discussion générale ...................................................................................................................................106
V. Conclusion générale ....................................................................................................................................118
VI. Perspectives ................................................................................................................................................120
Bibliographie ....................................................................................................................................................122
Annexes ............................................................................................................................................................163

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 10
LISTE DES FIGURES
FIGURE 1. DISTRIBUTION DU PALUDISME À P. FALCIPARUM EN FONCTION DE L'ENDÉMICITÉ EN 2007. ...........22 FIGURE 2. CYCLE DE PLASMODIUM FALCIPARUM ET MÉCANISMES IMMUNITAIRES DE L’HÔTE À CHAQUE PHASE DU
CYCLE DU PARASITE. ............................................................................................................24 FIGURE 3. RÉCEPTEURS DE L’HÔTE IMPLIQUÉS DANS LA SÉQUESTRATION DU GLOBULE ROUGE INFECTÉ PAR P.
FALCIPARUM. ....................................................................................................................26 FIGURE 4. REPRÉSENTATION SCHÉMATIQUE DE MSP1 DE P. FALCIPARUM...............................................28 FIGURE 5. ASSEMBLAGE ET PROCESSING DU COMPLEXE MSP1. ............................................................28 FIGURE 6. REPRÉSENTATION SCHÉMATIQUE D’AMA1. .......................................................................31 FIGURE 7. REPRÉSENTATION SCHÉMATIQUE DE CSP ET DU VACCIN RTS, S. .............................................33 FIGURE 8. EFFET DU PALUDISME PENDANT LA GROSSESSE SUR LA SANTÉ MATERNELLE, NÉONATALE ET INFANTILE.
......................................................................................................................................36 FIGURE 9. REPRÉSENTATION SCHÉMATIQUE DU POSSIBLE MÉCANISME D’ACTION DE L’IMMUNITÉ À MÉDIATION
CELLULAIRE DIRIGÉE CONTRE LE STADE SANGUIN DE PLASMODIUM ..................................................49 FIGURE 10. MÉCANISMES DE BASE UTILISÉS PAR LES TREGS .................................................................62 FIGURE 11. (A) CONTRÔLE DE QUALITÉ DE LA DÉPLÉTION DES CELLULES T CD4+CD25+
(B) EXPRESSION DE
L’ARNM FORKHEAD BOX P3 (FOXP3) ET (C) PRODUCTION DE L’IL-10 APRÈS STIMULATION PAR LA PHA
PENDANT 18 H...................................................................................................................79 FIGURE 12. POURCENTAGE DES CELLULES TREGS CHEZ LES CMCS PARASITÉS OU NON PARASITÉS. .................80 FIGURE 13. PRODUCTION DE CYTOKINES PAR LES CMSCS EN RÉPONSE À LA PHYTOHÉMAGGLUTININE (PHA). ..81 FIGURE 14. PRODUCTION DE CYTOKINES PAR LES CMSCS EN RÉPONSE AUX GLOBULES PARASITÉS ET AUX
ANTIGÈNES DE P. FALCIPARUM. ..............................................................................................83 FIGURE 15. LE TRAITEMENT DES CULTURES CELLULAIRES AVEC DES ANTICORPS ANTI-IL-10 RESTAURE LA
PRODUCTION D’IFN- DANS LE GROUPE DE PLACENTAS PARASITÉS. .................................................84 FIGURE 16. PRODUCTION DE CYTOKINES PAR LES PBMC EN RÉPONSE AUX PROTÉINES MSP119. ..................89
FIGURE 17. (A) FRÉQUENCE DES DONNEURS ET DES CELLULES SÉCRÉTRICES D’IFN- EN RÉPONSE AUX PEPTIDES DE
MSP-119 CHEZ 43 ADULTES GAMBIENS EXPOSÉS AU PLASMODIUM. ...............................................90 FIGURE 18. PRODUCTION DE CYTOKINES PAR LES PBMCS EN RÉPONSE AUX PEPTIDES MSP-119. ..................92 FIGURE 19. CONCENTRATION EN IGG, IGG1 ET IGG3 ANTI-CSP, ANTI-AMA1 ET ANTI-MSP119 CHEZ LES
PEULS, WOLOFS ET MANDINGUES. ....................................................................................... 102
FIGURE 20. COURBE DE SURVIE DE KAPLAN-MEIER REPRÉSENTANT LES RÉPONSES HUMORALES ET L’INCIDENCE
DU PALUDISME DANS LES 3 GROUPES ETHNIQUES PEUL , MANDINGUE ET WOLOF . ........................... 104

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 11
LISTE DES TABLEAUX
TABLEAU I : DONNÉES ÉPIDÉMIOLOGIQUES DES FEMMES GAMBIENNES DE SUKUTA. ...................................78
TABLEAU II. SÉQUENCE DES PEPTIDES MUTANTS ET SAUVAGES DE MSP119 .............................................88
TABLEAU III. ALLÈLES HLA-DR CLASS II DÉTECTÉS DANS LA POPULATION D’ÉTUDE. ...................................93
TABLEAU IV. CARACTÉRISTIQUES DES PARTICIPANTS DE L’ÉTUDE ET DE LA RÉPONSE HUMORALE AUX ANTIGÈNES
DE P. FALCIPARUM CSP, AMA1 ET MSP119 À L’INCLUSION .........................................................97
TABLEAU V. CARACTÉRISTIQUES DES PARTICIPANTS DU GROUPE À L’ENRÔLEMENT ET INCIDENCE DU PALUDISME
EN ASSOCIATION AVEC LES TAUX D’IGG ANTI-AMA1, ANTI-MSP119 ET ANTI-CSP AJUSTÉS POUR LES
FACTEURS CONFONDANTS. ....................................................................................................99
TABLEAU VI. RÉPONSES IMMUNITAIRES HUMORALES À P. FALCIPARUM À L'ENRÔLEMENT DANS LES 3 GROUPES
ETHNIQUES ..................................................................................................................... 101
TABLEAU VII. INCIDENCE DU PALUDISME EN ASSOCIATION AVEC LES TAUX DES SOUS-CLASSES D’IGG ANTI-
AMA1, CSP ET MSP119 AJUSTÉS POUR TOUS LES FACTEURS CONFONDANTS .................................. 103

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 12
SIGLES ET ABREVIATIONS
Ac
ADCI
Anticorps
Immunité cellulaire dépendante d’anticorps
ADN Acide désoxyribonucléique ADNc Acide désoxyribonucléique complémentaire ARN Acide Ribonucléique
AMA1 Antigène apical de la membrane du mérozoite
AMPc Adénosine monophosphate cyclique
BCR Récepteur du lymphocyte B
CD Groupe de différenciation
CERBA Centre de recherche Biomoléculaire Pietro
CIDR Région inter-domaine riche en cystéine
CMH Complexe majeur d’histocompatibilité
CMSC Cellules mononuclées du sang de cordon
CPA Cellule présentatrice d’antigène
CSP Protéine circumsporozoite
CSA Sulfate de Chondroïtine A
CR1 Récepteur du complément 1
CTLA-4 Antigène du lymphocyte T Cytotoxique
DI Domaine I
DII Domaine II
DIII Domaine III
DBL Duffy Binding-Like
DC Cellule dendritique
DO Densité optique
DHFR Dihydrofolate réductase
DHPS Dihydropteroate synthétase
EBA Erythrocyte Binding Antigen

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 13
EDTA acide éthylène diamine tétracétique
EGF Eperdimal growth factor
ELISA Enzyme-Linked immunosorbent assay
ELISPOT Enzyme-Linked immunospot
EIR Taux d'inoculation entomologique
FITC Fluorescéine isothiocyanate
FOXP3 Forkhead box protein 3
GADPH Glycéraldéhyde 3 phosphate déshydrogénase
GM-CSF Facteur de croissance de colonies de granulocytes et de macrophages
GLURP Protéine riche en Glutamine
GPI Glycosylphosphatidylinositol
G6PD Glucose 6 phosphate déshydrogénase
GYP Glycophorine
Hb Hémoglobine
HLA antigènes des leucocytes humains
ICAM1 molécule d’adhésion intercellulaire
IDO Indolamine 2,3 dioxygénase
IFN Interféron
Ig immunoglobuline
IL Interleukine
kDa Kilo dalton
LAG-3 Gène d’activation du Lymphocyte
MSP1 Antigène de Surface du Merozoïte 1
MSP2 Antigène de Surface du Merozoïte 2
MSP3 Antigène de Surface du Merozoïte 3
MSP119 Fragment de 19 kilo daltons de MSP1
NK Cellule tueuse naturelle
NO Monoxyde d’azote
NOS Oxyde nitrique synthétase
iNOS Oxyde nitrique synthétase inductible

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 14
PBMC cellules mononuclées du sang circulant
PCR Réaction en Chaîne par Polymérase
PE Phycoérythrine
PHA Phytohémagglutinine
Pf Plasmodium falciparum
PfAMA1 AMA1 de Plasmodium falciparum
PfCRT Plasmodium falciparum Chloroquine resistance transporter
PfEMP1 Plasmodium falciparum erythrocyte membrane protein 1
Pfmdr1 Plasmodium falciparum multidrug resistance 1
RT-PCR PCR en temps réel
SERA5 Serine repeat Antigen 5
STAT4 Facteur de transcription 4
T CD4+ Lymphocytes T CD4+
T CD8+ Lymphocytes T CD8+
TCR Récepteur du lymphocyte T
TGF Facteur de croissance transformant
Th Lymphocyte T auxiliaire
TNF facteur de nécrose tumorale
Tregs Cellules T régulatrices
VSA Antigène variable de surface

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 15
RESUME Le paludisme demeure un problème majeur de santé publique en Afrique subsaharienne. Du
fait de la résistance du parasite aux antimalariques, et du vecteur aux insecticides, le
contrôle voire même l’éradication de la maladie passe par la mise au point d’un vaccin
efficace. À ce stade de la recherche aucun vaccin efficace n’a été mis sur le marché. La mise
au point d’un vaccin se heurte à la complexité du parasite. Différents antigènes du parasite
sont des candidats vaccins en phase d’essai clinique. La présente thèse est une contribution
à l’étude de l’immunité humorale et cellulaire contre les antigènes candidats vaccins AMA1,
CSP et MSP119 dans une zone d’endémie palustre à transmission saisonnière.
Nous avons étudié le rôle des cellules T régulatrices dans la réponse Th1/Th2 des cellules T
néonatales. Nous avons montré que l’infection placentaire influence la différentiation Th1
des cellules T néonatales en impliquant les cellules T régulatrices à travers la production d’IL-
10.
La réponse immunitaire cellulaire a été aussi évaluée en étudiant l’antigénicité d’une
protéine triple mutant de MSP119 chez des Gambiens adultes naturellement exposés au
paludisme. Cette protéine MSP119 triple mutant est responsable d’une production plus
élevée d’IFN-γ que la protéine native par les PBMCs de Gambiens adultes.
La réponse humorale dirigée contre les antigènes de P. falciparum a été évaluée en étudiant
les différences interethniques des réponses humorales dirigées contres ces derniers chez des
Peuls de Gambie et des groupes ethniques sympatriques Wolofs et Mandingues.
Nous avons montré, contrairement aux études précédentes, que l’incidence du paludisme, la
prévalence parasitaire et les réponses humorales dirigées contre les antigènes de P.
falciparum étaient les mêmes chez les Peuls de Gambie comparativement aux groupes
ethniques Wolofs et Mandingues.
Mots-clés : Immunité, Lymphocyte T, anticorps, groupe ethnique, Tregs.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 16
ABSTRACT
Malaria remains a major public health problem in sub-Saharan Africa. Because of parasite
resistance to antimalarial drugs and vectors to insecticides; control even eradication of the
disease necessites the development of an effective vaccine.
At this stage of researches, no effective vaccine is available on the market. The development
of a vaccine is hampered by the complexity of the parasite. Different parasite antigens which
are vaccine candidates are actually tested clinically.
The present thesis is a contribution to the humoral and cellular immunity against antigens
vaccine candidates AMA1, MSP119 and CSP in an area of malaria seasonal transmission.
We studied the role of regulatory T cells in the neonatal T cells Th1/Th2 bias and found that
Plasmodium placental infection influence the Th1 differentiation of neonatal T cells through
regulatory T cells and IL-10.
The cellular immune response was also assessed by studying the antigenicity of a triple
mutant MSP119 protein among Gambian adults naturally exposed to malaria.
The triple mutant MSP119 protein showed a higher production of IFN-γ than the native
protein by PBMCs of adult Gambians.
The antibody responses directed against P. falciparum antigens were assessed by studying
the inter-ethnic humoral differences in the Gambian Fulani and the sympatric ethnic groups
Wolof and Mandinka. In contrast to previous reports, we found that the prevalence and the
incidence of malaria parasite as well as humoral responses directed against P. falciparum
antigens were the same in the Gambian Fulani compared to Wolof and Mandinka.
Keywords: immunity, T cells, antibodies, ethnic group, Tregs.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 17

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 18
INTRODUCTION
Le paludisme demeure un problème majeur de santé publique en Afrique subsaharienne. En
effet, le paludisme continue d’y tuer les enfants, essentiellement ceux de moins de cinq ans.
Les femmes enceintes payent aussi un lourd tribut dans la morbidité palustre. Du fait de la
complexité du parasite à travers son cycle de vie, de sa résistance aux antipaludiques, des
échecs observés dans la lutte anti vectorielle et d’une compréhension incomplète des
mécanismes immunitaires assurant la protection de l’hôte contre la maladie; le
développement d’un vaccin anti palustre est l’outil qui permettrait le contrôle de l’infection
voire même son éradication.
Le développement d’un vaccin anti palustre se heurte non seulement à la complexité du
parasite à travers, son polymorphisme et sa variation antigénique, mais aussi à la
complexité des interactions entre le parasite et le système immunitaire de l’hôte.
Dans les régions hyper endémiques pour le paludisme, les adultes sont mieux protégés que
les enfants contre les formes sévères de la maladie. Cela, met en évidence un mécanisme de
protection des adultes, protection acquise au cours de longues années d’exposition au
parasite. Les mécanismes immunitaires responsables de cette protection ne sont pas bien
connus. Il a été montré que le parasite induit chez l’hôte de fortes réponses immunitaires
humorales et cellulaires. Ces réponses sont dirigées contre les antigènes du parasite dont
certains sont considérés comme de potentiels candidats vaccins. La présente thèse est une
contribution à une meilleure compréhension des mécanismes immunitaires humoraux et
cellulaires dirigés contre les antigènes de Plasmodium falciparum candidats vaccins. Les
différentes études qui font l’objet de cette thèse ont été menées en Gambie, dans une
région où la transmission du paludisme est saisonnière.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 19
Objectifs de la thèse
Objectif principal
Le principal objectif de ce travail est l’étude des réponses immunes humorales et cellulaires
dirigées contre quelques antigènes de P. falciparum candidats vaccins dans une zone
d’endémie palustre à transmission saisonnière.
Objectifs spécifiques
Ce travail se décompose en trois principaux objectifs spécifiques qui sont :
L’étude du rôle des cellules T régulatrices dans le biais Th1/Th2 des cellules T
néonatales
L’évaluation de l’antigénicité des peptides et des protéines du candidat vaccin
MSP119.
L’évaluation des réponses humorales dirigées contre les antigènes candidats vaccins
MSP119, AMA1 et CSP chez des sujets appartenant aux principaux groupes ethniques
de Gambie : Peul, Wolof et Mandinka.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 20

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 21
I. GENERALITES
I.1. Situation globale et historique
I.1.1. Situation globale du paludisme
Selon les estimations mondiales en 2009, 225 millions de personnes étaient malades
de paludisme et parmi elles, 781.000 personnes ont décédé de la maladie (OMS, 2010). Le
nombre de personnes malades dans les zones endémiques d’Afrique subsaharienne et d’Asie
du Sud-Est était respectivement de 176 et 49 milliions. L’Afrique subsaharienne avec
709.000 décès représentait 90% de la mortalité due au paludisme (OMS, 2010).
La majorité d’épisodes cliniques à Plasmodium falciparum (P. falciparum), se produit en
Afrique tropicale et en Asie du Sud-est avec respectivement 70% et 25% d’épisodes cliniques
(Snow et al, 2005) (figure 1).
Le paludisme est responsable en Afrique de 3000 décès par jour, la plupart des décès
concernent les enfants de moins de cinq ans. La distribution du paludisme dans les régions
endémiques du monde est définie en fonction du degré d'endémicité. Le paludisme est
considéré comme endémique dans les zones où il y a une transmission constante au cours
d’années successives. Le paludisme est dit hypo endémique dans les zones où la prévalence
de l'infection est inférieure à 10%, et méso endémique dans les zones où la prévalence de
l'infection se situe entre 11% et 50%. Dans les régions où la prévalence de l'infection est
supérieure à 50%, les zones sont définies comme holo endémiques et hyper endémiques.
I.1.2 Historique et pathologie
Le parasite du paludisme, P. falciparum, a d'abord été décrit par Laveran en 1880
dans le sang d'un patient présentant des fièvres intermittentes.
Les parasites du paludisme de l'homme appartiennent au genre Plasmodium, à la famille
des Plasmodiidae, au sous-ordre des Haemospondiidae et à l’ordre des coccidies. Les
parasites de l'homme appartiennent à deux sous-genres : Laverania et Plasmodium. Quatre
espèces distinctes de Plasmodium sont traditionnellement reconnues comme étant des
parasites de l’homme, il s’agit de: Plasmodium falciparum, Plasmodium malariae,
Plasmodium ovale et Plasmodium vivax.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 22
Figure 1. Distribution du paludisme à P. falciparum en fonction de l'endémicité en 2007.
La couleur rouge claire indique les zones hypo endémiques, le brun rougeâtre les zones méso endémiques et le rouge foncé les zones holo et hyper endémiques (Hay et al, 2009)
Trois parasites du paludisme sont exclusivement trouvés chez l’Homme, ce sont P.
falciparum (Welch, 1897); P. vivax (Grassi, 1890); et P.ovale (Stephens, 1922). Le paludisme à
P. malariae a été décrit aussi bien chez l’homme que chez les singes africains (Grassi, 1890).
Toutefois, il est à noter que les infections par P. knowlesi (parasite simien) ont été
rapportées chez l’homme à Bornéo (Singh et al, 2004).
Les quatre espèces responsables du paludisme chez l'homme se retrouvent dans les régions
tropicales et subtropicales du monde, mais leur répartition est variable: P. falciparum est le
parasite répandu en Afrique subsaharienne; P. vivax est plutôt le parasite le plus
fréquemment trouvé en Asie, en Amérique du Sud et Central alors qu'il est absent de
l'Afrique de l'Ouest, la majorité de la population ne portant pas le facteur Duffy qu’ utilise le
parasite pour entrer dans les globules rouges de l’hôte.
P. malariae et P. ovale sont des parasites beaucoup moins fréquents dans la plupart des
régions d'Afrique. La grande majorité des cas cliniques de la maladie et notamment la quasi-
totalité des décès liés au paludisme sont dues à P. falciparum.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 23
I.2. Le parasite
I.2.1. Cycle de vie de Plasmodium
Tous les parasites du paludisme ont besoin de deux hôtes dans leur cycle de vie: l'hôte
définitif où a lieu le développement sexuel (anophèle femelle) et l'hôte intermédiaire
(l'homme par exemple) où se produit le développement sexuel (Figure 2). Le plasmodium,
parasite haploïde adopte trois stratégies au cours de son cycle de vie pour proliférer.
La première stratégie est sa capacité de croissance et de réplication intensive,
réalisée grâce à trois stades différents. Le premier stade est l'oocyste qui se produit chez le
moustique dans un processus appelé sporogonie. Le deuxième stade est la schizogonie pré-
érythrocytaire (ou exo-érythrocytaire ou héatique) et le troisième stade est la schizogonie
érythrocytaire qui se produisent tous les deux chez l’hôte intermédiaire.
La deuxième stratégie adoptée par le parasite est la dispersion et l'invasion des cellules
hôtes. Cette stratégie inclut : les stades mérozoïte, sporozoïte et ookinète du parasite.
La troisième stratégie est la reproduction sexuée impliquant la formation des gamétocytes
chez l'hôte vertébré, elle est complétée lors de la formation de l'ookinète chez l’anophèle
femelle après un repas sanguin.
Les trois stratégies du parasite sont indiquées dans le cycle de vie ci-dessous chez le
moustique et chez l’hôte humain (Figure 2).
Les sporozoïtes sont injectés dans la circulation sanguine avec la salive du moustique, et
circulent pendant une courte période (2-30 min) avant d'entrer dans les hépatocytes.
Dans les hépatocytes, les sporozoïtes se transforment en schizontes pré-érythrocytaires qui
engendrent des mérozoïtes (stade hépatique de 5-15 jours). Un sporozoïte unique qui se
divise peut donner naissance à un schizonte multinuclé de 30.000 mérozoïtes filles. Les
mérozoïtes issus des schizontes hépatiques sont libérés dans la circulation et la phase
érythrocytaire du cycle peut commencer.
Chez P. falciparum, le cycle érythrocytaire dure 48 heures en fonction de la souche du
parasite. Les mérozoïtes envahissent les globules rouges et se développent jusqu’au stade
« ring » (anneaux) (0-24h), puis au stade trophozoïte (24-35h) et enfin au stade schizonte
(35-48h) où les mérozoïtes sont produits. Les mérozoïtes sont libérés dans la circulation
sanguine après la rupture des schizontes pour commencer un nouveau cycle érythrocytaire.
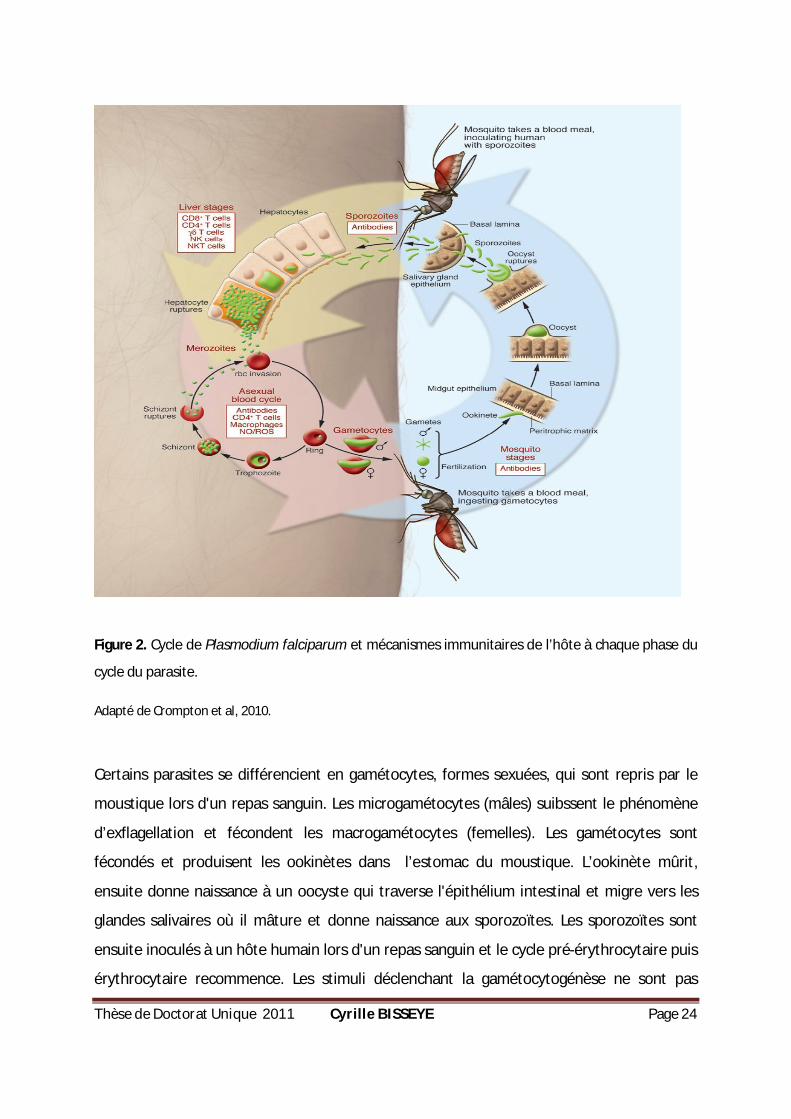
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 24
Figure 2. Cycle de Plasmodium falciparum et mécanismes immunitaires de l’hôte à chaque phase du
cycle du parasite.
Adapté de Crompton et al, 2010.
Certains parasites se différencient en gamétocytes, formes sexuées, qui sont repris par le
moustique lors d'un repas sanguin. Les microgamétocytes (mâles) suibssent le phénomène
d’exflagellation et fécondent les macrogamétocytes (femelles). Les gamétocytes sont
fécondés et produisent les ookinètes dans l’estomac du moustique. L’ookinète mûrit,
ensuite donne naissance à un oocyste qui traverse l'épithélium intestinal et migre vers les
glandes salivaires où il mâture et donne naissance aux sporozoïtes. Les sporozoïtes sont
ensuite inoculés à un hôte humain lors d'un repas sanguin et le cycle pré-érythrocytaire puis
érythrocytaire recommence. Les stimuli déclenchant la gamétocytogénèse ne sont pas

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 25
encore entièrement compris, mais résulteraient très probablement de diverses formes de
stress, y compris la pression exercée par le système immunitaire de l'hôte.
I.2.2. Pathogenèse du paludisme
Comme le montre son cycle de vie, P. falciparum survit chez le moustique où sa
reproduction sexuée s’effectue, et le moustique vecteur transmet le parasite à l'hôte
humain.
Chez l’homme le parasite a besoin de proliférer et de survivre sans être détruit par la
réponse immunitaire de l'hôte. Pour ce faire, P. falciparum utilise différentes techniques afin
de contourner le système immunitaire de l'hôte et cela aboutit au développement d’une
maladie qui peut devenir grave.
Les techniques les mieux connues incluent notamment la cyto-adhérence et la
séquestration, la formation de rosettes et la variation antigénique.
I.2.2.1. La Séquestration
La séquestration est définie comme l’élimination des globules rouges infectés de la
circulation périphérique grâce à leur liaison à l'endothélium vasculaire des veinules post-
capillaires des tissus profonds. La surface du globule rouge infecté est recouverte
d'excroissances appelées Knobs qui sont le point de contact avec les cellules hôtes.
Une protéine unique du parasite, P. falciparum Erythrocyte Membrane Protein 1 (PfEMP1),
assure sa liaison avec divers récepteurs (Baruch et al, 1995).
PfEMP1 est codée par la grande famille des gènes variables var, comprenant environ 60 loci
polymorphes dans le génome de P. falciparum (Su et al, 1995, Gardner et al, 2002).
La séquestration du globule rouge infecté dans la micro vascularisation des organes vitaux
est un trait marquant du paludisme à P. falciparum. La séquestration massive des globules
rouges parasités dans le cerveau est considérée comme la principale cause du coma dans le
paludisme cérébral souvent rapidement mortel (Turner et al, 1994). La séquestration des
parasites dans le cerveau impliquerait des récepteurs de l’hôte tels que : le CD36 et l’ICAM1
(Intercellular Adhesion molecule 1) (Figure 3).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 26
Figure 3. Récepteurs de l’hôte impliqués dans la séquestration du globule rouge infecté par P.
falciparum.
CIDR (cysteine-rich inter-domain regions); DBL (Duffy-binding-like); PfEMP-1; RBC (red blood cell); TM
(transmembrane region). Adapté de Mackintosh et al, 2004
En outre, la séquestration des parasites dans le placenta est assurée par l'adhésion au
sulfate de chondroïtine A (CSA).
I.2.2.2.Les Rosettes
Les globules rouges infectés peuvent se lier aux globules non infectés pour former
des rosettes. Si la liaison se fait entre globules rouges infectés, on parle d’agglutination.
Agglutination et rosettes sont associées au paludisme sévère et à l'anémie (Carlson et al,
1990; Newbold et al, 1995; Rowe et al, 1995). Le récepteur du complément (CR1) a été
identifié comme un important récepteur de PfEMP1 dans la formation des rosettes (Rowe et
al, 1997).
I.2.2.3. La Variation antigénique
La variation antigénique a été découverte chez P. knowlesi (Craig et al, 2001). PfEMP1
est impliquée dans la variation antigénique du paludisme à P. falciparum. Avec environ 60
gènes var codant pour PfEMP1 et un seul gène var dominant exprimé au stade adulte du

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 27
parasite, PfEMP1 a atteint une forme de variabilité qui permet au parasite de contourner le
système immunitaire de l'hôte. De plus, les fréquentes recombinaisons et remaniements
génétiques au cours des processus de fusion et de division dans le moustique et les
érythrocytes humains peuvent entraîner une grande diversité génétique et antigénique du
parasite (Flick et al, 2004).
L'affinité d'un PfEMP1 à certains récepteurs de l'hôte pourrait déterminer la virulence du
parasite (Roberts et al, 1992). Cette hypothèse est appuyée dans le cas du paludisme
placentaire par le biais de DBL et β (Duffy Binding-Like) qui assurent l’adhésion du parasite
au tissu placentaire à travers le CSA ou les immunoglobulines non-immunes. Un autre
exemple appuyant la même hypothèse est l’existence chez le parasite 3D7 des domaines
CIDRα (Cysteine-rich Inter-domain regions) qui ont une affinité pour les récepteurs CD36
(Flick et al, 2001; Scherf et al, 2001). Ce qui revient à dire que certains parasites peuvent être
plus virulents que d'autres et des souches de parasites tels que 3D7 qui ont plus de gènes
var conservés dans leur centromère pourraient être plus virulents (Flick et al, 2004).
I.2.3. Antigènes de surface de P. falciparum
I.2.3.1. Antigène de surface du mérozoïte 1 (MSP1)
Structure et processing de MSP-1
MSP-1 est une grosse protéine qui varie en taille et en séquence d'acides aminés
dans différentes lignées parasitaires. La protéine est synthétisée au cours de la schizogonie
comme un polypeptide précurseur unique d’environ 200 kDa et exprimé à la surface du
parasite intracellulaire. La structure primaire de la protéine a été déduite de l'analyse des
séquences des gènes de plusieurs clones de P. falciparum (Tanabe et al, 1987; Miller et al,
1993). La séquence de la protéine est divisée en blocs numérotés de 1 à 17 (figure 4). Il
existe trois types de blocs : conservés, semi-conservés et variables.
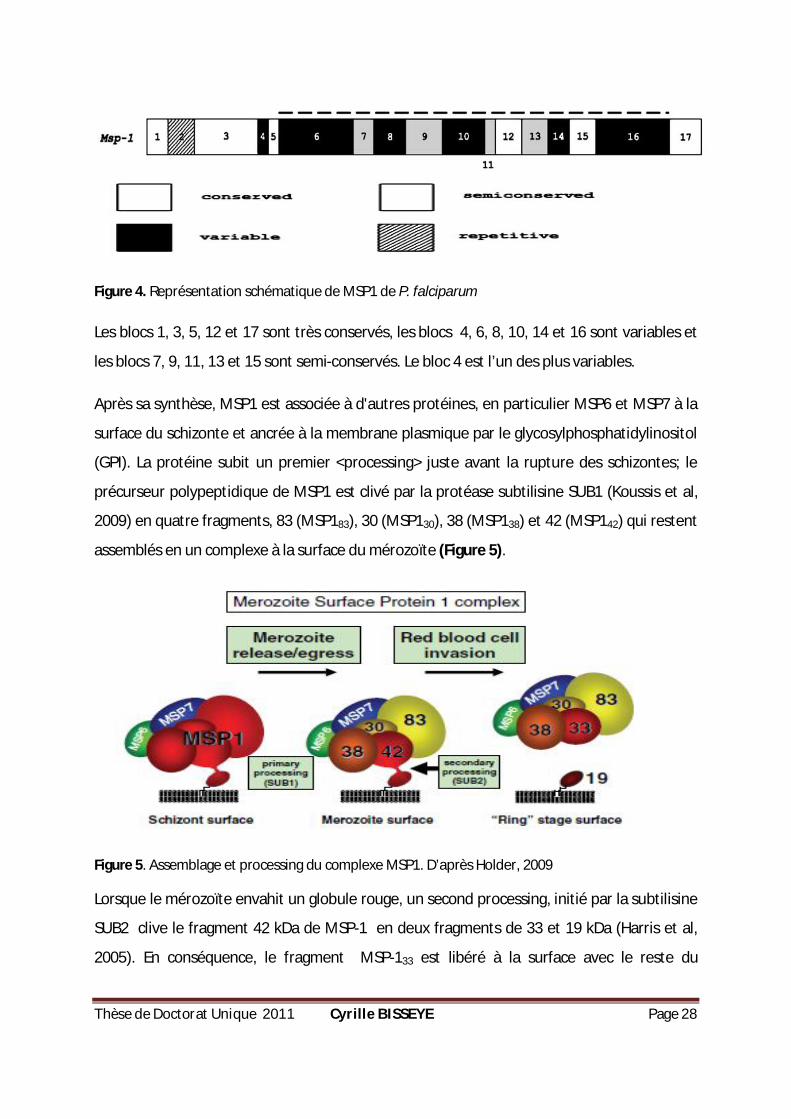
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 28
Figure 4. Représentation schématique de MSP1 de P. falciparum
Les blocs 1, 3, 5, 12 et 17 sont très conservés, les blocs 4, 6, 8, 10, 14 et 16 sont variables et
les blocs 7, 9, 11, 13 et 15 sont semi-conservés. Le bloc 4 est l’un des plus variables.
Après sa synthèse, MSP1 est associée à d'autres protéines, en particulier MSP6 et MSP7 à la
surface du schizonte et ancrée à la membrane plasmique par le glycosylphosphatidylinositol
(GPI). La protéine subit un premier <processing> juste avant la rupture des schizontes; le
précurseur polypeptidique de MSP1 est clivé par la protéase subtilisine SUB1 (Koussis et al,
2009) en quatre fragments, 83 (MSP183), 30 (MSP130), 38 (MSP138) et 42 (MSP142) qui restent
assemblés en un complexe à la surface du mérozoïte (Figure 5).
Figure 5. Assemblage et processing du complexe MSP1. D’après Holder, 2009
Lorsque le mérozoïte envahit un globule rouge, un second processing, initié par la subtilisine
SUB2 clive le fragment 42 kDa de MSP-1 en deux fragments de 33 et 19 kDa (Harris et al,
2005). En conséquence, le fragment MSP-133 est libéré à la surface avec le reste du

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 29
complexe MSP-1 et MSP119 est retenu par son ancre GPI à la surface du parasite (Blackman
et al, 1990) (Figure 5).
Réponse immunitaire protectrice dirigée contre l’extrémité C-terminal de MSP1
De nombreuses études ont montré que la région C-terminale de MSP1 est la cible des
réponses immunes protectrices. Ces réponses sont médiées par des anticorps. En effet, les
expériences d’immunisation dans le modèle d’infection de P. yoelii montrent le rôle de
certains anticorps monoclonaux et polyclonaux dirigés contre l’extrémité C-terminal de
MSP1 dans la protection d’un challenge par les parasites du stade sanguin (Ling et al, 1997;
Spencer-Valero et al, 1998). Les études in vitro confirment que les anticorps monoclonaux ou
polyclonaux spécifiques de l’extrémité C-terminal de P. falciparum peuvent inhiber l’invasion
des érythrocytes (Blackman et al, 1990; Chang et al, 1992).
De plus, la vaccination avec les protéines recombinantes de cette région de MSP1, induit
une réponse immunitaire protectrice, dans différents modèles tels que, P. yoelii (Ling et al,
1994); P. cynomolgi (Perera et al, 1998) et P. falciparum (Chang et al, 1996).
La réponse dirigée contre MSP119 induit trois classes différentes d’anticorps monoclonaux et
polyclonaux (Blackman et al, 1994; Guevara Patino et al, 1997) : anticorps inhibiteurs,
anticorps bloquants et anticorps neutres.
Les anticorps inhibiteurs sont des anticorps qui inhibent le processing secondaire de MSP1 et
inhibent ainsi l'invasion des érythrocytes. Les anticorps neutres n’inhibent ni le processing ni
l’invasion. Enfin, les anticorps bloquants n’inhibent pas le processing ou l’invasion mais
facilitent l’invasion des érythrocytes en rentrant en compétition avec les anticorps
inhibiteurs dans la liaison à l’antigène. Alors que tous les anticorps inhibiteurs se fixent à
MSP119, certains anticorps bloquants sont spécifiques d'épitopes formés à partir d'acides
aminés qui sont éloignés dans la séquence primaire de la protéine (Guevara Patino et al,
1997). La présence de ces différentes classes d'anticorps dans les sérums d’enfants
développant une immunité anti palustre en zone endémique met en exergue l'importance
de la compréhension de la fine spécificité de la liaison de ces anticorps (Nwuba et al, 2002;
Corran et al, 2004; Okech et al, 2004; Omosun et al, 2008). Il a été postulé que l'induction

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 30
d'anticorps bloquants par P. falciparum représenterait un mécanisme d'évasion du système
immunitaire de l’hôte (Holder et al, 1999).
Modifications de MSP119 visant à améliorer l’antigène
Malgré le fait que la région C-terminale de MSP1 soit la cible d'une réponse
immunitaire protectrice, les résultats d’un essai vaccinal en phase 2b utilisant cet antigène,
ont été très décevants ; les sujets vaccinés ayant produit des anticorps sans effet sur la
sensibilité à la maladie (Ogutu et al, 2009). Il est possible que les anticorps produits chez les
sujets vaccinés fussent des anticorps neutres ou bloquants.
Il existe différentes approches pour améliorer l’immunogénicité de MSP119.
Une de ces approches pour améliorer l’antigène serait d’enlever de la protéine les épitopes
d’anticorps bloquants sans affecter les épitopes des anticorps inhibiteurs; en limitant la taille
de la protéine ou par mutagenèse dirigée.
Les substitutions d’acides aminés affectant l’apprêtement et la présentation de l’antigène
sont importantes. Dans un modèle rongeur, il a été montré que la protéine sauvage MSP119
est un mauvais substrat pour les protéases dans l’apprêtement de l’antigène dans les
lysosomes des cellules dendritiques. En effet, les protéines réduites ou alkylées (ayant
perdues leurs ponts disulfures) induisent une réponse immunitaire plus rapide mais non
protectrice, en l’absence d’anticorps dirigés contre les épitopes conformationnels de la
protéine (Hensmann et al, 2004). Une étude similaire menée chez P. falciparum a permis de
mettre en évidence l’importance des ponts disulfures dans l’existence d’épitopes des
cellules T et B de la protéine (Egan et al, 1997).
Une étude plus récente confirme la très grande résistance de MSP119 aux protéases, la
protéine est à la fois présente et intacte dans la vacuole alimentaire (lysosomale) tout au
long du développement intracellulaire (Dluzewski et al. 2008).
Une autre approche possible pour améliorer l’antigénicité de MSP119 serait l'introduction
d’acides aminés qui favorisent le processing sans avoir un effet considérable sur la structure
de la protéine ou sur la présence d'épitopes des cellules B et T. En effet, l'insertion
d’asparagines supplémentaires reconnues par l'asparagine endoprotéinase des lysosomes
des cellules dendritiques (Hensmann et al, 2004) pourrait accélérer le processing de MSP119.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 31
Un certain nombre de variants MSP119 qui sont sans effets sur les anticorps inhibiteurs mais
affectent plutôt les anticorps bloquants ont été décrits (Uthaipibul et al, 2001). Plusieurs de
ces variants MSP119 construits ne se lient plus aux anticorps bloquants, tout en continuant à
se fixer aux anticorps inhibiteurs (Annexe Table 1) et il serait intéressant de voir quel est le
résultat d'une immunisation par ces protéines.
I.2.3.2. Apical membrane Antigen-1 (AMA1)
Structure et fonction d’AMA1
AMA1 de P. falciparum (PfAMA1) est localisé dans les micronèmes du mérozoïte,
organite contenant des récepteurs de l’invasion, ce qui suggère qu’AMA1 pourrait jouer un
rôle dans ce processus. La protéine est exprimée en fin de schizogonie au cours du
développement érythrocytaire asexué. PfAMA1 est synthétisée sous forme d’un précurseur
de 83 kDa (PfAMA183) qui est converti en PfAMA166 (Narum et Thomas, 1994). La protéine se
subdivise en trois domaines (DI, DII et DIII) sur la base des ponts disulfures (Hodder et al,
1996) et de sa structure cristalline (Pizarro et al, 2005) (Figure 6). La protéine est apprêtée
au stade tardif du mérozoïte ou au cours de l'invasion érythrocytaire (Howell et al, 2001).
Elle jouerait un rôle lors de la réorientation du mérozoïte au cours de laquelle, les organites
apicaux sont alignés sur la membrane des érythrocytes (Mitchell et al, 2004). PfAMA1
pourrait également être impliqué dans l'invasion des hépatocytes (Sylvie et al, 2004).
PfAMA1 jouerait un rôle dans l’invasion érythrocytaire par l’intermédiaire de son domaine
DIII (Kato et al, 2005). PfAMA1 présente de nombreux polymorphismes dans ses trois
domaines DI, DII et DIII.
Figure 6. Représentation schématique d’AMA1.
Adapté de Kato et al. (2005) TM : domaine transmembranaire; Cyt : domaine cytoplasmique

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 32
Réponses immunes naturelles dirigées contre AMA1
La plupart des personnes exposées au Plasmodium produisent des anticorps anti-
AMA1, leur prévalence augmentant avec l’âge (Polley et al, 2004; Cortes et al, 2005; Chelimo
et al, 2005; Johnson et al, 2004). Chez les individus naturellement infectés par P. falciparum
la réponse immunitaire humorale est caractérisée par la production d’IgG1 et une faible
réponse IgG3. La production d’IgG2 et IgG4 est rarement observée. Les anticorps anti-AMA1
sont essentiellement dirigés contre les domaines DI et DII de la protéine (Polley et al,
2004; Cortes et al, 2005) et ces anticorps réagissent avec plusieurs variants alléliques (Polley
et al, 2004; Cortes et al, 2005). La réponse anticorpale dirigée contre le domaine DIII de la
protéine est généralement faible mais augmente chez les adultes. Il a été montré que les IgG
dirigées contre une construction d’AMA1 contenant au moins les domaines DI et DII sont
associées à un risque réduit de paludisme clinique chez les sujets présentant une
parasitémie avant la saison de transmission (Polley et al, 2004). Les anticorps anti-AMA1
purifiés de sujets immuns inhibent aussi la croissance des parasites in vitro (Hodder et al,
2001). Dans les populations vivant en zone d’endémie palustre les réponses immunes anti-
AMA1 sont souvent beaucoup plus élevées que celles d’autres antigènes de P. falciparum
tels que MSP119, GLURP, MSP3 (Chelimo et al, 2005). La réponse immune cellulaire dirigée
contre AMA1 fait l’objet de peu d’études. Une étude suggère que la réponse cellulaire T anti-
AMA1 est de courte durée et qu’elle est associée à la protection contre l’infection
(Udhayakumar et al, 2001).
I.2.3.3. Protéine Circumsporozoite (CSP)
La protéine circumsporozoite est l'antigène qui prédomine à la surface des
sporozoïtes. La protéine est composée d'une région N-terminal qui se lie aux protéoglycanes
de sulfate d'héparine (RI), une région centrale contenant une répétition de quatre acides
aminés (Asn-Ala-Asn-Pro), et une région C-terminal contenant un domaine
thrombospondine (RII) ancrée à une GPI (Figure 7). La région centrale répétée contient
l’épitope immunodominant des cellules B. Cette région centrale induit la production
d’anticorps qui inhibent l’invasion des hépatocytes par les sporozoïtes in vitro (Gysin et al,
1984 ; Plassmeyer et al, 2009).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 33
Les anticorps dirigés contre la région centrale répétée sont associés à la protection contre
l’infection par P. falciparum (Guinovart et al, 2009 ; Aponte et al, 2007) mais ne protègent
pas contre la maladie (Alonso et al, 2004 ; Bejon et al, 2008). Les anticorps monoclonaux
dirigés contre la thrombospondine bloquent également l'invasion des hépatocytes par les
sporozoïtes, mais à un degré moindre que les anticorps de la région centrale répétée
(Plassmeyer et al, 2009). La CSP induit aussi une réponse immune cellulaire T. En effet, elle
comporte trois épitopes de lymphocytes T connus: un épitope des cellules T CD4+ très
variable en amont du domaine de la thrombospondine (Guttinger et al, 1988), un épitope de
cellules T CD8+ très variable à l’intérieur du domaine de la thrombospondine (Kumar et al,
1988), et un épitope"universel" conservé des cellules T CD4+ à l'extrémité C-terminal de la
protéine (Sinigaglia et al, 1988).
Figure 7. Représentation schématique de CSP et du vaccin RTS, S.
Adapté de Crompton et al, 2010
I.3. La maladie
I.3.1. Caractéristiques cliniques
Les premiers symptômes du paludisme lors d’une primo-infection peuvent être
observés 8 à 10 jours après que le parasite ait été introduit dans l'hôte humain par la piqure
du moustique femelle. La maladie se manifeste essentiellement par des fièvres qui
deviennent peu à peu récurrentes. Elles sont dues à la libération de toxines du parasite lors
de la rupture des érythrocytes infectés. Différentes types de fièvres ont été décrites en
fonction de l’espèce plasmodiale responsable de l’infection. Certaines formes de paludisme
présentent des fièvres quotidiennes (fièvre semi-tierce) tandis que d'autres sont
responsables des fièvres tous les deux jours (fièvre tierce) et il y a encore d'autres formes qui

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 34
présentent des fièvres tous les trois jours (fièvre quarte). Le paludisme à P. falciparum
présente deux formes de fièvre, tierce et semi-tierce, tandis que P. vivax et P. ovale
présentent une forme de paludisme à fièvre tierce. Le paludisme à P. malariae est
responsable d’une forme de paludisme à fièvre quarte.
P. falciparum est responsable de deux formes de paludisme : le paludisme simple et le
paludisme grave (Berendt et al, 1992). Dans les zones endémiques, le paludisme simple se
produit chez les personnes semi-immunes alors que le paludisme grave et celui associé à la
grossesse affectent les personnes non-immunes et notamment les enfants de moins de
cinq ans et les femmes enceintes (les primipares principalement).
I.3.1.1. Le paludisme simple
Les principaux symptômes cliniques de l’accès palustre simple sont : la fièvre, les
frissons et sueurs, céphalées, vomissements, diarrhée, l'anémie, la jaunisse et souvent la
splénomégalie. Parfois les enfants peuvent présenter des expériences de convulsions.
I.3.1.2. Paludisme grave
La définition du paludisme grave est difficile, il résulte probablement d'une
combinaison de facteurs spécifiques du parasite : adhérence, séquestration dans le système
vasculaire et libération de molécules bioactives ; mais aussi de la réponse inflammatoire de
l'hôte avec notamment la production de cytokines, chémokines et d’infiltrats cellulaires
(Mackintosh et al, 2004).
Deux syndromes majeurs caractérisent le paludisme sévère: l’anémie sévère et le paludisme
cérébral. La pathogénie du paludisme cérébral est attribuée à la cytoadhérence des
érythrocytes infectés aux cellules endothéliales dans les micros vaisseaux du cerveau
entrainant l’obstruction puis la rupture des vaisseaux sanguins. Un des mécanismes de la
pathogenèse du paludisme grave est l'acidose métabolique qui entraîne la détresse
respiratoire. Différentes études ont montré que l’acidose métabolique est un bon prédicteur
de la mortalité due au paludisme sévère (Taylor et al, 1993; Marsh et al, 1995; English et al,
1996). La pathogénie de l'acidose métabolique est mal connue.
Les cytokines pro-inflammatoires jouent un rôle central dans les manifestations cliniques du
paludisme cérébral. L’interleukine 10 (IL-10) aurait un rôle protecteur dans le paludisme

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 35
cérébral (Clark et Cowden, 2003). Par contre, une cytokine comme le facteur de nécrose
tumorale alpha (TNF-) est plutôt associée à la sensibilité au paludisme cérébral (Grau et al,
1989; Kwiatkowski et al, 1990; Brown et al, 1999). Le rôle de l’interféron gamma (IFN-) est
ambigu, elle est associée à la fois à la pathogenèse et à la protection contre le paludisme
chez l'homme (Day et al, 1999; Luty et al, 1999). Un autre mécanisme impliqué dans la
pathogenèse du paludisme cérébral est l’induction du monoxyde d’azote (NO), il serait
responsable non seulement d’une hypotension intracrânienne due à la dilatation excessive
des vaisseaux sanguins (vasodilatation) mais aussi à une neurotransmission aberrante (Clark
et al, 1994).
I.3.2. Paludisme et grossesse
Plus de 50 millions de femmes sont exposées au risque de paludisme pendant la
grossesse chaque année. Le paludisme est responsable d’une morbidité maternelle et
infantile substantielle, causant de 75.000 à 200.000 décès de nourrissons chaque année
(Steketee et al, 2001; Desai et al, 2007).
Les effets du paludisme pendant la grossesse sur la santé maternelle, néonatale et infantile
sont présentés dans la figure 8. Les infections par P. falciparum et P. vivax pendant la
grossesse entraînent l'anémie maternelle et un faible poids des nouveau-nés en relation
avec les accouchements prématurés et un retard de croissance fœtale (Nosten et al, 1999).
Les femmes enceintes sont plus sensibles que les femmes non enceintes au paludisme, et
cette sensibilité est plus grande chez les primipares comparativement aux multipares.
Le paludisme pendant la grossesse se manifeste par une parasitémie plus élevée dans
l’espace intervilleux du placenta que dans la circulation périphérique (Rogerson et al, 2007);
une absence de trophozoïtes et de schizontes matures dans le sang périphérique, car
séquestrés dans le placenta (Beeson et al, 2002). Ainsi, la relation entre parasitémie
placentaire élevée et accouchement prématuré a été établie (Menendez et al, 2000).
L’infection placentaire est souvent accompagnée d’une infiltration intervilleuse de
monocytes et de macrophages, certains contenant l’hémozoïne (pigment malarique ou
hémoglobine oxydée).
Une densité élevée de monocytes dans l’espace intervilleux du placenta est associée au
faible de poids chez le nouveau-né et à l’anémie (Ordi et al, 1998; Rogerson et al, 2003).
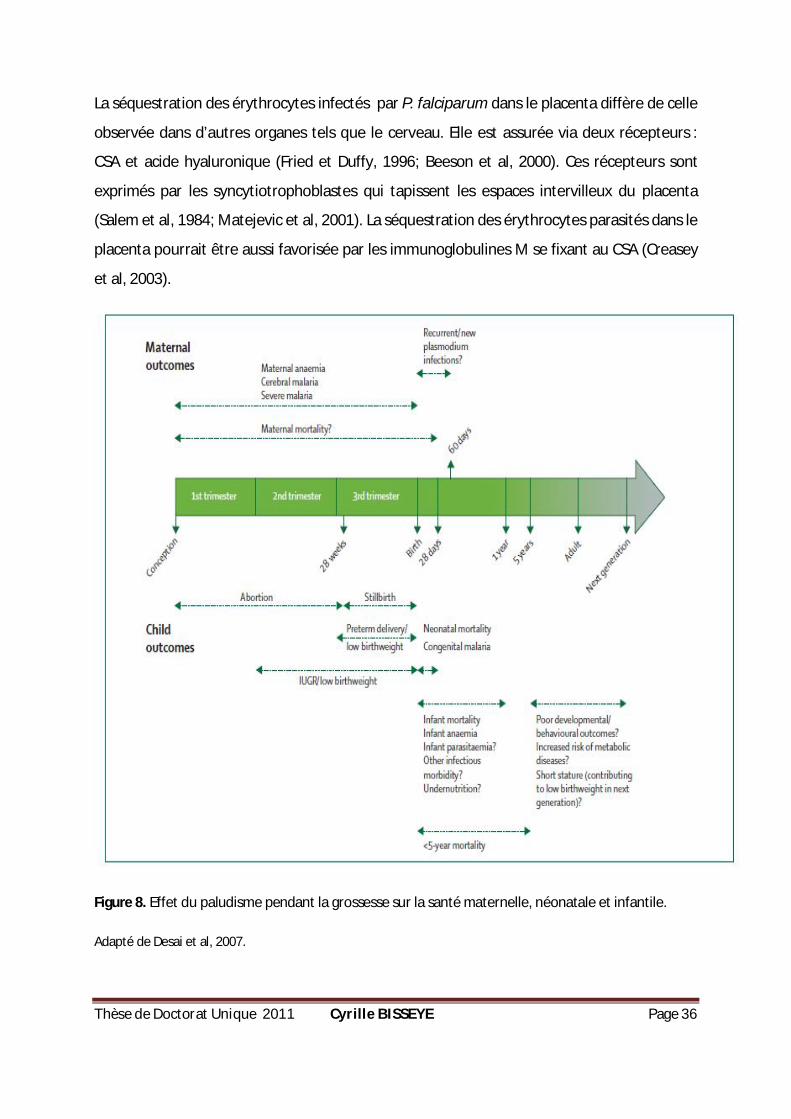
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 36
La séquestration des érythrocytes infectés par P. falciparum dans le placenta diffère de celle
observée dans d’autres organes tels que le cerveau. Elle est assurée via deux récepteurs :
CSA et acide hyaluronique (Fried et Duffy, 1996; Beeson et al, 2000). Ces récepteurs sont
exprimés par les syncytiotrophoblastes qui tapissent les espaces intervilleux du placenta
(Salem et al, 1984; Matejevic et al, 2001). La séquestration des érythrocytes parasités dans le
placenta pourrait être aussi favorisée par les immunoglobulines M se fixant au CSA (Creasey
et al, 2003).
Figure 8. Effet du paludisme pendant la grossesse sur la santé maternelle, néonatale et infantile.
Adapté de Desai et al, 2007.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 37
La formation des rosettes qui est importante dans la pathogénèse du paludisme cérébral,
joue un rôle mineur dans l’infection du placenta (Maubert et al, 1998; Rogerson et al, 2000).
Le ligand de P. falciparum au CSA est la PfEMP1 une protéine codée par la famille des gènes
multiples var. Un gène var particulier appelé var2csa est exprimé dans la plupart des isolats
se liant au placenta (Salanti et al, 2003; Duffy et al, 2005).
I.3.3. Epidémiologie du paludisme
Les facteurs environnementaux, l’intensité de la transmission, la virulence du parasite
et les différents traits génétiques de l’hôte confèrent une complexité au paludisme
(Mackinnon et al, 2005). Les études épidémiologiques montrent que, la sensibilité au
paludisme est très variable entre les individus d’une population. Ainsi, charge parasitaire,
incidence et gravité de la maladie (Greenwood et al, 1991); ampleur et type de réponses
immunitaires dirigées contre les antigènes du parasite diffèrent selon les individus (Good et
al, 1988; Troye-Blomberg et al, 1989; Riley et al, 1990; Modiano et al, 1996).
Les manifestations cliniques du paludisme sont liées à l’âge des individus en zone endémique
(Marsh, 1992 ; Brewster et al, 1990). Les enfants de moins de cinq ans souffrent du
paludisme cérébral alors que les adultes souffrent du paludisme simple. Le risque de décès
est aussi plus élevé chez les enfants que chez les adultes vivant en zone endémique. Les
facteurs génétiques de l’hôte affectent aussi l’évolution de la maladie. Parmi ces facteurs
génétiques, les polymorphismes de l’hémoglobine jouent un rôle essentiel.
Hémoglobines S et C (HbS et HbC)
Les polymorphismes génétiques de HbS et HbC assurent la protection contre les
formes simples et sévères du paludisme. En effet, les individus hétérozygotes HbAS
présentent de faibles parasitémies (Allison, 1954). Plus de 70% de porteurs HbAS sont
protégés contre le paludisme sévère alors que 60% sont protégés contre le paludisme
simple (Allison, 1954; Flint et al, 1986).
HbC est aussi associée à la protection contre le paludisme en Afrique de l’ouest (Agarwal et
al, 2000; Modiano et al, 2001). La protection est d'environ 90% chez les homozygotes HbCC
et de 30% chez les hétérozygotes HbAC (Modiano et al, 2001). Des résultats contrastés sur la

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 38
protection contre le paludisme conférée par HbC ont été signalés au Mali (Guinet et al,
1997). D’autres études menées au Ghana (Mockenhaupt et al, 2004) et au Burkina Faso
(Rihet et al, 2004) ont confirmé la protection contre le paludisme conférée par HbC.
HbAS joue un rôle dans la mise en place d’une immunité acquise. Une étude réalisée au
Kenya a montré l’acquisition accélérée de l'immunité contre le paludisme simple chez les
enfants âgés de moins de 10 ans porteurs de HbAS (Williams et al, 2005).
Alpha et Béta-Thalassémies
Deux autres polymorphismes de l’hémoglobine qui affectent la susceptibilité au
paludisme sont les α et β-Thalassémies. et β-thalassémies sont associées à la protection
contre le paludisme (Allen et al, 1997; Udomsangpetch et al, 1993). L’-thalassémie
protégerait contre le paludisme en limitant les manifestations cliniques de la maladie plutôt
que la multiplication du parasite. Par contre, la β-thalassémie protégerait contre le
paludisme sévère en altérant la cytoadhérence et la formation de rosettes par les
érythrocytes β-Thalassémiques infectés (Udomsangpetch et al, 1993).
Déficience en Glucose 6 phosphate déshydrogénase (G6PD)
Un autre type de polymorphismes génétiques de l’hôte influençant la sensibilité au
paludisme est la déficience en G6PD. Le variant G6PD A- (A376G et G202A) est le plus
fréquent en Afrique sub-saharienne. La déficience en G6PD a été associée à la protection
contre le paludisme grave chez les enfants africains (Allison and Clyde, 1961). Cette
protection concerne les femmes hétérozygotes (Bienzle et al, 1972). Cependant, une autre
étude a montré que la déficience en G6PD protégeait aussi bien les hommes hémizygotes
que les femmes homozygotes et hétérozygotes (Ruwende et al, 1995). Une étude plus
récente menée au Mali confirme, que la protection conférée contre le paludisme par la
G6PD ne concerne que les hommes hémizygotes et les femmes homozygotes (Guindo et al,
2007).
Antigène Duffy
Le polymorphisme de la membrane des érythrocytes de l’hôte joue un rôle dans la
susceptibilité ou la protection contre le paludisme. La fréquence élevée du polymorphisme

