Embed Size (px)
Citation preview

BIOCttlMICA ET BIOPI-IYSICA ACTA 325
BBA 95767
ACTIVITE DE DONATEURS D'AZOTE DES DERIVES FORMIMINES AU
COURS DE LA BIOSYNTHESE DES PURINES CHEZ ESCHERICHIA COLI B ~
Y V E S L E G A L , J E N S H E D E G A A R D ET J E A N R O C H E
Laboratoire de Biochimie Gdndrale et Comparde, Coll~ge de France, Paris (France)
(Re~u le 6 juil let , 1967)
SUMMARY
Nitrogen donor activity o/ the /ormimino compounds during purine biosynthesis in Escherichia coli B
The nitrogen donor activity of the natural formimino-compounds, formimino- glutamic and -aspartic acids and formimino-glycine during purine biosynthesis has been studied in Escherichia coli B.
I. The imino-nitrogen of the formimino-compounds is actively incorporated into 5-amino-4-imidazolecarboxamide (AICA) by partially sulfadiazine-inhibited growing cells of E. coli B. This incorporation is enhanced while that of free ammonia ions is decreased by glutamine. These results indicate that the incorporation of the imino-nitrogen into AICA does not proceed with glutamine as an intermediate and that the mechanism of utilization of the infino-nitrogen is different from that of ammonia ions.
2. The incorporation of E14CJglycine into the precursor of AICA, 5'-phospho- ribosyl-formylglycinamide, and into nucleic acid purines of azaserine-inhibited non- proliferating cells of E. coli shows that the formimino-derivatives exert their nitrogen donor activity specifically during the reaction catalyzed by the 5'-phosphoribosyl- formylglycinamide: L-glutamine amido-ligase (ADP) (EC 6.3.5.3). Results obtained with cells of which the activity of glutamine synthetase is strongly repressed and additionally inhibited by methionine sulfoximine further confirm that the imino- nitrogen is used directly in the synthesis of phosphoribosyl-formylglycinamidine and not with glutamine as an intermediate. The nitrogen donor activities of formimino- glutamic acid and glutamine are approximately identical.
INTRODUCTION
Les 6tapes successives de la biosynth~se de l'acide inosinique A partir de pr6- curseurs simples ont 6t6 bien 6tablies et les origines des diff6rents atomes d'azote du noyau purique d6termindes 1,~. Ainsi, l'azote amid6 de la glutamine est retrouv6
* Ce t r ava i l es t ex t r a i t d ' u n e th~se de doc to ra t d ' E t a t qu i sera prdsentde d e v a n t la Facu l t6 des Sciences de l 'Un ive r s i td de Par i s pa r Y v e s Le Gal.
Abrdvia t ion : AICA, 5 -amino-4 - imidazo leca rboxamide .
Biochim. Biophys. Acta, 149 (1967) 325-334

326 Y. LE GAL, J. HEDEGAARD, J. ROCHE
clans la position N-3 de l'acide inosinique ~t la suite de son incorporation au niveau de la formation de 5'-phosphoribos~l-formylglycinamidine (formylglycimidine ribo- tide). De plus, cet azote est 6galement utilis6 lors de la synth~se de ribos:ylamine-5'- phosphate ~ partir de 5'-phosphoribosyl-pyrophosphate et d 'ATP; il devient ainsi le pr6curseur de l'azote N- 9 d'acide inosinique. L'azote ammoniacal peut aussi s'in- corporer dans le ribosylamine-5'-phosphate par une deuxi~me voie de synth&se, qvi proc~de directement ~ partir de ribose-5'-phosphate, d 'ATP et d'ions ammonium et dont la nature enzymatique a r6cemment 6t6 mise en 6vidence ~,4.
L'azas6rine, analogue de la L-glutamine, inhibe comp6titivement l'activit6 de donateur d'azote de la glutamine lors de la r6action catalys6e par la 5'-phosphoribo- syl-formylglycinamide: L-glutamine amido-ligase (ADP) (EC 6.3-5.3). Cette inhibi- tion peut cependant 8tre antagonisde chez E. coli B in vivo par la formiminoglycine et par les m6tabolites de l'histidine, les acides formimino-glutamique et -aspartique 5. Ces d6riv~s formimin6s, qui ne semblent pourtant pas ~tre des produits naturels de ]a d6gradation de l'histidine chez E. coli ~, pr6sentent certaines analogies structurales avec la glutamine et possddent, comme cette derni~re, un atome d'azote labile, l'azote imin6. L'antagonisme entre l'azas6rine et les d6riv6s formimin6s pent 6tre dfi simplement ~ un effet structural ou bien ~t une activit6 m6tabolique de ces compos6s en tant que donateurs d'azote; ce probl~me demeurait ouvert.
Au cours du pr6sent travail, le r61e et les activit6s respectives de donateurs d'azote des d6riv6s formimin6s, de la glutamine et des ions ammonium dans la bio- synth~se des purines ont 6t6 6tudi6s chez E. coli B. Les r6sultats obtenus confirment que les ddriv6s formimin6s activent la synth~se de ribosylamine-5'-phosphate 4 o/1 leur action pourrait s'6tablir avec les ions ammonium comme interm6diaires. De plus, il ressort des faits observ6s que les acides formimino-glutamique et -aspartique et la formiminoglycine peuvent participer, directement et en tant que donateurs de leur azote imin6, ~ la formation de 5'-phosphoribosyl-formylglycinamidine, et cela par l 'action de la phosphoribosyl-formylglycinamidine synth~tase.
