Embed Size (px)
Citation preview

Mutation Research Elsevier Publishing Company, Amsterdam Printed in The Netherlands
491
ACTION MUTAGI);NE DES UV SUR LES OVOGONIES DE DROSOPHILES
II. EFFETS DU DOUBLE I~CLAIREMENT ET DU FRACTIONNEMENT SUR
LE POURCENTAGE DE MUTATIONS
JACQUELINE PROUST E'r ANNE FLAMAND
Laboratoire de Biologie Expdrimentale, Facultd des Sciences d'Orsay, Universitd de Paris (France)
(Re~u le 3 ° juin 1966 ) (Manuscrit revis6 re~u le 16 novembre 1966)
SUMMARY
Mutagenic action of UV on the oogonia of Drosophila
II. Effects of double-illumination and of dose-fractionation on mutation rate
Recessive lethal mutations are induced by UV light in Drosophila oogonia by irradiating in vitro ovary primordia which afterwards are implanted into host larvae. A constant dose of 2000 ergs/mm 2 is used with the following 3 methods of irradiation: (a) The whole dose is given at once in one direction (control). (b) The dose is given in 2 halves separated by a 2-h interval without change of direction. (c) Half of the dose is given in one direction and the other half in the opposite direction.
As compared with the control, method (b) leads to a decrease both in the muta- tion frequency and in the absolute size of the cluster of identical lethals. Method (c) leads to an increase ill the mutation frequency and a decrease in both the probability of success of the implantation and the size of the total progeny produced by the implanted ovary, Differences between effects after t reatment according to (c) and the control favour the assumption that the different cell layers of the primordium do not receive the same amount of radiation. At high doses the recovered eggs tend to arise only from the less damaged oogonia. This mechanism can account for part of the decrease in the frequency of recovered mutations which is observed at high doses.
Differences between the effects after t reatment according to (b) and the control point to an additional mechanism. The correlation between the size of the clusters and the mutation frequency which is obtained is quite general. I t is also observed when high and low doses are compared. The reduction of cluster size is probably the outcome of a delay imposed by the irradiation on the resumption of DNA synthesis. With this delay, the probability that premutational lesions undergo "dark repair" is increased.
Mutation Res., 4 (1967) 491-5Ol

492 JACQUELINE PROUST, ANNE FLAMAND
INTRODUCTION
Apr6s irradiation aux UV d'6bauches ovariennes pr61ev6es sur des larves, nous avons observ6 une relation dose-fr6quence non lin6aire pour les mutat ions 16tales r6cessives induites darts le second chromosome 20,21.
La courbe repr6sentative passe par un max imum qui se situe aux environs de Iooo ergs/mm 2 (r6f. 2o, 2I). Cette non lin6arit6 a 6t6 retrouv6e par d 'autres exp6ri- mentateurs sur divers organismes avec les UV 4,18 mais aussi, pour de trhs faibles doses, avec les radiations ionisantes 19. Outre cette non lin6arit6, dans certains cas, un effet d' intensit6 et un effet de fract ionnement de la dose ont 6t6 observ6s quelle que soit la radiation utilis6e 2,53 (UV, R.X., R.7). Tous ces faits tont ressortir la complexit6 du processus de la mutag6n6se. Parmi toutes les hypoth6ses propos6es pour leur inter- pr6tation, 2 d 'entre elles ont 6t6 retenues.
La premiere fait appel ~ un m6canisme se si tuant au niveau de la population des cellules irradi6es. Toutes les cellules du massif ovogonial ne sont pas 6clair6es d 'une 6gale mani~re. Cette in6galit6 dans la dose reCue introduit une h6t6rog6n6it6 20,51 qui se manifeste ~ 2 niveaux: au moment de la diff6renciation des ovafioles et apr6s isolement des ovogonies dans les ovarioles 2%21.
La seconde hypoth6se fait intervenir des processus jouant au niveau de la cel- lule individuelle. I1 s 'agit des proeessus de restauration qui entrainent une gu6rison des 16sions pr6mutationnelles et se manifestent pendant une p6riode de temps limit6e qui correspond sensiblement "a la premiere moiti~ de l ' interphase 7,14-17,a2. Quelles que soient les modalit4s du processus de restaurationa,s-t°,22,2¢,25,34, une alternative existe pour le sort des ldsions pr6mutationnelles; elles peuvent, soit se stabiliser sous forme de mutat ions ddfinitives, soit revenir ?t l 'dtat initial, c'est-~-dire gudrirT,14,16, a3. En outre, le jeu du processus de restaurat ion permet un second type de photorestau- ration que l 'on qualifie d'indirecte11. On sait que les longueurs d'ondes voisines des UV provoquent aussi bien en prd- qu 'en post- t ra i tement un allongement de la pdriode sensible qui permet donc une restauration spontan6e plus efficace 15. Le mode d 'act ion de ce type de photorestaurat ion indirecte serait, donc, de favoriser la restauration sombre 1°. Sa part icipation peut ~tre envisag6e en tant que cause majeure de la redescente de la courbe. En effet, nous avons observd qu 'aux doses fortes les mitoses repartent plus tard, ainsi, les l~sions pr6mutationnelles induites ont davantage de chances d 'etre restaur4es ~0,2~.
Ces 2 m~canismes, hdt6rogdndi% de position et photorestaurat ion indirecte ne s 'exeluent pas et la redescente de la courbe peut ~tre due ~ leur intervention eonju- gude. Pour essayer de d6terminer la part de responsabilit6 qui revient ~ chacnn d'eux, nous avons effectud des expdriences en adminis t rant une m~ine dose de 20o0 ergs/mm ~ de 3 mani~res diff4rentes. Les dbauches re¢oivent cette dose: (I) dans un seul sens et en une seule fois; (2) successivement, sans ehangement de position, dans 2 directions oppos~es; (3) dans un seul sens, en 2 fractions 6gales, sdpardes par un intervalle de temps.