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 39
de l’antigène Duffy prévenant l’invasion des globules rouges par P. vivax dans la population
ouest africaine explique l’absence de P. vivax dans cette région (Miller et al, 1976).
Glycophorines A et B
Chez l’homme, les globules rouges déficients en Glycophorines A et B (GYPA et GYPB)
sont résistants à l'invasion par P. falciparum (Bejon et al, 2007). EBA175, protéine du
micronème présente sur le mérozoïte a des domaines Duffy Binding-Like (DBL) spécifiques
qui reconnaissent les résidus d'acide sialique sur la molécule de GYPA, ainsi l'absence de
GYPA et GYPB affecterait la reconnaissance de la protéine par le globule rouge. L’absence de
l'exon 3 au niveau de la GYPC est responsable du groupe sanguin Gerbich que l’on trouve
chez 50% d’hommes vivant sur les côtes de Papouasie-Nouvelle Guinée. Chez les individus
Gerbich négatifs, la protéine EBA140 ne se fixe pas aux globules rouges et l'invasion de P.
falciparum est bloquée (Maier et al, 2003).
Groupes sanguins ABO
Le polymorphisme du complexe des groupes sanguins ABO joue un rôle dans
l’évolution du paludisme. Il a été montré que les individus du groupe sanguin O étaient plus
protégés contre le paludisme grave comparativement aux individus des autres groupes
sanguins. Une faible séquestration et un taux réduit de rosettes par les érythrocytes du
groupe sanguin O infectés par P. falciparum assureraient cette protection (Rowe et al, 2007).
Le groupe sanguin A, a quant à lui été associé au paludisme grave (Rowe et al, 2007; Fischer
and Boone, 1998).
Récepteur du Complément (CR1)
Le CR1 est une protéine immunorégulatrice présente à la surface des érythrocytes.
Certaines populations présentent une déficience en CR1. C’est le cas des populations
mélanésiennes. La déficience en CR1 induit une réduction de la formation des rosettes par
les érythrocytes infectés par P. falciparum (Rowe et al, 1997). Elle a été associée à la
protection contre le paludisme grave dans les zones endémiques de Papouasie-Nouvelle
Guinée (Cockburn et al, 2004). Une étude réalisée au Kenya associe la déficience en CR1 à la
protection contre l’anémie sévère (Waitumbi et al, 2000).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 40
Un autre facteur qui complique le pronostic du paludisme est le développement de la
résistance des parasites aux antimalariques.
Cette résistance est responsable de l’échec thérapeutique des médicaments de première
intention dans le traitement du paludisme simple dans différentes régions du monde.
I.3.4. Résistance des parasites aux médicaments antipaludiques
La résistance à un médicament antipaludique est la capacité du parasite à survivre et
à se multiplier dans l’organisme de l’hôte en présence de la dose thérapeutique de ce
médicament. La résistance aux antipaludiques concerne essentiellement P. falciparum et P.
vivax. Elle n’a pas été décrite avec P. ovale et P. malariae. La résistance de P. falciparum aux
antipaludiques est celle qui est la plus étudiée en Afrique sub-saharienne.
La résistance aux antipaludiques se produirait dans les zones de faible ou de haute
transmission de paludisme (Hastings et D’Alessandro, 2000; Molyneux et al, 1999). En effet,
la résistance s’est développée en premier dans les zones de faible transmission (Thaïlande et
Brésil) et elle demeure plus fréquente dans ces zones que dans celles de transmission plus
élevée, appuyant l'hypothèse de la faible transmission.
La résistance concerne à des degrés variables la plupart des antipaludiques actuels sur le
marché compliquant l’efficacité du traitement contre le paludisme.
I.3.4.1. Résistance à la chloroquine
La résistance à la chloroquine est apparue à la fin des années 50 à la frontière entre la
Thaïlande et le Cambodge et en Colombie (Wernsdorfer, 1994). En Afrique, la résistance à la
chloroquine s’est propagée d’Est en Ouest. Les polymorphismes de deux gènes de P.
falciparum sont responsables de cette résistance: Pfcrt (P. falciparum chloroquine resistance
transporter) (Wellems et al, 1991) et Pfmdr1 (P. falciparum multidrug resistance 1). Le gène
Pfcrt est situé sur le chromosome 7, et code pour pfCRT, une protéine membranaire de
transport vacuolaire.
De nombreux polymorphismes de Pfcrt associés à la résistance à la chloroquine ont été
identifiés. La mutation sur le codon 76 (Thr→Lys) de Pfcrt est essentielle dans la résistance à

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 41
la chloroquine. La mutation Thr76Lys est associée à une résistance absolue à la chloroquine
in vitro (Djimdé et al, 2001; Fidock et al, 2000).
De nombreuses études cliniques réalisées dans diverses zones géographiques ont confirmé
l'association entre la mutation Thr76Lys et la résistance à la chloroquine notamment en
Afrique , au Mali (Djimdé et al, 2001), au Cameroun (Basco et Ringwald, 2001), au Soudan
(Babiker et al, 2001), et au Mozambique (Mayor et al, 2001) ; en Asie, au Laos (Pillai et al,
2001) et en Thaïlande (Cheng et al, 2001); ainsi qu’en Amérique du Sud, au Brésil (Vieira et
al, 2001).
Pfmdr1 est situé sur le chromosome 5 et code pour une glycoprotéine. Le polymorphisme de
Pfmdr1 est aussi associé à la résistance à la chloroquine. Le changement d’acide aminé sur le
codon 86 (Asp86Tyr) de Pfmdr1 a été associé à la résistance à la chloroquine dans certaines
études in vitro ou cliniques : en Gambie (Von Seidlein et al, 1997), en Ouganda (Flueck et al,
2000), en Thaïlande (Price et al, 1999) et au Brésil (Zalis et al, 1998). En revanche, ce
polymorphisme Asp86Tyr n’était pas associé à la résistance à la chloroquine dans d’autres
études : en Ouganda (Dorsey et al, 2001), au Laos (Pillai et al, 2001), en Thaïlande (Chaiyaroj
et al, 1999) et au Brésil (Povoa et al, 1998). Plusieurs autres polymorphismes de pfmdr1
notamment, Phe184, Cys1034, Asp1042 et Tyr1246, sont impliqués à des degrés divers dans
la résistance à la chloroquine.
I.3.4.2. Résistance à la pyrimethamine-sulfadoxine
La résistance à la pyrimethamine-sulfadoxine est apparue à la frontière entre la
Thaïlande et le Cambodge dans le milieu des années 60. En Afrique, la sensibilité de la
pyrimethamine- sulfadoxine a commencé à baisser dans les années 1980. Cette résistance
s’est répandue plus rapidement à l’Est qu’à l’Ouest du continent. La résistance in vitro à la
pyrimethamine-sulfadoxine a été reportée dans plusieurs pays d’Afrique subsaharienne,
variant de 13 à 30 % (Alin, 1997; Benito et al, 1995; Philipps et al, 1998; Landgraf et al, 1994).
De toutes les résistances aux antipaludiques, la base moléculaire de la résistance à la
pyrimethamine-sulfadoxine est la mieux caractérisée. Les mutations spécifiques de P.
falciparum qui conduisent à la résistance à la pyrimethamine et à la sulfadoxine ont été
identifiées. Sulfadoxine et pyrimethamine agissent en synergie.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 42
La sulfadoxine inhibe la dihydropteroate synthétase (dhps) et la pyrimethamine, la
dihydrofolate réductase (dhfr) du parasite. Ces deux enzymes sont impliquées dans la
synthèse des folates.
Les mutations sur cinq codons du gène dhps sont associées à la résistance à la sulfadoxine en
diminuant l'affinité de liaison de l'enzyme au médicament: ce sont les codons 436 (Ser→ Ala
ou Phe), 437 (Ala→Gly), 540 (Lys→Glu), 581 (Ala→Gly) et 613 (Ala→Ser ou Thr).
La résistance à la pyrimethamine est due à des mutations ponctuelles spécifiques dans le
gène dhfr. Il en résulte une réduction de l’affinité de liaison entre dhfr et la pyrimethamine.
Ces mutations ont lieu sur les codons : 16 (Ala→Val), 51 (Asn→Ile), 59 (Cys→Arg), 108
(Ser→Asn ou Thr) et 164 (Ile→Leu).
La mutation sur le codon 108 (Ser→Asn) est connue pour être la mutation clé de la
résistance à la pyrimethamine. Les mutations ponctuelles supplémentaires dans trois autres
codons, Ile51, Arg59, et Leu164, sont connues pour augmenter progressivement le degré de
la résistance.
I.3.4.3. Résistance à la quinine
La résistance à la quinine a été documentée au milieu des années 60 à la frontière de
La Thaïlande et du Cambodge.
Les données des tests in vitro, ont montré que cette résistance est moins fréquente en
Amérique du sud (Zalis et al, 1998) et en Afrique (Jelinek et al, 2001). La quinine est réservée
comme traitement de deuxième ou troisième ligne et est utilisée dans les cas de paludisme
grave. Dans une étude menée au Brésil les mutations de Pfmdr1 (Asn86, Phe184, Cys1034,
Asp1042 et Tyr1246) ont été associées à la réduction de la sensibilité à la quinine (Zalis et al,
1998). En Gambie une faible association entre la mutation Tyr86 de Pfmdr1 et la réduction
de la sensibilité à la quinine a été observée (Duraisingh et al, 2000).
I.3.4.4. La résistance à la méfloquine
La résistance à la méfloquine a été observée près de la frontière de la Thaïlande et du
Cambodge à la fin des années 80 (Wongsrichanalai et al, 2001).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 43
Des études in vitro ont suggéré la présence de souches de P. falciparum ayant une faible
sensibilité à la méfloquine en Afrique (Jelinek et al, 2001; Brasseur et al, 1992). Bien que
rares, quelques résistances cliniques à la méfloquine ont été décrites en Afrique (Witkowski
et al, 2010; Brasseur et al, 1990; Gay et al, 1990). Les résultats contrastés ont été obtenus
sur le rôle du polymorphisme de Pfmdr1 dans la résistance à la méfloquine. Des études
réalisées en Thaïlande (Price et al, 1999; Wilson et al, 1993) et récemment en Afrique de
l’Ouest (Witkowski et al, 2010) ont suggéré l’association entre le nombre élevé de copies de
Pfmdr1 et la résistance à la méfloquine.
Une autre forme de résistance de P. falciparum est la multi résistance aux antipaludiques. La
multi résistance a été définie comme étant une résistance à plus de deux composés
antipaludiques opérationnels appartenant à des classes chimiques différentes (Wernsdorfer,
1994). Du fait de cette multi résistance, et de la résistance à la plupart des médicaments de
première ligne tels que la chloroquine, la pyrimethamine-sulfadoxine et dans une moindre
mesure la méfloquine; de nouveaux schémas thérapeutiques ont été mis en place : ce sont
des associations médicamenteuses impliquant souvent l’arthémisinine. Aucune résistance à
l’arthémisinine n’a encore été formellement documentée en Afrique subsaharienne.
I.4. Paludisme à P. falciparum et vaccination
L’outil clé pour le contrôle, l'élimination, voire même l'éradication du paludisme, en
plus des médicaments antipaludiques et de la lutte anti vectorielle, est un vaccin efficace. À
ce jour, aucun vaccin efficace contre le paludisme n’est disponible.
Le développement d'un vaccin contre le paludisme est entravé par la complexité du parasite
et de son cycle de vie (Gardner et al, 2002; Florens et al, 2002), une très grande variation
antigénique (Scherf et al, 2008), et une compréhension limitée des interactions entre P.
falciparum et le système immunitaire de l’hôte humain (Langhorne et al, 2008).
Dans l’infection par P. falciparum, Le développement d'un vaccin est possible parce que
l'infection induit une immunité clinique.
Dans les zones hyper endémiques à P. falciparum, l'immunité contre le paludisme sévère est
acquise dès la petite enfance, alors que l'immunité contre une maladie bénigne n'est pas
acquise avant la fin de l'adolescence (Marsh et al, 2006, Gupta et al, 1999).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 44
L’immunité est acquise plus rapidement par les adultes que par les enfants (Baird et al, 1991;
Baird et al, 1993), cette immunité est rarement complète et des épisodes occasionnels de
paludisme peuvent se produire (Marsh et al, 2006).
Une meilleure compréhension des mécanismes immunitaires qui assurent la protection chez
l’adulte et l’identification de leurs cibles sur P. falciparum, pourraient servir au
développement d’un vaccin qui induirait chez l’enfant une immunité mimant celle de
l’adulte.
Différents vaccins contre le paludisme sont testés chez l’homme. Nous nous limiterons à la
présentation des vaccins des stades pré-érythrocytaire et érythrocytaire.
I.4.1. Vaccins pré-érythrocytaires
L’immunité du stade pré-érythrocytaire ne semble pas acquise naturellement dans
les zones endémiques où les adultes sont souvent infectés par les stades asexués du
parasite.
L'idée d'un vaccin pré-érythrocytaire a pris forme avec l'observation historique faite par Ruth
Nussenzweig que la vaccination de souris avec des sporozoïtes irradiés les protégeait contre
le paludisme (Nussenzweig et al, 1967). En outre, cette protection pouvait être obtenue par
l’immunisation avec la CSP (Circumsporozoite protein) seule (Nussenzweig et Nussenzweig,
1986).
Le clonage de la CSP de P. falciparum (Dame et al, 1984) a accéléré la mise au point du
vaccin RTS, S. Le RTS, S se compose de particules de l'antigène de surface de l’hépatite B
(HBsAg) fusionnés au segment CSP dans l’adjuvant AS01 (Casares et al, 2010, Gordon et
al, 1995). Le vaccin RTS,S/AS02A est composé de deux polypeptides (RTS and S)
provenant de la circumsporozoite protein (CSP) de P. falciparum, exprimés dans des
cellules de la levure Saccharomyces cerevisiae (la levure de boulanger) et fusionné à
l’HBsAg, (le vaccin contre l’hépatite B);
• Il est présenté sous une forme particulaire associée à un adjuvant, AS02A, une
émulsion d’huile dans de l’eau avec deux immunostimulants : le monophosphoryl
lipid A et le QS21dérivé de la saponine;
• Il induit une forte réponse anticorps et il stimule l’immunité cellulaire de type 1.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 45
RTS, S est le vaccin du stade pré-érythrocytaire le plus avancé en ce qui concerne les essais
cliniques.
Les essais cliniques en phase II ont montré que 30 à 50% d’adultes naïfs vaccinés avec RTS, S
étaient protégés contre un challenge par des moustiques infestés par un clone homologue
de P. falciparum (Gordon et al, 1995; Stoute et al, 1997; Kester et al, 2001; Kester et al,
2008, Kester et al, 2009).
Les essais cliniques en phase II menés en zones endémiques ont montré qu’environ 30 à 50%
d’enfants et nourrissons vaccinés avec RTS, S étaient protégés contre le paludisme clinique
(Alonso et al, 2004 ; Sacarlal et al, 2009 ; Abdulla et al, 2008 ; Bejon et al, 2008), mais que la
protection induite par la vaccination était de courte durée. Aussi, ce vaccin, induit des
anticorps qui sont corrélés à la protection contre l'infection par P. falciparum (Guinovart et
al, 2009 ; Aponte et al, 2007) mais qui n’assurent pas la protection contre la maladie clinique
(Alonso et al, 2004 ; Bejon et al, 2008 ; Guinovart et al, 2009).
Le mécanisme par lequel un vaccin qui cible le sporozoïte et le stade hépatique du parasite,
protège contre le stade sanguin de la maladie reste inconnu.
Il est probable que le vaccin RTS, S induise une protection contre le paludisme clinique en
réduisant temporairement le nombre de mérozoïtes qui sortent du foie; ce qui conduirait
l’hôte à une exposition prolongée aux parasites du stade sanguin à des niveaux sub-cliniques
qui, stimulerait à son tour l'immunité acquise (Guinovart et al, 2009).
I.4.2. Vaccins érythrocytaires
Les candidats vaccins du stade érythrocytaire comprennent essentiellement les
antigènes exprimés à la surface du mérozoïte.
À l’heure actuelle, quelques antigènes du stade sanguin du parasite sont à l’étape clinique de
développement vaccinal. Il s'agit notamment d’AMA1 (Sagara et al, 2009), de EBA175 (El
Sahly et al, 2010), de GLURP (Glutamate rich protein) (Esen et al, 2009; Hermsen et al, 2007),
de MSP1 (Ogutu et al, 2009 ), de MSP2 (Genton et al, 2002), de MSP3 (Esen et al, 2009;
Audran et al, 2005; Sirima et al, 2009; Druilhe et al, 2005), et de SERA5 (serine repeat
antigen 5) (Horii et al, 2010), qui sont tous fortement exprimés à la surface du mérozoïte.
Contrairement au stade pré-érythrocytaire avec le vaccin RTS, S actuellement testé en phase

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 46
III, aucun vaccin du stade érythrocytaire n’a atteint à l’heure actuelle la phase III de
développement.
Les récents essais en phase II des candidats vaccins du stade érythrocytaire les plus avancés,
AMA1 et MSP1, n'ont pas démontré d’efficacité chez les enfants en Afrique (Sagara et al,
2009; Ogutu et al, 2009).
Cependant, sont en cours, les efforts visant à l’amélioration des vaccins AMA1 et MSP1, soit
par l’adjonction de nouveaux adjuvants (Ellis et al, 2010; Sagara et al, 2009), soit par le
boosting dans des vecteurs viraux (Hill et al, 2010), ou par leur combinaison (Malkin et al,
2008). L’obstacle majeur pour la réalisation d’un vaccin du stade sanguin du parasite
demeure, sa grande diversité génétique découlant de la pression sélective exercée par la
réponse immunitaire de l'hôte (Weedall et al, 2010; Takala et al, 2009).
I.5. Immunité antipalustre
I.5.1. Mécanismes effecteurs de protection contre le paludisme
Les êtres humains vivant dans les zones endémiques développent une immunité
contre le paludisme. En effet, les enfants âgés de plus de cinq ans et les adultes sont
résistants au paludisme grave et à la mortalité, tout en demeurant sensibles à l'infection
(Marsh, 1992). L’immunité anti palustre se développe dans tous les stades du cycle
parasitaire et notamment aux stades pré-érythrocytaire et érythrocytaire.
I.5.1.1 Immunité au stade pré-érythrocytaire
L’immunité au stade pré-érythrocytaire du parasite pourrait être dirigée contre le
sporozoïte libre, du site de son injection dans la peau à son entrée dans l'hépatocyte, ou
sous sa forme de schizonte intra-hépatique. Bien que les humains vivant dans les zones
endémiques soient exposés aux sporozoïtes à intervalles réguliers, il est peu probable que
l’immunité pré-érythrocytaire y soit acquise naturellement.
Chez l’homme et dans les modèles expérimentaux d’infection, Il a été montré que l’injection
de sporozoïtes atténués par irradiation entraîne une résistance complète contre l’infection
pendant une durée variable (Nardin et al, 1999).
Il n’est pas encore compris pourquoi la vaccination avec les sporozoïtes atténués provoque
une plus forte réponse immune que celle induite par l’infection naturelle.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 47
La différence pourrait cependant s’expliquer d’une part par le grand nombre de sporozoïtes
utilisés (des centaines ou des milliers par la vaccination par rapport à 1 à 100 sporozoïtes par
piqûre dans les infections naturelles) (Beier et al, 1991; Hoffman et al, 2002) et d’autre part,
par la persistance de sporozoïtes atténués dans le foie (Scheller et al, 1995) et enfin, par
l'induction de l’apoptose d’hépatocytes observée en présence de sporozoïtes atténués, mais
absente en présence de sporozoïtes normaux, permettant l'exposition d’antigènes au
système immunitaire (Leiriao et al, 2005; James, 2005).
La protection contre le stade pré-érythrocytaire du parasite est assurée par les anticorps. En
effet, les anticorps dirigés contre le sporozoïte permettraient de protéger, soit par
opsonisation en entraînant l’élimination du parasite avant d'atteindre l'hépatocyte, ou bien
en interférant avec l'invasion des hépatocytes. La réponse humorale anti-CSP est dirigée
contre une région immunodominante composée d'une séquence répétée d’acides aminés
(Asn-Ala-Asn-Pro) (Esposito et al, 1988). Les anticorps anti-CSP inhibent aussi l’invasion des
hépatocytes par les sporozoïtes in vitro (Pasquetto et al, 1997; Silvie et al, 2004).
L'immunité au stade pré-érythrocytaire est assurée aussi par une réponse immunitaire à
médiation cellulaire (Tsuji et al, 2003). Les modèles murins ont permis une meilleure
compréhension des mécanismes effecteurs mis en jeu, en révélant leur complexité.
Doolan et Hoffman ont montré au moins cinq mécanismes de protection distincts
avec les lignées de souris consanguines d’origines génétiques diverses (Doolan et Hoffman,
2000).
Chez certaines souris (C57BL/6) la protection contre la maladie était associée aux cellules T
CD4+ et NK. Dans la plupart des cas, la protection dépendait des cellules T CD8+. La
protection assurée par les cellules T CD8+ ne dépendait pas de la voie cytotoxique classique
comprenant, la synthèse de perforine et de granzyme, mais plutôt par l'induction de
cascades cellulaires et cytokiniques incluant : les cellules T CD4+ ou NK et les cytokines IL-12
et IFN-; déclenchant la production d'effecteurs finaux tels que le NO.
La complexité de la réponse cellulaire décrite chez les souris existe aussi chez l’homme, avec
un mécanisme de protection différent. Dans une étude faite en Gambie, Reece et al. (2004),
ont montré que la sécrétion de l’IFN- par les cellules T CD4+ en réponse à une séquence

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 48
conservée de CSP, était associée à la fois à la protection contre l’infection et contre la
maladie.
I.5.1.2. Immunité au stade érythrocytaire
Le stade érythrocytaire du parasite est marqué par l’invasion des globules rouges.
C’est une étape clé dans l’établissement de l’infection palustre. Elle est susceptible d’être
aussi une importante cible pour les réponses immunes protectrices. La preuve la plus directe
que les anticorps sont des médiateurs importants de l'immunité palustre provient d'études
de transfert passif d'anticorps dans lesquelles les sérums d’adultes immuns ont été utilisés
avec succès pour traiter les patients atteints de paludisme grave (Cohen et al, 1961;
Sabchareon et al, 1991).
Les anticorps pourraient empêcher l'invasion des érythrocytes par les mérozoïtes par
différents mécanismes : opsonisation favorisant leur phagocytose par les macrophages,
destruction par le complément, inhibition du processing des protéines d'invasion ou blocage
de leurs sites de liaison sur les érythrocytes.
En outre, les anticorps cytophiliques (IgG1 et IgG3 chez l'homme) peuvent en collaboration
avec les monocytes détruire les globules rouges infectés par le mécanisme d’immunité
cellulaire dépendante d’anticorps (ADCI).
Osier et al, (2008) ont montré qu’une forte réponse humorale était associée à la protection
contre le paludisme et que les associations les plus fortes étaient obtenues en combinant les
réponses dirigées contre MSP2 et MSP3.
Le rôle des anticorps dirigés contre les antigènes du mérozoïte dans la protection contre le
paludisme demeure controversé. En effet, certaines études suggèrent que les anticorps
dirigés contre des épitopes des protéines telles que MSP1, MSP2 et AMA1 (Polley et al,
2004; Egan et al, 1996; Conway et al, 2000; Metzger et al, 2003) mesurée par ELISA peuvent
être des marqueurs de la protection contre le paludisme clinique, d'autres études ne
corroborent pas ces résultats (Dodoo et al, 1999; Okech et al, 2004 ; Corran et al, 2004).
Les mérozoïtes libres et les hématies infectées, n’exprimant pas à leurs surfaces de
molécules HLA, sont souvent négligés comme cibles pour des réponses immunes
protectrices à médiation cellulaire. Cependant l’un des résultats les plus étonnants sur le
rôle de l’immunité à médiation cellulaire dans la protection contre le paludisme a été la

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 49
démonstration faite par Pombo et al, (2002) que les personnes non immunes infectées à
maintes reprises par des doses ultra faibles de parasites du stade sanguin et soignés,
développent une immunité subséquente en l'absence de réponse anticorpale détectable.
Dans cette étude, les personnes protégées présentaient de fortes proliférations cellulaires T
CD4+ et T CD8+, une réponse cytokinique composée d’IFN-, mais une absence d’interleukine
4 ou d’interleukine 10 et la production de fortes concentrations de NO dans les cellules
mononuclées du sang des sujets (Pombo et al, 2002).
Figure 9. Représentation schématique du possible mécanisme d’action de l’immunité à médiation
cellulaire dirigée contre le stade sanguin de Plasmodium. Good, 2001.
Le rôle des cellules T spécifiques de Plasmodium a été mis en évidence grâce au transfert
adoptif de la protection en absence de réponse humorale détectable (van der Heyde et al,
1994, von der Weid et al, 1996). Les cellules T ont la capacité d'inhiber la croissance du
parasite in vitro (Taylor-Robinson et al, 1993; Fell et al, 1994; Amante et al, 1997).
Le modèle de l'immunité cellulaire ne dépendant pas d’anticorps au stade sanguin du
parasite est résumé sur la figure 9 ci-dessus.
Dans ce modèle, la réponse immune cellulaire débute par l'activation des cellules T CD4+
dans la rate, après la présentation antigénique par les Cellules dendritiques (DC). L’immunité

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 50
cellulaire T est régulée par l'IL-12 et fait intervenir IFN- et TNF- qui, induisent la
phagocytose des globules rouges infectés ainsi que la mort des parasites intracellulaires via
la synthèse de NO et des radicaux oxygénés (Ferrante et al, 1990, Stevenson et al, 1995) par
les neutrophiles et les macrophages (Ockenhouse et al, 1984, Stevenson et al, 1989).
La destruction des parasites a lieu essentiellement dans la rate (Favila-Castillo et al, 1996).
L’IFN- produite par les cellules T peut également contribuer à produire des anticorps
cytophiliques qui interviennent dans l’immunité cellulaire dépendante d’anticorps (ADCI)
(Bouharoun-Tayoun et al, 1995). La contribution relative de l'immunité humorale et
cellulaire dépend à la fois parasite et de l’hôte, et de la complexité de la relation hôte-
parasite. Chez les souris, l'immunité chez Plasmodium yoelii est principalement assurée par
les anticorps alors que chez P. chabaudi, elle est essentiellement à médiation cellulaire. Il
est probable que les humains diffèrent entre eux par le type de mécanisme effecteur mis en
jeu dans la réponse immune protectrice contre le parasite. Il est possible que différents
mécanismes effecteurs opèrent pour différentes souches de parasites.
I.5.1.3. Immunité dirigée contre les antigènes variables de surface
Les antigènes variables de surface (VSA) exprimés à la surface des globules rouges
parasités par P. falciparum sont des cibles importantes de l'immunité protectrice acquise qui
se développe après une exposition au parasite (Bull et al, 1999; Fried et al, 1998; Marsh et al,
1989; Nielsen et al, 2004).
Il a été montré que chez les femmes ayant souffert de paludisme placentaire, la présence
d’anticorps antiadhésifs spécifiques de VSA est associée à la protection contre le paludisme
lors de grossesses ultérieures (Staalsoe et al, 2004; Duffy et Fried, 2003).
Ces anticorps différencient les femmes primipares et multipares dans leur sensibilité au
paludisme. Les anticorps protecteurs sont dirigés contre des antigènes conservés du
parasite. En effet, des réactions croisées entre sérums et parasites provenant de régions
endémiques différentes ont été montrées (Fried et Duffy, 1998).
La protection contre le paludisme pendant la grossesse est associée au var2csa. L’acquisition
d’anticorps dirigés contre ce VSA est faite pendant la grossesse en réponse au parasite
(Tuike Ndam et al, 2006). Les taux plasmatiques élevés d'anticorps anti-VAR2CSA en début

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 51
de grossesse sont associés à un risque réduit de faible poids à la naissance (Salanti et al,
2004) et d’infections placentaires de longue durée (Tuike Ndam et al, 2006).
Le mécanisme humoral de protection contre le paludisme placentaire implique à la fois, les
anticorps bloquant l'adhérence, tout comme des processus cytophiliques tels que la
phagocytose et l’activation du complément. En effet, il a été montré que les IgG secrétées
pendant la grossesse chez les femmes camerounaises impaludées sont essentiellement
composés de sous-classes IgG1 et IgG3 (Megnekou et al, 2005).
L’immunité à médiation cellulaire joue aussi un rôle dans le paludisme placentaire, mais sa
contribution dans la protection demeure ambigüe. Au début de la grossesse il y a un biais
dans la polarisation Th1/Th2 pour faciliter le développement de l'allogreffe fœtale, en
favorisant une diminution des cytokines de type Th1 (TFN- et IFN-) (Krishnan et al, 1996)
et une augmentation des cytokines de type Th2 (IL-4, IL-10, TGF-β) (Wegmann et al, 1993;
Chaouat et al, 1999).
Les cytokines placentaires modulent la fonction des cellules présentatrices d’antigènes (CPA)
en inhibant ou en stimulant l’expression de molécules à la surface des monocytes, des
cellules dendritiques (DC) et des macrophages. La sécrétion d’IFN- par les cellules
mononuclées du sang intervilleux est associée à la protection contre le paludisme
placentaire (Moore et al, 1999), montrant le rôle de l’implication de la réponse cellulaire.
I.5.1.4. Mémoire immunologique
L’immunité anti palustre se développe lentement, et n’est jamais totale, elle est
perdue chez les adultes qui quittent une zone d’endémie palustre, suggérant que
l’exposition constante aux antigènes du parasite est nécessaire pour la mise en place de la
mémoire immunologique mais aussi de son entretien.
Il est généralement admis que les réponses immunes anti palustres et notamment les
réponses humorales sont de très courte durée (Achtman et al, 2005). Cependant, la notion
de mémoire immunologique dans le paludisme reste controversée. Chez les adultes, on a
mis en évidence, la présence de cellules B mémoires spécifiques de P. falciparum pendant
une période supérieure à 8 années en absence évidente d’exposition au paludisme (Deloron
et Chougnet, 1992). Une étude plus récente confirme la persistance des cellules B mémoires

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 52
spécifiques de P. vivax et P. falciparum pendant une période de 6 ans en absence de toute
transmission dans une zone endémique à faible transmission (Wipasa et al, 2010). De même,
une autre étude récente a montré que le compartiment des cellules B mémoires spécifiques
de P. falciparum augmente avec l’exposition au paludisme (Weiss et al, 2010). En revanche,
une étude précédente avait rapporté de très faibles fréquences de cellules B mémoires
spécifiques du parasite chez les enfants exposés au paludisme (Dorfman et al, 2005).
I.5.2. Rôle des cytokines dans l’infection palustre
Dans l’infection par Plasmodium la réponse inflammatoire est nécessaire pour
éliminer le parasite. Cependant, elle peut conduire à des lésions tissulaires considérables.
L’activation des phagocytes pour l’élimination des parasites intracellulaires ou
extracellulaires, nécessite la production des cytokines inflammatoires, ce qui peut provoquer
des effets systémiques tels que l’anémie sévère ou le paludisme cérébral (McGuire et al,
1994; Luty et al, 2000). Le pronostic d’une infection palustre dépend d’un délicat équilibre
entre une induction appropriée ou inappropriée de ces médiateurs.
I.5.2.1 Cytokines pro-inflammatoires et paludisme
I.5.2.1.1. TNF-
TNF- est la première cytokine caractérisée, induite par le parasite. La production de
TNF- est induite chez les macrophages par les érythrocytes infectés par Plasmodium,
l’hémozoïne (Pichyangkul et al, 1994) et certains glycolipides comme le GPI (Scholfield et al,
1993). Le GPI stimule aussi la synthèse d’oxyde nitrique synthétase (NOS) par les
macrophages (Tachado et al, 1996) et active les cellules endothéliales par la transduction
d’un signal médiée par la tyrosine-kinase (Schofield et al, 1996). Les anticorps dirigés contre
le GPI bloquent la production de TNF- par les cellules mononuclées en réponse à
l’activation de lysats de globules rouges infectés par différents clones de parasites (Bate et
al, 1994).
TNF- joue un rôle important dans la phagocytose des érythrocytes infectés en augmentant
l’expression des récepteurs Fc sur les monocytes.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 53
TNF- agit également sur les lymphocytes et monocytes en augmentant l’inhibition de P.
falciparum par un mécanisme indépendant de la phagocytose.
TNF- joue aussi un rôle dans la régulation de la production d’autres cytokines. En effet,
TNF- régule la production de l’interleukine-12 (IL-12) par les macrophages. C’est aussi un
important cofacteur de l’IL-12 dans la production d’IFN- par les cellules NK (Tripp et al,
1993).
Les concentrations plasmatiques de TNF- et NO sont associées à la résolution rapide de la
fièvre et à l’élimination des parasites. Toutefois, il convient de noter que TNF- semble
également avoir, chez environ 1% des individus atteints de paludisme, des propriétés
néfastes telles que la fièvre, les courbatures et douleurs en relation avec la phase aiguë de la
maladie, une hypoglycémie, des hémorragies et un coma réversible (Beutler et al, 1993).
En outre, une étude a montré une étroite association entre l’anémie sévère, des
concentrations élevées de TNF- et un grand nombre de monocytes circulants contenant
l’hémozoïne. Ceci suggère que la production de TNF- induite par l’hémozoïne joue un rôle
dans l’initiation ou l’exacerbation de l’anémie comme pronostic clinique d’une parasitémie
chronique incontrôlée (Luty et al, 2000).
I.5.2.1.2. Interféron-gamma (IFN-)
L'interféron est un facteur d'activation des macrophages impliqué dans la réponse
immunitaire innée au paludisme. Il est produit principalement dans une réponse
immunitaire spécifique par les lymphocytes T CD4+ et T CD8+et par les cellules NK dans une
réponse immunitaire non spécifique (Weiss et al, 1993).
Les études faites dans les modèles murins expérimentaux ainsi que chez l'homme suggèrent
un rôle important de l’IFN- dans les réponses immunitaires protectrices contre le paludisme
au stade sanguin. La production d’IFN- par les cellules T CD4+ spécifiques d’antigènes du
stade sanguin est associée à une protection contre la réinfection (Luty et al, 1999). La
sécrétion cellulaire de l’IFN- pourrait également contribuer à stimuler la synthèse des
immunoglobulines cytophiliques spécifiques du stade sanguin et jouer un rôle dans les
mécanismes cellulaires inhibiteurs dépendant d’anticorps (Bouharoun-tayoun et al, 1995).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 54
Les cellules cibles de l’IFN- lors d'une infection à P. falciparum sont des monocytes/
macrophages (Bate et al, 1988), les neutrophiles (Kumaratilake et al, 1991), les cellules Th2
(Taverne et al, 1993) et les hépatocytes infectés par le parasite (Klotz et al, 1995).
Les Macrophages activés par l’IFN- libèrent le TNF-, le facteur de croissance transformant-
beta (TGF-β), l'interleukine 1 (IL-1), l’interleukine 6 (IL-6), les radicaux libres oxygéné et le NO
(Clark et al, 1997). L’IFN- par l’intermédiaire de signaux de transducteurs associés à la
transcription, active l’oxyde nitrique synthétase inductible (iNOS) et induit la L-arginine par
la voie dépendante du NO, entrainant l'élimination d’hépatocytes ou de schizontes
hépatiques infectés (Snounou et al, 2000).
Le NO jouerait donc un rôle important dans la destruction des parasites intra hépatiques en
réponse à l’IFN- et à d'autres cytokines libérées par les cellules T et les cellules NK.
Le traitement d’hépatocytes infectés par Plasmodium in vitro avec de l’IFN- entraîne
l’élimination de P. falciparum de la culture.
Chez l’homme, il a été montré que les enfants ayant une hyper parasitémie à P. falciparum
avaient des concentrations plus faibles en cellules T CD4+ sécrétrices d’IFN-
comparativement à ceux ayant un paludisme simple (Winkler et al, 1999). Il semblerait que
l’IFN- soit essentiel dans la résolution de l'infection primaire en limitant la phase initiale de
la réplication du parasite, mais contribuerait également aux symptômes de la phase aigue
du paludisme, comme la fièvre, les nausées, des maux de tête à travers l’induction de TNF-
et l’IL-1(Riley et al, 1999)
I.5.2.1.3. L’interleukine-12 (IL-12)
L’IL-12 est une cytokine immunomodulatrice impliquée dans la protection contre les
infections virales, bactériennes et parasitaires. Cette cytokine stimule non seulement une
réponse immunitaire à médiation cellulaire, mais elle affecte également l’immunité
humorale en induisant la commutation isotypique des immunoglobulines par des
mécanismes qui sont à la fois dépendants ou indépendants de l’IFN- (Trinchieri, 1995). L’IL-
12 stimulerait aussi la production d’anticorps par les lymphocytes B.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 55
Dans un modèle murin, Il a été montré que l'IL-12 induisait une immunité protectrice contre
le stade sanguin de l'infection plasmodiale (Crutcher et al, 1995). Il est probable que la
production d’IL-12 soit stimulée par la phagocytose du parasite.
L’IL-12 agit sur les cellules T CD4+ activées par l'antigène, en stimulant les signaux
transducteurs, en activant le facteur de transcription 4 (STAT4) et en induisant la
différenciation des cellules T en sous-type Th1 (O’Garra et al, 2000).
Les cellules effectrices Th1 produisent l’IFN- qui agit sur les macrophages en stimulant leur
fonction microbicide et en augmentant leur production d'IL-12. Les concentrations élevées
d'IL- 12 modulent l'activité des macrophages, qui est associée à une forte destruction des
érythrocytes (Luty et al, 2000, Crutcher et al, 1995). Il semblerait que les événements
précoces de la réponse immunitaire à médiation cellulaire nécessaires dans la défense
contre le paludisme, sont initiés par la libération de l'IL- 12 par les monocytes/macrophages,
les cellules B, et d'autres types cellulaires (Luty et al, 2000, Crutcher et al, 1995) et par
conséquent la concentration d'IL-12, révèle une signification pronostique dans l’infection
palustre.
L'induction de l’IFN- est une conséquence directe de l’activation des cellules T CD4+ et T
CD8+: la production d’IFN- précède et initie celle de l’IL-12, qui à son tour induit la
production de l’IFN- par les cellules NK dans une boucle rétroactive positive qui représente
un important mécanisme d'amplification (O’Garra et al, 2000). Les concentrations réduites
d’IL-12 chez les patients atteints d’hyper parasitémie et de paludisme sévère pourraient être
liées à l’activité réduite de l’interféron médiée par les cellules T. Au cours du paludisme aigu
la production constitutive de l'IL-12 par les monocytes est inhibée après la phagocytose de
l’hémozoïne ou après production d'IL-10 qui antagonise l'activité de l'IL- 12 (O’Garra et al,
2000).
I.5.2.2. Cytokines anti-inflammatoires
Les réponses précoces des cytokines pro-inflammatoires semblent assurer une
immunité protectrice, tandis que les réponses tardives contribuent à la pathologie. Ceci
suggère qu’un équilibre crucial devrait exister au cours de la réponse inflammatoire dans
l’infection palustre ; une réponse asymétrique conduisant à une maladie grave. Dans le

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 56
paludisme simple, la réponse inflammatoire devrait être régulée par les cytokines anti-
inflammatoires telles que l’IL-4, l’IL-10 et TGF-β.
I.5.2.2.1. Interleukine-4
L’IL-4 est produite par les lymphocytes de type Th2 et les basophiles/mastocytes
activés. Elle est impliquée dans l’activation des lymphocytes T cytotoxiques (LTC), les cellules
NK et les macrophages.
L’IL-4 est une composante importante de la réponse immunitaire. Elle stimule la croissance
des lymphocytes Th2 et inhibe la réponse Th1 par la réduction de la production d'IFN- (Luty
et al, 2000). L’IL-4 et les lymphocytes Th2 sont importants dans la réponse immunitaire
humorale contre Plasmodium. Les cellules T CD4+ sont cruciales pour le développement de
réponses T CD8+ dirigées contre les hépatocytes infectés (Carvalho et al, 2002). Il a été
montré que la production d’IL-4 stimulée chez les cellules T par les antigènes du parasite in
vitro était associée à l’augmentation d’anticorps sériques en réponse aux mêmes antigènes
plasmodiaux in vivo. Cependant, une autre étude a montré que l’IL-4 inhibait la capacité des
macrophages de donneurs naïfs à éliminer P. falciparum (Kumaratilake et al, 1992). Cette
étude est en contradiction avec celle montrant que l’IL-4 contribue à la réponse humorale
dirigée contre les parasites (Troye-Blomberg et al, 1990).
I.5.2.2.2. Facteur de croissance transformant-beta (TGF-β)
TGF-β produit par un large éventail de cellules comme les macrophages, les cellules
NK et les cellules T et B, a un rôle central dans le contrôle de la transition entre la réponse
pro-inflammatoire (Th1) et la réponse anti-inflammatoire (Th2) pendant les phases aigue et
de résolution de l’infection palustre (Tsunawaki et al, 1988).
Le rôle de TGF-β a été mis en évidence dans un modèle expérimental d’infection où, sa
concentration est cruciale pour l’activation des macrophages (Tsunawaki et al, 1988). Effet,
les monocytes / macrophages immatures ont de fortes concentrations de récepteurs TGF-β
et sont sensibles à de faibles concentrations de TGF-β, qui favorisent la maturation des
macrophages et les rendent plus sensibles à l'activation par ce dernier.
De plus, le TGF-β inhibe la production d'IFN- et de TNF-, augmente celle de l 'IL-10
(Maeda et Shiraishi, 1996) et diminue l’expression des molécules d'adhésion (Nakabayashi et

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 57
al, 1997). Une production précoce de TGF-β dans l'infection, active les
monocytes/macrophages provoquant la phagocytose des globules rouges parasités et la
mort des parasites ingérés (Ferrante et al, 1990). La cinétique de TGF-β semble être cruciale
pour un contrôle efficace de la densité parasitaire dans l’infection plasmodiale. En effet, trop
de TGF-β trop tôt va empêcher la mise en place de l’immunité cellulaire de type Th1 par
l’inhibition de l’IFN- et de TNF-. Trop peu de TGF-β trop tard conduit à une hyper
parasitémie et à la mort liée à une surproduction des cytokines de type Th1 (Omer et al,
2000).
TGF-β jouerait deux rôles importants dans le paludisme qui varient selon le moment de
l'infection. Au début de l'infection, le TGF-β pourrait induire les réponses de type Th1 qui
contrôlent la croissance du parasite. Plus tard dans l'infection TGF-β pourrait réguler
négativement les réponses Th1 pour limiter la pathologie associée à l’inflammation. TGF-β
joue aussi un rôle dans le pronostic de l’infection plasmodiale à travers ses effets sur les
lymphocytes B. En effet, de faibles concentrations de TGF-β stimulent la sécrétion de sous-
classes d'immunoglobulines par les cellules B (Snapper et al, 1993), mais à des
concentrations plus élevées la production d'anticorps est inhibée (Stavnezer et al, 1995).
Le rôle de TGF-β a été aussi étudié dans les infections palustres chez l’homme. Deux études
de l'infection naturelle chez les patients au Gabon (Perkins et al, 2000) et en Thaïlande
(Wenisch et al, 1995) et une étude expérimentale de l'infection humaine (Walther et al,
2005 et Walther et al, 2006) corroborent les résultats obtenus dans les modèles animaux.
Une étude longitudinale d’une infection induite par les sporozoïtes de P. falciparum chez
l’homme a montré qu’une régulation positive précoce du taux de TGF-β1 et des taux élevés
de TGF-β1 bioactifs au moment de l'émergence du parasite du foie étaient associés à une
croissance plus rapide du parasite (Walther et al, 2005 et Walther et al, 2006).
I.5.2.2.3. L’interleukine-10 (IL-10)
L’Interleukine-10 a été trouvé dans le plasma des patients atteints de paludisme aigu
(Wenisch et al, 1995). L’IL-10 est produite par les monocytes, les lymphocytes Th2 et les
cellules B. Cette cytokine inhibe la production des cytokines des lymphocytes Th1 et des
cellules T CD8+, mais pas celle des lymphocytes Th2. Néanmoins, l'IL-10 n'affecte pas la

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 58
prolifération des lymphocytes Th1 et T CD8 +, mais induit la prolifération des cellules B et la
production d'immunoglobulines, qui est essentiel pour le développement et la maturation
des anticorps antipaludéens. L’IL-10 semble avoir un rôle important dans la définition de la
réponse des cellules T auxiliaires du paludisme.
En outre, l'IL-10 réduit l’expression du CMH II sur les macrophages, conduisant à la réduction
de la présentation d’antigènes (Akdis et al, 1999), inhibe la production des radicaux libres
oxygénés et le monoxyde d’azote (NO), empêche le priming et la prolifération des cellules T',
et supprime la production par les cellules T d'IFN-, d’IL-6, TNF- et de GM-CSF (Akdis et al,
1999). L'inhibition de la sécrétion de l’IFN- et de TNF- par l'IL-10 a été reportée comme un
mécanisme pour contrecarrer le rôle pathologique des macrophages dans le paludisme
cérébral (Kossodo et al, 1997). Une étude réalisée au Gabon chez des enfants et des adultes
infectés par P. falciparum, a montré de nombreuses cellules T CD4+ et T CD8+ productrices
d’IL-10 co-exprimant l’IFN- (Winkler et al, 1999).
Enfin, l'augmentation de l'IL-10 est plus prononcée et plus spécifique que l'IL-6 et l'IL-8 chez
les patients ayant une infection palustre par rapport à d'autres infections (Jason et al, 2001).
Toutefois, il n'est pas encore clair si l'augmentation de la concentration d’IL-10 a un rôle
bénéfique en réduisant la réponse inflammatoire induite par le parasite, ou est préjudiciable
en diminuant les réponses immunitaires cellulaires.
I.5.3. Les cellules présentatrices d’antigènes (CPA)
La présentation d’antigènes d’un agent pathogène peut être faite aux lymphocytes B
mais également aux lymphocytes T.
La présentation de l’antigène aux lymphocytes B se fait via l’interaction entre leur récepteur
d’antigène (BCR) et des épitotes d’antigènes solubles, particulaires ou situés sur des
membranes cellulaires. Les CPA des lymphocytes B sont essentiellement les cellules
dendritiques folliculaires (CDF) des ganglions et de la rate.
Les antigènes sont aussi présentés aux lymphocytes T. Cette présentation est faite au TCRβ
sous forme de peptides par les molécules de classe I ou de classe II du complexe majeur
d’histocompatibilité (CMH). La présentation par les CMH I aux lymphocytes T CD8+ est
assurée par presque toutes les cellules de l’organisme. Elle intervient dans la lyse des

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 59
parasites intracellulaires. Les CPA comprennent les cellules dendritiques, les lymphocytes B,
les macrophages/monocytes, les cellules endothéliales et les cellules épithéliales.
I.5.3.1 Les cellules dendritiques (DC)
Les cellules dendritiques sont les principales CPA dans la majorité des organes. Ce
sont des cellules qui ont la particularité unique d’activer les cellules T naïves. Elles expriment
à leur surface des molécules de CMH II et présentent les antigènes aux cellules T CD4+. Elles
interagissent avec les cellules T et cette interaction se fait via la relation entre leurs
molécules co-stimulatrices B7 (CD80 et CD86) et le CD28 des cellules T. Les cellules
dendritiques captent les antigènes, assurent leur apprêtement, et leur présentation sous
formes de peptides associés aux CMH II, puis leur transport vers les zones T des ganglions
lymphoides. Elles ont la particularité d’orienter la réponse immunitaire vers une réponse de
type Th1 par la production d’IL-12. Elles perdent leur capacité de phagocytose par la
migration et la maturation (augmentation à leur surface de molecules CMH II, CD80 et CD86,
CD40). Leur maturation est stimulée par divers parasites et agents pathogènes et par
certains composés bactériens tels que les lipopolysaccharides (LPS) ; mais elle est inhibée en
présence d’IL-10. Le rôle des cellules dendritiques a été décrit dans divers modèles
d’infections plasmodiales. Certaines études ont montré que leur maturation est inhibée en
présence d’érythrocytes inféctés par P. falciparum, alors que d’autres études au contraire,
montrent qu’elles demeurent fonctionnelles.
I.5.3.2. Les lymphocytes B
Les lymphocytes B reconnaissent l’antigène par leurs immunoglobulines
membranaires, internalisent le complexe BCR-Antigène et le dégrade en peptides dans la
voie des endosomes ; ces peptides sont associés aux CMH II et exprimés sur la membrane
cellulaire. Cela permet l’activation des cellules T qui, à leur tour par contact membranaire et
synthèse de cytokines contrôlent l’activation des cellules B. Ce mode de présentation de
l’antigène intervient surtout lors de la réponse anticorps à une injection de rappel d’un
antigène T-dépendant.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 60
I.5.3.3. Les macrophages/monocytes
Les macrophages et monocytes, qui expriment les molecules de classe II du CMH,
présentent des peptides d’antigènes aux lymphocytes T CD4+. Du fait de leur localisation
anatomique hors des zones T des organes lymphoïdes périphériques, ces cellules
n’interviennent pas dans l’induction d’une réponse immunitaire. Elles interagissent par
contre avec des lymphocytes T CD4+ différenciés au stade effecteur des réponses T
dépendantes.
La présentation de l’antigène aux lymphocytes T CD4+ peut aussi être assurée par les cellules
endothéliales ou des cellules épithéliales exprimant des molécules de classe II du CMH après
stimulation par l’IFN-.
I.5.4. Stratégies d’évasion immunitaire
Les relations entre l’hôte et son parasite sont complexes. Pour survivre P. falciparum
a développé plusieurs stratégies d’évasion du système immunitaire de son hôte. Ces
stratégies vont du parasitisme intracellulaire, simple mécanisme d’échappement à la
reconnaissance antigénique, à la diversité ou la variation antigénique. Le génome de P.
falciparum compte plus de 5300 protéines, et nombreuses d’entre elles sont polymorphes
(Gardner et al, 2002).
La plupart des réponses induites par les antigènes polymorphiques du parasite pourraient ne
pas être protectrices mais servir de leurre au système immunitaire de l’hôte.
La variation antigénique apparaît alors comme un mécanisme efficace pour échapper à
l’immunité humorale. Les stratégies d’évasion immunitaire de P. falciparum sont
nombreuses. Nous présentons deux de ces mécanismes dans la section ci-dessous.
I.5.4.1. Altération des ligands peptidiques
Les épitopes variants de CSP provoquent un antagonisme induit par l’altération des
ligands peptidiques qui inhibe le priming des cellules T par les cellules présentatrices
d’antigènes de class I (Gilbert et al, 1998; Young et al, 2005). Ainsi, les cellules T sont
capables de proliférer en réponse à l’antigène, mais sont incapables d’éliminer le parasite ou
de produire des cytokines protectrices telles que l’IFN-. Il a été montré aussi que les mêmes
épitopes variants de CSP sont capables d’interagir avec les cellules T cytotoxiques mémoires

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 61
de sujets exposés au paludisme, abolissant ainsi leur activité lytique (Plebanski et al, 1999).
L’altération des ligands peptidiques interfère avec les réponses T protectrices. Cet
antagonisme pourrait être une stratégie pour maintenir une population de cellules T de
l’hôte exposée au paludisme tout en demeurant fonctionnellement naïve.
L’infection par P. falciparum induit l’anergie et la suppression des cellules T spécifiques du
parasite et non celle des cellules T de spécificité différente, c’est une autre stratégie
développée par le parasite pour échapper à son hôte et de retarder le développement de
l’immunité dirigée contre lui (Hirunpetcharat et Good, 1998).
I.5.4.2. Altération des cellules présentatrices d’antigènes
L’infection des globules rouges par P. falciparum altère la maturation des cellules
dendritiques aussi bien chez l’homme (Urban et al, 1999) que chez les souris
(Ocana‐Morgner et al, 2003). Chez l’homme, l’inhibition des cellules dendritiques est
probablement médiée par l’interaction entre PfEMP-1 exprimé à la surface des globules
rouges parasités et le récepteur CD36 exprimé à la surface des cellules dendritiques (Urban
et al, 2001).
Le parasite peut aussi échapper au système immunitaire de l’hôte en inhibant la fonction des
monocytes/macrophages par les globules rouges infectés (Leitner et Krzych, 1997) et/ ou par
l’hémozoïne (Skorokhod et al, 2004). L’inhibition de la maturation des cellules dendritiques
par les globules rouges parasités demeure une controverse. En effet, différentes études ont
montré des cellules dendritiques entièrement fonctionnelles en réponse à l’infection
plasmodiale (Seixas et al, 2001; Coban et al, 2002; Perry et al, 2004).
PfEMP-1 peut aussi atténuer la réponse immune de l’hôte en supprimant la production
d’IFN- par les PBMCs par un mécanisme indépendant du CD36 (D’Ombrain et al, 2007).
I.5.5. Rôle des cellules T régulatrices naturelles dans l’infection palustre
Les lymphocytes T régulatrices (Tregs) appartiennent à une population de cellules
lymphoïdes exerçant une activité immunosuppressive. Elles jouent un rôle critique dans
l’équilibre entre les réponses immunes protectrices et les pathologies auto-immunes
(Kullberg et al, 2002; Maloy et al, 2003; Hori et al, 2002; Montagnoli et al, 2002).
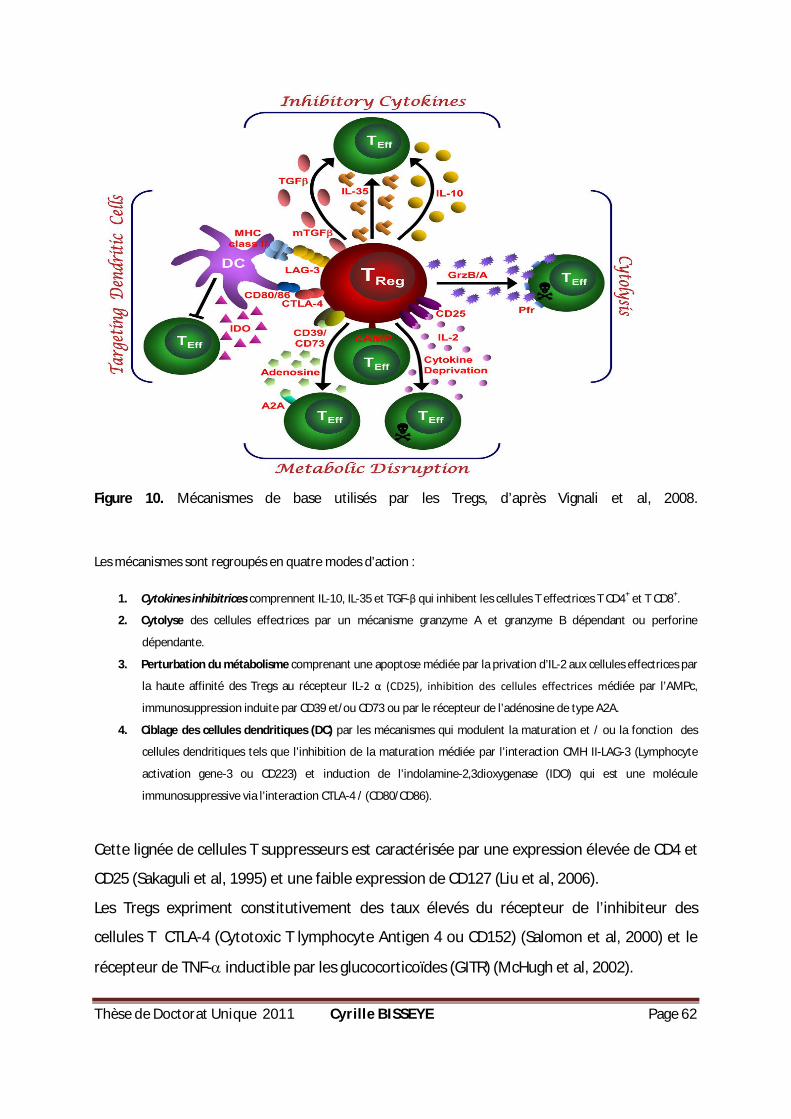
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 62
Figure 10. Mécanismes de base utilisés par les Tregs, d’après Vignali et al, 2008.
Les mécanismes sont regroupés en quatre modes d’action :
1. Cytokines inhibitrices comprennent IL-10, IL-35 et TGF-β qui inhibent les cellules T effectrices T CD4+ et T CD8+.
2. Cytolyse des cellules effectrices par un mécanisme granzyme A et granzyme B dépendant ou perforine
dépendante.
3. Perturbation du métabolisme comprenant une apoptose médiée par la privation d’IL-2 aux cellules effectrices par
la haute affinité des Tregs au récepteur IL-2 α (CD25), inhibition des cellules effectrices médiée par l’AMPc,
immunosuppression induite par CD39 et/ou CD73 ou par le récepteur de l’adénosine de type A2A.
4. Ciblage des cellules dendritiques (DC) par les mécanismes qui modulent la maturation et / ou la fonction des
cellules dendritiques tels que l’inhibition de la maturation médiée par l’interaction CMH II-LAG-3 (Lymphocyte
activation gene-3 ou CD223) et induction de l’indolamine-2,3dioxygenase (IDO) qui est une molécule
immunosuppressive via l’interaction CTLA-4 / (CD80/CD86).
Cette lignée de cellules T suppresseurs est caractérisée par une expression élevée de CD4 et
CD25 (Sakaguli et al, 1995) et une faible expression de CD127 (Liu et al, 2006).
Les Tregs expriment constitutivement des taux élevés du récepteur de l’inhibiteur des
cellules T CTLA-4 (Cytotoxic T lymphocyte Antigen 4 ou CD152) (Salomon et al, 2000) et le
récepteur de TNF- inductible par les glucocorticoïdes (GITR) (McHugh et al, 2002).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 63
Forkhead box P3 (Foxp3) est le seul facteur de transcription requis pour la génération des
Tregs (Hori et al, 2003) et c’est le marqueur le plus spécifique de cette population de
cellules.
Le rôle des Tregs est bien établi dans l’inhibition de l’immunité anti-tumorale (Shimizu et al,
1999) mais aussi dans la prévention des maladies auto-immunes comme le diabète (Salomon
et al, 2000).
Les mécanismes de base utilisés par les cellules Tregs sont résumés dans la figure 10.
Les cellules T regs naturelles sont capables d'inhiber à la fois la production d'IL-2 et la
prolifération cellulaire des lymphocytes TCD4+ classiques (Thornton et al, 1998) par des
mécanismes contact-dépendants et/ou par la production de cytokines anti-inflammatoires
comme le TGF-β (Nakamura et al, 2001) et l'IL-10 (Asseman et al, 1999).
Les T regs naturelles ont une capacité de prolifération limitée et sont incapables de sécréter
l’IL-2, elles exercent leur fonction suppressive à travers un récepteur des cellules T (CTLA-4),
un processus qui semble nécessiter la présence de cellules présentatrices d’antigène comme
les cellules dendritiques (Thornton et al, 1998).
Après activation, les Tregs naturelles ne nécessitent aucune stimulation supplémentaire et
sont capables de supprimer les réponses des lymphocytes T de manière non spécifique.
En plus des Tregs naturelles, les Tregs inductibles constituent un autre sous-type de
lymphocytes T suppresseurs (Bluestone et al, 2003). Contrairement aux Tregs naturelles qui
se développent dans le thymus, les Tregs inductibles proviennent des lymphocytes T CD4+ et
T CD8+ classiques qui acquièrent leur activité suppressive par l’activation par les cytokines
immunosuppressives ou les cellules dendritiques immatures.
La raison pour laquelle les parasites du paludisme persistent dans l'état de prémunition est
inconnue. Il est probable que des mécanismes immunosuppresseurs modulant l’immunité
protectrice conduisent à la persistance des parasites. Par leur action immunosuppressive, les
Tregs naturelles pourraient entraver l’induction de l’immunité anti plasmodiale.
I.5.5.1. Rôle des Tregs naturelles dans le contrôle de l’infection palustre chez les rongeurs
Chez les rongeurs, de nombreuses études ont investigué le rôle des Tregs naturelles
dans la modulation de l’induction des mécanismes immunitaires responsables du contrôle
de la charge parasitaire. En effet, Il existe deux sous-lignées de P.yoelii 17X. L'une d’entre

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 64
elle est très virulente (PyL) et provoque des infections létales, tandis que l’autre (PyNL) se
traduit par une auto-résolution d’infections non létales.
Chez les souris BALB/c infectées par les parasites virulents PyL, la déplétion des cellules
Tregs naturelles provoque une hyper parasitémie suivie de la mort, faisant ainsi apparaître le
rôle des cellules Tregs dans un mécanisme d’échappement essentiel utilisé par les parasites
pour se soustraire à l'immunité de l'hôte (Hisaeda et al, 2004).
Dans une autre étude utilisant les parasites virulents et non virulents de P. yoelii, les
expériences de déplétion in vivo n’ont pas confirmé le rôle des cellules Tregs naturelles dans
le contrôle de la parasitémie chez les souris (Couper et al, 2008).
L'impact de la déplétion des cellules Tregs naturelles a été aussi évalué dans les modèles
murins d’infection par les lignées létales (DS) et non létales (DK) de P. chabaudi adami. Dans
ces modèles, La déplétion des cellules Tregs entraîne une production accrue d'IFN- et de
TNF- au cours d’infections létale et non létale par P. chabaudi adami. La déplétion des
cellules Tregs entraîne des réponses pro-inflammatoires accrues chez les souris infectées par
la lignée non létale; cependant, elle ne modifie pas le cours de leur infection (Cambos et al,
2008). Dans la même étude, il a été montré que La déplétion des Tregs naturelles aggravait
la charge parasitaire et accélérait la mort des souris infectées par la lignée létale DS.
L'anémie hémolytique qui résulte d’une hyper parasitémie a également été renforcée chez
les souris en l'absence de cellules Tregs naturelles (Cambos et al, 2008).
I.5.5.2. Rôle des cellules Tregs naturelles dans le contrôle du paludisme cérébral expérimental
Plasmodium berghei-ANKA induit chez les lignées de souris sensibles le paludisme
cérébral. Cette infection murine a de nombreuses caractéristiques communes avec
l’infection humaine et est donc acceptée comme un modèle pour certains aspects
importants du paludisme cérébral (Brain de Souza, 2002; Miller et al, 2002; Schofield et
Grau, 2005; Clark et Schofield, 2000).
Les cytokines anti-inflammatoires comme le TGF-β et l'IL-10 régulant d’une part les
réponses immunitaires de type Th1 au cours de l’infection palustre (Omer et al, 1998), et
d’autre part, jouant un rôle protecteur contre le paludisme cérébral à P. berghei (Kossodo et

