Embed Size (px)
Citation preview

UNIVERSITÉ DE GENÈVE
TRAVAIL DE MAITRISE UNIVERSITAIRE (master) EN BIOLOGIE
ETUDE FLORISTIQUE ET BIOGÉOGRAPHIQUE DES ALTITUDES SUPÉRIEURES
DE LA MONTAGNE D’AMBRE (NORD DE MADAGASCAR)
Par
Sonia Mélissa TRIGUI
Août 2010
Effectué au Laboratoire de Botanique Systématique et Biodiversité
Conservatoire et Jardin botaniques de la Ville de Genève DIRECTEUR DU TRAVAIL RESPONSABLE DU TRAVAIL Dr Laurent GAUTIER Prof. Rodolphe SPICHIGER Dpt Botanique et Biologie Végétale Dpt Botanique et Biologie Végétale Faculté des Sciences Faculté des Sciences Conservateur Herbiers Phanérogamies Directeur Honoraire CJBG ENCADREURS Louis NUSBAUMER, CJBG Dr Patrick RANIRISON, Université d’Antananarivo DBEV Dr Cyrille CHATELAIN, CJBG
© CJBG 2010

1
Table des matières Résumé ...................................................................................................................................... 7
Remerciements ......................................................................................................................... 9
1. Introduction .................................................................................................................. 11
1.1. Objectifs ......................................................................................................................... 12
1.2. Milieu d’étude ................................................................................................................ 14
1.2.1. Milieu abiotique ................................................................................................... 14
1.2.1.1. Géographie..................................................................................................... 14
1.2.1.2. Volcanisme et géologie ................................................................................. 14
1.2.1.3. Climat ............................................................................................................ 15
1.2.1.4. Hydrologie et ressources ............................................................................... 17
1.2.2. Milieu biotique ..................................................................................................... 18
1.2.2.1. Flore et végétation ......................................................................................... 18
1.2.2.2. Faune ............................................................................................................. 21
1.2.3. Milieu humain ...................................................................................................... 24
1.3. Hypothèses du projet ...................................................................................................... 24
1.3.1. Colonisation de la flore humide ........................................................................... 24
1.3.2. Endémiques locales et type de différenciation ..................................................... 26
2. Méthodologie................................................................................................................. 27
Première partie : INVENTAIRE FLORISTIQUE ................................................................... 27
2.1. Liste floristique préliminaire.......................................................................................... 27
2.2. Préparation d’une carte du massif .................................................................................. 27
2.3. Travaux réalisés sur le terrain ........................................................................................ 28
2.4. Gestion et détermination des spécimens d’herbiers ....................................................... 31
2.5. Elaboration de la liste floristique ................................................................................... 33
2.6. Système de classification utilisé..................................................................................... 34
Seconde partie : ANALYSE BIOGEOGRAPHIQUE ............................................................. 36
2.7. Répartition géographique des espèces............................................................................ 36
2.7.1. Répartition des espèces sur l’île de Madagascar................................................. 37

2
2.7.2. Répartition mondiale des espèces ........................................................................ 39
2.8. Mode de dispersion des diaspores.................................................................................. 40
2.9. Analyse de la répartition des espèces du massif ............................................................ 40
3. Résultats et interprétations ......................................................................................... 42
3.1. Prospection botanique de la Montagne d’Ambre ........................................................... 42
3.1.1. Données générales sur les échantillons d’herbier ............................................... 42
3.1.2. Chronologie des récoltes et augmentation de la connaissance de la diversité.... 43
3.1.3. Répartition saisonnière des échantillons du massif ............................................. 44
3.1.4. Répartition altitudinale des échantillons ............................................................. 45
3.1.5. Espèces nouvelles trouvées durant notre prospection botanique ........................ 46
3.1.6. Les espèces menacées du massif d’Ambre ........................................................... 47
3.2. Richesse spécifique ........................................................................................................ 48
3.2.1. Répartition de la flore selon la phylogénie .......................................................... 49
3.2.2. Richesse spécifique des familles et des genres..................................................... 50
3.2.3. Différences floristiques entre le massif d’Ambre et le Manongarivo................... 52
3.3. La flore du massif et ses affinités mondiales ................................................................. 54
3.3.1. Distribution mondiale des plantes vasculaires .................................................... 54
3.3.2. Distribution des espèces selon le spectre morphologique ................................... 56
3.3.3. Les espèces non endémiques à distribution malgache restreinte au massif
d’Ambre ............................................................................................................................. 58
3.3.4. Cas particulier des espèces situées à proximité des lacs d’anciens cratères ...... 59
3.4. La flore du massif dans le contexte malgache................................................................ 62
3.4.1. Distribution à Madagascar .................................................................................. 62
3.4.2. La flore d’Ambre et ses affinités avec deux régions du Nord de l’île .................. 64
3.5. Etude des étages de végétation au sein du massif d’Ambre........................................... 66
3.5.1. Répartition altitudinale des catégories phytogéographiques............................... 66
3.5.2. Répartition altitudinale des domaines phytogéographiques ................................ 68
3.5.3. Répartition altitudinale des groupes d’espèces appartenant à une même catégorie
phytogéographique ............................................................................................................ 70
3.5.4. Distribution altitudinale des domaines phytogéographiques............................... 71
3.6. Les taxons endémiques du massif d’Ambre................................................................... 73

3
3.6.1. Historique et évolution des connaissances........................................................... 73
3.6.2. Situation actuelle.................................................................................................. 78
3.6.3. Répartition altitudinale des espèces endémiques................................................. 78
3.6.4. Comparaisons avec trois massifs du nord de l’île ............................................... 79
3.6.5. Considérations sur les phénomènes de spéciation impliqués .............................. 80
4. Discussion...................................................................................................................... 83
4.1. Prospection botanique et richesse floristique du massif d’Ambre ................................. 83
4.2. Effets du volcanisme et recolonisation de la végétation ................................................ 83
4.3. Colonisation des altitudes supérieures par la flore humide............................................ 84
4.4. Endémiques locales et phénomènes de spéciation ......................................................... 88
5. Contraintes et analyses de la méthodologie ............................................................... 89
6. Conclusion..................................................................................................................... 91
7. Bibliographie................................................................................................................. 94
8. Annexes ....................................................................................................................... 101
8.1. Liste floristique de la Montagne d’Ambre ................................................................... 101
8.1.1. Phanérogames .................................................................................................... 101
8.1.1.1. Index des genres de Phanérogames ............................................................. 175
8.1.2. Ptéridophytes...................................................................................................... 178
8.1.3. Bryophytes, Ascomycètes lichenisés et Champignons........................................ 189
8.2. Liste des taxons endémiques du Massif de la Montagne d’Ambre.............................. 191
8.3. Liste des phanérogames du massif d’Ambre présents dans la liste rouge de l'IUCN, dans
son état actuel (17% de la flore couverte) .............................................................................. 193
8.4. Liste des espèces situées en bordure des lacs du massif .............................................. 196
8.5. Liste de quelques Mammifères, Reptiles, et Amphibiens de la Montagne d’Ambre... 198

4
Liste des tableaux Tableau 1 : Répartition altitudinale des quatre principaux domaines phytogéographiques àu massif d’Ambre et
récapitulatif des types d’espèces qui les représentent........................................................................................... 73
Tableau 2 : Nombre d’espèces endémiques dans trois régions du Nord de Madagascar..................................... 79
Tableau 3 : Analyse des taxons les plus proches des endémiques locales. ........................................................... 82
Liste des cartes Carte 1 : Situation de la Montagne d'Ambre dans l'extrémité Nord de Madagascar. .......................................... 14
Carte 2 : Les domaines phytogéogaphiques selon HUMBERT (1955). ................................................................ 19
Carte 3 : Carte de la végétation de HUMBERT & COURS DARNE (1965), Nord de Madagascar. ................... 20
Carte 4 : Représentation schématique des deux hypothèses concernant la colonisation en taxons humides des
altitudes supérieures du massif d’Ambre. ............................................................................................................. 26
Carte 5 : Carte topographique du massif d’Ambre et localisation des sites d’études. ......................................... 28
Carte 6 : Localisation des points de récoltes des échantillons qui possèdent dans leur description de la station de
récolte le vocable « prairie ». ............................................................................................................................... 60
Carte 7 : distribution de 9 espèces Autrefois considérées comme endémiques de la montagne d’ambre............. 77
Carte 8 : Hypothèse de spéciation pour les endémiques issues d’un taxon-sœur humide. ................................... 80
Carte 9 : Hypothèse de spéciation pour les endémiques issues d’un taxon-sœur sec. .......................................... 80
Carte 10 : Représentation schématique des deux hypothèses concernant la colonisation en taxons humides des
altitudes supérieures du massif d’Ambre. ............................................................................................................. 85
Liste des photos Photo 1 (gauche) : Traces de roches Basaltiques brunes du massif d’Ambre au milieu du massif de l’Ankarana
(massif calcaire, roches de couleurs grises). ........................................................................................................ 15
Photo 2 (droite) : Agrandissement de la partie ouest du massif. Les traces de coulées de laves brunes forment
deux bras (Google Earth 2009)............................................................................................................................. 15
Photo 3 : Les lacs et cascades du massif d’Ambre................................................................................................ 17
Photo 4 : Illustration de quelques organismes de la faune du massif d’Ambre. ................................................... 23
Photo 5 : Photos de récoltes fertiles effectuées..................................................................................................... 30
Photo 6 : Masque de saisie pour la phytogéographie de chaque espèce du massif d’Ambre. .............................. 36
Photo 7 : Espèces menacées dites « en danger critique d’extinction » et endémiques du massif d’Ambre. ......... 48
Liste des figures Figure 1 : Organigramme présentant le parcours des parts réalisées pour chaque récolte fertile effectuée dans le
cadre de ce projet.................................................................................................................................................. 33
Figure 2 : Nombre d’échantillons d’herbier récoltés par année au massif d’Ambre. .......................................... 44
Figure 3 : Evolution du nombre de taxons déterminés et du nombre d’échantillons d’herbier............................ 44
Figure 4 : Répartition selon les mois de l’année des échantillons d’herbier du massif d’Ambre rescensés......... 45
Figure 5 : Répartition selon des tranches altitudinales de 100m des récoltes effectuées au massif d’Ambre. ..... 46

5
Figure 6 : Les 912 espèces du massif et leur répartition dans les catégories de la liste rouge de l’IUCN........... 48
Figure 7 : La flore d’Ambre répartie dans les groupes de végétaux et leurs embranchements selon APG 3....... 49
Figure 8 : Richesse spécifique des familles présentes au massif d’Ambre............................................................ 51
Figure 9A (gauche) : Richesse spécifique des genres présents dans le massif d’Ambre. ..................................... 51
Figure 9B (droite) : Détail des 19% de genres les plus riches en espèces au massif d’AMbre. ........................... 51
Figure 10 : Distribution mondiale des phanérogames du massif d’Ambre........................................................... 54
Figure 11 : Distribution mondiale des ptéridophytes du massif d’Ambre. ........................................................... 55
Figure 12 : Pourcentage d’espèces du massif d’Ambre présentes sur les différents continents ou la région
malgache. .............................................................................................................................................................. 56
Figure 13A : Distribution des espèces du massif d’Ambre selon leur type morphologique.................................. 58
Figure 13B : répartition mondiale des espèces d’après leur type morphologique. .............................................. 58
Figure 14 : Distribution mondiale des espèces situées en bordure des lacs d’anciens cratères du massif d’Ambre
comparée à la distribution mondiale de tous les phanérogames (droite). ............................................................ 61
Figure 15 : Distribution malgache des Phanérogames du massif d’Ambre. ........................................................ 63
Figure 16 : Distribution malgache des ptéridophytes du massif d’Ambre............................................................ 63
Figure 17 : Pourcentage d’espèces du massif d’Ambre dans les 6 domaines phytogéographique de Madagascar.
.............................................................................................................................................................................. 64
Figure 18 : Répartition des espèces présentes à la Montagne d’Ambre et au Daraina selon les domaines
phytogéogaphiques de l’île. .................................................................................................................................. 66
Figure 19 : Répartition des espèces présentes à la Montagne d’Ambre et au Manongarivo selon les domaines
phytogéogaphiques de l’île. .................................................................................................................................. 66
Figure 20 : Répartition des espèces de plantes vasculaires du massif d’Ambre selon l’altitude et les domaines
phytogéographiques de Humbert. ......................................................................................................................... 68
Figure 21 : Répartition des espèces de plantes vasculaires du massif d’Ambre selon l’altitude pour montrer
l’importance de chaque domaine phytogéographique. ......................................................................................... 69
Figure 22 : Ecologie des espèces selon l’altitude. ................................................................................................ 70
Figure 23 : amplitude altitudinale des groupes d’espèces appartenant à une même catégorie phytogéographique.
.............................................................................................................................................................................. 71
Figure 24 : Année de description des 41 taxons dont la diagnose ne cite qu’un échantillon du massif d’Ambre et
comportant le radical « ambrens » et leur actuelle considération........................................................................ 75
Figure 25 : Distribution altitudinale des taxons endémiques du massif d’Ambre. ............................................... 79
Figure 26 : Répartition des taxons-sœurs des 46 endémiques du massif d’Ambre selon les 4 domaines
phytogéographiques de Humbert (1955)............................................................................................................... 81

6
Liste des abréviations, organismes et bases de données ANGAP : Association Nationale pour la Gestion des Aires Protégées de Madagascar, ancien
nom du MNP.
BD Ambre : Base de données Ambre, répertoriant tous les échantillons d’herbiers effectués à
la Montagne d’ Ambre que nous avons pu rassembler.
CJBG : Conservatoire et Jardin botaniques de la Ville de Genève.
DBEV : Département de Biologie et Ecologie Végétale, Université d’Antananarivo,
Madagascar.
JSTOR : « Organisation à but non lucratif visant à aider la communauté universitaire à
utiliser les technologies numériques pour préserver les travaux scientifiques et pour faire
progresser la recherche et l’enseignement de manière durable. ». Base de données répertoriant
des scans de spécimens types d’herbiers du monde entier. Disponible sur Internet :
<http://plants.jstor.org/>.
Index Herbariorum : Base de données répertoriant les institutions et herbiers du monde
entier ainsi que leurs membres, permettant une recherche par l’acronyme des herbiers.
Disponible sur Internet : <http://sweetgum.nybg.org/ih/>.
MNHN : Muséum National d’Histoire Naturelle de Paris, France.
MNP : Madagascar National Parks, nouveau nom de l’organisme étatique en charge
notamment de la gestion du parc national et de la réserve spéciale d’Ambre.
MBG : Missouri Botanical Garden, St Louis, Etats-Unis
Sonnerat : Base de données des spécimens d’herbiers du MNHN, disponible sur Internet :
<http://coldb.mnhn.fr/colweb/form.do?model=SONNERAT.wwwsonnerat.wwwsonnerat.ww
wsonnerat>.
TROPICOS : Base de données des spécimens d’herbiers du MBG, disponible sur Internet :
<http://www.tropicos.org/Home.aspx>.

Résumé
7
ÉTUDE FLORISTIQUE ET BIOGÉOGRAPHIQUE DES ALTITUDES SUPÉRIEURES
DE LA MONTAGNE D’AMBRE (NORD DE MADAGASCAR)
RÉSUMÉ
Ce travail de Master est le fruit d’une collaboration entre les Conservatoire et Jardin
botaniques de la Ville de Genève (CJBG), le laboratoire de Systématique Végétale et
Biodiversité de l’Université de Genève et l’Université d’Antananarivo à Madagascar. La
présente étude du massif de la Montagne d’Ambre, à l’extrême Nord de Madagascar, s’inscrit
dans le cadre des recherches entreprises depuis 1994 dans divers massifs forestiers du nord de
l’île par les CJBG.
La Montagne d’Ambre est un ancien volcan, dont les altitudes supérieures sont peuplées
d’une forêt dense humide sempervirente isolée d’autres formations végétales homologues par
un piedmont de forêt dense sèche. Malgré son statut d’aire protégée depuis plus de 50 ans, la
connaissance floristique est restée sommaire et dirigée sur l’étude de taxons ou de groupes
déterminés.
Des travaux de terrain conséquents, qui ont ciblé les altitudes supérieures du massif, ont été
entrepris en collaboration avec l’Université d’Antananarivo dans le but de dresser la première
liste floristique du massif et la première liste des espèces endémiques, puis d’effectuer une
étude phytogéographique de cette flore. La répartition de toutes les espèces du massif qui ont
pu être identifiées a fait l’objet d’une étude détaillée, tant sur le plan mondial, qu’au niveau
des domaines phytogéographiques de Madagascar. Notre étude a permis de mettre en
évidence différentes caractéristiques qui rend ce massif unique. La présence d’espèces
connues sur l’île de Madagascar uniquement au massif d’Ambre mais possédant une
distribution géographique mondiale ont été recensées. La répartition des espèces selon les
domaines phytogéographiques de l’île a montré que quatre domaines phytogéographiques
sont principalement impliqués : le Centre pour les altitudes supérieures, l’Ouest au niveau du
piedmont, ainsi que l’Est et le Sambirano à des altitudes intermédiaires. Des hypothèses
concernant la colonisation de la flore des altitudes supérieures ont pu être émises. En ce qui
concerne les endémiques locales, des hypothèses quant à leur mode de spéciation ont été
formulées et évaluées dans cette étude.
Les prospections botaniques effectuées dans le cadre de ce travail, ont non seulement permis
d’augmenter le nombre d’espèces connues pour ce massif, mais aussi permis la découverte de
nouvelles espèces pour la Science, dont une PANDANACEAE qui a récemment été publiée.

Résumé
8
MOTS-CLEFS Madagascar, Montagne d’Ambre, flore, biogéographie, endémisme, biodiversité.
CITATION TRIGUI, S. M. (2010). Etude floristique et biogéographique des altitudes supérieures de la
Montagne d’Ambre (Nord de Madagascar). Travail de Maîtrise universitaire (Master) en
Biologie. Université de Genève / Laboratoire de Systématique végétale et biodiversité et
Conservatoire et Jardin botaniques de la Ville de Genève, 199pp.

Remerciements
9
REMERCIEMENTS
De nombreuses personnes et institutions ont contribués à la réussite de ce travail de Master
aussi bien à Genève, en Suisse, qu’à Madagascar.
Tout d’abord, je remercie le laboratoire de Botanique Systématique et Biodiversité rattaché au
département de Biologie Végétale de la section de Biologie, Faculté des Sciences de
l’Université de Genève, situé au Conservatoire et Jardin botaniques de la Ville de Genève
(CJBG) où ce projet a été mis en place et effectué ; et plus particulièrement les personnes
suivantes :
- Professeur Rodolphe SPICHIGER, directeur honoraire des CJBG, professeur
responsable du travail,
- Docteur Laurent GAUTIER, conservateur principal des herbiers de Phanérogamie
CJBG, directeur de ce présent travail de Master, qui m’a donné la chance de
commencer un travail de recherche dans ce sujet passionnant et qui a encadré ce
travail par ses conseils toujours très précis et rigoureux,
- Louis NUSBAUMER, étudiant en doctorat, encadreur, qui m’a soutenue, encouragée,
motivée, et beaucoup aidée,
- Docteur Cyrille CHATELAIN, conservateur au CJBG, encadreur, toujours disponible
pour aider et répondre aux questions.
Je tiens à remercier également les personnes suivantes au sein des CJBG : Docteur Martin
CALLMANDER, Nicolas FUMEAUX, pour ses judicieux conseils, les monteurs d’herbier en
particulier Carlo MULLER qui a fait un travail magnifique, les collègues du « labo Scan-
Saisie » du Projet Mellon ainsi que tout le personnel des CJBG qui entretiennent une
ambiance très chaleureuse et accueillante dans ce cadre magnifique qu’est le jardin botanique
de Genève.
Je souhaite également remercier les personnes suivantes de l’Université d’Antananarivo à
Madagascar :
- Docteur Edmond ROGER, maître de conférences, qui a été présent sur le terrain et à
Antananarivo, toujours disponible pour d’éventuelles questions,
- Professeur Charlotte RAJERIARISON qui a également été présente sur le terrain,
- Docteur Patrick RANIRISON, encadreur du projet à Madagascar, qui a permis une
bonne mise en marche du projet de part son soutien logistique et ses précieux conseils,

Remerciements
10
qui a également été présent à plusieurs reprises sur le terrain et a toujours été
disponible pour l’encadrement et le partage de sa propre expérience. Il m’a accueillie
chaleureusement dans sa famille durant les semaines passées à la capitale et m’a fait
découvrir une autre facette de cette ville,
- Solotiana Deraharilanto RAMANDIMBIMANANA et Mialy Harindra
RAZANAJATOVO, les étudiants avec qui les travaux de terrain ont été réalisés. Ils
ont été très accueillants, ils m’ont fait découvrir leur culture avec plein
d’enthousiasme. Durant les phases de travaux de terrain, parfois difficile, notre très
bonne entente a été un moteur et ils m’ont toujours soutenue.
Je remercie aussi pour leur soutien les différents professeurs et techniciens de laboratoire de
l’Université d’Antananarivo (avec une pensée particulière à la famille de M. Lalao).
Je tiens aussi à remercier, d’une part à Antananarivo les différents étudiants que j’ai eu la
chance de rencontrer, et d’autre part à Diego-Suarez, dans le nord de Madagascar, la famille
RAVAOZANANY qui nous a généreusement accueillit entre les missions de terrain. Sans
oublier les personnes avec qui nous avons passé des fêtes de fin d’année inoubliables. Je tiens
à remercier également nos assistants présents sur le terrain, Justin, Claude, Florant ainsi que
Ferdinand et sa famille.
Les institutions suivantes ont également contribués au bon déroulement de ce travail :
- L’Initiative Sud Expert Plantes « SEP 348», ainsi que l’IRD qui ont soutenu
financièrement le projet,
- Le Fond Marc Birkigt, de l’Académie de l’Université de Genève, ainsi que la Faculté
des Sciences, département de Biologie végétale, pour leur soutien financier,
- Le Missouri Botanical Garden « MBG », à Antananarivo et Diego-Suarez pour leur
soutien logistique, et plus particulièrement Jimmy Lalao RAZAFINTSALAMA,
- Les Eaux et Forêts de Madagascar ainsi que Madagascar National Park « MNP » à
Antananarivo et Diego-Suarez, pour les autorisations de recherche, ainsi que M.
Soulouf directeur du Parc National de la Montagne d’Ambre et toute son équipe pour
leur précieux conseils et leur disponibilité.
Je tiens à remercier également, et tout particulièrement, tous les membres de ma famille. Ils
ont constamment été présents et ce malgré la distance lors des travaux de terrain.

1. Introduction
11
1. INTRODUCTION
Madagascar est situé au large des côtes du Mozambique dans l’océan Indien. Cette île
d’environ 600 000 km2 (plus de 14 fois la superficie de la Suisse) présente une diversité
biologique remarquable et est considérée comme l’un des plus importants « hot-spot » de
biodiversité de la planète (MYERS, 2000). Des estimations récentes démontrent que plus de
90% des plantes vasculaires, 92% des reptiles, 44% des oiseaux et 74% des papillons présents
à Madagascar, se retrouvent uniquement sur cette île (GOODMAN & BENSTEAD, 2003 ;
KRÜGER, 2007 ; PHILLIPSON, 2006 cité dans VENCE & AL., 2009). Une grande
variabilité au sein des facteurs environnementaux tels que le climat, la géologie, ou la
géographie ainsi que l’insularité expliquent en partie cette diversité. Toutefois cet
environnement est soumis à des pressions anthropiques grandissantes. Concernant la flore de
l’île, le nombre d’espèces de plantes vasculaires était estimé, il y a quarante ans, à 10 000
espèces alors que 7 800 était connues (HUMBERT, 1959). Dernièrement PHILLIPSON &
AL. (2006) estiment ce chiffre à 14 000 espèces, alors que nous en connaissons que 11 200.
Ces chiffres nous montrent qu’il est donc primordial d’effectuer des inventaires floristiques
pour d’une part mieux comprendre cette diversité et d’autre part pour permettre une gestion
durable de cet environnement exceptionnel.
Pour ces raisons, les CJBG ont commencé à étudier la flore et la végétation du nord de l’île.
Ces études ont été menées notamment dans les massifs du Manongarivo, du Marojejy et dans
la région du Loky-Manambato ainsi que plus récemment dans la région de la presqu'île
d’Ampasindava.
L’étude du massif de la Montagne d’Ambre s’inscrit dans ce cadre de recherche. Les CJBG et
l’Université d’Antananarivo ont collaboré pour permettre la mise en place du présent projet.
La Montagne d’Ambre est un massif volcanique situé à l’extrême nord de l’île de
Madagascar. Une flore humide en peuple les altitudes supérieures. Elle se trouve isolée
d’autres formations homologues car elle est positionnée dans une flore environnante de type
sèche. Une partie importante du massif est protégée. En effet, 18 200 hectares de forêt dense
humide composent le Parc National de la Montagne d’Ambre. De plus, 4 810 hectares de
forêts de transition entre la forêt dense humide sempervirente et la forêt sèche décidue font
partie de la Réserve Spéciale de la Forêt d’Ambre. Ce complexe d’aires protégées a été
inauguré il y a plus de 50 ans, le 28 octobre 1958. Malgré ce statut, le massif ne semble pas

1. Introduction
12
avoir fait l'objet d'un inventaire floristique, ni d’une liste des espèces endémiques, d’après nos
recherches.
L’intérêt pour ce massif remonte au 19ème siècle; les premières récoltes d'échantillons
d’herbier ont été effectuées par J. M. Hildebrandt en 1880. Ensuite, durant l’époque coloniale
les collecteurs importants sont: H. Humbert, H. Perrier de la Bâthie, R. Capuron., J. Bosser, et
A.-M. Homolle. A l’herbier de Genève, des récoltes effectuées par L. Bernardi, datant de
1967, ont également été retrouvées. Durant les années 1992 et 1993, des chercheurs de
l’Université d’Antananarivo ont étudié la végétation de la Réserve Spéciale d’Ambre au nord-
est du massif. Divers botanistes spécialisés sur des groupes particuliers ont également fait des
récoltes d’échantillons d’herbiers durant les années suivantes, notamment des équipes du
Missouri Botanical Garden avec : S.T. Malcomber et O. Andrianantoanina. Toutefois, les
études floristiques effectuées dans ce massif sont jusqu’alors restées ciblées sur des familles
ou des groupes précis comme celui des Ptéridophytes (RAKOTONDRAINBE, 2003).
A la faveur d’un financement provenant d’une initiative du Ministère Français des affaires
Etrangères : Sud Expert Plantes, ce projet a pu être mis en place. Des travaux préliminaires
ont débuté en 2006 sur le terrain par L. Gautier et C. Chatelain. Des contacts avec les autorités
locales ont été établis et quelques récoltes d’échantillons fertiles ont été réalisées. De plus,
une recherche bibliographique restreinte aux ressources disponibles sur Internet et à la
bibliothèque des CJBG a permis de créer une base de données de 59 références (TRIGUI,
2007). En outre, un travail d’élaboration d’une base de données des spécimens d’herbiers
récoltés dans le massif nommé « BD Ambre » a été réalisé (BETTINELLI, 2007). Ce travail
regroupe les informations floristiques provenant des bases de données disponibles sur Internet
de « Sonnerat » du Muséum National d’Histoire Naturelle de Paris et de « Tropicos » du
Missouri Botanical Garden (USA). Un travail de recherche a permis également de dégager les
premières tendances phytogéographiques du massif d’Ambre pour 8 familles caractéristiques
des altitudes supérieures à 800 m (CHRISTE, 2009).
1.1. OBJECTIFS
La présente étude a été menée dans le but d’augmenter les connaissances floristiques du
massif de la Montagne d’Ambre afin d’en dresser la première liste floristique et d’en chiffrer
le nombre d’endémiques locales. Une analyse phytogéographique de cette flore permettra
d’en déterminer les modes de colonisation les plus probables concernant la flore des altitudes

1. Introduction
13
supérieures, ainsi que d’en évaluer les phénomènes de spéciation concernant la formation des
endémiques locales. Les hypothèses de colonisation et de spéciation sont présentées au
chapitre 1.3, p. 24.

1. Introduction
14
1.2. MILIEU D’ÉTUDE
1.2.1. MILIEU ABIOTIQUE
1.2.1.1. GÉOGRAPHIE
Le massif de la Montagne d’Ambre présente un allongement nord-sud de 30 km de longueur,
et une distance de 15 km de largeur d’est en ouest. Il est situé entre 12°25’ et 12°43’ de
latitude sud et 49°03’ et 49°15’ de longitude est. Le sommet culmine à 1 475 m d’altitude. Le
relief est très accentué dans la partie ouest du massif. En général, les pentes sont plus douces
sur les versants est. La grande ville la plus proche de la Montagne d’Ambre est Diego-Suarez
(ou Antsiranana en malgache) qui se situe à une vingtaine de km du massif en direction du
nord-est. Les massifs et sites d’étude des CJBG les plus proches de la Montagne d’Ambre
ainsi que la ville de Diego-Suarez sont indiqués sur la carte 1 (milieu).
CARTE 1 : SITUATION DE LA MONTAGNE D'AMBRE DANS L'EXTRÉMITÉ NORD DE MADAGASCAR.
A GAUCHE : CARTE DE MADASCAR.
AU MILIEU : DÉTAIL DU NORD DE L’ÎLE, AVEC NOMS DES PRINCIPAUX MASSIFS FORESTIERS ÉTUDIÉS PAR LES
CJBG ET LOCALISATION DE LA VILLE DE DIEGO-SUAREZ.
A DROITE : VUE EN PERSPECTIVE. (SOURCE : SRTM).
1.2.1.2. VOLCANISME ET GÉOLOGIE
Une caractéristique de ce massif est son ancienne activité volcanique. C’est en effet l’un des
plus grands et des plus anciens volcans de Madagascar. Les dernières éruptions volcaniques
datent du Quaternaire, il y a moins de deux millions d’années (BRENON, 1972). Des traces
de ce passé sont encore visibles aujourd’hui sur la végétation du versant ouest du massif
(Photo 2, p. 15). La Montagne d’Ambre est recouverte d’une roche mère de type basaltique.
NN
Montagne d’Ambre
Montagne d’AmbreDaraina
Ampasindava Manongarivo
Diego-Suarez

1. Introduction
15
Des projectiles de cette roche issue du massif d’Ambre ont été retrouvés à plus de 30 km en
direction du sud dans la région de l’Ankarana (Photo 1).
PHOTO 1 (GAUCHE) : TRACES DE ROCHES BASALTIQUES BRUNES DU MASSIF D’AMBRE AU MILIEU DU MASSIF DE
L’ANKARANA (MASSIF CALCAIRE, ROCHES DE COULEURS GRISES).
PHOTO 2 (DROITE) : AGRANDISSEMENT DE LA PARTIE OUEST DU MASSIF. LES TRACES DE COULÉES DE LAVES
BRUNES FORMENT DEUX BRAS (GOOGLE EARTH 2009).
La partie minérale du sol est composée de deux types d’éléments : des tufs basaltiques (roche
formée par l’accumulation de projections volcaniques très friables) et des basaltes (roches
basiques). Les sols qui en dérivent sont légers, spongieux avec des éléments assez gros et
perméables. Ils sont riches en potassium (K), azote (N), acide phosphorique (H3PO4) et
pauvres en calcaire (CaCO3) (BARAT 1958).
1.2.1.3. CLIMAT
Le climat de Madagascar est contrôlé par deux courants aériens importants dans la zone
intertropicale. Le premier est l’alizé qui est orienté dans un axe sud-est à nord-ouest et qui
domine l'hiver austral, de juin à décembre. L’alizé humidifie principalement la côte orientale
de l’île et s’assèche par effet de foehn en redescendant vers la côte occidentale après avoir
traversé les hauts plateaux du centre. Le même phénomène s'observe à une échelle locale à la
Montagne d’Ambre: le versant oriental et la partie sommitale sont constamment arrosés et
l'effet de foehn accentue considérablement la sécheresse au niveau du versant occidental. Les
orages et fortes précipitations arrivent avec la mousson, qui fait barrière à l’alizé et se dirige
© CJBG 2010

1. Introduction
16
du nord au sud de l’île. Cet air humide s’installe pendant l'été austral. A la Montagne
d’Ambre il se fait sentir de janvier à avril.
Sur le carton bioclimatique de la carte de HUMBERT & COURS DARNE (1965) l’essentiel
de la Montagne d’Ambre est considéré comme ayant des précipitations de plus de 2 000 mm
d’eau/an, avec aucun mois sec. Les précipitations sont donc très abondantes durant toute
l’année, quelques soit la saison. La température moyenne du mois le plus frais est comprise
entre 0 °C et 10 °C. A l'échelle de cette carte, aucune transition vers les climats secs du
piedmont n’est visible.
CORNET (1974) classifie les régions climatiques selon différents étages dans sa carte
bioclimatique. Le piedmont de la Montagne d'Ambre fait partie de l'étage sec, les altitudes
intermédiaires sont rattachées à l'étage humide et les hautes altitudes à l'étage subhumide,
sous-étage à saison sèche atténuée par des brouillards. La classification des deux ceintures
supérieures surprend, tant il apparaît évident qu'il y a un accroissement massif des
précipitations avec l'altitude (voir ci-dessous). Elle reflète le parti pris des auteurs de classer
toute la végétation de montagne dans leur étage subhumide.
A l’échelle du terrain et en périphérie du massif d’Ambre, les stations météorologiques
suivantes : Sakaramy, Ambahivahibe, Anivorano, Joffreville, Andrafiamadinika et
Andranofanjava enregistrent au minimum des précipitations de 1 500 mm d’eau/an (ROSSI &
ROSSI 1998). Ce chiffre augmente d’environ 300 mm par tranche de 100 m sur le versant est,
au vent. Au Roussettes, à 1 000 m d'altitude, la moyenne est de 3 600 mm d’eau/an. Aucune
des stations de mesure n’a jamais enregistré des mois entiers sans précipitations. La saison
sèche est de 6 mois à 200 m d’altitude, de 2 mois vers 700 m d’altitude et disparaît dès
1 000 m d’altitude où se trouve la zone de brouillard permanent (ROSSI & ROSSI 1998).
Pour les températures, on remarque logiquement leur diminution avec l’augmentation de
l’altitude. A 400 m, la moyenne annuelle est de 22,8 °C, l’amplitude annuelle de 4 °C et
l’amplitude diurne de 10-12 °C selon la saison. A 1 000 m, à la station des Roussettes, la
moyenne annuelle est de 19,6 °C, l’amplitude annuelle reste à 4 °C, et l’amplitude diurne
reste également à 10-12 °C. Toujours à cette altitude, le mois le plus froid est juillet avec
17,6 °C et le mois le plus chaud présente une moyenne de température de 21 °C. Le maximum
absolu a été atteint en février (29,8 °C), et le minimum absolu en juillet (9 °C) (ROSSI &
ROSSI 1998).

1. Introduction
17
Un phénomène climatique qui a une grande importance sur la végétation est la période
cyclonique. L’île de Madagascar se trouve sur la « ceinture » des cyclones tropicaux, qui
emmagasinent leur énergie avec la température chaude de la surface de la terre et l’humidité
venant de l’évaporation de l’océan (JURY, 2003). Selon JURY (2003) il y a, en moyenne, 10
cyclones tropicaux qui touchent l’île de Madagascar chaque année, principalement durant la
période de Décembre à Mars. Les dégâts les plus importants sont ressentis sur la côte Est de
l’île, et sont principalement engendrés par des précipitations massives qui entraînent
notamment des inondations et des glissements de terrain. Les zones pluvieuses peuvent
« s’étendre à plusieurs centaines de kilomètres du centre du météore, provoquant sur des
régions entières des crues destructrices » (RAVET, 1952). BARAT (1958) cite un cyclone
tropical qui, en 1937, aurait produit un éboulement de pans entiers au massif de la Montagne
d’Ambre.
1.2.1.4. HYDROLOGIE ET RESSOURCES
Le massif compte 6 lacs de cratère, 32 cours d’eau annuels et 36 cours d’eau saisonniers. De
plus, on y dénombre 9 cascades ainsi que plusieurs prairies inondables formant des lacs
temporaires de diverses grandeurs en pleine forêt (photo 3).
PHOTO 3 : LES LACS ET CASCADES DU MASSIF D’AMBRE.
DE GAUCHE À DROITE : LAC MAUDIT, LAC TEXIER ET LA CASCADE DES ROUSSETTES.
Les ressources hydrauliques qui résultent de ce massif sont un apport fondamental pour toute
la région, car elles permettent d’irriguer non seulement les cultures environnantes mais
également les villages et villes du nord de Madagascar. Ce massif, dont les altitudes
supérieures sont composées d’une forêt dense humide sempervirente, est donc d’une
importance incontestable pour la stabilité des ressources hydrauliques de cette région.
© CJBG 2010 © CJBG 2010 © CJBG 2010

1. Introduction
18
1.2.2. MILIEU BIOTIQUE
1.2.2.1. FLORE ET VÉGÉTATION
D’une manière générale, sur l’île de Madagascar « ce n’est pas du Nord au Sud que s’accusent
le plus fortement les différences, tant physionomiques que floristiques, entre les types de
végétation autochtone liés aux bioclimats, mais suivant les profils du relief, d’Est en Ouest »
(HUMBERT, 1965). Cette observation semble être également valable pour la flore et la
végétation du massif d’Ambre qui est une chaîne de montagne avec un allongement Nord-
Sud. Ainsi, la végétation peut être modifiée selon les versants Est-Ouest du massif d’Ambre.
La flore et la végétation du massif de la Montagne d’Ambre ont été classées différemment
selon les auteurs et leurs modèles.
Du point de vue phytogéographique, PERRIER DE LA BÂTHIE (1936) a proposé une
classification de Madagascar en cinq régions, qui deviendront pour HUMBERT (1955) des
domaines (carte 2, p. 19). La flore sèche est constituée du Domaine de l’Ouest et de celui du
Sud. La flore humide comprend: le Domaine oriental (au-dessous de 800 m), le Domaine
central (altitudes supérieures à 800 m), le Domaine des Hautes Montagnes (au-dessus de
2 000 m) et le Domaine du Sambirano (petite zone du nord-ouest, humide, comprise entre 0 et
800 m d’altitude). Concernant le domaine du Centre, il est subdivisé en trois sous-domaines
(Centre-Nord, Centre-Moyen et Centre-Sud), par deux seuils où l’altitude s’abaisse : seuil de
Mandritsara (au Nord) et le seuil de Menaharaka (au Sud). De plus HUMBERT (1965)
distingue pour les versants ouest des trois sous-domaines du Centre un secteur des pentes
occidentales, une « forêt basse sclérophylle, série à Uapaca Bojeri et Chlaenacées ». La
Montagne d’Ambre est comprise dans le Domaine du Centre sous-domaine du Centre-Nord
pour les altitudes supérieures à 800 m, et le piedmont se rattache par contre au domaine de
l’Ouest. Une description détaillée des domaines phytogéographiques selon Humbert est
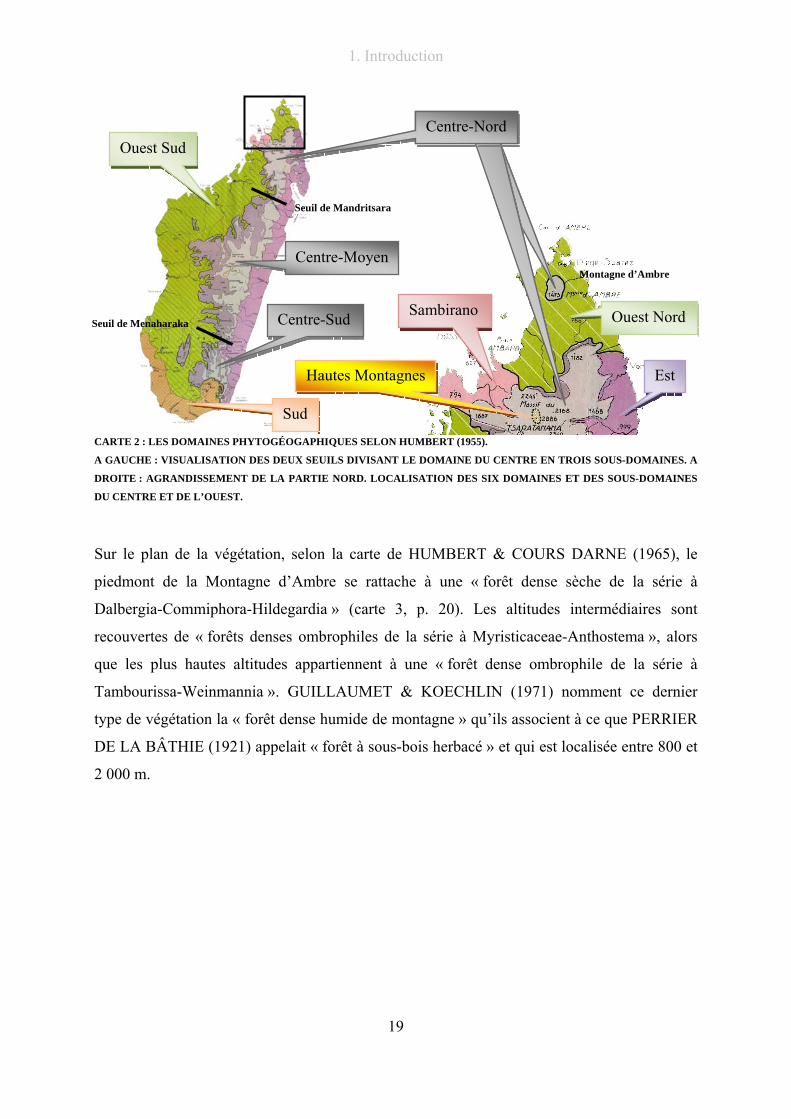
proposée dans la partie méthodologie de ce travail (chapitre 2.7.1, p. 37).
1. Introduction
19
CARTE 2 : LES DOMAINES PHYTOGÉOGAPHIQUES SELON HUMBERT (1955).
A GAUCHE : VISUALISATION DES DEUX SEUILS DIVISANT LE DOMAINE DU CENTRE EN TROIS SOUS-DOMAINES. A
DROITE : AGRANDISSEMENT DE LA PARTIE NORD. LOCALISATION DES SIX DOMAINES ET DES SOUS-DOMAINES
DU CENTRE ET DE L’OUEST.
Sur le plan de la végétation, selon la carte de HUMBERT & COURS DARNE (1965), le
piedmont de la Montagne d’Ambre se rattache à une « forêt dense sèche de la série à
Dalbergia-Commiphora-Hildegardia » (carte 3, p. 20). Les altitudes intermédiaires sont
recouvertes de « forêts denses ombrophiles de la série à Myristicaceae-Anthostema », alors
que les plus hautes altitudes appartiennent à une « forêt dense ombrophile de la série à
Tambourissa-Weinmannia ». GUILLAUMET & KOECHLIN (1971) nomment ce dernier
type de végétation la « forêt dense humide de montagne » qu’ils associent à ce que PERRIER
DE LA BÂTHIE (1921) appelait « forêt à sous-bois herbacé » et qui est localisée entre 800 et
2 000 m.
Centre-NordCentre-Nord
Est
Sambirano
Sud
Hautes Montagnes
Ouest Nord
Montagne d’Ambre
Ouest Sud
Centre-Moyen
Centre-Sud
Seuil de Mandritsara
Seuil de Menaharaka
Centre-Nord

1. Introduction
20
CARTE 3 : CARTE DE LA VÉGÉTATION DE HUMBERT & COURS DARNE (1965), NORD DE MADAGASCAR.
La Montagne d’Ambre est rattachée aux formations forestières d’altitude de Madagascar dont
GUILLAUMET & KOECHLIN (1971) donnent une description plus précise, basée sur la
morphologie des arbres. La « forêt dense humide de montagne » culmine généralement à 5 m
de haut et est composée d’arbres à port ramifié bas et à feuilles à tendance sclérophylle. Les
strates arborescentes et arbustives sont réduites alors que la strate herbacée est dense. Les
épiphytes, mousses et lichens sont très abondants. Les auteurs différencient la « forêt dense
sclérophylle de montagne », ou « sylve à lichens » selon PERRIER DE LA BÂTHIE (1921),
par sa plus petite taille, environ 10 m de hauteur. Les arbres sont très ramifiés et ont des
petites feuilles coriaces. La strate moyenne arbustive est dans ce cas constituée de nombreuses
Composées et d’Ericacées. Les mousses et lichens forment parfois une strate continue au
niveau du sol et sont également très abondants en tant qu’épiphytes. Enfin, les parties du
massif les plus hautes sont nommées « fourré de montagne » par GUILLAUMET &
KOECHLIN (1971) ou « brousse à éricoïde » par PERRIER DE LA BÂTHIE (1921). C’est
une végétation buissonnante de moins de 6 m de haut. La microphyllie est caractéristique et se
retrouve chez les Ericacées mais également chez les Composées, Rubiacées, etc.
Sur la carte de FARAMALALA & al. (1998), dressée à partir d'images du satellite
LANDSAT, la partie centrale de la Montagne d’Ambre est uniformément rattachée à la
végétation de forêt dense humide sempervirente saisonnière.
Forêt dense sèche
série à Dalbergia-Commiphora-
Hildegardia (<800 m)
Forêt dense ombrophile
série à Myristicaceae-Anthostema
(altitudes intermédiaires)
Forêt dense ombrophile
série à Tambourissa-Weinmannia / versant
occidental: forêt basse sclérophylle Série à
Uapaca Bojeri et Chlaenacées (>800 m)

1. Introduction
21
Pour finir, selon les dernières cartes proposées par DU PUY & MOAT (1996) combinant la
végétation primaire restante et la géologie, le piémont est rattaché à une forêt dense humide
sempervirente de basse altitude tandis que les altitudes supérieures font parties d’une forêt
dense humide sempervirente de moyenne altitude, le tout reposant sur un sol volcanique de
laves (basaltes et gabbros).
1.2.2.2. FAUNE
Mammifères
- Primates
En ce qui concerne les lémuriens, sept espèces sont recensées d’après MITTERMEIER & AL.
(2007) et GLAW & VENCES (1994) se sont : Phaner electromontris Groves & Tattersall,
Cheirogaleus major É. Geoffroy, Eulemur sanfordi Archbold, Eulemur coronatus Gray,
Daubentonia madagascariensis Gmelin, Lepilemur septentrionalis Rumpler & Albignac (sud
du massif uniquement) et Microcebus tavaratra Rasoloarison, Goodman & Ganzhorn.
L’Aye-aye (Daubentonia madagascariensis Gmelin) est protégé depuis 1927 à Madagascar. Il
a été localisé dans de nombreuses aires protégées et notamment dans le Parc National ainsi
que dans la Réserve Spéciale de la Montagne d’Ambre (STERLING, 2003).
- Chiroptères
D’après GLAW & VENCES (1994), les chiroptères sont parmi les groupes de mammifères
malgaches les moins bien connus, car aucune révision récente n’a été effectuée. Pour cette
raison, nous n’avons pas le nombre d’espèces de chiroptères présentes dans le massif
d’Ambre.
- Carnivores
Les deux carnivores connus de Madagascar ont été recensés dans le massif d’Ambre. Il s’agit
de : Galidia elegans Saint-Hilaire subsp. dambrensis Tate & Rand et Cryptoprocta ferox
Bennett (RASOLONANDRASANA, 1994).
- Rodentiens
Eliurus majori Thomas et Eliurus minor Major sont présent dans le massif d’Ambre d’après
CARLETON (2003), ainsi que le rat invasif Rattus rattus L.

1. Introduction
22
- Afrosoricidés
Durant nos missions de terrain nous avons aperçu le Tenrec ecaudatus Schreber, qui est une
espèce commune à Madagascar.
Une étude sur les musaraignes de différents parc nationaux dénombre six espèces à la
Montagne d’Ambre dont la plus répandue est Microgale drouhardi Grandidier (JENKINS,
2003).
- Artiodactyles
Le Potamochère (omnivore) est le plus grand mammifère de Madagascar. Il se trouve
également sur le continent africain. Des troupeaux de zébus qui traversent l’aire protégée
laissent des traces dans la forêt du massif d’Ambre.
Oiseaux
D’après la « Check List of the Birds of Amber Mountain National Park, Madagascar » de
LANGRAND (1995), 77 espèces d’oiseaux ont été enregistrés dans ce massif. Sur ce total, 70
sont visibles durant toute l’année à la Montagne d’Ambre, 4 espèces migrent sur le continent
africain durant l’hiver austral et 3 espèces migrent localement dans la région Nord de l’île et
font quelques visites à la Montagne d’Ambre.
Deux espèces recensées ne nidifient pas à la Montagne d’Ambre. Il y a le Falco eleonorae
Gené qui est un exemple d’un « grand migrateur » (COLLIN, 2010 en ligne). Il nidifie dans
les îles de la méditerranée et migre jusqu’à Madagascar et notamment à la Montagne d’Ambre
durant l’hiver boréal. De plus, le Apus affinis J.E. Gray, est une espèce répandue
principalement en Afrique. OLIOSO (2010 en ligne) mentionne que cette espèce « a besoin de
plans d'eau permanents pour pouvoir boire ».
Reptiles et Amphibiens
La Montagne d’Ambre est une des régions de Madagascar ayant une grande diversité de
reptiles (RAXWORTHY, 2003), car des conditions écologiques favorables à leur
développement y sont présentes. Une compilation des espèces de reptiles et d’amphibiens
citées du massif de la Montagne d’Ambre et recensées par GLAW & VENCES (1994) et
RAXWORTHY & NUSSBAUM (1994) a été effectuée et est donnée en annexe 8.5, p. 198.
Nous pouvons donc dénombrer pour le massif d’Ambre 46 espèces de reptiles dont 9 espèces
de lézards, 15 espèces de serpents, 10 espèces de caméléons, 11 espèces de la famille de
Gekkonidae (Gecko), 1 espèce de la famille des Gerrhosauridae. Le nombre d’espèces

1. Introduction
23
d’amphibiens est de 24 dans le massif d’Ambre. Aucune espèce d’iguanes, ni de tortues ne
sont recensées à la Montagne d’Ambre.
Poissons
Une espèce Pachypanchax sakaramyi Holly est commune dans le « Grand lac » du massif
d’Ambre (GLAW & VENCES, 1994). Des anguilles ont également été aperçues dans la
rivière sortant du « lac Texier » au sud-est du massif vers le village d’Antsalaka durant nos
travaux de terrain.
Invertébrés
Les invertébrés aquatiques ont une forte tendance au micro-endémisme dans les forêts
humides (Montagne d’Ambre, Tsaratanana, etc.). Toutefois les causes de ce processus sont
mal connues (ELOUARD & GIBON, 2003). Des listes d’espèces de divers groupes
d’invertébrés présents à Madagascar ont été publiées notamment pour : les Gastéropodes
(PEARCE, 2003) ; les Mille-pattes (ENGHOFF, 2003) ; les Collemboles (BETSCH, 2003) ;
les Papillons (LEES & AL., 2003) et les Taons (CHAINEY, 2003).
PHOTO 4 : ILLUSTRATION DE QUELQUES ORGANISMES DE LA FAUNE DU MASSIF D’AMBRE.
DE GAUCHE À DROITE DEPUIS LE HAUT : EULEMUR SANFORDI ARCHBOLD, UN CAMÉLÉON, BOOPHIS SP., UNE
LIBELLULE.
© CJBG 2010 © CJBG 2010
© CJBG 2010© CJBG 2010

1. Introduction
24
1.2.3. MILIEU HUMAIN
A la lisière de la forêt du massif d’Ambre il y a 38 villages, ce qui représente environs
40 000 habitants (RAKOTONDRASANA, 1997). La population de Madagascar est composée
de différents groupes ethniques. Dans les zones périphériques du massif on retrouve
principalement les « Antakarana » et les « Betsimisaraka ». Ces populations vivent
notamment de la culture du riz, qui est l’élément de base de leur nourriture. La culture sur
brûlis, appelée localement « Tavy » est une pratique courante à Madagascar. Toutefois ces
feux ne sont pas toujours maîtrisés et peuvent parfois empiéter sur la végétation environnante
et créer des répercussions irréversibles sur la faune ou la flore. La culture du khat (Catha
edulis Forssk. CELASTRACEAE) dont les feuilles sont mâchées comme du tabac à chiquer,
est une source de revenu importante dans cette région. Des canaux creusés par les populations
locales sont observés à l’intérieur du parc national pour arroser les cultures environnantes.
Certains villageois vivent de l’élevage de bovins et des troupeaux traversent les forêts du
massif. En lisière de l’aire protégée, dans la partie nord et sud-est du massif, les pressions
anthropiques sont importantes et diverses, notamment pour l’acquisition de ressources
alimentaires (récolte de miel, pêche), et des coupes de bois d’œuvre laissent des traces qui
sont visibles sur la végétation. De plus, des feux de forêts ont également été observés à la
limite du parc. L’environnement se retrouve ainsi modifié par l’action de l’homme et la
végétation primaire est secondarisée. Malgré la présence des autorités locales, les limites de
l’aire protégée sont parfois franchies, pour accéder aux diverses ressources offertes par ce
massif imposant de la Montagne d’Ambre.
1.3. HYPOTHÈSES DU PROJET
1.3.1. COLONISATION DE LA FLORE HUMIDE
La Montagne d’Ambre n’est pas le seul massif forestier dont la flore humide est isolée au sein
d’une flore sèche du domaine phytogéographique de l’Ouest. Dans la partie nord du domaine
de l’Ouest nous retrouvons notamment les plus hauts sommets de la région du Daraina et dans
la partie sud du domaine de l’Ouest : le massif de l’Anavelona ou de l’Isalo.
Deux différentes classifications de cette flore humide du massif d’Ambre ont été retrouvées
dans la littérature. D’une part, celle de HUMBERT (1965) qui rattache la flore humide de ce
massif au domaine phytogéographique du Centre, sous-domaine du Centre-Nord, et d’autre
part celle de CORNET & GUILLAUMET (1976) qui en font un secteur d’altitude du

1. Introduction
25
domaine de l’Ouest. Ils expliquent que « les montagnes d’Ambre et de l’Anavelona qui ont
certes, nombre d’espèces communes avec le Domaine du Centre, n’en ont pas moins une
grande dépendance vis-à-vis de la flore occidentale et devraient vraisemblablement être
regardées comme des secteurs d’altitude du Domaine de l’Ouest ».
Ces deux classifications de la flore humide du massif d’Ambre sous-tendent implicitement
deux hypothèses concernant la colonisation des altitudes supérieures :
- La première (flèches bleues, carte 4, p. 26) qui se trouve liée au mode de classification
de HUMBERT (1965) suppose une dispersion de taxons humides depuis d’autres
massifs humides d’altitude du nord de l’île possédant une flore typique du domaine du
Centre comme le Tsaratanana, le Manongarivo ou le Marojejy. Les diaspores des
espèces présentes dans ces régions auraient été transportées sur de longues distances
par différents moyens comme la zoochorie, l’anémochorie, ou encore qu’il y aurait eu
des connexions directes entre ces massifs humides durant un passé climatique plus
humide que la situation actuelle. Il n’est pas à exclure également que la région du
Loky-Manambato ait pu faciliter la dispersion à longue distance en servant de relais,
au vu de sa position géographique intermédiaire entre les massifs humides du nord de
la chaîne principale de l’île et la Montagne d’Ambre à l’extrême nord.
- La seconde (flèche orange, carte 4, p. 26), qui reprend la classification de CORNET &
GUILLAUMET (1976), fait intervenir une colonisation à partir des taxons de la flore
sèche du piedmont par une adaptation progressive aux conditions environnementales
plus humides présentes dans les altitudes supérieures du massif, soit une colonisation
dite « de proche en proche ».

1. Introduction
26
CARTE 4 : REPRÉSENTATION SCHÉMATIQUE DES DEUX HYPOTHÈSES CONCERNANT LA COLONISATION EN
TAXONS HUMIDES DES ALTITUDES SUPÉRIEURES DU MASSIF D’AMBRE.
CARTE DU NORD DE L’ÎLE VUE DE PROFIL ET EN RELIEF, MONTRANT LES 4 DOMAINES PHYTOGÉOGRAPHIES
IMPORTANTS DU NORD. LES FLÈCHE BLEUES REPRÉSENTENT L’HYPOTHÈSE DE COLONISATION PAR LONGUE
DISTANCE DES DIFFÉRENTS DOMAINES HUMIDES DE L’ÎLE SITUÉS PLUS AU SUD. LA FLÈCHE ORANGE
REPRÉSENTE L’HYPOTHÈSE D’UNE ADAPTATION PROGRESSIVE DES TAXONS DE LA FLORE SÈCHE DU PIEDMONT
AUX CONDITIONS HUMIDES TROUVÉES EN ALTITUDE. DESSIN DE C. CHATELAIN.
1.3.2. ENDÉMIQUES LOCALES ET TYPE DE DIFFÉRENCIATION
D’après SPICHIGER & AL. 2004, « selon le concept de l’espèce que l’on admet, la
spéciation sera définie comme un processus adaptatif induisant des barrières contre le flux de
gènes entre des populations étroitement liées, par le développement de mécanismes
d’isolement à la reproduction ou encore, plus généralement, comme un processus de
changements génétiques produisant la naissance d’une nouvelle espèce. » Concernant les
endémiques locales de la Montagne d’Ambre et leur mode de spéciation, nous avons émis
deux hypothèses concurrentes qui sont liées à deux types d’isolement reproductif :
- D’une par la différenciation s’est effectuée à partir d’un taxon issu de la flore humide,
ce qui suppose un isolement géographique puis une dérive génétique.
- D’autre part, la différenciation s’est effectuée à partir d’un taxon issu de la flore sèche
du piedmont, adapté aux conditions humides d’altitude, ce qui suppose dans ce cas un
isolement écologique puis une dérive génétique.
Une recherche des taxons-sœurs (= espèce ou genre le plus proche retrouvé en consultant les
diagnoses des endémiques), ainsi qu’une analyse de leur répartition à Madagascar a été
effectuée.
Montagne d’Ambre
30 km

2. Méthodologie
27
2. MÉTHODOLOGIE La méthodologie comprend cinq points principaux. Premièrement, des travaux de terrain ont
été effectués au sein de cette forêt tropicale humide durant une période de 9 mois, pour y
réaliser un inventaire floristique des altitudes supérieures à 800 m d’altitude. Des récoltes
intenses d’échantillons d’herbiers ont été réalisées. Deuxièmement, les échantillons récoltés
ont été déterminés à l’herbier de Genève, et certains spécialistes de taxons malgaches ont été
consultés. Troisièmement, une liste floristique a été établie en ajoutant les récoltes de ce
présent projet aux plus anciennes récoltes qui ont pu être recensées. Quatrièmement, une
analyse de la distribution géographique de tous les taxons déterminés a été effectuée pour
l’ensemble des plantes vasculaires recensées. Enfin, cinquièmement, une analyse des données
a été effectuée pour répondre aux hypothèses de cette recherche et présenter la flore de la
Montagne d’Ambre. Ces analyses ont par la suite permis la rédaction de ce travail mais
également d’articles scientifiques.
PREMIÈRE PARTIE : INVENTAIRE FLORISTIQUE
2.1. LISTE FLORISTIQUE PRÉLIMINAIRE
Une première liste floristique a été constituée à partir des échantillons d’herbiers recensés
pour le massif d’Ambre dans les bases de données disponibles sur Internet: Sonnerat (MNHN,
Paris) et Tropicos (MBG, Missouri). Cette liste a été complétée par des récoltes retrouvées à
l’herbier de Genève (notamment un certain nombre de récoltes de L. Bernardi), et des
échantillons cités dans la Flore de Madagascar et des Comores (HUMBERT & AL., 1936).
Une première BD Ambre a donc été créée avant les travaux de terrain, ce qui a permis à
l’équipe d'approcher l'historique de la prospection de la flore du massif d’Ambre.
2.2. PRÉPARATION D’UNE CARTE DU MASSIF
Une cartographie du massif a été crée à l’aide du Logiciel ArcView (GIS version 3.2), ce qui
a permis de superposer différentes cartes topographiques du massif, dont une vue aérienne tiré
de Google Earth. Les points GPS des récoltes antérieures à notre étude ont également été
ajoutés. Les zones d’études ont été choisies après superpositions de ces différentes données.
Des cartes topographiques du massif ont été imprimées et ont pu être utilisées sur le terrain.
2. Méthodologie
28
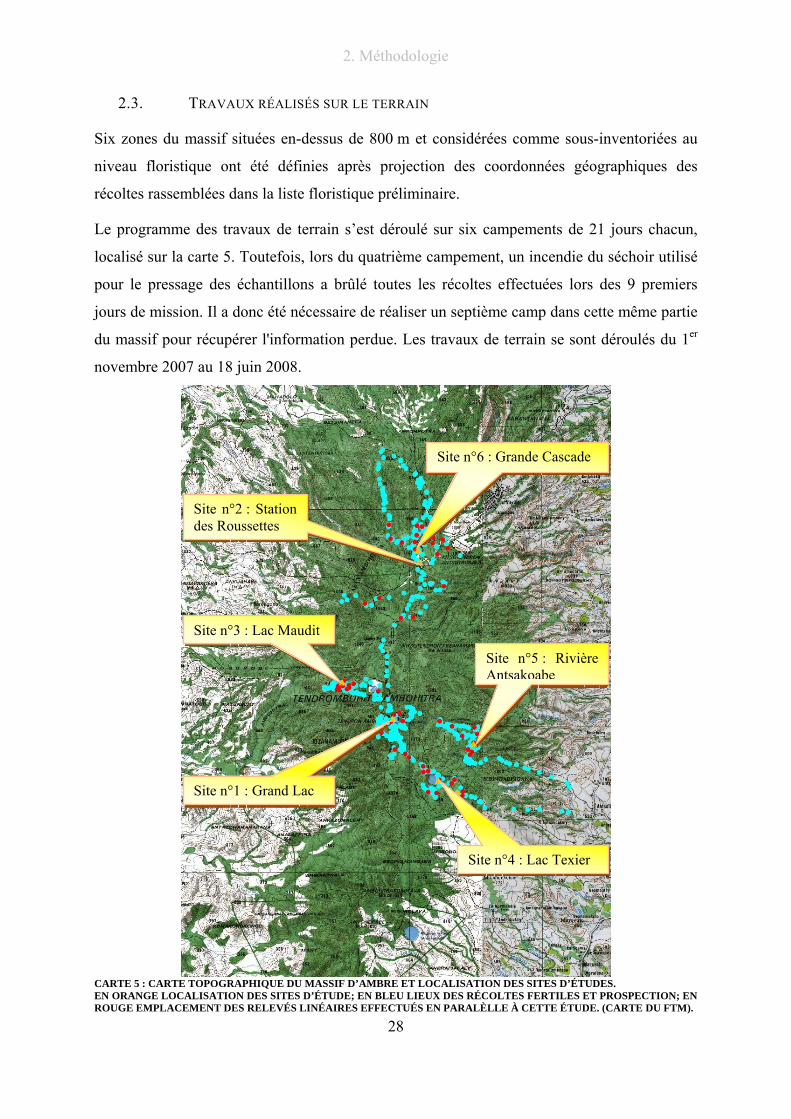
2.3. TRAVAUX RÉALISÉS SUR LE TERRAIN
Six zones du massif situées en-dessus de 800 m et considérées comme sous-inventoriées au
niveau floristique ont été définies après projection des coordonnées géographiques des
récoltes rassemblées dans la liste floristique préliminaire.
Le programme des travaux de terrain s’est déroulé sur six campements de 21 jours chacun,
localisé sur la carte 5. Toutefois, lors du quatrième campement, un incendie du séchoir utilisé
pour le pressage des échantillons a brûlé toutes les récoltes effectuées lors des 9 premiers
jours de mission. Il a donc été nécessaire de réaliser un septième camp dans cette même partie
du massif pour récupérer l'information perdue. Les travaux de terrain se sont déroulés du 1er
novembre 2007 au 18 juin 2008.
CARTE 5 : CARTE TOPOGRAPHIQUE DU MASSIF D’AMBRE ET LOCALISATION DES SITES D’ÉTUDES. EN ORANGE LOCALISATION DES SITES D’ÉTUDE; EN BLEU LIEUX DES RÉCOLTES FERTILES ET PROSPECTION; EN ROUGE EMPLACEMENT DES RELEVÉS LINÉAIRES EFFECTUÉS EN PARALÈLLE À CETTE ÉTUDE. (CARTE DU FTM).
Site n°6 : Grande Cascade
Site n°2 : Station des Roussettes
Site n°3 : Lac Maudit
Site n°1 : Grand Lac
Site n°5 : Rivière Antsakoabe
Site n°4 : Lac Texier

2. Méthodologie
29
Pour chacun des camps effectués dans ce massif d’Ambre, les premiers jours ont été
consacrés à une reconnaissance des alentours du camp. Ces journées ont permis d’ouvrir des
chemins et de repérer les sites potentiels pour les relevés linéaires de la végétation effectués
en parallèle à cette étude floristique (RAZANAJATOVO, 2009 ; RAMANDIMBIMANANA,
2009).
La recherche de récoltes fertiles s’est déroulée de manière aléatoire en ouvrant des chemins au
travers de la forêt, et en essayant de trouver et de repérer, à l’aide de jumelles, toutes les
plantes fertiles. Des récoltes de bryophytes et de lichens ont aussi été réalisées en plus des
ptéridophytes et phanérogames. Il y a toutefois des groupes ou familles pour lesquelles le
repérage des fleurs de par leurs grandeurs, couleurs, ou encore hauteurs, est plus facile à
distinguer que d’autres.
Une gaule ou échenilloir permettant d’atteindre les parties fertiles des grands arbres, ainsi
qu’une presse permettant le pressage sur le terrain des récoltes fragiles nous ont accompagnés
durant chaque journée. La liste du matériel de terrain utilisé lors des prospections et des
récoltes itinérantes est la suivante :
Matériels porté durant la journée
- Pour la récolte : un carnet de récolte, une gaule (ou échenilloir), un sécateur, un
appareil photo, une paire de jumelle, une loupe.
- Pour noter les différents paramètres stationnels : un GPS (Global Positioning System),
un altimètre, une boussole.
- Pour transporter les récoltes : un sac de jute (gony) pour les récoltes non fragiles, une
presse pour les récoltes fragiles à presser sur place, des sangles, des papiers journaux,
des cartons, un sac plastique (protégeant les cartons et journaux de la pluie), des
flacons avec de l’alcool pour y conserver les fleurs fragiles.
Matériels restant au camp
- Silicagel, sachets plastiques et boîtes hermétiques, bouteilles de gaz (séchage des
échantillons), une grande presse/séchoir fonctionnant jour et nuit, des sacs plastiques.
Lorsqu’une récolte est effectuée, différentes informations ont été relevées dans la mesure du
possible, à l’aide du matériel cité ci-dessus. Nous avons noté : la détermination faite sur le
terrain (famille, genre, espèce) ; le numéro de récolte permettant une distinction entre les
récoltes effectuées ; la localisation de la récolte et de son alentour : la date, le pays, le massif,

2. Méthodologie
30
la forêt, le substrat, la station, la pente, l’orientation, l’altitude, ainsi que les coordonnées
géographiques précises données par un GPS. Nous avons également pu indiquer grâce à nos
assistants sur le terrain, quelques noms vernaculaires et les modes d’utilisation de certaines
plantes que nous avons récoltées. Nous relevons également la biologie de la plante permettant
de définir notamment son type biologique ainsi que les caractères morphologiques visibles sur
le terrain et pas forcément sur la plante séchée (couleur, odeur, présence de latex, etc.).
Une photo de la récolte et de son environnement a également été effectuée dans la majorité
des cas, permettant lors des déterminations en herbier une visualisation de la récolte dans son
état naturel (quelques exemples ci-dessous, photo 5).
PHOTO 5 : PHOTOS DE RÉCOLTES FERTILES EFFECTUÉES.
DEPUIS LE HAUT DE GAUCHE À DROITE : COLEOTRYPE GOUDOTII H.PERRIER VAR. GOUDOTII ; TAMBOURISSA
CAPURONII CAVACO, PHYLLARTHRON BOJERANUM A.DC., ONCOSTEMUM SP.
Le séchage des échantillons pressés a été effectué par air chaud directement sur le terrain.
Cette méthode permet une bonne conservation des échantillons, (un meilleur rendu des
couleurs que la méthode de séchage par alcool) et la possibilité d'utiliser le matériel pour des
© CJBG 2010 © CJBG 2010
© CJBG 2010© CJBG 2010

