Embed Size (px)
Citation preview

Analyse des isomères de F2-isoprostanes par chromatographie liquide couplée à la spectrométrie de
masse dans la circulation maternelle en fin de grossesse
Mémoire
Jessica Larose
Maîtrise en physiologie-endocrinologie
Maître ès sciences (M. Sc.)
Québec, Canada
© Jessica Larose, 2013


III
Résumé
Un stress oxydatif survient lorsqu’il y a surproduction de dérivés actifs de l’oxygène par rapport aux
défenses antioxydantes. Ce déséquilibre est associé, entre autres, à la prééclampsie, une pathologie
de la grossesse. Les F2-isoprostanes regroupent soixante-quatre isomères issus de la peroxydation
de l’acide arachidonique. Ceux-ci sont reconnus comme étant les biomarqueurs les plus fiables du
stress oxydatif in vivo. Une méthode d’analyse par chromatographie liquide couplée à la
spectrométrie de masse en tandem pour le dosage de sept isomères de F2-isoprostanes dans des
échantillons de plasma, de sang et de membranes érythrocytaires a été mise au point et validée. Les
F2-isoprostanes dans le plasma ont été corrélés positivement avec plusieurs acides gras trans
plasmatiques au troisième trimestre de la grossesse. Contre toute attente, les F2-isoprostanes du
plasma, du sang et des membranes érythrocytaires sont moins abondants en prééclampsie par
rapport aux contrôles en fin de grossesse.


V
Abstract
An oxidative stress is defined as an imbalance between the production of reactive oxygen species
and antioxidant defenses of the organism. This imbalance has been associated with preeclampsia, a
pathology of the mid-to-late pregnancy. Peroxidation of arachidonic acid generates sixty-four isomers
of F2-isoprostanes. The latter are recognized as the most reliable biomarkers of oxidative stress in
vivo. A method using liquid chromatography tandem mass spectrometry (HPLC-MS/MS) for the
determination of seven isomers of F2-isoprostanes in plasma samples, whole blood and erythrocyte
membranes has been developed and validated. The F2-isoprostanes correlated positively with
several trans fatty acids in plasma at end of the pregnancy. Unexpectedly, F2-isoprostanes were less
abundant in preeclampsia than in control pregnancies at the third trimester.


VII
Table des matières
Résumé .............................................................................................................................................. III
Abstract .............................................................................................................................................. V
Table des matières .......................................................................................................................... VII
Liste des tableaux ............................................................................................................................. XI
Liste des figures ............................................................................................................................... XI
Liste des abréviations .................................................................................................................... XIII
Remerciements ............................................................................................................................... XIX
Avant-Propos .................................................................................................................................. XXI
Chapitre 1 – Introduction .................................................................................................................. 1
1.1. Le stress oxydatif ...................................................................................................................... 1
1.1.1. Les prooxydants ................................................................................................................ 1 1.1.2. Les antioxydants ................................................................................................................ 3
1.2. Les lipides ................................................................................................................................. 9
1.2.1. Définitions et fonctions biologiques .................................................................................... 9 1.2.2. Digestion, absorption et transport dans l’organisme .......................................................... 9
1.2.3. Acide arachidonique ........................................................................................................ 12 1.2.4. Acides gras trans ............................................................................................................. 14 1.2.5. Acides gras et stress oxydatif .......................................................................................... 16
1.3. Les F2-isoprostanes ................................................................................................................ 19 1.3.1. Formation des F2-isoprostanes ........................................................................................ 20
1.3.2. Transport et métabolisme des F2-isoprostanes ................................................................ 22 1.3.3. Mesure des F2-isoprostanes ............................................................................................ 23 1.3.4. Effets biologiques des F2-isoprostanes ............................................................................ 25
1.4. Grossesse et désordre hypertensif avec protéinurie ............................................................... 26 1.4.1. La grossesse, un évènement oxydatif .............................................................................. 26 1.4.2. Désordre hypertensif avec protéinurie (prééclampsie) ..................................................... 27
1.5. Problématique et objectifs ....................................................................................................... 31 1.5.1. Problématique .................................................................................................................. 31 1.5.2. Hypothèses ...................................................................................................................... 32
1.5.3. Objectifs ........................................................................................................................... 32
Chapitre 2 – F2-isoprostanes analysis in plasma of pregnant women by HPLC-MS/MS using a column packed with core-shell particles.......................................................................... 35
2.1. Résumé .................................................................................................................................. 35
2.2. Abstract ................................................................................................................................... 37 2.3. Introduction ............................................................................................................................. 38 2.4. Material and methods ............................................................................................................. 39
2.4.1. Materials .......................................................................................................................... 39 2.4.2. Patient selection .............................................................................................................. 39 2.4.3. Blood collection and processing ...................................................................................... 40 2.4.4. Preparation of standards for F2-isoPs analysis ................................................................ 40

VIII
2.4.5. Extraction of F2-isoPs from plasma .................................................................................. 40 2.4.6. Chromatography .............................................................................................................. 41 2.4.7. Mass Spectrometry .......................................................................................................... 41 2.4.8. Method validation ............................................................................................................. 42 2.4.9. Statistical analyses ........................................................................................................... 42
2.5. Results .................................................................................................................................... 42 2.5.1. Analysis of F2-isoPs in plasma by selected reaction monitoring mass spectrometry ........ 42 2.5.2. Chromatographic separation ............................................................................................ 43
2.5.3. Method validation ............................................................................................................. 43 2.5.4. F2-isoPs in the plasma of pregnant women ...................................................................... 44
2.6. Discussion ............................................................................................................................... 44 2.7. Acknowledgments ................................................................................................................... 46
2.8. References .............................................................................................................................. 46 2.9. Figure legends ........................................................................................................................ 50
Chapitre 3 – F2-isoprostanes are correlated with trans fatty acids in plasma of pregnant women............................................................................................................................................... 57
3.1. Résumé ................................................................................................................................... 57
3.2. Abstract ................................................................................................................................... 59 3.3. Introduction ............................................................................................................................. 60 3.4. Material and methods .............................................................................................................. 61
3.4.1. Materials .......................................................................................................................... 61 3.4.2. Patient recruitment ........................................................................................................... 62 3.4.3. Blood collection and processing ....................................................................................... 62
3.4.4. F2-isoPs analysis by HPLC-MS/MS ................................................................................. 62
3.4.5. Determination of plasma fatty acid profile by gas chromatography .................................. 63 3.4.6. Statistical analyses ........................................................................................................... 63
3.5. Results .................................................................................................................................... 63
3.5.1. F2-isoPs in the plasma of pregnant women ...................................................................... 63 3.5.2. Analysis of plasma fatty acids .......................................................................................... 64 3.5.3. Relationship between F2-isoPs and trans fatty acids........................................................ 64
3.6. Discussion ............................................................................................................................... 64 3.7. Acknowledgments ................................................................................................................... 68 3.8. References .............................................................................................................................. 68
Chapitre 4 – Étude du stress oxydatif en prééclampsie ............................................................... 79
4.1. Introduction ............................................................................................................................. 79 4.2. Matériel et méthode ................................................................................................................ 80
4.2.1. Matériel ............................................................................................................................ 80 4.2.2. Sélection des patientes .................................................................................................... 80 4.2.3. Récolte et traitement des échantillons de sang ................................................................ 80 4.2.4. Préparation des standards pour l’analyse des F2-isoprostanes ....................................... 81 4.2.5. Extraction des F2-isoprostanes totaux contenus dans le plasma ..................................... 81 4.2.6. Extraction des membranes érythrocytaires ...................................................................... 81 4.2.7. Extraction des F2-isoprostanes totaux contenus dans les membranes érythrocytaires .... 81 4.2.8. Extraction des F2-isoprostanes totaux contenus dans le sang total ................................. 82 4.2.9. Analyse des F2-isoprostanes par HPLC-MS/MS .............................................................. 82

IX
4.2.10. Analyses statistiques ..................................................................................................... 82 4.3. Résultats ................................................................................................................................. 82
4.3.1. F2-isoprostanes dans le plasma ....................................................................................... 82 4.3.2. F2-isoprostanes dans le sang........................................................................................... 83 4.3.3. F2-isoprostanes dans les membranes érythrocytaires ..................................................... 84
4.4. Discussion .............................................................................................................................. 85 4.5. Bibliographie ........................................................................................................................... 87
Chapitre 5 – Discussion générale .................................................................................................. 89
Bibliographie .................................................................................................................................... 97
Annexe 1 ......................................................................................................................................... 109


XI
Liste des tableaux
Chapitre 1
Tableau 1. Caractéristiques des différents types de lipoprotéines ..................................................... 10 Tableau 2. Facteurs de risques pour la prééclampsie liés à la mère ................................................. 30
Tableau 3. Facteurs de risques pour la prééclampsie liés au père .................................................... 31 Tableau 4. Facteurs de risques pour la prééclampsie liés à la grossesse ......................................... 31
Chapitre 2
Table 1. Selected reaction monitoring parameters optimized for each classes of F2-isoPs ............... 51 Table 2. Results for the method validation of F2-isoPs. ..................................................................... 52 Table 3. F2-isoPs in the plasma of third trimester pregnant women. .................................................. 53
Chapitre 3
Table 1. F2-isoprostanes in the plasma of third trimester pregnant women. ...................................... 75 Table 2. Fatty acids of interest in plasma of third trimester pregnant women. ................................... 76 Table 3. Correlations between F2-isoprostanes and trans fatty acids in plasma of third
trimester pregnant women. .................................................................................................. 77
Chapitre 4
Tableau 1. F2-isoprostanes totaux mesurés dans le plasma de femmes au troisième trimestre d’une grossesse normotensive ou prééclamptique. .......................................... 83
Tableau 2. F2-isoprostanes totaux mesurés dans le sang entier de femmes au troisième trimestre d’une grossesse normotensive ou prééclamptique. .......................................... 84
Tableau 3. F2-isoprostanes totaux mesurés dans les membranes érythrocytaires de femmes au troisième trimestre d’une grossesse normotensive ou prééclamptique. ............................................................................................................... 85
Liste des figures
Chapitre 1
Figure 1. Cycle du glutathion. .............................................................................................................. 4 Figure 2. Système de la thiorédoxine. Adaptée de [24]. ...................................................................... 5 Figure 3. Exemples de petites molécules antioxydantes. .................................................................... 6 Figure 4. Structures des tocophérols et des tocotriénols. .................................................................... 7 Figure 5. Neutralisation du radical peroxyle en hydroperoxyde organique par l’α-tocophérol
qui est à son tour recyclée par la vitamine C (ascorbate). Adaptée de [31]. ......................... 8 Figure 6. Schémas général d'une lipoprotéine. Tirée de [41]. ............................................................ 10 Figure 7. Transport des lipides par les lipoprotéines. Tirée de [45]. .................................................. 12 Figure 8. Voies de synthèse des eicosanoïdes dérivés de l’acide arachidonique. Tirée de
[46]. .................................................................................................................................... 13 Figure 9. Conversion de l'acide linoléique en acide arachidonique. ................................................... 14

XII
Figure 10. Distribution des t-18:1 dans (A) l’huile végétale partiellement hydrogénée (margarine, n = 46 échantillons) et dans (B) le gras de lait de vache (n = 1765 échantillons). Adapté de [57]. ........................................................................................... 15
Figure 11. Mécanisme général de la peroxydation des lipides. .......................................................... 17 Figure 12. Structure des différents types d'isoprostanes dérivés de l'acide arachidonique.
Tirée de [9]. ...................................................................................................................... 18 Figure 13. Mécanisme général de la réaction d'isomérisation cis-trans induite par les
radicaux libres. X• représente n’importe quel radical capable d’induire cette réaction. Tirée de [22]. ...................................................................................................... 19
Figure 14. Structures de la Prostaglandine F2α (PGF2α) et du 8-iso PGF2α. ....................................... 20 Figure 15. Mécanisme de formation des F2-isoprostanes. Adaptée de [92]. ...................................... 21 Figure 16. Structure d’un peroxyde dioxolane-isoprostane (classe IV). R = -(CH2)3COOH................ 22
Figure 17. Structures du 2,3-dinor-8-iso-PGF2α et le 2,3-dinor-5,6-dihydro-8-iso-PGF2α. ................... 23 Figure 18. Dérivation chimique pour l'analyse des F2-isoPs par GC-MS. Tirée de [104]. .................. 24 Figure 19. Signalisation impliquant le 8-iso-PGF2α dans la fonction plaquettaire. Tirée de
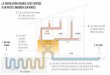
[115]. ................................................................................................................................ 25 Figure 20. Invasion des artères spiralées par les cytotrophoblastes dans une grossesse
sans complications. Tirée de [130]. .................................................................................. 27 Figure 21. Invasion des artères spiralées par les cytotrophoblastes dans une grossesse
prééclamptique. Tirée de [130]. ........................................................................................ 29
Chapitre 2
Figure 1. Chemical structures of analytes investigated by LC-MS/MS. .............................................. 54 Figure 2. Mass chromatograms of a standard solution (A-F) and of a typical plasma sample
spiked with 50 pg of each internal standard (G-L). ............................................................. 55
Chapitre 5
Figure 1. Compétition entre la formation des F2-isoprostanes et des isofuranes ............................... 94

XIII
Liste des abréviations
11t-18:1 : Acide trans-vaccénique
1O2 : Oxygène singulet
9t-18:1 : Acide élaïdique
AA : Acide arachidonique
AA-CoA : Acide arachidonique estérifié à la coenzyme A
ALA : Acide alpha-linolénique
AMPc : Adénosine monophosphate cyclique
Asc•- : Radical ascorbyl
AscH- : Ascorbate
BHT : Hydroxytoluène butylé; butylated hydroxytoluene
CAT : Catalase
CCl4 : Tétrachlorométhane
CETP : Protéine de transfert des esters de cholestérol; cholesteryl ester transfer protein
CM : Chylomicrons
CoA : Coenzyme A
COX : Cyclo-oxygénases
Cu : Cuivre
CYP : Cytochromes P450
DHA : Acide docosahexaénoïque
DHAsc : Dehydroacscorbate
ELISA : Méthodes immuno-enzymatiques
F2-isoPs : F2-isoprostanes
Fe : Fer
GC-MS/MS : Chromatographie en phase gazeuse couplée à la spectrométrie de masse en tandem
GC-NICI-MS : Chromatographie en phase gazeuse couplée à la spectrométrie de masse en mode « ionisation chimique négative »
GPR : Glutathion réductase
GPx : Glutathion peroxydases
GSH : Glutathion réduit
GSSG : Glutathion oxydé
GST : Glutathion-s-transférase
H• : Hydrogène

XIV
H2O2 : Peroxyde d’hydrogène
HDL : Lipoprotéines de haute densité; high density lipoprotein
HELLP syndrome : Hemolysis, Elevated Liver enzymes and Low Platelets count syndrome
HL : Lipase hépatique; hepatic lipase
HO• : Radical hydroxyle
HOCl : Acide hypochloreux
HPLC-MS/MS : Chromatographie liquide couplée à la spectrométrie de masse en tandem
IDL : Lipoprotéines de densité intermédiaire
isoFs : Isofuranes
isoP : Isoprostanes
ISR : Récepteur des isoprostanes
iTFA : Acides gras trans d'origine industrielle
L• : Lipide radicalaire
LA : Acide linoléique; linoleic acid
LCAT : Lécithin:cholestérol acyl transférase
LDL : Lipoprotéines de basse densité; low density lipoprotein
LOO• : Radical peroxyle de lipide
LOOH : Hydroperoxyde de lipide
LOX : Lipoxygénase
LPL : Lipase lipoprotéique
LPLAT : Lysophospholipide acyltransférase
Lp-PLA2 : PLA2 associée aux lipoprotéines
NADPH : Forme réduite du nicotinamide adénine dinucléotide phosphate
NO• : Oxyde nitrique
NO2• : Dioxyde d’azote
NOS : Synthétases d’oxyde nitrique
NOX : Nicotinamide adénine dinucléotide phosphate oxydase
O2 : Oxygène moléculaire
O2•- : Superoxyde
O3 : Ozone
OMS : Organisation mondiale de la santé
ONOO- : Peroxynitrite
PGF2α : Prostaglandine F2α
PL : Phospholipides
PLA2 : Phospholipase A2

XV
PUFA : Acide gras polyinsaturé; polyunsaturated fatty acid
R• : Radicaux alkyles
RIA : Méthodes radio-immunologiques
RO• : Radicaux alkoxyles
ROO• : Radicaux peroxyles
ROOH : Hydroperoxydes organiques
ROS : Dérivés actifs de l'oxygène; reactive oxygen species
RS• : Radicaux thiyle
RSH : Thiols
rTFA : Acides gras trans retrouvés dans les aliments dérivés des ruminants
SOD : Superoxyde dismutases
TAG : Triacylglycérol
TBA : Acide thiobarbiturique
TFA : Acide gras trans; trans fatty acid
TPR : Récepteur de la thromboxane A2
Trx : Thiorédoxine
TXA2 : Thromboxane A2
VLDL : Lipoprotéines de très basse densité; very low density lipoprotein
α-TO• : α-tocophérol oxydé
α-TOH : α-tocophérol
α-TTP : α-tocopherol transfer protein


XVII
«Tu ne traverseras jamais l’océan si tu as
peur de perdre de vue le rivage»
Christophe Colomb


XIX
Remerciements
Premièrement, j’aimerais remercier mon directeur de recherche, le Dr Jean-François Bilodeau, pour
son encadrement tout au long de ma maîtrise. Celui-ci m’a offert une occasion en or de découvrir ce
domaine passionnant qu’est la recherche dans le secteur médical. Il a également su faire preuve de
patience et de compréhension à mon égard puisque, étant chimiste, je n’étais pas très familière avec
les domaines de la physiologie et de l’endocrinologie. Malgré cela, le Dr Bilodeau a eu confiance en
mes capacités et m’a accordé beaucoup d’autonomie pour la réalisation de mon projet de maîtrise. Il
m’a également donné l’opportunité de présenter et d’assister à plusieurs congrès et conférences. J’ai
entre autres présenté mes résultats au Eastern Canadian Perinatal Investigators Meeting le 16
novembre 2012 à Toronto.
Ensuite, j’aimerais remercier la Dre Vanessa Moisan tant pour son support technique que moral. Son
aide a été particulièrement précieuse alors que j’avais de la difficulté à saisir certaines notions
enseignées pendant les cours que j’ai dû suivre dans le cadre de ma maîtrise et pour lesquels je
n’avais pas toutes les connaissances nécessaires pour comprendre.
Les conseils du Dr Pierre Julien ont été grandement appréciés. J’aimerais également remercier ce
dernier pour le dosage des lipides fait dans les échantillons sur lesquels j’ai travaillé ainsi que pour
les corrections apportées aux manuscrits des articles qui seront présentés dans ce mémoire.
Je suis reconnaissante envers le Fonds de recherche en santé du Québec (FRSQ) pour le support
financier accordé par l’entremise d’une bourse de formation de maîtrise ainsi qu’envers le Centre de
recherche en biologie de la reproduction (CRBR) pour la bourse À la poursuite de l’excellence.
Enfin, je tiens à remercier chaleureusement mes parents, Denise et Michel, mon conjoint, Sébastien,
ainsi que tous les autres membres de ma famille pour leur écoute et leurs encouragements. Leur
support a été très précieux tout au long de mes études.


XXI
Avant-Propos
Dans le cadre de ma maîtrise, je devais développer et valider une méthode d’analyse des F2-
isoprostanes, des biomarqueurs du stress oxydatif, par chromatographie liquide couplée à la
spectrométrie de masse en tandem (HPLC-MS/MS). Je devais ensuite utiliser cette technique pour
étudier le stress oxydatif dans la circulation maternelle en fin de grossesse. Le présent mémoire
résume mes travaux effectués au cours des deux dernières années. Il contient cinq chapitres et une
annexe. Voici une brève description de chacune de ces sections :
Le premier chapitre met en place toutes les notions théoriques nécessaires à la
compréhension du reste du document.
Le deuxième chapitre est le manuscrit d’un article scientifique qui a été accepté pour
publication dans le Journal of Lipid Research en février 2013. En tant que première auteure,
j’ai mis au point et validé la technique d’analyse des F2-isoprostanes par HPLC-MS/MS. J’ai
ensuite procédé au dosage de ces molécules dans le plasma de femmes au troisième
trimestre d’une grossesse normotensive avec l’aide de la Dre Vanessa Moisan. Enfin, j’ai
analysé les résultats et rédigé l’article sous la supervision de mon directeur de recherche, le
Dr Jean-François Bilodeau. Le manuscrit a été révisé avant sa soumission au journal par ce
dernier ainsi que par le Dr Pierre Julien.
Le troisième chapitre est le manuscrit d’un article scientifique dont je suis la première
auteure et qui sera soumis sous peu dans le journal Biochimica et Biophysica Acta. J’ai
utilisé la méthode que j’ai mise au point pour le dosage des F2-isoprostanes. La mesure des
acides gras a été faite par Karine Greffard qui est professionnelle de recherche dans le
laboratoire du Dr Pierre Julien. J’ai analysé les résultats et rédigé ce manuscrit sous la
supervision de mon directeur de recherche. Le manuscrit a été révisé par celui-ci et par le Dr
Pierre Julien.
Le quatrième chapitre résume les résultats que j’ai obtenus suite au dosage des F2-
isoprostanes dans le plasma, le sang entier et les membranes érythrocytaires de femmes au
troisième trimestre d’une grossesse normotensive ou prééclamptique. Ces résultats n’ont
pas fait l’objet d’un article scientifique pour le moment.

XXII
Dans le dernier chapitre du présent mémoire, les résultats présentés précédemment sont
discutés.
L’article présenté en annexe a été publié en 2012 dans le journal Molecular Reproduction &
Development. Il traite de la voie des polyols dans l’oviducte bovin. En tant que première
auteure, j’ai optimisé une technique d’analyse des polyols par HPLC-MS/MS et j’ai ensuite
procédé au dosage de ces molécules dans des sections d’oviductes bovin. Ces travaux ont
été faits lors d’un stage effectué dans le laboratoire du Dr Jean-François Bilodeau alors que
j’étais encore étudiante au baccalauréat, mais la rédaction de l’article à laquelle j’ai participé
a été faite pendant ma maîtrise.