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 65
al, 1997), il a été postulé que la résistance à la maladie pourrait être associée à l'élaboration
de mécanismes de défense visant à inhiber directement les réponses inflammatoires
pathogènes.
Chez les souris BALB/c résistantes au paludisme cérébral, les cellules T CD4+ présentent une
sécrétion d’IL-2 réduite et une prolifération cellulaire fortement inhibée en réponse aux
stimuli polyclonaux et spécifiques du parasite. Les cellules Tregs joueraient un rôle dans
l’inhibition de la prolifération des cellules T se produisant au cours de l’infection (Nie et al,
2007). Dans cette même étude, la déplétion des cellules Tregs a montré qu’ils empêchaient
la mise en place de cellules mémoires de type Th1 spécifiques du parasite, démontrant le
rôle des Tregs dans le contrôle des réponses pathogéniques aboutissant à une maladie fatale
(Nie et al, 2007).
I.5.5.3. Rôle des cellules T régulatrices dans le contrôle du paludisme chez l’homme
Quelques études se sont intéressées au rôle des cellules Tregs dans l’infection
palustre chez l’homme. Ces études sont complexes du fait de la variété de facteurs à
prendre en compte: design de l’étude, taille de l’échantillon, utilisation d’échantillons frais
ou cryopréservés qui pourraient avoir un impact sur les résultats.
Walther et al, (2005) ont montré chez des volontaires infectés expérimentalement par les
piqûres de moustique, une augmentation du nombre de cellules Tregs dans leur sang
périphérique. Ils ont observé chez les personnes ayant un nombre élevé de cellules Tregs, de
plus fortes densités parasitaires que chez celles ayant de faibles fréquences de cellules Tregs;
suggérant que la présence de cellules Tregs dans le sang périphérique de personnes
infectées pourrait faciliter la réplication du parasite. Dans cette même étude, ces auteurs
ont montré qu’une réduction des cytokines pro-inflammatoires telles qu’IFN-, IL-6 et IL-12
ainsi qu’une production significative de TGF-β étaient associées à une parasitémie élevée
(Walther et al, 2005). En outre, les monocytes et non les cellules Tregs étaient la source
cellulaire de TGF-β produit en réponse à l'infection (Walther et al, 2005). Ces auteurs ont de
ce fait postulé que le TGF-β produit par les monocytes pouvait favoriser la différenciation
des lymphocytes T CD4+ naïfs périphériques en cellules Tregs en induisant leur expression de

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 66
Foxp3. Cette augmentation du nombre de cellules Tregs entraîne une croissance accrue du
parasite.
Dans une étude réalisée au Kenya, l’augmentation du nombre de cellules Tregs CD4+
CD25high dans le sang périphérique des sujets a été associée à un risque accru de paludisme
clinique (Todryk et al, 2008), suggérant que les lymphocytes Tregs naturelles peuvent
affecter négativement l'immunité naturelle acquise contre le paludisme.
Il a été montré que, malgré le niveau similaire d’exposition au paludisme, les Peuls du
Burkina Faso semblent être plus résistants à la maladie que les autres groupes ethniques
(Modiano et al, 1996). Au Burkina Faso, le rôle des cellules T régulatrices a été étudié dans
une approche comparative interethnique intégrant la susceptibilité au paludisme (Torcia et
al, 2008). Dans cette étude, le pourcentage de cellules Tregs était supérieur chez les Mossis
que chez les Peuls. Une forte résistance au paludisme chez les personnes d’origine Peule
était associée non seulement à un déficit fonctionnel des cellules Tregs mais aussi à une
réduction de l’activité immunosuppressive par rapport aux Mossis (Torcia et al, 2008).
Le rôle des cellules Tregs a été aussi exploré chez les enfants gambiens atteints de paludisme
simple ou sévère (Walther et al, 2009). Walther et al, ont montré que la réponse
immunitaire de type Th1 était plus élevée chez les enfants présentant un paludisme grave.
Cependant, le nombre et la fréquence des cellules Tregs étaient identiques dans les deux
groupes cliniques (Walther et al, 2009).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 67

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 68
II. Matériels et méthodes
II.1. Populations d’étude
Groupe de Sukuta
Le groupe d’étude de Sukuta a été recruté en collaboration avec le ministère
gambien de la santé de septembre 2001 à janvier 2002. La ville de Sukuta (banlieue de
Banjul) est située à 11 km du Medical Research Council (MRC). La ville de Sukuta compte
environ 20.000 habitants, le nombre d’accouchements annuels est d’environ 800. Soixante
quinze (75) femmes ayant consenti librement à participer à l’étude ont été recrutées. Le
sang du cordon (jusqu'à 50 ml) a été prélevé à l’accouchement dans les tubes héparines. Les
gouttes épaisses ont été préparées, colorées au Giemsa pour la recherche de l’infection
active par P. falciparum dans le sang périphérique des mères, le sang du cordon prélevé dans
les tubes EDTA, et les empreintes de placentas. Toutes les lames ont été lues à deux reprises
par deux microscopistes expérimentés et les différences de lecture ont été résolues par un
troisième lecteur. Une lame n’était déclarée négative qu’après lecture de 100 champs au
grossissement x1000 (limite de détection, 2 parasites/µL) (Greenwood et al, 1991)
Groupe de Brefet
Quarante-trois (43) donneurs adultes asymptomatiques pour le paludisme ont été
recrutés à Brefet (village situé à 85 km de Banjul) en Gambie. Des échantillons de sang (50
ml) ont été collectés dans des tubes héparine après consentement éclairé de tous les
donneurs. Les gouttes épaisses ont été préparées pour déterminer la parasitémie des
donneurs. Le taux d’hémoglobine des donneurs a été aussi mesuré. Des échantillons de sang
ont ensuite été traités pour séparer les plasmas des cellules mononuclées du sang
périphérique (PBMC). Cette étude a été approuvée par le Comité d'éthique mixte
gouvernement de Gambie/ MRC.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 69
Groupe de Farafenni
Pour étudier la réponse immunitaire dirigée contre les antigènes MSP119, AMA1 et
CSP de P. falciparum, un groupe de 658 adultes et enfants (303 enfants et 234 adultes) a été
recruté à Farafenni, située sur la rive nord du fleuve Gambie à près de 300 kms de Banjul. La
transmission du paludisme y est saisonnière, elle correspond à la saison pluvieuse comprise
entre août et décembre. La transmission connaît un pic entre octobre et novembre. Les
sujets ont été recrutés dans 8 villages environnant Farafenni.
Le système démographique d’enregistrement des ménages du MRC a été utilisé pour
recruter un échantillon d’enfants et d’adultes âgés de 6 mois à 45 ans. Les enfants et les
adultes ont été recrutés parmi les trois principales ethnies de la région, Wolof, Mandinka et
Peul.
Le consentement éclairé a été obtenu de tous les participants adultes et des parents ou
tuteurs des enfants. Les individus ont été inclus dans l’étude du 14 au 28 juillet 2003. Un
prélèvement de sang veineux a été effectué dans des tubes EDTA et héparine. Culots
sanguins et plasmas ont été séparés. Les plasmas ont été congelés et conservés à -20°C pour
l’analyse des réponses immunitaires humorales. Les culots sanguins ont servis pour
l’extraction d’ADN génomique humain. Les gouttes épaisses ont été préparées et colorées au
Giemsa pour la détection des parasites à l’inclusion.
Après l’échantillonnage initial en juillet 2003, une détection active des cas de paludisme était
faite par un suivi clinique hebdomadaire de chaque participant pendant près de 23 semaines
(jusqu’à janvier 2004). À chaque visite, la température axillaire du patient était prise avec un
thermomètre digital. En cas de fièvre (température > 37,5 ° C), un test rapide de paludisme
(ICT Malaria Pf/PvTM: HRP, AMRAD, Australia) était effectué. Si le test était positif, une
goutte épaisse était aussi faite pour déterminer la densité parasitaire par microscopie.
Toutes les personnes malades ont été traitées conformément aux directives nationales en
vigueur (Pyrimethamine-sulfadoxine ou Amodiaquine). Tous les participants ayant eu des
épisodes de fièvre en dehors du suivi actif étaient référés à l’hôpital provincial de Farafenni
et pris en charge par un médecin.
Deux enquêtes transversales impliquant l’ensemble des participants ont été réalisées, pour
estimer le taux d’infection des sujets pendant les périodes de haute transmission (fin

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 70
octobre début novembre 2003) et de faible transmission (mi-décembre 2003) du paludisme.
Les informations concernant l’utilisation ou non des moustiquaires imprégnées et l’état de
l’habitat ont été aussi recueillies à l’inclusion. Une enquête entomologique a été réalisée
dans les villages pour estimer le taux d'inoculation entomologique (EIR). Un questionnaire a
été rempli pour recueillir les informations sur l’origine ethnique de chaque participant
(groupe ethnique des parents, et des grands parents paternels et maternels). Le paludisme a
été défini comme une fièvre (température axillaire > 37,5 ° C) avec densité parasitaire >
5000/µl (Migot-Nabias et al, 2000). L'étude a été approuvée par le comité d’éthique mixte
gouvernement de Gambie / MRC.
II.2. Mesure des anticorps dirigés contre P. falciparum par Enzyme Linked Immunosorbent Assay (ELISA)
II.2.1. ELISA MSP119 et AMA1
Les réponses immunitaires humorales dirigées contre MSP119 et AMA1 ont été
mesurées dans les plasmas par (ELISA). Brièvement, les plaques ELISA (Immunlon4,
Dynatech) sont recouvertes avec 1µg/ml d’antigène (MSP119 ou AMA1) dans du tampon
carbonate et incubées pendant 16 heures à 4 ˚ C. Les plaques sont lavées trois fois avec une
solution de PBS-Tween (0,05%), puis on y ajoute 150 µL d’une solution de blocage de PBS-
BSA à 1%. Les plaques sont incubées pendant 3 heures à température ambiante. La solution
de blocage est retirée des plaques. Une dilution sériée des plasmas est ajoutée dans les
plaques (1/1000; 1/2000; 1/4000; 1/8000 pour MSP119 et 1/1000 et 1/2000 pour AMA1).
Une dilution sériée d’IgG humain purifiée et d’anti-IgG-POD (Dako A/S, Danemark) ont été
utilisés pour servir de standard dans chaque plaque pour déterminer les concentrations
relatives d’anticorps anti-MSP119 et AMA1.
Les plaques sont incubées pendant 16 heures à 4°C, ensuite elles sont lavées 6 fois avec la
solution de lavage de PBS-Tween (0,05%). Les anticorps anti-IgG-HRP, anti-IgG1-HRP et anti-
IgG3-HRP (The Binding Site, UK) sont ajoutés dans les plaques à la dilution de 1/5000 dans
une solution de PBS-BSA à 1%. Les plaques sont incubées à température ambiante pendant 3
heures, puis lavées 6 fois. La réaction est développée avec 100 µL d’une solution de

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 71
tetramethyl-benzidine (TMB) (Sigma, UK). Les réactions sont stoppées avec 100 µL de
solution d’acide sulfurique 2M et elles sont lues le plus tôt possible à 450 nm.
Les plasmas d’adultes vivants en zone d’endémie (Brefet) ont servi de contrôles positifs. Un
pool de plasmas d’adultes européens n’ayant jamais eu de contact avec le paludisme a servi
de contrôle négatif. Le seuil de détection a été défini comme étant la moyenne ± 3
déviations standard de la concentration du contrôle négatif.
II.2.2. ELISA pour mesurer les anticorps anti-R32LR (CSP)
Pour mesurer la réponse immunitaire humorale dirigée contre la séquence répétitive
de CSP de P. falciparum, les plaques ELISA (Immunlon 4, Dynatech) sont enduites avec 0,1
µg d’antigène R32LR dans 50 µL de PBS et caséine [4 µL de caséine bouillie (0,5% de caséine,
rouge de phénol, Thimérosal, PBS, 5N NaOH)] pour 5 mL d’antigène dilué.
Cinquante (50) microlitres de l’antigène dilué sont mis dans les plaques ELISA en triplicata.
Les plaques sont incubées pendant 16 heures à température ambiante. Elles sont ensuite
bloquées par 250 µL d’une solution de blocage de caséine bouillie-Tween20 à 0,5% pendant
une heure à température ambiante. Le contenu de la plaque est vidé, et on y ajoute 50 µL
des plasmas dilués à la concentration appropriée. Chaque plaque comprend un contrôle
négatif, un contrôle positif (DGP) et un blanc. Les échantillons sont mis dans les puits en
triplicata à la dilution 1/50, suivi de dilutions sériées de 1/2.
Les plaques sont incubées pendant 2 heures à température ambiante puis lavées avec du
PBS-Tween20 à 0,5%. Dans chaque puits un volume de 50 µL d’anticorps anti-IgG humain
couplé à la peroxydase est ajouté. Les plaques sont incubées à température ambiante
pendant 1 heure. Les plaques sont ensuite lavées 4 fois avec du PBS-Tween20 à 0,5%.
Les IgG de chèvres anti-humaines couplées au HRP (Kirkegaard et Perry Laboratories) sont
utilisées à une dilution de 1/1000. La révélation de la réaction est faite en utilisant 100 µL
d’une solution de substrat ABTS HRP (Kirkegaard et Perry Laboratories) après une incubation
pendant 1h à température ambiante. La réaction est arrêtée avec 10 µL de SDS à 10%
(Sigma, Allemagne). La lecture des plaques est faite à 414 nm.
La moyenne de la densité optique (DO) du sérum testé en triplicata est calculée, cette
moyenne est multipliée par le facteur de dilution pour obtenir la moyenne pondérée (MP).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 72
La moyenne pondérée de chaque plaque est calculée de la façon suivante :
MPcorrigée = moyenne MPDGP x MPéchantillon / MPDGP
Les concentrations en µg/ml pour chaque plaque sont calculées selon la formule ci dessous:
Concentration échantillon (µg/ml)= concentration DGP (9,8) x MPcorrigée échantillon/MPcorrigée DGP
II.3. Culture in vitro de P. falciparum et préparation d’extraits de
schizontes
La souche Pf164 de P. falciparum a été maintenue en culture par la technique
classique de Trager et Jensen (1976). Les globules rouges non parasités de donneurs de
sang de groupe sanguin O ont été collectés dans les tubes citrate (ACD) [0,6% citrate
d'hydrogène disodique, 0,4% de dextrose (D-glucose), Sigma, St Louis, MO, USA].
Les cultures des parasites sont repiquées par l’ajout de globules rouges frais chaque
semaine. Le mélange des globules rouges parasités avec ceux non parasités a permis
d’obtenir les parasitémies désirées. Les flacons sont mis en culture sous jarre à 37°C. Le
milieu de culture est changé tous les jours et une goutte épaisse est faite pour déterminer la
parasitémie et le stade parasitaire de la culture.
Quand dans la culture la parasitémie est suffisamment élevée, elle est lavée et les schizontes
sont concentrés dans une solution de Percoll à 72% (Sigma) et resuspendus à un taux
d’hématocrite de 20%. Les schizontes subissent 4 fois un cycle de congélation et
décongélation et la concentration protéique est évaluée par la lecture de la densité optique
à 280 nm. Les extraits sont titrés avec des PBMCs d’adultes semi immuns.
L’activation maximale induite par l’extrait de schizonte à travers la production d’IFN- par les
PBMCs a été obtenue à une concentration de 10 µg/ml. L’extrait de globules rouges non
parasités de concentration identique a servi de contrôle négatif. Les cultures de P.
falciparum ont été testées à intervalles réguliers et déclarées exemptes de contamination
par des mycoplasmes en utilisant un kit à base d’une réaction en chaîne par polymérase
(PCR) (American Type Culture Collection; ATCC, Manassas, USA).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 73
II.4. Séparation et analyse des cellules T CD4+CD25+
Les cellules mononuclées du sang de cordon (CMSC) sont délestées en cellules T CD4+
CD25+ en utilisant des billes Miltenyi (Miltenyi Biotec, Bergisch Gladbach, Allemagne), en
suivant les instructions du fabricant. Brièvement, la déplétion des cellules T CD4+CD25+ est
réalisée en deux étapes. Dans la première étape , les cellules non TCD4+ sont marquées avec
un cocktail d’anticorps biotinylés anti- CD16, CD14, CD8, CD19, CD56 et TCRγδ et des
microbilles anti-biotine pour séparer cellules T CD4+ des non T CD4+. Les cellules marquées
sont déplétées sur les colonnes MACS®. Dans la seconde étape, la fraction pré-enrichie en T
CD4+ est marquée avec un anticorps anti-CD25+ biotinylé et des billes anti-biotine pour
séparer les cellules T CD4+CD25+ des T CD4+ CD25-. La fraction des cellules T CD4+CD25-
résultant de la séparation des cellules T CD4+CD25+ est ensuite ajoutée aux cellules non CD4+
pour reconstituer cette population lors d’expériences de stimulation in vitro. Les cellules T
CD4+CD25+ obtenues par sélection positive sont éluées des colonnes et mises en culture
(106/ml) pendant 24h en présence de phytohémagglutinine (PHA) pour évaluer leur
production d’IL-10 comparativement aux cellules T CD4+CD25-.
Les anticorps anti-CD25 et anti-CD4 couplés respectivement à la phycoérythrine (anti-CD25-
PE) et à la fluorescéine isothiocyanate (anti-CD4-FITC) (Becton-Dickinson, UK) ont été utilisés
pour contrôler la qualité des séparations mais aussi pour comparer la fréquence des cellules
T CD4+CD25+ dans les groupes de placentas parasités ou non parasités.
Pour les analyses de contrôle de qualité, les cellules mononuclées du sang de cordon ont été
décongelées et colorées pendant 20 mn à 4°C. Elles sont ensuite lavées avec un tampon de
PBS-BSA à 2% et analysées par un cytomètre de flux (FACS Calibur, Becton-Dickinson, UK).
II.5. Culture des cellules mononuclées du sang de cordon
Une sélection de CMSCs congelées du groupe de participants de Sukuta, a été
décongelée et lavée avec du RPMI-1640 (Sigma, St Louis, MO, USA). Les cellules ont été
remises en suspension dans du RPMI-1640 supplémenté avec 20 µg/ml de gentamycine
(Sigma), 2 mM de L-glutamine (Sigma) et 10% de sérum d’anticorps humains (Sigma).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 74
La viabilité des CMSCs était de 85%. Les CMSCs ont été cultivées (2 x 105 cellules dans 200 µl
de milieu complet) pendant 3 jours en présence de PHA (PHA-L 2,5µg/ml; Sigma) ou pendant
6 jours en présence d'érythrocytes infectés par P. falciparum (Pf164) ayant une parasitémie
de 10% ou en présence de d’extrait de schizontes de 10 µg/ml. Les érythrocytes non infectés
ou les extraits provenant de ces derniers ont été utilisées comme témoins négatifs.
L’antigène MSP119 (Morgan et al, 1999) a été utilisé à une concentration de 5 µg/ml pour
étudier la réponse des lymphocytes T. Tous les essais ont été effectués en triplicata. Les
cytokines (IFN-, IL-10 et sCD30 solubles) ont été mesurées dans les surnageants de culture
en utilisant la technique ELISA.
II.6. Détection des cellules T CD4+ productrices d’IFN-γ par culture
ELISPOT (Enzyme Linked Immunoassay Spot)
Les PBMCs ont été séparés du sang veineux par centrifugation dans un gradient de
densité sur Ficoll (Pharmacia, Oslo, Norvège) à 2200 tr/min pendant 20 min dans une
centrifugeuse de paillasse. Les cellules ont été lavées dans du milieu RPMI 1640 (Sigma, Saint
Louis, USA), puis remises en suspension dans le même milieu supplémenté avec 2 mM L-
Glutamine (Life Technologies), 100 UI/ml de pénicilline, 100 µg/ml de streptomycine (Life
Technologies) et 10% de sérum humain du groupe sanguin AB+ (Sigma, Saint Louis, USA)
inactivé par la chaleur (30 min à 56°C).
2 x 105 PBMCs par puits ont été cultivés à 37 ° C dans un incubateur à CO2 en présence de
formes sauvages ou modifiées des peptides MSP119 (25 µg/ml) ou de protéines (10 µg/ml).
La PHA à 2,5 µg/ml et 10 µg/ml d’un dérivé protéique purifié de Mycobacterium tuberculosis
(PPD RT49, States Serum Institute, Denmark), ont été utilisés comme contrôles positifs.
Les surnageants sont collectés 72 heures après le début de la culture et congelés à -20°C
pour le dosage des cytokines. Aux jours 3 et 7 sont ajoutés aux cultures 20U/ml d’IL-2
(Proleukine). Au jour 10 de la culture, les cellules sont lavées et transférées dans les plaques
ELISPOT (Millipore MAIPS45 PVDF-backed plates).
L'activation des PBMCs a été réalisée pendant 24 heures en présence de peptides ou
protéines de MSP119; le test ELISPOT a été réalisé en utilisant le kit ELISPOT IFN- (MABTECH,
Stockholm, Suède) en suivant les recommandations du fabricant. Brièvement, 4 x 105

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 75
PBMCs/puits sont mis en culture dans les plaques ELISPOT (MAIP 45S; Millipore, Molsheim,
France) contenant des anti-IFN- (Mabtech, Nadra, Sweden). Les plaques sont lavées 6 fois
avec du PBS-Tween 20 (Sigma, Irvine, CA, USA) à 0,05%. On ajoute dans les plaques le
second anticorps biotinylé puis elles sont incubées à température ambiante pendant 2
heures. Les plaques sont lavées 6 fois avec la même solution de lavage puis sont incubées en
présence de streptavidine couplée à la phosphatase alcaline. Après un lavage, le substrat de
la phosphatase alcaline est ajouté. On incube 5 à 10 min jusqu'à l’apparition des spots. La
réaction est arrêtée par lavage à l’eau. Les cellules productrices d’IFN- sont dénombrées
par un système informatisé et exprimées en spot-forming units (SFU) par million de PBMC.
II.7. Extraction de l'ARN et transcription inverse de Foxp3
L’ARN total a été extrait des fractions des T CD4+CD25+ et T CD4+CD25- en utilisant un
kit d’extraction de l’ARN total (Promega Corporation, Madison, WI, USA) en suivant les
instructions du fabricant. La synthèse de L’ADN complémentaire est faite par transcription
inverse à 37°C pendant 1h30mn, suivie d’incubation à 95°C pendant 5 min pour désactiver la
transcriptase inverse. Foxp3 et Glycéraldéhyde-3-phosphate-déshydrogénase (GAPDH)
(Contrôle positif) ont été amplifiés par PCR dans un volume réactionnel de 25 µl contenant 1
µl d’ADNc, 0,2 µmol de chaque amorce, 0,5unité de Taq Polymérase (Boehringer,
Mannheim, Germany), 0,1 µmol de chaque dNTP et 1X de Tampon Polymérase. Les
séquences des amorces pour Foxp3 et GADPH sont les suivantes : GADPH (sens) 5’-
GCAAATTC CATGGCACCGT-3’ ; GADPH (anti-sens) 5’-TCGCCCCACTTGATTTTGG-3’ ; Foxp3
(sens) 5’- AGCTGGAGTTCCGCAAGAAAC-3’ et Foxp3 (anti-sens) 5’-
TGTTCGTCCATCCTCCTTTCC-3’.
II.8. Génotypage de la G6PD et de l’hémoglobine S par PCR en temps
réel
L'ADN a été extrait à partir de PBMCs en utilisant le kit d'extraction d'ADN Puregene
(Flowgen, Ashby de la Zouch, Royaume-Uni) ou la méthode standard salting-out. Le déficit
en G6PD et l’'hémoglobine S ont été génotypés en utilisant les sondes Taqman (ABI, Applera

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 76
International Inc, Foster City, CA, USA) suivantes, rs1050828 (G6PD 202G/A), rs1050829
(G6PD 376A/G) et rs334 (HbS), dans un volume réactionnel de 10 µl, en utilisant soit le
Rotor-Gene 3000 (Corbett Robotics Pty Ltd, Brisbane, Queensland, Australie ) ou le système
PCR en Temps réel 7500 ABI. Les courbes de fluorescence ont été analysés avec la version 6
du logiciel Rotor-Gene et le système de détection de séquence 7500 ABI version 1.1.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 77

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 78
III. Résultats
III.1. Article I. L’infection placentaire par Plasmodium falciparum interfère sur l’équilibre Th1/Th2 des cellules T néonatales à travers les cellules T régulatrices CD4+CD25+FoxP3+ et la production d’IL-10
C. Bisseye*, M. van der Sande*, W. D. Morgan†, A. A. Holder†, M. Pinder* and J. Ismaili*
*Medical Research Council Laboratories, Banjul, The Gambia, and †Division of Parasitology,
National Institute for Medical Research, Mill Hill, London, U
Clinical and Experimental Immunology (2009), 158: 287–293
III.1.1. But de l’étude
Des études antérieures ont montré que Plasmodium falciparum induit un biais Th2 dans
l’infection palustre au cours de la grossesse. L’infection placentaire affecte aussi l’équilibre
Th1/Th2 des cellules T néonatales. Nous avons étudié le rôle potentiel des cellules T
régulatrices dans cet équilibre en comparant les réponses des lymphocytes T des cellules
mononuclées du sang de cordon (CMSCs) provenant de placentas de femmes gambiennes
parasitées et non parasitées par Plasmodium falciparum.
III.1.2. Population
La description du groupe de femmes gambiennes utilisées dans la présente étude a déjà été
faite par Ismaili et al, (2003). Le tableau I résume les données épidémiologiques de ce
groupe de femmes de l’étude.
Tableau I : Données épidémiologiques des femmes gambiennes de Sukuta.
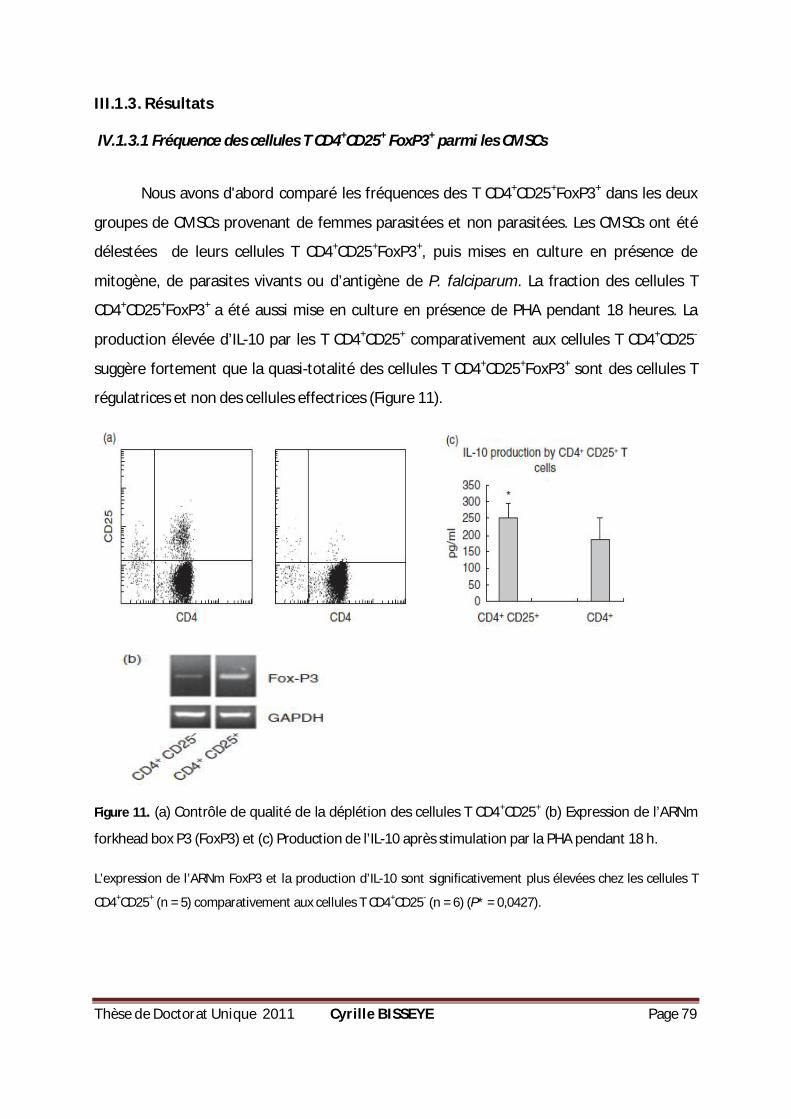
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 79
III.1.3. Résultats
IV.1.3.1 Fréquence des cellules T CD4+CD25+ FoxP3+ parmi les CMSCs
Nous avons d'abord comparé les fréquences des T CD4+CD25+FoxP3+ dans les deux
groupes de CMSCs provenant de femmes parasitées et non parasitées. Les CMSCs ont été
délestées de leurs cellules T CD4+CD25+FoxP3+, puis mises en culture en présence de
mitogène, de parasites vivants ou d’antigène de P. falciparum. La fraction des cellules T
CD4+CD25+FoxP3+ a été aussi mise en culture en présence de PHA pendant 18 heures. La
production élevée d’IL-10 par les T CD4+CD25+ comparativement aux cellules T CD4+CD25-
suggère fortement que la quasi-totalité des cellules T CD4+CD25+FoxP3+ sont des cellules T
régulatrices et non des cellules effectrices (Figure 11).
Figure 11. (a) Contrôle de qualité de la déplétion des cellules T CD4+CD25+ (b) Expression de l’ARNm
forkhead box P3 (FoxP3) et (c) Production de l’IL-10 après stimulation par la PHA pendant 18 h.
L’expression de l’ARNm FoxP3 et la production d’IL-10 sont significativement plus élevées chez les cellules T
CD4+CD25+ (n = 5) comparativement aux cellules T CD4+CD25- (n = 6) (P* = 0,0427).
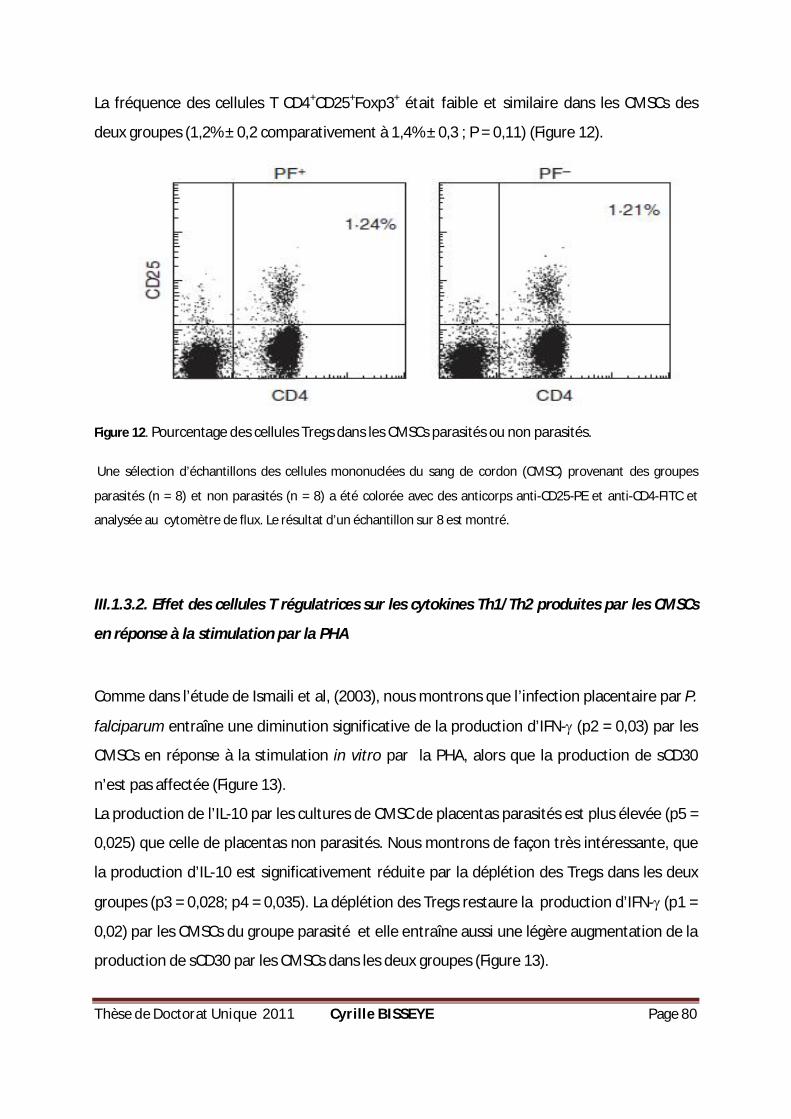
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 80
La fréquence des cellules T CD4+CD25+Foxp3+ était faible et similaire dans les CMSCs des
deux groupes (1,2% ± 0,2 comparativement à 1,4% ± 0,3 ; P = 0,11) (Figure 12).
Figure 12. Pourcentage des cellules Tregs dans les CMSCs parasités ou non parasités.
Une sélection d’échantillons des cellules mononuclées du sang de cordon (CMSC) provenant des groupes
parasités (n = 8) et non parasités (n = 8) a été colorée avec des anticorps anti-CD25-PE et anti-CD4-FITC et
analysée au cytomètre de flux. Le résultat d’un échantillon sur 8 est montré.
III.1.3.2. Effet des cellules T régulatrices sur les cytokines Th1/Th2 produites par les CMSCs
en réponse à la stimulation par la PHA
Comme dans l’étude de Ismaili et al, (2003), nous montrons que l’infection placentaire par P.
falciparum entraîne une diminution significative de la production d’IFN- (p2 = 0,03) par les
CMSCs en réponse à la stimulation in vitro par la PHA, alors que la production de sCD30
n’est pas affectée (Figure 13).
La production de l’IL-10 par les cultures de CMSC de placentas parasités est plus élevée (p5 =
0,025) que celle de placentas non parasités. Nous montrons de façon très intéressante, que
la production d’IL-10 est significativement réduite par la déplétion des Tregs dans les deux
groupes (p3 = 0,028; p4 = 0,035). La déplétion des Tregs restaure la production d’IFN- (p1 =
0,02) par les CMSCs du groupe parasité et elle entraîne aussi une légère augmentation de la
production de sCD30 par les CMSCs dans les deux groupes (Figure 13).
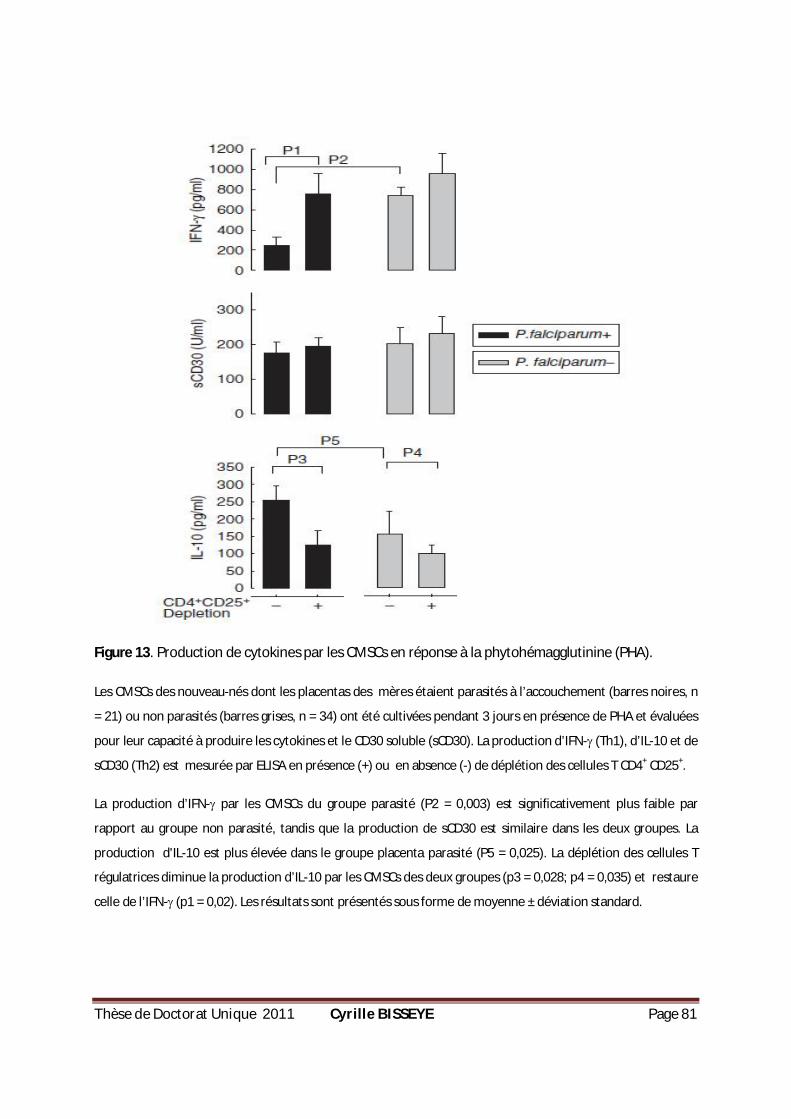
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 81
Figure 13. Production de cytokines par les CMSCs en réponse à la phytohémagglutinine (PHA).
Les CMSCs des nouveau-nés dont les placentas des mères étaient parasités à l’accouchement (barres noires, n
= 21) ou non parasités (barres grises, n = 34) ont été cultivées pendant 3 jours en présence de PHA et évaluées
pour leur capacité à produire les cytokines et le CD30 soluble (sCD30). La production d’IFN- (Th1), d’IL-10 et de
sCD30 (Th2) est mesurée par ELISA en présence (+) ou en absence (-) de déplétion des cellules T CD4+ CD25+.
La production d’IFN- par les CMSCs du groupe parasité (P2 = 0,003) est significativement plus faible par
rapport au groupe non parasité, tandis que la production de sCD30 est similaire dans les deux groupes. La
production d'IL-10 est plus élevée dans le groupe placenta parasité (P5 = 0,025). La déplétion des cellules T
régulatrices diminue la production d’IL-10 par les CMSCs des deux groupes (p3 = 0,028; p4 = 0,035) et restaure
celle de l’IFN- (p1 = 0,02). Les résultats sont présentés sous forme de moyenne ± déviation standard.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 82
III.1.3.3. Effets des Tregs sur les cytokines Th1/Th2 produites par les CMSCs en réponses
aux globules rouges intacts parasités et aux antigènes de P. falciparum
Les CMSCs ont également été testés pour leur capacité à produire in vitro les cytokines en
réponse à leur stimulation par les globules rouges infectés par P. falciparum, les extraits de
schizontes Pf164 et la protéine purifiée MSP119 après 6 jours en culture (Figure 14).
Nous montrons une baisse de la production d'IFN- (schizontes Pf164 p1 = 0,0485; MSP119
p2 = 0,00497) et une augmentation de celle de l'IL-10 (schizontes Pf164 p12 = 0,048;
MSP119 p13 = 0,0478) par les CMSCs en réponse à la stimulation par les extraits de
schizontes Pf164, les globules rouges infectés (Live Pf) et l’antigène MSP119 dans le groupe
parasité par rapport au groupe non parasité.
En outre, la production de sCD30 observée en réponse à la stimulation par les antigènes de
P. falciparum (schizontes Pf164 p7 = 0,0245; MSP119 p8 = 0,021) est plus élevée dans le
groupe parasité.
La déplétion des Tregs affecte la production des cytokines par les CMSCs. Elle restaure la
production de l’IFN-, réduit celle de l’IL-10 et augmente la production du sCD30 dans les
deux groupes (Figure 14). Dans le cas du sCD30, l’augmentation reste significative en
réponse aux antigènes de P. falciparum (schizontes Pf164 p10 = 0,032; MSP119 p11 = 0,041).
Les globules rouges infectés par P. falciparum (Live Pf) induisent des concentrations les plus
élevées de toutes les cytokines testées par rapport aux antigènes (schizontes Pf164 et
MSP119). Une plus faible concentration d’IFN- (p6 = 0,015) et de plus fortes productions
d’IL-10 (p15 = 0,038) et de sCD30 (p9 = 0,028) sont observées dans le groupe parasité.
En outre, la déplétion des cellules T CD4+CD25+ réduit la production d’IL-10 (p14 = 0,049),
elle restaure celle de l'IFN-(p3 = 0,0209) et augmente la production de sCD30 dans les deux
groupes conduisant à une production similaire dans ces deux derniers.
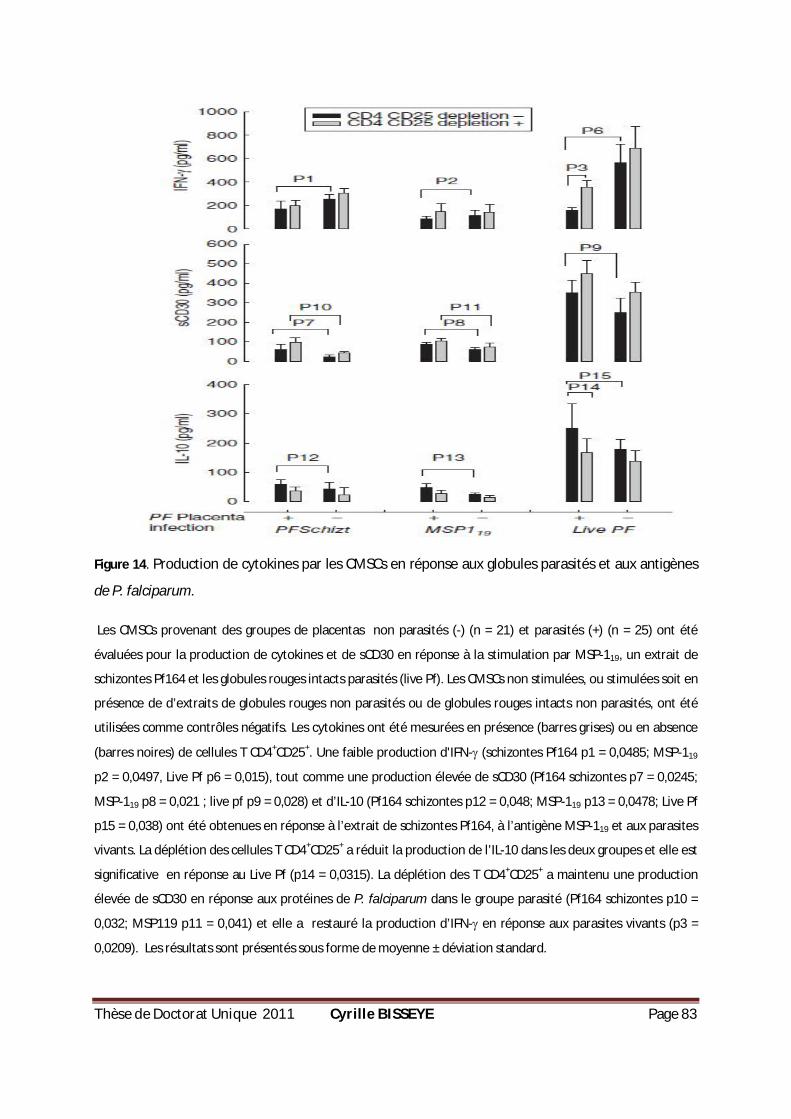
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 83
Figure 14. Production de cytokines par les CMSCs en réponse aux globules parasités et aux antigènes
de P. falciparum.
Les CMSCs provenant des groupes de placentas non parasités (-) (n = 21) et parasités (+) (n = 25) ont été
évaluées pour la production de cytokines et de sCD30 en réponse à la stimulation par MSP-119, un extrait de
schizontes Pf164 et les globules rouges intacts parasités (live Pf). Les CMSCs non stimulées, ou stimulées soit en
présence de d’extraits de globules rouges non parasités ou de globules rouges intacts non parasités, ont été
utilisées comme contrôles négatifs. Les cytokines ont été mesurées en présence (barres grises) ou en absence
(barres noires) de cellules T CD4+CD25+. Une faible production d'IFN- (schizontes Pf164 p1 = 0,0485; MSP-119
p2 = 0,0497, Live Pf p6 = 0,015), tout comme une production élevée de sCD30 (Pf164 schizontes p7 = 0,0245;
MSP-119 p8 = 0,021 ; live pf p9 = 0,028) et d’IL-10 (Pf164 schizontes p12 = 0,048; MSP-119 p13 = 0,0478; Live Pf
p15 = 0,038) ont été obtenues en réponse à l’extrait de schizontes Pf164, à l’antigène MSP-119 et aux parasites
vivants. La déplétion des cellules T CD4+CD25+ a réduit la production de l’IL-10 dans les deux groupes et elle est
significative en réponse au Live Pf (p14 = 0,0315). La déplétion des T CD4+CD25+ a maintenu une production
élevée de sCD30 en réponse aux protéines de P. falciparum dans le groupe parasité (Pf164 schizontes p10 =
0,032; MSP119 p11 = 0,041) et elle a restauré la production d’IFN- en réponse aux parasites vivants (p3 =
0,0209). Les résultats sont présentés sous forme de moyenne ± déviation standard.
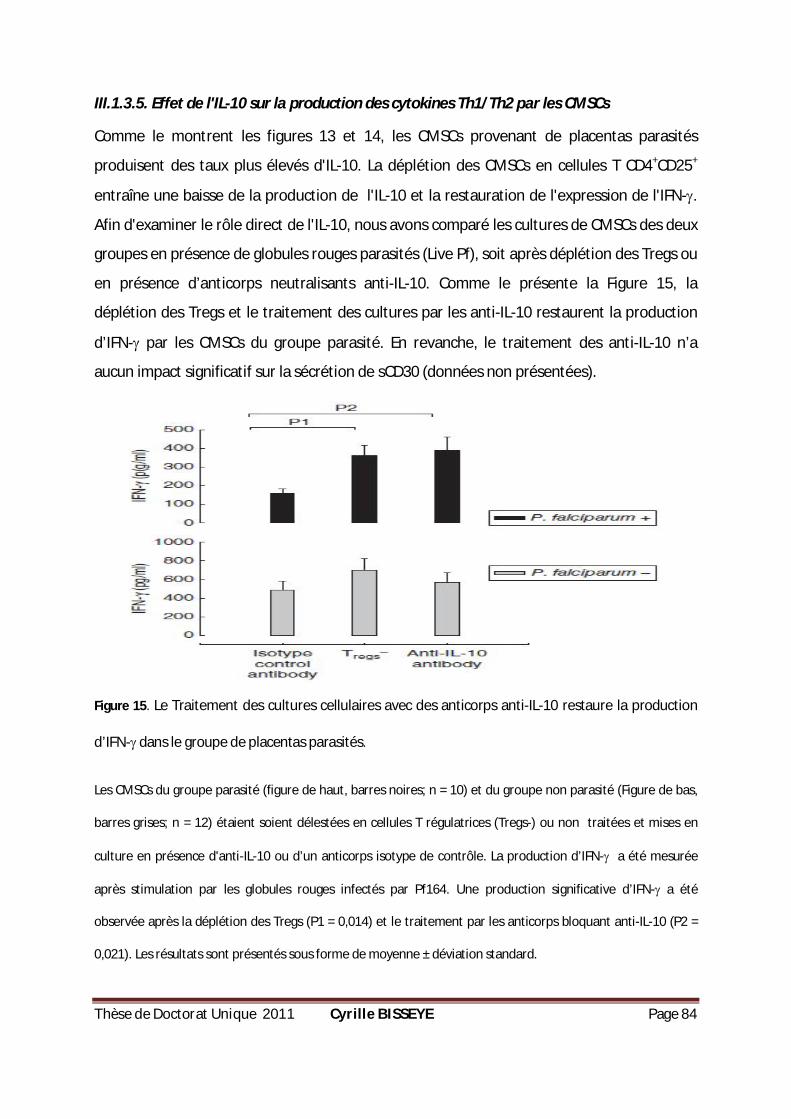
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 84
III.1.3.5. Effet de l'IL-10 sur la production des cytokines Th1/Th2 par les CMSCs
Comme le montrent les figures 13 et 14, les CMSCs provenant de placentas parasités
produisent des taux plus élevés d'IL-10. La déplétion des CMSCs en cellules T CD4+CD25+
entraîne une baisse de la production de l'IL-10 et la restauration de l'expression de l'IFN-.
Afin d'examiner le rôle direct de l'IL-10, nous avons comparé les cultures de CMSCs des deux
groupes en présence de globules rouges parasités (Live Pf), soit après déplétion des Tregs ou
en présence d’anticorps neutralisants anti-IL-10. Comme le présente la Figure 15, la
déplétion des Tregs et le traitement des cultures par les anti-IL-10 restaurent la production
d’IFN- par les CMSCs du groupe parasité. En revanche, le traitement des anti-IL-10 n’a
aucun impact significatif sur la sécrétion de sCD30 (données non présentées).
Figure 15. Le Traitement des cultures cellulaires avec des anticorps anti-IL-10 restaure la production
d’IFN- dans le groupe de placentas parasités.
Les CMSCs du groupe parasité (figure de haut, barres noires; n = 10) et du groupe non parasité (Figure de bas,
barres grises; n = 12) étaient soient délestées en cellules T régulatrices (Tregs-) ou non traitées et mises en
culture en présence d'anti-IL-10 ou d’un anticorps isotype de contrôle. La production d’IFN- a été mesurée
après stimulation par les globules rouges infectés par Pf164. Une production significative d’IFN- a été
observée après la déplétion des Tregs (P1 = 0,014) et le traitement par les anticorps bloquant anti-IL-10 (P2 =
0,021). Les résultats sont présentés sous forme de moyenne ± déviation standard.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 85
III.1.4. Conclusion
Le déséquilibre Th1/Th2 des cellules T néonatales dû à l'infection active du placenta par P.
falciparum, est une stratégie possible utilisée par le parasite pour échapper à la réponse
immunitaire de l’hôte. Elle implique les Tregs à travers leur production d’IL-10, comme le
révèlent leur déplétion et l’inhibition de l’IL-10 qui restaurent la production d’IFN-. Nos
résultats suggèrent que la réduction de la production de l’IFN- pourrait être incriminée à la
fois aux CPA par leur faible production d’IL-12 et aux Tregs à travers l’IL-10, d’où une
exacerbation de la polarisation Th2.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 86
III.2. Article II. Un variant de la protéine de 19 kilodaltons de
l’extrémité C-terminal du Merozoite Surface Protein 1 de Plasmodium
falciparum candidat vaccin induit des taux élevés d’IFN-gamma
associés à des réponses immunitaires cellulaires à des séquences
peptidiques spécifiques chez les adultes gambiens naturellement
exposés au paludisme
Cyrille Bisseye*‡, Louis M. Yindom*, Jacques Simporé‡, William D. Morgan†, Anthony A.
Holder† and Jamila Ismaili*2#
*Medical Research Council Laboratories, PO Box 273, Banjul, The Gambia; †Division of Parasitology,
MRC National Institute for Medical Research, Mill Hill, London NW7 1AA, United Kingdom; ‡Centre de
Recherche Biomoléculaire Pietro Annigoni, Ouagadougou, Burkina Faso.
Clinical and Experimental Immunology (2011), 166 (3):366-73
III.2.1. But de l’étude
La protéine de 19 kDa de MSP119 de P. falciparum est un important candidat vaccin du stade
sanguin du paludisme. Elle est la cible des réponses immunitaires humorales et cellulaires
chez les individus naturellement infectés par P. falciparum. Des variants génétiquement
modifiés de cette protéine ont été conçus pour être potentiellement de meilleurs candidats
vaccins. Cependant, la réponse immunitaire humaine à ces protéines n'a pas encore été
entièrement caractérisée. Nous avons donc étudié d’une part, l'antigénicité d'une protéine
triple mutant de MSP119 par rapport à celle de la protéine sauvage, et d’autre part
l’antigénicité des peptides mutés par rapport aux peptides non mutés chez les adultes vivant
dans une zone endémique en Gambie.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 87
III.2.2. Population
Quarante trois (43) adultes gambiens ont été recrutés dans cette étude. L’antigénicité des
protéines et peptides de MSP119 a été évaluée par ELISPOT en culture et par ELISA en
mesurant respectivement, les cellules T CD4+ sécrétrices d’IFN- et les cytokines Th1 et Th2
produites en réponses à ces derniers.
III.2.3. Résultats
III.2.3.1. Réponses des adultes gambiens aux protéines MSP119 par ELISPOT en culture
La réponse des cellules des adultes gambiens aux protéines MSP119 a été évaluée en premier
par ELISPOT en culture, en mesurant le nombre de cellules T CD4+ productrices d’IFN- après
stimulation par ces protéines (mutant et sauvage). Nous avons obtenu 27% (11/43) de
répondeurs pour la protéine normale contre 30% (13/43) pour la protéine triple mutant, la
différence n’était pas significative (654 ± 189 et 721 ± 299 SFU/106 PBMCs respectivement,
pour la protéine sauvage et le triple mutant).
III.2.3.2. Production des cytokines en réponses aux protéines MSP119
Nous avons ensuite mesuré la production de l’IFN-, de l’IL-13 et du marqueur Th2 CD30
soluble par les PBMCs d’adultes gambiens en réponse aux protéines normale et triple
mutant. Les réponses obtenues sont indiquées sur les figures 16 ci- dessous.
Une production significativement élevée d’IFN- par les PBMCs a été obtenue en réponse à
la protéine triple mutant par rapport à la protéine sauvage (4500 ± 256 et 3186 ± 267, P* =
0,015) (figure 16a). La production d’IL-13 et de CD30 soluble induite en réponse aux deux
protéines était identique (Figures 16b et 16c).
III.2.3.3. Réponses des adultes gambiens aux peptides MSP119 par ELISPOT en culture
Nous avons évalué la fréquence des répondeurs aux peptides normaux ou mutants de
MSP119 (Tableau II) chez les adultes gambiens pour déterminer le ou les peptides
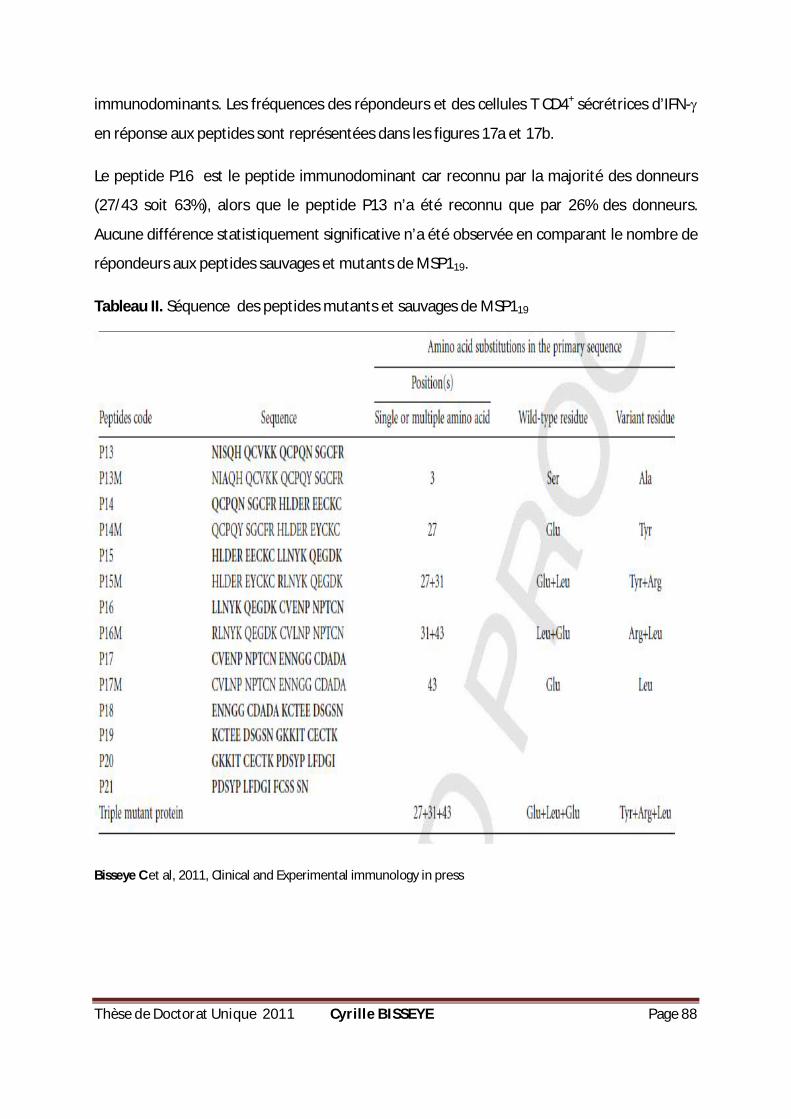
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 88
immunodominants. Les fréquences des répondeurs et des cellules T CD4+ sécrétrices d’IFN-
en réponse aux peptides sont représentées dans les figures 17a et 17b.
Le peptide P16 est le peptide immunodominant car reconnu par la majorité des donneurs
(27/43 soit 63%), alors que le peptide P13 n’a été reconnu que par 26% des donneurs.
Aucune différence statistiquement significative n’a été observée en comparant le nombre de
répondeurs aux peptides sauvages et mutants de MSP119.
Tableau II. Séquence des peptides mutants et sauvages de MSP119
Bisseye C et al, 2011, Clinical and Experimental immunology in press
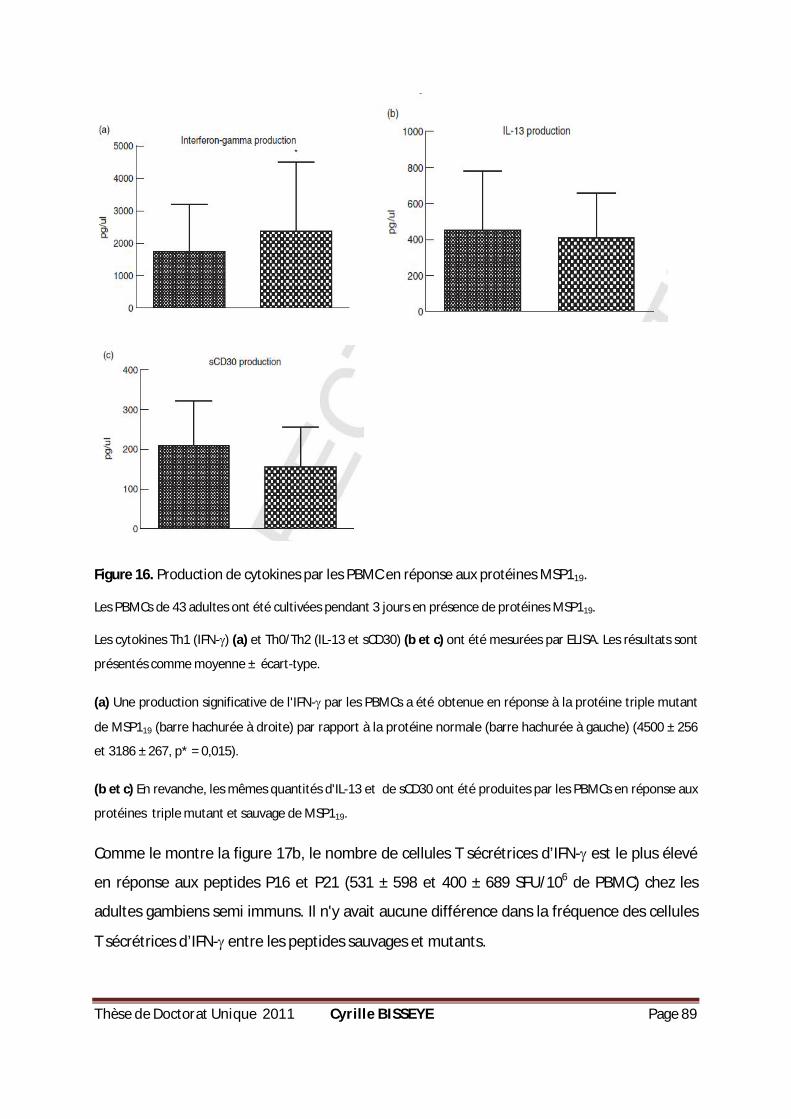
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 89
Figure 16. Production de cytokines par les PBMC en réponse aux protéines MSP119.
Les PBMCs de 43 adultes ont été cultivées pendant 3 jours en présence de protéines MSP119.
Les cytokines Th1 (IFN-) (a) et Th0/Th2 (IL-13 et sCD30) (b et c) ont été mesurées par ELISA. Les résultats sont
présentés comme moyenne ± écart-type.
(a) Une production significative de l'IFN- par les PBMCs a été obtenue en réponse à la protéine triple mutant
de MSP119 (barre hachurée à droite) par rapport à la protéine normale (barre hachurée à gauche) (4500 ± 256
et 3186 ± 267, p* = 0,015).
(b et c) En revanche, les mêmes quantités d'IL-13 et de sCD30 ont été produites par les PBMCs en réponse aux
protéines triple mutant et sauvage de MSP119.
Comme le montre la figure 17b, le nombre de cellules T sécrétrices d’IFN- est le plus élevé
en réponse aux peptides P16 et P21 (531 ± 598 et 400 ± 689 SFU/106 de PBMC) chez les
adultes gambiens semi immuns. Il n'y avait aucune différence dans la fréquence des cellules
T sécrétrices d’IFN- entre les peptides sauvages et mutants.
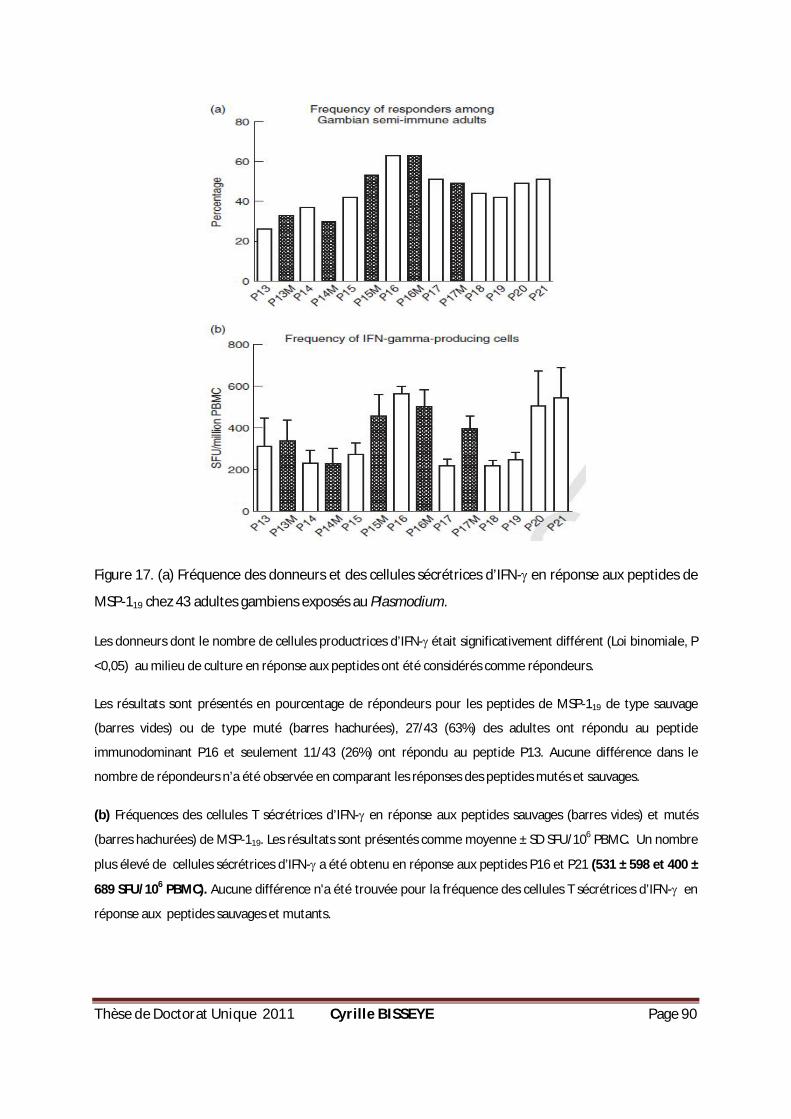
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 90
Figure 17. (a) Fréquence des donneurs et des cellules sécrétrices d’IFN- en réponse aux peptides de
MSP-119 chez 43 adultes gambiens exposés au Plasmodium.
Les donneurs dont le nombre de cellules productrices d’IFN- était significativement différent (Loi binomiale, P
<0,05) au milieu de culture en réponse aux peptides ont été considérés comme répondeurs.
Les résultats sont présentés en pourcentage de répondeurs pour les peptides de MSP-119 de type sauvage
(barres vides) ou de type muté (barres hachurées), 27/43 (63%) des adultes ont répondu au peptide
immunodominant P16 et seulement 11/43 (26%) ont répondu au peptide P13. Aucune différence dans le
nombre de répondeurs n’a été observée en comparant les réponses des peptides mutés et sauvages.
(b) Fréquences des cellules T sécrétrices d’IFN- en réponse aux peptides sauvages (barres vides) et mutés
(barres hachurées) de MSP-119. Les résultats sont présentés comme moyenne ± SD SFU/106 PBMC. Un nombre
plus élevé de cellules sécrétrices d’IFN- a été obtenu en réponse aux peptides P16 et P21 (531 ± 598 et 400 ±
689 SFU/106 PBMC). Aucune différence n'a été trouvée pour la fréquence des cellules T sécrétrices d’IFN- en
réponse aux peptides sauvages et mutants.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 91
III.2.3.4. Production de cytokines en réponse aux peptides mutants et sauvages de MSP119
Nous avons également mesuré la sécrétion d'IFN- (Th1), d’IL-13 et de CD30 soluble
(Th0/Th2) en réponse aux peptides sauvages et mutants de MSP119 comme indiquée sur les
figures 18a à 18c. Aucune différence n’a été observée dans la production d’IFN- entre les
peptides sauvages et mutants. La production d’IFN- était significativement plus élevée en
réponse au peptide P16 comparativement aux autres peptides (sauf pour les peptides P14,
P14M, P15M, P16M, P17M, et P21) (Figure 18a). La production d’IL-13 était plus élevée en
réponse aux peptides mutants par rapport aux peptides sauvages (P13M vs P13, P = 0,025;
P15M vs P15, P = 0,001; P17M vs P17, P = 0,03) (Figure 18b). La production d’IL-13 a été
significativement plus élevée en réponse au peptide P19 comparativement aux autres
peptides [P13M (p = 0,033); P14 (p = 0,046); P14M (p = 0,003); P15M (p < 0,001); P16 (p =
0,030); P16M (p = 0,003); P17 (p < 0,0001); P17M (p = 0,019); P18 (p = 0,002) et P21 (p =
0,019)], sauf pour les peptides P13, P15 et P20. La production de sCD30 par les PBMCs
d’adultes gambiens était identique en réponse aux peptides mutants et sauvages de MSP119.
Elle a été significativement plus élevée en réponse au P19 comparativement à tous les autres
peptides (Figure 18c). Les peptides P16 et P19 induisent deux types distincts de réponses
cytokiniques. P16 induit une forte sécrétion d’IFN- (Th1) alors P19 provoque une forte
production d’IL-13 et de CD30 soluble (Th0/Th2).
III.2.3.5.Typage HLA-DR et présentation des peptides MSP119 aux cellules T CD4+
La réponse cellulaire T CD4+ est souvent restreinte par les molécules HLA classe II. Les loci
HLA-DRB1, HLA-DRB3, HLA-DRB4, HLA-DR5 et HLA-DQB1 ont été typés chez 30 adultes
gambiens. Le tableau III montre la fréquence des allèles dans la population étudiée.
Aucun participant de l’étude n’était porteur d’allèles HLA DRB4 et DRB5.
Onze allèles DRB1 ont été trouvés parmi les participants de l'étude: 19 sur 30 (63%), 10 sur
30 (33%) et 7 sur 30 (23%) des donneurs portaient respectivement les allèles DRB1*13,
DRB1*11 et DRB1*10.
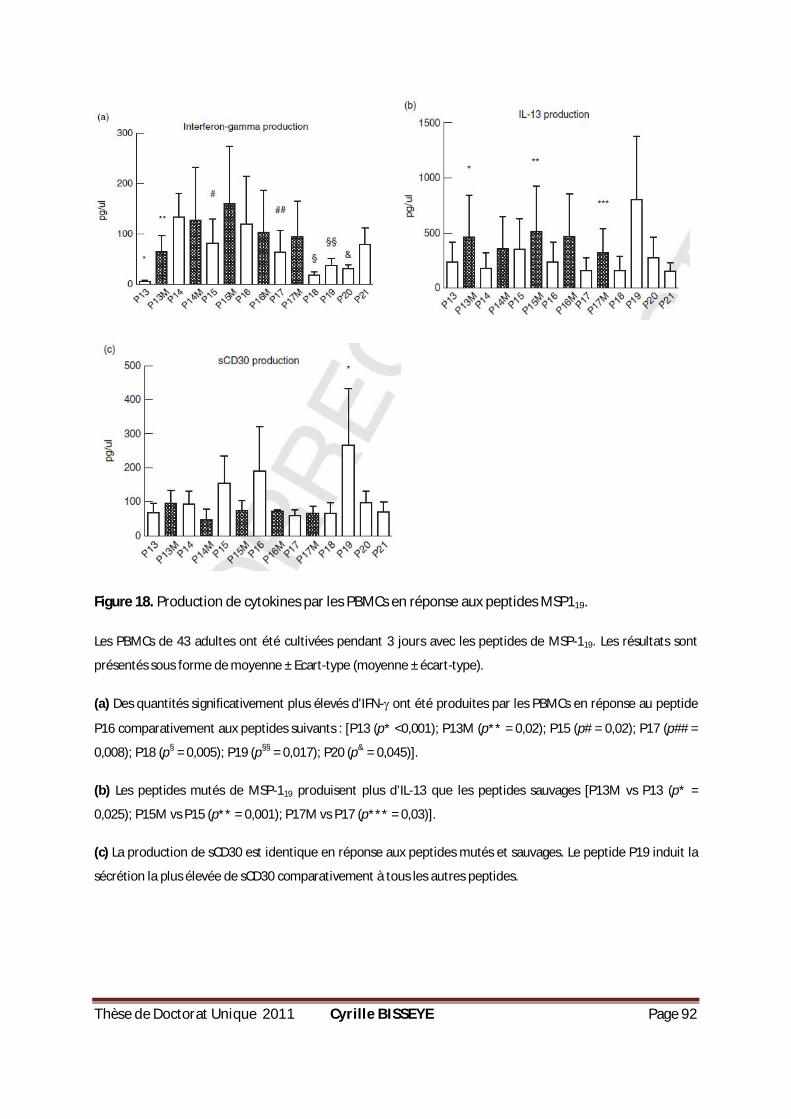
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 92
Figure 18. Production de cytokines par les PBMCs en réponse aux peptides MSP119.
Les PBMCs de 43 adultes ont été cultivées pendant 3 jours avec les peptides de MSP-119. Les résultats sont
présentés sous forme de moyenne ± Ecart-type (moyenne ± écart-type).
(a) Des quantités significativement plus élevés d'IFN- ont été produites par les PBMCs en réponse au peptide
P16 comparativement aux peptides suivants : [P13 (p* <0,001); P13M (p** = 0,02); P15 (p# = 0,02); P17 (p## =
0,008); P18 (p§ = 0,005); P19 (p§§ = 0,017); P20 (p& = 0,045)].
(b) Les peptides mutés de MSP-119 produisent plus d’IL-13 que les peptides sauvages [P13M vs P13 (p* =
0,025); P15M vs P15 (p** = 0,001); P17M vs P17 (p*** = 0,03)].
(c) La production de sCD30 est identique en réponse aux peptides mutés et sauvages. Le peptide P19 induit la
sécrétion la plus élevée de sCD30 comparativement à tous les autres peptides.
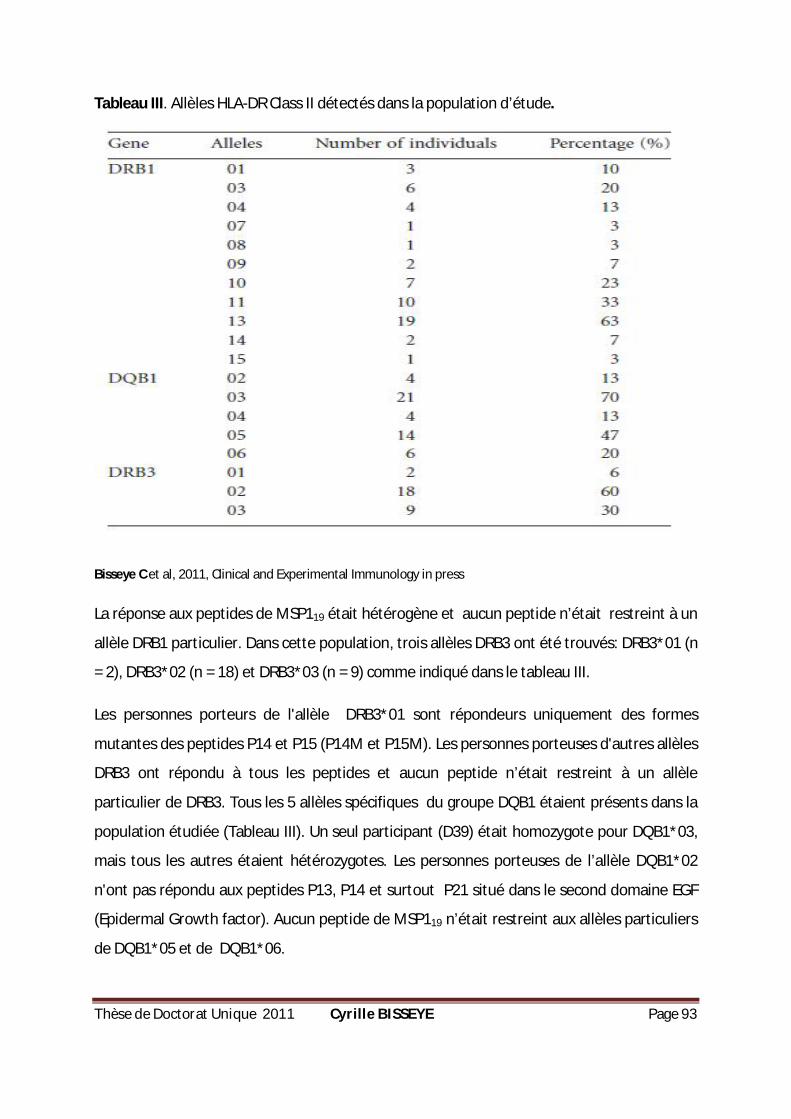
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 93
Tableau III. Allèles HLA-DR Class II détectés dans la population d’étude.
Bisseye C et al, 2011, Clinical and Experimental Immunology in press
La réponse aux peptides de MSP119 était hétérogène et aucun peptide n’était restreint à un
allèle DRB1 particulier. Dans cette population, trois allèles DRB3 ont été trouvés: DRB3*01 (n
= 2), DRB3*02 (n = 18) et DRB3*03 (n = 9) comme indiqué dans le tableau III.
Les personnes porteurs de l'allèle DRB3*01 sont répondeurs uniquement des formes
mutantes des peptides P14 et P15 (P14M et P15M). Les personnes porteuses d'autres allèles
DRB3 ont répondu à tous les peptides et aucun peptide n’était restreint à un allèle
particulier de DRB3. Tous les 5 allèles spécifiques du groupe DQB1 étaient présents dans la
population étudiée (Tableau III). Un seul participant (D39) était homozygote pour DQB1*03,
mais tous les autres étaient hétérozygotes. Les personnes porteuses de l’allèle DQB1*02
n'ont pas répondu aux peptides P13, P14 et surtout P21 situé dans le second domaine EGF
(Epidermal Growth factor). Aucun peptide de MSP119 n’était restreint aux allèles particuliers
de DQB1*05 et de DQB1*06.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 94
III.2.4. Conclusion
Nous avons montré que les adultes gambiens naturellement exposés au Plasmodium
présentent de fortes réponses cytokiniques Th1 et Th2 en réponses aux formes sauvages et
mutantes des protéines et peptides MSP119. Aucune différence n’a été trouvée dans notre
étude, en termes de nombre de répondeurs pour la production d’IFN-. Cependant, la
protéine triple mutant de MSP119 a induit une réponse cytokinique Th1 (IFN-) plus élevée
que la protéine sauvage. En outre, nous avons identifié le peptide P16 comme étant
immunodominant parce que reconnu par 63% des participants de l'étude. Aucun peptide
MSP119 n’était restreint par les allèles DRB1, DRB3 et DQB1 particuliers. Fait intéressant, les
peptides P16 et P19 de MSP119 sont responsables de réponses différentes; P16 induit une
plus forte réponse Th1 (IFN-) et P19 stimule des taux élevés d'IL-13 et sCD30. Nos résultats
pourraient être utiles dans le développement de futurs vaccins et dans les études
d’immunomodulation Th1/Th2, en utilisant soit les formes sauvages ou mutantes des
protéines ou en association avec leurs peptides.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 95
III.3. Article III. Comparaison des réponses humorales dirigées contre
CSP, AMA1 et MSP119 et la susceptibilité au paludisme chez les Peuls,
Wolofs et Mandingues de Gambie
Jamila Ismaili1*†, Cyrille Bisseye1,4‡, Dongmei liu2, Hanif Ismael1, Musa Jawara1, Mathurin
Diatta1, Giorgio Sirugo1, Margaret Pinder1, Sam dunyo1, William D. Morgan3, Michael J.
Blackman3, Anthony A. Holder3, Jacques Simporé4 and Paul Milligan1,2.
Medical Research Council Laboratories, PO Box 273, Banjul, The Gambia1; Department of Infectious and Tropical
Diseases, London School of Hygiene and Tropical Medicine, Keppel Street, London, WC1E 7HT, United Kingdom2;
Division of Parasitology, MRC National Institute for Medical Research, Mill Hill, London NW7 1AA, United
Kingdom3; and Centre de Recherche Biomoléculaire Pietro Annigoni, Ouagadougou, Burkina Faso4
Article soumis pour publication
III.3.1. But de l’étude
Les Peuls du Burkina Faso sont plus résistants au paludisme que les groupes ethniques
sympatriques Mossi et Rimaibé. Cette résistance a été associée à une réponse humorale
immunitaire plus élevée dirigée contre les antigènes de P. falciparum. Peu d’études sur la
résistance au paludisme par les Peuls ont été faites ailleurs en Afrique de l’Ouest. Par
conséquent, nous avons mené cette étude en Gambie sur un grand groupe de participants
appartenant aux groupes ethniques Peul, Wolof et Mandingue afin d’étudier la réponse
humorale interethnique, la prévalence parasitaire et la résistance au paludisme. Nous avons
étudié les réponses humorales aux protéines CSP, AMA1 et MSP119. Toutes ces protéines
sont des candidats vaccins contre le paludisme à P. falciparum en cours de développement.
Notre étude avait pour but de tester l’hypothèse de la résistance des Peuls de Gambie au
paludisme comparativement aux groupes ethniques sympatriques Wolof et Mandingue.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 96
III.3.2. Population
L’étude a été réalisée dans un groupe stratifiée de 658 enfants et adultes âgés de 6 mois à
45 ans recrutés parmi les trois groupes ethniques Peul, Wolof et Mandingue. Les participants
ont bénéficié d’un suivi clinique actif de 23 semaines à partir de la date de leur inclusion
dans l’étude.
III.3.3. Résultats
III.3.3.1. Caractéristiques des participants de l’étude
Les caractéristiques démographiques des participants de l’étude sont résumées dans le
tableau II. La cohorte a été constituée pour être représentative des trois principaux groupes
ethniques dans chaque tranche d’âge. Les Wolofs, les Peuls et les Mandingues
représentaient respectivement 31,26%, 36,39% et 32,35% des participants à l’étude. Ils ont
été classés en trois catégories d’âge < 5ans, 6-15 ans et > 15 ans avec respectivement
20.88%, 42.39% et 36.73% des sujets. Parmi les participants de l’étude, près de 60% étaient
de sexe féminin, 25% utilisaient une moustiquaire et 1% une moustiquaire imprégnée
d’insecticide. Le taux d'inoculation entomologique (EIR) était supérieur à 50 dans 18% des
sites de résidence. En ce qui concerne les facteurs génétiques de l’hôte affectant la
protection contre le paludisme, 1/5 des sujets étaient hétérozygotes HbAS et environ 1%
étaient homozygotes HbSS. La déficience en G6PD affectait respectivement environ 4% et
1% des hommes hémizygotes et des femmes hétérozygotes. L'immunité acquise à P.
falciparum a été évaluée lors de l’enrôlement dans l’étude. Les anticorps dirigés contre les
antigènes CSP, MSP119 et AMA1 ont été retrouvés respectivement chez 76,75%, 64,29% et
50% des participants à l’étude (Tableau IV).
III.3.3.2. Comparaisons interethniques de l'incidence du paludisme
Nous avons étudié les différences interethniques de l'incidence du paludisme chez les trois
principaux groupes ethniques de la Gambie, Peul (n = 234), mandingue (n = 208) et wolof (n
= 201). Le taux d'incidence du paludisme était similaire dans les trois groupes ethniques.
Cependant, les EIRs enregistrés dans les villages et les génotypes G6PD et Hb ont été
associés à un risque accru de paludisme et à l'ethnicité. En effet, les Peuls vivaient plus
souvent dans les villages où l’EIR était faible.
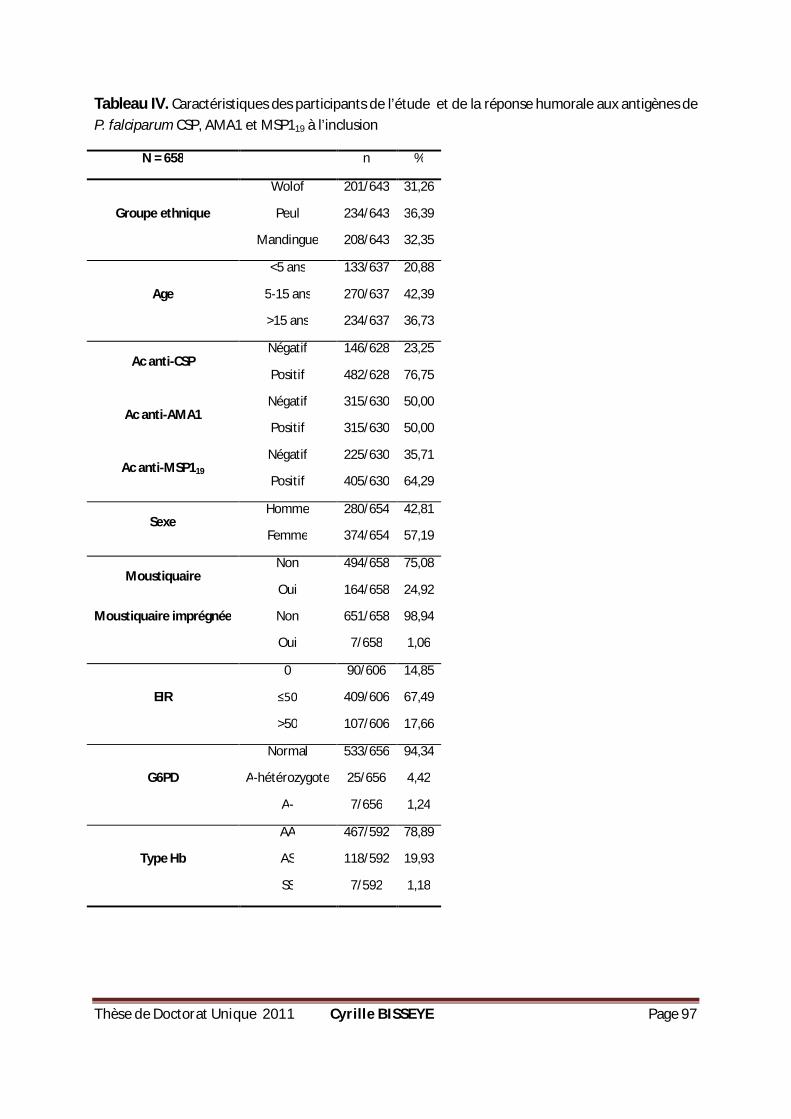
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 97
Tableau IV. Caractéristiques des participants de l’étude et de la réponse humorale aux antigènes de
P. falciparum CSP, AMA1 et MSP119 à l’inclusion
N = 658 n %
Groupe ethnique
Wolof 201/643 31,26
Peul 234/643 36,39
Mandingue 208/643 32,35
Age
<5 ans 133/637 20,88
5-15 ans 270/637 42,39
>15 ans 234/637 36,73
Ac anti-CSP Négatif 146/628 23,25
Positif 482/628 76,75
Ac anti-AMA1 Négatif 315/630 50,00
Positif 315/630 50,00
Ac anti-MSP119 Négatif 225/630 35,71
Positif 405/630 64,29
Sexe Homme 280/654 42,81
Femme 374/654 57,19
Moustiquaire Non 494/658 75,08
Oui 164/658 24,92
Moustiquaire imprégnée Non 651/658 98,94
Oui 7/658 1,06
EIR
0 90/606 14,85
≤50 409/606 67,49
>50 107/606 17,66
G6PD
Normal 533/656 94,34
A-hétérozygote 25/656 4,42
A- 7/656 1,24
Type Hb
AA 467/592 78,89
AS 118/592 19,93
SS 7/592 1,18