2. Méthodologie
31
études sur l’ADN. Il y a toutefois des inconvénients dont nous avons pu faire les frais durant
l’incendie du séchoir sur le quatrième camp.
Sur demande de différents spécialistes, des feuilles ont été mises en silicagel pour des études
en laboratoire lors des récoltes fertiles de certains genres. Les genres récoltés en silicagel
appartiennent aux familles suivantes : ASPARAGACEAE Dracaena, EUPHORBIACEAE
Acalypha, MYRSINACEAE Oncostemum, OLEACEAE Noronhia, SAPINDACEAE tout les
genres, SAPOTACEAE tout les genres, VIOLACEAE Rinorea.
D’autres récoltes dont les fleurs sont fragiles et très difficiles à identifier ou décrire après le
séchage ont été mises en alcool.
Certains fruits impossibles à presser de par leur taille trop imposante ont été séchés à part et
ont permis de constituer une collection carpologique.
2.4. GESTION ET DÉTERMINATION DES SPÉCIMENS D’HERBIERS
Les données recueillies sur le terrain pour chaque récolte, ont été saisies informatiquement
dans la base de données Ambre (BD Ambre).
Pour chacune des récoltes fertiles accomplies dans le cadre de ce projet, 6 parts au maximum
ont été effectuées dans la mesure du possible. Ceci a permis de constituer des parts pour
échange avec les grands herbiers mondiaux impliqués dans la botanique de Madagascar, ainsi
qu’une part en don contre détermination pour les différents spécialistes s’ils existent.
La distribution des parts pour échange se fait uniquement lorsque la récolte est déterminée et
de la manière suivante : 1 part a été laissée à Madagascar (TEF) (acronyme d’herbier selon
HOLMGREN & AL., 1990), 1 part reste à Genève puis les 4 autres (exportés à l’herbier de
Genève) sont distribuées dans l’ordre aux herbiers de : Paris (P), Missouri (MO), Kew (K) et
Wageningen (WAG). Les parts pour échange non encore distribuées, en attente d’une
détermination, restent provisoirement stockées à l’herbier de Genève (cf. figure 1, p. 33).
La distribution de certaines parts en don contre détermination a été effectuée pour 32
familles, en utilisant les contacts précédemment établis dans le cadre des projets antérieurs
des CJBG dans le nord de Madagascar. Les photos des récoltes faites sur le terrain ont été
classées selon les familles et gravées sur des CD pour être envoyées aux différents spécialistes
concernés et ainsi aider à la détermination des échantillons d’herbiers. Lorsque les parts en

2. Méthodologie
32
double pour échange n’étaient pas en nombre suffisant, la part de l’herbier de Genève a été
envoyée en prêt, après des contacts avec certains spécialistes.
Nous tenons à remercier chaleureusement les spécialistes suivants qui nous ont fait part de
leurs déterminations. Pour certains groupes ou certaines familles sans révisions récentes, les
déterminations deviennent très difficiles et leur aide a donc été précieuse. Ci-dessous, se
trouve la liste des spécialistes par ordre alphabétique avec mention de l’acronyme de l’herbier
de référence entre parenthèse :
Abdul-Salim, K. (OSU); Archer, R.H. (PRE); Breteler, F.J. (WAG); Buerki, S. (NEU);
Callmander, M.W. (G); Clerc, P. (G); Cribb, P.J. (K); Davis, A.P. (K); De Block, P. (BR); De
Wilde, J.J.F.E (WAG); Dessein, S. (BR); Faden, R.B. (US); Gautier, L. (G); Goyder, D.J. (K);
Groeninckx, I. (BR); Hong-Wa, C. (MO); Klackenberg, J. (S); Labat, J.-N. (P); Lorence, D.H.
(PTBG); Lowry II, P.P. (MO); Liede, S. (UBT); Malécot, V. (ANGERS); Mathieu, G. (BR);
McPherson, G. (MO); Meve, U. (UBT); Miller, J. (MO); Price, M. (G); Rakotondrainibe, F.
(P); Razafimandimbison, S.G. (SUNIV); Rogers, Z.S. (MO); Schatz, G.E. (MO); Stone, D.
(NU); et Van der Werff, H. (MO).
Les déterminations des échantillons fertiles se sont déroulées à l’herbier de Genève. Ce travail
a été effectué à l’aide de la Flore de Madagascar et des Comores (HUMBERT & AL., 1936),
ou avec l’aide des révisions récentes. Ensuite, les déterminations ont été comparées dans la
mesure du possible avec les spécimens d’herbier présent à Genève, ou à des scans de planches
d’herbier disponibles sur Internet (Sonnerat, TROPICOS et JSTOR).
La BD Ambre, initialement élaborée sous Microsoft Excel a été introduite dans Microsoft
Access afin d’en optimiser la gestion. Ce logiciel a permis d’imprimer les étiquettes
d’herbiers et les nouvelles déterminations (ou déterminavits) pour toutes les parts de chacun
des échantillons d’herbiers récoltés dans le cadre de ce projet, par la réalisation de formulaires
compilant les données recherchées. De plus, pour situer nos récoltes dans ce massif, des
localités ont été créées dans le logiciel ArcView. Les étiquettes d’herbier contiennent donc,
les coordonnées du point GPS précis, et l'attribution de la récolte à l'une des 6 localités
suivantes : partie centrale (12°34'S - 49°10'E), partie nord (12°28'S - 49°10'E), versants ouest
(12°34'S - 49°7'E), versants est (12°34'S - 49°13'E), foret de Sahantanana (12°27'S - 49°13'E),
partie sud (12°40'S - 49°9'E).
Chaque nouvelle détermination a été notée dans la BD Ambre. Un nouveau déterminavit
comprenant le nom du premier récolteur, le numéro de récolte, la nouvelle détermination,
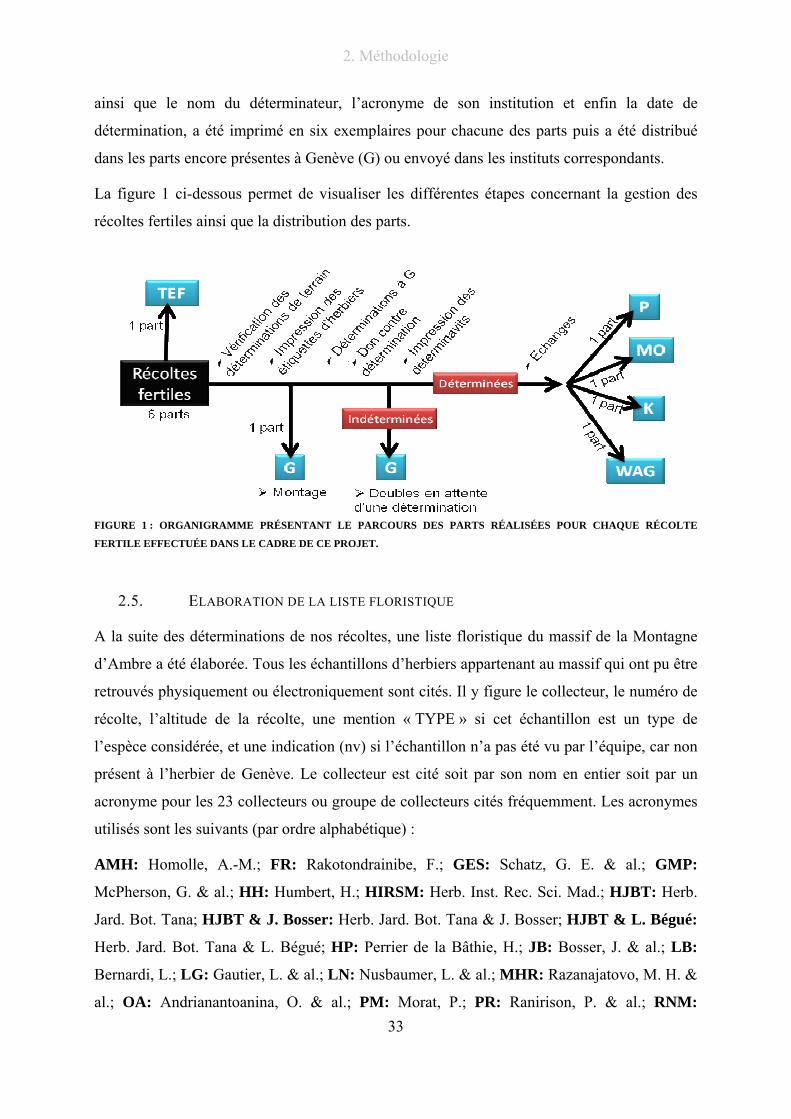
2. Méthodologie
33
ainsi que le nom du déterminateur, l’acronyme de son institution et enfin la date de
détermination, a été imprimé en six exemplaires pour chacune des parts puis a été distribué
dans les parts encore présentes à Genève (G) ou envoyé dans les instituts correspondants.
La figure 1 ci-dessous permet de visualiser les différentes étapes concernant la gestion des
récoltes fertiles ainsi que la distribution des parts.
FIGURE 1 : ORGANIGRAMME PRÉSENTANT LE PARCOURS DES PARTS RÉALISÉES POUR CHAQUE RÉCOLTE
FERTILE EFFECTUÉE DANS LE CADRE DE CE PROJET.
2.5. ELABORATION DE LA LISTE FLORISTIQUE
A la suite des déterminations de nos récoltes, une liste floristique du massif de la Montagne
d’Ambre a été élaborée. Tous les échantillons d’herbiers appartenant au massif qui ont pu être
retrouvés physiquement ou électroniquement sont cités. Il y figure le collecteur, le numéro de
récolte, l’altitude de la récolte, une mention « TYPE » si cet échantillon est un type de
l’espèce considérée, et une indication (nv) si l’échantillon n’a pas été vu par l’équipe, car non
présent à l’herbier de Genève. Le collecteur est cité soit par son nom en entier soit par un
acronyme pour les 23 collecteurs ou groupe de collecteurs cités fréquemment. Les acronymes
utilisés sont les suivants (par ordre alphabétique) :
AMH: Homolle, A.-M.; FR: Rakotondrainibe, F.; GES: Schatz, G. E. & al.; GMP:
McPherson, G. & al.; HH: Humbert, H.; HIRSM: Herb. Inst. Rec. Sci. Mad.; HJBT: Herb.
Jard. Bot. Tana; HJBT & J. Bosser: Herb. Jard. Bot. Tana & J. Bosser; HJBT & L. Bégué:
Herb. Jard. Bot. Tana & L. Bégué; HP: Perrier de la Bâthie, H.; JB: Bosser, J. & al.; LB:
Bernardi, L.; LG: Gautier, L. & al.; LN: Nusbaumer, L. & al.; MHR: Razanajatovo, M. H. &
al.; OA: Andrianantoanina, O. & al.; PM: Morat, P.; PR: Ranirison, P. & al.; RNM:

2. Méthodologie
34
Réserves Naturelles de Madagascar; SDR: Ramandimbimanana, S. D. & al.; SF: Service
Forestier; SM: Malcomber, S. & al.; SMT: Trigui, S. M. & al.
L’altitude de récolte est donnée entre parenthèses. Si celle-ci a été notée à l'origine sous forme
d'intervalle (avec une différence inférieure à 500 m) ou que l’altitude a pu être estimée à partir
des indications précisées dans la localité de récolte, nous mentionnons sa valeur entre
crochets. Pour les échantillons type, on donne généralement des informations concernant la
distribution des différentes parts de l’échantillon dans les herbiers internationaux pour les HT
(holotypes), IT (isotypes), ST (syntypes), et LT (lectotypes).
2.6. SYSTÈME DE CLASSIFICATION UTILISÉ
Classiquement, les Angiospermes sont séparées en Monocotylédones et Dicotylédones.
Toutefois, à la faveur d'études moléculaires nombreuses effectuées dans les 20 dernières
années, cette classification s'est avérée artificielle. « Un groupe de botanistes le « Angiosperm
Phylogeny Group » [a proposé] en 1998 une classification ordinale des plantes à fleurs (APG,
1998) » (SPICHIGER & AL., 2004). Ensuite, deux nouvelles classifications se sont créées
APG II puis APG III, se dernier étant utilisé dans ce travail, il est disponible sur Internet
(STEVENS, 2001 en ligne).
Deux grands groupes apparaissent: les Euangiospermes-monoaperturées et les
Euangiospermes-triaperturées. L’une des caractéristiques principales distinguant ces deux
groupes est le nombre d'ouverture du pollen. L’appellation monoaperturées regroupe les
plantes à une seule ouverture; les triaperturées celles qui en ont trois. Toutes les
Euangiospermes-Triaperturées sont des anciennes Dicotylédones, mais si les
Euangiospermes-Monoaperturées sont en majorité des anciennes Monocotylédones, on y
trouve aussi un important d'anciennes Dicotylédones.
Dans la liste floristique des phanérogames du massif d’Ambre (8.1.1., p. 99) nous avons
choisis de distinguer les Gymnospermes des Angiospermes, puis les Angiospermes sont
rangées par ordre alphabétique des familles indifféremment de leur appartenance aux
Monocotylédones-Dicotylédones ou Monoaperturées-Triaperturées. Au niveau des genres, ils
ont été regroupés dans les familles selon le système proposé par APG 3. Par exemple les
Dracaena se trouvent dans la famille des ASPARAGACEAE, les Anthocleista sont dans la
famille des GENTIANACEAE, les Nuxia sont dans la famille des STILBACEAE, les
Buddleja dans les SCROPHULARIACEAE, et les Mostuea dans celle des

2. Méthodologie
35
GELSEMIACEAE. De plus, les Grewia, Dombeya, Byttneria, Adansonia sont regroupés dans
la famille des MALVACEAE au sens large. Un index répertoriant les genres de
phanérogames se trouve à la fin de la liste, pour faciliter les recherches (cf. p. 175).
Concernant la liste floristique des ptéridophytes du massif d’Ambre (Annexe 8.1.2., p. 178),
les espèces sont rangées par ordre alphabétique des genres indépendamment de leur
appartenance aux familles.
Enfin, une liste des Bryophytes, des Ascomycètes lichenisés, et des Champignons que nous
avons récoltés est proposée en Annexe 8.1.3., p. 189.

2. Méthodologie
36
SECONDE PARTIE : ANALYSE BIOGEOGRAPHIQUE
2.7. RÉPARTITION GÉOGRAPHIQUE DES ESPÈCES
Des données sur la répartition géographique de toutes les espèces recensées du massif de la
Montagne d’Ambre ont été recueillies, afin de répondre aux hypothèses concernant
l’installation de cette flore humide au sein du massif de la Montagne d’Ambre. Pour chaque
espèce déterminée, sa répartition géographique à Madagascar et également sur les autres
continents a été prise en compte et saisie. Si des éventuelles variétés, sous-espèces ou formes
sont présentes, leur distribution géographique est également considérées et annotées, mais en
addition à la distribution de l’espèce. Le concept d’espèce est clairement défini comme « un
ensemble de populations dont les membres sont en mesure de se reproduire entre eux dans un
environnement naturel et de donner naissance à une descendance féconde » (CAMPBELL &
REECE, 2004).
Pour effectuer une synthèse des informations concernant la distribution géographique des
espèces, différentes ressources ont été consultées :
- la Flore de Madagascar et des Comores (HUMBERT & AL. 1936),
- les révisions publiées sur les taxons concernés présentes à la bibliothèque des CJBG,
- la base de données en ligne du Missouri Botanical Garden « Tropicos »,
- la base de données en ligne du MNHN de Paris « Sonnerat »,
- la consultation des spécimens d’herbiers présents à l’herbier de Genève.
Pour saisir les données recueillies, un masque de saisie a été créé dans Microsoft Access
(photo 6) sur le modèle de celui utilisé par GAUTIER (2002) pour l’inventaire floristique du
Manongarivo.
PHOTO 6 : MASQUE DE SAISIE POUR LA PHYTOGÉOGRAPHIE DE CHAQUE ESPÈCE DU MASSIF D’AMBRE.
PHOTO DE L’ÉCRAN DU FORMULAIRE DE SAISIE ÉLABORÉ DANS MICROSOFT ACCESS.

2. Méthodologie
37
2.7.1. RÉPARTITION DES ESPÈCES SUR L’ÎLE DE MADAGASCAR
Pour l’île de Madagascar, la répartition géographique des espèces a été notée par
présence/absence dans les 6 domaines phytogéographiques proposés par HUMBERT (1955,
1965). Nous utilisons ici le système de classification de Humbert, car il reste encore
aujourd'hui à la base de tous les systèmes de classifications phytogéographiques proposés.
Pour localiser les récoltes effectuées au massif d’Ambre parmis les domaines
phytogéographiques, deux catégories définies par l’altitude de récoltes ont été créés pour
affiner la saisie : le « Centre Montagne d’Ambre », pour des altitudes de récoltes supérieures à
1 000 m, et l’« Ouest Montagne d’Ambre », pour des altitudes de récoltes inférieures à 600 m.
Si une espèce est présente entre 600 et 1 000 m d’altitude, ou à moins de 600 m et également
à plus de 1 000 m d’altitude, ou si l’altitude n’est pas connue, les deux catégories ont été
cochées comme « Présence ». Cependant ces données n’ont pas été utilisées dans ce présent
travail, afin de laisser apparaître les tendances phytogéographiques des espèces de ce massif,
sans y introduire les points de vue existants sur la classification phytogéographique de cette
montagne. Les espèces connues à Madagascar uniquement de la Montagne d’Ambre sont
identifiées par l’acronyme « MdA », pour Montagne d’Ambre dans la liste floristique.
Les domaines phytogéographiques selon HUMBERT (1965) sont définis ci-dessous et
symbolisés dans les résultats et la liste floristique (Annexe 8.1., p. 101) par les lettres
suivantes :
‐ E : Domaine de l’Est
Ce domaine se situe sur la côte orientale de l’île sur plus de 1 100 km de long, et large de
60 km, du niveau du littoral jusqu’aux altitudes de 800-900 m. La pluviosité annuelle est de
2 000 mm en moyenne et diminue au fur et à mesure du nord au sud du domaine. Une
pluviosité exceptionnelle de 6 000 mm de pluie par an est relevée dans certaines localités de
la presqu’île de Masoala. La végétation est de type humide. Cette « série à Myristicacées et
Anthostema » composant la forêt dense ombrophile orientale est secondarisée par des
défrichements et incendies répétés qui ont eu lieux dans ces zones relativement faciles
d’accès. Des grandes villes du pays se trouve sur cette côte Est (Tamatava, Fianarantsoa, Fort-
Dauphin, etc.).
‐ Sb : Domaine du Sambirano

2. Méthodologie
38
D’après Humbert (1965) « Ce domaine forme une enclave dans la partie NW du Domaine de
l’Ouest. » Cette partie de l’île correspond au bassin du Sambirano qui est un fleuve de cette
région. Les hautes montagnes du Tsaratanana à proximité dévient les courants de vents
humides du sud-est, pour conférer à cette région une pluviosité annuelle de 2 000 à 2 500 mm
par an. C’est une végétation humide de basse altitude. La forêt dense ombrophile est
composée d’une « série à Chlaenacées – Myristicacées – Anthostema ».
‐ C : Domaine du Centre
Ce domaine est caractérisé par l’ensemble des territoires situés au-delà de 800 m d’altitude
jusqu’à 2 000 m environ. Il est composé principalement du massif montagneux allant du nord
au sud de l’île, mais également des altitudes supérieures de la Montagne d’Ambre, des
plateaux de l’Isalo et des quelques petits massifs isolés dans le domaine du Sud. HUMBERT
(1965) subdivise ce domaine en trois sous-domaines : Centre nord, Centre moyen, et Centre
sud, à cause de deux seuils où l’altitude diminue pour n’atteindre que 900 m et ou la largeur
du domaine d’est en ouest diminue fortement. Ce sont les seuils de Mandritsara (dans le nord)
et celui de Menarahaka (dans le sud). La pluviosité est d’environ 1 500 mm vers 1 400 m
d’altitude. On y trouve également de la végétation de type humide. La végétation primaire de
ce domaine est, d’après HUMBERT (1965), celle qui a été le plus largement détruite sur la
plus grande partie des territoires qu’elle recouvrait avant l’action directe ou indirecte de
l’homme. La végétation est partagée en trois étages. L’étage de moyenne altitude (800 -
1 300 m.) est une « série à Tambourissa et Weinmannia » composé d’une forêt dense
ombrophile. L’étage montagnard ou « sylve à lichens » qui se situe au-delà de 1300 m
d’altitude, avec une forêt basse. Puis, l’étage des pentes occidentales ou « série à Uapaca
Bojeri et Chlaenacées », une forêt basse sclérophylle dont la végétation primaire recouvrait
des altitudes situées entre 800 et 2 000 m des versants Ouest du Domaine du Centre.
‐ HM : Domaine des Hautes Montagnes
Il comprend des massifs disséminés du nord au sud de l’île qui ont une altitude supérieure à
2000 m. Ce domaine comprend notamment le Tsaratanana et ses satellites (2 886 m), le
Marojejy (2 137 m), l’Ankaratra (2 638 m), l’Andringitra (2 658 m), et l’Andohahela
(1 979 m). La pluviosité annuelle est de 1 200 à 2 000 mm. On y trouve une végétation
humide d’altitude appelée « fourré dense et savane d’altitude » par HUMBERT (1965).
‐ W : Domaine de l’Ouest

2. Méthodologie
39
Le domaine de l’Ouest englobe une grande partie des altitudes inférieures à 800 m du versant
occidental de l’île, jusqu’au littoral. La particularité de ce domaine est la présence de taxons à
feuilles caduques durant la saison sèche. C’est donc un type de végétation sec, ou « série à
Dalbergia – Commiphora – Hildegardia » pour HUMBERT (1965). Ce domaine est divisé en
deux secteurs : extrême Nord et Sud, séparés géographiquement par le domaine du
Sambirano. Durant la saisie des données phytogéographiques, nous avons distingués deux
sous-secteurs l’ouest-nord et l’ouest-sud. (W: symbole utilisé si le taxon est présent dans les
deux secteurs, sinon utilisation des abréviations de W(N) pour le secteur nord et W(S) pour le
secteur sud).
‐ S : Domaine du Sud
Situé à l’extrême sud de l’île, ce domaine présente un climat semi-aride. Une différenciation
presque inexistante entre les saisons permet de différencier ce domaine de celui de l’Ouest. La
pluviométrie annuelle est inférieure à 700 mm. La végétation est de type sèche, xérophile.
HUMBERT (1965) place cette végétation dans la « série à Didiéracées et Euphorbia ».
2.7.2. RÉPARTITION MONDIALE DES ESPÈCES
Lorsque l’espèce considérée n’est pas endémique de Madagascar, les informations ont été
enregistrées et codées de la manière suivante:
‐ RM : Région Malgache, comprenant les différentes îles de l’Océan Indien à proximité
de Madagascar (Les Mascareignes, Les Comores, Les Seychelles),
‐ Aft : Afrique tropicale (pour les ptéridophytes uniquement il est précisé si l’espèce est
présente en Afrique du Sud : SAf),
‐ Amt : Amérique tropicale,
‐ Oc : Océanie,
‐ As : Asie,
‐ Eur : Europe.
Lors des analyses phytogéographiques des regroupements de catégories ont été effectués,
pour une meilleure compréhension. Les catégories associées sont les suivantes :
‐ M (espèces endémiques de Madagascar),

2. Méthodologie
40
‐ M+RM (espèces se trouvant à Madagascar et dans la Région Malgache),
‐ M+Aft (espèces retrouvées sur le continent africain, mais absente de la Région
Malgache),
‐ M+RM+Aft (espèces présentes à la fois en Région Malgache et sur le continent
Africain),
‐ M+RM+As/Oc (espèces distribuées en Région Malgache et à l’Est soit en Asie ou/et
en Océanie),
‐ PALEO (espèces paléotropicales: retrouvées en Afrique et en Asie/Océanie mais pas
en Amérique),
‐ PAN (espèces pantropicales retrouvées en Amérique tropicale, mais pas dans l'espace
Holarctique),
‐ COSMOPOLITES (espèces cosmopolites se retrouvant sur tous les continents).
2.8. MODE DE DISPERSION DES DIASPORES
Une étude sur le mode de dispersion des diaspores de certaines espèces du massif permet
d’amener des éléments d'explication à leur présence au sein du massif. Toutefois, malgré la
grande importance du mode de transport des diaspores, ce caractère n’a pas pu être identifié
pour toutes les espèces. Il n’existe aucune base de données, ou de listes qui traitent
uniquement de ce sujet. Toutefois dans diverses révisions ou des livres de phytogéographie
générale (SCHNELL, 1970; SCHNELL, 1971) il est possible de retrouver cette information
pour certaines espèces caractéristiques.
Nous avons recherché le mode de dispersion des diaspores pour les endémiques locales
uniquement.
2.9. ANALYSE DE LA RÉPARTITION DES ESPÈCES DU MASSIF
Pour l’analyse biogéographique, différents aspects de la flore du massif d’Ambre ont été mis
en évidence par nos résultats. Tout d’abord, une présentation de la distribution mondiale puis
malgache des espèces du massif d’Ambre est proposée, pour les phanérogames et pour les
ptéridophytes. Ces figures permettent d’observer les affinités phytogéographiques des espèces
du massif d’une manière générale.

2. Méthodologie
41
Ensuite, divers sujets ont été approfondis tout en utilisant les données de distributions des
espèces de phanérogames et de ptéridophytes comprises.
Nous avons utilisés les outils disponible sur Microsoft Access et Excel afin d’obtenir ces
résultats. Des tableaux croisés ont été créés à l’aide de Microsoft Excel pour rassembler les
données.

3. Résultats et Interprétations
42
3. RÉSULTATS ET INTERPRÉTATIONS Cette étude ce base sur toutes les espèces que nous avons pu inventorier du massif de la
Montagne d’Ambre. Notre travail sur le terrain a ciblé l’approche de la flore humide des
altitudes supérieures du massif, mais les résultats que nous analyserons sont issus de
l'échantillonnage global du massif, qui reste toutefois plus important au niveau des altitudes
supérieures. Pour ce qui concerne les analyses phytogéographiques, elles ont été réalisées sur
les 912 espèces identifiées du massif. Toutefois deux espèces dont les déterminations nous ont
parues douteuses, ainsi que les espèces cultivées (au nombre de trois) n’ont pas été prises en
compte.
3.1. PROSPECTION BOTANIQUE DE LA MONTAGNE D’AMBRE
Le massif d’Ambre a toujours intrigué les naturalistes et notamment les botanistes de par son
isolement géographique actuel par rapport à la grande matrice de montagnes du Centre de la
Grande Ile. Toutefois les prospections botaniques sont restées, jusqu’à cette présente étude,
centrées sur une petite partie de la réserve ou ciblées sur des groupes particuliers de plantes.
Parmi tous les collecteurs et groupes de collecteurs recensés qui ont récoltés des échantillons
d’herbier dans ce massif, nous sommes l’équipe qui en a réalisé le plus grand nombre.
3.1.1. DONNÉES GÉNÉRALES SUR LES ÉCHANTILLONS D’HERBIER
Actuellement, nous recensons 3 521 récoltes pour le massif d’Ambre, dont 34% ont été
récoltés dans le cadre de ce présent projet. Tous ces échantillons forment la « BD Ambre ».
Ce chiffre est certainement encore sous-estimé, car il ne tient compte que des données
d’autres herbiers disponibles en ligne, de ce qui a été trouvé à l’herbier de Genève et des
échantillons citées dans la Flore de Madagascar, ainsi que dans les différentes révisions
consultées. L’herbier de Genève possède une part de plus de 41% des récoltes du massif
d’Ambre, selon nos recherches actuelles.
Le pourcentage de taxons déterminés est de plus de 74% pour toute la BD Ambre. Ce chiffre
est de 67,4% pour les récoltes réalisées lors de ce présent projet.
Au cours des travaux de terrain accomplis dans le cadre de cette étude, 1 266 récoltes de
spécimens d’herbiers ont été effectuées. En additionnant le nombre de parts réalisées pour
chaque récolte cela représente 5 826 parts. Lors d’un incendie sur le quatrième campement,

3. Résultats et Interprétations
43
nous avons perdus 65 récoltes fertiles. Ces échantillons perdus ne sont pas pris en compte,
cependant quelques déterminations ont tout de même été entreprises à partir des
photographies effectuées sur le terrain.
Des feuilles mises en silicagel ont été préparées sur le terrain pour 43 récoltes, et 27 récoltes
dont les fleurs sont fragiles ont été gardées dans de l’alcool. Une collection carpologique
comprenant 22 échantillons a aussi été constituée.
La majorité des récoltes effectuées, ainsi que la végétation et les paysages ont été
photographiés et composent une base de données de 7 088 photos de terrain.
3.1.2. CHRONOLOGIE DES RÉCOLTES ET AUGMENTATION DE LA CONNAISSANCE DE LA DIVERSITÉ
Le graphique ci-dessous montre la répartition des 3 521 échantillons selon les années de
récolte. Les premières récoltes faites dans ce massif datent de 1880 et ont été effectuées par J.
M. Hildebrandt. C’est seulement à partir des années 1950 que les récoltes d’échantillons
d’herbier commencent à croître de manière significative, ce qui coïncide avec la création de
l’aire protégée en 1958. Les collecteurs importants de cette période sont: H. Humbert, H.
Perrier de la Bâthie, R. Capuron., J. Bosser, L. Bernardi et A.-M. Homolle. Entre 1992 et
1993 des collecteurs du Missouri Botanical Garden (Andrianantoanina, O. & al., Malcomber,
S. & al.) ont fait d’importantes récoltes à la Montagne d’Ambre. En 2007 et 2008 (entouré en
bleu sur la figure 2, p. 44), ce sont les récoltes réalisées durant cette étude, qui ont permis
l’acquisition d’un grand nombre d’échantillons d’herbiers. Toutefois, l’augmentation du
nombre d’échantillons d’herbier n’est pas forcément corrélée avec une augmentation du
nombre d’espèces. La figure 3, p. 44 est une comparaison de l’évolution avec le temps du
nombre d’espèces pour le massif d’Ambre (en rouge) en parallèle avec l’évolution du nombre
d’échantillons d’herbiers (en bleu). On remarque logiquement que plus le temps passe, plus
l’écart entre le nombre d’échantillons et le nombre d’espèces se creuse. Toutefois le nombre
d’espèces nouvelles pour le massif continue d’augmenter, et augmente même plus fortement
depuis les années 1990. Il reste donc encore beaucoup d’espèces à découvrir dans ce massif,
notamment dans les lieux les plus difficiles d’accès. Durant nos travaux de terrain, en
seulement 6 mois, nous avons récoltés plus de 47% des taxons que nous recensons du massif
d’Ambre, le reste provenant uniquement de récoltes antérieures. Plus de 21% des échantillons
que nous avons récoltés représentent des espèces inconnues du massif avant notre étude.
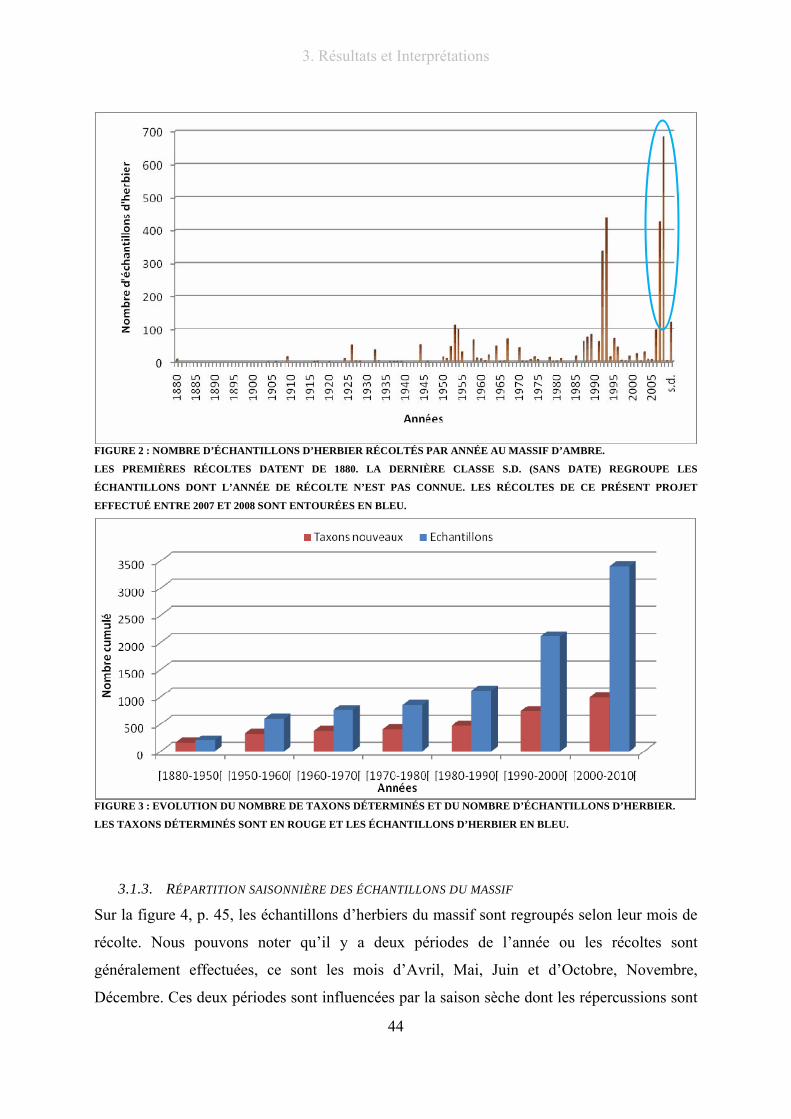
3. Résultats et Interprétations
44
FIGURE 2 : NOMBRE D’ÉCHANTILLONS D’HERBIER RÉCOLTÉS PAR ANNÉE AU MASSIF D’AMBRE.
LES PREMIÈRES RÉCOLTES DATENT DE 1880. LA DERNIÈRE CLASSE S.D. (SANS DATE) REGROUPE LES
ÉCHANTILLONS DONT L’ANNÉE DE RÉCOLTE N’EST PAS CONNUE. LES RÉCOLTES DE CE PRÉSENT PROJET
EFFECTUÉ ENTRE 2007 ET 2008 SONT ENTOURÉES EN BLEU.
FIGURE 3 : EVOLUTION DU NOMBRE DE TAXONS DÉTERMINÉS ET DU NOMBRE D’ÉCHANTILLONS D’HERBIER.
LES TAXONS DÉTERMINÉS SONT EN ROUGE ET LES ÉCHANTILLONS D’HERBIER EN BLEU.
3.1.3. RÉPARTITION SAISONNIÈRE DES ÉCHANTILLONS DU MASSIF
Sur la figure 4, p. 45, les échantillons d’herbiers du massif sont regroupés selon leur mois de
récolte. Nous pouvons noter qu’il y a deux périodes de l’année ou les récoltes sont
généralement effectuées, ce sont les mois d’Avril, Mai, Juin et d’Octobre, Novembre,
Décembre. Ces deux périodes sont influencées par la saison sèche dont les répercussions sont
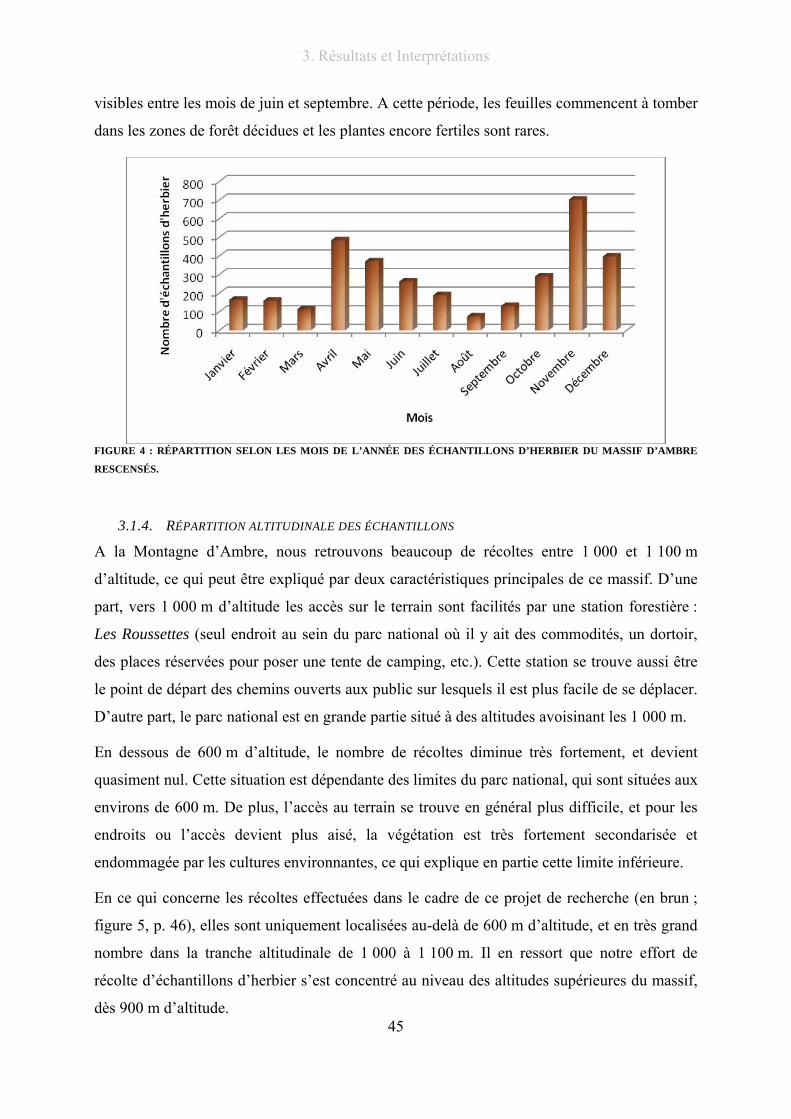
3. Résultats et Interprétations
45
visibles entre les mois de juin et septembre. A cette période, les feuilles commencent à tomber
dans les zones de forêt décidues et les plantes encore fertiles sont rares.
FIGURE 4 : RÉPARTITION SELON LES MOIS DE L’ANNÉE DES ÉCHANTILLONS D’HERBIER DU MASSIF D’AMBRE
RESCENSÉS.
3.1.4. RÉPARTITION ALTITUDINALE DES ÉCHANTILLONS
A la Montagne d’Ambre, nous retrouvons beaucoup de récoltes entre 1 000 et 1 100 m
d’altitude, ce qui peut être expliqué par deux caractéristiques principales de ce massif. D’une
part, vers 1 000 m d’altitude les accès sur le terrain sont facilités par une station forestière :
Les Roussettes (seul endroit au sein du parc national où il y ait des commodités, un dortoir,
des places réservées pour poser une tente de camping, etc.). Cette station se trouve aussi être
le point de départ des chemins ouverts aux public sur lesquels il est plus facile de se déplacer.
D’autre part, le parc national est en grande partie situé à des altitudes avoisinant les 1 000 m.
En dessous de 600 m d’altitude, le nombre de récoltes diminue très fortement, et devient
quasiment nul. Cette situation est dépendante des limites du parc national, qui sont situées aux
environs de 600 m. De plus, l’accès au terrain se trouve en général plus difficile, et pour les
endroits ou l’accès devient plus aisé, la végétation est très fortement secondarisée et
endommagée par les cultures environnantes, ce qui explique en partie cette limite inférieure.
En ce qui concerne les récoltes effectuées dans le cadre de ce projet de recherche (en brun ;
figure 5, p. 46), elles sont uniquement localisées au-delà de 600 m d’altitude, et en très grand
nombre dans la tranche altitudinale de 1 000 à 1 100 m. Il en ressort que notre effort de
récolte d’échantillons d’herbier s’est concentré au niveau des altitudes supérieures du massif,
dès 900 m d’altitude.
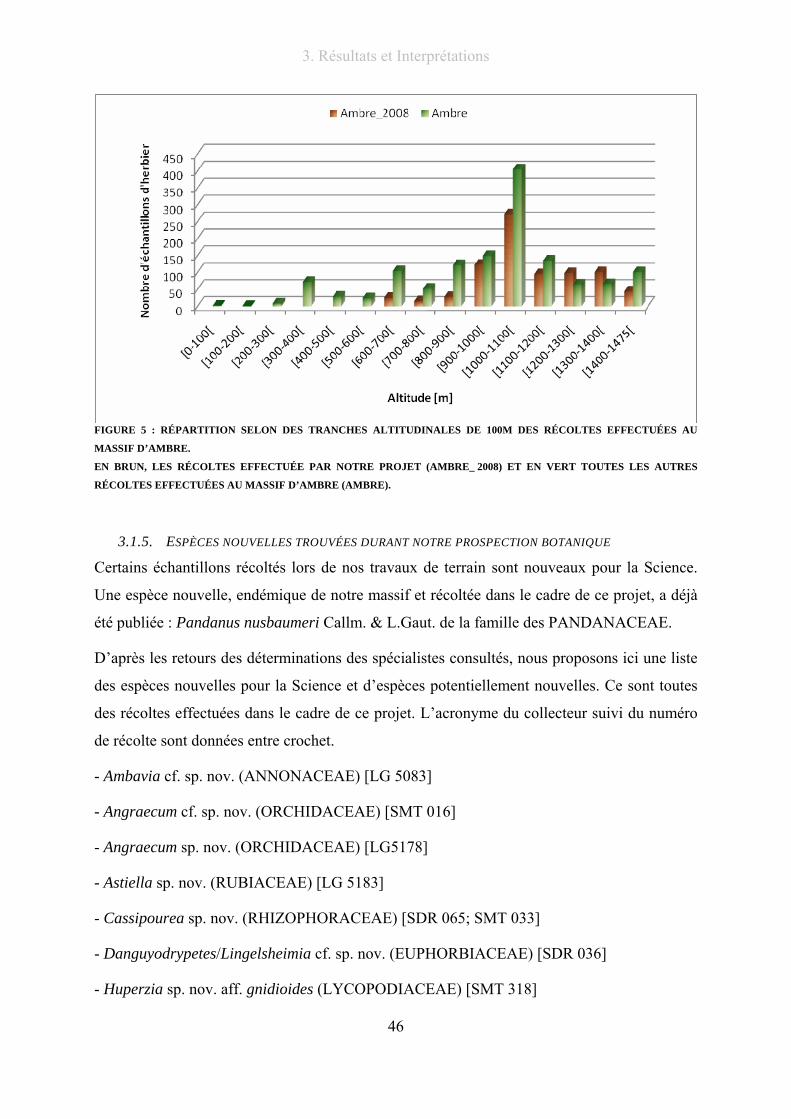
3. Résultats et Interprétations
46
FIGURE 5 : RÉPARTITION SELON DES TRANCHES ALTITUDINALES DE 100M DES RÉCOLTES EFFECTUÉES AU
MASSIF D’AMBRE.
EN BRUN, LES RÉCOLTES EFFECTUÉE PAR NOTRE PROJET (AMBRE_ 2008) ET EN VERT TOUTES LES AUTRES
RÉCOLTES EFFECTUÉES AU MASSIF D’AMBRE (AMBRE).
3.1.5. ESPÈCES NOUVELLES TROUVÉES DURANT NOTRE PROSPECTION BOTANIQUE
Certains échantillons récoltés lors de nos travaux de terrain sont nouveaux pour la Science.
Une espèce nouvelle, endémique de notre massif et récoltée dans le cadre de ce projet, a déjà
été publiée : Pandanus nusbaumeri Callm. & L.Gaut. de la famille des PANDANACEAE.
D’après les retours des déterminations des spécialistes consultés, nous proposons ici une liste
des espèces nouvelles pour la Science et d’espèces potentiellement nouvelles. Ce sont toutes
des récoltes effectuées dans le cadre de ce projet. L’acronyme du collecteur suivi du numéro
de récolte sont données entre crochet.
- Ambavia cf. sp. nov. (ANNONACEAE) [LG 5083]
- Angraecum cf. sp. nov. (ORCHIDACEAE) [SMT 016]
- Angraecum sp. nov. (ORCHIDACEAE) [LG5178]
- Astiella sp. nov. (RUBIACEAE) [LG 5183]
- Cassipourea sp. nov. (RHIZOPHORACEAE) [SDR 065; SMT 033]
- Danguyodrypetes/Lingelsheimia cf. sp. nov. (EUPHORBIACEAE) [SDR 036]
- Huperzia sp. nov. aff. gnidioides (LYCOPODIACEAE) [SMT 318]

3. Résultats et Interprétations
47
- Ophiocolea sp. nov. (BIGNONIACEAE) [MHR 006]
- Ouratea cf. sp. nov. (OCHNACEAE) [SMT 496]
- Pandanus nusbaumeri Callm. & L. Gaut. (PANDANACEAE) (CALLMANDER & AL.,
2009) [LG 5217]
- Pyrrosia sp. nov. (POLYPODIACEAE) est une nouvelle espèce du groupe des
ptéridophytes, qui a également été récoltée récemment dans la région du Daraina. Une
publication va être préparée par l’un des encadreurs de ce projet avec la spécialiste de cette
famille F. Rakotondrainbe. [LN 2429]
- Tambourissa sp. nov (MONIMIACEAE) [SMT 340]
3.1.6. LES ESPÈCES MENACÉES DU MASSIF D’AMBRE
Une liste provisoire donnant la catégorie de menaces des espèces de phanérogames de l’île de
Madagascar est en cours d'élaboration par le Missouri Botanical Garden. Elle a été comparée
à notre liste floristique des espèces du massif d’Ambre. Il en ressort que 126 espèces, soit
13,8% du total de toutes les espèces répertoriées actuellement dans ce massif, figurent dans
cette liste (Annexe 8.3., p. 193). 64 espèces du massif d’Ambre sont actuellement considérées
comme menacées, c'est-à-dire rattachées aux catégories suivantes : vulnérable (VU), en
danger (EN) ou en danger critique d’extinction (CR). Les espèces appartenant à cette dernière
catégorie sont confrontées « à un risque extrêmement élevé d’extinction à l’état sauvage »
(IUCN, 2001). Au massif d’Ambre, ce sont en tout 14 espèces, dont 8 qui sont des
endémiques locales et 9 appartiennent à la famille des ORCHIDACEAE. Ce sont par
exemple : Aeranthes ambrensis Toill.-Gen., Ursch & Bosser, Angraecum ambrense H.Perrier,
Gastrorchis geffrayi (Bosser) Senghas. Il y a également une LAURACEAE : Aspidostemon
reticulatum van der Werff, une PIPERACEAE : Peperomia pluvisilvatica G.Mathieu, deux
APOCYNACEAE et une BALSAMINACEAE (figure 6 et photo 7 page suivante).

3. Résultats et Interprétations
48
FIGURE 6 : LES 912 ESPÈCES DU MASSIF ET LEUR RÉPARTITION DANS LES CATÉGORIES DE LA LISTE ROUGE DE
L’IUCN.
DONNÉES PROVENANT DU MISSOURI BOTANICAL GARDEN (EXTRACTION DE JUIN 2010).
PHOTO 7 : ESPÈCES MENACÉES DITES « EN DANGER CRITIQUE D’EXTINCTION » ET ENDÉMIQUES DU MASSIF
D’AMBRE.
PEPEROMIA PLUVISILVATICA G.MATHIEU (GAUCHE) ET SECAMONE MARSUPIATA KLACK (DROITE).
3.2. RICHESSE SPÉCIFIQUE
La liste floristique du massif de la Montagne d’Ambre dénombre 1 450 taxons de plantes
vasculaires. Sur ce total, 200 taxons sont des ptéridophytes. Il y a en tout 912 espèces
déterminées et réparties dans 564 genres et dans 151 familles différentes.
34 taxons de Bryophytes, Ascomycètes lichenisés et de Champignons récoltés dans le cadre
de ce projet faisant également partie de la flore du massif sont listés en Annexe (8.1.3.,
p. 189). Ils ne sont toutefois pas pris en compte pour les analyses, ni dans les chiffres ci-
dessus.
© CJBG 2010 © CJBG 2010
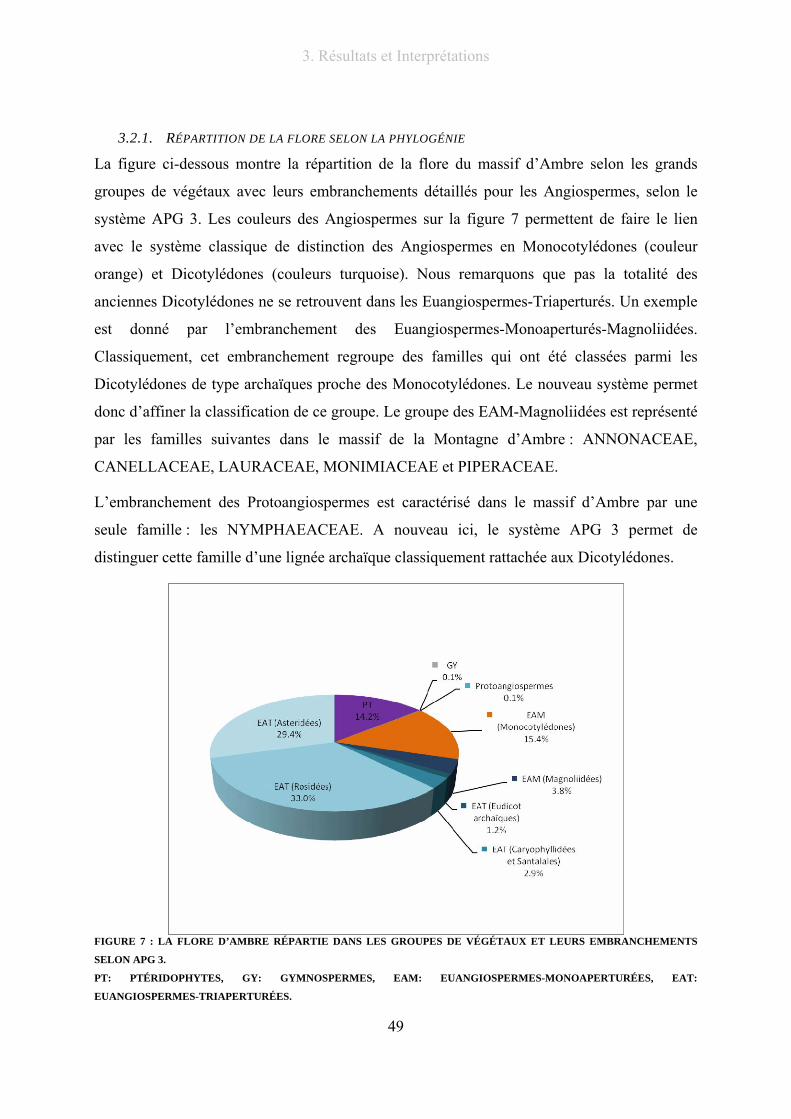
3. Résultats et Interprétations
49
3.2.1. RÉPARTITION DE LA FLORE SELON LA PHYLOGÉNIE
La figure ci-dessous montre la répartition de la flore du massif d’Ambre selon les grands
groupes de végétaux avec leurs embranchements détaillés pour les Angiospermes, selon le
système APG 3. Les couleurs des Angiospermes sur la figure 7 permettent de faire le lien
avec le système classique de distinction des Angiospermes en Monocotylédones (couleur
orange) et Dicotylédones (couleurs turquoise). Nous remarquons que pas la totalité des
anciennes Dicotylédones ne se retrouvent dans les Euangiospermes-Triaperturés. Un exemple
est donné par l’embranchement des Euangiospermes-Monoaperturés-Magnoliidées.
Classiquement, cet embranchement regroupe des familles qui ont été classées parmi les
Dicotylédones de type archaïques proche des Monocotylédones. Le nouveau système permet
donc d’affiner la classification de ce groupe. Le groupe des EAM-Magnoliidées est représenté
par les familles suivantes dans le massif de la Montagne d’Ambre : ANNONACEAE,
CANELLACEAE, LAURACEAE, MONIMIACEAE et PIPERACEAE.
L’embranchement des Protoangiospermes est caractérisé dans le massif d’Ambre par une
seule famille : les NYMPHAEACEAE. A nouveau ici, le système APG 3 permet de
distinguer cette famille d’une lignée archaïque classiquement rattachée aux Dicotylédones.
FIGURE 7 : LA FLORE D’AMBRE RÉPARTIE DANS LES GROUPES DE VÉGÉTAUX ET LEURS EMBRANCHEMENTS
SELON APG 3.
PT: PTÉRIDOPHYTES, GY: GYMNOSPERMES, EAM: EUANGIOSPERMES-MONOAPERTURÉES, EAT:
EUANGIOSPERMES-TRIAPERTURÉES.

3. Résultats et Interprétations
50
Plus généralement, pour la flore du massif d’Ambre, cette figure montre qu’il y a plus de 70%
d’EAT ou Eudicotylédones dans cette flore. Le tiers restant se partage équitablement entre les
Euangiospermes-monoaparturées et les ptéridophytes.
3.2.2. RICHESSE SPÉCIFIQUE DES FAMILLES ET DES GENRES
La figure 8, p. 51 montre les familles les plus riches en espèces dans la flore de la Montagne
d’Ambre. La classification utilisée est celle proposée par APG III (STEVENS, 2001 en ligne).
Les 18 familles les plus riches en espèces sont illustrées en détail. La catégorie « autres »
regroupe le reste, soit 133 familles qui possèdent moins de 21 espèces. Il y a en tout 34
familles qui ne sont représentées que par une seule espèce.
Les RUBIACEAE sont avec les ORCHIDACEAE, les deux familles les plus riches en
espèces. Puis viennent les FABACEAE, EUPHORBIACEAE, MALVACEAE,
ASTERACEAE, POACEAE, APOCYNACEAE, ACANTHACEAE, SAPINDACEAE,
ASPLENIACEAE avec plus de 30 espèces. Parmi les familles les plus riches en espèces, la
majorité sont typiques des formations végétales humides qui composent une grande partie du
massif. De plus, 4 familles sur les 18 les plus riches en espèces, appartiennent au groupe de
ptéridophytes, qui sont toujours mieux représentées dans les végétations humides. Les
endroits les plus accessibles, et les plus étudiés de ce massif sont surtout les plus hautes
altitudes composées d’une flore humide, ce qui permet d’expliquer en partie cette dominance
des familles typiques des formations végétales humides.
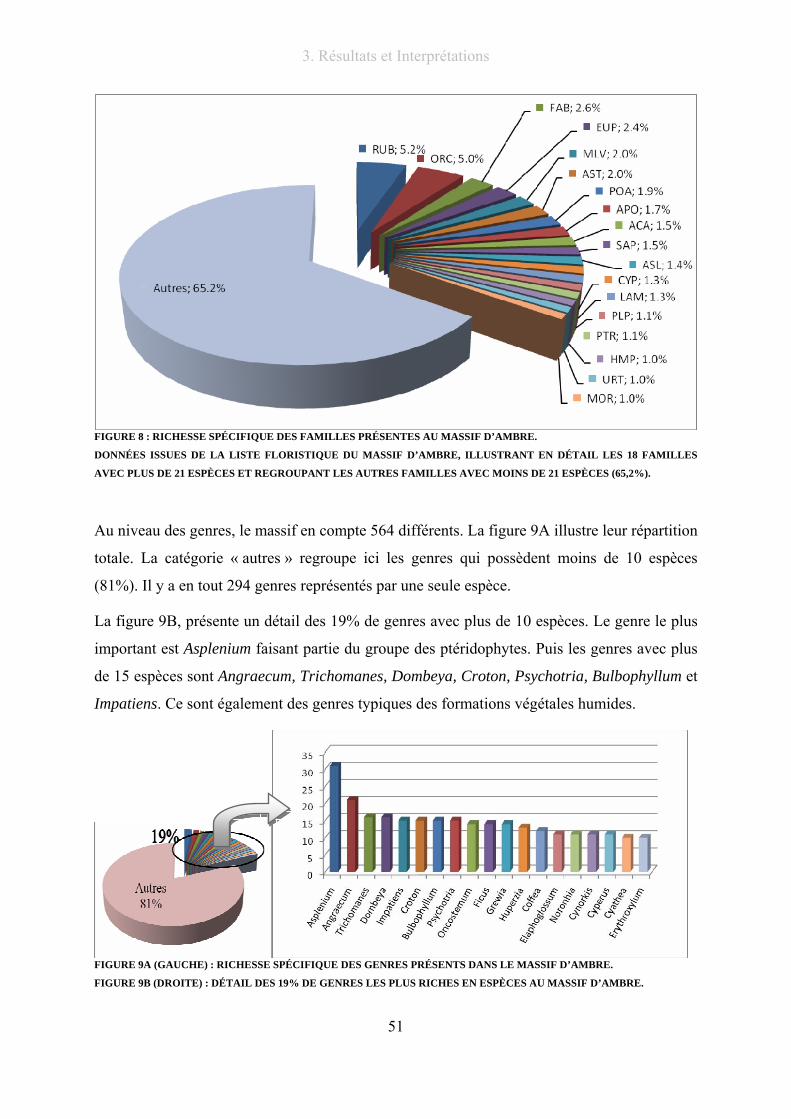
3. Résultats et Interprétations
51
FIGURE 8 : RICHESSE SPÉCIFIQUE DES FAMILLES PRÉSENTES AU MASSIF D’AMBRE.
DONNÉES ISSUES DE LA LISTE FLORISTIQUE DU MASSIF D’AMBRE, ILLUSTRANT EN DÉTAIL LES 18 FAMILLES
AVEC PLUS DE 21 ESPÈCES ET REGROUPANT LES AUTRES FAMILLES AVEC MOINS DE 21 ESPÈCES (65,2%).
Au niveau des genres, le massif en compte 564 différents. La figure 9A illustre leur répartition
totale. La catégorie « autres » regroupe ici les genres qui possèdent moins de 10 espèces
(81%). Il y a en tout 294 genres représentés par une seule espèce.
La figure 9B, présente un détail des 19% de genres avec plus de 10 espèces. Le genre le plus
important est Asplenium faisant partie du groupe des ptéridophytes. Puis les genres avec plus
de 15 espèces sont Angraecum, Trichomanes, Dombeya, Croton, Psychotria, Bulbophyllum et
Impatiens. Ce sont également des genres typiques des formations végétales humides.
FIGURE 9A (GAUCHE) : RICHESSE SPÉCIFIQUE DES GENRES PRÉSENTS DANS LE MASSIF D’AMBRE.
FIGURE 9B (DROITE) : DÉTAIL DES 19% DE GENRES LES PLUS RICHES EN ESPÈCES AU MASSIF D’AMBRE.

3. Résultats et Interprétations
52
3.2.3. DIFFÉRENCES FLORISTIQUES ENTRE LE MASSIF D’AMBRE ET LE MANONGARIVO
Nous pouvons comparer la flore du massif d’Ambre et celle du Manongarivo. Ce massif situé
dans la partie nord de l’île à la confluence entre le domaine du Centre, du Sambirano et de
l’Ouest, possède certaines caractéristiques rappelant la Montagne d’Ambre : l’amplitude
altitudinale varie jusqu’à 1 876 m, la végétation est composée en dessous de 800 m d’une
forêt dense ombrophile de basse altitude, puis entre 800 et 1 600 m d’une forêt dense humide
de montagne, et au-delà de 1 600 m d’une forêt d’altitude (GAUTIER & GOODMAN, 2002).
De plus, nous avons toutes les données nécessaires sur ce massif pour effectuer cette
comparaison. La flore des altitudes supérieures du Manongarivo est, tout comme pour le
massif d’Ambre, rattachée au domaine phytogéographique du Centre. Cependant une
différence importante est à signaler : au Manongarivo cette flore n’est pas isolée mais en
connexion directe avec la flore du domaine du Centre qui se prolonge du nord au sud de
Madagascar.
Comparaison au niveau de la richesse spécifique
Au Manongarivo, 1 095 espèces sont recensées, alors que pour la Montagne d’Ambre nous en
avons 912 espèces. L’amplitude altitudinale qui est beaucoup plus importante au
Manongarivo offre une plus large variabilité de niches écologiques pour les espèces, ce qui
explique en partie cette différence.
Au niveau de la répartition de la flore selon les groupes de végétaux, la Montagne d’Ambre
possède une richesse spécifique des ptéridophytes plus importante que celle du Manongarivo.
Dans le massif du Manongarivo, 5,9% de la flore fait partie du groupe des ptéridophytes alors
qu’à la Montagne d’Ambre ce chiffre est de 14,2%, égalant presque celui des
Monocotylédones. Dans le massif du Manongarivo, les Dicotylédones sont plus importantes
qu’à la Montagne d’Ambre avec 77,2% et 70,4% respectivement. Pour les Monocotylédones
les chiffres sont de même ordre de grandeur avec 16,7% pour Manongarivo et 15,3% pour le
massif d’Ambre.
Les familles dominantes pour le massif du Manongarivo sont les RUBIACEAE largement
dominantes, puis les FABACEAE, ORCHIDACEAE, ASTERACEAE, POACEAE. Nous
notons une plus grande richesse spécifique des FABACEAE au Manongarivo qu’à la
Montagne d’Ambre et une plus grande importance des RUBIACEAE.
Au niveau générique, le massif du Manongarivo compte 630 genres différents contre 564 pour
la Montagne d’Ambre, et seulement 360 sont communs au deux massifs.

3. Résultats et Interprétations
53
Il y a 35 genres qui comptent plus de 10 espèces, alors qu’à la Montagne d’Ambre seulement
19 genres possèdent plus de 10 espèces.
Les 10 premiers genres les plus riches en espèces sont les suivants pour le Manongarivo (les
genres communs aux deux listes sont mis en évidence, en gras):
Impatiens, Bulbophyllum, Oncostemum, Psychotria, Mapouria, Diospyros, Vernonia, Ficus,
Panicum, Peperomia, Cyathea, Weinmannia, Dypsis, Gravesia, Asplenium, Garcinia,
Erythroxylum, Croton, Hypoestes.
Et pour la Montagne d’Ambre :
Asplenium, Angraecum, Trichomanes, Dombeya, Impatiens, Croton, Bulbophyllum,
Psychotria, Oncostemum, Ficus, Grewia, Huperzia, Coffea, Elaphoglossum, Noronhia,
Cynorkis, Cyperus, Cyathea, Erythroxylum.
Ces deux listes confirment la grande diversité de ptéridophytes rencontrées dans le massif
d’Ambre. Le genre Asplenium est en effet le plus riche en espèces pour la flore du massif
d’Ambre, puis on y retrouve le genre Trichomanes, Elaphoglossum, et Cyathea, compris
également dans le groupe des ptéridophytes. Ce groupe est connu pour avoir des diaspores
pouvant se déplacer facilement sur de longue distance, c’est une raison qui pourrait expliquer
cette richesse plus importante en ptéridophytes au massif d’Ambre qu’au Manongarivo.
Nous avons recensé un total de 270 genres du Manongarivo inexistants à la Montagne
d’Ambre. Parmi ces genres nous en retrouvons un certains nombre qui sont caractéristiques de
l’étage des pentes occidentales du domaine du Centre de HUMBERT (1965). Ces genres
caractéristiques, absents de la Montagne d’Ambre, mais présents au Manongarivo sont les
suivants : Uapaca, Leptolaena, Sarcolaena, Asteropeia, Faurea, Protorhus. HUMBERT
(1965) nomme par ailleurs cette végétation la « Série à Uapaca bojeri et Chlaenacées
[= Sarcolaenacées]». La famille des Sarcolaenacées est également absente du massif
d’Ambre. Il semble donc que ce soit précisément cet étage des pentes occidentales du
domaine du Centre qui soit inexistant dans le massif d’Ambre.
De plus, au Manongarivo il existe une plus grande diversité de Weinmannia, Senecio,
Vernonia, Conyza, Helichrysum et légèrement plus d’espèces de Psiadia qui sont des genres
abondants dans le domaine du Centre et celui des Hautes Montagnes.