La dose 2000 ergs/mm 2 a ~td choisie parce qu'elle est situ6e dans la partie descendante de la courbe.
3lulation Res., 4 (1967) 49I-5°~

ACTION U V SUR LES OVOGONIES DE DROSOPHILES. I [ . 493
MATI~RIEL ET M]~THODE
L'irradiation est appliqu~e in ~,itro ~ des ~bauches ovariennes pr61ev6es par dissection sur des larves. Elles sont ensuite implant6es dans des larves h6tes selon ]a technique raise au point par EPHRUSSI ET BEADLE.
Les larves donneuses sont h6t6rozygotes pour ]es g6nes r~cessifs black et purple et les larves h6tes sont homozygotes Bar.
Les 2 lampes utilis6es sont des lampes germicides ~ faible pression de mercure dont 90% de l'~nergie 6mise correspond ~ ]a longueur d'onde de 2537 ,/k. Les tech- niques d ' implantation et d'irradiation ont 6t6 d6crites dans ]e mdmoire prdc6dent 5, L'6clairement dans 2 directions oppos6es est obtenu en pla~ant les 6bauches ovariennes dans une lame creuse de quartz entre 2 lampes identiques qui se font face. Nous avons manipul~ en lumi6re rouge pour 6viter l ' intervention du processus de photorestaura- tion. L'intensit6 a 6t6 choisie constante, soit 60 ergs/mm ~ par sec.
Nous avons recherch6 les l~taux induits sur le chromosome I I en utilisant la mdthode 'Sifter' et les tests d'all61isme intra-descendances. Ces techniques g6n6tiques ont 6t6 d6crites pr6c6demment.
Nous avons, dgalement, conserv6 dans ce travail, les r~gles d6j~ adopt~es dans le premier m6moire concernant le d6nombrement des clones 16taux induits.
PLAN DES EXPI~RIENCES
Double dclairement Deux s6ries d'exp6riences sont ex6cut6es parall~lement. Dans la premiere s6rie, les ~bauches ovariennes re~oivent 2000 ergs/mm 2 dans
une seule direction, seule, i des 2 faces de celles-ci est expos6e aux UV. Dans la seconde s6rie, les 6bauches ovariennes re~oivent IOOO ergs/mm 2 dans
une direction, imm6diatement apr~s et sans avoir subi aucun changement, elles re~oivent ~t nouveau iooo ergs/mm 2 dans une direction oppos6e. Les 2 faces des 6bauches sont ainsi expos6es aux UV.
Fractionnement Une premiere s6rie d'6bauches ovariennes est exposde ~ une dose de IOOO ergs/
m m 2 aussit6t apr~s la dissection. A la suite d 'un s6jour de 2 h dans le liquide physiolo- gique, elles re~oivent {L nouveau iooo ergs/mm 2, puis sont imm6diatement implant6es.
Parall~lement, dans une seconde sdrie, les ~bauches ovariennes, issues de la m~me population larvaire, re~oivent une dose de 2000 ergs/mmL en une seule fois. Elles s6journent 2 h darts le Ringer avant d'etre implant6es.
Un t~moin, non implant6, permet de connaltre le taux de mutabilit6 spontan6e et la fr6quence des l~taux prdexistants. Pour diminuer cette cat6gorie de 16taux, les souches b pr et b + pr+, ont dt6 rendues homozygotes pour le second chromosome, avant d'etre utilis6es.
RI%ULTATS ET DISCUSSION
Fiddlitd des rdsultats obtenus dans 3 rdpdtitions du m~me type d'irradiation Parmi les r6sultats obtenus ~ la dose de 2000 ergs/mm 2 distribute dans une
Mutation Res., 4 (1967) 491-5°1
4 9 4 JACQUELINE PROUST, ANNE FLAMAND
<
<
~a ©
r, -a.I m
N Z
~2
N "N
Z Z ©
<
L
;< ~2
Z
.<
<
N ©
~2 N
N
Z ©
m
m M
m
Z
N <
<
,~ ~ . ~
~~
7 7 oo4 , I I
b-. c~.
5 o !! -+i
r-i ~0
d 6
% ~-
d ~
,2 ~ ~
~ c
~ ~
O o 6 ~
~ d
"~ o ~-
ee~
©
6
oo
6
oc
i
6
©
o
Mutation Res., 4 (1967) 491-5Ol

ACTION U V SUR LES OVOGONIES DE DROSOPHILES. I I . 495
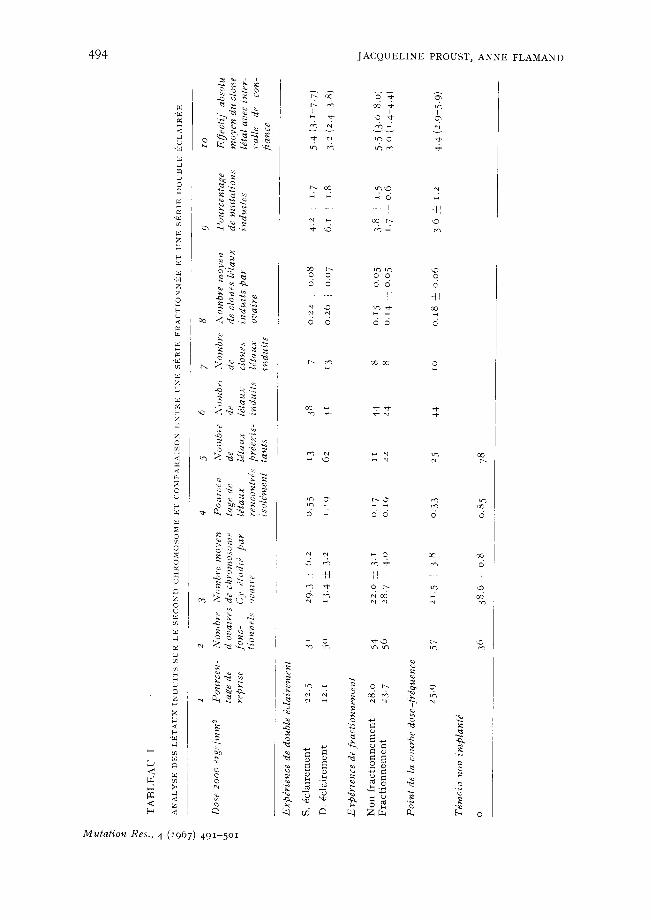
seule direction et en une seule fois, le premier d'entre eux constitue l 'un des points de la courbe dose-fr6quence, les 2 autres sont les t6moins des exp6riences 'double 6clairement' et ' fractionnement ' rapport6es dans cet article.
