Embed Size (px)
Citation preview

Activation of the Hippo pathway by CTLA-4 regulatesthe expression of Blimp-1 in the CD8+ T cellJames E. D. Thaventhirana,1, Anja Hoffmannb,1, Lukasz Magierab,c, Maike de la Roched, Holger Lingele,Monika Brunner-Weinzierle, and Douglas T. Fearonb,c,2
aDepartment of Immunology, Division of Infection and Immunity, University College London, Royal Free Hospital, London NW3 2QG, United Kingdom;bDepartment of Medicine, University of Cambridge School of Clinical Medicine, Cambridge CB2 2QH, United Kingdom; cCancer Research UK CambridgeResearch Institute, Li Ka Shing Centre, Cambridge CB2 0RE, United Kingdom; dCambridge Institute for Medical Research, University of Cambridge, CambridgeCB2 0XY, United Kingdom; and eDepartment of Experimental Pediatrics, University Hospital, Otto-von-Guericke University Magdeburg, 39120 Magdeburg,Germany
Contributed by Douglas T. Fearon, June 1, 2012 (sent for review April 27, 2012)
During the primary response, the commitment of the CD8+ T cell toBlimp-1 expression and the terminal differentiation that Blimp-1induces must be timed so as not to impair the process of clonalexpansion. We determined whether the Hippo pathway, whichlinks cell–cell contact to differentiation in other cell lineages, con-trols Blimp-1 expression. Activating the CD8+ T cell with antigenand IL-2 causes expression of the core Hippo pathway compo-nents, including the pivotal transcriptional cofactor Yap. Contactbetween activated CD8+ T cells induces Hippo pathway-mediatedYap degradation and Blimp-1 expression; a Hippo-resistant, stableform of Yap suppresses Blimp-1 expression. Cytotoxic T lympho-cyte antigen 4 (CTLA-4) and CD80 comprise the receptor–ligandpair that mediates contact-dependent Hippo pathway activation.In vivo, CD8+ T cells expressing Hippo resistant-Yap or lackingCTLA-4 have diminished expression of the senescence marker,KLRG1, during a viral infection. The CTLA-4/Hippo pathway/Blimp-1 system may couple terminal differentiation of CD8+ T cellwith the magnitude of clonal expansion.
During the first week of a viral infection, virus-specific CD8+
T cells must coordinate two interdependent but mutuallyexclusive processes: rapid replication to generate the precursorsof effector cells and development of effector cells from theseprecursors. These two cellular responses must be appropriatelytimed, because full effector differentiation, so-called terminaldifferentiation, is associated with the diminution or loss of rep-licative function. If terminal differentiation occurs too earlyduring the process of clonal expansion, then paradoxically, therate at which effector cells would be generated would be in-sufficient for host defense. The ability of the CD8+ T cell tosuspend terminal differentiation until clonal expansion has oc-curred is documented by the observation that CD62Llo, CD127lo
CD8+ T cells specific for Listeria monocytogenes did not appearuntil day 4 postinfection (1). Conversely, inappropriate delay ofterminal differentiation would also impair control of an infection.Recent studies that have focused on identifying the determi-
nants of terminal differentiation of the CD8+ T cell have ad-vanced our understanding of this developmental step. Thetranscriptional repressor, Blimp-1, and the T-box transcriptionfactors, T-bet (2) and Eomesodermin (Eomes) (3), have beenshown to regulate the differentiation of effector CD8+ T cells.Absence of Blimp-1 is associated with virus-specific CD8+ T cellshaving impaired cytolytic function, lower expression of the se-nescence-associated marker killer cell lectin-like receptor G1(KLRG1), and enhanced development of the central memoryphenotype (4, 5). The acquisition of effector functions [butmaintenance of replicative function in both CD4+ (6) and CD8+
(7) T cells] is consistent with the capacity of Blimp-1–deficientCD8+ T cells to express IFN-γ and cytolytic activities, althoughat suboptimal levels. These loss-of-function studies (4, 5) in-dicate that Blimp-1 is required for terminal differentiation by theactivated CD8+ T cell, which it for two other lymphocytic line-ages, the B (8) and NK (9) cells. T-bet promotes Blimp-1 ex-
pression in the natural killer (NK) cell (9), and a similar role maybe expected in the CD8+ cell, because T-bet– (2) and Blimp-1–deficient (4, 5) CD8+ T cells have similar differentiation phe-notypes. Moreover, in vitro studies have shown that T-bet ex-pression enables signaling by the IL-2 receptor to induce Blimp-1transcription (10). Although T-bet levels are always higher interminally differentiated, KLRG1+ cells than at other stages ofdifferentiation, it may be that the ratio of T-bet and Eomes isalso critical (11). Eomes shares with T-bet a capacity to mediatethe development of effector function (12), but it differs from T-bet in being necessary for the development of memory CD8+ Tcells having secondary proliferative capability (3) in response tothe mammalian homolog of the target of rapamycin (mTOR)(13) or T-cell factor (TCF-1) (14) signaling.The extracellular signals that regulate the expression of these
transcription factors include antigen and the cytokines IL-2, IL-12, IL-21, and IL-27 (15–19). The inflammatory cytokines, IL-12 and IL-27, are produced by dendritic cells and cells of theinnate immune system that have been stimulated by microbialproducts, whereas IL-2 and IL-21 are secreted by antigen-stimulated CD4+ and CD8+ T cells. Although both types ofcytokines may increase during the course of an infection andtherefore, provide temporal elements by which the CD8+ T cellcould time its commitment to terminal differentiation, thesesignals also would be coupled to the extent of microbial repli-cation, which may not reflect the magnitude of clonal expan-sion. IL-2, acting in a paracrine manner, could meet the re-quirement of being directly correlated with the number ofantigen-stimulated T cells, but this cytokine has dual functionsin promoting the development of both terminally differentiatedeffector cells and replication-competent memory cells (20–24).Thus, neither inflammatory- nor T-cell–derived cytokines arelikely to be the ultimate determinants of Blimp-1 expression andterminal differentiation.In multiple cell types of invertebrates and vertebrates, termi-
nal differentiation is directed by a highly conserved develop-mental system, termed the Hippo pathway. The Hippo pathwayis triggered when replicating cells come into contact with eachother, enabling a receptor–ligand pair to activate a serine/threoninekinase cascade (25). This activation results in the phosphoryla-tion and proteosomal degradation of the transcriptional cofac-
Author contributions: J.E.D.T. and D.T.F. designed research; J.E.D.T., A.H., L.M., M.d.l.R.,and H.L. performed research; M.B.-W. contributed new reagents/analytic tools; J.E.D.T.,A.H., L.M., M.d.l.R., H.L., and D.T.F. analyzed data; and J.E.D.T. and D.T.F. wrote the paper.
The authors declare no conflict of interest.
Freely available online through the PNAS open access option.1J.E.D.T. and A.H. contributed equally to this work.2To whom correspondence should be addressed. E-mail: [email protected].
See Author Summary on page 13152 (volume 109, number 33).
