Embed Size (px)
Citation preview

Ventilatory response to exercise does not evidence
electroencephalographical respiratory-related activation of
the cortical premotor circuitry in healthy humans
L. Jutand,1,2 L. Tremoureux,2 A. Pichon,3 N. Delpech,1 A. Denjean,4 M. Raux,2,5
C. Straus2,6,† and T. Similowski7†
1 Universite de Poitiers, Laboratoire des Adaptations Physiologiques aux Activites Physiques, Faculte des Sciences du Sport, UPRES
EA 3813, 4 Allee Jean Monnet, 86000, Poitiers, France
2 UPMC Univ Paris 6, ER10 UPMC, 75013, Paris, France
3 Universite Paris 13, UFR SMBH, STAPS, UPRES EA 2363, Laboratoire Reponses Cellulaires et Fonctionnelles a l’Hypoxie, 74 rue
Marcel Cachin, 93017, Bobigny, France
4 Assistance Publique – Hopitaux de Paris, Hopital Robert Debre, Service de physiologie, Explorations Fonctionnelles, 75019, Paris,
France
5 Assistance Publique – Hopitaux de Paris, Groupe Hospitalier Pitie-Salpetriere, Departement d’Anesthesie-Reanimation, 75013,
Paris, France
6 Assistance Publique – Hopitaux de Paris, Groupe Hospitalier Pitie-Salpetriere, Service Central d’Explorations Fonctionnelles Respi-
ratoires, 75013, Paris, France
7 Assistance Publique – Hopitaux de Paris, Groupe Hospitalier Pitie-Salpetriere, Service de Pneumologie et Reanimation, 75013,
Paris, France.
Received 18 August 2011,
revision requested 10 October
2011,
revision received 9 February
2012,
accepted 12 February 2012
Correspondence: Dr Christian
Straus, Service Central d’Explora-
tions Fonctionnelles Respiratoires,
Groupe Hospitalier Pitie-Salpetri-
ere, 47-83 Bd de l’Hopital, 75651
Paris Cedex 13, France.
E-mail: [email protected]
†Both last authors.
Abstract
Aim: The neural structures responsible for the coupling between ventila-
tory control and pulmonary gas exchange during exercise have not been
fully identified. Suprapontine mechanisms have been hypothesized but not
formally evidenced. Because the involvement of a premotor circuitry in the
compensation of inspiratory mechanical loads has recently been described,
we looked for its implication in exercise-induced hyperpnea.Methods: Electroencephalographical recordings were performed to iden-
tify inspiratory premotor potentials (iPPM) in eight physically fit normal
men during cycling at 40 and 70% of their maximal oxygen consumption
( _VO2max). Relaxed pedalling (0 W) and voluntary sniff manoeuvres were
used as negative and positive controls respectively.Results: Voluntary sniffs were consistently associated with iPPMs. This
was also the case with voluntarily augmented breathing at rest (in three
subjects tested). During the exercise protocol, no respiratory-related
activity was observed whilst performing bouts of relaxed pedalling. Exer-
cise-induced hyperpnea was also not associated with iPPMs, except in one
subject.Conclusion: We conclude that if there are cortical mechanisms involved
in the ventilatory adaptation to exercise in physically fit humans, they are
distinct from the premotor mechanisms activated by inspiratory load com-
pensation.
Keywords cerebral cortex, control of breathing, exercise, humans.
© 2012 The AuthorsActa Physiologica © 2012 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2012.02427.x356
Acta Physiol 2012, 205, 356–362

Mammalian breathing depends on brainstem neurons
that produce the ventilatory rhythm and adjust venti-
lation to meet metabolic demands. In humans, partic-
ularly powerful suprapontine commands can disrupt
metabolic ventilation, for example during speech.
Motor cortical structures engage during volitional
breathing (Macefield & Gandevia 1991, Smejkal et al.
2000) and during certain types of inspiratory load
compensation (Raux et al. 2007a). Whether or not
they are involved in the control of breathing during
exercise is not known.