En outre, il est ~ noter que les 2 premi6res exp6riences relatives au double 6clairement ont ~t6 r6alis6es ~ des moments diff6rents par le m~me manipulateur et celles relatives au fractionnement par un autre manipulateur.
Dans les Tableaux I et II, il apparalt que les 3 r6pdtitions du m6me type d'irradiation ont conduit aux m~mes valeurs k la lois pour le pourcentage de mutations et l'effectif absolu du clone 16tal. Les r6sultats obtenus par la m6thode d' implantation des 6bauches ovariennes sont donc relativement fid~les, malgr6 la grande variabilit6 observ6e dans l'effectif des descendances 6tudi6es.
Double dclairement Reprise et fdconditd des ovaires implantds. Le double 6clairement diminue le
pourcentage de reprise de l ' implant, il passe de 22.5 ~ 12.1% (Tableau I, colonne i). D'autre part, il conduit ~ une baisse importante de la f6condit6 moyenne de l'ovaire implant6 (Tableau I, colonne 3). Cette f6condit6 est 6valude el1 hombre de chromo- somes Cy; comme il a 6t6 expliqu~ dans le precedent m6moire, il suffit de multiplier ce hombre par un facteur constant 2.06, pour obtenir le volume r6el de la descendance.
L'action d'un simple dclairement sur la fdcondit~ des ovaires implant6s a d6j~ 6t6 discutde dans la publication pr~c6dente. Les points principaux de cette discussion seront rappel6s ici. La greffe, par elle-m~me sans l ' intervention d'aucune irradiation, entraine une baisse de f6condit6 de l 'ovaire implant6 fonctionnel, due au fait que seul un tr~s petit nombre d'ovarioles r6ussissent k s 'aboucher correctement ~ l'oviducte. A 2000 ergs/mmL /~ ce ph6nom6ne m6canique du blocage des ovarioles, s 'ajoute l 'action cytocide de l'irradiation.
Dans le cas du double 6clairement, nous avons observ6, malgr6 l'identit6 de la dose totale revue, une action beaucoup plus accentu6e de l 'irradiation k la fois sur la fr6quence de r6cup6ration d'un ovaire fonctionnel et sur la f6condit6 moyenne de celui-ci. On dolt donc penser que l'6clairement de l'6bauche sur ses 2 faces, sans doute parce qu'il atteint plus uniform6ment l 'ensemble des cellules somatiques responsables de la formation des ovarioles, entraine de fortes perturbations dans le d6veloppement de l'ovaire. Ces perturbations sont souvent trop importantes pour que l'ovaire puisse ~tre fonctionnel; lorsqu'il l'est, il ne poss~de, sans doute, que peu d'ovarioles nor- maux correctement abouch6s ~ l'oviducte. Ces observations sont en faveur de l 'hypo- th&se, selon laquelle, dans le cas de l%clairement sur une seule face, les 616ments fonctionnels r6cup6r6s doivent essentiellement d6river des parties prot6g6es de l'6- bauche. Malgr6 la faible 6paisseur de celle-ci, les cellules ne seraient donc qu'irr6gu- li~rement atteintes par le rayonnement.
Dimensions des clones ldtaux. Les 16taux sont rencontr6s dans la descendance sous forme de clones dont la taille tant absolue que relative est variable. Le Tableau I I pr6sente les r6sultats relatifs aux d6nombrements de ces clones (moyenne et variance).
Tout d'abord, on volt que dans la s6rie double ~clairement, l'effectif absolu moyen des clones est plus petit que dans la s6rie simple 6clairement. La diff6rence est significative au niveau 0:oi (t ---- 2.2, d = 18, P < o,oi). D'autre part, dans chaque cas, le nombre" d'ovogonies qui sont fi l'origine de la descendance a 6t6 calcul6 comme il a 6t6 indiqu6 prdc6demment 2o,21. Ce calcul r6v~le que les 2 s6ries different ~galement
Mutation Res., 4 (1967) 491-5Ol
496 J A C Q U E L I N E PROUST, A N N A F L A M A N D
5 ̀ce poin t de vue. Dans le cas du double 6clairement, le nombre moyen d 'ovogonies originelles est 2.2 contre 4.9 pour le s imple 6clairement. L '6car t n 'es t cependant pas significatif (t - - 1.49, d = 18, o.2o > P > o.Io). La baisse observ6e dans le nombre d 'ovogonies 6chanti l lonn6es n 'es t pas forcdment due 5, une des t ruc t ion directe des ovogonies par le r ayonnement , elle peut tout aussi bien 6tre la cons6quence de 16sions provoqu6es dans les cellules somat iques de l '6bauche ovarienne. Celles-ci, en effet, peuvent en t ra iner de graves pe r tu rba t ions dans la s t ruc ture de l 'ovai re et l '61imination de nombreuses ovogonies dans des ovarioles non fonctionnels.