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1209115109/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1209115109 PNAS | Published online June 27, 2012 | E2223–E2229
IMMUNOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Oct
ober
20,
202
1
tor, Yap, which permits differentiation to proceed (26). A po-tential role for the Hippo pathway in regulating the expressionof Blimp-1 and Eomes is suggested by the independent obser-vations that Blimp-1 and the Hippo pathway, respectively, arerequired for the normal differentiation of skin (27, 28) and thatYap, by interacting with the transcription factor TEA domain(TEAD), induces Eomes expression in trophoectoderm (29, 30).Because cell–cell contact triggers the Hippo pathway, the like-lihood of which would be proportional to the relative frequencyof the interacting cells, this pathway would fulfill the hypotheticalrequirement of linking the magnitude of clonal expansion byactivated CD8+ T cells to their commitment to terminal differ-entiation. Accordingly, we have examined the role of the Hippopathway in regulating Blimp-1 expression by the CD8+ T cell.
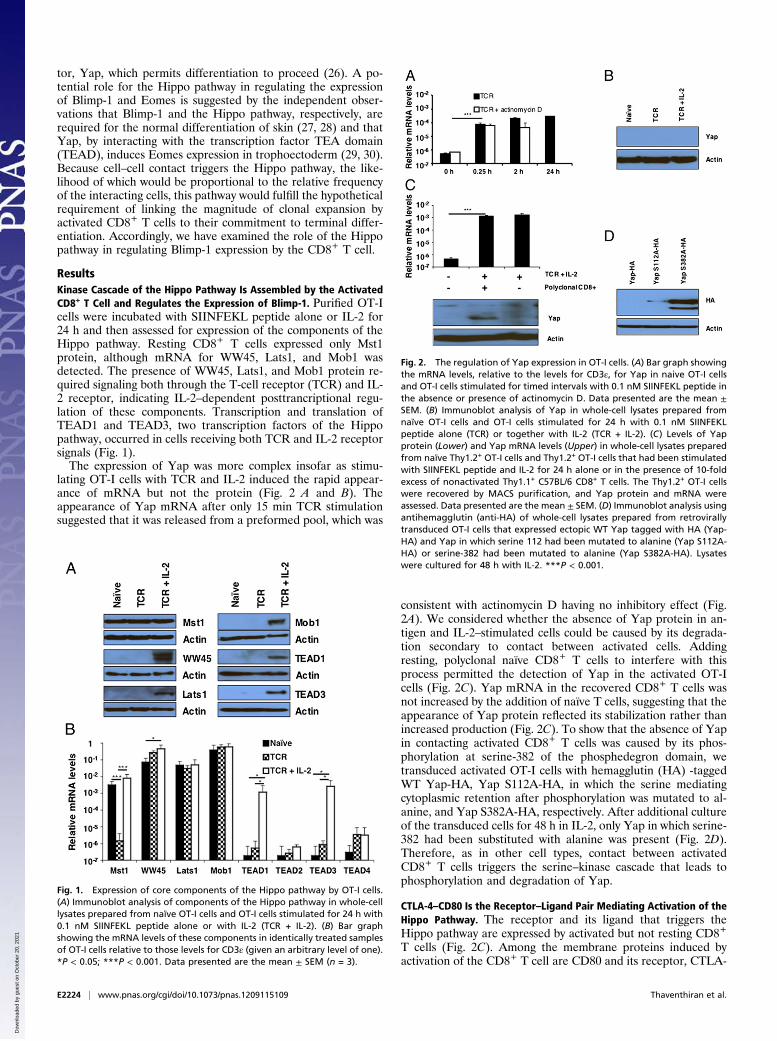
ResultsKinase Cascade of the Hippo Pathway Is Assembled by the ActivatedCD8+ T Cell and Regulates the Expression of Blimp-1. Purified OT-Icells were incubated with SIINFEKL peptide alone or IL-2 for24 h and then assessed for expression of the components of theHippo pathway. Resting CD8+ T cells expressed only Mst1protein, although mRNA for WW45, Lats1, and Mob1 wasdetected. The presence of WW45, Lats1, and Mob1 protein re-quired signaling both through the T-cell receptor (TCR) and IL-2 receptor, indicating IL-2–dependent posttrancriptional regu-lation of these components. Transcription and translation ofTEAD1 and TEAD3, two transcription factors of the Hippopathway, occurred in cells receiving both TCR and IL-2 receptorsignals (Fig. 1).The expression of Yap was more complex insofar as stimu-
lating OT-I cells with TCR and IL-2 induced the rapid appear-ance of mRNA but not the protein (Fig. 2 A and B). Theappearance of Yap mRNA after only 15 min TCR stimulationsuggested that it was released from a preformed pool, which was
consistent with actinomycin D having no inhibitory effect (Fig.2A). We considered whether the absence of Yap protein in an-tigen and IL-2–stimulated cells could be caused by its degrada-tion secondary to contact between activated cells. Addingresting, polyclonal naïve CD8+ T cells to interfere with thisprocess permitted the detection of Yap in the activated OT-Icells (Fig. 2C). Yap mRNA in the recovered CD8+ T cells wasnot increased by the addition of naïve T cells, suggesting that theappearance of Yap protein reflected its stabilization rather thanincreased production (Fig. 2C). To show that the absence of Yapin contacting activated CD8+ T cells was caused by its phos-phorylation at serine-382 of the phosphedegron domain, wetransduced activated OT-I cells with hemagglutin (HA) -taggedWT Yap-HA, Yap S112A-HA, in which the serine mediatingcytoplasmic retention after phosphorylation was mutated to al-anine, and Yap S382A-HA, respectively. After additional cultureof the transduced cells for 48 h in IL-2, only Yap in which serine-382 had been substituted with alanine was present (Fig. 2D).Therefore, as in other cell types, contact between activatedCD8+ T cells triggers the serine–kinase cascade that leads tophosphorylation and degradation of Yap.
CTLA-4–CD80 Is the Receptor–Ligand Pair Mediating Activation of theHippo Pathway. The receptor and its ligand that triggers theHippo pathway are expressed by activated but not resting CD8+
T cells (Fig. 2C). Among the membrane proteins induced byactivation of the CD8+ T cell are CD80 and its receptor, CTLA-
Fig. 1. Expression of core components of the Hippo pathway by OT-I cells.(A) Immunoblot analysis of components of the Hippo pathway in whole-celllysates prepared from naïve OT-I cells and OT-I cells stimulated for 24 h with0.1 nM SIINFEKL peptide alone or with IL-2 (TCR + IL-2). (B) Bar graphshowing the mRNA levels of these components in identically treated samplesof OT-I cells relative to those levels for CD3ε (given an arbitrary level of one).*P < 0.05; ***P < 0.001. Data presented are the mean ± SEM (n = 3).
Fig. 2. The regulation of Yap expression in OT-I cells. (A) Bar graph showingthe mRNA levels, relative to the levels for CD3ε, for Yap in naive OT-I cellsand OT-I cells stimulated for timed intervals with 0.1 nM SIINFEKL peptide inthe absence or presence of actinomycin D. Data presented are the mean ±SEM. (B) Immunoblot analysis of Yap in whole-cell lysates prepared fromnaïve OT-I cells and OT-I cells stimulated for 24 h with 0.1 nM SIINFEKLpeptide alone (TCR) or together with IL-2 (TCR + IL-2). (C) Levels of Yapprotein (Lower) and Yap mRNA levels (Upper) in whole-cell lysates preparedfrom naïve Thy1.2+ OT-I cells and Thy1.2+ OT-I cells that had been stimulatedwith SIINFEKL peptide and IL-2 for 24 h alone or in the presence of 10-foldexcess of nonactivated Thy1.1+ C57BL/6 CD8+ T cells. The Thy1.2+ OT-I cellswere recovered by MACS purification, and Yap protein and mRNA wereassessed. Data presented are the mean ± SEM. (D) Immunoblot analysis usingantihemagglutin (anti-HA) of whole-cell lysates prepared from retrovirallytransduced OT-I cells that expressed ectopic WT Yap tagged with HA (Yap-HA) and Yap in which serine 112 had been mutated to alanine (Yap S112A-HA) or serine-382 had been mutated to alanine (Yap S382A-HA). Lysateswere cultured for 48 h with IL-2. ***P < 0.001.