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 98
Après ajustement pour les effets des covariables, l’ethnicité Peule a été associée à un risque
légèrement accru de paludisme par rapport aux deux autres groupes ethniques.
Des estimations corrigées de l'incidence du paludisme, en association avec les
caractéristiques de base décrites dans le tableau V et en particulier la présence de la
parasitémie lors de l'enrôlement abolissent les différences interethniques de susceptibilité
au paludisme clinique et aucune différence significative de l'incidence du paludisme a été
trouvée entre les trois groupes ethniques [estimations brutes (4.226; ST dev: 0,325; Pr (> | z
|): 0.000) et les estimations ajustées pour tous les facteurs confondants (3,972; ST dev:
0,344; Pr (> | z |): 0)].
III.3.3.3. Comparaisons interethniques dans la réponse humorale à CSP, AMA1 et MSP119
La réponse humorale anti-CSP, MSP119 et AMA1 (IgG, IgG1 et IgG3), augmente avec l'âge.
Aucune association significative n'a été trouvée d’une part, entre la réponse humorale et
l'utilisation de moustiquaires et d’autre part, entre la réponse humorale et l'EIR, ni en
utilisant la régression logistique, ou la régression log-linéaire (non représentée).
La proportion de patients ayant répondu à AMA1 était similaire dans les 3 groupes
ethniques. Elle était cependant plus faible chez les Mandingues pour CSP et MSP119
comparativement aux 2 autres groupes ethniques (Tableau VI).
Les concentrations en IgG totales et sous-classes (IgG1 et IgG3) anti-CSP, AMA1 et MSP119
ont été comparées chez les participants des différents groupes ethniques. Les
concentrations en IgG et IgG1 anti-MSP119 étaient plus élevées chez les Peuls (figure 19)
tandis que les IgG, IgG1 et IgG3 anti- AMA1 étaient similaires dans les 3 groupes ethniques.
Comme le montre la figure 19, la concentration en IgG anti-CSP était plus faible chez les
Mandingues comparativement aux Peuls et Wolofs, qui avaient des concentrations
anticorpales similaires. Les participants des 3 groupes ethniques avaient les mêmes
concentrations en IgG1 et IgG3 anti-CSP.
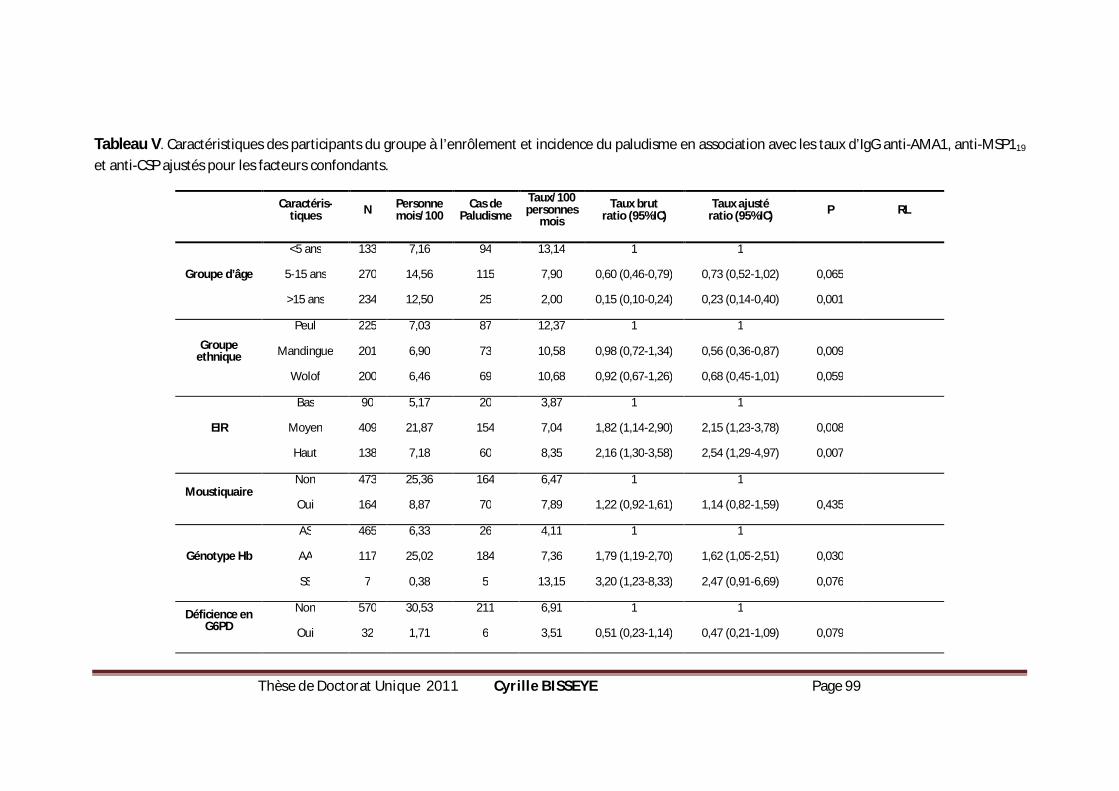
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 99
Tableau V. Caractéristiques des participants du groupe à l’enrôlement et incidence du paludisme en association avec les taux d’IgG anti-AMA1, anti-MSP119
et anti-CSP ajustés pour les facteurs confondants.
Caractéris-tiques N Personne
mois/100 Cas de
Paludisme
Taux/100 personnes
mois
Taux brut ratio (95%IC)
Taux ajusté ratio (95%IC) P RL
Groupe d’âge
<5 ans 133 7,16 94 13,14 1 1
5-15 ans 270 14,56 115 7,90 0,60 (0,46-0,79) 0,73 (0,52-1,02) 0,065
>15 ans 234 12,50 25 2,00 0,15 (0,10-0,24) 0,23 (0,14-0,40) 0,001
Groupe ethnique
Peul 225 7,03 87 12,37 1 1
Mandingue 201 6,90 73 10,58 0,98 (0,72-1,34) 0,56 (0,36-0,87) 0,009
Wolof 200 6,46 69 10,68 0,92 (0,67-1,26) 0,68 (0,45-1,01) 0,059
EIR
Bas 90 5,17 20 3,87 1 1
Moyen 409 21,87 154 7,04 1,82 (1,14-2,90) 2,15 (1,23-3,78) 0,008
Haut 138 7,18 60 8,35 2,16 (1,30-3,58) 2,54 (1,29-4,97) 0,007
Moustiquaire Non 473 25,36 164 6,47 1 1
Oui 164 8,87 70 7,89 1,22 (0,92-1,61) 1,14 (0,82-1,59) 0,435
Génotype Hb
AS 465 6,33 26 4,11 1 1
AA 117 25,02 184 7,36 1,79 (1,19-2,70) 1,62 (1,05-2,51) 0,030
SS 7 0,38 5 13,15 3,20 (1,23-8,33) 2,47 (0,91-6,69) 0,076
Déficience en G6PD
Non 570 30,53 211 6,91 1 1
Oui 32 1,71 6 3,51 0,51 (0,23-1,14) 0,47 (0,21-1,09) 0,079
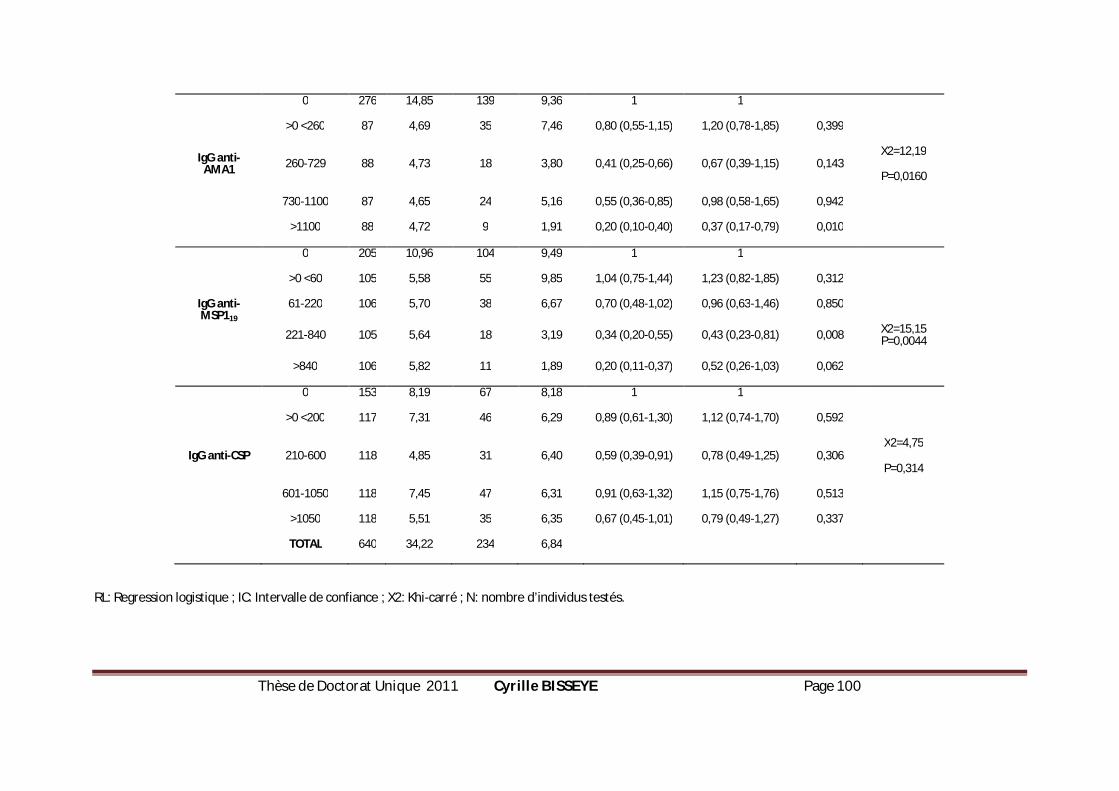
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 100
IgG anti-AMA1
0 276 14,85 139 9,36 1 1
>0 <260 87 4,69 35 7,46 0,80 (0,55-1,15) 1,20 (0,78-1,85) 0,399
260-729 88 4,73 18 3,80 0,41 (0,25-0,66) 0,67 (0,39-1,15) 0,143 X2=12,19
P=0,0160
730-1100 87 4,65 24 5,16 0,55 (0,36-0,85) 0,98 (0,58-1,65) 0,942
>1100 88 4,72 9 1,91 0,20 (0,10-0,40) 0,37 (0,17-0,79) 0,010
IgG anti-MSP119
0 205 10,96 104 9,49 1 1
>0 <60 105 5,58 55 9,85 1,04 (0,75-1,44) 1,23 (0,82-1,85) 0,312
61-220 106 5,70 38 6,67 0,70 (0,48-1,02) 0,96 (0,63-1,46) 0,850
221-840 105 5,64 18 3,19 0,34 (0,20-0,55) 0,43 (0,23-0,81) 0,008 X2=15,15 P=0,0044
>840 106 5,82 11 1,89 0,20 (0,11-0,37) 0,52 (0,26-1,03) 0,062
IgG anti-CSP
0 153 8,19 67 8,18 1 1
>0 <200 117 7,31 46 6,29 0,89 (0,61-1,30) 1,12 (0,74-1,70) 0,592
210-600 118 4,85 31 6,40 0,59 (0,39-0,91) 0,78 (0,49-1,25) 0,306 X2=4,75
P=0,314
601-1050 118 7,45 47 6,31 0,91 (0,63-1,32) 1,15 (0,75-1,76) 0,513
>1050 118 5,51 35 6,35 0,67 (0,45-1,01) 0,79 (0,49-1,27) 0,337
TOTAL 640 34,22 234 6,84
RL: Regression logistique ; IC: Intervalle de confiance ; X2: Khi-carré ; N: nombre d’individus testés.
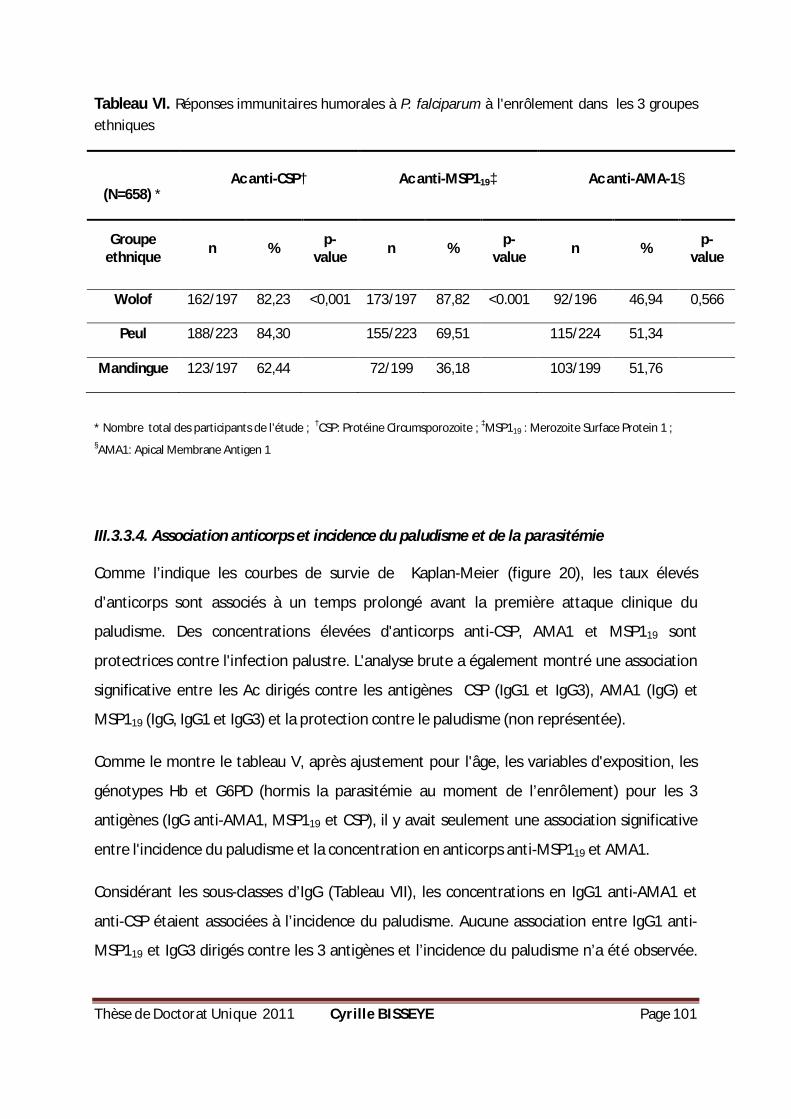
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 101
Tableau VI. Réponses immunitaires humorales à P. falciparum à l'enrôlement dans les 3 groupes
ethniques
(N=658) * Ac anti-CSP† Ac anti-MSP119‡ Ac anti-AMA-1§
Groupe ethnique
n % p-
value n %
p-value
n % p-
value
Wolof 162/197 82,23 <0,001 173/197 87,82 <0.001 92/196 46,94 0,566
Peul 188/223 84,30 155/223 69,51 115/224 51,34
Mandingue 123/197 62,44 72/199 36,18 103/199 51,76
* Nombre total des participants de l’étude ; †CSP: Protéine Circumsporozoite ; ‡MSP119 : Merozoite Surface Protein 1 ; §AMA1: Apical Membrane Antigen 1
III.3.3.4. Association anticorps et incidence du paludisme et de la parasitémie
Comme l’indique les courbes de survie de Kaplan-Meier (figure 20), les taux élevés
d’anticorps sont associés à un temps prolongé avant la première attaque clinique du
paludisme. Des concentrations élevées d'anticorps anti-CSP, AMA1 et MSP119 sont
protectrices contre l'infection palustre. L'analyse brute a également montré une association
significative entre les Ac dirigés contre les antigènes CSP (IgG1 et IgG3), AMA1 (IgG) et
MSP119 (IgG, IgG1 et IgG3) et la protection contre le paludisme (non représentée).
Comme le montre le tableau V, après ajustement pour l'âge, les variables d'exposition, les
génotypes Hb et G6PD (hormis la parasitémie au moment de l’enrôlement) pour les 3
antigènes (IgG anti-AMA1, MSP119 et CSP), il y avait seulement une association significative
entre l'incidence du paludisme et la concentration en anticorps anti-MSP119 et AMA1.
Considérant les sous-classes d’IgG (Tableau VII), les concentrations en IgG1 anti-AMA1 et
anti-CSP étaient associées à l’incidence du paludisme. Aucune association entre IgG1 anti-
MSP119 et IgG3 dirigés contre les 3 antigènes et l’incidence du paludisme n’a été observée.
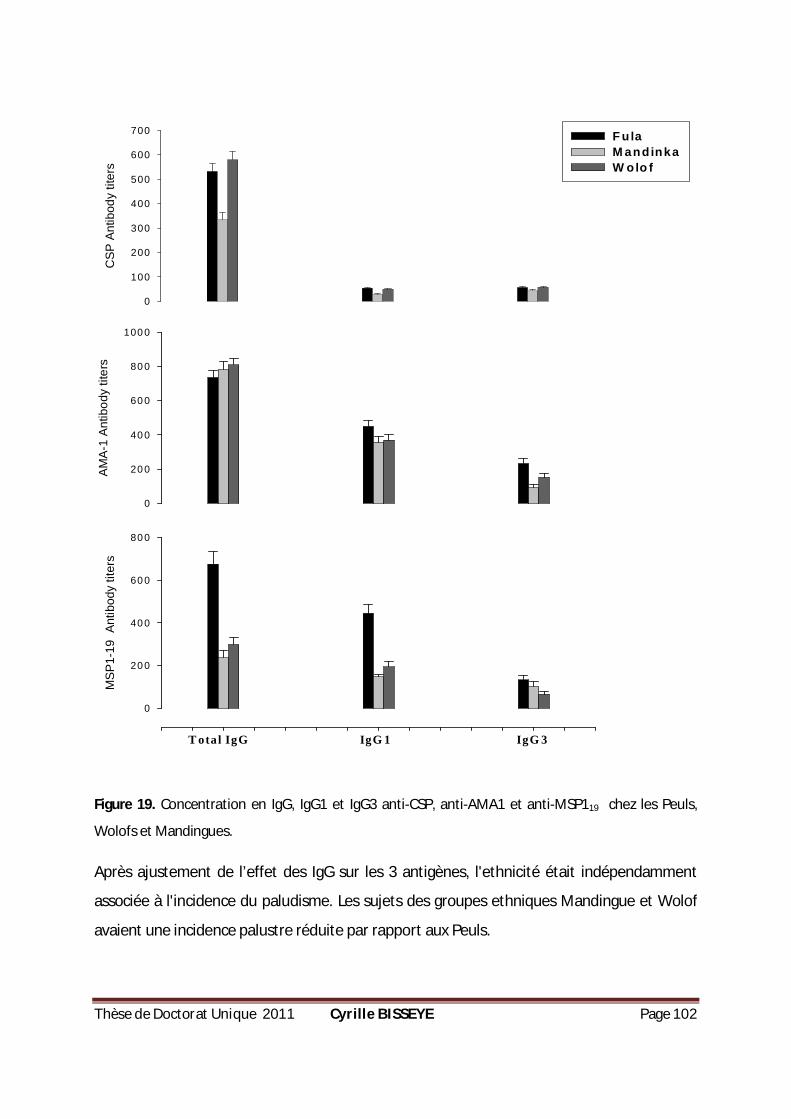
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 102
CSP
Ant
ibod
y tit
ers
0
100
200
300
400
500
600
700
T otal IgG IgG 1 IgG 3
MSP
1-19
Ant
ibod
y tit
ers
0
20 0
40 0
60 0
80 0
F ulaM andinkaW olo f
AMA-
1 An
tibod
y tit
ers
0
20 0
40 0
60 0
80 0
100 0
Figure 19. Concentration en IgG, IgG1 et IgG3 anti-CSP, anti-AMA1 et anti-MSP119 chez les Peuls,
Wolofs et Mandingues.
Après ajustement de l’effet des IgG sur les 3 antigènes, l'ethnicité était indépendamment
associée à l'incidence du paludisme. Les sujets des groupes ethniques Mandingue et Wolof
avaient une incidence palustre réduite par rapport aux Peuls.
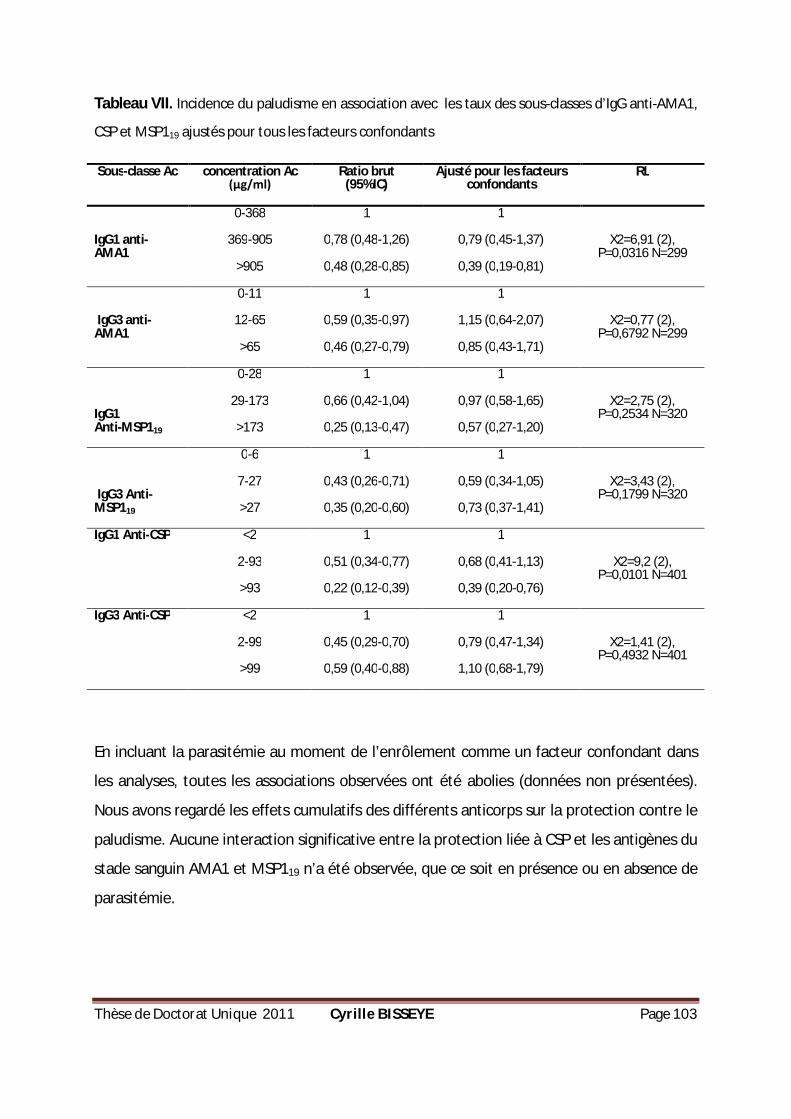
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 103
Tableau VII. Incidence du paludisme en association avec les taux des sous-classes d’IgG anti-AMA1,
CSP et MSP119 ajustés pour tous les facteurs confondants
Sous-classe Ac concentration Ac (µg/ml)
Ratio brut (95%IC)
Ajusté pour les facteurs confondants
RL
IgG1 anti-AMA1
0-368
1
1
X2=6,91 (2), P=0,0316 N=299
369-905
0,78 (0,48-1,26)
0,79 (0,45-1,37)
>905
0,48 (0,28-0,85)
0,39 (0,19-0,81)
IgG3 anti-AMA1
0-11
1 1
X2=0,77 (2), P=0,6792 N=299
12-65
0,59 (0,35-0,97)
1,15 (0,64-2,07)
>65
0,46 (0,27-0,79)
0,85 (0,43-1,71)
IgG1 Anti-MSP119
0-28
1 1
X2=2,75 (2), P=0,2534 N=320
29-173
0,66 (0,42-1,04)
0,97 (0,58-1,65)
>173
0,25 (0,13-0,47)
0,57 (0,27-1,20)
IgG3 Anti-MSP119
0-6
1 1
X2=3,43 (2), P=0,1799 N=320
7-27
0,43 (0,26-0,71)
0,59 (0,34-1,05)
>27
0,35 (0,20-0,60)
0,73 (0,37-1,41)
IgG1 Anti-CSP <2
1 1
X2=9,2 (2), P=0,0101 N=401
2-93
0,51 (0,34-0,77)
0,68 (0,41-1,13)
>93
0,22 (0,12-0,39)
0,39 (0,20-0,76)
IgG3 Anti-CSP <2
1 1
X2=1,41 (2), P=0,4932 N=401
2-99
0,45 (0,29-0,70)
0,79 (0,47-1,34)
>99
0,59 (0,40-0,88)
1,10 (0,68-1,79)
En incluant la parasitémie au moment de l’enrôlement comme un facteur confondant dans
les analyses, toutes les associations observées ont été abolies (données non présentées).
Nous avons regardé les effets cumulatifs des différents anticorps sur la protection contre le
paludisme. Aucune interaction significative entre la protection liée à CSP et les antigènes du
stade sanguin AMA1 et MSP119 n’a été observée, que ce soit en présence ou en absence de
parasitémie.
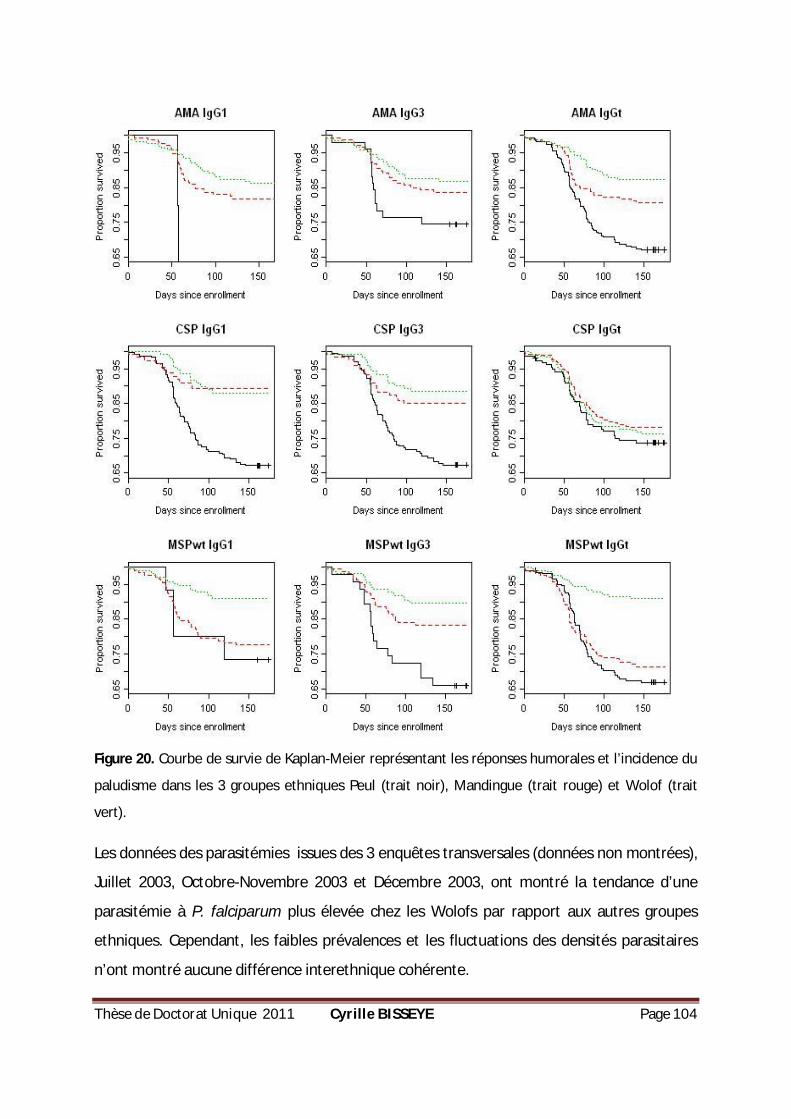
Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 104
Figure 20. Courbe de survie de Kaplan-Meier représentant les réponses humorales et l’incidence du
paludisme dans les 3 groupes ethniques Peul (trait noir), Mandingue (trait rouge) et Wolof (trait
vert).
Les données des parasitémies issues des 3 enquêtes transversales (données non montrées),
Juillet 2003, Octobre-Novembre 2003 et Décembre 2003, ont montré la tendance d’une
parasitémie à P. falciparum plus élevée chez les Wolofs par rapport aux autres groupes
ethniques. Cependant, les faibles prévalences et les fluctuations des densités parasitaires
n’ont montré aucune différence interethnique cohérente.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 105
III.3.4. Conclusion
Nous rapportons ici la première étude sur les différences interethniques sur l’incidence du
paludisme et les réponses humorales chez lez Peuls, les Wolofs et les Mandingues de
Gambie.
Nous avons prouvé que la réponse humorale dirigée contre les antigènes de P. falciparum
est plus élevée chez les Peuls de Gambie comparativement aux Wolofs et aux Mandingues;
cependant l’incidence du paludisme et la prévalence et la densité parasitaires étaient les
mêmes dans les trois (3) groupes ethniques.
Contrairement aux études antérieures, l’inclusion de la parasitémie comme facteur
confondant a aboli l’association entre la réponse humorale aux antigènes AMA1, CSP et
MSP119 et la protection contre le paludisme clinique.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 106
IV. Discussion générale
La présente thèse avait trois principaux objectifs spécifiques à savoir :
L’étude du rôle des cellules T régulatrices dans le biais Th1/Th2 des cellules T
néonatales
L’évaluation de l’antigénicité des peptides et des protéines du candidat vaccin
MSP119.
La comparaison des réponses humorales dirigées contre les antigènes candidats
vaccins MSP119, AMA1 et CSP chez des sujets appartenant aux principaux groupes
ethniques de Gambie : Peul, Wolof et Mandinka.
Cette thèse a permis de préciser un rôle possible joué par les Tregs dans le biais Th1/Th2 des
cellules T néonatales dans l’infection placentaire par P. falciparum.
Rôle des cellules T régulatrices dans le biais Th1/Th2 des cellules T néonatales
De nombreuses études avaient déjà montré que l’infection placentaire par P.
falciparum affecte les réponses immunitaires du nouveau-né en agissant sur la
différentiation Th1 et Th2 des lymphocytes T néonatales (Ismaili et al, 2003; Malhotra et al,
2008).
Dans la présente thèse, les échantillons des femmes gambiennes primipares et multipares
utilisés provenaient d’une étude ayant montré que l’infection active du placenta par P.
falciparum entraînait une diminution de la production d’IFN- et d’IL-12 par les cellules T
néonatales (Ismaili et al, 2003). L’étude d’Ismaili et coll., ainsi que celles montrant
l’altération de la fonction de l’IL-12 chez les nouveau-nés (Goriely et al, 2001) et l'effet
inhibiteur de P. falciparum sur la maturation des cellules dendritiques (CD) (Urban et al,
1999), pourraient attribuer la réduction de la production de l’IFN- à une inhibition des
cellules présentatrices d'antigène (CPA) dans les cultures des sujets parasités.
Des études précédentes ont montré que les Tregs pouvaient être impliquées dans le
maintien de l’infection palustre à travers la production élevée d’IL-10 et de TGF-β (Walter et

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 107
al, 2005, 2006). Ces cytokines connues pour être communes aux Tregs et aux CPA, sont des
molécules immunomodulatrices et pourraient être impliquées dans l’inhibition de la
production de cytokines Th1 par les CMSCs dans le groupe de placenta parasité.
Nous avons donc évalué le rôle possible des Tregs dans le déséquilibre Th1/Th2 observé. En
Utilisation le même groupe de femmes qu’Ismaili et coll. (2003), nous avons comparé les
fréquences des cellules T CD4+CD25+Foxp3+ et les cultures de CMSCs d’échantillons
sélectionnés avant et après déplétion des cellules T CD4+CD25+FoxP3low en présence de
PHA, d'extraits de schizontes Pf164, MSP119, parasites vivants et nous avons testé leur
capacité à sécréter l'IFN-, le sCD30 et l'IL-10.
Nos résultats confirment les observations précédentes d’Ismaili et al (2003) que l’infection
active du placenta par P. falciparum exacerbe le biais Th2 des lymphocytes T néonatales,
comme le témoigne la baisse de sécrétion d’IFN- en réponse à tous les antigènes, et
l’augmentation de la production de sCD30 en réponse aux antigènes protéiques et au
parasite vivant par les cellules du groupe parasité.
Nous avons trouvé une production d'IL-10 plus élevée en réponse à tous les antigènes ainsi
que l'augmentation de l'IL-10 par la population de cellules T CD4+CD25+ en réponse à la PHA,
ce qui confirme leur phénotype de cellules T régulatrices.
La production de l’IL-10 pouvant être assurée aussi bien par les CPA que par les Tregs
CD4+CD25+Foxp3+ (Walther et al, 2005; Hisaeda et al, 2004), le rôle de chaque population
de cellules a été établi en délestant les CMSCs en cellules T CD4+CD25+ et leur production en
cytokines Th1/Th2 réévaluée in vitro. Nous avons montré que la déplétion des Tregs conduit
à la baisse de la production de l'IL-10 et à la restauration de la production d'IFN- en réponse
à tous les stimuli testés.
La production des cytokines a été aussi évaluée en stimulant les cellules T néonales par le
parasite vivant (éthrocytes infectés) et par les antigènes plasmodiaux (extrait de schizonte et
antigène MSP119). Nous avons observé une production plus élevée de cytokines induites par
les parasites vivants comparativement à celle provoquée par les antigènes protéiques ainsi
que d’une différentiation Th2 antigène-dépendant suggérant un mécanisme complexe des
CPA avec tous ces différents stimuli.
Dans la présente thèse, contrairement à l’étude de Brustoski et coll. (2006), nous avons
montré que la fréquence des Tregs est la même dans les CMSCs des placentas parasités et