During physical exercise, ventilation must adapt to
sequential changes in the interior milieu. Early on,
exercise increases the production of carbon dioxide
(CO2): in response, ventilation augments to regulate
the arterial partial pressure of CO2. When exercise
intensifies and the lactate threshold is crossed, meta-
bolic acidosis sets in, requiring additional ventilatory
compensation. A constant work rate exercise results
in an immediate increase in ventilation that rises
exponentially until an isocapnic steady state is
reached (Whipp & Ward 1998). Recent experiments
have identified a subset of neurons within the
mesencephalic locomotor region that sends direct
inputs to neurons in the respiratory generator (Gari-
epy et al. 2012) and thus seems intimately involved in
the coupling between respiration and locomotion
(Gariepy et al. 2012). However, the neural mecha-
nisms responsible for the coupling between ventilatory
control and pulmonary gas exchange during exercise
have not been fully identified (review in Haouzi
2006). This coupling is bound to be complex, insofar
as exercise proceeds from voluntary commands acti-
vating automatic programs. Krogh and Lindhard
(1913) suggested early on that exercise-related hyper-
ventilation can depend on cortical structures, at least
in the early stages of effort. A cortical mechanism has
also been proposed to account for the increase in ven-
tilation that occurs during anticipation of exercise
and at its very onset (Tobin et al. 1986), where venti-
lation exceeds the actual exercise-related ventilatory
demand. Cerebral imaging studies have lent support
to this concept. For example, Fink et al. (1995)
observed increases in regional blood flow during exer-
cise in areas associated with volitional breathing.
During imagined exercise, Thornton et al. (2001) also
described activation in areas of the brain that relate
to voluntary breathing during imagined exercise under
hypnosis. Yet, contradictory to these elements that
support the role of a cortical input to the ventilatory
response to exercise, it has been shown that during
sinusoidal work, the ventilatory response to exercise
is dynamically linked to metabolism through carbon
dioxide production (Casaburi et al. 1977, 1978,
Haouzi et al. 1992). This suggests that the rate at
which CO2 leaves the tissues and is exchanged at the
lungs during exercise is a major determinant of exer-
cise-related hyperpnea (Haouzi et al. 2004) and that,
in contrast, cortical commands play a minor role. In
line with this, in a patient suffering from an Arnold
Chiari malformation who completely lacked a ventila-
tory response to carbon dioxide, Haouzi et al. (2000)
observed that the ventilatory response to exercise was
profoundly abnormal inspite of an apparently normal
voluntary control of breathing. Although this observa-
tion contrasted with the persistence of a ventilatory
response to aerobic exercise observed in patients with
Ondine’s curse also lacking chemosensitivity (Shea
et al. 1993), Haouzi et al. (2000) concluded that the
corticospinal respiratory pathway did not play a
major role in exercise-related ventilatory adaptation.
This case report indicates that cortical mechanisms
may not be mandatory for an appropriate ventilatory
response to exercise, but it is, however, not sufficient
to rule out cooperative mechanisms involving brain-
stem and suprapontine structures. Such cooperative
mechanisms probably account for the active compen-
sation of a mechanical, inspiratory load that occurs in
awake humans but not during sleep, as illustrated by
the presence of cortical respiratory-related premotor
potentials in such circumstances (Raux et al. 2007b,
Tremoureux et al. 2010).
We therefore set out to test the hypothesis that an
exercise-related activation of the cortical premotor
respiratory circuitry exists. To do so, we performed
inspiratory-triggered retro-averagings of the electroen-
cephalogram (Raux et al. 2007a,b) in a population of
physically active, normal volunteers during exercise.
Material and methods
Subjects
Eight physically active, male, healthy subjects (age
27 ± 5 years, _VO2max 55 ± 6 mL min�1 per kg, body
mass index 23 ± 6 kg m�2) were studied. They were
instructed to refrain from sedative intake and alcohol
consumption, to avoid sleep deprivation and to avoid
heavy exercise during the 24 h preceding the tests.
The study was approved by the appropriate ethical
committee, and all subjects gave their written,
informed consent.