E2224 | www.pnas.org/cgi/doi/10.1073/pnas.1209115109 Thaventhiran et al.
Dow
nloa
ded
by g
uest
on
Oct
ober
20,
202
1
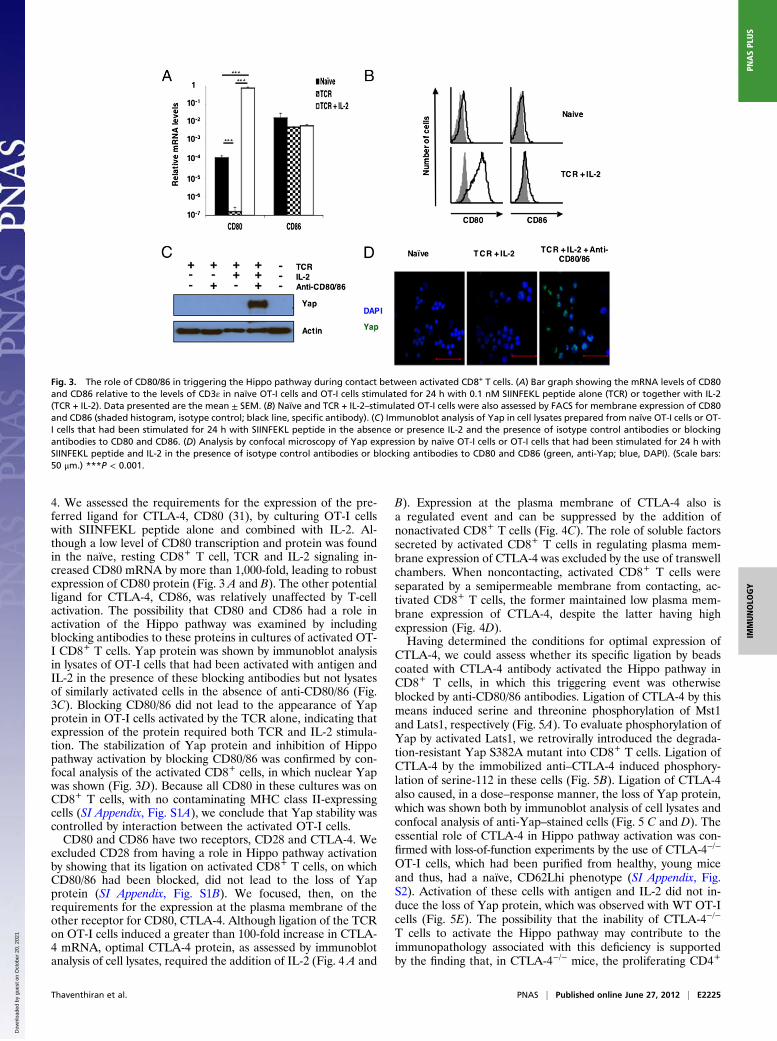
4. We assessed the requirements for the expression of the pre-ferred ligand for CTLA-4, CD80 (31), by culturing OT-I cellswith SIINFEKL peptide alone and combined with IL-2. Al-though a low level of CD80 transcription and protein was foundin the naïve, resting CD8+ T cell, TCR and IL-2 signaling in-creased CD80 mRNA by more than 1,000-fold, leading to robustexpression of CD80 protein (Fig. 3 A and B). The other potentialligand for CTLA-4, CD86, was relatively unaffected by T-cellactivation. The possibility that CD80 and CD86 had a role inactivation of the Hippo pathway was examined by includingblocking antibodies to these proteins in cultures of activated OT-I CD8+ T cells. Yap protein was shown by immunoblot analysisin lysates of OT-I cells that had been activated with antigen andIL-2 in the presence of these blocking antibodies but not lysatesof similarly activated cells in the absence of anti-CD80/86 (Fig.3C). Blocking CD80/86 did not lead to the appearance of Yapprotein in OT-I cells activated by the TCR alone, indicating thatexpression of the protein required both TCR and IL-2 stimula-tion. The stabilization of Yap protein and inhibition of Hippopathway activation by blocking CD80/86 was confirmed by con-focal analysis of the activated CD8+ cells, in which nuclear Yapwas shown (Fig. 3D). Because all CD80 in these cultures was onCD8+ T cells, with no contaminating MHC class II-expressingcells (SI Appendix, Fig. S1A), we conclude that Yap stability wascontrolled by interaction between the activated OT-I cells.CD80 and CD86 have two receptors, CD28 and CTLA-4. We
excluded CD28 from having a role in Hippo pathway activationby showing that its ligation on activated CD8+ T cells, on whichCD80/86 had been blocked, did not lead to the loss of Yapprotein (SI Appendix, Fig. S1B). We focused, then, on therequirements for the expression at the plasma membrane of theother receptor for CD80, CTLA-4. Although ligation of the TCRon OT-I cells induced a greater than 100-fold increase in CTLA-4 mRNA, optimal CTLA-4 protein, as assessed by immunoblotanalysis of cell lysates, required the addition of IL-2 (Fig. 4 A and
B). Expression at the plasma membrane of CTLA-4 also isa regulated event and can be suppressed by the addition ofnonactivated CD8+ T cells (Fig. 4C). The role of soluble factorssecreted by activated CD8+ T cells in regulating plasma mem-brane expression of CTLA-4 was excluded by the use of transwellchambers. When noncontacting, activated CD8+ T cells wereseparated by a semipermeable membrane from contacting, ac-tivated CD8+ T cells, the former maintained low plasma mem-brane expression of CTLA-4, despite the latter having highexpression (Fig. 4D).Having determined the conditions for optimal expression of