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 108
non parasités, et que la déplétion de cette population influe sur les cytokines Th1/Th2 in
vitro. Cette différence entre les fréquences des Tregs pourrait être attribuée à des
différences au niveau des taux de parasitémie placentaire, du type d’infection placentaire,
active et/ou chronique entre les deux groupes de femmes (Brustoski et al, 2006) et que les
effets post-déplétions observés dans notre groupe de femmes pourraient être le fait d’un
état différent d’activation des cellules T CD4+CD25+, d’un effet synergique avec les CPA ou
d’une interaction avec d’autres populations de lymphocytes T.
Dans un modèle murin d’infection, il a été montré que les Tregs maternelles traversent le
placenta (Mold et al, 2008). Cependant, nos expériences de déplétion des Tregs n’ont pas eu
d’incidence sur les réactions allo leucocytaires mixtes (allo-MLR) dirigés contre les
lymphocytes T des mères tel que rapporté par Mold et coll., suggérant que les antigènes de
P. falciparum et les cytokines inflammatoires placentaires amorcent principalement les
Tregs des CMSCs.
Nous avons ensuite évalué le rôle de l'IL-10 sur la sécrétion des cytokines ou des marqueurs
Th1 et Th2, en privilégiant les parasites vivants par rapport aux antigènes protéiques, car ils
ont induit des taux plus élevés de cytokines et une différence de production significative en
présence ou en absence des Tregs.
Le blocage de l'IL-10 par un anticorps monoclonal anti-IL-10 produit le même effet que la
déplétion des Tregs, notamment, la restauration de la production d’IFN-. Ce résultat
suggère que les cellules T CD4+CD25+Foxp3+ du groupe parasité, pourrait par leur forte
production d’IL-10 avoir réduit la production d’IL-12 (Peng et al, 2006) par les CPA qui, à son
tour, diminue la production de l’IFN- et augmente celle du sCD30 par les cellules T.
Dans la présente thèse, le rôle de la réponse immune cellulaire dans l’infection palustre par
P. falciparum a été aussi évalué en étudiant la réponse cellulaire anti-MSP119 (peptides et
protéines) chez les adultes gambiens naturellement exposés au paludisme.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 109
Immunité cellulaire induite contre les peptides et protéines du candidat
vaccin MSP119 chez les adultes gambiens
Nous avons évalué l’antigenicité de la protéine MSP119 triple mutant qui est un
candidat vaccin synthétisé pour améliorer l’efficacité et la spécificité de la réponse
immunitaire humorale induite chez l’hôte. Toutefois, sa capacité à induire une réponse
cellulaire T CD4+ par rapport à la protéine de type sauvage était inconnue.
En effet, Il est possible que la substitution d'acides aminés dans une protéine affecte
l’apprêtement et la présentation des peptides par les CPA, elle peut aussi créer ou détruire
des épitopes de lymphocytes T; ces épitopes reconnus par des individus naturellement
exposés au parasite sont particulièrement intéressants. Les Modifications de MSP119 qui
améliorent à la fois la qualité des réponses cellulaire et humorale de la protéine pourraient
être bénéfiques.
Nous avons donc étudié les réponses immunitaires cellulaires dirigées contre la protéine
MSP119 triple mutant comparée à la protéine de type sauvage et identifié les peptides
immunodominants chez les individus naturellement exposés au paludisme à P. falciparum.
Comme il a été rapporté que l'IFN- est la cytokine dominante induite en réponse à
l'antigène MSP119 aussi bien chez les adultes naturellement exposés au Plasmodium que
chez les volontaires vaccinés (Lee et al, 2002); nous avons testé en premier par la méthode
ELISPOT, la production d’IFN- par les PBMCs d’adultes gambiens en réponse à la stimulation
par les formes sauvage et triple mutant de la protéine MSP119.
Nous avons constaté qu'environ un tiers d’adultes gambiens avait répondu aux deux types
de protéines. Nos résultats sont similaires à ceux obtenus dans deux études antérieures
réalisées en Gambie et au Kenya où la prolifération des cellules T en réponse à des peptides
chevauchants de MSP119 avait été mesurée (Egan et al, 1997; Udhayakumar et al, 1995).
Un nombre égal de cellules T sécrétrices d'IFN- a aussi été obtenu en réponse aux deux
types de protéines MSP119 chez les adultes naturellement exposés au Plasmodium.
Nous avons également évalué la production d'IFN- par les PBMCs d’adultes Gambiens par
ELISPOT en réponse aux peptides sauvages ou mutants chevauchants de MSP119. Nos

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 110
résultats ont permis de mettre en évidence que le nombre de répondeurs aux peptites
(mutants et sauvages) variait de 26% (pour le peptide P13) à 63% (pour le peptide P16).
La séquence de MSP119 de P. falciparum est très largement conservée, avec une diversité
portant seulement sur quatre acides aminés sur les 96 (Miller et al, 1993) qui précèdent
l’ancrage GPI C-terminal (Gerold et al, 1996). Dans le premier domaine EGF, le résidu 14 est
soit une Gln ou une Glu et dans le second domaine EGF, les résidus 61, 70 et 71 sont soient
des Lys, Asn, Gly ou des Thr, Ser, Arg, respectivement. À partir du variant Q-KNG un panel de
protéines modifiées a été conçu dans le but d'améliorer l'immunogénicité de MSP119. Le
premier aspect du changement d’un ou plusieurs résidus d’acides aminés de la protéine a
été d’examiner les conséquences sur la liaison d’anticorps inhibiteurs ou bloquants. Le
deuxième aspect envisageable est l’amélioration de l’apprêtement et de la présentation des
peptides en supprimant un ou plusieurs ponts disulfure ou en introduisant des sites de
clivage de la protéase. Cependant de telles modifications peuvent modifier ou supprimer les
épitopes des lymphocytes T. Dans la présente thèse le deuxième aspect concernant
l’amélioration de l’apprêtement et de la présentation des protéines et peptides ont été
évaluées. Nous montrons qu’aucune différence significative n’a été trouvée dans les
réponses aux protéines sauvage ou triple mutant et aux peptides sauvage ou mutant de
MSP119.
Le nombre de cellules T sécrétrices d'IFN- était également similaire après stimulation soit
par les protéines ou les peptides de type sauvage ou mutant de MSP119. Nous avons
démontré ainsi, que les épitopes antigéniques linéaires de MSP119 des peptides et des
protéines n'ont pas été modifiés par les substitutions d’acides aminés choisis.
Dans une précédente étude, une plus forte prolifération des lymphocytes T a été induite
en réponse aux protéines modifiées de MSP119 comparativement à la protéine sauvage
chez les individus naturellement exposés au Plasmodium (Okafor et al, 2009). Ce résultat
diffère du nôtre et pourrait s'expliquer par quelques différences entre les deux études :
notamment par le type de sujets inclus (asymptomatique contre symptomatique), la nature
des protéines MSP119 choisies et le type d’essai utilisé pour mesurer la prolifération
cellulaire T.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 111
Nous avons également mesuré la production des cytokines de type Th1 (IFN-) et Th0/Th2
(IL-13 et sCD30) par les lymphocytes T stimulées par les formes sauvages et modifiées des
protéines et peptides de MSP119.
Différents profiles de réponses cytokiniques ont été observés en utilisant les protéines et les
peptides. Une plus forte production d'IFN- a été obtenue avec la protéine triple mutant
comparativement au type sauvage de MSP119. Une production plus élevée d'IFN- pourrait
augmenter la résistance à la réinfection par Plasmodium (Luty et al, 1999).
En revanche, les protéines de type sauvage et triple mutant ont montré les mêmes
tendances pour la production d’IL-13 et de sCD30.
En ce qui concerne les réponses aux stimulations par les peptides de MSP119, aucune
différence n'a été observée pour la production d’IFN- et de sCD30 par les peptides mutants
par rapport aux peptides de type sauvage.
Cependant, il a été intéressant de constater que les individus naturellement exposés au
Plasmodium produisaient plus d'IL-13 en réponse aux peptides mutants par rapport aux
peptides de type sauvage. Les peptides mutants suscitant une réponse de type Th0 à travers
l'IL-13 pourrait activer plus efficacement la production d'anticorps (Hirunpetcharat et al,
1999).
Nos résultats ont montré également que deux peptides conservés de MSP119 (P16 et P19)
induisaient différents modèles de réponses cytokiniques. Le peptide conservé P16 et sa
forme modifiée P16M ont stimulé la plus forte production d'IFN- parmi les peptides de
MSP119, tandis que la plus grande induction d’IL-13 a été obtenue en réponse au peptide
P19.
Ces différences pourraient être utiles pour les expérimentations immunologiques dans
l’optique de la vaccination. Il est possible que la substitution d'acides aminés modifie la
cinétique de l’apprêtement et de la présentation des peptides MSP119 associés aux
molécules HLA de classe II par les CPA. Les molécules HLA de classe II sont importantes dans
la protection ou la sensibilité à l'infection palustre (Beck et al, 1995; May et al, 2001; Osafo-
Addo et al, 2008). Les molécules HLA de classe II peuvent affecter le taux d’anticorps dirigé

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 112
contre le Plasmodium. Dans une précédente étude, il a été montré que les personnes
porteuses d’allèles HLA DRB1*1201 avaient des taux plus élevés d’anticorps dirigés contre
un variant recombinant d’AMA1 que les individus porteurs des autres types d’allèles HLA
(John et al, 2004). Nous avons étudié la prévalence des locus HLA-DRB1, HLA-DRB3, HLA-
DR4, HLA-DR5 et HLA-DQB1 dans notre population d'étude. Aucun des sujets inclus dans
notre étude n’était porteur des allèles HLA-DR4 et HLA-DR5. L’allèle DRB1*13 était le plus
représenté avec 63% des individus porteurs de ce dernier. L’allèle HLA-DRB1*04 associé à
l'anémie palustre (Osafo-Addo et al, 2008) a été retrouvé chez 13% d’individus.
Les réponses des individus aux peptides MSP119 n’étaient pas restreintes par les allèles HLA
DRB1, DRB3 et DQB1 particuliers.
L’infection par P. falciparum induit une forte réponse humorale. Différents antigènes du
parasite sont des candidats vaccins à diffirents stades d’essais cliniques. Nous avons évalué
Les réponses humorales de trois antigènes candidats vaccins (AMA1, CSP et MSP119) dans
les principaux groupes ethniques de Gambie : Peul, Wolof et Mandingue.
Comparaison interethnique des réponses immunitaires humorales dirigées
contre les antigènes candidats vaccins AMA1, CSP et MSP119 dans les
principaux ethniques de Gambie
Le but de cette étude étude était de tester l’hypothèse de la différence interethnique des
réponses humorales dirigées contre les antigènes de P. falciparum candidats vaccins (AMA1,
CSP et MSP119) chez les Peuls de Gambie et les groupes ethniques Wolof et Mandingue.
En effet, une précédente étude avait montré que les Peuls du Burkina Faso avaient de plus
fortes réponses humorales contre le Plasmodium tout en étant moins fréquemment
parasitées et moins sensibles au paludisme clinique (Modiano et al, 1996) par rapport aux
groupes ethniques sympatriques Mossi et Rimaibé.
Les différentes études immunologiques menées sur différents sites, ont donné des résultats
contradictoires sur l'association entre les réponses humorales anti-AMA1, anti-CSP et anti-
MSP119 et l’immunité clinique (Egan et al, 1996; Polley et al, 2004; Dodoo et al, 1999).

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 113
La présente thèse a donc réevalué l'association des réponses anticorpales contre les
antigènes AMA1, CSP et MSP119 et l’immunité clinique en intégrant les facteurs génétiques
et la parasitémie comme facteurs confondants dans les différentes analyses.
Nous avons montré que l’incidence du paludisme était plus élevée chez les Peuls de Gambie
comparativement aux groupes ethniques Mandingue et Wolof. Toutefois, après ajustement
par tous les facteurs confondants, aucune différence significative n’était plus observée entre
les 3 groupes ethniques.
La comparaison du risque de paludisme entre les groupes ethniques est souvent compliquée
du fait de l’existence de nombreux facteurs confondants. Il est important de tenir compte
des différences dans l'EIR et d'autres facteurs associés à l'exposition comme le type
d’habitat. Nous avons trouvé une différence géographique dans l’exposition au
Plasmodium; les Peuls vivant généralement dans les villages à faible EIR.
La concentration moyenne d'IgG anti-MSP119 qui présentait la plus forte association avec
l’incidence du paludisme était la plus élevée chez les Peuls. Après ajustement pour les IgG
totales et des facteurs environnementaux et génétiques associés au risque de paludisme, les
groupes ethniques Mandingue et wolof ont montré une plus faible incidence contre le
paludisme. Ce résultat obtenu pourrait refléter des différences génétiques (Koch et al, 2002),
ou l’effet résiduel des facteurs confondants. En effet, nous n’avons pas observé que les
sujets vivant dans les zones où l’EIR est élevé avaient de plus fortes concentrations d‘IgG
contre l’un des 3 antigènes, ou étaient de meilleurs répondeurs aux antigènes. La réponse
était similaire dans les 3 groupes ethniques ; cependant, après ajustement pour ces facteurs,
les Peuls avaient une incidence élevée et une plus forte réponse immunitaire. Les réponses
immunitaires pourraient refléter en partie une plus grande exposition au parasite.
Une précédente comparaison interethnique faite au Burkina Faso (Modiano et al, 1996) a
montré que les réponses humorales étaient constamment plus élevées chez les Peuls que
dans les groupes ethniques Mossi et Rimaibé. Dans notre étude, l'analyse brute de la
réponse humorale a révélé des taux plus élevés d'anticorps chez les Peuls comparativement
aux 2 autres groupes ethniques, ce qui est en accord avec les conclusions de l’étude
conduite au Burkina Faso.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 114
En effet, les concentrations d’IgG anti-CSP étaient plus faibles chez les Mandingues
comparativement aux Peuls et aux Wolofs qui avaient des concentrations équivalentes.
Toutefois, des taux similaires d’IgG1 et d’IgG3 anti-CSP ont été trouvés dans les 3 groupes
ethniques. Les taux d’IgG et IgG1 étaient similaires dans les 3 groupes ethniques ; alors que
les taux d’IgG3 étaient plus élevés chez les Peuls, moyens chez les Wolofs et faibles chez les
Mandingues. Ceci est en accord avec l’étude de Bolad et coll. (2005) qui avaient montré des
taux élevés d’IgG3 chez les Peuls du Burkina Faso et du Mali comparativement aux Mossis et
Dogons.
En revanche, les taux d’IgG, d’IgG1 et d’IgG3 anti-MSP119 étaient plus élevés chez les Peuls
par rapport aux groupes ethniques Mandingue et Wolof, confortant ainsi les conclusions de
Modiano et coll. (1996). En effet, dans leur étude une plus grande résistance des Peuls à P.
falciparum a été associée à leur forte réactivité humorale contre les antigènes du parasite
tels que: TRAP, MSP-1, Pf155 (RESA) et Pf332.
Nous avons examiné la réponse humorale dirigée contre les antigènes CSP, MSP119 et AMA1
et leur association avec la protection contre le paludisme clinique, en incluant les traits
génétiques de l’hôte comme un nouveau facteur confondant.
L’analyse brute des données a révélé une association entre les IgG dirigées contre les 3
antigènes (CSP, AMA1 et MSP119), les IgG1 et IgG3 (CSP et MSP119) et la protection contre le
paludisme. L’ajustement des résultats avec toutes les covariables, hormis la parasitémie lors
de l’enrôlement réduit cette association pour l’antigène CSP.
Aucune interaction significative ou protection supplémentaire n’a été observée chez les
sujets ayant une double ou une triple réponse aux Ig ou sous-classes dirigées contre les
antigènes CSP, AMA1 et MSP119. La parasitémie au moment du recrutement a été associée
soit à des taux réduits d’anticorps soit à l’incidence du paludisme; mais aucune association
entre les anticorps anti-CSP, AMA1 et MSP119 avec la protection et la parasitémie n’a été
trouvée à l’inclusion.
En incluant la parasitémie à l’enrôlement dans notre étude comme facteur confondant, elle
a aboli toutes les autres associations observées. Ceci est en divergence avec les résultats

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 115
d’une étude réalisée au Kenya, où la présence d'anticorps anti-AMA1 avait été associée à la
protection de sujets porteurs de P. falciparum au moment du recrutement (Polley et al,
2004).
Nous avons également analysé la prévalence et la densité parasitaire d’échantillons issus des
trois enquêtes transversales : au début de la saison de transmission (Juillet), la période de
haute transmission (Octobre-Novembre) et à la fin de la période de transmission
(Décembre). La prévalence d’une parasitémie à P. falciparum légèrement plus élevée a été
observée chez les Wolofs. Toutefois, nous n’avons pas pu établir de différence interethnique
claire du fait de la faible parasitémie observée dans notre groupe. Des études antérieures
avaient également signalé une faible prévalence parasitaire dans les échantillons
majoritairement constitués de Peuls du nord du Nigeria (Oomen et al, 1979) et de Gambie
(Riley et al, 1992b).
Nos résultats diffèrent de ceux rapportés antérieurement au Burkina Faso. Cette différence
peut s’expliquer d’une part, par le fait que les Peuls du Burkina Faso et de Gambie sont
génétiquement très éloignés et d’autre part, par une grande distance génétique entre les
Peuls du Burkina Faso et les groupes ethniques sympatriques Mossi et Rimaibé alors qu’il
existe une faible distance génétique entre les Peuls de Gambie et les groupes ethniques
sympatriques Mandingue et Wolof (Modiano et al, 2001). En effet, les études du
polymorphisme de GM dans l'Est du Sénégal (Allsopp et al, 1992) et des molécules HLA de
classe I (Blanc et al, 1990) et de classe II (Riley et al, 1992a) en Gambie indiquent plutôt de
faibles distances génétiques entre les Peuls et leurs voisins; au contraire l'analyse
préliminaire des individus du Burkina Faso suggère un degré relativement élevé de
divergence génétique. La réponse anticorpale élevée observée chez les Peuls peut être
attribuée à des facteurs codés par des gènes situés en dehors de la région HLA de classe II et
impliqués dans la régulation des réponses immunitaires humorales dirigées contre les
antigènes de P. falciparum (Sjoberg et al, 1992).
Dans notre étude, de fortes réponses humorales ont également été observées chez les Peuls
de Gambie, même si elles n’étaient pas corrélées avec la protection contre le paludisme.
Ainsi, d’autres mécanismes immunologiques impliquant des gènes liés à la fonction des

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 116
cellules B, l’apprêtement de l’antigène (Farouk et al, 2005 ; Arama et al, 2011), la production
d'IL-4 (Luoni et al, 2001), ou la différenciation des Tregs (Torcia et al, 2008) pourraient
expliquer les différences interethniques et être partagés par les Peuls d’Afrique occidentale.
En conséquence, les études interethniques en cours sur les polymorphismes des cytokines
Th1/Th2 pourraient apporter des réponses sur les gènes impliqués dans la régulation des
réponses immunitaires humorales (G. Sirugo, en préparation), communs aux Peuls d’Afrique
de l'Ouest.
Il est intéressant d’observer notamment que la parasitémie à l'inclusion en tant que facteur
de confusion abolit l'association des réponses humorales à tous les candidats vaccins,
MSP119, CSP et AMA1, ce qui est en contradiction avec une étude antérieure.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 117

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 118
V. Conclusion générale
L’objectif principal de cette thèse est l’étude des réponses immunes humorales et cellulaires
dirigées contre les antigènes de P. falciparum candidats vaccins dans une zone d’endémie
palustre à transmission saisonnière.
Le premier objectif était d’évaluer le rôle des Tregs dans le biais Th1/Th2 des cellules T
néonatales provenant du sang de cordon de placentas parasités par P. falciparum. Nous
avons montré comme dans l’étude d’Ismaili et al, (2003) que l’infection placentaire par P.
falciparum entraîne une diminution de la production d’IFN- et une augmentation de celle
du sCD30 par les cellules T néonatales. Cette infection est aussi responsable de
l’augmentation de la production d’IL-10 par les cellules mononuclées du sang de cordon
provenant de placentas parasités. Cette production d’IL-10 est assurée par les Tregs comme
le révèle les expériences de déplétion de ces dernières. En effet, la déplétion des Tregs
entraîne la réduction de la production d’IL-10 et par conséquent la restauration de celle de
l’IFN- par les CMSCs en réponse aux antigènes du parasite (extrait de schizontes,
érythrocytes parasités et la protéine MSP119). Un effet similaire à la déplétion des Tregs sur
la production des cytokines par les CMSCs a été obtenu en utilisant un anticorps monoclonal
anti-IL-10. Nos résultats suggèrent que l’infection par P. falciparum influence la
différenciation Th1 des cellules T néonatales en impliquant les Tregs à travers leur
production d’IL-10.
Le second objectif de cette thèse était d’évaluer l’antigénicité d’une protéine triple mutant
et des peptides synthétisés à partir des séquences normale et mutée du candidat vaccin
MSP119. En effet, la protéine triple mutant a été synthétisée afin d’améliorer l’antigénicité
du candidat vaccin MSP119. Cependant, la réponse antigénique de cette protéine n’était pas
connue dans une population vivant en zone d’endémie palustre. Nous avons montré que les
adultes gambiens produisaient à la fois des taux élevés de cytokines Th1 (IFN-) et Th0/Th2
(IL-13 et sCD30) en réponses aux protéines normale et triple mutant et aux peptides dérivant
des ces deux formes. La protéine MSP119 triple mutant (Glu27Tyr, Leu31Arg et Glu43Leu) a
induit une plus forte production d’IFN- par rapport au type sauvage chez les adultes

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 119
gambiens naturellement exposés au Plasmodium. Nous avons aussi montré que les peptides
modifiés induisent une forte production d'IL-13 que les peptides sauvages, tandis que les
peptides conservés P16 et P19 stimulent les taux les plus élevés d'IFN- et d'IL-13 et / ou de
sCD30 comparativement à la majorité des peptides MSP119. Nous avons identifié le peptide
P16 comme porteur d’un épitope immuno dominant parce que reconnu par 63% de la
population étudiée. Le peptide P16 n’était restreint par un type de HLA-DR particulier. Nos
résultats ont fourni des informations nouvelles et très utiles pour le développement futur
d’un vaccin ainsi que l’immunomodulation Th1/Th2 en utilisant soit la protéine sauvage ou
mutée en combinaison avec leurs peptides.
Le dernier objectif de cette thèse était d’évaluer les différences interethniques des réponses
humorales dirigées contre les antigènes candidats vaccins CSP, AMA1 et MSP119 chez les
Peuls, les Wolofs et les Mandingues de Gambie. En effet, les études antérieures ont montré
que les Peuls du Burkina Faso et du Mali semblaient plus résistants au paludisme que les
autres groupes ethniques ceci, en raison de leurs fortes réponses humorales à plusieurs
antigènes de P. falciparum. Cependant, peu d’études avaient étudié la différence
interethnique de la réponse humorale impliquant les Peuls dans un autre pays d’Afrique de
l’Ouest. Contrairement à la situation observée au Burkina Faso, nous avons trouvé la même
incidence du paludisme, la même prévalence parasitaire chez les Peuls de Gambie
comparativement aux groupes ethniques sympatriques Wolofs et Mandingues. Nous avons
montré que de fortes réponses humorales anti- MSP119 et anti-AMA1, et non anti-CSP
étaient associées à la protection contre le paludisme Clinique. Le fait nouveau est que la
prise en compte de la parasitémie à l’inclusion a annulé toutes les associations entre
réponses humorales dirigées contres tous les antigènes candidats vaccins et la protection
contre le paludisme clinique.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 120
VI. Perspectives
La présente thèse ouvre différentes perspectives à savoir :
Etudier l’immunogénicité de la protéine triple mutant de MSP119 et des peptides
immunodominants dans des populations résidant dans les zones palustres d’endémie
différentes.
Etudier la corrélation entre la réponse immunitaire humorale dirigée contre la
protéine triple mutant de MSP119 conçue pour améliorer la production d’anticorps
inhibiteurs de P. falciparum et réduire la production d’anticorps bloquants et
augmenter la protection contre le paludisme clinique à P. falciparum.
Recherche des facteurs génétiques liés aux différences interethniques des réponses
immunitaires humorales observées entre les Peuls de Gambie et les groupes
ethniques Wolof et Mandingue.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 121

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 122
Bibliographie
ABDULLA, S., OBERHOLZER, R., JUMA, O., KUBHOJA, S., MACHERA, F., MEMBI, C., OMARI, S.,
URASSA, A., MSHINDA, H., JUMANNE, A., SALIM, N., SHOMARI, M., AEBI, T.,
SCHELLENBERG, D. M., CARTER, T., VILLAFANA, T., DEMOITIE, M. A., DUBOIS, M. C.,
LEACH, A., LIEVENS, M., VEKEMANS, J., COHEN, J., BALLOU, W. R. & TANNER, M.
(2008) Safety and immunogenicity of RTS,S/AS02D malaria vaccine in infants. N Engl J
Med, 359, 2533-44.
ACHTMAN, A. H., BULL, P. C., STEPHENS, R. & LANGHORNE, J. (2005) Longevity of the
immune response and memory to blood-stage malaria infection. Curr Top Microbiol
Immunol, 297, 71-102.
AGARWAL, A., GUINDO, A., CISSOKO, Y., TAYLOR, J. G., COULIBALY, D., KONE, A., KAYENTAO,
K., DJIMDE, A., PLOWE, C. V., DOUMBO, O., WELLEMS, T. E. & DIALLO, D. (2000)
Hemoglobin C associated with protection from severe malaria in the Dogon of Mali, a
West African population with a low prevalence of hemoglobin S. Blood, 96, 2358-63.
AKDIS, C. A. & BLASER, K. (1999) IL-10-induced anergy in peripheral T cell and reactivation by
microenvironmental cytokines: two key steps in specific immunotherapy. FASEB J, 13,
603-9.
ALIN, M. H. (1997) In vitro susceptibility of Tanzanian wild isolates of Plasmodium falciparum
to artemisinin, chloroquine, sulfadoxine/pyrimethamine and mefloquine.
Parasitology, 114 ( Pt 6), 503-6.
ALLEN, S. J., O'DONNELL, A., ALEXANDER, N. D., ALPERS, M. P., PETO, T. E., CLEGG, J. B. &
WEATHERALL, D. J. (1997) alpha+-Thalassemia protects children against disease
caused by other infections as well as malaria. Proc Natl Acad Sci U S A, 94, 14736-41.
ALLISON, A. C. (1954) Protection afforded by sickle-cell trait against subtertian malareal
infection. Br Med J, 1, 290-4.
ALLISON, A. C. & CLYDE, D. F. (1961) Malaria in African children with deficient erythrocyte
glucose-6-phosphate dehydrogenase. Br Med J, 1, 1346-9.
ALLSOPP, C. E., HARDING, R. M., TAYLOR, C., BUNCE, M., KWIATKOWSKI, D., ANSTEY, N.,
BREWSTER, D., MCMICHAEL, A. J., GREENWOOD, B. M. & HILL, A. V. (1992)