Maximal oxygen consumption ( _VO2max)
_VO2max was measured in the subjects exercising on a
braked cycle ergometer (Ergoselect 100P; Ergoline,
Bitz, Germany). After a 5 min warm up at 90 W, the
load was increased by 25 W every minute until the
subject was unable to carry on despite vigorous
© 2012 The AuthorsActa Physiologica © 2012 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2012.02427.x 357
Acta Physiol 2012, 205, 356–362 L. Jutand et al. · Cortical respiratory control during exercise

encouragement. The subjects breathed through a face-
mask (Hans Rudolph, Kansas City, MO, USA) con-
nected in series to a low-resistance pneumotachograph
(Sensormedics; VIASYS Healthcare, Palm Springs, CA,
USA). Inspiratory and expiratory gas was analysed,
breath by breath, using an automated system (Vmax
Encore; Viasys Healthcare, Palm Srings, CA, USA).
Three criteria were required to be met before accept-
ing the highest _VO2 recorded: (i) _VO2 stabilization
despite an increase in exercise intensity, (ii) respiratory
exchange ratio >1.1 and (iii) attainment of the age-
predicted maximal heart rate (220 – age).
Electroencephalographical recordings (EEG)
Electroencephalographical activity was recorded with
a sterile single-use subcutaneous needle electrode
inserted into the scalp at Cz (international 10–20 sys-
tem) (12 mm, diameter 0.35 mm, ref. MF3OE1S3512;
Comepa, Bagnolet, France) with two reference elec-
trodes on the mastoids. The impedance of the EEG
electrodes was regularly checked during the experi-
ments and remained between 2 and 5 kOhms. These
electrodes were connected to an EEG preamplifier
(2000-fold gain, band-pass frequency 0.05–500 Hz;
Electronique du Mazet, Le Mazet St Voy, France) out
of which the signals were fed to an analog–digital con-
verter and digitized at a sampling frequency of
2000 Hz (ML 780 PowerLab 8s, 16 bits resolution,
maximum range ± 10 V, recording range set on ±1 V;
Chart v5.2 software; AD Instruments, Castle Hill,
Australia) with a 1000-Hz low-frequency band-pass
anti-aliasing filter, then stored as computer files for sub-
sequent analysis (same as in Tremoureux et al. 2010).
Protocol
During all the experiments, the subjects watched a
movie on a computer screen placed at the centre of
their visual field, to deflect as much as possible their
attention from the physiological recordings. They
wore a facemask connected in series to a pneumotacho-
graph (MLT 1000 L, AD Instrument, dead space
350 mL, flow resistance 0.002 cm H2O L�1 s�1).
Mouth pressure was recorded with a linear differential
transducer (DP-45-18, ±2 cmH20; Validyne, North-
ridge, CA, USA) connected to the mask. End-tidal
CO2 partial pressure (PET, CO2) was measured from
another side port of the mouthpiece, using an infrared
CO2 gas analyser (IR1505; Servomex, Plaine Saint
Denis, France).
First, voluntary sniffs were performed. Then, the
exercise protocol began with a 15-min control run
during which the subjects breathed freely, seated on
the bicycle. They then exercised at an intensity corre-
sponding to 40% of their maximal oxygen uptake
( _VO2max) during 15 min. This was followed by a sec-
ond control run consisting of 15 min cycling at
0 Watts. The final part of the experiment included a
bout of cycling for 15 min at an intensity correspond-
ing to 70% _VO2max. No instruction was given to the
subjects regarding their pedalling rate, to avoid their
focusing on the experiment.
EEG processing
This was performed according to the method previ-
ously described (Raux et al. 2007a,b). In each of the
study conditions, 80 EEG epochs of 4-s duration
(from 2.5 s before to 1.5 s after the onset of mechani-
cal inspiration defined as the point of zero flow) were
created once a steady-state _VO2 had been reached.
