Embed Size (px)
Citation preview

20
LES COMPARTIMENTS DES CELLULES EUCARYOTES
Contrairement aux bactéries, les cellules eucaryotes sont organisées en
compartiments limités par une membrane et fonctionnellement distincts. Les
protéines jouent un rôle essentiel dans la compartimentation d'une cellule eucaryote.
Elles catalysent les réactions qui se produisent dans chaque compartiment (organite).
Elles servent aussi de marqueurs spécifiques de l'organite. La synthèse des protéines
commence dans le cytosol, puis chaque molécule néo synthétisée est ensuite
transmise spécifiquement au compartiment cellulaire qui en a besoin.
I. Généralités.
A. Les différents compartiments des cellules eucaryotes.
Les principaux compartiments des cellules eucaryotes sont les suivants (Figure 1) :
- le noyau, qui contient le génome et où se déroule la synthèse de l'ADN et de
l'ARN ;
- le cytoplasme, qui l'entoure, est composé du cytosol et des organites
cytoplasmiques qui y sont en suspension ;
- le cytosol représente environ la moitié du volume total de la cellule ; il est le site
de la synthèse des protéines et du métabolisme cellulaire intermédiaire ;
- le réticulum endoplasmique constitue un espace labyrinthique limité par une
membrane, dont la surface représente environ 50 % de la surface membranaire totale
de la cellule. Des ribosomes peuvent être liés à la surface cytosolique du réticulum
endoplasmique ; ces ribosomes interviennent dans la synthèse des protéines
membranaires et solubles (présentes dans la lumière des différents organites ou
destinées à être sécrétées) ;
- l'appareil de Golgi est composé d'un empilement d'organites en forme de
disques appelés saccules. Il intervient dans la maturation des protéines et des lipides
provenant du réticulum endoplasmique ;
- les mitochondries sont responsables de la production d'énergie ;

21
- les lysosomes sont des organites riches en enzymes hydrolytiques qui
dégradent les organites intracellulaires arrivés au terme de leur vie ainsi que les
molécules prélevées dans le milieu extracellulaire par endocytose ;
- les peroxysomes sont de petits organites vésiculaires utilisés pour un certain
nombre de réactions oxydatives ;
- la cellule contient également de nombreuses vésicules qui servent de
transporteurs entre les organites ou qui communiquent avec la membrane plasmique
lors du processus d'endocytose ou d'exocytose.
Compartiments intracellulaires
(hépatocyte)
% du volume
total de la cellule
Nombre /
cellule
Cytosol
Mitochondries
Réticulum endoplasmique rugueux
Réticulum endoplasm. lisse + Golgi
Noyau
Peroxysomes
Lysosomes
Endosomes
54
22
9
6
6
1
1
1
1
1700
1
1
1
400
300
200

22
B. Etude de la circulation protéique intracellulaire par autoradiographie.
Cette méthode a été développée dans les années 60 pour étudier la voie que
suivent les protéines sécrétées par les cellules du pancréas dans le milieu
extracellulaire. L'avantage de cette cellule est que la plus grande partie des protéines
synthétisées est destinée à l'exportation dans le milieu extracellulaire (amylase,
lipase, désoxyribonucléase, trypsinogène, chymotrypsinogène).
Le principe de la méthode est le suivant : les cellules sont brièvement incubées en
présence d'un acide aminé radioactif (tritium : 3H) qui est éliminé par lavage. Ainsi,
après lavage, la seule radioactivité qui reste dans les cellules est celle qui a été
incorporée dans les protéines nouvellement synthétisées durant la période
d'exposition à l'isotope. Les cellules sont ensuite incubées pendant des temps variés,
puis fixées et enfin recouvertes d'une émulsion photographique. La désintégration
du tritium active les grains d'argent qui vont apparaître sous la forme de points
noirs, visibles en microscopie électronique. Ainsi, en fonction du temps, les grains
d'argent activés apparaissent successivement :
- sur le réticulum endoplasmique à 3 min,
- sur l'appareil de Golgi à 20 min,
- dans des vésicules de sécrétion immatures de taille importante situées près du
Golgi à 90 min,
- et enfin dans des vésicules de sécrétion matures où les protéines sont
concentrées.
Figure 1.

23
De nombreuses protéines sécrétées dans le milieu extracellulaire ont été étudiées
sur d'autres types cellulaires où elles suivent toutes une voie identique :
ribosome ⇒⇒⇒⇒ réticulum endoplasm. ⇒⇒⇒⇒ Golgi ⇒⇒⇒⇒ vésicule secrétoire
⇒⇒⇒⇒ milieu extracellulaire.
C. Les différentes voies de la circulation protéique.
Toutes les protéines prennent naissance sur les ribosomes du cytosol et, de là, sont
dirigées vers 2 embranchements principaux (Figure 2).
Dans le premier embranchement, les protéines sont initialement libérées dans le
cytosol après leur synthèse. La majorité de ces protéines va rester dans le cytosol,
tandis que d'autres sont exportées vers les mitochondries, le noyau ou les
peroxysomes. Le passage des protéines au travers de la membrane des
mitochondries ou des peroxysomes se fait grâce à une protéine membranaire de
translocation. Le passage des protéines dans le noyau se fait, quant à lui, au travers
de pores nucléaires qui ont une perméabilité sélective.
Dans le deuxième embranchement, les protéines sont initialement transférées dans
le réticulum endoplasmique. Le transfert de ces protéines dans le réticulum
Figure 2.

24
endoplasmique se fait après fixation, sur le réticulum, des ribosomes qui synthétisent
ces protéines. La translocation nécessite une protéine de transport. Les protéines
transloquées peuvent rester dans le réticulum endoplasmique, ou bien être dirigées
vers l'appareil de Golgi. Dans l'appareil de Golgi, les protéines peuvent là encore
rester dans ce compartiment, ou être dirigées soit vers les lysosomes, soit vers la
membrane plasmique.
Le transfert des protéines à partir du réticulum vers les autres organites (Golgi,
lysosomes) et la membrane plasmique se fait par l'intermédiaire de vésicules de
transport. Il en est de même en ce qui concerne le transport des protéines d'un
saccule golgien à un autre. Dans ce type de transport, les vésicules se forment par
bourgeonnement du compartiment donneur. Au cours de ce processus, les vésicules
ainsi formées s'emparent du matériel présent dans le compartiment donneur et le
libèrent dans le compartiment cible, après fusion de la membrane vésiculaire avec
celle du compartiment cible (Figure 3).
Dans un organite donné, les protéines peuvent se trouver sous forme soluble dans
la lumière du compartiment, ou bien être intégrées dans sa membrane. Dans un
Figure 3.

25
transport de type vésiculaire : les protéines sécrétées dans le milieu extracellulaire et
celles présentes dans la lumière des saccules golgiens ou des lysosomes proviennent
à l'origine de protéines libérées dans la lumière du réticulum endoplasmique ; les
protéines intégrées dans la membrane de ces organites (Golgi, lysosomes), ou dans la
membrane plasmique, proviennent de protéines ancrées dans la membrane du
réticulum endoplasmique.
Le tri des protéines vers les différents embranchements doit être parfaitement
sélectif. Le mécanisme de tri est complexe et dépend en partie de signaux de tri (ou
d'adressage) présents dans les protéines, qui sont reconnus par des protéines
réceptrices spécifiques. Une protéine à destinée nucléaire possède un signal de tri qui
est reconnu par une protéine réceptrice associée au complexe du pore nucléaire. Une
protéine qui doit être transférée au travers d'une membrane possède un signal
d'adressage qui est reconnu par une protéine de translocation située dans la
membrane. Enfin, les protéines qui doivent être retenues dans certains organites ou
encore être transportées dans des vésicules de transport possèdent également des
signaux de tri spécifiques reconnus par des protéines membranaires.
D. Les signaux de tri.
On distingue 2 types de signaux de tri, les peptides signal et les régions signal.
1. Les peptides signal (Figure 4).
Ils sont constitués d'une séquence continue d'acides aminés (15 à 60 résidus).
Lorsque le processus de tri est terminé, ces peptides signal sont le plus souvent clivés
de la protéine mature par une signal peptidase. Les peptides signal sont utilisés pour
le transport des protéines du cytosol vers le réticulum endoplasmique, les
mitochondries, les peroxysomes et le noyau, et sont aussi impliqués dans la rétention
des protéines dans le réticulum endoplasmique.
Séquences typiques de peptides signal :
- Importation dans le réticulum endoplasmique : peptide situé à l'extrémité amino-
terminale de la protéine, comportant une séquence centrale de 5 à 10 acides aminés
hydrophobes.

26
Segments contribuant à la région signal
- Maintien dans le réticulum endoplasmique : séquence de 4 acides aminés
spécifiques à l'extrémité carboxy-terminale de la protéine.
- Importation dans la matrice mitochondriale : peptides signal situés à l'extrémité
amino-terminale de la protéine, possédant des acides aminés chargés positivement
(Lys, Arg) et alternant avec des acides aminés hydrophobes.
- Importation dans le noyau : séquence d'acides aminés chargés positivement au
sein de la protéine.
L'importance de ces peptides signal a été démontrée par des expériences de
recombinaison génétique, à l'aide de protéines de fusion . En effet, si l'on place le
peptide signal d'importation dans le réticulum endoplasmique à l'extrémité amino-
terminale d'une protéine cytosolique, on observe que cette protéine est
alors transportée dans le réticulum endoplasmique.
2. Les régions signal (Figure 4).
Elles correspondent à un arrangement tridimensionnel particulier de la molécule.
Les acides aminés qui forment une région signal peuvent être très distants les uns des
autres sur la molécule déployée. De telles régions signal sont impliquées dans le
transport des protéines du Golgi vers les lysosomes.
E. Un exemple de mécanisme de tri : le transport des protéines vers les
mitochondries.
Chez l’homme, 13 protéines seulement sont codées par le génome mitochondrial.
La majorité des protéines mitochondriales (1000 à 1500) est codée dans l’ADN du
Figure 4.