MATt~RIEL ET MI~THODES
La glycine, les acides glntamique et aspartique, et la glutamine utilis6s sont des produits Hoffman-La Roche et British Drug House. Les d6riv6s formimin6s de la glycine et des acides glutamique et aspartique ont 6t6 synth6tis6s par conden- sation de la formamidine avec l'acide amin6 correspondant ~. L'utilisation de la [15N]- formamidine s a permis la pr6paration des d6riv6s formimin6s marqu6s dans l'azote imin6 par le [15NJazote. La [15N~amide-glutamine est un produit Merck, Sharp & Dohme et le nitrate de [~SN]ammonium provient de l'Office National Industriel de l'Azote (France). La i14C]glycine a 6t6 fournie par le C.E.A. (France). La sulfadiazine est un produit Struers (Danemark) et l'azas6rine nous a 6t6 g6n6reusement offerte par le Dr. Dice (Parke, Davis & Co., Michigan, U.S.A.). La m6thionine sulfoximine est un produit Sigma (U.S.A.).
Cultures bactdriennes La souche d'E. coli B, No 54.125 utilis6e provient de la collection de l 'Institut
Pasteur, Paris. Le microorganisme a 6t6 cultiv6 avec agitation, ~ 30°, dans un milieu
Biochim. Biophys. Acta, 149 (1967) 325-334

DONATEURS D'AZOTE DANS LA BIOSYNTHI~SE PURIQUE 327
minfral glucos6 contenant 2.7. lO-8 M d'i0Es ammonium 9. L'inoculation a 6t~ faite ~t partir d'une adaptation vieille de 16 h e t la croissance des cellules arrSt6e aprils I2-~4 h de culture par centrifugation ~ froid. La densit6 bact6rienne a 6t6 suivie par des mesures au photom~tre Klett-Summerson, ~ 54 ° m # .
Inhibition d'E. coli B par la sul[adiazine La culture d 'E. coli B e n sulfamide bact6riostase a 6t6 effectufe dans le milieu
glucos~ 9 suppl6ment6 en o.75.Io-~M glycine et contenant la sulfadiazine (conc. finale: 0.5" Io -4 M). L'inhibition de la croissance est alors de 60 % environ et permet une accumulation optima dans ce milieu du pr6curseur purique, I'AICA riboside 1°. Le glucose ainsi que les divers produits azot6s (acide formiminoglutamique, acide formiminoaspartique, formiminoglycine, glutamine), dont l 'incorporation int6grale ou partielle dans I'AICA riboside a 6t6 6tudi~e, ont 6t6 introduits stdrilement dans le milieu (conc. finale: o.75-1o -~ M) par filtration (Millipore, 0.45 #). Dans certains cas, les d6riv6s formimin6s ont 6t6 hydrolys6s avant d'Stre ajout6s au milieu de culture (hydrolyse en cellule de Conway, (pH II) , 24 h, 20 °, captation de l 'ammoniaque lib~r6 par H2SO4). Les produits de cette hydrolyse (l'acide amin6 correspondant au ddriv6 formimin6, l 'azote ammoniacal et le formiate) ont alors 6t6 rfunis et neutra- lis6s (pH 7.2) avant d'Stre aiout6s au milieu de culture.
L'accumulation de I'AICA riboside dans le milieu de culture a 6t6 mesur~.e par la m6thode de BRATTON ET MARSHALL 11 et I'AICA riboside a ensuite 6t6 isol6 et purifid 12. Les rapports ~5 N/14N de I'AICA et de l'azote labile du milieu ont 6t6 mesur6s s6par6ment par spectrom6trie de masse apr~s min6ralisation et transformation de l 'azote ammoniacal en azote gazeux 13.
Inhibition d'E. coli B par l'azasdrine L'inhibition d'E. coli B par l 'antagoniste de la L-glutamine, l'azas6rine, r6-
sulte d 'un bloquage pr6f6rentiel de la 5'-phosphoribosyl-formylglycinamid¢: L-glu- tamine amido-ligase (ADP) (EC 6.3.5.3) avec l 'accumulation intracellulaire conse- cutive de 5'-phosphoribosyl-formylglycinamide (formylglycinamide ribotide). L'6- tude de l'activit6 des d~rivfis formimin~s (acide formiminoglutamique, acide formimi- noaspartique, formiminoglycine), de la glutamine et des ions ammonium en tant que donateurs d'azote lors de la r6action catalys6e par la phosphoribosyl-formylglycina- midine synth6tase a 6t6 effectude avec des cellules d 'E. coli B obtenues apr~s culture en milieu min6ral glucos69.