Epochs exhibiting obvious artifacts were discarded,
and the rest was ensemble averaged. On the averaged
tracings, an inspiratory premotor activity was sought
in the form of a slow upward shift of the EEG signal
starting between 2 and 0.5 s before inspiration. When
this occurred, a first-order least-square regression
equation was fitted to the corresponding segment. A
premotor potential was considered present if and only
if the slope of this equation was positive and signifi-
cantly different from zero according to the F-test for
equality of variance. Motor potentials, defined as an
increase in negativity synchronous with the onset of
inspiration, were looked for in a similar manner.
Statistical analysis
This was conducted with the Prism 4 (GraphPad Soft-
ware Inc., La Jolla, CA, USA) and Excel (Microsoft
Corporation, Redmond, WA, USA) software. Data
distributions being consistently normal according to a
Shapiro–Wilk, the values are reported in terms of
mean ± SD. The effects of exercise on respiratory pat-
tern data were assessed using a one-way analysis of
variance with a post hoc Newman–Keuls test. The
proportions of the subjects exhibiting a premotor
activity in the various conditions were compared with
Fisher’s exact test. The results were considered signifi-
cant when the risk of a first type error was below 5%
(P < 0.05).
Additional experiments
The absence of respiratory-related premotor potentials
during exercise in our subjects (see Results below)
raised the issue of the sensitivity of the technique. To
address this, we conducted complementary experi-
ments using exactly the same recording technique and
signal processing method in three subjects who were
© 2012 The AuthorsActa Physiologica © 2012 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2012.02427.x358
Cortical respiratory control during exercise · L. Jutand et al. Acta Physiol 2012, 205, 356–362

asked to voluntarily augment their breathing indepen-
dently of any exercise.
Results
Respiratory pattern
Exercise significantly increased minute ventilation,
breathing frequency, tidal volume and mouth pressure.
These increases were in proportion to exercise inten-
sity (Table 1). Inspiratory and expiratory times
decreased with the intensity of exercise (Table 1).
Premotor potentials
Typical inspiratory premotor potentials were present
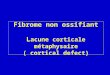
during sniff manoeuvres (Fig. 1).
No EEG changes suggestive of respiratory-related
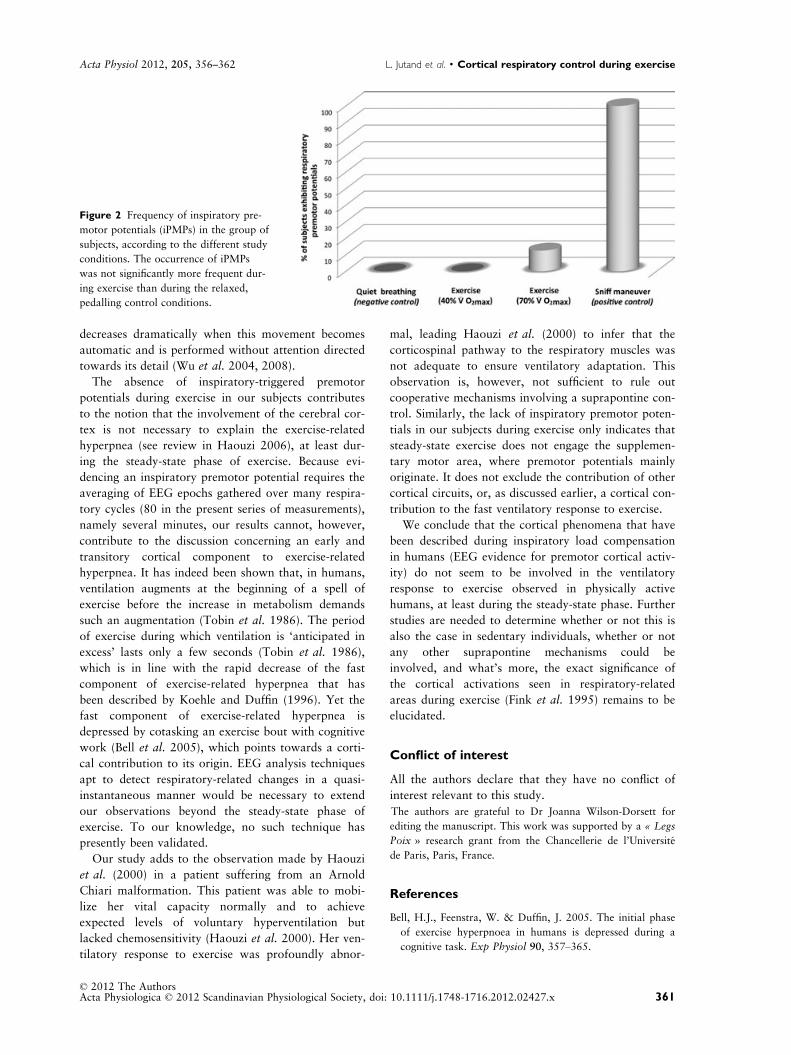
premotor activity were found during the two control
sessions in any of the subjects (Fig. 2). Except in one
subject, inspiratory premotor potentials were not iden-
tified during exercise, either at 40% _VO2max or at
70% _VO2max (Fig. 2).