27
noyau, synthétisée sous forme précurseur dans le cytosol et acheminée en 1 à 2 min
dans la mitochondrie. L’importation des protéines mitochondriales est donc post-
traductionnelle. Ces protéines ont plusieurs localisations finales : la membrane
mitochondriale externe (MME), l’espace intermembranaire, la membrane
mitochondriale interne (MMI) ou la matrice mitochondriale. Elles ont un signal
d’adressage différent selon leur localisation dans la mitochondrie. Le mieux connu
est le peptide signal des protéines destinées à la matrice mitochondriale ou de
certaines protéines ancrées dans la MMI qui est constitué d’acides aminés chargés
positivement (Lys, Arg) alternant avec des acides aminés hydrophobes et situé à
l’extrémité amino-terminale de la protéine. Beaucoup de protéines destinées à la
mitochondrie comporte un peptide signal interne à la molécule.
Le processus d'importation des protéines mitochondriales comporte plusieurs
étapes (Figure 5). 1) La protéine mitochondriale nouvellement synthétisée s'associe
rapidement à des protéines de choc thermique cytosoliques (famille hsp 70). Les
protéines de choc thermique (également appelées hsp = heat shock protein ou
protéines chaperonnes) sont généralement impliquées dans le contrôle de la
conformation, c'est à dire de l'état de repliement des protéines auxquelles elles
s'associent. Leur association aux protéines mitochondriales permet le maintien de
celles-ci dans une conformation dépliée qui facilite leur transport dans la
mitochondrie au travers des systèmes de translocation. 2) Toutes les protéines
mitochondriales codées dans l’ADN nucléaire se fixent par leur signal d’adressage à
un récepteur spécifique situé dans la MME, intégré dans le complexe protéique
TOM (Translocator of Outer Membrane). TOM participe à la translocation de toutes
les protéines dans la mitochondrie et assure l’insertion des protéines
transmembranaires de la MME, avec, pour certaines d’entre elles, l’intervention d’un
deuxième complexe protéique de la MME, SAM. 3) Lors de leur translocation, les
protéines se dissocient des protéines chaperonnes cytosoliques par un processus qui
nécessite l'hydrolyse de l'ATP. 4) La translocation des précurseurs protéiques dans la
matrice mitochondriale s’effectue grâce au complexe protéique TIM23 (Translocator
of Inner Membrane) présent dans la MMI. Le transfert de la protéine au travers de la

28
membrane mitochondriale interne (complexe TIM23) fait intervenir le gradient
électrochimique d’ions H+ et les protéines hsp 70 mitochondriales. Ces protéines
chaperonnes sont liées à TIM23, se fixent sur la protéine dès son apparition dans la
matrice et participent à son entrée dans la matrice mitochondriale grâce à l’énergie
fournie par l’hydrolyse de l’ATP. Dans la matrice, une signal peptidase clive le
peptide signal amino-terminal, libérant une protéine mature. La protéine
mitochondriale s'associe immédiatement à d'autres protéines chaperonnes (hsp 60)
qui la maintiennent dans une conformation adéquate permettant d'assurer leur
insertion dans les complexes multiprotéiques du cycle de Krebs situés dans la
matrice. 5) L’insertion des protéines dans la MMI fait intervenir le complexe
protéique TIM22 ou le complexe TIM23. Les protéines de la MMI qui utilisent
TIM23 pour leur insertion ont, après le peptide signal amino-terminal d’importation
vers la matrice, une séquence d’acides aminés hydrophobes qui interrompt leur
transfert au travers de TIM23. Le reste de la molécule passe dans l’espace
intermembranaire et le peptide signal amino-terminal est clivé. La séquence
hydrophobe sort alors de TIM23 et ancre la protéine dans la MMI. Si ce second
peptide hydrophobe est clivé, la protéine est alors libérée en totalité dans l’espace
intermembranaire. 6) Le complexe OXA est principalement utilisé pour ancrer les
protéines synthétisées dans la mitochondrie dans la MMI.
II. Le cytosol.
Le cytosol occupe la moitié environ du volume de la cellule. Il est le siège du
métabolisme intermédiaire (cf. cours de Biochimie) et contient les ribosomes qui sont
responsables de la synthèse des protéines.
A. Composition.
Le cytosol est très riche en eau (85 %). Il contient un grand nombre de molécules
(sucres, acides aminés, acides gras...) de provenance diverse : i) captées du milieu
extracellulaire par diffusion passive ou grâce à des perméases ; ii) issues de la
digestion dans les lysosomes de macromolécules endocytées ou phagocytées ; iii)
provenant du métabolisme qui se déroule dans le cytosol. Certaines molécules sont

29
Figure 5.
stockées dans le cytosol : les sucres sous forme de glycogène, les lipides sous forme
d'inclusions lipidiques. Le cytosol contient de nombreux filaments protéiques
organisés en filaments qui forment le cytosquelette. Le cytosquelette donne sa forme
à la cellule et permet des mouvements cytoplasmiques cohérents.
B. Propriétés.
Les protéines représentent 20 % du poids du cytosol et sont liées les unes aux
autres par des forces d'intensité variable. Pour cette raison, le cytosol a une
consistance hétérogène, fluide ou gélatineuse. Toutefois, il a été montré que les
petites molécules et les petites protéines diffusaient dans le cytosol presque aussi
rapidement que dans l'eau pure. En revanche, les grosses particules, comme les
vésicules, diffusent très lentement, notamment parce qu'elles entrent en collision
avec les éléments du cytosquelette. Par conséquent, pour se déplacer à des vitesses
utiles, les particules sont transportées activement par des protéines motrices
associées au cytosquelette.

30
C. Principales fonctions.
1. De nombreuses voies métaboliques débutent dans le cytosol pour se
terminer dans d'autres compartiments après adressage des métabolites.
2. Le cytosol joue un rôle essentiel dans le métabolisme des glucides (cf
cours de biochimie) par la voie de la glycolyse à partir du glucose, celle des pentoses
et de la synthèse de glycogène à partir du glucose-6-phosphate. Par la glycolyse qui
produit de l'ATP, le cytosol est avec la mitochondrie l'un des 2 sites de production
d'énergie.
3. La synthèse protéique se déroule dans le cytosol à l'exception de
certaines protéines mitochondriales (cf. cours de Biochimie). La synthèse protéique
s'effectue par traduction de la molécule d'ARN messager au niveau d'un ribosome.
Chez les eucaryotes, plusieurs ribosomes peuvent être fixés sur un même ARNm
pour former un polysome (ou polyribosome) permettant la synthèse de plusieurs
copies du même polypeptide. Des facteurs présents dans le cytosol reconnaissent les
signaux de tri présents sur certaines protéines et interviennent dans leur orientation
vers leur destination finale.
Exemples : la SRP (Signal Recognition Particle) intervient dans l'adressage des
protéines destinées au réticulum endoplasmique (cf. paragraphe III) ; la NLSBP
(Nuclear Localization Signal Binding Protein) se lie au signal peptide d'adressage au
noyau et permet la traversée des pores nucléaires.
4. Devenir des protéines cytosoliques.
a) Modifications post-traductionnelles.
Les protéines cytosoliques subissent fréquemment des modifications post-
traductionnelles :

31
- modifications permanentes, nécessaires à l'activité, telles que la liaison à un
coenzyme ;
- modifications transitoires, qui permettent de réguler l'activité de nombreuses
protéines (phosphorylation, méthylation, acétylation). Les réactions de
phosphorylation ou de déphosphorylation sont très fréquentes et concernent près de
10 % des protéines animales ;
- les glycosylations sont rares par comparaison aux protéines exportées vers le
réticulum endoplasmique. Elles consistent seulement en l'addition d'une seule
molécule de N-acétylglucosamine à une protéine occasionnelle.
b) Liaison à la face cytoplasmique des membranes par une
chaine d'acide gras.
Certaines protéines cytoplasmiques sélectionnées sont dirigées vers les
membranes, et se fixent de façon covalente à un acide gras (acide palmitique, acide
myristique...) de la monocouche cytoplasmique de la membrane. Ces modifications
sont dues à des enzymes qui reconnaissent un signal peptide spécifique. L'ancrage
aux membranes est indispensable aux fonctions biologiques des protéines ainsi
modifiées. Certaines protéines s’ancrent dans la membrane plasmique après addition
d’un groupement prényl.
Exemples : membrane plasmique : l'ankyrine et les protéines G trimériques, la
protéine Ras ; face nucléoplasmique de l'enveloppe nucléaire : l'une des lamines.
c) Dégradation des protéines par les protéasomes.
- Protéines cytosoliques normales. La majorité des protéines cytosoliques a une
durée de vie assez longue, de plusieurs jours. D'autres sont programmées pour être
rapidement détruites après leur synthèse. C'est le cas notamment des protéines
régulant la progression dans le cycle cellulaire. Ces protéines programmées pour être
rapidement détruites portent des signaux reconnus par un système protéolytique.
L'un de ces signaux est constitué par la nature du premier acide aminé amino-
terminal de la chaîne polypeptidique. Met, Ser, thr, Ala, Val, Cys, Gly et Pro sont
stabilisants (demi-vie protéique > 20 h), alors que les 12 autres acides aminés sont
responsables d'une dégradation rapide de la protéine (demi-vie de 2 à 10 min).

32
D'autres signaux de destruction ont été identifiés. Ainsi, certains enchaînements
d'acides aminés raccourcissent la durée de vie des protéines qui les portent. A titre
d'exemple, les cyclines, des protéines impliquées dans la progression du cycle
cellulaire, portent une séquence de 9 acides aminés appelée boite de dégradation (D-
box). La dégradation des protéines peut être constitutionnelle ou conditionnelle
(régulée) après démasquage d’un signal de dégradation : dans ce dernier cas, divers
mécanismes régulateurs existent : phosphorylation de la protéine ou coupure
protéolytique permettant soit le démasquage d’une boîte de dégradation, soit
l’apparition en position N-terminale (après coupure) d’un acide aminé déstabilisant.
Chez les eucaryotes, le système protéolytique responsable de la dégradation sélective
des protéines cytosoliques fait intervenir une protéine chaperonne, l'ubiquitine. Le
mécanisme de liaison des molécules d’ubiquitine fait intervenir une enzyme E1
responsable de l’activation d’une molécule d’ubiquitine (en présence d’ATP) et le
complexe E2-E3 (appelé ubiquitine ligase) permettant la fixation covalente de la
molécule d’ubiquitine activée sur la protéine à dégrader (Figure 6). Comme le
processus se répète, plusieurs molécules d’ubiquitine vont former une chaîne fixée
sur la protéine. Il existe environ une trentaine d’enzymes E2 similaires mais
distinctes et des centaines de molécules E3, permettant ainsi la destruction de
protéines aux signaux très variés. Les protéines portant une chaîne d’ubiquitine sont
alors reconnues par le protéasome qui est responsable de la dégradation de la
protéine. De multiples copies du protéasome existent dans le cytosol et aussi dans le
noyau. Le protéasome est constitué d’un cylindre creux fait de sous-unités protéiques
dont certaines sont des protéases dont le site catalytique est orienté vers l’intérieur
du cylindre. A chaque extrémité du cylindre se trouve un large complexe protéique
dont le rôle est de reconnaître les protéines marquées par un chaîne d’ubiquitine, de
les dérouler et de les insérer dans le cylindre afin qu’elles soient dégradées. Ce
processus nécessite de l’ATP et se fait à pH neutre. Les produits de l'hydrolyse
partielle par le protéasome ont plusieurs destinations possibles :
- le compartiment lysosomal via des perméases spécifiques, où leur dégradation
sera achevée ;