1
Chapitre 1 – Introduction
1.1. Le stress oxydatif
Les dérivés actifs de l’oxygène (ROS) sont des molécules chimiquement réactives contenant au
moins un atome d’oxygène. Plusieurs d’entre eux sont radicalaires, c’est-à-dire qu’ils possèdent un
électron non apparié sur la couche électronique externe [1]. Dans les conditions physiologiques
normales, les ROS occupent des fonctions biologiques importantes. Entre autres, ils agissent comme
seconds messagers dans différentes voies de signalisation [2, 3] et sont impliqués dans la défense
immunitaire [4]. Les ROS sont constamment produits dans l’organisme, mais étant donné leur très
grande réactivité, leurs niveaux sont contrôlés par des antioxydants enzymatiques et non
enzymatiques. Un stress oxydatif survient lorsqu’il y a surproduction de ROS par rapport aux
défenses antioxydantes de l’organisme [5]. Ce déséquilibre est observé dans le processus du
vieillissement ainsi que dans certaines situations physiologiques normales comme la grossesse [6,
7]. Il est également impliqué dans la physiopathologie de plusieurs maladies [8]. Par exemple, le
stress oxydatif est observé dans plusieurs maladies cardiovasculaires (ex. : athérosclérose,
insuffisance cardiaque), inflammatoires (ex. : arthrite, asthme) et neurodégénératives (ex. : maladie
d'Alzheimer).
1.1.1. Les prooxydants
Parmi les ROS figurent le superoxyde (O2•-), le peroxyde d’hydrogène (H2O2), le radical hydroxyle
(HO•), radicaux alkoxyles (RO•), les radicaux peroxyles (ROO•), les hydroperoxydes (ROOH),
l’oxygène singulet (1O2) et l’ozone (O3) [9].
Le superoxyde est principalement produit dans les mitochondries où le transfert des électrons entre
les différents constituants de la chaîne de transport des électrons n’est pas complètement efficace
[10]. Une faible proportion des électrons qui circulent dans cette chaîne peuvent s’échapper par les
complexes I et III et causer la réduction directe de l’oxygène moléculaire pour former le superoxyde.
Entre 1 et 2 % de l’oxygène consommé chaque jour est ainsi réduit dans les conditions
physiologiques normales [11, 12]. Outre les mitochondries, le réticulum endoplasmique constitue une
autre source majeure de la production du superoxyde dans la cellule. En effet, jusqu’à 25 % du

2
superoxyde produit dans une cellule pourrait provenir de cet organite [13]. Pendant le repliement des
protéines, la formation de ponts disulfures est une réaction d’oxydation et les électrons libérés
peuvent être récupérés par l’oxygène moléculaire et ainsi le réduire en superoxyde. Certaines
enzymes, comme les cytochromes P450, la xanthine oxydase et les nicotinamide adénine
dinucléotide phosphate (NADPH) oxydases (NOX) sont également responsables de la production de
ce ROS. Les NOX catalysent la production de superoxyde à partir de l’oxygène et du NADPH [14].
Ces enzymes sont impliquées dans la signalisation cellulaire et l’une d’entre elles, NOX2, est
retrouvée dans les cellules phagocytaires qui produisent le superoxyde après exposition à des
microorganismes étrangers ou à des médiateurs de l’inflammation afin de détruire l’élément étranger.
Le superoxyde est une espèce radicalaire hautement réactive, mais à cause de sa charge anionique,
il lui est impossible de traverser les membranes lipidiques. Sa dismutation, catalysée par les
enzymes superoxyde dismutases (SOD), forme du peroxyde d’hydrogène (H2O2). Cet autre ROS
peut également être généré par les oxydases des peroxysomes, par la xanthine oxydase [15], par les
monoamines-oxydases [16] et par la p66shc dans les mitochondries [17]. Le peroxyde d’hydrogène
peut circuler dans l’organisme sur de plus longues distances, car il ne porte pas de charge anionique,
ce qui lui permet de traverser les membranes lipidiques. De plus, cet oxydant est beaucoup moins
réactif que le superoxyde, car il n’est pas radicalaire. Cependant, en oxydant le fer (II) non chélaté
selon la réaction de Fenton (1), il forme le radical hydroxyle, une espèce encore plus réactive que le
superoxyde et qui a la possibilité de traverser les membranes lipidiques. Le superoxyde peut quant à
lui réduire le fer (III) non chélaté en fer (II) (réaction (2) plus bas). En combinant les réactions (1) et
(2), la réaction de Haber-Weiss (3) est obtenue et celle-ci démontre que les ions de fer non chélatés
agissent comme des catalyseurs dans la production du radical hydroxyle. D’autres ions métalliques
non chélatés, comme le cuivre, peuvent également servir de catalyseur dans cette réaction [18].
(Réaction de Fenton) (1)
(2)
(Réaction de Haber-Weiss) (3)
H2O
2 + Fe
2+
O2
•- + Fe
3+
O2
•- + H
2O
2
HO• + HO
- + Fe
3+
O2 + Fe
2+
HO• + HO
- + O
2
Fe3+
, Fe2+

3
La réaction des ROS avec d’autres constituants biologiques peut causer la formation d'autres
molécules réactives. Par exemple, la réaction du radical hydroxyle avec d’autres molécules
biologiques mène à la formation d'autres ROS (RO•, R-O-O•, R-O-O-H) ou encore d'autres espèces
radicalaires (R•, RS•). Le superoxyde peut quant à lui réagir avec l’oxyde nitrique (NO•), une espèce
radicalaire synthétisée par les synthétases d’oxyde nitrique (NOS) à partir de la L-arginine (réaction
(4)). Cette réaction mène à la formation du peroxynitrite, un autre prooxydant qui peut se
décomposer en radical hydroxyle (réaction (5)) [19]. L’acide hypochloreux (HOCl) peut aussi être
formé lors de la réaction entre un chlorure (Cl-) et le peroxyde d’hydrogène [20]. Finalement, si la
production de ROS n’est pas suffisamment ralentie dans l’organisme par les antioxydants, ils
peuvent induire de sérieux dommages aux différents constituants cellulaires (ex. : protéines, ADN,
glucides et lipides). La réaction des ROS avec les lipides se nomme la peroxydation lipidique [21].
Les acides gras insaturés sont particulièrement sujets à réagir avec les ROS et d’autres radicaux
libres. Certains radicaux libres tels que les radicaux thiyles (RS•) ont la propriété de catalyser leur
isomérisation cis-trans [22]. La peroxydation lipidique et l’isomérisation cis-trans seront deux
réactions traitées plus en détail ultérieurement.
(4)
(5)
1.1.2. Les antioxydants
Les ROS causent des dommages importants aux différents constituants cellulaires, mais l’organisme
peut compter sur l’action des antioxydants enzymatiques et non enzymatiques.
1.1.2.1. Les antioxydants enzymatiques
Plusieurs enzymes sont impliquées à différents degrés dans les la défense de l’organisme contre
l’assaut des ROS [23]. En première ligne, les superoxyde dismutases (SOD) catalysent la
dismutation du superoxyde en peroxyde d’hydrogène selon la réaction (6). Ces enzymes existent
sous trois formes. La SOD1, ou CuZn-SOD, est retrouvée dans le cytosol alors que la SOD2, ou Mn-
SOD, est retrouvée dans les mitochondries. Il existe également une SOD extracellulaire, la SOD3.
ONOO-
ONOO- + H
+
NO• + O
2
•-
HO• +
•NO
2

4
Le peroxyde d’hydrogène peut quant à lui être décomposé par la catalase (CAT) qui est retrouvée
dans les peroxysomes (réaction (7)) ou encore par les glutathion peroxydases (GPx). Il existe cinq
GPx qui utilisent le glutathion (GSH) comme réducteur afin de décomposer le peroxyde d’hydrogène
et les hydroperoxydes organiques (réactions (8) et (9)) [24]. Cependant, ces cinq GPx ne possèdent
pas la même spécificité pour chacun des substrats. Ainsi, GPx1 et GPx2 peuvent réduire le peroxyde
d’hydrogène ainsi que les hydroperoxydes organiques solubles (ex. : hydroperoxydes d’acides gras).
GPx3 et GPx4 peuvent réduire des hydroperoxydes organiques plus complexes, mais GPx4 est le
seul enzyme pouvant utiliser les hydroperoxydes de phospholipides, de cholestérol et d'ester de
cholestérol encore intégrés dans les membranes lipidiques, et cela sans aide des phospholipases
[25]. GPx5, quant à elle, est seulement retrouvée dans l’épididyme. Après avoir été utilisé par les
GPx, le glutathion oxydé (GSSG) est réduit par une glutathion réductase (GPR) avec NADPH comme
agent réducteur (réaction (10)) pour compléter le cycle du glutathion (Figure 1). Une autre famille
d’enzyme utilisant le glutathion, les glutathion-S-transférases (GST), sont également capable
d’éliminer les ROS de la même manière que les GPx [26].
(6)
(7)
(8)
(9)
(10)
Figure 1. Cycle du glutathion.
2 O2
•- + 2 H
+
H2O + O
2
2 H2O + GSSG
H2O + GSSG + ROH
GSH + NADP+
SOD
CAT
GPx
GPx
GPR
O2 + H
2O
2
2 H2O
2
H2O
2 + 2 GSH
ROOH + 2 GSH
GSSG + NADPH
ROOH 2 GSH NADPH
GPx GPR
ROH + H2O GSSG NADP
+

5
Les enzymes appartenant au système de la thiorédoxine (Trx) sont aussi impliquées dans la
détoxication directe des ROS (Figure 2) [24]. Brièvement, les thiorédoxine peroxydases (TrxP), aussi
appelées peroxyrédoxines, peuvent décomposer le peroxyde d’hydrogène et les hydroperoxydes
organiques. La thiorédoxine peroxydase oxydée (oxTrxP) est recyclée par la thiorédoxine (Trx), une
autre enzyme capable de réduire les ROS. Il se crée alors des ponts disulfures entre deux cystéines
situées dans la portion N-terminale de l’enzyme oxydée (oxTrx) qui peuvent être réduits grâce à la
thiorédoxine réductase (TrxR) qui utilise le NADPH comme agent réducteur. Cette dernière enzyme
participe également dans la détoxication des ROS et elle est capable de réduire la vitamine C oxydée
ainsi que l’acide α-lipoïque [20]. À noter, aucune enzyme n’est capable de réduire le radical
hydroxyle, les radicaux alkoxyles (RO•) et les radicaux peroxyles (ROO•) car ceux-ci sont trop réactifs
et endommageraient la protéine impliquée dans leur détoxication. Le système enzymatique tente
plutôt d’empêcher la formation de ces espèces hautement réactives en éliminant leurs précurseurs :
les peroxydes et le superoxyde.
Figure 2. Système de la thiorédoxine. Adaptée de [24].
1.1.2.2. Les antioxydants non enzymatiques
Les enzymes antioxydantes utilisent principalement les peroxydes ou le superoxyde comme
substrats. L’organisme a également recourt à de nombreuses petites molécules hydrosolubles et
liposolubles capables de neutraliser les ROS, dont les ROS radicalaires. Par exemple, le glutathion,
l’acide urique et la vitamine C (acide ascorbique) sont des antioxydants hydrosolubles alors que
ROOH
Vitamine C oxydée ROOH
ROOH TrxP oxTrx TrxR NADPH
ROH + H2O oxTrxP Trx oxTrxR NADP
+
Acide α-lipoïque

6
l’ubiquinone (Coenzyme Q), l’acide α-lipoïque, les caroténoïdes (β-carotène, la vitamine A), et les
vitamines E sont des antioxydants liposolubles (Figure 3 et Figure 4).
Figure 3. Exemples de petites molécules antioxydantes.
Les vitamines E sont produites par les plantes et regroupent huit molécules de structures similaires,
c’est-à-dire quatre tocophérols (forme α-, β-, γ-, et δ-) et quatre tocotriénols (forme α-, β-, γ-, et δ-)
[27]. Ces molécules possèdent toutes un noyau chromanol et une longue chaîne latérale de 16
carbones (Figure 4). Pour les tocophérols, cette longue chaîne est saturée et compte trois centres
chiraux. Ces carbones asymétriques sont retrouvés seulement dans la configuration R dans la
nature. Pour les tocotriénols, cette chaîne contient trois liaisons doubles et le seul carbone

7
asymétrique est en configuration R dans la nature. Les huit types de vitamine E sont absorbés par
l’intestin, mais une fois acheminées dans le foie, la α-tocopherol transfer protein (α-TTP) favorise la
sécrétion de l’α-tocophérol (α-TOH) dans le plasma par le biais des lipoprotéines de très basse
densité (VLDL) et cela au détriment des autres formes de vitamines E [28]. Il en résulte que l’α-
tocophérol est la vitamine E la plus abondante dans les tissus et dans le plasma alors que les
tocotriénols sont les moins abondants [29]. C’est également la forme de tocophérol la plus efficace
comme antioxydant in vivo, suivi par les formes β-, γ-, et finalement δ- [27].
Figure 4. Structures des tocophérols et des tocotriénols.
À cause de sa nature hydrophobe, l’α-tocophérol est retrouvé dans les membranes lipidiques qu’il
protège contre la peroxydation. Cette molécule est capable de donner son hydrogène (H•) phénolique
aux différents ROS radicalaires (HO•, RO•, ROO•, O2-•) afin de les convertir en espèces moins
réactives (Figure 5). Elle empêche ainsi de façon très efficace la propagation de la réaction
radicalaire en chaîne qu’est la peroxydation lipidique. En effet, le radical peroxyle réagit 1000 fois
plus rapidement avec l’α-tocophérol qu’avec les acides gras polyinsaturés [30]. L’α-tocophérol
radicalaire (α-TO•) obtenu est moins réactif que les ROS, car le radical est délocalisé par résonnance
dans la molécule et donc moins accessible pour réagir. Cependant, ce radical peut être prooxydant
s’il n’est pas rapidement neutralisé [27]. Étant confiné dans les membranes lipidiques, il peut initier
de nouvelles réactions radicalaires (réactions (11) et (12)). L’α-tocophérol et l’α-tocophérol oxydé

8
peuvent également réduire les ions métalliques tels que le fer (III) (réactions (13) et (14)), favorisant
ainsi la réaction de Fenton (1).
(11)
(12)
(13)
(14)
Figure 5. Neutralisation du radical peroxyle en hydroperoxyde organique par l’α-tocophérol qui est à
son tour recyclée par la vitamine C (ascorbate). Adaptée de [31].
La vitamine C (ou ascorbate, AscH-) est une petite molécule hydrosoluble qui a la capacité de
neutraliser directement les ROS radicalaires (HO•, RO•, ROO•, O2-•). Cependant, sa principale
fonction comme antioxydant est de recycler l’α-tocophérol oxydé (Figure 5) [31]. Le radical ascorbyl
formé (Asc•-) peut ensuite être réduit soit par dismutation (réaction (15)), soit grâce à la thiorédoxine
réductase (Figure 2). Le dehydroacscorbate (DHAsc) formé lors de la dismutation de Asc•- peut être
réduit grâce à la DHAsc réductase dépendante du GSH (réaction (16)).
α-TO• + RH
α-TO• + ROOH
α-TOH + Fe3+
α-TO• + Fe
3+
α-TOH + R•
α-TOH + ROO•
α-TO• + Fe
2+ + H
+
α-TO+ + Fe
2+
ROOH ROO•
α-tocophérol
Radical ascorbyl Ascorbate

9
(15)
(16)
1.2. Les lipides
1.2.1. Définitions et fonctions biologiques
Les lipides sont de petites molécules hydrophobes ou amphiphiles divisées en huit catégories : les
acides gras, les acylglycérols, les glycérophospholipides (ou phospholipides), les sphingolipides, les
stérols (ex. : cholestérol, esters de cholestérols), les prénols, les glycolipides et les polycétides [32].
Ils occupent d’importantes fonctions biologiques [33]. Les phospholipides (PL) et le cholestérol sont
des constituants majeurs des membranes biologiques alors que les triacylglycérols (TAG) servent à
emmagasiner de l’énergie. En effet, l’oxydation complète de 1 g d’acides gras génère environ 38 kJ.
De plus, les cellules adipeuses sous-cutanées riches en TAG servent d’isolant thermique. Certains
lipides ou dérivés de lipides sont également impliqués dans la signalisation cellulaire.
1.2.2. Digestion, absorption et transport dans l’organisme
Les lipides représentent environ 35% de l’énergie quotidienne fournie par une diète américaine
typique [34]. Les TAG sont les principaux lipides ingérés. Les PL, le cholestérol et les esters de
cholestérol sont également retrouvés dans la diète, mais en quantité beaucoup plus faible. La
digestion et l’absorption des lipides se font dans l’intestin grêle où ils sont mélangés à la bile qui est
composée entre autres de sels biliaires, de phospholipides et de cholestérol [35]. Les TAG sont
hydrolysés en 2-monoacylglycérols et en acides gras grâce à la lipase pancréatique alors que les
phospholipides sont hydrolysés en 2-lysophospholipides et en acides gras grâce à une
phospholipase A2 (PLA2) [36, 37]. Les esters de cholestérols sont hydrolysés par la cholestérol
estérase [38-40]. Les acides gras, le 2-monoacylglycérol, le 2-lysophospholipide et le cholestérol
entrent ensuite dans les entérocytes principalement par diffusion passive où ils sont réestérifiés puis
assemblés pour former des chylomicrons, un type de lipoprotéine. Les lipoprotéines sont des
2 Asc•-
DHAsc
AcsH- + DHAsc
AscH-
DHAsc réductase
dépendante du GSH

10
particules sphériques dont la surface est composée d’apolipoprotéines, de PL et de cholestérol et
dont le cœur est composé de TAG et d’esters de cholestérol (Figure 6). Outre les chylomicrons (CM),
il existe trois autres types de lipoprotéines : les lipoprotéines de très basse densité (VLDL), les
lipoprotéines de basse densité (LDL) et les lipoprotéines de haute densité (HDL). Ces particules
diffèrent par leur densité, leur composition lipidique (Tableau 1) ainsi que par le type et la quantité
d’apolipoprotéines qui y sont associées (Figure 7).
Figure 6. Schémas général d'une lipoprotéine. Tirée de [41].
Tableau 1. Caractéristiques des différents types de lipoprotéines
CM VLDL LDL HDL
Densité (g/mL) < 0.94 0.94 - 1.006 1.006 - 1.063 1.063 - 1.210
Lipides (g/100 g lipoprotéine) 98 – 99 90 – 92 75 - 80 40 – 48
Acylglycérols (g/100 g lipides) 81 – 89 50 – 58 7 - 11 6 – 7
Esters de Cholestérol (g/100 g lipides) 2 – 4 15 – 23 47 - 51 24 – 45
Cholestérol (g/100 g lipides) 1 – 3 4 – 9 10 - 12 6 – 8
Phophoslipides (g/100 g lipides) 7 – 9 19 – 21 28 - 30 42 – 51
Adapté de [42]
La principale fonction des lipoprotéines est le transport des lipides dans l’organisme (Figure 7) [41,
43]. Le transport des TAG d’origine alimentaire vers les tissus est fait par les chylomicrons. Ceux-ci
sont formés dans les entérocytes puis sécrétés dans la lymphe avant de rejoindre la circulation
sanguine au niveau du canal thoracique. La lipase lipoprotéique (LPL) endothéliale hydrolyse les

11
TAG contenus dans ces particules en acides gras. Ceux-ci traversent ensuite l’endothélium et entrent
dans les cellules sous-jacentes où ils seront réestérifiés et entreposés sous forme de TAG ou encore
oxydés pour produire de l’énergie. Au cours de l’hydrolyse des TAG, la taille du chylomicron diminue
et le surplus de phospholipides, de cholestérol et de protéines retrouvé à sa surface est transféré aux
particules de HDL. Les résidus de chylomicrons sont captés et dégradés par le foie où les acides
gras obtenus sont soit oxydés, soit réorganisés en TAG, en PL et en esters de cholestérol pour être
sécrétés sous forme de VLDL. Les VLDL sont également constituées de lipides provenant des acides
gras emmagasinés dans les cellules adipeuses ou encore des acides gras synthétisés par le foie.
Tout comme les chylomicrons, ces lipoprotéines assurent le transport des TAG vers les tissus où ils
sont hydrolysés par la LPL alors que les PL, le cholestérol et certaines protéines membranaires sont
transférés aux HDL. Les VLDL résiduelles se nomment lipoprotéines de densité intermédiaire (IDL).
Certaines IDL sont éliminées par le foie alors que d’autres sont converties en LDL en interagissant
avec la lipase hépatique (HL), une enzyme capable d’hydrolyser les TAG et, à plus petite échelle, les
PL [44]. Les LDL sont des lipoprotéines enrichies en esters de cholestérol et assurent le transport de
ces lipides dans le sang vers les tissus jusqu’à ce qu’elles soient éliminées par le foie. Le transport
inverse du cholestérol des tissus vers le foie est quant à lui pris en charge par les HDL. Ces
dernières lipoprotéines sont assemblées dans les hépatocytes et les entérocytes et finissent leur
maturation dans le sang en captant le cholestérol non estérifié provenant des membranes cellulaires
et des autres lipoprotéines. Elles captent également les PL et les protéines larguées lors de la
lipolyse des chylomicrons et des VLDL. Le cholestérol situé à la surface des HDL est estérifié grâce
à la lécithin:cholestérol acyl transférase (LCAT) puis introduit à l’intérieur de la particule. Les esters
de cholestérols ainsi formés sont soit acheminés au foie ou transférés vers les autres lipoprotéines
grâce à la protéine de transfert des esters de cholestérol (CETP).

12
Figure 7. Transport des lipides par les lipoprotéines. Tirée de [45].
1.2.3. Acide arachidonique
Certains lipides et dérivés de lipides jouent un rôle dans la signalisation cellulaire. C’est le cas de
l’acide arachidonique (AA), un acide gras polyinsaturé (PUFA) oméga-6 de 20 carbones (20:4ω6)
précurseur d’un très grand nombre de molécules bioactives faisant partie de la famille des
eicosanoïdes (Figure 8) [46]. Ces molécules régulent de nombreux processus physiologiques et sont
impliquées de près dans l’inflammation [47]. Trois voies enzymatiques contribuent à la synthèse des
eicosanoïdes: celle des cyclooxygénases (COX), celle des lipoxygénases (LOX) et celle des
cytochromes P450 (CYP). Pour initier ces voies de biosynthèse, l’acide arachidonique doit d’abord
être libéré des phospholipides par une PLA2. Il existe également une voie non enzymatique menant à
la formation d'autres eicosanoïdes appelés les isoprostanes (isoPs). Cette dernière sera décrite plus
en détail ultérieurement. L’acide dihomo-gamma-linolénique (20:3ω6) et l’acide eicosapentaénoïque
(EPA, 20:5ω3) sont également des précurseurs des eicosanoïdes.

13
Figure 8. Voies de synthèse des eicosanoïdes dérivés de l’acide arachidonique. Tirée de [46].
Une partie de l’AA retrouvé dans les tissus et en circulation dans le sang est fournie directement par
la diète (entre 100 et 300 mg AA/jour) [48]. L’organisme peut également synthétiser cet acide gras à
partir de l’acide linoléique (18:2ω6, LA), un acide gras essentiel qui ne peut être synthétisé chez
l’humain et qui doit donc absolument être fourni par l’alimentation. Dans une diète occidentale, entre
10 et 20 g de LA est consommé chaque jour, ce qui représente plus de 85 % des acides gras
polyinsaturés ingérés [48, 49]. La conversion de cet acide gras en AA est faite dans plusieurs tissus,
mais principalement dans le foie grâce aux enzymes Δ6-désaturase, élongase et Δ5-désaturase
(Figure 9) [50]. Il a été estimé qu’environ 677 mg AA/jour peuvent être synthétisés de cette façon
lorsque 20 g LA/jour sont ingérés dans une diète américaine typique [51]. L’acide arachidonique
provenant de la diète ou synthétisé à partir de LA est ensuite acheminé aux tissus par le biais des
lipoprotéines. Pour parvenir aux tissus moins accessibles par ces dernières, il est transporté en étant
lié à l’albumine soit sous forme de 1-lyso-2-arachidonyl-phospholipide soit sous forme d’acide gras
non estérifié [48, 52].¸
La catégorie de lipides qui contient la plus forte proportion d’AA est celle des phospholipides [53]. En
effet, dans le plasma, l’AA représente 8.3 mol% des acides gras contenus dans les phospholipides
alors qu’il représente 5.1 mol% des acides gras estérifiés au cholestérol et 0.8 mol% des acides gras
dans les TAG. L’intégration de ce PUFA en position sn-2 des phospholipides se fait principalement
lors de leur remodelage (cycle de Lands) plutôt que lors de leur synthèse de novo (voie de Kennedy)

14
[54-56]. Pour ce faire, l’acide gras en position sn-2 d’un phospholipide quelconque doit préalablement
avoir été hydrolysé par une PLA2 avant que le 1-acyl-2-lyso-phospholipide obtenu soit réacétylé avec
l’AA estérifié à la coenzyme A (AA-CoA) grâce à une lysophospholipide acyltransférase (LPLAT).
Figure 9. Conversion de l'acide linoléique en acide arachidonique.
1.2.4. Acides gras trans
Les désaturases sont des enzymes qui insèrent des liaisons doubles dans la chaîne aliphatique des
acides gras. Chez l’humain, ces nouvelles liaisons doubles sont seulement retrouvées en
configuration cis. Des acides gras insaturés contenant au moins une liaison double en configuration
trans (acides gras trans, TFA) sont toutefois présents dans les différentes classes de lipides de
l’organisme. Ceux-ci proviennent majoritairement de la diète. Ils sont retrouvés naturellement dans
les produits laitiers et la viande provenant de ruminants. Ils y représentent 2 à 5 % de tous les acides
gras [57]. Ce sont des bactéries situées dans le rumen de ces animaux qui sont responsables de la
production des TFA lors de la biohydrogénation partielle des acides gras insaturés. Les TFA sont
également produits industriellement lors de l’hydrogénation partielle des huiles végétales. Avant les
années 2000, les TFA d’origine industrielle pouvaient représenter entre 40 et 65 % de tous les acides
gras contenus dans la margarine et le shortening [58, 59]. Le profil d’isomères de TFA retrouvé dans
les aliments dérivés des ruminants (rTFA) diffère beaucoup de celui retrouvé dans les huiles
végétales partiellement hydrogénées (iTFA). Dans les deux cas, les acides gras mono-insaturés
trans comptant dix-huit carbones sont les plus abondants [60]. Cependant, les rTFA sont
principalement composés d’acide trans-vaccénique (11t-18:1) alors que l’isomère le plus abondant
Acide linoléique
(18:2ω6, LA)
Acide γ-linoléique
(18:3ω6, GLA)
Δ6-désaturase
Acide arachidonique
(20:4ω6, AA)
Δ5-désaturase
Acide dihomo-γ-linoléique
(20:3ω6, DGLA)
Élongase

15
parmi les iTFA est l’acide élaïdique (9t-18:1), suivi de près par le 10t-18:1 et le 11t-18:1 (Figure 10)
[57, 60, 61].
Figure 10. Distribution des t-18:1 dans (A) l’huile végétale partiellement hydrogénée (margarine, n =
46 échantillons) et dans (B) le gras de lait de vache (n = 1765 échantillons). Adapté de [57].
Il a été démontré par de nombreuses études que la consommation d’une grande quantité de TFA
augmente les risques de maladies cardiovasculaires [62-68]. En effet, ils altèrent le profil lipidique du
sang (ex. : augmentation du cholestérol associé aux LDL et diminution du cholestérol associé aux
HDL, augmentation du rapport cholestérol total sur HDL-cholestérol) et modifient le niveau
d’apolipoprotéines. Ils provoquent également une réponse inflammatoire systémique ainsi que la
dysfonction de l’endothélium. Plusieurs études cliniques ont également démontré qu’une
consommation plus importante de TFA est associée à une augmentation du stress oxydatif [69-71].
Jusqu’à maintenant, les études portant sur l’impact de la consommation de TFA sur la santé n’ont
pas réussi à démontrer clairement si les rTFA (ex. : 11t-18:1) et les iTFA (ex. : 9t-18:1, 10t-18:1, 12t-
18:1) sont aussi nuisibles les uns que les autres. En fait, il semble plutôt que les effets néfastes
observés sur la santé sont plus liés à la quantité ingérée qu’à l’origine des TFA. Si les rTFA semblent
moins dommageables pour la santé, c’est qu’ils sont consommés en moins grande quantité que les
iTFA [62, 72, 73]. Cependant, les t-18:2 semblent plus nocifs pour la santé que les t-18:1 alors que
les t-16:1 semblent n’avoir aucun impact sur les facteurs de risque des maladies cardiovasculaires
[64, 68, 74].
Il a été estimé qu’au milieu des années 1990, les Canadiens consommaient en moyenne 8.4 g
TFA/jour, dont 81 % provenait des iTFA [58, 60]. À cause des préoccupations émises au sujet des
impacts néfastes de ces gras sur la santé, il est devenu obligatoire à partir de décembre 2005 au
B A