3. Résultats et Interprétations
54
Le massif du Manongarivo, de part sa position géographique, et son amplitude altitudinale
offre une plus grande diversité de milieu qu’au massif de la Montagne d’Ambre, dont la flore
humide est actuellement en position isolée.
3.3. LA FLORE DU MASSIF ET SES AFFINITÉS MONDIALES
44,5% de toutes les plantes vasculaires du massif d’Ambre ont une distribution dépassant les
frontières de l’île de Madagascar. Les affinités mondiales de cette flore varient selon divers
critères comme les groupes considérés (Phanérogames, Ptéridophytes) et les zones du massif
(pleine forêt, bordure des lacs d’anciens cratères).
3.3.1. DISTRIBUTION MONDIALE DES PLANTES VASCULAIRES
La répartition des espèces de phanérogames au niveau mondial montre un nombre important
d’espèces endémiques de Madagascar (61,4%) (figure 10). Les espèces avec une répartition
étendue à la région malgache (Mascareignes, Comores, et Seychelles) sont la deuxième
catégorie la plus importante avec 11,1%. Le pourcentage d’espèces communes avec l’Afrique
(M+Aft) ou avec l’Afrique et la région malgache (M+RM+Aft) est presque identique avec
5,6% et 6,5% respectivement. Les espèces partagées avec la région malgache, l’Asie et
l’Océanie sont peu nombreuses avec seulement 0,7%. Les espèces paléotropicales sont
légèrement plus nombreuses mais restent relativement faible avec 2,6%.
Les espèces pantropicales sont presque aussi nombreuses que les espèces de la catégorie
M+RM. Enfin, les espèces cosmopolites sont rares avec seulement 1,7% de la flore des
phanérogames du massif d’Ambre.
FIGURE 10 : DISTRIBUTION MONDIALE DES PHANÉROGAMES DU MASSIF D’AMBRE.
M : MADAGASCAR, RM : RÉGION MALGACHE, AFT : AFRIQUE TROPICALE, AS/OC : ASIE-OCÉANIE, PALEO :
PALÉOTROPICALE, PAN : PANTROPICALE, COSMOPOLITES.
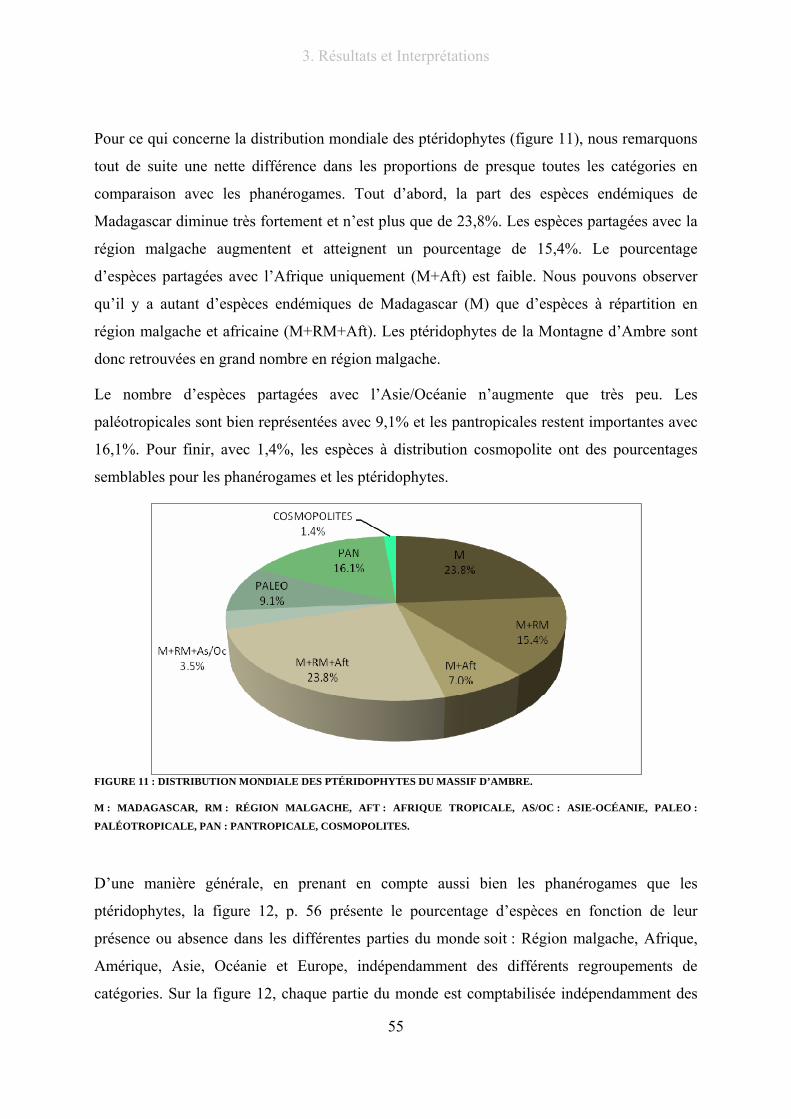
3. Résultats et Interprétations
55
Pour ce qui concerne la distribution mondiale des ptéridophytes (figure 11), nous remarquons
tout de suite une nette différence dans les proportions de presque toutes les catégories en
comparaison avec les phanérogames. Tout d’abord, la part des espèces endémiques de
Madagascar diminue très fortement et n’est plus que de 23,8%. Les espèces partagées avec la
région malgache augmentent et atteignent un pourcentage de 15,4%. Le pourcentage
d’espèces partagées avec l’Afrique uniquement (M+Aft) est faible. Nous pouvons observer
qu’il y a autant d’espèces endémiques de Madagascar (M) que d’espèces à répartition en
région malgache et africaine (M+RM+Aft). Les ptéridophytes de la Montagne d’Ambre sont
donc retrouvées en grand nombre en région malgache.
Le nombre d’espèces partagées avec l’Asie/Océanie n’augmente que très peu. Les
paléotropicales sont bien représentées avec 9,1% et les pantropicales restent importantes avec
16,1%. Pour finir, avec 1,4%, les espèces à distribution cosmopolite ont des pourcentages
semblables pour les phanérogames et les ptéridophytes.
FIGURE 11 : DISTRIBUTION MONDIALE DES PTÉRIDOPHYTES DU MASSIF D’AMBRE.
M : MADAGASCAR, RM : RÉGION MALGACHE, AFT : AFRIQUE TROPICALE, AS/OC : ASIE-OCÉANIE, PALEO :
PALÉOTROPICALE, PAN : PANTROPICALE, COSMOPOLITES.
D’une manière générale, en prenant en compte aussi bien les phanérogames que les
ptéridophytes, la figure 12, p. 56 présente le pourcentage d’espèces en fonction de leur
présence ou absence dans les différentes parties du monde soit : Région malgache, Afrique,
Amérique, Asie, Océanie et Europe, indépendamment des différents regroupements de
catégories. Sur la figure 12, chaque partie du monde est comptabilisée indépendamment des
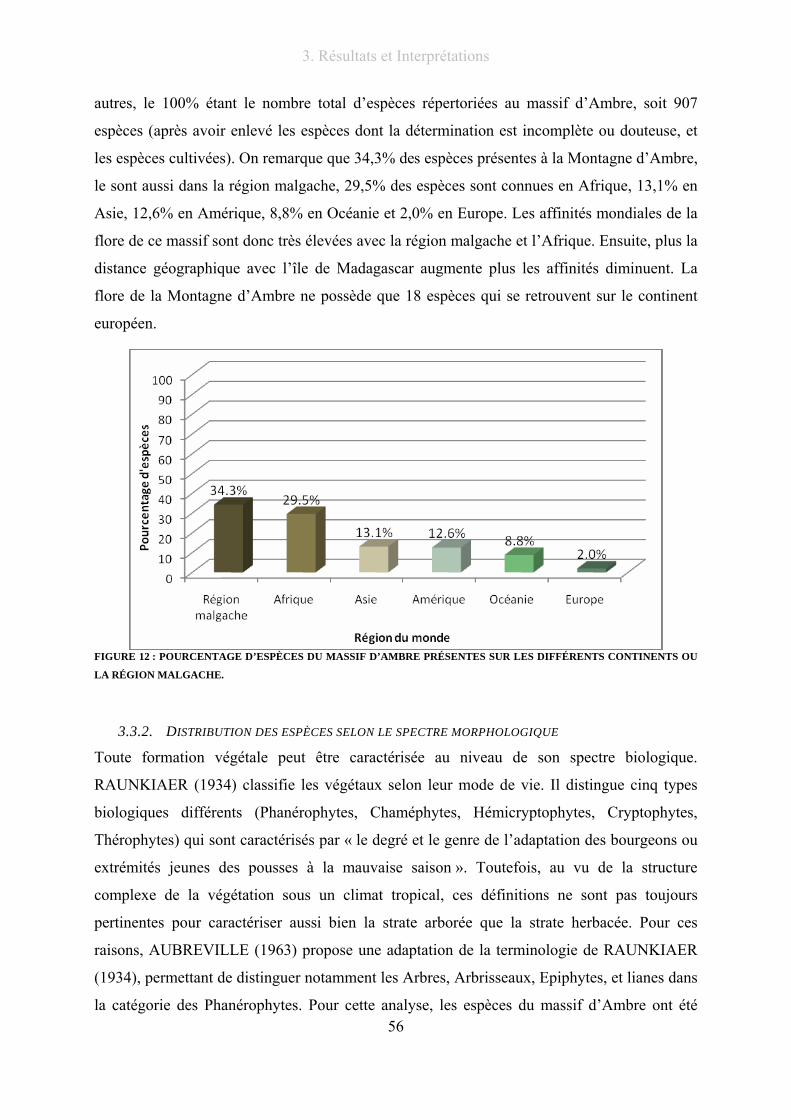
3. Résultats et Interprétations
56
autres, le 100% étant le nombre total d’espèces répertoriées au massif d’Ambre, soit 907
espèces (après avoir enlevé les espèces dont la détermination est incomplète ou douteuse, et
les espèces cultivées). On remarque que 34,3% des espèces présentes à la Montagne d’Ambre,
le sont aussi dans la région malgache, 29,5% des espèces sont connues en Afrique, 13,1% en
Asie, 12,6% en Amérique, 8,8% en Océanie et 2,0% en Europe. Les affinités mondiales de la
flore de ce massif sont donc très élevées avec la région malgache et l’Afrique. Ensuite, plus la
distance géographique avec l’île de Madagascar augmente plus les affinités diminuent. La
flore de la Montagne d’Ambre ne possède que 18 espèces qui se retrouvent sur le continent
européen.
FIGURE 12 : POURCENTAGE D’ESPÈCES DU MASSIF D’AMBRE PRÉSENTES SUR LES DIFFÉRENTS CONTINENTS OU
LA RÉGION MALGACHE.
3.3.2. DISTRIBUTION DES ESPÈCES SELON LE SPECTRE MORPHOLOGIQUE
Toute formation végétale peut être caractérisée au niveau de son spectre biologique.
RAUNKIAER (1934) classifie les végétaux selon leur mode de vie. Il distingue cinq types
biologiques différents (Phanérophytes, Chaméphytes, Hémicryptophytes, Cryptophytes,
Thérophytes) qui sont caractérisés par « le degré et le genre de l’adaptation des bourgeons ou
extrémités jeunes des pousses à la mauvaise saison ». Toutefois, au vu de la structure
complexe de la végétation sous un climat tropical, ces définitions ne sont pas toujours
pertinentes pour caractériser aussi bien la strate arborée que la strate herbacée. Pour ces
raisons, AUBREVILLE (1963) propose une adaptation de la terminologie de RAUNKIAER
(1934), permettant de distinguer notamment les Arbres, Arbrisseaux, Epiphytes, et lianes dans
la catégorie des Phanérophytes. Pour cette analyse, les espèces du massif d’Ambre ont été

3. Résultats et Interprétations
57
classées selon les principaux groupements proposés par AUBREVILLE (1963), que nous
appellerons ici des types morphologiques : Arbres s.l. > 7m ; Arbrisseaux s.l. ≤ 7m ; Palmiers
s.l. ; Lianes ; Epiphytes ; Saproparasites ; Herbacées s.l. (comprenant des herbacées terrestres,
aquatiques, annuelles, mais aussi les fougères arborescentes).
La figure 13A, p. 58 montre la répartition des espèces du massif d’Ambre selon les types
morphologiques d’AUBREVILLE. Le type morphologique de chaque espèce a été attribué
après observation des descriptions faites sur le terrain pour chaque récolte de l’espèce
considérée, ou par observation des indications issues de la littérature.
La figure 13B, p.58 est un tableau croisé représentant le spectre morphologique des espèces
du massif d’Ambre et leur distribution mondiale. Les données de distribution mondiale des
espèces ont été simplifiées en 2 catégories : les espèces endémiques de Madagascar (vert) et
les espèces non endémiques (brun).
Nous pouvons remarquer que le type morphologique le mieux représenté dans la flore de ce
massif est la catégorie des herbacées avec 28,8%. Puis il y a les arbres (22,8%), les
arbrisseaux (20,6%) et les épiphytes avec 17,2%. Les lianes ne représentent que 9% de toutes
les espèces recensées du massif d’Ambre. Les saproparasites et les palmiers sont minoritaires
avec respectivement 0,8 et 0,1%.
Nous observons sur la figure 13B, que le type morphologique qui regroupe le plus d’espèces
endémiques de Madagascar est la catégorie des arbres, puis vient les arbrisseaux. Les lianes
regroupent aussi légèrement plus d’espèces endémiques de Madagascar. Par contre, pour les
herbacées le nombre d’espèces endémiques est nettement inférieur au nombre d’espèces non
endémiques de Madagascar. Les espèces de ce type morphologique possèdent donc une plus
forte capacité de dispersion de leurs diaspores sur de longues distances que les espèces
d’Arbres ou Arbrisseaux. Les espèces épiphytes sont dans une moindre mesure comparables
aux herbacées avec légèrement plus d’espèces non endémiques. Pour ce qui concerne les
saproparasites (LORANTHACEAE et CUSCUTACEAE) et les palmiers, leur nombre est
relativement faible au sein du massif d’Ambre.
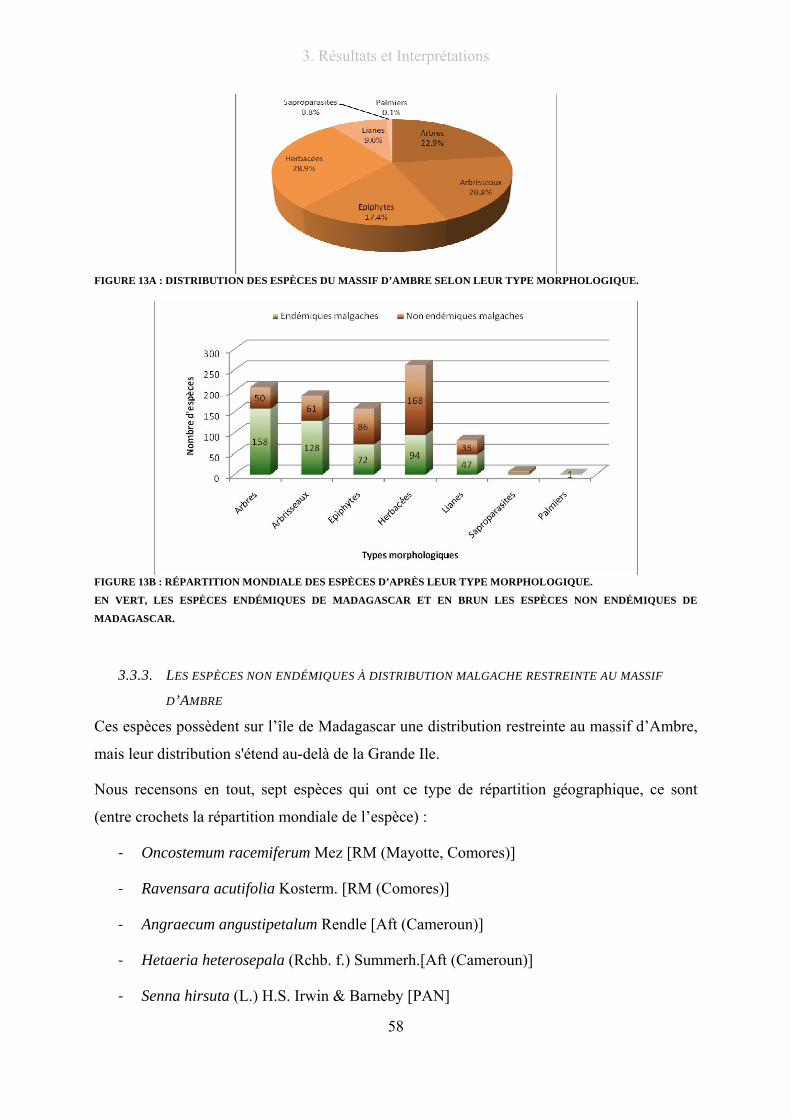
3. Résultats et Interprétations
58
FIGURE 13A : DISTRIBUTION DES ESPÈCES DU MASSIF D’AMBRE SELON LEUR TYPE MORPHOLOGIQUE.
FIGURE 13B : RÉPARTITION MONDIALE DES ESPÈCES D’APRÈS LEUR TYPE MORPHOLOGIQUE.
EN VERT, LES ESPÈCES ENDÉMIQUES DE MADAGASCAR ET EN BRUN LES ESPÈCES NON ENDÉMIQUES DE
MADAGASCAR.
3.3.3. LES ESPÈCES NON ENDÉMIQUES À DISTRIBUTION MALGACHE RESTREINTE AU MASSIF
D’AMBRE
Ces espèces possèdent sur l’île de Madagascar une distribution restreinte au massif d’Ambre,
mais leur distribution s'étend au-delà de la Grande Ile.
Nous recensons en tout, sept espèces qui ont ce type de répartition géographique, ce sont
(entre crochets la répartition mondiale de l’espèce) :
‐ Oncostemum racemiferum Mez [RM (Mayotte, Comores)]
‐ Ravensara acutifolia Kosterm. [RM (Comores)]
‐ Angraecum angustipetalum Rendle [Aft (Cameroun)]
‐ Hetaeria heterosepala (Rchb. f.) Summerh.[Aft (Cameroun)]
‐ Senna hirsuta (L.) H.S. Irwin & Barneby [PAN]

3. Résultats et Interprétations
59
‐ Pueraria thunbergiana (Willd.) Ohwi [PAN]
‐ Cardamine trichocarpa Hochst. ex A.Rich. [PALEO]
Les trois dernières espèces de cette liste possèdent une distribution pantropicale et
paléotropicale. Elles possèdent donc de fortes capacités de dispersion. Leur distribution
actuelle à Madagascar, restreinte à la Montagne d’Ambre, peut suggérer qu’elles sont
certainement arrivées récemment sur l’île. Il est donc fort probable que leur dispersion se
poursuive. Inversement, pour les espèces possédant une distribution restreinte à la région
malgache ou au continent africain, leur dispersion peut être plus ancienne, car elles sont
limitées à quelques régions particulières du monde. Les deux espèces retrouvées sur le
continent africain ont été récoltées uniquement au Cameroun, pays d’Afrique de l’Ouest, situé
géographiquement à l’opposé de Madagascar. Elles sont très probablement établies en
Afrique de l’Est, mais n’ont peut-être pas encore été recensées.
L’espèce Cardamine trichocarpa Hochst. ex A.Rich. a été récoltée par Perrier de la Bâthie,
qui précisait dans la Flore de Madagascar (HUMBERT & AL., 1936) que cette récolte avait
été effectuée « autour du Lac Maudit ». Il en avait conclu que cette « espèce [était] très
probablement introduite dans cette localité (isolée au milieu d’une forêt dense) par les oiseaux
aquatiques ». Il semble que certaines espèces situées en bordure des lacs de cratères aient pu
être acheminées au sein du massif d’Ambre par les oiseaux comme le suppose également
Perrier de la Bâthie.
Du point de vue biogéographique, l’idée qui ressort de cette conclusion de Perrier de la
Bâthie, est que les espèces situées en bordure des lacs possèdent certainement une répartition
géographique plus large, et que l’endémisme insulaire doit diminuer fortement. Dans le
chapitre ci-dessous, nous avons analysé la répartition géographique d’espèces situées en
bordure de lacs d’anciens cratères.
3.3.4. CAS PARTICULIER DES ESPÈCES SITUÉES À PROXIMITÉ DES LACS D’ANCIENS CRATÈRES
Afin d’observer la répartition mondiale de ces espèces, nous avons pris en compte tous les
échantillons d’herbiers qui possédaient dans leur description de la station de récolte le vocable
« prairie » ou « lac maudit » ou encore « grand lac », puis nous avons éliminé les stations
décrites comme éloignée des lacs. Le nombre d’échantillons d’herbiers pris en compte pour
cette analyse est de 52 (Annexe 8.4., p. 196). Les points GPS de ces échantillons (sauf pour 1
échantillon sans point GPS) sont indiqués sur la carte 6, p. 60 par un point bleu. Ils sont situés

3. Résultats et Interprétations
60
autour de deux lacs différents : le Lac Maudit, le Grand Lac, mais aussi autour d’un lac
temporairement inondé (lac temporaire sur la carte). La physionomie de ces deux lacs permet
d’avoir des prairies aux alentours. En effet, les bords de ces lacs ne sont pas abrupts comme
c’est le cas pour le Lac Texier, autour duquel aucune prairie n’a pu s’installer.
CARTE 6 : LOCALISATION DES POINTS DE RÉCOLTES DES ÉCHANTILLONS QUI POSSÈDENT DANS LEUR
DESCRIPTION DE LA STATION DE RÉCOLTE LE VOCABLE « PRAIRIE ».
ILS SONT REPRÉSENTÉS PAR UN POINT BLEU CIEL ET SONT SITUÉS AUTOUR DE 2 LACS (GRAND LAC, LAC
MAUDIT) ET D’UN LAC TEMPORAIRE. LE NOM DES CINQ LACS SITUÉS À L’INTÉRIEUR DU PARC NATIONAL EST
ÉGALEMENT MENTIONNÉ (PETIT LAC, LAC MAUDIT, GRAND LAC, LAC TEXIER, LAC FANTANY), LE SIXIÈME LAC
ÉTANT SITUÉ DANS LA RÉSERVE SPÉCIALE À L’EXTRÊME NORD DU MASSIF.
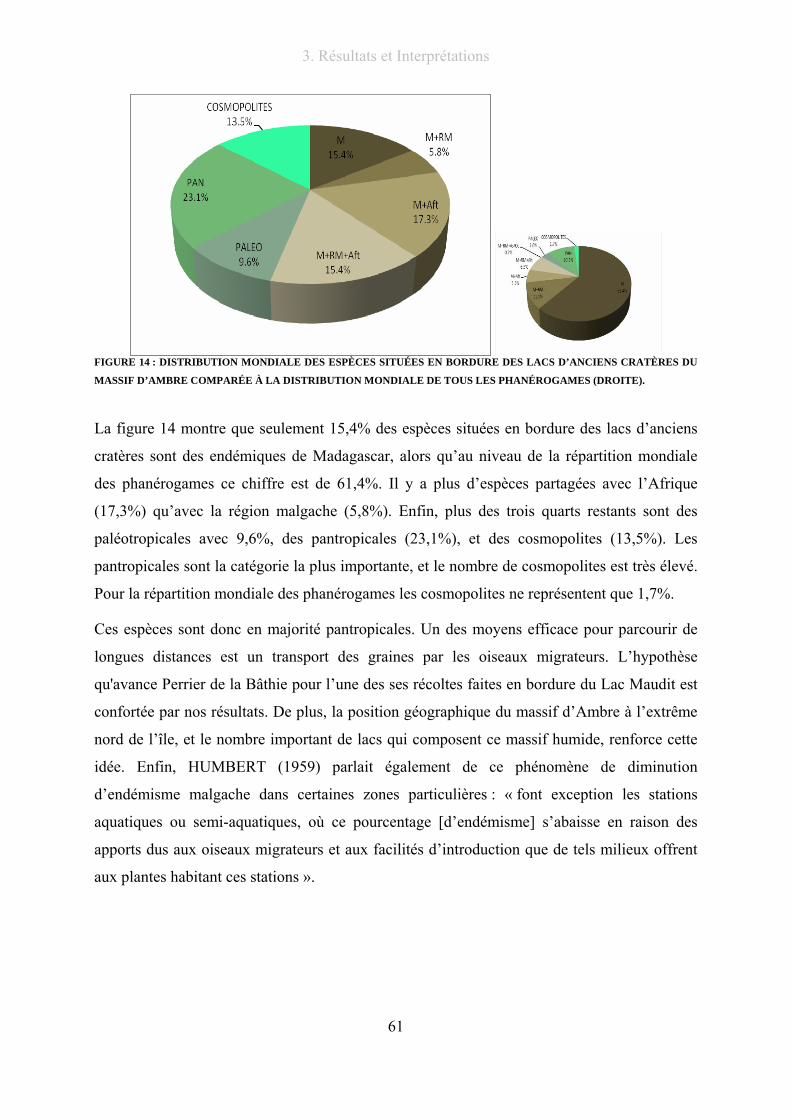
3. Résultats et Interprétations
61
FIGURE 14 : DISTRIBUTION MONDIALE DES ESPÈCES SITUÉES EN BORDURE DES LACS D’ANCIENS CRATÈRES DU
MASSIF D’AMBRE COMPARÉE À LA DISTRIBUTION MONDIALE DE TOUS LES PHANÉROGAMES (DROITE).
La figure 14 montre que seulement 15,4% des espèces situées en bordure des lacs d’anciens
cratères sont des endémiques de Madagascar, alors qu’au niveau de la répartition mondiale
des phanérogames ce chiffre est de 61,4%. Il y a plus d’espèces partagées avec l’Afrique
(17,3%) qu’avec la région malgache (5,8%). Enfin, plus des trois quarts restants sont des
paléotropicales avec 9,6%, des pantropicales (23,1%), et des cosmopolites (13,5%). Les
pantropicales sont la catégorie la plus importante, et le nombre de cosmopolites est très élevé.
Pour la répartition mondiale des phanérogames les cosmopolites ne représentent que 1,7%.
Ces espèces sont donc en majorité pantropicales. Un des moyens efficace pour parcourir de
longues distances est un transport des graines par les oiseaux migrateurs. L’hypothèse
qu'avance Perrier de la Bâthie pour l’une des ses récoltes faites en bordure du Lac Maudit est
confortée par nos résultats. De plus, la position géographique du massif d’Ambre à l’extrême
nord de l’île, et le nombre important de lacs qui composent ce massif humide, renforce cette
idée. Enfin, HUMBERT (1959) parlait également de ce phénomène de diminution
d’endémisme malgache dans certaines zones particulières : « font exception les stations
aquatiques ou semi-aquatiques, où ce pourcentage [d’endémisme] s’abaisse en raison des
apports dus aux oiseaux migrateurs et aux facilités d’introduction que de tels milieux offrent
aux plantes habitant ces stations ».

3. Résultats et Interprétations
62
3.4. LA FLORE DU MASSIF DANS LE CONTEXTE MALGACHE
Les domaines phytogéographiques de HUMBERT (1965) ont été utilisés pour classer la flore
du massif sur l’île de Madagascar. Un regroupement des répartitions géographiques des
espèces a permis de distinguer 16 catégories phytogéographiques différentes. Le domaine du
Centre et celui des Hautes Montagnes sont comptabilisés ensemble tout comme le domaine de
l’Ouest et celui du Sud.
3.4.1. DISTRIBUTION À MADAGASCAR
La répartition malgache de toutes les espèces inventoriées a fait l’objet d’une analyse. Des
catégories phytogéographiques ont été obtenues après regroupement des données de
distribution. Pour uniformiser les graphiques, un code de couleur a été adopté et se retrouve
dans toutes les figures suivantes. D’une part, les catégories phytogéographiques représentées
par une couleur de ton bleu sont celles qui sont composées uniquement des domaines humides
de l’île (Centre, Est, Sambirano), ce sont les espèces dites humides. D’autre part, les
catégories phytogéographiques symbolisées par une couleur de ton rouge sont celles où le
domaine de l’Ouest et du Sud sont présents se sont donc des espèces généralistes, à
l’exception de l’une de ces catégories qui est constituée d'espèces présentes uniquement dans
les domaines secs de l’Ouest et/ou du Sud, correspondant aux espèces dites sèches. De plus,
une catégorie de couleur verte M [MdA] réunit les endémiques locales ainsi que les espèces
dont la répartition malgache n’est connue qu’au massif d’Ambre mais possédant une
répartition dans d’autres régions du monde.
Concernant la distribution des phanérogames du massif d’Ambre (figure 15, p. 63), il y a plus
de 47% de la flore qui est composée des catégories de la flore humide de Madagascar. Les
catégories incluant le domaine de l’ouest et du sud sont également dans les mêmes
proportions avec plus de 45%. Les espèces endémiques locales au sens large représenté par M
[MdA] sont présentes à raison de 6,3%. Au sein des catégories typiques de la flore humide,
trois sont dominantes. Ce sont les espèces du domaine du Centre (C), les espèces du Centre et
de l’Est (E-C), et les espèces retrouvées au Sambirano, à l’Est et au Centre (Sb, E, C). Cette
dernière représente la catégorie des espèces à large répartition en milieu humide. Quatre
autres catégories ne sont représentées que faiblement ce sont : E ; Sb ; Sb-C, et la catégorie du
Sb-E.
Pour les catégories de tons rouges, trois d’entre elles se démarquent des autres. Il y a tout
d’abord la catégorie des espèces malgaches, retrouvées dans tous les domaines de l’île (Sb-E-
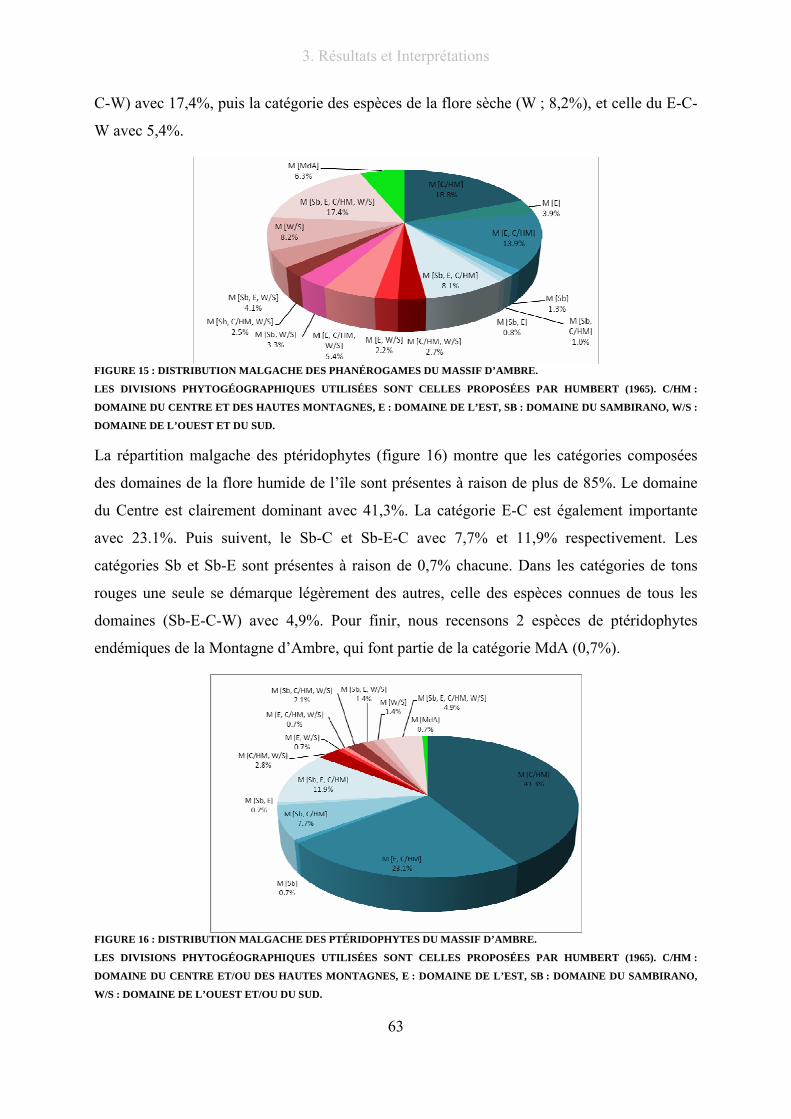
3. Résultats et Interprétations
63
C-W) avec 17,4%, puis la catégorie des espèces de la flore sèche (W ; 8,2%), et celle du E-C-
W avec 5,4%.
FIGURE 15 : DISTRIBUTION MALGACHE DES PHANÉROGAMES DU MASSIF D’AMBRE.
LES DIVISIONS PHYTOGÉOGRAPHIQUES UTILISÉES SONT CELLES PROPOSÉES PAR HUMBERT (1965). C/HM :
DOMAINE DU CENTRE ET DES HAUTES MONTAGNES, E : DOMAINE DE L’EST, SB : DOMAINE DU SAMBIRANO, W/S :
DOMAINE DE L’OUEST ET DU SUD.
La répartition malgache des ptéridophytes (figure 16) montre que les catégories composées
des domaines de la flore humide de l’île sont présentes à raison de plus de 85%. Le domaine
du Centre est clairement dominant avec 41,3%. La catégorie E-C est également importante
avec 23.1%. Puis suivent, le Sb-C et Sb-E-C avec 7,7% et 11,9% respectivement. Les
catégories Sb et Sb-E sont présentes à raison de 0,7% chacune. Dans les catégories de tons
rouges une seule se démarque légèrement des autres, celle des espèces connues de tous les
domaines (Sb-E-C-W) avec 4,9%. Pour finir, nous recensons 2 espèces de ptéridophytes
endémiques de la Montagne d’Ambre, qui font partie de la catégorie MdA (0,7%).
FIGURE 16 : DISTRIBUTION MALGACHE DES PTÉRIDOPHYTES DU MASSIF D’AMBRE.
LES DIVISIONS PHYTOGÉOGRAPHIQUES UTILISÉES SONT CELLES PROPOSÉES PAR HUMBERT (1965). C/HM :
DOMAINE DU CENTRE ET/OU DES HAUTES MONTAGNES, E : DOMAINE DE L’EST, SB : DOMAINE DU SAMBIRANO,
W/S : DOMAINE DE L’OUEST ET/OU DU SUD.
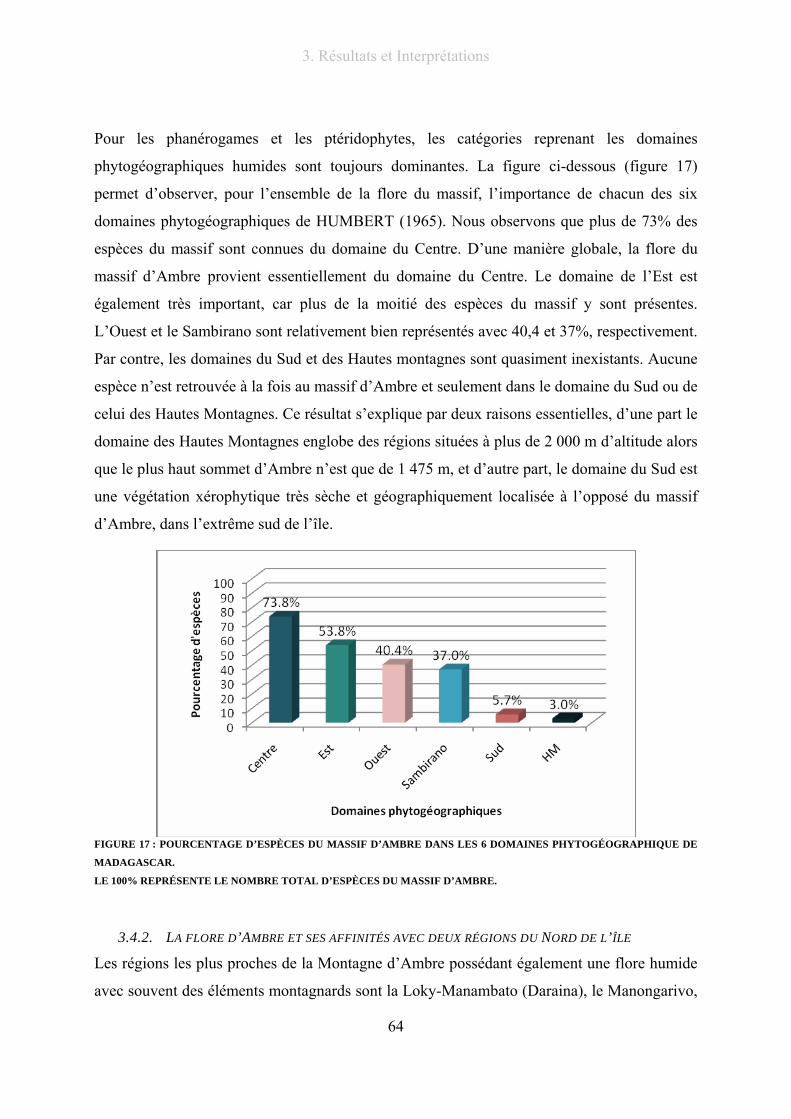
3. Résultats et Interprétations
64
Pour les phanérogames et les ptéridophytes, les catégories reprenant les domaines
phytogéographiques humides sont toujours dominantes. La figure ci-dessous (figure 17)
permet d’observer, pour l’ensemble de la flore du massif, l’importance de chacun des six
domaines phytogéographiques de HUMBERT (1965). Nous observons que plus de 73% des
espèces du massif sont connues du domaine du Centre. D’une manière globale, la flore du
massif d’Ambre provient essentiellement du domaine du Centre. Le domaine de l’Est est
également très important, car plus de la moitié des espèces du massif y sont présentes.
L’Ouest et le Sambirano sont relativement bien représentés avec 40,4 et 37%, respectivement.
Par contre, les domaines du Sud et des Hautes montagnes sont quasiment inexistants. Aucune
espèce n’est retrouvée à la fois au massif d’Ambre et seulement dans le domaine du Sud ou de
celui des Hautes Montagnes. Ce résultat s’explique par deux raisons essentielles, d’une part le
domaine des Hautes Montagnes englobe des régions situées à plus de 2 000 m d’altitude alors
que le plus haut sommet d’Ambre n’est que de 1 475 m, et d’autre part, le domaine du Sud est
une végétation xérophytique très sèche et géographiquement localisée à l’opposé du massif
d’Ambre, dans l’extrême sud de l’île.
FIGURE 17 : POURCENTAGE D’ESPÈCES DU MASSIF D’AMBRE DANS LES 6 DOMAINES PHYTOGÉOGRAPHIQUE DE
MADAGASCAR.
LE 100% REPRÉSENTE LE NOMBRE TOTAL D’ESPÈCES DU MASSIF D’AMBRE.
3.4.2. LA FLORE D’AMBRE ET SES AFFINITÉS AVEC DEUX RÉGIONS DU NORD DE L’ÎLE
Les régions les plus proches de la Montagne d’Ambre possédant également une flore humide
avec souvent des éléments montagnards sont la Loky-Manambato (Daraina), le Manongarivo,

3. Résultats et Interprétations
65
le Sambirano, le Tsaratanana, le Marojejy, et d’autres petits massifs forestiers des alentours.
Nous avons eu accès à des listes floristiques pour les deux premières régions: le Daraina et le
Manongarivo que nous avons utilisées ici pour notre comparaison.
Le nombre de genres communs entre le massif d’Ambre et le Daraina est de 211, tandis que
ce chiffre est de 360 avec le Manongarivo (chapitre 3.2.3., p. 51). Le nombre d’espèces
partagées entre la Montagne d’Ambre et le Daraina est de 273, alors qu’entre le massif
d’Ambre et le Manongarivo ce chiffre est de 289. Ces résultats sont comparables, toutefois il
y a légèrement plus de ressemblance au niveau de la composition floristique entre le massif
d’Ambre et le Manongarivo, qu’entre le massif d’Ambre et le Daraina. Des comparaisons au
niveau phytogéographique permettent de distinguer le Manongarivo du Daraina en
comparaison avec le massif d’Ambre.
La répartition selon les domaines phytogéographiques de l’île des espèces d’Ambre partagées
avec la région du Daraina est illustrée sur la figure 18, p. 66 et celles partagées avec le
Manongarivo sur la figure 19, p. 66. La comparaison entre ces deux figures montre que les
espèces retrouvées dans la région du Daraina sont plutôt des généralistes où des espèces
sèches, tandis que les espèces partagées avec le Manongarivo sont plus humides. Les massifs
humides de la région du Daraina sont, tout comme le massif d’Ambre, situés dans une flore
environnante de type sèche appartenant au domaine de l’Ouest secteur Nord. Pour cette
raison, un nombre important des espèces communes entre ces deux régions sont réparties dans
ce domaine de l’Ouest qui relie ces deux régions. Concernant le Manongarivo, ce massif est
en connexion directe avec la chaîne de montagne du Centre de l’île de Madagascar composée
notamment d’une flore humide du domaine du Centre. C’est ce qui explique que les espèces
partagées entre la Montagne d’Ambre et le Manongarivo sont surtout recensées dans les
domaines humides de l’île.

3. Résultats et Interprétations
66
FIGURE 18 : RÉPARTITION DES ESPÈCES PRÉSENTES À LA MONTAGNE D’AMBRE ET AU DARAINA SELON LES
DOMAINES PHYTOGÉOGAPHIQUES DE L’ÎLE.
FIGURE 19 : RÉPARTITION DES ESPÈCES PRÉSENTES À LA MONTAGNE D’AMBRE ET AU MANONGARIVO SELON
LES DOMAINES PHYTOGÉOGAPHIQUES DE L’ÎLE.
3.5. ETUDE DES ÉTAGES DE VÉGÉTATION AU SEIN DU MASSIF D’AMBRE
Les figures du spectre chorologique des phanérogames et ptéridophytes (ci-dessus) ont permis
d’observer, d’une manière générale, la proportion des catégories phytogéographiques et
d’observer dans quels domaines phytogéographiques nous retrouvons le plus d’espèces de la
Montagne d’Ambre. Cependant, si nous observons ce spectre à des altitudes différentes, il est
probable que la composition des catégories phytogéographiques évolue également. La
question à laquelle ce chapitre se propose de répondre est comment cette composition varie
avec l’altitude.
3.5.1. RÉPARTITION ALTITUDINALE DES CATÉGORIES PHYTOGÉOGRAPHIQUES
Pour la figure 20, p. 68 les données altitudinales des espèces sont issues des échantillons
d’herbiers du massif d’Ambre. L’altitude de chaque échantillon figure dans la liste floristique
(Annexe 8.1.1., p. 101 et Annexe 8.1.2., p. 178). Les altitudes approximées ont également
été prises en compte.

3. Résultats et Interprétations
67
Chaque espèce possède une répartition altitudinale plus ou moins restreinte. Les espèces à
large répartition peuvent donc figurer dans différentes tranches altitudinales. Toutefois, au
sein d’une tranche altitudinale, chaque espèce n’est comptabilisée qu’une seule fois, même si
celle-ci a été récoltée plusieurs fois.
Pour délimiter le domaine du Centre, HUMBERT (1965) cite une limite altitudinale inférieure
à 800 m car en-dessous « l’influence des vents descendants échauffés détermine un climat à
longue saison sèche ». La figure 20 montre que les espèces retrouvées uniquement au
domaine du Centre et dans celui des Hautes Montagnes (C/HM) sont plus importantes en-
dessus de 1 000 m d’altitude, et que les espèces appartenant aux domaines de l’Ouest et du
Sud sont plus importantes aux altitudes inférieures. Les espèces humides de basses altitudes
(E, Sb et Sb-E) sont les mieux représentées à des altitudes intermédiaires entre 1 000 et
800 m. Ces espèces sont restreintes à ces altitudes intermédiaires, car les conditions
climatiques sont trop humides dans les altitudes supérieures et à l’inverse trop sèches aux
altitudes inférieures du massif. Nous remarquons que le spectre chorologique varie selon
l’altitude. Cependant, il y a une « infiltration » des flores entre les étages de végétation. Nous
observons par exemple que les espèces restreintes au domaine de l’Ouest sont encore en grand
nombre à des altitudes avoisinant les 1 000 m et même au-dessus, de même que des espèces
réparties uniquement dans le domaine du Centre et Hautes Montagnes sont présentes en
dessous de 800 m. GUILLAUMET & KOECHLIN (1971) soulignent que «la flore sera
évidemment très variable en fonction des conditions climatiques locales, de l’altitude, de la
nature de la roche ». Les conditions climatiques et édaphiques modulent ces limites
altitudinales et des milieux favorables aux espèces du domaine sec de l’Ouest peuvent se
rencontrer au-dessus de 800 m, et inversement pour les espèces appartenant au domaine
humide d’altitude du Centre.
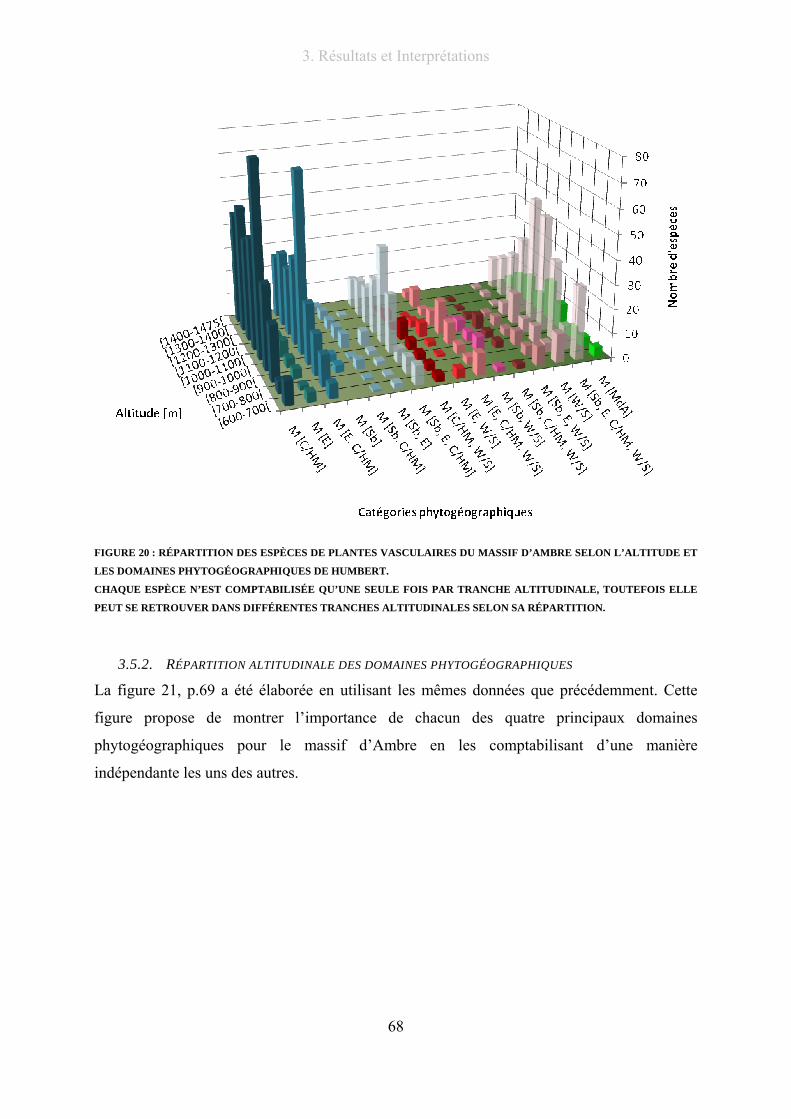
3. Résultats et Interprétations
68
FIGURE 20 : RÉPARTITION DES ESPÈCES DE PLANTES VASCULAIRES DU MASSIF D’AMBRE SELON L’ALTITUDE ET
LES DOMAINES PHYTOGÉOGRAPHIQUES DE HUMBERT.
CHAQUE ESPÈCE N’EST COMPTABILISÉE QU’UNE SEULE FOIS PAR TRANCHE ALTITUDINALE, TOUTEFOIS ELLE
PEUT SE RETROUVER DANS DIFFÉRENTES TRANCHES ALTITUDINALES SELON SA RÉPARTITION.
3.5.2. RÉPARTITION ALTITUDINALE DES DOMAINES PHYTOGÉOGRAPHIQUES
La figure 21, p.69 a été élaborée en utilisant les mêmes données que précédemment. Cette
figure propose de montrer l’importance de chacun des quatre principaux domaines
phytogéographiques pour le massif d’Ambre en les comptabilisant d’une manière
indépendante les uns des autres.
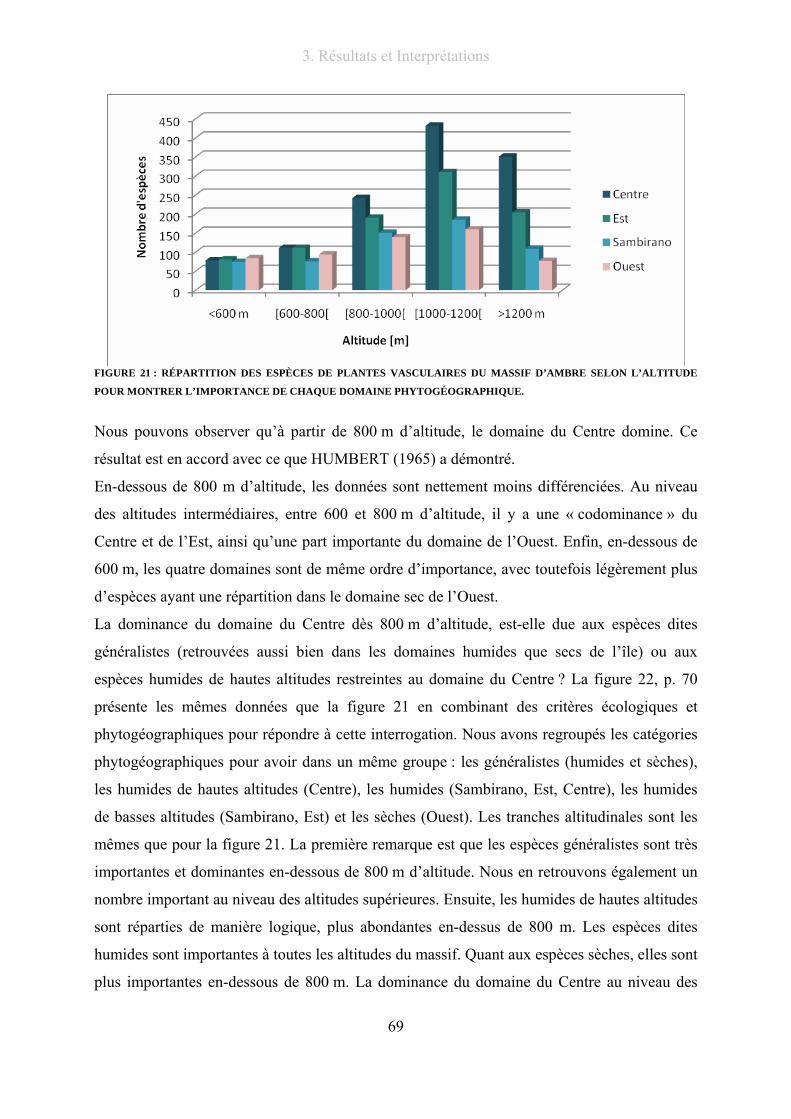
3. Résultats et Interprétations
69
FIGURE 21 : RÉPARTITION DES ESPÈCES DE PLANTES VASCULAIRES DU MASSIF D’AMBRE SELON L’ALTITUDE
POUR MONTRER L’IMPORTANCE DE CHAQUE DOMAINE PHYTOGÉOGRAPHIQUE.
Nous pouvons observer qu’à partir de 800 m d’altitude, le domaine du Centre domine. Ce
résultat est en accord avec ce que HUMBERT (1965) a démontré.
En-dessous de 800 m d’altitude, les données sont nettement moins différenciées. Au niveau
des altitudes intermédiaires, entre 600 et 800 m d’altitude, il y a une « codominance » du
Centre et de l’Est, ainsi qu’une part importante du domaine de l’Ouest. Enfin, en-dessous de
600 m, les quatre domaines sont de même ordre d’importance, avec toutefois légèrement plus
d’espèces ayant une répartition dans le domaine sec de l’Ouest.
La dominance du domaine du Centre dès 800 m d’altitude, est-elle due aux espèces dites
généralistes (retrouvées aussi bien dans les domaines humides que secs de l’île) ou aux
espèces humides de hautes altitudes restreintes au domaine du Centre ? La figure 22, p. 70
présente les mêmes données que la figure 21 en combinant des critères écologiques et
phytogéographiques pour répondre à cette interrogation. Nous avons regroupés les catégories
phytogéographiques pour avoir dans un même groupe : les généralistes (humides et sèches),
les humides de hautes altitudes (Centre), les humides (Sambirano, Est, Centre), les humides
de basses altitudes (Sambirano, Est) et les sèches (Ouest). Les tranches altitudinales sont les
mêmes que pour la figure 21. La première remarque est que les espèces généralistes sont très
importantes et dominantes en-dessous de 800 m d’altitude. Nous en retrouvons également un
nombre important au niveau des altitudes supérieures. Ensuite, les humides de hautes altitudes
sont réparties de manière logique, plus abondantes en-dessus de 800 m. Les espèces dites
humides sont importantes à toutes les altitudes du massif. Quant aux espèces sèches, elles sont
plus importantes en-dessous de 800 m. La dominance du domaine du Centre au niveau des
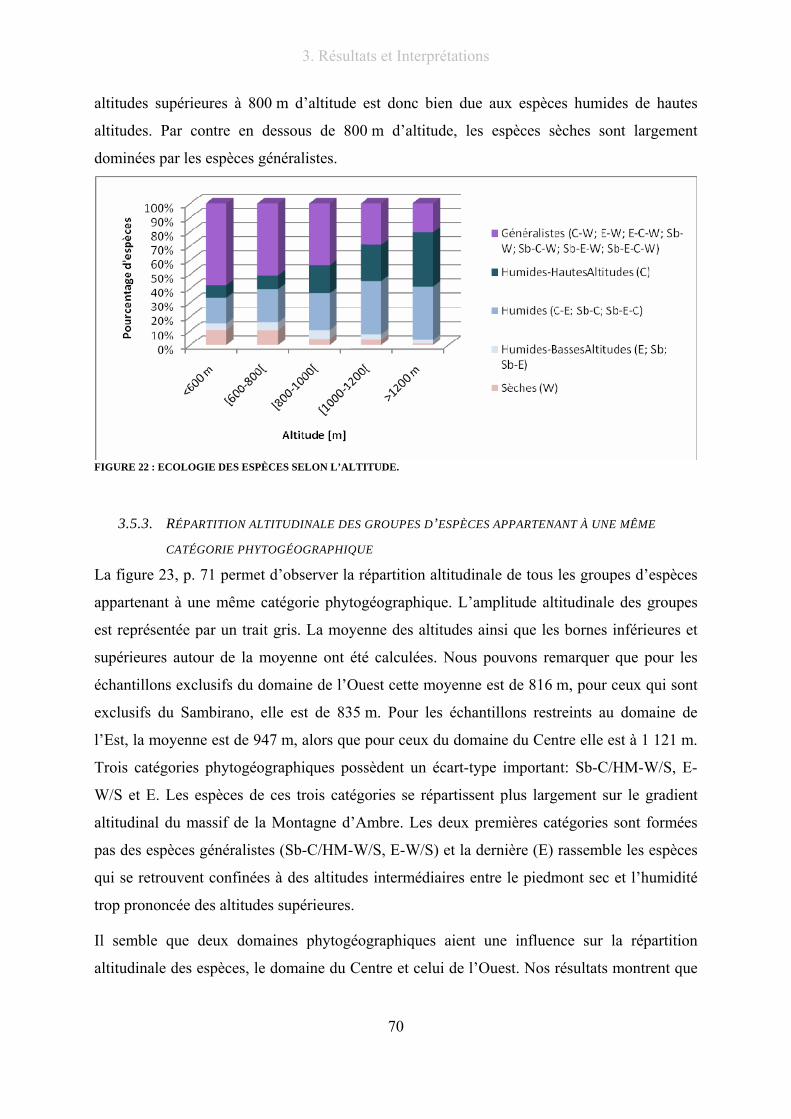
3. Résultats et Interprétations
70
altitudes supérieures à 800 m d’altitude est donc bien due aux espèces humides de hautes
altitudes. Par contre en dessous de 800 m d’altitude, les espèces sèches sont largement
dominées par les espèces généralistes.
FIGURE 22 : ECOLOGIE DES ESPÈCES SELON L’ALTITUDE.
3.5.3. RÉPARTITION ALTITUDINALE DES GROUPES D’ESPÈCES APPARTENANT À UNE MÊME
CATÉGORIE PHYTOGÉOGRAPHIQUE
La figure 23, p. 71 permet d’observer la répartition altitudinale de tous les groupes d’espèces
appartenant à une même catégorie phytogéographique. L’amplitude altitudinale des groupes
est représentée par un trait gris. La moyenne des altitudes ainsi que les bornes inférieures et
supérieures autour de la moyenne ont été calculées. Nous pouvons remarquer que pour les
échantillons exclusifs du domaine de l’Ouest cette moyenne est de 816 m, pour ceux qui sont
exclusifs du Sambirano, elle est de 835 m. Pour les échantillons restreints au domaine de
l’Est, la moyenne est de 947 m, alors que pour ceux du domaine du Centre elle est à 1 121 m.
Trois catégories phytogéographiques possèdent un écart-type important: Sb-C/HM-W/S, E-
W/S et E. Les espèces de ces trois catégories se répartissent plus largement sur le gradient
altitudinal du massif de la Montagne d’Ambre. Les deux premières catégories sont formées
pas des espèces généralistes (Sb-C/HM-W/S, E-W/S) et la dernière (E) rassemble les espèces
qui se retrouvent confinées à des altitudes intermédiaires entre le piedmont sec et l’humidité
trop prononcée des altitudes supérieures.
Il semble que deux domaines phytogéographiques aient une influence sur la répartition
altitudinale des espèces, le domaine du Centre et celui de l’Ouest. Nos résultats montrent que
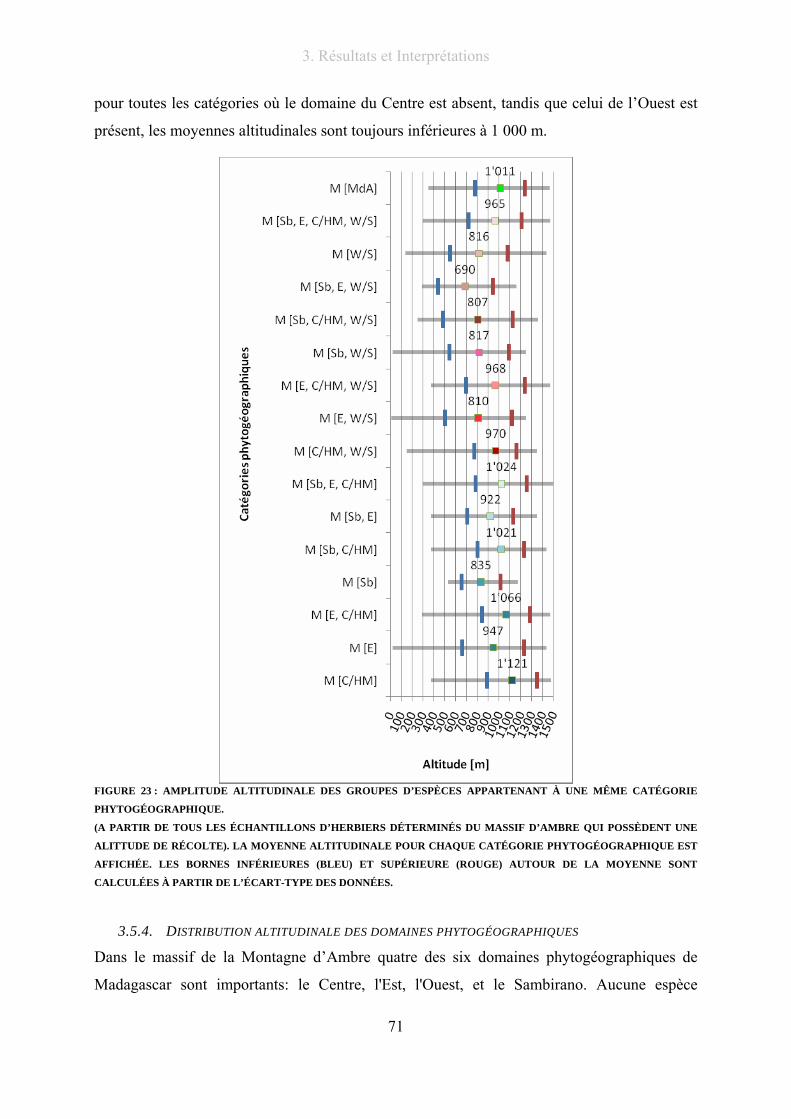
3. Résultats et Interprétations
71
pour toutes les catégories où le domaine du Centre est absent, tandis que celui de l’Ouest est
présent, les moyennes altitudinales sont toujours inférieures à 1 000 m.
FIGURE 23 : AMPLITUDE ALTITUDINALE DES GROUPES D’ESPÈCES APPARTENANT À UNE MÊME CATÉGORIE
PHYTOGÉOGRAPHIQUE.
(A PARTIR DE TOUS LES ÉCHANTILLONS D’HERBIERS DÉTERMINÉS DU MASSIF D’AMBRE QUI POSSÈDENT UNE
ALITTUDE DE RÉCOLTE). LA MOYENNE ALTITUDINALE POUR CHAQUE CATÉGORIE PHYTOGÉOGRAPHIQUE EST
AFFICHÉE. LES BORNES INFÉRIEURES (BLEU) ET SUPÉRIEURE (ROUGE) AUTOUR DE LA MOYENNE SONT
CALCULÉES À PARTIR DE L’ÉCART-TYPE DES DONNÉES.
3.5.4. DISTRIBUTION ALTITUDINALE DES DOMAINES PHYTOGÉOGRAPHIQUES
Dans le massif de la Montagne d’Ambre quatre des six domaines phytogéographiques de
Madagascar sont importants: le Centre, l'Est, l'Ouest, et le Sambirano. Aucune espèce

3. Résultats et Interprétations
72
présente à la Montagne d’Ambre n’est distribuée exclusivement au domaine du Sud ni dans
celui des Hautes Montagnes.
Concernant le domaine du Centre, nous avons pu remarquer qu’il domine au-dessus de 800 m
et que cette dominance est due aux espèces dites humides (C-E, C-Sb et C-E-Sb) et aux
humides de hautes altitudes (C). Entre 600 et 800 m le Centre est également important cette
fois-ci à cause du poids considérable des espèces généralistes. En moyenne, les espèces
distribuées au domaine du Centre et Hautes Montagnes se trouvent à 1 121 m d’altitude. Nous
pouvons donc conclure que le domaine du Centre domine les altitudes supérieures à 800 m,
conformément à la théorie de Humbert.
En dessous de 800 m, les espèces généralistes sont largement dominantes. Il en ressort que les
domaines phytogéographiques sont répartis équitablement dans chaque tranche altitudinale en
dessous de 800 m. Aucune tendance phytogéographique n’est visible. Toutefois, nous avons
pu remarquer une augmentation des espèces sèches à ces altitudes inférieures, et une légère
dominance du domaine de l’Ouest aux altitudes inférieures à 600 m.
Nous pouvons donc effectuer une première synthèse de la répartition altitudinale des
différents domaines. Du sommet au piedmont, nous retrouvons les domaines du : Centre, de
l’Est, quelques éléments du Sambirano et l’Ouest. Le tableau 1, p. 73 présente un résumé de
cette synthèse au niveau de la répartition altitudinale des domaines phytogéographiques pour
la Montagne d’Ambre. L’écologie des espèces dominantes selon chaque tranche altitudinale
(figure 22, p. 70) est également notée. Cette synthèse permet notamment de mettre en
évidence le poids des espèces généralistes. Concernant les altitudes inférieures à 800 m, un
échantillonnage plus important serait toutefois nécessaire pour confirmer ou modifier nos
résultats.
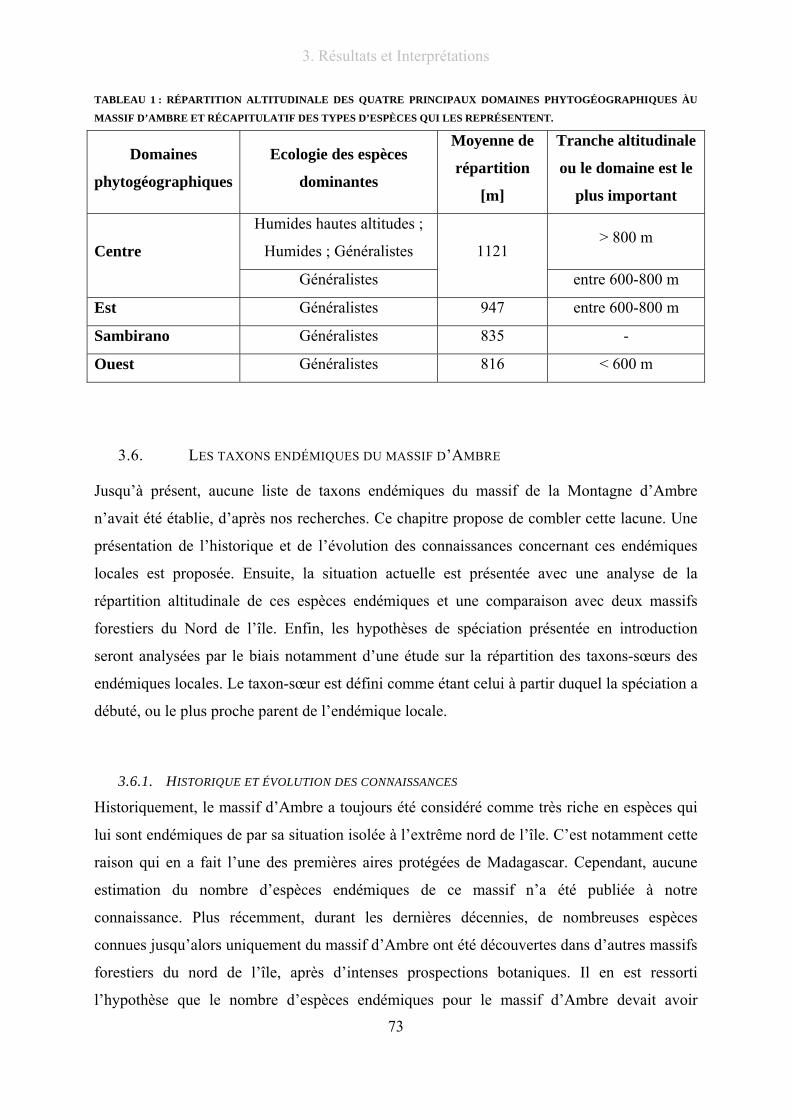
3. Résultats et Interprétations
73
TABLEAU 1 : RÉPARTITION ALTITUDINALE DES QUATRE PRINCIPAUX DOMAINES PHYTOGÉOGRAPHIQUES ÀU
MASSIF D’AMBRE ET RÉCAPITULATIF DES TYPES D’ESPÈCES QUI LES REPRÉSENTENT.
Domaines
phytogéographiques
Ecologie des espèces
dominantes
Moyenne de
répartition
[m]
Tranche altitudinale
ou le domaine est le
plus important
Humides hautes altitudes ;
Humides ; Généralistes > 800 m
Centre
Généralistes
1121
entre 600-800 m
Est Généralistes 947 entre 600-800 m
Sambirano Généralistes 835 -
Ouest Généralistes 816 < 600 m
3.6. LES TAXONS ENDÉMIQUES DU MASSIF D’AMBRE
Jusqu’à présent, aucune liste de taxons endémiques du massif de la Montagne d’Ambre
n’avait été établie, d’après nos recherches. Ce chapitre propose de combler cette lacune. Une
présentation de l’historique et de l’évolution des connaissances concernant ces endémiques
locales est proposée. Ensuite, la situation actuelle est présentée avec une analyse de la
répartition altitudinale de ces espèces endémiques et une comparaison avec deux massifs
forestiers du Nord de l’île. Enfin, les hypothèses de spéciation présentée en introduction
seront analysées par le biais notamment d’une étude sur la répartition des taxons-sœurs des
endémiques locales. Le taxon-sœur est défini comme étant celui à partir duquel la spéciation a
débuté, ou le plus proche parent de l’endémique locale.
3.6.1. HISTORIQUE ET ÉVOLUTION DES CONNAISSANCES
Historiquement, le massif d’Ambre a toujours été considéré comme très riche en espèces qui
lui sont endémiques de par sa situation isolée à l’extrême nord de l’île. C’est notamment cette
raison qui en a fait l’une des premières aires protégées de Madagascar. Cependant, aucune
estimation du nombre d’espèces endémiques de ce massif n’a été publiée à notre
connaissance. Plus récemment, durant les dernières décennies, de nombreuses espèces
connues jusqu’alors uniquement du massif d’Ambre ont été découvertes dans d’autres massifs
forestiers du nord de l’île, après d’intenses prospections botaniques. Il en est ressorti
l’hypothèse que le nombre d’espèces endémiques pour le massif d’Ambre devait avoir

3. Résultats et Interprétations
74
nettement diminué. Toutefois, d’intenses prospections botaniques dans le massif d’Ambre, ont
récemment aboutis à la découverte de nouveaux taxons, dont certains y sont probablement
endémiques. Le fait qui en découle est le suivant : plus il y a de prospections à la Montagne
d’Ambre, plus le nombre d’endémiques augmente et parallèlement, plus il y a de prospections
dans d’autres régions du nord de l’île, plus ce nombre diminue.
Pour appréhender ce phénomène, nous avons observé la répartition actuelle des taxons
comportant le radical « ambrens* » dans leur épithète spécifique ou infraspécifique. Après
analyse de notre liste des espèces présentes à Madagascar, nous avons obtenu un
échantillonnage de 50 taxons. Les échantillons possédant dans leur publication originale la
citation de récoltes effectuées dans d’autres régions qu’à la Montagne d’Ambre ont été
écartés. Les 41 taxons que nous avons conservés étaient donc bien considérés à l’origine
comme endémiques de la Montagne d’Ambre. La figure 24, p. 75 représente ces 41 taxons en
fonction de leur année de publication et de leur considération actuelle selon trois catégories :
‐ taxons toujours considérés comme endémiques actuellement (vert),
‐ taxons tombés en synonymie d'autres taxons à plus large répartition (en rouge),
‐ taxons trouvés depuis dans d’autres régions de l’île (en bleu).
Nous pouvons remarquer que les synonymies, qui nécessitent une étude approfondie du
groupe considéré, sont effectuées en large majorité pour des taxons décrits il y a plus de
60 ans. Pour les taxons décrits entre 1951 et 2000 et considérés maintenant comme non
endémiques: un seul a été mis en synonymie, et 5 autres ont été récoltés dans d’autres régions
de l’île. Sur ces 41 taxons, 25 se trouvent aujourd'hui encore dans notre liste des endémiques
du massif d’Ambre.