Apr~s 2 lavages successifs avec o.oi M de tampon phosphate (pH 7.2, o-4°), les cellules ont 6t6 remises en suspension dans le milieu de culture glucos6 (4 mg de poids sec par ml) et pr6incub6es pendant ~o rain ~ 30 ° avant addition de l'azas6rine. La [14C~glycine et le formiate (conc. finales lO -3 M) ainsi que le donateur d'azote (acide formiminoglutamique, acide formiminoaspartique, formiminoglycine, gluta- mine on ions ammonium) ont 6t6 ajout6s 5~ la suspension de cellules 30 mill apr~s l'azas~rine. Apr~s 12o min d'incubation ~ 3 °0 avec agitation, la suspension des cel- lules a 6t~ rapidement refroidie et trait6e par l'acide perchlorique (conc. finale 0.55 M). Les nucl6otides libres (y compris le formylglycinamide ribotide) ainsi solubilis6s ont
Biochim. Biophys. Acta, 149 (1967) 325-334

328 Y. LEGAL, J. HEDEGAARD, J. ROCHE
6t6 fix6s sur une colonne de Dowex I X 8, 1OO--2OO mesh HCOO-, et la [14C]glycine compl~tement 61imin6e par lavage avec o. 5 M d'acide formique 14,a5. Le ribotide de formylglycinamide a ensuite 6t6 61u6 h l'6tat chromatographiquement et radiogra- phiquement pur par 4 M d'acide formique. L'incorporation de la [14C3glycine dans le formylglycinamide ribotide a 6t6 mesur6e au compteur ~t scintillation liquide e t a permis de d6terminer le taux d'accumulation intracellulaire de ce dernier.
Les acides nucl6iques des cellules d 'E. coli B incub6es en pr6sence d'azas6rine et de diff6rents donateurs d'azote ont 6t6 paral]~lement isol6s. Apr~s 61imination du surnageant d'incubation par centrifugation A froid, les cellules ont 6t6 remises en suspension dans 0.5 ml d'eau et extraites par 8 vol. d'~thanol (5 rain, IOO°). Le r6sidu cellulaire a 6t6 trait6 par le solvant 6thanol-6ther (I : 1, v/v) pendant 15 rain A 4 °0 avant extraction des acides nucl6iques par l'acide trichloroac6tique ~t 20 % (20 min, 90°). Apr~s 61imination d'acide trichloroac6tique par des extractions successives ~t l'6ther, les acides nuel6iques ont 6t6 hydrolys6s (HC1, I M; IOO°; 60 min) et les bases puriques s6par6es et isoldes ~ l'6tat pur p~r chromatographie ascendante sur papier Whatman No. I (solvant: 6thanol, 95 o/ , .... ,)-acetate d'ammonium, I M; 75:3 ° v/v), L'incorporation de la ~laC]glycine dans l'ad6nine et dans la guanine ~ 6t6 d6termin6e par comptage en scintillation liquide.
L'activit6 de la glutamine synth6tase (EC 6.3.1.2 ) d 'E. coli B a 6t6 mesur6e d'apr6s la m6thode de KOHLHAW, DR/~GERT ET HOLZER ~6.
RESULTATS
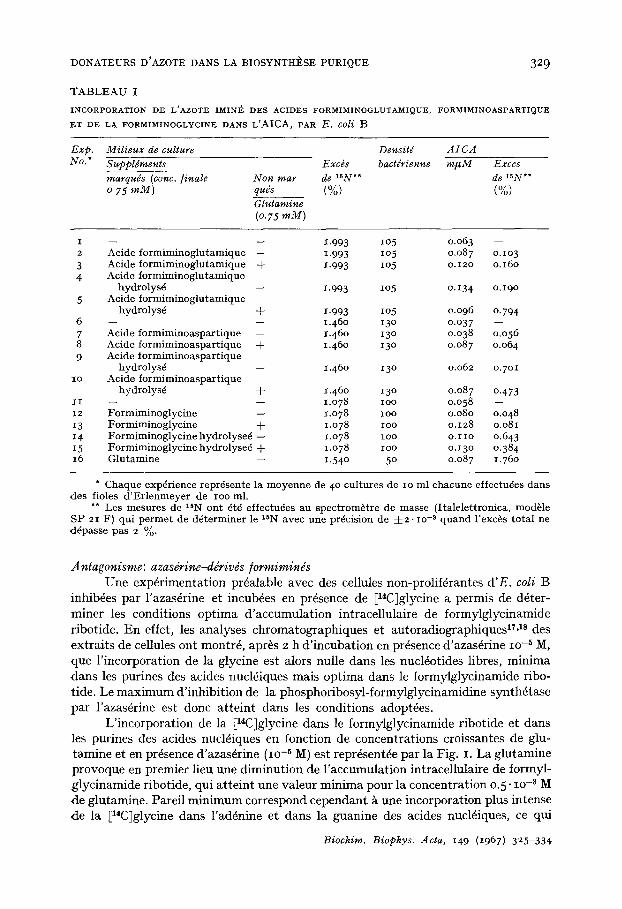
Incorporation dans l'AICA-riboside de l'azote imind des ddrivds /ormiminds Quand E. coti B est cultiv6, en sulfamide bact6riostase dans un milieu min6ral
glucos6 suppl6ment6 en glycine et en un des d6riv6s [15N]imine-formimin6s, on ob- serve une nette incorporation de 15N dans I'AICA (Tableau I). I1 convient de souligner que les taux d'incorporation sont donn6s par rapport ~ l'ensemble des atomes d'azote de la mol6cule d'AICA. Or, seuls les atomes d'azote N-3 et N- 9 peuvent provenir des azotes imin6s, tandis que les origines des atomes d'azote N-I et N- 7 sont respec- tivement et sp6cifiquement l'acide aspartique et la glycine. Aussi l 'incorporation r6elle de l'azote imin6 dans I'AICA serait-elle soit deux, soit quatre fois sup6rieure
celle indiqu6e dans les tableaux. Les taux d'incorporation de l'azote imin6 de l'acide formiminoglutamique et de la formiminoglycine sont sensiblement6 quivalents, tandis que celui de l'azote imin6 de l'acideformiminoaspartique est ldg+rement plus faible.