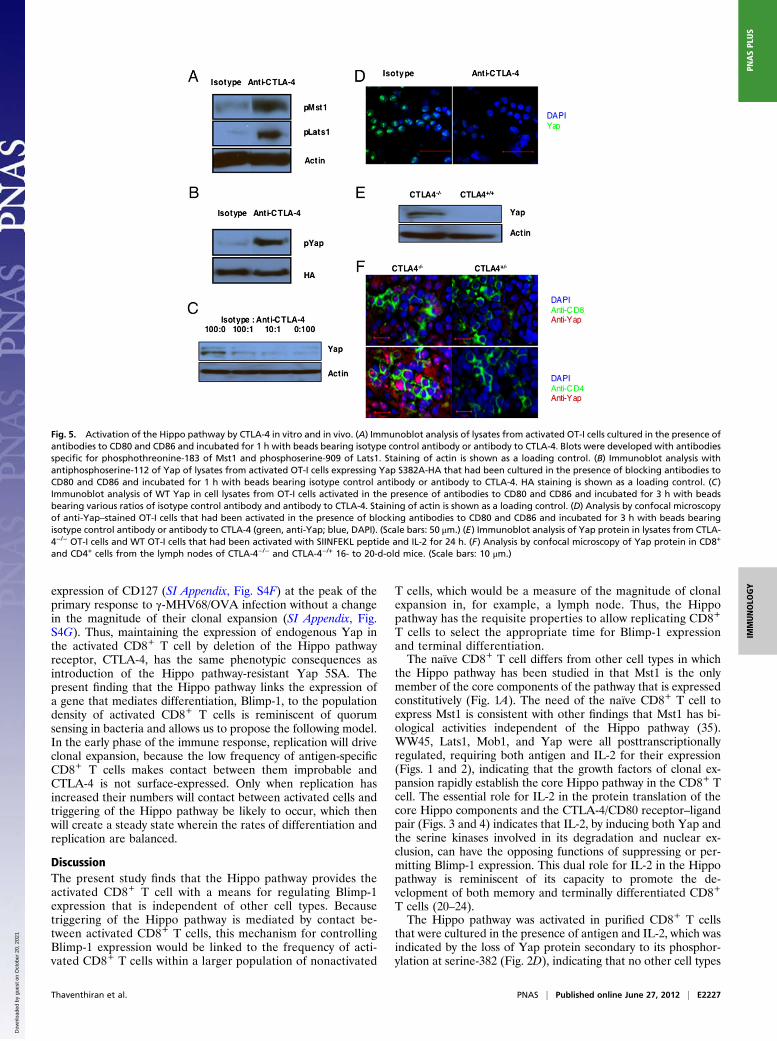
CTLA-4, we could assess whether its specific ligation by beadscoated with CTLA-4 antibody activated the Hippo pathway inCD8+ T cells, in which this triggering event was otherwiseblocked by anti-CD80/86 antibodies. Ligation of CTLA-4 by thismeans induced serine and threonine phosphorylation of Mst1and Lats1, respectively (Fig. 5A). To evaluate phosphorylation ofYap by activated Lats1, we retrovirally introduced the degrada-tion-resistant Yap S382A mutant into CD8+ T cells. Ligation ofCTLA-4 by the immobilized anti–CTLA-4 induced phosphory-lation of serine-112 in these cells (Fig. 5B). Ligation of CTLA-4also caused, in a dose–response manner, the loss of Yap protein,which was shown both by immunoblot analysis of cell lysates andconfocal analysis of anti-Yap–stained cells (Fig. 5 C and D). Theessential role of CTLA-4 in Hippo pathway activation was con-firmed with loss-of-function experiments by the use of CTLA-4−/−
OT-I cells, which had been purified from healthy, young miceand thus, had a naïve, CD62Lhi phenotype (SI Appendix, Fig.S2). Activation of these cells with antigen and IL-2 did not in-duce the loss of Yap protein, which was observed with WT OT-Icells (Fig. 5E). The possibility that the inability of CTLA-4−/−
T cells to activate the Hippo pathway may contribute to theimmunopathology associated with this deficiency is supportedby the finding that, in CTLA-4−/− mice, the proliferating CD4+
Fig. 3. The role of CD80/86 in triggering the Hippo pathway during contact between activated CD8+ T cells. (A) Bar graph showing the mRNA levels of CD80and CD86 relative to the levels of CD3ε in naïve OT-I cells and OT-I cells stimulated for 24 h with 0.1 nM SIINFEKL peptide alone (TCR) or together with IL-2(TCR + IL-2). Data presented are the mean ± SEM. (B) Naïve and TCR + IL-2–stimulated OT-I cells were also assessed by FACS for membrane expression of CD80and CD86 (shaded histogram, isotype control; black line, specific antibody). (C) Immunoblot analysis of Yap in cell lysates prepared from naïve OT-I cells or OT-I cells that had been stimulated for 24 h with SIINFEKL peptide in the absence or presence IL-2 and the presence of isotype control antibodies or blockingantibodies to CD80 and CD86. (D) Analysis by confocal microscopy of Yap expression by naïve OT-I cells or OT-I cells that had been stimulated for 24 h withSIINFEKL peptide and IL-2 in the presence of isotype control antibodies or blocking antibodies to CD80 and CD86 (green, anti-Yap; blue, DAPI). (Scale bars:50 μm.) ***P < 0.001.
Thaventhiran et al. PNAS | Published online June 27, 2012 | E2225
IMMUNOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Oct
ober
20,
202
1
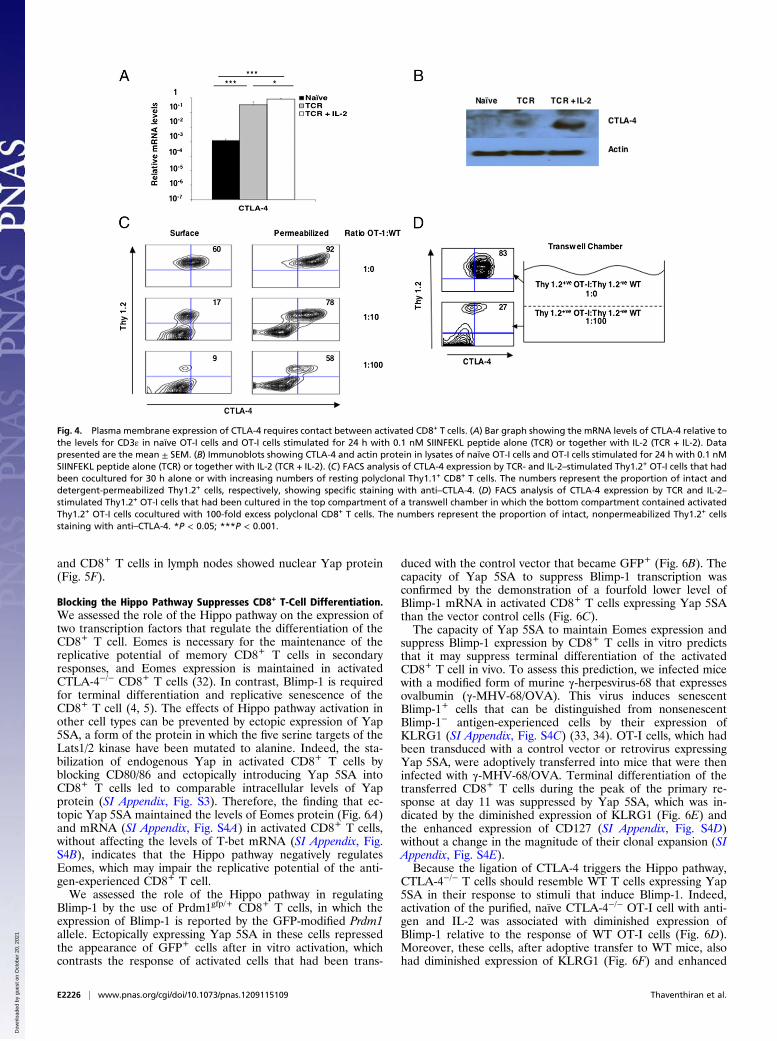
and CD8+ T cells in lymph nodes showed nuclear Yap protein(Fig. 5F).