3. Résultats et Interprétations
75
FIGURE 24 : ANNÉE DE DESCRIPTION DES 41 TAXONS DONT LA DIAGNOSE NE CITE QU’UN ÉCHANTILLON DU
MASSIF D’AMBRE ET COMPORTANT LE RADICAL « AMBRENS » ET LEUR ACTUELLE CONSIDÉRATION.
Neuf espèces et une variété ont été retrouvées ailleurs à Madagascar. Les neuf espèces sont
listées ci-dessous:
‐ A : Monanthotaxis ambrensis (Cavaco & Keraudren) Verdc. ; M [W(N)]
‐ B : Lepidotrichilia ambrensis J.-F.Leroy ; M [C, W(N)] - Aft
‐ C : Nuxia ambrensis Jovet ; M [C]
‐ D : Pittosporum ambrense Cufod. ; M [C]
‐ E : Tricalysia ambrensis Randriamb. & De Block. ; M [C]
‐ F : Garcinia ambrensis (H. Perrier) P. Sweeney & Z.S. Rogers ; M [C]
‐ G : Bulbophyllum ambrense H.Perrier ; M [E]
‐ H : Olea ambrensis H.Perrier ; M [E, C]
‐ I : Faucherea ambrensis Capuron ex Aubrév. ; M [E, C]
Leur répartition actuelle sur l’île de Madagascar est illustrée sur la carte 7, p. 77.
Nous remarquons que deux espèces sont retrouvées dans le domaine de l’Ouest. Leur
répartition est toutefois restreinte au secteur Nord du domaine de l’Ouest.
Une seule espèce possède une distribution partagée avec le domaine de l’Est uniquement,
c’est Bulbophyllum ambrense H.Perrier. Nous remarquons que la carte de distribution de cette
espèce (G) montre que les échantillons récoltés hors du massif étudié sont localisés dans la

3. Résultats et Interprétations
76
partie Nord du domaine de l’Est. Toutes les autres espèces distribuées à la fois dans le
domaine du Centre et de l’Est, sont présentes à de plus basses latitudes, et présentent donc une
répartition géographique plus large. Quatre espèces sont exclusives du domaine du Centre.
Une seule possède une répartition allant au-delà du seuil de Mandritsara, dans le Centre
moyen, c’est Garcinia ambrensis (H. Perrier) P. Sweeney & Z.S. Rogers (F). La répartition
géographique de Tricalysia ambrensis Randriamb. & De Block. est sur ce point
caractéristique d’une espèce qui n’a pas réussi à traverser ce seuil situé dans le nord. Pour
cette espèce, l’altitude et le rétrécissement du domaine du Centre à cet endroit ont
certainement contribué à cette distribution restreinte au domaine du Centre sous-domaine du
Centre nord.
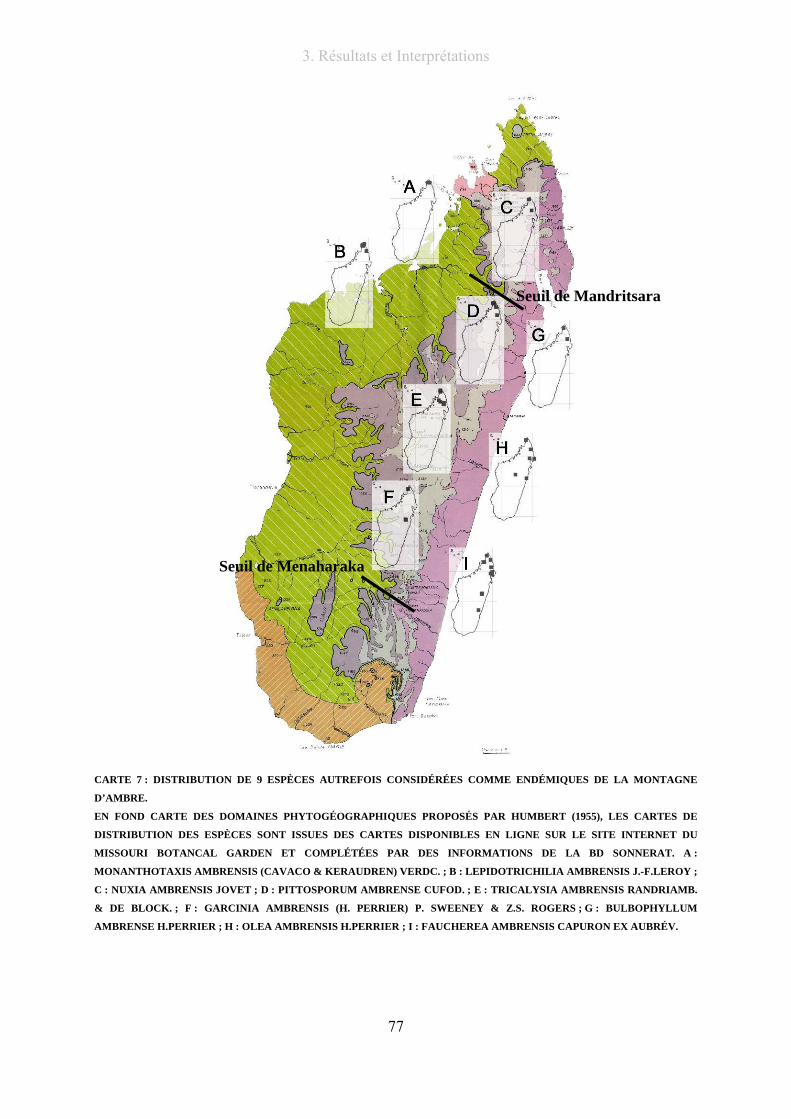
3. Résultats et Interprétations
77
CARTE 7 : DISTRIBUTION DE 9 ESPÈCES AUTREFOIS CONSIDÉRÉES COMME ENDÉMIQUES DE LA MONTAGNE
D’AMBRE.
EN FOND CARTE DES DOMAINES PHYTOGÉOGRAPHIQUES PROPOSÉS PAR HUMBERT (1955), LES CARTES DE
DISTRIBUTION DES ESPÈCES SONT ISSUES DES CARTES DISPONIBLES EN LIGNE SUR LE SITE INTERNET DU
MISSOURI BOTANCAL GARDEN ET COMPLÉTÉES PAR DES INFORMATIONS DE LA BD SONNERAT. A :
MONANTHOTAXIS AMBRENSIS (CAVACO & KERAUDREN) VERDC. ; B : LEPIDOTRICHILIA AMBRENSIS J.-F.LEROY ;
C : NUXIA AMBRENSIS JOVET ; D : PITTOSPORUM AMBRENSE CUFOD. ; E : TRICALYSIA AMBRENSIS RANDRIAMB.
& DE BLOCK. ; F : GARCINIA AMBRENSIS (H. PERRIER) P. SWEENEY & Z.S. ROGERS ; G : BULBOPHYLLUM
AMBRENSE H.PERRIER ; H : OLEA AMBRENSIS H.PERRIER ; I : FAUCHEREA AMBRENSIS CAPURON EX AUBRÉV.
Seuil de Mandritsara
Seuil de Menaharaka

3. Résultats et Interprétations
78
3.6.2. SITUATION ACTUELLE
Dans l'état actuel des connaissances, nous dénombrons 1 genre monospécifique (Ambrella
H.Perrier), 39 espèces et 7 taxons infraspécifiques endémiques du massif de la Montagne
d’Ambre (Annexe 8.2, p. 191). Nous pouvons ajouter cependant que ce chiffre semble être
pour l’instant sous-estimé, car la liste floristique du massif contient encore des espèces
indéterminées qui sont potentiellement nouvelles pour la Science. De plus, une nouvelle
espèce endémique du massif a récemment été décrite sur la base d’un échantillon récolté par
l’équipe du présent projet c’est le Pandanus nusbaumeri Callm. & L. Gaut (CALLMANDER
& AL., 2009). A cela s’ajoute trois espèces considérées comme des endémiques probables. En
effet, pour chacune de ces trois espèces (Malleastrum pseudodepauperatum J.-F.Leroy,
Peperomia terebinthina G.Mathieu, et Angraecum bicallosum H.Perrier), la majorité des
échantillons répertoriés dans les diverses sources d’informations consultées proviennent de la
Montagne d’Ambre à l'exception d'un seul échantillon d’herbier, auquel nous n'avons pas eu
accès et dont la détermination n'a pas été vérifiée par un spécialiste de la famille concernée.
3.6.3. RÉPARTITION ALTITUDINALE DES ESPÈCES ENDÉMIQUES
Dans le massif de la Montagne d’Ambre, les espèces d’endémiques se trouvent
principalement à plus de 900 m d’altitude. La figure 25, p. 79, illustre la répartition
altitudinale des récoltes des 46 taxons endémiques. Pour l’histogramme de gauche, chaque
taxon n’est comptabilisé qu’une seule fois par tranche altitudinale, mais peut se trouver dans
plusieurs tranches d’altitude différentes. Plus de 40% des taxons endémiques du massif
d’Ambre se situent entre 900 et 1 100 m d’altitude. En dessus de 1 100 m, la présence
d’endémiques reste toutefois plus importante, qu’en dessous de 900 m d’altitude. Il faut
néanmoins remarquer que l’allure de cet histogramme porte fortement l’empreinte du nombre
de récoltes. L’histogramme de droite est une représentation de ces mêmes chiffres par tranche
altitudinale, pondérés par le nombre total de récoltes dans chaque tranche altitudinale. Nous y
constatons que néanmoins, d’une manière générale, le nombre d’endémiques est plus
important aux altitudes supérieures, au-delà de 700 m d’altitude, et semble diminuer vers
1 300 m.
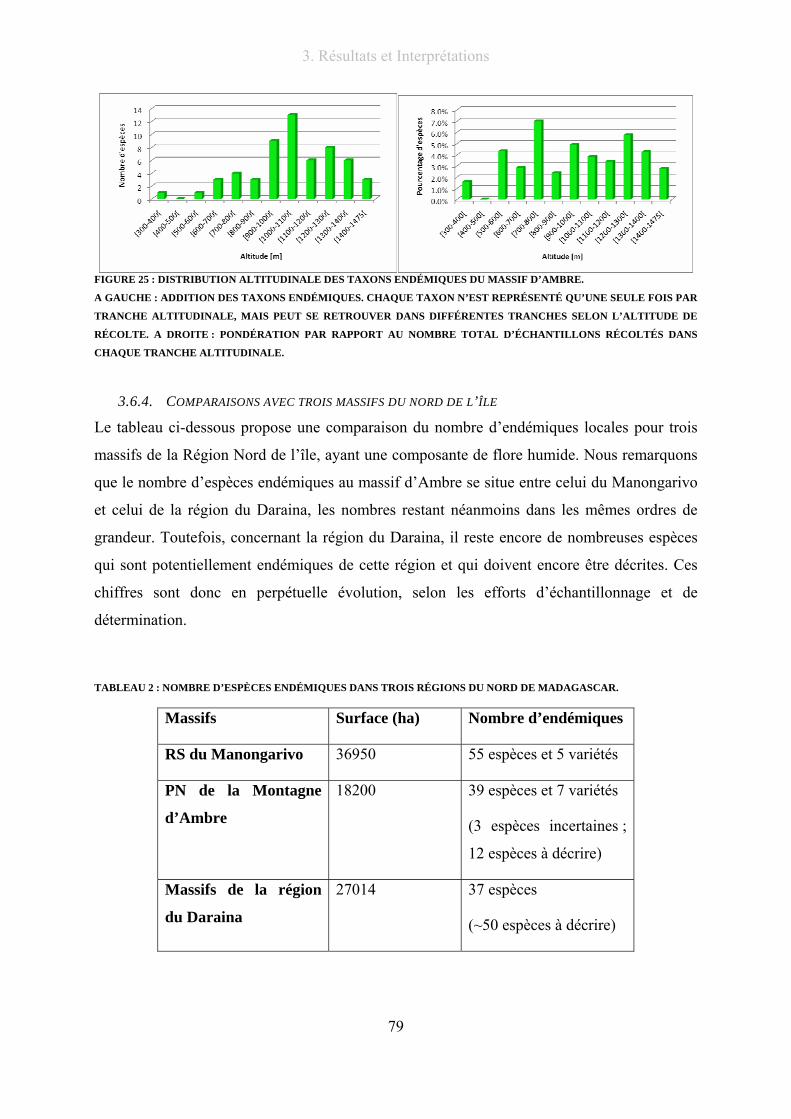
3. Résultats et Interprétations
79
FIGURE 25 : DISTRIBUTION ALTITUDINALE DES TAXONS ENDÉMIQUES DU MASSIF D’AMBRE.
A GAUCHE : ADDITION DES TAXONS ENDÉMIQUES. CHAQUE TAXON N’EST REPRÉSENTÉ QU’UNE SEULE FOIS PAR
TRANCHE ALTITUDINALE, MAIS PEUT SE RETROUVER DANS DIFFÉRENTES TRANCHES SELON L’ALTITUDE DE
RÉCOLTE. A DROITE : PONDÉRATION PAR RAPPORT AU NOMBRE TOTAL D’ÉCHANTILLONS RÉCOLTÉS DANS
CHAQUE TRANCHE ALTITUDINALE.
3.6.4. COMPARAISONS AVEC TROIS MASSIFS DU NORD DE L’ÎLE
Le tableau ci-dessous propose une comparaison du nombre d’endémiques locales pour trois
massifs de la Région Nord de l’île, ayant une composante de flore humide. Nous remarquons
que le nombre d’espèces endémiques au massif d’Ambre se situe entre celui du Manongarivo
et celui de la région du Daraina, les nombres restant néanmoins dans les mêmes ordres de
grandeur. Toutefois, concernant la région du Daraina, il reste encore de nombreuses espèces
qui sont potentiellement endémiques de cette région et qui doivent encore être décrites. Ces
chiffres sont donc en perpétuelle évolution, selon les efforts d’échantillonnage et de
détermination.
TABLEAU 2 : NOMBRE D’ESPÈCES ENDÉMIQUES DANS TROIS RÉGIONS DU NORD DE MADAGASCAR.
Massifs Surface (ha) Nombre d’endémiques
RS du Manongarivo 36950 55 espèces et 5 variétés
PN de la Montagne
d’Ambre
18200 39 espèces et 7 variétés
(3 espèces incertaines ;
12 espèces à décrire)
Massifs de la région
du Daraina
27014 37 espèces
(~50 espèces à décrire)
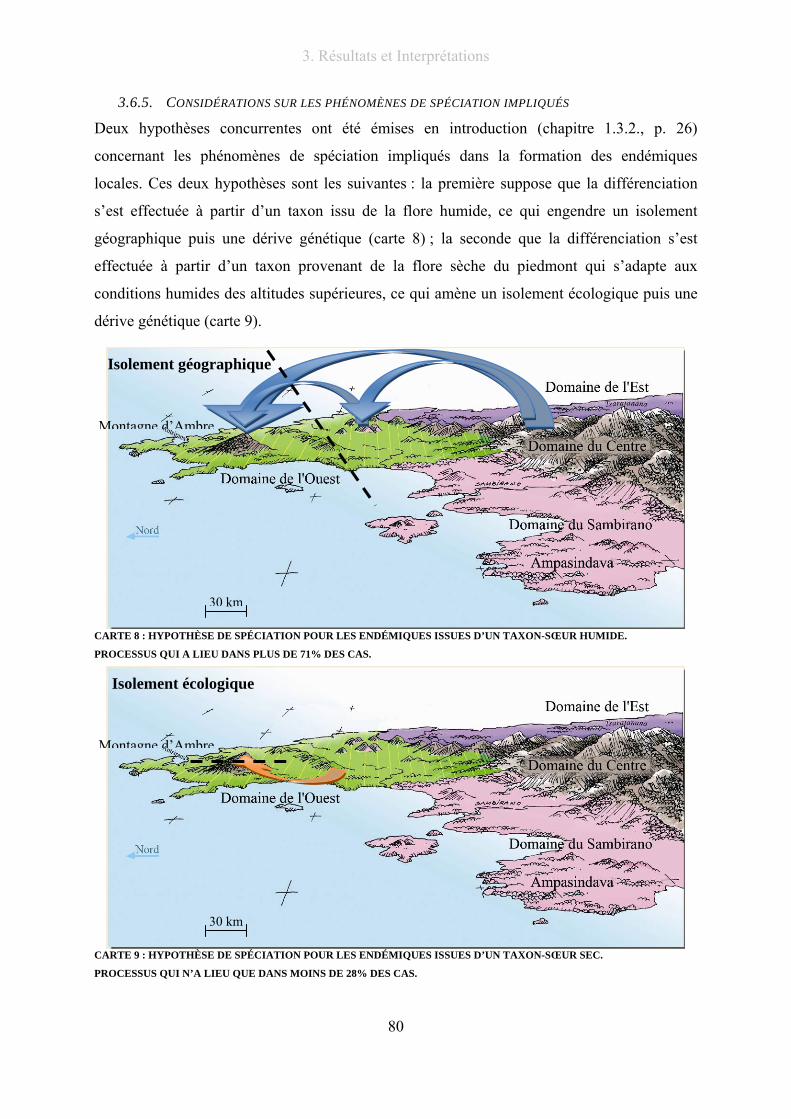
3. Résultats et Interprétations
80
3.6.5. CONSIDÉRATIONS SUR LES PHÉNOMÈNES DE SPÉCIATION IMPLIQUÉS
Deux hypothèses concurrentes ont été émises en introduction (chapitre 1.3.2., p. 26)
concernant les phénomènes de spéciation impliqués dans la formation des endémiques
locales. Ces deux hypothèses sont les suivantes : la première suppose que la différenciation
s’est effectuée à partir d’un taxon issu de la flore humide, ce qui engendre un isolement
géographique puis une dérive génétique (carte 8) ; la seconde que la différenciation s’est
effectuée à partir d’un taxon provenant de la flore sèche du piedmont qui s’adapte aux
conditions humides des altitudes supérieures, ce qui amène un isolement écologique puis une
dérive génétique (carte 9).
CARTE 8 : HYPOTHÈSE DE SPÉCIATION POUR LES ENDÉMIQUES ISSUES D’UN TAXON-SŒUR HUMIDE.
PROCESSUS QUI A LIEU DANS PLUS DE 71% DES CAS.
CARTE 9 : HYPOTHÈSE DE SPÉCIATION POUR LES ENDÉMIQUES ISSUES D’UN TAXON-SŒUR SEC.
PROCESSUS QUI N’A LIEU QUE DANS MOINS DE 28% DES CAS.
Montagne d’Ambre
30 km
Montagne d’Ambre
30 km
Isolement géographique
Isolement écologique

3. Résultats et Interprétations
81
Pour répondre à ces deux hypothèses de spéciation, nous nous sommes intéressés aux taxons-
sœurs des endémiques locales, qui se trouve être le(s) taxon(s) le plus proche de l’endémique,
idéalement celui à partir duquel la dérive génétique a débuté. Dans un deuxième temps, nous
avons recherché les modes de dispersion de ces espèces, afin d’observer leur potentialité de
dispersion à longue distance.
Les taxons-sœurs
Pour chaque taxon endémique (39 espèces et 7 infra-spécifiques) les taxons les plus proches
ont été recherchés. Pour ce faire, les descriptions originales, la Flore de Madagascar et des
Comores (HUMBERT & AL., 1936) et les révisions récentes ont été consultées pour chaque
taxon. La liste des taxons-sœurs est donnée en Annexe 8.2, p. 191.
La figure 26 montre la répartition au sein des domaines phytogéographiques de HUMBERT
(1965) des taxons-sœurs des 46 endémiques. La majorité des taxons endémiques est issue
d’un « taxon-sœur » de la flore humide. En effet, pour 33 taxons contre 13, le(s) taxon(s)-
sœur(s) possède(nt) une distribution phytogéographique restreinte aux domaines humides de
l’île (Centre ; Est ; Sambirano). Le reste des espèces endémiques a pour taxon-sœur des
espèces à plus large répartition, qui croissent également dans les domaines secs. Il est
important de remarquer qu’aucun taxon-sœur n’a été retrouvé comme étant exclusif des
domaines secs (Ouest/Sud). Nous pouvons donc dire que l’hypothèse vraisemblablement la
plus fréquente, concernant la spéciation des endémiques locales, est liée à un phénomène
d’isolement géographique puis d’une différenciation issue d’un taxon-sœur de la flore
humide.
FIGURE 26 : RÉPARTITION DES TAXONS-SŒURS DES 46 ENDÉMIQUES DU MASSIF D’AMBRE SELON LES 4
DOMAINES PHYTOGÉOGRAPHIQUES DE HUMBERT (1955).
LA FLÈCHE BLEUE MARQUE LES 33 TAXONS-SŒURS PROVENANT DE LA FLORE HUMIDE, ET LA FLÈCHE ROUGE
LES 13 TAXONS-SŒURS DE LA FLORE HUMIDE OU SÈCHE.
3. Résultats et Interprétations
82
L’arrivée sur le site des taxons-sœurs
Les taxons-sœurs qui possèdent une distribution large aussi bien dans les domaines humides
que dans le domaine sec de l’Ouest étaient vraisemblablement présents sur le site et n’ont pas
eu de difficulté à coloniser la Montagne d'Ambre lors de sa surrection. En effet, le piedmont
est composé d’une végétation sèche, dans laquelle ces taxons ont la capacité de s’adapter et de
coloniser de proche en proche le massif. Par contre, pour les taxons-sœurs restreints
actuellement aux domaines phytogéographiques humides, deux hypothèses concernant leur
arrivée à la Montagne d’Ambre peuvent être envisagées : soit par dispersion à longue distance
en franchissant la barrière de la flore sèche du piedmont qui isole actuellement les altitudes
supérieures humides du massif, soit par isolation d'une population arrivée de proche en proche
lors d'épisodes plus humides du passé climatique. Pour décider laquelle des deux hypothèses
présentées ci-dessus est la plus vraisemblable, il faudrait mettre en place une étude
phylogéographique fine sur les populations qui dépasse de loin le cadre de notre travail. Nous
nous sommes contentés d'évaluer le potentiel de dispersion à longue distance (LDD : Long
Distance Dispersal) des 46 taxons endémiques en se basant sur le type de diaspore des espèces
tel que nous le connaissons par les échantillons d'herbier, notre expérience de terrain et la
littérature.
Nous avons obtenus que 48,5% des endémiques locales qui possèdent un taxon-sœur
provenant de la flore humide, présentent des syndromes de dispersion compatibles avec une
LDD. Le tableau 3 récapitule les différentes étapes de cette analyse des endémiques.
TABLEAU 3 : ANALYSE DES TAXONS LES PLUS PROCHES DES ENDÉMIQUES LOCALES.
Taxons-sœurs des endémiques locales
Distribution phytogéographique
malgache
Présence à la Montagne d’Ambre
Non Oui
Moyens pour arriver dans le massif
(étude des espèces endémiques) 48,5% avec potentiel de
dispersion par longue distance 71,7% domaines humides (C-E-Sb)
23 10 51,5% sans potentiel de dispersion par longue distance
28,2% domaines humides et secs (W et C-E-Sb)
5 8 Capacité d’adaptation au piedmont sec

4. Discussion
83
4. DISCUSSION
4.1. PROSPECTION BOTANIQUE ET RICHESSE FLORISTIQUE DU MASSIF D’AMBRE
La Montagne d’Ambre est une aire protégée depuis 1958, et c’est seulement depuis les années
1990 que le nombre d’espèces connues pour ce massif commence à augmenter
significativement. Les prospections botaniques ont jusqu’à présent ciblé les altitudes
supérieures du massif, notamment au-dessus de 800 m d’altitude. Des récoltes en-dessous de
cette limite ont également été effectuées mais restent toutefois anecdotiques.
Cette présente étude a permis d’augmenter de plus de 21% le nombre d’espèces recensées
pour ce massif. Il reste toutefois encore des espèces indéterminées et d’autres qui sont
potentiellement nouvelles pour la Science. Des prospections botaniques sont encore
nécessaires afin de poursuivre l’effort entamé et de continuer à faire augmenter la
connaissance de la diversité floristique pour la Montagne d’Ambre.
Les 13,8% du total des espèces recensées du massif d’Ambre figurent dans la liste de Rouge
de l’IUCN, et 64 espèces sont dites menacées. Ces chiffres sont sous-estimés, car
actuellement 17% de la flore malgache a été analysée au regard des critères de l’IUCN.
Une comparaison entre la richesse spécifique de la Montagne d’Ambre et de celle du
Manongarivo a permis de mettre en évidence quelques particularités de la Montagne
d’Ambre : la richesse spécifique des ptéridophytes est importante, et les espèces et familles
caractéristiques des pentes occidentales du domaine du Centre n’ont pas été retrouvées au
massif d’Ambre. Ce qui signifie que d’après nos résultats, l’étage des pentes occidentales de
HUMBERT (1965) n’est pas présent au massif de la Montagne d’Ambre.
4.2. EFFETS DU VOLCANISME ET RECOLONISATION DE LA VÉGÉTATION
Un événement essentiel qui a fortement conditionné le processus d’installation de cette flore
des altitudes supérieures dans le massif d’Ambre est l’activité volcanique qui a durée jusqu’au
Quaternaire. D’une manière générale, lors d’activités volcaniques deux actions différentes ont
des effets sur la végétation ce sont : les cendres et les coulées de laves (SCHNELL, 1971).
Au niveau de la recolonisation, SCHNELL (1971) fait remarquer qu’il n’y a pas un ordre bien
défini pour la colonisation d’une surface, les lichens, bryophytes ou plantes vasculaires
apparaissent plus ou moins spontanément, dès le stade pionnier. LEGRIS (1969) cité dans
SCHNELL (1971) signale, qu’à la Grande Comore les arbres suivant recolonisent les laves :

4. Discussion
84
Nuxia pseudodentata, Weinmannia, Olea, etc. Nous avons rencontré ces genres dans les forêts
du massif d’Ambre.
A la Montagne d’Ambre, nous avons retrouvé sur la végétation des traces d’anciennes coulées
de laves, qui ont certainement eu lieu durant les dernières périodes d'activité volcanique du
Quaternaire (cf. photo 2, chapitre 1.2.1.2, p. 14). Ces coulées de laves ont modifié le sol dans
la partie ouest du massif, à plus de 800 m d’altitude. La végétation s’en trouve également
modifiée, et à cet endroit, malgré l’altitude, une flore sèche décidue typique des altitudes
inférieures à 800 m a pu s’y installer. En plus, dans ce milieu, nous y avons récolté une
nouvelle espèce déjà publiée, appartenant à la famille des PANDANACEAE
(CALLMANDER & AL. 2009). SCHNELL (1971) précise que « des roches volcaniques plus
ou moins anciennes peuvent constituer des milieux spécialisés permettant un maintien localisé
d’éléments floristiques ailleurs non représentés dans un territoire et peuvent devenir des
stations refuges d’un grand intérêt biogéographique. » C’est ce que nous avons pu observer
sur une partie du versant occidental du massif de la Montagne d’Ambre.
4.3. COLONISATION DES ALTITUDES SUPÉRIEURES PAR LA FLORE HUMIDE
Après le volcanisme, la végétation s’est reconstruite. Deux hypothèses liées implicitement à
deux différentes classifications de la flore humide trouvées dans la littérature ont été
formulées concernant la colonisation des altitudes supérieures (cf. chapitre 1.3.1, p. 24). Ces
deux classifications sont les suivantes :
‐ d’une part HUMBERT (1965) propose de rattacher la flore des altitudes supérieures au
domaine du Centre, sous-domaine du Centre-Nord,
‐ d’autre part CORNET & GUILLAUMET (1976) rattachent les altitudes supérieures à
un secteur d’altitude du domaine de l’Ouest.
Les deux hypothèses sont les suivantes (carte 10, page suivante) :
‐ soit une colonisation par longue distance (flèches bleues),
‐ soit une colonisation de proche en proche (flèche orange).
Notre première remarque est que ces deux hypothèses ne sont pas exclusives l’une par rapport
à l’autre, et que les deux phénomènes se produisent en parallèle, pour constituer la flore dans
son ensemble.

4. Discussion
85
CARTE 10 : REPRÉSENTATION SCHÉMATIQUE DES DEUX HYPOTHÈSES CONCERNANT LA COLONISATION EN
TAXONS HUMIDES DES ALTITUDES SUPÉRIEURES DU MASSIF D’AMBRE.
CARTE DU NORD DE L’ÎLE VUE DE PROFIL ET EN RELIEF, MONTRANT LES 4 DOMAINES PHYTOGÉOGRAPHIES
IMPORTANTS DU NORD. LES FLÈCHE BLEUES REPRÉSENTENT L’HYPOTHÈSE DE COLONISATION PAR LONGUE
DISTANCE. LA FLÈCHE ORANGE REPRÉSENTE L’HYPOTHÈSE D’UNE ADAPTATION PROGRESSIVE DES TAXONS DE
LA FLORE SÈCHE DU PIEDMONT. (DESSIN DE C. CHATELAIN).
Nos résultats ont permis d’observer que selon l’altitude, la proportion des espèces colonisant
selon l’un ou l’autre mode était différente. Dans l’état actuel de nos recherches, et parce que
nos connaissances sont encore insuffisantes au niveau des altitudes intermédiaires ou zones de
transition entre la flore sèche du piedmont et la flore humides des altitudes supérieures, nous
présentons une synthèse des résultats obtenus pour les altitudes supérieures à 800 m (qui est la
limite inférieure selon HUMBERT (1965) du domaine du Centre) en parallèle avec ce qui se
passe au niveau des altitudes intermédiaires aux environ de 800 m.
Altitudes supérieures à 800 m
‐ Nous avons observé une dominance des espèces présentes dans le domaine du Centre,
qu'elles soient exclusives à ce domaine (espèces de hautes altitudes) ou qu'elles aient
une distribution plus large au sein de l'ensemble des domaines humides (Centre, Est,
Sambirano). Les espèces généralistes (présentes dans les domaines humides et secs à
la fois) occupent une place secondaire et deviennent de moins en moins importantes
avec l’altitude qui augmente. En parallèle, les espèces exclusives au domaine du
Centre augmentent. Un travail antérieur (CHRISTE, 2009) traitant de la répartition au
sein du domaine du Centre, d’espèces croissant exclusivement à des altitudes
supérieures à 800 m, a montré que sur l'échantillon analysé, seul 10% des espèces ont
une répartition qui inclut la Montagne d'Ambre. Ces quelques espèces sont pour
Montagne d’Ambre
30 km

4. Discussion
86
moitié « rencontrées sur l’ensemble du domaine du Centre, du nord au sud, et
présentent donc vraisemblablement de grandes facultés de dispersion » (CHRISTE,
2009). Ces résultats confortent l'hypothèse que la flore des altitudes supérieures est
constituée en priorité d'espèces arrivées par dispersion à longue distance.
‐ Nous avons remarqué l’importante richesse spécifique des ptéridophytes en
comparaison avec le massif du Manongarivo. Pour ce groupe particulier, qui est
répartit essentiellement dans les domaines phytogéographiques humides de l’île, la
facilité de dispersion à longue distance a certainement favorisé l’arrivée de ses
représentants à la Montagne d’Ambre et explique cette richesse.
‐ De plus, nous avons pu relever que les espèces caractéristiques de l’étage des pentes
occidentales du domaine du Centre (HUMBERT, 1965) sont absentes de la Montagne
d’Ambre. Ce type de végétation semble donc absent dans le massif isolé de la
Montagne d’Ambre. Nous pouvons y voir plusieurs explications. Une des possibilités
est que la niche climatique où cette végétation se développe dans le reste de l'île,
associée à un phénomène de foehn à des altitudes supérieures à 800 m, soit absente de
la Montagne d'Ambre, en raison de sa relativement faible altitude et de son étendue
limitée. Une autre possibilité est que la niche existe, mais que les espèces qui la
remplissent ne sont pas parvenues à la Montagne d'Ambre. A ce titre, l'absence de la
famille des SARCOLAENACEAE, abondante dans ce type de végétation mérite d'être
soulignée. De fait, le versant occidental de la Montagne d'Ambre présente des stations
moins humides que le versant oriental (RAMANDIMBIMANANA, 2009), mais abrite
une forêt transitionnelle sans rapport avec celle des pentes occidentales.
‐ En bordure des lacs d’anciens cratères, la flore comporte une plus grande proportion
d'espèces qui ne sont pas endémiques de Madagascar. Dans ces zones particulières du
massif, les oiseaux migrateurs qui rencontrent les premiers milieux d'eau douce après
leur traversée s'y arrêtent et déposent vraisemblablement des diaspores d’espèces
provenant d’autres continents. De par sa situation géographique et ses ressources
hydrauliques importantes, la Montagne d’Ambre possède un fort potentiel attracteur
pour ce type de processus. Un des oiseaux recensés qui migre régulièrement dans le
massif d’Ambre est le Apus affinis J.E. Gray qui « a besoin de plans d'eau permanents
pour pouvoir boire » (OLIOSO, 2010 en ligne) (chapitre 1.2.2.2, p. 21).

4. Discussion
87
En conclusion, au niveau des altitudes supérieures à 800 m, on retrouve le domaine du
Centre sous-domaine du Centre Nord qui domine, ainsi que des espèces humides comme
la théorie de HUMBERT (1965) le propose. De plus, les espèces du domaine du Centre
sont très certainement des généralistes d’altitude, possédant une forte capacité de
dispersion. C’est donc certainement une colonisation par longue distance qui explique le
peuplement des altitudes supérieures à 800 m.
Altitudes intermédiaires (<800 m)
‐ Contrairement aux altitudes supérieures où la composition floristique reflétait
clairement la dominance des espèces du Centre, nous avons observé aux altitudes
intermédiaires un mélange d'espèces appartenant à toutes sortes de catégories
phytogéographiques.
‐ Les espèces dites généralistes, présentes à la fois dans les domaines secs et les
domaines humides de l’île, sont très largement dominantes. Les espèces sèches sont en
augmentation, alors que l’altitude décroît. Cependant ces dernières restent en faible
proportion.
‐ Nous avons pu noter la présence d’espèces avec une distribution restreinte au domaine
du Sambirano ou de celui de l’Est. Toutefois, la proportion des espèces de ces cortèges
floristiques restent suffisamment faibles et n'ont pas d'influence sur une cartographie
phytogéographique comme celle de HUMBERT (1955), à plus forte raison à l’échelle
utilisée. Toutefois, une ceinture de végétation rattachée au domaine de l’Est est visible
au niveau des altitudes intermédiaires de la Montagne d'Ambre sur la feuille "Baie
d’Ampasindava" de la carte de la végétation de HUMBERT & COURS DARNE
(1965).
‐ A ces altitudes, nous sommes face à une zone de transition entre la flore sèche du
piedmont et la flore humide des altitudes supérieures, particulièrement riche du point
de vue de la diversité. Les relevés linéaires effectués en parallèle à ce travail ont
permis de révéler cette richesse, vers 1 000 m d’altitude et en dessous
(RAMANDIMBIMANANA, 2009). Les forêts y sont peuplées de « taxa provenant
des deux types de végétation [humides et sèches] » (RAZANAJATOVO, 2009).
‐ Les données sont encore lacunaires au niveau des ces altitudes intermédiaires. Des
prospections botaniques ciblant ces altitudes clefs sont nécessaires.

4. Discussion
88
En conclusion pour les altitudes intermédiaires, aucun des quatre domaines
phytogéographiques n’est dominant. Il est à relever toutefois l’importance et l’augmentation
des espèces généralistes, ainsi que l’apparition d’espèces sèches restreintes au domaine sec de
l’Ouest. D’après les résultats des relevés linéaires effectués en parallèle à cette étude, ces
zones sont les plus riches en espèces. A ces altitudes intermédiaires, la colonisation de proche
en proche doit certainement augmenter de par la proximité avec le domaine sec de l’Ouest,
mais le phénomène de colonisation dominant reste toutefois celui de la longue distance.
4.4. ENDÉMIQUES LOCALES ET PHÉNOMÈNES DE SPÉCIATION
Historiquement, le nombre d’endémiques locales pour le massif d’Ambre a toujours été
surestimé, de par la position isolée de sa flore humide. Nous avons pu l’observer sur un
échantillonnage d’espèces présentant le suffixe « ambrens* » et décrit à l’origine comme
endémique de la Montagne d’Ambre. Plus du tiers de ces espèces ne sont plus des
endémiques locales. Cependant, le chiffre actuel des endémiques (1 genre monospécifique, 39
espèces et 7 taxons infraspécifique) reste comparable aux chiffres d’endémiques d’autres
massifs forestiers du Nord de Madagascar comme par exemple le Daraina ou le Manongarivo.
Nous avons aussi remarqué que les endémiques locales sont principalement réparties entre
900 et 1 100 m d’altitude à la Montagne d’Ambre.
Concernant leurs modes de spéciation, d’après notre analyse sur les taxons-sœurs, ils sont
distribués à plus de 70% dans des domaines humides de l’île (Centre, Est, Sambirano). La
spéciation se déroule donc majoritairement à partir d’un taxon issu de la flore humide. Il
semble donc qu’il y ait une influence entre ce que nous observons pour la colonisation des
altitudes supérieures du massif par l’ensemble de la flore (dispersion par longue distance) et
pour la spéciation des endémiques locales.

5. Contraintes et analyses de la méthodologie
89
5. CONTRAINTES ET ANALYSES DE LA MÉTHODOLOGIE Au cours des travaux de terrain, des contraintes logistiques, notamment dues au déblocage de
fonds tardifs, ont retardé l’exécution des trois dernières missions de terrain. L’effet a été
ressenti sur les récoltes fertiles qui sont en nombre relativement faible pour le dernier
campement qui a eu lieu en juin avec trois mois de retard sur le planning initial. A cette
période les fruits sont déjà passés, et certains arbres commencent déjà à perdre leurs feuilles.
Il a donc été très difficile de trouver des végétaux encore fertiles.
Sur le quatrième campement, nous avons eu un problème d’incendie du séchoir. Les plantes
sont séchées par air chaud avec du gaz qui fonctionnent jour et nuit, afin d’éviter qu’elles se
ré-humidifient, étant donné l’humidité atmosphérique trop importante. Une nuit, au bout de 9
jours de mission, un incendie s’est déclenché, et le feu s’est propagé aux récoltes entreposées
non loin de là, ainsi qu’au matériel nécessaire pour le séchage (journaux et cartons). Nous
avons été contraints de sortir plus rapidement du camp, et une septième mission a du être
réalisée dans cette même zone pour récupérer les données manquantes. Heureusement, à
cause de cette humidité atmosphérique, le feu s’est rapidement arrêté, et n’a pas pu se
propager dans la forêt. La méthode de séchage par air chaud reste malgré tout un très bon
moyen de séchage des plantes, comparé à la méthode de séchage par alcool qui est aussi
utilisée dans ces régions tropicales. Par air chaud, les couleurs des plantes ressortent mieux, et
des analyses sur l’ADN sont encore possibles sur les feuilles d’un échantillon. Ainsi, des
feuilles d’URTICACEAE ont été prélevées à l’herbier de Genève par un spécialiste de cette
famille, pour en effectuer des analyses moléculaires.
Après les missions de terrain, le travail à fournir pour la gestion des spécimens d’herbiers
entrepris à l’herbier de Genève a pris un temps considérable. En effet, il a fallut : saisir les
données récoltées sur le terrain, gérer la base de données dans Microsoft Access, imprimer les
étiquettes d’herbiers et les déterminavits, puis distribuer les parts pour les différents herbiers
internationaux. Des formulaires ont été spécialement créés dans Access pour optimiser la
saisie des données. Concernant les déterminations des échantillons, pour certaines familles
non parues dans la Flore de Madagascar et des Comores (HUMBERT & AL., 1936) et qui
n’ont pas fait l’objet de révision récente, il a été difficile d’arriver à des déterminations par
nous-mêmes. Les spécialistes travaillant sur les représentants malgaches de ces groupes ont
été consultés et ont été d’une grande aide. Il reste toutefois encore des échantillons qui ont été

5. Contraintes et analyses de la méthodologie
90
envoyés à des spécialistes, dont nous n’avons pas encore reçu de déterminations, ou pour
lesquels les déterminations ont été reçues trop tardivement pour être inclues dans ce travail.
Lorsque les déterminations ont été considérées comme terminées, afin d’obtenir une analyse
de la distribution géographique de toutes les espèces recensées du massif d’Ambre, un long
travail de recherche bibliographique a été effectué. Au niveau de la méthodologie, il aurait été
préférable d’effectuer l’étape de saisie de la phytogéographie des espèces en même temps que
la phase de détermination, pour ne pas avoir à rechercher l’espèce deux fois dans un même
ouvrage. En effet, une importante source d’information consultée pour l’analyse de la
distribution des espèces et pour les déterminations est la Flore de Madagascar et des Comores
(HUMBERT & AL., 1936).

6. Conclusion
91
6. CONCLUSION La Montagne d’Ambre possède des caractéristiques aussi bien environnementales (isolement
géographique, orographie, ressources hydrauliques, climat) que biologiques uniques
(zoologique, botanique).
L’histoire de ce massif est liée à son ancienne activité volcanique qui lui a donné sa forme
actuelle et un sol fertile. Des lacs formés par des puits d’anciens cratères, ainsi que de
nombreux lacs temporaires, permettent à la Montagne d’Ambre d’offrir des environnements
différents et ouverts qui permettent à diverses espèces transportées par des oiseaux migrateurs
de s’y installer.
Une analyse des prospections botaniques effectuées à la Montagne d’Ambre permet de relever
que nous sommes la première équipe de botanistes à avoir effectué un inventaire floristique
sur une longue durée. Nos spécimens d’herbiers représentent plus de 34% de tout ce qui y a
été récolté depuis le premier collecteur recensé en 1880, J. M. Hildebrandt. Durant nos
travaux de terrain, 21% des échantillons que nous avons récoltés représentaient des espèces
encore non citées du massif de la Montagne d’Ambre.
Au niveau de la richesse spécifique des familles, HUMBERT (1959) a écrit que les
ORCHIDACEAE sont la plus grande famille de la Flore Malgache. Il en est de même dans
nos résultats avec une très grande proportion d’ORCHIDACEAE et de RUBIACEAE dans la
flore du massif d’Ambre.
La Montagne d’Ambre possède des caractéristiques floristiques qui n’ont pas été retrouvées
dans d’autres massifs forestiers du Nord de l’île, notamment la richesse spécifique des
ptéridophytes qui est beaucoup plus importante qu’au massif du Manongarivo. L’isolement de
la flore humide des altitudes supérieures de la Montagne d’Ambre aurait favorisé les espèces
capables de dispersion à longue distance comme c’est le cas pour les ptéridophytes. Sur les
versants occidentaux, une humidité moins marquée que sur les versants orientaux a été
remarquée au niveau des altitudes supérieures à 800 m. Toutefois, les espèces caractéristiques
de l’étage des pentes occidentales décrit par HUMBERT (1965), qui remplissent ces niches
écologiques dans la chaîne de montagne principale du domaine du Centre n’ont pas été
retrouvées à la Montagne d’Ambre.
Pour ce qui concerne les affinités mondiales de cette flore, la région malgache et l’Afrique qui
sont géographiquement les plus proches de Madagascar ont une influence importante sur la

6. Conclusion
92
flore du massif d’Ambre. Selon les zones du massif ou les groupes considérés, les affinités
mondiales varient. Au niveau des zones où la canopée est plus ouverte, comme sur les crêtes
ou à proximité des lacs d’anciens cratères, des espèces qui sont présentes à Madagascar
uniquement à la Montagne d’Ambre mais existant ailleurs dans le monde ont été retrouvées.
L’analyse de la répartition des espèces sur l’île de Madagascar, étudiée par leur présence dans
les domaines phytogéographiques de HUMBERT (1965) a montré d’une manière générale
une très forte affinité avec le domaine du Centre. Nous avons également retrouvé à la
Montagne d’Ambre, des espèces connues jusqu’alors uniquement du domaine de l’Est ou du
Sambirano, ce que la carte des domaines phytogéographiques de HUMBERT (1965) ne
permet pas d’observer. Au niveau du piedmont, nous retrouvons un mélange entre le domaine
de l’Ouest, du Sambirano, de l’Est et du Centre. Pour les altitudes supérieures, par contre,
nous pouvons dire que les espèces du domaine du Centre dominent largement en-dessus de
800 m d’altitude, conformément aux travaux de HUMBERT (1965). Aucune espèce restreinte
au domaine du Sud ou au domaine des Hautes Montagnes n’a été trouvée.
Deux hypothèses ont été formulées concernant la colonisation de la Montagne d'Ambre
depuis sa surrection: dispersion à longue distance ou dispersion de proche en proche depuis le
domaine de l’Ouest du piedmont. Au niveau des altitudes supérieures du massif c’est
certainement une colonisation par longue distance qui domine. En revanche, au niveau des
altitudes intermédiaires, la colonisation de proche en proche paraît plus probable.
Nous avons recensé 46 taxons et 1 genre monospécifique endémiques de la Montagne
d’Ambre. Toutefois ce chiffre est en perpétuelle évolution, en fonction du développement de
nos connaissances de la flore malgache. Plus il y a d’études au massif d’Ambre, plus ce
chiffre augmente (découverte de nouveaux taxons), et parallèlement, plus il y a d’études dans
d’autres massif du Nord de l’île, plus ce chiffre diminue (découverte ailleurs de taxons qu'on
pensait endémiques du massif). Il semble toutefois que globalement ce chiffre se maintienne à
une valeur élevée en comparaison avec d’autres massifs forestiers de la région Nord de l’île.
Des hypothèses relatives au mode de spéciations des endémiques locales ont également été
formulées. Il semble que l’hypothèse la plus probable soit une différenciation qui s’est
déroulée à partir d’un taxon issu de la flore humide, ce qui suppose un isolement
géographique puis une dérive génétique. Des études au niveau génétique, qui dépassent le
cadre de ce travail, devraient permettre d’identifier avec précision pour chaque taxon
endémique, le mode de spéciation qui s’est effectivement produit. D’après nos connaissances

6. Conclusion
93
du mode de transport de leurs diaspores nous pouvons déduire que la moitié des espèces
endémiques font partie de groupes ayant un potentiel de dispersion à longue distance.
Le nombre d’espèces endémiques de Madagascar reste important à la Montagne d’Ambre
mais se situe en-dessous du chiffre de PHILLIPSON (2006) concernant toute la Flore de
Madagascar. Ce phénomène s’observe sur toute région de la Grande Ile. Le poids apporté par
chaque massif forestier et son nombre d’endémiques locales propres est additionné lorsque
nous considérons l’ensemble de la flore de Madagascar. C’est ce qui explique le taux énorme
de 90% d’endémisme de la flore malgache d’une manière générale.
En ce qui concerne les familles endémiques de Madagascar, on en retrouve deux à la
Montagne d’Ambre : les SPHAEROCEPALACEAE et les PHYSENACEAE sur les cinq
familles endémiques malgaches qui ont récemment été recensées par
RAMANANJANAHARY & AL. (2010 in prep.).
Enfin, dans notre liste floristique, il reste encore des espèces indéterminées et pour certaines
d’entre elles, les spécialistes consultés ont pu dire qu’elles étaient nouvelles pour la Science.
Il en ressort que cette Montagne recèle encore beaucoup de diversité et qu’il est nécessaire de
continuer son étude notamment au niveau des zones de transition entre la flore humide des
altitudes supérieures et la flore sèche du piedmont où les prospections botaniques restent
insuffisantes. Les pressions anthropiques grandissantes qui pèsent sur cette forêt et
spécialement sur ces zones de transition rendent urgente et impérative la poursuite des travaux
d'inventaire floristique pour mieux connaître, protéger et valoriser la biodiversité de la
Montagne d’Ambre.

7. Bibliographie
94
7. BIBLIOGRAPHIE
ANGIOSPERM PHYLOGENY GROUP (1998). An ordinal classification of the families of
flowering plants. Ann. Missouri Bot. Gard. 85 : 531-553.
AUBREVILLE, A. (1963). Classification des formes biologiques des plantes vasculaires en
milieu tropical. In Adansonia. Muséum National d’Histoire Naturelle de Paris. Nouvelle Série
Tome III, Fascicule 2: 221-226.
BARAT, C. (1958). La Montagne d'Ambre (Nord de Madagascar). Revue de géographie
alpine 46: 629-681.
BETTINELLI, S. (2007). Recherche sur les récoltes botaniques de la Montagne d’Ambre,
Nord de Madagascar. Monographie du Baccalauréat (Bachelor). Université de
Genève/Laboratoire de botanique systématique et biodiversité. Conservatoire et Jardin
botaniques de la Ville de Genève. 14pp.
BRENON, P. (1972). The Geology of Madagascar. in Biogeography and Ecology in
Madagascar. R. Battistini and G. Richard-Vindard (eds.). The Hague, Dr. W. Junk B. V.,
Publishers: 27-86.
BETSCH, J.-M. (2003). Collembola, Springtails. in The Natural History of Madagascar. S.
M. Goodman and J. P. Benstead (eds.). Chicago, The University of Chicago Press.
CALLMANDER, M. C., GAUTIER, L., TRIGUI, S. M. (2009). Pandanus nusbaumeri
(Pandanaceae), a new endemic species from the Montagne d’Ambre (northern Madagascar).
Candollea 64(2): 213-218.
CAMPBELL, N. A. & REECE, J. B. (2004). Biologie. Adaptation et révision scientifique de
Richard Mathieu 2e édition. Editions De Boeck Université, Bruxelles. 1364pp.
CARLETON, M. D. (2003). Eliurus, Tufted-tailed Rats. in The Natural History of
Madagascar. S. M. Goodman and J. P. Benstead (eds.). Chicago, The University of Chicago
Press: 1373-1380.
CHAINEY, J. E. (2003). Tabanidae, Horse Flies. in The Natural History of Madagascar. S.
M. Goodman and J. P. Benstead (eds.). Chicago, The University of Chicago Press: 721-727.

7. Bibliographie
95
CHRISTE, C. (2009). Analyse de la distribution d’espèces d’altitude de Madagascar, dans la
perspective du peuplement forestier de la Montagne d’Ambre. Monographie du Baccalauréat
(Bachelor). Université de Genève/Laboratoire de botanique systématique et biodiversité.
Conservatoire et Jardin botaniques de la Ville de Genève. 31pp.
COLLIN, D. (2010 en ligne). Oiseaux.net, fiche descriptive du Faucon d’Eléonore.
Disponible sur Internet : <http://www.oiseaux.net/oiseaux/faucon.d.eleonore.html> (consulté
le 14.07.2010).
CORNET, A. (1974). Essai cartographique bioclimatique à Madagascar, carte à 1/2'000'000 et
notice explicative N° 55. Paris, ORSTOM.
CORNET, A. & GUILLAUMET, J.-L. (1976). Divisions floristiques et étages de végétation à
Madagascar. Cah. ORSTOM, sér. Biol., vol. XI, n°1 :35-40.
DU PUY, D.J & MOAT, J.F. (1996). A refined classification of the primary vegetation of
Madagascar based on the underlying geology: using GIS to map its distribution and to assess
its conservation status. in W.R. Lourenço (editor). Proceedings of the International
Symposium on the Biogeography of Madagascar. Editions de l’ORSTOM, Paris: 205-218, +
3 maps.
ELOUARD, J.-M. & F.-M. GIBON (2003). Ecology of Aquatic Invertebrates. in The Natural
History of Madagascar. S. M. Goodman and J. P. Benstead (eds.). Chicago, The University of
Chicago Press: 511-517.
ENGHOFF, H. (2003). Diplopoda, Millipedes. in The Natural History of Madagascar. S. M.
Goodman and J. P. Benstead (eds.). Chicago, The University of Chicago Press: 617-627.
FARAMALALA, M. H., M. VINCELETTE, & AL. (1998). Formations végétales et domaine
forestier national de Madagascar. Antananarivo, CI, DEF, CNRE, FTM.
GAUTIER, L. (2002). Chapitre 6. Liste commentée des phanérogames de la Réserve Spéciale
de Manongarivo, Madagascar. In Inventaire floristique et faunistique de la Réserve Spéciale
de Manongarivo (NW Madagascar). Boissiera 59, Conservatoire et Jardin botaniques de
Genève : 105-239.

7. Bibliographie
96
GAUTIER, L. & S. M. GOODMANN (2002). Chapitre 2. Description de la Réserve Spéciale
de Manongarivo, Madagascar. In Inventaire floristique et faunistique de la Réserve Spéciale
de Manongarivo (NW Madagascar). Boissiera 59, Conservatoire et Jardin botaniques de
Genève : 21-40.
GLAW, F. & VENCES, M. (1994). A Fieldguide to the Amphibians and Reptiles of
Madagascar. Second edition including mammals and freshwater fish. Zoologisches
Forschungsinstitut und Museum Alexander Koenig, Bonn. Germany. 480pp.
GOODMAN, S. M. & J. P. BENSTEAD (2003). The Natural History of Madagascar.
Chicago, The University of Chicago Press.
GUILLAUMET, J.-L. & J. KOECHLIN (1971). Contribution à la définition des types de
végétation dans les régions tropicales (exemple de Madagascar). Candollea 26/2: 263-277.
HOLMGREN, P. K., N. H. HOLMGREN, & L. C. BARNETT (1990). Index herbariorum.
Part 1: The Herbaria of the World. 8th ed. New York, New York botanical Garden.
HUMBERT, H. & AL. (1936 -). Flore de Madagascar et des Comores. Muséum National
d’Histoire Naturelle, Paris.
HUMBERT, H. (1955). Les territoires phytogéographiques de Madagascar. In Colloques
internationaux du Centre National de la Recherche Scientifique, LIX : les divisions
écologiques du monde, moyen d’expression, nomenclature, cartographie. Paris, 1954. Année
Biologique, série 3-31 : 439-48.
HUMBERT, H. (1959). Origines de la flore de Madagascar. In Mém. Inst. Sc. Mad. Série B.
Biologie Végétale. Tome 9:149-187.
HUMBERT, H., (1965). Description des types de végétation. In Notice de la carte
Madagascar. Carte internationale du tapis végétal et des conditions écologiques. Humbert, H.
& G. Cours Darne Toulouse, France, CNRS et O.R.S.T.O.M : 46-78.
HUMBERT, H. & G. COURS DARNE (1965). Carte internationale du tapis végétal et des
conditions écologiques. N°10 Baie d’Ampasindava. Toulouse, France, CNRS et
O.R.S.T.O.M.

7. Bibliographie
97
UICN (2001). Catégories et Critères de l’UICN pour la Liste Rouge : Version 3.1.
Commission de la sauvegarde des espèces de l’UICN. UICN, Gland, Suisse et Cambridge,
Royaume-Uni. ii + 32 pp.
JENKINS, P. D. (2003). Microgale, Shrew Tenrecs. in The Natural History of Madagascar. S.
M. Goodman and J. P. Benstead (eds.). Chicago, The University of Chicago Press: 1273-
1278.
JURY, M. R. (2003). The Climate of Madagascar. In The Natural History of Madagascar. S.
M. Goodman and J. P. Benstead (eds.). Chicago, The University of Chicago Press: 75-87.
KRÜGER, M. (2007). Composition and origin of the Lepidoptera faunas of southern Africa,
Madagascar and Réunion (Insecta: Lepidoptera). Ann. Transvaal Mus. 44 : 123–178.
LANGRAND, O. (1995). Check List of the Birds of Amber Mountain National Park,
Madagascar. Anatananarivo.
LEES, D. C., C. KREMEN, ET AL. (2003). Classification, Diversity, and Endemism of the
Butterflies (Papilionoidea and Hesperioidea): A Revised Species Checklist. in The Natural
History of Madagascar. S. M. Goodman and J. P. Benstead (eds.). Chicago, The University of
Chicago Press: 762-793.
LEGRIS, P. (1969). La Grande Comore. Climats et végétation. Travaux de la Section
Scientifique et Technique. Institut Français de Pondichéry, 3(5) : 1-28.
OLIOSO, G. (2010 en ligne). Oiseaux.net, fiche descriptive du martinet des maisons.
Disponible sur Internet : < http://www.oiseaux.net/oiseaux/martinet.des.maisons.html>
(consulté le 14.07.2010).
MITTERMEIER, R, A. & AL. (2007). Lemurs of Madagascar Pocket Identification Guide.
Conservation International (Tropical pocket Guide Series).
MYERS, N. & AL. (2000). Biodiversity hotspots for conservation priorities. in Nature. N°
403: 853-858.

7. Bibliographie
98
PEARCE, T. A. (2003). Gasteropoda, Terrestrial Snails. in The Natural History of
Madagascar. S. M. Goodman and J. P. Benstead (eds.). Chicago, The Universtiy of Chicago
Press: 529-574.
PERRIER DE LA BÂTHIE, H. (1921). La végétation malgache. Annales du Musée colonial
de Marseille, série 3 9: 1-268.
PERRIER DE LA BÂTHIE, H. (1936). Biogéographie des plantes de Madagascar. Paris,
Société d'éditions géographiques, maritimes et coloniales.
PHILLIPSON, P. B. & AL. (2006). A catalogue of the vascular plants of Madagascar. in
Taxonomy and Ecology of African Plants, their Conservation and sustainable Use.
Proceedings of the 17th AETFAT Congress Addis Ababa, Ethiopia. Ghazanfar, S. A. and
Beentje, H., (eds.). Royal Botanic Gardens, Kew : 613–627.
RAKOTONDRAINIBE, F. (2003). Diversity, Ecology, and Distribution of the Pteridophyte
Flora. in The Natural History of Madagascar. S. M. Goodman and J. P. Benstead (eds.).
Chicago, The University OF Chigaco Press: 282-313.
RAKOTONDRASANA, S. R. (1997). Approche à l'analyse structurale de la forêt d'Ambre,
Réserve Spéciale n°5: Typologie - Cartographie - Inventaire forestier. Mémoire de DEA.
Faculté des Sciences Antananarivo. 89p.
RAMANDIMBIMANANA, S. D. (2009). Analyse floristique de quelques de quelques
groupements végétaux des forêts de la Montagne d’Ambre (Nord de Madagascar, Région
DIANA). Mémoire de DEA. Faculté des Sciences Antananarivo. 112p.
RAMANANJANAHARY, R. H., FRASIER C. L., LOWRY II, P. P., RAJAONARY, F. A. &
G. E. SCHATZ (2010). Madagascar's Endemic Plant Families Species Guide / Guide des
espèces de Plantes appartenant aux Familles Endémiques de Madagascar.
RASOLONANDRASANA, B. P. N. (1994). Contribution à l'étude de l'alimentation de
Cryptoprocta ferox Bennett (1833) dans son milieu naturel. Mémoire de DEA. Service de
Paléontologie. Faculté des Sciences Antananarivo.

7. Bibliographie
99
RAUNKIAER, C. (1934). Type biologique pour la géographie botanique. Kun. Gdansk
Vidensk. Selsk.Copenhague. 1905, p. 347-438.
RAVET, J. (1952). Notice sur la climatologie de Madagascar et des Comores. Mémoires de
l’institut scientifique de Madagascar. Série D. Tome IV. Fascicule 1: 1-36.
RAXWORTHY, C. J. (2003). Introduction to the Reptiles. in The Natural History of
Madagascar. S. M. Goodman and J. P. Benstead (eds.). Chicago: 934-949.
RAXWORTHY, C. J. & R. A. NUSSBAUM (1994). A rainforest survey of amphibians,
reptiles and small mammals at Montagne d'Ambre, Madagascar. Biological Conservation 69:
65-73.
RAZANAJATOVO, M. H. (2009). Caractérisation écologique et analyse structurale des
forêts humides de la Montagne d’Ambre (Région DIANA). Mémoire de DEA. Faculté des
Sciences Antananarivo. 123p.
ROSSI, G. & M. ROSSI (1998). La Montagne d'Ambre (extrême Nord de Madagascar)
espace à protéger? Cahiers d'analyse spatiale quantitative et appliquée. N° 38-39, Université
de Nice.
SCHNELL, R. (1970). Introduction à la Phytogéographie des pays tropicaux. Les problèmes
généraux (vol. I : Les flores. Les structures). Gauthier-Villars Editeur. Paris : 1-503.
SCHNELL, R. (1971). Introduction à la Phytogéographie des pays tropicaux. Les problèmes
généraux (vol. II : Les milieux. Les groupements végétaux). Gauthier-Villars Editeur. Paris :
504-951.
SPICHIGER, R.-E., SAVOLAINEN, V. V., FIGEAT, M. & D. JEANMONOD (2004). Une
approche phylogénétique nouvelle des Angiospermes des régions tempérées et tropicales.
Troisième édition revue et corrigée. Presses polytechniques et universitaires romandes,
Suisse. 413pp.
STERLING, E. (2003). Daubentonia madagascariensis, Aye-Aye, Aye-Aye. in The Natural
History of Madagascar. S. M. Goodman and J. P. Benstead (eds.). Chicago, The University of
Chicago Press: 1348-1351.

7. Bibliographie
100
STEVENS, P. F. (2001, en ligne). Angiosperm Phylogeny Website. Version 9, Juin 2008.
Disponible sur Internet : <http://www.mobot.org/mobot/research/apweb/>.
TRIGUI, S. M. (2007). Etude bibliographique d’un massif forestier du Nord de Madagascar:
La Montagne d’Ambre. Monographie du Baccalauréat (Bachelor). Université de
Genève/Laboratoire de botanique systématique et biodiversité. Conservatoire et Jardin
botaniques de la Ville de Genève. 24pp.
VENCE, M. & AL. (2009). Madagascar as a model region of species diversification. In
Trends in Ecology and Evolution. Vol.24 No.8 : 456-465.
Liste des sites Internet consultés Index herbariorum : <http://sweetgum.nybg.org/ih/>
JSTOR : <http://plants.jstor.org/>
Sonnerat :<http://coldb.mnhn.fr/colweb/form.do?model=SONNERAT.wwwsonnerat.wwwso
nnerat.wwwsonnerat>
Tropicos : <http://www.tropicos.org/Home.aspx>.

8. Annexes
101
8. ANNEXES
8.1. LISTE FLORISTIQUE DE LA MONTAGNE D’AMBRE
8.1.1. PHANÉROGAMES
GYMNOSPERMES
PODOCARPACEAE Podocarpus L'Hér. ex Pers.
Podocarpus madagascariensis Baker M [E, C] LG 5013 (1320 m)
ANGIOSPERMES
ACANTHACEAE Acanthopale C.B.Clarke
Acanthopale sp. OA 136 [800 m] (nv)
Asystasia Blume Asystasia sp. 1 MHR 119 (1070 m)
Barleria L. Barleria vincaefolia Baker M [Sb, C, W(N)] MHR 151 (1040 m); OA 93 [950 m]; OA 987 (809 m) (nv)
Brachystephanus Nees Brachystephanus africanus S. Moore M [C] - Aft Brachystephanus africanus S. Moore var. madagascariensis Figueiredo HH 32074 (1000 m); HH 3976 (1100 m), TYPE: HT:G; IT: TAN; LB 11994 [1150 m]; SMT 126 (1080 m); SMT 395 (990 m)
Brachystephanus lyallii Nees M [C, HM] HH 32157 (1340 m); LG 5041 (1240 m); LG 5210 (1230 m); SDR 007 (1230 m); SMT 410 (1020 m)
Brillantaisia P.Beauv. Brillantaisia madagascariensis T. Anderson ex Lindau M [Sb, E, C] - Aft AMH s.n. [1100 m] (nv); Gentry, A. 11948 (1200 m) (nv); JB 20359 (nv); JB 5944 (nv); LN 2384 (1250 m); MHR 103 (980 m)
Crossandra Salisb. Crossandra multidentata Vollesen M [W(N)] OA 88 [950 m] (nv); SM 1229 (610 m) (nv)
Dicliptera Juss. Dicliptera sp. JB 5906 (nv)
Dyschoriste Nees Dyschoriste gracilicaulis (Benoist) Benoist M [MdA] HP 17702 (500 m) (nv), TYPE: HT:P, IT:P

8. Annexes
102
Hypoestes Sol. ex R.Br. Hypoestes brachiata Baker M [C] LG 5142 (1250 m); OA 135 [800 m]; SM 977 (1300 m)
Hypoestes erythrostachya Benoist M [Sb, C, W(N)] JB 5626 (nv); JB 5937 (nv)
Hypoestes loniceroides Baker M [Sb] OA 152 [1175 m] (nv); OA 217 [900 m]
Hypoestes macilenta Benoist M [C] SMT 468 (985 m)
Hypoestes tubiflora Benoist M [C] Du Puy, D. & al. 215 (1120 m) (nv); OA 137 [1175 m]
Hypoestes verticillaris R. Br. M [C] - Aft MHR 149 (1030 m); SMT 442 (650 m)
Hypoestes verticillaris var. modesta Benoist JB 5983 (nv)
Hypoestes sp. Cremers, G. 2545 (1000 m) (nv); Cremers, G. 2546 (1000 m) (nv); Du Puy, D. & al. MB215 (1300 m) (nv); JB 5946 (nv); JB 5979 (nv); OA
345 (1475 m) (nv); Rakotozafy, A. 1481 (nv)
Isoglossa Oerst. Isoglossa melleri Baker M [C] MHR 044 (1050 m); SM 1276 (710 m); SMT 032 (1010 m); SMT 281 (1130 m)
Justicia L. Justicia delicatula Scott-Elliot M [E, C] Cremers, G. 2569 (1000 m) (nv); MHR 097 [1040 m]; MHR 118 (1070 m); SMT 116 (1050 m); SMT 405 (1050 m)
Justicia haplostachya (Nees) T. Anderson M [Sb, E, C, W(S)] - RM LG 5144 (1250 m); SM 955 (1300 m); SMT 041 (1270 m)
Justicia rhodoptera Baker M [C] JB 5649 (nv)
Justicia tenella (Nees) T. Anderson M [C, HM, W(N)] - Aft, As SMT 399 (990 m)
Justicia sp. 1 SMT 494 (865 m)
Justicia sp. GES 1509 (1000 m) (nv); GES 2393 (1000 m) (nv)
Mendoncia Vell. ex Vand. Mendoncia cowanii (S.Moore) Benoist M [E, C] OA 213 [900 m]; OA 221 [375 m]; OA 364 [375 m]; OA 777 (874 m); SDR 047 (1080 m); SMT 220 (1000 m); SMT 306 (1065 m)
Mendoncia flagellaris (Baker) Benoist M [Sb, E, C, W(N)] - RM OA 123 [800 m]; OA 259 [375 m]; OA 280 [375 m]; OA 309 (1475 m); OA 74 [950 m]; SMT 246 (1000 m); SMT 315 (975 m)
Mendoncia sp. AMH 86 [1100 m] (nv)
Mimulopsis Schweinf. Mimulopsis sp. 1 LN 2497 (1161 m); SMT 124

8. Annexes
103
Ruellia L. Ruellia cyanea (Nees) T. Anderson M [Sb, E, C, W] LG 5050 (900 m); LN 2391 (1250 m); MHR 116 (1060 m); SMT 180 (1140 m)
Ruellia sp. 1 SMT 346 (1035 m)
Ruellia sp. 2 SMT 184 (1065 m); SMT 261 (1040 m)
Ruellia sp. 3 SM 961 (1300 m)
aff. Ruellia sp. SMT 371 (1010 m)
ACANTHACEAE indéterminées Miller, J. S. & al. 10678 (1050 m) (nv)
AGAVACEAE Chlorophytum Ker Gawl.
Chlorophytum chloranthum Baker M [E, C] GES 2395 (1000 m) (nv)
AMARANTHACEAE Achyranthes L.
Achyranthes aspera L. M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt SMT 432 (885 m)
Achyranthes mauritiana Moq. M [Sb, C] - RM LG 5187 (1350 m)
Alternanthera Forssk. Alternanthera sessilis (L.) DC. M [Sb, E, C, W] - RM, Aft, As, Oc, Amt, Eur SMT 397 (990 m)
Cyathula Blume Cyathula achyranthoides (Kunth) Moq. M [E, C, W(N)] - Aft, Amt HP 17714 (nv); JB 5645 (nv)
Cyathula prostrata (L.) Blume M [Sb, E, C, W] - RM, Aft, As, Oc, Amt JB 5954 (1300 m) (nv); MHR 107 (970 m); SMT 359 (970 m)
Cyathula triuncinella Schinz M [E] SDR 105 (1030 m)
Lagrezia Moq. Lagrezia ambrensis Cavaco M [MdA] HH 3979 (1100 m) (nv), TYPE: HT:P; PM 1186 (nv)
AMARYLLIDACEAE
Crinum L. Crinum firmifolium Baker M [Sb, E, C, W, S] SDR 117 (975 m)

8. Annexes
104
ANACARDIACEAE Poupartia Comm. ex Juss.
Poupartia silvatica H.Perrier M [W] SF & R. Capuron 20073 (nv). Echantillon rattaché avec doute : SM 1237 (620 m) (nv)
Rhus L. Rhus perrieri (Courchet) H. Perrier M [W, S] Miller, J. S. & al. 3625 (1050 m) (nv)
Sclerocarya Hochst. Sclerocarya birrea (A.Rich.) Hochst. M [W] - Aft Sclerocarya birrea (A.Rich.) Hochst. subsp. caffra (Sond.) Kokwaro AMH 9 [1050 m] (nv); Harder, D. K. & al. 1621 (660 m) (nv)
Sorindeia Thouars Sorindeia madagascariensis Thouars ex DC. M [Sb, E, W] - RM, Aft GES 1538 (1000 m) (nv); HP 18820 (800 m) (nv); OA 873 [900 m] (nv); Van Nek, F. I. 1866 (715 m) (nv)
ANNONACEAE Ambavia Le Thomas
Ambavia capuronii (Cavaco & Keraudren) Le Thomas M [E, C, W(N)] SF & R. Capuron 11304 (600 m) (nv); SF & R. Capuron 20045 (nv); SMT 519 (715 m)
Ambavia sp. 1 LG 5083 (750 m)
Artabotrys R.Br. Artabotrys mabifolius Diels M [E, C, W] Harder, D. K. & al. 1646 (840 m) (nv)
Artabotrys scytophyllus (Diels) Cavaco & Keraudren M [Sb, C, W(S)] Rakotonandrasana, S. & al. 912 (290 m) (nv)
Artabotrys sp. OA 859 (533 m) (nv); OA 985 (533 m) (nv)
Isolona Engl. Isolona sp. OA 70 [950 m] (nv)
Monanthotaxis Baill. Monanthotaxis ambrensis (Cavaco & Keraudren) Verdc. M [W(N)] HP 18826 (1000 m) (nv), TYPE: IT: P; SMT 544 (1000 m)
Monanthotaxis cf. valida (Diels) Verdc. Harder, D. K. & al. 1650 (840 m) (nv)
Polyalthia Blume Polyalthia oligosperma (Danguy) Diels M [C, W(N)] GMP 14513 (1000 m) (nv); SDR 112 (970 m); SF & R. Capuron 11278 (nv); SM 1812 [1045 m]
Polyalthia sp. 1 SDR 040 (1041 m)
Xylopia L. Xylopia buxifolia Baill. M [Sb, E, C, W(S)] SF & N. Boanarisolo 8126 [1050 m] (nv); SF & R. Capuron 11511 (nv); SF & R. Rabevohitra 29394 (nv)
Xylopia perrieri Diels M [E, C] Rakotondrafara, A. & al. 280 (717 m) (nv)

8. Annexes
105
Xylopia sp. Misch 63 (nv); SF & R. Capuron 20041 (nv); SF & R. Capuron 20051 (nv)
APIACEAE Agrocharis Hochst.
Agrocharis melanantha Hochst. M [C, HM] - Aft LG 5150 (1350 m); PM 1204 (nv); PM 1391 (nv)
Centella L. Centella asiatica (L.) Urb. M [E, C, W] - RM, Aft, As, Amt HP 6832 (nv); LG 5164 (1350 m)
Hydrocotyle L. Hydrocotyle cf. mannii Hook.f. LN 2459 (1248 m)
Hydrocotyle ranunculoides L.f. M [C] - Aft, Amt HP 6891 (nv)
Hydrocotyle sp. 1 LG 5168 (1350 m)
Sanicula L. Sanicula elata D.Don M [Sb, E, C, W] - RM, Aft, As, Amt Leeuwenberg, A. J. M. & al. 14284 (1000 m) (nv); LN 2413 (1230 m); SM 1726 [1045 m] (nv); SMT 378 (1035 m)
APOCYNACEAE Asclepias L.
Asclepias curassavica L. M [E, C] - As, Amt SM 1724 [1045 m] (nv)
Calyptranthera Klack. Calyptranthera gautieri Klack. M [Sb, W(N)] SMT 048 (985 m)
Camptocarpus Decne. Camptocarpus acuminatus (Choux) Venter M [C] GMP 14494 [1150 m] (nv); Miller, J. S. & al. 3615 (1050 m) (nv); OA 228 [375 m] (nv); SMT 090 (1030 m); SMT 187 (1080 m); SMT 307
(1065 m); SMT 446 (1010 m)
Camptocarpus mauritianus (Lam.) Decne. M [C, W(S)] - RM SMT 066 (1070 m)
Carissa L. Carissa boivinianum (Baill.) Leeuwenb. M [Sb, E, C, W] HP 18823 (800 m) (nv), TYPE: HT; OA 946 (662 m) (nv); SF 17 (nv); SM 1233 (610 m)
Carissa spinarum L. M [Sb, E, C, W] - RM, Aft, As, Oc GMP 14502 [1150 m] (nv); HP 18856 (800 m) (nv); LB 11950 [1050 m] (nv); LB 11980 [1150 m]; Leeuwenberg, A. J. M. & al. 14243
(1000 m) (nv); Leeuwenberg, A. J. M. & al. 14312 (1000 m) (nv); LG 5044 (1180 m); Schedl, K. E. 195 (1150 m) (nv); SDR 013 (975 m);
SDR 018 (1250 m); SF 211 [1050 m] (nv); SF 36 (nv); SF 62 (nv)
Ceropegia L. Ceropegia saxatilis Jum. & H.Perrier M [W] SM 1741 [1045 m] (nv); SMT 061 (1075 m)
Ceropegia sp. 1 LN 2462 (1300 m)