33
- le cytosol : dans ce cas, des exopeptidases cytosoliques achèvent la dégradation
des peptides ;
- la lumière du réticulum endoplasmique granulaire via des transporteurs
spécifiques présents dans la membrane du réticulum. Ces petits peptides (une
dizaine d’acides aminés) se lient alors à des protéines transmembranaires, les
complexes majeurs d'histocompatibilité de type I (CMH I). Les CMH I liés aux
peptides sont transportés vers la membrane plasmique où le peptide antigénique est
présenté au lymphocytes T de type CD8 (cf. cours d'immunologie).
Figure 6.
- Protéines anormales. Le protéasome dégrade également des protéines
cytosoliques anormales, soit du fait d’erreur lors de leur synthèse ou de l’oxydation
secondaires de certains acides aminés, ou des protéines adressées au réticulum
endoplasmique et reconnues comme anormales (cf. chapitre réticulum
endoplasmique).
III. Le réticulum endoplasmique.
Le réticulum endoplasmique (RE) est un espace labyrinthique limité par une
membrane, et s'étendant dans tout le cytoplasme de la cellule (Figure 7). Cet espace
aux multiples circonvolutions est appelé lumière du RE ou citerne du RE. Il occupe

34
souvent plus de 10 % du volume cellulaire. Il est en continuité avec l'enveloppe
nucléaire. Le RE joue un rôle essentiel dans la biosynthèse cellulaire des protéines et
des lipides.
Figure 7.
A. Le réticulum endoplasmique rugueux ou granulaire.
Le RE capte de façon sélective certaines protéines cytosoliques, dès qu'elles sont
synthétisées. On distingue 2 types de protéines :
- les protéines trans-membranaires qui sont enchassées dans la membrane du RE
et dont certaines seront acheminées vers la membrane plasmique ou celle du Golgi
ou des lysosomes ;
- les protéines solubles qui sont entièrement transloquées à travers la membrane
du RE et libérées dans sa lumière ; ces protéines seront destinées à la lumière d'un
organite en aval du réticulum dans le flux vectoriel, à la sécrétion dans le milieu
extracellulaire, ou à l'ancrage à la face externe de la membrane plasmique sous forme
de protéines extrinsèques.
Dans les cellules de mammifères, l'importation de la plupart des protéines dans le
RE commence avant que la chaîne polypeptidique ne soit entièrement synthétisée

35
(importation co-traductionnelle). De ce fait, des ribosomes sont liés à la membrane
du RE sur son versant cytosolique. Ces régions du RE liées à des ribosomes portent le
nom de réticulum endoplasmique rugueux ou granulaire.
Dans le cytoplasme, il existe donc 2 types de ribosomes (Figure 8) :
- les ribosomes liés à la membrane du RE,
- les ribosomes libres qui fabriquent les protéines cytosoliques, dont certaines sont
acheminées vers les mitochondries, le noyau ou les peroxysomes.
Les 2 types de ribosomes sont identiques. L'attachement d'un ribosome au RE
dépend de la nature de la protéine synthétisée, et notamment de l'existence d'un
peptide signal d'importation dans le RE. Le RE rugueux est situé près du noyau et sa
membrane est en continuité avec l'enveloppe du noyau.
Figure 8.
B. Réticulum endoplasmique lisse.
Le RE lisse correspond aux régions du RE dépouvues de ribosomes (Figure 7). La
grande majorité des cellules contient peu de véritable RE lisse. Au lieu de cela, une
petite région du RE est partiellement rugueuse et partiellement lisse, et c'est au
niveau des régions de RE lisse que les vésicules de transport bourgeonnent pour se
diriger vers l'appareil de Golgi. Dans ce cas, ce réticulum s’appelle aussi réticulum
transitionnel. A l’inverse, dans certaines cellules, le RE lisse est abondant et

36
correspond à une véritable entité anatomique ; dans ces cellules, le RE lisse a des
fonctions spécialisées :
- cellules responsables de la synthèse des hormones stéroïdes, où le RE lisse
participe à la synthèse du cholestérol et à sa transformation en hormones.
- l'hépatocyte (principale cellule du foie), où le RE lisse est le site principal de
production des particules lipoprotéiques destinées à l'exportation. De plus, dans
l'hépatocyte, le RE lisse est le siège d'enzymes métabolisant les médicaments (famille
des cytochromes P450), les rendant plus hydrosolubles, avant leur élimination dans
la bile ou l'urine.
- les cellules musculaires, où le RE lisse est appelé réticulum sarcoplasmique. Le
réticulum sarcoplasmique est capable de complexer les ions Ca++ ou de les libérer
dans le cytosol, et, pour cette raison, joue un rôle essentiel dans la contraction
musculaire.
C. Méthodes d'isolement du réticulum endoplasmique.
La fonction du RE ne peut être étudiée que si les membranes du RE sont séparées
des autres constituants cellulaires. Pour cela, il est nécessaire de faire éclater les
cellules et de les homogénéiser. Au cours de l'homogénéisation, le RE est fragmenté
en petites vésicules fermées, appelées microsomes, qui sont relativement faciles à
purifier (Figure 9).
Figure 9.

37
On distingue 2 types de microsomes :
- les microsomes rugueux, qui sont recouverts de ribosomes sur leur surface
externe, et dont l'intérieur à une composition biochimique identique à celle de la
lumière du RE ;
- les microsomes lisses, qui proviennent en partie du RE lisse, et en partie de
fragments vésiculés de la membrane plasmique, de l'appareil de Golgi, des
endosomes, et des mitochondries. L'hépatocyte est une exception, car du fait de
l'abondance du RE lisse dans cette cellule, la majorité des microsomes lisses provient
du RE lisse.
En raison des grandes quantités d'ARN qu'ils contiennent, les microsomes rugueux
sont plus denses que les microsomes lisses. Ils peuvent donc être séparés par
centrifugation dans un gradient continu de densité (Figure 10).
D. Rôle du réticulum endoplasmique.
1. Transport et maturation des protéines.
a) Mécanisme d'importation co-traductionnelle des protéines
dans le RE.
Les protéines importées dans le RE comportent un peptide signal composé
d'acides aminés hydrophobes à leur extrémité aminée (la première synthétisée). Le
transfert dans le RE comporte plusieurs étapes (Figure 10) :
- Une particule de reconnaissance du signal (SRP) se fixe sur le peptide signal qui
émerge du ribosome libre dans le cytosol. La SRP est une ribonucléo-protéine
cytosolique constituée de 6 chaînes polypeptidiques et d'une molécule d'ARN et
possède une activité d'hydrolyse du GTP (activité GTPasique). L'une de ses
extrémités possède un site de fixation sur le peptide signal, tandis que l'autre se fixe
sur le ribosome, ce qui a pour effet d'interrompre temporairement la traduction de
l'ARNm.

38
- La SRP se fixe sur un récepteur appelé protéine d'ancrage ou récepteur de la
SRP, exposé à la face cytosolique de la membrane du RE rugueux et qui possède une
activité d'hydrolyse du GTP.
- Le récepteur de la SRP permet de positionner le complexe SRP-grosse sous-unité
ribosomale sur un complexe protéique voisin, le translocon (complexe Sec61), qui
constitue un pore aqueux entre le cytosol et la lumière du RE, et dont le versant
luminal est fermé. En fin de positionnement, la SRP se détache du peptide signal, la
synthèse de la protéine redémarre et la chaîne peptidique naissante s’insère dans la
lumière du translocon dont l’extrémité cytosolique est obturée par la grosse sous-
unité ribosomale ; la SRP se sépare de son récepteur.
- La fixation du peptide signal sur un site spécifique du translocon est responsable
de l’ouverture de ce pore du côté luminal.
- La progression de la chaîne protéique au travers du dispositif de translocation
fait probablement intervenir, d'une part l'allongement de la chaîne protéique en
cours de synthèse, et d'autre part des molécules chaperonnes localisées dans la
lumière du RE, en particulier la BiP (Binding Protein), qui "tirent" le peptide en cours
de synthèse.
Figure 10.

39
b) Mécanisme de libération des protéines solubles dans la lumière
du RE.
Dans le cas des protéines solubles libérées dans la lumière du RE, le peptide
signal reste ancré dans le translocon alors que la protéine s’enfile de façon continue
dans la lumière du RE, sous forme d'une boucle. Le signal peptide est clivé par une
signal peptidase liée à la face luminale de la membrane du RE et la protéine est
libérée dans la lumière du RE (Figure 11) où elle prend sa conformation définitive
sous l'action de protéines chaperonnes. Le signal peptide clivé sort latéralement du
translocon (par une ouverture latérale) dans la membrane du RE où il est dégradé
localement par d’autres protéases.
Figure 11.
c) Mécanismes d'insertion des protéines dans la membrane du RE.
Le processus de translocation des protéines membranaires intrinsèques est plus
complexe que celui des protéines solubles car :
- certaines parties de la protéine ne sont pas transloquées,
- certaines protéines transmembranaires traversent plusieurs fois la double couche
lipidique,

40
- l'extrémité amino-terminale de la protéine peut être située du côté cytosolique ou
dans la lumière du RE.
Nous décrirons 3 mécanismes d’insertion de protéines à traversée unique dans la
membrane du RE.
- Premier mécanisme. La translocation est initiée par un peptide signal amino-
terminal qui s’ancre dans le translocon. Un second peptide hydrophobe, appelé
peptide de terminaison de transfert, situé dans la protéine, interrompt la
translocation avant qu'elle ne soit terminée. Après clivage du peptide signal amino-
terminal, ce peptide de terminaison de transfert sert de point d'ancrage de la protéine
dans le translocon puis dans la membrane du RE. La protéine trans-membranaire est
à traversée unique et son extrémité amino-terminale est située du côté luminal du RE
(Figure 12).
- Dans les deux autres mécanismes, le peptide signal est plus interne dans la
protéine. Ce signal est également reconnu par la SRP et s’insère dans le translocon. Il
n’est pas clivé et sert donc d’ancrage de la protéine dans la membrane du RE. Selon
l’orientation du peptide signal dans le translocon, la protéine aura son extrémité N-
terminale dans la lumière du RE ou dans le cytosol (Figure 13).
Figure 12.