16
Canada d’inscrire la quantité de TFA contenu dans les aliments sur leur emballage [75]. L’étiquetage
nutritionnel obligatoire ainsi qu’une plus grande sensibilisation de la population canadienne ont fait
diminuer à 4.9 g/jour leur consommation moyenne de TFA en 2005 [58]. Cependant, cette valeur
était nettement supérieure à la quantité recommandée par l’Organisation mondiale de la santé
(OMS). Selon l’OMS, les gras trans ne doivent pas représenter plus de 1 % de l’apport énergétique
quotidien. C’est-à-dire que pour une diète de 2000 kCal/jour, la quantité maximale de TFA pouvant
être ingérée est de 2 g/jour. Afin d’atteindre cet objectif de consommation, un groupe d’étude sur les
gras trans a été créé en 2005 par Santé Canada. Celui-ci a remis en juin 2006 son rapport final dans
lequel il recommande de limiter la teneur totale en acides gras trans à 2 % de la teneur totale en
graisses pour les huiles végétales et les margarines molles et tartinables et de limiter cette teneur à 5
% pour tous les autres aliments, y compris les ingrédients vendus aux restaurants [75].
1.2.5. Acides gras et stress oxydatif
Les radicaux libres générés en période de stress oxydatifs sont hautement réactifs et ils peuvent
induire de sérieux dommages aux différents constituants cellulaires comme il a été expliqué
précédemment. Les acides gras insaturés sont particulièrement susceptibles à réagir avec eux. Deux
réactions sont possibles : la peroxydation lipidique et l’isomérisation cis-trans.
1.2.5.1. Peroxydation lipidique
La peroxydation lipidique est une réaction radicalaire en chaîne initiée lorsqu’un radical (HO•, RO•,
ROO•, ONOO-) vient arracher un hydrogène situé sur la chaîne aliphatique d’un acide gras ou d’un
ester d’acide gras (ex : TAG, PL, ester de cholestérol, etc) pour former un lipide radicalaire (L•)
(Figure 11) [1, 9, 21]. Les hydrogènes les plus susceptibles d’être arrachés sont les hydrogènes bis-
allyliques et allyliques puisque le nouveau radical obtenu est stabilisé par résonnance. L’énergie de
liaison d’un hydrogène allylique étant plus élevée de 10 kCal/mol en comparaison à celle d’un
hydrogène bis-allylique, ce dernier réagira donc plus rapidement [76]. D’ailleurs, plus le nombre
d’hydrogènes bis-allyliques est important dans un acide gras ou dans un ester d’acide gras, plus il
sera susceptible à être peroxydé [77]. Ainsi, la peroxydabilité d’acides gras communs décroit selon
l’ordre qui suit : acide docosahexaénoïque (DHA, 22:6ω3) > acide eicosapentaénoïque (EPA,
20:5ω3) > acide arachidonique (AA, 20:4ω6) > acide linoléique (LA, 18:2ω6). Le lipide radicalaire, L•,
se combine ensuite avec l’oxygène moléculaire (O2), un diradical, pour former un radical peroxyle de
lipide (LOO•) qui pourra à son tour arracher un hydrogène sur un autre acide gras ou ester d’acide

17
gras et ainsi propager la réaction radicalaire. La réaction est arrêtée grâce aux défenses
antioxydantes de l’organisme. Par exemple, l’α-tocophérol retrouvé dans les membranes lipidiques
est un inhibiteur de radicaux libres très efficace. En effet, comme mentionné précédemment, LOO•
réagit 1000 fois plus rapidement avec l’α-tocophérol qu’avec les acides gras polyinsaturés qui
l’entoure [30].
La peroxydation lipidique génère un vaste éventail de produits pouvant perturber l’organisation des
membranes lipidiques, leur fluidité ainsi que leur perméabilité. En effet, L• et LOO• peuvent se
réarranger, se fragmenter, se réduire, se cycliser ou encore réagir avec d’autres molécules de leur
environnement (ex. : protéines). Parmi les produits issus de la peroxydation lipidique figurent de
nombreux hydroperoxydes et aldéhydes (ex. : malondialdehyde (MDA)) [1, 9]. La cyclisation est un
processus particulièrement important pour les LOO• comptant trois doubles liaisons et plus [21].
Ainsi, la peroxydation de l’acide arachidonique peut mener à une série de produits cycliques
semblables aux prostaglandines, les isoprostanes (isoPs, Figure 12) et d’isofuranes [9].
Figure 11. Mécanisme général de la peroxydation des lipides.

18
Figure 12. Structure des différents types d'isoprostanes dérivés de l'acide arachidonique.
Tirée de [9].
1.2.5.2. Isomérisation cis-trans
Les TFA retrouvés dans les différentes catégories de lipides proviennent principalement de la diète.
Cependant, il a également été démontré qu’ils peuvent être générés in vivo en présence de stress
oxydatif. Ce phénomène a, entre autres, été observé dans les membranes érythrocytaires et les reins
de rats âgés nourris avec des aliments ne contenant pas de TFA et exposés au tétrachlorométhane
(CCl4), un inducteur de stress oxydatif [78]. En fait, certains radicaux libres catalysent, selon un
mécanisme d’addition-élimination, l’isomérisation d’acides gras cis vers la configuration trans qui est
thermodynamiquement plus stable (Figure 13). Plusieurs radicaux sont capables de réagir de cette
façon et les plus importants retrouvés in vivo sont les radicaux thiyles (RS•) et le dioxyde d’azote
(NO2•) [78-80]. Par contre, selon les données cinétiques disponibles, les RS• semblent plus efficaces
que le NO2• pour isomériser les acides gras insaturés [79]. Les RS• sont produits en période de
stress oxydatif à partir des thiols (RSH) retrouvés en abondance dans l’organisme (ex. : glutathion).
Ces derniers sont des inhibiteurs de la peroxydation lipidique à cause de leur facilité à donner un
hydrogène (H•) [22]. NO2• provient quant à lui de sources exogènes (ex. : fumée de cigarette, smog)

19
et de sources endogènes (ex. : oxydation de NO•, oxydation des nitrites (NO2-), décomposition du
peroxynitrite selon la réaction (5) à la page 3) [81]. Outre l’isomérisation cis-trans, ce radical est
également capable de causer la peroxydation et la nitration des lipides [81]. La réaction favorisée
dépend beaucoup des conditions du milieu.
Figure 13. Mécanisme général de la réaction d'isomérisation cis-trans induite par les radicaux libres.
X• représente n’importe quel radical capable d’induire cette réaction. Tirée de [22].
Un peu comme pour la peroxydation lipidique, plus le nombre d’insaturations est important dans un
acide gras et plus celui-ci sera facilement isomérisé [82]. Les TFA obtenus rigidifient les membranes
lipidiques et diminuent leur perméabilité. Ils peuvent également modifier le métabolisme des lipides et
influencer différentes réactions enzymatiques [79]. Afin de limiter ces effets néfastes, l’organisme
peut compter sur des inhibiteurs d’isomérisation tels que la vitamine A, les caroténoïdes et l’α-
tocophérol. Ce dernier est cependant moins efficace que les deux autres [83].
1.3. Les F2-isoprostanes
Afin d’étudier le rôle des ROS dans la physiopathologie de plusieurs maladies, l’utilisation de
biomarqueurs est essentielle. En effet, mesurer directement les ROS in vivo est impensable à cause
de leur haute réactivité et de l’instrumentation inadaptée. Les lipides sont des cibles faciles pour ces
petites molécules et c’est donc pourquoi différents produits issus de la peroxydation lipidique sont
utilisés comme marqueur du stress oxydatif. Pendant longtemps, la méthode la plus utilisée a été le
dosage du malondialdehyde (MDA) à l’aide de l’acide thiobarbiturique (TBA). Cependant, malgré sa
simplicité, cette technique manque de spécificité pour le MDA puisque le TBA peut réagir avec
d’autres composés dont plusieurs sont générés lors du traitement des échantillons, menant ainsi à
des résultats erronés [84-86]. En 1990, Morrow et son équipe démontrent que la peroxydation non
enzymatique de l’acide arachidonique mène à la formation in vitro et in vivo de composés analogues
à la prostaglandine F2α (PGF2α) (Figure 14) [87]. Ces composés, nommés les F2-isoprostanes (F2-

20
isoPs), sont des biomarqueurs très fiables du stress oxydatif comme l’a mis en évidence une étude
réalisée en 2005 au cours de laquelle différents biomarqueurs potentiels ont été mesurés dans le
plasma et l’urine de rats exposés au tétrachlorométhane (CCl4) [88]. En effet, les F2-isoPs possèdent
de nombreuses qualités leur permettant de jouer ce rôle. Ils sont produits spécifiquement par la
peroxydation lipidique, ils sont stables chimiquement et détectables dans tous les tissus et fluides
biologiques même en condition non pathologique. De plus, leur concentration augmente en réponse
à un stress oxydatif [89]. L’isomère de F2-isoPs le plus utilisé jusqu’à maintenant comme
biomarqueur du stress oxydatif est le 8-iso-PGF2α (Figure 14) [8].
Figure 14. Structures de la Prostaglandine F2α (PGF2α) et du 8-iso PGF2α.
1.3.1. Formation des F2-isoprostanes
Lors de l’initiation de la biosynthèse de PGF2α par la voie des cyclooxygénases (COX), l’acide
arachidonique est d’abord libéré des phospholipides par une PLA2. Bien que les F2-isoPs possèdent
une structure très similaire à ce médiateur lipidique et qu’ils dérivent eux aussi de l’acide
arachidonique, les étapes menant à leur synthèse in vivo sont très différentes et n’impliquent pas
d’enzymes (Figure 15). Premièrement, la synthèse de ces composés se fait in situ sur les
phospholipides et débute par l’abstraction d’un des hydrogènes bis-allyliques de l’acide
arachidonique par un ROS [90]. La probabilité d’abstraction de ces hydrogènes situés sur les
carbones C7, C10 ou C13 est à peu près la même [9]. Selon l’hydrogène arraché, un radical peroxyle
(ROO•) conjugué est ensuite formé par l’addition d’une molécule d’oxygène sur l’un des carbones
suivants : C5, C8, C9, C11, C12 ou C15. L’oxygène peut également s’additionner sur les carbones
C7, C10 ou C13 et ainsi former un ROO• non conjugué, mais ce composé est instable. En effet, la
constante de vitesse pour l’élimination de la molécule d’oxygène nouvellement additionné (kβ = 1.9 x
PGF2α
8-iso-PGF2α

21
106 s-1) est beaucoup plus grande que celle mesurée dans le cas des ROO• conjugués (kβ = 70 s-1)
[89]. L’étape suivante de la formation des F2-isoPs consiste en une cyclisation 5-exo du ROO•
conjugué alors qu’une deuxième molécule d’oxygène est additionnée pour ultimement mener à la
formation d’un bicycloendoperoxyde (Figure 15, avant dernière rangée de molécules). Dans le cas
des ROO• où l’oxygène est situé sur les carbones C5 ou C15, le mécanisme de cyclisation est un
peu plus complexe, car ces derniers ne peuvent pas cycliser directement. L’oxygène doit d'abord se
détacher de la molécule puis se réadditionner sur les carbones C9 ou C11, respectivement, pour que
la cyclisation 5-exo soit possible [91]. Finalement, le bicycloendoperoxyde est réduit et un F2-isoPs
est obtenu.
Figure 15. Mécanisme de formation des F2-isoprostanes. Adaptée de [92].
ROS ROS
ROS
O2 O
2 O
2 O
2
O2 O
2 O
2 O
2
7
13 10
5 9 8 12
15 11
5
12 8
15
Acide arachidonique

22
La peroxydation de l’acide arachidonique peut générer 64 isomères de F2-isoPs subdivisés en quatre
classes de régioisomères qui contiennent chacune 8 diastéréoisomères racémiques [89]. Les
isomères de classe IV et V sont les moins abondants de tous, car leur précurseur
bicycloendoperoxyle peut s’oxyder davantage et mener à la formation de peroxydes dioxolane-
isoprostane (Figure 16) [93]. Ceux de classe VI sont les plus abondants, suivis par ceux de classe III
[94]. Parmi les isomères de classe III figurent le populaire 8-iso-PGF2α et la PGF2α d’origine non
enzymatique [91]. Par contre, la formation de la PGF2α n’est pas favorisée, car les chaînes latérales
de cet isomère sont en configuration trans par rapport au cyclopentane alors que c’est la
configuration cis qui est privilégiée lors de la synthèse des F2-isoPs [91]. Malgré cela, la PGF2α
retrouvée chez l’humain provient d’avantage de la peroxydation lipidique que de la synthèse
enzymatique par le biais des COX [9].
Figure 16. Structure d’un peroxyde dioxolane-isoprostane (classe IV). R = -(CH2)3COOH.
1.3.2. Transport et métabolisme des F2-isoprostanes
Les F2-isoPs sont générés in situ en position sn-2 sur les phospholipides (PL) et peuvent être
hydrolysés par l’action de certaines PLA2 dont la PLA2 associée aux lipoprotéines (Lp-PLA2) [90].
D’ailleurs, les PLA2 retrouvées dans le venin de serpent préfèrent cliver les lipides oxydés,
protégeant ainsi les membranes lipidiques des dommages causés par la peroxydation lipidique [95,
96]. Malgré cela, les F2-isoPs circulent principalement estérifiés au PL des HDL dans le plasma [97].
En effet, ils sont quatre fois plus abondants sous forme estérifiée au PL des lipoprotéines que sous
forme libre dans le plasma [98]. La Lp-PLA2 est plus active en présence des LDL, ce qui fait de ces
lipoprotéines de moins bons transporteurs d’isoprostanes [97].
La demi-vie du 8-iso-PGF2α libre, le F2-isoPs le plus étudié jusqu’à maintenant, est d’environ 16
minutes chez l’humain [8]. Ce composé, tout comme les autres isomères formés lors de la
peroxydation de l’acide arachidonique, est filtré tel quel par les reins et excrété dans l’urine où il est

23
retrouvé en très grande quantité. Il peut également être métabolisé par le foie. Les principaux
métabolites du 8-iso-PGF2α formés sont le 2,3-dinor-8-iso-PGF2α et le 2,3-dinor-5,6-dihydro-8-iso-
PGF2α (Figure 17) [99]. Ces derniers composés sont également retrouvés en très grande quantité
dans l’urine.
Figure 17. Structures du 2,3-dinor-8-iso-PGF2α et le 2,3-dinor-5,6-dihydro-8-iso-PGF2α.
1.3.3. Mesure des F2-isoprostanes
Les F2-isoPs sont retrouvés dans tous les tissus et fluides biologiques. Le plasma et l’urine sont les
deux matrices biologiques les plus utilisées pour évaluer le stress oxydatif in vivo, car leur collection
est facile et peu ou pas invasive. Certaines précautions doivent être prises lors de l’entreposage et
de l’analyse des échantillons de plasma afin d’éviter la peroxydation ex vivo des lipides qu’ils
contiennent [98]. En effet, ces échantillons doivent être conservés à -80°C en présence d’un
antioxydant comme le hydroxytoluène butylé (BHT) [100]. L’ajout de BHT n’est pas nécessaire pour
les échantillons d’urine, car ce liquide biologique ne contient qu’une quantité négligeable lipides.
Dans le plasma, les F2-isoPs sont principalement retrouvés estérifiés aux PL et en une plus faible
proportion sous la forme libre [98]. Afin d’évaluer le stress oxydatif, les F2-isoPs libres peuvent être
mesurés, mais aussi les F2-isoPs totaux, c’est-à-dire la somme de ces composés libres et estérifiés.
Dans ce cas, les F2-isoPs doivent être hydrolysés des PL soit par hydrolyse alcaline, soit par
hydrolyse enzymatique à l’aide d’une PLA2 [101]. Dans l’urine, une telle hydrolyse n’est pas
nécessaire. Par contre, en plus des F2-isoPs, mieux vaut y mesurer aussi leurs métabolites dans le
but d’évaluer la peroxydation lipidique endogène réelle puisque cette information peut être faussée
par la production locale de F2-isoPs dans les reins [102]. Les concentrations mesurées sont
normalisées en fonction de la quantité de créatinine contenue dans l’urine.
2,3-dinor-8-iso-PGF2α
2,3-dinor-5,6-dihydro-8-iso-PGF2α

24
De nombreuses méthodes ont été mises au point pour mesurer les F2-isoPs dans les différentes
matrices biologiques. La première technique à avoir été développée utilise la chromatographie en
phase gazeuse couplée à la spectrométrie de masse en mode « ionisation chimique négative » (GC-
NICI-MS) [101, 103]. Pour être mesurés par cette technique, les F2-isoPs estérifiés sont hydrolysés
des PL puis extraits de la matrice biologique avant d’être soumis à une dérivation chimique afin de
les rendre compatibles avec la chromatographie gazeuse (Figure 18). Cette méthode très sensible
mesure les F2-isoPs, c’est-à-dire le 8-iso-PGF2α et les autres F2-isoPs qui coéluent avec ce composé
[104]. L’utilisation de la chromatographie en phase gazeuse couplée à la spectrométrie de masse en
tandem (GC-MS/MS) est possible afin d’améliorer la sélectivité de la méthode [105, 106]. Dans le but
de simplifier la préparation des échantillons, des méthodes utilisant la chromatographie liquide
couplée à la spectrométrie de masse en tandem (HPLC-MS/MS) ont été mises au point. Ces
méthodes ne nécessitent pas la dérivation chimique des F2-isoPs. De plus, elles permettent de doser
simultanément les quatre classes de regioisomères et de doser séparément plusieurs
diastéréoisomères dans une même classe [107, 108]. Finalement, des méthodes immuno-
enzymatiques (ELISA) [109] et radio-immunologiques (RIA) ont également été mises au point [110].
Ces méthodes ne nécessitent pas l’utilisation d’instruments dispendieux comme les GC-MS et
HPLC-MS/MS, mais elles sont moins spécifiques à cause des réactions croisées de l’anticorps avec
d’autres eicosanoïdes apparentés.
Figure 18. Dérivation chimique pour l'analyse des F2-isoPs par GC-MS. Tirée de [104].

25
1.3.4. Effets biologiques des F2-isoprostanes
Les F2-isoPs ne sont pas que des biomarqueurs du stress oxydatif, ce sont également des molécules
bioactives. La plupart des études en lien avec la bioactivité des F2-isoPs ont été faites à l’aide du 8-
iso-PGF2α. Il a été démontré que ce composé est vasoconstricteur dans la plupart des tissus, dont
l’aorte, le cerveau, les reins et le poumon [111-113]. De plus, il induit la prolifération des cellules
musculaires lisses [114]. Il exerce ces actions biologiques principalement en interagissant avec le
récepteur du thromboxane A2 (TPR). 8-iso-PGF2α est également un médiateur dans la fonction
plaquettaire [115]. Cette molécule stimule l’agrégation des plaquettes en interagissant avec le
récepteur TP mais il peut également inhiber cette agrégation en interagissant avec un récepteur
inconnu (ISR, récepteur des isoprostanes) jusqu’à maintenant qui implique l’adénosine
monophosphate cyclique (AMPc) (Figure 19) [115, 116]. Enfin, le 8-iso-PGF2α possède d’autres
activités biologiques et entre autres, il peut induire la synthèse de TXA2 dans certains lits vasculaires
tels que les microvaisseaux du cerveau de porc [117].
Figure 19. Signalisation impliquant le 8-iso-PGF2α dans la fonction plaquettaire. Tirée de [115].

26
Les études concernant l’activité biologique des autres F2-isoPs sont limitées. Une étude faite chez le
porc sur des microvaisseaux cérébraux et rétiniens a démontré que d’autres isomères de la classe
III, c’est-à-dire 8-iso-15(R)-PGF2α, ent-8-iso-PGF2α et ent-8-iso-15(S)-PGF2α, sont également des
vasoconstricteurs comme le 8-iso-PGF2α. Il en va de même pour les deux isomères de la classe V
qui ont été testés. Cependant, deux des trois isomères de la classe VI testés (iPF2α-VI et 5-iPF2α-VI)
ne possèdent pas d’effets vasomoteurs alors que le ent-iPF2α-VI cause une faible vasoconstriction.
Ces résultats confirment ce qui avait déjà été démontré dans des vaisseaux sanguins humains
concernant le iPF2α-VI et le 5-iPF2α-VI [118].
1.4. Grossesse et désordre hypertensif avec protéinurie
1.4.1. La grossesse, un évènement oxydatif
Le placenta est un organe hémochorial qui se développe pendant la grossesse et qui lie la mère à
son enfant par le biais du cordon ombilical. Il occupe plusieurs fonctions essentielles au bon
développement du fœtus [119]. Premièrement, il assure le transport de l’oxygène, de l’eau et des
nutriments de la mère vers le fœtus et expulse le dioxyde de carbone de même que les autres
déchets métaboliques vers la circulation maternelle. Ensuite, il protège le fœtus contre les infections
et maladies maternelles ainsi que contre certains xénobiotiques. Enfin, il occupe également une
fonction endocrine puisqu’il relâche autant dans la circulation sanguine maternelle que fœtale
différents types d’hormones dont des œstrogènes, de la progestérone, des facteurs de croissance,
des cytokines et des eicosanoïdes.
Le développement du placenta débute 6 à 7 jours après la fécondation de l’ovule [120]. Lors de la
placentation, les cytotrophoblastes d’origine fœtales envahissent les artères spiralées utérines où ils
remplacent les cellules endothéliales maternelles et détruisent les cellules musculaires lisses sous-
jacentes jusqu’au premier tiers du myomètre (Figure 20) [120, 121]. Ces artères ainsi remodelées ont
un diamètre au moins quatre fois plus large qu’initialement et ne répondent plus aux stimuli
vasomoteurs [121]. Ces nouvelles caractéristiques permettent au placenta de recevoir, à partir du
deuxième trimestre de la grossesse, un haut débit sanguin non pulsé et de faible pression [7, 122].
Pendant le premier trimestre, les artères spiralées sont obstruées par un bouchon de

27
cytotrophoblastes et le placenta se développe dans un environnement pauvre en oxygène [7]. Dix à
douze semaines après le début de la grossesse, le sang maternel commence à circuler dans le
placenta à partir de la périphérie de celui-ci et la pression partielle en oxygène y passe de moins de
20 mm Hg (8e semaine) à plus de 50 mm Hg (12e semaine) [7, 123]. Cet évènement est à l’origine
d’un choc oxydatif [124, 125]. Ensuite, plus la grossesse avance plus la mère est soumise à un
stress oxydatif placentaire et systémique [126, 127]. En effet, afin de soutenir la croissance rapide du
fœtus, le métabolisme du placenta nécessite beaucoup d’oxygène et par conséquent ce tissu est
riche en mitochondries, une source majeure de ROS [125]. La grossesse est également associée à
une réponse inflammatoire systémique qui se manifeste principalement par le biais de l’activation de
monocytes et de granulocytes ainsi que de l’endothélium [128]. L’activation de ces cellules mène à la
production de ROS pouvant contribuer au stress oxydatif maternel. Le stress oxydatif atteint son
paroxysme au moment de l’accouchement vaginal alors que les contractions de l’utérus coupent par
intermittence l’apport en sang du placenta (ischémie-réperfusion). Ce phénomène amplifie la
production de ROS par les mitochondries et les xanthines oxydases [123, 129].
Figure 20. Invasion des artères spiralées par les cytotrophoblastes dans une grossesse sans
complications. Tirée de [130].
1.4.2. Désordre hypertensif avec protéinurie (prééclampsie)
La prééclampsie est un syndrome pouvant être observé après la 20e semaine de grossesse chez
l’humain et qui est associé à une tension artérielle élevée (pression diastolique > 90 mm Hg) et à une

28
protéinurie (plus de 0.3 g de protéines par jour dans un échantillon d’urine collecté sur 24h) [131]. Le
seul traitement efficace est l’accouchement [132]. Elle complique entre 3 et 5 % des grossesses dans
le monde et est une cause majeure de mortalité et de morbidité maternelle et fœtale [133]. Parmi les
complications maternelles probables de la prééclampsie figurent l’éclampsie, le syndrôme HELLP
(Hemolysis, Elevated Liver enzymes and Low Platelets count), l’insuffisance rénale, l’œdème
pulmonaire, l'hématome rétroplacentaire et la mort [132]. Aux États-Unis, 20 % des décès reliés à la
grossesse sont dus à ce syndrome [134]. De plus, les femmes qui ont souffert de prééclampsie ont
plus de chances de développer des maladies cardiovasculaires plus tard dans leur vie [135, 136].
Parmi les complications fœtales probables figurent la restriction de croissance intra-utérine,
l’asphyxie périnatale et la mort [132]. Comme le seul traitement à la prééclampsie est
l’accouchement, ce syndrome est la cause de nombreuses naissances prématurées. Selon le degré
de prématurité, les conséquences pour le bébé peuvent être importantes. Finalement, un bébé né
d’une grossesse prééclamptique est plus à risque de développer certaines maladies à l’âge adulte,
par exemple des maladies cardiovasculaires (hypothèse de Barker) [132, 137].
1.4.2.1. Physiopathologie de la prééclampsie
Les causes exactes de la prééclampsie sont inconnues. Toutefois, il est possible d’expliquer la
physiopathologie de ce syndrome grâce à un modèle en deux étapes.
Au cours de la première étape, qui se déroule avant la 20e semaine de grossesse, la placentation est
inadéquate. L’invasion des artères spiralées par les cytotrophoblastes est limitée à la décidue (Figure
21) [138]. De plus, 30 à 50 % des artères échappent totalement au remodelage imposé par la
grossesse [139]. Ces vaisseaux sanguins conservent donc leur petit diamètre et leur rigidité. Ils
demeurent également sensibles aux molécules vasoactives. Le débit sanguin résultant est par
conséquent irrégulier (ischémie-réperfusion), pulsé et de forte pression. La pression élevée du débit
sanguin est susceptible de causer des dommages hydrostatiques aux villosités placentaires alors
que l’ischémie-réperfusion est responsable de la production accrue de ROS [121, 122]. En fait, la
prééclampsie est associée à un stress oxydatif et à une réponse inflammatoire systémique plus
importants que ce qui est observé au cours d’une grossesse normale [140, 141].