8. Annexes
106
Ceropegia sp. OA 82 [950 m] (nv)
Cynanchum L. Cynanchum bosseri Liede M [E] SM 1796 [1045 m] (nv)
Cynanchum membranaceum (Liede & Meve) Liede & Meve M [W(S)] HP 18862 (800 m) (nv)
Cynanchum messeri (Buchenau) Jum. & H.Perrier M [E, W, S] LN 2466 (1120 m)
Cynanchum obovatum (Decne.) Choux M [C] HP 11658 (800 m) (nv); HP 17540 (1200 m) (nv); PM 1185 (nv); SM 2369 [1438 m] (nv)
Gomphocarpus R.Br. Gomphocarpus fruticosus (L.) W. T. Aiton M [Sb, E, C, W] - RM, Aft SM 1718 [1045 m] (nv)
Landolphia P.Beauv. Landolphia myrtifolia (Poir.) Markgr. M [Sb, E, C, W] - RM HP 18866 (800 m) (nv); Leeuwenberg, A. J. M. & al. 14295 (900 m) (nv)
Mascarenhasia A.DC. Mascarenhasia arborescens A.DC. M [Sb, E, C] - RM, Aft Harder, D. K. & al. 1603 (450 m) (nv); JB 5923 (800 m) (nv); Leeuwenberg, A. J. M. & al. 14315 (1000 m) (nv); SF & R. Capuron 20044
(nv), TYPE: HT: P; IT: K, MO, P, TEF, WAG; SM 1266 (590 m); SMT 050 (985 m); SMT 457 (1000 m)
Mascarenhasia angustifolia A.DC. var. keraudreniana Markgr. SF & Andrianandianina 30725 (nv)
Oncinotis Benth. Oncinotis tomentella Radlk. M [Sb, E, C, W] HP 17566 (1000 m) (nv); Leeuwenberg, A. J. M. & al. 14235 (1000 m) (nv)
Pentopetia Decne. Pentopetia androsaemifolia Decne. M [Sb, C, W, S] - RM Randrianaivo, R. & al. 796 (400 m) (nv)
Petchia Livera Petchia erythrocarpa (Vatke) Leeuwenb. M [Sb, E, C, W] - RM AMH 161 (nv); AMH 174 (nv); AMH 45 (nv); Harder, D. K. & al. 1588 (990 m) (nv); HH 32094 (1000 m) (nv); Leeuwenberg, A. J. M. &
al. 14241 (970 m) (nv); Leeuwenberg, A. J. M. & al. 14287 (1000 m) (nv); Leeuwenberg, A. J. M. & al. 14287 (1000 m) (nv); Leeuwenberg,
A. J. M. & al. 14758 (400 m) (nv); MHR 141 (975 m); OA 886 (874 m); SF & R. Capuron 22056 [900 m]; SF & R. Capuron 24056 [900 m]
(nv); SF & R. Rabevohitra 32606 (nv); SMT 015 (1119 m); SMT 075 (1150 m); SMT 465 (1020 m)
Rauvolfia L. Rauvolfia obtusiflora A.DC. M [Sb, E, C, W] Keraudren, M. 1726 (nv)
Roupellina (Baill.) Pichon Roupellina boivinii (Baill.) Pichon M [W, S] PM 1426 (nv)
Secamone R.Br. Secamone marsupiata Klack. M [MdA] Labat, J.-N. 2870 (900 m) (nv); Leeuwenberg, A. J. M. & al. 14293 (900 m) (nv), TYPE: HT: WAG; IT: MO; LG 5057 (870 m); Van Nek,
F. I. 1777 (nv); Van Nek, F. I. 1864 (785 m) (nv)

8. Annexes
107
Secamone pinnata Choux M [E] Secamone pinnata Choux subsp. septentrionalis Klack. HP 18824 (800 m) (nv), TYPE; HP 18840 (nv); LG 5234 (1100 m)
Secamone varia Klack. M [C] LB 12026; LG 5009 (1310 m); SMT 024 (1280 m)
Secamone aff. varia Klack. SMT 251 (970 m); SMT 355 (975 m)
Secamone sp. Labat, J.-N. & al. 2870 (900 m) (nv)
Stephanostegia Baill. Stephanostegia hildebrandtii Baill. M [Sb, W] SF & R. Capuron 20079 (nv)
Tabernaemontana L. Tabernaemontana coffeoides Bojer ex A.DC. M [Sb, E, C, W, S] - RM AMH 129 (nv)
Tabernaemontana eusepala Aug.DC. M [E] AMH 74 [1050 m] (nv); SF & R. Rabevohitra 32600 [1050 m] (nv)
Tabernaemontana phymata Leeuwenb. M [E, C] Allorge, L. & al. 2723 (nv); Allorge, L. 2754 (nv); AMH 138 (nv); Debray, M. M. 1529 [950 m] (nv); GES 1520 (1000 m) (nv); GES 2391
(1000 m) (nv); GMP 14488 [1050 m] (nv); GMP 14507 [1150 m] (nv); Harder, D. K. & al. 1584 (935 m) (nv); HH 32044 [950 m] (nv); HH
3971 (1100 m); HP 18845 (900 m) (nv); HP 8933 (1000 m) (nv); Keraudren, M. & al. 25572 (nv); Keraudren, M. 25572 (nv); Labat, J.-N. &
al. 2866 (900 m); LB 11950 (1000 m); Leeuwenberg, A. J. M. & al. 14233 (1000 m) (nv); Leeuwenberg, A. J. M. & al. 14233 (1000 m) (nv);
Leeuwenberg, A. J. M. & al. 14239 (1000 m) (nv); Leeuwenberg, A. J. M. & al. 14239 (1000 m) (nv); LG 5027 (900 m); MHR 065 (1050
m); OA 362 [375 m]; OA 871 [900 m]; Rakotozafy, A. 2305 (nv); Razafimandimbison, S. G. 120 [1150 m] (nv); Schedl, K. E. 171 (1100 m)
(nv); SF & J. de D. Rakoto 11017 [1050 m] (nv); SF & J. de D. Rakoto 11036 [1050 m] (nv); SF & R. Capuron 29183 (1000 m) (nv), TYPE:
HT: P; IT: P; SF 11017 [1050 m] (nv); SF 11036 [1050 m] (nv); SF 13222 (1000 m) (nv); SM 1181 (670 m); SM 989 (700 m); SMT 264
(1050 m); SMT 407 (1040 m); Van Nek, F. I. 1814 (950 m) (nv); Van Nek, F. I. 1859 (775 m) (nv); Van Nek, F. I. 1861 (780 m) (nv)
Tabernaemontana sp. Miller, J. S. & al. 10674 (1050 m) (nv); OA 787 (874 m) (nv)
Tylophora R.Br. Tylophora sylvatica Decne. M [Sb, E, C, W] - Aft JB 20322 [850 m] (nv)
APONOGETONACEAE Aponogeton L.f.
Aponogeton boivinianus Jum. M [Sb, E, W(N)] - RM AMH 190 (nv); HP 17669 (500 m) (nv); HP 7145 (nv); JB 20326 [850 m] (nv); PM 1385 (nv); PM 1401 [850 m] (nv); SF 24467bis (nv);
SMT 532 (650 m)
AQUIFOLIACEAE Ilex L.
Ilex mitis (L.) Radlk. M [C] - Aft, Amt GMP 14493 [1150 m] (nv); GMP 14516 (1000 m) (nv); Harder, D. K. & al. 1574 (890 m) (nv); LG 5040 (1260 m); OA 115 [800 m] (nv);
OA 776 (874 m) (nv); Phillipson, P. B. 1963 (900 m) (nv); SM 1758 [1045 m] (nv); SM 1797 [1045 m] (nv); SM 2361 [1438 m] (nv); SM
939 (700 m) (nv); SMT 224 (1090 m); SMT 285 (1065 m)
Ilex mitis Radlk. var. mitis Radlk. GES 2386 (1000 m) (nv)

8. Annexes
108
ARACEAE Pothos L.
Pothos scandens L. M [Sb, E, C, W] - RM, As OA 1003 (507 m) (nv)
ARALIACEAE Polyscias J.R.Forst. & G.Forst.
Polyscias aculeata (Decne. & Planch.) Harms M [E] HIRSM 176 (1100 m) (nv)
Polyscias amplifolia (Baker) Harms M [C] Du Puy, D. & al. 216 (1300 m) (nv); Du Puy, D. & al. MB216 (1300 m) (nv); GES 2401 (1000 m) (nv); LB 11953 (1000 m) (nv); Phillipson,
P. B. 1957 (900 m) (nv)
Polyscias boivinii (Seem.) Bernardi M [Sb, W] SF & J. Belin 265-R-6 (450 m) (nv)
Polyscias briquetiana (Bernardi) Lowry & G.Plunkett M [C] LN 2388 (1260 m); MHR 073 (1100 m); SM 959 (1300 m) (nv); SMT 276 (1100 m)
Polyscias duplicata (Thouars ex Baill.) Lowry & G.Plunkett M [E, C] Phillipson, P. B. 1972 (900 m) (nv); SDR 005 (1090 m); SM 1769 [1045 m] (nv)
Polyscias ornifolia (Baker) Harms M [C] HH 32160 (1400 m) (nv); LB 11995 (1100 m) (nv); LG 5011 (1310 m); LG 5103 (1430 m); OA 310 (1475 m) (nv); OA 32 [1438 m] (nv);
SDR 008 (1300 m); SM 974 (1470 m) (nv)
Polyscias cf. ornifolia (Baker) Harms SMT 164 (1080 m); SMT 328 (955 m)
Polyscias sp. 1 SDR 087 (970 m)
Polyscias sp. Labat, J.-N. & al. 2864 (1100 m) (nv); Schedl, K. E. 154 (1100 m) (nv)
Schefflera J.R.Forst. & G.Forst. Schefflera longipedicellata (Lecomte) Bernardi M [C] Harder, D. K. & al. 1582 (935 m) (nv); Miller, J. S. & al. 3634 (1050 m) (nv); Phillipson, P. B. 1973 (900 m) (nv); SDR 080 (1110 m); SMT
321 (970 m); SMT 478 (1000 m)
Schefflera monophylla (Baker) Bernardi M [C] LG 5106 (1470 m); SM 968 (1470 m)
Schefflera myriantha (Baker) Drake M [C, HM] - Aft GES 2398 (1000 m) (nv); OA 147 [1175 m] (nv); SDR 041 (1200 m); SMT 230 (1005 m)
Schefflera sp. OA 35 [1438 m] (nv)
ARECACEAE Dypsis Noronha ex Mart.
Dypsis baronii (Becc.) Beentje & J.Dransf. M [C] HP 18870 (1000 m) (nv), TYPE: HT; JB 20367 [1350 m] (nv); SM 976 (1470 m) (nv)
Dypsis sp. 1 LG 5125 (1470 m)
Dypsis sp. 2 SDR 064 (1090 m)

8. Annexes
109
Dypsis sp. 3 SMT 368 (1020 m)
Dypsis sp. Du Puy, D. & al. MB217 (1300 m) (nv); Du Puy, D. & al. MB222 (1300 m) (nv)
ASPARAGACEAE Asparagus L.
Asparagus simulans Baker M [E, C, W(S)] Bernier, J. 341 (nv); LG 5092 (1030 m); MHR 080 (1100 m); MHR 105 (990 m); OA 59 [950 m] (nv); SM 1260 (630 m)
Dracaena Vand. ex L. Dracaena angustifolia Roxb. M [E, C] - RM, As, Oc SDR 102 (1140 m); SMT 222 (1115 m); SMT 479 (1000 m)
Dracaena reflexa Lam. M [Sb, E, C, W, S] - RM, Aft Leeuwenberg, A. J. M. & al. 14761 [400 m] (nv); OA 306 (1475 m) (nv)
Dracaena reflexa Lam. var. angustifolia Baker Harder, D. K. & al. 1591 (965 m) (nv); MHR 009 (1300 m)
Dracaena reflexa Lam. var. lanceolata H.Perrier HP 18878 (1000 m) (nv); Labat, J.-N. & al. 2867 (900 m) (nv); MHR 091
Dracaena reflexa Lam. var. linearifolia Baker Harder, D. K. & al. 1649 (840 m) (nv); MHR 024 (1050 m); SM 1199 (620 m); SMT 419 (670 m)
Dracaena reflexa Lam. var. nervosa H.Perrier MHR 067 (1040 m); MHR 104 (990 m)
Dracaena xiphophylla Baker M [E, C, W(S)] SM 1234 (610 m)
Dracaena sp. OA 33 [1438 m] (nv)
ASPHODELACEAE Aloe L.
Aloe capitata Baker M [C] Aloe capitata Baker var. silvicola H.Perrier SMT 393 (990 m)
Aloe sakoankenke J.-B.Castillon M [W(N)] Castillon, J. B. 10 (nv), TYPE: HT: P; IT: P, MO; Castillon, J. B. 11 (nv)
Lomatophyllum Willd. Lomatophyllum cf. roseum H.Perrier LN 2468 (1120 m); SM 1185 (680 m) (nv)
ASTERACEAE Acmella Rich.
Acmella caulirhiza Delile M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt LN 2448 (1248 m); SMT 396 (990 m)
Adenostemma J.R.Forst. & G.Forst. Adenostemma viscosum J.R.Forst. & G.Forst. M [Sb, E, C, W] - RM, Aft, As, Oc, Amt LN 2415 (1332 m); MHR 051 (1080 m); OA 42 [1438 m] (nv); SMT 433 (885 m)

8. Annexes
110
Bidens L. Bidens pilosa L. M [Sb, E, C, W] - RM, Aft, As, Oc, Amt LN 2455 (1248 m)
Brachylaena R.Br. Brachylaena perrieri (Drake) Humbert M [Sb, C, W] SF & J. Belin 10522 (nv)
Centauropsis Bojer ex DC. Centauropsis aff. decaryi Humbert SMT 160 (1080 m); SMT 324 (955 m)
Centauropsis rhaponticoides (Baker) Drake M [E, C] GES 2406 (1000 m) (nv); GMP 14496 [1150 m] (nv); HP 17772 (nv); HP 2906 (nv); LG 5128 (1450 m)
Chrysanthellum Pers. Chrysanthellum americanum (L.) Vatke M [C, W] - Aft, As, Amt LN 2446 (1248 m)
Conyza Less. Conyza sp. JB 5966 (nv)
Cosmos Cav. Cosmos caudatus Kunth M [Sb, E, C, W] - RM, Aft, As, Oc, Amt SMT 232
Crassocephalum Moench Crassocephalum crepidioides (Benth.) S.Moore M [Sb, E, C, W] - RM, Aft LG 5194 (1350 m); LN 2442 (1248 m)
Crassocephalum luteum (Humbert) Humbert M [C] - Aft HH 3981 (1100 m); LG 5130 (1370 m); SMT 282 (1100 m)
Crassocephalum sarcobasis (DC.) S.Moore M [Sb, E, C, W(S)] - RM, Aft SM 1715 [1045 m] (nv)
Crassocephalum sp. JB 5970 (nv)
Dichrocephala L'Hér. ex DC. Dichrocephala latifolia (Poir.) DC. M [E, C] - Aft, As LG 5196 (1350 m)
Elephantopus L. Elephantopus scaber L. M [Sb, E, C] - RM, Aft, As, Oc, Amt SMT 114
Emilia Cass. Emilia bathiei Humbert M [W(N)] HP 17564 (800 m) (nv), TYPE: HT: P; LG 5155 (1350 m)
Emilia citrina DC. M [Sb, E, C] SMT 137 (1005 m)
Emilia humifusa DC. M [Sb, E, C] MHR 086; SMT 009 (1300 m)
Gerbera L. Gerbera petasitifolia Humbert M [C] LG 5172 (1200 m)

8. Annexes
111
Helichrysum Mill. Helichrysum globosum A.Rich M [C, HM] - RM, Aft HH 3977 (1100 m); HP 18872 (nv); HP 2907 (800 m) (nv), TYPE
Helichrysum triplinerve DC. M [C, W(S)] - RM MHR 085
Helichrysum sp. JB 5900 (nv)
Mikania Willd. Mikania scandens (L.) Willd. M [Sb, E, C, W] - RM, Aft, As, Oc, Amt SMT 505 (910 m)
Mikania sp. JB 5933 (nv); JB 5968 (nv)
Pseudognaphalium Kirp. Pseudognaphalium luteoalbum (L.) Hilliard & B.L.Burtt M [C] - RM, Aft, As, Oc, Amt LG 5162 (1350 m)
Psiadia Jacq. Psiadia altissima (DC.) Drake M [Sb, E, C] SDR 069
Psiadia altissima (DC.) Drake var. altissima HH 3973 [1000 m] (nv)
Psiadia cf. coursii Humbert LG 5127 (1470 m)
Psiadia nigrescens Humbert M [Sb, E, C, W(N)] SM 1795 [1045 m] (nv)
Psiadia sp. JB 5973 (nv); JB 6002 (nv)
Senecio L. Senecio cf. alleizettei (Humbert) Humbert SMT 029 (1300 m)
Siegesbeckia L. Siegesbeckia abyssinica Stapf M [C] - Aft AMH 5 [1050 m] (nv)
Sonchus L. Sonchus oleraceus L. M [C, S] - RM, Aft, As, Oc, Amt LN 2461 (1248 m)
Vernonia Schreb. Vernonia ambrensis Humbert M [MdA] HP 18829 (800 m) (nv), TYPE: HT: P, IST; Poisson, H. 84 (nv)
Vernonia appendiculata Less. M [E, C, W, S] AMH 85 [1100 m] (nv); LN 2483 (1248 m)
Vernonia baronii Baker M [C, HM] OA 312 (1475 m) (nv)
Vernonia cinerea (L.) Less. M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt SMT 513 (750 m)
Vernonia colorata (Willd.) Drake subsp. grandis (DC.) C.Jeffrey Du Puy, D. & al. MB212 (800 m) (nv)

8. Annexes
112
Vernonia grandis (DC.) Bojer ex Vatke M [Sb, W(S)] - RM, Aft Du Puy, D. & al. 212 (1120 m) (nv)
Vernonia lastellei Drake M [E, C] MHR 130 (1080 m); SMT 348 (1045 m)
Vernonia secundifolia DC. M [C] LG 5132 (1410 m); SM 969 (1470 m); SMT 341 (1025 m)
Vernonia sp. SF & Andrianandianina 30731 (nv)
ASTERACEAE indéterminées LB 11949 [1025 m] (nv)
BALANOPHORACEAE Langsdorffia Mart.
Langsdorffia malagasica (Fawc.) B.Hansen M [E, C] SM 2384 [1438 m] (nv)
BALSAMINACEAE Impatiens L.
Impatiens bicaudata H.Perrier M [MdA] De Block, P. 972 (nv); Du Puy, D. & al. MB204 (1325 m) (nv); GES 1516 (1000 m) (nv); HH 3980 (nv); HP 17547 (1200 m) (nv), TYPE:
HT:P, IT:P; HP 5782bis (nv); Leeuwenberg, A. J. M. & al. 14310 (1000 m) (nv); OA 915 (662 m) (nv); SM 1721 [1045 m] (nv); SM 926
(700 m) (nv)
Impatiens bisaccata Warb. M [W(N)] Du Puy, D. & al. 200 (1325 m) (nv); GES 1531 (1000 m) (nv); Hildebrandt, J. M. 3382a (nv); HP 17546 [1100 m] (nv); HP 17752 (1200 m)
(nv); HP 18853 (900 m) (nv); HP 5782 (1000 m) (nv); JB 20345 (nv); Keraudren, M. 1648 [1050 m] (nv); Labat, J.-N. & al. 2873 [850 m]
(nv); LB 11954 (1000 m); Leeuwenberg, A. J. M. & al. 14302 (950 m) (nv); LG 5062 (760 m); SM 1200 (620 m) (nv); SM 1727 [1045 m]
(nv); SM 934 (700 m); SMT 125 (1050 m)
Impatiens elatostemmoides H.Perrier M [C] LN 2472 (1380 m)
Impatiens lyallii Baker M [C, HM] OA 160 [1175 m] (nv)
Impatiens malcomberi Eb.Fisch. & Raheliv. M [MdA] LG 5105 (1430 m); SM 2356 [1438 m] (nv), TYPE: HT: TAN; IT: MO
Impatiens sacculata Warb. M [MdA] Hildebrandt, J. M. 3382 (nv), TYPE
Impatiens sp. 1 LG 5085 (1070 m); MHR 033 (1021 m); SMT 295 (1065 m)
Impatiens sp. 2 LG 5135 (1360 m); LG 5148 (1350 m); SDR 048 (1080 m)
Impatiens sp. 3 LN 2496 (1300 m); SDR 095 (1085 m)
Impatiens sp. 4 SMT 291 (1065 m)
Impatiens sp. 5 MHR 059 (990 m)
Impatiens sp. 6 LG 5181 (1350 m)

8. Annexes
113
Impatiens sp. 7 SDR 097 (1120 m)
Impatiens sp. 8 MHR 120 (1075 m); SMT 171 (1090 m)
Impatiens sp. JB 5981 (nv); Miller, J. S. & al. 10679 (1050 m) (nv); Miller, J. S. & al. 10681 (1050 m) (nv); OA 913 (662 m) (nv); OA 916 (662 m) (nv);
PM 1169 (1000 m) (nv); PM 1395 [1100 m] (nv); Rakotozafy, A. 1490 (nv)
BASELLACEAE Basella L.
Basella madagascariensis Boivin ex H. Perrier M [Sb, W(S)] LB 12010 [900 m]; SMT 498 (840 m)
BEGONIACEAE Begonia L.
Begonia francoisii Guillaumin M [Sb, E, C] SM 2367 [1438 m] (nv)
Begonia humbertii Keraudren M [C] PM 1392 [1150 m] (nv)
Begonia lyallii A.DC. M [Sb, E, C] Begonia lyallii A.DC. var. pubescens Keraudren HH 32156 (1200 m) (nv); LG 5131 (1370 m); SDR 045 (1080 m)
Begonia madecassa Keraudren M [C] HJBT 57 (nv); JB 20333 (800 m) (nv); OA 282 [375 m] (nv)
Begonia sp. SM 1814 [1045 m] (nv)
BIGNONIACEAE Colea Bojer ex Meisn.
Colea sp. Gentry, A. 11944 (1200 m) (nv); SM 1790 [1045 m] (nv)
Jacaranda Juss. Jacaranda sp. OA 195 [900 m] (nv)
Ophiocolea H.Perrier Ophiocolea ambrensis Callm. & Phillipson M [MdA] AMH 193 (nv); Miller, J. S. & al. 3620 (1050 m) (nv), TYPE: IT: TAN; HT: MO; SF & R. Capuron 11280 (nv); SF & R. Capuron 20032
[1050 m] (nv); SF 32001 [1050 m] (nv)
Ophiocolea schatzii A.H. Gentry (syn. Ophiocolea ambrensis Callm. & Phillipson) GES 1524 (1000 m) (nv), TYPE: HT: MO
Ophiocolea floribunda (Bojer ex Lindl.) H.Perrier M [Sb, E, C, W(N)] - RM HIRSM 174 (1100 m) (nv)
Ophiocolea ornithophila A.H.Gentry M [MdA] SM 1197 (630 m)
Ophiocolea velutina H.Perrier M [W(N)] SF & R. Capuron 20050 (nv)
Ophiocolea sp. 1 MHR 006 (1070 m)

8. Annexes
114
Ophiocolea sp. 2 LG 5093 (1030 m)
Phyllarthron DC. Phyllarthron bojeranum A.DC. M [E, C] LG 5022 (1310 m); LG 5119 (1400 m); SM 1753 [1045 m] (nv); SM 981 (700 m); SMT 203 (1175 m); SMT 316 (975 m)
Phyllarthron sp. OA 250 [375 m] (nv)
Stereospermum Cham. Stereospermum cf. euphorioides (Bojer) A.DC. Randrianaivo, R. & al. 792 (400 m) (nv)
BORAGINACEAE Cynoglossum L.
Cynoglossum lanceolatum Forssk. M [Sb, E, C, W(S)] - RM, Aft, As HP 9083 (nv); Leeuwenberg, A. J. M. & al. 14311 (1000 m) (nv); LN 2431 (1250 m); Miller, J. S. & al. 3633 (1050 m)
Ehretia P.Browne Ehretia cymosa Thonn. M [Sb, E, C, W] - RM, Aft SF & R. Capuron 20058 (nv); SMT 105 (1123 m)
Heliotropium L. Heliotropium indicum L. M [E, C, W] - RM, Aft, Amt JB 5901 (nv)
Hilsenbergia Tausch ex Rchb. Hilsenbergia schatziana J.S.Mill. M [E, W(N)] SF & R. Capuron 11276 (1000 m) (nv); SF & R. Capuron 11302 (600 m) (nv), TYPE: HT: P, IT: P, MO; SF 7210 (nv)
Tournefortia L. Tournefortia puberula Baker M [Sb, E, C] SMT 153 (1060 m)
BRASSICACEAE Cardamine L.
Cardamine africana L. M [E, C, HM, W] - RM, Aft, As, Oc, Amt Allorge, L. & al. 2721 (nv); HH 32066 (1000 m) (nv); LN 2359 (1300 m)
Cardamine trichocarpa Hochst. ex A.Rich. M [MdA] - Aft, As HP 17710 (1200 m) (nv)
BURMANNIACEAE Gymnosiphon Blume
Gymnosiphon danguyanus H.Perrier M [Sb, E, C, W(S)] Allorge, L. & al. 2720 (nv)
BURSERACEAE Canarium L.
Canarium madagascariense Engl. M [Sb, E, C, W] OA 94 [950 m] (nv); Rakotondrafara, A. & al. 303 (400 m) (nv); SF & R. Rabevohitra 29393 (nv); SF 7191 (nv); SMT 101 (1170 m); SMT
223 (1100 m); SMT 527 (650 m)

8. Annexes
115
Canarium madagascariense Engl. subsp. obtusifolium (Scott-Elliot) Leenh. Harder, D. K. & al. 1586 (990 m) (nv); SF & R. Capuron 684 [1050 m] (nv)
Canarium sp. SF 7209 (nv)
CACTACEAE Rhipsalis Gaertn.
Rhipsalis baccifera (J.S. Muell.) Stearn M [Sb, E, C, W] - RM, Aft, As, Amt AMH 46 [1100 m] (nv); LG 5242 (1100 m); SM 1206 (670 m) (nv); SMT 014 (1050 m); SMT 320 (970 m)
CALOPHYLLACEAE Mammea L.
Mammea glaucifolia (H. Perrier) Kosterm. M [C] Ursch, E. 143 (1200 m) (nv), TYPE: HT: P, IT: P
Mammea punctata (H. Perrier) P.F. Stevens M [Sb, E, W(S)] Harder, D. K. & al. 1604 (450 m) (nv); SM 1253 (630 m) (nv)
CANELLACEAE Cinnamosma Baill.
Cinnamosma sp. OA 861 (533 m) (nv); SF & R. Capuron 20068 (nv)
CANNABACEAE Aphananthe Planch.
Aphananthe sakalava J.-F.Leroy M [Sb, E, C, W] HIRSM 227 (600 m) (nv); SF & R. Capuron 20046 (nv); SF & R. Capuron 23290 (598 m) (nv); SF & R. Rabevohitra 29994 (nv)
Celtis L. Celtis gomphophylla Baker M [Sb, E, C, W(S)] - RM, Aft SF & J. Belin 5845 (nv); SF 5845 (nv)
Celtis aff. gomphophylla Baker SM 1257 (640 m) (nv)
Celtis mildbraedii Engl. M [Sb, W] - RM, Aft SF & R. Capuron 20052 (nv), TYPE: HT: P, IT: P; SF & R. Capuron s.n. (600 m) (nv)
Celtis sp. LB 12007 (nv); SF & N. Boanarisolo 12871 [1050 m] (nv)
Trema Lour. Trema orientalis (L.) Blume M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt OA 261 [375 m] (nv); SMT 079 (1078 m)
CAPPARACEAE Crateva L.
Crateva excelsa Bojer M [Sb, E, C, W] - RM Miller, J. S. & al. 3631 (1050 m) (nv); Rakotozafy, A. 2290 (nv)
Thylachium DC. Thylachium sp. JB 5910 (nv); OA 948 (662 m) (nv)

8. Annexes
116
CARDIOPTERIDACEAE Leptaulus Benth.
Leptaulus citroides Baill. M [Sb, E, C, W] Harder, D. K. & al. 1634 (840 m) (nv); LG 5051 (900 m); SM 1246 (650 m) (nv)
Leptaulus cf. citroides Baill. SMT 414 (985 m)
CARYOPHYLLACEAE Cerastium L.
Cerastium cf. glomeratum Thuill. LG 5157 (1350 m)
Drymaria Willd. ex Schult. Drymaria cordata (L.) Willd. ex Roem. & Schult. M [E, C, W(S)] - RM, Aft, As, Oc, Amt LG 5192 (1350 m)
Stellaria L. Stellaria mannii Hook.f. M [E, C] - RM, Aft LG 5226 (1350 m); SMT 418 (920 m)
Stellaria media (L.) Vill. M [C] - RM, Aft, As, Oc, Amt, Eur LG 5195 (1350 m)
CELASTRACEAE Brexia Noronha ex Thouars
cf. Brexia sp. OA 72 [950 m] (nv)
Brexiella H.Perrier Brexiella acutifolia H. Perrier M [E] SF & R. Capuron 11296 (900 m) (nv)
Brexiella ilicifolia H. Perrier M [C] SMT 530 (650 m)
Brexiella sp. SF & R. Capuron 20057 (nv); SF & R. Capuron 20060 (nv)
Elaeodendron Jacq. Elaeodendron sp. SF 5468 [1050 m] (nv)
Euonymus L. Euonymus elaeodendroides Loes. M [E, C, W] Euonymus elaeodendroides Loes. subsp. elaeodendroides Capuron, R. 20054 (nv), TYPE
Evonymopsis H.Perrier Evonymopsis cf. Humbertii H.Perrier MHR 166 (980 m); SMT 537 (650 m)
Maytenus Molina Maytenus fasciculata (Tul.) Loes. M [Sb, E, W(N)] - RM, Aft, Amt SDR 032 (1162 m)
Maytenus undata (Thunb.) Blakelock M [Sb, E, W(N)] - RM, Aft, Amt SMT 520 (710 m)

8. Annexes
117
Mystroxylon Eckl. & Zeyh. Mystroxylon aethiopicum (Thunb.) Loes. M [Sb, E, C, W, S] - RM, Aft GES 1523 (1000 m) (nv); GMP 14514 (1000 m) (nv); Harder, D. K. & al. 1587 (990 m) (nv); LN 2385 (1260 m); Miller, J. S. & al. 3616
(1050 m) (nv); Phillipson, P. B. 1971 (900 m) (nv); PM 1199 (1000 m) (nv); SDR 004 (1060 m); SDR 011 (1300 m); SDR 067 (1100 m); SF
& N. Boanarisolo 11104 (nv); SF & R. Rabevohitra 29387 (nv); SF 30733 (nv); SM 1265 (590 m) (nv)
Polycardia Juss. Polycardia lateralis O. Hoffm. M [Sb, E, C, W] AMH 423 (nv); SF 1 (nv)
Salacia L. Salacia madagascariensis (Lam.) DC. M [Sb, E, C, W] SMT 091 (995 m)
Salacia sp. Miller, J. S. & al. 3622 (1050 m) (nv)
CHRYSOBALANACEAE Grangeria Comm. ex Juss.
Grangeria porosa Boivin ex Baill. M [Sb, W] SF & R. Capuron 11344 [25 m] (nv)
Hirtella L. Hirtella zanzibarica Oliv. M [Sb, E, W(N)] - Aft OA 898 (533 m) (nv); Randrianaivo, R. & al. 788 (399 m) (nv)
CLUSIACEAE Garcinia L.
Garcinia ambrensis (H. Perrier) P. Sweeney & Z.S. Rogers M [C] HP 17730 (1200 m) (nv), TYPE: IT: P; LG 5086 (1030 m); OA 1000 (507 m) (nv); SDR 141 (980 m)
Garcinia calcicola (Jum. & H. Perrier) P. Sweeney & Z.S. Rogers M [C, W] GES 1517 (1000 m) (nv)
Garcinia aff. calcicola (Jum. & H. Perrier) P. Sweeney & Z.S. Rogers SM 1196 (620 m)
Garcinia decipiens (Baill.) Vesque M [Sb, E, C, W(N)] SF & J. de D. Rakoto 11022 (nv)
Garcinia disepala Vesque M [Sb] OA 204 [900 m] (nv)
Garcinia orthoclada Baker M [E, C] HP 17548 (1000 m) (nv), TYPE: IT: TAN; HP 18892 (nv)
Garcinia aff. orthoclada Baker GES 1498 (1000 m) (nv); OA 119 [800 m] (nv); OA 146 [1175 m] (nv); OA 331 (1475 m) (nv); SM 937 (700 m) (nv)
Garcinia aff. verrucosa Jum. & H.Perrier OA 91 [950 m] (nv)
Garcinia sp. 1 SMT 425 (905 m)
Garcinia sp. OA 277 [375 m] (nv); SF & R. Capuron 20033 (nv)
Ochrocarpos Noronha ex Thouars Ochrocarpos parvifolius Scott-Elliot M [E, C] Harder, D. K. & al. 1581 (935 m) (nv); Harder, D. K. & al. 1602 (450 m) (nv); SF & R. Capuron 20024 [1050 m] (nv)

8. Annexes
118
Ochrocarpos sp. HP 12548 (1000 m) (nv), TYPE: P, TAN; SF & R. Capuron 688 (nv); SF & R. Rabevohitra 29385 (nv); SF & R. Rabevohitra 32011 (nv)
Symphonia L.f. Symphonia oligantha Baker M [Sb] LG 5071 (820 m); Ursch, E. 251 (nv)
COMBRETACEAE Combretum Loefl.
Combretum coursianum (H.Perrier) Jongkind M [C] Miller, J. S. & al. 3618 (1050 m) (nv)
Terminalia L. Terminalia calophylla Tul. M [Sb, W(N)] Harder, D. K. & al. 1629 (840 m) (nv); Harder, D. K. & al. 1643 (840 m) (nv)
Terminalia fatraea (Poir.) DC. M [E, W(S)] Randrianaivo, R. & al. 798 (400 m) (nv)
Terminalia sp. SF & R. Capuron 20040 (nv)
COMMELINACEAE Coleotrype C.B.Clarke
Coleotrype baroni Baker M [E, C] Coleotrype baroni Baker var. ambrensis H.Perrier SM 1738 [1045 m] (nv)
Coleotrype cf. baronii Baker MHR 043 (1050 m)
Coleotrype goudotii H.Perrier M [Sb, E, C] Coleotrype goudotii H.Perrier var. glabrescens H.Perrier Hildebrandt, J. M. 3378; LB 12028 [1225 m]; SM 1809 [1045 m] (nv)
Coleotrype goudotii H.Perrier var. goudotii SMT 044 (1060 m); SMT 064 (1021 m)
Coleotrype goudotii H.Perrier var. orientalis H.Perrier LG 5038 (1060 m); SMT 279 (1090 m); SMT 403 (1030 m)
Coleotrype madagascarica C.B.Clarke M [Sb, C] GMP 14485 [1050 m] (nv); HH 3966 (1100 m); LN 2436 (1248 m); SM 1736 [1045 m]; SM 983 (700 m)
Coleotrype sp. JB 5642 (nv)
Commelina L. Commelina diffusa Burm. f. M [Sb, E, C, W(S)] - RM, Aft, Amt LN 2438 (1248 m)
CONNARACEAE Agelaea Sol. ex Planch.
Agelaea pentagyna (Lam.) Baill. M [Sb, E, C, W(N)] - RM, Aft LG 5080 (750 m)
Ellipanthus Hook.f. Ellipanthus madagascariensis (G.Schellenb.) Capuron ex Keraudren M [Sb, E, C, W] - Aft Randrianaivo, R. & al. 794 (400 m) (nv)

8. Annexes
119
CONVOLVULACEAE Cuscuta L.
Cuscuta campestris Yunck. M [Sb, C] - RM, Aft, As, Oc, Amt, Eur LN 2390 (1260 m)
Cuscuta planiflora Ten. M [C, HM] - Aft, Amt, Eur Cuscuta planiflora var. madagascarensis (Yunck.) Verdc. HP 17725 (1200 m) (nv)
Ipomoea L. Ipomoea trifida (Kunth) G.Don M [Sb, E, W] - Aft, Amt Keraudren, M. & al. 25586 (nv)
CORNACEAE Alangium Lam.
Alangium grisolleoides Capuron M [C] Debray, M. M. 1324 (800 m) (nv); SF & R. Capuron 20056 (1000 m); SF & R. Capuron 29189 [700 m]
CRASSULACEAE Kalanchoe Adans.
Kalanchoe aromatica H.Perrier M [C, W(N)] HJBT 2063 (nv)
Kalanchoe aff. campanulata (Baker) Baill. SMT 028 (1280 m)
Kalanchoe mortagei Raym.-Hamet & H.Perrier M [W(N)] - Amt Francois, E. s.n (nv)
Kalanchoe uniflora (Stapf) Raym.-Hamet M [C] GES 1515 (1000 m) (nv); HP 17735 (1200 m) (nv), TYPE; JB 16558 (nv); JB 18446 (nv); LG 5034 (1280 m); LG 5098 (1070 m); LN 2348
(1150 m); OA 322 (1475 m) (nv); SMT 213 (1000 m)
Kalanchoe uniflora x gracilipes M [C] AMH 30 [1100 m] (nv)
Kalanchoe sp. 1 LG 5247 (1100 m)
CUCURBITACEAE Cucumis L.
Cucumis hirsutus Sond. M [C, W(N)] - Aft Keraudren, M. 1652 [1050 m] (nv)
Diplocyclos (Endl.) Post & Kuntze Diplocyclos palmatus (L.) C. Jeffrey M [W(N)] - Aft, As, Amt GES 1514 (1000 m) (nv); JB 20358 (nv); OA 275 [375 m] (nv)
Momordica L. Momordica charantia L. M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt JB 20321 [850 m] (nv); JB 5921 (nv); MHR 162 (975 m)
Raphidiocystis Hook.f. Raphidiocystis brachypoda Baker M [E, C] HP 17726 [1300 m] (nv); Keraudren, M. & al. 25590 (nv); Keraudren, M. 1650 (nv); LG 5179 (1070 m)
cf. Raphidiocystis sp. MHR 070 (1085 m)

8. Annexes
120
Zehneria Endl. Zehneria emirnensis (Baker) Keraudren M [Sb, E, C, HM] - RM, Aft OA 141 [1175 m] (nv)
Zehneria madagascariensis Keraudren M [C] SDR 134 (875 m); SMT 506 (910 m)
Zehneria perrieri Keraudren M [E, C] - RM JB 20338 (nv)
Zehneria aff. perrieri Keraudren SMT 277 (1080 m)
Zehneria polycarpa (Cogn.) Keraudren M [Sb, E, W] JB 20320 [1050 m] (nv)
Zehneria sp. OA 986 (533 m) (nv)
CUNONIACEAE Weinmannia L.
Weinmannia decora Tul. M [Sb, E, C, W(N)] LB 11961 (1000 m); SF & L. Bégué 9959 (nv); SF & N. Boanarisolo 8125 [1050 m]; SF & N. Boanarisolo 8125bis [1050 m] (nv); SF &
Randriamanalina 3493 (nv); SMT 474 (970 m)
Weinmannia sanguisugarum Bernardi M [C] LB 11999 [1175 m], TYPE: HT: G; IT: P; LG 5015 (1320 m); MHR 012 (1300 m)
Weinmannia sp. SM 2363 [1438 m] (nv)
CYPERACEAE Actinoschoenus Benth.
Actinoschoenus thouarsii (Kunth) Benth. M [Sb, E] - RM LG 5185 (1350 m)
Carex L. Carex baronii Baker M [E, C, HM] GES 2396 (1000 m) (nv); LN 2471 (1250 m)
Carex renschiana Boeck. M [Sb, E, C] LG 5122 (1420 m); MHR 100 (1100 m)
Carex sphaerogyna Baker M [C, HM] LG 5224 (1350 m)
Carex sp. Harder, D. K. & al. 1651 (840 m) (nv)
Cyperus L. Cyperus aurifer Cherm. M [C, W(N)] HP 17658 (150 m) (nv)
Cyperus balfouri C.B.Clarke M [E, C, W] - RM, Amt LG 5145 (1400 m); MHR 047 (1080 m)
Cyperus baronii C.B.Clarke M [C] - RM, Aft HP 17563 (1200 m) (nv); SM 2353 [1438 m] (nv)
Cyperus chamaecephalus Cherm. M [Sb, E, C] LG 5141 (1160 m)
Cyperus distans L.f. M [Sb, E, C, W(N)] - RM, Aft, As, Oc, Amt LG 5160 (1350 m)

8. Annexes
121
Cyperus haspan L. M [Sb, E, C, W(S)] - RM, Aft, As, Oc, Amt LG 5159 (1350 m)
Cyperus iria L. M [Sb] - RM, Aft, As, Oc, Amt JB 5913 (nv)
Cyperus laxus Lam. M [Sb, E, C, W] - RM, Aft, As, Oc, Amt SM 1775 [1045 m] (nv)
Cyperus longifolius Poir. M [E, C, ] - RM Cyperus longifolius Poir. var. condensatus Cherm. HP 18884 (nv)
Cyperus prolifer Lam. M [Sb, E, C, W(S)] - RM, Aft LN 2439 (1248 m)
Cyperus sp. JB 5351 (nv); JB 5368 (nv); PM 1172 (nv); PM 1195 (nv)
Fimbristylis Vahl Fimbristylis squarrosa Vahl M [Sb, E, C, W] - Aft, As, Oc, Amt, Eur LN 2451 (1248 m)
Kyllinga Rottb. Kyllinga intricata Cherm. M [C] - Aft, AS, Amt LN 2454 (1248 m)
Kyllinga nemoralis (J.R.Forst. & G.Forst.) Hutch. & Dalziel M [C, W(N)] - RM, Aft, As, Amt LN 2457 (1248 m)
Kyllinga sp. 1 LG 5191 (1350 m)
Kyllinga sp. JB 5547 (nv)
Machaerina Vahl Machaerina sp. SM 2371 [1438 m] (nv)
Mariscus Vahl Mariscus sp. JB 5647 (nv)
Pycreus P.Beauv. Pycreus mundtii Cherm. M [E, C, W(S), S] - RM, Aft, Eur LG 5151 (1350 m)
Pycreus polystachyos (Rottb.) P.Beauv. M [Sb, E, C, W, S] - RM, Aft, As, Oc MHR 087
Pycreus sp. 1 LN 2458 (1248 m)
Schoenoplectus (Rchb.) Palla Schoenoplectus corymbosus (Roem. & Schult.) J. Raynal M [C] - Aft LG 5171 (1350 m)
Scleria P.J.Bergius Scleria madagascariensis Boeck. M [E, C] LG 5123 (1420 m); MHR 057 (1110 m)
Scleria racemosa Poir. M [Sb, E, W(N)] - Aft JB 5892 (nv)

8. Annexes
122
DICHAPETALACEAE Dichapetalum Thouars
Dichapetalum leucosia (Spreng.) Engl. M [Sb, E, C] SM 1202 (630 m) (nv); Van Nek, F. I. 1862 (785 m) (nv); Van Nek, F. I. 1899 (635 m) (nv); Van Nek, F. I. 1930 (780 m) (nv)
Dichapetalum madagascariense Poir. M [Sb, E, C, W] - RM, Aft LG 5025 (950 m); OA 264 [375 m] (nv); SDR 026 (1180 m); SMT 077 (1150 m); Van Nek, F. I. 1923 (1025 m) (nv)
Dichapetalum madagascariense Poir. var. brevistylum Breteler GMP 14511 (1000 m) (nv)
Dichapetalum madagascariense Poir. var. madagascariense GES 1521 (1000 m) (nv)
DIDYMELACEAE Didymeles Thouars
Didymeles integrifolia J. St.-Hil. M [E] SF 7190 (nv)
Didymeles perrieri Leandri M [E, C] GES 2403 (1000 m) (nv); LG 5219 (1270 m); OA 114 [800 m]; OA 301 (1475 m); Rakotozafy, A. & al. 2300 (nv); Razafimandimbison, S.
G. 119 [1150 m] (nv); Schedl, K. E. 195 (1100 m) (nv); SF & R. Capuron 20013 [1050 m] (nv); SF & R. Capuron 20013bis [1050 m] (nv);
SM 1740 [1045 m]; SM 967 (1300 m); Ursch, E. 218 (nv)
Didymeles cf. perrieri Leandri GMP 14495 [1150 m] (nv)
DIOSCOREACEAE Dioscorea L.
Dioscorea sambiranensis R.Knuth M [Sb, W(N)] Dioscorea sambiranensis R.Knuth subsp. ambrensis H.Perrier HP 17551 [900 m] (nv)
Dioscorea sp. 1 SDR 139 (1000 m)
Dioscorea sp. 2 LN 2467 (1120 m)
EBENACEAE Diospyros L.
Diospyros gracilipes Hiern M [Sb, E, C] Du Puy, D. & al. 213 (1120 m) (nv); Du Puy, D. & al. MB213 (1300 m) (nv); MHR 031 (1015 m); MHR 155 (1000 m); OA 281 [375 m]
(nv); OA 81 [950 m] (nv); Phillipson, P. B. 1959 (900 m) (nv); SF & R. Rabevohitra 32601 [1050 m] (nv); SF & R. Rabevohitra 32637
[1050 m] (nv)
Diospyros tropophylla (H.Perrier) G.E.Schatz & Lowry M [W(S), S] Hildebrandt, J. M. 3374, TYPE: ST : G
Diospyros urschii H.Perrier M [W(N)] SF & R. Rabevohitra 32617 (nv)
Diospyros sp. 1 LG 5243 (1100 m)
Diospyros sp. 2 LG 5087 (1030 m); SDR 075 [1040 m]; SMT 387 (980 m)
Diospyros sp. 3 SMT 409 (1020 m)

8. Annexes
123
Diospyros sp. 4 LG 5033 (1200 m)
Diospyros sp. GES 1506 (1000 m) (nv); GES 1518 (1000 m) (nv); OA 64 [950 m] (nv); SF & R. Capuron 20066 (nv); SF & R. Capuron 29192 [700 m]
(nv); SF & R. Rabevohitra 32599 [1050 m] (nv); SM 1802 [1045 m] (nv)
ELAEOCARPACEAE Elaeocarpus L.
Elaeocarpus subserratus Baker M [Sb, E, C, W] AMH 95 [1100 m] (nv); GES 1502 (1000 m) (nv); Harder, D. K. & al. 1576 (935 m) (nv); LB 11947 (1000 m); LG 5072 (820 m); LG 5134
(1410 m); MHR 127 (1120 m); Rakotondrafara, A. & al. 281 (717 m) (nv); SDR 016 (1040 m); SDR 070; SF 13234 (1000 m); SF 3491; SF
3494 (nv); SF 7205 (nv); SM 1267 [645 m]; SM 1270 [645 m]
Elaeocarpus sp. OA 314 (1475 m) (nv); OA 357 [375 m] (nv); SF & R. Rabevohitra 29389 (nv); SM 1815 [1045 m] (nv)
ERICACEAE Agauria (DC.) Benth. & Hook.f.
Agauria salicifolia (Comm. ex Lam.) Hook. f. ex Oliv. M [E, C, W] - RM, Aft Agauria salicifolia var. pyrifolia (Pers.) Oliv. SM 938 (700 m)
Agauria sp. Miller, J. S. & al. 3627 (1050 m) (nv); SF & N. Boanarisolo 8124 [1050 m] (nv)
Erica L. Erica sp. OA 289 [375 m] (nv); OA 339 (1475 m) (nv)
Vaccinium L. Vaccinium madagascariense (Poir.) Sleumer M [C] LG 5133 (1410 m); MHR 003 (1430 m)
ERYTHROXYLACEAE Erythroxylum P.Browne
Erythroxylum coffeifolium Baill. M [Sb, C, W(N)] Harder, D. K. & al. 1633 (840 m) (nv)
Erythroxylum aff. coffeifolium Baill. LG 5246 (1100 m)
Erythroxylum cf. ferrugineum Cav. LB 11975 (1000 m); LG 5008 (1310 m); LG 5068 (950 m); LG 5203 (1360 m); SM 960 (1300 m); SMT 022 (1300 m)
Erythroxylum firmum Baker M [Sb, E, C] GMP 14500 [1150 m] (nv)
Erythroxylum nitidulum H. Perrier M [E, C] Erythroxylum nitidulum H. Perrier var. ambrense H. Perrier HP 5680 (nv)
Erythroxylum sphaeranthum H.Perrier M [Sb, E, C, W(N)] SMT 421 (875 m)
Erythroxylum sp. 1 SM 1194 (750 m)
Erythroxylum sp. 2 SMT 067 (1070 m)

8. Annexes
124
Erythroxylum sp. 3 SM 1209 (600 m)
Erythroxylum sp. OA 265 [375 m] (nv); PM 1187 (nv); SF & J. de D. Rakoto 11021 [1050 m] (nv)
EUPHORBIACEAE Acalypha L.
Acalypha filiformis Poir. M [Sb, E, C, W(S)] - RM Harder, D. K. & al. 1598 (965 m) (nv); LG 5199 (1350 m); MHR 027 (1020 m); MHR 064 (1045 m); SMT 147 (1040 m); SMT 150 (1055
m); SMT 258 (1020 m)
Acalypha aff. filiformis Poir. LG 5064 (840 m)
Acalypha sp. 1 LG 5076 (670 m); LG 5214 (1090 m)
Acalypha sp. AMH 443 (nv); Gillespie, L. 4096 [900 m] (nv); GMP 14478 [1050 m] (nv); JB 5951 (nv); SM 1746 [1045 m] (nv); SM 2351 [1438 m] (nv)
Anomostachys Hurus. Anomostachys lastellei (Müll. Arg.) Kruijt M [Sb, E, C] SF & J. de D. Rakoto 10948 [1050 m] (nv); SF & R. Capuron 20053 (nv)
Benoistia H.Perrier & Leandri Benoistia orientalis Radcl.-Sm. M [E, C] LG 5052 (870 m); SF & J. de D. Rakoto 11034 [1050 m] (nv); SF 11034 [1050 m] (nv); SF 201 (760 m) (nv)
Benoistia sp. SF & R. Capuron 11301 (600 m) (nv)
Claoxylon A.Juss. Claoxylon lancifolium (Prain) Leandri M [C] GES 1505 (1000 m); Labat, J.-N. & al. 2857 (1100 m) (nv); LG 5117 (1410 m); LG 5118 (1410 m); SMT 309 (1065 m)
Claoxylon sp. 1 LB 12019 [900 m]; SMT 408 (1040 m)
Claoxylon sp. 2 LG 5029 (1025 m); LG 5030 (1025 m); SMT 417 (920 m)
Claoxylon sp. GES 2394 (1000 m) (nv); Gillespie, L. 4094 [900 m] (nv); Gillespie, L. 4095 [900 m] (nv); OA 305 (1475 m) (nv); Rakotozafy, A. 1480
(nv); Rakotozafy, A. 2298bis (nv); SM 1747 [1045 m] (nv); SM 1756 [1045 m] (nv); SM 1785 [1045 m] (nv)
Cordemoya Baill. Cordemoya capuronii (Baill.) Baill. M [E, C] SF & R. Capuron 20015 [1050 m]; SMT 374 (1020 m)
Croton L. Croton argyrodaphne Baill. M [Sb, E, W] Debray, M. M. 1318 (800 m) (nv)
Croton boivinianus (Baill.) Baill. M [Sb, E] AMH 19 [1050 m] (nv)
Croton cassinoides Lam. M [E, C] AMH 94 [1100 m] (nv)
Croton fianarantsoae Leandri M [C] OA 36 [1438 m] (nv); SM 975 (1360 m)

8. Annexes
125
Croton goudotii Baill. M [C, HM] Cours, G. & al. 5370 (1100 m) (nv); HH 32143 (1000 m) (nv); LG 5167 (1350 m)
Croton goudotii var. tsaratananae (Leandri) Radcl.-Sm. AMH 105 [1100 m] (nv)
Croton jennyanus Baill. M [Sb, C, W] - RM Harder, D. K. & al. 1590 (990 m) (nv)
Croton cf. jennyanus Gris. ex Baill. SMT 027 (1280 m)
Croton minimimarginiglandulosus Radcl.-Sm. M [MdA] OA 283 [375 m]
Croton nitidulus Baker M [E, C, W(S)] Croton nitidulus Baker var. eglandulosus LB 12038 [1225 m]
Croton noronhae Baill. M [E, C] AMH 104 [1100 m] (nv)
Croton sp. 1 LG 5253 (1100 m)
Croton sp. 2 MHR 026 (1020 m)
Croton sp. 3 LG 5031 (1025 m); LG 5180 (1400 m); SMT 035 (1010 m); SMT 134 (1005 m); SMT 162 (1080 m); SMT 350 (1025 m); SMT 469 (970 m)
Croton sp. Boiteau, P. 2572 (nv); Cremers, G. 2555 (1000 m) (nv); Du Puy, D. & al. 218 (1120 m) (nv); Du Puy, D. & al. MB218 (1300 m) (nv);
Gentry, A. 11952 (1200 m) (nv); SM 1810 [1045 m] (nv)
Danguyodrypetes Leandri Danguyodrypetes sp. 1 (vel Lingelsheimia sp.) SDR 036 (1030 m)
Danguyodrypetes sp. SF & R. Capuron 20065 (nv)
Euphorbia L. Euphorbia leuconeura Boiss. M [E, C] SMT 402 (1000 m)
Euphorbia mandravioky Leandri M [W(N), S] AMH 425 (nv)
Euphorbia milii Des Moul. M [C, W] - Aft, Oc, Amt HP 17666 (nv)
Euphorbia tetraptera Baker M [Sb, E, C] AMH 413 (nv)
Euphorbia sp. 1 SMT 404 (1050 m)
Euphorbia sp. 2 LG 5215 (1100 m)
Excoecaria L. Excoecaria goudotiana (Baill.) Müll. Arg. M [C] HH 32046 (nv); HH 32079 (nv); HP 18887 (nv); SMT 415 (1010 m)

8. Annexes
126
Excoecaria thouarsiana (Baill.) Müll. Arg. M [E, C] - RM LG 5213 (1100 m); SMT 094 (1065 m)
Excoecaria sp. SM 1806 [1045 m] (nv)
Givotia Griff. Givotia cf. stipularis Radcl.-Sm. OA 998 (507 m) (nv)
Lautembergia Baill. Lautembergia sp. SF & R. Capuron 20049 (nv); SF & R. Capuron 20249 (nv); SF & R. Capuron 22049 (nv)
Macaranga Thouars Macaranga alnifolia Baker M [Sb, E, C, W] - RM AMH s.n. [1100 m] (nv); GES 1519 (1000 m) (nv); GES 1528 (1000 m) (nv); GMP 14481 [1050 m] (nv); HP 17721 (1200 m) (nv); LG
5124 (1420 m); OA 196 [900 m] (nv); OA 236 [375 m] (nv); OA 298 (1475 m) (nv); Phillipson, P. B. 1960 (900 m) (nv);
Razafimandimbison, S. G. 113 [1050 m] (nv); SM 1761 [1045 m] (nv); SM 1778 [1045 m] (nv); SM 940 (700 m) (nv); SMT 344 (1035 m);
SMT 437 (985 m); SMT 438 (985 m); SMT 470 (970 m)
Macaranga boutonioides Baill. M [Sb, E, C, W] - RM SF & J. Belin 5084 (nv); SF 14247 (nv)
Macaranga cuspidata Baill. M [Sb, E, C, W] SDR 131 (990 m)
Orfilea Baill. Orfilea sp. Rakotondrafara, A. & al. 282 (717 m) (nv)
Suregada Roxb. ex Rottler Suregada eucleoides Radcl.-Sm. M [W(S)] Harder, D. K. & al. 1569 (840 m) (nv)
Suregada sp. 1 MHR 037
Tannodia Baill. Tannodia cordifolia Baill. M [E, W] - RM SF & J. Belin 13235 (nv)
Tannodia grandiflora Radcl.-Sm. M [W(N)] GMP 14487 [1050 m] (nv); SF & J. Ch. Rabearivo 7192 (nv); SF & J. Ch. Rabearivo 7213 (nv); SF 1-R-246 (nv)
Tannodia grandiflora Radcl.-Sm.var. grandiflora RNM 1 [1050 m] (nv); SF & R. Capuron 20021 [1050 m] (nv); SF & R. Capuron 20025 [1050 m] (nv); SF & R. Capuron 20031 [1050 m]
(nv); SF 7213 (nv); SM 1189 (700 m) (nv); SM 944 (700 m) (nv), TYPE: HT: K; IT: MO
Tannodia perrieri (Leandri) Radcl.-Sm. M [E, C] - RM SM 928 (700 m) (nv)
Tannodia cf. perrieri (Leandri) Radcl.-Sm. LG 5139 (1160 m); SMT 152 (1060 m); SMT 495 (865 m); SMT 525 (655 m)
FABACEAE Acacia Mill.
Acacia pervillei Benth. M [W, S] Acacia pervillei Benth. var. pervillei SMT 480 (1010 m)

8. Annexes
127
Aeschynomene L. Aeschynomene americana L. M [Sb, W] - RM, Aft, As, Oc, Amt SMT 108
Albizia Durazz. Albizia adianthifolia (Schumach.) W.Wight M [Sb, E, C, W(N)] - Aft Albizia adianthifolia var. intermedia (De Wild. & T. Durand) Villiers Schedl, K. E. 228 (600 m) (nv)
Albizia bernieri E.Fourn. ex Villiers M [Sb, C, W, S] - RM SF 3577 (nv)
Albizia boivinii E.Fourn. M [Sb, E, C, W] SF & N. Boanarisolo 8132 [1050 m] (nv); SF 8132 (nv)
Albizia mainaea Villiers M [Sb, E, C, W] Labat, J.-N. & al. 2875 [850 m]
Albizia polyphylla E.Fourn. M [W, S] Harder, D. K. & al. 1611 (450 m) (nv)
Albizia viridis E.Fourn. M [Sb, E, C, W] Albizia viridis E.Fourn. var. viridis Du Puy, D. & al. 251 (1200 m) (nv); LG 5240 (1100 m)
Baudouinia Baill. Baudouinia fluggeiformis Baill. M [Sb, E, C, W, S] SF & N. Boanarisolo 15062 [1050 m] (nv); SF & R. Capuron 20078 (nv); SF 15062 [1050 m] (nv)
Bauhinia L. Bauhinia monandra Kurz M [W, S] - RM, Amt OA 899 (533 m) (nv); SF 528 (350 m) (nv)
Cassia L. Cassia hippophallus Capuron M [Sb, W] HP 17659 (400 m) (nv); SF 10382 (nv)
Cassia sp. JB 5942 (nv)
Centrosema (DC.) Benth. Centrosema pubescens Benth. M [Sb, E, C] - RM, Aft, As, Oc, Amt MHR 090
Chamaecrista Moench Chamaecrista telfairiana (Hook.f.) Lock M [Sb, W(N)] - RM JB 5941 (800 m) (nv)
Clitoria L. Clitoria lasciva Bojer ex Benth. M [Sb, E, W] Allorge, L. 2753 (nv); Harder, D. K. & al. 1606 (450 m) (nv); Peltier, M. 2406 (nv); SMT 109
Clitoria rubiginosa Juss. ex Pers. M [E, W] - Amt Clitoria rubiginosa Juss. ex Pers. var. rubiginosa Allorge, L. 2752 (nv)
Crotalaria L. Crotalaria alata Buch.-Ham. ex D.Don M [C] Du Puy, D. & al. 254 (1150 m) (nv); SM 1717 [1045 m] (nv)
Crotalaria bernieri Baill. M [Sb, E, C, W(N)] - RM JB 5935 (800 m) (nv); JB 5971 (800 m) (nv)

8. Annexes
128
Crotalaria toamasinae M.Pelt. M [E, W(N)] JB 5360 (nv)
Crotalaria sp. JB 5898 (nv)
Cynometra L. Cynometra abrahamii Du Puy & R.Rabev. M [E, W] AMH 428 (nv)
Cynometra sakalava Du Puy & R.Rabev. M [W] SF & J. Belin 508 (nv)
Dalbergia L.f. Dalbergia bracteolata Baker M [Sb, C, W] - Aft Leeuwenberg, A. J. M. & al. 14759 (400 m) (nv)
Dalbergia hildebrandtii Vatke M [Sb, W(N)] Louvel, M. 135 (nv)
Dalbergia madagascariensis Vatke M [Sb, E, W(N)] SF & J. Belin 13215 (300 m) (nv); SF 10668 (nv)
Dalbergia pseudobaronii R.Vig. M [Sb, W(N)] Du Puy, D. & al. 253 (1050 m) (nv); SF & L. Rajery 30553 (nv); SF & R. Capuron 20036 (nv); SF 11012 [1050 m] (nv); SF 3552 (nv); SF
3553 (nv)
Delonix Raf. Delonix boiviniana (Baill.) Capuron M [W, S] Mortages s.n. (nv)
Desmodium Desv. Desmodium adscendens (Sw.) DC. M [Sb, E, C, W] - RM, Aft, As, Oc, Amt Leeuwenberg, A. J. M. & al. 14322 (1000 m) (nv); SM 1720 [1045 m] (nv)
Desmodium incanum DC. M [Sb, E, C, W] - RM, Aft, Oc, Amt Labat, J.-N. & al. 2865 (1100 m) (nv); Peltier, M. 2405 (nv)
Desmodium ramosissimum G.Don M [Sb, E, C, W] - RM, Aft Peltier, M. 2407 (nv)
Desmodium repandum (Vahl) DC. M [Sb, E, C] - RM, Aft, As, Oc Du Puy, D. & al. 250 (1500 m) (nv); JB 5634 (1200 m) (nv); Leroy, J.-F. 48 (nv); SMT 149 (1055 m); SMT 293 (1065 m)
Desmodium triflorum (L.) DC. M [Sb, E, W] - RM, Aft, As, Oc, Amt Peltier, M. 2408 (nv)
Desmodium velutinum (Willd.) DC. M [Sb, E, C, W] - RM, Aft, As, Oc, Amt SMT 198
Dupuya J.H.Kirkbr. Dupuya madagascariensis (R.Vig.) J.H.Kirkbr. M [Sb, E, W, S] Dupuya madagascariensis (R.Vig.) J.H.Kirkbr. subsp. tamarindoides (Capuron) J.H.Kirkbr. SF 30575 (nv)
Gagnebina Neck. ex DC. Gagnebina commersoniana (Baill.) R.Vig. M [Sb, C, W, S] Du Puy, D. & al. 252 (1080 m) (nv); Harder, D. K. & al. 1601 (450 m) (nv); HH 32009 (950 m) (nv); SF & R. Capuron 29202 (nv)
Gagnebina pterocarpa (Lam.) Baill. M [W] - RM JB 5651 (nv); SF & R. Capuron 29201 (nv)

8. Annexes
129
Haematoxylum L. Haematoxylum campechianum L. M [W] - RM, Amt SF 5592 (nv)
Hymenaea L. Hymenaea verrucosa Gaertn. M [Sb, E, W] - RM, Aft Rakotonandrasana, S. & al. 911 (290 m) (nv)
Macrotyloma (Wight & Arn.) Verdc. Macrotyloma axillare (E.Mey.) Verdc. M [Sb, E, C, W, S] - RM, Aft, As, Amt Macrotyloma axillare (E.Mey.) Verdc. var. glabrum (E.Mey.) Verdc. SM 1714 [1045 m] (nv)
Millettia Wight & Arn. Millettia lenneoides Vatke M [C, W] Hildebrandt, J. M. 3370 (nv), TYPE: ST: K
Millettia richardiana (Baill.) Du Puy & Labat M [W, S] SF 12052 (nv)
Mimosa L. Mimosa capuronii Villiers M [W(N)] SF 5532 (nv)
Mucuna Adans. Mucuna paniculata Baker M [Sb, E, C, W] Du Puy, D. & al. 256 (900 m) (nv); JB 20372 (nv); OA 271 [375 m] (nv); SMT 338 (910 m)
Pachyrhizus Rich. ex DC. Pachyrhizus erosus (L.) Urb. M [C] - Aft, Amt JB 20394 (nv)
Phylloxylon Baill. Phylloxylon xylophylloides (Baker) Du Puy, Labat & Schrire M [E, C] Labat, J.-N. & al. 2872 [850 m] (nv); SF 162 (nv)
Pithecellobium Mart. Pithecellobium dulce (Roxb.) Benth. M [C, W(N), S] - RM, Aft, As, Amt PM 1400 (nv)
Pueraria DC. Pueraria lobata (Willd.) Ohwi M [MdA] - As, Amt JB 1001 (nv)
Senna Mill. Senna hirsuta (L.) H.S. Irwin & Barneby M [MdA] - RM, Aft, Amt SMT 118
Senna lactea (Vatke) Du Puy M [Sb, E, W(N)] Harder, D. K. & al. 1625 (660 m) (nv); Rakotonandrasana, S. & al. 915 (nv); SF & R. Capuron 11349 (300 m) (nv)
Senna petersiana (Bolle) Lock M [Sb, E, C, W] - Aft Harder, D. K. & al. 1626 (660 m) (nv); JB 5882 (800 m) (nv); JB 5940 (nv); Randrianaivo, R. 1189 (nv)
Strongylodon Vogel Strongylodon craveniae Baron & Baker M [Sb, E, C, W] Du Puy, D. & al. 255 (1100 m) (nv); GES 2400 (1000 m) (nv); Keraudren, M. & al. 25598bis (nv); Labat, J.-N. & al. 2862 (1100 m) (nv);
LG 5208 (1160 m); MHR 022 (1050 m); PM 1399 (1000 m) (nv); SM 1744 [1045 m] (nv); SMT 148 (1040 m); SMT 375 (1020 m); SMT
459 (1000 m)

8. Annexes
130
Sylvichadsia Du Puy & Labat Sylvichadsia grandifolia (R.Vig.) Du Puy & Labat M [Sb, E] HP 18860 (1000 m) (nv); OA 870 [900 m] (nv); SF & R. Capuron 11283 [850 m] (nv); SF & R. Capuron 29191 [700 m] (nv); SF & R-152
203 (630 m) (nv); SF 203-R-15 (630 m) (nv); Ursch, E. 204 (nv)
Viguieranthus Villiers Viguieranthus glaber Villiers M [W] LG 5079 (670 m)
Xanthocercis Baill. Xanthocercis madagascariensis Baill. M [E, W] SF & J. Belin 10287 (nv); SF 10287 (nv); SF 14876 (400 m) (nv); SF 15 (250 m) (nv)
Xylia Benth. Xylia hoffmannii (Vatke) Drake M [Sb, E, C, W] Hildebrandt, J. M. 3373 (nv), TYPE (Parkia hoffmannii Vatke) IS; SF 10286 (300 m) (nv); SF 6618
FLACOURTIACEAE Aphloia (DC.) Benn.
Aphloia theiformis (Vahl) Benn. M [Sb, E, C, W] - RM, Aft GMP 14491 [1150 m] (nv); LG 5104 (1430 m); SDR 133 (875 m); SM 1804 [1045 m] (nv); SMT 004 (1430 m); SMT 092 (1015 m)
Aphloia theiformis (Vahl) Benn. var. angusta Boivin SM 1259 (630 m) (nv)
FLACOURTIACEAE indéterminées LB 11944 [1025 m] (nv)
FLAGELLARIACEAE Flagellaria L.
Flagellaria indica L. M [Sb, E, C, W, S] - RM, As, Oc, Amt OA 68 [950 m] (nv); SMT 463 (1020 m)
GELSEMIACEAE Mostuea Didr.
Mostuea brunonis Didr. M [Sb, E, C, W] - Aft OA 76 [950 m]
Mostuea brunonis Didr. var. brunonis AMH 214 (nv); De Block, P. & al. 945; GES 2399 (1000 m) (nv); LB 11991 [1150 m]; Leeuwenberg, A. J. M. & al. 14236 (1000 m) (nv);
Leeuwenberg, A. J. M. & al. 14237 (1000 m) (nv); MHR 007 (1040 m); Razafimandimbison, S. G. 108 [1050 m] (nv); SM 1264 (590 m)
(nv); SM 932 (700 m); SMT 255 (990 m); SMT 412 (1000 m)
GENTIANACEAE Anthocleista Afzel. ex R.Br.
Anthocleista amplexicaulis Baker M [E, C] Razafimandimbison, S. G. 114 [1050 m] (nv); SF & R. Capuron 686 [1050 m] (nv); SF & R. Rabevohitra 32638 [1050 m] (nv); SM 2375
[1438 m] (nv)
Anthocleista madagascariensis Baker M [C] Harder, D. K. & al. 1570 (840 m) (nv); MHR 013 (1050 m); OA 245 [375 m] (nv); Phillipson, P. B. 1961 (900 m) (nv); PR 1165 (1430 m);
SMT 323 (965 m)
Anthocleista vogelii Planch. Détermination douteuse : OA 354 [375 m] (nv)

8. Annexes
131
GERANIACEAE Geranium L.
Geranium cf. simense Hochst. ex A.Rich. LG 5154 (1350 m)
GESNERIACEAE Hovanella A.Weber & B.L.Burtt
Hovanella madagascarica (C.B.Clarke) A.Weber & B.L.Burtt M [E, C] AMH 115 [1100 m] (nv); HP 17749 (1200 m) (nv)
Streptocarpus Lindl. Streptocarpus hildebrandtii Vatke M [Sb, C] HH 32065 (850 m) (nv)
Streptocarpus oliganthus B.L. Burtt M [E, C] Du Puy, B. MB206 (1325 m) (nv); LN 2373 (1275 m); MHR 124 (1120 m); SMT 132 (1105 m)
Streptocarpus thompsonii R. Br. M [E, C] - RM Du Puy, D. & al. 206 (1325 m) (nv)
Streptocarpus thompsonii R. Br. var. thompsonii JB 5982 (nv)
Streptocarpus sp. 1 LG 5101 (1380 m)
HALORAGACEAE Laurembergia P.J.Bergius
Laurembergia tetrandra (Spreng.) Kanitz M [E, C] - RM, Aft, Amt Laurembergia tetrandra var. numidica (Durieu ex Batt. & Trab.) A.Raynal LN 2389 (1260 m); LN 2441 (1248 m)
HEMEROCALLIDACEAE Dianella Lam. ex Juss.
Dianella ensifolia (L.) DC. M [Sb, E, C] - RM, As, Oc MHR 115 (1050 m); SMT 011 (1300 m); SMT 458 (1000 m)
Dianella sp. SM 1773 [1045 m] (nv)
HYACINTHACEAE Rhodocodon Baker
Rhodocodon urgineoides Baker M [E, C] LN 2382 (1310 m)
HYPERICACEAE Harungana Lam.
Harungana madagascariensis Lam. ex Poir. M [Sb, E, C, W] - RM, Aft OA 121 [800 m] (nv); OA 205 [900 m] (nv); OA 235 [375 m] (nv); SDR 051 (1105 m); SF & J. Ch. Rabearivo 7200 (nv); SMT 377 (990 m)
Psorospermum Spach Psorospermum androsaemifolium Baker M [E, C, W] - RM SF & Andrianandianina 30729 (nv); SF & J. Belin 13231 (900 m) (nv); SF & L. Bégué 9962 (nv)
Psorospermum aff. cerasifolium Baker Randrianaivo, R. & al. 799 (400 m) (nv)