41
Figure 13.
d) Liaison d’une protéine à un glycosylphosphatidylinositol (GPI).
La liaison a lieu dans la lumière du RE par attachement covalent d’un GPI
préformé à l’extrémité C-terminale de protéines destinées à la membrane plasmique.
Dans le RE, la protéine est liée à la membrane mais ses acides aminés sont exposés
dans la lumière de cet organite.
e) N-glycosylation des protéines solubles ou transmembranaires.
Une des principales fonctions de biosynthèse du RE est l'addition de glucides aux
protéines. La grande majorité des protéines présentes dans la lumière du RE et
exportées vers les organites, la membrane plasmique ou le milieu extracellulaire, sont
des glycoprotéines. A l'inverse, peu de protéines cytosoliques sont glycosylées.
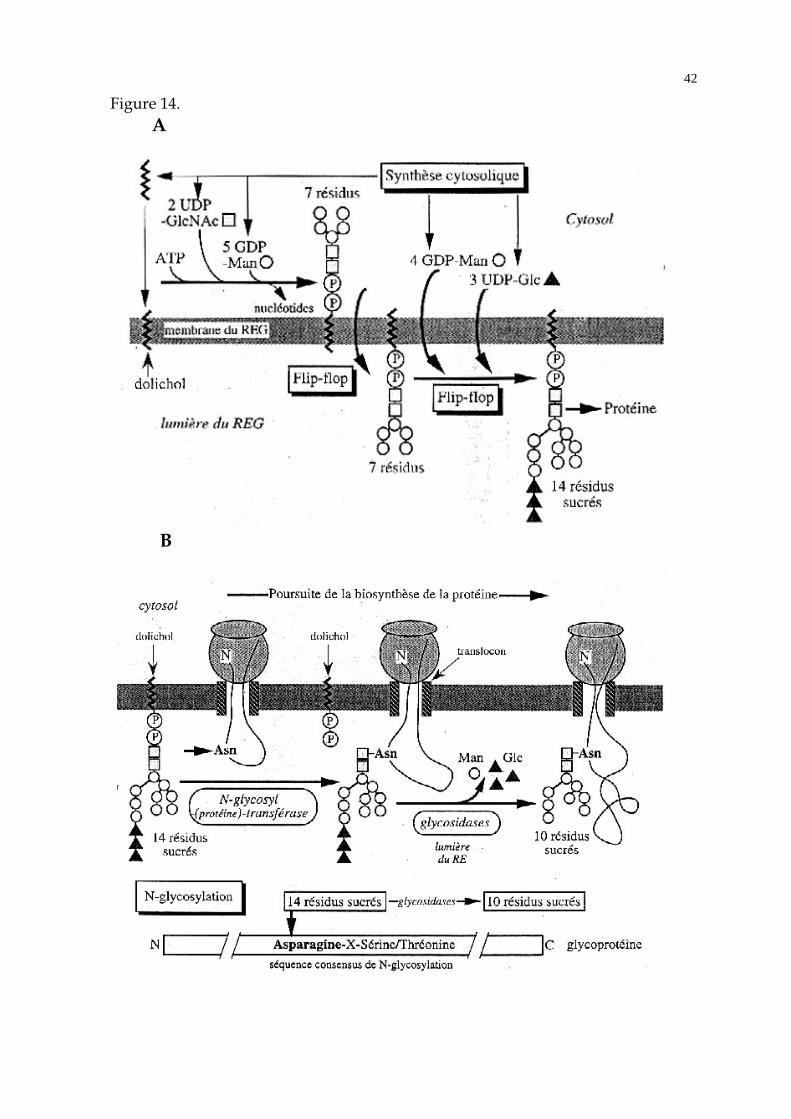
La N-glycosylation des protéines se déroule de façon co-traductionnelle, par
transport en bloc d'une seule espèce d'oligosaccharide, constituée de N-
acétylglucosamine, de mannose et de glucose (Figure 14). Le processus comporte
plusieurs étapes :
42
Figure 14.
A
B

43
- Synthèse sur la face cytosolique de la membrane du RE d'un oligosaccharide
comportant de la N-acétylglucosamine et du mannose, fixé à un dolichol de la
monocouche cytosolique de la membrane du RE (Figure 14 A).
- Basculement de la molécule de dolichol glycosylée dans la monocouche luminale
de la membrane du RE (Figure 14 A).
- Accrochage supplémentaire de mannose et de glucose sur l'oligosaccharide
(Figure 14 A).
- Transfert en bloc de l'oligosaccharide sur le groupement -NH2 présent sur la
chaîne latérale d'un résidu asparagine (N-glycosylation), sous l'action d'une
oligosaccharide-protéine transférase de la membrane du RE dont le site catalytique est
exposé du côté luminal de la membrane (Figure 14 B). Cette réaction concerne les
résidus asparagine situés au sein d'une séquence consensus Asn-X-Ser ou Thr, où X
est un acide aminé quelconque.
- L'arborisation sucrée fixée à la protéine est ensuite modifiée par des glycosidases
présentes dans la lumière du RE qui l'élaguent en partie.
f) Repliement des protéines membranaires ou solubles.
La conformation spatiale des protéines du RE est assurée par plusieurs types de
protéines résidant dans la lumière du RE. Le repliement consomme de l'énergie
fournie par l'ATP.
- Des protéines chaperonnes, parmi lesquelles la BiP (binding protein ou protéine
de liaison). Ces protéines assurent le bon repliement des protéines synthétisées. La
BiP se lie dans la lumière du RE aux segments hydrophobes des protéines en cours
de translocation, évitant ainsi à ces protéines de s’agréger entre-elles ou de mal se
configurer. Par des cycles successifs de liaison-libération ATP-dépendant, la BiP
permet à chaque protéine d’acquérir la bonne conformation. En restant liée aux
protéines mal configurées, la BiP les empêche de quitter le RE.
- Les disulfides isomérases. Ces enzymes catalysent la formation de ponts
disulfures S-S intramoléculaires entre les groupements sulfhydryl (-SH) des
protéines. Elles sont également capables de reconnaître des ponts disulfures
anormaux qui se seraient formés et de les corriger. En règle générale, les résidus

44
cystéine des protéines localisées dans la lumière des organites ou dans le milieu
extracellulaire forment pour la majorité d’entre eux des ponts disulfures,
contrairement à ceux des protéines cytosoliques.
- la calnexine (ou la calréticuline) et la glucosyltransférase (GT). Le rôle de ces
protéines est de vérifier la conformation spatiale des protéines dans la lumière du RE
(Figure 15).
Figure 15.
Les protéines N-glycosylées subissent un premier élagage dans le RE qui conduit
initialement à l’élimination de 2 résidus glucose par les glucosidases I et II (Figure
15). Les protéines qui ne possèdent plus qu’un résidu glucose terminal se lient à la
calnexine (protéine membranaire) par l’intermédiaire de ce sucre (la calréticuline est
une protéine soluble du RE dont le mécanisme d’action est identique à celui de la
calnexine). Une thioloxydoréductase associée à la calnexine catalyse la formation des
ponts disulfures ou leur réarrangement. Après clivage du 3ème glucose par la
glucosidase II, la protéine se libère de la calnexine. Si la bonne configuration est

45
obtenue, la protéine peut quitter le RE. Dans le cas contraire, la protéine est reconnue
par la glucosyltransférase (GT), glucosylée puis à nouveau liée à la calnexine. Si après
plusieurs cycles, le bon repliement n’est pas obtenu, la protéine est détruite. On ne
connaît pas le mécanisme par lequel la GT reconnaît les protéines anormales. Les
protéines anormales du RE qui ne peuvent être corrigées refluent dans le cytosol à
travers le translocon et sont dégradées par le système ubiquitine-protéasome.
- La réponse du réticulum endoplasmique au stress. Cette réponse s’observe
lorsque des protéines mal configurées s’accumulent dans le RE. Elle se traduit par
une augmentation au niveau nucléaire de la transcription de gènes codant pour des
protéines chaperonnes du RE. Cette réponse est due à l’activation, par les protéines
mal configurées, de protéines insérées dans le membrane du RE qui, par leur
domaine cytosolique, transmette l’information au noyau.
2. Synthèse des lipides.
La membrane du RE produit presque tous les lipides nécessaires à la fabrication
des nouvelles membranes de la cellule : phospholipides (phosphatidylcholine,
phosphatidylsérine, phosphatidyléthanolamine et phosphatidylinositol), céramide et
cholestérol.
a) Phospholipides.
La synthèse des phospholipides présente des points communs avec la N-
glycosylation des protéines. Elle débute sur le versant cytosolique de la membrane ;
les enzymes nécessaires sont situées dans la membrane du RE et leurs sites
catalytiques sont exposés sur son versant cytosolique. Un facteur de translocation, la
scramblase, transfert ensuite des phospholipides vers la monocouche luminale de la
membrane du RE. Les différents phospholipides se retrouvent également distribués
entre les 2 feuillets de la membrane du RE. L’asymétrie de distribution des
phospholipides au niveau de la membrane plasmique est due à la présence, dans
cette membrane, d’une flippase qui transfert la phosphatidylsérine et la
phosphatidyléthanolamine de la monocouche externe vers la monocouche interne

46
(Figure 16). Il existe une scramblase dans la membrane plasmique. Contrairement à
la scramblase du RE qui est toujours active, la scramblase de la membrane plasmique
n’est active que dans certaines situations comme l’apoptose.
b) Le RE produit également du cholestérol et du céramide.
Le céramide est exporté dans la membrane de l'appareil de Golgi, où il sert de
précurseur pour la synthèse des glycolipides (réaction de glycosylation) et de la
sphingomyéline (transfert d'un groupement phosphocholine d'une molécule de
phosphatidylcholine sur un céramide). Ces modifications sont catalysées par des
enzymes situées dans la lumière de l'appareil de Golgi et intéressent donc
exclusivement la couche luminale de la bi-couche lipidique de la membrane de
l'appareil de Golgi.
c) Devenir des constituants membranaires.
Par l'intermédiaire des vésicules de transport, les lipides et les protéines
transmembranaires entrent dans le flux vectoriel permanent entre le RE, le Golgi, les
lysosomes et la membrane plasmique. Ce mécanisme permet le renouvellement de la
membrane de ces différents compartiments.
En revanche, la croissance des mitochondries et des peroxysomes fait intervenir un
mécanisme différent. Les protéines des membranes de ces organites proviennent
directement du cytosol. Les lipides de ces organites sont synthétisés dans le RE puis
arrachés de la monocouche cytosolique de la membrane du RE par des protéines de
Figure 16.

47
transfert des phospholipides qui les acheminent jusqu'aux mitochondries ou aux
peroxysomes (Figure 17). Toutefois, la phosphatidyléthanolamine peut être produite
dans la mitochondrie par modification de la phosphatidylsérine provenant du RE.
En plus des protéines de transfert des phospholipides, les phospholipides
pourraient aussi être transférés directement de la membrane du RE vers la
membrane mitochondriale au niveau de zones de contact étroit entre ces deux types
d’organites. Il est également possible qu’un transfert direct de phospholipides puisse
aussi se produire entre le RE et d’autres organites comme l’appareil de Golgi ou le
système endosome/lysosome.
IV. Transport vésiculaire intracellulaire. Mécanismes moléculaires.
Le transfert des protéines à partir du réticulum vers les autres organites (Golgi,
lysosomes) et la membrane plasmique (exocytose) se fait par l'intermédiaire de
vésicules de transport. Il en est de même pour le transport des protéines d'un saccule
golgien à un autre. Dans ce type de transport, les vésicules se forment par
bourgeonnement du compartiment donneur. Au cours de ce processus, les vésicules
ainsi formées s'emparent du matériel présent dans le compartiment donneur et le
libèrent dans le compartiment cible, après fusion de la membrane vésiculaire avec
celle du compartiment cible. Toujours grâce à des vésicules, les cellules eucaryotes
captent des macromolécules du milieu extracellulaire pour les diriger vers les
lysosomes (endocytose).
Figure 17.