29
Figure 21. Invasion des artères spiralées par les cytotrophoblastes dans une grossesse
prééclamptique. Tirée de [130].
Le stress oxydatif joue un rôle important dans la transition vers la deuxième étape de la
physiopathologie de la prééclampsie, c’est-à-dire vers l’apparition des symptômes maternels. Ces
symptômes sont principalement associés à la dysfonction de l’endothélium [142-145]. Dans ses
conditions non pathologiques, l’endothélium agit entre autres comme une barrière sélective pour
empêcher la diffusion de macromolécules de la lumière artérielle vers l'espace interstitiel. Ce tissu
joue également un rôle dans la régulation du tonus vasculaire, dans la modulation de l'inflammation,
dans la promotion, ainsi que dans l'inhibition de la croissance vasculaire et dans la modulation de
l'agrégation plaquettaire et dans la coagulation [146]. Les ROS peuvent directement induire la
dysfonction de l'endothélium et ainsi causer l’hypertension et la protéinurie caractéristiques à la
prééclampsie [146]. Ils peuvent également provoquer la dysfonction de l’endothélium indirectement
en favorisant l’inflammation systémique observée en prééclampsie. En effet, les ROS peuvent
favoriser l’inflammation de diverses façons notamment en activant NF-κB, en produisant des
biomolécules oxydées pro-inflammatoires (ex. : LDL oxydées) ou encore en provoquant la nécrose
des cellules [2, 147]. De plus, les ROS activent les leucocytes [148] ainsi que de la production de
cytokines inflammatoires (ex. : Tumor Necrosis Factor-α (TNF-α), interleukine-1β (IL-1β), IL-6) en
prééclampsie [2, 130, 149-152]. Ces cytokines sont susceptibles de causer la dysfonction de
l’endothélium, tout comme les facteurs antiangiogéniques (ex. : soluble vascular endothelial growth
factor receptor-1 (sFlt-1) and soluble endoglin) libérés par le placenta en ischémie-réperfusion [130,
153].

30
1.4.2.2. Facteurs de risque
De nombreux facteurs de risques sont connus pour la prééclampsie. Ils sont résumés dans les
tableaux suivants (Tableau 2, Tableau 3, Tableau 4). Ils peuvent être séparés en trois catégories :
ceux liés à la mère, ceux liés au père et ceux liés à la grossesse. Outre ces facteurs, le statut
socioéconomique de la mère peut également influencer ses chances de développer ce syndrome au
cours d’une grossesse. En effet, plus son statut socioéconomique est faible, plus le risque est élevé
[154]. Il semble que la diète, le mode de vie et l’environnement contribuent à l’augmentation des cas
de prééclampsie dans les pays pauvres [155]. Curieusement, les fumeuses ont 30 à 40 % moins de
chances d’avoir une grossesse prééclamptique [156, 157]. Par contre, ce bénéfice est annulé par
l'effet négatif substantiel du tabagisme sur la croissance du fœtus, le risque de décollement
placentaire, et la santé en générale. [158].
Tableau 2. Facteurs de risques pour la prééclampsie liés à la mère
Facteurs liés à la mère Risque relatif
Réf.
Âge maternel > 40 ans (primipare) 1.681 [159]
Âge maternel > 40 ans (multipare) 1.961 [159]
Nulliparité 2.911 [159]
Prééclampsie lors d’une grossesse précédente 7.191 [159]
Intervalle entre les grossesses 1.12/année2 [160]
Antécédents familiaux de prééclampsie 2.901 [159]
Hypertension artérielle préexistante (diastolique > 80 mm Hg) 1.381 [159]
Diabète de type 1 3.561 [159]
Obésité 2.471 [159]
Thrombophilie n.d. [161]
Maladies rénales n.d. [159]
Maladies auto-immunes chroniques 6.91 [159]
Syndrome des antiphospholipides 9.721 [159]
n.d. : non disponible 1Risque relatif non-ajusté (intervalle de confiance à 95%) 2Rapport des cotes (odds ratio) (intervalle de confiance à 95%)

31
Tableau 3. Facteurs de risques pour la prééclampsie liés au père
Facteurs liés au père Risque relatif
Réf.
Exposition limité au sperme (0 à 4 mois) 7.81 [162]
Exposition limité au sperme (5 à 8 mois) 4.51 [162]
Exposition limité au sperme (9 à 12 mois) 2.91 [162]
Exposition limité au sperme (12 mois et plus) 1.01 [162]
Père impliqué dans une grossesse prééclamptique antérieure 1.82 [163] 1Risque relatif non-ajusté (intervalle de confiance à 95%) 2Odds ratio non-ajusté (intervalle de confiance à 95%)
Tableau 4. Facteurs de risques pour la prééclampsie liés à la grossesse
Facteurs liés à la grossesse Risque relatif
Réf.
Diabète gestationnel n.d. [158]
Grossesse gémellaire 2.931 [159]
Mole hydatiforme n.d. [164]
Anomalies chromosomiques n.d. [158]
n.d. : non disponible 1Risque relatif non-ajusté (intervalle de confiance à 95%)
1.5. Problématique et objectifs
1.5.1. Problématique
Le stress oxydatif est impliqué dans la grossesse en tant qu’événement physiologique normal [165].
Cependant, la prééclampsie, une pathologie de la grossesse, est associée à un stress oxydatif plus
important que celui qui est observé au cours d’une grossesse dite « normale » [140].
Plusieurs biomarqueurs ont été utilisés afin d’évaluer le stress oxydatif in vivo. Depuis quelques
années, les F2-isoprostanes, des produits de la peroxydation de l’acide arachidonique, sont reconnus
comme étant les biomarqueurs les plus fiables du stress oxydatif [88]. Bien qu’il existe 64 isomères
de F2-isoprostanes, le plus étudié est un isomère de la classe III, le 8-iso-PGF2α (Figure 14) [8]. Cet

32
isomère a été mesuré dans plusieurs matrices biologiques (placenta, plasma, urine, salive) afin
d’étudier le stress oxydatif en prééclampsie [166-178]. Certaines études ont démontré une
augmentation du niveau de 8-iso-PGF2α en prééclampsie [166, 171], d’autres non [169, 179]. Une
seule étude s’est attardée à mesurer un autre F2-isoprostane que le 8-iso-PGF2α. En effet, Regan et
al. ont mesuré le 8,12-iso-iPF2α-VI dans des échantillons d’urine de femmes prééclamptiques et
normales obtenus à différents moments de la grossesse, mais ils n’ont pas trouvés de différences
significatives entre les deux groupes [180]. Aucune étude ne s’est penchée sur la mesure d’autres
isomères que le 8-iso-PGF2α dans le sang, le plasma, et les membranes érythrocytaires de femmes
au troisième trimestre d’une grossesse normale ou prééclamptique.
1.5.2. Hypothèses
Les F2-isoprostanes de la classe VI sont généralement les plus abondants de tous les F2-
isoprostanes. Ceux-ci pourraient être de meilleurs biomarqueurs du stress oxydatif durant la
grossesse sans complications et en prééclampsie que le 8-iso-PGF2α.
Les acides gras trans peuvent provoquer un stress oxydatif in vivo et le stress oxydatif peut
induire la formation d’acides gras trans. Des corrélations positives entre les F2-isoprostanes
et les acides gras trans pourraient donc être observées dans le sang de femmes ayant eu
une grossesse sans complications.
1.5.3. Objectifs
Développer et valider une méthode d’analyse par HPLC-MS/MS permettant le dosage
simultané de plusieurs F2-isoprostanes.
Mesurer les F2-isoprostanes totaux (estérifiés aux phospholipides + libres) dans le plasma, le
sang total et les membranes érythrocytaires de femmes au troisième trimestre de la
grossesse par HPLC-MS/MS.
Étudier la relation entre les F2-isoprostanes et les acides gras trans dans le plasma de
femmes au troisième trimestre de la grossesse par HPLC-MS/MS.

33
Mesurer les F2-isoprostanes totaux (estérifiés aux phospholipides + libres) dans le plasma, le
sang total et les membranes érythrocytaires de femmes au troisième trimestre d’une
grossesse prééclamptique par HPLC-MS/MS.
Comparer le niveau de chaque F2-isoprostane mesuré dans les trois matrices biologiques
(plasma, sang total et membranes érythrocytaires) entre les grossesses contrôles et
prééclamptiques.


35
Chapitre 2 – F2-isoprostanes analysis in plasma of
pregnant women by HPLC-MS/MS using a column
packed with core-shell particles
2.1. Résumé
Les F2-isoprostanes contenus dans le plasma sont des biomarqueurs fiables du stress oxydatif. Il
existe 64 isomères de F2-isoprostanes pouvant être générés par la peroxydation de l’acide
arachidonique estérifié aux phospholipides. Analyser individuellement et rapidement ces isomères
représente un défi technique. Nous avons développé une méthode d’analyse par HPLC-MS/MS pour
la détermination simultanée de sept F2-isoprostanes dans le plasma, c’est-à-dire le 8-iso-15(R)-
PGF2α, le 8-iso-PGF2α, le 15(R)-PGF2α, le iPF2α-IV, le iPF2α-VI, le 5-iPF2α-VI et le (±)5-8,12-iso-iPF2α-
VI. Cette méthode a été validée et utilisée pour le dosage de ces molécules dans le plasma au
troisième trimestre de grossesse (n = 20). Les échantillons de plasma ont été soumis à une
hydrolyse alcaline puis les F2-isoprostanes totaux ont été isolés par extraction liquide-liquide. Ils ont
été séparés et analysés par HPLC-MS/MS en 16.5 minutes grâce à une colonne chromatographique
C18 remplie avec des particules partiellement poreuses. Les F2-isoprostanes de la classe VI sont les
plus abondants à avoir été mesurés dans les échantillons, représentant 65 % de tous les F2-
isoprostanes mesurés par la méthode HPLC-MS/MS. Le 15(R)-PGF2α est le plus abondant parmi les
isomères de la classe III dans le plasma des femmes enceintes.

36
F2-isoprostanes analysis in plasma of pregnant women by HPLC-MS/MS using a column
packed with core-shell particles
Jessica Larose1, Pierre Julien2,4 and Jean-François Bilodeau1,3,4*
1Axe reproduction, santé de la mère et de l'enfant, et Centre de Recherche en Biologie de la
Reproduction (CRBR), Centre de recherche du CHU de Québec, Québec, Canada G1V 4G2.
2Axe endocrinologie et néphrologie, et Centre de Recherche en endocrinologie moléculaire et
oncologique et génomique humaine (CREMOGH), Centre de recherche du CHU de Québec,
Québec, Canada G1V 4G2.
3Département d'Obstétrique, Gynécologie et Reproduction.
4Département de Médecine, Faculté de médecine, Université Laval, Québec, Canada G1K 7P4.
Running title: F2-isoPs separation using core-shell particles technology
*Correspondence and reprint request:
Centre de recherche du CHU de Québec
Axe reproduction, santé périnatale et santé de l'enfant, Local T-1-49
2705 Boulevard Laurier
Québec (Qc) G1V 4G2
Tel: (418) 525-4444 x 46153
Fax: (418) 654-2765
Email:[email protected]
Publié dans J lipid Res, février 2013

37
2.2. Abstract
Plasma F2-isoprostanes (F2-isoPs) are reliable biomarkers of oxidative stress. Several possible F2-
isoPs are generated by the oxidation of arachidonic acid esterified in phospholipids. The separation
of these isomers represents a technical challenge for rapid and selective determination. We have
developed a HPLC-MS/MS method for the simultaneous determination of seven plasma F2-isoPs,
namely 8-iso-15(R)-PGF2α, 8-iso-PGF2α, 15(R)-PGF2α, iPF2α-IV, iPF2α-VI, 5-iPF2α-VI and (±)5-8,12-
iso-iPF2α-VI. We have validated this method in plasma of pregnant women, a mild physiological
oxidative stress known to increase F2-isoPs. Thus, plasma samples of women collected at the third
trimester of pregnancy (n = 20) were subjected to alkaline hydrolysis followed by liquid-liquid
extraction in order to extract total F2-isoPs. The F2-isoPs were separated within 16.5 min using a
column packed with core-shell particles. The class VI isomers were the most abundant, accounting
for 65% of the total level of all quantified F2-isoPs in plasma of pregnant women (p < 0.05). The
15(R)-PGF2α was the most abundant of the class III isomers quantified. This method allowed fast and
selective separation of seven isomers from three different classes of F2-isoPs regioisomers.
Supplementary keywords: oxidative stress, pregnancy, mass spectrometry, 8-iso-PGF2α.
Abbreviations: F2-isoP, F2-isoprostane; IS, internal standard; LLOD, lower limit of detection; LLOQ,
lower limit of quantification; PGF2α, prostaglandin F2α; PLA2, phospholipase A2; ROS, Reactive
oxygen species;

38
2.3. Introduction
Oxidative stress is an independent risk factor for coronary heart diseases (1, 2) and is a feature of
many other pathologies (3) including hypercholesterolemia (4), inflammatory diseases (5) and
endothelial dysfunctions (6, 7). F2-isoPs found in tissues and biological fluids are reliable biomarkers
of oxidative stress (8, 9). F2-isoPs result from the free radical-mediated peroxidation of arachidonic
acid esterified in phospholipids. This reaction potentially generates 64 isomers of F2-isoPs divided
into four classes of regioisomers, each composed of eight racemic diastereoisomers (10). Once
formed, these compounds can be released from phospholipids by the phospholipases A2 (PLA2) or by
the platelet activating factor acetylhydrolase (PAF-AH or lipoprotein (Lp)-PLA2) (11). The F2-isoPs
exert their biological activity, such as vasoconstriction, in their free form in plasma and are excreted
in urine (12).
The 8-iso-propstaglandin F2α (PGF2α) has been the most studied isomer known as an in vivo
biomarker of oxidative stress (3). Urinary level of F2-isoPs is a good indicator of oxidative stress but
rely on other factors such as the rate of hydrolysis of F2-isoPs from phospholipids, their metabolism
as well as their excretion (13). However, valuable information could be obtained from the
simultaneous analyses of the plasma concentrations of several isomers of F2-isoP as they may be
formed and eliminated differentially according to the physiological state or disorder (14-17).
Several methods have been developed to measure F2-isoPs including enzyme immuno-assays and
gas or liquid chromatography coupled to mass spectrometry. The methods using HPLC-MS/MS are
usually the most selective for F2-isoPs regioisomers and diastereoisomers (18). Generally, a C18
stationary phase composed of fully porous particles were used in these methods (18-22). However,
new chromatographic columns packed with core-shell particles improve separation efficiency for
several analytes in comparison to classical porous particles (23, 24). The benefits of these new
columns for F2-isoPs analysis were not demonstrated so far to our knowledge.
Thus, the aim of this study was to develop a rapid and specific method to quantify F2-isoPs isomers in
plasma by HPLC-MS/MS using a column filled with core-shell particles. To test this new method, the
plasma total levels (esterified + free) of several F2-isoPs isomers were determined in women at the
third trimester of pregnancy. In order to detect a wide range of physiologically produced F2-isoPs

39
isomers above the baseline, we chose to use plasma of pregnant women since pregnancy is a mild
oxidative stress known to increase the levels of 8-iso-PGF2α (25-28).
2.4. Material and methods
2.4.1. Materials
The 8-iso-15(R)-PGF2α, 8-iso-PGF2α, 8-iso-PGF2β, 11β-PGF2α, 15(R)-PGF2α, 5-trans-PGF2α, PGF2α,
iPF2α-IV, (±)5-iPF2α-VI, (±)5-8,12-iso-iPF2α-VI and their deuterated counterparts 8-iso-PGF2α-d4,
PGF2α-d4, iPF2α-IV-d4, iPF2α-VI-d4, (±)5-iPF2α-VI-d11, and (±)5-8,12-iso-iPF2α-VI-d11 were
purchased from Cayman Chemical (Ann Arbor, MI, USA). Butylated hydroxytoluene (BHT) was
bought from Sigma-Aldrich (Oakville, ON, Canada) and sodium chloride was obtained from
Laboratoire Mat (Québec, QC, Canada). All other reagents and solvents were HPLC grade and were
purchased from VWR International (Ville Mont-Royal, QC, Canada).
2.4.2. Patient selection
We recruited twenty normotensive pregnant women at the Centre Mère-Enfant du Centre Hospitalier
Universitaire de Québec. The institution approved the protocol and informed consents were obtained
from all pregnant women participating to the study. Normotensive pregnancy was defined as a
pregnancy in which the mother had normal blood pressure (≤140/90 mm Hg), absence of proteinuria
and absence of other medical complications such as thrombocytopenia (platelet count <
100000×109/L), oliguria (< 500 mL/d), pulmonary edema, elevated liver enzyme levels, severe
nausea and vomiting, frontal headache, visual disturbances, persistent abdominal pain in right upper
quadrant, chest pain or shortness of breath, suspected abruption placentae, hemolysis, elevated liver
enzymes syndrome, intrauterine growth retardation and oligohydramnios. Patients presenting any
pre-existing medical conditions such as chronic hypertension, diabetes mellitus, obesity (body mass
index > 30 prior to pregnancy), kidney diseases, inflammatory intestinal diseases and blood clotting
disorders or any concurrent medical obstetrical complications, such as gestational diabetes, were
excluded. Other exclusion factors were age (< 18 years old or > 40 years old) and the intake of
anticoagulant drugs or drugs affecting lipid metabolism. No patient had symptoms associated with

40
gestational hypertension with proteinuria (preeclampsia) as it is defined by the Canadian
Hypertension Society Consensus (29). All women were non-smokers.
2.4.3. Blood collection and processing
Twenty mL of blood were collected in heparinized tubes before the active phase of labor. The
processing of blood samples was made within 2 hours. Five hundred µL of whole blood was
immediately frozen on dry ice and kept at -80°C for further analysis. The remaining blood was
centrifuged at 180 x g for 10 minutes at 20°C. The supernatant (plasma) was recentrifuged at 1300 x
g for 25 min to remove platelets from plasma, which was kept at -80°C for further analyses.
2.4.4. Preparation of standards for F2-isoPs analysis
A solution of internal standards (IS) containing 50 ng/mL of each deuterated analyte (8-iso-PGF2α-d4,
PGF2α-d4, iPF2α-IV-d4, iPF2α-VI-d4, (±)5-iPF2α-VI-d11, and (±)5-8,12-iso-iPF2α-VI-d11) in 0.01 %
acetic acid was prepared to be added to samples and standards. A stock solution containing 1 µg/mL
of each compound (8-iso-15(R)-PGF2α, 8-iso-PGF2α, 15(R)-PGF2α, 5-trans-PGF2α, PGF2α, iPF2α-IV,
(±)5-iPF2α-VI and (±)5-8,12-iso-iPF2α-VI) was prepared in 0.01 % acetic acid. This last solution was
used to prepare two sets of working solutions with concentrations ranging from 2 ng/mL to 80 ng/mL
in 0.01 % acetic acid for the preparation of the standard curve, quality control and method validation.
The first set of working solutions was diluted to obtain the standard curves for each analyte (10 µL of
working solution, 10 µL of internal standard, 80 µL of water containing 10% (v/v) acetonitrile and
0.01% (v/v) acetic acid). The second set of working solutions was diluted the same way to obtain
quality controls.
2.4.5. Extraction of F2-isoPs from plasma
F2-isoPs were extracted from plasma using a modified version of the method described by Taylor
(19). Ten µL of a BHT solution (1% in ethanol) and 10 µL of the internal standard were added to 250
µL of freshly thawed plasma. Samples were completed to 500 µL with water. Then, 500 µL of
hydrolysis solution (1 mL of 50% (w/w) KOH, 1 mL of water and 10 mL methanol) were added. The
resulting mixture was vortexed and incubated at 37°C for 60 minutes. The reaction was stopped with
100 µL of formic acid 0.05% (v/v) and acidified with 90 µL of hydrochloric acid 5 N. The tubes were
then extracted twice with 1.5 mL of hexane. The organic phase was discarded. The aqueous phase

41
was then extracted three times with 1.5 mL of 3:1 ethyl acetate:hexane. The resulting organic phase
of the three extractions were combined and evaporated to dryness under nitrogen and reconstituted
to 100 µL in 10% (v/v) acetonitrile and 0.01% (v/v) acetic acid in water.
2.4.6. Chromatography
The chromatography was carried out using a Shimadzu Prominence system (Columbia, MD, USA). A
Kinetex XB-C18 100 Å column (100 x 3.0 mm, 2.6 µm) was used preceded by a 4.0 x 2.0 mm C18
SecurityGuard Cartridges. Both were from Phenomenex (Torrance, CA, USA). The column oven
temperature was controlled at 30°C. The injection volume was 40 µL. The separation was done
using a gradient of three solvents at a flow rate of 0.45 mL/min. Solvent A was composed of 0.01 %
(v/v) acetic acid in water, solvent B consisted of 0.01 % (v/v) acetic acid in acetonitrile and solvent C
was composed of 0.01 % (v/v) acetic acid in methanol. First, solvent B was held at 17% for 1 min
while solvent C was at 33%. The latter was followed by a linear gradient for over 8.9 min to 13.5% of
B and 58.9% of C. The next step was a linear gradient over 0.5 min to 47.5% B and 47.5% C
respectively. The previous conditions were maintained for 1.6 min and the solvent B and C were
decreased to 17% and 33% in 0.1 min respectively. This last condition was maintained for an
additional 4.4 min to complete the 16.5 min run.
2.4.7. Mass Spectrometry
The HPLC was coupled to a 3200 QTRAP® LC/MS/MS system from AB Sciex (Concord, ON,
Canada) through a Turbo VTM ion source using the electrospray ionization probe. The mass
spectrometer was operated in negative mode. Curtain gas (CUR), collision gas (CAD), ion source
gas 1 (GS1) and ion source gas 2 (GS2) were set at 37, 7, 45 and 55 respectively. The ions spray
voltage (IS) was set at -4100 V and source temperature was set at 700°C. Class III F2-isoPs and their
internal standard, 8-iso-PGF2α-d4 and PGF2α-d4 (class III-d4), were monitored in the multiple-reaction
monitoring (MRM) mode using the transitions 353.3 → 193.2 and 357.3 → 197.2 m/z respectively.
Class IV F2-isoPs and their internal standard, iPF2α-IV-d4 (class IV-d4), were monitored using the
transitions 353.3 → 127.0 and 357.0 → 127.0 m/z. Finally, class VI F2-isoPs and their internal
standard, (±)5-iPF2α-VI-d11, and (±)5-8,12-iso-iPF2α-VI-d11 (class VI-d11), were analyzed using the
transitions 353.0 → 115.0 and 364.6 → 115.0 m/z respectively. Quantification was performed using
Analyst® 1.4.2 Software (AB Sciex).