The occurrence of inspiratory premotor potentials
during exercise was therefore not significantly more
frequent than during the relaxed, pedalling control
conditions (P = 0.23).
Voluntarily augmented breathing
In the three subjects tested, voluntarily augmented
breathing was associated with a clear upward depar-
ture of the Cz-EEG trace from baseline that occurred
circa 1.5 s before the start of inspiration and met all
the characteristics of a respiratory-related premotor
potential.
Discussion
In this study, healthy fit volunteers failed to exhibit
inspiratory premotor potentials during exercise-related
hyperpnea.
As in any instance of a negative result, the first
hypothesis to consider is an insufficient statistical
power, related to the small size of the study popula-
tion. However, in other experimental models involv-
ing inspiratory load compensation paradigms (Raux
et al. 2007a,b, Tremoureux et al. 2010), inspiratory
premotor potentials have been evidenced with a statis-
tically significant frequency in populations as small as
the present one. Therefore, we can reasonably con-
sider that exercise is a much less potent stimulus than
inspiratory mechanical loading to activate the respira-
tory premotor circuitry, if at all. Of note, we consis-
tently found premotor potentials during voluntary
sniff manoeuvres in our subjects, in agreement with
previous studies (Macefield & Gandevia 1991, Raux
et al. 2007b), which validates our methodology. We
also found premotor potentials before inspiration dur-
ing voluntarily augmented breathing at rest in the
three subjects who were tested in this manner. This
goes against our technique not being sensitive enough
to detect the signal if present during exercise.
One of our subjects did exhibit EEG evidence of a
respiratory-related premotor activity during exercise.
After the experiments, this subject reported that,
according to his habit, he had concentrated on breath-
ing in synchrony with his running rhythm. This proba-
bly explains why this subject deviated from the general
pattern, all the more so that he did not exhibit a pre-
motor potential during the control conditions. It is
tempting to relate this observation to the study per-
formed by Wuyam et al. (1995), during which it was
Table 1 Ventilatory pattern in the study population
Control‡ 40% _VO2max 70% _VO2max
Ventilation (L min�1) 13.8 ± 3.6 44.4 ± 5.1* 77.1 ± 10.5*†
Breathing frequency (cycle min�1) 14.4 ± 2.8 23.2 ± 2.0* 30.6 ± 3.6*†
Tidal volume (VT) (L) 0.94 ± 0.12 1.94 ± 0.25* 2.54 ± 0.36*†
Inspiratory time (TI) (s) 1.75 ± 0.31 1.18 ± 0.13* 0.92 ± 0.09*†
Expiratory time (s) 2.57 ± 0.99 1.30 ± 0.32* 1.03 ± 0.25*
TI/TT 0.39 ± 0.04 0.44 ± 0.02* 0.46 ± 0.02*
VT/TI 0.57 ± 0.15 1.67 ± 0.24* 2.76 ± 0.37*†
PETCO2 44.0 ± 4.3 48.0 ± 5.3 46.3 ± 5.2
Mouth pressure (cmH2O) �0.24 ± 0.07 �0.81 ± 0.22 �1.75 ± 0.49*
All values expressed as mean ± SD.
TT, breathing period.