48
Les vésicules transportant les protéines sont recouvertes d’une enveloppe
protéique. Le revêtement protéique joue un rôle dans le bourgeonnement de la
vésicule et dans la sélection des protéines à transporter. Il a été montré qu'il existait
principalement 2 types de vésicules enveloppées : i) les vésicules enveloppées de
clathrine qui transportent du matériel destiné à l’exocytose contrôlée (Figure 36) ou
aux endosomes [de l’appareil de Golgi vers le compartiment endosomal tardif
(Figure 30) ou de la membrane plasmique vers le compartiment endosomal précoce
(endocytose)] ; ii) les vésicules enveloppées de COP (COat Proteins) sont impliquées
dans le transport vésiculaire non sélectif du RE au Golgi et dans les transports
vésiculaires de l’appareil de Golgi au RE et d'un compartiment du Golgi au suivant
(Figure 19). Le revêtement protéique des vésicules impliquées dans l’exocytose
constitutive n’a pas été caractérisé biochimiquement.
A. Vésicules enveloppées de clathrine.
Les molécules de clathrine s'auto-assemblent pour former les puits recouverts sur la
face cytosolique des membranes. En microscopie électronique, les puits recouverts
apparaissent comme des dépressions membranaires recouvertes sur leur face
cytosolique par un matériel protéique dense correspondant aux complexes de
clathrine. Les molécules de clathrine sont liées à la membrane par l'intermédiaire de
complexes protéiques appelés adaptateurs. Il existe différents types d’adaptateurs,
certains étant formés de 4 sous-unités protéiques, d’autres d’une seule chaîne
protéique. Ces différents types d’adaptateurs identifient des vésicules d’origine et de
destination différentes. Les adaptateurs permettent de lier le squelette de clathrine à
la membrane. Leur rôle est aussi de capturer des récepteurs transmembranaires dans
les puits recouverts en reconnaissant des peptides signal spécifiques situés dans la
partie cytosolique du récepteur (Figure 18). Ces récepteurs se concentrent dans les
puits recouverts avant ou seulement après avoir fixé leur ligand spécifique. Ce
processus permet ainsi de capter et de concentrer dans un petit volume des
macromolécules présentes en très faibles concentrations dans la lumière des

49
organites ou le milieu extracellulaire. Les puits recouverts ont une durée de vie
Figure 18
courte. La formation de l’enveloppe de clathrine provoque l'invagination de la
membrane qui se pince pour former une vésicule recouverte de clathrine. Le
pincement des puits recouverts fait intervenir une protéine à activité GTPasique, la
dynamine. En quelques secondes, la vésicule perd son revêtement protéique par un
processus actif qui fait intervenir une ATPase et devient une vésicule non recouverte
ou vésicule lisse.
Figure 19.

50
B. Vésicules enveloppées de COP.
Il existe deux groupes de COP : COP-I et COP-II (Figure 19). Le revêtement des
vésicules recouvertes de COP-I est constitué de complexes protéiques constitués de 7
protéines majeures. Les vésicules recouvertes de COP-I assurent le transport de
protéines de l’appareil de Golgi au RE ou d’un compartiment Golgien au suivant. Le
revêtement des vésicules recouvertes de COP-II est constitué de complexes
protéiques constitués de 4 protéines. Les vésicules recouvertes de COP-II assurent le
transport des protéines du RE à l’appareil de Golgi.
C. Vésicules recouvertes de rétromères.
Les rétromères s’assemblent sur l’endosome et forment des vésicules qui assurent
le retour des récepteurs des hydrolases acides (récepteurs du mannose-6-phosphate)
vers l’appareil de Golgi (Figure 30). Un rétromère est constitué de 4 sous-unités
protéiques dont l’une se lie au domaine cytosolique des récepteurs au mannose-6-
phosphate et dont une autre se lie à un phosphatidylinositol phosphaté en 3’ de
l’inositol [PI(3)P].
D. Assemblage des protéines de revêtement.
L’assemblage des protéines de revêtement dépend d’une famille de GTPases
monomériques ; les protéines ARF, responsables de l’assemblage du revêtement de
COP-I et de clathrine, et la protéine Sar1 responsable de l’assemblage du revêtement
de COP-II (Figure 20). Ces GTPases existent sous deux formes, actives lorsqu’elles
sont liées au GTP, inactives lorsqu’elles sont liées au GDP. Le « switch » entre les
deux formes est régulé par deux classes de protéine, les facteurs d’échange des
nucléotides guanyliques (GEF) qui sont activateurs car ils catalysent le remplacement
du GDP par du GTP et les GAP (protéines activatrices de la GTPase) qui sont
inactivatrices car elles stimulent l’hydrolyse du GTP en GDP. Les protéines ARF et
Sar1 sont présentes à de fortes concentrations dans le cytosol sous forme inactive,
liées au GDP.

51
Avant le bourgeonnement membranaire, des GEF membranaires activent cette
famille de GTPase (remplacement d’un GDP par un GTP). Sous forme active, ces
molécules exposent des chaînes hydrophobes leur permettant de s’insérer dans les
membranes et de recruter les protéines de revêtement. Après hydrolyse du GTP en
GDP, les protéines ARF et Sar1 inactives se décrochent des membranes provoquant
le désassemblage du revêtement vésiculaire (Figure 20).
Figure 20

52
E. Sélectivité du transport vésiculaire.
Le processus de reconnaissance des membranes cibles des vésicules de transport
est hautement spécifique et fait intervenir deux classes de protéine, les SNARE et les
protéines rab.
1. Les protéines SNARE.
Les protéines SNARE jouent un rôle essentiel dans la spécificité de l’amarrage
d’une vésicule à la membrane cible et dans la fusion des deux membranes (Figure
21). Ils existent plus de 35 protéines SNARE différentes dans l’espèce animale qui se
regroupent en paires complémentaires où chaque élément d’une paire s’amarre
spécifiquement à l’autre élément ; les v-SNARE sont présentes sur la membrane des
vésicules qui bourgeonnent et les t-SNARE sur la membrane ciblent et chaque couple
identifie une voie bien définie de transport vésiculaire. Une v-SNARE est constituée
d’une seule chaîne polypeptidique ; une t-SNARE est constituée de 2 à 3 chaînes
polypeptidiques. Les SNARE sont des protéines transmembranaires ; le domaine
cytosolique d’une v-SNARE s’enroule en hélice avec le domaine cytosolique de son
partenaire t-SNARE. Cet enroulement permet le rapprochement des deux
membranes puis leur fusion. Après fusion membranaire, chaque couple peut se
dissocier pour que les deux éléments qui le constituent soient réutilisés. La
dissociation est catalysée par des protéines accessoires et une ATPase appelée NSF.
2. Les protéines rab.
Les protéines rab sont des GTPases monomériques qui contribuent également à la
sélectivité du transport vésiculaire. Il existe plus de 60 protéines rab différentes,
chacune ayant une distribution caractéristique sur les membranes cellulaires. Leur
rôle est de faciliter et de réguler l’amarrage d’une vésicule à une membrane et
l’appariement d’une v-SNARE avec son partenaire t-SNARE. Comme les protéines
ARF et Sar1, les protéines rab existent sous deux formes, inactives dans le cytosol
(liées au GDP), actives membranaires (liées à du GTP). Le passage de la forme

53
inactive à la forme active est dû à des facteurs d’échange (GEF) ancrés dans les
membranes. L’activation des protéines rab se caractérise par l’exposition d’un
ancrage lipidique permettant la liaison de la protéine à une membrane. En se liant à
un effecteur de rab présent sur une membrane cible, la protéine rab intégrée dans la
membrane d’une vésicule permet l’amarrage de ces deux membranes l’une à l’autre,
facilitant ainsi l’appariement des SNARE puis la fusion membranaire (Figure 22).
Figure 21
Figure 22

54
V. Mécanismes d'adressage et de rétention des protéines spécifiques de chaque
compartiment.
Ils assurent le maintien des caractéristiques spécifiques de chaque compartiment.
L’adressage et la rétention d’une protéine dans un compartiment spécifique (RE,
appareil de Golgi, lysosome) fait intervenir un signal d’adressage et de rétention
(différent du signal d’entrée dans le RE) spécifique de ce compartiment. Ces signaux
diffèrent pour les protéines transmembranaires et solubles :
- Les protéines transmembranaires portent un peptide signal dans leur domaine
cytosolique ou dans leur premier segment transmembranaire (si celles-ci en
comportent plusieurs).
- Les protéines solubles ont un peptide signal dans leur séquence (cas des protéines
résidant dans le RE comme la BiP et la disulfide isomérase) ou un signal porté par
leurs chaînes glycosylées (cas des enzymes lysosomales). Le signal d'adressage de ces
protéines solubles est reconnu par une protéine transmembranaire qui porte elle-
même, dans son domaine cytosolique, un signal peptide de tri et de rétention dans
un compartiment donné.
VI. L’appareil de Golgi
A. Structure.
L'appareil de Golgi est habituellement localisé près du noyau et, dans les cellules
animales, il est proche du centrosome. Il est constitué de saccules aplatis, limités par
une membrane et organisés par groupes de 4 à 6 en pile d'assiettes constituant des
dictyosomes. Le nombre et la taille de ces empilements peuvent varier de façon
importante d'un type cellulaire à un autre.
Chaque dictyosome présente 2 faces et 3 compartiments (figure 23) :
- la face cis, qui est située du côté du RE, correspond à la face d'entrée dans
le Golgi ;
- la face trans, située vers la membrane plasmique ou les lysosomes, est la face de
sortie de l'appareil de Golgi ;

55
- le compartiment cis (dont le réseau cis) est constitué par les saccules situés à
proximité du RE et reçoit les protéines exportées du RE ;
- le compartiment trans (dont le réseau trans) est formé par les saccules les plus
périphériques ; c'est là que se termine la maturation de la majorité des protéines,
avant leur exportation vers les lysosomes ou la membrane plasmique ;
- le compartiment médian est situé entre les compartiments cis et trans.
De nombreuses vésicules assurent le transport des protéines, du RE vers le
compartiment cis, de l’appareil de Golgi vers le RE, d'un saccule golgien à l'autre, et
du compartiment trans vers les lysosomes ou la membrane plasmique. Cette
circulation des protéines a pu être mise en évidence grâce aux expériences
d'autoradiographie.
B. L'appareil de Golgi est au carrefour de 4 flux membranaires (Figure 24).
- Un flux entrant provenant du RE.
- Un flux rétrograde du Golgi vers le RE.
- Le flux vectoriel permanent à partir du compartiment trans, par des vésicules
destinées aux endosomes et à la membrane plasmique (cf. paragraphe exocytose).
- Un flux rétrograde des endosomes vers le Golgi trans.
Figure 23