42
2.4.8. Method validation
The lower limit of quantification (LLOQ) was defined as the concentration to which the signal-to-noise
ratio was higher than 5 with a precision below 20% and an accuracy of ± 20% of the nominal
concentration (30). Determination of intra-day precision was done by analyzing a pool of plasma
samples from three nonpregnant women (Innovative Research, Novi, MI) spiked with 10 µL of
working solutions containing either 0 ng/mL, 7 ng/mL and 20 ng/mL of each analyte (n = 4 per
concentration). This validation procedure was carried out on three consecutive days in order to
evaluate inter-day precision (n = 12 per concentration). Accuracy and recovery was determined using
plasma samples spiked with the 7 and 20 ng/mL working solutions. The recovery was evaluated by
comparing signal obtained for plasma spiked before extraction with signal obtained for plasma spiked
after extraction with the corresponding working solutions. Matrix effects were evaluated by
postcolumn infusion at 10 µL/min of a solution containing 100 ng/mL of each following molecules: 8-
iso-PGF2α, 8-iso-PGF2α-d4, iPF2α-IV, iPF2α-IV-d4, (±)5-iPF2α-VI, (±)5-iPF2α-VI-d11. During
postcolumn infusion, an extract of plasma was injected concomitantly using the described HPLC-
MS/MS method above.
2.4.9. Statistical analyses
Statistical analyses were performed with SigmaPlot 12.3 (Systat Software, Inc., 2011, San Jose,
USA). The normality was tested using the Shapiro-Wilk test. The Kruskal-Wallis one-way ANOVA
was used to compare levels of F2-isoPs. All pairwise multiple comparisons were done according to
Student-Newman-Keuls’ method. In all cases, a p-value lower than 0.05 was considered significant
and a p-value between 0.05 and 0.1 was considered a tendency.
2.5. Results
2.5.1. Analysis of F2-isoPs in plasma by selected reaction monitoring mass spectrometry
Compound-dependent mass spectrometric parameters were optimized for each class of regioisomers
using split infusion at conditions close to the chromatographic separation. The 8-iso-PGF2α was use
to optimize class III F2-isoPs parameters (transition 353.3 → 193.2 and 357.3 → 197.2 m/z for

43
deuterated standard respectively). The iPF2α-IV was used to define which parameters to employ for
class IV F2-isoPs. The (±)5-iPF2α-VI was used to determine parameters for class VI regioisomers. It
was not possible to measure class V F2-isoPs because no commercial standards were available.
Declustering potential (DP), entrance potential (EP), collision energy (CE), collision cell entrance
potential (CEP) and collision cell exit potential (CXP) for each transition are shown in Table 1.
2.5.2. Chromatographic separation
HPLC conditions were optimized to obtain a baseline separation of all F2-isoPs in plasma (Fig. 1).
Mass chromatograms are shown for a pure standard solution (Fig. 2 A-F) and for a typical plasma
sample (Fig. 2 G-L). All class III isomers were well resolved (Fig. 2A) except for the 8-iso-PGF2β. This
last isomer, as well as the 11β-PGF2α isomer, were not detected in the plasma of third trimester
pregnant women (Fig. 2G). The peak observed at 7.78 min (Fig. 2G) was considered an unknown
plasma compound since it has the same retention time as the 5-trans-PGF2α, but eluted
inconsistently under different chromatographic conditions (data not shown). The 8-iso-15(R)-PGF2α,
8-iso-PGF2α, 15(R)-PGF2α and PGF2α were measurable in all plasma samples analyzed. The iPF2α-IV
was well separated from impurities (Fig. 2I), but was not always detectable in our samples. Both
iPF2α-VI and 5-iPF2α-VI were found in equal proportion in the (±)5-iPF2α-VI bought from the supplier.
The iPF2α-VI-d4 (transition 357.0 → 115.0 m/z) was used to identify corresponding unlabeled
isomers (data not shown). All class VI isomers were well resolved in pure standard solution and were
all detected in the plasma samples analyzed (Fig. 2E, K). Both, iPF2α-VI and 5-iPF2α-VI are well
separated from plasma impurities (Fig. 2K). Only the (±)5-8,12-iso-iPF2α-VI barely coeluted with an
unknown compound.
2.5.3. Method validation
Quantification of F2-isoPs was done using the ratio of peak area of each analyte to the peak area of
corresponding internal standards. Class III isomers, 8-iso-15(R)-PGF2α, 8-iso-PGF2α and 15(R)-
PGF2α, were quantified using 8-iso-PGF2α-d4. The iPF2α-IV was calculated using iPF2α-IV-d4. The
iPF2α-VI-d11, 5-iPF2α-VI-d11 and (±)5-8,12-iso-iPF2α-VI-d11 were used to measure respectively the
iPF2α-VI, 5-iPF2α-VI and (±)5-8,12-iso-iPF2α-VI. The calibration curves for each F2-isoPs covered 0.2
ng/mL to 8 ng/mL and were linear throughout this range (r2 > 0.99). Lower limits of quantification
(LLOQ) ranged between 11.3 and 21.0 pg injected on column (Table. 2). At these concentrations, the

44
precision was < 20 % (% coefficient of variation (CV)) and accuracy was below ± 20 % of the nominal
concentration. Mean intra-day coefficients of variation (% CV) fluctuated from 2.6 to 5.2 % and mean
inter-day CVs varied between 3.4 and 8.2 %. Mean intra-day and inter-day accuracy were below ± 15
% of the nominal concentration except for the 8-iso-PGF2α. The extraction recovery was also
evaluated and ranged from 57.9 to 73.3 %. No matrix effect was detected at the retention times of F2-
isoPs except for 8-iso-PGF2α, which showed low ionic suppression (data not shown).
2.5.4. F2-isoPs in the plasma of pregnant women
The plasma from twenty non-smoking women was retrieved at an average of 40.6 weeks of
pregnancies before the active phase of labor. The total F2-isoPs were extracted from plasma by
liquid-liquid extraction and measured by HPLC-MS/MS using the new validated method described
above. The respective plasma median levels for 8-iso-15(R)-PGF2α, 8-iso-PGF2α, 15(R)-PGF2α, iPF2α-
IV, iPF2α-VI, 5-iPF2α-VI and 5-8,12-iso-iPF2α-VI are shown in Table 3. Statistical analyses indicated
that class VI isomers were the most abundant, accounting for 65 % of the total level of quantified F2-
isoPs (p < 0.05, Table 3) and (±)5-8,12-iso-iPF2α-VI is the most abundant of all isomers (35 % of the
total level). The 15(R)-PGF2α is the most abundant of the class III isomers quantified. However, no
significant differences were observed between 8-iso-15(R)-PGF2α, 8-iso-PGF2α and iPF2α-IV. No
significant differences were observed between iPF2α-VI and 5-iPF2α-VI as well.
2.6. Discussion
F2-isoPs found in tissues and biological fluids are reliable biomarkers of oxidative stress (8, 9). We
have developed a new HPLC-MS/MS method for the simultaneous determination of seven F2-isoPs
isomers, namely the 8-iso-15(R)-PGF2α, 8-iso-PGF2α, 15(R)-PGF2α, iPF2α-IV, iPF2α-VI, 5-iPF2α-VI and
(±)5-8,12-iso-iPF2α-VI in plasma. The liquid chromatography coupled to mass spectrometry offers
many advantages over immunoassays, allowing the analysis of many compounds in complex
matrices with excellent selectivity, sensitivity and dynamic range. The simultaneous analysis of
several isomers of F2-isoPs can better characterize the oxidative stress of specific pathologies or
physiological conditions. Indeed, in a comparative study, more than eight F2-isoP isomers were

45
increased in urine of hypercholesterolemic patients, while only three isomers, including the 8-iso-
PGF2α, were found to be increased in cases of congestive heart failure (17).
Several methods using HPLC-MS/MS have already been published for the determination of F2-isoPs
(15, 18-22, 31, 32). Most chromatographic columns reported in these methods used a C18 stationary
phase although, other types of stationary phases have also been used such as C8 or porous
graphitic carbon. However, all columns used in previous methods were filled with fully porous
particles. Our method is the first to use a chromatographic column filled with core-shell particles. This
type of particles offers better efficiency than fully porous particles. In fact, as shown by the Van
Deemter equation, many parameters contribute to peak broadening in HPLC, namely the longitudinal
diffusion, the eddy dispersion and the solid-liquid mass transfer resistance. Those three parameters
are reduced when core-shell particles are used instead of the fully porous particles of about the same
diameters (the eddy dispersion being the most affected) (33). This allowed us to achieve a better F2-
isoPs separation in only 16.5 minutes without loss of precision and accuracy. To our knowledge, the
present chromatographic separation is the fastest among previously published methods.
The HPLC conditions were optimized in order to obtain a baseline separation of F2-isoPs for each
class of regioisomers. The separation was performed using a gradient of three solvents, namely
water, methanol and acetonitrile, each containing 0.01% of acetic acid. The methanol allowed for a
better selectivity of the F2-isoPs separation, but the use of acetonitrile was essential in order to avoid
co-elution between 8-iso-PGF2α and iPF2α-VI. Indeed, it is important to prevent co-elution of isomers
from class VI with isomers of the class III since class VI isomers, when fragmented in the collision
cells, produce both the main daughter ion 115 m/z, known to be specific to class VI, and the daughter
ion 193 m/z that characterizes class III F2-isoPs. The signal for the 193 m/z ion represents about 10%
of the signal of the 115 m/z ion produced from class VI fragmentation (22). That is why the proportion
of acetonitrile in the eluent was higher at the beginning than at the end of the gradient.
In this study, we reported, the total levels of the seven F2-isoPs isomers in the plasma of twenty
healthy women at the end of third trimester of pregnancy. The 8-iso-PGF2α is the most studied isomer
used to characterize oxidative stress in vivo (3). The median level of 8-iso-PGF2α found in our study
was 195 pg/mL of plasma. Literature values for total level of 8-iso-PGF2α in plasma from healthy
subjects ranged between 40 and 170 pg/mL (34, 35). Our measurement is slightly higher but this was
expected since pregnancy, per se, is known to be a mild oxidative event (3, 25-28, 36). The class VI

46
isomers were the most abundant by two to three folds when compared to the 8-iso-PGF2α. The class
IV and VI may be underestimated marker of oxidative stress. This the first study that quantified a
wide array of F2-isoPs in blood of pregnant women. This is why, except for 8-iso-PGF2α, it is not
possible to compare our results with other studies.
In sum, we have developed a new HPLC-MS-MS method using a C18 core-shell column for the
determination of seven F2-isoP isomers among classes III, IV and VI in blood plasma. This new
method has many advantages over the previously published method for the determination F2-isoPs
such as speed and selectivity.
2.7. Acknowledgments
This work was supported by a grant from the Canadian Institutes of Health research (CIHR, grant#
MOP-84219 to J.-F.B). Jessica Larose is a recipient of a Fonds de Recherche en Santé du Québec
(FRSQ) award.
2.8. References
1. Davies, S. S., and L. J. Roberts, 2nd. 2011. F2-isoprostanes as an indicator and risk factor
for coronary heart disease. Free Radic Biol Med 50: 559-566.
2. Schwedhelm, E., A. Bartling, H. Lenzen, D. Tsikas, R. Maas, J. Brummer, F. M. Gutzki, J.
Berger, J. C. Frolich, and R. H. Boger. 2004. Urinary 8-iso-prostaglandin F2alpha as a risk marker in
patients with coronary heart disease: a matched case-control study. Circulation 109: 843-848.
3. Basu, S. 2008. F2-isoprostanes in human health and diseases: from molecular mechanisms
to clinical implications. Antioxid Redox Signal 10: 1405-1434.
4. Reilly, M. P., D. Pratico, N. Delanty, G. DiMinno, E. Tremoli, D. Rader, S. Kapoor, J. Rokach,
J. Lawson, and G. A. FitzGerald. 1998. Increased formation of distinct F2 isoprostanes in
hypercholesterolemia. Circulation 98: 2822-2828.

47
5. Pratico, D., J. Rokach, J. Lawson, and G. A. FitzGerald. 2004. F2-isoprostanes as indices of
lipid peroxidation in inflammatory diseases. Chem Phys Lipids 128: 165-171.
6. Arcaro, G., C. Fava, R. Dagradi, G. Faccini, S. Gaino, M. Degan, C. Lechi, A. Lechi, and P.
Minuz. 2004. Acute hyperhomocysteinemia induces a reduction in arterial distensibility and
compliance. J Hypertens 22: 775-781.
7. Schulz, E., T. Gori, and T. Munzel. 2011. Oxidative stress and endothelial dysfunction in
hypertension. Hypertens Res 34: 665-673.
8. Basu, S. 2010. Bioactive eicosanoids: role of prostaglandin F(2alpha) and F-isoprostanes in
inflammation and oxidative stress related pathology. Mol Cells 30: 383-391.
9. Kadiiska, M. B., B. C. Gladen, D. D. Baird, D. Germolec, L. B. Graham, C. E. Parker, A.
Nyska, J. T. Wachsman, B. N. Ames, S. Basu, N. Brot, G. A. Fitzgerald, R. A. Floyd, M. George, J.
W. Heinecke, G. E. Hatch, K. Hensley, J. A. Lawson, L. J. Marnett, J. D. Morrow, D. M. Murray, J.
Plastaras, L. J. Roberts, 2nd, J. Rokach, M. K. Shigenaga, R. S. Sohal, J. Sun, R. R. Tice, D. H. Van
Thiel, D. Wellner, P. B. Walter, K. B. Tomer, R. P. Mason, and J. C. Barrett. 2005. Biomarkers of
oxidative stress study II: are oxidation products of lipids, proteins, and DNA markers of CCl4
poisoning? Free radical biology & medicine 38: 698-710.
10. Morrow, J. D., K. E. Hill, R. F. Burk, T. M. Nammour, K. F. Badr, and L. J. Roberts, 2nd.
1990. A series of prostaglandin F2-like compounds are produced in vivo in humans by a non-
cyclooxygenase, free radical-catalyzed mechanism. Proc Natl Acad Sci U S A 87: 9383-9387.
11. Stafforini, D. M., J. R. Sheller, T. S. Blackwell, A. Sapirstein, F. E. Yull, T. M. McIntyre, J. V.
Bonventre, S. M. Prescott, and L. J. Roberts, 2nd. 2006. Release of free F2-isoprostanes from
esterified phospholipids is catalyzed by intracellular and plasma platelet-activating factor
acetylhydrolases. J Biol Chem 281: 4616-4623.
12. Milne, G. L., H. Yin, K. D. Hardy, S. S. Davies, and L. J. Roberts, 2nd. 2011. Isoprostane
generation and function. Chem Rev 111: 5973-5996.
13. Halliwell, B., and C. Y. Lee. 2010. Using isoprostanes as biomarkers of oxidative stress:
some rarely considered issues. Antioxid Redox Signal 13: 145-156.

48
14. Praticò, D., O. P. Barry, J. A. Lawson, M. Adiyaman, S.-W. Hwang, S. P. Khanapure, L.
Iuliano, J. Rokach, and G. A. FitzGerald. 1998. IPF2α-I: An index of lipid peroxidation in humans.
Proceedings of the National Academy of Sciences 95: 3449-3454.
15. Zhang, B., and K. Saku. 2007. Control of matrix effects in the analysis of urinary F2-
isoprostanes using novel multidimensional solid-phase extraction and LC-MS/MS. J Lipid Res 48:
733-744.
16. Basu, S. 2004. Isoprostanes: novel bioactive products of lipid peroxidation. Free Radic Res
38: 105-122.
17. Li, H., J. A. Lawson, M. Reilly, M. Adiyaman, S. W. Hwang, J. Rokach, and G. A. FitzGerald.
1999. Quantitative high performance liquid chromatography/tandem mass spectrometric analysis of
the four classes of F(2)-isoprostanes in human urine. Proc Natl Acad Sci U S A 96: 13381-13386.
18. Yin, H., N. A. Porter, and J. D. Morrow. 2005. Separation and identification of F2-isoprostane
regioisomers and diastereomers by novel liquid chromatographic/mass spectrometric methods. J
Chromatogr B Analyt Technol Biomed Life Sci 827: 157-164.
19. Taylor, A. W., R. S. Bruno, B. Frei, and M. G. Traber. 2006. Benefits of prolonged gradient
separation for high-performance liquid chromatography-tandem mass spectrometry quantitation of
plasma total 15-series F-isoprostanes. Analytical biochemistry 350: 41-51.
20. Haschke, M., Y. L. Zhang, C. Kahle, J. Klawitter, M. Korecka, L. M. Shaw, and U. Christians.
2007. HPLC-atmospheric pressure chemical ionization MS/MS for quantification of 15-F2t-
isoprostane in human urine and plasma. Clin Chem 53: 489-497.
21. Korecka, M., C. M. Clark, V. M. Lee, J. Q. Trojanowski, and L. M. Shaw. 2010. Simultaneous
HPLC-MS-MS quantification of 8-iso-PGF(2alpha) and 8,12-iso-iPF(2alpha) in CSF and brain tissue
samples with on-line cleanup. J Chromatogr B Analyt Technol Biomed Life Sci 878: 2209-2216.
22. Yan, W., G. D. Byrd, and M. W. Ogden. 2007. Quantitation of isoprostane isomers in human
urine from smokers and nonsmokers by LC-MS/MS. J Lipid Res 48: 1607-1617.

49
23. Gritti, F., I. Leonardis, J. Abia, and G. Guiochon. 2010. Physical properties and structure of
fine core-shell particles used as packing materials for chromatography Relationships between particle
characteristics and column performance. Journal of chromatography. A 1217: 3819-3843.
24. Gritti, F., I. Leonardis, D. Shock, P. Stevenson, A. Shalliker, and G. Guiochon. 2010.
Performance of columns packed with the new shell particles, Kinetex-C18. Journal of
chromatography. A 1217: 1589-1603.
25. Myatt, L., and X. Cui. 2004. Oxidative stress in the placenta. Histochem Cell Biol 122: 369-
382.
26. Agarwal, A., S. Gupta, and R. K. Sharma. 2005. Role of oxidative stress in female
reproduction. Reprod Biol Endocrinol 3: 28.
27. Morris, J. M., N. K. Gopaul, M. J. Endresen, M. Knight, E. A. Linton, S. Dhir, E. E. Anggard,
and C. W. Redman. 1998. Circulating markers of oxidative stress are raised in normal pregnancy and
pre-eclampsia. Br J Obstet Gynaecol 105: 1195-1199.
28. Siddiqui, I. A., A. Jaleel, W. Tamimi, and H. M. Al Kadri. 2010. Role of oxidative stress in the
pathogenesis of preeclampsia. Arch Gynecol Obstet 282: 469-474.
29. Helewa, M. E., R. F. Burrows, J. Smith, K. Williams, P. Brain, and S. W. Rabkin. 1997.
Report of the Canadian Hypertension Society Consensus Conference: 1. Definitions, evaluation and
classification of hypertensive disorders in pregnancy. Canadian Medical Association journal 157: 715-
725.
30. Food and Drug Administration. 2001. Guidance for Industry - Bioanalytical Method Validation.
31. Bohnstedt, K. C., B. Karlberg, H. Basun, and S. Schmidt. 2005. Porous graphitic carbon
chromatography-tandem mass spectrometry for the study of isoprostanes in human cerebrospinal
fluid. J Chromatogr B Analyt Technol Biomed Life Sci 827: 39-43.
32. Bohnstedt, K. C., B. Karlberg, L. O. Wahlund, M. E. Jonhagen, H. Basun, and S. Schmidt.
2003. Determination of isoprostanes in urine samples from Alzheimer patients using porous graphitic

50
carbon liquid chromatography-tandem mass spectrometry. J Chromatogr B Analyt Technol Biomed
Life Sci 796: 11-19.
33. Gritti, F., and G. Guiochon. 2012. Facts and Legends on Columns Packed with Sub-3-µm
Core-Shell Particles. LCGC North America 30: 586-595.
34. Bastani, N. E., T. E. Gundersen, and R. Blomhoff. 2009. Determination of 8-epi PGF(2alpha)
concentrations as a biomarker of oxidative stress using triple-stage liquid chromatography/tandem
mass spectrometry. Rapid Commun Mass Spectrom 23: 2885-2890.
35. Schwedhelm, E., and R. H. Boger. 2003. Application of gas chromatography-mass
spectrometry for analysis of isoprostanes: their role in cardiovascular disease. Clin Chem Lab Med
41: 1552-1561.
36. Ishihara, O., M. Hayashi, H. Osawa, K. Kobayashi, S. Takeda, B. Vessby, and S. Basu.
2004. Isoprostanes, prostaglandins and tocopherols in pre-eclampsia, normal pregnancy and non-
pregnancy. Free radical research 38: 913-918.
2.9. Figure legends
Fig. 1. Chemical structures of analytes investigated by LC-MS/MS.
Fig. 2. Mass chromatograms of a standard solution (A-F) and of a typical plasma sample spiked with
50 pg of each internal standard (G-L). Peak identification (see Figure 1 for structure): 1 = 8-iso-15(R)-
PGF2α; 2 = 8-iso-PGF2α; 3 = 8-iso-PGF2β; 4 = 11β-PGF2α; 5 = 15(R)-PGF2α; 6 = 5-trans-PGF2α; 7 =
PGF2α; 8 = 8-iso-PGF2α-d4; 9 = PGF2α-d4; 10 = iPF2α-IV; 11 = iPF2α-IV-d4; 12 = iPF2α-VI; 13 = 5-
iPF2α-VI; 14 = (±)5-8,12-iso-iPF2α-VI; 15 = iPF2α-VI-d11; 16 = 5-iPF2α-VI-d11; 17 = (±)5-8,12-iso-
iPF2α-VI-d11; ? = unknown compounds. IS = deuterated internal standards.

51
Table 1. Selected reaction monitoring parameters optimized for each classes of F2-isoPs
F2-isoPs Transitions
(m/z)
DP
(V)
EP
(V)
CEP
(V)
CE
(V)
CXP
(V)
Class III 353.3 → 193.2 -50.0 -7.0 -20.0 -34.0 -4.0
Class III-d4 357.3 → 197.2 -50.0 -7.0 -20.0 -34.0 -4.0
Class IV 353.0 → 127.0 -45.0 -7.0 -17.0 -33.0 -2.0
Class IV-d4 357.0 → 127.0 -45.0 -7.0 -17.0 -33.0 -2.0
Class VI 353.0 → 115.0 -47.0 -7.0 -21.0 -30.0 -2.0
Class VI-d11 364.6 → 115.0 -47.0 -7.0 -21.0 -30.0 -2.0
DP : declustering potential; EP : entrance potential; CEP : collision cell entrance potential; CE :
collision energy; CXP : collision cell exit potential

52
Table 2. Results for the method validation of F2-isoPs.
Intraday4 Interday5
LLOD
(pg inj.) LLOQ
(pg inj.) Recovery1
(%) Precision2
(% CV) Accuracy3
(%)
Precision (% CV)
Accuracy (%)
8-iso-15(R)-PGF2α
9.1 13.8 62.6 3.1 -3.9 4.7 -5.8
8-iso-PGF2α 10.6 16.1 71.1 4.6 -21.6 5.3 -22.6
15(R)-PGF2α 11.0 16.6 73.3 2.9 1.6 4.1 0.3
iPF2α-IV 7.5 11.3 59.1 2.6 -2.6 3.4 -4.6
iPF2α-VI 9.2 13.9 58.4 5.2 1.5 8.2 2.3
5-iPF2α-VI 8.2 12.4 57.9 3.4 1.8 6.5 0.1
(±)5-8,12-iso-iPF2α-VI
13.8 21.0 71.2 4.1 9.2 8.1 4.1
LLOD, lower limit of detetion; LLOQ, lower limit of quantification; inj., injected.
1Recovery was expressed as the mean value of data obtained from plasma samples spiked with 10
µL of working solutions containing 7 ng/mL or 20 ng/mL of each analyte.
2Precision is expressed as the mean value of data obtained from plasma samples spiked with 10 µL
of working solutions containing 0 ng/mL, 7 ng/mL or 20 ng/mL of each analyte.
3Accuracy is expressed as the mean value of data obtained from plasma samples spiked with the 7
or 20 ng/mL working solutions.
4n = 4 per concentration.
5n = 12 per concentration.

53
Table 3. F2-isoPs in the plasma of third trimester pregnant women.
F2-IsoPs levels1
(pg/ml plasma)
Class III
8-iso-15(R)-PGF2α 177 [150, 839]a
8-iso-PGF2α 195 [164, 551]a
15(R)-PGF2α 338 [285, 1758]b
Class IV
iPF2α-IV 137 [102, 984]a
Class VI
iPF2α-VI 416 [358, 4159]c
5-iPF2α-VI 393 [245, 5598]c
(±)5-8,12-iso-iPF2α-VI 879 [487, 2643]d
1Values are medians and quartiles [Q1, Q3] (n = 20).
Medians with different superscript letters (a-d) are statistically different (Kruskal-Wallis test followed
by Student-Newman-Keuls, p < 0.05).

54
Figure 1

55
Figure 2
5 ?


57
Chapitre 3 – F2-isoprostanes are correlated with
trans fatty acids in plasma of pregnant women
3.1. Résumé
Le stress oxydatif peut induire l’isomérisation cis-trans des acides gras insaturés cis et ainsi mener à
la formation d’acides gras trans in vivo. Les F2-isoprostanes (F2-isoPs) contenus dans le plasma sont
des biomarqueurs fiables du stress oxydatif in vivo. Nous avons émis comme hypothèse que le léger
stress oxydatif associé à la grossesse pourrait à la fois augmenter le niveau des F2-isoPs et celui des
acides gras trans. Des échantillons de plasma obtenus au troisième trimestre de grossesse (n = 21)
ont donc été soumis à une hydrolyse alcaline puis les F2-isoPs totaux ont été isolés par extraction
liquide-liquide. Sept isomères de F2-isoPs (8-iso-15(R)-PGF2α, 8-iso-PGF2α, 15(R)-PGF2α, iPF2α-IV,
iPF2α-VI, 5-iPF2α-VI et (±)5-8,12-iso-iPF2α-VI) ont été mesurés par HPLC-MS-MS. Les acides gras
trans dans les phospholipides du plasma ont été mesurés par GC-FID. Plusieurs corrélations
positives ont été trouvées entre certains F2-isoPs et certains acides gras trans tels que le 6t-18:1, le
9t-18:1 et le 9c,12t-18:2 (r > 0.442; p < 0.045). Le 9c,12t-18:2 était l’acide gras trans qui corrèle le
plus avec les F2-isoPs malgré sa faible concentration dans les phospholipides du plasma. Aucune
corrélation n’a été observée entre les F2-isoPs et le 9t-16:1 ni entre les F2-isoPs et le 11t-18:1. Cette
étude a démontré un lien entre les acides gras trans et les marqueurs du stress oxydatif, les F2-
isoPs, pendant la grossesse. Cela suggère aussi une compétition entre la peroxydation lipidique et
l’isomérisation cis-trans des acides gras cis.

58
F2-isoprostanes are correlated with trans fatty acids in plasma of pregnant women
Jessica Larose1,2, Karine Greffard3,5, Pierre Julien3,4,5 and Jean-François Bilodeau1,2,4,5,*
1Axe reproduction, santé de la mère et de l'enfant, Centre de recherche du CHU de Québec,
Québec, Canada G1V 4G2.
2Centre de Recherche en Biologie de la Reproduction (CRBR), Université Laval, Québec, G1V 4G2.
3Axe endocrinologie et néphrologie, Centre de recherche du CHU de Québec, Québec, Canada G1V
4G2.
4Département d'Obstétrique, Gynécologie et Reproduction, et Département de Médecine, Faculté de
médecine, Université Laval, Québec, Canada G1K 7P4
5Centre de Recherche en endocrinologie moléculaire et oncologique et génomique humaine
(CREMOGH), Université Laval, Québec, G1V 4G2.
Running title: F2-isoprostanes correlated with trans fatty acids
*Correspondence and reprint request:
Centre de recherche du CHU de Québec
Axe reproduction, santé périnatale et santé de l'enfant, Local T-1-49
2705 Boulevard Laurier
Québec (Qc) G1V 4G2
Tel: (418) 525-4444 x 46153
Fax: (418) 654-2765
Email: [email protected]

59
3.2. Abstract
Trans fatty acids (TFA) can be formed by oxidative stress through cis-trans isomerization. Plasma F2-
isoprostanes (F2-isoPs) are reliable biomarkers of oxidative stress. We hypothesized that the mild
physiological oxidative stress found in pregnancy could increase both, F2-isoPs and TFA accordingly.
Thus, plasma samples of women collected at the third trimester of pregnancy (n = 21) were subjected
to alkaline hydrolysis followed by liquid-liquid extraction in order to extract total F2-isoPs. Seven
plasma F2-isoPs (8-iso-15(R)-PGF2α, 8-iso-PGF2α, 15(R)-PGF2α, iPF2α-IV, iPF2α-VI, 5-iPF2α-VI and
(±)5-8,12-iso-iPF2α-VI) were measured by HPLC-MS-MS. TFA levels in plasma phospholipids were
measured by GC-FID. Several positive correlations were found between specific F2-isoPs and TFA,
such as 6t-18:1, 9t-18:1 and 9c,12t-18:2 (r > 0.442; p < 0.045). The 9c,12t-18:2 was the mostly
correlated TFA with F2-isoPs despite its low level in plasma phospholipids. No correlation was
observed between F2-isoPs and 9t-16:1 or 11t-18:1. This study demonstrated a link between TFA
and oxidative stress markers, F2-isoPs, during pregnancy and also suggested a competition between
lipid peroxidation and cis-trans isomerisation of the cis precursor fatty acid.
Supplementary keywords: conjugated linoleic acid, elaidic acid, oxidative stress, pregnancy, mass
spectrometry, vaccenic acid, F2-isoprostanes, 8-iso-PGF2α.
Abbreviations: 9t-16:1, palmitelaidic acid; 9c,12c-18:2, linoleic acid; 9c,12t-18:2, cis-9, trans-12
octadecadienoic acid; 9t,12t-18:2, linolenelaidic acid; 9t-18:1, elaidic acid; 11t-18:1, vaccenic acid;
F2-isoP, F2-isoprostane; GC-FID, gas chromatography coupled to a flame ionisation detector; IS,
internal standard; iTFA, industrial trans fatty acids; PGF2α, prostaglandin F2α; PLA2, phospholipase
A2; ROS, Reactive oxygen species; rTFA, ruminant trans fatty acids; TFA, trans fatty acids.