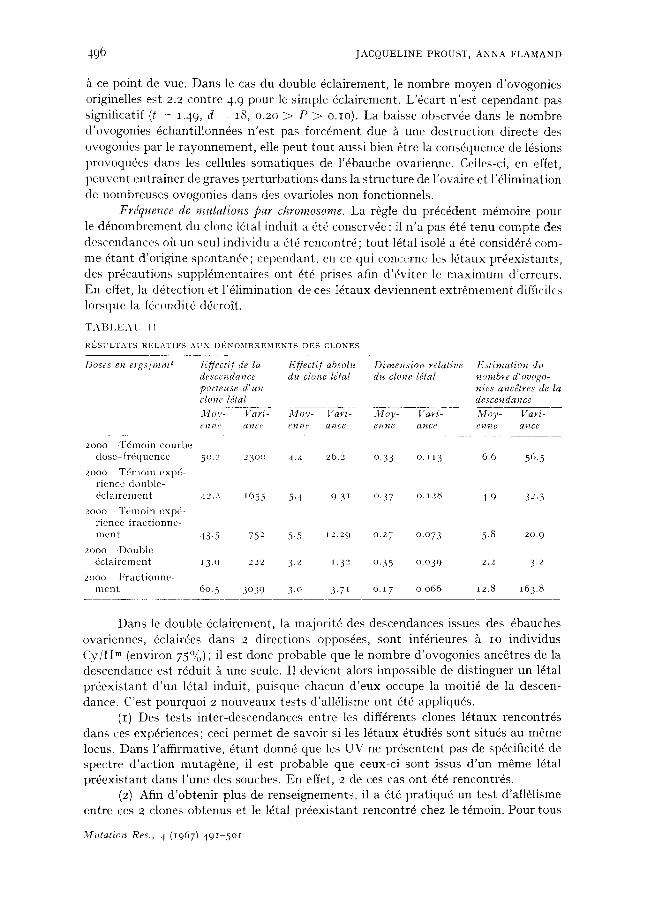
Frdquence de mutations par chromosome. La r6gle du prdc6dent m4moire pour le d6nolnbrement du clone 16tal indui t a 6t6 conserv6e : il n ' a pas 6t6 tenu compte des descendances oh un seul indiv idu a 6t6 rencontr6; tou t 16tal isol6 a 4t6 consid6r6 corn- me 6tant d 'or igine spontan6e; cependant , en ce qui concerne les 16taux pr6exis tants , des prdcaut ions suppl6menta i res ont 6t6 prises afin d '6v i te r le m a x i m u m d 'erreurs . En effet, la d6tect ion et l '61imination de ces Idtaux dev iennent ex t r6mement difflcilcs lorsque la f6condit6 ddcrolt .
T A B L E A U 1 I
RI 'SULTATS RELATIFS AUX DI~2NOMBREMENTS DES CLONES
Doses en ergs/mm 2 F.ffectif de la Effeclif absoh~ Dimension relative Estimation du descendance du clone ldtal d~* clone h;lal ~ombre d'ovogo- porteuse d'un hies a~cdlres de la clone Idtal descendance
Mo v- Vari- Moy- Vari- Moy- Vari- Moy- Vari- eJz t~e a t l c e e n t ? e a~' lce e~'lFIC a n t e e n l q e a ~ c e
2 0 0 0 - T 6 m o i n c o u r b e d o s e - f r 6 q u e n c e 50.2 2300 4.4 26 .2 0 .33 o. ~ 13 6 .6 56.5
2o00 T d m o i n e x p d - r i e n c e d o u b l e - d c l a i r e m e n t 42.2 1655 5 .4 9.31 0 .37 o .~28 4 .9 32.3
2 o o o - - ' F d m o i n e x p d - r i e n c e f r a c t i o n n e m e n t 43 .5 752 5.5 12.29 o .27 o . o 7 3 5.8 20 .9
2 o o o - - D o u b l e 6 c l a i r e m e n t 13.9 222 3.2 1.32 o .35 o . o 3 9 2.2 3.2
2ooo F r a c t i o n n e - n l e n t 6o. 5 3o39 3 .o 3.71 o . i 7 o . o 6 6 12.8 163 .8
Dans le double 6clairement, la major i t6 des descendances issues des 6bauches ovariennes, 6clair6es dans 2 direct ions oppos6es, sont inf6rieures 5 ̀ IO individus C y / I I m (environ 75°/~) ; il est donc probable que le nombre d 'ovogonies anc~tres de la descendance est r6dui t 5 ̀une seule. I1 devient alors impossible de d is t inguer un 16tal p r6ex is tan t d ' un 16tal induit , puisque chacun d ' eux occupe la moit i6 de la descen- dance. C'est pourquoi 2 nouveaux tests d'all61isme ont 6t6 appliqu6s.
(I) Des tes ts in ter -deseendances entre les diff6rents clones 16taux reneontr6s dans ces exp6rienees; ceei pe rme t de savoir si les 16taux 6tudi6s sont situ6s au m~me locus. Dans l 'af f i rmat ive, 6rant donn6 que les UV ne pr6sentent pas de sp6cificit6 de spectre &ac t ion mutag6ne, il est probable que ceux-ci sont issus d 'un m~me 16tal p r6exis tan t dans l 'une des souches. En effet, 2 de ces eas ont 6t6 rencontr fs .
(2) Afin d 'ob ten i r plus de renseignements , il a 6t4 pra t iqu6 un tes t d'all61isme entre ces 2 clones obtenus et le 16tal p r6exis tan t rencontr6 chez le t6moin. Pour tous
Mutation Res., 4 (1967) 4 9 I - 5 ° z

ACTION U V SUR LES OVOGONIES DE DROSOPHILES. I I 497
ces tests la r@onse a 6t6 affirmative, tous ces 16taux 6taient identiques, Ils ont donc 6t6 consid6r6s comme un clone unique de 16taux pr6existants.