*Significantly different from control.
†Significantly different from 40% _VO2max.
‡Average of the two control periods.
© 2012 The AuthorsActa Physiologica © 2012 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2012.02427.x 359
Acta Physiol 2012, 205, 356–362 L. Jutand et al. · Cortical respiratory control during exercise
shown that imagination of exercise produced signifi-
cant alterations in cardioventilatory variables in ath-
letes, but not in untrained normal subjects. Of note,
the subjects who participated in our study were all fit
and physically active. This necessitates a word of cau-
tion regarding the general validity of our observations:
we cannot exclude the hypothesis that a control group
of sedentary subjects would have behaved differently.
Indeed, training can modify cortical processes both in
general and regarding respiratory muscle control
(Demoule et al. 2008). Comparing athletes and nonath-
letes in terms of their ventilatory response to exercise,
Wuyam et al. (1995) found that only trained individu-
als exhibited breathing pattern changes and hypocap-
nia when imagining efforts. The coupling between
breathing and pedalling, for instance, is also known to
occur differently in athletes vs. nonathletes (Kohl et al.
1981). This could stem from mechanisms resembling,
or related to, ‘classical respiratory conditioning’ as it is
known to occur in humans (Gallego & Perruchet
1991). In view of this, the lack of cortical involvement
in the response to exercise of trained subjects would
not be surprising. In addition, it is known that the cor-
tical activation related to a volitional movement
Figure 1 Results of the inspiratory-triggered back averaging of the electroencephalographical (EEG) signal in one of the study
subjects. In each panel, the top trace is the mouth pressure signal, and the bottom trace is the EEG signal. The top panel
corresponds to a positive control experiment (voluntary sniffing), showing the clear presence of a premotor potential (horizontal
double arrow) starting about 0.9 s before inspiration (vertical line) and followed by a clearly visible motor potential attesting to
a cortical contribution to the inspiratory command. On the middle row, the left panel corresponds to the pre-exercise control
period, whereas the right panel corresponds to exercise at 40% _VO2max. On the bottom row, the left panel corresponds to the
post-exercise control period (following immediately the previous exercise bout), whilst the right one corresponds to exercise at
70% _VO2max. In these four cases, there is no indication of a pre-inspiratory EEG activity and no motor potential.
© 2012 The AuthorsActa Physiologica © 2012 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2012.02427.x360
Cortical respiratory control during exercise · L. Jutand et al. Acta Physiol 2012, 205, 356–362
decreases dramatically when this movement becomes
automatic and is performed without attention directed
towards its detail (Wu et al. 2004, 2008).
The absence of inspiratory-triggered premotor
potentials during exercise in our subjects contributes
to the notion that the involvement of the cerebral cor-
tex is not necessary to explain the exercise-related
hyperpnea (see review in Haouzi 2006), at least dur-
ing the steady-state phase of exercise. Because evi-
dencing an inspiratory premotor potential requires the
averaging of EEG epochs gathered over many respira-
tory cycles (80 in the present series of measurements),
namely several minutes, our results cannot, however,
contribute to the discussion concerning an early and
transitory cortical component to exercise-related
hyperpnea. It has indeed been shown that, in humans,
ventilation augments at the beginning of a spell of
exercise before the increase in metabolism demands
such an augmentation (Tobin et al. 1986). The period
of exercise during which ventilation is ‘anticipated in
excess’ lasts only a few seconds (Tobin et al. 1986),
which is in line with the rapid decrease of the fast
component of exercise-related hyperpnea that has
been described by Koehle and Duffin (1996). Yet the
fast component of exercise-related hyperpnea is
depressed by cotasking an exercise bout with cognitive
work (Bell et al. 2005), which points towards a corti-
cal contribution to its origin. EEG analysis techniques
apt to detect respiratory-related changes in a quasi-
instantaneous manner would be necessary to extend
our observations beyond the steady-state phase of
exercise. To our knowledge, no such technique has
presently been validated.