56
Figure 23 (suite)
Figure 24.

57
1. Transport vésiculaire du réticulum endoplasmique vers l’appareil de
Golgi.
Au niveau du réticulum endoplasmique, les protéines destinées à l’appareil de
Golgi et au-delà sont empaquetées dans des vésicules recouvertes de COP-II qui
bourgeonnent à partir du RE transitionnel (Figure 19). Contrairement à ce qui était
admis, ce processus est sélectif : les protéines transmembranaires destinées à
l’exportation hors du RE possèdent dans leur domaine cytosolique un signal de
sortie qui leur permet de se fixer sur les complexes COP-II ; les protéines solubles
destinées à l’exportation hors du RE se fixent sur des récepteurs membranaires
possédant dans leur domaine cytosolique un signal leur permettant également de se
fixer sur les complexes COP-II. Ces derniers, après avoir libéré leurs ligands dans
l’appareil de Golgi, doivent être réadressées vers le RE. Enfin, des protéines solubles
ou membranaires résidentes du RE sont « par erreur » incorporées dans les vésicules
lors de leur bourgeonnement dans le RE et, comme les précédents devront être
réacheminées vers le RE (transport rétrograde).
2. Formation d’un compartiment vésiculo-tubulaire intermédiaire.
Après leur bourgeonnement et la perte de leur revêtement protéique, les vésicules
provenant du RE fusionnent entre elles (fusion homotypique) pour former un
compartiment vésiculo-tubulaire intermédiaire (Figure 19). Ce compartiment se
déplace le long des microtubules vers l’appareil de Golgi (face cis) avec lequel il
fusionne.
3. Transport rétrograde des protéines de l’appareil de Golgi vers le RE.
Du compartiment vésiculo-tubulaire et de l’appareil de Golgi, des vésicules
recouvertes de COP-1 bourgeonnent en permanence pour réadresser vers le RE les
récepteurs transmembranaires et les protéines solubles résidents du RE (Figure 19).
Ces protéines portent un signal d’adressage (ou de rétention dans le RE) différent
selon qu’elles sont solubles ou membranaires. Les protéines transmembranaires
possèdent dans leur domaine cytosolique le peptide signal d’adressage et de

58
rétention : KKXX. Ce signal se lie aux complexes protéiques COP-1. Les protéines
solubles résidentes du RE portent le peptide signal spécifique de rétention dans le
RE : KDEL (Figure 25). Lorsqu’elles sortent du RE, ce signal de rétention leur permet
de se fixer sur des récepteurs membranaires spécifiques (récepteurs KDEL) situés
dans la membrane des structures vésiculo-tubulaires et des compartiments cis,
median et trans de l'appareil de Golgi. Ces récepteurs se fixent par leur domaine
cytosolique sur les complexes protéiques COP-1 permettant ainsi de réacheminer les
protéines solubles résidentes du RE vers le RE. Les récepteurs KDEL circulent donc
entre le RE et l’appareil de Golgi. Les protéines résidentes du RE doivent donc avoir
une faible affinité pour les récepteur KDEL dans le RE (pour ne pas se fixer ou s’en
libérer) et une forte affinité dans l’appareil de Golgi (pour s’y fixer). Cette
modification d’affinité est due à la variation de pH entre ces deux compartiments, pH
neutre dans la lumière du RE (faible affinité de la protéine pour son récepteur), pH
faiblement acide dans l’appareil de Golgi (forte affinité de la protéine pour son
récepteur). Ainsi, dans l’appareil de Golgi, les protéines résidentes du RE se fixent à
leur récepteur et s’en détachent lorsqu’elles se retrouvent dans le RE. De plus, pour
rapatrier vers le RE une surface membranaire importante dans un faible volume, les
vésicules qui assurent le transport rétrograde de l’appareil de Golgi vers le RE ont
une structure tubulaire aplatie (« canalicule », voir Figure 24).
C. Fonctions de l'appareil de Golgi.
1. O-glycosylation des protéines.
Il s'agit de la fixation de sucres sur le groupement OH des chaines latérales de
certaines sérines et thréonines (O-glycosylation) (Figure 26). Elle concerne le
domaine luminal des protéines transmembranaires et les protéines solubles dans le
Golgi. Elle a lieu dans les compartiments médian et trans de l’appareil de Golgi : i)
les sucres sont synthétisés dans le cytosol et pénètrent dans le Golgi par une
perméase spécifique ; ii) ils sont accrochés aux protéines par une O-glycosyl-protéine
transférase.

59
Figure 25.
L'appareil de Golgi participe en particulier à la synthèse des protéoglycannes. Un
protéoglycanne est formé d’une chaîne protéique sur laquelle est (sont) fixé(s) un ou
plusieurs glycosaminoglycanne(s). Les glycosaminoglycannes sont des chaînes
linéaires sucrées constituées d'unités disaccharidiques répétitives fixées à la protéine
par O-glycosylation d'un résidu sérine ; ils se forment par addition successive de
résidus glucidiques. Dans le compartiment trans, les résidus sucrés des
glycosaminoglycannes sont abondamment sulfatés. De nombreux protéoglycannes
sont sécrétés dans le milieu extracellulaire où ils sont des composants de la matrice
extracellulaire. D'autres restent fixés à la face externe de la membrane plasmique.
Figure 26

60
2. Modification des chaînes oligosaccharidiques des protéines N-
glycosylées.
Les chaînes oligosaccharidiques N-glycosylées dans le RE peuvent ou non être
modifiées dans l’appareil de Golgi. Les modifications, quand elles se produisent,
consistent en :
- la phosphorylation, dans le compartiment cis, des résidus mannose
indispensables à la maturation des enzymes destinées à la lumière des lysosomes (cf
chapitre lyosomes) ;
- l'élagage de résidus mannoses et l’addition de nouveau résidus sucrés provenant
du cytosol (N-acétylglucosamine, galactose, acide sialique) sur les chaînes arborisées
N-glycosylées.
Ces réactions enzymatiques se déroulent de façon séquentielle et ordonnée dans
les différents compartiments de l’appareil de Golgi et aboutissent à la formation
d’oligosaccharides complexes : i) compartiment cis : élimination de résidus mannose
(mannosidase I) ; ii) compartiment médian : élimination de résidus mannose,
(mannosidase II), addition de N-acétylglucosamine ; iii) compartiment trans :
addition de galactose, d’acide sialique.
Le grand nombre de glycoprotéines et la complexité de leur synthèse suggèrent
que les chaîne glycosylées des protéines ont des fonctions importantes. Les chaînes
sucrées sont relativement inflexibles et, de ce fait, font saillie à la surface de la
protéine. Elles peuvent ainsi protéger les protéines de la digestion par les protéases.
Les chaînes oligosaccharidiques ont d'autres rôles, notamment dans l’adhésion
intercellulaire (via les sélectines).
4. Maturation protéolytique des protéines.
De nombreuses hormones polypeptidiques et des neuropeptides sont synthétisés
sous forme de précurseurs inactifs, à partir desquels les molécules actives sont
progressivement produites par protéolyse. Ces clivages débutent dans le
compartiment trans du Golgi et se poursuivent dans les vésicules de sécrétion, voire
parfois même dans le milieu extracellulaire.
Exemple : dans le pancréas, la proinsuline est clivée en insuline (figure 27).

61
VII. Transport du compartiment trans du Golgi à la surface de la cellule :
l'exocytose.
Des vésicules de transport destinées à la membrane plasmique quittent le
compartiment trans du Golgi. Les protéines et les lipides membranaires de ces
vésicules renouvellent la membrane plasmique, tandis que les protéines solubles de
la lumière de ces vésicules sont sécrétées dans le milieu extracellulaire. Ce processus
d’exportation et de fusion des vésicules avec la membrane plasmique est appelé
exocytose. On distingue deux types d’exocytose : l’exocytose constitutive et
l’exocytose controlée.
Figure 27.
Figure 28.

62
A. L'exocytose constitutive.
L’exocytose constitutive est continue et s’observe dans toutes les cellules (Figure
28). Il s’agit d’un processus par défaut qui concerne des protéines qui ne sont pas
sélectionnées pour une destination spécifique ; ces protéines n’ont pas de signal
spécifique ; elles sont empaquetées dans des vésicules qui bourgeonnent du réseau
trans de l’appareil de Golgi et qui fusionnent avec la membrane plasmique pour
libérer leur contenu dans le milieu extracellulaire.
B. L'exocytose contrôlée.
L’exocytose contrôlée s’observe dans certains types cellulaires, notamment dans
les cellules spécialisées dans la sécrétion d'hormones, de neurotransmetteurs ou
d'enzymes digestives (figure 28).
1. Formation des vésicules de sécrétion.
Les protéines destinées aux vésicules de sécrétion s'agrègent dans la lumière du
réseau trans de l’appareil de Golgi grâce à une région signal commune à ce type de
protéine. Les protéines agrégées sont probablement reconnues par des récepteurs
situés dans la membrane de l’appareil de Golgi. La membrane de ces vésicules porte
un revêtement de clathrine. Après bourgeonnement de la vésicule (vésicule
sécrétoire immature), le contenu vésiculaire se condense sous l'effet : i) d'une
acidification de la lumière vésiculaire induite par une pompe à proton membranaire
couplée à l'ATP et ii) la diminution de taille des vésicules sécrétoires due au
bourgeonnement à leur niveau de vésicules recouvertes de clathrine qui regagnent
l’appareil de Golgi (Figure 29). Les processus d'agrégation et de condensation
permettent de concentrer de façon importante les substances stockées dans les
vésicules de sécrétion (vésicule sécrétoire mature).
2. Maturation des protéines dans les vésicules de sécrétion.
La maturation protéolytique de nombreuses protéines qui débute dans le réseau
trans du Golgi se poursuit dans les vésicules de sécrétion (cf. paragraphe sur
l'appareil de Golgi).

63
3. Stockage des vésicules de sécrétion.
Les vésicules sont acheminées jusqu'à leur site de sécrétion. Dans certaines cellules,
ce site est très éloigné de l'appareil de Golgi. Ainsi, dans les neurones, les vésicules
de sécrétion sont formées dans le corps cellulaire et acheminées jusqu'à l'extrémité de
l'axone, située parfois à plus d'un mètre. Le transport des vésicules jusqu'à leur site
de sécrétion se fait grâce aux microtubules du cytosquelette et à des protéines
motrices. Les vésicules sécrétoires s’accumulent à proximité de la membrane
plasmique ; leur contenu n’est libéré qu’en réponse à un signal extracellulaire
spécifique (stimulation hormonale, électrique …), après fusion de la membrane
vésiculaire avec la membrane plasmique (Figure 28).
4. Les composants membranaires des vésicules de sécrétion sont
recyclés.
Lorsqu'une vésicule de sécrétion fusionne avec la membrane plasmique,
l'augmentation de surface de la membrane plasmique qui en résulte est
contrebalancée par le processus d'endocytose.
Figure 29.