Blocking the Hippo Pathway Suppresses CD8+ T-Cell Differentiation.We assessed the role of the Hippo pathway on the expression oftwo transcription factors that regulate the differentiation of theCD8+ T cell. Eomes is necessary for the maintenance of thereplicative potential of memory CD8+ T cells in secondaryresponses, and Eomes expression is maintained in activatedCTLA-4−/− CD8+ T cells (32). In contrast, Blimp-1 is requiredfor terminal differentiation and replicative senescence of theCD8+ T cell (4, 5). The effects of Hippo pathway activation inother cell types can be prevented by ectopic expression of Yap5SA, a form of the protein in which the five serine targets of theLats1/2 kinase have been mutated to alanine. Indeed, the sta-bilization of endogenous Yap in activated CD8+ T cells byblocking CD80/86 and ectopically introducing Yap 5SA intoCD8+ T cells led to comparable intracellular levels of Yapprotein (SI Appendix, Fig. S3). Therefore, the finding that ec-topic Yap 5SA maintained the levels of Eomes protein (Fig. 6A)and mRNA (SI Appendix, Fig. S4A) in activated CD8+ T cells,without affecting the levels of T-bet mRNA (SI Appendix, Fig.S4B), indicates that the Hippo pathway negatively regulatesEomes, which may impair the replicative potential of the anti-gen-experienced CD8+ T cell.We assessed the role of the Hippo pathway in regulating
Blimp-1 by the use of Prdm1gfp/+ CD8+ T cells, in which theexpression of Blimp-1 is reported by the GFP-modified Prdm1allele. Ectopically expressing Yap 5SA in these cells repressedthe appearance of GFP+ cells after in vitro activation, whichcontrasts the response of activated cells that had been trans-
duced with the control vector that became GFP+ (Fig. 6B). Thecapacity of Yap 5SA to suppress Blimp-1 transcription wasconfirmed by the demonstration of a fourfold lower level ofBlimp-1 mRNA in activated CD8+ T cells expressing Yap 5SAthan the vector control cells (Fig. 6C).The capacity of Yap 5SA to maintain Eomes expression and
suppress Blimp-1 expression by CD8+ T cells in vitro predictsthat it may suppress terminal differentiation of the activatedCD8+ T cell in vivo. To assess this prediction, we infected micewith a modified form of murine γ-herpesvirus-68 that expressesovalbumin (γ-MHV-68/OVA). This virus induces senescentBlimp-1+ cells that can be distinguished from nonsenescentBlimp-1− antigen-experienced cells by their expression ofKLRG1 (SI Appendix, Fig. S4C) (33, 34). OT-I cells, which hadbeen transduced with a control vector or retrovirus expressingYap 5SA, were adoptively transferred into mice that were theninfected with γ-MHV-68/OVA. Terminal differentiation of thetransferred CD8+ T cells during the peak of the primary re-sponse at day 11 was suppressed by Yap 5SA, which was in-dicated by the diminished expression of KLRG1 (Fig. 6E) andthe enhanced expression of CD127 (SI Appendix, Fig. S4D)without a change in the magnitude of their clonal expansion (SIAppendix, Fig. S4E).Because the ligation of CTLA-4 triggers the Hippo pathway,
CTLA-4−/− T cells should resemble WT T cells expressing Yap5SA in their response to stimuli that induce Blimp-1. Indeed,activation of the purified, naïve CTLA-4−/− OT-I cell with anti-gen and IL-2 was associated with diminished expression ofBlimp-1 relative to the response of WT OT-I cells (Fig. 6D).Moreover, these cells, after adoptive transfer to WT mice, alsohad diminished expression of KLRG1 (Fig. 6F) and enhanced
Fig. 4. Plasma membrane expression of CTLA-4 requires contact between activated CD8+ T cells. (A) Bar graph showing the mRNA levels of CTLA-4 relative tothe levels for CD3ε in naïve OT-I cells and OT-I cells stimulated for 24 h with 0.1 nM SIINFEKL peptide alone (TCR) or together with IL-2 (TCR + IL-2). Datapresented are the mean ± SEM. (B) Immunoblots showing CTLA-4 and actin protein in lysates of naïve OT-I cells and OT-I cells stimulated for 24 h with 0.1 nMSIINFEKL peptide alone (TCR) or together with IL-2 (TCR + IL-2). (C) FACS analysis of CTLA-4 expression by TCR- and IL-2–stimulated Thy1.2+ OT-I cells that hadbeen cocultured for 30 h alone or with increasing numbers of resting polyclonal Thy1.1+ CD8+ T cells. The numbers represent the proportion of intact anddetergent-permeabilized Thy1.2+ cells, respectively, showing specific staining with anti–CTLA-4. (D) FACS analysis of CTLA-4 expression by TCR and IL-2–stimulated Thy1.2+ OT-I cells that had been cultured in the top compartment of a transwell chamber in which the bottom compartment contained activatedThy1.2+ OT-I cells cocultured with 100-fold excess polyclonal CD8+ T cells. The numbers represent the proportion of intact, nonpermeabilized Thy1.2+ cellsstaining with anti–CTLA-4. *P < 0.05; ***P < 0.001.
E2226 | www.pnas.org/cgi/doi/10.1073/pnas.1209115109 Thaventhiran et al.
Dow
nloa
ded
by g
uest
on
Oct
ober
20,
202
1
expression of CD127 (SI Appendix, Fig. S4F) at the peak of theprimary response to γ-MHV68/OVA infection without a changein the magnitude of their clonal expansion (SI Appendix, Fig.S4G). Thus, maintaining the expression of endogenous Yap inthe activated CD8+ T cell by deletion of the Hippo pathwayreceptor, CTLA-4, has the same phenotypic consequences asintroduction of the Hippo pathway-resistant Yap 5SA. Thepresent finding that the Hippo pathway links the expression ofa gene that mediates differentiation, Blimp-1, to the populationdensity of activated CD8+ T cells is reminiscent of quorumsensing in bacteria and allows us to propose the following model.In the early phase of the immune response, replication will driveclonal expansion, because the low frequency of antigen-specificCD8+ T cells makes contact between them improbable andCTLA-4 is not surface-expressed. Only when replication hasincreased their numbers will contact between activated cells andtriggering of the Hippo pathway be likely to occur, which thenwill create a steady state wherein the rates of differentiation andreplication are balanced.