8. Annexes
132
HYPOXIDACEAE Hypoxis L.
Hypoxis angustifolia Lam. M [Sb, E, C, W] - RM, Aft LN 2433 (1400 m)
ICACINACEAE Apodytes E.Mey. ex Arn.
Apodytes dimidiata E.Mey. ex Arn. M [E, C, W, S] - RM, Aft LB 11965 [1025 m] (nv); LG 5252 (1050 m); SF & R. Capuron 20038 (nv); SM 1212 (630 m) (nv)
Desmostachys Planch. ex Miers Desmostachys sp. Van Nek, F. I. 1954 (555 m) (nv)
Pyrenacantha Hook. Pyrenacantha ambrensis Labat, El-Achkar & R.Rabev. M [MdA] OA 62 [950 m] (nv), TYPE: HT
IRIDACEAE Aristea Aiton
Aristea cladocarpa Baker M [E, C] LG 5021 (1310 m); SMT 021 (1300 m)
Geosiris Baill. Geosiris aphylla Baill. M [E, C] SMT 229 (1085 m)
Sisyrinchium L. Sisyrinchium micranthum Cav. M [C] - Oc, Amt HH 32018 (800 m) (nv)
Sisyrinchium rosulatum E.P.Bicknell M [C] - Amt Rakotozafy, A. 1489 (nv)
LAMIACEAE Achyrospermum Blume
Achyrospermum fruticosum Benth. M [Sb, C] OA 134 [800 m]
Achyrospermum purpureum Phillipson M [Sb, E, C, HM] - RM, Aft, As, Oc LB 11955 (1000 m); Phillipson, P. B. 1969 (900 m) (nv), TYPE: HT: MO; IT: E; SMT 420 (875 m)
Achyrospermum purpureum Phillipson x urens Baker JB 5895 (1100 m) (nv)
Achyrospermum urens Baker M [E, C, HM] AMH 26 (nv); AMH 40 (nv); Du Puy, D. & al. MB219 (1120 m) (nv); GES 1508 (1000 m) (nv); HP 17748 (nv); LG 5177 (1270 m);
Phillipson, P. B. 1970 (900 m) (nv); SDR 090 (1030 m); Seigler, D. S. 12892 (nv); SM 1780 [1045 m] (nv); SMT 143 (1050 m)
Ajuga L. Ajuga oocephala Baker M [C] LG 5149 (1350 m); PM 1179 (nv); SM 1713 [1045 m] (nv)
Clerodendrum L. Clerodendrum bosseri Capuron M [MdA] SF & R. Capuron 29198 [975 m] (nv), TYPE: HT: P; IT: TEF

8. Annexes
133
Clerodendrum aff. heterophyllum SM 1274 (700 m) (nv)
Clerodendrum micans Gürke M [C, HM] OA 101 [800 m] (nv). Echantillon rattaché avec doute: Harder, D. K. & al. 1575 (935 m) (nv)
Clerodendrum putre Schau M [E, C, W] - RM Du Puy, D. & al. 202 (1325 m) (nv); MHR 129 (1030 m); MHR 152 (1040 m); SMT 362 (975 m)
Clerodendrum pyrifolium Baker M [E, C, W(S)] SDR 098 (1120 m); SF & R. Rabevohitra 32609 (nv); SMT 131 (1080 m)
Clerodendrum sp. 1 SMT 502 (775 m)
Clerodendrum sp. OA 142 [1175 m] (nv); OA 358 [375 m] (nv); Rakotozafy, A. 1488 (nv); SM 2354 [1438 m] (nv)
Madlabium Hedge Madlabium magenteum Hedge M [W(N)] AMH 434 (nv)
Ocimum L. Ocimum gratissimum L. M [E, C] - RM, Aft, As, Amt Van Nek, F. I. 1934 (720 m) (nv)
Plectranthus L'Hér. Plectranthus bojeri (Benth.) Hedge M [E, C, W] - Aft OA 45 [1438 m]
Plectranthus emirnensis (Baker) Hedge M [E, C, HM, W] GES 1503 (1000 m) (nv); HH 32027 (800 m); HH 32041 (800 m) (nv); JB 20346 (nv); LG 5046 (1240 m); Miller, J. S. & al. 3613 (1050 m)
(nv); OA 346 (1475 m); SM 1732 [1045 m]; SM 2377 [1438 m] (nv); SM 965 (1300 m)
Plectranthus vinaceus Hedge M [E, C, HM] HH 31750 (1300 m) (nv)
Plectranthus sp. 1 LG 5143 (1250 m); MHR 150 (1050 m)
Plectranthus sp. 2 LG 5115 (1420 m)
Plectranthus sp. 3 SDR 127
Plectranthus sp. 4 LG 5116 (1420 m); SMT 273 (1085 m); SMT 280 (1010 m); SMT 300 (1095 m)
Plectranthus sp. PM 1387 (1000 m) (nv); PM 1398 [1100 m] (nv)
Premna L. Premna corymbosa (Burm.f.) Rottler & Willd. M [E, C, W(N)] SM 1223 (610 m) (nv)
Pycnostachys Hook. Pycnostachys coerulea Hook. M [C] - Aft LN 2440 (1248 m)
Stachys L. Stachys reptans Hedge M [E] HH 32025 (800 m) (nv), TYPE: HT: P; IT: MO; HP 17536 (1100 m) (nv); SM 2378 [1438 m] (nv)

8. Annexes
134
Vitex L. Vitex chrysomallum Steud. M [Sb, E, C] SF & Randriamanalina 4088 (nv)
Vitex sp. OA 893 (533 m) (nv); OA 918 (662 m) (nv)
LAURACEAE Aspidostemon Rohwer & H.G.Richt.
Aspidostemon reticulatum van der Werff M [MdA] SF & R. Capuron 20011 (nv), TYPE: HT: P; IT: MO
Cryptocarya R.Br. Cryptocarya capuronii Kosterm. M [C] Harder, D. K. & al. 1642 (840 m) (nv)
Cryptocarya rotundifolia Kosterm. M [MdA] SMT 054 (1030 m)
Cryptocarya sp. 1 LG 5047 (1050 m); PR 1167 (1249 m); SMT 460 (1015 m)
Cryptocarya sp. GMP 14517 (1000 m) (nv); OA 166 [1175 m] (nv); OA 229 [375 m] (nv); OA 327 (1475 m) (nv); OA 872 [900 m] (nv)
Ocotea Aubl. Ocotea ambrensis van der Werff M [MdA] OA 270 [375 m] (nv); OA 60 [950 m] (nv); SF & R. Capuron 11290 (1000 m) (nv); SF & Randriamanalina 5467 [900 m] (nv); SM 1817
[1045 m] (nv); SMT 518 (715 m)
Ocotea aff. cymosa (Nees) Palacky Rakotondrafara, A. & al. 278 (717 m) (nv)
Ocotea laevis Kosterm. M [E, C] PM 5474 (nv); SF & N. Boanarisolo 13018 [1050 m] (nv); SF & N. Boanarisolo 13020 [1050 m] (nv); SF & N. Boanarisolo 13021 [1050 m]
(nv); SF & N. Boanarisolo 8131 [1050 m] (nv); SF & R. Rabevohitra 29383 (nv); SF & R. Rabevohitra 32615 (nv)
Ocotea racemosa (P. Danguy) Kosterm. M [E, C] HIRSM 196 (1150 m) (nv); LG 5205 (1260 m)
Ocotea sp. 1 SDR 114 (1025 m); SMT 347 (1040 m)
Ocotea sp. 2 LG 5066 (870 m); OA 165 [1175 m]; OA 365 [375 m]; SM 982 (700 m)
Ocotea sp. 3 MHR 167 (970 m)
Ocotea sp. 4 SMT 088 (1065 m); SMT 354 (975 m); SMT 357 (975 m)
Ocotea sp. GES 1497 (1000 m) (nv); SF & J. Belin 5844 (nv)
Ravensara Sonn. Ravensara acutifolia Kosterm. M [MdA] - RM (Comores) SF & J. de D. Rakoto 11014 [1050 m] (nv), TYPE: IT: TEF; SF & N. Boanarisolo 12417 (800 m) (nv); SF & N. Boanarisolo 13019 [1050
m] (nv)
Ravensara sp. SF & N. Boanarisolo 13017 (nv)

8. Annexes
135
LINDENIACEAE Lindernia All.
Lindernia rotundifolia (L.) Alston M [C] - RM, Aft HP 8500 (600 m) (nv); OA 163 [1175 m] (nv); OA 57 [1438 m] (nv); PM 1388 (1000 m) (nv); SM 1711 [1045 m] (nv)
Lindernia aff. rotundifolia (L.) Alston LG 5161 (1350 m)
LOGANIACEAE Strychnos L.
Strychnos henningsii Gilg M [E, C, W, S] - Aft SM 1235 (610 m) (nv)
Strychnos madagascariensis Poir. M [Sb, E, C, W, S] - Aft SMT 524 (655 m)
LORANTHACEAE Bakerella Tiegh.
Bakerella ambongoensis Balle M [E, C, W] SMT 487 (930 m)
Bakerella clavata (Desr.) Balle M [Sb, E, C, HM, W] - RM Bakerella clavata (Desr.) Balle var. baronii (Baker) Balle Harder, D. K. & al. 1563 (840 m) (nv)
Bakerella clavata var. aldabrensis (Turrill) Balle LG 5018 (1320 m); LG 5111 (1470 m); LG 5170 (1350 m); MHR 016 (660 m); SDR 044 (1050 m); SMT 447 (1010 m)
Bakerella clavata var. tsaratanensis (Lecomte) Balle LB 11960 (1000 m); LB 11960 bis (1000 m); LG 5032 (1200 m)
Bakerella cf. clavata (Desr.) Balle LG 5043 (1180 m)
Bakerella hoyifolia (Baker) Balle M [E, C, HM, W(S)] SMT 436 (985 m)
Bakerella sp. GMP 14520 (1000 m) (nv); PM 1189 (nv); SF & J. Belin 6620 (nv); SM 1249 [675 m] (nv); SM 1777 [1045 m] (nv); SM 2362 [1438 m]
(nv)
Taxillus Tiegh. Taxillus sp. OA 786 (874 m) (nv); OA 80 [950 m] (nv)
LYTHRACEAE Woodfordia Salisb.
Woodfordia fruticosa (L.) Kurz M [Sb, C, W] - RM, Aft, As, Oc GES 1539 (1000 m) (nv); Miller, J. S. & al. 3629 (1050 m) (nv)
MALPIGHIACEAE Acridocarpus Guill. & Perr.
Acridocarpus perrieri Arènes M [W(N)] HP 18819 (800 m) (nv), TYPE: ST
Tristellateia Thouars Tristellateia madagascariensis Poir. M [Sb, E, C, W] Labat, J.-N. & al. 2861 (1100 m) (nv); SMT 461 (1015 m)

8. Annexes
136
Tristellateia cf. pubescens Baill. SDR 137 (660 m)
Tristellateia sp. SM 1757 [1045 m] (nv)
MALVACEAE Adansonia L.
Adansonia madagascariensis Baill. M [Sb, W] - RM HP 18849 (800 m) (nv)
Adansonia perrieri Capuron M [W(N)] ? & s.n. (nv); Baum, D. & al. 298 (650 m) (nv); Baum, D. & al. 308 (650 m) (nv); Baum, D. & al. 314 (475 m) (nv); Baum, D. & al. 340
(650 m) (nv); Baum, D. & al. 341 (650 m) (nv); GMP 14539 (400 m) (nv); HH 17560 (nv); HP 17560 (900 m) (nv); LG 5078 (670 m); SF &
R. Capuron 20076 (nv); SMT 443 (650 m)
Byttneria Loefl. Byttneria heterophylla Hook. M [Sb, C, W(N)] Harder, D. K. & al. 1610 (450 m) (nv)
Byttneria melleri Baker M [E, C] - RM Byttneria melleri Baker var. melleri OA 240 [375 m]; PM 1183 (nv); SDR 125
Byttneria nitidula Baker M [C, W(N)] SM 1271 (640 m) (nv)
Byttneria sp. 1 SMT 493 (875 m)
Corchorus L. Corchorus sp. 1 LG 5255 (1080 m)
Dombeya Cav. Dombeya ankazobeensis Arènes M [C] Dombeya ankazobeensis Arènes subsp. ankazobeensis Du Puy, D. & al. 205 (1325 m) (nv)
Dombeya brevistyla Arènes M [MdA] AMH 103 (nv); AMH 79 (nv); Labat, J.-N. & al. 2863 (1100 m) (nv); SM 949 (1360 m) (nv); Ursch, E. 29 (nv), TYPE
Dombeya capuroniana Arènes M [E] SF & R. Capuron 11271 (nv), TYPE: IT: P, IST: P
Dombeya macropoda Hochr. M [E, C] AMH 80 [1100 m] (nv); GES 1500 (1000 m) (nv); GES 2397 (1000 m) (nv)
Dombeya perrieri Arènes M [MdA] HP 17771 [1200 m] (nv), TYPE: IST: P
Dombeya pubescens (Hochr.) Arènes M [C, W(S)] GES 1534 (1000 m) (nv); GMP 14510 (1000 m) (nv)
Dombeya sp. 1 SDR 132 (990 m); SMT 464 (1020 m)
Dombeya sp. 2 SDR 043 (1250 m)
Dombeya sp. 3 SDR 012 (1300 m)

8. Annexes
137
Dombeya sp. 4 LG 5169 (1350 m)
Dombeya sp. 5 SMT 516 (750 m)
Dombeya sp. 6 LG 5037 (1095 m)
Dombeya sp. 7 LN 2495 (1360 m)
Dombeya sp. 8 SDR 066 (1110 m); SMT 542 (950 m)
Dombeya sp. Gentry, A. 11950 (1200 m) (nv); GMP 14510A (1000 m) (nv); OA 151 [1175 m] (nv); OA 302 (1475 m) (nv); OA 347 (1475 m) (nv); OA
348 (1475 m) (nv); OA 356 [375 m] (nv); OA 875 [900 m] (nv); SF & R. Capuron 11269 [1050 m] (nv); SF & R. Capuron 11279 [850 m]
(nv); SF & R. Capuron 11288 [1050 m] (nv); SF & R. Capuron 29195 (1300 m) (nv); SM 1748 [1045 m] (nv); SM 1816 [1045 m] (nv); SM
2366 [1438 m] (nv)
Grewia L. Grewia cf. antsiranensis Capuron MHR 039 (1070 m)
Grewia brideliifolia Baill. M [E, C, W(N)] SF & R. Capuron 20071 (nv); SF & R. Capuron 20029 [1050 m] (nv); SF & R. Capuron 20048 (nv)
Grewia brideliifolia Baill. var. brideliifolia SF & R. Capuron 20063 (nv)
Grewia cuneifolia Juss. M [E, C] LG 5014 (1320 m); LG 5221 (1260 m); MHR 010 (1300 m); SF 11004 [1050 m] (nv); AMH 117 [1100 m] (nv); SF & R. Capuron 20034
(nv)
Grewia humblotii Baill. M [E, C] Randrianaivo, R. & al. 793 (400 m) (nv)
Grewia madagascariensis Capuron M [W] Harder, D. K. & al. 1627 (660 m) (nv); Hildebrandt, J. M. 3375, TYPE: HT: P; IT: G; LG 5250 (1100 m); SMT 510 (975 m)
Grewia triflora (Bojer) Walp. M [Sb, W, S] - RM, Aft SF & Andrianandianina 30734 (nv)
Grewia sp. AMH 440 (nv); GMP 14483 [1050 m] (nv); HP 18567 (1000 m) (nv); OA 874 [900 m] (nv)
cf. Grewia sp. LB 11969 (1000 m)
Helmiopsis H.Perrier Helmiopsis boivinii (Baill.) Arènes M [W(S)] SF & Andrianandianina 30730 (nv)
Helmiopsis pseudo-populus (Baill.) Arènes M [W(N)] Rakotonandrasana, S. & al. 910 (290 m) (nv)
Helmiopsis sphaerocarpa L.C. Barnett M [W(N)] SF & R. Capuron 29194 [700 m] (nv), TYPE: HT: P; IT: MO, P
Helmiopsis sp. SF & Andrianandianina 30726 (nv)
Hibiscus L. Hibiscus megistanthus Hochr. M [E, W(N)] SF & R. Capuron 20018 [1050 m]; SF & R. Capuron 29199

8. Annexes
138
Hibiscus sp. 1 LG 5249 (1100 m)
Nesogordonia Baill. Nesogordonia normandii Capuron M [W(N)] Schedl, K. E. 218 [1080 m] (nv); SF & Randriamanalina 2223 (nv), TYPE: ILT: TAN; SF & Randriamanalina 2780 (nv), TYPE: LT: P; ILT:
P; SF 2233 (nv)
Triumfetta L. Triumfetta pilosa Roth M [Sb, E, C] - RM, Aft, As, Oc SMT 441 (985 m); SMT 507 (930 m)
MARANTACEAE Marantochloa Brongn. ex Gris
Marantochloa comorensis Gris M [Sb, E, C, W] - RM Harder, D. K. & al. 1648 (840 m) (nv); MHR 140 (910 m); Phillipson, P. B. 1975 (900 m) (nv); SM 1261 (620 m); SMT 209 (1000 m)
MELASTOMATACEAE Dichaetanthera Endl.
Dichaetanthera oblongifolia Baker M [E, C] LG 5121 (1400 m); SDR 062 (1090 m); SM 986 (700 m)
Dichaetanthera oblongifolia Baker var. ambrensis HP 17779 (nv), TYPE: HT: P
Dichaetanthera sp. OA 118 [800 m] (nv); OA 159 [1175 m] (nv); OA 299 (1475 m) (nv); SM 1772 [1045 m] (nv); SM 2376 [1438 m] (nv)
Gravesia Naudin Gravesia ambrensis H.Perrier M [MdA] HP 17533 (1200 m) (nv), TYPE: HT; LG 5102 (1390 m)
Gravesia macrophylla (Naudin) Baill. M [Sb, E, C] HP 17535 (nv); LG 5227 (1290 m); SDR 046 (1080 m)
Medinilla Gaudich. ex DC. Medinilla ambrensis Jum. & H.Perrier M [MdA] HP 11488 (1000 m) (nv), TYPE: HT; HP 18861 (1000 m) (nv), TYPE: HT: P, syntype: 2xP; Labat, J.-N. & al. 2876 (980 m) (nv); LB 11993
[1150 m]; LB 12005 [900 m]; LG 5036 (1095 m); LG 5129 (1260 m); MHR 018 (1050 m)
Medinilla cf. baronii Baker GMP 14504 [1150 m] (nv)
Medinilla basaltarum Jum. & H.Perrier M [C] GMP 14486 [1050 m] (nv); HP 11489 (800 m) (nv), TYPE: HT; Labat, J.-N. & al. 2860 (1100 m); LB 12029 [1225 m]; LG 5035 (1095 m);
SM 980 (700 m)
Medinilla cf. humbertiana H.Perrier SM 1735 [1045 m] (nv)
Medinilla ibityensis H.Perrier M [C] SM 945 (1360 m) (nv)
Medinilla papillosa Baker M [C, HM] HP 17780 (1200 m) (nv), TYPE: HT: P, IT: P
Medinilla rotundiflora H.Perrier M [MdA] LG 5017 (1320 m); LG 5108 (1470 m)
Medinilla sphaerocarpa Hochr. M [E, C] GES 2388 (1000 m) (nv)

8. Annexes
139
Medinilla sp. OA 326 (1475 m) (nv); OA 778 (874 m) (nv); OA 884 (874 m) (nv); PM 1192 (nv); SM 1731 [1045 m] (nv)
Memecylon L. Memecylon ambrense Jacq.-Fél. M [MdA] HH 32099 (1000 m) (nv); OA 210 [900 m]; SF & R. Capuron 681 [1050 m] (nv), TYPE: IT: TEF; SF 9961 (nv); SMT 521 (1010 m).
Echantillon rattaché avec doute: Harder, D. K. & al. 1594 (965 m) (nv)
Memecylon louvelianum H.Perrier M [E, C] De Block, P. & al. 1338; Harder, D. K. & al. 1564 (840 m) (nv); OA 208 [900 m] (nv); OA 239 [375 m] (nv); OA 360
[375 m] (nv); SF 10670 (nv)
Memecylon sp. OA 276 [375 m] (nv); SF & L. Bégué 9961 (nv); SF & R. Capuron s.n. (nv); SF & R. Rabevohitra 29391 (nv); SM 1782 [1045 m] (nv)
Tristemma Juss. Tristemma mauritianum J.F.Gmel. M [Sb, E, C, W] - RM, Aft SDR 074
MELIACEAE Lepidotrichilia (Harms) J.-F.Leroy
Lepidotrichilia ambrensis J.-F.Leroy M [C, W(N)] - Aft AMH 72 [1050 m] (nv); GES 1499 (1000 m) (nv); GMP 14474 [1050 m] (nv); Harder, D. K. & al. 1562 (840 m) (nv); HH 32082 (1000 m)
(nv); HH 32083 (nv); LB 11951 (1000 m); LG 5090 (1030 m); MHR 002 (1210 m); OA 116 [800 m] (nv); OA 199 [900 m]; OA 877 [900
m]; Razafimandimbison, S. G. 102 [1050 m] (nv); Schedl, K. E. 158 (1100 m) (nv); SDR 129 (1040 m); SF & Randriamanalina 2226 (nv);
SF & Randriamanalina 5469 [1050 m] (nv); SF 10007 [1050 m] (nv); SF 10308 [1050 m] (nv); SF 11040 [1050 m] (nv); SF 11273 (1000 m)
(nv); SF 12416 [1050 m] (nv), TYPE: HT: P; IT: P; SF 20016 [1050 m] (nv); SF 7032 (nv); SF 7215 (nv); SM 987 (700 m); SMT 193 (1000
m); SMT 302 (1060 m); SMT 424 (905 m); SMT 508 (965 m)
Malleastrum J.-F.Leroy Malleastrum cf. antsingyense J.-F.Leroy Harder, D. K. & al. 1619 (660 m) (nv)
Malleastrum pseudodepauperatum J.-F.Leroy M [MdA] Harder, D. K. & al. 1617 (660 m) (nv); SF & J. Belin 10280 (450 m) (nv), TYPE: IT: TEF; SF & J. Belin 14049 (350 m) (nv); SF & J. de D.
Rakoto 11027 [1050 m] (nv); SF & N. Boanarisolo 12873 (450 m) (nv); SF & R. Capuron 11298 (650 m) (nv); SF & R. Capuron 11305 (650
m) (nv)
Malleastrum sp. 1 LB 11968 (1000 m)
Malleastrum sp. 2 MHR 138 (970 m); SMT 540 (990 m)
Malleastrum sp. 3 SDR 030 (1050 m)
Malleastrum sp. 4 SDR 071
Malleastrum sp. OA 148 [1175 m] (nv); SF & N. Boanarisolo 10419 (nv); SF & N. Boanarisolo 13022 (nv); SF & R. Rabevohitra 29399 (nv)
Trichilia P.Browne Trichilia cf. cauliflora J.-F.Leroy & Lescot LB 12016 [900 m]; LG 5059 (840 m)
Trichilia mucronata (Cav.) Harms M [E] - RM SM 935 (700 m); SMT 422 (785 m)

8. Annexes
140
Trichilia sp. GES 1537 (1000 m) (nv); JB 5924 (nv); OA 257 [375 m] (nv); OA 65 [950 m] (nv); Razafimandimbison, S. G. 109 [1050 m] (nv); SF & J.
de D. Rakoto 11025 [1050 m] (nv); SF & R. Capuron 11295 [650 m] (nv); SF & R. Rabevohitra 29390 (nv)
Turraea L. Turraea sericea Sm. M [Sb, E, W] Harder, D. K. & al. 1600 (450 m) (nv)
Turraea sp. Rakotozafy, A. 1774 (1100 m) (nv)
MENISPERMACEAE Burasaia Thouars
Burasaia madagascariensis DC. M [Sb, E, C, W] SF 6621 (nv)
Burasaia sp. OA 894 (533 m) (nv)
Cyclea Arn. ex Wight Cyclea madagascariensis Baker M [Sb, W(N)] SDR 060 (1085 m)
Triclisia Benth. Triclisia sp. 1 SDR 028
MONIMIACEAE Ephippiandra Decne.
Ephippiandra madagascariensis (Danguy) Lorence M [E, C] Gentry, A. 11951 (1200 m); Gentry, A. 11962 (1200 m) (nv); GMP 14519 (1000 m) (nv); Harder, D. K. & al. 1580 (935 m) (nv); LB 11977
[1050 m]; LN 2381 (1290 m); LN 2499 (1160 m); OA 203 [900 m] (nv); OA 917 (662 m) (nv); PM 1182 (nv); Schedl, K. E. 170 (1150 m)
(nv); Schedl, K. E. 177 (1100 m) (nv); SDR 020 (1020 m); SF & R. Capuron 689 [1050 m] (nv); SF & R. Rabevohitra 32602 [1050 m] (nv);
SF & Randriamanalina 4 (nv); SF 8-R-246 (nv); SM 1765 [1045 m]; SM 2365 [1438 m] (nv); SMT 197 (1150 m); SMT 360 (975 m); Ursch,
E. 66 [850 m] (nv)
Tambourissa Sonn. Tambourissa capuronii Cavaco M [E, C] HP 11741 (1200 m) (nv); LN 2353 (1300 m)
Tambourissa aff. capuronii Cavaco SM 970 (1470 m) (nv)
Tambourissa hildebrandtii Perkins M [E, C, W] Labat, J.-N. & al. 2859 (1100 m); OA 351 [375 m]; Razafimandimbison, S. G. 116 [1050 m] (nv)
Tambourissa madagascariensis Cavaco M [E, C] OA 333 (1475 m)
Tambourissa purpurea (Tul.) A.DC. M [Sb, E, C, W] Harder, D. K. & al. 1596 (965 m) (nv)
Tambourissa thouvenotii Danguy M [E, C] GES 2389 (1000 m) (nv)
Tambourissa cf. trichophylla Baker LG 5094 (1070 m); LG 5095 (1070 m); SMT 541 (910 m)
Tambourissa sp. 1 SMT 340 (930 m)

8. Annexes
141
Tambourissa sp. Rakotozafy, A. 2306bis (nv)
MONTINIACEAE Grevea Baill.
Grevea madagascariensis Verdc. M [W] AMH 156 (nv)
MORACEAE Ficus L.
Ficus bojeri Baker M [Sb, C, W(N)] - RM PR 1166 (1360 m)
Ficus aff. bojeri Baker SDR 076 (1090 m)
Ficus botryoides Baker M [Sb, E, C, W] Harder, D. K. & al. 1623 (660 m) (nv); MHR 088 [1005 m]; SM 1226 (600 m) (nv); SMT 401 (990 m)
Ficus brachyclada Baker M [Sb, E, C] GES 2390 (1000 m) (nv); Harder, D. K. & al. 1592 (965 m) (nv); HIRSM 147 (1100 m) (nv); MHR 075 (1105 m); Razafimandimbison, S.
G. 117 [1150 m] (nv); SDR 118 (975 m); SF & R. Capuron 20020 [1050 m] (nv); SM 1734 [1045 m] (nv); SM 1779 [1045 m] (nv); SM 927
(700 m) (nv); SM 929 (700 m) (nv); SMT 151 (1065 m); SMT 333 (930 m); SMT 369 (1010 m)
Ficus cf. brachyclada Baker MHR 110 (1030 m); SDR 001 (1360 m); SMT 337 (910 m)
Ficus lutea Vahl M [Sb, E, C, W] - RM, Aft OA 253 [375 m] (nv); Phillipson, P. B. 1968 (900 m) (nv); SDR 138 (650 m); SMT 391 (990 m)
Ficus politoria Lam. M [Sb, E, C, W] GES 1530 (1000 m) (nv); GMP 14479 [1050 m] (nv); HIRSM 147 (1100 m) (nv); LB 11957 (1000 m); LB 12000 [1150 m]; LG 5096 (1070
m); LG 5140 (1050 m); LG 5204 (1260 m); Miller, J. S. & al. 3619 (1050 m) (nv); OA 108 [800 m]; OA 262 [375 m] (nv); OA 29 [1438 m];
OA 363 [375 m] (nv); OA 66 [950 m]; OA 87 [950 m] (nv); PM 1418 [1100 m] (nv); SF 7204 (nv); SF & J. Ch. Rabearivo 7197 (nv); SM
1754 [1045 m]; SM 1762 [1045 m] (nv); SM 1781 [1045 m]; SMT 008 (1230 m); SMT 269 (1070 m); SMT 314 (1060 m); SMT 413 (990
m); SMT 484 (1000 m). Echantillon rattaché avec doute : Gentry, A. 11945 (1200 m) (nv)
Ficus reflexa Thunb. M [Sb, E, C, W] - RM LG 5245 (1100 m); MHR 035; MHR 036; MHR 147 (990 m); PR 1170 (1249 m); SDR 006 (1090 m); SDR 017 (1250 m); SMT 533 (650 m)
Ficus reflexa Thunb. subsp. reflexa Harder, D. K. & al. 1589 (990 m) (nv); OA 140 [1175 m] (nv); OA 254 [375 m] (nv); OA 290 (1475 m) (nv); OA 332 (1475 m) (nv); OA
781 (874 m) (nv); OA 991 (1105 m) (nv); SMT 191 (1080 m)
Ficus tiliifolia Baker M [Sb, E, C, W(N)] - RM Harder, D. K. & al. 1622 (660 m) (nv)
Ficus sp. Gentry, A. 11947 (1200 m) (nv); HIRSM 202 (1100 m) (nv); Miller, J. S. & al. 10680 (1050 m) (nv); Rakotozafy, A. 1479 (nv); Rakotozafy,
A. 2281 (nv)
Streblus Lour. Streblus dimepate (Bureau) C.C. Berg M [Sb, E, C] Cours, G. 956 [900 m] (nv); Rakotondrafara, A. & al. 277 (717 m) (nv); SF & R. Rabevohitra 29396 [1050 m] (nv)
Streblus mauritianus (Jacq.) Blume M [Sb, E, C, W] - RM SF 6615 (nv)
Trilepisium Thouars Trilepisium madagascariense Thouars ex DC. M [Sb, E, C, W] - RM, Aft SF & R. Capuron 11306 (600 m) (nv); SMT 501 (780 m)

8. Annexes
142
Trophis P.Browne Trophis montana (Leandri) C.C. Berg M [Sb, E, C, W] - RM AMH 89 [1100 m] (nv); GES 1532 (1000 m) (nv); GES 2387 (1000 m) (nv); GMP 14518 (1000 m) (nv); HH 32045 [950 m] (nv); HH 32101
(1000 m) (nv); HP 18858 (800 m) (nv); LB 12011 [900 m]; LG 5081 (720 m); LG 5091 (1030 m); MHR 015 (975 m); OA 912 (662 m) (nv);
OA 914 (662 m) (nv); Randrianaivo, R. & al. 795 (400 m) (nv); SF & R. Capuron 11289 [1050 m] (nv); SF & R. Capuron 20017 [1050 m];
SF & Ratsirarison 30106 [1050 m] (nv); SF 11016 [1050 m] (nv); SF 11023 [1050 m] (nv); SF 7198 (nv); SM 1178 (800 m) (nv); SM 1808
[1045 m] (nv); SM 936 (700 m) (nv)
Trophis sp. SF & J. Ch. Rabearivo 7189 [1050 m] (nv)
MUSACEAE Musa L.
Musa sp. 1 SMT 233 (1005 m); SMT 288 (1065 m)
MYRSINACEAE Ardisia Sw.
Ardisia crenata Sims M [C] - RM, Aft, As, Amt GES 1536 (1000 m) (nv); OA 876 [900 m]; Phillipson, P. B. 1964 (900 m) (nv); SM 1818 (800 m) (nv); SMT 056 (1030 m)
Embelia Burm.f. Embelia concinna Baker M [C] LG 5110 (1470 m)
Embelia sp. OA 55 [1438 m] (nv); OA 989 (809 m) (nv)
Maesa Forssk. Maesa lanceolata Forssk. M [Sb, E, C, W] - RM, Afr LB 11942 (1000 m); LG 5190 (1350 m); OA 122 [800 m]; OA 157 [1175 m]; OA 194 [900 m]; OA 243 [375 m]; OA 274 [375 m]; OA 28
[1438 m] (nv); OA 297 (1475 m); OA 330 (1475 m); PR 1168 (1249 m); SDR 015 (1050 m); SM 1192 (740 m); SM 942 [720 m]; SM 946
(1360 m) (nv); SMT 095 (1200 m); SMT 146 (1040 m)
Oncostemum A.Juss. Oncostemum boivinianum H.Perrier M [C] SF & R. Rabevohitra 32614 (nv)
Oncostemum leprosum Mez M [C] MHR 132 (1020 m); MHR 142 (975 m); SDR 121 (1025 m); SMT 239 [1005 m]; SMT 267 (1055 m)
Oncostemum longipes (Baker) Mez M [E, C] Harder, D. K. & al. 1568 (840 m) (nv); Harder, D. K. & al. 1632 (840 m) (nv)
Oncostemum racemiferum Mez M [MdA] - RM LB 12032 [1225 m]
Oncostemum reflexum Mez M [E, C] Rakotonandrasana, S. & al. 913 (290 m) (nv)
Oncostemum aff. richardianum H.Perrier Du Puy, D. & al. 220 (1120 m) (nv)
Oncostemum umbellatum (Baker) Mez M [E, C] LB 11970 (1000 m); LB 11978 [1150 m]; LB 12031 [1225 m]; LG 5042 (1240 m); SMT 036 (1270 m)
Oncostemum sp. 1 LB 11990 [1150 m]; LG 5136 (1360 m); MHR 076 (1105 m); SMT 278; SMT 454 (910 m)

8. Annexes
143
Oncostemum sp. 2 SMT 471 (970 m)
Oncostemum sp. 3 SMT 089 (1150 m)
Oncostemum sp. 4 SDR 115 (975 m); SDR 119 (1025 m); SDR 120 (1025 m); SMT 072 (1025 m); SMT 238 [1005 m]; SMT 249 (1000 m); SMT 329 (975 m);
SMT 373 (1010 m)
Oncostemum sp. 5 MHR 125 (1100 m); SDR 107 (1060 m); SMT 365 (1025 m); SMT 376 (1020 m)
Oncostemum sp. 6 SMT 483 (1000 m)
Oncostemum sp. Du Puy, D. & al. MB220 (1300 m) (nv); Gentry, A. 11955 (1200 m) (nv); GMP 14477 [1050 m] (nv); JB 5723 (nv); Miller, J. S. & al. 10676
(1050 m) (nv); Miller, J. S. & al. 3630 (1050 m) (nv); OA 103 [800 m] (nv); OA 120 [800 m] (nv); OA 155 [1175 m] (nv); OA 156 [1175 m]
(nv); OA 241 [375 m] (nv); OA 248 [375 m] (nv); OA 267 [375 m] (nv); OA 300 (1475 m) (nv); OA 303 (1475 m) (nv); OA 323 (1475 m)
(nv); OA 350 [375 m] (nv); OA 56 [1438 m] (nv); OA 61 [950 m] (nv); OA 63 [950 m] (nv); OA 784 (874 m) (nv); OA 992 (1105 m) (nv);
PM 1194 (nv); Razafimandimbison, S. G. 122 [1150 m] (nv); SF & J. Ch. Rabearivo 7206 [1050 m] (nv); SF & R. Capuron 20042 (nv); SM
2359 [1438 m] (nv)
MYRTACEAE Eugenia L.
Eugenia cf. coursiana H.Perrier LG 5176 (1140 m); MHR 041 (1085 m)
Eugenia aff. oligantha Baker Randrianaivo, R. & al. 786 (399 m) (nv)
Eugenia sp. 1 SMT 428 (890 m)
Eugenia sp. OA 51 [1438 m] (nv)
Syzygium Gaertn. Syzygium cumini (L.) Skeels M [Sb, E, C, W] - RM, Aft, As, Amt HH 3969 (900 m) (nv)
Syzygium emirnense (Baker) Labat & G.E.Schatz M [E, C] SMT 504 (760 m)
Syzygium micropodum (Baker) Labat & G.E.Schatz M [E, C] LG 5012 (1310 m)
Syzygium parkeri (Baker) Labat & G.E.Schatz M [E, C] Harder, D. K. & al. 1583 (935 m) (nv)
Syzygium cf. parkeri (Baker) Labat & G.E.Schatz MHR 069 (1040 m)
Syzygium sp. OA 145 [1175 m] (nv); OA 164 [1175 m] (nv)
NYMPHAEACEAE Nymphaea L.
Nymphaea madagascariensis DC. M [E, W] - Aft LN 2445 (1248 m)

8. Annexes
144
OCHNACEAE Campylospermum Tiegh.
Campylospermum dependens (DC.) H.Perrier M [Sb, E, C, W(N)] - RM SDR 136 (650 m)
Ochna L. Ochna sp. OA 897 (533 m) (nv)
Ouratea Aubl. Ouratea dependens (DC.) Baill. M [Sb, E, C, W(N)] - RM SM 1216 (630 m) (nv)
Ouratea sp. 1 SMT 496 (840 m)
Ouratea sp. Harder, D. K. & al. 1561 (840 m) (nv); OA 1005 (507 m) (nv); OA 207 [900 m] (nv); SF & J. Belin 13237 (800 m) (nv); SF & R.
Rabevohitra 29402 (nv)
OLACACEAE Anacolosa (Blume) Blume
Anacolosa pervilleana Baill. M [Sb, W] Randrianaivo, R. & al. 784 (399 m) (nv)
Olax L. Olax madagascariensis (DC.) Valeton M [Sb, E, W] Harder, D. K. & al. 1567 (840 m) (nv); SF & R. Capuron 11292 (750 m) (nv); SM 1263 (580 m) (nv)
OLEACEAE Chionanthus L.
Chionanthus incurvifolius (H.Perrier) Stearn M [E, C, W(N)] OA 366 [375 m] (nv)
Chionanthus sp. Leeuwenberg, A. J. M. & al. 14309 (1000 m) (nv)
Jasminum L. Jasminum elegans Knobl. M [Sb, E, W] Razafimandimbison, S. G. 107 [1050 m] (nv); SM 1784 [1045 m] (nv); SMT 512 (840 m); Van Nek, F. I. 1875 (655 m) (nv)
Noronhia Stadman ex Thouars Noronhia ambrensis H.Perrier M [W(N)] GES 1501 (1000 m) (nv); Hong-Wa, C. 573 (1000 m) (nv); HP 18839 (900 m) (nv), TYPE
Noronhia boivinii Dubard M [Sb, E, C] LG 5084 (670 m); SM 931 (700 m)
Noronhia cf. boivinii Dubard SM 943 (700 m)
Noronhia brevituba H.Perrier M [E, C, W(N)] SM 1811 [1045 m] (nv)
Noronhia cf. candicans H.Perrier SMT 535 (650 m)
Noronhia capuronii Bosser M [W(N)] SF & R. Capuron 20072 (nv), TYPE: HT: P, IT: P; SMT 536 (650 m); Van Nek, F. I. 1874 (660 m) (nv)

8. Annexes
145
Noronhia crassinodis H.Perrier M [W(N)] SDR 034 (1162 m)
Noronhia gracilipes H.Perrier M [E, C] Besnard, G. 46-2006 (800 m); GES 1504 (1000 m); Hong-Wa, C. & al. 581 (nv); Hong-Wa, C. & al. 582 (1084 m) (nv); SDR 038; SDR 104
(1030 m); SDR 128 (1040 m); SMT 335 (910 m)
Noronhia sp. 1 SMT 383 (1035 m); SMT 543 (1030 m)
Noronhia sp. 2 OA 851 (533 m)
Noronhia sp. GMP 14512 (1000 m) (nv); OA 130 [800 m] (nv); OA 191 [900 m] (nv); OA 269 [375 m] (nv); OA 273 [375 m] (nv); OA 313 (1475 m)
(nv); OA 983 (533 m) (nv); Razafimandimbison, S. G. 121 [1150 m] (nv); SF & J. Belin 14248 [1050 m] (nv); SF & R. Rabevohitra 29382
(nv); SF & R. Rabevohitra 32612 (nv)
Olea L. Olea ambrensis H.Perrier M [E, C] HIRSM 187 (1150 m) (nv); HP 17706 [1100 m] (nv), TYPE: HT; LG 5097 (1070 m)
Olea capensis L. M [Sb, E, C, W(N)] - Aft LB 11967 (nv); OA 201 [900 m] (nv)
Olea madagascariensis H.Perrier M [Sb, E, C, W(N)] - Aft GES 1525 (1000 m); GMP 14515 (1000 m) (nv); Harder, D. K. & al. 1579 (935 m) (nv); SMT 511 (970 m)
Olea sp. SF & J. Belin 13236 [1050 m] (nv); SF & L. Bégué 9958 (nv); SF & N. Boanarisolo 8129 [1050 m] (nv); SF & R. Capuron 11287 (1000 m)
(nv)
ONAGRACEAE Ludwigia L.
Ludwigia octovalvis (Jacq.) P.H. Raven M [Sb, W] Ludwigia octovalvis (Jacq.) P.H. Raven subsp. octovalvis LN 2449 (1248 m); SMT 214 (1000 m); SMT 431 (885 m)
ORCHIDACEAE Aerangis Rchb.f.
Aerangis citrata (Thouars) Schltr. M [Sb, E, C] Harder, D. K. & al. 1585 (935 m) (nv); SDR 083 (970 m)
Aerangis hyaloides (Rchb.f.) Schltr. M [E] Toilliez, J. s.n. (nv)
Aerangis modesta (Hook.f.) Schltr. M [Sb, E, C] AMH s.n. [1100 m] (nv); HP 18881 (1000 m) (nv)
Aerangis stylosa (Rolfe) Schltr. M [E, C] - RM HP 18833 [800 m] (nv); HP 18879 (1000 m) (nv)
Aerangis sp. LG 5232 (1160 m)
Aeranthes Lindl. Aeranthes ambrensis Toill.-Gen., Ursch & Bosser M [MdA] HJBT & L. Bégué 246 (nv), TYPE: HT
Aeranthes cf. ambrensis Toill.-Gen., Ursch & Bosser SDR 109 (1060 m)

8. Annexes
146
Aeranthes caudata Rolfe M [C] - RM MHR 145 (1020 m); SDR 108 (1060 m)
Aeranthes ecalcarata H.Perrier M [C] HP 17715 (1200 m) (nv), TYPE: HT
Aeranthes grandiflora Lindl. M [E, C] - RM Bernier, J. 350 (nv)
Aeranthes moratii Bosser M [E] OA 84 [900 m] (nv)
Aeranthes schlechteri Bosser M [Sb, W(N)] SM 1240 (620 m) (nv)
Aeranthes cf. schlechteri Bosser SMT 526 (650 m)
cf. Aeranthes sp. SDR 079 (1110 m)
Ambrella H.Perrier Ambrella longituba H.Perrier M [MdA] Du Puy, D. s.n (nv); HP 18811 (700 m) (nv), TYPE: HT
Angraecum Bory Angraecum aloifolium J.Hermans & P.J.Cribb M [W(S)] LN 2492 (1100 m)
Angraecum ambrense H.Perrier M [MdA] HP 17719 (1200 m) (nv), TYPE: HT
Angraecum angustipetalum Rendle M [MdA] - Aft MHR 074 (1100 m)
Angraecum bicallosum H.Perrier M [MdA] HP 18874 (1000 m) (nv), TYPE: HT; SMT 010 (1300 m)
Angraecum caricifolium H.Perrier M [E, C] MHR 094 (1100 m)
Angraecum conchoglossum Schltr. M [C] - RM MHR 034 (1070 m); SM 2369 [1425 m] (nv)
Angraecum cornucopiae H.Perrier M [C] LG 5178 (1380 m)
Angraecum cf. curvicaule Schltr. SMT 016 (1280 m); SMT 363 (1025 m)
Angraecum drouhardii H.Perrier M [E] HP 19289 (200 m) (nv), TYPE: HT
Angraecum eburneum Bory M [Sb, E, W(S), S] - RM SM 1211 (600 m)
Angraecum florulentum Rchb. f. M [C] - RM SMT 055 (1030 m)
Angraecum germinyanum Hook. f. M [E, C] - RM HP 18876 (1000 m) (nv), TYPE: ST; JB s.n (nv)
Angraecum mauritianum (Poir.) Frapp. M [E, C] - RM MHR 106 (980 m); SDR 089 (1000 m); SMT 237 [1005 m]
Angraecum musculiferum H.Perrier M [C] HP 17737 (1200 m) (nv), TYPE: ST; MHR 135 (1060 m); SMT 345 (1035 m)

8. Annexes
147
Angraecum pseudodidieri H.Perrier M [C] HP 18864 (1000 m) (nv), TYPE
Angraecum pterophyllum H.Perrier M [C] LN 2425 (1140 m); LN 2490 (1100 m)
Angraecum ramosum Thouars M [E, C] - RM JB 1495 (nv); Keraudren, M. s.n. (nv)
Angraecum sacculatum Schltr. M [C] MHR 095 [1040 m]
Angraecum subcordatum (H.Perrier) Bosser M [C] SMT 252 (970 m)
Angraecum urschianum Toill.-Gen. & Bosser M [C] SMT 326 (955 m)
Angraecum sp. JB s.n. (nv); SMT 097 (1200 m)
Beclardia A.Rich. Beclardia macrostachya (Thouars) A.Rich. M [C] - RM JB 20373 (nv); PM 1231 (nv)
Bulbophyllum Thouars Bulbophyllum ambrense H.Perrier M [E] HP 17718 (1200 m) (nv), TYPE: HT
Bulbophyllum coriophorum Ridl. M [E, C] - RM SMT 023 (1300 m)
Bulbophyllum longiflorum Thouars M [Sb, E, C] - RM, Aft, As, Oc Cremers, G. 2549 (1000 m) (nv)
Bulbophyllum molossus Rchb.f. M [C] SMT 096 (1200 m); SMT 317 (970 m)
Bulbophyllum multiligulatum H.Perrier M [C] SMT 020 (1300 m)
Bulbophyllum nutans Thouars M [C] - RM Bulbophyllum nutans Thouars var. variifolium (Schltr.) Bosser HP 17716 (1200 m) (nv), TYPE: HT
Bulbophyllum occlusum Ridl. M [Sb, E, C] LN 2408 (1332 m)
Bulbophyllum occultum Thouars M [Sb, E, C] - RM SMT 388 (980 m)
Bulbophyllum pervillei Rolfe M [Sb, E] HP 18832 (1000 m) (nv), TYPE: HT
Bulbophyllum septatum Schltr. M [C, HM] HP 18873 (1000 m) (nv), TYPE: HT
Bulbophyllum subcrenulatum Schltr. M [C] Labat, J.-N. & al. 2869 (900 m) (nv)
Bulbophyllum vulcanorum H.Perrier M [MdA] HP 18877 (1000 m) (nv), TYPE: HT: P, IT: P
Bulbophyllum sp. 1 LN 2400 (1300 m); SMT 042 (1270 m)
Bulbophyllum sp. 2 SMT 053 (1000 m)

8. Annexes
148
Bulbophyllum sp. 3 LG 5019 (1320 m); MHR 011 (1300 m); SMT 427 (905 m)
Bulbophyllum sp. Du Puy, D. & al. MB208 (1325 m) (nv); SM 1794 [1045 m] (nv)
Calanthe R.Br. Calanthe madagascariensis Rolfe M [E, C] HP 18472 (nv); SMT 172 (1100 m)
Calanthe sylvatica (Thouars) Lindl. M [Sb, E, C] - RM, Aft, As LG 5137 (1400 m); MHR 099 (1100 m); SMT 385 (1040 m)
Calanthe sp. OA 154 [1175 m] (nv); OA 783 (874 m) (nv)
Cheirostylis Blume Cheirostylis gymnochiloides (Ridl.) Rchb.f. M [E, C, W(S)] - RM LG 5230 (1310 m)
Cheirostylis nuda (Thouars) Ormerod M [E, C, W(S)] - RM LG 5231 (1320 m)
Corymborkis Thouars Corymborkis corymbosa (Ridl.) Kuntze M [E] - Aft SM 1221 (620 m) (nv)
Cynorkis Thouars Cynorkis boinana Schltr. M [W] Veyret, Y. s.n. (nv)
Cynorkis fastigiata Thouars M [Sb, E, C] - RM SM 1207 (950 m) (nv)
Cynorkis nutans (Ridl.) H. Perrier M [C] LG 5113 (1460 m)
Cynorkis quinquepartita H.Perrier M [E] HP 17738 (1200 m) (nv), TYPE: HT
Cynorkis ridleyi T.Durand & Schinz M [C] - RM HP 17720 (1200 m) (nv); LG 5138 (1400 m)
Cynorkis rosellata (Thouars) Bosser M [C] - RM AMH 77 [1100 m] (nv); HP 17739 (1200 m) (nv)
Cynorkis schlechterii H.Perrier M [E] LN 2375 (1400 m); SMT 168 (1100 m)
Cynorkis stenoglossa Kraenzl. M [E, C] Cynorkis stenoglossa Kraenzl. var. pallens H.Perrier HP 17733 (1200 m) (nv), TYPE: HT:P, IT:P; SM 2331 [1438 m] (nv)
Cynorkis stenoglossa Kraenzl. var. stenoglossa AMH 44 [1100 m] (nv); AMH s.n. [1100 m] (nv); Saboureau, P. s.n. (nv)
Cynorkis stolonifera (Schltr.) Schltr. M [C] LG 5153 (1350 m); SM 950 (1360 m)
Cynorkis tryphioides Schltr. M [Sb, W] HH 32147 (1000 m) (nv); HP 17558 (900 m) (nv)
Disperis Sw. Disperis hildebrandtii Rchb.f. M [Sb, E, C] - RM Allorge, L. & al. 2722 (nv); HH 32144 (1000 m) (nv)

8. Annexes
149
Disperis humblotii Rchb.f. M [C] - RM HP 17740 (nv)
Disperis oppositifolia Sm. M [Sb, E] - RM SMT 046 (1060 m)
Gastrorchis Schltr. Gastrorchis geffrayi (Bosser) Senghas M [MdA] Peyrot, J. P. 50 (nv), TYPE: HT: P, IT: P
Gastrorchis humblotii (Rchb.f.) Schltr. M [C] AMH s.n. [1100 m] (nv); HH 32075 [1050 m] (nv); HH 32115 [1050 m] (nv); HJBT & J. Bosser 30 (nv); JB s.n (nv); MHR 050 (1080 m);
OA 949 (662 m) (nv); SM 1208 (880 m) (nv); SMT 386 (1035 m)
Gastrorchis sp. SM 1208 (880 m) (nv)
Habenaria Willd. Habenaria saprophytica Bosser & P.J.Cribb M [MdA] SM 1214 (610 m) (nv), TYPE: IT: MO
Hetaeria Blume Hetaeria heterosepala (Rchb. f.) Summerh. M [MdA] - Aft SMT 390 (990 m)
Jumellea Schltr. Jumellea ambrensis H.Perrier M [MdA] HP 19015 (1000 m) (nv), TYPE: HT
Jumellea flavescens H.Perrier M [E, C] HP 17543 (1200 m) (nv), TYPE: ST
Jumellea cf. punctata H.Perrier SMT 372 (1015 m)
Jumellea sp. 1 SMT 406 (1040 m)
Jumellea sp. AMH 215 (nv); HH 32116 (1000 m) (nv); JB 19614 (nv); JB 20178 (1300 m) (nv); Keraudren, M. s.n. (nv)
Lemurella Schltr. Lemurella pallidiflora Bosser M [C] HH 32146 (1000 m) (nv); MHR 134 (1060 m)
Liparis Rich. Liparis anthericoides H.Perrier M [E, C] JB 13539 (nv); SMT 384 (1055 m)
Liparis clareae Hermans M [E, C] MHR 143 (1025 m); SDR 049 (1070 m)
Liparis listeroides Schltr. M [C] SDR 050 (1070 m); SMT 263 (1040 m)
Liparis cf. panduriformis H.Perrier SDR 106 (1080 m); SMT 304 (1060 m)
Liparis purpurascens Lindl. M [C] - RM HP 17724 (1200 m) (nv)
Liparis salassia (Pers.) Summerh. M [C] - RM LN 2362 (1200 m); SDR 056 (1040 m)

8. Annexes
150
Liparis sp. JB 21170 (nv); OA 52 [1438 m] (nv)
Malaxis Sol. ex Sw. Malaxis physuroides (Schltr.) Summerh. M [C] LN 2374 (1400 m)
Nervilia Comm. ex Gaudich. Nervilia affinis Schltr. M [E, C] LG 5120 (1400 m)
Nervilia cf. affinis Schltr. LG 5209 (1170 m)
Oberonia Lindl. Oberonia disticha (Lam.) Schltr. M [Sb, E, C] - RM, Aft AMH 9 [1050 m] (nv); LN 2447 (1248 m); Poisson, H. 135 (nv); SDR 086 (970 m); SMT 207 (1000 m)
Oeceoclades Lindl. Oeceoclades ambrensis (H.Perrier) Bosser & Morat M [MdA] HH 32154 (1000 m) (nv); HH 32175; HH 3964 (1100 m) (nv), TYPE: HT: P, TYPE; JB 14732 [1050 m] (nv); LG 5070 (1030 m); MHR
021 (1010 m); SMT 274 (1085 m)
Oeceoclades pulchra (Thouars) M.a.Clem. & P.J.Cribb M [Sb, E, C] - RM, Aft, As, Oc HP 18455 (nv)
Oeonia Lindl. Oeonia rosea Ridl. M [E, C] MHR 096 [1040 m]; MHR 131 (1080 m); SMT 445 (985 m)
Phaius Lour. Phaius pulchellus Kraenzl. M [E, C] LN 2378 (1420 m)
Phaius pulchellus Kraenzl. var. ambrensis Bosser JB 20785 (nv), TYPE: HT: P, IT: P
Phaius pulchellus Kraenzl. var. pulchellus HH 32076 [1050 m] (nv)
Platylepis A.Rich. Platylepis densiflora Rolfe M [E, C] SMT 361 (975 m)
Polystachya Hook. Polystachya cultriformis (Thouars) Spreng. M [Sb, E, C] - RM, Aft AMH 220 (nv); Harder, D. K. & al. 1578 (935 m) (nv); MHR 098 (1080 m); MHR 144 (990 m); SM 2346 [1438 m] (nv); SMT 159 (1050
m); SMT 259 (1040 m)
Polystachya sp. 1 SDR 057 (1060 m)
Satyrium Sw. Satyrium trinerve Lindl. M [E, C] - RM, Aft HH 32148 (900 m) (nv)
Sobennikoffia Schltr. Sobennikoffia poissoniana H.Perrier M [W(N)] Poisson, H. 43 (nv), TYPE: HT

8. Annexes
151
Solenangis Schltr. Solenangis cornuta (Rchb.f.) Summerh. M [W] - RM AMH 429 (nv)
ORCHIDACEAE indéterminées SDR 078 (1110 m)
OROBANCHACEAE Alectra Thunb.
Alectra cf. sessiliflora (Vahl) Kuntze LG 5061 (760 m)
OXALIDACEAE Biophytum DC.
Biophytum sp. Labat, J.-N. & al. 2871 [850 m] (nv)
Oxalis L. Oxalis sp. 1 LG 5193 (1350 m)
Oxalis sp. 2 LG 5248 (1100 m)
PANDANACEAE Pandanus Parkinson
Pandanus connatus H.St.John M [MdA] Callmander, M. W. 45 (940 m) (nv); Callmander, M. W. 46 (1000 m) (nv); Cremers, G. 2583 (nv); OA 220 [900 m]; SDR 063 (1090 m);
SMT 472 (970 m); St.John, H. 26578 (1000 m) (nv), TYPE: HT: BISH
Pandanus latistigmaticus Huynh M [W(N)] Callmander, M. W. 47 (940 m) (nv); MHR 092; Phillipson, P. B. 1974 (900 m) (nv), TYPE: HT: MO; Ranaivojaona, R. & al. 1575 (140 m)
(nv); SMT 342 (1025 m); SMT 466 (1000 m)
Pandanus nusbaumeri Callm. & L.Gaut. M [MdA] LG 5217 (1170 m), TYPE: HT:G, IT: P, MO
Pandanus spicatus H.St.John M [E] Callmander, M. W. 44 (990 m) (nv); LN 2428 (1230 m); St.John, H. 26577 (1000 m) (nv)
Pandanus sp. LG 5254 (1100 m); MHR 038
PASSIFLORACEAE Erblichia Seem.
Erblichia madagascariensis O. Hoffm. M [W(S)] Hildebrandt, J. M. 3376 (nv)
PEDALIACEAE Sesamum L.
Sesamum indicum L. M [Sb, E, C, W] - RM, Aft, As, Amt Harder, D. K. & al. 1612 (450 m) (nv)
Uncarina Stapf Uncarina peltata (Baker) Stapf, in Engler M [W] HP 18851 (700 m) (nv); OA 1001 (507 m) (nv)

8. Annexes
152
PHYLLANTHACEAE Antidesma L.
Antidesma madagascariense Lam. M [Sb, E, C, W, S] - RM Harder, D. K. & al. 1616 (660 m) (nv); LB 12020 [1225 m] (nv); Miller, J. S. & al. 3621 (1050 m) (nv); SDR 021 (1000 m); SF 26 (nv); SM
1241 (610 m) (nv); SMT 382 (1035 m); SMT 389 (980 m); SMT 476 (970 m)
Antidesma sp. SF & R. Capuron 29200 (nv)
Bridelia Willd. Bridelia pervilleana Baill. M [Sb, C, W] Harder, D. K. & al. 1613 (450 m) (nv)
Cleistanthus Hook.f. ex Planch. Cleistanthus capuronii Leandri M [Sb, E, W(N)] Randrianaivo, R. & al. 789 (399 m) (nv)
Meineckia Baill. Meineckia madagascariensis (Humbert) G.L. Webster M [C] SDR 019 (1250 m)
Phyllanthus L. Phyllanthus cf. andalangiensis Leandri LN 2473 (1400 m); SDR 123
Phyllanthus sp. 1 LG 5225 (1350 m)
Thecacoris A.Juss. Thecacoris sp. 1 SMT 003 (1210 m)
Wielandia Baill. Wielandia elegans Baill. M [Sb, E, W] - RM HH 32072 (1000 m) (nv); Miller, J. S. & al. 3617 (1050 m) (nv); SF & R. Capuron 11282 (1000 m) (nv)
Wielandia platyrachis (Baill.) Petra Hoffm. & McPherson M [Sb, E, C] Du Puy, D. & al. 214 (1120 m) (nv); Du Puy, D. & al. MB214 (1300 m) (nv); Harder, D. K. & al. 1620 (660 m) (nv)
PHYSENACEAE Physena Noronha ex Thouars
Physena madagascariensis Tul. M [Sb, E, C, W(N)] AMH 154 (nv); AMH 157 (nv); AMH 176 (nv); LG 5058 (840 m); OA 255 [375 m] (nv); OA 361 [375 m] (nv); Razafimandimbison, S. G.
111 [1050 m] (nv); SF 11009 [1050 m] (nv); SM 1217 (630 m) (nv); Van Nek, F. I. 1779 (850 m) (nv)
PHYTOLACCACEAE Hilleria Vell.
Hilleria latifolia (Lam.) H.Walter M [E, C] - Aft, Amt HH 32098 (1000 m); HP 17539 (900 m) (nv); HP 18847 (900 m) (nv); HP 8669 (900 m) (nv); LN 2430 (1230 m); SM 1807 [1045 m] (nv);
SMT 085 (1065 m)
Phytolacca L. Phytolacca dodecandra L'Hér. M [E, C, W] - Aft SMT 236

8. Annexes
153
PIPERACEAE Peperomia Ruiz & Pav.
Peperomia bangroana C.DC. M [E, C, W(S)] - Aft SMT 058 (1015 m)
Peperomia glabrilimba C.DC. M [E, C, HM] MHR 045 (1050 m)
Peperomia imerinae C.DC. M [Sb, C] AMH 111 [1100 m] (nv); HH 32036 (nv); Mathieu, G. 455 (1400 m) (nv); Mathieu, G. 483 (nv)
Peperomia pluvisilvatica G.Mathieu M [MdA] AMH 112 [1100 m] (nv); AMH 199 (nv); HH 32035 (nv); Mathieu, G. 452 (1100 m) (nv); Mathieu, G. 456 (1400 m), TYPE: HT: BR; IT:
B, G, MO, P, TAN; PM 1396 (nv); SMT 019 (1300 m)
Peperomia terebinthina G.Mathieu M [MdA] AMH 32 (nv); Cours, G. & al. 5360 (nv); LG 5147 (1350 m); LN 2370 (1200 m); Mathieu, G. 457 (1352 m) (nv), TYPE: HT: BR; IT: B;
PM 1205 (nv)
Peperomia tetraphylla Hook. & Arn. M [C] - RM, Aft, As, Oc, Amt LG 5237 (1100 m); SMT 226 (1000 m)
Peperomia thomeana C.DC. M [C] - Aft AMH 113 [1100 m] (nv); AMH 114 [1100 m] (nv); AMH 17 [1050 m] (nv); HH 32034 (800 m) (nv); LB 11996 (nv); LN 2357 (1320 m);
Mathieu, G. 451 (1000 m) (nv); Mathieu, G. 454 (1400 m) (nv); MHR 082 (1060 m); SM 2357 [1438 m] (nv); SM 973 (1470 m) (nv)
Peperomia sp. 1 SMT 256 (1000 m)
Peperomia sp. Mathieu, G. 453 (1100 m) (nv)
Piper L. Piper capense L. f. M [C, HM] - RM, Aft LG 5201 (1320 m)
Piper guineense Schumach. & Thonn. M [C] - Aft LG 5207 (1280 m); SDR 003 (1180 m); SDR 100 (1100 m); SM 958 (1300 m); SMT 038 (1270 m); SMT 305 (1065 m); SMT 308 (1065 m)
Piper subpeltatum Willd. M [Sb, E, C, W(S)] - RM, Aft, As, Oc, Amt HH 3968 (1100 m) (nv)
Piper umbellatum L. M [Sb, E, C, W(S)] - RM, Aft, As, Oc, Amt MHR 049 (1080 m); MHR 111 (1030 m); OA 50 [1438 m] (nv); SM 1182 (670 m) (nv); SM 1728 [1045 m] (nv); SMT 005 (1040 m)
Piper sp. GMP 14480 [1050 m] (nv); OA 288 [375 m] (nv); OA 353 [375 m] (nv)
PITTOSPORACEAE Pittosporum Banks ex Gaertn.
Pittosporum ambrense Cufod. M [C] GMP 14475 [1050 m] (nv); GMP 14489 [1050 m] (nv); HP 2305 (800 m) (nv), TYPE; Labat, J.-N. & al. 2858 (1100 m); OA 249 [375 m];
SF & R. Rabevohitra 32607 (nv); SM 1771 [1045 m]
Pittosporum verticillatum Bojer M [E, C] Harder, D. K. & al. 1577 (935 m) (nv)
Pittosporum verticillatum Bojer var. microphyllum Cufod. HP 17709 (1000 m) (nv), TYPE; SF & R. Capuron 11270 (nv)
Pittosporum verticillatum Bojer var. tsaratananae Cufod. SF & R. Capuron 22057 [900 m]

8. Annexes
154
Pittosporum viridiflorum Sims M [Sb, E, C, W, S] - Aft OA 268 [375 m]
Pittosporum sp. GES 2392 (1000 m) (nv); HIRSM 198 (1150 m) (nv); OA 113 [800 m] (nv); OA 161 [1175 m] (nv); OA 202 [900 m] (nv); OA 853 (533 m)
(nv); OA 90 [950 m] (nv); SF & R. Capuron 24057 [900 m] (nv)
PLUMBAGINACEAE Plumbago L.
Plumbago dawei Rolfe M [C, S] - Aft JB 20332 [850 m] (nv)
Plumbago zeylanica L. M [Sb] - RM, Aft, As, Oc, Amt HP 18841 (800 m) (nv)
POACEAE Acroceras Stapf
Acroceras bosseri A. Camus M [E, C, W(S), S] - RM JB 5330 [1100 m] (nv), TYPE: ST
Bothriochloa Kuntze Bothriochloa glabra (Roxb.) A. Camus M [C] - Aft, As, Amt JB 5347 (nv)
Brachiaria (Trin.) Griseb. Brachiaria tsiafajavonensis A.Camus M [C] JB 5338 (nv)
Brachiaria sp. JB 5515 [1050 m] (nv)
Chloris Sw. Chloris pycnothrix Trin. M [C] - Aft, Amt PM 1174 [1050 m] (nv)
Coix L. Coix lacryma-jobi L. M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt, Eur PM 1165 (nv)
Cynodon Rich. Cynodon sp. JB 5310 (nv); JB 5525 (nv)
Cyrtococcum Stapf Cyrtococcum deltoideum (Hack.) A. Camus M [C] JB 5540 (nv)
Cyrtococcum patens (L.) A. Camus M [E] - RM, As Besnard, G. 42-2006 (800 m)
Digitaria Haller Digitaria debilis (Desf.) Willd. M [Sb, E, C, W] - Aft LN 2460 (1248 m)
Digitaria horizontalis Willd. M [Sb, E, C, W] - RM, Aft, As, Amt JB 5449 (nv); JB 5639 (nv); PM 1159 [1150 m] (nv)
Digitaria longiflora (Retz.) Pers. M [C] - RM, Aft, As, Amt JB 5331 (nv)

8. Annexes
155
Digitaria thouaresiana (Flüggé) A.Camus M [E] - RM, Aft LG 5165 (1350 m)
Digitaria sp. PM 1171 (nv)
Eragrostis Wolf Eragrostis ambrensis A.Camus M [MdA] HP 17713 (1200 m) (nv)
Eragrostis sp. JB 5528 (nv); PM 1176 [1050 m] (nv)
Isachne R.Br. Isachne mauritiana Kunth M [Sb, E, C] - RM, Aft SMT 194 (1100 m); SMT 245 (1000 m)
Isachne sp. JB 5366 (nv)
Nastus Juss. Nastus ambrensis A.Camus M [MdA] HP 17765 (1000 m) (nv), TYPE: HT: P, IT: P
Olyra L. Olyra latifolia L. M [Sb, E, C, W] - RM, Aft, As, Oc Harder, D. K. & al. 1631 (840 m) (nv); SDR 126; SM 1183 (680 m) (nv)
Oplismenus P.Beauv. Oplismenus hirtellus (L.) P.Beauv. M [E] - Aft, Amt Gentry, A. 11961 (1200 m) (nv); SMT 289 (1065 m)
Oplismenus sp. JB 5365 (nv); JB 5550 (nv); JB 5554 (nv); JB 5555 (nv)
Panicum L. Panicum humbertii A. Camus M [Sb] JB 5342 (nv)
Panicum muscicola A. Camus M [MdA] HP 19300 (1200 m) (nv), TYPE: HT
Panicum umbellatum Trin. M [Sb, E, C, W] - RM, Aft LG 5200 (1350 m)
Panicum uvulatum Stapf M [Sb, E, C] - RM, Aft PM 1175 (1000 m) (nv); SMT 127 (1080 m)
Panicum sp. 1 LG 5188 (1350 m)
Panicum sp. 2 SMT 481 (1000 m)
Panicum sp. JB 5302 (nv); JB 5448 (nv); JB 5536 (nv); JB 5545 (nv); JB 5667 (nv)
Paspalum L. Paspalum conjugatum P.J. Bergius M [Sb, E, C, W] - RM, Aft, As, Oc, Amt SMT 440 (985 m)
Paspalum sp. Miller, J. S. & al. 10683 (1050 m) (nv); PM 1173 [1050 m] (nv)

8. Annexes
156
Poecilostachys Hack. Poecilostachys bromoides Stapf M [C] Besnard, G. 43-2006 (800 m)
Poecilostachys sp. 1 LG 5146 (1400 m); SMT 034 (1010 m)
Poecilostachys sp. JB 5313 (nv); JB 5537 (nv)
Pseudolasiacis (A.Camus) A.Camus Pseudolasiacis leptolomoides (A. Camus) A. Camus M [C] JB 5530 (nv)
Setaria P.Beauv. Setaria chevalieri Stapf M [Sb, E, C] - RM, Aft LN 2500 (1160 m)
Sporobolus R.Br. Sporobolus centrifugus (Trin.) Nees M [C] - Aft LG 5166 (1350 m)
Sporobolus aff. paniculatus (Trin.) T.Durand & Schinz LG 5189 (1350 m)
Stenotaphrum Trin. Stenotaphrum dimidiatum (L.) Brongn. M [E, C] - RM, Aft LN 2444 (1248 m)
Stenotaphrum oostachyum Baker M [C] HP 11272 (nv)
Themeda Forssk. Themeda quadrivalvis (L.) Kuntze M [C, W] - Aft, Amt Besnard, G. 40-2006 (700 m)
POLYGALACEAE Polygala L.
Polygala compressa H.Perrier M [C] LG 5216 (1100 m); MHR 032
Polygala hildebrandtii Baill. M [Sb, W(N)] Hildebrandt, J. M. 3359a (nv)
POLYGONACEAE Polygonum L.
Polygonum minus Huds. M [C, W(S)] - As, Eur LG 5163 (1350 m)
Polygonum mite Schrank M [Sb, E, C] - Eur MHR 158 (990 m); SMT 398 (990 m)
Polygonum nepalense Meisn. M [C] - Aft, As LG 5197 (1350 m)
Polygonum sp. OA 162 [1175 m] (nv); OA 206 [900 m] (nv); SM 1719 [1045 m] (nv)
Rumex L. Rumex nepalensis Spreng. M [C] - Aft, As LG 5156 (1350 m)