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 123
Interethnic genetic differentiation in Africa: HLA class I antigens in The Gambia. Am J
Hum Genet, 50, 411-21.
ALONSO, P. L., SACARLAL, J., APONTE, J. J., LEACH, A., MACETE, E., AIDE, P., SIGAUQUE, B.,
MILMAN, J., MANDOMANDO, I., BASSAT, Q., GUINOVART, C., ESPASA, M.,
CORACHAN, S., LIEVENS, M., NAVIA, M. M., DUBOIS, M. C., MENENDEZ, C.,
DUBOVSKY, F., COHEN, J., THOMPSON, R. & BALLOU, W. R. (2005) Duration of
protection with RTS,S/AS02A malaria vaccine in prevention of Plasmodium
falciparum disease in Mozambican children: single-blind extended follow-up of a
randomised controlled trial. Lancet, 366, 2012-8.
ALONSO, P. L., SACARLAL, J., APONTE, J. J., LEACH, A., MACETE, E., MILMAN, J.,
MANDOMANDO, I., SPIESSENS, B., GUINOVART, C., ESPASA, M., BASSAT, Q., AIDE, P.,
OFORI-ANYINAM, O., NAVIA, M. M., CORACHAN, S., CEUPPENS, M., DUBOIS, M. C.,
DEMOITIE, M. A., DUBOVSKY, F., MENENDEZ, C., TORNIEPORTH, N., BALLOU, W. R.,
THOMPSON, R. & COHEN, J. (2004) Efficacy of the RTS,S/AS02A vaccine against
Plasmodium falciparum infection and disease in young African children: randomised
controlled trial. Lancet, 364, 1411-20.
AMANTE, F. H. & GOOD, M. F. (1997) Prolonged Th1-like response generated by a
Plasmodium yoelii-specific T cell clone allows complete clearance of infection in
reconstituted mice. Parasite Immunol, 19, 111-26.
AMANTE, F. H., STANLEY, A. C., RANDALL, L. M., ZHOU, Y., HAQUE, A., MCSWEENEY, K.,
WATERS, A. P., JANSE, C. J., GOOD, M. F., HILL, G. R. & ENGWERDA, C. R. (2007) A role
for natural regulatory T cells in the pathogenesis of experimental cerebral malaria.
Am J Pathol, 171, 548-59.
APONTE, J. J., AIDE, P., RENOM, M., MANDOMANDO, I., BASSAT, Q., SACARLAL, J., MANACA,
M. N., LAFUENTE, S., BARBOSA, A., LEACH, A., LIEVENS, M., VEKEMANS, J.,
SIGAUQUE, B., DUBOIS, M. C., DEMOITIE, M. A., SILLMAN, M., SAVARESE, B., MCNEIL,
J. G., MACETE, E., BALLOU, W. R., COHEN, J. & ALONSO, P. L. (2007) Safety of the
RTS,S/AS02D candidate malaria vaccine in infants living in a highly endemic area of
Mozambique: a double blind randomised controlled phase I/IIb trial. Lancet, 370,
1543-51.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 124
ASSEMAN, C., MAUZE, S., LEACH, M. W., COFFMAN, R. L. & POWRIE, F. (1999) An essential
role for interleukin 10 in the function of regulatory T cells that inhibit intestinal
inflammation. J Exp Med, 190, 995-1004.
AUDRAN, R., CACHAT, M., LURATI, F., SOE, S., LEROY, O., CORRADIN, G., DRUILHE, P. &
SPERTINI, F. (2005) Phase I malaria vaccine trial with a long synthetic peptide derived
from the merozoite surface protein 3 antigen. Infect Immun, 73, 8017-26.
ARAMA, C., GIUSTI, P., BOSTRÖM, S., DARA, V., TRAORE, B., DOLO, A., DOUMBO, O., VARANI,
S. & TROYE-BLOMBERG M. (2011) Interethnic differences in antigen-presenting cell
activation and TLR responses in Malian children during Plasmodium falciparum
malaria. PlosOne, 6 (3):e18319
BABIKER, H. A., PRINGLE, S. J., ABDEL-MUHSIN, A., MACKINNON, M., HUNT, P. & WALLIKER,
D. (2001) High-level chloroquine resistance in Sudanese isolates of Plasmodium
falciparum is associated with mutations in the chloroquine resistance transporter
gene pfcrt and the multidrug resistance Gene pfmdr1. J Infect Dis, 183, 1535-8.
BAIRD, J. K., JONES, T. R., DANUDIRGO, E. W., ANNIS, B. A., BANGS, M. J., BASRI, H.,
PURNOMO & MASBAR, S. (1991) Age-dependent acquired protection against
Plasmodium falciparum in people having two years exposure to hyperendemic
malaria. Am J Trop Med Hyg, 45, 65-76.
BAIRD, J. K., PURNOMO, BASRI, H., BANGS, M. J., ANDERSEN, E. M., JONES, T. R., MASBAR, S.,
HARJOSUWARNO, S., SUBIANTO, B. & ARBANI, P. R. (1993) Age-specific prevalence of
Plasmodium falciparum among six populations with limited histories of exposure to
endemic malaria. Am J Trop Med Hyg, 49, 707-19.
BARUCH, D. I., PASLOSKE, B. L., SINGH, H. B., BI, X., MA, X. C., FELDMAN, M., TARASCHI, T. F.
& HOWARD, R. J. (1995) Cloning the P. falciparum gene encoding PfEMP1, a malarial
variant antigen and adherence receptor on the surface of parasitized human
erythrocytes. Cell, 82, 77-87.
BASCO, L. K. & RINGWALD, P. (2001) Analysis of the key pfcrt point mutation and in vitro and
in vivo response to chloroquine in Yaounde, Cameroon. J Infect Dis, 183, 1828-31.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 125
BATE, C. A. & KWIATKOWSKI, D. (1994) A monoclonal antibody that recognizes
phosphatidylinositol inhibits induction of tumor necrosis factor alpha by different
strains of Plasmodium falciparum. Infect Immun, 62, 5261-6.
BATE, C. A., TAVERNE, J. & PLAYFAIR, J. H. (1988) Malarial parasites induce TNF production
by macrophages. Immunology, 64, 227-31.
BEESON, J. G., AMIN, N., KANJALA, M. & ROGERSON, S. J. (2002) Selective accumulation of
mature asexual stages of Plasmodium falciparum-infected erythrocytes in the
placenta. Infect Immun, 70, 5412-5.
BEESON, J. G., ROGERSON, S. J., COOKE, B. M., REEDER, J. C., CHAI, W., LAWSON, A. M.,
MOLYNEUX, M. E. & BROWN, G. V. (2000) Adhesion of Plasmodium falciparum-
infected erythrocytes to hyaluronic acid in placental malaria. Nat Med, 6, 86-90.
BEIER, J. C., ONYANGO, F. K., KOROS, J. K., RAMADHAN, M., OGWANG, R., WIRTZ, R. A.,
KOECH, D. K. & ROBERTS, C. R. (1991) Quantitation of malaria sporozoites
transmitted in vitro during salivation by wild Afrotropical Anopheles. Med Vet
Entomol, 5, 71-9.
BEJON, P., BERKLEY, J. A., MWANGI, T., OGADA, E., MWANGI, I., MAITLAND, K., WILLIAMS,
T., SCOTT, J. A., ENGLISH, M., LOWE, B. S., PESHU, N., NEWTON, C. R. & MARSH, K.
(2007) Defining childhood severe falciparum malaria for intervention studies. PLoS
Med, 4, e251.
BEJON, P., LUSINGU, J., OLOTU, A., LEACH, A., LIEVENS, M., VEKEMANS, J., MSHAMU, S.,
LANG, T., GOULD, J., DUBOIS, M. C., DEMOITIE, M. A., STALLAERT, J. F., VANSADIA, P.,
CARTER, T., NJUGUNA, P., AWUONDO, K. O., MALABEJA, A., ABDUL, O., GESASE, S.,
MTURI, N., DRAKELEY, C. J., SAVARESE, B., VILLAFANA, T., BALLOU, W. R., COHEN, J.,
RILEY, E. M., LEMNGE, M. M., MARSH, K. & VON SEIDLEIN, L. (2008) Efficacy of
RTS,S/AS01E vaccine against malaria in children 5 to 17 months of age. N Engl J Med,
359, 2521-32.
BEJON, P., MWACHARO, J., KAI, O., TODRYK, S., KEATING, S., LOWE, B., LANG, T., MWANGI,
T. W., GILBERT, S. C., PESHU, N., MARSH, K. & HILL, A. V. (2007) The induction and
persistence of T cell IFN-gamma responses after vaccination or natural exposure is
suppressed by Plasmodium falciparum. J Immunol, 179, 4193-201.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 126
BENITO, A., ROCHE, J., MOLINA, R., AMELA, C. & ALVAR, J. (1995) In vitro susceptibility of
Plasmodium falciparum to chloroquine, amodiaquine, quinine, mefloquine, and
sulfadoxine/pyrimethamine in Equatorial Guinea. Am J Trop Med Hyg, 53, 526-31.
BERENDT, A. R., MCDOWALL, A., CRAIG, A. G., BATES, P. A., STERNBERG, M. J., MARSH, K.,
NEWBOLD, C. I. & HOGG, N. (1992) The binding site on ICAM-1 for Plasmodium
falciparum-infected erythrocytes overlaps, but is distinct from, the LFA-1-binding site.
Cell, 68, 71-81.
BEUTLER, B. & GRAU, G. E. (1993) Tumor necrosis factor in the pathogenesis of infectious
diseases. Crit Care Med, 21, S423-35.
BIENZLE, U., AYENI, O., LUCAS, A. O. & LUZZATTO, L. (1972) Glucose-6-phosphate
dehydrogenase and malaria. Greater resistance of females heterozygous for enzyme
deficiency and of males with non-deficient variant. Lancet, 1, 107-10.
BLACKMAN, M. J., HEIDRICH, H. G., DONACHIE, S., MCBRIDE, J. S. & HOLDER, A. A. (1990) A
single fragment of a malaria merozoite surface protein remains on the parasite
during red cell invasion and is the target of invasion-inhibiting antibodies. J Exp Med,
172, 379-82.
BLACKMAN, M. J., SCOTT-FINNIGAN, T. J., SHAI, S. & HOLDER, A. A. (1994) Antibodies inhibit
the protease-mediated processing of a malaria merozoite surface protein. J Exp Med,
180, 389-93.
BLUESTONE, J. A. & ABBAS, A. K. (2003) Natural versus adaptive regulatory T cells. Nat Rev
Immunol, 3, 253-7.
BOLAD, A., FAROUK, S. E., ISRAELSSON, E., DOLO, A., DOUMBO, O. K., NEBIÉ, I., MAIGA, B.,
KOURIBA, B., LUONI, G., SIRIMA, B. S., MODIANO, D., BERZINS, K. & TROYE-
BLOMBERG, M. (2005) Distinct Interethnic Differences in Immunoglobulin G
Class/Subclass and Immunoglobulin M Antibody Responses to Malaria Antigens but
not in Immunoglobulin G Responses to Nonmalarial Antigens in Sympatric Tribes
Living in West Africa. Scandinavian Journal of immunology, 61:380-386
BOUHAROUN-TAYOUN, H., OEUVRAY, C., LUNEL, F. & DRUILHE, P. (1995) Mechanisms
underlying the monocyte-mediated antibody-dependent killing of Plasmodium
falciparum asexual blood stages. J Exp Med, 182, 409-18.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 127
BRABIN, B. J. (1983) An analysis of malaria in pregnancy in Africa. Bull World Health Organ,
61, 1005-16.
BRASSEUR, P., KOUAMOUO, J., MOYOU, R. S. & DRUILHE, P. (1990) Emergence of
mefloquine-resistant malaria in Africa without drug pressure. Lancet, 336, 59.
BRASSEUR, P., KOUAMOUO, J., MOYOU, R. S. & DRUILHE, P. (1992) Mefloquine resistant
malaria in Cameroon and correlation with resistance to quinine. Mem Inst Oswaldo
Cruz, 87 Suppl 3, 271-3.
BROWN, H., TURNER, G., ROGERSON, S., TEMBO, M., MWENECHANYA, J., MOLYNEUX, M. &
TAYLOR, T. (1999) Cytokine expression in the brain in human cerebral malaria. J Infect
Dis, 180, 1742-6.
BRUSTOSKI, K., MOLLER, U., KRAMER, M., HARTGERS, F. C., KREMSNER, P. G., KRZYCH, U. &
LUTY, A. J. (2006) Reduced cord blood immune effector-cell responsiveness mediated
by CD4+ cells induced in utero as a consequence of placental Plasmodium falciparum
infection. J Infect Dis, 193, 146-54.
BULL, P. C., LOWE, B. S., KORTOK, M. & MARSH, K. (1999) Antibody recognition of
Plasmodium falciparum erythrocyte surface antigens in Kenya: evidence for rare and
prevalent variants. Infect Immun, 67, 733-9.
CALLE, J. M., NARDIN, E. H., CLAVIJO, P., BOUDIN, C., STUBER, D., TAKACS, B., NUSSENZWEIG,
R. S. & COCHRANE, A. H. (1992) Recognition of different domains of the Plasmodium
falciparum CS protein by the sera of naturally infected individuals compared with
those of sporozoite-immunized volunteers. J Immunol, 149, 2695-701.
CAMBOS, M., BELANGER, B., JACQUES, A., ROULET, A. & SCORZA, T. (2008) Natural
regulatory (CD4+CD25+FOXP+) T cells control the production of pro-inflammatory
cytokines during Plasmodium chabaudi adami infection and do not contribute to
immune evasion. Int J Parasitol, 38, 229-38.
CARLSON, J., HELMBY, H., HILL, A. V., BREWSTER, D., GREENWOOD, B. M. & WAHLGREN, M.
(1990) Human cerebral malaria: association with erythrocyte rosetting and lack of
anti-rosetting antibodies. Lancet, 336, 1457-60.
CARVALHO, L. H., SANO, G., HAFALLA, J. C., MORROT, A., CUROTTO DE LAFAILLE, M. A. &
ZAVALA, F. (2002) IL-4-secreting CD4+ T cells are crucial to the development of CD8+
T-cell responses against malaria liver stages. Nat Med, 8, 166-70.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 128
CASARES, S., BRUMEANU, T. D. & RICHIE, T. L. (2010) The RTS,S malaria vaccine. Vaccine, 28,
4880-94.
CHAIYAROJ, S. C., BURANAKITI, A., ANGKASEKWINAI, P., LOORESSUWAN, S. & COWMAN, A.
F. (1999) Analysis of mefloquine resistance and amplification of pfmdr1 in multidrug-
resistant Plasmodium falciparum isolates from Thailand. Am J Trop Med Hyg, 61, 780-
3.
CHANG, S. P., CASE, S. E., GOSNELL, W. L., HASHIMOTO, A., KRAMER, K. J., TAM, L. Q.,
HASHIRO, C. Q., NIKAIDO, C. M., GIBSON, H. L., LEE-NG, C. T., BARR, P. J., YOKOTA, B.
T. & HUT, G. S. (1996) A recombinant baculovirus 42-kilodalton C-terminal fragment
of Plasmodium falciparum merozoite surface protein 1 protects Aotus monkeys
against malaria. Infect Immun, 64, 253-61.
CHANG, S. P., GIBSON, H. L., LEE-NG, C. T., BARR, P. J. & HUI, G. S. (1992) A carboxyl-terminal
fragment of Plasmodium falciparum gp195 expressed by a recombinant baculovirus
induces antibodies that completely inhibit parasite growth. J Immunol, 149, 548-55.
CHAOUAT, G., CAYOL, V., MAIROVITZ, V. & DUBANCHET, S. (1999) Localization of the Th2
cytokines IL-3, IL-4, IL-10 at the fetomaternal interface during human and murine
pregnancy and lack of requirement for Fas/Fas ligand interaction for a successful
allogeneic pregnancy. Am J Reprod Immunol, 42, 1-13.
CHELIMO, K., OFULLA, A. V., NARUM, D. L., KAZURA, J. W., LANAR, D. E. & JOHN, C. C. (2005)
Antibodies to Plasmodium falciparum antigens vary by age and antigen in children in
a malaria-holoendemic area of Kenya. Pediatr Infect Dis J, 24, 680-4.
CHEN, N., RUSSELL, B., STALEY, J., KOTECKA, B., NASVELD, P. & CHENG, Q. (2001) Sequence
polymorphisms in pfcrt are strongly associated with chloroquine resistance in
Plasmodium falciparum. J Infect Dis, 183, 1543-5.
CLARK, I. A., AL YAMAN, F. M. & JACOBSON, L. S. (1997) The biological basis of malarial
disease. Int J Parasitol, 27, 1237-49.
CLARK, I. A. & COWDEN, W. B. (2003) The pathophysiology of falciparum malaria. Pharmacol
Ther, 99, 221-60.
CLARK, I. A. & ROCKETT, K. A. (1994) Sequestration, cytokines, and malaria pathology. Int J
Parasitol, 24, 165-6.
CLARK, I. A. & SCHOFIELD, L. (2000) Pathogenesis of malaria. Parasitol Today, 16, 451-4.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 129
COBAN, C., ISHII, K. J., SULLIVAN, D. J. & KUMAR, N. (2002) Purified malaria pigment
(hemozoin) enhances dendritic cell maturation and modulates the isotype of
antibodies induced by a DNA vaccine. Infect Immun, 70, 3939-43.
COCKBURN, I. A., MACKINNON, M. J., O'DONNELL, A., ALLEN, S. J., MOULDS, J. M., BAISOR,
M., BOCKARIE, M., REEDER, J. C. & ROWE, J. A. (2004) A human complement receptor
1 polymorphism that reduces Plasmodium falciparum rosetting confers protection
against severe malaria. Proc Natl Acad Sci U S A, 101, 272-7.
COHEN, S., MC, G. I. & CARRINGTON, S. (1961) Gamma-globulin and acquired immunity to
human malaria. Nature, 192, 733-7.
CONWAY, D. J., CAVANAGH, D. R., TANABE, K., ROPER, C., MIKES, Z. S., SAKIHAMA, N.,
BOJANG, K. A., ODUOLA, A. M., KREMSNER, P. G., ARNOT, D. E., GREENWOOD, B. M.
& MCBRIDE, J. S. (2000) A principal target of human immunity to malaria identified by
molecular population genetic and immunological analyses. Nat Med, 6, 689-92.
CORTES, A., MELLOMBO, M., MASCIANTONIO, R., MURPHY, V. J., REEDER, J. C. & ANDERS, R.
F. (2005) Allele specificity of naturally acquired antibody responses against
Plasmodium falciparum apical membrane antigen 1. Infect Immun, 73, 422-30.
COUPER, K. N., BLOUNT, D. G., WILSON, M. S., HAFALLA, J. C., BELKAID, Y., KAMANAKA, M.,
FLAVELL, R. A., DE SOUZA, J. B. & RILEY, E. M. (2008) IL-10 from CD4CD25Foxp3CD127
adaptive regulatory T cells modulates parasite clearance and pathology during
malaria infection. PLoS Pathog, 4, e1000004.
CRAIG, A. & SCHERF, A. (2001) Molecules on the surface of the Plasmodium falciparum
infected erythrocyte and their role in malaria pathogenesis and immune evasion. Mol
Biochem Parasitol, 115, 129-43.
CREASEY, A. M., STAALSOE, T., RAZA, A., ARNOT, D. E. & ROWE, J. A. (2003) Nonspecific
immunoglobulin M binding and chondroitin sulfate A binding are linked phenotypes
of Plasmodium falciparum isolates implicated in malaria during pregnancy. Infect
Immun, 71, 4767-71.
CROMPTON, P. D., PIERCE, S. K. & MILLER, L. H. (2010) Advances and challenges in malaria
vaccine development. J Clin Invest, 120, 4168-78.
CRUTCHER, J. M., STEVENSON, M. M., SEDEGAH, M. & HOFFMAN, S. L. (1995) Interleukin-12
and malaria. Res Immunol, 146, 552-9.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 130
DAME, J. B., WILLIAMS, J. L., MCCUTCHAN, T. F., WEBER, J. L., WIRTZ, R. A., HOCKMEYER, W.
T., MALOY, W. L., HAYNES, J. D., SCHNEIDER, I., ROBERTS, D. & ET AL. (1984) Structure
of the gene encoding the immunodominant surface antigen on the sporozoite of the
human malaria parasite Plasmodium falciparum. Science, 225, 593-9.
DAY, N. P., HIEN, T. T., SCHOLLAARDT, T., LOC, P. P., CHUONG, L. V., CHAU, T. T., MAI, N. T.,
PHU, N. H., SINH, D. X., WHITE, N. J. & HO, M. (1999) The prognostic and
pathophysiologic role of pro- and antiinflammatory cytokines in severe malaria. J
Infect Dis, 180, 1288-97.
DE SOUZA, J. B. & RILEY, E. M. (2002) Cerebral malaria: the contribution of studies in animal
models to our understanding of immunopathogenesis. Microbes Infect, 4, 291-300.
DELORON, P. & CHOUGNET, C. (1992) Is immunity to malaria really short-lived? Parasitol
Today, 8, 375-8.
DESAI, M., TER KUILE, F. O., NOSTEN, F., MCGREADY, R., ASAMOA, K., BRABIN, B. &
NEWMAN, R. D. (2007) Epidemiology and burden of malaria in pregnancy. Lancet
Infect Dis, 7, 93-104.
DJIMDE, A., DOUMBO, O. K., CORTESE, J. F., KAYENTAO, K., DOUMBO, S., DIOURTE, Y.,
DICKO, A., SU, X. Z., NOMURA, T., FIDOCK, D. A., WELLEMS, T. E., PLOWE, C. V. &
COULIBALY, D. (2001) A molecular marker for chloroquine-resistant falciparum
malaria. N Engl J Med, 344, 257-63.
DLUZEWSKI, A. R., LING, I. T., HOPKINS, J. M., GRAINGER, M., MARGOS, G., MITCHELL, G. H.,
HOLDER, A. A. & BANNISTER, L. H. (2008) Formation of the food vacuole in
Plasmodium falciparum: a potential role for the 19 kDa fragment of merozoite
surface protein 1 (MSP1(19)). PLoS One, 3, e3085.
DODOO, D., THEANDER, T. G., KURTZHALS, J. A., KORAM, K., RILEY, E., AKANMORI, B. D.,
NKRUMAH, F. K. & HVIID, L. (1999) Levels of antibody to conserved parts of
Plasmodium falciparum merozoite surface protein 1 in Ghanaian children are not
associated with protection from clinical malaria. Infect Immun, 67, 2131-7.
D'OMBRAIN, M. C., VOSS, T. S., MAIER, A. G., PEARCE, J. A., HANSEN, D. S., COWMAN, A. F. &
SCHOFIELD, L. (2007) Plasmodium falciparum erythrocyte membrane protein-1
specifically suppresses early production of host interferon-gamma. Cell Host Microbe,
2, 130-8.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 131
DOOLAN, D. L. & HOFFMAN, S. L. (2000) The complexity of protective immunity against liver-
stage malaria. J Immunol, 165, 1453-62.
DORFMAN, J. R., BEJON, P., NDUNGU, F. M., LANGHORNE, J., KORTOK, M. M., LOWE, B. S.,
MWANGI, T. W., WILLIAMS, T. N. & MARSH, K. (2005) B cell memory to 3 Plasmodium
falciparum blood-stage antigens in a malaria-endemic area. J Infect Dis, 191, 1623-30.
DRUILHE, P., SPERTINI, F., SOESOE, D., CORRADIN, G., MEJIA, P., SINGH, S., AUDRAN, R.,
BOUZIDI, A., OEUVRAY, C. & ROUSSILHON, C. (2005) A malaria vaccine that elicits in
humans antibodies able to kill Plasmodium falciparum. PLoS Med, 2, e344.
DUFFY, M. F., BYRNE, T. J., ELLIOTT, S. R., WILSON, D. W., ROGERSON, S. J., BEESON, J. G.,
NOVIYANTI, R. & BROWN, G. V. (2005) Broad analysis reveals a consistent pattern of
var gene transcription in Plasmodium falciparum repeatedly selected for a defined
adhesion phenotype. Mol Microbiol, 56, 774-88.
DUFFY, P. E. & FRIED, M. (2003) Antibodies that inhibit Plasmodium falciparum adhesion to
chondroitin sulfate A are associated with increased birth weight and the gestational
age of newborns. Infect Immun, 71, 6620-3.
DUFFY, P. E. & FRIED, M. (2005) Malaria in the pregnant woman. Curr Top Microbiol
Immunol, 295, 169-200.
DURAISINGH, M. T., JONES, P., SAMBOU, I., VON SEIDLEIN, L., PINDER, M. & WARHURST, D.
C. (2000) The tyrosine-86 allele of the pfmdr1 gene of Plasmodium falciparum is
associated with increased sensitivity to the anti-malarials mefloquine and
artemisinin. Mol Biochem Parasitol, 108, 13-23.
EGAN, A., WATERFALL, M., PINDER, M., HOLDER, A. & RILEY, E. (1997) Characterization of
human T- and B-cell epitopes in the C terminus of Plasmodium falciparum merozoite
surface protein 1: evidence for poor T-cell recognition of polypeptides with
numerous disulfide bonds. Infect Immun, 65, 3024-31.
EGAN, A. F., MORRIS, J., BARNISH, G., ALLEN, S., GREENWOOD, B. M., KASLOW, D. C.,
HOLDER, A. A. & RILEY, E. M. (1996) Clinical immunity to Plasmodium falciparum
malaria is associated with serum antibodies to the 19-kDa C-terminal fragment of the
merozoite surface antigen, PfMSP-1. J Infect Dis, 173, 765-9.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 132
ENGLISH, M., WARUIRU, C., AMUKOYE, E., MURPHY, S., CRAWLEY, J., MWANGI, I., PESHU, N.
& MARSH, K. (1996) Deep breathing in children with severe malaria: indicator of
metabolic acidosis and poor outcome. Am J Trop Med Hyg, 55, 521-4.
ESEN, M., KREMSNER, P. G., SCHLEUCHER, R., GASSLER, M., IMOUKHUEDE, E. B., IMBAULT,
N., LEROY, O., JEPSEN, S., KNUDSEN, B. W., SCHUMM, M., KNOBLOCH, J., THEISEN, M.
& MORDMULLER, B. (2009) Safety and immunogenicity of GMZ2 - a MSP3-GLURP
fusion protein malaria vaccine candidate. Vaccine, 27, 6862-8.
ESPOSITO, F., LOMBARDI, S., MODIANO, D., ZAVALA, F., REEME, J., LAMIZANA, L., COLUZZI,
M. & NUSSENZWEIG, R. S. (1988) Prevalence and levels of antibodies to the
circumsporozoite protein of Plasmodium falciparum in an endemic area and their
relationship to resistance against malaria infection. Trans R Soc Trop Med Hyg, 82,
827-32.
FAROUK, S. E., DOLO, A., BERECZKY, S., KOURIBA, B., MAIGA, B., FARNERT, A., PERLMANN,
H., HAYANO, M., MONTGOMERY, S. M., DOUMBO, O. K. & TROYE-BLOMBERG, M.
(2005) Different antibody- and cytokine-mediated responses to Plasmodium
falciparum parasite in two sympatric ethnic tribes living in Mali. Microbes Infect, 7,
110-7.
FAVILA-CASTILLO, L., MONROY-OSTRIA, A., KOBAYASHI, E., HIRUNPETCHARAT, C., KAMADA,
N. & GOOD, M. F. (1996) Protection of rats against malaria by a transplanted immune
spleen. Parasite Immunol, 18, 325-31.
FELL, A. H., CURRIER, J. & GOOD, M. F. (1994) Inhibition of Plasmodium falciparum growth in
vitro by CD4+ and CD8+ T cells from non-exposed donors. Parasite Immunol, 16, 579-
86.
FERRANTE, A., KUMARATILAKE, L., RZEPCZYK, C. M. & DAYER, J. M. (1990) Killing of
Plasmodium falciparum by cytokine activated effector cells (neutrophils and
macrophages). Immunol Lett, 25, 179-87.
FIDOCK, D. A., NOMURA, T., TALLEY, A. K., COOPER, R. A., DZEKUNOV, S. M., FERDIG, M. T.,
URSOS, L. M., SIDHU, A. B., NAUDE, B., DEITSCH, K. W., SU, X. Z., WOOTTON, J. C.,
ROEPE, P. D. & WELLEMS, T. E. (2000) Mutations in the P. falciparum digestive
vacuole transmembrane protein PfCRT and evidence for their role in chloroquine
resistance. Mol Cell, 6, 861-71.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 133
FISCHER, P. R. & BOONE, P. (1998) Short report: severe malaria associated with blood group.
Am J Trop Med Hyg, 58, 122-3.
FLICK, K., AHUJA, S., CHENE, A., BEJARANO, M. T. & CHEN, Q. (2004) Optimized expression of
Plasmodium falciparum erythrocyte membrane protein 1 domains in Escherichia coli.
Malar J, 3, 50.
FLICK, K., SCHOLANDER, C., CHEN, Q., FERNANDEZ, V., POUVELLE, B., GYSIN, J. & WAHLGREN,
M. (2001) Role of nonimmune IgG bound to PfEMP1 in placental malaria. Science,
293, 2098-100.
FLINT, J., HILL, A. V., BOWDEN, D. K., OPPENHEIMER, S. J., SILL, P. R., SERJEANTSON, S. W.,
BANA-KOIRI, J., BHATIA, K., ALPERS, M. P., BOYCE, A. J. & ET AL. (1986) High
frequencies of alpha-thalassaemia are the result of natural selection by malaria.
Nature, 321, 744-50.
FLORENS, L., WASHBURN, M. P., RAINE, J. D., ANTHONY, R. M., GRAINGER, M., HAYNES, J. D.,
MOCH, J. K., MUSTER, N., SACCI, J. B., TABB, D. L., WITNEY, A. A., WOLTERS, D., WU,
Y., GARDNER, M. J., HOLDER, A. A., SINDEN, R. E., YATES, J. R. & CARUCCI, D. J. (2002)
A proteomic view of the Plasmodium falciparum life cycle. Nature, 419, 520-6.
FLUECK, T. P., JELINEK, T., KILIAN, A. H., ADAGU, I. S., KABAGAMBE, G., SONNENBURG, F. &
WARHURST, D. C. (2000) Correlation of in vivo-resistance to chloroquine and allelic
polymorphisms in Plasmodium falciparum isolates from Uganda. Trop Med Int
Health, 5, 174-8.
FREEMAN, R. R. & HOLDER, A. A. (1983) Surface antigens of malaria merozoites. A high
molecular weight precursor is processed to an 83,000 mol wt form expressed on the
surface of Plasmodium falciparum merozoites. J Exp Med, 158, 1647-53.
FRIED, M. & DUFFY, P. E. (1996) Adherence of Plasmodium falciparum to chondroitin sulfate
A in the human placenta. Science, 272, 1502-4.
FRIED, M. & DUFFY, P. E. (1998) Maternal malaria and parasite adhesion. J Mol Med, 76, 162-
71.
FRIED, M., NOSTEN, F., BROCKMAN, A., BRABIN, B. J. & DUFFY, P. E. (1998) Maternal
antibodies block malaria. Nature, 395, 851-2.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 134
GARDNER, J. P., PINCHES, R. A., ROBERTS, D. J. & NEWBOLD, C. I. (1996) Variant antigens and
endothelial receptor adhesion in Plasmodium falciparum. Proc Natl Acad Sci U S A,
93, 3503-8.
GARDNER, M. J., HALL, N., FUNG, E., WHITE, O., BERRIMAN, M., HYMAN, R. W., CARLTON, J.
M., PAIN, A., NELSON, K. E., BOWMAN, S., PAULSEN, I. T., JAMES, K., EISEN, J. A.,
RUTHERFORD, K., SALZBERG, S. L., CRAIG, A., KYES, S., CHAN, M. S., NENE, V.,
SHALLOM, S. J., SUH, B., PETERSON, J., ANGIUOLI, S., PERTEA, M., ALLEN, J.,
SELENGUT, J., HAFT, D., MATHER, M. W., VAIDYA, A. B., MARTIN, D. M., FAIRLAMB, A.
H., FRAUNHOLZ, M. J., ROOS, D. S., RALPH, S. A., MCFADDEN, G. I., CUMMINGS, L. M.,
SUBRAMANIAN, G. M., MUNGALL, C., VENTER, J. C., CARUCCI, D. J., HOFFMAN, S. L.,
NEWBOLD, C., DAVIS, R. W., FRASER, C. M. & BARRELL, B. (2002) Genome sequence
of the human malaria parasite Plasmodium falciparum. Nature, 419, 498-511.
GAY, F., BUSTOS, D. G., DIQUET, B., ROJAS RIVERO, L., LITAUDON, M., PICHET, C., DANIS, M.
& GENTILINI, M. (1990) Cross-resistance between mefloquine and halofantrine.
Lancet, 336, 1262.
GENTON, B., BETUELA, I., FELGER, I., AL-YAMAN, F., ANDERS, R. F., SAUL, A., RARE, L.,
BAISOR, M., LORRY, K., BROWN, G. V., PYE, D., IRVING, D. O., SMITH, T. A., BECK, H. P.
& ALPERS, M. P. (2002) A recombinant blood-stage malaria vaccine reduces
Plasmodium falciparum density and exerts selective pressure on parasite populations
in a phase 1-2b trial in Papua New Guinea. J Infect Dis, 185, 820-7.
GILBERT, S. C., PLEBANSKI, M., GUPTA, S., MORRIS, J., COX, M., AIDOO, M., KWIATKOWSKI,
D., GREENWOOD, B. M., WHITTLE, H. C. & HILL, A. V. (1998) Association of malaria
parasite population structure, HLA, and immunological antagonism. Science, 279,
1173-7.
GOOD, M. F., POMBO, D., QUAKYI, I. A., RILEY, E. M., HOUGHTEN, R. A., MENON, A., ALLING,
D. W., BERZOFSKY, J. A. & MILLER, L. H. (1988) Human T-cell recognition of the
circumsporozoite protein of Plasmodium falciparum: immunodominant T-cell
domains map to the polymorphic regions of the molecule. Proc Natl Acad Sci U S A,
85, 1199-203.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 135
GORDON, D. M., MCGOVERN, T. W., KRZYCH, U., COHEN, J. C., SCHNEIDER, I., LACHANCE, R.,
HEPPNER, D. G., YUAN, G., HOLLINGDALE, M., SLAOUI, M. & ET AL. (1995) Safety,
immunogenicity, and efficacy of a recombinantly produced Plasmodium falciparum
circumsporozoite protein-hepatitis B surface antigen subunit vaccine. J Infect Dis,
171, 1576-85.
GORIELY, S., VINCART, B., STORDEUR, P., VEKEMANS, J., WILLEMS, F., GOLDMAN, M. & DE
WIT, D. (2001) Deficient IL-12(p35) gene expression by dendritic cells derived from
neonatal monocytes. J Immunol, 166, 2141-6.
GRAU, G. E., MACKENZIE, C. D., CARR, R. A., REDARD, M., PIZZOLATO, G., ALLASIA, C.,
CATALDO, C., TAYLOR, T. E. & MOLYNEUX, M. E. (2003) Platelet accumulation in brain
microvessels in fatal pediatric cerebral malaria. J Infect Dis, 187, 461-6.
GRAU, G. E., TAYLOR, T. E., MOLYNEUX, M. E., WIRIMA, J. J., VASSALLI, P., HOMMEL, M. &
LAMBERT, P. H. (1989) Tumor necrosis factor and disease severity in children with
falciparum malaria. N Engl J Med, 320, 1586-91.
GREENWOOD, B., MARSH, K. & SNOW, R. (1991) Why do some African children develop
severe malaria? Parasitol Today, 7, 277-81.
GREENWOOD, B. M. & ARMSTRONG, J. R. (1991) Comparison of two simple methods for
determining malaria parasite density. Trans R Soc Trop Med Hyg, 85, 186-8.
GREENWOOD, B. M., GROENENDAAL, F., BRADLEY, A. K., GREENWOOD, A. M., SHENTON, F.,
TULLOCH, S. & HAYES, R. (1987) Ethnic differences in the prevalence of splenomegaly
and malaria in The Gambia. Ann Trop Med Parasitol, 81, 345-54.
GUEVARA PATINO, J. A., HOLDER, A. A., MCBRIDE, J. S. & BLACKMAN, M. J. (1997) Antibodies
that inhibit malaria merozoite surface protein-1 processing and erythrocyte invasion
are blocked by naturally acquired human antibodies. J Exp Med, 186, 1689-99.
GUINDO, A., FAIRHURST, R. M., DOUMBO, O. K., WELLEMS, T. E. & DIALLO, D. A. (2007) X-
linked G6PD deficiency protects hemizygous males but not heterozygous females
against severe malaria. PLoS Med, 4, e66.
GUINET, F., DIALLO, D. A., MINTA, D., DICKO, A., SISSOKO, M. S., KEITA, M. M., WELLEMS, T.
E. & DOUMBO, O. (1997) A comparison of the incidence of severe malaria in Malian
children with normal and C-trait hemoglobin profiles. Acta Trop, 68, 175-82.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 136
GUINOVART, C., APONTE, J. J., SACARLAL, J., AIDE, P., LEACH, A., BASSAT, Q., MACETE, E.,
DOBANO, C., LIEVENS, M., LOUCQ, C., BALLOU, W. R., COHEN, J. & ALONSO, P. L.
(2009) Insights into long-lasting protection induced by RTS,S/AS02A malaria vaccine:
further results from a phase IIb trial in Mozambican children. PLoS One, 4, e5165.
GUPTA, S., SNOW, R. W., DONNELLY, C. A., MARSH, K. & NEWBOLD, C. (1999) Immunity to
non-cerebral severe malaria is acquired after one or two infections. Nat Med, 5, 340-
3.
GUTTINGER, M., CASPERS, P., TAKACS, B., TRZECIAK, A., GILLESSEN, D., PINK, J. R. &
SINIGAGLIA, F. (1988) Human T cells recognize polymorphic and non-polymorphic
regions of the Plasmodium falciparum circumsporozoite protein. EMBO J, 7, 2555-8.
GUYATT, H. L. & SNOW, R. W. (2004) Impact of malaria during pregnancy on low birth weight
in sub-Saharan Africa. Clin Microbiol Rev, 17, 760-9, table of contents.
GYSIN, J., BARNWELL, J., SCHLESINGER, D. H., NUSSENZWEIG, V. & NUSSENZWEIG, R. S.
(1984) Neutralization of the infectivity of sporozoites of Plasmodium knowlesi by
antibodies to a synthetic peptide. J Exp Med, 160, 935-40.
HANSEN, D. S., BERNARD, N. J., NIE, C. Q. & SCHOFIELD, L. (2007) NK cells stimulate
recruitment of CXCR3+ T cells to the brain during Plasmodium berghei-mediated
cerebral malaria. J Immunol, 178, 5779-88.
HARRIS, P. K., YEOH, S., DLUZEWSKI, A. R., O'DONNELL, R. A., WITHERS-MARTINEZ, C.,
HACKETT, F., BANNISTER, L. H., MITCHELL, G. H. & BLACKMAN, M. J. (2005) Molecular
identification of a malaria merozoite surface sheddase. PLoS Pathog, 1, 241-51.
HASTINGS, I. M. & D'ALESSANDRO, U. (2000) Modelling a predictable disaster: the rise and
spread of drug-resistantmalaria. Parasitol Today, 16, 340-7.
HAY, S. I., GUERRA, C. A., GETHING, P. W., PATIL, A. P., TATEM, A. J., NOOR, A. M., KABARIA,
C. W., MANH, B. H., ELYAZAR, I. R., BROOKER, S., SMITH, D. L., MOYEED, R. A. &
SNOW, R. W. (2009) A world malaria map: Plasmodium falciparum endemicity in
2007. PLoS Med, 6, e1000048.
HENSMANN, M., LI, C., MOSS, C., LINDO, V., GREER, F., WATTS, C., OGUN, S. A., HOLDER, A.
A. & LANGHORNE, J. (2004) Disulfide bonds in merozoite surface protein 1 of the
malaria parasite impede efficient antigen processing and affect the in vivo antibody
response. Eur J Immunol, 34, 639-48.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 137
HEPPNER, D. G., JR., KESTER, K. E., OCKENHOUSE, C. F., TORNIEPORTH, N., OFORI, O., LYON,
J. A., STEWART, V. A., DUBOIS, P., LANAR, D. E., KRZYCH, U., MORIS, P., ANGOV, E.,
CUMMINGS, J. F., LEACH, A., HALL, B. T., DUTTA, S., SCHWENK, R., HILLIER, C.,
BARBOSA, A., WARE, L. A., NAIR, L., DARKO, C. A., WITHERS, M. R., OGUTU, B.,
POLHEMUS, M. E., FUKUDA, M., PICHYANGKUL, S., GETTYACAMIN, M., DIGGS, C.,
SOISSON, L., MILMAN, J., DUBOIS, M. C., GARCON, N., TUCKER, K., WITTES, J., PLOWE,
C. V., THERA, M. A., DUOMBO, O. K., PAU, M. G., GOUDSMIT, J., BALLOU, W. R. &
COHEN, J. (2005) Towards an RTS,S-based, multi-stage, multi-antigen vaccine against
falciparum malaria: progress at the Walter Reed Army Institute of Research. Vaccine,
23, 2243-50.
HERMSEN, C. C., VERHAGE, D. F., TELGT, D. S., TEELEN, K., BOUSEMA, J. T., ROESTENBERG,
M., BOLAD, A., BERZINS, K., CORRADIN, G., LEROY, O., THEISEN, M. & SAUERWEIN, R.
W. (2007) Glutamate-rich protein (GLURP) induces antibodies that inhibit in vitro
growth of Plasmodium falciparum in a phase 1 malaria vaccine trial. Vaccine, 25,
2930-40.
HILL, A. V., REYES-SANDOVAL, A., O'HARA, G., EWER, K., LAWRIE, A., GOODMAN, A.,
NICOSIA, A., FOLGORI, A., COLLOCA, S., CORTESE, R., GILBERT, S. C. & DRAPER, S. J.
(2010) Prime-boost vectored malaria vaccines: progress and prospects. Hum Vaccin,
6, 78-83.
HIRUNPETCHARAT, C. & GOOD, M. F. (1998) Deletion of Plasmodium berghei-specific CD4+ T
cells adoptively transferred into recipient mice after challenge with homologous
parasite. Proc Natl Acad Sci U S A, 95, 1715-20.
HISAEDA, H., MAEKAWA, Y., IWAKAWA, D., OKADA, H., HIMENO, K., KISHIHARA, K.,
TSUKUMO, S. & YASUTOMO, K. (2004) Escape of malaria parasites from host
immunity requires CD4+ CD25+ regulatory T cells. Nat Med, 10, 29-30.
HODDER, A. N., CREWTHER, P. E. & ANDERS, R. F. (2001) Specificity of the protective
antibody response to apical membrane antigen 1. Infect Immun, 69, 3286-94.
HODDER, A. N., CREWTHER, P. E., MATTHEW, M. L., REID, G. E., MORITZ, R. L., SIMPSON, R. J.
& ANDERS, R. F. (1996) The disulfide bond structure of Plasmodium apical membrane
antigen-1. J Biol Chem, 271, 29446-52.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 138
HOFFMAN, S. L., GOH, L. M., LUKE, T. C., SCHNEIDER, I., LE, T. P., DOOLAN, D. L., SACCI, J., DE
LA VEGA, P., DOWLER, M., PAUL, C., GORDON, D. M., STOUTE, J. A., CHURCH, L. W.,
SEDEGAH, M., HEPPNER, D. G., BALLOU, W. R. & RICHIE, T. L. (2002) Protection of
humans against malaria by immunization with radiation-attenuated Plasmodium
falciparum sporozoites. J Infect Dis, 185, 1155-64.
HOLDER, A. A. (2009) The carboxy-terminus of merozoite surface protein 1: structure,
specific antibodies and immunity to malaria. Parasitology, 136, 1445-56.
HOLDER, A. A., GUEVARA PATINO, J. A., UTHAIPIBULL, C., SYED, S. E., LING, I. T., SCOTT-
FINNIGAN, T. & BLACKMAN, M. J. (1999) Merozoite surface protein 1, immune
evasion, and vaccines against asexual blood stage malaria. Parassitologia, 41, 409-14.
HORI, S., CARVALHO, T. L. & DEMENGEOT, J. (2002) CD25+CD4+ regulatory T cells suppress
CD4+ T cell-mediated pulmonary hyperinflammation driven by Pneumocystis carinii
in immunodeficient mice. Eur J Immunol, 32, 1282-91.
HORI, S., NOMURA, T. & SAKAGUCHI, S. (2003) Control of regulatory T cell development by
the transcription factor Foxp3. Science, 299, 1057-61.
HORII, T., SHIRAI, H., JIE, L., ISHII, K. J., PALACPAC, N. Q., TOUGAN, T., HATO, M., OHTA, N.,
BOBOGARE, A., ARAKAKI, N., MATSUMOTO, Y., NAMAZUE, J., ISHIKAWA, T., UEDA, S.
& TAKAHASHI, M. (2010) Evidences of protection against blood-stage infection of
Plasmodium falciparum by the novel protein vaccine SE36. Parasitol Int, 59, 380-6.
HOWELL, S. A., WITHERS-MARTINEZ, C., KOCKEN, C. H., THOMAS, A. W. & BLACKMAN, M. J.
(2001) Proteolytic processing and primary structure of Plasmodium falciparum apical
membrane antigen-1. J Biol Chem, 276, 31311-20.
ISMAILI, J., VAN DER SANDE, M., HOLLAND, M. J., SAMBOU, I., KEITA, S., ALLSOPP, C., OTA,
M. O., MCADAM, K. P. & PINDER, M. (2003) Plasmodium falciparum infection of the
placenta affects newborn immune responses. Clin Exp Immunol, 133, 414-21.
JAMES, E. (2005) Apoptosis: key to the attenuated malaria vaccine? J Infect Dis, 191, 1573-5.
JASON, J., ARCHIBALD, L. K., NWANYANWU, O. C., BELL, M., BUCHANAN, I., LARNED, J.,
KAZEMBE, P. N., DOBBIE, H., PAREKH, B., BYRD, M. G., EICK, A., HAN, A. & JARVIS, W.
R. (2001) Cytokines and malaria parasitemia. Clin Immunol, 100, 208-18.
JELINEK, T., GROBUSCH, M. P. & LOSCHER, T. (2001) Patterns of Plasmodium falciparum drug
resistance in nonimmune travellers to Africa. Eur J Clin Microbiol Infect Dis, 20, 284-6.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 139
JOHN, C. C., ZICKAFOOSE, J. S., SUMBA, P. O., KING, C. L. & KAZURA, J. W. (2003) Antibodies
to the Plasmodium falciparum antigens circumsporozoite protein, thrombospondin-
related adhesive protein, and liver-stage antigen 1 vary by ages of subjects and by
season in a highland area of Kenya. Infect Immun, 71, 4320-5.
JOHNSON, A. H., LEKE, R. G., MENDELL, N. R., SHON, D., SUH, Y. J., BOMBA-NKOLO, D.,
TCHINDA, V., KOUONTCHOU, S., THUITA, L. W., VAN DER WEL, A. M., THOMAS, A.,
STOWERS, A., SAUL, A., ZHOU, A., TAYLOR, D. W. & QUAKYI, I. A. (2004) Human
leukocyte antigen class II alleles influence levels of antibodies to the Plasmodium
falciparum asexual-stage apical membrane antigen 1 but not to merozoite surface
antigen 2 and merozoite surface protein 1. Infect Immun, 72, 2762-71.
KATO, K., MAYER, D. C., SINGH, S., REID, M. & MILLER, L. H. (2005) Domain III of Plasmodium
falciparum apical membrane antigen 1 binds to the erythrocyte membrane protein
Kx. Proc Natl Acad Sci U S A, 102, 5552-7.
KESTER, K. E., CUMMINGS, J. F., OCKENHOUSE, C. F., NIELSEN, R., HALL, B. T., GORDON, D.
M., SCHWENK, R. J., KRZYCH, U., HOLLAND, C. A., RICHMOND, G., DOWLER, M. G.,
WILLIAMS, J., WIRTZ, R. A., TORNIEPORTH, N., VIGNERON, L., DELCHAMBRE, M.,
DEMOITIE, M. A., BALLOU, W. R., COHEN, J. & HEPPNER, D. G., JR. (2008) Phase 2a
trial of 0, 1, and 3 month and 0, 7, and 28 day immunization schedules of malaria
vaccine RTS,S/AS02 in malaria-naive adults at the Walter Reed Army Institute of
Research. Vaccine, 26, 2191-202.
KESTER, K. E., CUMMINGS, J. F., OFORI-ANYINAM, O., OCKENHOUSE, C. F., KRZYCH, U.,
MORIS, P., SCHWENK, R., NIELSEN, R. A., DEBEBE, Z., PINELIS, E., JUOMPAN, L.,
WILLIAMS, J., DOWLER, M., STEWART, V. A., WIRTZ, R. A., DUBOIS, M. C., LIEVENS,
M., COHEN, J., BALLOU, W. R. & HEPPNER, D. G., JR. (2009) Randomized, double-
blind, phase 2a trial of falciparum malaria vaccines RTS,S/AS01B and RTS,S/AS02A in
malaria-naive adults: safety, efficacy, and immunologic associates of protection. J
Infect Dis, 200, 337-46.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 140
KESTER, K. E., MCKINNEY, D. A., TORNIEPORTH, N., OCKENHOUSE, C. F., HEPPNER, D. G.,
HALL, T., KRZYCH, U., DELCHAMBRE, M., VOSS, G., DOWLER, M. G., PALENSKY, J.,
WITTES, J., COHEN, J. & BALLOU, W. R. (2001) Efficacy of recombinant
circumsporozoite protein vaccine regimens against experimental Plasmodium
falciparum malaria. J Infect Dis, 183, 640-7.
KESTER, K. E., MCKINNEY, D. A., TORNIEPORTH, N., OCKENHOUSE, C. F., HEPPNER, D. G., JR.,
HALL, T., WELLDE, B. T., WHITE, K., SUN, P., SCHWENK, R., KRZYCH, U., DELCHAMBRE,
M., VOSS, G., DUBOIS, M. C., GASSER, R. A., JR., DOWLER, M. G., O'BRIEN, M.,
WITTES, J., WIRTZ, R., COHEN, J. & BALLOU, W. R. (2007) A phase I/IIa safety,
immunogenicity, and efficacy bridging randomized study of a two-dose regimen of
liquid and lyophilized formulations of the candidate malaria vaccine RTS,S/AS02A in
malaria-naive adults. Vaccine, 25, 5359-66.
KINYANJUI, S. M., CONWAY, D. J., LANAR, D. E. & MARSH, K. (2007) IgG antibody responses
to Plasmodium falciparum merozoite antigens in Kenyan children have a short half-
life. Malar J, 6, 82.
KLOTZ, F. W., SCHELLER, L. F., SEGUIN, M. C., KUMAR, N., MARLETTA, M. A., GREEN, S. J. &
AZAD, A. F. (1995) Co-localization of inducible-nitric oxide synthase and Plasmodium
berghei in hepatocytes from rats immunized with irradiated sporozoites. J Immunol,
154, 3391-5.
KOCH, O., AWOMOYI, A., USEN, S., JALLOW, M., RICHARDSON, A., HULL, J., PINDER, M.,
NEWPORT, M. & KWIATKOWSKI, D. (2002) IFNGR1 gene promoter polymorphisms
and susceptibility to cerebral malaria. J Infect Dis, 185, 1684-1687
KOSSODO, S., MONSO, C., JUILLARD, P., VELU, T., GOLDMAN, M. & GRAU, G. E. (1997)
Interleukin-10 modulates susceptibility in experimental cerebral malaria.
Immunology, 91, 536-40.
KOUSSIS, K., WITHERS-MARTINEZ, C., YEOH, S., CHILD, M., HACKETT, F., KNUEPFER, E.,
JULIANO, L., WOEHLBIER, U., BUJARD, H. & BLACKMAN, M. J. (2009) A
multifunctional serine protease primes the malaria parasite for red blood cell
invasion. EMBO J, 28, 725-35.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 141
KRISHNAN, L., GUILBERT, L. J., WEGMANN, T. G., BELOSEVIC, M. & MOSMANN, T. R. (1996) T
helper 1 response against Leishmania major in pregnant C57BL/6 mice increases
implantation failure and fetal resorptions. Correlation with increased IFN-gamma and
TNF and reduced IL-10 production by placental cells. J Immunol, 156, 653-62.
KULLBERG, M. C., JANKOVIC, D., GORELICK, P. L., CASPAR, P., LETTERIO, J. J., CHEEVER, A. W.
& SHER, A. (2002) Bacteria-triggered CD4(+) T regulatory cells suppress Helicobacter
hepaticus-induced colitis. J Exp Med, 196, 505-15.
KUMAR, S., MILLER, L. H., QUAKYI, I. A., KEISTER, D. B., HOUGHTEN, R. A., MALOY, W. L.,
MOSS, B., BERZOFSKY, J. A. & GOOD, M. F. (1988) Cytotoxic T cells specific for the
circumsporozoite protein of Plasmodium falciparum. Nature, 334, 258-60.
KUMARATILAKE, L. M. & FERRANTE, A. (1992) IL-4 inhibits macrophage-mediated killing of
Plasmodium falciparum in vitro. A possible parasite-immune evasion mechanism. J
Immunol, 149, 194-9.
KUMARATILAKE, L. M., FERRANTE, A. & RZEPCZYK, C. (1991) The role of T lymphocytes in
immunity to Plasmodium falciparum. Enhancement of neutrophil-mediated parasite
killing by lymphotoxin and IFN-gamma: comparisons with tumor necrosis factor
effects. J Immunol, 146, 762-7.
KWIATKOWSKI, D., HILL, A. V., SAMBOU, I., TWUMASI, P., CASTRACANE, J., MANOGUE, K. R.,
CERAMI, A., BREWSTER, D. R. & GREENWOOD, B. M. (1990) TNF concentration in
fatal cerebral, non-fatal cerebral, and uncomplicated Plasmodium falciparum malaria.
Lancet, 336, 1201-4.
LAMBROS, C. & VANDERBERG, J. P. (1979) Synchronization of Plasmodium falciparum
erythrocytic stages in culture. J Parasitol, 65, 418-20.
LANDGRAF, B., KOLLARITSCH, H., WIEDERMANN, G. & WERNSDORFER, W. H. (1994)
Plasmodium falciparum: susceptibility in vitro and in vivo to chloroquine and
sulfadoxine-pyrimethamine in Ghanaian schoolchildren. Trans R Soc Trop Med Hyg,
88, 440-2.
LANGHORNE, J., NDUNGU, F. M., SPONAAS, A. M. & MARSH, K. (2008) Immunity to malaria:
more questions than answers. Nat Immunol, 9, 725-32.
LEIRIAO, P., MOTA, M. M. & RODRIGUEZ, A. (2005) Apoptotic Plasmodium-infected
hepatocytes provide antigens to liver dendritic cells. J Infect Dis, 191, 1576-81.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 142
LEITNER, W. W. & KRZYCH, U. (1997) Plasmodium falciparum malaria blood stage parasites
preferentially inhibit macrophages with high phagocytic activity. Parasite Immunol,
19, 103-10.
LING, I. T., OGUN, S. A. & HOLDER, A. A. (1994) Immunization against malaria with a
recombinant protein. Parasite Immunol, 16, 63-7.
LING, I. T., OGUN, S. A., MOMIN, P., RICHARDS, R. L., GARCON, N., COHEN, J., BALLOU, W. R.
& HOLDER, A. A. (1997) Immunization against the murine malaria parasite
Plasmodium yoelii using a recombinant protein with adjuvants developed for clinical
use. Vaccine, 15, 1562-7.
LIU, W., PUTNAM, A. L., XU-YU, Z., SZOT, G. L., LEE, M. R., ZHU, S., GOTTLIEB, P. A.,
KAPRANOV, P., GINGERAS, T. R., FAZEKAS DE ST GROTH, B., CLAYBERGER, C., SOPER,
D. M., ZIEGLER, S. F. & BLUESTONE, J. A. (2006) CD127 expression inversely correlates
with FoxP3 and suppressive function of human CD4+ T reg cells. J Exp Med, 203,
1701-11.
LUONI, G., VERRA, F., ARCA, B., SIRIMA, B. S., TROYE-BLOMBERG, M., COLUZZI, M.,
KWIATKOWSKI, D. & MODIANO, D. (2001) Antimalarial antibody levels and IL4
polymorphism in the Fulani of West Africa. Genes Immun, 2, 411-4.
LUTY, A. J., LELL, B., SCHMIDT-OTT, R., LEHMAN, L. G., LUCKNER, D., GREVE, B., MATOUSEK,
P., HERBICH, K., SCHMID, D., MIGOT-NABIAS, F., DELORON, P., NUSSENZWEIG, R. S. &
KREMSNER, P. G. (1999) Interferon-gamma responses are associated with resistance
to reinfection with Plasmodium falciparum in young African children. J Infect Dis, 179,
980-8.
LUTY, A. J., PERKINS, D. J., LELL, B., SCHMIDT-OTT, R., LEHMAN, L. G., LUCKNER, D., GREVE,
B., MATOUSEK, P., HERBICH, K., SCHMID, D., WEINBERG, J. B. & KREMSNER, P. G.
(2000) Low interleukin-12 activity in severe Plasmodium falciparum malaria. Infect
Immun, 68, 3909-15.
MACKINNON, M. J., MWANGI, T. W., SNOW, R. W., MARSH, K. & WILLIAMS, T. N. (2005)
Heritability of malaria in Africa. PLoS Med, 2, e340.
MACKINTOSH, C. L., BEESON, J. G. & MARSH, K. (2004) Clinical features and pathogenesis of
severe malaria. Trends Parasitol, 20, 597-603.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 143
MAEDA, H. & SHIRAISHI, A. (1996) TGF-beta contributes to the shift toward Th2-type
responses through direct and IL-10-mediated pathways in tumor-bearing mice. J
Immunol, 156, 73-8.
MAIER, A. G., DURAISINGH, M. T., REEDER, J. C., PATEL, S. S., KAZURA, J. W., ZIMMERMAN, P.
A. & COWMAN, A. F. (2003) Plasmodium falciparum erythrocyte invasion through
glycophorin C and selection for Gerbich negativity in human populations. Nat Med, 9,
87-92.
MALHOTRA, I., WAMACHI, A. N., MUNGAI, P. L., MZUNGU, E., KOECH, D., MUCHIRI, E.,
MOORMANN, A. M. & KING, C. L. (2008) Fine specificity of neonatal lymphocytes to
an abundant malaria blood-stage antigen: epitope mapping of Plasmodium
falciparum MSP1(33). J Immunol, 180, 3383-90.
MALKIN, E., HU, J., LI, Z., CHEN, Z., BI, X., REED, Z., DUBOVSKY, F., LIU, J., WANG, Q., PAN, X.,
CHEN, T., GIERSING, B., XU, Y., KANG, X., GU, J., SHEN, Q., TUCKER, K., TIERNEY, E.,
PAN, W., LONG, C. & CAO, Z. (2008) A phase 1 trial of PfCP2.9: an AMA1/MSP1
chimeric recombinant protein vaccine for Plasmodium falciparum malaria. Vaccine,
26, 6864-73.
MALOY, K. J., SALAUN, L., CAHILL, R., DOUGAN, G., SAUNDERS, N. J. & POWRIE, F. (2003)
CD4+CD25+ T(R) cells suppress innate immune pathology through cytokine-
dependent mechanisms. J Exp Med, 197, 111-9.
MARSH, K. (1992) Malaria--a neglected disease? Parasitology, 104 Suppl, S53-69.
MARSH, K., FORSTER, D., WARUIRU, C., MWANGI, I., WINSTANLEY, M., MARSH, V., NEWTON,
C., WINSTANLEY, P., WARN, P., PESHU, N. & ET AL. (1995) Indicators of life-
threatening malaria in African children. N Engl J Med, 332, 1399-404.
MARSH, K. & KINYANJUI, S. (2006) Immune effector mechanisms in malaria. Parasite
Immunol, 28, 51-60.
MARSH, K., OTOO, L., HAYES, R. J., CARSON, D. C. & GREENWOOD, B. M. (1989) Antibodies to
blood stage antigens of Plasmodium falciparum in rural Gambians and their relation
to protection against infection. Trans R Soc Trop Med Hyg, 83, 293-303.
MATEJEVIC, D., NEUDECK, H., GRAF, R., MULLER, T. & DIETL, J. (2001) Localization of
hyaluronan with a hyaluronan-specific hyaluronic acid binding protein in the placenta
in pre-eclampsia. Gynecol Obstet Invest, 52, 257-9.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 144
MAUBERT, B., FIEVET, N., TAMI, G., BOUDIN, C. & DELORON, P. (1998) Plasmodium
falciparum-isolates from Cameroonian pregnant women do not rosette. Parasite, 5,
281-3.
MAYOR, A. G., GOMEZ-OLIVE, X., APONTE, J. J., CASIMIRO, S., MABUNDA, S., DGEDGE, M.,
BARRETO, A. & ALONSO, P. L. (2001) Prevalence of the K76T mutation in the putative
Plasmodium falciparum chloroquine resistance transporter (pfcrt) gene and its
relation to chloroquine resistance in Mozambique. J Infect Dis, 183, 1413-6.
MCGREGOR, I. A., WILSON, M. E. & BILLEWICZ, W. Z. (1983) Malaria infection of the placenta
in The Gambia, West Africa; its incidence and relationship to stillbirth, birthweight
and placental weight. Trans R Soc Trop Med Hyg, 77, 232-44.
MCGUIRE, W., HILL, A. V., ALLSOPP, C. E., GREENWOOD, B. M. & KWIATKOWSKI, D. (1994)
Variation in the TNF-alpha promoter region associated with susceptibility to cerebral
malaria. Nature, 371, 508-10.
MCHUGH, R. S., WHITTERS, M. J., PICCIRILLO, C. A., YOUNG, D. A., SHEVACH, E. M., COLLINS,
M. & BYRNE, M. C. (2002) CD4(+)CD25(+) immunoregulatory T cells: gene expression
analysis reveals a functional role for the glucocorticoid-induced TNF receptor.
Immunity, 16, 311-23.
MEGNEKOU, R., STAALSOE, T., TAYLOR, D. W., LEKE, R. & HVIID, L. (2005) Effects of
pregnancy and intensity of Plasmodium falciparum transmission on immunoglobulin
G subclass responses to variant surface antigens. Infect Immun, 73, 4112-8.
MENENDEZ, C., ORDI, J., ISMAIL, M. R., VENTURA, P. J., APONTE, J. J., KAHIGWA, E., FONT, F.
& ALONSO, P. L. (2000) The impact of placental malaria on gestational age and birth
weight. J Infect Dis, 181, 1740-5.
METZGER, W. G., OKENU, D. M., CAVANAGH, D. R., ROBINSON, J. V., BOJANG, K. A., WEISS,
H. A., MCBRIDE, J. S., GREENWOOD, B. M. & CONWAY, D. J. (2003) Serum IgG3 to the
Plasmodium falciparum merozoite surface protein 2 is strongly associated with a
reduced prospective risk of malaria. Parasite Immunol, 25, 307-12.
MIGOT-NABIAS, F., MOMBO, L. E., LUTY, A. J., DUBOIS, B., NABIAS, R., BISSEYE, C., MILLET, P.,
LU, C. Y. & DELORON, P. (2000) Human genetic factors related to susceptibility to
mild malaria in Gabon. Genes Immun, 1, 435-41.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 145
MILLER, L. H., BARUCH, D. I., MARSH, K. & DOUMBO, O. K. (2002) The pathogenic basis of
malaria. Nature, 415, 673-9.
MILLER, L. H., MASON, S. J., CLYDE, D. F. & MCGINNISS, M. H. (1976) The resistance factor to
Plasmodium vivax in blacks. The Duffy-blood-group genotype, FyFy. N Engl J Med,
295, 302-4.
MILLER, L. H., ROBERTS, T., SHAHABUDDIN, M. & MCCUTCHAN, T. F. (1993) Analysis of
sequence diversity in the Plasmodium falciparum merozoite surface protein-1 (MSP-
1). Mol Biochem Parasitol, 59, 1-14.
MITCHELL, G. H., THOMAS, A. W., MARGOS, G., DLUZEWSKI, A. R. & BANNISTER, L. H. (2004)
Apical membrane antigen 1, a major malaria vaccine candidate, mediates the close
attachment of invasive merozoites to host red blood cells. Infect Immun, 72, 154-8.
MOCKENHAUPT, F. P., EHRHARDT, S., CRAMER, J. P., OTCHWEMAH, R. N., ANEMANA, S. D.,
GOLTZ, K., MYLIUS, F., DIETZ, E., EGGELTE, T. A. & BIENZLE, U. (2004) Hemoglobin C
and resistance to severe malaria in Ghanaian children. J Infect Dis, 190, 1006-9.
MODIANO, D., LUONI, G., PETRARCA, V., SODIOMON SIRIMA, B., DE LUCA, M., SIMPORE, J.,
COLUZZI, M., BODMER, J. G. & MODIANO, G. (2001) HLA class I in three West African
ethnic groups: genetic distances from sub-Saharan and Caucasoid populations. Tissue
Antigens, 57, 128-37.
MODIANO, D., LUONI, G., SIRIMA, B. S., SIMPORE, J., VERRA, F., KONATE, A., RASTRELLI, E.,
OLIVIERI, A., CALISSANO, C., PAGANOTTI, G. M., D'URBANO, L., SANOU, I.,
SAWADOGO, A., MODIANO, G. & COLUZZI, M. (2001) Haemoglobin C protects against
clinical Plasmodium falciparum malaria. Nature, 414, 305-8.
MODIANO, D., PETRARCA, V., SIRIMA, B. S., NEBIE, I., DIALLO, D., ESPOSITO, F. & COLUZZI, M.
(1996) Different response to Plasmodium falciparum malaria in west African
sympatric ethnic groups. Proc Natl Acad Sci U S A, 93, 13206-11.
MOLD, J. E., MICHAELSSON, J., BURT, T. D., MUENCH, M. O., BECKERMAN, K. P., BUSCH, M.
P., LEE, T. H., NIXON, D. F. & MCCUNE, J. M. (2008) Maternal alloantigens promote
the development of tolerogenic fetal regulatory T cells in utero. Science, 322, 1562-5.
MOLYNEUX, D. H., FLOYD, K., BARNISH, G. & FEVRE, E. M. (1999) Transmission control and
drug resistance in malaria: a crucial interaction. Parasitol Today, 15, 238-40.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 146
MONTAGNOLI, C., BACCI, A., BOZZA, S., GAZIANO, R., MOSCI, P., SHARPE, A. H. & ROMANI, L.
(2002) B7/CD28-dependent CD4+CD25+ regulatory T cells are essential components
of the memory-protective immunity to Candida albicans. J Immunol, 169, 6298-308.
MOORE, J. M., NAHLEN, B. L., MISORE, A., LAL, A. A. & UDHAYAKUMAR, V. (1999) Immunity
to placental malaria. I. Elevated production of interferon-gamma by placental blood
mononuclear cells is associated with protection in an area with high transmission of
malaria. J Infect Dis, 179, 1218-25.
MORGAN, W. D., BIRDSALL, B., FRENKIEL, T. A., GRADWELL, M. G., BURGHAUS, P. A., SYED, S.
E., UTHAIPIBULL, C., HOLDER, A. A. & FEENEY, J. (1999) Solution structure of an EGF
module pair from the Plasmodium falciparum merozoite surface protein 1. J Mol Biol,
289, 113-22.
NAKABAYASHI, T., LETTERIO, J. J., GEISER, A. G., KONG, L., OGAWA, N., ZHAO, W., KOIKE, T.,
FERNANDES, G., DANG, H. & TALAL, N. (1997) Up-regulation of cytokine mRNA,
adhesion molecule proteins, and MHC class II proteins in salivary glands of TGF-beta1
knockout mice: MHC class II is a factor in the pathogenesis of TGF-beta1 knockout
mice. J Immunol, 158, 5527-35.
NAKAMURA, K., KITANI, A. & STROBER, W. (2001) Cell contact-dependent
immunosuppression by CD4(+)CD25(+) regulatory T cells is mediated by cell surface-
bound transforming growth factor beta. J Exp Med, 194, 629-44.
NARDIN, E., ZAVALA, F., NUSSENZWEIG, V. & NUSSENZWEIG, R. S. (1999) Pre-erythrocytic
malaria vaccine: mechanisms of protective immunity and human vaccine trials.
Parassitologia, 41, 397-402.
NARUM, D. L. & THOMAS, A. W. (1994) Differential localization of full-length and processed
forms of PF83/AMA-1 an apical membrane antigen of Plasmodium falciparum
merozoites. Mol Biochem Parasitol, 67, 59-68.
NEWBOLD, C. I., CRAIG, A. G., KYES, S., BERENDT, A. R., SNOW, R. W., PESHU, N. & MARSH, K.
(1997) PfEMP1, polymorphism and pathogenesis. Ann Trop Med Parasitol, 91, 551-7.
NIE, C. Q., BERNARD, N. J., NORMAN, M. U., AMANTE, F. H., LUNDIE, R. J., CRABB, B. S.,
HEATH, W. R., ENGWERDA, C. R., HICKEY, M. J., SCHOFIELD, L. & HANSEN, D. S. (2009)
IP-10-mediated T cell homing promotes cerebral inflammation over splenic immunity
to malaria infection. PLoS Pathog, 5, e1000369.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 147
NIE, C. Q., BERNARD, N. J., SCHOFIELD, L. & HANSEN, D. S. (2007) CD4+ CD25+ regulatory T
cells suppress CD4+ T-cell function and inhibit the development of Plasmodium
berghei-specific TH1 responses involved in cerebral malaria pathogenesis. Infect
Immun, 75, 2275-82.
NIELSEN, M. A., VESTERGAARD, L. S., LUSINGU, J., KURTZHALS, J. A., GIHA, H. A., GREVSTAD,
B., GOKA, B. Q., LEMNGE, M. M., JENSEN, J. B., AKANMORI, B. D., THEANDER, T. G.,
STAALSOE, T. & HVIID, L. (2004) Geographical and temporal conservation of antibody
recognition of Plasmodium falciparum variant surface antigens. Infect Immun, 72,
3531-5.
NOSTEN, F., ROGERSON, S. J., BEESON, J. G., MCGREADY, R., MUTABINGWA, T. K. & BRABIN,
B. (2004) Malaria in pregnancy and the endemicity spectrum: what can we learn?
Trends Parasitol, 20, 425-32.
NUSSENZWEIG, R. S., VANDERBERG, J., MOST, H. & ORTON, C. (1967) Protective immunity
produced by the injection of x-irradiated sporozoites of plasmodium berghei. Nature,
216, 160-2.
NUSSENZWEIG, V. & NUSSENZWEIG, R. S. (1986) Development of a sporozoite malaria
vaccine. Am J Trop Med Hyg, 35, 678-88.
NWUBA, R. I., SODEINDE, O., ANUMUDU, C. I., OMOSUN, Y. O., ODAIBO, A. B., HOLDER, A. A.
& NWAGWU, M. (2002) The human immune response to Plasmodium falciparum
includes both antibodies that inhibit merozoite surface protein 1 secondary
processing and blocking antibodies. Infect Immun, 70, 5328-31.
OCANA-MORGNER, C., MOTA, M. M. & RODRIGUEZ, A. (2003) Malaria blood stage
suppression of liver stage immunity by dendritic cells. J Exp Med, 197, 143-51.
OCKENHOUSE, C. F. & SHEAR, H. L. (1984) Oxidative killing of the intraerythrocytic malaria
parasite Plasmodium yoelii by activated macrophages. J Immunol, 132, 424-31.
O'GARRA, A. & ARAI, N. (2000) The molecular basis of T helper 1 and T helper 2 cell
differentiation. Trends Cell Biol, 10, 542-50.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 148
OGUTU, B. R., APOLLO, O. J., MCKINNEY, D., OKOTH, W., SIANGLA, J., DUBOVSKY, F., TUCKER,
K., WAITUMBI, J. N., DIGGS, C., WITTES, J., MALKIN, E., LEACH, A., SOISSON, L. A.,
MILMAN, J. B., OTIENO, L., HOLLAND, C. A., POLHEMUS, M., REMICH, S. A.,
OCKENHOUSE, C. F., COHEN, J., BALLOU, W. R., MARTIN, S. K., ANGOV, E., STEWART,
V. A., LYON, J. A., HEPPNER, D. G. & WITHERS, M. R. (2009) Blood stage malaria
vaccine eliciting high antigen-specific antibody concentrations confers no protection
to young children in Western Kenya. PLoS One, 4, e4708.
OKECH, B. A., CORRAN, P. H., TODD, J., JOYNSON-HICKS, A., UTHAIPIBULL, C., EGWANG, T. G.,
HOLDER, A. A. & RILEY, E. M. (2004) Fine specificity of serum antibodies to
Plasmodium falciparum merozoite surface protein, PfMSP-1(19), predicts protection
from malaria infection and high-density parasitemia. Infect Immun, 72, 1557-67.
OKOKO, B. J., OTA, M. O., YAMUAH, L. K., IDIONG, D., MKPANAM, S. N., AVIEKA, A., BANYA,
W. A. & OSINUSI, K. (2002) Influence of placental malaria infection on foetal outcome
in the Gambia: twenty years after Ian Mcgregor. J Health Popul Nutr, 20, 4-11.
OMER, F. M., KURTZHALS, J. A. & RILEY, E. M. (2000) Maintaining the immunological balance
in parasitic infections: a role for TGF-beta? Parasitol Today, 16, 18-23.
OMER, F. M. & RILEY, E. M. (1998) Transforming growth factor beta production is inversely
correlated with severity of murine malaria infection. J Exp Med, 188, 39-48.
OMOSUN, Y. O., ADORO, S., ANUMUDU, C. I., ODAIBO, A. B., UTHIAPIBULL, C., HOLDER, A. A.,
NWAGWU, M. & NWUBA, R. I. (2009) Antibody specificities of children living in a
malaria endemic area to inhibitory and blocking epitopes on MSP-1 19 of
Plasmodium falciparum. Acta Trop, 109, 208-12.
OMS (2010). World Malaria report, 2010 [www.document]
http://whqlibdoc.who.int/publications/2010/9789241564106_eng.pdf
OOMEN, J. M., MEUWISSEN, J. H. & GEMERT, W. (1979) Differences in blood status of three
ethnic groups inhabiting the same locality in Northern Nigeria. Anaemia,
splenomegaly and associated causes. Trop Geogr Med, 31, 587-606.
ORDI, J., ISMAIL, M. R., VENTURA, P. J., KAHIGWA, E., HIRT, R., CARDESA, A., ALONSO, P. L. &
MENENDEZ, C. (1998) Massive chronic intervillositis of the placenta associated with
malaria infection. Am J Surg Pathol, 22, 1006-11.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 149
OSIER, F. H., FEGAN, G., POLLEY, S. D., MURUNGI, L., VERRA, F., TETTEH, K. K., LOWE, B.,
MWANGI, T., BULL, P. C., THOMAS, A. W., CAVANAGH, D. R., MCBRIDE, J. S., LANAR,
D. E., MACKINNON, M. J., CONWAY, D. J. & MARSH, K. (2008) Breadth and magnitude
of antibody responses to multiple Plasmodium falciparum merozoite antigens are
associated with protection from clinical malaria. Infect Immun, 76, 2240-8.
PASQUETTO, V., FIDOCK, D. A., GRAS, H., BADELL, E., ELING, W., BALLOU, W. R., BELGHITI, J.,
TARTAR, A. & DRUILHE, P. (1997) Plasmodium falciparum sporozoite invasion is
inhibited by naturally acquired or experimentally induced polyclonal antibodies to
the STARP antigen. Eur J Immunol, 27, 2502-13.
PASVOL, G., WILSON, R. J., SMALLEY, M. E. & BROWN, J. (1978) Separation of viable schizont-
infected red cells of Plasmodium falciparum from human blood. Ann Trop Med
Parasitol, 72, 87-8.
PENG, J. C., ABU BAKAR, S., RICHARDSON, M. M., JONSSON, J. J., FRAZER, I. H., NIELSEN, L. K.,
MORAHAN, G. & THOMAS, R. (2006) IL10 and IL12B polymorphisms each influence IL-
12p70 secretion by dendritic cells in response to LPS. Immunol Cell Biol, 84, 227-32.
PERERA, K. L., HANDUNNETTI, S. M., HOLM, I., LONGACRE, S. & MENDIS, K. (1998)
Baculovirus merozoite surface protein 1 C-terminal recombinant antigens are highly
protective in a natural primate model for human Plasmodium vivax malaria. Infect
Immun, 66, 1500-6.
PERKINS, D. J., WEINBERG, J. B. & KREMSNER, P. G. (2000) Reduced interleukin-12 and
transforming growth factor-beta1 in severe childhood malaria: relationship of
cytokine balance with disease severity. J Infect Dis, 182, 988-92.
PERRY, J. A., RUSH, A., WILSON, R. J., OLVER, C. S. & AVERY, A. C. (2004) Dendritic cells from
malaria-infected mice are fully functional APC. J Immunol, 172, 475-82.
PEYRON, F., VUILLEZ, J. P., BARBE, G., BOUDIN, C., PICOT, S. & AMBROISE-THOMAS, P. (1990)
Plasma levels of tumor necrosis factor during a longitudinal survey in an endemic
area of malaria. Acta Trop, 47, 47-51.
PHILIPPS, J., RADLOFF, P. D., WERNSDORFER, W. & KREMSNER, P. G. (1998) Follow-up of the
susceptibility of Plasmodium falciparum to antimalarials in Gabon. Am J Trop Med
Hyg, 58, 612-8.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 150
PICHYANGKUL, S., SAENGKRAI, P. & WEBSTER, H. K. (1994) Plasmodium falciparum pigment
induces monocytes to release high levels of tumor necrosis factor-alpha and
interleukin-1 beta. Am J Trop Med Hyg, 51, 430-5.
PILLAI, D. R., LABBE, A. C., VANISAVETH, V., HONGVANGTHONG, B., POMPHIDA, S.,
INKATHONE, S., ZHONG, K. & KAIN, K. C. (2001) Plasmodium falciparum malaria in
Laos: chloroquine treatment outcome and predictive value of molecular markers. J
Infect Dis, 183, 789-95.
PIZARRO, J. C., VULLIEZ-LE NORMAND, B., CHESNE-SECK, M. L., COLLINS, C. R., WITHERS-
MARTINEZ, C., HACKETT, F., BLACKMAN, M. J., FABER, B. W., REMARQUE, E. J.,
KOCKEN, C. H., THOMAS, A. W. & BENTLEY, G. A. (2005) Crystal structure of the
malaria vaccine candidate apical membrane antigen 1. Science, 308, 408-11.
PLASSMEYER, M. L., REITER, K., SHIMP, R. L., JR., KOTOVA, S., SMITH, P. D., HURT, D. E.,
HOUSE, B., ZOU, X., ZHANG, Y., HICKMAN, M., UCHIME, O., HERRERA, R., NGUYEN, V.,
GLEN, J., LEBOWITZ, J., JIN, A. J., MILLER, L. H., MACDONALD, N. J., WU, Y. & NARUM,
D. L. (2009) Structure of the Plasmodium falciparum circumsporozoite protein, a
leading malaria vaccine candidate. J Biol Chem, 284, 26951-63.
PLEBANSKI, M., FLANAGAN, K. L., LEE, E. A., REECE, W. H., HART, K., GELDER, C., GILLESPIE,
G., PINDER, M. & HILL, A. V. (1999) Interleukin 10-mediated immunosuppression by a
variant CD4 T cell epitope of Plasmodium falciparum. Immunity, 10, 651-60.
PLEBANSKI, M., LEE, E. A., HANNAN, C. M., FLANAGAN, K. L., GILBERT, S. C., GRAVENOR, M.
B. & HILL, A. V. (1999) Altered peptide ligands narrow the repertoire of cellular
immune responses by interfering with T-cell priming. Nat Med, 5, 565-71.
POLLEY, S. D., MWANGI, T., KOCKEN, C. H., THOMAS, A. W., DUTTA, S., LANAR, D. E.,
REMARQUE, E., ROSS, A., WILLIAMS, T. N., MWAMBINGU, G., LOWE, B., CONWAY, D.
J. & MARSH, K. (2004) Human antibodies to recombinant protein constructs of
Plasmodium falciparum Apical Membrane Antigen 1 (AMA1) and their associations
with protection from malaria. Vaccine, 23, 718-28.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 151
POMBO, D. J., LAWRENCE, G., HIRUNPETCHARAT, C., RZEPCZYK, C., BRYDEN, M., CLOONAN,
N., ANDERSON, K., MAHAKUNKIJCHAROEN, Y., MARTIN, L. B., WILSON, D., ELLIOTT,
S., EISEN, D. P., WEINBERG, J. B., SAUL, A. & GOOD, M. F. (2002) Immunity to malaria
after administration of ultra-low doses of red cells infected with Plasmodium
falciparum. Lancet, 360, 610-7.
POVOA, M. M., ADAGU, I. S., OLIVEIRA, S. G., MACHADO, R. L., MILES, M. A. & WARHURST,
D. C. (1998) Pfmdr1 Asn1042Asp and Asp1246Tyr polymorphisms, thought to be
associated with chloroquine resistance, are present in chloroquine-resistant and -
sensitive Brazilian field isolates of Plasmodium falciparum. Exp Parasitol, 88, 64-8.
PRICE, R. N., CASSAR, C., BROCKMAN, A., DURAISINGH, M., VAN VUGT, M., WHITE, N. J.,
NOSTEN, F. & KRISHNA, S. (1999) The pfmdr1 gene is associated with a multidrug-
resistant phenotype in Plasmodium falciparum from the western border of Thailand.
Antimicrob Agents Chemother, 43, 2943-9.
RANDALL, L. M., AMANTE, F. H., MCSWEENEY, K. A., ZHOU, Y., STANLEY, A. C., HAQUE, A.,
JONES, M. K., HILL, G. R., BOYLE, G. M. & ENGWERDA, C. R. (2008) Common strategies
to prevent and modulate experimental cerebral malaria in mouse strains with
different susceptibilities. Infect Immun, 76, 3312-20.
REECE, W. H., PINDER, M., GOTHARD, P. K., MILLIGAN, P., BOJANG, K., DOHERTY, T.,
PLEBANSKI, M., AKINWUNMI, P., EVERAERE, S., WATKINS, K. R., VOSS, G.,
TORNIEPORTH, N., ALLOUECHE, A., GREENWOOD, B. M., KESTER, K. E., MCADAM, K.
P., COHEN, J. & HILL, A. V. (2004) A CD4(+) T-cell immune response to a conserved
epitope in the circumsporozoite protein correlates with protection from natural
Plasmodium falciparum infection and disease. Nat Med, 10, 406-10.
RIHET, P., FLORI, L., TALL, F., TRAORE, A. S. & FUMOUX, F. (2004) Hemoglobin C is associated
with reduced Plasmodium falciparum parasitemia and low risk of mild malaria attack.
Hum Mol Genet, 13, 1-6.
RILEY, E. M., ALLEN, S. J., WHEELER, J. G., BLACKMAN, M. J., BENNETT, S., TAKACS, B.,
SCHONFELD, H. J., HOLDER, A. A. & GREENWOOD, B. M. (1992a) Naturally acquired
cellular and humoral immune responses to the major merozoite surface antigen
(PfMSP1) of Plasmodium falciparum are associated with reduced malaria morbidity.
Parasite Immunol, 14, 321-37.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 152
RILEY, E. M., OLERUP, O., BENNETT, S., ROWE, P., ALLEN, S. J., BLACKMAN, M. J., TROYE-
BLOMBERG, M., HOLDER, A. A. & GREENWOOD, B. M. (1992b) MHC and malaria: the
relationship between HLA class II alleles and immune responses to Plasmodium
falciparum. Int Immunol, 4, 1055-63.
RILEY, E. M., ONG, C. S., OLERUP, O., EIDA, S., ALLEN, S. J., BENNETT, S., ANDERSSON, G. &
TARGETT, G. A. (1990) Cellular and humoral immune responses to Plasmodium
falciparum gametocyte antigens in malaria-immune individuals. Limited response to
the 48/45-kilodalton surface antigen does not appear to be due to MHC restriction. J
Immunol, 144, 4810-6.
ROBERTS, D. J., CRAIG, A. G., BERENDT, A. R., PINCHES, R., NASH, G., MARSH, K. & NEWBOLD,
C. I. (1992) Rapid switching to multiple antigenic and adhesive phenotypes in malaria.
Nature, 357, 689-92.
ROGERSON, S. J., BEESON, J. G., MHANGO, C. G., DZINJALAMALA, F. K. & MOLYNEUX, M. E.
(2000) Plasmodium falciparum rosette formation is uncommon in isolates from
pregnant women. Infect Immun, 68, 391-3.
ROGERSON, S. J., HVIID, L., DUFFY, P. E., LEKE, R. F. & TAYLOR, D. W. (2007) Malaria in
pregnancy: pathogenesis and immunity. Lancet Infect Dis, 7, 105-17.
ROGERSON, S. J., MWAPASA, V. & MESHNICK, S. R. (2007) Malaria in pregnancy: linking
immunity and pathogenesis to prevention. Am J Trop Med Hyg, 77, 14-22.
ROGERSON, S. J., POLLINA, E., GETACHEW, A., TADESSE, E., LEMA, V. M. & MOLYNEUX, M. E.
(2003) Placental monocyte infiltrates in response to Plasmodium falciparum malaria
infection and their association with adverse pregnancy outcomes. Am J Trop Med
Hyg, 68, 115-9.
ROWE, A., OBEIRO, J., NEWBOLD, C. I. & MARSH, K. (1995) Plasmodium falciparum rosetting
is associated with malaria severity in Kenya. Infect Immun, 63, 2323-6.
ROWE, J. A., HANDEL, I. G., THERA, M. A., DEANS, A. M., LYKE, K. E., KONE, A., DIALLO, D. A.,
RAZA, A., KAI, O., MARSH, K., PLOWE, C. V., DOUMBO, O. K. & MOULDS, J. M. (2007)
Blood group O protects against severe Plasmodium falciparum malaria through the
mechanism of reduced rosetting. Proc Natl Acad Sci U S A, 104, 17471-6.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 153
ROWE, J. A., MOULDS, J. M., NEWBOLD, C. I. & MILLER, L. H. (1997) P. falciparum rosetting
mediated by a parasite-variant erythrocyte membrane protein and complement-
receptor 1. Nature, 388, 292-5.
RUWENDE, C., KHOO, S. C., SNOW, R. W., YATES, S. N., KWIATKOWSKI, D., GUPTA, S., WARN,
P., ALLSOPP, C. E., GILBERT, S. C., PESCHU, N. & ET AL. (1995) Natural selection of
hemi- and heterozygotes for G6PD deficiency in Africa by resistance to severe
malaria. Nature, 376, 246-9.
SABCHAREON, A., BURNOUF, T., OUATTARA, D., ATTANATH, P., BOUHAROUN-TAYOUN, H.,
CHANTAVANICH, P., FOUCAULT, C., CHONGSUPHAJAISIDDHI, T. & DRUILHE, P. (1991)
Parasitologic and clinical human response to immunoglobulin administration in
falciparum malaria. Am J Trop Med Hyg, 45, 297-308.
SACARLAL, J., AIDE, P., APONTE, J. J., RENOM, M., LEACH, A., MANDOMANDO, I., LIEVENS,
M., BASSAT, Q., LAFUENTE, S., MACETE, E., VEKEMANS, J., GUINOVART, C.,
SIGAUQUE, B., SILLMAN, M., MILMAN, J., DUBOIS, M. C., DEMOITIE, M. A.,
THONNARD, J., MENENDEZ, C., BALLOU, W. R., COHEN, J. & ALONSO, P. L. (2009)
Long-term safety and efficacy of the RTS,S/AS02A malaria vaccine in Mozambican
children. J Infect Dis, 200, 329-36.
SAGARA, I., DICKO, A., ELLIS, R. D., FAY, M. P., DIAWARA, S. I., ASSADOU, M. H., SISSOKO, M.
S., KONE, M., DIALLO, A. I., SAYE, R., GUINDO, M. A., KANTE, O., NIAMBELE, M. B.,
MIURA, K., MULLEN, G. E., PIERCE, M., MARTIN, L. B., DOLO, A., DIALLO, D. A.,
DOUMBO, O. K., MILLER, L. H. & SAUL, A. (2009) A randomized controlled phase 2
trial of the blood stage AMA1-C1/Alhydrogel malaria vaccine in children in Mali.
Vaccine, 27, 3090-8.
SAGARA, I., ELLIS, R. D., DICKO, A., NIAMBELE, M. B., KAMATE, B., GUINDO, O., SISSOKO, M.
S., FAY, M. P., GUINDO, M. A., KANTE, O., SAYE, R., MIURA, K., LONG, C., MULLEN, G.
E., PIERCE, M., MARTIN, L. B., RAUSCH, K., DOLO, A., DIALLO, D. A., MILLER, L. H. &
DOUMBO, O. K. (2009) A randomized and controlled Phase 1 study of the safety and
immunogenicity of the AMA1-C1/Alhydrogel + CPG 7909 vaccine for Plasmodium
falciparum malaria in semi-immune Malian adults. Vaccine, 27, 7292-8.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 154
SAKAGUCHI, S., SAKAGUCHI, N., ASANO, M., ITOH, M. & TODA, M. (1995) Immunologic self-
tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains
(CD25). Breakdown of a single mechanism of self-tolerance causes various
autoimmune diseases. J Immunol, 155, 1151-64.
SALANTI, A., DAHLBACK, M., TURNER, L., NIELSEN, M. A., BARFOD, L., MAGISTRADO, P.,
JENSEN, A. T., LAVSTSEN, T., OFORI, M. F., MARSH, K., HVIID, L. & THEANDER, T. G.
(2004) Evidence for the involvement of VAR2CSA in pregnancy-associated malaria. J
Exp Med, 200, 1197-203.
SALANTI, A., STAALSOE, T., LAVSTSEN, T., JENSEN, A. T., SOWA, M. P., ARNOT, D. E., HVIID, L.
& THEANDER, T. G. (2003) Selective upregulation of a single distinctly structured var
gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in
pregnancy-associated malaria. Mol Microbiol, 49, 179-91.
SALEM, H. H., MARUYAMA, I., ISHII, H. & MAJERUS, P. W. (1984) Isolation and
characterization of thrombomodulin from human placenta. J Biol Chem, 259, 12246-
51.
SALOMON, B., LENSCHOW, D. J., RHEE, L., ASHOURIAN, N., SINGH, B., SHARPE, A. &
BLUESTONE, J. A. (2000) B7/CD28 costimulation is essential for the homeostasis of
the CD4+CD25+ immunoregulatory T cells that control autoimmune diabetes.
Immunity, 12, 431-40.
SCHELLER, L. F. & AZAD, A. F. (1995) Maintenance of protective immunity against malaria by
persistent hepatic parasites derived from irradiated sporozoites. Proc Natl Acad Sci U
S A, 92, 4066-8.
SCHERF, A., LOPEZ-RUBIO, J. J. & RIVIERE, L. (2008) Antigenic variation in Plasmodium
falciparum. Annu Rev Microbiol, 62, 445-70.
SCHERF, A., POUVELLE, B., BUFFET, P. A. & GYSIN, J. (2001) Molecular mechanisms of
Plasmodium falciparum placental adhesion. Cell Microbiol, 3, 125-31.
SCHOFIELD, L. & GRAU, G. E. (2005) Immunological processes in malaria pathogenesis. Nat
Rev Immunol, 5, 722-35.
SCHOFIELD, L. & HACKETT, F. (1993) Signal transduction in host cells by a
glycosylphosphatidylinositol toxin of malaria parasites. J Exp Med, 177, 145-53.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 155
SCHOFIELD, L., NOVAKOVIC, S., GEROLD, P., SCHWARZ, R. T., MCCONVILLE, M. J. &
TACHADO, S. D. (1996) Glycosylphosphatidylinositol toxin of Plasmodium up-
regulates intercellular adhesion molecule-1, vascular cell adhesion molecule-1, and E-
selectin expression in vascular endothelial cells and increases leukocyte and parasite
cytoadherence via tyrosine kinase-dependent signal transduction. J Immunol, 156,
1886-96.
SEIXAS, E., CROSS, C., QUIN, S. & LANGHORNE, J. (2001) Direct activation of dendritic cells by
the malaria parasite, Plasmodium chabaudi chabaudi. Eur J Immunol, 31, 2970-8.
SHIMIZU, J., YAMAZAKI, S. & SAKAGUCHI, S. (1999) Induction of tumor immunity by
removing CD25+CD4+ T cells: a common basis between tumor immunity and
autoimmunity. J Immunol, 163, 5211-8.
SILVIE, O., FRANETICH, J. F., CHARRIN, S., MUELLER, M. S., SIAU, A., BODESCOT, M.,
RUBINSTEIN, E., HANNOUN, L., CHAROENVIT, Y., KOCKEN, C. H., THOMAS, A. W., VAN
GEMERT, G. J., SAUERWEIN, R. W., BLACKMAN, M. J., ANDERS, R. F., PLUSCHKE, G. &
MAZIER, D. (2004) A role for apical membrane antigen 1 during invasion of
hepatocytes by Plasmodium falciparum sporozoites. J Biol Chem, 279, 9490-6.
SINGH, B., KIM SUNG, L., MATUSOP, A., RADHAKRISHNAN, A., SHAMSUL, S. S., COX-SINGH, J.,
THOMAS, A. & CONWAY, D. J. (2004) A large focus of naturally acquired Plasmodium
knowlesi infections in human beings. Lancet, 363, 1017-24.
SINIGAGLIA, F., GUTTINGER, M., KILGUS, J., DORAN, D. M., MATILE, H., ETLINGER, H.,
TRZECIAK, A., GILLESSEN, D. & PINK, J. R. (1988) A malaria T-cell epitope recognized in
association with most mouse and human MHC class II molecules. Nature, 336, 778-
80.
SIRIMA, S. B., TIONO, A. B., OUEDRAOGO, A., DIARRA, A., OUEDRAOGO, A. L., YARO, J. B.,
OUEDRAOGO, E., GANSANE, A., BOUGOUMA, E. C., KONATE, A. T., KABORE, Y.,
TRAORE, A., CHILENGI, R., SOULAMA, I., LUTY, A. J., DRUILHE, P., COUSENS, S. &
NEBIE, I. (2009) Safety and immunogenicity of the malaria vaccine candidate MSP3
long synthetic peptide in 12-24 months-old Burkinabe children. PLoS One, 4, e7549.
SJOBERG, K., LEPERS, J. P., RAHARIMALALA, L., LARSSON, A., OLERUP, O., MARBIAH, N. T.,
TROYE-BLOMBERG, M. & PERLMANN, P. (1992) Genetic regulation of human anti-
malarial antibodies in twins. Proc Natl Acad Sci U S A, 89, 2101-4.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 156
SKOROKHOD, O. A., ALESSIO, M., MORDMULLER, B., ARESE, P. & SCHWARZER, E. (2004)
Hemozoin (malarial pigment) inhibits differentiation and maturation of human
monocyte-derived dendritic cells: a peroxisome proliferator-activated receptor-
gamma-mediated effect. J Immunol, 173, 4066-74.
SNAPPER, C. M., WAEGELL, W., BEERNINK, H. & DASCH, J. R. (1993) Transforming growth
factor-beta 1 is required for secretion of IgG of all subclasses by LPS-activated murine
B cells in vitro. J Immunol, 151, 4625-36.
SNOUNOU, G., JARRA, W. & PREISER, P. R. (2000) Malaria multigene families: the price of
chronicity. Parasitol Today, 16, 28-30.
SNOW, R. W., GUERRA, C. A., NOOR, A. M., MYINT, H. Y. & HAY, S. I. (2005) The global
distribution of clinical episodes of Plasmodium falciparum malaria. Nature, 434, 214-
7.
SNOW, R. W., OMUMBO, J. A., LOWE, B., MOLYNEUX, C. S., OBIERO, J. O., PALMER, A.,
WEBER, M. W., PINDER, M., NAHLEN, B., OBONYO, C., NEWBOLD, C., GUPTA, S. &
MARSH, K. (1997) Relation between severe malaria morbidity in children and level of
Plasmodium falciparum transmission in Africa. Lancet, 349, 1650-4.
SPENCER VALERO, L. M., OGUN, S. A., FLECK, S. L., LING, I. T., SCOTT-FINNIGAN, T. J.,
BLACKMAN, M. J. & HOLDER, A. A. (1998) Passive immunization with antibodies
against three distinct epitopes on Plasmodium yoelii merozoite surface protein 1
suppresses parasitemia. Infect Immun, 66, 3925-30.
STAALSOE, T., SHULMAN, C. E., BULMER, J. N., KAWUONDO, K., MARSH, K. & HVIID, L. (2004)
Variant surface antigen-specific IgG and protection against clinical consequences of
pregnancy-associated Plasmodium falciparum malaria. Lancet, 363, 283-9.
STAVNEZER, J. (1995) Regulation of antibody production and class switching by TGF-beta. J
Immunol, 155, 1647-51.
STEKETEE, R. W., NAHLEN, B. L., PARISE, M. E. & MENENDEZ, C. (2001) The burden of malaria
in pregnancy in malaria-endemic areas. Am J Trop Med Hyg, 64, 28-35.
STEVENSON, M. M., GHADIRIAN, E., PHILLIPS, N. C., RAE, D. & PODOBA, J. E. (1989) Role of
mononuclear phagocytes in elimination of Plasmodium chabaudi AS infection.
Parasite Immunol, 11, 529-44.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 157
STEVENSON, M. M., TAM, M. F., WOLF, S. F. & SHER, A. (1995) IL-12-induced protection
against blood-stage Plasmodium chabaudi AS requires IFN-gamma and TNF-alpha and
occurs via a nitric oxide-dependent mechanism. J Immunol, 155, 2545-56.
STOUTE, J. A., SLAOUI, M., HEPPNER, D. G., MOMIN, P., KESTER, K. E., DESMONS, P., WELLDE,
B. T., GARCON, N., KRZYCH, U. & MARCHAND, M. (1997) A preliminary evaluation of a
recombinant circumsporozoite protein vaccine against Plasmodium falciparum
malaria. RTS,S Malaria Vaccine Evaluation Group. N Engl J Med, 336, 86-91.
SU, X. Z., HEATWOLE, V. M., WERTHEIMER, S. P., GUINET, F., HERRFELDT, J. A., PETERSON, D.
S., RAVETCH, J. A. & WELLEMS, T. E. (1995) The large diverse gene family var encodes
proteins involved in cytoadherence and antigenic variation of Plasmodium
falciparum-infected erythrocytes. Cell, 82, 89-100.
TACHADO, S. D., GEROLD, P., MCCONVILLE, M. J., BALDWIN, T., QUILICI, D., SCHWARZ, R. T.
& SCHOFIELD, L. (1996) Glycosylphosphatidylinositol toxin of Plasmodium induces
nitric oxide synthase expression in macrophages and vascular endothelial cells by a
protein tyrosine kinase-dependent and protein kinase C-dependent signaling
pathway. J Immunol, 156, 1897-1907.
TAKALA, S. L. & PLOWE, C. V. (2009) Genetic diversity and malaria vaccine design, testing and
efficacy: preventing and overcoming 'vaccine resistant malaria'. Parasite Immunol,
31, 560-73.
TANABE, K., MACKAY, M., GOMAN, M. & SCAIFE, J. G. (1987) Allelic dimorphism in a surface
antigen gene of the malaria parasite Plasmodium falciparum. J Mol Biol, 195, 273-87.
TAYLOR, T. E., BORGSTEIN, A. & MOLYNEUX, M. E. (1993) Acid-base status in paediatric
Plasmodium falciparum malaria. Q J Med, 86, 99-109.
TAYLOR-ROBINSON, A. W., PHILLIPS, R. S., SEVERN, A., MONCADA, S. & LIEW, F. Y. (1993) The
role of TH1 and TH2 cells in a rodent malaria infection. Science, 260, 1931-4.
THORNTON, A. M. & SHEVACH, E. M. (1998) CD4+CD25+ immunoregulatory T cells suppress
polyclonal T cell activation in vitro by inhibiting interleukin 2 production. J Exp Med,
188, 287-96.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 158
TODRYK, S. M., BEJON, P., MWANGI, T., PLEBANSKI, M., URBAN, B., MARSH, K., HILL, A. V. &
FLANAGAN, K. L. (2008) Correlation of memory T cell responses against TRAP with
protection from clinical malaria, and CD4 CD25 high T cells with susceptibility in
Kenyans. PLoS One, 3, e2027.
TORCIA, M. G., SANTARLASCI, V., COSMI, L., CLEMENTE, A., MAGGI, L., MANGANO, V. D.,
VERRA, F., BANCONE, G., NEBIE, I., SIRIMA, B. S., LIOTTA, F., FROSALI, F., ANGELI, R.,
SEVERINI, C., SANNELLA, A. R., BONINI, P., LUCIBELLO, M., MAGGI, E., GARACI, E.,
COLUZZI, M., COZZOLINO, F., ANNUNZIATO, F., ROMAGNANI, S. & MODIANO, D.
(2008) Functional deficit of T regulatory cells in Fulani, an ethnic group with low
susceptibility to Plasmodium falciparum malaria. Proc Natl Acad Sci U S A, 105, 646-
51.
TRINCHIERI, G. (1995) Interleukin-12: a proinflammatory cytokine with immunoregulatory
functions that bridge innate resistance and antigen-specific adaptive immunity. Annu
Rev Immunol, 13, 251-76.
TRIPP, C. S., WOLF, S. F. & UNANUE, E. R. (1993) Interleukin 12 and tumor necrosis factor
alpha are costimulators of interferon gamma production by natural killer cells in
severe combined immunodeficiency mice with listeriosis, and interleukin 10 is a
physiologic antagonist. Proc Natl Acad Sci U S A, 90, 3725-9.
TROYE-BLOMBERG, M., RILEY, E. M., KABILAN, L., HOLMBERG, M., PERLMANN, H.,
ANDERSSON, U., HEUSSER, C. H. & PERLMANN, P. (1990) Production by activated
human T cells of interleukin 4 but not interferon-gamma is associated with elevated
levels of serum antibodies to activating malaria antigens. Proc Natl Acad Sci U S A, 87,
5484-8.
TROYE-BLOMBERG, M., RILEY, E. M., PERLMANN, H., ANDERSSON, G., LARSSON, A., SNOW,
R. W., ALLEN, S. J., HOUGHTEN, R. A., OLERUP, O., GREENWOOD, B. M. & ET AL.
(1989) T and B cell responses of Plasmodium falciparum malaria-immune individuals
to synthetic peptides corresponding to sequences in different regions of the P.
falciparum antigen Pf155/RESA. J Immunol, 143, 3043-8.
TSUJI, M. & ZAVALA, F. (2003) T cells as mediators of protective immunity against liver stages
of Plasmodium. Trends Parasitol, 19, 88-93.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 159
TSUNAWAKI, S., SPORN, M., DING, A. & NATHAN, C. (1988) Deactivation of macrophages by
transforming growth factor-beta. Nature, 334, 260-2.
TUIKUE NDAM, N. G., SALANTI, A., LE-HESRAN, J. Y., COTTRELL, G., FIEVET, N., TURNER, L.,
SOW, S., DANGOU, J. M., THEANDER, T. & DELORON, P. (2006) Dynamics of anti-
VAR2CSA immunoglobulin G response in a cohort of senegalese pregnant women. J
Infect Dis, 193, 713-20.
TURNER, G. D., MORRISON, H., JONES, M., DAVIS, T. M., LOOAREESUWAN, S., BULEY, I. D.,
GATTER, K. C., NEWBOLD, C. I., PUKRITAYAKAMEE, S., NAGACHINTA, B. & ET AL.
(1994) An immunohistochemical study of the pathology of fatal malaria. Evidence for
widespread endothelial activation and a potential role for intercellular adhesion
molecule-1 in cerebral sequestration. Am J Pathol, 145, 1057-69.
UDHAYAKUMAR, V., KARIUKI, S., KOLCZACK, M., GIRMA, M., ROBERTS, J. M., OLOO, A. J.,
NAHLEN, B. L. & LAL, A. A. (2001) Longitudinal study of natural immune responses to
the Plasmodium falciparum apical membrane antigen (AMA-1) in a holoendemic
region of malaria in western Kenya: Asembo Bay Cohort Project VIII. Am J Trop Med
Hyg, 65, 100-7.
UDOMSANGPETCH, R., SUEBLINVONG, T., PATTANAPANYASAT, K., DHARMKRONG-AT, A.,
KITTIKALAYAWONG, A. & WEBSTER, H. K. (1993) Alteration in cytoadherence and
rosetting of Plasmodium falciparum-infected thalassemic red blood cells. Blood, 82,
3752-9.
URBAN, B. C., FERGUSON, D. J., PAIN, A., WILLCOX, N., PLEBANSKI, M., AUSTYN, J. M. &
ROBERTS, D. J. (1999) Plasmodium falciparum-infected erythrocytes modulate the
maturation of dendritic cells. Nature, 400, 73-7.
URBAN, B. C., MWANGI, T., ROSS, A., KINYANJUI, S., MOSOBO, M., KAI, O., LOWE, B.,
MARSH, K. & ROBERTS, D. J. (2001) Peripheral blood dendritic cells in children with
acute Plasmodium falciparum malaria. Blood, 98, 2859-61.
UTHAIPIBULL, C., AUFIERO, B., SYED, S. E., HANSEN, B., GUEVARA PATINO, J. A., ANGOV, E.,
LING, I. T., FEGEDING, K., MORGAN, W. D., OCKENHOUSE, C., BIRDSALL, B., FEENEY,
J., LYON, J. A. & HOLDER, A. A. (2001) Inhibitory and blocking monoclonal antibody
epitopes on merozoite surface protein 1 of the malaria parasite Plasmodium
falciparum. J Mol Biol, 307, 1381-94.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 160
VAN DER HEYDE, H. C., HUSZAR, D., WOODHOUSE, C., MANNING, D. D. & WEIDANZ, W. P.
(1994) The resolution of acute malaria in a definitive model of B cell deficiency, the
JHD mouse. J Immunol, 152, 4557-62.
VIEIRA, P. P., DAS GRACAS ALECRIM, M., DA SILVA, L. H., GONZALEZ-JIMENEZ, I. & ZALIS, M.
G. (2001) Analysis of the PfCRT K76T mutation in Plasmodium falciparum isolates
from the Amazon region of Brazil. J Infect Dis, 183, 1832-3.
VIGARIO, A. M., GORGETTE, O., DUJARDIN, H. C., CRUZ, T., CAZENAVE, P. A., SIX, A.,
BANDEIRA, A. & PIED, S. (2007) Regulatory CD4+ CD25+ Foxp3+ T cells expand during
experimental Plasmodium infection but do not prevent cerebral malaria. Int J
Parasitol, 37, 963-73.
VIGNALI, D. A., COLLISON, L. W. & WORKMAN, C. J. (2008) How regulatory T cells work. Nat
Rev Immunol, 8, 523-32.
VON DER WEID, T., HONARVAR, N. & LANGHORNE, J. (1996) Gene-targeted mice lacking B
cells are unable to eliminate a blood stage malaria infection. J Immunol, 156, 2510-6.
VON SEIDLEIN, L., DURAISINGH, M. T., DRAKELEY, C. J., BAILEY, R., GREENWOOD, B. M. &
PINDER, M. (1997) Polymorphism of the Pfmdr1 gene and chloroquine resistance in
Plasmodium falciparum in The Gambia. Trans R Soc Trop Med Hyg, 91, 450-3.
WAITUMBI, J. N., OPOLLO, M. O., MUGA, R. O., MISORE, A. O. & STOUTE, J. A. (2000) Red cell
surface changes and erythrophagocytosis in children with severe plasmodium
falciparum anemia. Blood, 95, 1481-6.
WALTHER, M., JEFFRIES, D., FINNEY, O. C., NJIE, M., EBONYI, A., DEININGER, S., LAWRENCE,
E., NGWA-AMAMBUA, A., JAYASOORIYA, S., CHEESEMAN, I. H., GOMEZ-ESCOBAR, N.,
OKEBE, J., CONWAY, D. J. & RILEY, E. M. (2009) Distinct roles for FOXP3 and FOXP3
CD4 T cells in regulating cellular immunity to uncomplicated and severe Plasmodium
falciparum malaria. PLoS Pathog, 5, e1000364.
WALTHER, M., TONGREN, J. E., ANDREWS, L., KORBEL, D., KING, E., FLETCHER, H.,
ANDERSEN, R. F., BEJON, P., THOMPSON, F., DUNACHIE, S. J., EDELE, F., DE SOUZA, J.
B., SINDEN, R. E., GILBERT, S. C., RILEY, E. M. & HILL, A. V. (2005) Upregulation of TGF-
beta, FOXP3, and CD4+CD25+ regulatory T cells correlates with more rapid parasite
growth in human malaria infection. Immunity, 23, 287-96.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 161
WALTHER, M., WOODRUFF, J., EDELE, F., JEFFRIES, D., TONGREN, J. E., KING, E., ANDREWS,
L., BEJON, P., GILBERT, S. C., DE SOUZA, J. B., SINDEN, R., HILL, A. V. & RILEY, E. M.
(2006) Innate immune responses to human malaria: heterogeneous cytokine
responses to blood-stage Plasmodium falciparum correlate with parasitological and
clinical outcomes. J Immunol, 177, 5736-45.
WEEDALL, G. D. & CONWAY, D. J. (2010) Detecting signatures of balancing selection to
identify targets of anti-parasite immunity. Trends Parasitol, 26, 363-9.
WEGMANN, T. G., LIN, H., GUILBERT, L. & MOSMANN, T. R. (1993) Bidirectional cytokine
interactions in the maternal-fetal relationship: is successful pregnancy a TH2
phenomenon? Immunol Today, 14, 353-6.
WEISS, G. E., TRAORE, B., KAYENTAO, K., ONGOIBA, A., DOUMBO, S., DOUMTABE, D., KONE,
Y., DIA, S., GUINDO, A., TRAORE, A., HUANG, C. Y., MIURA, K., MIRCETIC, M., LI, S.,
BAUGHMAN, A., NARUM, D. L., MILLER, L. H., DOUMBO, O. K., PIERCE, S. K. &
CROMPTON, P. D. (2010) The Plasmodium falciparum-specific human memory B cell
compartment expands gradually with repeated malaria infections. PLoS Pathog, 6,
e1000912.
WEISS, W. R., SEDEGAH, M., BERZOFSKY, J. A. & HOFFMAN, S. L. (1993) The role of CD4+ T
cells in immunity to malaria sporozoites. J Immunol, 151, 2690-8.
WELLEMS, T. E., WALKER-JONAH, A. & PANTON, L. J. (1991) Genetic mapping of the
chloroquine-resistance locus on Plasmodium falciparum chromosome 7. Proc Natl
Acad Sci U S A, 88, 3382-6.
WENISCH, C., PARSCHALK, B., BURGMANN, H., LOOAREESUWAN, S. & GRANINGER, W.
(1995) Decreased serum levels of TGF-beta in patients with acute Plasmodium
falciparum malaria. J Clin Immunol, 15, 69-73.
WENISCH, C., PARSCHALK, B., NARZT, E., LOOAREESUWAN, S. & GRANINGER, W. (1995)
Elevated serum levels of IL-10 and IFN-gamma in patients with acute Plasmodium
falciparum malaria. Clin Immunol Immunopathol, 74, 115-7.
WERNSDORFER, W. H. (1994) Epidemiology of drug resistance in malaria. Acta Trop, 56, 143-
56.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 162
WILLIAMS, T. N., MWANGI, T. W., ROBERTS, D. J., ALEXANDER, N. D., WEATHERALL, D. J.,
WAMBUA, S., KORTOK, M., SNOW, R. W. & MARSH, K. (2005) An immune basis for
malaria protection by the sickle cell trait. PLoS Med, 2, e128.
WILSON, C. M., VOLKMAN, S. K., THAITHONG, S., MARTIN, R. K., KYLE, D. E., MILHOUS, W. K.
& WIRTH, D. F. (1993) Amplification of pfmdr 1 associated with mefloquine and
halofantrine resistance in Plasmodium falciparum from Thailand. Mol Biochem
Parasitol, 57, 151-60.
WINKLER, S., WILLHEIM, M., BAIER, K., SCHMID, D., AICHELBURG, A., GRANINGER, W. &
KREMSNER, P. G. (1999) Frequency of cytokine-producing T cells in patients of
different age groups with Plasmodium falciparum malaria. J Infect Dis, 179, 209-16.
WIPASA, J., SUPHAVILAI, C., OKELL, L. C., COOK, J., CORRAN, P. H., THAIKLA, K., LIEWSAREE,
W., RILEY, E. M. & HAFALLA, J. C. (2010) Long-lived antibody and B Cell memory
responses to the human malaria parasites, Plasmodium falciparum and Plasmodium
vivax. PLoS Pathog, 6, e1000770.
WITKOWSKI, B., IRIART, X., SOH, P. N., MENARD, S., ALVAREZ, M., NANEIX-LAROCHE, V.,
MARCHOU, B., MAGNAVAL, J. F., BENOIT-VICAL, F. & BERRY, A. (2010) pfmdr1
amplification associated with clinical resistance to mefloquine in West Africa:
implications for efficacy of artemisinin combination therapies. J Clin Microbiol, 48,
3797-9.
WONGSRICHANALAI, C., SIRICHAISINTHOP, J., KARWACKI, J. J., CONGPUONG, K., MILLER, R.
S., PANG, L. & THIMASARN, K. (2001) Drug resistant malaria on the Thai-Myanmar
and Thai-Cambodian borders. Southeast Asian J Trop Med Public Health, 32, 41-9.
YODER, B. J. & GOODRUM, K. J. (2001) Plasmodium chabaudi chabaudi: B-1 cell expansion
correlates with semiresistance in BALB/cJ mice. Exp Parasitol, 98, 71-82.
YOUNG, K., FRODSHAM, A., DOUMBO, O. K., GUPTA, S., DOLO, A., HU, J. T., ROBSON, K. J.,
CRISANTI, A., HILL, A. V. & GILBERT, S. C. (2005) Inverse associations of human
leukocyte antigen and malaria parasite types in two West African populations. Infect
Immun, 73, 953-5.
ZALIS, M. G., PANG, L., SILVEIRA, M. S., MILHOUS, W. K. & WIRTH, D. F. (1998)
Characterization of Plasmodium falciparum isolated from the Amazon region of
Brazil: evidence for quinine resistance. Am J Trop Med Hyg, 58, 630-7.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 163
Annexes