Dans le cas oft le d6rivd [15N]imine-formimin6 a 6t6 ajout6 au milieu de culture simultan6ment ~t la glutamine, on observe une augmentation de l'incorporation de l'azote imin6 dans I'AICA (Tableau I). Cette augmentation indique clairement que l 'incorporation de l'azote imin6 ne s'effectue pas avec l'azote amid6 de la glutamine comme interm6diaire. Par contre, l'incorporation de l'azote imin6, lib6r6 par hydro- lyse sous forme d'ions ammonium, diminue sensiblement en pr6sence de la glutamine (Tableau I). Ce dernier r6sultat, qui indique que le m6canisme d'incorporation de N-imind dans I'AICA est diff6rent de celui des ions ammonium, pourrait cependant trouver son explication dans l'utilisation de glutamine et d'ions ammonium ~ la synth+se de ribosylamine-5'-phosphate, pr6curseur d'AICA ribosidO.
Biochim. Biophys. Acla, I49 (I967) 325-334
DONATEURS D'AZOTE DANS LA BIOSYNTHI~SE PURIQUE 329
T A B L E A U I
I N C O R P O R A T I O N D E L ' A Z O T E I M I N I ~ D E S A C I D I ~ S F O R M I M I N O G L U T A M I Q U E . F O R M I M I N O A S P A R T I Q U E
E T D E L& F O R M I M I N O G L Y C I N E D A N S L'AICA, P A R E. coli B
Exp. Milieux de culture Densitd AICA No. * Suppldments Exc~s bactdrienne m#M Exces
marquds (conc. finale Non mar de 15N*" de 15N'~ o 75 raM) quds (%) (%)
Glutamine (o.75 mM)
I - -
2 Acide formiminoglu tamique -- 3 Acide formiminoglu tamique + 4 Acide formiminoglutamique
hydrolys6 5 Acide formiminoglutamique
hydrolys6 +
7 Acide formiminoaspar t ique -- 8 Acide formiminoaspar t ique + 9 Acide to rmiminoaspar t ique
hydrolysd Io Acide formiminoaspar t ique
kydrolys6 + I 1 - -
I2 Formiminoglycine 13 Formiminoglycine + 14 Formiminoglycine hydrolysed -- 15 Formiminoglycine hydrolyse6 + 16 Glutamine
1-993 lO5 0.063 -- 1.993 lO5 o.o87 O.lO3 1.993 lO5 o.12o o.16o
1.993 lO5 o.134 o.19 °
1.993 lO5 0.o96 o.794 1.46o 13o o.o37 -- 1.46o 13o o.038 o.o56 1.46o 13o o.o87 o.o64
1.46o 13o 0.062 o.7oi
1.46o 13o 0.087 o.473 Lo78 lOO 0.058 -- 1.o78 lOO 0.080 0.048 1.o78 IOO o.128 o.o81 i.o78 ioo o.I iO 0.643 1.o78 ioo o.13o 0.384 1.54 ° 5 ° o.087 1.76o
* Chaque exp6rience repr6sente la moyenne de 4 ° cultures de IO ml chacune effectu6es darts des fioles d 'Er lenmeyer de ioo ml.
** Les mesures de 15N ont ~t6 effectu~es au spectrom~tre de masse (Italelettronica, module SP 21 F) qui pe rmet de d~terminer le lSN avec une precision de -4-2. IO -3 quand l'exc~,s total ne d6passe pas 2 %.
A ntagonisme: azasdrine-ddrivds [ormiminds Une exp6rimentation pr6alable avec des cellules non-prolifdrantes d'E. coli B
inhib~es par l'azas~rine et incub6es en pr6sence de [14C~glycine a permis de d6ter- miner les conditions optima d'accumulation intracellulaire de formylglycinamide ribotide. En effet, les analyses chromatographiques et autoradiographiques 1~,1s des extraits de cellules ont montr6, apr~s 2 h d'incubation en pr6sence d'azas~rine lO -5 M,
que l'incorporation de la glycine est alors nulle dans les nucl6otides libres, minima dans les purines des acides nucl6iques mais optima dans le formylglycinamide ribo- tide. Le maximum d'inhibition de la phosphoribosyl-formylglycinamidine synth6tase par l'azas~rine est donc atteint dans les conditions adopt6es.