Dans les descendances de tr6s faibles effectifs, le clone 16tal induit peut se r6duire ~t un individu. Ces cas extrfimes viendront augmenter la classe des 16taux spontan6s; de ce faR, le pourcentage de mutations induites se trouvera sous-estim6. Cependant, cette erreur n'est pas tr6s importante, 6tant donn6 que le pourcentage de 16taux spontan4s rencontr6s dans les exp6riences du double 6clairement ne diff~re pas significativement de celui du tdmoin (Tableau I). L 'examen de la colonne 9, du Tableau I, montre que le pourcentage de mutations 16tales r6cessives est plus 61ev6 dans le cas du double 6clairement que dans eelui du simple 6clairement. Ceci rdsulte, non pas d'une variation dans la fr6quence des clones 16taux par ovaire, qui est sensi- blement la m~me dans les 2 cas (colonne 8), mais de la rfduetion du nombre moyen d'ovogonies participant ~ la descendance. La diff6rence est ~ la limite du seuil de signification. Si cette diff6rence est r6elle, elle apporte, 4videmment, un argument en faveur de l ' intervention de l'h6t6rogfn6it6 de position des eellules irradi6es dans la cin6tique dose-effet observ6e. Elle pourrait ~tre une cause majeure de la redescente de la courbe.
Fr actionneme~zt Reprise el fdconditd des ovaires implantds. On voit sur le Tableau I que le pour-
centage de reprise de l 'ovaire implant6 (colonne I) et la f6condit6 moyenne de cet ovaire (colonne 3) sont sensiblement les m6mes darts les 2 s6ries.
F.tude des clones ldtaux. L'effectif absolu moyen du clone 16tal est plus petit~dans la s6rie fractionn6e (colonne IO, Tableau I). La diff6rence est statist iquement signi- ficative (t - - 2.97, d ~ 14, P < o.oi). Le fractionnement de la dose 2000 ergs/mm 2 a donc diminu6 le hombre moyen de mitoses que les ovogonies irradi6es subissent avant d'en arriver ~t la production des ceufs. Cette baisse du rendement moyen d'une ovo- gonie individuelle ne s 'accompagne pas d'un ehangement comparable dans la f6con- dit6 globale de l'ovaire. Celle-ci serait plut6t augment6e quoique d'une mani6re non significative (colonne 3). I1 en r6sulte, fividemment, que le nombre d'ovogonies 6chan- tillonn6es est plus grand dans la sdrie oh la dose a 6t6 fractionn6e que dans le t6moin. I1 passe de 5.8 ~t 12.8; cet 6cart n'est toutefois pas significatif (t = 1.36, d = 14, 0.20 ..'> P > o.Io). Darts la mesure oh il est r6el, il pourrait indiquer que le fractionne- merit de la dose diminue l'effet eytocide des UV.
Frdquence de mutations par chromosome. L'examen de la colonne 9 du Tableau I montre que le poureentage de mutations 16tales rdcessives est plus bas dans la s6rie fractionn6e que dans la sdrie non fractionn6e. La diff6rence est tr6s pros du seuil de signification. Toutefois, si eette diff6rence est r6elle, l ' interprdtation la plus simple est que la prolongation du temps d'arrbt de la premi6re division qui suit l 'irradiation provoqu6e par le fraetionnement favorise la restauration sombre.
CONCLUSION
Les exp6riences rapport6es ci-dessus ont 6t6 entreprises dans le but de tenter d'expliquer le caract~re non lin6aire de la relation dose-frdquence trouv6 pr6c6dem- ment pour la production des 16taux r6cessifs par l 'irradiation UV d'ovogonies de Dro- sophiles. Elles ont apport6, tout d'abord, la d6monstration de la fid61it6 des r6sultats.
Mulalion Res., 4 (1967) 491-5°1

498 JACQUELINE PROUST, ANNE FLAMAND
En ce qui concerne le caract~re non lindaire de la relation dose-effet, l'interven- tion de 2 m6canismes distincts avait 6t6 sugg6rde 5 titre d'hypoth~se de travail.
(I) L'irrdgularit~ dans la dose recue par les diffdrentes cellules d'une ~bauche. (2) La photorestauration indirecte qui favorise le processus spontan6 de restau-
ration sombre. Si la redescente de la courbe est due uniquement ~ l'h6t6rogfin6it~ de position
des cellules irradides (hypoth~se I), nous devons nous attendre, dans le cas du double dclairement, /~ observer davantage de clones ldtaux. En effet, 2 fois plus de cellules supcrficielles sont expos~es aux UV; mais la dose reCue par chacune d'elles n'est que de I000 ergs/mm 2. L'effet cytocide ~ I000 ergs/mm 2 est n~gligeable, par cons6quent les cellules mut6es subsistent, alors que dans l'hypoth~se envisagde, elles auraient ~t~ ~limindes si elles avaient re~u la dose de 2ooo ergs/mm ~. Dans le cadre de cette hypoth~se, le fractionnement, par contre, n'a aucune raison d'influer sur la forme de la courbe.
Si la redescente de la courbe n'est, au contraire, que le r6sultat du jeu de la photorestauration indirecte (hypoth~se 2), nous ne devons pas observer d'augmenta- tion du pourcentage de mutations dans le cas du double 6clairement dans deux direc- tions.~En effet, toutes les cellules auraient re~u la m~me dose, avec la m~me intensitd, quelle soit donn~e dans une seule ou dans 2 directions. Par contre, le fractionnement de la dose doit conduire ~ une modification du pourcentage de mutations. En effet, la seconde dose administr6e pendant la p6riode sensible peut ~tre considdrde comme une sorte de post-traitement et aura comme r6sultat d'allonger encore la p6riode de temps pendant laquelle peut s'effectuer la restauration sombre spontan6e.
Les r6sultats rapportds ci-dessus semblent bien montrer que ces 2 m6canismes interviennent l'un et l'autre. La forme de la relation dose-effet serait la consdquence globale de leur jeu simultand. Les exp6riences d'6clairement dans 2 directions oppo- s6es mettent en 6vidence l'existence du premier m6canisme. On observe, en effet, que ce type d'irradiation conduit ~ un rfisultat qui diff~re de celui du t~moin de 2 mani~res : (i) Le pourcentage de 16taux r~cessifs induits est augment6. (2) La probabilit6 de reprise de l'6bauche et sa fertilit6 moyenne sont diminu6es.