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 164
Liste des publications de cette thèse
Bisseye C, van der Sande M, Morgan WD, Holder AA, Pinder M, Ismaili J: Plasmodium
falciparum infection of the placenta impacts on the T helper type 1 (Th1)/Th2 balance of neonatal
T cells through CD4 (+) CD25 (+) forkhead box P3 (+) regulatory T cells and interleukin-10. Clin
Exp Immunol. 2009; 158: 287-93.
Cyrille Bisseye, Louis M. Yindom, Jacques Simporé, William D. Morgan, Anthony A. Holder and
Jamila Ismaili. An engineered Plasmodium falciparum C-terminal 19-kilodalton merozoite surface
protein 1 vaccine candidate induces high levels of interferon-gamma production associated with
cellular immune responses to specific peptide sequences in Gambian adults naturally exposed to
malaria. Clin Exp Immunolo. 2011; 166 (3):366-73
Jamila Ismaili, Cyrille Bisseye, Dongmei liu, Hanif Ismael, Musa Jawara, Mathurin Diatta,
Giorgio Sirugo, Margaret Pinder, Sam dunyo, William D. Morgan, Michael J. Blackman, Anthony
A. Holder, Jacques Simpore and Paul Milligan. Comparison of the humoral responses to CSP,
AMA1 and MSP119 and susceptibility to malaria in Fulani, and the sympatric Wolof and
Mandinka groups in The Gambia (Manuscrit soumis pour publication)
Articles sur le paludisme non compris dans la présente thèse
Marek Cyrklaff, Cecilia P. Sanchez, Nicole Kilian, Cyrille Bisseye, Jacques Simpore, Friedrich
Frischknecht, Michael Lanzer. Hemoglobins S and C Interfere with Actin Remodeling in
Plasmodium falciparum–Infected Erythrocytes. Science 2011; 334 (6060): 1283-1286.
Dunyo S, Sirugo G, Sesay S, Bisseye C, Njie F, Adiamoh M, Nwakanma D, Diatta M, Janha R,
Sisay Joof F, Temple B, Snell P, Conway D, Walton R, Cheung YB, Milligan P. Randomized trial
of safety and effectiveness of chlorproguanil-dapsone and lumefantrine-artemether for
uncomplicated malaria in children in the Gambia. PLoSOne. 2011; 6(6):e17371.
Touré FS, Bisseye C, Mavoungou E. Imbalanced distribution of Plasmodium falciparum EBA-175
genotypes related to clinical status in children from Bakoumba, Gabon. Clin Med Res. 2006 Mar;
4(1):7-11.
Mombo LE, Ntoumi F, Bisseye C, Ossari S, Lu CY, Nagel RL, Krishnamoorthy R. Human genetic
polymorphisms and asymptomatic Plasmodium falciparum malaria in Gabonese schoolchildren.
Am J Trop Med Hyg. 2003 Feb; 68(2):186-90.

Thèse de Doctorat Unique 2011 Cyrille BISSEYE Page 165
Migot-Nabias F, Mombo LE, Luty AJ, Dubois B, Nabias R, Bisseye C, Millet P, Lu CY, Deloron P.
Human genetic factors related to susceptibility to mild malaria in Gabon. Genes Immun. 2000
Oct; 1(7):435-41.
Autres publications significatives en 2011
Morris GA, Edwards DR, Hill PC, Wejse C, Bisseye C, Olesen R, Edwards TL, Gilbert JR, Myers
JL, Stryjewski ME, Abbate E, Estevan R, Hamilton CD, Tacconelli A, Novelli G, Brunetti E,
Aaby P, Sodemann M, Østergaard L, Adegbola R, Williams SM, Scott WK, Sirugo G. Interleukin
12B (IL12B) genetic variation and pulmonary tuberculosis: a study of cohorts from The Gambia,
Guinea-Bissau, United States and Argentina. PLoS One. 2011 Feb 9; 6(2):e16656. Erratum in:
PLoS One. 2011;6(2). doi:
Nagalo MB, Sanou M, Bisseye C, Kaboré MI, Nebie YK, Kienou K, Kiba A, Dahourou H, Ouattara
S, Zongo JD, Simporé J. Seroprevalence of human immunodeficiency virus, hepatitis B and C
viruses and syphilis among blood donors in Koudougou (Burkina Faso) in 2009. Blood Transfus.
2011 Jul 18;4(1):1-6. doi: 10.2450/2011.0112-10.
Nagalo BM, Bisseye C, Sanou M, Nebié YK, Kiba A, Kienou K, Zongo JD, Simporé J. [Molecular
diagnosis of acquired human immunodeficiency virus (HIV) in pooled plasma from blood donors
at the Regional Blood Transfusion Center in Ouagadougou, Burkina Faso]. Med Trop (Mars).
2011 Apr; 71(2):137-41.
Hennig BJ, Velez-Edwards DR, Schim van der Loeff MF, Bisseye C, Edwards TL, Tacconelli A,
Novelli G, Aaby P, Kaye S, Scott WK, Jaye A, Whittle HC, Williams SM, Hill AV, Sirugo G.
CD4 intragenic SNPs associate with HIV-2 plasma viral load and CD4 count in a community-
based study from Guinea-Bissau, West Africa. J Acquir Immune Defic Syndr. 2011 Jan; 56(1):1-
8.
8

166

167
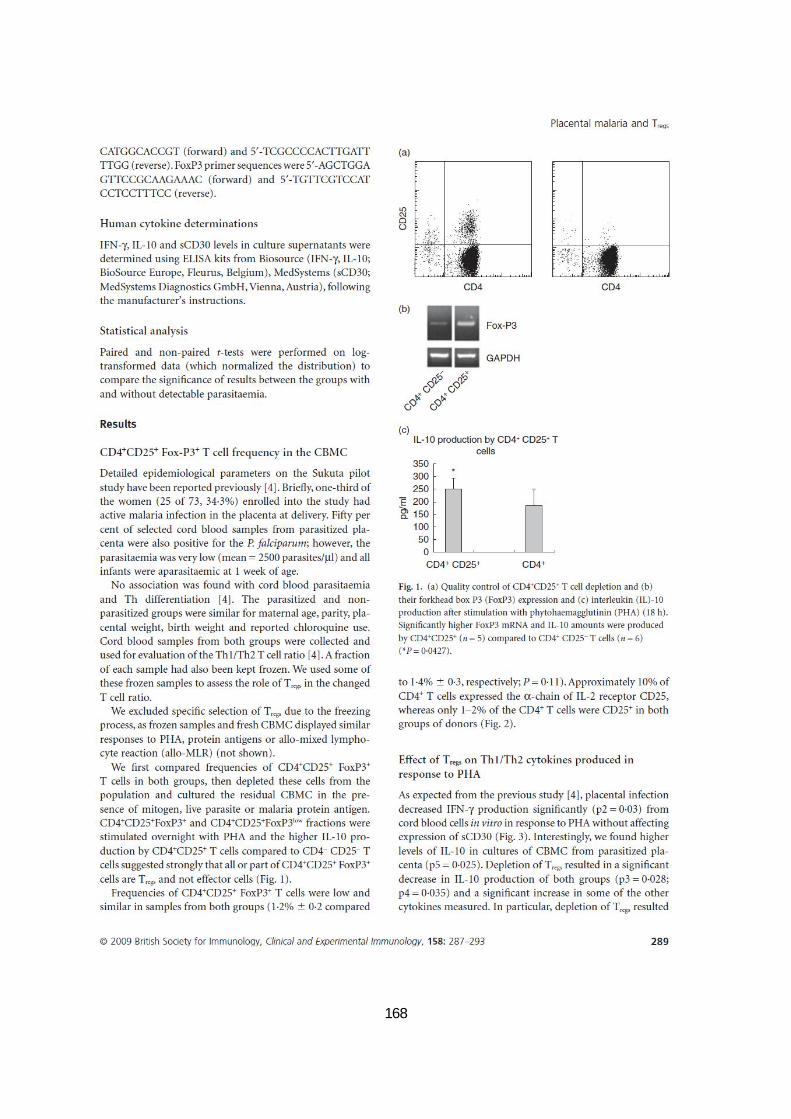
168
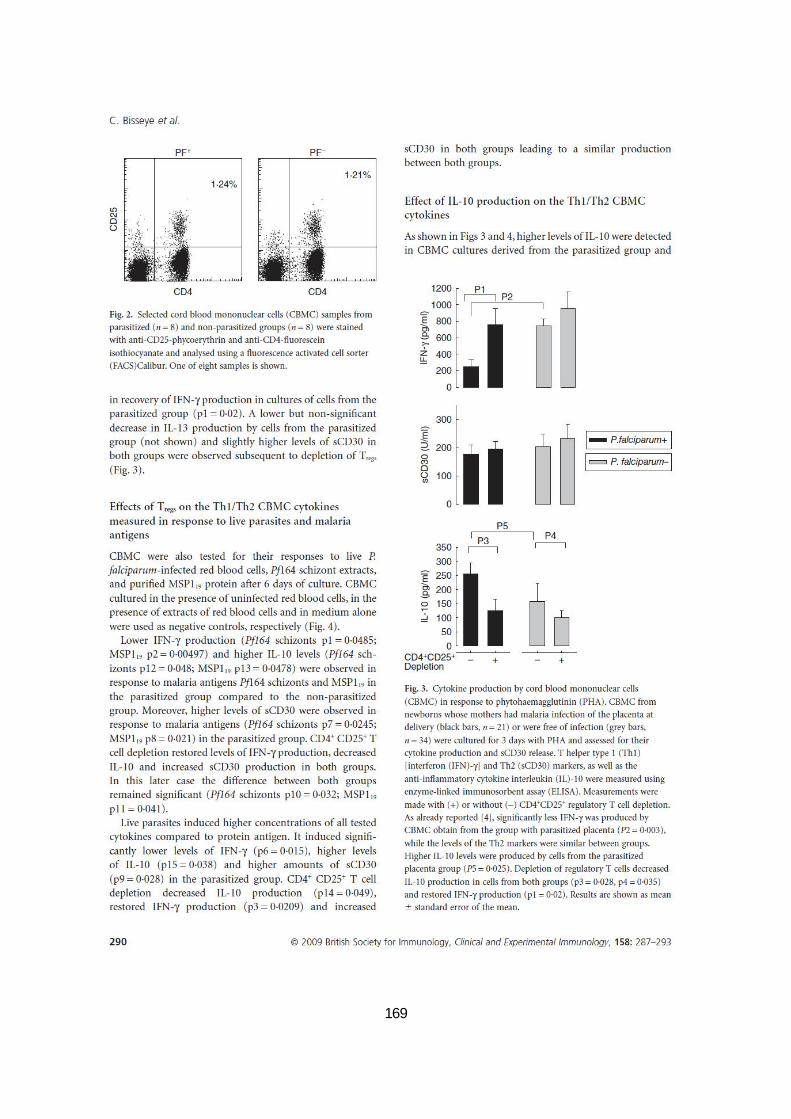
169

170
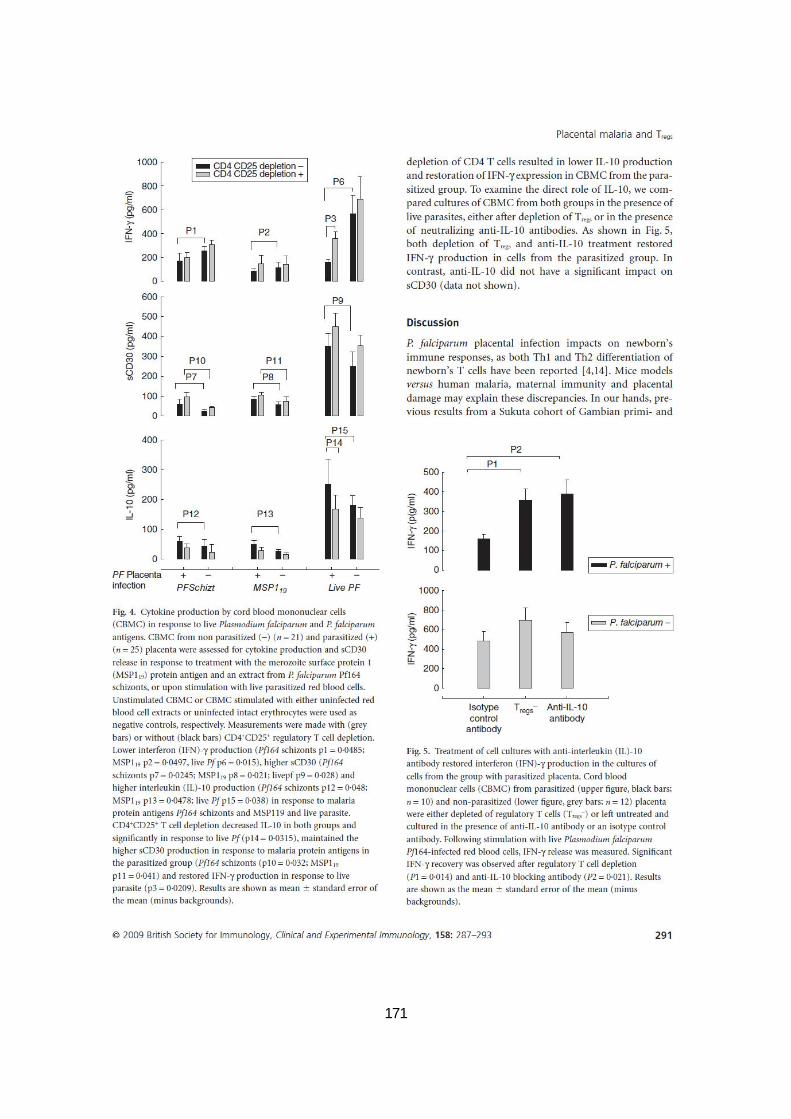
171

172

173
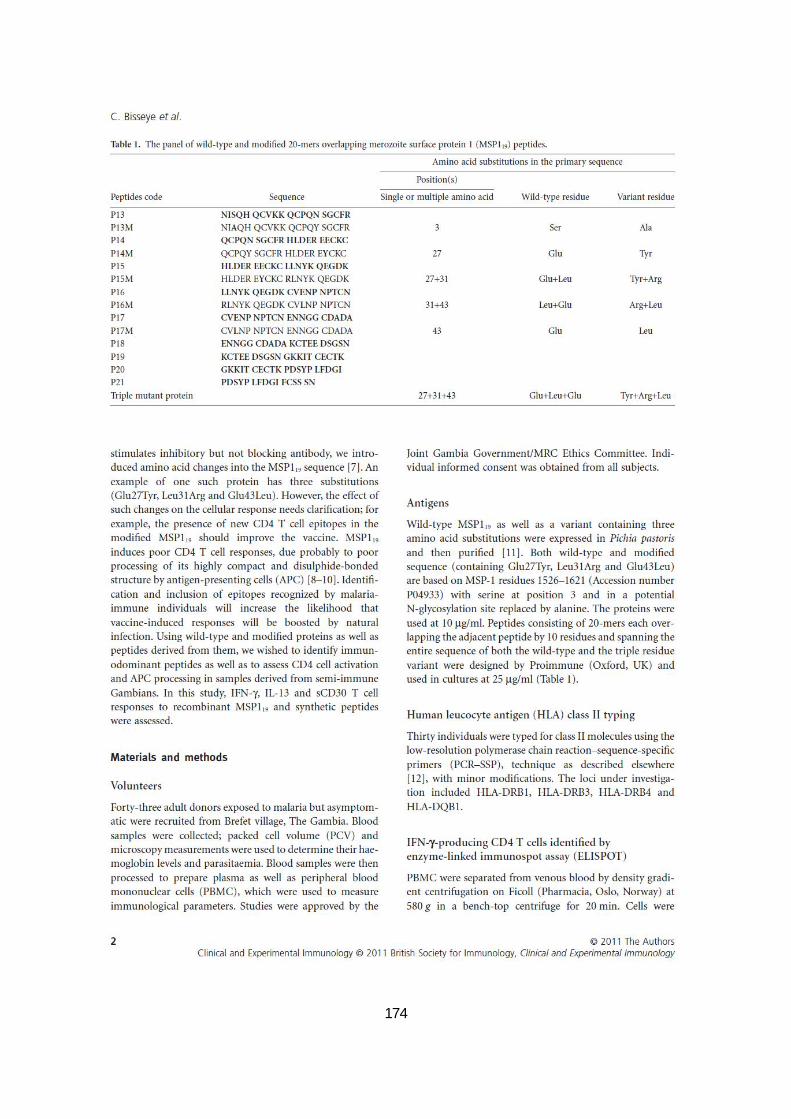
174

175
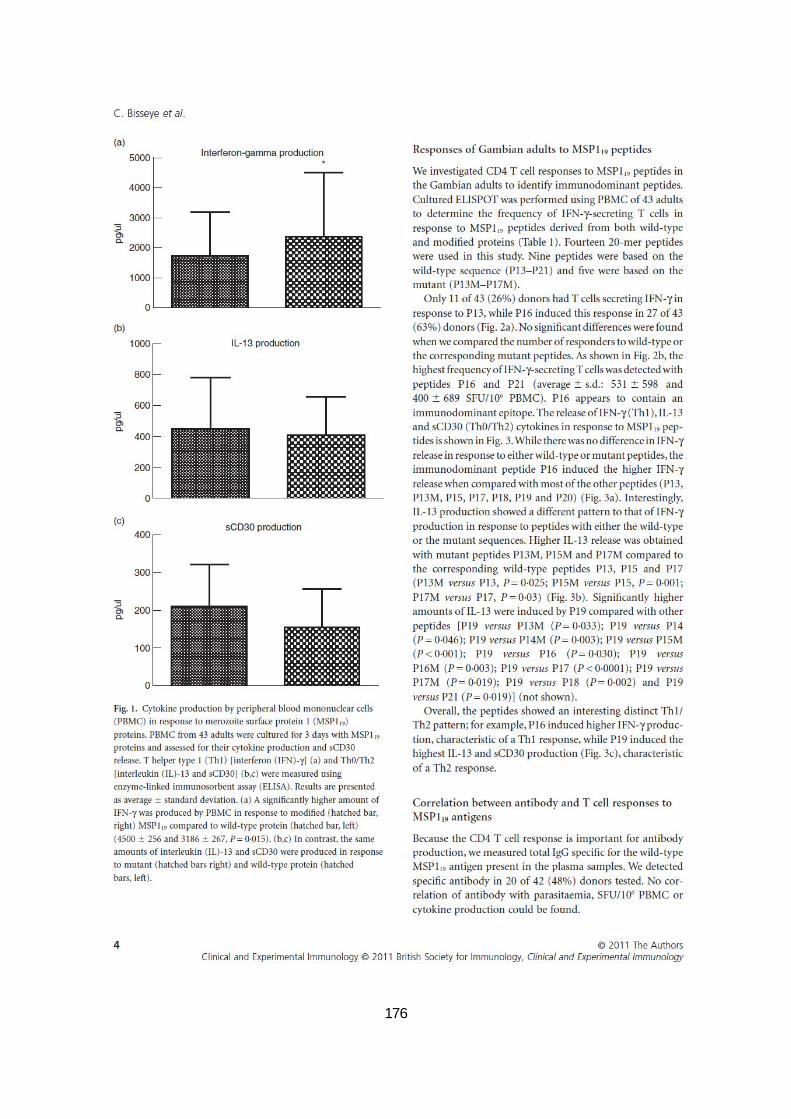
176
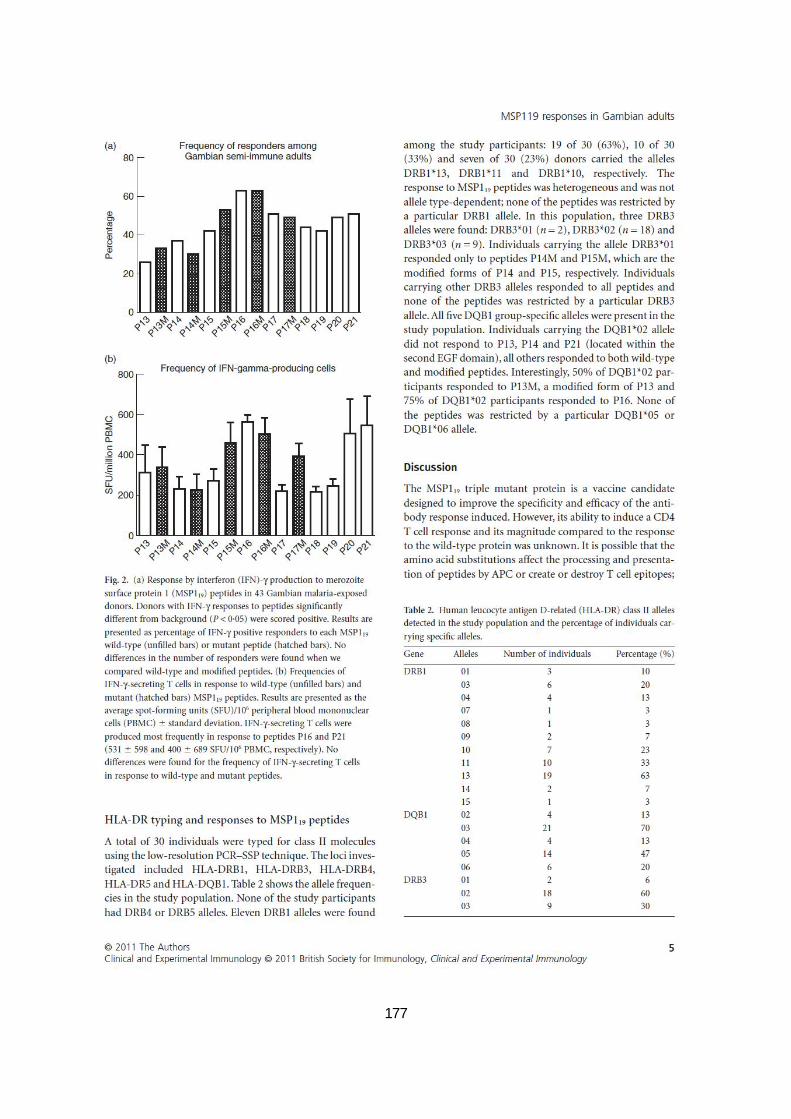
177
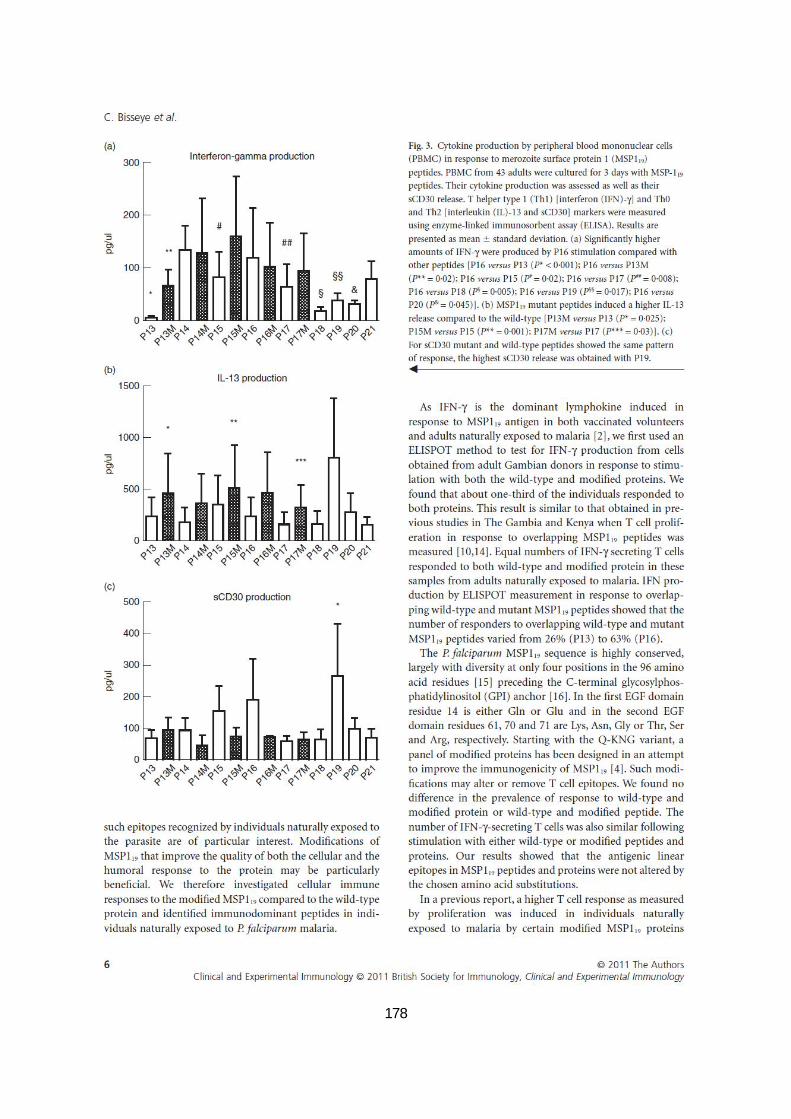
178

179

180

181
Comparison of the humoral responses to CSP, AMA1 and MSP119 and susceptibility to
malaria in Fulani, and the sympatric Wolof and Mandinka groups in The Gambia
Jamila Ismaili1*†, Cyrille Bisseye1,4‡, Dongmei liu2, Hanif Ismael1, Musa Jawara1, Mathurin
Diatta1, Giorgio Sirugo1, Margaret Pinder1, Sam dunyo1, William D. Morgan3,
Michael J. Blackman3, Anthony A. Holder3, Jacques Simpore4 and Paul Milligan1,2.
Medical Research Council Laboratories, PO Box 273, Banjul, The Gambia1; Department of
Infectious and Tropical Diseases, London School of Hygiene and Tropical Medicine, Keppel
Street, London, WC1E 7HT, United Kingdom2; Division of Parasitology, MRC National
Institute for Medical Research, Mill Hill, London NW7 1AA, United Kingdom3; and Centre
de Recherche Biomoléculaire Pietro Annigoni, Ouagadougou, Burkina Faso4
Running Title: vaccine candidates and malaria incidence in Gambian Fulani
Key words: Malaria, Antibody, Ethnic group, CSP, AMA1 and MSP119
Footnotes:
Corresponding author*. Mailing address: IBL, Institut Pasteur, Lille, France.
Phone: 33-320 871121
Fax: 33-320 871158
E-mails: [email protected]; [email protected]
Present addresses: †IBL, Institut Pasteur, Lille, France.
‡Centre de Recherche Biomoléculaire Pietro Annigoni (CERBA), Ouagadougou, Burkina
Faso

182
Abstract
The Fulani of Burkina Faso and Mali seem to be more resistant to malaria than other groups
due to their higher levels of antibody responses to several malaria antigens. Few studies have
investigated this finding elsewhere in West Africa. We used a large cohort of individuals aged
6 months to 45 years and living in The Gambia to assess interethnic differences in malaria
incidence and humoral responses to malaria antigens (CSP, MSP119 and AMA1) between
Fulani and the sympatric Mandinka and Wolof. In contrast to the situation in Burkina Faso
and Mali, we found similar malaria incidence, and parasite prevalence in the three groups,
although the humoral response to MSP119 in the Gambian Fulani was higher than that of their
sympatric groups. High responses to MSP119 and AMA1, but not to CSP, were associated
with protection against malaria. Interestingly, adjusting for parasitaemia at enrolment
abolished the association of responses to all vaccine candidates with protection against
clinical malaria.

183
Introduction
It is well established that Fulani from Burkina Faso and Mali, have lower parasite density and
higher levels of antibodies to a variety of malaria antigens when compared to the other
sympatric Mossi, Rimaibe and Dogon ethnic groups (Modiano et al, 1996; Dolo et al, 2005).
Very few studies are available for Fulani from other West African countries. Riley et al.
(1992a) reported a higher antibody response of the Gambian Fulani to the Pf155 ring-infected
erythrocyte surface antigen (RESA), associated this immune response with resistance to
clinical malaria and correlated it with class II DQA-DQB, which is common in the Fulani
ethnic group. Other reports have referred to splenomegaly and higher IgM levels as more
frequent in the Fulani from Nigeria (Oomen et al, 1979), Burkina Faso and Mali (Bolad et al,
2005) and The Gambia (Greenwood et al, 1987) compared to other sympatric ethnic groups.
However, the latter studies did not provide parasitological or specific immunological data and
did not verify to what extent the high antibody response to Plasmodium falciparum is a
common trait of the Fulani ethnic group across West Africa. Consequently we have conducted
a cohort study in the Gambia in a wide age range of participants from the three major ethnic
groups: Mandinka, Fulani and Wolof, to investigate humoral responses, parasite prevalence,
and resistance to malaria. We investigated the humoral response to the pre-erythrocytic stage
circumsporozoite protein (CSP) and to the erythrocytic stage merozoite antigens, apical
membrane antigen 1 (AMA1), and the 19 kDa C-terminal region of merozoite surface protein
1 (MSP119). All proteins are candidates in development for inclusion in a vaccine against P.
falciparum malaria. Immune responses to these antigens may prevent or reduce malaria–
related morbidity and mortality by inhibiting sporozoites invasion and development within
hepatocytes or merozoite invasion of erythrocytes, thus eliminating or reducing the parasite
load, as supported by evidence drawn from studies of malaria in humans and several animal

184
models (Heppner et al, 2005; Egan et al, 1996; Polley et al, 2004). However, there is
conflicting evidence for the protective effects of responses to CSP and MSP119 in studies
from different West African countries (Dodoo et al, 1999; Riley et al, 1992b). In addition,
further evidence is needed for a broad protective effect of AMA1-specific antibodies, for
example because of extensive antigenic polymorphism. Here we have used a cohort study to
reassess the protective effects of responses to each of the vaccine candidates. We have re-
examined antigen-specific IgG1/IgG3 subclass levels present before the malaria season to
predict the proteins' potential efficacy as vaccine candidates, adjusting for the confounding
effects of exposure and age, as well as the prevalence of parasites and ethnicity. This is also
the first report of a cohort study designed to compare humoral responses to these antigens in
blood samples from Gambians of Fulani, Mandinka and Wolof ethnicity and with reference to
their susceptibility to clinical malaria.
Materials and methods
Study Design
A cohort of 658 adults and children was recruited from 8 rural villages surrounding Farafenni,
a town on the North bank of the River Gambia which experiences annual peak malaria
transmission from August to December (October to November being the highest transmission
period). We used the MRC Household Registration System to recruit a sample of children and
adults aged from 6 months to 45 years, stratified by age and ethnicity so that the age
distribution was the same in each group (Wolof, Mandinka and Fulani). Informed consent
was obtained from all adult participants or the parent or guardian for the children below 18-

185
years old. Venous blood samples were collected during the period 14th to 28th July 2003 from
each participant for analysis of their immune responses and parasitaemia.
500 µl of blood collected in EDTA tubes was used for sickle cell haemoglobin genotyping,
packed cell volume (PCV) measurement and thick blood smear microscopy for parasite
detection. The remainder of the blood was collected into a heparin tube for subsequent
measurement of humoral and cell immune responses. Plasma samples were stored at –20°C
prior to measurement of antibody levels.
Following initial sampling in July 2003, each participant in the cohort was visited weekly by a
field worker for a period of 23 weeks (up to January 2004). Axillary temperature was
measured with a digital thermometer, and in cases of fever (temp >37.5°C), a rapid diagnostic
test (ICT Malaria Pf/PvTM: histidine-rich protein antigen II- AMRAD, Australia) was
performed and the person treated for malaria according to national guidelines (at that time,
SP+CQ), if the test was positive. A blood film was also taken to determine parasite density by
microscopy. Clinical malaria incidence for any cohort participant, who attended the Farafenni
APFRC hospital or outpatients clinic, was also recorded. Cross-sectional surveys of all
participants were also carried out; finger prick samples were collected at the end of October
and the beginning of November (the peak of highest transmission) and during mid December
(low transmission) for preparation of thick blood films. Parasitaemia was assessed by
microscopic examination of Giemsa-stained smears; all slides were read twice by experienced
microscopists and any discrepancy resolved by a third reader. One hundred fields at 1000x
magnification were examined before declaring a slide negative (limit of detection,
approximately 2 parasites/l). Malaria was defined as fever (axillary temperature >37.5oC)
with a parasite density > 5000/µL (Migot-Nabias et al, 2000). Details of bed-net use and
condition, insecticide use, and type of house were collected in July 2003. A surveillance was

186
carried out in the villages to estimate the entomological inoculation rate (EIR). The ethnicity
of participants was determined from questionnaires to collect information on the ethnicity of
their parents and grandparents.
The study was approved by the Joint Gambia Government/MRC Ethical Committee.
Sickle cell haemoglobin and G6PD genotyping
Genomic DNA was extracted from the buffy coat cell fraction or whole blood using a
standard salting out procedure, and sickle cell trait genotyping was performed by PCR, using
an amplification refractory mutation system (ARMS), to detect the sickle cell mutation (an A
to T transition in codon six of the β-globin gene). Primer sequences for typing HbS were as
follows:
HbS allele - 5'GGCAGTAACGGCAGACTTCTCCA 3'
Normal HbA allele - 5' GGCAGTAACGGCAGACTTCTCCT 3'
Reverse primer - 5' GCCAGTGCCAGAAGAGCCAA 3'
Sixty 60 ng genomic DNA was mixed with 15 pmol of each forward and reverse primer, in a
15 µl reaction containing 16mM (NH4)2SO4, 67mM Tris-HCl (pH 8.8 at 25ºC), 3.45 mM
MgCl2, 0.6 mM each dNTP, and 0.5 U Taq DNA polymerase (Bioline Ltd., London, UK).
Cycling conditions were 96oC 1 min; 5 cycles of 96oC 35 sec, 70oC 45 sec and 72oC 35 sec;
then 21 cycles of 96oC 25 sec, 65oC 50 sec and 72oC 40 sec; then 6 cycles of 96oC 35sec,
and 55ºC 1 min; followed by 72oC 90 sec. PCR products were electrophoresed on a 2%
agarose gel, stained with ethidium bromide and digitally photographed using a Gel Doc
System 2000 apparatus (Bio-Rad Laboratories Ltd. Hertfordshire, UK).

187
For G6PD status determination, samples (4 ng of DNA) were genotyped by Taqman assays
(ABI, Applera International Inc, Foster City, CA, USA) in 10 µl reaction volume using the
ABI 7500 real-time PCR systems. Fluorescence curves were analyzed with the 7500
Sequence Detection Software version 1.2.1 for allelic discrimination.
Antibody responses to MSP119, AMA1 and R32LR
Levels of antibody to MSP119 and AMA1 were measured in the plasma samples using ELISA.
Briefly, ELISA plates (Immunlon4, Dynatech Laboratories Inc.) were coated with 1µg/ml of
MSP119 or AMA1 in 100 µl carbonate buffer and incubated overnight at 4 ˚C, then washed
with PBS-Tween (0.05%), flicked dry and 150 µl of PBS-1% BSA blocking solution was
added. Plates were incubated at room temperature for 3 hours, then flicked dry and diluted
plasma samples were added. The plasma samples in duplicate were assayed at 1:1000;
1:2000, 1:4000, and 1:8000 dilutions for MSP119 and at 1:1000 and 1: 2000 dilutions for
AMA1. Serial dilutions of purified human IgG and anti human IgG-POD were used as
standards on each plate to assess the relative concentration of antibody (Ab) to AMA1 and
MSP119. The plates were incubated overnight at 4˚C, washed 6 times and horseradish-
peroxidase (HRP)-goat anti-human IgG (Dako A/S, Denmark) or horseradish-peroxidase
(HRP)-rabbit anti-human IgG1 or anti-IgG3 for the subtype determination (The Binding Site,
UK) were added at a 1:5000 dilution in PBS-1% BSA. The plates were incubated for 3 hours
at room temperature, washed 6 times and the reaction developed by adding 100 µl
tetramethyl-benzidine (TMB) (Sigma, UK). The reaction was stopped with 100 µl 2M H2SO4
and the plates read at 450 nm.
Samples from donors resident in Brefet were used as positive controls and sera of European
tourists with no previous exposure to malaria were used as negative controls. The mean + 3

188
standard deviation (SD) of the negative control European sera was used as the end point titre
cut-off to define ‘responders’.
To quantify IgG binding to the P. falciparum CSP NANP-repeat sequence, 0.1 µg of the
R32LR recombinant antigen in 50µl PBS and casein (4 µl of boiled casein [0.5% casein with
Phenol Red, Thimerosal, PBS, 5N NaOH; SIGMA] per 5 ml of diluted antigen) was used to
coat each well on Immunlon plates. Following incubation overnight at room temperature, the
wells were then blocked with 250 µl of blocking buffer (0.5% Boiled Casein and Tween) for
one hour at room temperature. The contents of the well were removed by aspiration and 50 µl
of an appropriate plasma dilution were added. Each plate contained the following wells: a
blank, negative and positive serum (DGP) controls and the samples (in triplicate) starting at a
1:50 dilution and with further two-fold dilutions. Samples were incubated in the wells for two
hours at room temperature and then the wells were washed with PBS-Tween 20. Goat
antihuman IgG conjugated to horse radish peroxidase (Kierkegaard and Perry Laboratories) at
a dilution of 1: 1000 was added at 50 µl/well and incubated for one hour at room temperature.
Wells were washed four times with PBS-Tween 20 and 100ul of substrate solution (ABTS
HRP Substrate Kit, Kierkegaard and Perry Laboratories) were added and incubated for one
hour at room temperature. Ten µl of 20% SDS (SIGMA) were used to stop the reaction and
then the absorbance was read at 414 nm. For each plasma sample tested three readings of a
dilution which gave an OD value of around 1.0 were selected. The mean OD reading was
calculated and this mean was multiplied by the dilution factor to obtain the weighted mean
(WM). The corrected weighted mean (CWM) for each plate was calculated: CWM =
Average WM DGP x WM sample / WM DGP. The IgG concentration (µg/ml) on each plate
was calculated by the formula: IgG in sample (µg/ml) = known concentration for DGP (9.8) x
CWM of sample / CWM of DGP standard.

189
Statistical analysis
Incidence rate ratios for the association of immunological variables and ethnicity with malaria
incidence were estimated by fitting a weibull model to the times of malaria episodes, this
model was chosen because it allows for seasonality in the incidence of malaria. To allow for
the lack of independence among repeated observations on the same individual, a random
effect was included to allow for variation in individual baseline risk, which was assumed to
follow a gamma distribution. Antibody concentrations were included in the model as category
variables, with 5 levels, the first category for zero values, and the remaining 4 categories
obtained by grouping the positive values according to quartiles; a likelihood ratio test was
used to obtain a P-value for the association of each antibody level with malaria incidence.
Covariates were specified in advance: EIR, net use, age group, sickle cell and glucose-6-
phosphate dehydrogenase (G6PD) genotype, and whether the house had open eaves. Several
models were fitted. Firstly, to assess the independent effects of total IgG to AMA1, MSP119
and CSP, all three variables were included in the model with the covariates. Secondly, to
examine the association with IgG subclasses, models were fitted for each antigen in turn,
including the concentration of IgG1 and IgG3 (as category variables with three levels
according to tertiles) and the same list of covariates.
Association between antibody level and various effects was analysed using both one way
analysis of variance and the Bernoulli lognormal model. In the Bernoulli lognormal model,
we modelled the event that the antibody levels from a subject were zero or non zero with a
Bernoulli variable, and modelled the non zero measurement on antibody level with a
lognormal variable. In this dataset, antibody levels are left truncated, i.e. measurements that
are less than 10 pg/ml are considered to be zero. The Bernoulli lognormal mixture model also
considers this by incorporating a left-censoring process in the lognormal component.