60
3.3. Introduction
Trans fatty acids (TFA) are unsaturated fatty acids containing at least one non-conjugated carbon-
carbon double bond in the trans configuration. Unsaturated fatty acids are synthetized only in the cis
configuration in human but a small amount of TFA are provided naturally from the diet. Indeed, TFA
are produced by bacteria in the rumen of ruminants and account for 2-5% of total fatty acids content
of their meat and milk (1). Before 2005 in North America, most dietary TFA were produced artificially
by partial hydrogenation of vegetable oil and were found in bakery products, deep-fried and frozen
food, packaged snack, margarines and crackers (2). Some partially hydrogenated vegetable oils
could contain up to 40–65% TFA as a percent of total fat (3, 4).
The distribution of TFA isomers from ruminants (rTFA) differs clearly from TFA of industrial sources
(iTFA). The iTFA are mainly represented by 9t-18:1 (elaidic acid) (5), while 11t-18:1 (vaccenic acid) is
the predominant rTFA (6). Consumption of iTFA has adverse effects on cardiovascular health by
altering blood lipid profile (increase of total to HDL cholesterol ratio, increase in LDL-cholesterol,
decrease in HDL-cholesterol), by modifying apolipoprotein levels, and by promoting systemic
inflammation as well as endothelial dysfunction (7-13). It is not clear yet whether rTFA induced similar
adverse effects as iTFA. Nevertheless, the outcomes of TFA on health seem more related to the
amount consumed than to their origin since rTFA are generally not consumed in sufficient quantity to
alter health (7, 14, 15). The various isomers of TFA may have different biological and physiological
effects in function of the length of the carbon chain, the number and the position of the carbon-carbon
double bonds. The trans-18:2 isomers seem more harmful for health than the trans-18:1 isomers
while the trans-16:1 isomers appeared to have no effect on risk factors for cardiovascular diseases
(8, 13, 16). Beside ingested TFA, there is also a possibility for TFA to be generated indirectly from
oxidative stress as shown by liposomes exposed to γ-radiation (17).
F2-isoprostanes (F2-isoPs) are reliable markers of oxidative stress that were associated to coronary
heart diseases (18, 19), hypercholesterolemia (20), inflammatory diseases (21), endothelial
dysfunctions (22, 23) and even pregnancy (24-27). The 8-iso-PGF2α has been the most studied
isomer so far (28). However, there exist more than 64 isomers of F2-isoPs divided into four classes of
regioisomers. They can be generated from the oxidation of arachidonic acid esterified to
phospholipids (29). F2-isoPs can be freed from phospholipids by the phospholipases A2 (PLA2) to

61
induce vasoconstriction in blood vessels (30, 31). The concomitant assessment of several F2-isoPs
can provide information on the balance between generation and elimination according to the
physiological state or the disorder studied (32-35).
The exact link between oxidative stress and TFA remains to be elucidated. However, we know that
TFA may elicit inflammation known to involve reactive oxygen species (ROS) (13, 16). Few controlled
dietary trials have been conducted in order to understand the link between TFA intake and levels of
urinary 8-iso-PGF2α. Generally, consumption of TFA was shown to increase excretion of the latter
(36-40). Urinary level of F2-isoPs is a good indicator of oxidative stress but rely on other factors such
as the rate of hydrolysis of F2-isoPs from phospholipids, their metabolism as well as their excretion
(41). However, valuable information could be obtained from the simultaneous analyses of the plasma
concentrations of several isomers of F2-isoPs.
Dietary intake of TFA can be estimated based on the TFA levels in plasma phospholipids (40, 42-45).
Thus, the aim of this study was to assess the relationship between the plasma concentrations of TFA
in phospholipids and the total (esterified + free) levels of several F2-isoPs isomers from women at the
third trimester of pregnancy, knowing that mild oxidative events have been shown to increase the
levels of 8-iso-PGF2α (24-27).
3.4. Material and methods
3.4.1. Materials
The 8-iso-15(R)-PGF2α, 8-iso-PGF2α, 8-iso-PGF2β, 11β-PGF2α, 15(R)-PGF2α, 5-trans-PGF2α, PGF2α,
iPF2α-IV, (±)5-iPF2α-VI, (±)5-8,12-iso-iPF2α-VI and their deuterated counterparts 8-iso-PGF2α-d4,
PGF2α-d4, iPF2α-IV-d4, iPF2α-VI-d4, (±)5-iPF2α-VI-d11, and (±)5-8,12-iso-iPF2α-VI-d11 were all
purchased from Cayman Chemical (Ann Arbor, MI, USA). Butylated hydroxytoluene (BHT) was
bought from Sigma-Aldrich (Oakville, ON, Canada) and sodium chloride (ACS grade) was obtained
from Laboratoire Mat (Québec, QC, Canada). All other reagents and solvents were HPLC grade and
were purchased from VWR International (Ville Mont-Royal, QC, Canada).

62
3.4.2. Patient recruitment
Twenty-one normotensive (blood pressure ≤140/90 mm Hg) pregnant women were recruited at the
Centre Mère-Enfant du Centre Hospitalier Universitaire de Québec. The institution approved the
protocol and informed consents were obtained from all pregnant women. Exclusion factors were age
(<18 years old or > 40 years old), intake of anticoagulant drugs or drugs affecting lipid metabolism,
chronic hypertension, diabetes mellitus, obesity (body mass index > 30 prior to pregnancy), kidney
diseases, inflammatory intestinal diseases, blood clotting disorders and gestational diabetes.
3.4.3. Blood collection and processing
Twenty mL of blood were collected in heparinized tubes before the active phase of labor. The
processing of blood samples was made within 2 hours. Five hundred µL of whole blood was
immediately frozen on dry ice and kept at -80°C for further analysis. The remaining blood was
centrifuged at 180 x g for 10 minutes at 20°C. The supernatant (plasma) was recentrifuged at 1300 x
g for 25 min to remove platelets from plasma, which was kept at -80°C for further analyses.
3.4.4. F2-isoPs analysis by HPLC-MS/MS
The F2-isoPs were extracted from plasma using a modified version (46) of the method originally
described by Taylor (47). The HPLC-MS/MS analysis of F2-isoPs was described by us previously
(46). Briefly, the chromatography was carried out using a Shimadzu Prominence system (Columbia,
MD, USA) coupled to a 3200 QTRAP® LC/MS/MS system from AB Sciex (Concord, ON, Canada)
operated in the negative mode. A Kinetex XB-C18 100 Å column (100 x 3.0 mm, 2.6 µm) was used
preceded by a 4.0 x 2.0 mm C18 SecurityGuard Cartridges. Both were from Phenomenex (Torrance,
CA, USA). The injection volume was 40 µL. The separation was done using a gradient of 3 solvents
(water, acetonitrile and methanol, each containing 0.01 % (v/v) acetic acid) at a flow rate of 0.45
mL/min (46).
The class III F2-isoPs and their internals standards, 8-iso-PGF2α-d4 and PGF2α-d4 (class III-d4), were
monitored in the multiple-reaction monitoring (MRM) mode at the transitions 353.3 → 193.2 and
357.3 → 197.2 m/z respectively. The Class IV F2-isoPs and their internal standard, iPF2α-IV-d4 (class
IV-d4), were monitored at the transitions 353.3 → 127.0 and 357.0 → 127.0 m/z. Lastly, class VI F2-
isoPs and their internals standards, (±)5-iPF2α-VI-d11, and (±)5-8,12-iso-iPF2α-VI-d11 (class VI-d11),

63
were analyzed at the transitions 353.0 → 115.0 and 364.6 → 115.0 m/z respectively. Quantification
was performed using Analyst® 1.4.2 Software (AB Sciex).
3.4.5. Determination of plasma fatty acid profile by gas chromatography
Plasma fatty acids were isolated according to a method previously described (48). Briefly, a solution
of chloroform:methanol (2:1, by volume) was used to extract lipids from plasma. Then, phospholipids
were separated by thin layer chromatography using a mix of isopropyl ether:acetic acid (96:4) as
eluant and phospholipid fatty acids were methylated following a transesterification reaction using a
mix of methanol:benzene (4:1) and acetyl chloride. Methylated fatty acids were finally analysed by
gas chromatography coupled with a flame ionisation detector (GC-FID) as explained elsewhere (49).
3.4.6. Statistical analyses
Statistical analyses were performed with SigmaPlot 12.3 (Systat Software, Inc., 2011, San Jose,
USA). The normality was tested using the Shapiro-Wilk test. The Kruskal-Wallis one-way ANOVA
was used to compare levels of F2-isoPs and of fatty acids. All pairwise multiple comparisons were
done according to Student-Newman-Keuls’ method. The Spearman's rank correlation coefficient was
used to study relationship between variables. In all cases, a p-value lower than 0.05 was considered
significant and a p-value between 0.05 and 0.1 was considered a tendency.
3.5. Results
3.5.1. F2-isoPs in the plasma of pregnant women
The F2-isoPs from twenty-one women, retrieved at the end of their pregnancies before the active
phase of labor, were separated from plasma by liquid-liquid extraction and measured by HPLC-
MS/MS. The plasma median levels for 8-iso-15(R)-PGF2α, 8-iso-PGF2α, 15(R)-PGF2α, iPF2α-IV, iPF2α-
VI, 5-iPF2α-VI and 5-8,12-iso-iPF2α-VI are reported in table 1. The statistical analysis revealed that
class VI isomers are the most abundant (p < 0.05, Table 1) and (±)5-8,12-iso-iPF2α-VI is the most
abundant of all isomers. The 15(R)-PGF2α is the most abundant of the class III isomers quantified.

64
3.5.2. Analysis of plasma fatty acids
Determination of plasma phospholipid fatty acids profile was done by GC-FID. This method allowed
for the determination of a large number of fatty acids. The median concentrations of trans fatty acids
of interest and of their corresponding cis isomer are presented in Table 2. The cis isomers are
generally found in greater quantities than trans isomers except for the 7c-18:1 + 6c-18:1, which level
was about the same as the 6t-18:1 (p > 0.05). The 9c,12c-18:1 (linoleic acid) is the most abundant of
all unsaturated fatty acids measured while 9c-18:1 is the most abundant of all monounsaturated fatty
acids, representing respectively 36 and 18 % of total unsaturated fatty acids (data not shown).
Monounsaturated trans fatty acids (6t-18:1, 9t-18:1 and 11t-18:1) are present in greater amounts than
the trans isomers of linoleic acid (9c,12t-18:2, 9t,12c-18:2 and 9t,12t-18:2). Among the
aforementioned trans isomers of linoleic acid, only the concentration of 9c,12t-18:2 was high enough
to be detected in the pregnant women.
3.5.3. Relationship between F2-isoPs and trans fatty acids
Spearman's rank correlation coefficient was used to study relations between F2-isoPs and fatty acids
(Table 3) in plasma of healthy women at the third trimester of pregnancy. Many positive and
significant correlations were found between F2-isoPs isomers and trans fatty acids. A correlation was
found between the 6t-18:1 and the 8-iso-PGF2α (r = 0.456, p = 0.038) and strong tendencies for
similar correlations were also observed for this fatty acid with the 5-iPF2α-VI (r = 0.419, p = 0.058)
and with the (±)5-8,12-iso-iPF2α-VI (r = 0.426, p = 0.053) respectively. The 9t-18:1 correlated
positively with both, the 5-iPF2α-VI (r = 0.461, p = 0.035) and the (±)5-8,12-iso-iPF2α-VI (r = 0.483, p =
0.026). The 9c,12t-18:2 correlated with 15(R)-PGF2α (r = 0.576, p = 0.006), iPF2α-IV (r = 0.442, p =
0.045) and iPF2α-VI (r = 0.469, p = 0.032) respectively. Of note, no correlation was found between F2-
isoPs and cis fatty acids, the 11t-18:1 or with the 9t-16:1.
3.6. Discussion
We recruited twenty-one healthy women at third trimester of pregnancy and measured the total levels
of the seven F2-isoPs isomers in their blood plasma. The median level of 8-iso-PGF2α, the most
studied isomer, was 184 pg/mL of plasma. Literature values for total level of 8-iso-PGF2α in plasma

65
from healthy subjects ranged between 40 and 170 pg/mL (50, 51). This slightly higher level was
expected since pregnancy can be considered as a mild oxidative event (24-28, 46, 52). The class VI
isomers were the most abundant by two to three folds when compared to the 8-iso-PGF2α. We
reported similar results for the seven F2-isoPs isomers in pregnant women recently (46).
Fatty acids associated to plasma phospholipids were also measured for each of the pregnant women.
The plasma fatty acids profile is known to be influenced by dietary habits (43). The 9t-18:1/11t-18:1
index can be used to evaluate whether the diet is high in iTFA or in rTFA. This ratio is reduced by
high dairy fat intake (53). A previous study in lactating women demonstrated the effect of a change in
diet on the fatty acid composition in different plasma lipid fractions (44). Thus, when women
consumed more butter, the 9t-18:1/11t-18:1 index in phospholipids was 0.5. If they consumed low-
TFA margarine, this ratio was lowered at 0.23 but, if they consumed regular margarine (high-TFA),
this ratio rose to 0.78. In our cohort, the 9t-18:1/11t-18:1 index was 0.39. Taken together, these
results suggest that the women in our cohort had a diet rich in rTFA, and/or low in TFA (44).
We studied in depth the relation between the seven F2-isoPs isomers measured in plasma and the
fatty acids content from the plasma phospholipids. Several significant and positive correlations were
found, demonstrating a clear link between oxidative stress and TFA in vivo. However, not all
measured F2-isoPs isomers correlated with a given TFA. This shows the importance of measuring a
large number of F2-isoPs isomers. Also, this indicates that F2-isoPs formation and elimination in vivo
differs from one isomer to another. Many controlled dietary trials have shown a relation between 8-
iso-PGF2α and higher consumption of TFA (36, 38, 39). Indeed, one study showed a 20% increase in
the level of urinary 8-iso-PGF2α in healthy subjects after three weeks of diet rich in iTFA in
comparison to the control diet (36). In contrast, another report showed no effect for of a 16-week diet
containing iTFA on the level of urinary 8-iso-PGF2α in fifty-two healthy overweight postmenopausal
women (37). Two other studies using a 1:1 mixture of 11t-18:1 (rTFA) and 12t-18:1 supplemented to
the daily diet, over a 6-week or a 9-day period, showed an increased level of urinary 8-iso-PGF2α in
healthy subjects. However, it was not clear from which TFA isomer was associated to the increase of
8-iso-PGF2α (38, 39). A dietary intervention study done to understand the effect of 11t-18:1 showed
no differences on urinary 8-iso-PGF2α level among twenty-two young men receiving a test diet for 5
weeks compared to twenty young men receiving the control diet (40). These two diets differed by the
amount of 11t-18:1 and of the cis isomer, 9c-18:1. This difference may explain the apparent lack of

66
effect of a higher consumption of 11t-18:1 on the 8-iso-PGF2α level. The previously mentioned
studies were all done on a limited time period (from 9 days to 16 weeks) and did not investigate the
consequences of trans-16:1 and trans-18:2 isomers on oxidative stress. Therefore, the relation
between 11t-18:1 and oxidative stress in these studies remains unclear. Furthermore, only urinary 8-
iso-PGF2α was used as biomarker of oxidative stress. Its level may depend on other factors than
oxidative stress such as the rate of hydrolysis of 8-iso-PGF2α from phospholipids, its metabolism and
its excretion. In our study, plasma 8-iso-PGF2α correlated exclusively with 6t-18:1. The use of a single
biomarker of oxidative stress would not have properly assessed the link with TFA.
Our study has shown that each TFA is differentially linked to oxidative stress. Both, the 6t-18:1 and
the 9t-18:1 are iTFA related to oxidative stress in our cohort. This is in accordance with results
obtained by most of the dietary trials discussed earlier. The 9c,12t-18:2, a trans isomer of linoleic
acid, is the TFA that present a higher number of tendency or correlations with F2-isoPs isomers than
any other TFA. Thus, among all TFA considered in our study, the 9c,12t-18:2 is clearly associated
with oxidative stress despite its very low level in plasma phospholipids. The trans-18:2 isomers have
generally been associated with a higher relative risk of cardiovascular diseases than trans-18:1
isomers (8, 54) but their impact on oxidative stress was unknown before the present study.
The 9t-16:1 is not associated to markers of oxidative stress in our work. Most studies that have
questioned the effects of trans-16:1 on the risk of cardiovascular disease, systemic inflammation or
endothelial dysfunction have shown that these TFA had no adverse effects on health (8). The
oxidative stress and 11t-18:1 were not associated either in our study. It has first been believed that
rTFA, such as 11t-18:1, were not harmful. However, a clinical and controlled nutritional study has
shown that a high intake of rTFA by healthy men induced deleterious changes in their plasma lipid
profile, just like a high intake of iTFA, while a moderate intake of rTFA had no effect (14). Another 3-
week study performed in healthy subjects has shown that both industrial and natural TFA affected the
cardiovascular risk factors in a sex-specific manner (55). A significant elevation was observed in total,
LDL- and HDL-cholesterol exclusively in women consuming rTFA rather than iTFA. Thus, according
to these studies, rTFA can modify the lipid profile as well as iTFA. This likely may also affect oxidative
stress. It appeared that the pregnant women in our cohort did not consume an amount sufficient of
11t-18:1 to induce an oxidative stress even if the 11t-18:1 is the most abundant TFA measured.

67
Several studies have shown that high consumption of both industrial and natural TFA causes
oxidative stress and induces the formation of F2-isoPs as discussed earlier, but discrepancies
remains. This suggests that free radicals formed during a typical pregnancy are able to induce the
formation of TFA in vivo via cis-trans isomerization of their cis precursor. This specific phenomenon
of increased TFA has been observed in erythrocyte membranes and kidney of aged rats fed with food
containing no TFA and exposed to carbon tetrachloride, as an oxidative stress inducer (56). It has
also been demonstrated that trans-arachidonic acid may originate from the NO2-mediated AA
isomerization in human (57). An in vitro study has shown the competition between lipid peroxidation
and isomerization (17). This competition could explain, at least in part, why correlations are observed
between 9t-18:1 and F2-isoPs but not with 11t-18:1 in our work. The cis precursor of 9t-18:1, the 9c-
18:1, is the most abundant monounsaturated fatty acid measured in the plasma phospholipids of third
trimester pregnant women. It accounts for 8.55% of total fatty acids content while 11c-18:1 accounts
for only for 0.24%. Thus, the 9c-18:1 fatty acid is more likely to be isomerized by free radicals than
11c-18:1. Same observation can be drawn from 9c,12t-18:1, which precursor, linoleic acid (9c,12c-
18:2), is the most abundant of all unsaturated fatty acids in our cohort, accounting for 19.26% of total
fatty acids. Knowing that polyunsaturated fatty acids are more easily isomerized than
monounsaturated (58), a stronger link between 9c,12t-18:2 and F2-isoPs was expected. However,
according to the in vitro study showing the competition between lipid peroxidation and isomerization
(17), the major trans isomer of 9c,12c-18:2 formed in liposomes is 9t,12c-18:2. However, we did not
detect this specific isomer in our cohort, neither for the 9t,12t-18:2. This suggests that the in vivo
oxidative stress is more complex and differs from that induced in vitro by γ-irradiation.
In sum, this is the first in vivo demonstration of the link between the levels of TFA and oxidative
stress markers, F2-isoPs, in the plasma of normal third trimester pregnant women, a condition
involving a mild, but physiological oxidative stress. The dietary TFA found in biological membranes
can induce oxidative stress. Also, TFA can be formed in vivo by the oxidative stress since ROS
generated can react with the unsaturated fatty acids to cause either a cis-trans isomerization, or a
peroxidation reaction yielding to F2-isoPs. These two pathways most likely explained the link between
TFA and F2-isoPs in the present report. In perspective, it would be of interest to investigate the profile
of the F2-isoPs isomers in pathophysiological conditions involving much higher oxidative challenges,
such as hypertension, preeclampsia and diabetes.

68
3.7. Acknowledgments
The authors would like to thank Ms. Line Berthiaume and M. François Cadelis for their help with the
extraction of fatty acids. This work was supported by a grant from the Canadian Institutes of Health
research (CIHR, grant# MOP-84219 to J.-F.B). Jessica Larose is a recipient of a Fonds de
Recherche en Santé du Québec (FRSQ) award.
3.8. References
1. List, G. R., D. Kritchevsky, and W. M. N. Ratnayake. 2007. Trans Fats in Foods. In. AOCS
Press. 97-126.
2. Allison, D. B., S. K. Egan, L. M. Barraj, C. Caughman, M. Infante, and J. T. Heimbach. 1999.
Estimated intakes of trans fatty and other fatty acids in the US population. J Am Diet Assoc 99: 166-
174; quiz 175-166.
3. Ratnayake, W. M., M. R. L'Abbe, S. Farnworth, L. Dumais, C. Gagnon, B. Lampi, V. Casey,
D. Mohottalage, I. Rondeau, L. Underhill, M. Vigneault, W. Lillycrop, M. Meleta, L. Y. Wong, T. Ng, Y.
Gao, K. Kwong, S. Chalouh, P. Pantazopoulos, H. Gunaratna, A. Rahardja, R. Blagden, V. Roscoe,
T. Krakalovich, G. Neumann, and G. A. Lombaert. 2009. Trans fatty acids: current contents in
Canadian foods and estimated intake levels for the Canadian population. J AOAC Int 92: 1258-1276.
4. Precht, D., and J. Molkentin. 1995. Trans fatty acids: Implications for health, analytical
methods, incidence in edible fats and intake. Food / Nahrung 39: 343-374.
5. Wolff, R., N. Combe, F. Destaillats, C. Boué, D. Precht, J. Molkentin, and B. Entressangles.
2000. Follow-up of the Δ4 to Δ16 trans-18:1 isomer profile and content in French processed foods
containing partially hydrogenated vegetable oils during the period 1995–1999. Analytical and
nutritional implications. Lipids 35: 815-825.
6. Mendis, S., C. Cruz-Hernandez, and W. M. Ratnayake. 2008. Fatty acid profile of Canadian
dairy products with special attention to the trans-octadecenoic acid and conjugated linoleic acid
isomers. J AOAC Int 91: 811-819.

69
7. Micha, R., and D. Mozaffarian. 2009. Trans fatty acids: effects on metabolic syndrome, heart
disease and diabetes. Nat Rev Endocrinol 5: 335-344.
8. Mozaffarian, D., A. Aro, and W. C. Willett. 2009. Health effects of trans-fatty acids:
experimental and observational evidence. Eur J Clin Nutr 63 Suppl 2: S5-21.
9. Brouwer, I. A., A. J. Wanders, and M. B. Katan. 2010. Effect of animal and industrial trans
fatty acids on HDL and LDL cholesterol levels in humans--a quantitative review. PLoS One 5: e9434.
10. Mozaffarian, D. 2006. Trans fatty acids - effects on systemic inflammation and endothelial
function. Atheroscler Suppl 7: 29-32.
11. Mozaffarian, D., M. B. Katan, A. Ascherio, M. J. Stampfer, and W. C. Willett. 2006. Trans
fatty acids and cardiovascular disease. N Engl J Med 354: 1601-1613.
12. Gebauer, S. K., T. L. Psota, and P. M. Kris-Etherton. 2007. The diversity of health effects of
individual trans fatty acid isomers. Lipids 42: 787-799.
13. Lopez-Garcia, E., M. B. Schulze, J. B. Meigs, J. E. Manson, N. Rifai, M. J. Stampfer, W. C.
Willett, and F. B. Hu. 2005. Consumption of trans fatty acids is related to plasma biomarkers of
inflammation and endothelial dysfunction. J Nutr 135: 562-566.
14. Motard-Belanger, A., A. Charest, G. Grenier, P. Paquin, Y. Chouinard, S. Lemieux, P.
Couture, and B. Lamarche. 2008. Study of the effect of trans fatty acids from ruminants on blood
lipids and other risk factors for cardiovascular disease. Am J Clin Nutr 87: 593-599.
15. Lacroix, E., A. Charest, A. Cyr, L. Baril-Gravel, Y. Lebeuf, P. Paquin, P. Y. Chouinard, P.
Couture, and B. Lamarche. 2012. Randomized controlled study of the effect of a butter naturally
enriched in trans fatty acids on blood lipids in healthy women. Am J Clin Nutr 95: 318-325.
16. Mozaffarian, D., T. Pischon, S. E. Hankinson, N. Rifai, K. Joshipura, W. C. Willett, and E. B.
Rimm. 2004. Dietary intake of trans fatty acids and systemic inflammation in women. Am J Clin Nutr
79: 606-612.