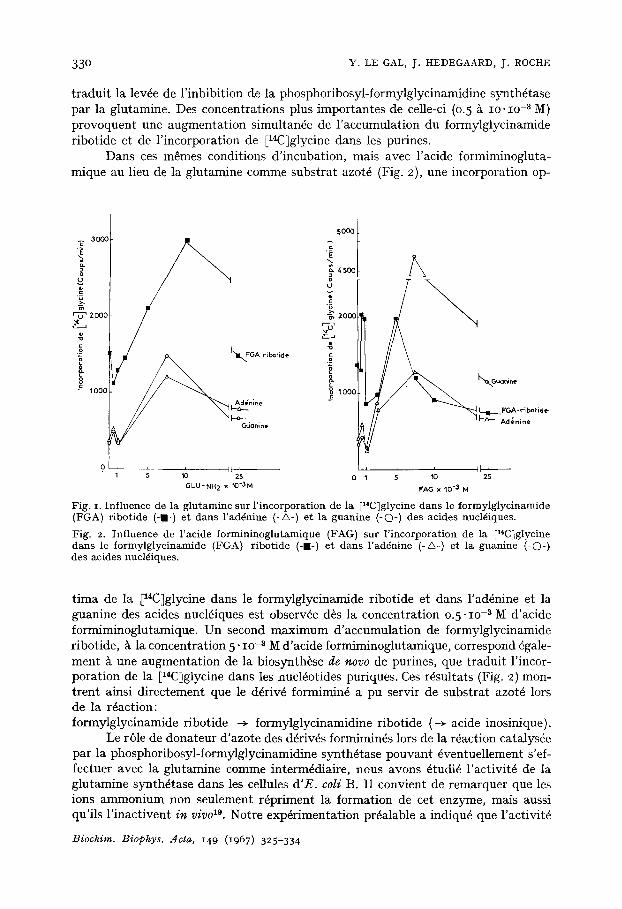
L'incorporation de la E14C~glycine dans le formylglycinamide ribotide et dans les purines des acides nucMiques en fonction de concentrations croissantes de glu- tamine et en pr6sence d'azas6rine (lO -5 M) est repr6sent6e par la Fig. I. La glutamine provoque en premier lieu une diminution de l 'accumulation intracellulaire de formyl- glycinamide ribotide, qui atteint une valeur minima pour la concentration 0.5" Io -~ M de glutamine. Pareil minimum correspond cependant ~ une incorporation plus intense de la ElaClglycine dans l'addnine et dans la guanine des acides nucl6iques, ce qui
Biochim. Biophys. Acta, 149 (1967) 325-334
330 Y. LE GAL, J. HEDEGAARD, J. ROCHE
traduit la lev6e de l'inhibition de la phosphoribosyl-formylglycinamidine synthftase par la glutamine. Des concentrations plus importantes de celle-ci (0.5 ~ IO. IO -z M} provoquent une augmentation simultan6e de l 'accumulation du formylglycinamide ribotide et de l'incorporation de [l~C~glycine dans les purines.
Dans ces m~mes conditions d'incubation, mais avec l'acide formiminogluta- mique au lieu de la glutamine comme substrat azot6 (Fig. 2), une incorporation op-
3OO0
u
~G, 2ooo
3 1000
~soo L o
~. 200C
7 , , ,-,
i100~ A "il
4/ ' ,i 5 10 25 0
GLU-NH 2 x "13-3M
, , I1 ' 5 10 25
FAG x 10 "3 M
Fig. i. Influence de la glutamine sur l'incorporation de la [14C]glycine dans le formylglycinamide (FGA) ribotide (-I-) et dans l'ad~nine (-A-) et la guanine (-O-) des acides nucl6iques. Fig. 2. Influence de l'acide formininoglutamique (FAG) sur l'incorporation de la E14C]glycine darts le formylglycinamide (FGA) ribotide (-B-) et dans l'ad6nine (-A-) et la guanine (-©-} des acides nucl6iques.
tima de la [14C]glycine dans le formylglycinamide ribotide et dans l'ad6nine et la guanine des acides nucl6iques est observ6e d~s la concentration 0.5" I0 -s M d'acide formiminoglutamique. Un second maximum d'accumulation de formylglycinamide ribotide, ~ la concentration 5" IO-S M d'acide formiminoglutamique, correspond 6gale- ment ~ une augmentation de la biosynth~se de novo de purines, que traduit l'incor- poration de la [14C~glycine dans les nucMotides puriques. Ces r~sultats (Fig. 2) mon- trent ainsi directement que le d6riv6 formimin6 a pu servir de substrat azot~ lors de la r6action: formylglycinamide ribotide -~ formylglycinamidine ribotide (-~ acide inosinique).