64
VIII. Transport vésiculaire de la membrane plasmique vers le système
endosomal : l’endocytose
On distingue deux types d’endocytose, la pinocytose qui s’observe dans tous les
types cellulaires et dont le principe général est celui décrit ci-dessus et la
phagocytose qui est la propriété de seulement certains types cellulaires et dont le
mécanisme diffère sensiblement de celui de la pinocytose.
A. La pinocytose.
On distingue deux types de pinocytose en fonction de la nature des molécules
internalisées et du revêtement situé sur la face cytosolique des vésicules : la
pinocytose dépendante de la clathrine et la pinocytose non-dépendante de la
clathrine.
1) La pinocytose dépendante de la clathrine.
La pinocytose dépendante de la clathrine se produit dans des régions spécialisées
de la membrane plasmique : les puits recouverts qui occupent environ 2% de la
surface de la membrane plasmique. Après la perte du revêtement de clathrine, les
vésicules lisses qui se forment fusionnent avec les endosomes précoces, structures les
plus superficielles du compartiment endosomal, situées sous la membrane
plasmique. Ce compartiment est constitué d'un ensemble complexe de structures
vésiculo-tubulaires entourées d'une membrane. Quant aux complexes de clathrine,
ceux-ci se réassemblent au niveau de la membrane plasmique pour reconstituer un
puits recouvert.
Après fusion des vésicules avec les endosomes précoces, le devenir des
macromolécules et de leurs récepteurs varie suivant la nature même de la molécule
transportée. Lorsque le devenir du ligand est différent de celui du récepteur, le
complexe ligand-récepteur doit se dissocier. Cette dissociation est liée au pH acide de
ce compartiment, qui, en induisant des modifications conformationnelles de

65
beaucoup de récepteurs, diminue leur affinité pour le ligand. Le pH acide de
l'endosome précoce est dû à l'existence d'une pompe H+ membranaire qui transfère
les ions H+ du cytosol dans l'endosome. Les ligands qui se dissocient de leurs
récepteurs sont donc habituellement dégradés dans les lysosomes alors que leurs
récepteurs sont recyclés vers la membrane plasmique. Les ligands qui restent fixés à
leurs récepteurs partagent leur sort : recyclage, dégradation ou transcytose.
a. Le récepteur peut être recyclé vers la membrane plasmique
alors que le ligand est dégradé (ex : Low Density Lipoprotein, LDL) (Figure 30).
Dans ce cas, le recyclage des récepteurs s'effectue par l'intermédiaire de vésicules
de transport bourgeonnant à partir du compartiment endosomal précoce. Des
portions tubulaires comportant les récepteurs destinés à être recyclés se détachent
également de ce compartiment et fusionnent entre-elles pour former l’endosome
recyclant. De l’endosome recyclant bourgeonnent des vésicules qui transportent les
récepteurs vers la membrane plasmique. L’endosome recyclant peut être considéré
comme un site de stockage de récepteurs membranaires dont la sortie vers la
membrane plasmique est régulée en fonction des besoins de la cellule. Quant au
ligand, celui-ci est transféré du compartiment endosomal précoce vers le
compartiment endosomal tardif (Figure 31). Au cours de ce processus, les
endosomes précoces migrent dans la profondeur de la cellule, le long des
microtubules, alors que leur membrane s’invagine pour former des vésicules
internes. Ainsi se forment des corps multivésiculaires qui fusionnent entre eux et
créent le compartiment endosomal tardif dont le pH interne est de 6 du fait d’une
acidification progressive de leur contenu. Les vésicules du compartiment endosomal
tardif fusionnent ensuite avec des lysosomes préexistants. Cette fusion est à l’origine
des endolysosomes qui maturent en lysosomes après acidification supplémentaire
(pH 5). La dégradation des ligands (ou des récepteurs) est due aux hydrolases acides
qui proviennent du réseau trans de l’appareil de Golgi et qui sont délivrées par
transport vésiculaire dans le compartiment endosomal précoce (cf. cours sur

66
lysosomes). Ces hydrolases ont un pH optimal d’activité de 5. Elles sont synthétisées
sous forme proenzyme inactive car elles possèdent à leur extrémité N-terminale un
domaine inhibiteur. Les hydrolases acides ne deviennent actives qu’après élimination
par clivage protéolytique de ce domaine inhibiteur. Ainsi, les hydrolases acides ne
deviennent effectivement fonctionnelles qu’à partir du stade endolysosome où le pH
est suffisamment acide et où les protéases délivrées par la fusion de lysosomes avec
les vésicules du compartiment endosomal tardif effectuent ce clivage.
Figure 30. Endocytose des LDL. Le cholestérol utilisé par les cellules est en partie
d’origine alimentaire. Dans le foie, les molécules de cholestérol sont empaquetées dans les
LDL, petits agrégats constitués de phospholipides, de cholestérol et d’une protéine qui est le
ligand spécifique du récepteur des LDL.
b. Le récepteur et le ligand peuvent être dégradés dans les
lysosomes (ex : Epidermal Growth Factor ou EGF) (Figures 31 et 32).
Le rôle des corps multivésiculaires est particulièrement important dans ce cas car
ils permettent la dégradation complète des récepteurs qui se concentrent
préalablement dans la membrane des vésicules internes (qui est elle-même

67
dégradée). Les récepteurs destinés à être dégradés dans les vésicules internes des
corps multivésiculaires sont marqués par une ou plusieurs molécules d’ubiquitine (et
dans ce deuxième cas, les molécules d’ubiquitine sont liées au récepteur sur des sites
différents)
Figure 31.
Figure 32. Endocytose de l’EGF

68
c. Le récepteur et le ligand peuvent être recyclés vers la membrane
plasmique (ex : transferrine) (Figure 33).
Comme précédemment pour les récepteurs des LDL, le recyclage des complexes
ligand-récepteur s'effectue par l'intermédiaire de vésicules de transport
bourgeonnant à partir de l'endosome précoce.
d. Le récepteur et son ligand peuvent être transportés d'une face de
la cellule à l'autre.
Ce processus appelé trancytose s'observe notamment pour le transport
transcellulaire des anticorps présents dans le lait maternel. Dans cet exemple, les
anticorps maternels présents dans la lumière intestinale du nouveau-né se fixent,
dans ce milieu acide, sur des récepteurs présents sur la surface apicale des
entérocytes (récepteurs Fc). Les anticorps sont endocytés dans des vésicules
recouvertes de clathrine puis délivrés dans le compartiment endosomal précoce. Les
complexes anticorps-récepteurs Fc sont ensuite transférés par transport vésiculaire
vers la membrane plasmique baso-latérale de l’entérocyte, via l’endosome recyclant.
Lorsque ces complexes s’exposent dans le milieu extracellulaire bordant la surface
basolatérale de la cellule, les anticorps se libèrent de leur récepteur du fait du pH
neutre de ce milieu.
Figure 33. Endocytose de la transferrine. La transferrine est une protéine plasmatique qui
transporte le fer dans les cellules. Elle existe sous deux formes, l’apotransferrine (forme
n’ayant pas d’atome de fer et une faible affinité pour son récepteur à pH 7,4) et la
ferrotransferrine (forme ayant deux molécules de Fe+++ par molécule de transferrine et une
forte affinité pour son récepteur à pH 7,4). Dans le milieu extracellulaire (pH 7,4), la
ferrotransferrine se lie à son récepteur de surface. Dans l’endosome, du fait du pH acide, le
fer se dissocie de la transferrine. L’apotransferrine reste liée à son récepteur car elle a une
forte affinité pour celui-ci à pH acide. Le fer est transporté hors du système vésiculaire sur
une protéine de stockage, la ferritine. L’apotransferrine fixée sur son récepteur est recyclée
vers la membrane plasmique. Exposée sur la face extracellulaire de la membrane plamique,
l’apotransferrine est libérée dans le milieu extracellulaire du fait de sa faible affinité pour

69
son récepteur à pH 7,4. L’apotransferrine libérée fixe à nouveau des ions ferriques et peut
alors se lier à son récepteur.
La formation des vésicules recouvertes conduit également à l’internalisation des
fluides extracellulaires et donc de toutes les molécules qui y sont dissoutes. Cette
endocytose obligatoire et non spécifique est appelée endocytose en phase fluide.
2) Pinocytose non-dépendante de la clathrine.
Un deuxième type d’endocytose s’observe au niveau de régions spécialisées de la
membrane plasmique riches en cholestérol, glycosphingolipides et protéines liées par
un glycosylphosphatidylinositol (« lipid rafts »). Certains de ces microdomaines sont
recouverts sur leur versant cytosolique par une protéine membranaire intégrée, la
cavéoline, d’où leur nom de cavéole. Qu’il y ait ou non un revêtement de cavéoline,
ces microdomaines peuvent s’invaginer et, grâce à la dynamine, former des vésicules
d’endocytose. La cavéoline, lorsqu’elle est présente, ne se dissocie pas de la vésicule.
Ce mode d’endocytose, non dépendant de la clathrine, fait intervenir des récepteurs.
Il permet l’endocytose de nombreux ligands : cytokines (interleukine 2), virus (SV 40,

70
papilloma virus), toxines bactériennes, albumine. Le devenir des vésicules est mal
connu. Les vésicules, qu’elles soient ou non recouvertes de cavéoline, transitent par
un compartiment intermédiaire, le cavéosome. Le contenu de ces vésicules est
finalement délivré à l’appareil de Golgi ou au réticulum endoplasmique (Figure 34).
Dans les cellules endothéliales, ce mode d’endocytose indépendant de la clathrine
participe au transfert transendothélial de molécules (transcytose).
Figure 34
B. La phagocytose.
La phagocytose est une fonction de certaines cellules, utilisée pour endocyter des
particules de taille importante (micro-organismes, globules rouges, débris cellulaires).
Chez les mammifères, la phagocytose est surtout la propriété d'un type de cellules,
les cellules phagocytaires qui regroupent les polynucléaires, les monocytes et les
macrophages. La vésicule formée, appelée phagosome, est de grande taille ; son
diamètre est égal à celui de la particule ingérée. Le devenir du phagosome est de
fusionner avec des lysosomes pour former un phagolysosome dans lequel le matériel
endocyté est dégradé (voir lysosomes). Lorsque le matériel n'est pas dégradable par
les enzymes lysosomales, il persiste dans les lysosomes sous forme de corps résiduels.

71
Dans le cas contraire, les produits de dégradation sont utilisés pour les besoins du
métabolisme cellulaire.
Figure 35
Le processus de phagocytose est déclenché par la liaison de la particule à la
membrane plasmique de la cellule phagocytaire (Figure 35). Il s'agit donc d'un
processus qui nécessite une stimulation de la cellule, stimulation passant par
l'activation de récepteurs de surface chargés de transmettre l'information à l'intérieur
de la cellule. Les stimuli les mieux caractérisés sont les anticorps qui, dans le cas des
micro-organismes, se fixent par une extrémité à leur surface et qui, par l'autre
extrémité (Fc) se fixent sur des récepteurs pour la région Fc, présents à la surface des
polynucléaires et des macrophages. La fixation de telles micro-organismes (par