70
17. Mihaljevic, B., I. Tartaro, C. Ferreri, and C. Chatgilialoglu. 2011. Linoleic acid peroxidation vs.
isomerization: a biomimetic model of free radical reactivity in the presence of thiols. Org Biomol
Chem 9: 3541-3548.
18. Davies, S. S., and L. J. Roberts, 2nd. 2011. F2-isoprostanes as an indicator and risk factor
for coronary heart disease. Free Radic Biol Med 50: 559-566.
19. Schwedhelm, E., A. Bartling, H. Lenzen, D. Tsikas, R. Maas, J. Brummer, F. M. Gutzki, J.
Berger, J. C. Frolich, and R. H. Boger. 2004. Urinary 8-iso-prostaglandin F2alpha as a risk marker in
patients with coronary heart disease: a matched case-control study. Circulation 109: 843-848.
20. Reilly, M. P., D. Pratico, N. Delanty, G. DiMinno, E. Tremoli, D. Rader, S. Kapoor, J. Rokach,
J. Lawson, and G. A. FitzGerald. 1998. Increased formation of distinct F2 isoprostanes in
hypercholesterolemia. Circulation 98: 2822-2828.
21. Pratico, D., J. Rokach, J. Lawson, and G. A. FitzGerald. 2004. F2-isoprostanes as indices of
lipid peroxidation in inflammatory diseases. Chem Phys Lipids 128: 165-171.
22. Arcaro, G., C. Fava, R. Dagradi, G. Faccini, S. Gaino, M. Degan, C. Lechi, A. Lechi, and P.
Minuz. 2004. Acute hyperhomocysteinemia induces a reduction in arterial distensibility and
compliance. J Hypertens 22: 775-781.
23. Schulz, E., T. Gori, and T. Munzel. 2011. Oxidative stress and endothelial dysfunction in
hypertension. Hypertens Res 34: 665-673.
24. Myatt, L., and X. Cui. 2004. Oxidative stress in the placenta. Histochem Cell Biol 122: 369-
382.
25. Agarwal, A., S. Gupta, and R. K. Sharma. 2005. Role of oxidative stress in female
reproduction. Reprod Biol Endocrinol 3: 28.
26. Morris, J. M., N. K. Gopaul, M. J. Endresen, M. Knight, E. A. Linton, S. Dhir, E. E. Anggard,
and C. W. Redman. 1998. Circulating markers of oxidative stress are raised in normal pregnancy and
pre-eclampsia. Br J Obstet Gynaecol 105: 1195-1199.

71
27. Siddiqui, I. A., A. Jaleel, W. Tamimi, and H. M. Al Kadri. 2010. Role of oxidative stress in the
pathogenesis of preeclampsia. Arch Gynecol Obstet 282: 469-474.
28. Basu, S. 2008. F2-isoprostanes in human health and diseases: from molecular mechanisms
to clinical implications. Antioxid Redox Signal 10: 1405-1434.
29. Morrow, J. D., K. E. Hill, R. F. Burk, T. M. Nammour, K. F. Badr, and L. J. Roberts, 2nd.
1990. A series of prostaglandin F2-like compounds are produced in vivo in humans by a non-
cyclooxygenase, free radical-catalyzed mechanism. Proc Natl Acad Sci U S A 87: 9383-9387.
30. Stafforini, D. M., J. R. Sheller, T. S. Blackwell, A. Sapirstein, F. E. Yull, T. M. McIntyre, J. V.
Bonventre, S. M. Prescott, and L. J. Roberts, 2nd. 2006. Release of free F2-isoprostanes from
esterified phospholipids is catalyzed by intracellular and plasma platelet-activating factor
acetylhydrolases. J Biol Chem 281: 4616-4623.
31. Milne, G. L., H. Yin, K. D. Hardy, S. S. Davies, and L. J. Roberts, 2nd. 2011. Isoprostane
generation and function. Chem Rev 111: 5973-5996.
32. Praticò, D., O. P. Barry, J. A. Lawson, M. Adiyaman, S.-W. Hwang, S. P. Khanapure, L.
Iuliano, J. Rokach, and G. A. FitzGerald. 1998. IPF2α-I: An index of lipid peroxidation in humans.
Proceedings of the National Academy of Sciences 95: 3449-3454.
33. Zhang, B., and K. Saku. 2007. Control of matrix effects in the analysis of urinary F2-
isoprostanes using novel multidimensional solid-phase extraction and LC-MS/MS. J Lipid Res 48:
733-744.
34. Basu, S. 2004. Isoprostanes: novel bioactive products of lipid peroxidation. Free Radic Res
38: 105-122.
35. Li, H., J. A. Lawson, M. Reilly, M. Adiyaman, S. W. Hwang, J. Rokach, and G. A. FitzGerald.
1999. Quantitative high performance liquid chromatography/tandem mass spectrometric analysis of
the four classes of F(2)-isoprostanes in human urine. Proc Natl Acad Sci U S A 96: 13381-13386.

72
36. Smit, L. A., M. B. Katan, A. J. Wanders, S. Basu, and I. A. Brouwer. 2011. A high intake of
trans fatty acids has little effect on markers of inflammation and oxidative stress in humans. J Nutr
141: 1673-1678.
37. Bendsen, N. T., S. Stender, P. B. Szecsi, S. B. Pedersen, S. Basu, L. I. Hellgren, J. W.
Newman, T. M. Larsen, S. B. Haugaard, and A. Astrup. 2011. Effect of industrially produced trans fat
on markers of systemic inflammation: evidence from a randomized trial in women. J Lipid Res 52:
1821-1828.
38. Kuhnt, K., A. Wagner, J. Kraft, S. Basu, and G. Jahreis. 2006. Dietary supplementation with
11trans- and 12trans-18:1 and oxidative stress in humans. Am J Clin Nutr 84: 981-988.
39. Turpeinen, A. M., M. Mutanen, A. Aro, I. Salminen, S. Basu, D. L. Palmquist, and J. M.
Griinari. 2002. Bioconversion of vaccenic acid to conjugated linoleic acid in humans. Am J Clin Nutr
76: 504-510.
40. Tholstrup, T., M. Raff, S. Basu, P. Nonboe, K. Sejrsen, and E. M. Straarup. 2006. Effects of
butter high in ruminant trans and monounsaturated fatty acids on lipoproteins, incorporation of fatty
acids into lipid classes, plasma C-reactive protein, oxidative stress, hemostatic variables, and insulin
in healthy young men. Am J Clin Nutr 83: 237-243.
41. Halliwell, B., and C. Y. Lee. 2010. Using isoprostanes as biomarkers of oxidative stress:
some rarely considered issues. Antioxid Redox Signal 13: 145-156.
42. Mansour, M. P., D. Li, and A. J. Sinclair. 2001. The occurrence of trans-18:1 isomers in
plasma lipids classes in humans. Eur J Clin Nutr 55: 59-64.
43. Seppanen-Laakso, T., I. Laakso, P. Backlund, H. Vanhanen, and J. Viikari. 1996. Elaidic and
trans-vaccenic acids in plasma phospholipids as indicators of dietary intake of 18:1 trans-fatty acids.
J Chromatogr B Biomed Appl 687: 371-378.
44. Shahin, A. M., M. K. McGuire, N. Anderson, J. Williams, and M. A. McGuire. 2006. Effects of
margarine and butter consumption on distribution of trans-18:1 fatty acid isomers and conjugated
linoleic acid in major serum lipid classes in lactating women. Lipids 41: 141-147.

73
45. Vessby, B. 2003. Dietary fat, fatty acid composition in plasma and the metabolic syndrome.
Curr Opin Lipidol 14: 15-19.
46. Larose, J., P. Julien, and J. F. Bilodeau. 2013. F2-isoprostanes analysis in plasma of
pregnant women by HPLC-MS/MS using a column packed with core-shell particles. J Lipid Res.
47. Taylor, A. W., R. S. Bruno, B. Frei, and M. G. Traber. 2006. Benefits of prolonged gradient
separation for high-performance liquid chromatography-tandem mass spectrometry quantitation of
plasma total 15-series F-isoprostanes. Analytical biochemistry 350: 41-51.
48. Nigam, A., N. Frasure-Smith, F. Lesperance, and P. Julien. 2009. Relationship between n-3
and n-6 plasma fatty acid levels and insulin resistance in coronary patients with and without
metabolic syndrome. Nutrition, metabolism, and cardiovascular diseases 19: 264-270.
49. Counil, E., P. Julien, B. Lamarche, M. L. Chateau-Degat, A. Ferland, and E. Dewailly. 2009.
Association between trans-fatty acids in erythrocytes and pro-atherogenic lipid profiles among
Canadian Inuit of Nunavik: possible influences of sex and age. Br J Nutr 102: 766-776.
50. Bastani, N. E., T. E. Gundersen, and R. Blomhoff. 2009. Determination of 8-epi PGF(2alpha)
concentrations as a biomarker of oxidative stress using triple-stage liquid chromatography/tandem
mass spectrometry. Rapid Commun Mass Spectrom 23: 2885-2890.
51. Schwedhelm, E., and R. H. Boger. 2003. Application of gas chromatography-mass
spectrometry for analysis of isoprostanes: their role in cardiovascular disease. Clin Chem Lab Med
41: 1552-1561.
52. Ishihara, O., M. Hayashi, H. Osawa, K. Kobayashi, S. Takeda, B. Vessby, and S. Basu.
2004. Isoprostanes, prostaglandins and tocopherols in pre-eclampsia, normal pregnancy and non-
pregnancy. Free radical research 38: 913-918.
53. Enke, U., A. Jaudszus, E. Schleussner, L. Seyfarth, G. Jahreis, and K. Kuhnt. 2011. Fatty
acid distribution of cord and maternal blood in human pregnancy: special focus on individual trans
fatty acids and conjugated linoleic acids. Lipids Health Dis 10: 247.

74
54. Sun, Q., J. Ma, H. Campos, S. E. Hankinson, J. E. Manson, M. J. Stampfer, K. M. Rexrode,
W. C. Willett, and F. B. Hu. 2007. A prospective study of trans fatty acids in erythrocytes and risk of
coronary heart disease. Circulation 115: 1858-1865.
55. Chardigny, J. M., F. Destaillats, C. Malpuech-Brugere, J. Moulin, D. E. Bauman, A. L. Lock,
D. M. Barbano, R. P. Mensink, J. B. Bezelgues, P. Chaumont, N. Combe, I. Cristiani, F. Joffre, J. B.
German, F. Dionisi, Y. Boirie, and J. L. Sebedio. 2008. Do trans fatty acids from industrially produced
sources and from natural sources have the same effect on cardiovascular disease risk factors in
healthy subjects? Results of the trans Fatty Acids Collaboration (TRANSFACT) study. Am J Clin Nutr
87: 558-566.
56. Zambonin, L., C. Prata, L. Cabrini, T. Maraldi, D. Fiorentini, F. Vieceli Dalla Sega, G. Hakim,
and L. Landi. 2008. Effect of radical stress and ageing on the occurrence of trans fatty acids in rats
fed a trans-free diet. Free Radic Biol Med 44: 594-601.
57. Zghibeh, C. M., V. Raj Gopal, C. D. Poff, J. R. Falck, and M. Balazy. 2004. Determination of
trans-arachidonic acid isomers in human blood plasma. Analytical biochemistry 332: 137-144.
58. Chatgilialoglu, C., and C. Ferreri. 2005. Trans lipids: the free radical path. Acc Chem Res 38:
441-448.

75
Table 1. F2-isoprostanes in the plasma of third trimester pregnant women.
Isoprostanes levels1
(pg/ml plasma)
Class III
8-iso-15(R)-PGF2α 173 [143, 760]a
8-iso-PGF2α 184 [149, 537]a
15(R)-PGF2α 322 [250, 1724]b
Class IV
iPF2α-IV 132 [98, 957]a
Class VI
iPF2α-VI 402 [330, 3885]c
5-iPF2α-VI 335 [241, 5594]d
(±)5-8,12-iso-iPF2α-VI 766 [480, 2440]e
1Values are medians and quartiles [Q1, Q3] (n = 21).
Medians with different superscript letters (a-e) are statistically different (Kruskal-Wallis test followed
by Student-Newman-Keuls, p < 0.05).

76
Table 2. Fatty acids of interest in plasma of third trimester pregnant women.
Fatty acids levels 1
(µg/mL plasma) (%)
Total trans fatty acids2 15.25 [12.40, 19.16] 0.90
9t-16:1 2.93 [2.24, 3.49]a 0.16
6t-18:1 4.36 [2.42, 5.05]b 0.26
9t-18:1 2.11 [1.40, 2.97]c 0.11
11t-18:1 5.40 [3.79, 6.77]d 0.30
9c,12t-18:2 1.72 [1.38, 1.90]e 0.08
9t,12c-18:2 n.d. n.d.
9t,12t-18:2 n.d. n.d.
Total cis fatty acids3 916.87 [876.43, 1003.23] 51.78
9c-16:1 10.26 [8.58, 15.27]f 0.61
7c-18:1 + 6c-18:1 3.87 [3.07, 5.23]b 0.23
9c-18:1 168.49 [151.91, 182.95]g 8.55
11c-18:1 23.74 [21.58, 26.02]h 0.24
9c,12c-18:2 340.83 [304.29, 374.75]i 19.26
1Values are medians and quartiles [Q1, Q3] (n = 21).
Medians with different superscript letters (a-i) are statistically different (Kruskal-Wallis test followed by
Student-Newman-Keuls, p < 0.05).
n.d. = not detected.
2Total trans fatty acids = 9t-14:1 + 9t-16:1 + 6t-18:1 + 9t-18:1 + 11t-18:1 + 9t,12t-18:2 + 9t,12c-18:2 +
9c,12t-18:2 + 9t,12t,15t-18:3
3Total cis fatty acids = 9c-14:1 + 9c-16:1 + (7c-18:1 + 6c-18:1; coelution) + 9c-18:1 + 11c-18:1 + 12c-
18:1 + 13c-18:1 + 9c,12c-18:2 + 6c,9c,12c-18:3 + 9c,12c,15c-18:3 + 8c-20:1 + 11c-20:1 +
6c,9c,12c,15c-18:4 + 11c,14c-20:2 + 8c,11c,14c-20:3 + 11c,14c,17c-20:3 + 5c,8c,11c,14c-20:4 +
13c-22:1 + 8c,11c,14c,17c-20:4 + 13c,16c-22:2 + 5c,8c,11c,14c,17c-20:5 + 13c,16c,19c-22:3 + 15c-
24:1 + 7c,10c,13c,16c-22:4 + 4c,7c,10c,13c,16c-22:5 + 7c,10c,13c,16c,19c-22:5 +
4c,7c,10c,13c,16c,19c-22:6

77
Table 3. Correlations between F2-isoprostanes and trans fatty acids in plasma of third trimester pregnant women.
9t-16:1 6t-18:1 9t-18:1 11t-18:1 9c,12t-18:2
8-iso-15(R)-PGF2α ns ns ns ns r = 0.394
(p = 0.076)
8-iso-PGF2α ns r = 0.456
(p = 0.038*) ns ns
r = 0.404
(p = 0.068)
15(R)-PGF2α ns r = 0.395
(p = 0.075) ns ns
r = 0.576
(p = 0.006*)
iPF2α-IV ns ns ns ns r = 0.442
(p = 0.045*)
iPF2α-VI ns ns ns ns r = 0.469
(p = 0.032*)
5-iPF2α-VI ns r = 0.419
(p = 0.058)
r = 0.461
(p = 0.035*) ns
r = 0.419
(p = 0.058)
(±)5-8,12-iso-iPF2α-VI ns r = 0.426
(p = 0.053)
r = 0.483
(p = 0.026*) ns
r = 0.397
(p = 0.074)
Values (r) are Spearman correlation coefficients (n=21).
ns = non-significant (p > 0.1).
*p < 0.05 is considered significant. 0.05 < p < 0.1 is considered a tendency.


79
Chapitre 4 – Étude du stress oxydatif en
prééclampsie
4.1. Introduction
La prééclampsie est un syndrome complexe de la grossesse caractérisé par une tension artérielle
élevée (pression diastolique > 90 mm Hg) et par la présence de protéines dans l’urine (protéinurie;
plus de 0.3 g de protéines par jour dans un échantillon d’urine collecté sur 24h) [131]. Elle complique
entre 3 et 5 % des grossesses dans le monde et est une cause majeure de mortalité et de morbidité
maternelle et fœtale [133]. Bien que les causes exactes de ce syndrome sont inconnues, un modèle
en deux étapes a été proposé pour expliquer sa physiopathologie [181]. La placentation inadéquate
entraine le rejet par le placenta de plusieurs facteurs pouvant induire la dysfonction de l’endothélium
maternel, ce qui conduit à l’apparition des symptômes caractéristiques à la prééclampsie après la 20e
semaine de grossesse. Le stress oxydatif semble jouer un rôle central dans l’évolution de ce
désordre [148].
Plusieurs biomarqueurs ont été utilisés afin d’étudier le stress oxydatif dans le contexte de la
prééclampsie et selon ces biomarqueurs, la prééclampsie est associée à un stress oxydatif plus
important qu’ au cours d’une grossesse normale [140]. Depuis quelques années, les F2-isoprostanes,
des produits de la peroxydation de l’acide arachidonique, sont reconnus comme étant les
biomarqueurs les plus fiables du stress oxydatif [88]. Le niveau de 8-iso-PGF2α a été mesuré dans le
placenta, le plasma, l’urine et la salive de femmes ayant une grossesse prééclamptique et comparé à
celui mesuré chez des femmes ayant une grossesse normotensive sans complications [166-171,
173-179]. En général, le niveau de 8-iso-PGF2α libre dans le plasma est plus élevé en prééclampsie
[166, 169-171, 173, 174]. Cependant, les résultats concernant le niveau total (libre + estérifié) de cet
isomère dans le plasma diffèrent d’une étude à l’autre. Une étude a démontré que ce niveau est plus
élevé en prééclampsie alors qu’une autre n’a trouvé aucune différence significative entre les deux
groupes [166, 179]. Enfin, une seule équipe s’est attardée à mesurer un autre F2-isoprostane que le
8-iso-PGF2α. En effet, Regan et al. ont mesuré le 8,12-iso-iPF2α-VI dans des échantillons d’urine de
femmes prééclamptiques et normales obtenus à différents moments de la grossesse, mais ils n’ont
pas trouvés de différences significatives entre les deux groupes [180].

80
Aucune étude n’a mesuré d’autres F2-isoprostanes que le 8-iso-PGF2α dans le plasma, le sang total
et les membranes érythrocytaires en prééclampsie. Le but de la présente étude était donc de
mesurer simultanément par HPLC-MS/MS le niveau total (libre + estérifié aux phospholipides) de
sept isomères de F2-isoprostanes dans ces trois matrices chez des femmes au troisième trimestre
d’une grossesse normale ou prééclamptique.
4.2. Matériel et méthode
4.2.1. Matériel
Le matériel utilisé était le même que celui du chapitre 2 (section 2.4.1, page 39).
4.2.2. Sélection des patientes
Les patientes au troisième trimestre d’une grossesse normale ont été recrutées selon les mêmes
critères que ceux énoncés dans le chapitre 2. Les patientes au troisième trimestre d’une grossesse
prééclamptique ont été sélectionnées selon les critères énoncés dans le rapport de la Canadian
Hypertension Society Consensus Conference (pression diastolique > 90 mm Hg et plus de 0.3 g de
protéines par jour dans un échantillon d’urine collecté sur 24h) [131]. Les patientes prééclamptiques
avec les conditions médicales préexistantes suivantes ont été exclues : obésité, hypertension
chronique, diabète, maladies du rein, maladies inflammatoires chroniques intestinales, troubles de la
coagulation sanguine. Les femmes ayant d’autres complications de la grossesse ont également été
exclues. Les autres facteurs d’exclusions étaient l’âge (< 18 ans ou > 40 ans), la prise de
médicaments anticoagulants ou encore affectant le métabolisme des lipides. Les échantillons ont été
prélevés avant la phase active du travail. Les caractéristiques des patientes au moment du
recrutement ont été publiées dans l’article de Roland et al. [182].
4.2.3. Récolte et traitement des échantillons de sang
Voir le chapitre 2 (section 2.4.3, page 40).

81
4.2.4. Préparation des standards pour l’analyse des F2-isoprostanes
La solution de standard interne, la solution stock et les solutions de travail ont été préparées comme
dans le chapitre 2 (section 2.4.4, page 40). La courbe standard et les contrôles de qualité pour la
mesure des F2-isoprostanes dans le plasma ont été dilués comme ceci : 10 µL de solution de travail,
10 µL de solution de standard interne et 80 µL d’eau contenant 10% (v/v) d’acétonitrile et 0.01 %
(v/v) d’acide acétique. La courbe standard et les contrôles de qualité pour la mesure des F2-
isoprostanes dans les membranes érythrocytaires et le sang total ont été dilués comme ceci : 10 µL
de solution de travail, 6 µL de solution de standard interne et 94 µL d’eau contenant 10% (v/v)
d’acétonitrile et 0.01 % (v/v) d’acide acétique.
4.2.5. Extraction des F2-isoprostanes totaux contenus dans le plasma
Voir le chapitre 2 (section 2.4.5, page 40).
4.2.6. Extraction des membranes érythrocytaires
À 250 µL de sang total a été ajouté 10 µL d’une solution de BHT (1 % dans l’éthanol) et 750 µL de
H2O HPLC. Les échantillons ont été mélangés au vortex et après 5 minutes d’attente, ils ont été
centrifugés à 21 000 g pendant 15 minutes. Ensuite, le surnageant a été jeté et 1 mL d’une solution
de NaCl 0.9 % (m/v) a été ajouté à l’échantillon. Celui-ci a été mélangé de nouveau puis centrifugé à
21 000 g pendant 12 minutes. Ce lavage avec la solution de NaCl a été refait une autre fois. À la fin,
le surnageant a été éliminé et 250 µL d’H2O HPLC ont été ajoutés aux membranes. L’échantillon
final a été congelé à -20°C jusqu’à l’extraction des F2-isoprostanes totaux.
4.2.7. Extraction des F2-isoprostanes totaux contenus dans les membranes érythrocytaires
La méthode d’extraction des F2-isoprostanes totaux contenus dans les membranes érythrocytaires
était très semblable à celle utilisée pour le plasma. Au lieu du 250 µL de plasma, la totalité de
l’échantillon obtenu suite à l’extraction des membranes érythrocytaires a été utilisée. Par contre,
comme la solution de BHT a été ajoutée avant cette extraction, celle-ci n’a pas été rajoutée. De plus,
6 µL de solution de standard interne ont été utilisés au lieu de 10 µL.

82
4.2.8. Extraction des F2-isoprostanes totaux contenus dans le sang total
La méthode d’extraction des F2-isoprostanes totaux contenus dans le sang total était très semblable
à celle utilisée pour le plasma. Par contre, lors de la préparation initiale de l’échantillon, 350 µL d’eau
ont été utilisés au lieu de 250 µL, 6 µL de solution de standard interne au lieu de 10 µL et 150 µL de
sang ont été utilisés au lieu de 250 µL de plasma. Ensuite, l’extraction avec l’hexane n’a été faite
qu’une seule fois au lieu de deux fois pour le plasma. Après la reconstitution de l’échantillon avec
100 µL d’eau contenant 10% (v/v) d’acétonitrile et 0.01 % (v/v) d’acide acétique, celui-ci a été filtré
grâce à des Nanosep MF GHP 0.45 µm (VWR International, Ville Mont-Royal, QC, Canada).
4.2.9. Analyse des F2-isoprostanes par HPLC-MS/MS
L’analyse des F2-isoprostanes par HPLC-MS/MS dans les trois matrices biologiques (plasma,
membranes érythrocytaires et sang total) a été faite selon la même méthode que celle décrite dans le
chapitre 2 (section 2.4.6, page 41 et section 2.4.7, page 41).
4.2.10. Analyses statistiques
Les analyses statistiques ont été faites grâce au logiciel SigmaPlot 12.3 (Systat Software, Inc., 2011,
San Jose, USA). La normalité a été testée selon la méthode Shapiro-Wilk. Les niveaux de F2-isoPs
ont été comparés à l’aide d’un ANOVA (Kruskal-Wallis) suivi du test de Student-Newman-Keuls pour
les comparaisons multiples. La comparaison entre les deux groupes (NOR et PE) pour un isomère
donné a été faite selon le test de Mann-Whitney. P < 0.05 est considéré comme étant significatif et
0.05 < P < 0.1 est considéré comme étant une tendance.
4.3. Résultats
4.3.1. F2-isoprostanes dans le plasma
Les résultats obtenus suite au dosage du niveau total de chacun des sept F2-isoprostanes dans le
plasma des femmes au troisième trimestre d’une grossesse normotensive ou prééclamptique sont
résumés dans le tableau 1 du présent chapitre. Les isomères de la classe VI sont les plus abondants
de tous les isomères dosés par la méthode dans les deux groupes et l’isomère le plus abondant de

83
tous est le (±)5-8,12-iso-iPF2α-VI. Le 8-iso-15(R)-PGF2α, le 8-iso-PGF2α et l’iPF2α-IV sont
significativement moins abondants dans le plasma des femmes prééclamptiques que dans le plasma
des femmes normotensives (P < 0.05). Il n’y a pas de différences significatives entre les deux
groupes pour le iPF2α-VI, le 5-iPF2α-VI et le (±)5-8,12-iso-iPF2α-VI.
Tableau 1. F2-isoprostanes totaux mesurés dans le plasma au troisième trimestre de grossesses normotensives ou prééclamptiques.
F2-isoprostanes (pg/mL plasma)1 P-value2
NOR (n = 22) PE (n = 19)
Classe III 8-iso-15(R)-PGF2α 173 [143, 760]a 144 [122, 162]a 0.012
8-iso-PGF2α 184 [149, 537]a 154 [135, 182]a 0.024
15(R)-PGF2α 322 [250, 1724]a 291 [255, 326]b 0.097
Classe IV iPF2α-IV 132 [98, 957]a 85 [80, 119]c 0.005
Classe VI iPF2α-VI 402 [33, 3885]b 368 [324, 433]d Ns
5-iPF2α-VI 335 [241, 5594]c 285 [232, 387]b Ns
(±)5-8,12-iso-iPF2α-VI 766 [480, 2440]d 608 [535, 833]e Ns
NOR : femmes normotensives; PE : femmes prééclamptiques; Ns : non significatif.
1Les valeurs sont exprimées en médiane et quartiles [Q1, Q3]. Dans une colonne, les médianes avec des lettres différentes en exposant sont statistiquement différentes (Kruskal-Wallis ANOVA à un facteur suivi d’un Student-Newman-Keuls à posteriori pour les comparaisons multiples, P < 0.05).
2Comparaison entre NOR et PE avec le test de Mann-Whitney. P < 0.05 est considéré comme étant significatif et 0.05 < P < 0.1 est considéré comme étant une tendance.
4.3.2. F2-isoprostanes dans le sang
Le tableau 2 du présent chapitre présente le niveau total de chacun des sept F2-isoprostanes dosés
dans le sang entier des femmes au troisième trimestre d’une grossesse normotensive ou
prééclamptique. Comme dans le plasma, les isomères de la classe VI sont les plus abondants de
tous les isomères dosés par la méthode dans les deux groupes. Le 15(R)-PGF2α est également très
abondant dans le sang entier. Il est aussi abondant que le (±)5-8,12-iso-iPF2α-VI dans les deux
groupes (NOR ou PE). Cinq F2-isoprostanes sur sept ont une concentration significativement plus
faible dans le sang entier des femmes prééclamptiques que dans le sang des femmes normotensives
(P < 0.05). Le niveau du 8-iso-PGF2α et le niveau du (±)5-8,12-iso-iPF2α-VI ne diffèrent pas entre les
deux groupes.