DiscussionThe present study finds that the Hippo pathway provides theactivated CD8+ T cell with a means for regulating Blimp-1expression that is independent of other cell types. Becausetriggering of the Hippo pathway is mediated by contact be-tween activated CD8+ T cells, this mechanism for controllingBlimp-1 expression would be linked to the frequency of acti-vated CD8+ T cells within a larger population of nonactivated
T cells, which would be a measure of the magnitude of clonalexpansion in, for example, a lymph node. Thus, the Hippopathway has the requisite properties to allow replicating CD8+
T cells to select the appropriate time for Blimp-1 expressionand terminal differentiation.The naïve CD8+ T cell differs from other cell types in which
the Hippo pathway has been studied in that Mst1 is the onlymember of the core components of the pathway that is expressedconstitutively (Fig. 1A). The need of the naïve CD8+ T cell toexpress Mst1 is consistent with other findings that Mst1 has bi-ological activities independent of the Hippo pathway (35).WW45, Lats1, Mob1, and Yap were all posttranscriptionallyregulated, requiring both antigen and IL-2 for their expression(Figs. 1 and 2), indicating that the growth factors of clonal ex-pansion rapidly establish the core Hippo pathway in the CD8+ Tcell. The essential role for IL-2 in the protein translation of thecore Hippo components and the CTLA-4/CD80 receptor–ligandpair (Figs. 3 and 4) indicates that IL-2, by inducing both Yap andthe serine kinases involved in its degradation and nuclear ex-clusion, can have the opposing functions of suppressing or per-mitting Blimp-1 expression. This dual role for IL-2 in the Hippopathway is reminiscent of its capacity to promote the de-velopment of both memory and terminally differentiated CD8+
T cells (20–24).The Hippo pathway was activated in purified CD8+ T cells
that were cultured in the presence of antigen and IL-2, which wasindicated by the loss of Yap protein secondary to its phosphor-ylation at serine-382 (Fig. 2D), indicating that no other cell types
Fig. 5. Activation of the Hippo pathway by CTLA-4 in vitro and in vivo. (A) Immunoblot analysis of lysates from activated OT-I cells cultured in the presence ofantibodies to CD80 and CD86 and incubated for 1 h with beads bearing isotype control antibody or antibody to CTLA-4. Blots were developed with antibodiesspecific for phosphothreonine-183 of Mst1 and phosphoserine-909 of Lats1. Staining of actin is shown as a loading control. (B) Immunoblot analysis withantiphosphoserine-112 of Yap of lysates from activated OT-I cells expressing Yap S382A-HA that had been cultured in the presence of blocking antibodies toCD80 and CD86 and incubated for 1 h with beads bearing isotype control antibody or antibody to CTLA-4. HA staining is shown as a loading control. (C)Immunoblot analysis of WT Yap in cell lysates from OT-I cells activated in the presence of antibodies to CD80 and CD86 and incubated for 3 h with beadsbearing various ratios of isotype control antibody and antibody to CTLA-4. Staining of actin is shown as a loading control. (D) Analysis by confocal microscopyof anti-Yap–stained OT-I cells that had been activated in the presence of blocking antibodies to CD80 and CD86 and incubated for 3 h with beads bearingisotype control antibody or antibody to CTLA-4 (green, anti-Yap; blue, DAPI). (Scale bars: 50 μm.) (E) Immunoblot analysis of Yap protein in lysates from CTLA-4−/− OT-I cells and WT OT-I cells that had been activated with SIINFEKL peptide and IL-2 for 24 h. (F) Analysis by confocal microscopy of Yap protein in CD8+
and CD4+ cells from the lymph nodes of CTLA-4−/− and CTLA-4−/+ 16- to 20-d-old mice. (Scale bars: 10 μm.)
Thaventhiran et al. PNAS | Published online June 27, 2012 | E2227
IMMUNOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Oct
ober
20,
202
1

were required for this process. The appearance of Yap protein inactivated CD8+ T cells after the addition of nonactivated CD8+
T cells (Fig. 2C) suggested that contact between the activatedCD8+ T cells triggers the Hippo pathway.Two independent studies reported that activation of lympho-
cyte function-associated antigen 1 (LFA-1) in lymphocytes in-volved Mst1 (36) and CTLA-4 (37), respectively, which raisedthe possibility that a Hippo core component and this lymphocytereceptor were in a common pathway. A triggering role forCTLA-4 was consistent with the up-regulation of CD80 by an-tigen- and IL-2–stimulated CD8+ T cells (Fig. 3 A and B), whichinduced Hippo pathway activation, and the absence of CD80 andCD86 on resting CD8+ T cells, which did not have this function(Fig. 2C). More importantly, blocking antibodies to CD80 andCD86 suppressed Yap degradation in contacting activated CD8+
T cells (Fig. 3 C and D), which identified CD28 and CTLA-4 asthe only two candidates for the Hippo pathway receptor. CTLA-4 was formally shown to be a Hippo pathway receptor by theability of immobilized anti–CTLA-4, an unambiguous ligand forCTLA-4, to induce the phosphorylation of Mst1, Lats1, and Yap(Fig. 5 A and B); the degradation of Yap (Fig. 5 C and D) by thestability of Yap in activated CTLA-4−/− CD8+ T cells in vitro(Fig. 5E) and in vivo (Fig. 5F); and the inability of agonistic anti-CD28 to induce degradation of Yap (SI Appendix, Fig. S1B).Two interactions between activated CD8+ T cells, therefore,
may be required for Hippo pathway activation. The first in-teraction involves undefined membrane components and leads toCTLA-4 expression at the plasma membrane of the antigen- andIL-2–stimulated cells (Fig. 4 C and D), which may indicate how,during the early phase of clonal expansion when the low fre-quency of activated CD8+ T cells would make contact betweenthem unlikely, the ligation of CD28 is favored over CTLA-4 byCD80 and CD86 on antigen-presenting cells. The second in-teraction involves CD80 and possibly, CD86 on one cell andCTLA-4 on the other cell, which initiates the Hippo pathwaykinase cascade. Effects of T-cell–T-cell interactions involvingCD80 and CTLA-4 have been observed in vivo (38) and sug-
gested previously to induce anergy (39). Furthermore, these ac-tivated T-cell–T-cell interactions have been directly visualized invivo and shown to occur through stable, multifocal synapse-likestructures (40) to which the microtubule-organizing complexpolarizes. Of interest in this regard is the observation thatCTLA-4 localizes to the microtubule-organizing complex (41).Blocking the transcriptional consequences of Hippo pathway
activation by expressing Yap 5SA at physiological levels (SIAppendix, Fig. S3) enhanced expression of Eomes and sup-pressed expression of Blimp-1 (Fig. 6 A–C), indicating that theHippo pathway enables contact between activated CD8+ T cellsto regulate the expression of these two transcription factors thatare involved in terminal differentiation. Two prior studies ex-amining the in vivo phenotype of Blimp-1−/− CD8+ T cells bothshowed impaired KLRG1 and enhanced CD127 expression,despite having conflicting conclusions regarding secondary mem-ory responses (4, 5). The present finding that Yap 5SA or CTLA-4 deficiency suppresses the development of the terminally differ-entiated KLRG1+/CD127lo CD8+ T cell during a viral infection(Fig. 6 E and F and SI Appendix, Fig. S4 D and F) is consistentwith the ability of Yap 5SA or CTLA-4 deficiency to suppressBlimp-1 expression in vitro.The relationship between the finding that CTLA-4 is the
triggering receptor of the Hippo pathway leading to Blimp-1expression and the phenotypic characteristics of the CTLA-4−/−
mouse may be complex. For example, the CTLA-4−/− mouse hasmassive lymphoproliferation (42, 43), and maintaining Yap ex-pression in hepatocytes causes an apparently similar phenotypeof marked hepatomegaly (25). However, the Yap 5SA-expressingCD8+ T cell and the CTLA-4−/− CD8+ T cell responding toa viral infection showed mainly impaired markers of terminaldifferentiation (Fig. 6 E and F and SI Appendix, Fig. S4 D and F)and not excessive clonal expansion (SI Appendix, Fig. S4 E andG). The marked lymphoproliferation of CTLA-4−/− mice, how-ever, may reflect the effects of persistent stimulation of effectorcells by self-antigen caused by the loss of regulatory function byCD4+ Foxp3+ T cells, whereas control of a viral infection would
Fig. 6. Preventing Hippo pathway activation in CD8+ T cells affects Blimp-1 expression and differentiation. (A) OT-I CD8+ T cells were transduced with the Yap5SA-expressing retrovirus or the pMig vector control and analyzed for Eomes protein by staining of permeabilized cells with anti-Eomes relative to naïve,unstimulated OT-I cells 16 h after restimulation with IL-2 and SIINFEKL peptide. Numbers represent the Eomes-specific mean fluorescence intensity ± SEM. (B)Prdm1gfp/+ CD8+ T cells that had been transduced with control pMig (black line) or the pMig vector-expressing Yap 5SA (red line) and WT CD8+ T cellstransduced with pMig (shaded) were restimulated for 48 h with anti-CD3ε and IL-2 and assessed for GFP fluorescence. Numbers are the GFP-specific meanfluorescence intensities ± SEM. (C and D) OT-I cells transduced with a retrovirus expressing the Lats-resistant Yap 5SA or pMig control vector (C) and CTLA-4+/+
or CTLA-4−/− OT-I cells (D) were analyzed for Blimp-1 mRNA 48 h after restimulation with IL-2 and SIINFEKL peptide. The levels of Blimp-1 mRNA relative to thelevels of naïve unstimulated OT-I cells are shown. Data presented are the mean ± SEM. (E and F) Thy1.2+ OT-I cells that had been transduced with the Yap 5SA-expressing retrovirus or control pMig (E), and Thy1.2+ CTLA-4−/− OT-I cells or CTLA-4+/+ OT-I cells (F) were adoptively transferred to Thy1.1+ C57BL/6 mice, and2 d later, the mice were infected with γ-MHV-68/OVA. On days 8 and 11 postinfection, peripheral blood Thy1.2+ OT-I cells were assessed for expression ofKLRG1. Numbers represent the percentage KLRG1+ cells (means ± SEM). *P < 0.05.