Ces 2 differences r6v~lent sans doute l'une et l'autre que l'irradiation dans une seule direction s'accompagne d'une h6t6rog6n6it6 de la dose revue par les cellules individuelles. Le fait de distribuer la dose en 2 parties, dans 2 directions oppos6es, diminue consid6rablement cette hdt6rog6n6it6. Ainsi, les cellules somatiques respon- sables du ddveloppement ultdrieur de l'6bauche, sont uniform6ment atteintes, ce qui a pour cons6quence d'entrainer davarttage de perturbations dans la fixation de l'6bauche et dans la formation des ovarioles. De m~me, la population d'ovogonies doit, dans le cas du double dclairement, avoir reCu une dose plus ou moins homog~ne, alors que, dans le cas de l'6clairement dans une seule direction, elle se sdpare sans doute en une fraction fortement endommag6e et une fraction faiblement atteinte. Si la premiere n 'a que peu de chance de participer 5 la descendance, il est normal d'observer, dans ce cas, une fr6quence de mutations plus faible.
Les r6sultats des expdriences de fractionnement de la dose en 2 moiti6s s6par6es par un intervalle de temps, sans changement de direction, paraissent bien, par contre, impliquer l'intervention du second m6canisme: la photorestauration indirecte. La baisse de la fr~quence de mutations qui s'observe dans la s6rie fractionn6e est en effet associ6e, dans ce cas, ~ une diminution de la dimension absolue des clones 16taux.
Mutation Res., 4 (1967) 491-5 °1

ACTION U V SUR LES OVOGONIES DE DROSOPHILES. II. 499
Cette double difffirence entre les 2 sdries d'expfiriences r6v~le, sans doute, l ' interven- tion d 'un processus de gudrison. La diminution absolue des clones 16taux correspond
une prolongation du temps d'arr~t de la premiere mitose qui suit l 'irradiation. Or, cette prolongation entralne probablement un allongement parall~le de la p6riode sen- sible pendant laquelle les pr6mutations ont plus de chances de gu6rir. Notons que le temps qui s@are la distribution des 2 fractions de la dose joue un r61e tr~s impor- tant et tr~s complexe dans ce processus de gu~rison 23.
La m~me corr6lation entre la dimension absolue des clones 16taux et le pourcen- tage de mutations s'observe lorsque l'on compare les r~sultats des irradiations ~ iooo et ~ 2oo0 ergs/mmL dans le cas oh la dose est appliqu6e dans une seule direction, sans fractionnement. La moyenne de la dimension absolue des clones passe, en effet, de 7.3 a 4.4 en m~me temps que le pourcentage de mutations tombe de 8.6 /~ 3.6. La photorestauration indirecte est donc, sans doute, un des facteurs responsable de la non lin6arit6 de la relation dose-effet; son action s'ajoute ~ celle qui d6coule de l'in6galit6 dans les doses revues par les diverses couches cellulaires.
On peut remarquer, au surplus, que l'~clairement dans 2 directions oppos6es, bien qu'il ait tendance ~ redresser la courbe, est encore loin de la rendre lindaire. I1 ne conduit, en effet, qu'~t une fr~quence de l~taux r~cessifs ~gale ~ 6.1% au lieu des 15.2% qui seraient attendus, si l 'on multipliait par 4 le r~sultat obtenu pour 500 ergs/mm 2. La photorestauration indirecte peut fitre responsable de cet 6cart, puisque les clones 16taux sont plus petits dans le cas oh les cellules re~oivent 2000 ergs/mm ~ dans 2 directions oppos6es qu'ils ne le sont aux faibles doses. Ils sont retiree, en fait, pour une raison qui n 'est pas bien comprise, plus petits dans ce cas que lorsque 2o00 ergs/ mm 2 sont donn6s dans une seule direction. La corr6lation fr6quence de mutations r6cupdrfes et dimension des clones est donc absolument g6n6rale.
Les exp6riences r~alis6es tendent donc/~ montrer qu'une fraction au moins des l~taux r6cessifs induits par les radiations chez la Drosophile, passe par un stade instable prfmutationnel dont la stabilisation est sous l'influence des variations du mftabolisme cellulaire.
Cette conclusion rejoint celle de SOBELS et coll. 26-al. Ces auteurs observent, chez le mfime organisme, que des pr6- et post-traitements divers modifient la frd- quence de mutations 16tales rdcessives induites par les rayons X. Ils ont montr6, en outre, en 6tudiant les diffdrents stades de la gam6tog6n~se ~" et par des exp6riences de fractionnement ~ que les spermatogonies ont une possibilit6 de restauration tr&s inf6- rieure ~ celle observ6e chez les spermatocytes. Effeetivement, en travaillant sur les ovogonies, nous n'avons observ6 qu'une faible diff6rence dans le pourcentage de muta- tions, non significative en fait, entre la s6rie ayant re~u une irradiation fractionn6e et la s6rie t6moin. De la m~me mani~re, le fair que BROWNING ET ALTENBORG n'obser- vent aucune modification dans la fr6quence des 16taux r6cessifs induits par les UV soit aprils fractionnement de la dose 1, soit apr~s trai tement au chloramphenicol 5 des cellules polaires, peut ~tre attribu6 ~ une tr&s faible aptitude ~ la restauration de ces cellules primordiales. Ainsi, il semble bien que selon les stades de division, le processus de restauration soit plus ou moins efficace 6.