Our study adds to the observation made by Haouzi
et al. (2000) in a patient suffering from an Arnold
Chiari malformation. This patient was able to mobi-
lize her vital capacity normally and to achieve
expected levels of voluntary hyperventilation but
lacked chemosensitivity (Haouzi et al. 2000). Her ven-
tilatory response to exercise was profoundly abnor-
mal, leading Haouzi et al. (2000) to infer that the
corticospinal pathway to the respiratory muscles was
not adequate to ensure ventilatory adaptation. This
observation is, however, not sufficient to rule out
cooperative mechanisms involving a suprapontine con-
trol. Similarly, the lack of inspiratory premotor poten-
tials in our subjects during exercise only indicates that
steady-state exercise does not engage the supplemen-
tary motor area, where premotor potentials mainly
originate. It does not exclude the contribution of other
cortical circuits, or, as discussed earlier, a cortical con-
tribution to the fast ventilatory response to exercise.
We conclude that the cortical phenomena that have
been described during inspiratory load compensation
in humans (EEG evidence for premotor cortical activ-
ity) do not seem to be involved in the ventilatory
response to exercise observed in physically active
humans, at least during the steady-state phase. Further
studies are needed to determine whether or not this is
also the case in sedentary individuals, whether or not
any other suprapontine mechanisms could be
involved, and what’s more, the exact significance of
the cortical activations seen in respiratory-related
areas during exercise (Fink et al. 1995) remains to be
elucidated.
Conflict of interest
All the authors declare that they have no conflict of
interest relevant to this study.
The authors are grateful to Dr Joanna Wilson-Dorsett for
editing the manuscript. This work was supported by a « Legs
Poix » research grant from the Chancellerie de l’Universite
de Paris, Paris, France.
References
Bell, H.J., Feenstra, W. & Duffin, J. 2005. The initial phase
of exercise hyperpnoea in humans is depressed during a
cognitive task. Exp Physiol 90, 357–365.
Figure 2 Frequency of inspiratory pre-
motor potentials (iPMPs) in the group of
subjects, according to the different study
conditions. The occurrence of iPMPs
was not significantly more frequent dur-
ing exercise than during the relaxed,
pedalling control conditions.
© 2012 The AuthorsActa Physiologica © 2012 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2012.02427.x 361
Acta Physiol 2012, 205, 356–362 L. Jutand et al. · Cortical respiratory control during exercise

Casaburi, R., Whipp, B.J., Wasserman, K., Beaver, W.L. &
Koyal, S.N. 1977. Ventilatory and gas exchange dynamics
in response to sinusoidal work. J Appl Physiol 42, 300–
301.
Casaburi, R., Whipp, B.J., Wasserman, K. & Koyal, S.N.
1978. Ventilatory and gas exchange responses to cycling
with sinusoidally varying pedal rate. J Appl Physiol 44, 97
–103.
Demoule, A., Verin, E., Montcel, S.T. & Similowski, T.
2008. Short-term training-dependent plasticity of the corti-
cospinal diaphragm control in normal humans. Respir Phy-
siol Neurobiol 160, 172–180.
Fink, G.R., Adams, L., Watson, J.D., Innes, J.A., Wuyam, B.,
Kobayashi, I., Corfield, D.R., Murphy, K., Jones, T. &
Frackowiak, R.S. 1995. Hyperpnoea during and immedi-
ately after exercise in man: evidence of motor cortical
involvement. J Physiol 489, 663–675.
Gallego, J. & Perruchet, P. 1991. Classical conditioning of
ventilatory responses in humans. J Appl Physiol 70, 676–
682.
Gariepy, J.F., Missaghi, K., Chevallier, S., Chartre, S., Rob-
ert, M., Auclair, F., Lund, J.P. & Dubuc, R. 2012. Specific
neural substrate linking respiration to locomotion. Proc
Natl Acad Sci USA 109, E84–E92.
Haouzi, P. 2006. Theories on the nature of the coupling
between ventilation and gas exchange during exercise.
Respir Physiol Neurobiol 151, 267–279.
Haouzi, P., Fukuba, Y., Peslin, R., Chalon, B., Marchal, F.