E2228 | www.pnas.org/cgi/doi/10.1073/pnas.1209115109 Thaventhiran et al.
Dow
nloa
ded
by g
uest
on
Oct
ober
20,
202
1

diminish the availability of antigen and terminate clonal expan-sion. Furthermore, with regard to whether the primary role ofthe Hippo pathway is to control proliferation or differentiation,recent reports emphasize the latter function. Overexpression ofYap in murine intestine or chick neural tubes results in loss ofdifferentiated cells (44, 45), and loss of WW45 leads to defectiveterminal differentiation in skin, intestine, and lung epithelia (46).Our results, moreover, are consistent with a previous study ofCTLA-4–deficient CD8+ T cells in mixed bone marrow chimericmice, which reported no difference in clonal expansion betweenWT and CTLA-4−/− CD8+ T cells (47, 48); unfortunately,markers of terminal differentiation, such as KLRG1 and CD127,were not evaluated. The relevance of the present studies to thequestion of the function of CTLA-4 on the CD4+ Foxp3+ T cellmay be that, in its absence, there is impaired Hippo pathway-mediated terminal differentiation and acquisition of regulatoryfunction, which has been shown for Blimp-1−/− regulatory T cells(17). At the very least, the present finding of a cell-intrinsic
function for CTLA-4 in the activation of a conserved, ancientdevelopmental pathway, when coupled with the recent demon-stration of its cell extrinsic function of transendocytosis of CD80and CD86 (49), is additional evidence of the range of biologicalactivities of CTLA-4 that account for its central role in theimmune system.
Materials and MethodsThe details of the mice strains used and their maintenance are contained inS1 Materials and Methods. Similarly, the preparation of tissue samples andcell culture, the antibodies used, the retroviral transduction, and the γ-MHV-68/OVA infection are described there. Also included in this section are theprotocols fro the qRT-PCR, immunoblot, flow cytometry and confocal assays.
ACKNOWLEDGMENTS. We thank F. Randow and E. Roberts for their helpfuldiscussions, F. Randow and A. Betz for their review of the manuscript, andL. Walker for providing lymph nodes from CTLA-4 mutant mice. M.B.-W. wassupported by Sonderforschungsbereich Grant 854. This research was sup-ported by the Wellcome Trust.
1. Khanna KM, McNamara JT, Lefrançois L (2007) In situ imaging of the endogenous CD8T cell response to infection. Science 318:116–120.
2. Joshi NS, et al. (2007) Inflammation directs memory precursor and short-lived effectorCD8(+) T cell fates via the graded expression of T-bet transcription factor. Immunity27:281–295.
3. Banerjee A, et al. (2010) Cutting edge: The transcription factor eomesoderminenables CD8+ T cells to compete for the memory cell niche. J Immunol 185:4988–4992.
4. Rutishauser RL, et al. (2009) Transcriptional repressor Blimp-1 promotes CD8(+) T cellterminal differentiation and represses the acquisition of central memory T cellproperties. Immunity 31:296–308.
5. Kallies A, Xin A, Belz GT, Nutt SL (2009) Blimp-1 transcription factor is required for thedifferentiation of effector CD8(+) T cells and memory responses. Immunity 31:283–295.
6. Harrington LE, Janowski KM, Oliver JR, Zajac AJ, Weaver CT (2008) Memory CD4 T cellsemerge from effector T-cell progenitors. Nature 452:356–360.
7. Bannard O, Kraman M, Fearon DT (2009) Secondary replicative function of CD8+ Tcells that had developed an effector phenotype. Science 323:505–509.
8. Turner CA, Jr., Mack DH, Davis MM (1994) Blimp-1, a novel zinc finger-containingprotein that can drive the maturation of B lymphocytes into immunoglobulin-secreting cells. Cell 77:297–306.
9. Kallies A, et al. (2011) A role for Blimp1 in the transcriptional network controllingnatural killer cell maturation. Blood 117:1869–1879.
10. Yeo CJ, Fearon DT (2011) T-bet-mediated differentiation of the activated CD8+ T cell.Eur J Immunol 41:60–66.
11. Joshi NS, et al. (2011) Increased numbers of preexisting memory CD8 T cells anddecreased T-bet expression can restrain terminal differentiation of secondary effectorand memory CD8 T cells. J Immunol 187:4068–4076.
12. Pearce EL, et al. (2003) Control of effector CD8+ T cell function by the transcriptionfactor Eomesodermin. Science 302:1041–1043.
13. Rao RR, Li Q, Odunsi K, Shrikant PA (2010) The mTOR kinase determines effectorversus memory CD8+ T cell fate by regulating the expression of transcription factorsT-bet and Eomesodermin. Immunity 32:67–78.
14. Zhou X, et al. (2010) Differentiation and persistence of memory CD8(+) T cells dependon T cell factor 1. Immunity 33:229–240.
15. Gong D, Malek TR (2007) Cytokine-dependent Blimp-1 expression in activated T cellsinhibits IL-2 production. J Immunol 178:242–252.
16. Pipkin ME, et al. (2010) Interleukin-2 and inflammation induce distinct transcriptionalprograms that promote the differentiation of effector cytolytic T cells. Immunity 32:79–90.
17. Cretney E, et al. (2011) The transcription factors Blimp-1 and IRF4 jointly control thedifferentiation and function of effector regulatory T cells. Nat Immunol 12:304–311.