8. Annexes
157
PORTULACACEAE Talinella Baill.
Talinella aff. ankaranensis Appleq. OA 999 (507 m) (nv)
Talinella pachypoda Eggli M [W(N)] JB 5928 (nv)
POTAMOGETONACEAE Potamogeton L.
Potamogeton nodosus Poir. M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt, Eur AMH 16; LN 2421 (1250 m)
PUTRANJIVACEAE Drypetes Vahl
Drypetes ambigua Leandri M [E] SF & R. Capuron 11275 [1050 m] (nv); SF & R. Capuron 20028 [1050 m] (nv); SF & R. Rabevohitra 32605 [1050 m] (nv)
Drypetes capuronii Leandri M [Sb, E, C, W] SF & N. Boanarisolo 185-R-152 (nv); SF & R. Capuron 11299 [600 m] (nv); SF & R. Capuron 11303 (600 m) (nv); SF & R. Capuron 11340
[300 m] (nv); SF & R. Capuron 20062 (nv); SF & R. Capuron 20070; SF & Randriamanalina 49-R-150 (nv); SM 1195 (670 m) (nv)
Drypetes madagascariensis (Lam.) Humbert & Leandri M [Sb, E, C, W] Harder, D. K. & al. 1635 (840 m) (nv); LB 12006 (nv); OA 860 (533 m); OA 883 (874 m) (nv); SDR 110 (990 m)
Drypetes perrieri Leandri M [Sb, E, C, W, S] - RM Randrianaivo, R. & al. 785 (399 m) (nv); SF & R. Capuron 11347 (300 m); SMT 528 (650 m)
Drypetes sp. OA 779 (874 m) (nv); SM 1743 [1045 m] (nv)
RANUNCULACEAE Clematis L.
Clematis mauritiana Lam. M [C] - RM OA 193 [900 m] (nv)
Clematis sp. OA 247 [375 m] (nv)
Ranunculus L. Ranunculus pinnatus Poir. M [E, C, S] - RM, Aft LN 2352 (1300 m)
RHAMNACEAE Bathiorhamnus Capuron
Bathiorhamnus dentatus (Capuron) Callm., Phillipson & Buerki M [Sb, E, W(N)] SF & R. Capuron 20075 (nv), TYPE: IT: TEF
Bathiorhamnus louvelii (H.Perrier) Capuron M [E, C, W] Harder, D. K. & al. 1641 (840 m) (nv); LG 5067 (920 m)
Colubrina Rich. ex Brongn. Colubrina faraloatra (H. Perrier) Capuron M [E, C] Colubrina faraloatra (H. Perrier) Capuron var. faraloatra SF & R. Rabevohitra 29388 (nv)

8. Annexes
158
Gouania Jacq. Gouania ambrensis Bürki, Callm. & Phillipson M [MdA] SMT 332 (930 m)
Gouania glandulosa Tul. M [Sb, E, C, W] JB 167 (1100 m) (nv)
Gouania laxiflora Tul. M [E, W, S] - RM LG 5251 (1100 m); MHR 030 (1021 m)
Gouania myriocarpa Tul. M [E, C] SMT 380 (1040 m)
Gouania sp. Van Nek, F. I. 1819 (1010 m) (nv); Van Nek, F. I. 1901 (700 m) (nv); Van Nek, F. I. 1904 (710 m) (nv); Van Nek, F. I. 1939 (635 m) (nv)
Lasiodiscus Hook.f. Lasiodiscus pervillei Baill. M [Sb, E, W] - RM, Aft Randrianaivo, R. & al. 790 (399 m) (nv)
RHIZOPHORACEAE Cassipourea Aubl.
Cassipourea cf. leptoclada Tul. LG 5045 (1180 m)
Cassipourea sp. 1 SDR 065 (1140 m)
Cassipourea sp. 2 SMT 033 (1250 m)
Cassipourea sp. SF & R. Rabevohitra 32608 (nv); SF & Ratsirarison 31104 [1050 m] (nv); SM 957 (1300 m)
Macarisia Thouars Macarisia lanceolata Baill. M [Sb, C, W] JB 5916 (nv); SF & Andrianandianina 30728 (nv); SF & R. Capuron 11339 [350 m] (nv)
Macarisia sp. GES 2404 (1000 m) (nv); Rakotozafy, A. & al. 2297 (nv)
ROSACEAE Alchemilla L.
Alchemilla cf. cryptantha Steud. ex A.Rich. LG 5186 (1350 m)
Rubus L. Rubus rosifolius Sm. M [E, C] - Aft, As, Oc, Amt LG 5182 (1350 m)
Rubus aff. rosifolius Sm. MHR 128 (1100 m); SDR 054 (990 m)
Rubus sp. 1 LG 5126 (1470 m); SMT 018 (1280 m)
RUBIACEAE Astiella Jovet
Astiella sp. nov. M [C] LG 5183 (1350 m)

8. Annexes
159
Bremeria Razafim. & Alejandro Bremeria sp. OA 984 (533 m) (nv)
Breonia A.Rich. Breonia capuronii Razafim. M [Sb, E, C, W] SF & R. Capuron 22059 [900 m] (nv), TYPE: HT: TEF; SM 1219 (630 m) (nv); SMT 477 (970 m)
Breonia perrieri Homolle M [W] SF 6616 (nv)
Breonia stipulata Havil. M [E, W] OA 885 (874 m) (nv)
Breonia sp. 1 SMT 456 (1000 m)
Breonia sp. SM 1788 [1045 m] (nv)
Canephora Juss. Canephora madagascariensis J.F.Gmel. M [E, C] LG 5055 (870 m)
Canthium Lam. Canthium sp. 1 LB 12033 [1225 m]; LG 5023 (1290 m); LG 5060 (840 m); LG 5229 (1220 m)
Canthium sp. 2 SM 951 (1410 m); SMT 017 (1280 m)
Canthium sp. 3 LG 5073 (820 m); MHR 159 (1000 m); SDR 025 (1180 m); SMT 057 (1015 m)
Canthium sp. 4 SMT 070 (1070 m)
Canthium sp. Miller, J. S. & al. 3623 (1050 m) (nv); Miller, J. S. & al. 3624 (1050 m) (nv); Miller, J. S. 3623 (nv); Randrianaivo, R. & al. 797 (400 m)
(nv); SF & R. Capuron 20055 (nv); SM 1786 [1045 m] (nv); SM 956 (1300 m) (nv)
Carphalea Juss. Carphalea kirondron Baill. M [Sb, C, W, S] SM 1184 (680 m) (nv); SMT 065 (1070 m)
Chassalia Comm. ex Poir. Chassalia sp. 1 LB 11946 [1025 m]; LB 12009 [900 m]; LG 5048 (1030 m); MHR 014 (975 m); MHR 060 (990 m); SDR 081 (1000 m)
Chassalia sp. 2 De Block, P. & al. 960; LB 12004 [900 m]; LG 5049 (1030 m); MHR 023 (1010 m)
Cinchona L. Cinchona calisaya Wedd. M [C] - Aft, Amt De Block, P. & al. 975; Leeuwenberg, A. J. M. 14304 (950 m) (nv)
Cinchona sp. OA 198 [900 m] (nv)
Coffea L. Coffea augagneuri Dubard M [MdA] HP 18846 (800 m) (nv), TYPE: ST: P, IST: P; Mogenet 4 (800 m) (nv), TYPE: ST

8. Annexes
160
Coffea bonnieri Dubard M [C] Mogenet 8699 (nv), TYPE: HT
Coffea dubardii Jumelle M [W] HP 18821 (800 m) (nv), TYPE: ST; HP 18834 (800 m) (nv), TYPE: ST
Coffea gallienii Dubard M [E] Mogenet s.n. (nv), TYPE: HT; SM 1252 (630 m)
Coffea jumellei J.-F.Leroy M [W(N)] AMH s.n. (nv)
Coffea mogeneti Dubard M [C, W(N)] 2258 [1175 m]; Davis, A. P. & al. 2229 (800 m); OA 209 [900 m]; SMT 534 (650 m)
Coffea pervilleana (Baill.) Drake M [Sb, W(S)] Miller, J. S. & al. 3614 (1050 m) (nv)
Coffea tetragona Jum. & H.Perrier M [Sb] SF & R. Capuron 20064 (nv)
Coffea tricalysioides J.F.Leroy M [C] LG 5218 (1250 m); SF & J. de J. F. Leroy 959A (1300 m); SM 954 (1300 m); SMT 002 (1250 m)
Coffea sp. 1 De Block, P. & al. 933; De Block, P. & al. 942
Coffea sp. 2 SMT 244 (1060 m); SMT 352 (975 m)
Coffea sp. Cremers, G. 2580 (80 m) (nv); GES 1529 (1000 m) (nv); OA 260 [375 m] (nv); Rakotozafy, A. 1484 (nv); SF & R. Capuron 29188bis (nv);
SM 1739 [1045 m] (nv); SM 1787 [1045 m] (nv)
Danais Comm. ex Vent. Danais cernua Baker M [Sb, E, C] SMT 177 (1125 m); SMT 439 (985 m)
Danais fragrans (Comm. ex Lam.) Pers. M [Sb, E, C] - RM Phillipson, P. B. 1967 (900 m) (nv)
Danais microcarpa Baker M [C] LG 5109 (1470 m)
Danais sp. 1 SMT 325 (955 m)
Danais sp. SM 1248 (640 m) (nv)
Enterospermum Hiern Enterospermum sp. 1 SDR 042 (1170 m); SMT 319 (970 m); SMT 467 (1000 m)
Enterospermum sp. 2 SM 1258 (630 m) (nv)
Enterospermum sp. 3 SDR 023 (1020 m)
Enterospermum sp. OA 223 [375 m] (nv); OA 359 [375 m] (nv)
Euclinia Salisb. Euclinia suavissima (Homolle ex Cavaco) J.-F.Leroy M [W, S] OA 895 (533 m) (nv)

8. Annexes
161
Gaertnera Lam. Gaertnera arenaria Baker M [Sb, E, C] De Block, P. & al. 951; Du Puy, D. & al. MB221 (1300 m) (nv); LG 5054 (870 m); MHR 137 (970 m); Miller, J. S. & al. 3632 (1050 m)
(nv); OA 238 [375 m]; OA 988 (809 m) (nv); Razafimandimbison, S. G. 104 [1050 m] (nv); SDR 072; SM 1799 [1045 m] (nv); SM 930
(700 m); SMT 444 (830 m)
Gaertnera sphaerocarpa Baker M [E, C] Harder, D. K. & al. 1565 (840 m) (nv)
Gaertnera sp. Phillipson, P. B. 1966 (900 m) (nv); SF & J. Belin 13238 [1050 m] (nv); SF & J. Belin 13288 [1050 m] (nv); SF & Rakotosihanaka 30736
[1050 m] (nv); SM 1275 (710 m) (nv); SM 1793 [1045 m] (nv)
Gyrostipula J.-F.Leroy Gyrostipula foveolata (Capuron) J.-F.Leroy M [C] Razafimandimbison, S. G. 270A; Razafimandimbison, S. G. 271 (nv); SDR 029 (1060 m); SF & R. Capuron 11284 (nv)
Hyperacanthus E.Mey. ex Bridson Hyperacanthus sp. SMT 529 (650 m)
cf. Hyperacanthus sp. Harder, D. K. & al. 1605 (450 m) (nv); OA 272 [375 m] (nv)
Ixora L. Ixora amplidentata De Block M [C] OA 292 (1475 m) (nv)
Ixora cremixora Drake M [E, W] - RM Harder, D. K. & al. 1615 (660 m) (nv)
Ixora cf. drakei Hochr. Harder, D. K. & al. 1638 (840 m) (nv)
Ixora sp. 1 Labat, J.-N. & al. 2868 (900 m); LG 5063 (840 m); LG 5174 (1200 m); MHR 029 (1026 m); MHR 164 (900 m); SMT 462 (1020 m); SMT
499 (800 m)
Ixora sp. 2 SMT 334 (910 m); SMT 336 (910 m)
Ixora sp. Davis, A. P. & al. 2222 (nv); Gentry, A. 11949 (1200 m) (nv); Gentry, A. 11958 (1200 m) (nv); SF & Andrianandianina 30732 (nv); SM
1204 (670 m) (nv); SM 1783 [1045 m] (nv)
Lemyrea A.Chev. & Beille Lemyrea sp. 1 LG 5053 (870 m); SMT 013 (660 m)
Morinda L. Morinda sp. SM 1273 (640 m) (nv)
Mussaenda L. Mussaenda arcuata Lam. ex Poir. M [Sb, E, C] - RM, Aft OA 242 [375 m] (nv); OA 774 (874 m) (nv); Phillipson, P. B. 1976 (900 m) (nv); SM 1247 (640 m) (nv); SMT 489 (925 m)
Neoleroya Cavaco Neoleroya verdcourtii Cavaco M [W(N)] OA 947 (662 m) (nv); SM 1243 (630 m)

8. Annexes
162
Oldenlandia L. Oldenlandia goreensis (DC.) Summerh. M [E, C] - RM, Aft LN 2456 (1248 m)
Oldenlandia lancifolia (Schumach.) DC. M [Sb, E, C] - RM, Aft, Amt Oldenlandia lancifolia var. scabridula Bremek. LN 2450 (1248 m)
Oldenlandia tsaratananensis Homolle M [C] HH 32085 (1000 m) (nv); HP 9403 (1000 m) (nv)
Oldenlandia sp. HP 17717 (1200 m) (nv); OA 48 [1438 m] (nv); SM 1749 [1045 m] (nv)
Pauridiantha Hook.f. Pauridiantha paucinervis (Hiern) Bremek. M [C] - Aft MHR 058 (1120 m); SM 1742 [1045 m] (nv); SMT 303 (1060 m)
Pauridiantha paucinervis (Hiern) Bremek. subsp. lyallii (Baker) Verdc. GES 1513 (1000 m) (nv); Harder, D. K. & al. 1595 (965 m) (nv); LB 11986 [1150 m]; LG 5107 (1470 m); OA 109 [800 m]; OA 232 [375
m]; OA 780 (874 m) (nv); SF 7216; SM 2352 [1438 m] (nv)
Peponidium (Baill.) Arènes Peponidium cf. humbertii Homolle ex Arènes SDR 068 (1110 m); SMT 062 (1075 m)
Peponidium sp. 1 OA 945 (662 m)
Peponidium sp. 2 LB 11973 (1000 m); SMT 106 (1082 m)
Peponidium cf. sp. 2 LG 5075 (670 m)
Peponidium velutinum Arènes M [Sb] Harder, D. K. & al. 1645 (840 m) (nv)
Phyllopentas (Verdc.) Karehed & K.Bremer Phyllopentas madagascariensis (Verdc.) Kårehed & B. Bremer M [C] MHR 040 (1162 m); MHR 136 (970 m); Phillipson, P. B. 1965 (900 m) (nv); SM 1231 (610 m) (nv); SMT 231 (1005 m); SMT 434 (885 m)
Phyllopentas aff. madagascariensis (Verdc.) Karehed & K.Bremer LG 5173 (1200 m)
Phyllopentas sp. Du Puy, D. & al. MB207 (1325 m) (nv); OA 73 [950 m] (nv); SM 1712 [1045 m] (nv)
Plectronia L. Plectronia micrantha Baker M [C] Harder, D. K. & al. 1593 (965 m) (nv)
Plectronia sp. Gentry, A. 11954 (1200 m) (nv)
Psychotria L. Psychotria aff. boivinii Bremek. Harder, D. K. & al. 1647 (840 m) (nv)
Psychotria expansissima K.Schum. M [Sb] SF & R. Capuron 22060 [900 m] (nv); SF & R. Capuron 24060 [900 m] (nv)

8. Annexes
163
Psychotria glaucifolia A.P.Davis & Govaerts M [C] De Block, P. & al. 967; De Block, P. & al. 974; De Block, P. & al. 980; LB 11982 [1150 m]; MHR 160 (920 m); SM 966 (1300 m); SMT
074 (1150 m); SMT 475 (975 m)
Psychotria homolleae Bremek. M [C] AMH 56 [1100 m] (nv), TYPE: HT
Psychotria aff. manambolensis A.P.Davis & Govaerts Harder, D. K. & al. 1599 (450 m) (nv)
Psychotria sambiranensis Bremek. Echantillon rattaché avec doute: Harder, D. K. & al. 1571 (890 m) (nv)
Psychotria tsiandrensis Bremek. M [E, W(S)] LB 11952bis (1000 m); LB 12022 [900 m]; OA 216 [900 m] (nv); Razafimandimbison, S. G. 106 [1050 m]; SMT 051 (985 m); SMT 416
(960 m)
Psychotria sp. 1 De Block, P. & al. 952; LB 11963 (1000 m); SMT 049 (985 m); SMT 509 (965 m)
Psychotria sp. 2 De Block, P. & al. 977; LN 2386 (1260 m); MHR 019 (1119 m); MHR 133 (1060 m); SDR 088 (990 m); SMT 301 (1060 m)
Psychotria sp. 3 2346 (1200 m); LB 11943 (1025 m); LG 5024 (950 m); MHR 008 (1190 m); MHR 025; SMT 031 (1010 m); SMT 073 (1025 m)
Psychotria sp. 4 De Block, P. & al. 948; MHR 146 (1020 m); SMT 086 (1065 m); SMT 254 (990 m)
Psychotria sp. 5 De Block, P. & al. 934; MHR 063 (1050 m); MHR 102 (980 m); MHR 109 (1020 m); Razafimandimbison, S. G. 101 [1050 m]; SDR 101
(1120 m); SDR 140 (1030 m)
Psychotria sp. 6 MHR 148 (1030 m)
Psychotria sp. 7 SMT 522 (650 m)
Psychotria sp. Gentry, A. 11956 (1200 m) (nv); GES 1533 (1000 m) (nv); OA 224 [375 m] (nv); OA 294 (1475 m) (nv); OA 782 (874 m) (nv); OA 852
(533 m) (nv); PM 1422 (nv); Razafimandimbison, S. G. 112 [1050 m] (nv)
cf. Psychotria sp. GMP 14506 [1150 m] (nv)
Pyrostria Comm. ex Juss. Pyrostria homolleae Arènes & Cavaco M [C] SF & R. Capuron 20077 (nv)
Pyrostria sp. 1 LG 5239 (1100 m); SM 1187 (700 m); SMT 071 (1070 m)
Pyrostria sp. 2 SM 1220 (610 m)
Randia L. Randia sp. SM 1203 (630 m) (nv)
Rothmannia Thunb. Rothmannia sp. Randrianaivo, R. & al. 787 (399 m) (nv); SF & R. Capuron 22062 (nv); SF & R. Capuron 24062 (nv)

8. Annexes
164
Rytigynia Blume Rytigynia sp. 1 SMT 063 (1021 m)
Sabicea Aubl. Sabicea sp. SM 1254 (640 m) (nv)
Saldinia A.Rich. ex DC. Saldinia aff. stenophylla Bremek. Harder, D. K. & al. 1597 (965 m) (nv)
Saldinia subacuminata Bremek. M [C] Saldinia subacuminata Bremek. var. subacuminata De Block, P. & al. 962; De Block, P. & al. 981; LB 11962 (1000 m); LG 5010 (1310 m); LG 5220 (1260 m); MHR 046 (1050 m);
Razafimandimbison, S. G. 469 (nv); Razafimandimbison, S. G. 472 (nv); SDR 111 (970 m); SDR 130 (1000 m); SMT 006 (1040 m); SMT
026 (1300 m)
Saldinia sp. Gentry, A. 11953 (1200 m) (nv); Gentry, A. 11957 (1200 m) (nv); Gentry, A. 11959 (1200 m) (nv); OA 110 [800 m] (nv)
Schizenterospermum Homolle ex Arènes Schizenterospermum cf. grevei Homolle ex Arènes Harder, D. K. & al. 1636 (840 m) (nv)
Spermacoce L. Spermacoce tenuior L. M [Sb, E, C] - Amt LG 5152 (1350 m); MHR 157 (990 m); SMT 400 (990 m)
Tarenna Gaertn. Tarenna capuroniana De Block M [C] LG 5175 (1140 m); MHR 078 (1100 m); MHR 139 (910 m); OA 256 [375 m]; SF & R. Capuron 24477 (900 m); SM 1180 (650 m) (nv),
TYPE; SM 933 (700 m); SMT 093 (1041 m); SMT 257 (1020 m); SMT 310 (1075 m)
Tarenna spiranthera (Drake) Homolle M [Sb, E, C] LG 5026 (950 m); MHR 153 (1010 m); MHR 156 (1000 m); SF & R. Capuron 22055 [900 m]; SF & R. Rabevohitra 32636 [1050 m] (nv);
SMT 538 (650 m). Echantillon rattaché avec doute : Harder, D. K. & al. 1572 (890 m) (nv)
Tarenna aff. spiranthera (Drake) Homolle OA 291 (1475 m) (nv)
Tarenna sp. 1 LG 5233 (1210 m); MHR 020 (1280 m); MHR 028 (1060 m); SDR 037 (1060 m); SMT 078 (1150 m); SMT 453 (1000 m)
Tarenna sp. GES 1522 (1000 m) (nv); GMP 14509 (1000 m) (nv); OA 112 [800 m] (nv); OA 211 [900 m] (nv); OA 92 [950 m] (nv);
Razafimandimbison, S. G. 103 [1050 m] (nv); Seigler, D. S. 12896 (nv); SF & R. Capuron 24478 (900 m) (nv)
Triainolepis Hook.f. Triainolepis sp. SF & Abraham 31597 (nv)
Tricalysia A.Rich. ex DC. Tricalysia ambrensis Randriamb. & De Block M [C] Cours, G. & al. 5373 (nv); Cours, G. & al. 5374 (nv); Cours, G. & al. 5375 (nv); Cours, G. & al. 5376 (nv); De Block, P. & al. 1313 (nv); De
Block, P. & al. 976 (nv); De Block, P. & al. 978 (nv); De Block, P. & al. 979 (nv); HH 32088 (nv); HH 32102 (nv); HH 32119 (1000 m)
(nv), TYPE: HT; HH 32132 (nv); HH 32133 (nv); HH 32135 (nv); HH 32137 (nv); OA 102 (nv); OA 304 (1475 m) (nv);
Razafimandimbison, S. G. 118 [1150 m] (nv); SF 7195 (nv); SF 9957 (nv)

8. Annexes
165
Trigonopyren Bremek. Trigonopyren aff. sambiranensis Bremek. Harder, D. K. & al. 1566 (840 m) (nv)
Vangueria Juss. Vangueria madagascariensis J.F.Gmel. M [Sb, E, W(N)] - RM, Aft, Amt Harder, D. K. & al. 1770 (450 m) (nv)
Vangueria sp. SM 1225 (600 m) (nv)
RUBIACEAE indéterminées LB 11941 [1025 m] (nv); LB 11945 [1025 m] (nv); Miller, J. S. & al. 10684 (1050 m) (nv); SF & R. Capuron 22061 [900 m] (nv)
RUTACEAE Citrus L.
Citrus aurantium L. M [Sb, E, C, W] - RM, As, Oc, Amt OA 355 [375 m] (nv); SDR 122 (965 m)
Citrus aurantium L. var. amara L. HP 17696 (500 m) (nv)
Melicope J.R.Forst. & G.Forst. Melicope aff. belahe (Baill.) T.G.Hartley OA 252 [375 m] (nv)
Toddalia Juss. Toddalia asiatica (L.) Lam. M [Sb, E, C, W] - RM, Aft, As, Oc OA 278 [375 m]; OA 86 [950 m]; SDR 033 (1162 m); SMT 503 (765 m)
Vepris Comm. ex A.Juss. Vepris cauliflora H.Perrier M [E, C] LG 5212 (1100 m); LG 5235 (1100 m); MHR 062 (1045 m); SF & R. Capuron 20074 (nv); SMT 076 (1150 m)
Vepris dicarpella H.Perrier M [C, HM] Harder, D. K. & al. 1637 (840 m) (nv)
Vepris aff. leandriana H.Perrier Harder, D. K. & al. 1639 (840 m) (nv)
Vepris madagascariensis (H.Perrier) Mziray M [E, W(N)] LG 5074 (670 m); SM 1256 (640 m) (nv); SM 1269 (630 m)
Vepris nitida (Baker) I.Verd. M [Sb, E, C] SDR 135 (630 m); SMT 515 (750 m)
Vepris punctata (I.Verd.) Mziray M [Sb, C, W(N)] HP 18825 (800 m) (nv); LG 5065 (870 m); SF 10281 (450 m) (nv)
Vepris sp. OA 75 [950 m] (nv); OA 77 [950 m] (nv)
SALICACEAE Bivinia Jaub. ex Tul.
Bivinia jalbertii Tul. M [Sb, C, W, S] - Aft Harder, D. K. & al. 1608 (450 m) (nv)
Calantica Jaub. ex Tul. Calantica biseriata H.Perrier M [W(S)] Harder, D. K. & al. 1624 (660 m) (nv)

8. Annexes
166
Calantica cerasifolia (Vent.) Tul. M [Sb, E, C, W] SF & R. Capuron 11281 (nv)
Casearia Jacq. Casearia nigrescens Tul. M [E, C] Casearia nigrescens Tul. var. lucida (Tul.) Sleumer SDR 039 (1041 m)
Homalium Jacq. Homalium micranthum (Boivin ex Tul.) O.Hoffm. M [Sb, E, C, W(N)] LB 11948 (1000 m); SF & R. Capuron 20039
Homalium nudiflorum (DC.) Baill. M [E, C] GMP 14484 [1050 m] (nv); HP 2307 (nv); SF & J. de D. Rakoto 11013 [1050 m] (nv); SF & R. Capuron 687 [1050 m] (nv); Ursch, E. 239
(nv)
Homalium nudiflorum (DC.) Baill. var. ciliolatum Sleumer SF & J. Belin 13239 [1050 m] (nv)
Homalium nudiflorum (DC.) Baill. var. nudiflorum LB 12003 [900 m]; LG 5020 (1320 m); SDR 009 (1300 m); SMT 485 (1000 m)
Homalium sp. OA 857 (533 m) (nv); SM 1805 [1045 m] (nv)
Ludia Comm. ex Juss. Ludia ambrensis H.Perrier M [E, C] - RM, Aft HP 18889 (1000 m) (nv), TYPE: LT; SF & R. Capuron 20035 (nv)
Ludia boinensis H.Perrier M [E, W] SF & E. Ngoloka 12750 (10 m) (nv); SF & J. Belin 6619 (nv); SF & J. Belin 6639 (nv); SF & R. Capuron 20061 (nv)
Ludia cf. boinensis H.Perrier LG 5088 (1030 m)
Ludia erosifolia Sleumer M [W(N)] SDR 024 (1020 m); SF & R. Capuron 20067 (nv), TYPE: HT: P; IT: TEF, P
Ludia aff. leandriana Sleumer vel sp. aff. SM 1224 (600 m)
Ludia sp. OA 85 [950 m] (nv)
Scolopia Schreb. Scolopia madagascariensis Sleumer M [E, C] SF & R. Capuron 29188 [700 m] (nv)
Tisonia Baill. Tisonia crenata Sleumer M [Sb, W] HH 32113 (nv); HH 32114 (nv)
SANTALACEAE Staufferia Z.S.Rogers, Nickrent & Malécot
Staufferia capuronii Z.S. Rogers & Nickrent & Malécot M [E, C, W(N)] LG 5244 (1100 m); SMT 069
Viscum L. Viscum cf. lophiocladum Baker SMT 473 (970 m)

8. Annexes
167
Viscum trachycarpum Baker M [Sb, C, HM, W, S] - RM Viscum trachycarpum var. douliotii (Lecomte) Balle LG 5238 (1100 m); SMT 068
Viscum aff. trachycarpum Baker LG 5236 (1100 m)
Viscum sp. OA 855 (533 m) (nv)
SAPINDACEAE Allophylus L.
Allophylus arboreus Choux M [Sb, E, C] Harder, D. K. & al. 1560 (840 m) (nv); SMT 435 (885 m)
Allophylus boinensis Choux M [W(S)] Harder, D. K. & al. 1628 (660 m) (nv); SM 2364 [1438 m] (nv)
Allophylus cobbe Raeusch. M [E, C, W] - RM, Aft, As Allophylus cobbe var. "antankarana" Capuron SF & R. Capuron 20069 (nv); SM 1228 (610 m)
Allophylus cobbe var. "boinensis" (Choux) Capuron SF & R. Rabevohitra 32604 [1050 m] (nv)
Allophylus macrocarpus Danguy & Choux M [C] Harder, D. K. & al. 1560A (840 m) (nv); LG 5039 (1180 m); PM 1419 [1150 m] (nv); PM 1424 (nv); PM 1924 (nv); Schedl, K. E. 201 (1150
m) (nv); SDR 022 (1050 m); SM 1242 (610 m)
Allophylus aff. macrocarpus Danguy & Choux SMT 045 (1060 m)
Allophylus macrocarpus Languy & Choux var. ambrensis Capuron SF 7211 (nv), TYPE
Allophylus trichodesmus Radlk. M [E, C] GMP 14498 [1150 m] (nv); Harder, D. K. & al. 1644 (840 m) (nv); SM 1230 (610 m) (nv)
Allophylus sp. Miller, J. S. & al. 10685 (1050 m) (nv); OA 139 [1175 m] (nv); OA 785 (874 m) (nv); SF & R. Capuron 685 [1050 m] (nv)
Camptolepis Radlk. Camptolepis grandiflora Capuron M [E] SF & R. Capuron 11294 (600 m) (nv); SF & R. Capuron 11348 [25 m] (nv), TYPE: HT: P; IT: TEF; SMT 497 (840 m)
Chouxia Capuron Chouxia borealis G.E.Schatz, Gereau & Lowry M [W(N)] SF & R. Capuron s.n. (nv)
Deinbollia Schumach. Deinbollia neglecta Radlk. M [E] - RM SF & R. Capuron 11291 (650 m) (nv); SF & R. Capuron 20010 (nv)
Deinbollia sp. Leeuwenberg, A. J. M. & al. 14300 (950 m) (nv); Van Nek, F. I. 1860 (780 m) (nv)
Doratoxylon Thouars ex Hook.f. Doratoxylon stipulatum Capuron M [E] PM 1405 [1100 m] (nv); PR 1169 (1249 m)
Filicium Thwaites ex Benth. & Hook.f. Filicium longifolium (H.Perrier) Capuron M [W, S] - RM SF & R. Capuron 20074bis (nv)

8. Annexes
168
Glenniea Hook.f. Glenniea pervillei (Baill.) Leenh. M [Sb, E, W(N)] Randrianaivo, R. & al. 791 (399 m) (nv)
Harpullia Roxb. cf. Harpullia sp. Miller, J. S. & al. 3626 (1050 m) (nv)
Lepisanthes Blume Lepisanthes perrieri (Choux) Buerki, Callm. & Lowry M [W] SF & R. Capuron 11423 [300 m] (nv)
Lepisanthes senegalensis (Juss. ex Poir.) Leenh. M [Sb] - Aft, As OA 896 (533 m) (nv)
Macphersonia Blume Macphersonia gracilis O.Hoffm. M [Sb, E, W] - RM Macphersonia gracilis O.Hoffm. var. gracilis SF & R. Capuron 11355 [350 m] (nv); Van Nek, F. I. 1955 (540 m) (nv)
Macphersonia gracilis O.Hoffm. var. hildebrandtii (O.Hoffm.) Capuron SM 1222 (610 m)
Macphersonia madagascariensis Blume M [Sb, E, C] Schedl, K. E. 194 (1150 m) (nv)
Neotina Capuron Neotina isoneura (Radlk.) Capuron M [E, C] LB 11959 bis (1000 m); SF & R. Capuron 22058 (nv); SF & R. Capuron 24058 (nv)
Plagioscyphus Radlk. Plagioscyphus aff. calciphilus Capuron LB 12025 [900 m]
Plagioscyphus louvelii Danguy & Choux M [Sb, E, C, W(N)] SM 1215 (630 m)
Plagioscyphus sp. Schedl, K. E. 229 (600 m) (nv); SF & R. Capuron 20012 (nv)
Stadmania Lam. Stadmania acuminata Capuron M [E] Schedl, K. E. 216 (1080 m) (nv)
Stadmania acuminata var. ambrensis Capuron LG 5069 (1030 m); SF & R. Capuron 20019 [1050 m] (nv), TYPE
Tina Schult. Tina chapelieriana (Cambess.) Kalkman M [E, C] SF & Andrianandianina 30727 (nv); SF 14211 (900 m) (nv); SMT 517 (740 m)
Tina striata Radlk. M [E, C] LG 5016 (1320 m); Rakotondrafara, A. & al. 279 (717 m) (nv); SDR 014 (1300 m); SDR 073
Tina striata subsp. multifoliolata Capuron LB 11958 (1000 m); Leeuwenberg, A. J. M. & al. 14294 (900 m) (nv); SF & J. de D. Rakoto 11011 [1050 m] (nv); SF & N. Boanarisolo
13489 (nv); SF & N. Boanarisolo 8127 [1050 m] (nv); SF & R. Capuron 20037 (nv); SM 1800 [1045 m]
Tina sp. SF & R. Capuron 11286 (nv)

8. Annexes
169
SAPOTACEAE Chrysophyllum L.
Chrysophyllum ambrense (Aubrév.) G.E.Schatz & L.Gaut. M [MdA] LB 12008 [900 m]; Leeuwenberg, A. J. M. & al. 14314 (1000 m); SF & R. Capuron 11277 (1000 m) (nv), TYPE: IT: P, HT: P; SF 14877
(1050 m) (nv)
Chrysophyllum boivinianum (Pierre) Baehni M [Sb, E, C] - RM HH 32060 (1000 m) (nv); LG 5089 (1030 m); LG 5206 (1260 m); Razafimandimbison, S. G. 115 [1050 m] (nv); SF & J. Ch. Rabearivo
7371 [1050 m] (nv); SF & R. Capuron 11274 (1000 m) (nv); SF 7372 [1050 m] (nv); SMT 490 (1040 m)
Chrysophyllum cf. lanceolatum (Blume) A.DC. SMT 364 (1025 m)
Chrysophyllum perrieri (Lecomte) G.E.Schatz & L.Gaut. M [Sb, E, W] LG 5082 (770 m)
Faucherea Lecomte Faucherea ambrensis Capuron ex Aubrév. M [E, C] LG 5007 (1310 m); SMT 349 (1025 m)
Faucherea cf. ambrensis Capuron ex Aubrév. SDR 116 (970 m)
Faucherea ambrensis Capuron ex Aubrév. var. ambrensis HP 18867 (1000 m) (nv), TYPE: HT: P; IT: P; Ursch, E. 208 (nv)
Faucherea sp. Schedl, K. E. 166 (1100 m) (nv); SF & R. Capuron 20043 (nv)
Labramia A.DC. Labramia ankaranaensis Aubrév. M [W(N)] Labramia ankaranaensis Aubrév. var. antsingensis Aubrév. SF 6617 (nv)
Labramia platanoides Capuron ex Aubrév. M [E, W(N)] LG 5211 (1100 m); MHR 163 (900 m); MHR 165 (1030 m); SF & R. Capuron 11350 [275 m] (nv), TYPE: HT: P; IT: P; SMT 451 (1040 m)
Labramia sp. SF & R. Capuron 29193 (nv)
Mimusops L. Mimusops capuronii Aubrév. M [Sb, E, W] Mimusops capuronii Aubrév. var. retusa Aubrév. SF & R. Capuron 11297 (600 m) (nv)
Mimusops sambiranensis Aubrév. M [W(N)] SF & R. Capuron 11358 [275 m] (nv)
Mimusops voalela Aubrév. M [E] SF 169-R-6 (nv)
Sideroxylon L. Sideroxylon beguei Capuron ex Aubrév. M [E] Sideroxylon beguei Capuron ex Aubrév. var. beguei LB 12024 [900 m]
Sideroxylon gerrardianum (Hook.f.) Aubrév. M [E] OA 856 (533 m) (nv); SF & R. Capuron 11293 (nv); SMT 514 (750 m)

8. Annexes
170
SCROPHULARIACEAE Buddleja L.
Buddleja acuminata Poir. M [Sb, E, C, W] - Aft Du Puy, B. MB210 (1325 m) (nv)
Buddleja axillaris Willd. ex Schult. M [Sb, E, C] - RM, Aft Du Puy, D. & al. 209 (1325 m) (nv); JB 5631 (nv); JB 5995 (nv); Leeuwenberg, A. J. M. & al. 14234 (1000 m) (nv); LG 5228 (1220 m); PM
1177 (1000 m) (nv); PM 5995 (nv); SM 947 (1360 m)
Buddleja indica Lam. M [Sb, E, C, W] - RM, ? au Sb AMH 39 [1100 m] (nv); LG 5241 (1100 m); MHR 077 (1100 m); SDR 027 (1070 m); SDR 031 (1106 m); SM 1792 [1045 m] (nv); SM 953
(1300 m); SMT 370 (1010 m); Van Nek, F. I. 1800 (1110 m) (nv); Van Nek, F. I. 1803 (1150 m) (nv)
Buddleja sp. Du Puy, B. & al. MB209 (1325 m) (nv); OA 158 [1175 m] (nv); Rakotozafy, A. 2296 (nv)
SMILACACEAE Smilax L.
Smilax anceps Willd. M [Sb, E, C, W] - RM, Aft MHR 112 (1030 m); Phillipson, P. B. 1958 (900 m) (nv); SMT 012 (1050 m)
Smilax sp. GES 1511 (1000 m) (nv); OA 78 [950 m] (nv)
SOLANACEAE Datura L.
Datura metel L. M [W(S)] - As, Amt SMT 423 (660 m)
Solanum L. Solanum americanum Mill. M [C] - RM, Aft, As, Oc, Amt, Eur SM 1716 [1045 m] (nv); LG 5198 (1350 m)
Solanum mauritianum Scop. M [Sb, E, C] - RM, Aft, As, Oc, Amt SMT 449 (1025 m)
Datura cf. suaveolens Humb. & Bonpl. ex Willd. Baum, D. & al. 342 (650 m) (nv)
Solanum torvum Sw. M [Sb, E] - RM, Aft, As, Oc, Amt HH 32124 (1000 m) (nv); HH 32125 (1000 m) (nv); HH 32153 (1000 m) (nv); JB 5632 (nv); PM 1257 (nv); PM 1393 (nv); SM 1722 [1045
m]; SMT 225 (1090 m); SMT 292 (1065 m)
Solanum sp. 1 SMT 227 (1090 m)
SPHAEROSEPALACEAE Rhopalocarpus Bojer
Rhopalocarpus alternifolius (Baker) Capuron M [Sb, E, W(N)] SF & R. Capuron 11346 [300 m]; SF & R-152 205-R-152 (nv); SF 10667 (nv); SF 205 (nv)
STEMONURACEAE Grisollea Baill.
Grisollea myriantha Baill. M [Sb, E, C] - RM SDR 035 (1030 m); SF & N. Boanarisolo 8133 (500 m) (nv); SF & R. Capuron 11356 [300 m] (nv); SF & R. Capuron 20026 [1100 m] (nv);
SF 14875 (nv)

8. Annexes
171
STILBACEAE Nuxia Comm. ex Lam.
Nuxia ambrensis Jovet M [C] HP 17695 (500 m) (nv), TYPE: HT: P; IT: P; SF & J. Ch. Rabearivo 7199 (nv); SF & R. Capuron 683 [1050 m] (nv); SF & R. Rabevohitra
32618 (nv); SMT 523 (655 m)
Nuxia coriacea Soler. M [C] GMP 14499 [1150 m] (nv); HH 32111 (1000 m) (nv); LB 11966 (1000 m); Leeuwenberg, A. J. M. & al. 14240 (1100 m) (nv); LG 5158
(1350 m); SDR 002 (1340 m); SMT 037 (1270 m)
Nuxia involucrata DC. M [Sb, E, C] LG 5223 (1350 m); MHR 042 (1200 m); PM 1420 (1300 m) (nv); PM 1425 (1000 m) (nv); SDR 010 (1300 m); SF & J. de D. Rakoto 11007
[1050 m] (nv); SF & R. Capuron 11285 (1000 m) (nv); SF & R. Capuron 29196 (1300 m) (nv); SMT 211 (1000 m)
Nuxia pachyphylla Baker M [Sb, E, C, W(N)] OA 990 (1105 m) (nv); Schedl, K. E. 188 (1150 m) (nv)
Nuxia sp. HIRSM 188 (1150 m) (nv); Miller, J. S. & al. 10675 (1050 m) (nv); SF & R. Capuron 29197 (nv)
THYMELAEACEAE Gnidia L.
Gnidia daphnifolia L.f. M [E, C, W] Rogers, Z. S. & al. 147 (960 m) (nv); Rogers, Z. S. 148 (960 m) (nv)
Lasiosiphon Fresen. Lasiosiphon madagascariensis (Lam.) Decne. M [E, C, W] AMH 167 (nv); Harder, D. K. & al. 1607 (450 m) (nv); Hildebrandt, J. M. 3369 (nv), TYPE: ST; Van Nek, F. I. 1942 (615 m) (nv)
Lasiosiphon madagascariensis var. baronii (Baker) Leandri JB 5929 (nv)
Stephanodaphne Baill. Stephanodaphne geminata H.Perrier ex Leandri M [Sb, C] Allorge, L. & al. 2719 (nv); Cours, G. & al. 5377 (1100 m) (nv); Harder, D. K. & al. 1630 (840 m) (nv); HP 17557 (900 m) (nv), TYPE:
HT:P, IT:2xP; OA 71 [950 m] (nv); Razafimandimbison, S. G. 105 [1050 m] (nv); Razafimandimbison, S. G. 110 [1050 m] (nv); Rogers, Z.
S. & al. 137 (938 m) (nv); Rogers, Z. S. & al. 138 (938 m) (nv); Rogers, Z. S. & al. 139 (938 m) (nv); Rogers, Z. S. & al. 140 (938 m) (nv);
Rogers, Z. S. & al. 141 (938 m) (nv); Rogers, Z. S. & al. 142 (938 m) (nv); Rogers, Z. S. & al. 143 (938 m) (nv); Rogers, Z. S. & al. 144
(938 m) (nv); Rogers, Z. S. & al. 145 (938 m) (nv); Rogers, Z. S. & al. 146 (938 m) (nv); SF 9963 (nv); SF & L. Bégué 9963 (nv); SF & R.
Capuron 22054 [900 m]; SMT 052 (985 m); SMT 500 (800 m)
Stephanodaphne cf. geminata H.Perrier ex Leandri HH 32022 (800 m) (nv); HH 32023 (800 m) (nv)
URTICACEAE Boehmeria Jacq.
Boehmeria tsaratananensis Leandri M [E, C] AMH 91 [1100 m] (nv); GES 2402 (1000 m) (nv); HP 17678 (1200 m) (nv), TYPE: ST: P; LN 2350 (1350 m); MHR 001 (1250 m); OA 343
(1475 m) (nv)
Boehmeria sp. Rakotozafy, A. 2294bis (nv)
Droguetia Gaudich. Droguetia cf. leptostachys (Juss. ex Pers.) Wedd. LG 5202 (1260 m)

8. Annexes
172
Elatostema J.R.Forst. & G.Forst. Elatostema goudotianum Wedd. M [E, C] HP 10008 (800 m) (nv); Keraudren, M. & al. 25562 (nv); LG 5114 (1420 m); SM 1723 [1045 m]; SMT 130 (1080 m)
Elatostema goudotianum var. weddellii Leandri Rakotozafy, A. 1487 (nv)
Laportea Gaudich. Laportea humblotii (Baill.) Friis M [C] GES 1510 (1000 m) (nv); LN 2349 (1150 m)
Laportea cf. humblotii (Baill.) Friis OA 321 (1475 m) (nv)
Phenax Wedd. Phenax perrieri Leandri M [Sb, C] HH 32024 (800 m) (nv)
Pilea Lindl. Pilea boiviniana Leandri M [Sb, E, C] HH 3978 (1100 m) (nv)
Pilea hypnophila Baker M [C] - Aft HP 10009 (1000 m) (nv); HP 17677 (1200 m) (nv)
Pilea aff. ivohibeensis Leandri LG 5112 (1470 m)
Pilea macropoda Baker M [C] - RM HH 32064 (1000 m) (nv); LN 2498 (1160 m)
Pilea rivularis Wedd. M [C] - RM OA 40 [1438 m] (nv); SMT 268 (1070 m)
Pilea tetraphylla (Steud.) Blume M [C] - Aft LG 5222 (1250 m)
Pilea tetraphylla var. hypnophila (Baker) Leandri Keraudren, M. & al. 25584 (nv)
Pilea sp. 1 LB 12017 [900 m]
Pilea sp. SM 1725 [1045 m] (nv); SM 1759 [1045 m] (nv); SM 2383 [1438 m] (nv)
Procris Comm. ex Juss. Procris crenata C.B.Rob. M [C] - Aft, As SMT 539 (980 m)
Procris pedunculata (J.R.Forst. & G.Forst.) Wedd. M [Sb, E, C] - RM, As, Oc SMT 040 (1270 m)
Urera Gaudich. Urera acuminata (Poir.) Gaudich. ex Decne. M [Sb, E, C] - RM GMP 14476 [1050 m] (nv); SM 979 (700 m)
Urera sp. SM 1770 [1045 m] (nv)
VERBENACEAE Duranta L.
Duranta erecta L. M [C] - RM, Aft, As, Oc, Amt, Eur OA 192 [900 m]

8. Annexes
173
Lantana L. Lantana camara L. M [E, C] - RM, Aft, As, Oc, Amt, Eur SMT 234
VIOLACEAE Rinorea Aubl.
Rinorea angustifolia (Thouars) Baill. M [Sb, E, C, W] - Aft LG 5056 (870 m); LG 5077 (670 m); SF & R. Capuron 20047 (nv)
Rinorea arborea (Thouars) Baill. M [Sb, E, W] - RM, Aft SMT 492 (875 m)
Rinorea pauciflora (Thouars) Baill. M [Sb, E, C, W(S)] SMT 491 (840 m)
Rinorea pugionifera (Oudem.) H.Perrier M [Sb, W(N)] SF 10671 (nv)
Rinorea squamosa (Boivin ex Tul.) Baill. M [Sb, E, W] - RM LG 5028 (900 m)
Rinorea aff. subintegrifolia (P.Beauv.) Kuntze Harder, D. K. & al. 1640 (840 m) (nv)
Rinorea sp. OA 237 [375 m] (nv)
Viola L. Viola abyssinica Steud. ex Oliv. M [C] - Aft LG 5184 (1350 m)
VITACEAE Cayratia Juss.
Cayratia imerinensis (Baker) Desc. M [Sb, E, C, W] - RM SDR 103 (1100 m); SM 985 (700 m); SMT 313 (1065 m)
Cayratia sp. SM 2360 [1438 m] (nv)
Cissus L. Cissus floribunda (Baker) Planch. M [E, C, W(N)] AMH 25 (nv); GES 2405 (1000 m) (nv); HP 6186 (800 m) (nv); Rakotozafy, A. & al. 2295 (nv)
Cissus microdonta (Baker) Planch. M [Sb, E, C, W, S] - RM SDR 099 (1100 m); SM 1179 (800 m); SMT 212 (1000 m); SMT 381 (1040 m); SMT 426 (905 m)
Cissus cf. microdonta (Baker) Planch. LN 2501 (1300 m); MHR 017 (1050 m)
Cissus sp. Leeuwenberg, A. J. M. & al. 14303 (950 m) (nv); OA 117 [800 m] (nv); OA 200 [900 m] (nv); SM 1730 [1045 m] (nv)
Leea D.Royen ex L. Leea guineensis G. Don M [Sb, E, C, W] - RM, Aft GES 1512 (1000 m) (nv); Harder, D. K. & al. 1573 (890 m) (nv); MHR 161 (875 m); PM 1180 (nv); Seigler, D. S. 12900 (nv); SMT 059
(1040 m); SMT 339 (910 m)
Leea guineensis var. cuspidifera (Baker) Desc. LB 11956 [1025 m] (nv); SM 1177 (850 m) (nv)
Leea guineensis fo. monticola Desc. HH 32095 (1000 m) (nv); HH 3965 (1100 m) (nv)

8. Annexes
174
Leea spinea Desc. M [E, C, W] - RM Harder, D. K. & al. 1618 (660 m) (nv); HP 17660 (600 m) (nv); OA 258 [375 m] (nv); OA 67 [950 m] (nv); OA 858 (533 m) (nv);
Rakotonandrasana, S. & al. 914 (nv); Rakotozafy, A. & al. 2292bis (nv); Rakotozafy, A. 2292 (nv); SF & J. Belin 10279 (nv); SM 1191 [785
m] (nv); SMT 488 (930 m)
Trois espèces cultivées :
AMARYLLIDACEAE
Furcraea Vent. Furcraea foetida (L.) Haw. Harder, D. K. & al. 1609 (450 m) (nv)
CELASTRACEAE Catha Forssk. ex Scop.
Catha edulis Forssk. SF & R. Capuron 29198bis (nv)
FABACEAE
Albizia Durazz. Albizia lebbeck (L.) Benth. Harder, D. K. & al. 1614 (450 m) (nv)

8. Annexes
175
8.1.1.1. INDEX DES GENRES DE PHANÉROGAMES Acacia Mill. ............................ 126 Acalypha L. ............................ 124 Acanthopale C.B.Clarke........ 101 Achyranthes L. ....................... 103 Achyrospermum Blume.......... 132 Acmella Rich. ......................... 109 Acridocarpus Guill. & Perr. .. 135 Acroceras Stapf ...................... 154 Actinoschoenus Benth. .......... 120 Adansonia L. .......................... 136 Adenostemma J.R.Forst. &
G.Forst. ............................. 109 Aerangis Rchb.f. .................... 145 Aeranthes Lindl...................... 145 Aeschynomene L. ................... 127 Agauria (DC.) Benth. & Hook.f.
........................................... 123 Agelaea Sol. ex Planch........... 118 Agrocharis Hochst. ................ 105 Ajuga L. .................................. 132 Alangium Lam. ...................... 119 Albizia Durazz. ............... 127, 174 Alchemilla L. .......................... 158 Alectra Thunb. ....................... 151 Allophylus L. .......................... 167 Aloe L. .................................... 109 Alternanthera Forssk. ............ 103 Ambavia Le Thomas .............. 104 Ambrella H.Perrier ................ 146 Anacolosa (Blume) Blume..... 144 Angraecum Bory .................... 146 Anomostachys Hurus. ............ 124 Anthocleista Afzel. ex R.Br. .. 130 Antidesma L............................ 152 Aphananthe Planch................ 115 Aphloia (DC.) Benn. .............. 130 Apodytes E.Mey. ex Arn. ....... 132 Aponogeton L.f....................... 107 Ardisia Sw............................... 142 Aristea Aiton .......................... 132 Artabotrys R.Br. ..................... 104 Asclepias L. ............................ 105 Asparagus L. .......................... 109 Aspidostemon Rohwer &
H.G.Richt. ......................... 134 Astiella Jovet .......................... 158 Asystasia Blume ..................... 101 Bakerella Tiegh. ..................... 135 Barleria L. .............................. 101 Basella L................................. 113 Bathiorhamnus Capuron....... 157 Baudouinia Baill. ................... 127 Bauhinia L. ............................ 127 Beclardia A.Rich. ................... 147 Begonia L. .............................. 113 Benoistia H.Perrier & Leandri
........................................... 124 Bidens L.................................. 110 Biophytum DC........................ 151 Bivinia Jaub. ex Tul............... 165 Boehmeria Jacq...................... 171 Bothriochloa Kuntze.............. 154 Brachiaria (Trin.) Griseb. ..... 154 Brachylaena R.Br. ................. 110
Brachystephanus Nees............101 Bremeria Razafim. & Alejandro
............................................159 Breonia A.Rich. ......................159 Brexia Noronha ex Thouars ..116 Brexiella H.Perrier.................116 Bridelia Willd. ........................152 Brillantaisia P.Beauv..............101 Buddleja L...............................170 Bulbophyllum Thouars ..........147 Burasaia Thouars...................140 Byttneria Loefl. .......................136 Calanthe R.Br.........................148 Calantica Jaub. ex Tul. ..........165 Calyptranthera Klack. ............105 Camptocarpus Decne. .............105 Camptolepis Radlk. ................167 Campylospermum Tiegh.........144 Canarium L.............................114 Canephora Juss. .....................159 Canthium Lam........................159 Cardamine L. ..........................114 Carex L. ..................................120 Carissa L. ................................105 Carphalea Juss........................159 Casearia Jacq..........................166 Cassia L...................................127 Cassipourea Aubl. ..................158 Catha Forssk. ex Scop............174 Cayratia Juss...........................173 Celtis L. ...................................115 Centauropsis Bojer ex DC......110 Centella L................................105 Centrosema (DC.) Benth. .......127 Cerastium L. ...........................116 Ceropegia L.............................105 Chamaecrista Moench............127 Chassalia Comm. ex Poir.......159 Cheirostylis Blume..................148 Chionanthus L. .......................144 Chloris Sw...............................154 Chlorophytum Ker Gawl........103 Chouxia Capuron...................167 Chrysanthellum Pers. .............110 Chrysophyllum L. ...................169 Cinchona L. ............................159 Cinnamosma Baill. .................115 Cissus L...................................173 Citrus L. ..................................165 Claoxylon A.Juss. ...................124 Cleistanthus Hook.f. ex Planch.
............................................152 Clematis L. ..............................157 Clerodendrum L......................132 Clitoria L.................................127 Coffea L. .................................159 Coix L......................................154 Colea Bojer ex Meisn. ............113 Coleotrype C.B.Clarke ...........118 Colubrina Rich. ex Brongn. ...157 Combretum Loefl....................118 Commelina L. .........................118 Conyza Less. ...........................110 Corchorus L. ...........................136
Cordemoya Baill..................... 124 Corymborkis Thouars ............ 148 Cosmos Cav............................ 110 Crassocephalum Moench ...... 110 Crateva L. ............................... 115 Crinum L. ............................... 103 Crossandra Salisb. ................. 101 Crotalaria L............................ 127 Croton L. ................................ 124 Cryptocarya R.Br. .................. 134 Cucumis L. ............................. 119 Cuscuta L. .............................. 119 Cyathula Blume ..................... 103 Cyclea Arn. ex Wight ............ 140 Cynanchum L......................... 106 Cynodon Rich......................... 154 Cynoglossum L....................... 114 Cynometra L........................... 128 Cynorkis Thouars .................. 148 Cyperus L. .............................. 120 Cyrtococcum Stapf ................. 154 Dalbergia L.f. ......................... 128 Danais Comm. ex Vent. ........ 160 Danguyodrypetes Leandri ..... 125 Datura L. ................................ 170 Deinbollia Schumach............. 167 Delonix Raf. ........................... 128 Desmodium Desv.................... 128 Desmostachys Planch. ex Miers
........................................... 132 Dianella Lam. ex Juss. .......... 131 Dichaetanthera Endl.............. 138 Dichapetalum Thouars .......... 122 Dichrocephala L'Hér. ex DC. 110 Dicliptera Juss. ....................... 101 Didymeles Thouars ................ 122 Digitaria Haller ...................... 154 Dioscorea L. ........................... 122 Diospyros L. ........................... 122 Diplocyclos (Endl.) Post &
Kuntze............................... 119 Disperis Sw. ............................ 148 Dombeya Cav. ........................ 136 Doratoxylon Thouars ex Hook.f.
........................................... 167 Dracaena Vand. ex L............. 109 Droguetia Gaudich. ............... 171 Drymaria Willd. ex Schult..... 116 Drypetes Vahl ......................... 157 Dupuya J.H.Kirkbr. .............. 128 Duranta L. .............................. 172 Dypsis Noronha ex Mart. ...... 108 Dyschoriste Nees .................... 101 Ehretia P.Browne .................. 114 Elaeocarpus L. ....................... 123 Elaeodendron Jacq. ............... 116 Elatostema J.R.Forst. & G.Forst.
........................................... 172 Elephantopus L. ..................... 110 Ellipanthus Hook.f................. 118 Embelia Burm.f...................... 142 Emilia Cass. ........................... 110 Enterospermum Hiern ........... 160 Ephippiandra Decne. ............. 140

8. Annexes
176
Eragrostis Wolf ...................... 155 Erblichia Seem. ...................... 151 Erica L. ................................... 123 Erythroxylum P.Browne ........ 123 Euclinia Salisb. ...................... 160 Eugenia L. .............................. 143 Euonymus L. .......................... 116 Euphorbia L. .......................... 125 Evonymopsis H.Perrier.......... 116 Excoecaria L. ......................... 125 Faucherea Lecomte ............... 169 Ficus L.................................... 141 Filicium Thwaites ex Benth. &
Hook.f................................ 167 Fimbristylis Vahl .................... 121 Flagellaria L........................... 130 Furcraea Vent. ....................... 174 Gaertnera Lam. ...................... 161 Gagnebina Neck. ex DC......... 128 Garcinia L. ............................. 117 Gastrorchis Schltr. ................. 149 Geosiris Baill. ......................... 132 Geranium L. ........................... 131 Gerbera L. .............................. 110 Givotia Griff. .......................... 126 Glenniea Hook.f. .................... 168 Gnidia L.................................. 171 Gomphocarpus R.Br. ............. 106 Gouania Jacq. ........................ 158 Grangeria Comm. ex Juss. .... 117 Gravesia Naudin .................... 138 Grevea Baill. ........................... 141 Grewia L. ................................ 137 Grisollea Baill......................... 170 Gymnosiphon Blume.............. 114 Gyrostipula J.-F.Leroy........... 161 Habenaria Willd. ................... 149 Haematoxylum L. ................... 129 Harpullia Roxb. ..................... 168 Harungana Lam. ................... 131 Helichrysum Mill. .................. 111 Heliotropium L. ...................... 114 Helmiopsis H.Perrier ............. 137 Hetaeria Blume ...................... 149 Hibiscus L............................... 137 Hilleria Vell. ........................... 152 Hilsenbergia Tausch ex Rchb.
........................................... 114 Hirtella L. ............................... 117 Homalium Jacq. ..................... 166 Hovanella A.Weber & B.L.Burtt
........................................... 131 Hydrocotyle L. ........................ 105 Hymenaea L. .......................... 129 Hyperacanthus E.Mey. ex
Bridson.............................. 161 Hypoestes Sol. ex R.Br. .......... 102 Hypoxis L. .............................. 132 Ilex L. ..................................... 107 Impatiens L............................. 112 Ipomoea L............................... 119 Isachne R.Br. ......................... 155 Isoglossa Oerst. ...................... 102 Isolona Engl. .......................... 104 Ixora L. ................................... 161 Jacaranda Juss. ...................... 113 Jasminum L............................ 144
Jumellea Schltr.......................149 Justicia L.................................102 Kalanchoe Adans....................119 Kyllinga Rottb. .......................121 Labramia A.DC. .....................169 Lagrezia Moq..........................103 Landolphia P.Beauv. ..............106 Langsdorffia Mart. .................112 Lantana L. ..............................173 Laportea Gaudich...................172 Lasiodiscus Hook.f. ................158 Lasiosiphon Fresen.................171 Laurembergia P.J.Bergius .....131 Lautembergia Baill. ................126 Leea D.Royen ex L. ................173 Lemurella Schltr.....................149 Lemyrea A.Chev. & Beille .....161 Lepidotrichilia (Harms) J.-
F.Leroy ..............................139 Lepisanthes Blume .................168 Leptaulus Benth......................116 Lindernia All...........................135 Liparis Rich. ...........................149 Lomatophyllum Willd.............109 Ludia Comm. ex Juss. ............166 Ludwigia L. .............................145 Macaranga Thouars...............126 Macarisia Thouars .................158 Machaerina Vahl ....................121 Macphersonia Blume .............168 Macrotyloma (Wight & Arn.)
Verdc. ................................129 Madlabium Hedge ..................133 Maesa Forssk..........................142 Malaxis Sol. ex Sw..................150 Malleastrum J.-F.Leroy .........139 Mammea L. .............................115 Mapouria Aubl. ......................161 Marantochloa Brongn. ex Gris
............................................138 Mariscus Vahl.........................121 Mascarenhasia A.DC. ............106 Maytenus Molina....................116 Medinilla Gaudich. ex DC. ....138 Meineckia Baill. ......................152 Melicope J.R.Forst. & G.Forst.
............................................165 Memecylon L. .........................139 Mendoncia Vell. ex Vand.......102 Mikania Willd.........................111 Millettia Wight & Arn. ..........129 Mimosa L. ...............................129 Mimulopsis Schweinf. ............102 Mimusops L. ...........................169 Momordica L. .........................119 Monanthotaxis Baill. ..............104 Morinda L. ..............................161 Mostuea Didr. .........................130 Mucuna Adans. ......................129 Musa L. ...................................142 Mussaenda L...........................161 Mystroxylon Eckl. & Zeyh. ....117 Nastus Juss. ............................155 Neoleroya Cavaco...................161 Neotina Capuron....................168 Nervilia Comm. ex Gaudich. .150 Nesogordonia Baill. ................138
Noronhia Stadman ex Thouars........................................... 144
Nuxia Comm. ex Lam. .......... 171 Nymphaea L. .......................... 143 Oberonia Lindl....................... 150 Ochna L.................................. 144 Ochrocarpos Noronha ex
Thouars............................. 117 Ocimum L............................... 133 Ocotea Aubl............................ 134 Oeceoclades Lindl. ................. 150 Oeonia Lindl. ......................... 150 Olax L..................................... 144 Oldenlandia L. ....................... 162 Olea L. .................................... 145 Olyra L. .................................. 155 Oncinotis Benth. .................... 106 Oncostemum A.Juss. .............. 142 Ophiocolea H.Perrier ............ 113 Oplismenus P.Beauv. ............. 155 Orfilea Baill............................ 126 Ouratea Aubl.......................... 144 Oxalis L. ................................. 151 Pachyrhizus Rich. ex DC. ...... 129 Pandanus Parkinson.............. 151 Panicum L. ............................. 155 Paspalum L. ........................... 155 Pauridiantha Hook.f. ............. 162 Pentopetia Decne.................... 106 Peperomia Ruiz & Pav. ......... 153 Peponidium (Baill.) Arènes ... 162 Petchia Livera........................ 106 Phaius Lour. .......................... 150 Phenax Wedd......................... 172 Phyllanthus L. ........................ 152 Phyllarthron DC..................... 114 Phyllopentas (Verdc.) Karehed
& K.Bremer...................... 162 Phylloxylon Baill. ................... 129 Physena Noronha ex Thouars152 Phytolacca L........................... 152 Pilea Lindl.............................. 172 Piper L. ................................... 153 Pithecellobium Mart. ............. 129 Pittosporum Banks ex Gaertn.
........................................... 153 Plagioscyphus Radlk.............. 168 Platylepis A.Rich.................... 150 Plectranthus L'Hér. ............... 133 Plectronia L............................ 162 Plumbago L. ........................... 154 Podocarpus L'Hér. ex Pers.... 101 Poecilostachys Hack. ............. 156 Polyalthia Blume.................... 104 Polycardia Juss....................... 117 Polygala L. ............................. 156 Polygonum L. ......................... 156 Polyscias J.R.Forst. & G.Forst.
........................................... 108 Polystachya Hook................... 150 Potamogeton L. ...................... 157 Pothos L. ................................ 108 Poupartia Comm. ex Juss...... 104 Premna L................................ 133 Procris Comm. ex Juss. ......... 172 Pseudognaphalium Kirp........ 111

8. Annexes
177
Pseudolasiacis (A.Camus) A.Camus ........................... 156
Psiadia Jacq............................ 111 Psorospermum Spach............. 131 Psychotria L............................ 162 Pueraria DC. .......................... 129 Pycnostachys Hook. ............... 133 Pycreus P.Beauv..................... 121 Pyrenacantha Hook. .............. 132 Pyrostria Comm. ex Juss. ...... 163 Randia L. ................................ 163 Ranunculus L. ........................ 157 Raphidiocystis Hook.f. ........... 119 Rauvolfia L. ............................ 106 Ravensara Sonn...................... 134 Rhipsalis Gaertn. ................... 115 Rhodocodon Baker................. 131 Rhopalocarpus Bojer ............. 170 Rhus L. ................................... 104 Rinorea Aubl. ......................... 173 Rothmannia Thunb................ 163 Roupellina (Baill.) Pichon ..... 106 Rubus L. ................................. 158 Ruellia L. ................................ 103 Rumex L. ................................ 156 Rytigynia Blume ..................... 164 Sabicea Aubl. ......................... 164 Salacia L................................. 117 Saldinia A.Rich. ex DC.......... 164 Sanicula L. ............................. 105 Satyrium Sw. .......................... 150 Schefflera J.R.Forst. & G.Forst.
........................................... 108 Schizenterospermum Homolle ex
Arènes ............................... 164 Schoenoplectus (Rchb.) Palla 121 Scleria P.J.Bergius................. 121 Sclerocarya Hochst. ............... 104 Scolopia Schreb...................... 166
Secamone R.Br. ......................106 Senecio L.................................111 Senna Mill...............................129 Sesamum L. ............................151 Setaria P.Beauv. .....................156 Sideroxylon L..........................169 Siegesbeckia L. .......................111 Sisyrinchium L........................132 Smilax L. .................................170 Sobennikoffia Schltr...............150 Solanum L...............................170 Solenangis Schltr....................151 Sonchus L. ..............................111 Sorindeia Thouars..................104 Spermacoce L..........................164 Sporobolus R.Br. ....................156 Stachys L.................................133 Stadmania Lam. .....................168 Staufferia Z.S.Rogers, Nickrent
& Malécot..........................166 Stellaria L. ..............................116 Stenotaphrum Trin. ................156 Stephanodaphne Baill.............171 Stephanostegia Baill. ..............107 Stereospermum Cham. ...........114 Streblus Lour. .........................141 Streptocarpus Lindl. ...............131 Strongylodon Vogel ................129 Strychnos L. ............................135 Suregada Roxb. ex Rottler.....126 Sylvichadsia Du Puy & Labat130 Symphonia L.f.........................118 Syzygium Gaertn. ...................143 Tabernaemontana L. ..............107 Talinella Baill. ........................157 Tambourissa Sonn. .................140 Tannodia Baill. .......................126 Tarenna Gaertn......................164 Taxillus Tiegh. ........................135
Terminalia L. ......................... 118 Thecacoris A.Juss. ................. 152 Themeda Forssk..................... 156 Thylachium DC. ..................... 115 Tina Schult. ............................ 168 Tisonia Baill. .......................... 166 Toddalia Juss.......................... 165 Tournefortia L........................ 114 Trema Lour. ........................... 115 Triainolepis Hook.f. ............... 164 Tricalysia A.Rich. ex DC. ...... 164 Trichilia P.Browne ................ 139 Triclisia Benth........................ 140 Trigonopyren Bremek............ 165 Trilepisium Thouars .............. 141 Tristellateia Thouars.............. 135 Tristemma Juss....................... 139 Triumfetta L. .......................... 138 Trophis P.Browne .................. 142 Turraea L. .............................. 140 Tylophora R.Br. ..................... 107 Uncarina Stapf....................... 151 Urera Gaudich. ...................... 172 Vaccinium L. .......................... 123 Vangueria Juss....................... 165 Vepris Comm. ex A.Juss........ 165 Vernonia Schreb. ................... 111 Viguieranthus Villiers............ 130 Viola L. ................................... 173 Viscum L. ............................... 166 Vitex L. ................................... 134 Weinmannia L........................ 120 Wielandia Baill....................... 152 Woodfordia Salisb. ................. 135 Xanthocercis Baill. ................. 130 Xylia Benth............................. 130 Xylopia L. ............................... 104 Zehneria Endl. ....................... 120

8. Annexes
178
8.1.2. PTÉRIDOPHYTES
PTÉRIDOPHYTES
Adiantum L. Adiantum madagascariense H.Rosend. M [E, W] SMT 531 (650 m)
Adiantum philippense L. M [Sb, E, C, W] - RM, Aft, As, Oc FR 1518 (nv)
Adiantum rhizophorum Sw. M [C] - RM FR 1519 (670 m) (nv)
Amauropelta Kunze cf. Amauropelta sp. SMT 215 (1000 m)
Antrophyum Kaulf. Antrophyum bivittatum C.Chr. M [Sb, C] FR 1776 (1250 m) (nv)
Antrophyum boryanum (Willd.) Spreng. M [Sb, C] - RM, Aft FR 1587 (1250 m) (nv); LN 2411 (1160 m); SDR 096 (1085 m)
Antrophyum immersum Willd. M [Sb, C] - RM, Aft SM 1210 (600 m) (nv)
Antrophyum malgassicum C.Chr. M [Sb, E, C] FR 1589 [1000 m] (nv); FR 1730 (1000 m) (nv); LN 2371 (1290 m); MHR 108 (990 m); OA 234 [375 m] (nv); OA 344 (1475 m) (nv); OA
367 [375 m] (nv); SMT 107 (1082 m); SMT 240 (1170 m)
Antrophyum sp. Badré, F. & al. 2113 [875 m] (nv); OA 34 [1438 m] (nv)
Arthropteris J.Smith ex Hook.f. Arthropteris monocarpa (Cordem.) C.Chr. M [C] - RM, Aft, SAf SM 2349 [1438 m] (nv); SMT 001 (1380 m); SMT 327 (955 m)
Asplenium L. Asplenium affine Sw. M [C] - RM, As Asplenium affine Sw. var. affine SMT 331 (975 m)
Asplenium aff. afzelii Rosenst. SMT 242 (1040 m)
Asplenium anisophyllum Kunze M [Sb, C] - Aft, SAf FR 1571 (nv)
Asplenium boltonii Hook. ex Schelpe ou eliotii ? SDR 082 (990 m)
Asplenium cancellatum Alston M [E, C] - Aft SM 1755 [1045 m] (nv)
Asplenium erectum Bory ex Willd. M [E, C] - RM, Aft SM 1227 (620 m) (nv)
Asplenium excisum C.Presl M [E, C] - RM, Aft, As, Oc SMT 155 (1040 m)