72
l'intermédiaire des anticorps) sur les cellules phagocytaires est responsable de
l'émission, par la cellule, de long prolongements appelés pseudopodes (ou voiles
hyaloplasmiques) qui vont entourer la particule puis fusionner à leur extrémité pour
former un phagosome. La formation des pseudopodes est due à la polymérisation
locale des filaments d’actine. D'autres classes de récepteurs cellulaires sont également
capables de promouvoir la phagocytose : il s'agit des récepteurs du complément ou
de récepteurs qui reconnaissent des oligosaccharides présents à la surface de certains
micro-organismes. Les macrophages sont également capables de phagocyter des
cellules en apoptose. Au cours de ce processus de mort cellulaire, la cellule perd la
distribution asymétrique des phospholipides de la membrane plasmique ; les
phosphatidylsérines, normalement présentes dans la monocouche cytosolique,
s’exposent sur la face extracellulaire de la membrane et déclenchent la phagocytose
de la cellule apoptotique.
IX. Les lysosomes.
A. Caractéristiques.
Les lysosomes sont des vésicules limitées par une membrane, dont la forme et la
taille sont très variées. On peut les détecter en microsocopie optique après marquage
cytoenzymologique de la phosphatase acide. Ils sont caractérisés par (Figure 36) :
- le pH acide de leur lumière, aux environ de 5 ;
- leur contenu riche en hydrolases acides, actives à pH 5, (protéases, nucléases,
lipases, glycosidases, phosphatases, sulfatases, phospholipases) ;
- une pompe à protons membranaire qui utilise l'énergie produite par l'hydrolyse
de l'ATP pour pomper des ions H+ du cytosol vers l'intérieur des lysosomes et
maintenir ainsi un pH acide ;
- des protéines membranaires fortement glycosylées ; la glycosylation protège ces
protéines de la protéolyse par les protéases contenues dans la lumière des
lysosomes ;
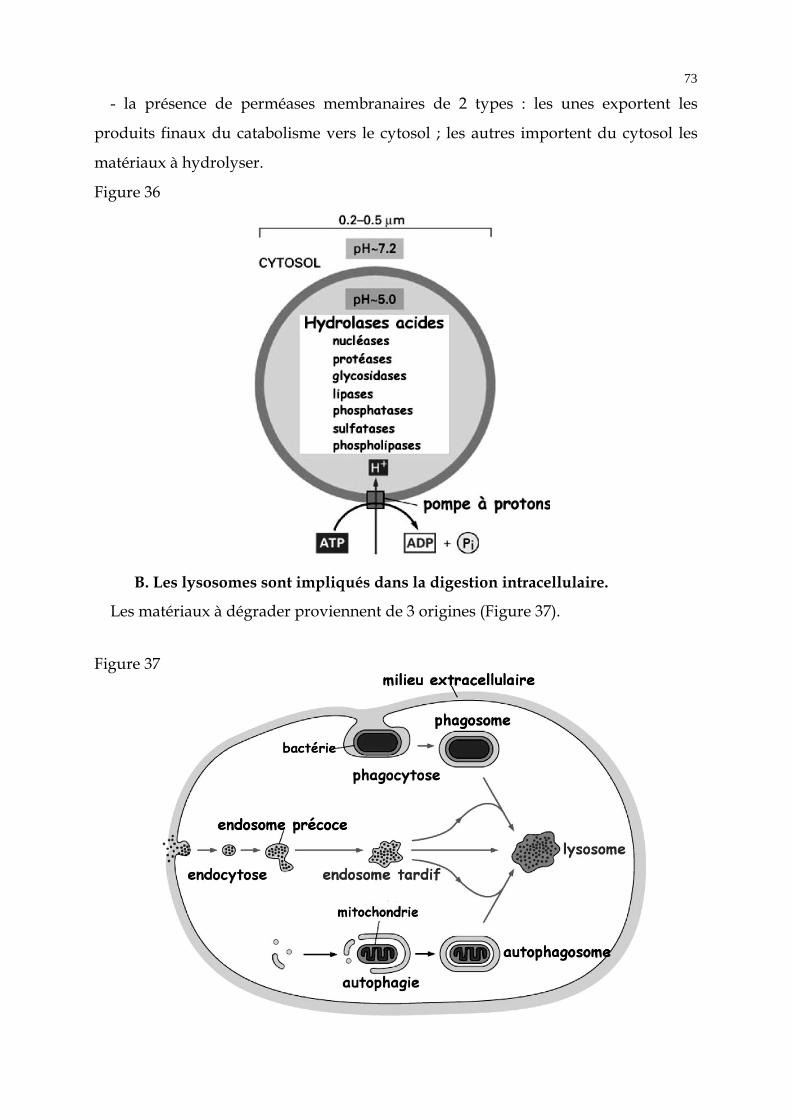
73
- la présence de perméases membranaires de 2 types : les unes exportent les
produits finaux du catabolisme vers le cytosol ; les autres importent du cytosol les
matériaux à hydrolyser.
Figure 36
B. Les lysosomes sont impliqués dans la digestion intracellulaire.
Les matériaux à dégrader proviennent de 3 origines (Figure 37).
Figure 37

74
1. La voie de l'endocytose.
La voie de l’endocytose (pinocytose, phagocytose) apporte des matériaux captés
dans le milieu extracellulaire.
2. La voie de l'autophagie.
Cette voie a pour rôle d'éliminer les organites cellulaires hors d'usage. Ainsi, dans
l'hépatocyte, la durée de vie moyenne d'une mitochondrie est d'environ 10 jours. Au
cours de ce processus, l'organite est enveloppé par une membrane dont l’origine
n’est pas connue, créant ainsi un autophagosome. L’autophagosome fusionne
ensuite avec un lysosome ou un endosome tardif. L’autophagie est un phénomène
également observé en cas de carence nutritive. Dans cette situation, des portions du
cytosol sont captées dans des autophagosomes ; après fusion avec des vésicules du
système lysosomal, les métabolites provenant de la dégradation des molécules
capturées sont alors réutilisés par les cellules pour leur survie. L’autophagie est aussi
impliquée dans le développement foetal.
3. Le passage direct de peptides cytosoliques.
Des peptides cytosoliques pénètrent dans les lysosomes grâce à des perméases
présentes dans la membrane de l’organite. C'est en particulier le cas des protéines
ubiquitinées et partiellement dégradées dans le cytosol par les protéasomes et qui
possèdent un signal de tri vers les lysosomes.
Toutes les familles de molécules biologiques sont dégradées dans les lysosomes en
métabolites élémentaires. Ceux-ci repassent ensuite dans le cytosol via des perméases
où ils sont réutilisés pour le métabolisme cellulaire.
C. Tri des enzymes lysosomales solubles de l'appareil de Golgi vers les
lysosomes (Figure 38).
La biosynthèse de ces enzymes obéit au schéma général de synthèse des
glycoprotéines solubles : elles sont importées et N-glycosylées dans le RE, puis
transportées dans l'appareil de Golgi. Les vésicules qui transportent ces protéines
vers le système endosomal bourgeonnent à partir du compartiment trans de
l'appareil de Golgi. C'est à ce niveau que s'effectue le tri car ces vésicules doivent

75
exclure les autres protéines dont les destinations sont différentes. Le mécanisme du
tri des hydrolases lysosomales est connu depuis quelques années (Figure 38).
1. Rôle du mannose-6-phosphate.
Les hydrolases lysosomales comportent un marqueur sélectif, sous la forme de
groupements mannose-6-phosphate (M6P) qui sont ajoutés aux oligosaccharides
(résultant de la N-glycosylation) de ces enzymes dans la lumière du compartiment
cis du Golgi. Cette réaction est catalysée par 2 enzymes :
- la N-acétylglucosamine phosphotransférase (NAcGlc-P-transférase) qui transfère
le groupement NAcGlc-P sur 1 ou 2 résidus mannose de l'oligosaccharide ;
- une phosphoglycosidase qui retire le résidu NAcGlc pour exposer le M6P.
Il a été suggéré que la NAcGlc-P-transférase reconnaissait spécifiquement les
hydrolases destinées au lysosome grâce à l'existence, sur ces enzymes, d'une région
signal.
Figure 38

76
2. Le récepteur du mannose-6-phosphate (Figure 38).
La membrane du compartiment trans de l'appareil de Golgi contient des
récepteurs du M6P qui reconnaissent donc spécifiquement les hydrolases
lysosomales phosphorylées. Ces récepteurs, sur lesquels se fixent les hydrolases
lysosomales, se lient par leur domaine cytosolique à des adaptateurs et se
concentrent dans des puits recouverts de clathrine qui bourgeonnent pour former
des vésicules recouvertes. Ces vésicules perdent ensuite leur enveloppe et fusionnent
de façon spécifique avec un endosome précoce qui, après acidification
supplémentaire, devient un endosome tardif. La libération des hydrolases
lysosomales dans l’endosome tardif est provoquée par le pH acide de ce
compartiment. En effet, le M6P se fixe sur son récepteur à pH 6,5-6,7 (pH du
compartiment trans) et s'en dissocie à pH 6, pH de l’endosome tardif. Les hydrolases
libérées sont déphosphorylées. Après avoir libéré leurs enzymes, les récepteurs sont
recyclés vers la membrane du compartiment trans du Golgi (recyclage membranaire)
dans des vésicules recouvertes de rétromères. Il a été montré qu'une petite fraction
des récepteurs du M6P était également présente dans la membrane plasmique. Ces
récepteurs pourraient servir à recapturer des enzymes lysosomales transportées par
erreur vers la membrane plasmique et libérées dans le milieu extracellulaire. Les
enzymes récupérées sont recyclées vers les endosomes par endocytose.
3. Les maladies lysosomales.
L'étude du mécanime de ces maladies a joué un rôle prépondérant dans la
caractérisation du mécanisme d'adressage des hydrolases lysosomales. Quatre
mécanismes différents peuvent conduire à l'accumulation de molécules non ou mal
dégradées dans les lysosomes.
a) Absence ou anomalie fonctionnelle d'une ou de plusieurs
enzymes.
Elles sont habituellement dues à une mutation dans un gène de structure qui code
pour une enzyme particulière, provoquant ainsi une accumulation de son substrat
non digéré dans les lysosomes.
Exemples :

77
- la sphingolipidose ou maladie de Gaucher, due à l'accumulation des
sphingolipides non hydrolysés dans les lysosomes, en particulier dans le système
nerveux.
- La forme la plus grave de ces maladies est la mucolipidose de type II (maladie
des cellules I). Elle est due à une anomalie ou à l'absence de la NAc Glc-P-
transférase. Dans cette maladie, les enzymes sont normalement synthétisées et
fonctionnelles mais ne sont pas délivrées aux lysosomes du fait de l'absence de M6P.
Ces enzymes sont alors sécrétées dans le milieu extracellulaire.
b) Anomalie congénitale des perméases membranaires qui ne
permettent plus le passage dans le cytosol des métabolites provenant de
l'hydrolyse.
Exemple : la cystinose par défaut de transport de la cystine vers le cytosol.
c) Une acidification insuffisante du lysosome qui altère le
fonctionnement des enzymes lysosomales, normalement actives à pH 5. Ce
mécanisme est utilisé en thérapeutique pour détruire l'agent du paludisme en
bloquant par la quinine le fonctionnement des lysosomes du parasite responsable de
la maladie.
d) L'incapacité des lysosomes à dégrader des matériaux non
biologiques.
Ce mécanisme n'intervient que dans les macrophages qui sont capables de
phagocyter de grosses particules minérales ou métalliques. Ce phénomène est
observé au cours des pneumoconioses, lorsque de telles particules inhalées sont
phagocytées par les macrophages des alvéoles pulmonaires. Cette accumulation, qui
provoque la libération du contenu enzymatique lysosomal dans le cytosol du
macrophage, est responsable de la mort de la cellule suivie de la destruction
progressive du tissu pulmonaire. Deux exemples en pathologie humaine sont
représentés par l'asbestose (accumulation de fibres d'amiante) et la silicose
(surcharge en particules de silice).