& Crance, J.P. 1992. Ventilatory dynamics in children and
adults during sinusoidal exercise. Eur J Appl Physiol
Occup Physiol 64, 410–418.
Haouzi, P., Marchal, J., Allioui, E.M., Hannhart, B., Chalon,
B. & Braun, M. 2000. Corticospinal pathway and exercise
hyperpnea: lessons from a patient with Arnold Chiari mal-
formation. Respir Physiol 123, 13–22.
Haouzi, P., Chenuel, B. & Chalon, B. 2004. Frequency
response of the input reaching the respiratory centres dur-
ing moderate intensity exercise. Adv Exp Med Biol 551,
287–290.
Koehle, M. & Duffin, J. 1996. The effect of exercise duration
on the fast component of exercise hyperpnoea at work
rates below the first ventilatory threshold. Eur J Appl
Physiol Occup Physiol 74, 548–552.
Kohl, J., Koller, E.A. & Jager, M. 1981. Relation between
pedalling- and breathing rhythm. Eur J Appl Physiol
Occup Physiol 47, 223–237.
Krogh, A. & Lindhard, J. 1913. The regulation of respiration
and circulation during the initial stages of muscular work.
J Physiol 47, 112–136.
Macefield, G. & Gandevia, S.C. 1991. The cortical drive to
human respiratory muscles in the awake state assessed by
premotor cerebral potentials. J Physiol 439, 545–558.
Raux, M., Ray, P., Prella, M., Duguet, A., Demoule, A. &
Similowski, T. 2007a. Cerebral cortex activation during
experimentally induced ventilator fighting in normal
humans receiving noninvasive mechanical ventilation.
Anesthesiology 107, 746–755.
Raux, M., Straus, C., Redolfi, S., Morelot-Panzini, C., Cou-
turier, A., Hug, F. & Similowski, T. 2007b. Electroenceph-
alographic evidence for pre-motor cortex activation during
inspiratory loading in humans. J Physiol 578, 569–578.
Shea, S.A., Andres, L.P., Shannon, D.C. & Banzett, R.B.
1993. Ventilatory responses to exercise in humans lacking
ventilatory chemosensitivity. J Physiol 468, 623–640.
Smejkal, V., Druga, R. & Tintera, J. 2000. Brain activation during
volitional control of breathing. Physiol Res 49, 659–663.
Thornton, J.M., Guz, A., Murphy, K., Griffith, A.R., Peder-
sen, D.L., Kardos, A., Leff, A., Adams, L., Casadei, B. &
Paterson, D.J. 2001. Identification of higher brain centres
that may encode the cardiorespiratory response to exercise
in humans. J Physiol 533, 823–836.
Tobin, M.J., Perez, W., Guenther, S.M., D’Alonzo, G. &
Dantzker, D.R. 1986. Breathing pattern and metabolic
behavior during anticipation of exercise. J Appl Physiol
60, 1306–1312.
Tremoureux, L., Raux, M., Jutand, L. & Similowski, T.
2010. Sustained preinspiratory cortical potentials during
prolonged inspiratory threshold loading in humans. J Appl
Physiol 108, 1127–1133.
Whipp, B.J. & Ward, S.A. 1998. Determinants and control
of breathing during muscular exercise. Br J Sports Med 32,
199–211.
Wu, T., Kansaku, K. & Hallett, M. 2004. How self-initiated
memorized movements become automatic: a functional
MRI study. J Neurophysiol 91, 1690–1698.
Wu, T., Chan, P. & Hallett, M. 2008. Modifications of the
interactions in the motor networks when a movement
becomes automatic. J Physiol 586, 4295–4304.
Wuyam, B., Moosavi, S.H., Decety, J., Adams, L., Lansing,
R.W. & Guz, A. 1995. Imagination of dynamic exercise
produced ventilatory responses which were more apparent
in competitive sportsmen. J Physiol 482(Pt 3), 713–724.
© 2012 The AuthorsActa Physiologica © 2012 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2012.02427.x362
Cortical respiratory control during exercise · L. Jutand et al. Acta Physiol 2012, 205, 356–362