La mutag6nfise par les UV est donc certainement un phdnom~ne complexe dont le mdcanisme n'est pas compl~tement 6clairci. Une difficult6 vient, sans doute, des diff6rences qui peuvent exister ~ ce point de rue entre les divers organismes et les diverses cellules. C'est ainsi qu'il n 'est probablement pas toujours 16gitime d 'extra-
Mutation Res., 4 (1967) 491-5 °1

500 JACQUELINE PROUST, ANNE FLAMAND
poler les r6sultats obtenus chez les micro-organismes o/a les connaissances dans ce do- maine sent plus avancdes, i~ la Drosophile oh la nature des photoproduits ~t l'origine des 16taux r6cessifs n'est pas connue. KADA et coll. ~'~ observent, en effet, chez E s c h e r i c h i a
cell , des faits oppos~s aux r6sultats rapportds iei. Recherchant l'apparition de proto- trophes b, la suite d'une irradiation UV, ils notent une augmentation de l'effet lors- qu'ils tractionnent la dose. Cette contradiction peut ~tre attribu6e, comme nous l'avons signal~ ci-dessus, ~ des possibili%s de restauration diff6rentes selon les organis- rues, mais aussi ~ la nature m~me des mutations ~tudi6es. I1 est 6vident que parmi la classe h6t6rog~ne des 16taux rdcessifs, des Idsions aussi diff6rentes que les mutations ponctuelles et les petites ddficiences peuvent ne pas exiger les m~mes proeessus de r@aration, ce qui complique beaucoup l'analyse des r6sultats.
REMERCIEMENTS
Nous exprimons notre reconnaissance au Professeur L'HERITIER pour sos suggestions et ses conseils utiles dans la r~alisation de ce travail.
Nous remercions Madame C. VIGIER et Mademoiselle M. C. I"ONTYNE pour leur prgcieuse assistance technique.
RESUME
Une m6me dose (2000 ergs/mm 2) est administr6e de 3 mani~res diffdrentes. (I) En une seule lois et dans un seul sens (t6moin). (2) Successivement, sans changement de position, dans e directions oppos6es. (3) Dans un seul sens, en 2 fractions ~gales sdpar~es par un intervalle de 2 h.
La comparaison entre les exp6riences I e t 2 montre que le double 6clairement conduit 5 augmenter le pourcentage de mutations, /t diminuer le pourcentage de reprise de l'implant et ~ baisser la f6condi% de l'ovaire implant6. Ces %sultats sent dus 5 une hd%rog6n6it6 de position des cellules irradides dans la population.
La comparaison entre les exp6riences i e t 3 montre clue le fractionnement entralne, it la fois, une baisse du pourcentage de mutations et une diminution de l'effectif absolu du clone 16tal. Ces faits s'expliquent par l'existence d 'un processus de gudrison, intervenant au niveau des cellules individuelles.
BI I:H.I ()(; I'~,\ Pt t 1 I'~
I AL'rENBU~RG, l~.., ET L. S. I~ROWNING, "l"hc mutagenic equivalence of cont inuous and intermit- tent ultraviolet in Drosophila melanogaster, Genetics, 47 (1962) 273-27t).
2 :\LTI.~NBURG, l.. ~., The effect of intensi ty on the mutagenic act ivi ty of ultraviolet light when administered in relatively low doses to Drosophila, Genetics, 41 (I956) 632 633.
3 I{OYCE, I{. P., I'T P. HOV~'ARD-FL,kNDERS, Release, of ultraviolet light induced thymine dimers from 1)NA in E. coli K I2, Proc. Natl. Acad. Sci. (U.S.), 51 (I964) 293 3oo.
4 BROWNING, L. S., ET E. ALTENBURG, The propor t ional i ty between muta t ion rate and ultra- violet dose after photoreact ivat ion in Drosophila, Genetics, 47 (I962) 36I-366"
5 |{RO\VNING, L. S., ET E. ALTF.NBUP.G, Failure to detect an ant i -mutagenie eiiect of chloram- phenicol in ultraviolet treated polar cap cells (early germ track) of Drosophila, Genetics, 48 (I9~,3) 525 52s.
6 I)AWI.:% D. R., Repair mechanisms and variat ions in [IV sensitivity within the cell cycle, Mu- tatifm Res., 2 (1965) 477 48()-
7 I)OU~N~-:Y, C. ()., Ultraviolet light induced muta t ion as an event in the physiology of the bacterial cell, in F. H. SOBFA~S (P2d.), tfepairfrom Genetic leadiatio~ Damage, Pergamon, London,
1963, p- J25 ~49.
Mutation Res., 4 (1967) 491-5 ° I

ACTION U V SUR LES OVOGONIES DE DROSOPHILES. I I . 5o i
DOUDNEY, C. O., Ult rav io le t l ight effects on deoxyr ibonuc le ic acid repl icat ion, Cellular Radia- tion Biology, Wil l i ams and Wilkins , Bal t imore , Md., 1965, p. I2O-I4I .
9 HOWARD-FLANDERS, P., E. SIMSON ET L. THERIOT, The excision of t h y m i n e d imers f rom D N A f i lament fo rma t ion and sens i t iv i ty to UV l ight in Escherichia coli I,ZI2, Mutation Res., i (1964) 219-226.
io JAGCER, J., P h o t o c h e m i s t r y of nucleic acids and its biological impl ica t ions , in E. J. BOWEN (Ed.), Recent Progress in Photobiology, Blaekwell , Oxford, I965, p. 59 8o.
11 JAOGER, J. , ET R. S. STAFFORD, Evidence for two m e c h a n i s m s of pho to reac t i va t i on in Escheri- chia coli B, Biophys. J., 5 (1965) 75 -88.
i2 JAGGER, J., W. C. WISE ET R. S. STAFFORD, Delay in g rowth and divis ion induced by near u l t rav io le t r ad ia t ion in Escherichia coli B and its role in pho top ro tec t ion and l iquid ho ld ing recovery, Photochem. Photobiol., 3 (1964) 11-24.
13 I'~ADA, T., C. O. DOUDNEY ET F. L. HAAS, Muta t ion f requency response of i r rad ia ted bac te r ia to a second rad ia t ion exposure , Mutation Res., 3 (1966) 118-128.