Le r61e de donateur d'azote des d6riv6s formimin6s lors de la r6action catalys6e par la phosphoribosyl-formylglycinamidine synth6tase pouvant 6ventuellement s'ef- fectuer avec la glutamine comme interm6diaire, nous avons 6tudi6 l'activit6 de la glutarnine synth~tase dans les cellules d 'E. coli B. I1 convient de remarquer que les ions ammonium non seulement r6priment la formation de cet enzyme, mais aussi qu'ils l 'inactivent in vivo 19. Notre exp6rimentation pr6alable a indiqu6 que l'activit6
Biochim. Biophys. Acta, 149 (1967) 325-334
DONATEURS D'AZOTE DANS LA BIOSYNTHESE PURIQUE 331
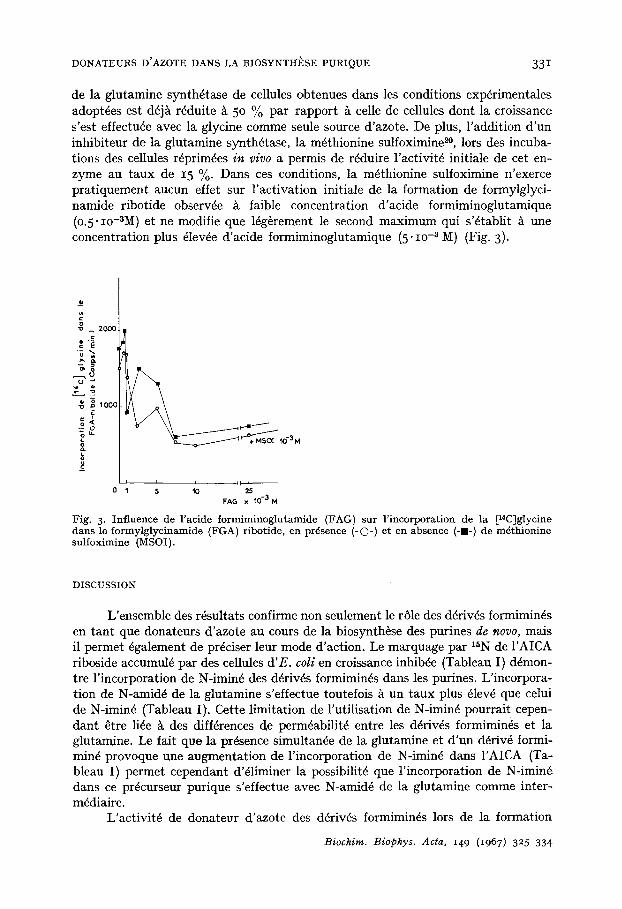
de la glutamine synth6tase de cellules obtenues darts les conditions exp6rimentales adopt6es est d6j~ r6duite A 50 % par rapport ~ celle de cellules dont la croissance s'est effectu6e avec la glycine comme seule source d'azote. De plus, l 'addition d'un inhibiteur de la glutamine synth~tase, la m6thionine sulfoximine 2°, lors des incuba- tions des cellules r6prim6es in vivo a permis de r~duire l'activit~ initiale de cet en- zyme au taux de 15 %. Dans ces conditions, la m6thionine sulfoximine n'exerce pratiquement aucun effet sur l 'activation initiale de la formation de formylglyci- namide ribotide observ6e ~ faible concentration d'acide formiminoglutamique (0.5" Io-~M) et ne modifie que 16g~rement le second maximum qui s'6tablit A une concentration plus 61evde d'acide formiminoglutamique (5" IO-~ M) (Fig. 3).
-~ 2ooo c
'--G, ~
~" ~ too(
=,? o
t , i i I
5 t O 25 FAG x 10 -3 M
Fig. 3. Influence de racide formiminoglutamide (FAG) sur l'incorporatioi1 de la [14C]glycine darts le formylglycinamide (FGA) ribotide, en pr6sence (-O-) et el1 absence (-I-) de m~thionine sulfoximine (MSOI).
DISCUSSION
L'ensemble des r6sultats confirme non seulement le r61e des d6riv6s formimin6s en rant que donateurs d'azote au cours de la biosynth~se des purines de novo, mais il permet 6galement de pr6ciser leur mode d'action. Le marquage par lsN de I'AICA riboside accumul6 par des cellules d 'E. coli en croissance inhib6e (Tableau I) d6mon- tre l 'incorporation de N-imin6 des d6riv6s formimin6s dans les purines. L'incorpora- tion de N-amid6 de la glutamine s'effectue toutefois A un taux plus 61ev6 que celui de N-imin6 (Tableau 11. Cette limitation de l'utilisation de N-imin6 pourrait cepen- dant 8tre li6e A des diff6rences de perm6abilit6 entre les d6riv6s formimin6s et la glutamine. Le fait ClUe la pr6sence simultan6e de la glutamine et d 'un d6riv6 formi- min6 provoque une augmentation de l'incorporation de N-imin6 dans I'AICA (Ta- bleau I) permet cependant d'61iminer la possibilit6 que l'incorporation de N-imin6 dans ce pr6curseur purique s'effectue avec N-amid6 de la glutamine comme inter- m6diaire.
L'activit6 de donateur d'azote des d6riv6s formimin6s lots de la formation
Biochim. Biophys. Acta, 149 (1967) 325-33¢

332 Y. LE GAL, J. HEDEGAARD, J. ROCHE
d'AICA riboside pourrait s'effectuer au niveau de l'une ou des deux rdactions pour laquelle la glutamine a gdndralement dtd reconnue comme dtant le donateur d'azote spdcifique: 5'-phosphoribosyl-pyrophosphate+glutamine ~ ribosylamine-5'-phosphate+acide glutamique (I) formylglycinamide r ibotide+glutamine --> formylglycinamidine r ibotide+acide glutamique (2) En ce qui concerne la rdaction I (synth~se de ribosylamine-5'-phosphate), nous avons rdcemment ddmontrd que le ribosylamine-5'-phosphate peut dgalement se former directement et par voie enzymatique (extrait d 'E. coli B) ~ partir de ribose-5'- phosphate et des ions ammonium 4. L'existence de cette double voie de synth~se de ribosylamine-5'-phosphate expliquerait ainsi que le taux d'incorporation dans AICA de 15N ammoniacal, ddrivd de l'azote imind apr~s hydrolyse, soit plus dlevd.