18. Kwon H, et al. (2009) Analysis of interleukin-21-induced Prdm1 gene regulationreveals functional cooperation of STAT3 and IRF4 transcription factors. Immunity 31:941–952.
19. Sun J, Dodd H, Moser EK, Sharma R, Braciale TJ (2011) CD4+ T cell help and innate-derived IL-27 induce Blimp-1-dependent IL-10 production by antiviral CTLs. NatImmunol 12:327–334.
20. Obar JJ, et al. (2010) CD4+ T cell regulation of CD25 expression controls developmentof short-lived effector CD8+ T cells in primary and secondary responses. Proc NatlAcad Sci USA 107:193–198.
21. Kalia V, et al. (2010) Prolonged interleukin-2Ralpha expression on virus-specific CD8+T cells favors terminal-effector differentiation in vivo. Immunity 32:91–103.
22. Williams MA, Tyznik AJ, Bevan MJ (2006) Interleukin-2 signals during priming arerequired for secondary expansion of CD8+ memory T cells. Nature 441:890–893.
23. BachmannMF, Wolint P, Walton S, Schwarz K, Oxenius A (2007) Differential role of IL-2R signaling for CD8+ T cell responses in acute and chronic viral infections. Eur JImmunol 37:1502–1512.
24. Feau S, Arens R, Togher S, Schoenberger SP (2011) Autocrine IL-2 is required forsecondary population expansion of CD8(+) memory T cells. Nat Immunol 12:908–913.
25. Dong J, et al. (2007) Elucidation of a universal size-control mechanism in Drosophilaand mammals. Cell 130:1120–1133.
26. Zhao B, Li L, Tumaneng K, Wang CY, Guan KL (2010) A coordinated phosphorylationby Lats and CK1 regulates YAP stability through SCF(beta-TRCP). Genes Dev 24:72–85.
27. Schlegelmilch K, et al. (2011) Yap1 acts downstream of α-catenin to control epidermalproliferation. Cell 144:782–795.
28. Magnúsdóttir E, et al. (2007) Epidermal terminal differentiation depends on Blymphocyte-induced maturation protein-1. Proc Natl Acad Sci USA 104:14988–14993.
29. Nishioka N, et al. (2009) The Hippo signaling pathway components Lats and Yappattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass. DevCell 16:398–410.
30. Russ AP, et al. (2000) Eomesodermin is required for mouse trophoblast developmentand mesoderm formation. Nature 404:95–99.
31. Pentcheva-Hoang T, Egen JG, Wojnoonski K, Allison JP (2004) B7-1 and B7-2selectively recruit CTLA-4 and CD28 to the immunological synapse. Immunity 21:401–413.
32. Hegel JK, Knieke K, Kolar P, Reiner SL, Brunner-Weinzierl MC (2009) CD152 (CTLA-4)regulates effector functions of CD8+ T lymphocytes by repressing Eomesodermin. EurJ Immunol 39:883–893.
33. Voehringer D, et al. (2001) Viral infections induce abundant numbers of senescentCD8 T cells. J Immunol 167:4838–4843.
34. Heffner M, Fearon DT (2007) Loss of T cell receptor-induced Bmi-1 in the KLRG1(+)senescent CD8(+) T lymphocyte. Proc Natl Acad Sci USA 104:13414–13419.
35. Choi J, et al. (2009) Mst1-FoxO signaling protects Naïve T lymphocytes from cellularoxidative stress in mice. PLoS One 4:e8011.
36. Katagiri K, ImamuraM, Kinashi T (2006) Spatiotemporal regulation of the kinase Mst1by binding protein RAPL is critical for lymphocyte polarity and adhesion. Nat Immunol7:919–928.
37. Schneider H, Valk E, da Rocha Dias S, Wei B, Rudd CE (2005) CTLA-4 up-regulation oflymphocyte function-associated antigen 1 adhesion and clustering as an alternatebasis for coreceptor function. Proc Natl Acad Sci USA 102:12861–12866.
38. Taylor PA, et al. (2004) B7 expression on T cells down-regulates immune responsesthrough CTLA-4 ligation via T-T interactions [corrections]. J Immunol 172:34–39.
39. Chai JG, Vendetti S, Amofah E, Dyson J, Lechler R (2000) CD152 ligation by CD80 on Tcells is required for the induction of unresponsiveness by costimulation-deficientantigen presentation. J Immunol 165:3037–3042.
40. Sabatos CA, et al. (2008) A synaptic basis for paracrine interleukin-2 signaling duringhomotypic T cell interaction. Immunity 29:238–248.
41. Egen JG, Allison JP (2002) Cytotoxic T lymphocyte antigen-4 accumulation in theimmunological synapse is regulated by TCR signal strength. Immunity 16:23–35.
42. Tivol EA, et al. (1995) Loss of CTLA-4 leads to massive lymphoproliferation and fatalmultiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4.Immunity 3:541–547.
43. Waterhouse P, et al. (1995) Lymphoproliferative disorders with early lethality in micedeficient in Ctla-4. Science 270:985–988.
44. Camargo FD, et al. (2007) YAP1 increases organ size and expands undifferentiatedprogenitor cells. Curr Biol 17:2054–2060.
45. Cao X, Pfaff SL, Gage FH (2008) YAP regulates neural progenitor cell number via theTEA domain transcription factor. Genes Dev 22:3320–3334.
46. Lee JH, et al. (2008) A crucial role of WW45 in developing epithelial tissues in themouse. EMBO J 27:1231–1242.
47. Bachmann MF, et al. (2001) Normal pathogen-specific immune responses mounted byCTLA-4-deficient T cells: A paradigm reconsidered. Eur J Immunol 31:450–458.
48. Bachmann MF, et al. (1998) Normal responsiveness of CTLA-4-deficient anti-viralcytotoxic T cells. J Immunol 160:95–100.
49. Qureshi OS, et al. (2011) Trans-endocytosis of CD80 and CD86: A molecular basis forthe cell-extrinsic function of CTLA-4. Science 332:600–603.
Thaventhiran et al. PNAS | Published online June 27, 2012 | E2229
IMMUNOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Oct
ober
20,
202
1




![Research Paper Connective tissue growth factor (CTGF) regulates … · 2020. 2. 21. · activation of osteoclasts based on the production of RANKL by activated lymphocytes [7]. Osteoclasts](https://img.pdfslide.fr/doc/110x75/5fed3349af9b1e1a2804e3b7/research-paper-connective-tissue-growth-factor-ctgf-regulates-2020-2-21-activation.jpg)









![[Webinaire] Connecteur Nuxeo / Hippo : Faire le lien entre la gestion de contenu et un CMS](https://img.pdfslide.fr/doc/110x75/559513581a28ab84598b464d/webinaire-connecteur-nuxeo-hippo-faire-le-lien-entre-la-gestion-de-contenu-et-un-cms.jpg)








![OsSHI1 Regulates Plant Architecture Through …OsSHI1 Regulates Plant Architecture Through Modulating the Transcriptional Activity of IPA1 in Rice[OPEN] ErchaoDuan,a,1 YihuaWang,a,1](https://img.pdfslide.fr/doc/110x75/5f6d82e92a27c416a935a6ad/osshi1-regulates-plant-architecture-through-osshi1-regulates-plant-architecture.jpg)