8. Annexes
179
Asplenium formosum Willd. M [C] - Aft, Amt, As FR 1569 (660 m) (nv); LN 2493 (1080 m)
Asplenium friesiorum C.Chr. M [C] - RM, Aft MHR 121 (1085 m); SM 2341 [1438 m] (nv)
Asplenium herpetopteris Baker M [E, C] SM 1767 [1045 m] (nv); SM 964 (1300 m) (nv)
Asplenium herpetopteris var. acutipinnata (Bonap.) Tardieu LN 2394 (1290 m)
Asplenium herpetopteris Baker var. herpetopteris FR 1566 (1100 m) (nv); FR 1584 [1000 m] (nv); SMT 144 (1050 m); SMT 243 (1040 m)
Asplenium herpetopteris var. masoulae (Bonap.) Tardieu FR 1567 (1300 m) (nv); FR 1568 (1300 m) (nv); FR 1777 (1150 m) (nv); LN 2396 (1412 m); SM 2347 [1438 m] (nv); SM 963 (1300 m)
(nv); SMT 166 (1080 m)
Asplenium inaequilaterale Willd. M [E, C] - RM, Aft, Amt, SAf, As FR 1563 (1100 m) (nv); FR 1564 (690 m) (nv); OA 284 [375 m] (nv)
Asplenium aff. inaequilaterale Willd. SMT 450 (1025 m)
Asplenium lokohoense Tardieu M [C] FR 1736 (1060 m) (nv); OA 227 [375 m] (nv)
Asplenium mangindranense Tardieu M [C] FR 1782 (1250 m) (nv)
Asplenium mannii Hook. M [C] - Aft FR 1721 (1050 m) (nv)
Asplenium masoulae Bonap. M [E, C] OA 315 (1475 m) (nv); OA 340 (1475 m) (nv)
Asplenium nidus L. M [Sb, E, C, W(S)] - RM, Aft, As, Oc LN 2494 (1260 m); MHR 068 (1040 m); SMT 158 (1040 m)
Asplenium obscurum Blume M [C] - Aft, As, Oc FR 1555 (nv); FR 1556 (1000 m) (nv); SM 2381 [1438 m] (nv); SMT 007 (1190 m)
Asplenium pellucidum Lam. M [Sb, E, C] FR 1557 (1000 m) (nv); FR 1558 (1000 m) (nv); MHR 122 (1085 m); SDR 085 (970 m); SMT 199 (1150 m)
Asplenium petiolulatum Mett. M [C] GES 1526 (1000 m) (nv)
Asplenium polyodon G. Forst. M [Sb, E, C] - RM, As, Oc SMT 219 (1000 m); SMT 455 (1000 m)
Asplenium poolii Baker M [E, C] Asplenium poolii Baker var. poolii SMT 163 (1080 m)
Asplenium protensum Schrad. M [C] - RM, Aft FR 1737 (1070 m) (nv); FR 1738 (1070 m) (nv)
Asplenium rutifolium (Bergius) Kunze M [E, C] - RM, Aft FR 1778 (980 m) (nv)
Asplenium sandersonii Hook. M [E, C] - RM, Aft, SAf GES 1527 (1000 m) (nv); LN 2412 (1230 m); MHR 101 (1000 m); OA 222 [375 m] (nv); OA 311 (1475 m) (nv); SM 1768 [1045 m] (nv);
SM 2345 [1438 m] (nv); SM 2358 [1438 m] (nv); SMT 135 (1005 m)
Asplenium thunbergii Kunze M [E, C] - RM, Aft, As FR 1783 (1240 m) (nv)

8. Annexes
180
Asplenium sp. 1 LN 2406 (1325 m)
Asplenium sp. Badré, F. & al. 2098 (1050 m) (nv); Badré, F. & al. 2109 [875 m] (nv); Badré, F. & al. 2115 [875 m] (nv); Badré, F. & al. 2116 [875 m] (nv);
Badré, F. & al. 2118 [875 m] (nv); OA 219 [900 m] (nv); OA 316 (1475 m) (nv); OA 96 [950 m] (nv); SM 2342 [1438 m] (nv)
Athyrium Roth Athyrium sp. Badré, F. & al. 2117 [875 m] (nv)
Belvisia Mirb. Belvisia spicata (L.f.) Mirb. M [E, C] - RM, Aft, As FR 1615 (1080 m) (nv); OA 233 [375 m] (nv); OA 295 (1475 m) (nv); SMT 122
Blechnum L. Blechnum attenuatum (Sw.) Mett. M [E, C] - RM, Aft, SAf FR 1574 (1200 m) (nv); GMP 14501 [1150 m] (nv); LN 2369 (1290 m); MHR 117 (1070 m); SM 1766 [1045 m] (nv); SMT 272 (1085 m)
Blechnum cf. punctulatum Sw. SMT 275 (1095 m)
Blechnum sp. 1 LN 2399 (1300 m)
Blechnum sp. Badré, F. & al. 2101 (1050 m) (nv); FR 1753 (1000 m) (nv)
Blotiella R.M.Tryon Blotiella madagascariensis R.M.Tryon M [E, C] FR 1595 (1200 m) (nv); OA 296 (1475 m) (nv)
Blotiella pubescens (Kaulf.) R.M.Tryon M [C] - RM FR 1594 (1475 m) (nv)
Blotiella aff. pubescens (Kaulf.) R.M.Tryon LN 2395 (1370 m); MHR 083 (1060 m)
Christella H.Lév. Christella dentata (Forssk.) Brownsey & Jermy M [E, C, W] - RM, Aft, As FR 1632 (1000 m) (nv); FR 1633 (960 m) (nv); FR 1750 (nv); FR 1751 (nv)
Christella distans (Hook.) Holttum M [Sb, E, C] - RM LN 2475 (1290 m); MHR 004 (1190 m); MHR 066 (1050 m); OA 127 (nv)
Coniogramme Fée Coniogramme madagascariensis C.Chr. M [C] - RM FR 1504 (1075 m) (nv); LN 2437 (1248 m); OA 129 [800 m] (nv); OA 341 (1475 m) (nv); SMT 043 (1270 m)
Ctenitis (C.Chr.) C.Chr. Ctenitis biformis (Mett.) Tardieu M [E, C] - RM FR 1626 (800 m) (nv); FR 1726 (600 m) (nv)
Ctenitis crinita (Poir.) Tardieu M [C, W] - RM Ctenitis crinita var. hispida (Kuhn) Tardieu FR 1742 (1020 m) (nv)
Ctenitis poolii (C.Chr.) Tardieu M [C] SMT 448 (1015 m)
Ctenopteris Blume ex Kunze Ctenopteris parvula (Willd.) J.Sm. M [C] - RM FR 1529 (1200 m) (nv); Janssen, T. 2455 (nv); Janssen, T. 2473 (nv); LN 2393 (1290 m)

8. Annexes
181
Ctenopteris zenkeri Tardieu M [C] - Aft SM 2332 [1438 m] (nv); SM 2336 [1438 m] (nv)
Cyathea Sm. Cyathea approximata Bonap. M [E, C] FR 1663 (1200 m) (nv); FR 1664 (1380 m) (nv); FR 1665 (1300 m) (nv); OA 37 [1438 m] (nv)
Cyathea bellisquamata Bonap. M [E, C] FR 1659 (1200 m) (nv)
Cyathea bellisquamata Bonap. var. ambrensis Janssen & Rakotondr. Janssen, T. 2462 (nv)
Cyathea decrescens Mett. M [Sb, E, C] LN 2463 (1400 m)
Cyathea hildebrandtii Kuhn M [E, C] - RM FR 1666 (1300 m) (nv); FR 1744 (1020 m) (nv); HP 17756 (1000 m) (nv); LB 12039 [1225 m] (nv); OA 128 [800 m] (nv); OA 30 [1438 m]
(nv); SM 984 (700 m) (nv)
Cyathea subincisa C.Chr. M [C] - Amt FR 1662 (1350 m) (nv)
Cyathea sp. 1 LN 2464 (1360 m); SDR 092 (1085 m); SDR 113 (970 m); SMT 204 (1170 m)
Cyathea sp. 2 LN 2418 (1400 m); SMT 205 (1170 m)
Cyathea sp. 3 LN 2465 (1280 m)
Cyathea sp. Badré, F. & al. 2100 (1050 m) (nv); Badré, F. & al. 2110 [875 m] (nv); FR 1745 (1050 m) (nv); FR 1786 (1160 m) (nv); HP 17757 (nv); OA
212 [900 m] (nv); OA 215 [900 m] (nv)
Cyclosorus Link Cyclosorus distans (Hook.) Ching M [Sb, E, C] - RM OA 318 (1475 m) (nv)
Cyclosorus samarensis Copel. Détermination douteuse : OA 230 [375 m] (nv)
Cyclosorus sp. SM 962 (1300 m) (nv)
Dennstaedtia Bernh. Dennstaedtia madagascariensis (Kunze) Tardieu M [E, C] OA 287 [375 m] (nv)
Deparia Hook. & Grev. Deparia boryana (Willd.) M. Kato M [C] - RM FR 1580 (nv); OA 124 (nv)
Dicranopteris Bernh. Dicranopteris linearis (Burm.f.) Underw. M [Sb, E, C, W] - RM, Aft, SAf, As, Oc, Amt LN 2480 (1320 m)
Didymochlaena Desv. Didymochlaena truncatula (Sw.) J.Sm. M [C] - RM, Aft, As, Oc, Amt FR 109 (nv); FR 1503 (nv); GMP 14490 [1150 m] (nv); LN 2417 (1400 m); OA 285 [375 m] (nv); OA 317 (1475 m) (nv); OA 342 (1475 m)
(nv); SDR 093 (1086 m); SM 2379 [1438 m] (nv)

8. Annexes
182
Diplazium Sw. Diplazium nemorale (Baker) Schelpe [complexe] M [C] - Aft LN 2380 (1290 m)
Diplazium proliferum (Lam.) Thouars M [C] - RM, Aft FR 1735 (980 m) (nv); SMT 353 (975 m)
Diplazium sp. Badré, F. & al. 2120 [875 m] (nv)
Doryopteris J.Sm. Doryopteris concolor (Langsd. & Fisch.) Kuhn M [Sb, C] - RM, Aft, SAf FR 1517 (nv)
Doryopteris kirkii (Hook.) Alston M [Sb, C, W] - RM, Aft FR 1515 (650 m) (nv)
Drymoglossum C.Presl Drymoglossum niphoboloides (Luerss.) Baker M [E, C] - RM FR 1616 (nv)
Drynaria (Bory) J.Sm. Drynaria willdenowii (Bory) T.Moore M [Sb, E, C] - RM SM 1268 [645 m] (nv); SMT 047 (985 m)
Dryopteris Adans. Dryopteris inaequalis (Schltdl.) Kuntze M [E, C] - RM, Aft FR 1622 [1000 m] (nv); FR 1739 (1070 m) (nv)
Dryopteris inaequalis (Schltdl.) Kuntze var. comorensis Tardieu FR 1740 (1070 m) (nv)
Dryopteris sp. Badré, F. & al. 2096 (1050 m) (nv)
Elaphoglossum Schott ex J.Sm. Elaphoglossum conforme (Sw.) Schott M [C] - RM, Aft, Amt FR 1772 (1160 m) (nv)
Elaphoglossum coriaceum Bonap. M [Sb, C] SM 1764 [1045 m] (nv)
Elaphoglossum deckenii (Kuhn) C.Chr. M [C] - RM, Aft, SAf SDR 077 [1040 m]
Elaphoglossum leucolepis (Baker) Krajina ex Tardieu M [C] AMH 15 [1050 m] (nv); FR 1606 (1240 m) (nv); GMP 14492 [1150 m] (nv); HH 32165 (nv); SM 2340 [1438 m] (nv)
Elaphoglossum aff. macropodium (Fée) T. Moore LN 2409 (1325 m)
Elaphoglossum scolopendriforme Tardieu M [C] FR 1605 (1350 m) (nv); LN 2479 (1320 m); MHR 052 (1110 m); SMT 133 (1005 m)
Elaphoglossum sieberi (Hook. & Grev.) T. Moore M [C] - RM FR 1741 (1050 m) (nv)
Elaphoglossum sp. 1 LN 2403 (1420 m)
Elaphoglossum sp. 2 LN 2489 (1100 m)
Elaphoglossum sp. Badré, F. & al. 2107 (700 m) (nv); Badré, F. & al. 2108 (700 m) (nv); SM 1763 [1045 m] (nv); SM 2350 [1438 m] (nv)

8. Annexes
183
Equisetum L. Equisetum ramosissimum Desf. M [C] - RM, Aft, As, Oc, Amt, Eur Labat, J.-N. & al. 2874 [850 m] (nv); LN 2422 (1250 m)
Grammitis Sw. Grammitis obtusa Kaulf. M [Sb, C] - RM FR 1532 (1400 m) (nv); LN 2402 (1350 m); SM 2335 [1438 m] (nv)
Huperzia Bernh. Huperzia gagnepainiana (Herter) Tardieu M [C] SMT 098 (1200 m)
Huperzia gnidioides (L.f.) Trevis. M [Sb, C] - RM, Aft FR 1645 (1100 m) (nv); FR 1747 (1040 m) (nv); FR 1793 (1050 m) (nv); LN 2995 (1300 m); Poisson, H. s.n. (nv)
Huperzia megastachya (Baker) Tardieu M [E, C] FR 1643 (1050 m) (nv); FR 1749 (1020 m) (nv); GMP 14482 [1050 m] (nv); LN 2355 (1300 m); MHR 055 (1110 m); MHR 114 (1040 m);
OA 336 (1475 m) (nv)
Huperzia obtusifolia (Sw.) Rothm. M [E, C, HM] - RM FR 1654 (1300 m) (nv); FR 1655 (1100 m) (nv); FR 1748 (1020 m) (nv); SMT 136 (1005 m); SMT 221 (1000 m)
Huperzia ophioglossoides (Lam.) Rothm. M [C] - RM, Aft FR 1651 (1400 m) (nv); LN 2474 (1430 m); SM 2343 [1438 m] (nv)
Huperzia pecten (Baker) Tardieu M [E, C] FR 1638 (1100 m) (nv); HP 17711 (nv); LN 2405 (1350 m)
Huperzia phlegmaria (L.) Rothm. M [Sb, E, C] - RM, Aft, As, Oc Huperzia phlegmaria var. tardieuae (Herter) Tardieu SMT 351 (1020 m)
Huperzia pichiana Tardieu M [MdA] FR 1641 (1300 m) (nv); HP 17773 (1200 m) (nv), TYPE: HT:Px2, IT:Px2; LN 2354 (1300 m)
Huperzia trigona (C.Chr.) Tardieu M [C] AMH 110 (nv); FR 1649 (nv); LN 2356 (1300 m); Poisson, H. s.n. (nv); SM 2337 [1438 m] (nv); SM 2355 [1438 m] (nv); SM 972 (1470 m)
(nv)
Huperzia verticillata (L.f.) Trevis. M [C] - RM, Aft, Amt FR 1637 (1300 m) (nv); HP 17694bis (nv); OA 324 (1475 m) (nv)
Huperzia sp. 1 LN 2996 (1310 m)
Huperzia sp. 2 SMT 217 (1000 m); SMT 318 (970 m)
Huperzia sp. OA 149 [1175 m] (nv); OA 150 [1175 m] (nv)
Hymenophyllum Sm. Hymenophyllum deltoideum C.Chr. M [E, C] FR 1552 (1400 m) (nv)
Hymenophyllum hirsutum (L.) Sw. M [C] - RM, Aft, As, Oc, Amt FR 1535 (1350 m) (nv); SMT 330 (975 m)
Hymenophyllum parvum C.Chr. M [C] FR 1542 (1050 m) (nv)
Hymenophyllum polyanthos (Sw.) Sw. M [C] - RM, Aft, SAf, As, Oc, Amt FR 1544 (1300 m) (nv)
Hymenophyllum poolii Baker M [C] LN 2397 (1360 m)

8. Annexes
184
Hymenophyllum sibthorpioides Mett. M [C] - RM, Aft, SAf FR 1533 (1250 m) (nv); FR 1534 (1200 m) (nv); GMP 14505 [1150 m] (nv); SM 2339 [1438 m] (nv)
Hypolepis Bernh. Hypolepis sparsisora (Schrad.) Kuhn M [C] - RM, Aft SM 2382 [1438 m] (nv)
Lepisorus (Sm.) Ching Lepisorus excavatus (Bory ex Willd.) Ching Echantillon rattaché avec doute: OA 226 [375 m] (nv)
Lindsaea Dryand. ex Sm. Lindsaea madagascariensis Baker M [C] - Aft FR 1582 (1390 m) (nv); LN 2434 (1400 m)
Lomariopsis Fée Lomariopsis cordata (Bonap.) Alston M [E, C] FR 1733 (1000 m) (nv); FR 1771 (1150 m) (nv); HP 7498 (800 m) (nv), TYPE: HT
Lomariopsis crassifolia Holttum M [C] FR 1576 (1300 m) (nv)
Lomariopsis pollicina (Willemet) Kuhn M [E, C] - RM FR 1732 (1000 m) (nv); FR 1770 (1250 m) (nv); LN 2414 (1175 m); SMT 039 (1270 m)
Lonchitis L. Lonchitis sp. Badré, F. & al. 2114 [875 m] (nv)
Loxogramme (Blume) C.Presl Loxogramme lanceolata (Sw.) C.Presl M [C] - RM, Aft FR 1577 (nv); LN 2404 (1480 m); MHR 093 (1080 m); OA 225 [375 m] (nv); OA 293 (1475 m) (nv); OA 325 (1475 m) (nv); OA 337 (1475
m) (nv); SM 1760 [1045 m] (nv); SMT 284 (1105 m)
Lycopodiella Holub Lycopodiella cernua (L.) Pic.Serm. M [Sb, E, C, W(S)] - RM, Aft, SAf, As, Oc, Amt LN 2482 (1310 m)
Lycopodium L. Lycopodium clavatum L. M [E, C] - RM, Aft, As, Oc, Amt, Eur Lycopodium clavatum var. borbonicum Bory FR 1505 (1100 m) (nv)
Lycopodium sp. OA 44 [1438 m] (nv)
Lygodium Sw. Lygodium kerstenii Kuhn M [Sb, C, W] - RM, Aft FR 1728 (250 m) (nv)
Lygodium lanceolatum Desv. M [Sb, E, C] - RM OA 246 [375 m] (nv); SDR 124
Marattia Sw. Marattia fraxinea Sm. M [Sb, E, C] - RM, Aft, SAf LN 2416 (1400 m); MHR 081 (1060 m)
Megalastrum Holttum Megalastrum lanuginosum (Willd. ex Kaulf.) Holttum M [C] - RM, Aft FR 1621 [1000 m] (nv)

8. Annexes
185
Microgramma C.Presl Microgramma lycopodioides (L.) Copel. M [Sb, E, W] - RM, Aft, SAf, Amt FR 1729 (600 m) (nv)
Microgramma mauritiana (Willd.) Tardieu M [Sb, E, W] - RM, Aft, SAf, Amt LN 2491 (1100 m)
Microlepia C.Presl Microlepia madagascariensis (Kunze) C.Presl M [Sb, E, C] LN 2477 (1320 m); OA 319 (1475 m) (nv); SDR 094 (1085 m)
Microsorum Link Microsorum pappei (Mett. ex Kuhn) Tardieu M [C] - RM, Aft, SAf FR 1731 (1000 m) (nv); SM 1733 [1045 m] (nv)
Microsorum polycarpon (Cav.) Tardieu M [Sb, E, C, W] - RM, Aft, SAf, As, Oc MHR 084 (1040 m)
Microsorum sp. Badré, F. & al. 2095 (1050 m) (nv)
Nephrolepis Schott Nephrolepis biserrata (Sw.) Schott M [Sb, E, W] - RM, Aft, SAf, As, Oc, Amt SMT 392 (990 m)
Nephrolepis undulata (Afzel. ex Sw.) J.Sm. M [Sb, E, C, W] - RM, Aft, As, Amt LN 2469 (1470 m); MHR 123 (1120 m); SMT 087 (1065 m); SMT 241 (1150 m); SMT 248 (980 m)
Nephrolepis sp. 1 SMT 366 (975 m)
Oleandra Cav. Oleandra distenta Kunze M [E, C] - RM, Aft, SAf FR 1572 (1300 m) (nv); LN 2432 (1400 m); OA 307 (1475 m) (nv); SMT 253 (970 m)
Ophioglossum L. Ophioglossum pendulum L. M [C] - RM, Amt, As, Oc FR 1746 (1050 m) (nv)
Pellaea Link Pellaea angulosa (Bory ex Willd.) Baker M [E, C] - RM, Aft FR 1512 (1000 m) (nv)
Phymatodes C. Presl Phymatodes scolopendria (Burm. f.) Ching M [Sb, E] - RM, Aft, SAf, As, Oc SM 1272 (640 m) (nv)
Phymatosorus Pic.Serm. Phymatosorus scolopendria (Burm.f.) Pic.Serm. M [Sb, E] - RM, Aft, SAf, As, Oc MHR 089 [1005 m]; OA 244 [375 m] (nv); OA 775 (874 m) (nv); SDR 084 (970 m); SMT 030 (1040 m)
Platycerium Desv. Platycerium quadridichotomum (Bonap.) Tardieu M [W] FR 1801 (540 m) (nv); FR 1802 (540 m) (nv)
Pleopeltis Humb. & Bonpl. ex Willd. Pleopeltis excavata (Bory ex Willd.) Sledge M [C] - RM, Aft, SAf LN 2372 (1400 m); MHR 054 (1110 m)
Pleopeltis macrocarpa (Bory ex Willd.) Kaulf. M [C, W(S)] - RM, Aft, Amt SM 988 (700 m) (nv)

8. Annexes
186
Pleopeltis schraderi (Mett.) Tardieu M [C] - Aft SM 2334 [1438 m] (nv)
Pneumatopteris Nakai Pneumatopteris remotipinna (Bonap.) Holttum M [C, W(N)] FR 1628 (1000 m) (nv); SMT 294 (1065 m)
Pneumatopteris subpennigera (C.Chr.) Holttum M [C] - RM, Aft FR 1635 [1300 m] (nv)
Pteris L. Pteris biaurita L. M [C, W(N)] - RM, As, Amt FR 1796 (680 m) (nv)
Pteris catoptera Kunze M [C] - RM, Aft FR 1754 (1020 m) (nv); FR 1797 (nv); FR 1798 (nv); FR 1799 (nv)
Pteris linearis Poir. M [Sb, E, C] - RM, Aft, As OA 214 [900 m] (nv)
Pteris cf. linearis Poir. OA 320 (1475 m) (nv)
Pteris mettenii Kuhn M [Sb] - RM FR 1756 (580 m) (nv)
Pteris quadriaurita Retz. M [C] - RM, Aft, Amt, As Pteris quadriaurita subsp. catoptera (Kunze) Schelpe LN 2379 (1290 m); MHR 154 (985 m)
Pteris woodwardioides Bory M [W] FR 1597 (800 m) (nv); FR 1598 (1185 m) (nv); FR 1755 (nv); OA 266 [375 m] (nv); SMT 482 (1000 m)
Pteris sp. 1 LN 2502 (1200 m)
Pteris sp. Badré, F. & al. 2089 (1050 m) (nv); Badré, F. & al. 2097 (1050 m) (nv); Badré, F. & al. 2104 (850 m) (nv); FR 1601 (1300 m) (nv); FR 1800
(1250 m) (nv)
Pyrrosia Mirb. Pyrrosia lanceolata (L.) Farw. M [Sb, C] - RM, Aft, As, Oc FR 1620 [1000 m] (nv); OA 279 [375 m]
Pyrrosia sp. 1 LN 2429 (1230 m)
Pyrrosia sp. SM 1198 (620 m) (nv)
Rumohra Raddi Rumohra adiantiformis (G.Forst.) Ching M [C] - RM, Aft, SAf, Oc, Amt LN 2420 (1380 m)
Rumohra lokohensis Tardieu M [C] FR 1573 (1330 m) (nv)
Schizaea Sm. Schizaea dichotoma (L.) J.Sm. M [E, C] - RM, Aft, As, Oc LN 2423 (1260 m); MHR 113 (1040 m)

8. Annexes
187
Selaginella P.Beauv. Selaginella goudotiana Spring M [C] Selaginella goudotiana Spring var. goudotiana FR 1522 (1000 m) (nv); FR 1523 (1300 m) (nv); FR 1524 (1000 m) (nv); FR 1554 (1300 m) (nv); JB s.n (nv); Keraudren, M. & al. 25568
(nv); Keraudren, M. 1716 [1000 m] (nv); SM 2374 [1438 m] (nv)
Selaginella kraussiana (Kunze) A. Braun M [C] - RM, Aft, Amt, Eur OA 189 [900 m] (nv)
Sphaerostephanos J.Sm. Sphaerostephanos unitus (L.) Holttum M [Sb, E, C, S] - RM SMT 430 (885 m)
Tectaria Cav. Tectaria gemmifera (Fée) Alston M [Sb, E, C] - RM, Aft, SAf FR 1509 (1050 m) (nv); FR 1727 (600 m) (nv); OA 231 [375 m] (nv); OA 286 [375 m] (nv); SMT 411 (985 m); SMT 452 (1040 m)
Tectaria sp. Badré, F. & al. 2092 (1050 m) (nv)
Thelypteris Schmidel Thelypteris confluens (Thunb.) C.V.Morton M [C] - Aft LN 2443 (1248 m)
Thelypteris interrupta (Willd.) K. Iwats. M [C] - Aft, Amt FR 1627 (nv)
Thelypteris subpennigera (C. Chr.) C.F. Reed M [C] - RM, Aft SM 2372 [1438 m] (nv)
Trichomanes L. Trichomanes bipunctatum Poir. M [Sb, E, C] - RM, As, Oc FR 1550 (1100 m) (nv); FR 1722 (1000 m) (nv); SMT 283 (1105 m)
Trichomanes borbonicum Bosch M [Sb, E, C] - RM, Aft FR 1545 (1360 m) (nv); FR 1546 (1240 m) (nv); FR 1791 (1160 m) (nv)
Trichomanes cupressoides Desv. M [Sb, E, C] - RM, Aft FR 1536 (1380 m) (nv); SMT 262 (1040 m)
Trichomanes cuspidatum Willd. M [Sb, E, C] - RM FR 1789 (1150 m) (nv)
Trichomanes cuspidatum Willd. f. rotundifolium Bonap. FR 1553 (1240 m) (nv)
Trichomanes digitatum Sw. M [Sb, C] - RM, As, Oc FR 1551 (1310 m) (nv)
Trichomanes fallax H.Christ M [C] - Aft FR 1734 (1000 m) (nv)
Trichomanes giganteum Willd. M [C] - RM, Aft SMT 250 (980 m)
Trichomanes lenormandii Bosch M [Sb, E, C] - RM FR 1540 (1380 m) (nv); FR 1541 (1250 m) (nv)
Trichomanes mannii Hook. M [Sb, C, W(S)] - RM, Aft FR 1725 (1040 m) (nv); SMT 311 (1075 m)
Trichomanes aff. mannii Hook. FR 1792 (1050 m) (nv)
Trichomanes meifolium Bory M [C] - RM FR 1538 (1390 m) (nv); LN 2377 (1420 m); SM 2373 [1438 m] (nv)

8. Annexes
188
Trichomanes melanotrichum Schltdl. M [E, C] - Aft FR 1548 (nv); FR 1549 (1300 m) (nv); FR 1724 (1000 m) (nv)
Trichomanes montanum Hook. M [E, C] - Aft, Amt FR 1539 (1300 m) (nv)
Trichomanes speciosum Willd. M [C] - RM, Aft, Amt, Eur FR 1537 (1250 m) (nv)
Trichomanes sp. 1 SMT 104 (1200 m)
Vittaria Sm. Vittaria elongata Sw. [complexe] M [Sb, C, ] - RM, As FR 1590 [1050 m] (nv); FR 1591 (960 m) (nv); SMT 322 (970 m)
Vittaria humblotii Hieron. M [E, C] - RM FR 1592 (1350 m) (nv); FR 1593 (1200 m) (nv); SM 2348 [1438 m] (nv); SMT 266 (1055 m)
Vittaria sp. Badré, F. & al. 2099 (1050 m) (nv)
Xiphopteris Kaulf. Xiphopteris myosuroides (Sw.) Kaulf. M [C] - RM, Aft JB 5932 (1200 m) (nv)
Xiphopteris serrulata (Sw.) Kaulf. M [E, C] - RM, Aft, Amt, Eur LN 2401 (1325 m)
Zygophlebia L.E. Bishop Zygophlebia aff. devoluta (Baker) Parris SM 2333 [1438 m] (nv); SM 2338 [1438 m] (nv)
Pteridophyta indéterminés sp. 1 MHR 005
Pteridophyta indéterminés sp. 2 LN 2503 (1200 m)
Pteridophyta indéterminés sp. Miller, J. S. & al. 10682 (1050 m) (nv)

8. Annexes
189
8.1.3. BRYOPHYTES, ASCOMYCÈTES LICHENISÉS ET CHAMPIGNONS
BRYOPHYTES
Bryophyta LN 2360 (1300 m); LN 2383 (1310 m); LN 2453 (1248 m); SMT 312 (1075 m); SMT 358 (970 m); SMT 367 (970 m)
Hepaticae LN 2368 (1400 m); SMT 287 (1035 m); SMT 290 (1065 m); SMT 296 (1065 m); SMT 297 (1065 m); SMT 429 (885 m)
Marchantiaceae LN 2476 (1290 m)
Rhizogoniaceae SMT 271 (1070 m)
Anthoceros sp. LN 2452 (1248 m)
Fissidens sp. MHR 079 (1065 m)
Hypopterygium sp. LN 2398 (1360 m)
Leucobryum sp. LN 2387 (1350 m)
Leucoloma sp. LN 2366 (1300 m)
Marchantia sp. LN 2351 (1250 m)
Metzgeria sp. LN 2367 (1250 m)
Neckera sp. LN 2365 (1300 m)
Polytrichum sp. LN 2376 (1400 m); LN 2481 (1320 m)
Pyrrhobryum sp. SMT 169 (1100 m)
Sematophyllum sp. SMT 270 (1070 m)
cf. Syrrhopodon sp. SMT 343 (1025 m)
Weymouthia sp. LN 2364 (1300 m)

8. Annexes
190
ASCOMYCÈTES LICHENISÉS
Cladonia sp. LN 2358 (1320 m); LN 2407 (1350 m)
Collema sp. LN 2486 (1100 m)
Heterodermia leucomela (L.) Poelt SMT 083 (1162 m)
Hypogymnia sp. LN 2427 (1250 m)
Leptogium azureum (Ach.) Mont. SMT 100 (1200 m)
Letrouitia muralis Hafellner SMT 082 (1162 m)
Lobanilla sp. LN 2419 (1400 m)
Parmelia sp. LN 2484 (1100 m)
Parmotrema sp. LN 2435 (1250 m)
Physcia sp. LN 2487 (1100 m)
Physma byrsinum (Ach.) Müll.Arg. SMT 099 (1200 m)
Sticta dichotoma (Bory) Delise SMT 060 (1030 m)
Sticta sp. SMT 025 (1300 m); SMT 181 (1065 m); SMT 265 (1055 m)
Usnea sp.
LN 2392 (1175 m); LN 2424 (1140 m); LN 2485 (1100 m); SMT 084 (1162 m) Ascomycètes lichenisés indéterminés LN 2361 (1300 m); LN 2363 (1300 m); LN 2410 (1250 m); LN 2426 (1170 m); LN 2478 (1320 m); LN 2488 (1100 m); MHR 126 (1120 m);
SMT 080 (1162 m); SMT 081 (1162 m); SMT 102 (1200 m); SMT 103 (1200 m); SMT 167 (1080 m); SMT 228 (1045 m); SMT 247 (1000
m); SMT 260 (1040 m); SMT 286 (1035 m); SMT 298 (1095 m); SMT 299 (1095 m); SMT 356 (975 m); SMT 379 (1035 m); SMT 394
(990 m)
CHAMPIGNONS
Champignons indéterminés LN 2470 (1120 m); LN 2657 (1260 m)
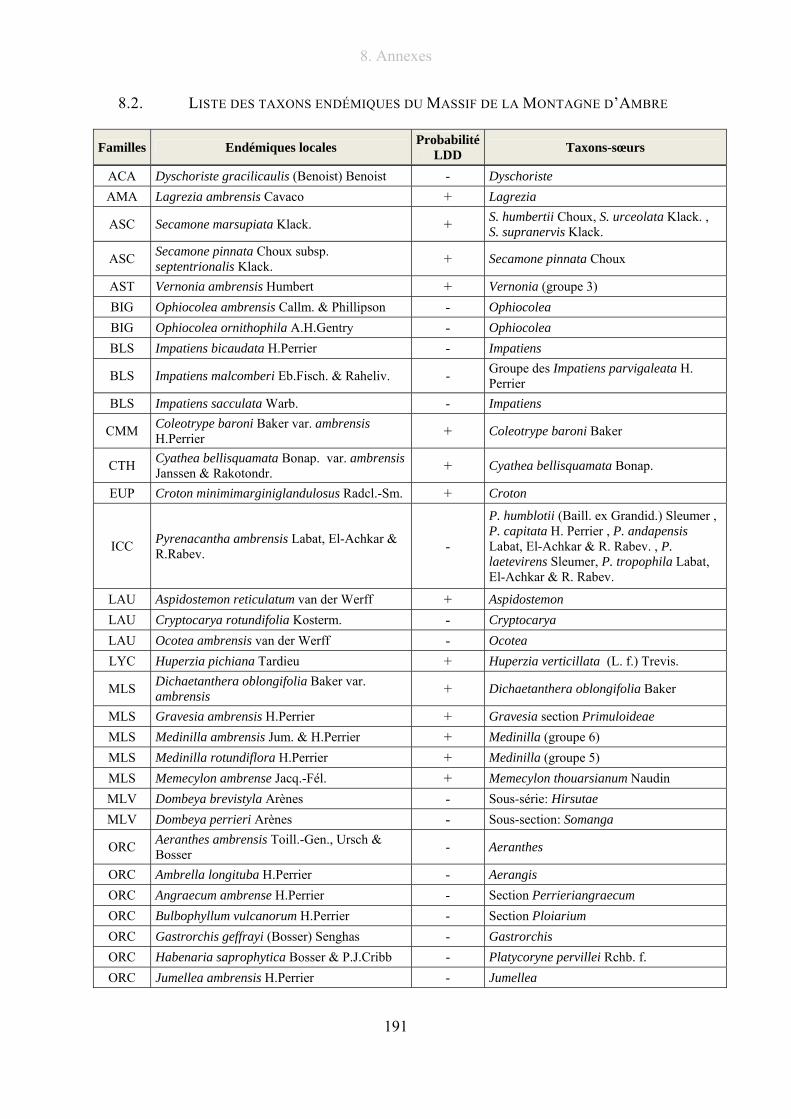
8. Annexes
191
8.2. LISTE DES TAXONS ENDÉMIQUES DU MASSIF DE LA MONTAGNE D’AMBRE
Familles Endémiques locales Probabilité LDD Taxons-sœurs
ACA Dyschoriste gracilicaulis (Benoist) Benoist - Dyschoriste AMA Lagrezia ambrensis Cavaco + Lagrezia
ASC Secamone marsupiata Klack. + S. humbertii Choux, S. urceolata Klack. , S. supranervis Klack.
ASC Secamone pinnata Choux subsp. septentrionalis Klack. + Secamone pinnata Choux
AST Vernonia ambrensis Humbert + Vernonia (groupe 3) BIG Ophiocolea ambrensis Callm. & Phillipson - Ophiocolea BIG Ophiocolea ornithophila A.H.Gentry - Ophiocolea BLS Impatiens bicaudata H.Perrier - Impatiens
BLS Impatiens malcomberi Eb.Fisch. & Raheliv. - Groupe des Impatiens parvigaleata H. Perrier
BLS Impatiens sacculata Warb. - Impatiens
CMM Coleotrype baroni Baker var. ambrensis H.Perrier + Coleotrype baroni Baker
CTH Cyathea bellisquamata Bonap. var. ambrensis Janssen & Rakotondr. + Cyathea bellisquamata Bonap.
EUP Croton minimimarginiglandulosus Radcl.-Sm. + Croton
ICC Pyrenacantha ambrensis Labat, El-Achkar & R.Rabev. -
P. humblotii (Baill. ex Grandid.) Sleumer , P. capitata H. Perrier , P. andapensis Labat, El-Achkar & R. Rabev. , P. laetevirens Sleumer, P. tropophila Labat, El-Achkar & R. Rabev.
LAU Aspidostemon reticulatum van der Werff + Aspidostemon LAU Cryptocarya rotundifolia Kosterm. - Cryptocarya LAU Ocotea ambrensis van der Werff - Ocotea LYC Huperzia pichiana Tardieu + Huperzia verticillata (L. f.) Trevis.
MLS Dichaetanthera oblongifolia Baker var. ambrensis + Dichaetanthera oblongifolia Baker
MLS Gravesia ambrensis H.Perrier + Gravesia section Primuloideae MLS Medinilla ambrensis Jum. & H.Perrier + Medinilla (groupe 6) MLS Medinilla rotundiflora H.Perrier + Medinilla (groupe 5) MLS Memecylon ambrense Jacq.-Fél. + Memecylon thouarsianum Naudin MLV Dombeya brevistyla Arènes - Sous-série: Hirsutae MLV Dombeya perrieri Arènes - Sous-section: Somanga
ORC Aeranthes ambrensis Toill.-Gen., Ursch & Bosser - Aeranthes
ORC Ambrella longituba H.Perrier - Aerangis ORC Angraecum ambrense H.Perrier - Section Perrieriangraecum ORC Bulbophyllum vulcanorum H.Perrier - Section Ploiarium ORC Gastrorchis geffrayi (Bosser) Senghas - Gastrorchis ORC Habenaria saprophytica Bosser & P.J.Cribb - Platycoryne pervillei Rchb. f. ORC Jumellea ambrensis H.Perrier - Jumellea
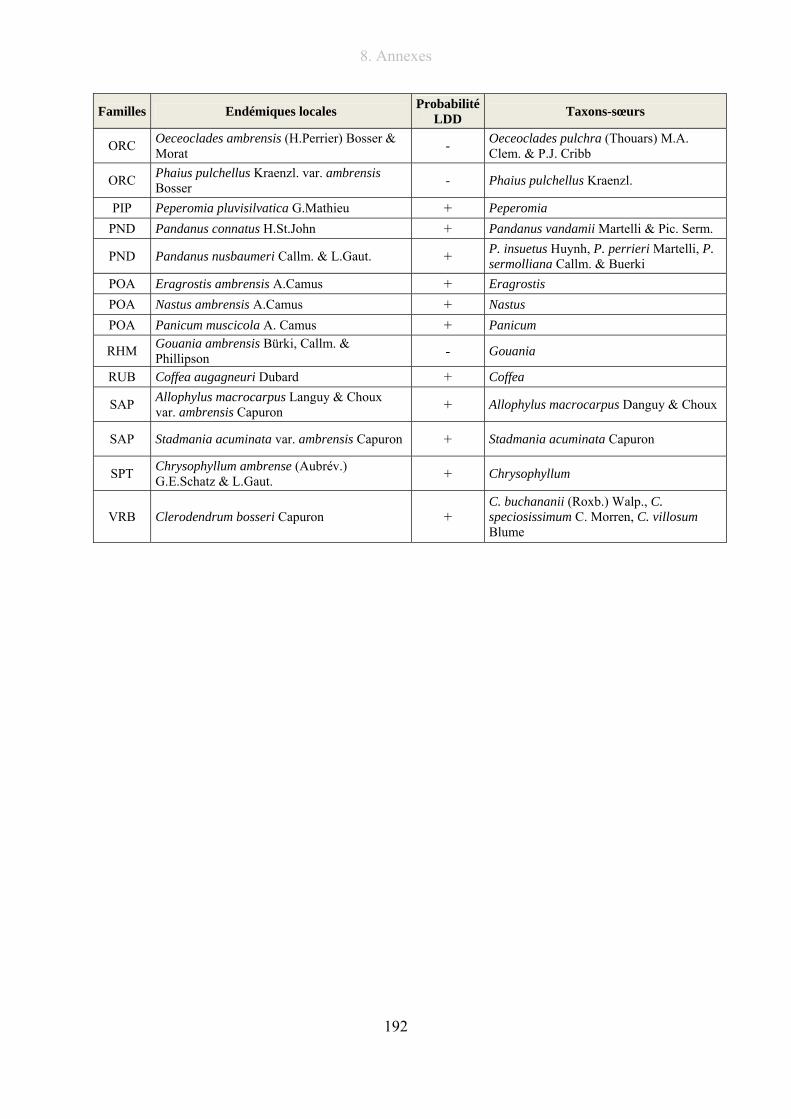
8. Annexes
192
Familles Endémiques locales Probabilité LDD Taxons-sœurs
ORC Oeceoclades ambrensis (H.Perrier) Bosser & Morat - Oeceoclades pulchra (Thouars) M.A.
Clem. & P.J. Cribb
ORC Phaius pulchellus Kraenzl. var. ambrensis Bosser - Phaius pulchellus Kraenzl.
PIP Peperomia pluvisilvatica G.Mathieu + Peperomia PND Pandanus connatus H.St.John + Pandanus vandamii Martelli & Pic. Serm.
PND Pandanus nusbaumeri Callm. & L.Gaut. + P. insuetus Huynh, P. perrieri Martelli, P. sermolliana Callm. & Buerki
POA Eragrostis ambrensis A.Camus + Eragrostis POA Nastus ambrensis A.Camus + Nastus POA Panicum muscicola A. Camus + Panicum
RHM Gouania ambrensis Bürki, Callm. & Phillipson - Gouania
RUB Coffea augagneuri Dubard + Coffea
SAP Allophylus macrocarpus Languy & Choux var. ambrensis Capuron + Allophylus macrocarpus Danguy & Choux
SAP Stadmania acuminata var. ambrensis Capuron + Stadmania acuminata Capuron
SPT Chrysophyllum ambrense (Aubrév.) G.E.Schatz & L.Gaut. + Chrysophyllum
VRB Clerodendrum bosseri Capuron + C. buchananii (Roxb.) Walp., C. speciosissimum C. Morren, C. villosum Blume
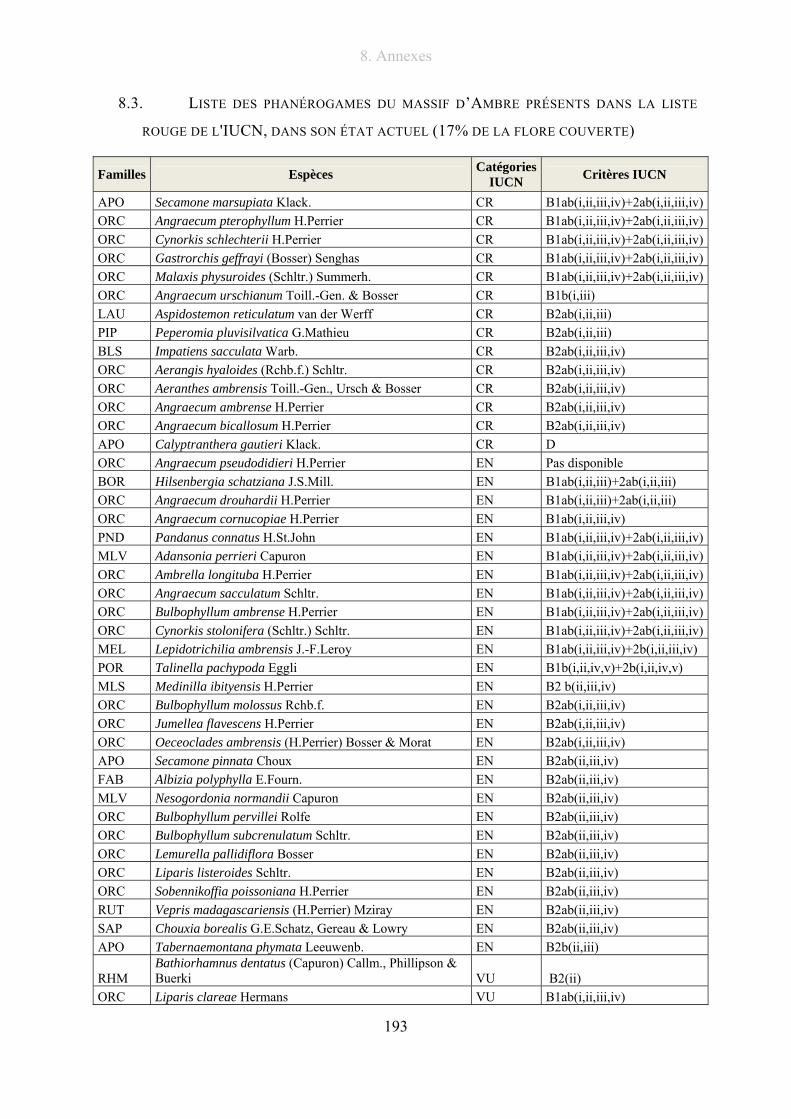
8. Annexes
193
8.3. LISTE DES PHANÉROGAMES DU MASSIF D’AMBRE PRÉSENTS DANS LA LISTE
ROUGE DE L'IUCN, DANS SON ÉTAT ACTUEL (17% DE LA FLORE COUVERTE)
Familles Espèces Catégories IUCN Critères IUCN
APO Secamone marsupiata Klack. CR B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Angraecum pterophyllum H.Perrier CR B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Cynorkis schlechterii H.Perrier CR B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Gastrorchis geffrayi (Bosser) Senghas CR B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Malaxis physuroides (Schltr.) Summerh. CR B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Angraecum urschianum Toill.-Gen. & Bosser CR B1b(i,iii) LAU Aspidostemon reticulatum van der Werff CR B2ab(i,ii,iii) PIP Peperomia pluvisilvatica G.Mathieu CR B2ab(i,ii,iii) BLS Impatiens sacculata Warb. CR B2ab(i,ii,iii,iv) ORC Aerangis hyaloides (Rchb.f.) Schltr. CR B2ab(i,ii,iii,iv) ORC Aeranthes ambrensis Toill.-Gen., Ursch & Bosser CR B2ab(i,ii,iii,iv) ORC Angraecum ambrense H.Perrier CR B2ab(i,ii,iii,iv) ORC Angraecum bicallosum H.Perrier CR B2ab(i,ii,iii,iv) APO Calyptranthera gautieri Klack. CR D ORC Angraecum pseudodidieri H.Perrier EN Pas disponible BOR Hilsenbergia schatziana J.S.Mill. EN B1ab(i,ii,iii)+2ab(i,ii,iii) ORC Angraecum drouhardii H.Perrier EN B1ab(i,ii,iii)+2ab(i,ii,iii) ORC Angraecum cornucopiae H.Perrier EN B1ab(i,ii,iii,iv) PND Pandanus connatus H.St.John EN B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)MLV Adansonia perrieri Capuron EN B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Ambrella longituba H.Perrier EN B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Angraecum sacculatum Schltr. EN B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Bulbophyllum ambrense H.Perrier EN B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Cynorkis stolonifera (Schltr.) Schltr. EN B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)MEL Lepidotrichilia ambrensis J.-F.Leroy EN B1ab(i,ii,iii,iv)+2b(i,ii,iii,iv) POR Talinella pachypoda Eggli EN B1b(i,ii,iv,v)+2b(i,ii,iv,v) MLS Medinilla ibityensis H.Perrier EN B2 b(ii,iii,iv) ORC Bulbophyllum molossus Rchb.f. EN B2ab(i,ii,iii,iv) ORC Jumellea flavescens H.Perrier EN B2ab(i,ii,iii,iv) ORC Oeceoclades ambrensis (H.Perrier) Bosser & Morat EN B2ab(i,ii,iii,iv) APO Secamone pinnata Choux EN B2ab(ii,iii,iv) FAB Albizia polyphylla E.Fourn. EN B2ab(ii,iii,iv) MLV Nesogordonia normandii Capuron EN B2ab(ii,iii,iv) ORC Bulbophyllum pervillei Rolfe EN B2ab(ii,iii,iv) ORC Bulbophyllum subcrenulatum Schltr. EN B2ab(ii,iii,iv) ORC Lemurella pallidiflora Bosser EN B2ab(ii,iii,iv) ORC Liparis listeroides Schltr. EN B2ab(ii,iii,iv) ORC Sobennikoffia poissoniana H.Perrier EN B2ab(ii,iii,iv) RUT Vepris madagascariensis (H.Perrier) Mziray EN B2ab(ii,iii,iv) SAP Chouxia borealis G.E.Schatz, Gereau & Lowry EN B2ab(ii,iii,iv) APO Tabernaemontana phymata Leeuwenb. EN B2b(ii,iii)
RHM Bathiorhamnus dentatus (Capuron) Callm., Phillipson & Buerki VU B2(ii)
ORC Liparis clareae Hermans VU B1ab(i,ii,iii,iv)

8. Annexes
194
Familles Espèces Catégories IUCN Critères IUCN
MLV Adansonia madagascariensis Baill. VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)BEG Begonia humbertii Keraudren VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)BLS Impatiens bisaccata Warb. VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)CLU Garcinia disepala Vesque VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)FAB Dalbergia hildebrandtii Vatke VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)FAB Sylvichadsia grandifolia (R.Vig.) Du Puy & Labat VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ORC Cynorkis tryphioides Schltr. VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)RUB Tricalysia ambrensis Randriamb. & De Block VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)RUT Vepris dicarpella H.Perrier VU B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv)ARL Polyscias aculeata (Decne. & Planch.) Harms VU B2ab(i,ii,iii) ANN Xylopia buxifolia Baill. VU B2ab(i,ii,iii,iv) ORC Angraecum caricifolium H.Perrier VU B2ab(i,ii,iii,iv) FAB Acacia pervillei Benth. VU B2ab(ii,iii,iv) ORC Liparis anthericoides H.Perrier VU B2ab(ii,iii,iv) RHM Bathiorhamnus louvelii (H.Perrier) Capuron VU B2ab(ii,iii,iv) SAN Staufferia capuronii Z.S. Rogers & Nickrent & Malécot VU B2ab(ii,iii,iv) SPT Sideroxylon gerrardianum (Hook.f.) Aubrév. VU B2ab(ii,iii,iv) APO Tabernaemontana coffeoides Bojer ex A.DC. VU B2b(iii) PND Pandanus spicatus H.St.John VU D1 ICC Pyrenacantha ambrensis Labat, El-Achkar & R.Rabev. VU D2 PND Pandanus latistigmaticus Huynh VU D2 APO Camptocarpus acuminatus (Choux) Venter NT APO Tabernaemontana eusepala Aug.DC. NT BEG Begonia francoisii Guillaumin NT CUN Weinmannia sanguisugarum Bernardi NT FAB Albizia boivinii E.Fourn. NT FAB Cynometra sakalava Du Puy & R.Rabev. NT LAM Plectranthus vinaceus Hedge NT ORC Aeranthes schlechteri Bosser NT PIP Peperomia glabrilimba C.DC. NT RUB Breonia capuronii Razafim. NT ACA Mendoncia cowanii (S.Moore) Benoist LC ACA Mendoncia flagellaris (Baker) Benoist LC APO Oncinotis tomentella Radlk. LC ARE Dypsis baronii (Becc.) Beentje & J.Dransf. LC ARL Polyscias amplifolia (Baker) Harms LC ARL Polyscias boivinii (Seem.) Bernardi LC ARL Polyscias ornifolia (Baker) Harms LC AST Brachylaena perrieri (Drake) Humbert LC AST Centauropsis rhaponticoides (Baker) Drake LC BEG Begonia madecassa Keraudren LC BIG Ophiocolea floribunda (Bojer ex Lindl.) H.Perrier LC BLS Impatiens lyallii Baker LC CLU Garcinia decipiens (Baill.) Vesque LC CLU Garcinia orthoclada Baker LC CLU Symphonia oligantha Baker LC CUN Weinmannia decora Tul. LC

8. Annexes
195
Familles Espèces Catégories IUCN Critères IUCN
DCH Dichapetalum leucosia (Spreng.) Engl. LC FAB Clitoria lasciva Bojer ex Benth. LC FAB Cynometra abrahamii Du Puy & R.Rabev. LC FAB Dalbergia madagascariensis Vatke LC FAB Millettia lenneoides Vatke LC FAB Millettia richardiana (Baill.) Du Puy & Labat LC FAB Mucuna paniculata Baker LC
FAB Phylloxylon xylophylloides (Baker) Du Puy, Labat & Schrire LC
FAB Xanthocercis madagascariensis Baill. LC IRI Aristea cladocarpa Baker LC LAM Ajuga oocephala Baker LC LAM Stachys reptans Hedge LC MNM Tambourissa purpurea (Tul.) A.DC. LC MOR Ficus politoria Lam. LC OLC Olax madagascariensis (DC.) Valeton LC ORC Aerangis citrata (Thouars) Schltr. LC ORC Aeranthes ecalcarata H.Perrier LC ORC Bulbophyllum septatum Schltr. LC PHY Physena madagascariensis Tul. LC PIP Peperomia terebinthina G.Mathieu LC PUT Drypetes ambigua Leandri LC PUT Drypetes madagascariensis (Lam.) Humbert & Leandri LC RUB Breonia perrieri Homolle LC RUB Breonia stipulata Havil. LC RUB Gaertnera arenaria Baker LC RUB Gaertnera obovata Baker LC RUT Vepris nitida (Baker) I.Verd. LC SAP Tina chapelieriana (Cambess.) Kalkman LC SPS Rhopalocarpus alternifolius (Baker) Capuron LC THY Gnidia daphnifolia L.f. LC THY Stephanodaphne geminata H.Perrier ex Leandri LC BLS Impatiens malcomberi Eb.Fisch. & Raheliv. DD ORC Aeranthes moratii Bosser DD ORC Bulbophyllum vulcanorum H.Perrier DD ORC Habenaria saprophytica Bosser & P.J.Cribb DD PUT Drypetes capuronii Leandri DD
LÉGENDE DES CATÉGORIES IUCN : CR : EN DANGER CRITIQUE D’EXTINCTION, EN : EN DANGER, VU :
VULNÉRABLE, NT : QUASI MENACÉ, LC : PRÉOCCUPATION MINEURE, DD : DONNÉES INSUFFISANTES. (IUCN (2001).

8. Annexes
196
8.4. LISTE DES ESPÈCES SITUÉES EN BORDURE DES LACS DU MASSIF
Familles Espece Répartition géographique AMA Achyranthes bidentata Blume M [Sb, C] - RM API Agrocharis melanantha Hochst. M [C, HM] - Aft API Centella asiatica (L.) Urb. M [E, C, W] - RM, Aft, As, Amt AST Acmella caulirhiza Delile M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt AST Bidens pilosa L. M [Sb, E, C, W] - RM, Aft, As, Oc, Amt AST Chrysanthellum americanum (L.) Vatke M [C, W] - Aft, As, Amt AST Crassocephalum crepidioides (Benth.) S.Moore M [Sb, E, C, W] - RM, Aft AST Dichrocephala latifolia (Poir.) DC. M [E, C] - Aft, As AST Emilia bathiei Humbert M [W(N)] AST Pseudognaphalium luteoalbum (L.) Hilliard & B.L.Burtt M [C] - RM, Aft, As, Oc, Amt AST Sonchus oleraceus L. M [C, S] - RM, Aft, As, Oc, Amt BOR Cynoglossum lanceolatum Forssk. M [Sb, E, C, W(S)] - RM, Aft, As BRA Cardamine trichocarpa Hochst. ex A.Rich. M [MdA] - Aft, As CMM Commelina diffusa Burm. f. M [Sb, E, C, W(S)] - RM, Aft, Amt CRY Drymaria cordata (L.) Willd. ex Roem. & Schult. M [E, C, W(S)] - RM, Aft, As, Oc, Amt CRY Stellaria media (L.) Vill. M [C] - RM, Aft, As, Oc, Amt, Eur CYP Actinoschoenus thouarsii (Kunth) Benth. M [Sb, E] - RM CYP Carex sphaerogyna Baker M [C, HM] CYP Cyperus distans L.f. M [Sb, E, C, W(N)] - RM, Aft, As, Oc, Amt CYP Cyperus haspan L. M [Sb, E, C, W(S)] - RM, Aft, As, Oc, Amt CYP Cyperus prolifer Lam. M [Sb, E, C, W(S)] - RM, Aft CYP Fimbristylis squarrosa Vahl M [Sb, E, C, W] - Aft, As, Oc, Amt, Eur CYP Kyllinga brevifolia Rottb. M [C] - Aft, As, Amt CYP Pycreus mundtii Cherm. M [E, C, W(S), S] - RM, Aft, Eur
CYP Schoenoplectus corymbosus (Roem. & Schult.) J. Raynal M [C] - Aft
EQU Equisetum ramosissimum Desf. M [C] - RM, Aft, As, Oc, Amt, Eur EUP Croton goudotii Baill. M [C, HM] LAM Ajuga oocephala Baker M [C] LAM Pycnostachys coerulea Hook. M [C] - Aft LAU Ocotea racemosa (P. Danguy) Kosterm. M [E, C] LID Lindernia rotundifolia (L.) Alston M [C] - RM, Aft MLS Medinilla ambrensis Jum. & H.Perrier M [MdA] MRS Maesa lanceolata Forssk. M [Sb, E, C, W] - RM, Afr NYM Nymphaea nouchali Burm.f. M [E, W] - Aft ORC Cynorkis stolonifera (Schltr.) Schltr. M [C] ORC Oberonia disticha (Lam.) Schltr. M [Sb, E, C] - RM, Aft PIP Piper guineense Schumach. & Thonn. M [C] - Aft PLG Persicaria minor (Huds.) Opiz M [C, W(S)] - As, Eur PLG Persicaria nepalensis (Meisn.) H.Gross M [C] - Aft, As PLG Rumex nepalensis Spreng. M [C] - Aft, As POA Digitaria debilis (Desf.) Willd. M [Sb, E, C, W] - Aft POA Digitaria thouaresiana (Flüggé) A.Camus M [E] - RM, Aft POA Sporobolus centrifugus (Trin.) Nees M [C] - Aft POA Stenotaphrum dimidiatum (L.) Brongn. M [E, C] - RM, Aft
POT Potamogeton nodosus Poir. M [Sb, E, C, W, S] - RM, Aft, As, Oc, Amt, Eur
RUB Oldenlandia goreensis (DC.) Summerh. M [E, C] - RM, Aft RUB Spermacoce tenuior L. M [Sb, E, C] - Amt
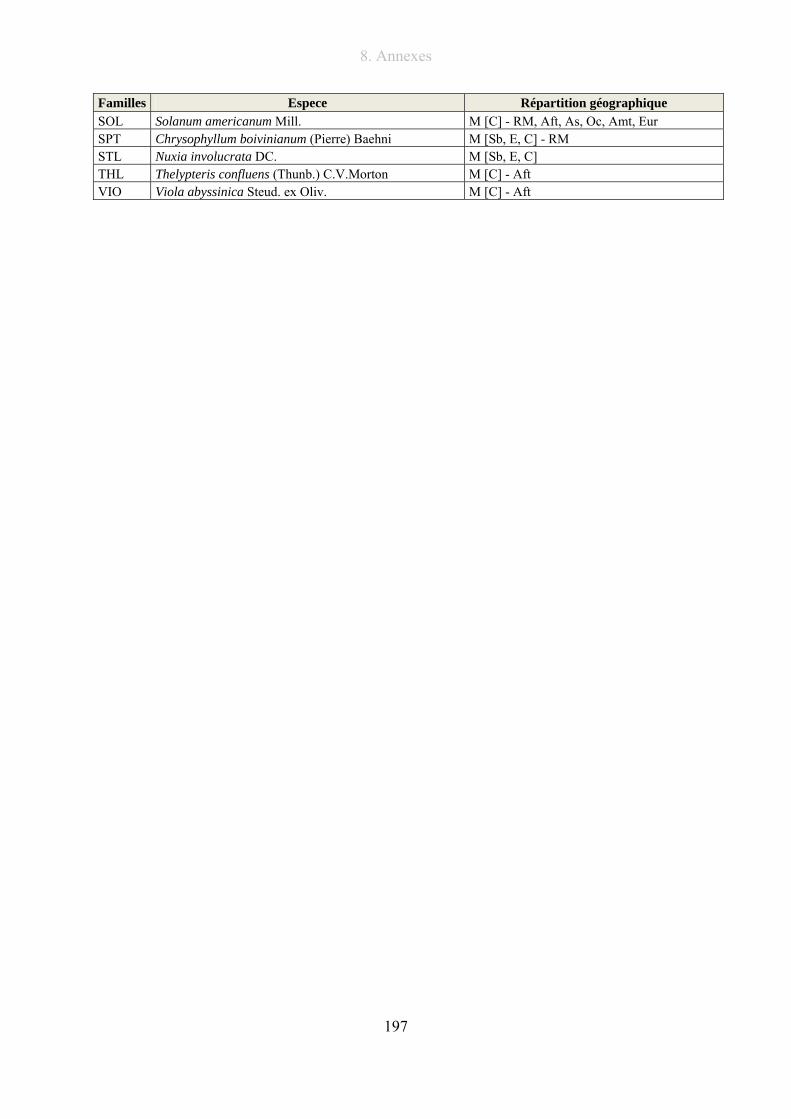
8. Annexes
197
Familles Espece Répartition géographique SOL Solanum americanum Mill. M [C] - RM, Aft, As, Oc, Amt, Eur SPT Chrysophyllum boivinianum (Pierre) Baehni M [Sb, E, C] - RM STL Nuxia involucrata DC. M [Sb, E, C] THL Thelypteris confluens (Thunb.) C.V.Morton M [C] - Aft VIO Viola abyssinica Steud. ex Oliv. M [C] - Aft
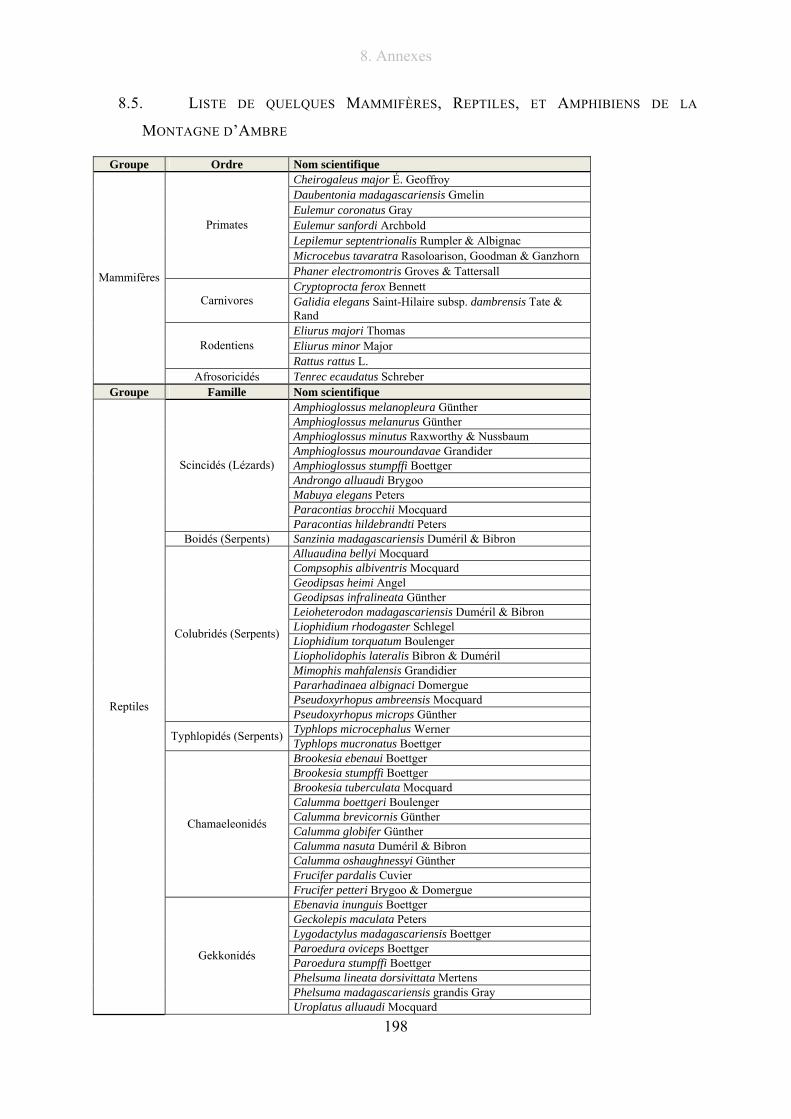
8. Annexes
198
8.5. LISTE DE QUELQUES MAMMIFÈRES, REPTILES, ET AMPHIBIENS DE LA
MONTAGNE D’AMBRE
Groupe Ordre Nom scientifique Cheirogaleus major É. Geoffroy Daubentonia madagascariensis Gmelin Eulemur coronatus Gray Eulemur sanfordi Archbold Lepilemur septentrionalis Rumpler & Albignac Microcebus tavaratra Rasoloarison, Goodman & Ganzhorn
Primates
Phaner electromontris Groves & Tattersall Cryptoprocta ferox Bennett
Carnivores Galidia elegans Saint-Hilaire subsp. dambrensis Tate & Rand Eliurus majori Thomas Eliurus minor Major Rodentiens Rattus rattus L.
Mammifères
Afrosoricidés Tenrec ecaudatus Schreber Groupe Famille Nom scientifique
Amphioglossus melanopleura Günther Amphioglossus melanurus Günther Amphioglossus minutus Raxworthy & Nussbaum Amphioglossus mouroundavae Grandider Amphioglossus stumpffi Boettger Androngo alluaudi Brygoo Mabuya elegans Peters Paracontias brocchii Mocquard
Scincidés (Lézards)
Paracontias hildebrandti Peters Boidés (Serpents) Sanzinia madagascariensis Duméril & Bibron
Alluaudina bellyi Mocquard Compsophis albiventris Mocquard Geodipsas heimi Angel Geodipsas infralineata Günther Leioheterodon madagascariensis Duméril & Bibron Liophidium rhodogaster Schlegel Liophidium torquatum Boulenger Liopholidophis lateralis Bibron & Duméril Mimophis mahfalensis Grandidier Pararhadinaea albignaci Domergue Pseudoxyrhopus ambreensis Mocquard
Colubridés (Serpents)
Pseudoxyrhopus microps Günther Typhlops microcephalus Werner Typhlopidés (Serpents) Typhlops mucronatus Boettger Brookesia ebenaui Boettger Brookesia stumpffi Boettger Brookesia tuberculata Mocquard Calumma boettgeri Boulenger Calumma brevicornis Günther Calumma globifer Günther Calumma nasuta Duméril & Bibron Calumma oshaughnessyi Günther Frucifer pardalis Cuvier
Chamaeleonidés
Frucifer petteri Brygoo & Domergue Ebenavia inunguis Boettger Geckolepis maculata Peters Lygodactylus madagascariensis Boettger Paroedura oviceps Boettger Paroedura stumpffi Boettger Phelsuma lineata dorsivittata Mertens Phelsuma madagascariensis grandis Gray
Reptiles
Gekkonidés
Uroplatus alluaudi Mocquard

8. Annexes
199
Groupe Famille Nom scientifique Uroplatus cf. ebenaui Boettger Uroplatus fimbriatus Schneider Gekkonidés Uroplatus sikorae Boettger
Reptiles
Gerrhosauridés Zonosaurus haraldmeieri Brygoo & Böhme Cophyla phyllodactyla Boettger Heterixalus betsileo Grandider Platypelis grandis Boulenger Plethodontohyla laevipes Mocquard Stumpffia gimmeli Glaw & Vence
Hyperoliidés
Stumpffia grandis Guibé Aglyptodactylus madagascariensis Duméril Boophis blommersae Glaw & Vence Boophis brachychir Boettger Boophis luteus septentrionalis Glaw & Vence Boophis madagascariensis Peters Mantidactylus ambohitra Vences & Glaw Mantidactylus ambreensis Mocquard Mantidactylus asper Boulenger Mantidactylus bicalcaratus Boettger Mantidactylus curtus Boulenger Mantidactylus femoralis Boulenger Mantidactylus granulatus Boettger Mantidactylus horridus Boettger Mantidactylus liber Peracca Mantidactylus luteus Methuen & Hewitt Mantidactylus pseudoasper Guibé Mantidactylus wittei Guibé
Amphibiens
Ranidés
Ptychadena m. mascareniensis Duméril &Bibron Poissons Pachypanchax sakaramyi Holly





