L'dtude de l'accumulation de formylglycinamide ribotide par des cellules non- prolifdrantes d 'E. coli B inhibdes par l'azasdrine permet de prdciser les modalitds d'utilisation de N-imind des ddrivds formiminds ~. la biosynth~se des purines. En effet, une augmentation de l 'accumulation intracellulaire de formylglycinamide ribo- tide indique que l 'activation par le donateur d'azote (ddrivd formimind, glutamine, ions ammonium) de la formation du prdcurseur de formylglycinamide ribotide, le ribo- bosylamine-5'-phosphate, est supdrieure au degrd de levde d'inhibition de la phos- phoribosyl-formylglycinamidine synthdtase. Cependant, quand cette augmentation est accompagnde par une activation simultande de l'incorporation de [14Clglycine dans les acides nucldiques, l'action du donateur d'azote doit consister dgalement en un antagonisme mdtabolique avec l'azasdrine permettant l 'amidation de formylgly- cinamide ribotide en formylglyeinamidine ribotide. Ainsi, l 'incorporation de la [i4C]- glycine dans l'addnine et dans la guanine des acides nucldiques devient-elle une me- sure de l'activitd mdtabolique des donateurs d'azote pour la phosphoribosyl-formyl- glycinamidine synthdtase. Par contre l 'accumulation de formylglycinaznide ribotide permet, ~t elle seule, d'estimer la synth~se de ribosylamine-5'-phosphate.
Les variations dans l'incorporation de la Ei4C]glycine dans le formylglycinalnide ribotide et dans les purines des acides nucldiques en fonction de concentrations crois- santes d'~cide formiminoglutamique (Fig. 2) refl&tent ainsi la double activitd de ce ddrivd formimind lors de la biosynth~se des purines. En effet, l'acide formiminoglutami- que sert de substrat azot6 pour la phosphoribosyl-formylglycinamidine synthdtase au m~me titre et au m6me degrd que la glutamine (Figs. I e t 2), comme l'indique, dans ces deux cas, la nette augmentation de la synth~se de novo des bases puriques (addnine et guanine) dans des cellules d 'E. coli B dont l'activit6 de la glutamine synthdtase est rdprimde k environ 50 %.
La formation de novo d'addnine et de guanine en prdsence de glutamine s'ef- fectue parall~lement (Fig. I), tandis que celle de la guanine est spdcifiquement et fortement augmentde en prdsence d'acide formiminoglutamique (Fig. 2). Ce rd- sultat pourrait trouver son explication dans une activitd du ddrivd formimind soit directe, soit indirecte (avec les ions ammonium comme intermddiaires) lors de l'ami- nation de XMP en GMP (ref. 21).
La diminution de l'incorporation de E14Clglycine aussi bien dans le formyl- glycinamide ribotide que dans les purines des acides nucldiques ~ des ccncentrations dlevdes d'acide formimninoglvtamique et de glutamine (Figs. I e t 2) pcurrait 8tre like
Biochim. Biophys. Acta, 149 (1967) 325-334

DONATEURS D'AZOTE DANS LA BIOSYNTH#.SE PURIQUE 333
~t une inhibition par le substrat (donateur d'azote) ~ de la syn. th~se de ribosylamine- 5'-phosphate, prdcurseur de formylglycinamide ribotide. En effet, les ions ammonium (IO -2 M) inhibent leur propre utilisation ~t la synth6se directe de ribosylamine-5'- phosphate ~ partir de ribose-5'-phosphate et d 'ATP par E. coli B in vitro*.
L'ensemble de nos r6sultats permet d 'attr ibuer aux ddrivds formiminds un r61e biologique nouveau de donateurs d'azote, r61e qui s'ajoute h celui, dd, j~ connu, de donateurs de carbone ("formyle actif") lors de la biosynth6se des purines. Le fait que cette activit6 nouvelle des ddrivds formiminds ait pu ~tre mise en dvidence chez E. coli, lequel ne d6grade cependant l'histidine ni en acide formiminoglutamique, ni en acide formiminoaspartique, laisserait supposer la prdsence d'une seule enzyme, la 5'-phosphoribosyl-formylglycinamide: L-glutamine amido-ligase, dont la large spdcificitd s'dtend de la glutamine aux d6rivds formiminds, plut6t que l'existence, dans ce microorganisme, d'un syst~me enzymatique spdcifique de ces derniers composds. La ddgradation de l'histidine par l'acide imidazolelactique en composds non h6t6ro- cycliques 6 permet cependant d'envisager la formation chez E. coli d'un analogue naturel des ddriv6s formiminds. Dans ce dernier cas, l'existence d'une activitd phos- phoribosyl-formylglycinamidine synth6tasique spdcifique des ddrives formiminds devrait 8tre envisagde.
R]~SUMI~
L'activit6 de donateurs d'azote des d6riv6s formiminds naturels, les acides formiminoglutamique et-aspartique et la formiminoglycine, dans la biosynth~se des purines a dt6 6tudide chez E. coli B.
I. L'azote imin6 des d6riv6s formimin6s est activement incorpor6 dans la 5- amino-4-imidazolecarboxamide (AICA) par E. coli, dont la croissance est partielle- ment inhib6e par la sulfadiazine. Cette incorporation est sensiblement augment6e tandis que celle des ions ammonium diminue en prdsence de la glutamine. L'incor- poration de N-imind dans I'AICA ne s'effectue donc pas par l'interm6diaire de la glutamine et le mdcanisme de l'utilisation de N-imind diff~re au moins partiellement de celui des ions ammonium.