190
To detect effects that are protective against malaria infection, we did Poisson regression
analysis on malaria incidence vs. various effects, including age, ethnicity, antibody level, Hb
type, entomology, and housing information. Antibody levels were included as an ordinal
variable in the analysis, i.e. subjects were categorized into high, medium, or low responders
based on the continuous antibody levels. To check how antibody levels were related to time
to the first malaria incidence, Kaplan-Meier plots were made for each antibody, with 3 strata,
high, medium, and low antibody levels.
Results
Baseline characteristics of study participants
Demographic and baseline characteristics of study subjects are shown in Table 1. The cohort
sample was made to be representative of each of the three main ethnic groups in each age
group, the distribution of participants across ethnicity (31.26%, 36.39% and 32.35% in the
Wolof, Fulani and Mandinka groups, respectively) and age categories (20.88%, 42.39% and
36.73% in the less than 5 years, 6-15 years and > 15 years old categories, respectively) was
similar. Nearly 60% of study subjects were female. Reported insecticide-treated net (ITN)
ownership at the beginning of the study was roughly 25%, whereas only 1% had had the net
impregnated with insecticide. The entomological inoculation rate (EIR) was found to be
higher than 50 in 18% of sites of residence and more than 50% of households reported having
an eaves gap in their housing. Acquired immunity to P. falciparum was assessed at enrolment.
Regarding genetic factors affecting protection against malaria, almost one fifth of subjects
were found to be heterozygous for sickle-cell and around 1% was homozygous. Around 4%
and 1% were identified as female heterozygous and male hemizygous for G6PD deficiency,

191
respectively. CSP, MSP119 and AMA1-specific antibodies were found to be present in
76.75%, 64.29% and 50% of participants, respectively.
Comparison of malaria incidence among ethnic groups
We investigated the interethnic differences of malaria incidence among the three major
groups in the Gambia, Fulani (n= 234), Mandinka (n=208) and Wolof (n=201). Malaria
incidence rates were similar in the three ethnic groups; however, the EIR recorded in the
village, and the G6PD and sickle cell genotype, were associated with malaria risk and
ethnicity. Fulani for example were more likely to live in villages where EIR was low, and
after adjustment for the effects of covariates, Fula ethnicity was associated with a slightly
increased risk of malaria compared to the two other ethnic groups. However, adjusted
estimates of malaria incidence, in association with the baseline characteristics described in
Table 2 and in particular parasitaemia at enrolment, abolished the interethnic differences of
clinical malaria and no significant difference of malaria incidence was found between the
three groups [Crude estimates (4.226; ST dev: 0.325; Pr(>|z|):0.000) and adjusted estimates
for all confounders (3.972; ST dev: 0.344; Pr(>|z|):0)].
Comparison of humoral responses to CSP, AMA1 and MSP119 among ethnic groups.
Antibody responses to CSP, MSP119 and AMA1 (total IgG, IgG1 and IgG3), increased with
age, while no significant association was found with bed net use or EIR, neither in logistic
regression nor in log linear regression (not shown). While the proportion of responders to
AMA1 was similar in all ethnic groups, for CSP and MSP119, the proportion of responders
was lower in the Mandinka group than in the other two groups (Table 3). The concentration of
total IgG among the responders was compared between ethnic groups; the concentration of
total IgG and IgG1 to MSP119 was higher in the Fulani group (Figure 1) while total IgG, IgG1

192
and IgG3 to AMA1 was similar in the three ethnic groups. As shown in Figure 1, the
concentrations of total IgG specific to CSP were lower within the Mandinka group compared
to Fulani and Wolof participants, who had similar levels of antibodies. In addition, the three
ethnic groups had similar levels of anti-CSP IgG1 or IgG3.
Antibody association with malaria incidence and parasitaemia
Kaplan-Meier analysis (Figure 2) showed that high antibody levels are associated with
prolonged time to the first malaria incidence, i.e. high antibody levels to CSP, AMA1 and
MSP119 are protective against malaria infection. The crude analysis also showed significant
association of antibodies to the individual antigens, CSP (IgG1 and IgG3), AMA1 (total IgG)
and MSP119 (total IgG, IgG1 and IgG3) with malaria protection (not shown).
As shown in Table 2, after adjustment for age, exposure variables, sickle cell genotype, and
G6PD genotype (but not parasitaemia at recruitment) for all three antigens (total IgG to
AMA1, MSP119 and CSP), there was only a significant association between malaria incidence
and the concentration of total IgG anti-MSP119 and AMA1 antibodies, but not anti-CSP
antibodies.
When IgG subclasses were considered, (Table 4), the concentration of IgG1 to AMA1, and of
IgG1 to CSP, were associated with the incidence of malaria, but there was no association of
malaria incidence with IgG1 to MSP119, and no association of IgG3 with malaria for any of
the antigens.
After adjusting for the effects of total IgG to the three antigens, ethnicity was independently
associated with malaria incidence; Mandinka and Wolof ethnicity was associated with
reduced incidence compared to Fulani.

193
When parasitaemia at enrolment was included as a confounder factor, this abolished the
observed associations (not shown). Looking at cumulative protective effects of the different
antibodies, no significant interaction between the protection from CSP and the blood stage
antigens AMA1 and MSP119 was observed, whether or not parasitaemia was included.
The parasitological data from the three cross-sectional surveys (Figure 3), in July, October-
November and December showed that amongst the three ethnic groups Wolof tended to have
a higher prevalence of P. falciparum. However, low parasite prevalence, and the fluctuations
of the parasite rates and densities show that there is no evidence of coherent interethnic
differences.
Discussion
Fulani populations from Burkina Faso and Mali were reported to make stronger antibody
responses to malaria while being less frequently parasitized and less susceptible to clinical
malaria (Modiano et al, 1996) compared to other sympatric groups (Mossi and Rimaibé).
Accordingly, our cohort study investigated interethnic differences in humoral responses, to
the CSP, AMA1 and MSP119 vaccine candidates between Fulani and the sympatric Mandinka
and Wolof in the Gambia. In addition, because different immunoepidemiological studies
conducted at several sites have yielded conflicting evidence on the association of anti-CSP or
anti-MSP119 antibodies with clinical immunity to malaria, and because the only cohort study
described for anti-AMA1 was conducted in Kenya where the pattern of malaria transmission
is different from that in the Gambia (Egan et al, 1996; Polley et al, 2004; Dodoo et al, 1999),
we have also reassessed the association of antibody responses to these antigens with clinical
immunity. To assess the reported discrepancies, with sufficient statistical power and to
examine whether or not there was a real association with protection, the genetic background

194
and parasitaemia at enrolment, which had not been considered in previous studies, were
included as confounders.
The comparison of malaria risk between ethnic groups is complicated by many confounding
factors. In this study, the crude analysis of malaria incidence in three ethnic groups showed
that the Fulani had the highest malaria incidence compared to other ethnic groups in The
Gambia. Whilst the Wolof group had the lowest malaria incidence, after adjustment for all
confounders no significant difference was found between the three ethnic groups. It is
important to allow for differences in EIR and other factors associated with exposure such as
house construction. Although malaria incidence was similar in all three ethnic groups, there
were local geographical differences in relation to EIR; and Fulani tended to live in villages
with lower EIR. There was no evidence that people living in areas of higher EIR had higher
IgG concentrations to any of the antigens or were more likely to be responders. The mean
concentration of IgG to MSP119, which had the strongest association with malaria incidence
of the antigens tested, was higher among Fulani. After adjusting for total IgG and
environmental and genetic factors associated with malaria risk, Mandinka and Wolof ethnicity
was associated with lower malaria incidence. This could reflect genetic differences (Koch et
al, 2002), or residual confounding factors. We found similar overall rates but after adjusting
for these factors Fulani were associated with higher incidence, and stronger immune
responses as reported previously (Bolad et al, 2005). Immune responses could partly reflect
greater exposure.
The previous interethnic comparison (Modiano et al, 1996) revealed that humoral responses
were consistently higher in the Fulani. In our study, the crude analysis of the level of humoral
response showed higher levels of antibody in Fulani compared to other sympatric groups,
which is in agreement with the findings in Burkina Faso. Concentrations of total IgG to CSP

195
were lower within the Mandinka compared to Fulani and Wolof groups, who had similar
levels of antibodies. However, similar levels of CSP-specific IgG1 and IgG3 were present in
all three groups. Levels of total anti AMA1 IgG and IgG1 were similar in the three groups,
whilst IgG3 levels were highest among Fulani, medium in Wolof, and lowest in the Mandinka
group. This finding is in accordance with a previous report which showed a high level of IgG3
among Fulani from Burkina Faso and Mali compared to Mossi and Dogon respectively (Bolad
et al, 2005). In contrast, levels of total anti-MSP119, IgG1 and IgG3 Abs were all higher in
Fulani compared to the Mandinka and Wolof groups, in support of Modiano’s findings
(Modiano et al, 1996).
In the previous study, parasitological and clinical evidence of a higher resistance of the Fulani
to P. falciparum was associated with their stronger humoral reactivity against a different
panel of parasite antigens: TRAP, MSP1, Pf155 (RESA), and Pf332 (Modiano et al, 1996).
We have examined humoral responses to CSP, MSP119 and AMA1 and their association with
protection against clinical malaria, including genetic traits as a new confounder. The crude
analysis associated total IgG to CSP, AMA1 and MSP119 as well as IgG1 and IgG3 to CSP
and MSP119 with protection against malaria. Adjusting for all covariates, except parasitaemia
at recruitment reduced this association for CSP. Furthermore, there was no significant
interaction or extra protection afforded to double or triple responders with total IgG, IgG1 or
IgG3 to CSP, AMA1 or MSP119. Parasitaemia at enrolment was either associated with lower
levels of antibodies or with malaria incidence; but no association of antibodies to CSP, AMA1
and MSP119 with protection and parasitaemia was found at enrolment. Interestingly, including
parasitaemia at enrolment as a confounder completely abolished all the observed associations.
This is in contrast to a study in Kenya, where presence of antibodies to AMA1 was associated
with protection in subjects carrying P. falciparum at recruitment (Polley et al, 2004).

196
We analysed the parasite prevalence and density, using samples from three cross sectional
surveys at the beginning of the transmission season (July), at the highest transmission period
(October-November) and at the end of the transmission season (December). We found that the
Wolof group had a slightly higher prevalence of P. falciparum. However, no clear conclusion
of interethnic differences could be drawn because of the low parasite prevalence in our cohort.
Previous studies had also reported low parasite prevalence in population samples mostly
constituted by Fulani in northern Nigeria (Oomen et al, 1979) and The Gambia (Riley et al,
1992b).
Our data are different from those reported previously from Burkina Faso. This discrepancy
may be attributed to large genetic differences between the Burkina Faso and Gambian Fulani,
or greater genetic differences between the Fulani and the sympatric Raimbé and Mossi groups
in Burkina Faso compared with the Fulani and the sympatric Mandinka and Wolof groups in
The Gambia (Modiano et al, 2001). Indeed, recent studies of GM polymorphism in Eastern
Senegal (Allsopp et al, 1992) and of HLA class I (Blanc et al, 1990) and II (Riley et al,
1992a) in The Gambia indicate rather small genetic distances between the Fulani and their
neighbours; in contrast preliminary analysis of individuals from Burkina Faso suggests a
relatively higher degree of genetic divergence.
The reported higher antibody response of the Fulani may be attributed to factors encoded by
genes outside the HLA class II region and involved in the regulation of humoral immune
responses against P. falciparum antigens (Sjoberg et al, 1992). Higher humoral responses
were also observed in the Gambian Fulani population, even though no association with
protection to malaria was found. Possible immunological mechanisms involving genes related
to B-cell function, antigen-presenting cell (Farouk et al, 2005; Arama et al, 2011), interleukin
4 production (Luoni et al, 2001), or regulatory T cell differentiation (Torcia et al, 2008) may

197
still show interethnic differences and be shared by Fulani across West Africa. Accordingly,
ongoing investigation into interethnic polymorphisms in Th1/Th2 cell cytokine genes may
provide insights into genes involved in regulation of humoral responses (G. Sirugo, in
preparation).
In summary, we report here for the first time studies on interethnic differences of malaria
incidence and humoral responses of Fulani, Mandinka and Wolof in The Gambia. We have
shown that Fulani compared to Mandinka and Wolof had a higher antibody response but a
similar pattern of malaria incidence and parasite prevalence and density. Interestingly,
including parasitaemia at enrolment as a confounder abolished the association of humoral
responses to all three vaccine candidates: MSP119, CSP and AMA1, which is in contrast to
reports of previous studies.
Acknowledgements: We are grateful to the all participants enrolled into the cohort study, to
the staff of Farafenni Health Centre and to the Department of State for Health. We thank all
Farfenni field workers as well as Simon Correa, Idrissa Sambou and Saikou Keita for the
technical support. The work was supported in part by the MRC through U117532067 and
U117532063

198
References
Arama C, Giusti P, Boström S, Dara V, Traore B, Dolo A, Doumbo O, Varani S, Troye-Blomberg M: Interethnic differences in antigen-presenting cell acrivation and TLR responses in Malian children during Plasmodium falciparum malaria. PlosOne 2011, 6 (3):e18319
Allsopp CE, Harding RM, Taylor C, Bunce M, Kwiatkowski D, Anstey N, Brewster D, McMichael AJ, Greenwood BM, Hill AV: Interethnic genetic differentiation in Africa: HLA class I antigens in The Gambia. Am J Hum Genet 1992, 50(2):411-421.
Blanc M, Sanchez-Mazas A, Van Blyenburgh NH, Sevin A, Pison G, Langaney A: Interethnic genetic differentiation: GM polymorphism in eastern Senegal. Am J Hum Genet 1990, 46(2):383-392.
Bolad A, Farouk SE, Israelsson E, Dolo A, Doumbo OK, Nebié I, Maiga B, Kouriba B, Luoni G, Sirima BS, Modiano D, Berzins K, Troye-Blomberg M: Distinct Interethnic Differences in Immunoglobulin G Class/Subclass and Immunoglobulin M Antibody Responses to Malaria Antigens but not in Immunoglobulin G Responses to Nonmalarial Antigens in Sympatric Tribes Living in West Africa. Scandinavian Journal of immunology 2005, 61:380-386
Dodoo D, Theander TG, Kurtzhals JA, Koram K, Riley E, Akanmori BD, Nkrumah FK, Hviid L: Levels of antibody to conserved parts of Plasmodium falciparum merozoite surface protein 1 in Ghanaian children are not associated with protection from clinical malaria. Infect Immun 1999, 67(5):2131-2137.
Dolo A, Modiano D, Maiga B, Daou M, Dolo G, Guindo H, Ba M, Maiga H, Coulibaly D, Perlman H, Blomberg MT, Touré YT, Coluzzi M, Doumbo O. Difference in susceptibility to malaria between two sympatric ethnic groups in Mali. Am J Trop Med Hyg. 2005 Mar;72(3):243-8.
Egan AF, Morris J, Barnish G, Allen S, Greenwood BM, Kaslow DC, Holder AA, Riley EM: Clinical immunity to Plasmodium falciparum malaria is associated with serum antibodies to the 19-kDa C-terminal fragment of the merozoite surface antigen, PfMSP-1. J Infect Dis 1996, 173(3):765-769.
Farouk SE, Dolo A, Bereczky S, Kouriba B, Maiga B, Farnert A, Perlmann H, Hayano M, Montgomery SM, Doumbo OK et al: Different antibody- and cytokine-mediated responses to Plasmodium falciparum parasite in two sympatric ethnic tribes living in Mali. Microbes Infect 2005, 7(1):110-117.
Greenwood BM, Groenendaal F, Bradley AK, Greenwood AM, Shenton F, Tulloch S, Hayes R: Ethnic differences in the prevalence of splenomegaly and malaria in The Gambia. Ann Trop Med Parasitol 1987, 81(4):345-354.

199
Heppner DG, Jr., Kester KE, Ockenhouse CF, Tornieporth N, Ofori O, Lyon JA, Stewart VA, Dubois P, Lanar DE, Krzych U et al: Towards an RTS,S-based, multi-stage, multi-antigen vaccine against falciparum malaria: progress at the Walter Reed Army Institute of Research. Vaccine 2005, 23(17-18):2243-2250.
Koch O, Awomoyi A, Usen S, Jallow M, Richardson A, Hull J, Pinder M, Newport M, Kwiatkowski D. IFNGR1 gene promoter polymorphisms and susceptibility to cerebral malaria. J Infect Dis 2002, 185:1684-1687
Luoni G, Verra F, Arca B, Sirima BS, Troye-Blomberg M, Coluzzi M, Kwiatkowski D, Modiano D: Antimalarial antibody levels and IL4 polymorphism in the Fulani of West Africa. Genes Immun 2001, 2(7):411-414.
Migot-Nabias F, Mombo LE, Luty AJ, Dubois B, Nabias R, Bisseye C, Millet P, Lu CY, Deloron P. Human genetic factors related to susceptibility to mild malaria in Gabon. Genes Immun. 2000 Oct;1(7):435-41.
Modiano D, Luoni G, Petrarca V, Sodiomon Sirima B, De Luca M, Simpore J, Coluzzi M, Bodmer JG, Modiano G: HLA class I in three West African ethnic groups: genetic distances from sub-Saharan and Caucasoid populations. Tissue Antigens 2001, 57(2):128-137.
Modiano D, Petrarca V, Sirima BS, Nebie I, Diallo D, Esposito F, Coluzzi M: Different response to Plasmodium falciparum malaria in west African sympatric ethnic groups. Proc Natl Acad Sci U S A 1996, 93(23):13206-13211.
Oomen JM, Meuwissen JH, Gemert W: Differences in blood status of three ethnic groups inhabiting the same locality in Northern Nigeria. Anaemia, splenomegaly and associated causes. Trop Geogr Med 1979, 31(4):587-606.
Polley SD, Mwangi T, Kocken CH, Thomas AW, Dutta S, Lanar DE, Remarque E, Ross A, Williams TN, Mwambingu G et al: Human antibodies to recombinant protein constructs of Plasmodium falciparum Apical Membrane Antigen 1 (AMA1) and their associations with protection from malaria. Vaccine 2004, 23(5):718-728.
Riley EM, Olerup O, Bennett S, Rowe P, Allen SJ, Blackman MJ, Troye-Blomberg M, Holder AA, Greenwood BM: MHC and malaria: the relationship between HLA class II alleles and immune responses to Plasmodium falciparum. Int Immunol 1992a, 4(9):1055-1063.
Riley EM, Allen SJ, Wheeler JG, Blackman MJ, Bennett S, Takacs B, Schonfeld HJ, Holder AA, Greenwood BM: Naturally acquired cellular and humoral immune responses to the major merozoite surface antigen (PfMSP1) of Plasmodium falciparum are associated with reduced malaria morbidity. Parasite Immunol 1992b, 14(3):321-337.
Sjoberg K, Lepers JP, Raharimalala L, Larsson A, Olerup O, Marbiah NT, Troye-Blomberg

200
M, Perlmann P: Genetic regulation of human anti-malarial antibodies in twins. Proc Natl Acad Sci U S A 1992, 89(6):2101-2104.
Torcia MG, Santarlasci V, Cosmi L, Clemente A, Maggi L, Mangano VD, Verra F, Bancone G, Nebie I, Sirima BS et al: Functional deficit of T regulatory cells in Fulani, an ethnic group with low susceptibility to Plasmodium falciparum malaria. Proc Natl Acad Sci U S A 2008, 105(2):646-651.
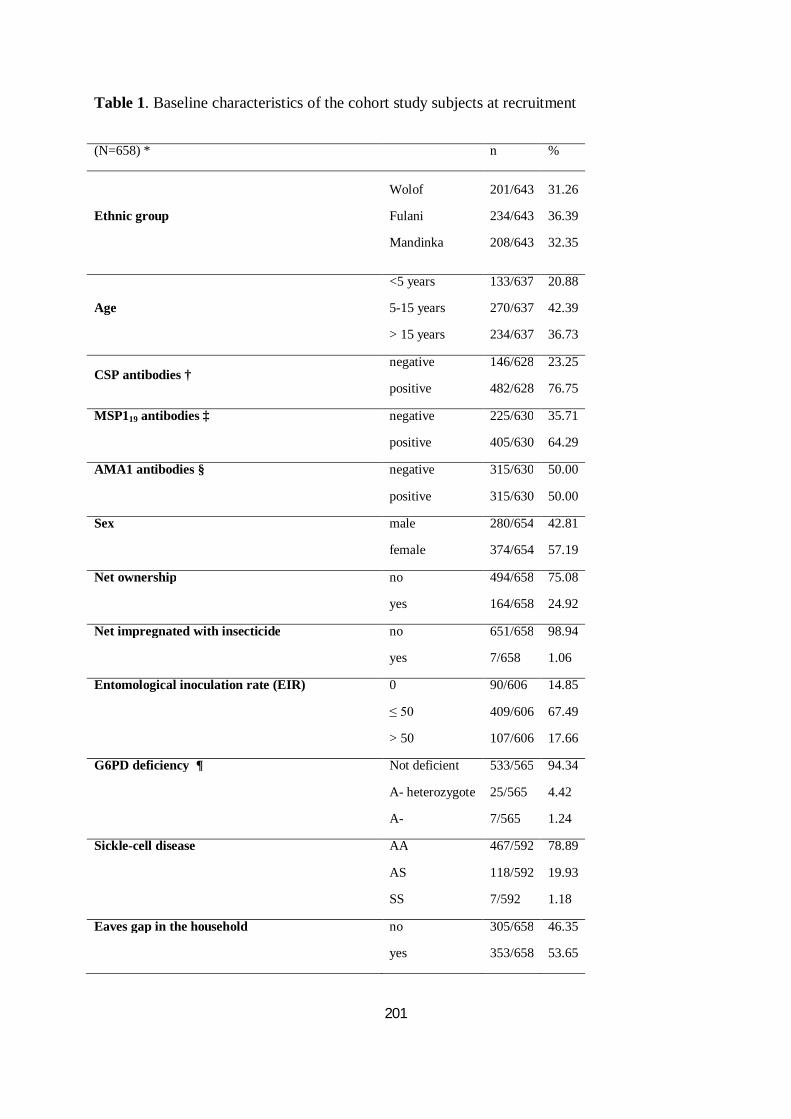
201
Table 1. Baseline characteristics of the cohort study subjects at recruitment
(N=658) * n %
Ethnic group
Wolof
Fulani
Mandinka
201/643
234/643
208/643
31.26
36.39
32.35
Age
<5 years
5-15 years
> 15 years
133/637
270/637
234/637
20.88
42.39
36.73
CSP antibodies † negative
positive
146/628
482/628
23.25
76.75
MSP119 antibodies ‡
negative
positive
225/630
405/630
35.71
64.29
AMA1 antibodies §
negative
positive
315/630
315/630
50.00
50.00
Sex
male
female
280/654
374/654
42.81
57.19
Net ownership
no
yes
494/658
164/658
75.08
24.92
Net impregnated with insecticide
no
yes
651/658
7/658
98.94
1.06
Entomological inoculation rate (EIR)
0
≤ 50
> 50
90/606
409/606
107/606
14.85
67.49
17.66
G6PD deficiency ¶
Not deficient
A- heterozygote
A-
533/565
25/565
7/565
94.34
4.42
1.24
Sickle-cell disease
AA
AS
SS
467/592
118/592
7/592
78.89
19.93
1.18
Eaves gap in the household
no
yes
305/658
353/658
46.35
53.65

202
Malaria treatment in the 30 days before cross 1 ║
no
yes
408/606
198/606
67.33
32.67
Malaria treatment in the 30 days before cross 2 no
yes
401/587
186/587
68.31
31.69
* Number of subjects included in the analysis
† CSP: Circumsporozoite Surface Protein
‡ MSP119: Conserved C terminal region of Merozoite Surface Protein 1
§ AMA1: Apical Membrane Antigen 1
¶ G6PD: Glucose-6-phosphate dehydrogenase
║ This information is not strictly baseline data
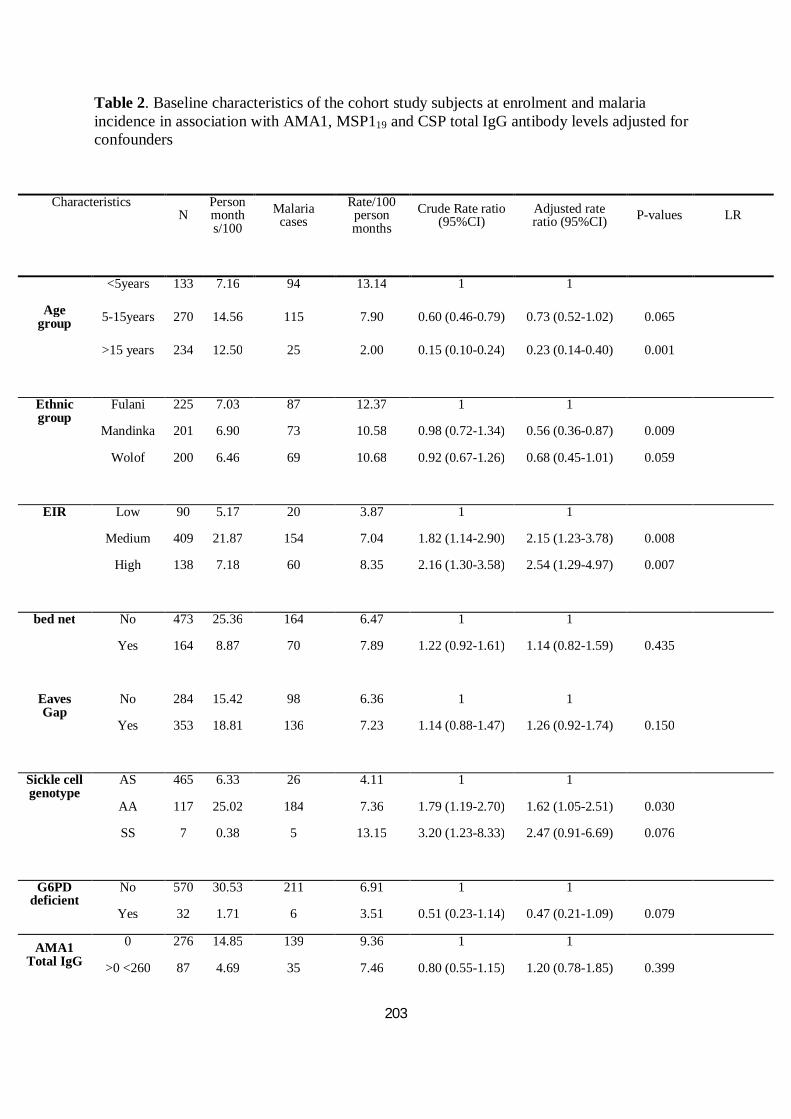
203
Table 2. Baseline characteristics of the cohort study subjects at enrolment and malaria incidence in association with AMA1, MSP119 and CSP total IgG antibody levels adjusted for confounders
Characteristics N
Person months/100
Malaria cases
Rate/100 person months
Crude Rate ratio (95%CI)
Adjusted rate ratio (95%CI) P-values LR
<5years 133 7.16 94 13.14 1 1
Age group 5-15years 270 14.56 115 7.90 0.60 (0.46-0.79) 0.73 (0.52-1.02) 0.065
>15 years 234 12.50 25 2.00 0.15 (0.10-0.24) 0.23 (0.14-0.40) 0.001
Ethnic group
Fulani 225 7.03 87 12.37 1 1
Mandinka 201 6.90 73 10.58 0.98 (0.72-1.34) 0.56 (0.36-0.87) 0.009
Wolof 200 6.46 69 10.68 0.92 (0.67-1.26) 0.68 (0.45-1.01) 0.059
EIR Low 90 5.17 20 3.87 1 1
Medium 409 21.87 154 7.04 1.82 (1.14-2.90) 2.15 (1.23-3.78) 0.008
High 138 7.18 60 8.35 2.16 (1.30-3.58) 2.54 (1.29-4.97) 0.007
bed net No 473 25.36 164 6.47 1 1
Yes 164 8.87 70 7.89 1.22 (0.92-1.61) 1.14 (0.82-1.59) 0.435
Eaves Gap
No 284 15.42 98 6.36 1 1
Yes 353 18.81 136 7.23 1.14 (0.88-1.47) 1.26 (0.92-1.74) 0.150
Sickle cell genotype
AS 465 6.33 26 4.11 1 1
AA 117 25.02 184 7.36 1.79 (1.19-2.70) 1.62 (1.05-2.51) 0.030
SS 7 0.38 5 13.15 3.20 (1.23-8.33) 2.47 (0.91-6.69) 0.076
G6PD deficient
No 570 30.53 211 6.91 1 1
Yes 32 1.71 6 3.51 0.51 (0.23-1.14) 0.47 (0.21-1.09) 0.079
AMA1 Total IgG
0 276 14.85 139 9.36 1 1
>0 <260 87 4.69 35 7.46 0.80 (0.55-1.15) 1.20 (0.78-1.85) 0.399
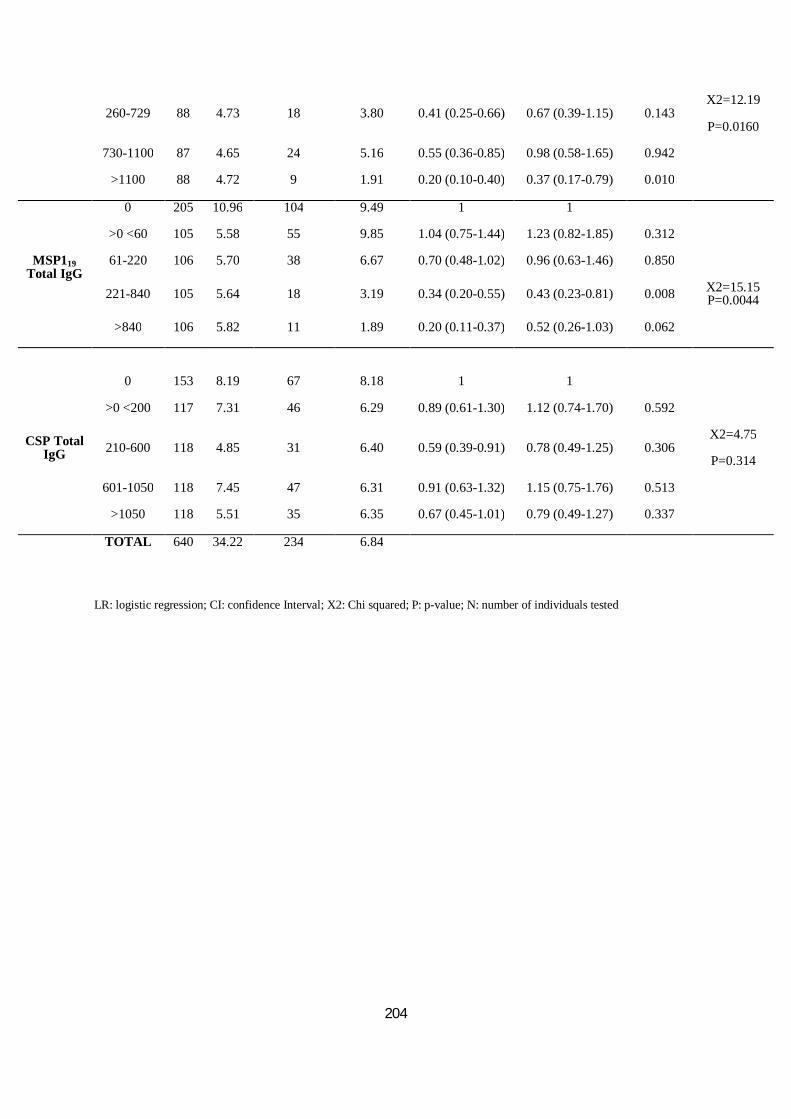
204
260-729 88 4.73 18 3.80 0.41 (0.25-0.66) 0.67 (0.39-1.15) 0.143 X2=12.19
P=0.0160
730-1100 87 4.65 24 5.16 0.55 (0.36-0.85) 0.98 (0.58-1.65) 0.942
>1100 88 4.72 9 1.91 0.20 (0.10-0.40) 0.37 (0.17-0.79) 0.010
MSP119 Total IgG
0 205 10.96 104 9.49 1 1
>0 <60 105 5.58 55 9.85 1.04 (0.75-1.44) 1.23 (0.82-1.85) 0.312
61-220 106 5.70 38 6.67 0.70 (0.48-1.02) 0.96 (0.63-1.46) 0.850
221-840 105 5.64 18 3.19 0.34 (0.20-0.55) 0.43 (0.23-0.81) 0.008 X2=15.15 P=0.0044
>840 106 5.82 11 1.89 0.20 (0.11-0.37) 0.52 (0.26-1.03) 0.062
CSP Total IgG
0 153 8.19 67 8.18 1 1
>0 <200 117 7.31 46 6.29 0.89 (0.61-1.30) 1.12 (0.74-1.70) 0.592
210-600 118 4.85 31 6.40 0.59 (0.39-0.91) 0.78 (0.49-1.25) 0.306 X2=4.75
P=0.314
601-1050 118 7.45 47 6.31 0.91 (0.63-1.32) 1.15 (0.75-1.76) 0.513
>1050 118 5.51 35 6.35 0.67 (0.45-1.01) 0.79 (0.49-1.27) 0.337
TOTAL 640 34.22 234 6.84
LR: logistic regression; CI: confidence Interval; X2: Chi squared; P: p-value; N: number of individuals tested
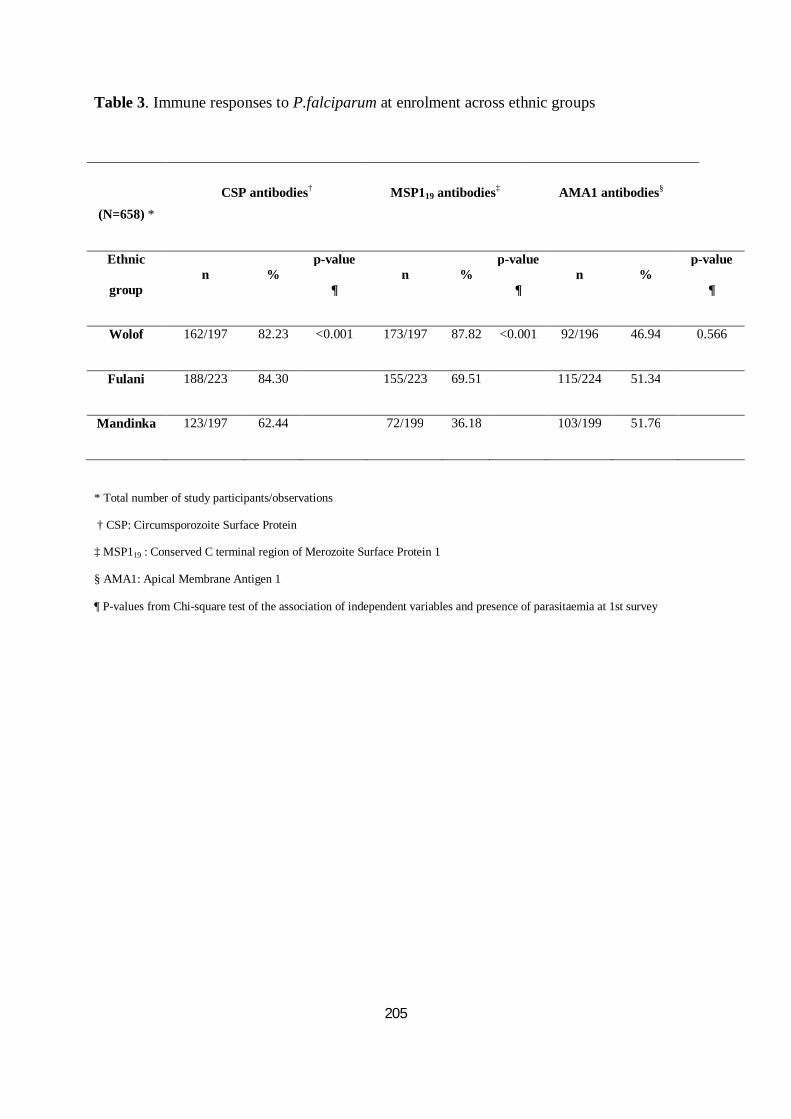
205
Table 3. Immune responses to P.falciparum at enrolment across ethnic groups
(N=658) * CSP antibodies† MSP119 antibodies‡ AMA1 antibodies§
Ethnic
group n %
p-value
¶ n %
p-value
¶ n %
p-value
¶
Wolof 162/197 82.23 <0.001 173/197 87.82 <0.001 92/196 46.94 0.566
Fulani 188/223 84.30 155/223 69.51 115/224 51.34
Mandinka 123/197 62.44 72/199 36.18 103/199 51.76
* Total number of study participants/observations
† CSP: Circumsporozoite Surface Protein
‡ MSP119 : Conserved C terminal region of Merozoite Surface Protein 1
§ AMA1: Apical Membrane Antigen 1
¶ P-values from Chi-square test of the association of independent variables and presence of parasitaemia at 1st survey
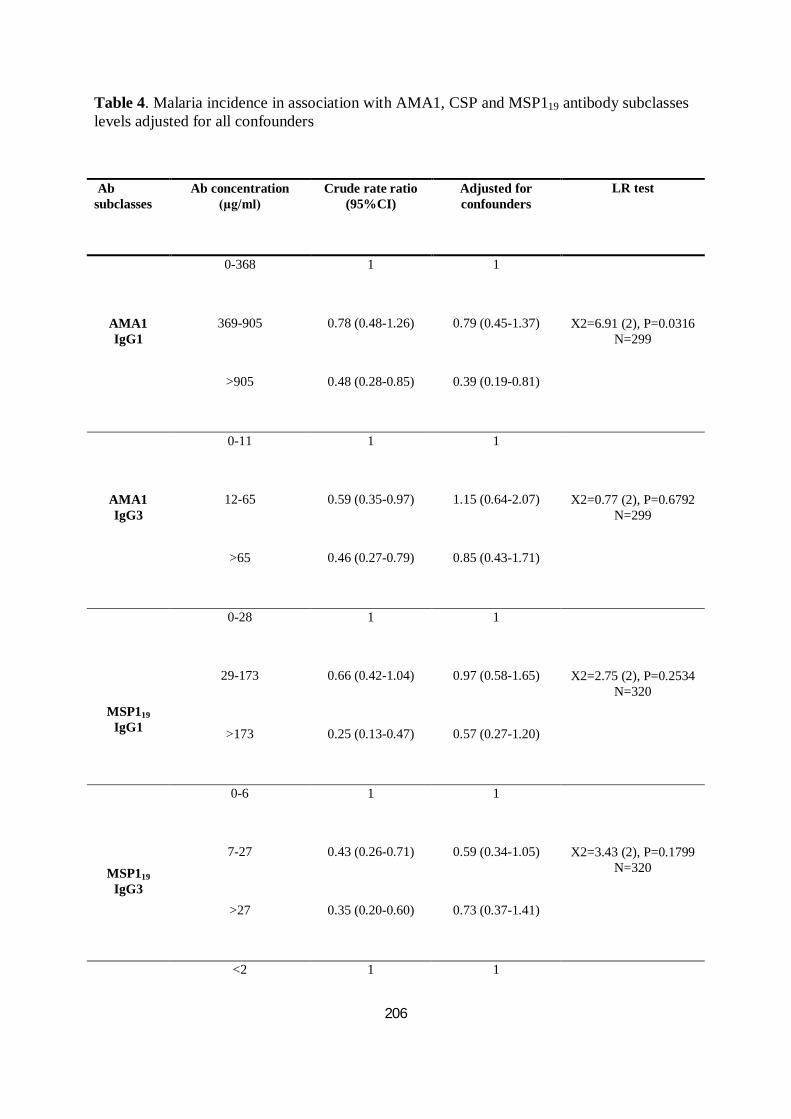
206
Table 4. Malaria incidence in association with AMA1, CSP and MSP119 antibody subclasses levels adjusted for all confounders
Ab subclasses
Ab concentration (µg/ml)
Crude rate ratio (95%CI)
Adjusted for confounders
LR test
AMA1 IgG1
0-368
1
1
X2=6.91 (2), P=0.0316 N=299
369-905
0.78 (0.48-1.26)
0.79 (0.45-1.37)
>905
0.48 (0.28-0.85)
0.39 (0.19-0.81)
AMA1 IgG3
0-11
1 1
X2=0.77 (2), P=0.6792 N=299
12-65
0.59 (0.35-0.97)
1.15 (0.64-2.07)
>65
0.46 (0.27-0.79)
0.85 (0.43-1.71)
MSP119 IgG1
0-28
1 1
X2=2.75 (2), P=0.2534 N=320
29-173
0.66 (0.42-1.04)
0.97 (0.58-1.65)
>173
0.25 (0.13-0.47)
0.57 (0.27-1.20)
MSP119 IgG3
0-6
1 1
X2=3.43 (2), P=0.1799 N=320
7-27
0.43 (0.26-0.71)
0.59 (0.34-1.05)
>27
0.35 (0.20-0.60)
0.73 (0.37-1.41)
<2 1 1

207
CSP IgG1
X2=9.2 (2), P=0.0101 N=401
2-93
0.51 (0.34-0.77)
0.68 (0.41-1.13)
>93
0.22 (0.12-0.39)
0.39 (0.20-0.76)
CSP IgG3
<2
1 1
X2=1.41 (2), P=0.4932 N=401
2-99
0.45 (0.29-0.70)
0.79 (0.47-1.34)
>99
0.59 (0.40-0.88)
1.10 (0.68-1.79)
LR: logistic regression
CI: confidence Interval
X2: Chi squared
P: p-value
N: number of individuals tested
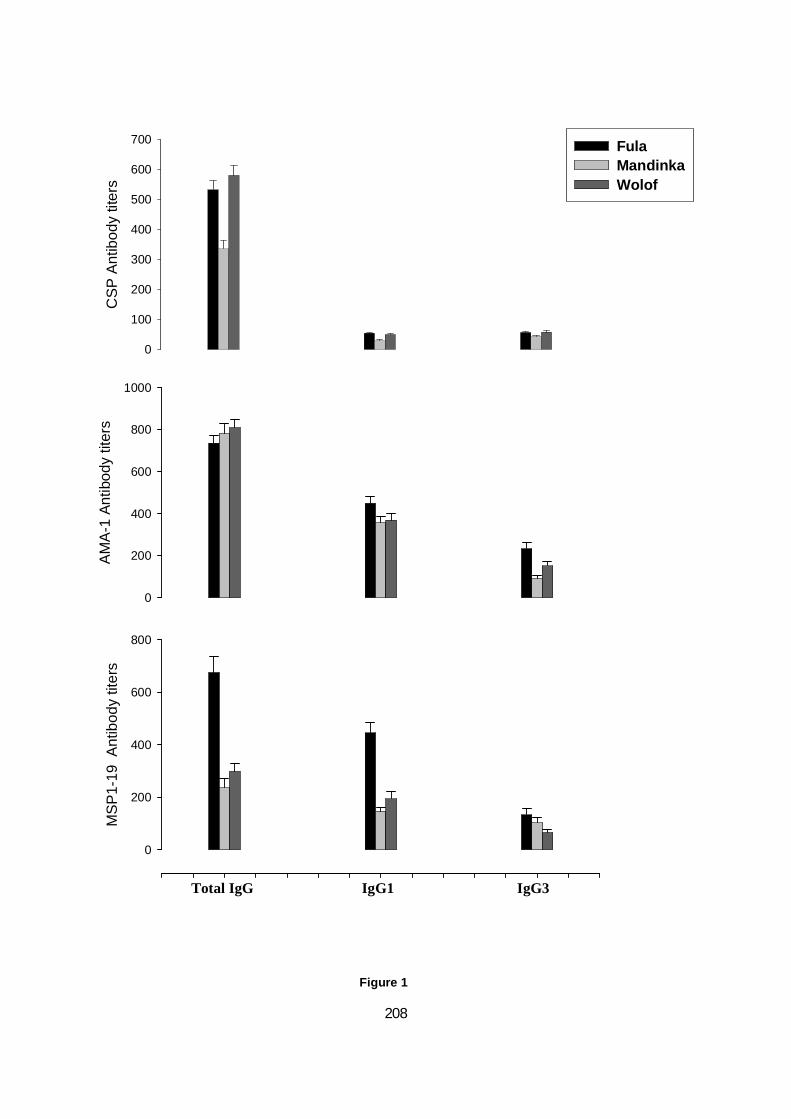
208
CS
P A
ntib
ody
titer
s
0
100
200
300
400
500
600
700
Total IgG IgG1 IgG3
MS
P1-
19 A
ntib
ody
titer
s
0
200
400
600
800
FulaMandinkaWolof
AM
A-1
Ant
ibod
y tit
ers
0
200
400
600
800
1000
Figure 1
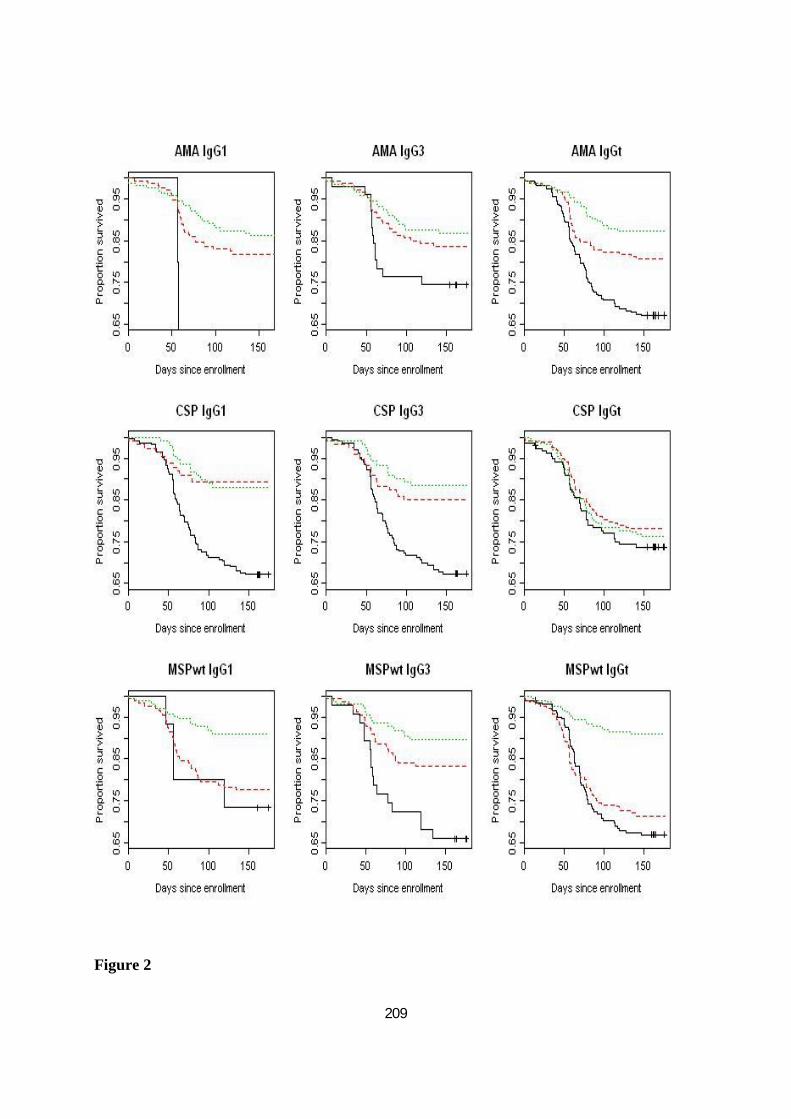
209
Figure 2
210
Figure 3A
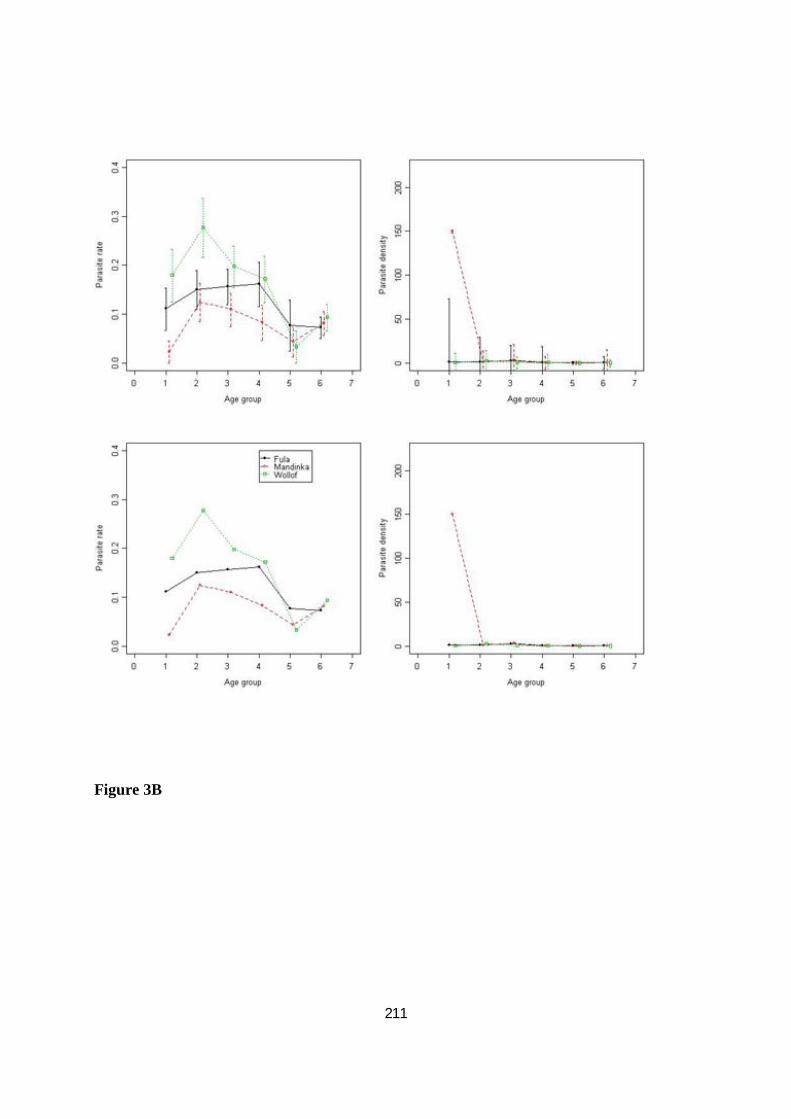
211
Figure 3B
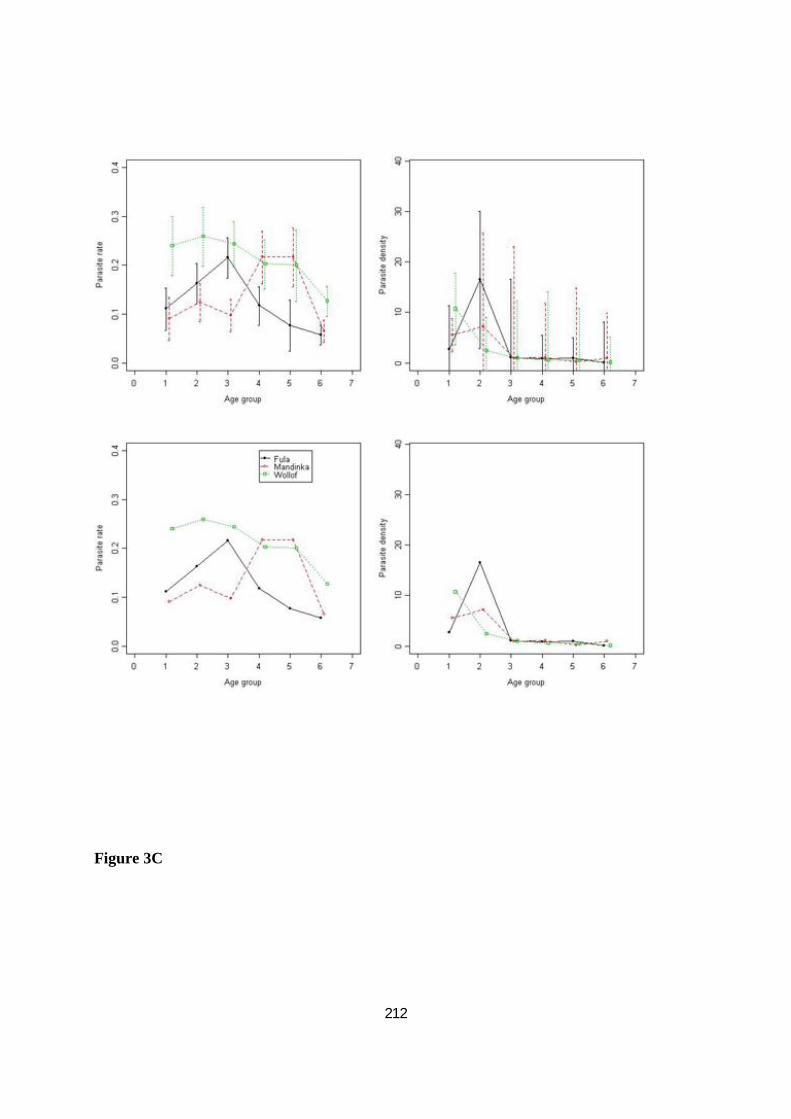
212
Figure 3C

213
Figure legends
Figure 1. A comparison of the total IgG, IgG1 and IgG3 levels to CSP, AMA1 and
MSP119 in plasma. The samples were from three groups: Fulani (black bar), Mandinka (light
gray bar) and Wolof (dark gray). Antibodies titres (pg/ml) were measured by ELISA. The
total anti-CSP IgG concentration was higher among Fulani and Wolof compared to Mandinka
samples, although there was no significant difference in the levels of IgG1 and IgG3. The
concentrations of anti-AMA1 total IgG, IgG1 and IgG3 were not significantly different
between the three groups. Higher concentrations of both anti-MSP119 total IgG and IgG1 were
observed for the Fulani compared to the Mandinka and Wolof samples.
Figure 2. Kaplan-Meier plot to show the relationship between time to first incidence of
malaria (days since enrolment) and level of specific antibodies. Dotted red (Mandinka), dotted
green (Wolof) and solid black (Fulani).
The analysis showed that high antibody levels were associated with prolonged time to the first
malaria incidence.
Figure 3A. Parsite rate and density among Fulani, Mandinka and Wolof at the time of the
inclusion in the study (July 2003).
Figure 3B. Parasite rate and density among Fulani, Mandinka and Wolof during the first
cross sectional survey (October 2003).
Figure 3C. Parasite rate and density during the second cross sectional survey (December
2003)
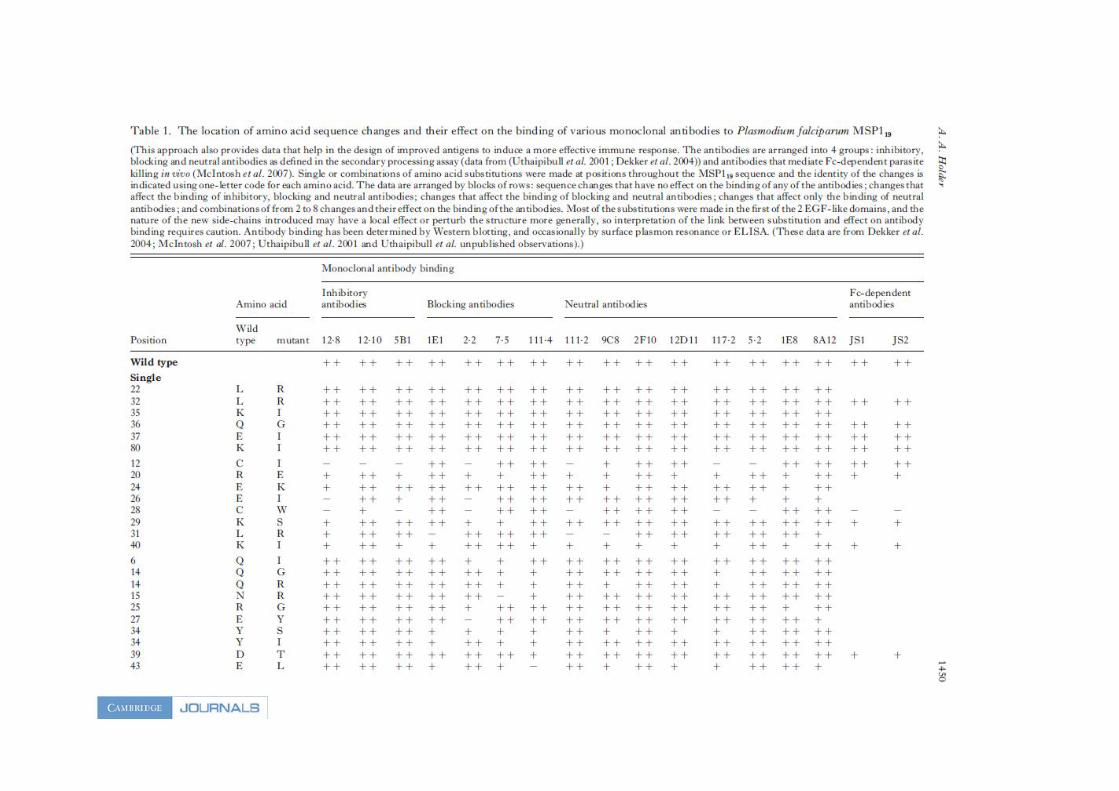
214
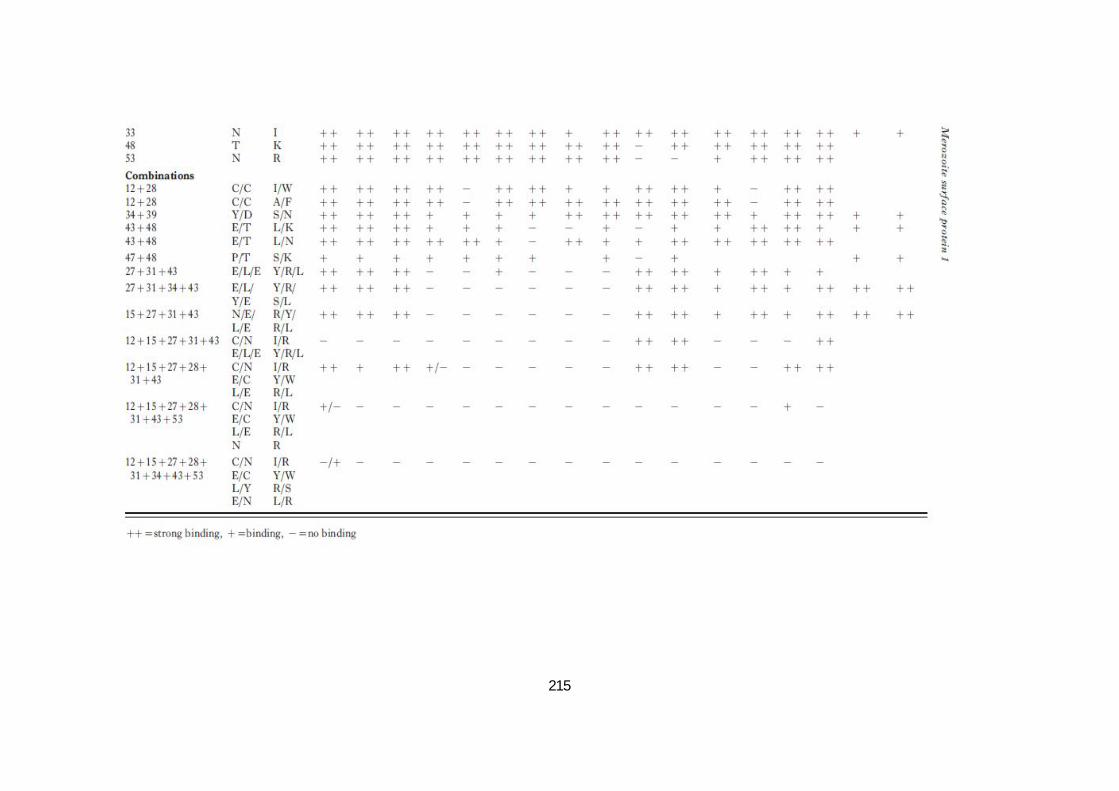
215





