14 I'~aDA, T., ET H. MARCOVlCH, Sur le si6ge initial de Fact ion m u t a g ~ n e des r ayons X et des u l t ra- violets chez E. coli KI2, Ann. Inst. Pasteur, lO 5 (I963) 989 Ioo6.
15 t'~IMBALL, R. F., The rela t ion of repair to differer~tial r ad iosens i t iv i ty in t he p roduc t ion of m u t a t i o n in Paramecium, in F. H. SOBELS (Ed.), Repair from Genetic Radiation Damage, Per- gamon , London , 1963, p. I67- I78 .
16 KIMBAI~L, R. F., N. GAITHER ET S. W. PERDUE, Metabolic repair of p ren lu t a t iona l d a m a g e in Paramecium, Intern. J. Radiation Biol., 3 (1961) I33- I47 .
17 LIEB, M., Deoxyr ibonucle ic acid syn thes i s and u l t rav io le t - induced m u t a t i o n , Biochim. Bio- phys. Acta, 37 (196o) I55- I57 .
18 MULLER, H. J., L. S. ALTENBURG, H. U. MEYER, M. EDMONDSON ET E. ALTENBURG, The lack of p ropor t iona l i ty be tween m u t a t i o n ra te and u l t rav io le t dose in Drosophila, Heredity, 8 (~954) 158-185 .
19 OFTEDAL, P., Radiosens i t iv i ty of Drosophila spermatogonia, I I I . Compar i son of acu te and p ro t r ac t ed i r radia t ion efflciencies in re la t ion to cell killing, Mutation Res., i (1964) 63-76.
20 PROUST, J., Etude de Faction gdndtique de la lumikre UV sur les ovogonies de Drosophiles, Th~se de Doc to ra t d ' E t a t , 1964, Facul t~ des Sciences d 'Orsay , Univers i t6 de Paris, France.
21 PROUST, J. , Action m u t a g b n e des UV sur ins ovogonies de Drosophile, I. Cin6t iques de la m u t a - g6n~se, Mutation Res., 4 (1967) 473-489 .
22 RUPERT, C. S., S. H. GOODGAL ET R. M. HEP, RIOTT, Pho to reac t i va t i on in vitro of u l t rav io le t i nac t iva t ed Hemophihts influenzae t r a n s f o r m i n g factor, J. Ge~. Physiol., 41 (1958) 451-471.
23 RUSSEL, V~ ~. L., The effect of r ad ia t ion dose ra te and f rac t iona t ion on m u t a t i o n in mice, in F. H. SOBELS (Ed.), Repair from Get~etie Radiation Damage, Pergamon , London , 1963, p. 205 217 •
2 4 SETLO~V, if. B., Phys ica l changes and n lu tagenes is , J. Cellular Comp. Physiol., 64, Suppl. i (1964) 51-68.
25 SETLOW, R. B., ET W. L. (;aRRIE~, The d i sappea rance of t h y m i n e d imers f rom DNA ; an error- cor rec t ing mec han i sm , Proc. Natl. Acad. Sci. (U.S.), 51 (1964) 226-231.
26 SOBELS, F. H., Modificat ion of p r e -mu t a t i ona l rad ia t ion danaage in Drosophila, Proc. Syrup. Radiation Effects and 3/Iilieu. Fritz Niggli, Strahle:~therapie, Suppl. , 51 (1962) 197 212.
27 SOBELS, F. H., Repa i r and differential rad iosens i t iv i ty in deve loping ge rm cells of Drosophila males, in F. H. SOBELS (Ed.), Repair from Genetic Radiation Damage, Pe rgamon , London , 1963, p. 179o-2o4 .
28 SOBELS, F. H., Peroxides and the induc t ion of m u t a t i o n s by X-rays , u l t rav io le t and formalde- hyde , Radiation Res., Suppl. 3 (1963) 171-183.
29 SOBELS, F. H., Rad iosens i t i v i ty and repair in different ge rm cell s tages of Drosophila, Proc. i I th Int. Contr. Genetics, The Hague, P e rgamon , Oxford, 1963, p. 235-255.
3 ° SOBELS, F. H., Pos t - r ad ia t ion reduc t ion of genet ic dan lage in m a t u r e Drosophila sperm by ni t rogen, Mutation Res., I (I964) 472-477 .
31 SOBELS, F. H., E T A . D. TATES, Recovery f rom p r e m u t a t i o n a l d a m a g e of X- i r rad ia t ion in Drosophila spermatogenes i s , J. Cellular Comp. Physiol., Suppl. I, 58 (I96I) 189-196.
32 WITKIN, E. M., ' D a r k repai r ' of m u t a t i o n s induced in Escherichia coli by u l t rav io le t l ight , in F. H. SOBELS (Ed.), Rebair from Genetic Radiation Damage, Pergamon , London , 1963, p. 151-165 .
33 WITKIN, E. M., Photo reve r sa l and ' da rk repai r ' of m u t a t i o n s to p r o t o t r o p h y induced by u l t rav io le t l ight in pho to reac t ivab le and nonpho to reac t i vab l e s t ra ins of Escherichia coli, Mu- tation Res., i (1964) 22-36.
34 ~vVITKIN, E. M., N. A. SICURELLA ET G. M. BENNETT, Photorevers ib i l i ty of induced m u t a t i o n s in a non pho to reac t ivab le s t ra in of Escherichia coli, Proc. Natl. Acad. Sci. (U.S.), 5 ° (1963) lO55-1o58.
Mutation Res., 4 (1967) 491-5°1











![SIMULATION DE PROCÉDÉS ET ACV … · SIMULATION DE PROCÉDÉS ET ACV ENVIRONNEMENTALE Eco-conception d’un procédé de fractionnement de micro-algues Rémi Julio Congrès [avniR]](https://img.pdfslide.fr/doc/110x75/5b9c84bb09d3f2194e8c6d24/simulation-de-procedes-et-acv-simulation-de-procedes-et-acv-environnementale.jpg)

