2, L'incorporation de la [laCJglycine dans le pr6curseur d'AICA, la 5'-phos- phoribosyl-formylglycinamide, et dans les purines des acides nucldiques de cellules non-prolif6rantes d 'E. coli inhib6es par l'azasdrine montre que les d~rivds formimi- n6s peuvent servir de donateurs d'azote sp6cifiquement au niveau de la r~action catalysde par la 5'-phosphoribosyl-formylglycinamide: L-glutamine amido-ligase. L'utilisation de cellules d'E. coli dont l'activit6 glutamine synthdtasique est fortement r6prim~e et, en outre, inhibde par la mdthionine sulfoximine a permis de confirmer que l'activitd de donateurs d'azote des d6riv6s formimin6s lors de la formation de 5'-phosphoribosyl-formylglycinamidine s'exerce directement et non par l'intermddi- aire de la glutamine. Les activitds de donateurs d'azote de l'acide formiminoglu- tamique et de la glutamine paraissent sensiblement 6gales. L'ensemble des r6sultats sugg~re l'existence chez E. coli d'une 5'-phosphoribosyl-formylgycinamide: L-glu- tamine amido-ligase dgalement active sur la glutamine et sur les d6rivds formiminds,
M. L. LEGAL, Y. LEGAL ET J. HED]~GAARD, rdsultats non publi6s.
Biochim. Biophys. Acta, 149 (1967) 325-334

334 Y. LEGAL, J. HEDEGAARD, J. ROCHE
plutht que la prdsence, dans ce microorganisme, d'un syst6me enzymatique spdci- fique de ces derniers.
REMERCIEMENTS
Nous remercions Mlle G. CHEVALIER de sa collaboration technique au cours de ce travail.
Les mesures de I~N au spectrom6tre de masse ont 6t6 effectudes A l'Istituto di Chimica Biologica, Universita di Napoli (Italie) (Dir.: Prof. F. CEDRANGOL0), grace ~ la subvention de I'0.T.A.N. No. 197. Nous remercions le Prof. CEDRANGOL0 d'avoir facilit6 cette coopdration.
BIBLIOGRAPHIE
I B. LEVENBERG, S. C. HARTMAN ET J. M. BUCHANAN, J. Biol. Chem., 220 (I956) 379. 2 J. M. BUCHANAN, in E. CHARGAFF ET J. N. DAVIDSON, The Nucleic Acids, Vol. I I I , Acad.
Press, New York, 196o, p. 303 . 3 M. KAPOOR ET E. R. WAYGOOD, Biochem. Biophys. Res. Commun., 9 (196o) 7. 4 M. L. LEGAL, Y. LEGAL, J. HEDEGAARD ET J. ROCHE, Biochem. Biophys. Res. Cornmun.,
27 (1967) 618. 5 J. HEDEGAARD, N. V. THOAI ET J. ROCHE, Arch. Biochern. Biophys., 83 (1959) 183. 6 J. HEDEGAARD, J. BREVET ET J. ROCHE, Biochem. Biophys. Res. Cornmun., 25 (1966) 335- 7 H. TABOR ET J. C. RABINOWITZ, Biochem. Prepn., 5 (1957) ioo. 8 Y. LEGAL, J. HEDEGAARD, D. CITTADINI ET J. ROCHE, Compt. Rend. Soc. Biol., 16o (1966) 299. 9 J. SPIZIZEN, J. C. KENNEY ET B. HAMPIL, J. Bacteriol., 62 (1951) 323.
io J. HEDEGAARD, Y. LEGAL ET J. ROCHE, Biochim. Biophys. zJcta, ioo (1965) 308. I I A. C. BRATTON ET E. MARSHALL, J. Biol. Chem., 128 (1939) 537. 12 D. A. GOLDTHWAIT ET G. n . GREENBERG, in S. P. COLOWICK ET N. O. KAPLAN, Methods in
Enzymology, Vol. II , Acad. Press, New York, 1955, P. 512. 13 D. B. SPRINSON ET D. RITTENBERG, J. Biol. Chem., 18o (1949) 707 . 14 C-. A. LE PAGE ET M. JONES, Cancer Res., 21 (1961) 642. 15 J. F. HENDERSON, J. Biol. Chem., 237 (1962) 2631. 16 G. KOHLHAW, W. DR~-GERT ET H. HOLZER, Biochem. Z., 341 (1965) 224. 17 A. J. TOMISEK, H. J, KELLY ET H. E. SKIPPER, Arch. Biochem. Biophys., 64 (1956) 437. i8 i . J. TOMISEK, M. R. REID ET H. E. SKIPPER, Cancer Res., 19 (1959) 489. 19 D. MECKE ET H. HOLZER, Biochim. Biophys. Acta, 122 (1966) 341. 20 C. LAMAR ET O. Z. SELLINGER, Biochem. Pharmacol., 14 (1965) 489 . 21 J. HEDEGAARD, A. BREVET, C. BALESTRIERI ET J. ROCHE, Compt. Rend. Soc. Biol., 16o (1966)
2o61. 22 M. DIXON ET E. C. WEBB, Enzymes, Longmans , Green and Co., Londres, 1958, p. 81.
Biochim. Biophys. Acta, 149 (1967) 325-334





