84
Tableau 2. F2-isoprostanes totaux mesurés dans le sang entier au troisième trimestre de grossesses normotensives et prééclamptiques.
F2-isoprostanes (pg/mL sang)1 P-value2
NOR (n = 25) PE (n = 25)
Classe III 8-iso-15(R)-PGF2α 606 [537, 803]a 530 [496, 632]a 0.014
8-iso-PGF2α 498 [461, 727]b 500 [456, 543]b Ns
15(R)-PGF2α 1537 [1386, 2417]c 1416 [1309, 1590]c 0.046
Classe IV iPF2α-IV 253 [241, 360]d 235 [216, 270]d 0.007
Classe VI iPF2α-VI 1016 [900, 1317]e 966 [516, 1044]e 0.048
5-iPF2α-VI 1139 [996, 1854]f 1009 [850, 1176]f 0.009
(±)5-8,12-iso-iPF2α-VI 1550 [1375, 2828]c 1520 [1400, 1764]c Ns
NOR : femmes normotensives; PE : femmes prééclamptiques; Ns : non significatif.
1Les valeurs sont exprimées en médiane et quartiles [Q1, Q3]. Dans une colonne, les médianes avec des lettres différentes en exposant sont statistiquement différentes (Kruskal-Wallis ANOVA à un facteur suivi d’un Student-Newman-Keuls à posteriori pour les comparaisons multiples, P < 0.05).
2Comparaison entre NOR et PE avec le test de Mann-Whitney. P < 0.05 est considéré comme étant significatif et 0.05 < P < 0.1 est considéré comme étant une tendance.
4.3.3. F2-isoprostanes dans les membranes érythrocytaires
Le tableau 3 du présent chapitre présente le niveau total de chacun des sept F2-isoprostanes dosés
dans les membranes érythrocytaires des femmes au troisième trimestre d’une grossesse
normotensive ou prééclamptique. Dans cette matrice biologique, la seule différence significative
observée entre les deux groupes est pour le 15(R)-PGF2α, lequel est significativement plus faible en
prééclampsie (P < 0.05). La concentration de deux isomères de la classe VI, c’est-à-dire du 5-iPF2α-
VI et du (±)5-8,12-iso-iPF2α-VI, tend à être plus faible chez les femmes prééclamptiques (0.05 < P <
0.1).

85
Tableau 3. F2-isoprostanes totaux mesurés dans les membranes érythrocytaires au troisième trimestre de grossesses normotensives et prééclamptiques.
F2-isoprostanes (pg/mL sang)1 P-value2
NOR (n = 24) PE (n = 27)
Classe III 8-iso-15(R)-PGF2α 152 [142, 163]a 142 [123, 160]a Ns
8-iso-PGF2α 187 [162, 198]b 175 [160, 202]b Ns
15(R)-PGF2α 332 [307, 363]c 310 [278, 335]c 0.014
Classe IV iPF2α-IV 126 [112, 135]d 122 [96, 134]d Ns
Classe VI iPF2α-VI 239 [219, 254]e 224 [204, 237]e Ns
5-iPF2α-VI 237 [211, 256]e 220 [196, 237]f 0.081
(±)5-8,12-iso-iPF2α-VI 352 [309, 385]c 336 [294, 368]g 0.072
NOR : femmes normotensives; PE : femmes prééclamptiques; Ns : non significatif.
1Les valeurs sont exprimées en médiane et quartiles [Q1, Q3]. Dans une colonne, les médianes avec des lettres différentes en exposant sont statistiquement différentes (Kruskal-Wallis ANOVA à un facteur suivi d’un Student-Newman-Keuls à posteriori pour les comparaisons multiples, P < 0.05).
2Comparaison entre NOR et PE avec le test de Mann-Whitney. P < 0.05 est considéré comme étant significatif et 0.05 < P < 0.1 est considéré comme étant une tendance.
4.4. Discussion
Dans la présente étude, les niveaux totaux (libre + estérifié aux phospholipides) de sept isomères de
F2-isoprostanes ont été mesurés par HPLC-MS/MS dans le plasma, le sang entier et les membranes
érythrocytaires de femmes au troisième trimestre d’une grossesse normale ou prééclamptique. Le 8-
iso-PGF2α est l’isomère le plus utilisé pour étudier le stress oxydatif in vivo. La concentration médiane
du 8-iso-PGF2α mesurée dans les échantillons de plasma provenant des femmes normotensives de
notre cohorte était de 184 pg/mL. Cette valeur est légèrement plus faible que celle de 218 pg/mL
rapportée par Harsem et al. en 2006 pour les femmes normotensives de leur cohorte [183]. Cette
équipe a mesuré le niveau total de 8-iso-PGF2α par GC-MS, une méthode moins sélective que le
HPLC-MS/MS. Cela peut expliquer pourquoi notre mesure est légèrement plus faible [104]. La
concentration médiane du 8-iso-PGF2α dans le sang entier et dans les membranes érythrocytaires
des femmes normotensives de notre cohorte étaient respectivement de 498 pg/mL et 187 pg/mL.
Très peu d’études ont mesuré cet isomère dans ces deux matrices et aucune ne l’a fait dans le cadre
de la grossesse. Bastani et al. ont utilisé la méthode HPLC-MS/MS qu’ils ont mise au point pour

86
mesurer la concentration totale du 8-iso-PGF2α dans le plasma, le sang entier et les membranes
érythrocytaires de sujets en santé. Les concentrations rapportées pour le sang entier varient entre 1
et 2 ng/mL alors que celles rapportées pour les membranes varient entre 0.4 et 1.1 ng/mL. Bien que
ces mesures soient plus élevées que les nôtres, il est difficile de comparer notre étude à la leur
puisqu’ils ne donnent aucun détail sur les sujets ayant fourni les échantillons. Enfin, il n’est pas
possible de comparer avec la littérature nos mesures pour les autres isomères de F2-isoprostanes
dosés par notre méthode à cause du manque de données disponibles pour le plasma, le sang total et
les membranes érythrocytaires.
Le but de la présente étude a été de caractériser le stress oxydatif en prééclampsie. Selon l’isomère
de F2-isoprostane et la matrice biologique étudiée dans notre cohorte, soit il n’y a pas de différence
significative observée entre les femmes normotensives et prééclamptiques, soit les femmes
prééclamptiques ont un niveau significativement plus faible de F2-isoprostanes. Le stress oxydatif
serait donc moins important chez les femmes prééclamptiques que chez les femmes normotensives
dans notre cohorte. Ces résultats sont très étonnants sachant que la prééclampsie a été associée à
un stress oxydatif plus important qu’au cours d’une grossesse normotensive [139]. Les équipes qui
ont mesuré le stress oxydatif grâce au 8-iso-PGF2α libre dans le plasma ont généralement démontré
un niveau plus élevé en prééclampsie [166, 169-171, 173, 174]. Cependant, lorsque c’est le niveau
plasmatique total du 8-iso-PGF2α qui est mesuré, les résultats sont moins concordants. Une étude a
démontré que ce niveau est plus élevé en prééclampsie alors qu’une autre n’a trouvé aucune
différence significative entre les deux groupes [166, 179]. Malgré ces divergences, nos résultats sont
à l’opposé de ce qui a déjà été publié. Cette divergence pourrait provenir d’une réponse antioxydante
différente en prééclampsie spécifique à notre cohorte. En effet, à cause de sa nature hydrophobe, la
vitamine E est retrouvée dans les membranes lipidiques qu’elle protège contre la peroxydation. C’est
ainsi un antioxydant très efficace. La concentration totale de vitamine E a été mesurée dans le
plasma des femmes de notre cohorte [182]. Les femmes prééclamptiques ont une concentration plus
élevée de vitamine E plasmatique que les femmes normotensives. Cette différence pourrait expliquer
pourquoi les F2-isoprostanes sont moins élevés en prééclampsie dans notre cohorte.
En résumé, sept isomères de F2-isoprostanes ont été mesurés dans trois matrices biologiques
provenant de femmes au troisième trimestre d’une grossesse normale ou prééclamptique. Les
concentrations obtenues pour le 8-iso-PGF2α chez les femmes normotensives sont comparables à ce

87
qui a déjà été publié dans la littérature. Cependant, les femmes prééclamptiques ont un niveau plus
faible de F2-isoprostanes que les femmes normotensives et ce peu importe la matrice biologique
étudiée. Comme la vitamine E totale plasmatique est augmentée en prééclampsie dans notre
cohorte, il est probable qu’un mécanisme compensatoire ait été mis en place pour diminuer le stress
oxydatif associé à ce syndrome [182].
4.5. Bibliographie
Voir la bibliographie générale à la page 97.


89
Chapitre 5 – Discussion générale
Les dérivés actifs de l’oxygène (ROS) jouent des rôles essentiels dans les conditions physiologiques
normales. En effet, ils agissent comme seconds messagers dans différentes voies de signalisation
[2, 3] et ils sont impliqués dans la défense immunitaire [4]. Des antioxydants enzymatiques et non
enzymatiques contrôlent la concentration de ces molécules hautement réactives afin de limiter les
dommages qu’elles pourraient causer aux biomolécules environnantes. Cependant, lorsque les
défenses antioxydantes de l’organisme ne sont pas suffisantes pour accomplir cette tâche ou lorsqu’il
y a surproduction de ROS par rapport à celles-ci, l’organisme subit un stress oxydatif. Le stress
oxydatif a été associé à la physiopathologie de nombreuses maladies (ex. : maladies
cardiovasculaires, maladies inflammatoires, maladies pulmonaires, maladies neurodégénératives,
prééclampsie, etc.) [8]. Il est également impliqué dans le processus du vieillissement ainsi que dans
certaines situations physiologiques normales comme la grossesse [6, 7].
Afin d’étudier le stress oxydatif in vivo, l’usage de biomarqueurs est essentiel. Pendant longtemps, le
biomarqueur le plus utilisé a été le malondialdehyde (MDA). Depuis quelques années, les F2-
isoprostanes sont reconnus comme étant les biomarqueurs les plus fiables du stress oxydatif [88]. Il
existe 64 isomères de F2-isoprostanes issus de la peroxydation de l’acide arachidonique estérifié aux
phospholipides. L’isomère le plus étudié jusqu’à maintenant est le 8-iso-PGF2α. Ce dernier a été
mesuré dans de nombreuses matrices biologiques afin de comprendre l’implication du stress oxydatif
dans la physiopathologie de nombreuses maladies [8]. Son métabolisme ainsi que ses effets
biologiques ont également été étudiés [99, 111-117]. Cependant, très peu d’informations sont
disponibles au sujet des autres isomères de F2-isoprostanes. Ceux-ci peuvent avoir un métabolisme
différent du 8-iso-PGF2α et ce métabolisme peut différer selon la maladie étudiée. L'analyse
simultanée de plusieurs isomères de F2-isoprostanes permettrait de mieux caractériser le stress
oxydatif associé à des pathologies spécifiques.
Dans le cadre de mon projet de maîtrise, j’ai développé et validé une méthode d’analyse par
chromatographie liquide couplée à la spectrométrie de masse en tandem (HPLC-MS/MS) permettant
le dosage simultané de plusieurs F2-isoprostanes. Cette technique offre de nombreux avantages par
rapport aux autres techniques couramment utilisées pour le dosage de ces molécules.

90
Premièrement, le HPLC-MS/MS offre une meilleure spécificité que la chromatographie gazeuse
couplée à la spectrométrie de masse (GC-MS) pour l’analyse de ces molécules. En effet, bien que
les méthodes utilisant le GC-MS soient très sensibles, celles-ci ne permettent pas d’analyser
séparément les quatre classes de F2-isoprostanes. Tous les F2-isoprostanes ont la même masse
moléculaire et cela les empêche d’être discriminés dans un spectromètre de masse avec un seul
filtre à ions (simple MS). Certaines équipes ont mis au point des techniques utilisant la
chromatographie gazeuse couplée à la spectrométrie de masse en tandem (GC-MS/MS) pour pallier
ce problème [105, 106]. En effet, les quatre classes de F2-isoprostanes ont un comportement
différent dans un tel instrument (MS/MS). La spectrométrie de masse en tandem consiste à
sélectionner un ion dans un premier temps, à le fragmenter dans une cellule de collision, puis à
sélectionner et analyser un des fragments générés. Tous les F2-isoprostanes ont le même
comportement dans le premier filtre à ions, mais les quatre classes fragmentent différemment dans la
cellule de collision. Cette propriété permet ainsi de générer des fragments spécifiques aux quatre
classes de régioisomères de F2-isoPs. Cependant, même si l’utilisation d’un spectromètre de masse
en tandem améliore la spécificité de l’analyse, la chromatographie gazeuse ne parvient pas à
résoudre certains isomères à l’intérieur d’une même classe. Ainsi, le 8-iso-PGF2α et le 8-iso-15(R)-
PGF2α coéluent si le GC-MS est utilisé alors que ces deux isomères sont facilement résolus par
HPLC-MS/MS [107, 184]. Un deuxième avantage de l’utilisation du HPLC-MS/MS pour le dosage des
F2-isoprostanes est que le traitement des échantillons est beaucoup plus simple et rapide que celui
nécessaire pour à une analyse GC-MS. Pour être analysées pas GC-MS, les molécules doivent être
volatiles, ce qui n’est pas le cas des F2-isoprostanes. Une dérivation chimique est donc essentielle
pour rendre ces molécules volatiles. Finalement, les F2-isoprostanes peuvent être mesurés grâce à
des méthodes immuno-enzymatiques (ELISA) [109] et radio-immunologiques (RIA) [110]. Ces
méthodes ne nécessitent pas l’utilisation d’instruments dispendieux comme le HPLC-MS/MS, mais
elles ne permettent le dosage de plusieurs isomères de F2-isoprostanes et leur spécificité pour
l’isomère dosé est couramment remise en doute à cause des réactions croisées possibles des
anticorps avec des molécules apparentées [107].
La chromatographie liquide couplée à la spectrométrie de masse en tandem a recourt à deux
procédés pour le dosage spécifique des F2-isoprostanes. Comme expliqué précédemment, le
spectromètre de masse en tandem permet l’analyse séparée des quatre classes de F2-isoprostanes.
La colonne chromatographique permet quant à elle de séparer les isomères à l’intérieur d’une même

91
classe. Afin de développer la méthode HPLC-MS/MS la plus spécifique possible pour le dosage
simultané de plusieurs F2-isoprostanes, j’ai testé plusieurs colonnes dont des colonnes de type C8 et
C18. Plus particulièrement, j’ai comparé deux types de colonne C18 ayant les mêmes dimensions
(diamètre interne: 3 mm, longueur : 100 mm). La différence principale entre celles-ci se situe au
niveau des particules. La colonne Shimpack XR-ODS est remplie de particules totalement poreuses
ayant un diamètre de 2.2 µm alors que la colonne Kinetex XB-C18 est remplie de particules
partiellement poreuses ayant un diamètre de 2.6 µm. Les particules partiellement poreuses rendent
la colonne Kinetex beaucoup plus efficace que la colonne Shimpack [185]. Plus une colonne est
efficace, plus elle est capable de résoudre deux composés ayant des propriétés chromatographiques
semblables. Ainsi l’utilisation de la Kinetex a permis la résolution du iPF2α-VI et du 5-iPF2α-VI alors
que ces deux composés coéluent si la colonne Shimpack est utilisée. Le 5-trans-PGF2α a également
pu être séparé du 15(R)-PGF2α. J’ai donc utilisé la colonne Kinetex XB-C18 (3 x 100 mm, 2.6 µm)
pour optimiser la chromatographie des F2-isoprostanes et la méthode d’analyse finale est décrite
dans le chapitre 2. De nombreuses méthodes ont déjà été publiées pour l’analyse des F2-
isoprostanes par HPLC-MS/MS, mais celle que j’ai mise au point est la première à utiliser une
colonne remplie avec des particules partiellement poreuses [107, 186-189]. L’efficacité de la colonne
Kinetex est telle qu’il serait possible d’utiliser une version plus courte de celle-ci tout en conservant
une résolution acceptable des F2-isoprostanes. Cela aurait pour effet d’augmenter la sensibilité et la
rapidité de la méthode d’analyse.
Le deuxième objectif qui a été fixé dans le cadre de mon projet de maîtrise était de mesurer les F2-
isoprostanes totaux (estérifiés aux phospholipides + libres) dans le plasma de femmes au troisième
trimestre d’une grossesse normale à l’aide de la méthode HPLC-MS/MS que j’ai mise au point. Le
but était de s’assurer du bon fonctionnement général de la méthode. Les résultats figurent dans le
chapitre 2. J’ai ensuite voulu étudier la relation entre les F2-isoprostanes et les acides gras trans
mesurés dans les mêmes échantillons de plasma. En effet, les acides gras trans peuvent provoquer
un stress oxydatif in vivo et le stress oxydatif peut induire la formation d’acides gras trans [22, 69].
Les résultats de cette étude figurent dans le chapitre 3. Des corrélations positives entre certains F2-
isoprostanes et certains acides gras trans ont été détectées dans le plasma de femmes ayant eu une
grossesse sans complications, ce qui confirme notre hypothèse de départ.

92
Les gras trans sont néfastes pour la santé. De nombreuses études ont démontré que la
consommation d’une grande quantité de TFA augmente les risques de maladies cardiovasculaires
[62-68]. Ils altèrent le profil lipidique du sang, ils modifient le niveau d’apolipoprotéines, ils provoquent
une réponse inflammatoire systémique ainsi que la dysfonction de l’endothélium. De plus, ils sont
susceptibles d’induire un stress oxydatif [69]. Certains groupes se sont également intéressés à
connaître l’impact des gras trans sur la grossesse. Une étude publiée en 1998 a démontré qu’une
concentration plus élevée d’acide élaïdique (9t-18:1) dans les membranes érythrocytaires est
associée à un risque plus grand de prééclampsie [190]. Cette étude a été faite sur une petite cohorte
et les résultats obtenus seraient à confirmer. Une autre équipe a démontré en 2007 qu’une plus
grande consommation de TFA au cours du premier trimestre de grossesse n’augmente pas le risque
de développer de l’hypertension gestationnelle ou une prééclampsie [191]. Enfin, le risque de faire du
diabète gestationnel n’est pas affecté par une plus grande consommation de TFA avant ou pendant
le premier trimestre de grossesse [192, 193]. Bref, les gras trans ne semblent pas avoir beaucoup
d’impacts sur la grossesse même si leur niveau est inversement relié à la durée de la gestation [194].
Par contre, il a été démontré qu’ils ont un impact négatif sur le développement et la croissance du
fœtus. En effet, il existe une relation inverse entre l’exposition du fœtus aux TFA et son poids à la
naissance [195, 196]. Un bébé ayant un petit poids à la naissance est plus à risque de développer
des maladies cardiovasculaires et du diabète à l’âge adulte. Il est aussi plus à risque de devenir
obèse [197-201]. De plus, une grande concentration de TFA dans les phospholipides des
membranes érythrocytaires maternelles est associée à une diminution de la concentration d’acide
docosahexaénoïque (DHA) et d’acide arachidonique (AA) dans ces mêmes lipides, deux acides gras
clés dans le développement du fœtus [202]. Les TFA inhibent aussi la conversion de l’acide
linoléique (LA) en AA et celle de l’acide alpha linolénique (ALA) en DHA [203]. Le DHA est
particulièrement important pour le développement du système nerveux central du fœtus [204, 205].
D’ailleurs, les nouveau-nés ayant un niveau sanguin de DHA relativement faible et ceux qui ont des
niveaux élevés d'acide gras trans ont une condition neurologique moins favorable à 18 mois [206].
Le dernier objectif qui a été fixé dans le cadre de mon projet de maîtrise a été de caractériser le
stress oxydatif en prééclampsie. Dans ce but, le niveau total (libre + estérifié aux phospholipides) de
chacun des sept isomères de F2-isoprostanes pouvant être mesurés par la méthode que j’ai mise au
point a été dosé dans le plasma, le sang total et les membranes érythrocytaires de femmes au
troisième trimestre d’une grossesse normale ou prééclamptique. Les résultats de cette étude se

93
retrouvent dans le chapitre 4. Selon l’isomère de F2-isoprostane et la matrice biologique étudiée dans
notre cohorte, soit il n’y a pas de différence significative observée entre les femmes normotensives et
prééclamptiques, soit les femmes prééclamptiques ont un niveau significativement plus faible de F2-
isoprostanes. Cela prouve l’importance de mesurer plus d’un isomère de F2-isoprostanes à la fois
sans quoi il serait possible de mal évaluer le stress oxydatif dans certaines situations. Je pense que
les isomères de la classe VI feraient de meilleurs biomarqueurs du stress oxydatif. En effet, ceux-ci
sont plus abondants que les isomères des autres classes et ils sont plus facilement détectables. Pour
une concentration égale, le ratio signal sur bruit pour ces composés est plus grand que pour les
isomères de la classe III dans la méthode HPLC-MS/MS que j’ai développée. Ce ratio est
directement lié à la sensibilité de la méthode d’analyse. De plus, les isomères de la classe VI ne
peuvent pas être synthétisés par la voie des cyclooxygénases alors que cette voie contribue à la
production d’une très faible quantité de 8-iso-PGF2α [207].
Contre toutes attentes, dans notre cohorte, les femmes prééclamptiques ont un niveau plus faible de
F2-isoprostanes que les femmes normotensives. Le contraire aurait été anticipé [140]. Le niveau plus
élevé de vitamine E dans le plasma des femmes prééclamptiques de notre cohorte pourrait expliquer
en partie ces résultats, la vitamine E étant un antioxydant membranaire très efficace. Outre les F2-
isoprostanes, il existe d’autres produits issus de la peroxydation de l’acide arachidonique qui
pourraient servir comme biomarqueurs du stress oxydatif dans certaines situations particulières.
C’est le cas des isofuranes (isoFs) dont la découverte a été rapportée récemment [208]. Comme les
F2-isoprostanes, ce sont des composés chimiquement stables détectables dans tous les tissus et
liquides biologiques et leur concentration augmente en présence de stress oxydatif. Il existe 256
isofuranes possibles divisés en huit classes de régioisomères composées chacune de seize
diastéréoisomères racémiques [209]. Leur formation est en compétition avec celle des F2-
isoprostanes et plus la concentration en oxygène moléculaire est élevée, plus la formation des
isofuranes est favorisée par rapport à celle des F2-isoprostanes (Figure 1). D’ailleurs, lorsque la
concentration en oxygène varie entre 1 % et 100 %, la peroxydation in vitro de l’acide arachidonique
favorise la formation des isofuranes plutôt que celle des F2-isoprostanes. Chez les rats exposés au
tétrachlorométhane (CCl4), un inducteur de stress oxydatif, le rapport isoFs/F2-isoPs varie en fonction
du degré d’oxygénation du tissu à l’étude [208]. Dans les organes très oxygénés comme le cerveau
et les reins, les isofuranes sont plus abondants alors que dans le foie, un organe faiblement oxygéné,
ce sont les F2-isoprostanes les plus abondants.

94
Figure 1. Compétition entre la formation des F2-isoprostanes et des isofuranes. Tirée de [210].
Il a été proposé que la mesure combinée des isofuranes et des F2-isoprostanes pourrait fournir un
meilleur indice du stress oxydatif que la mesure d’un seul d’entre eux [208, 209]. Jusqu’à maintenant,
ces deux types de molécules sont quantifiés en même temps à l’aide de la même méthode GC-MS
qui avait déjà été mise au point pour le dosage des F2-isoprostanes. Une équipe australienne a
utilisé ce type de méthode pour mesurer le niveau total des F2-isoprostanes et des isofuranes dans le
plasma de femmes au troisième trimestre d’une grossesse normale ou prééclamptique [211]. Ils ont
observé que les isofuranes étaient augmentés en prééclampsie, mais pas les F2-isoprostanes. Il
serait intéressant de mesurer les isofuranes dans notre cohorte afin de déterminer si ceux-ci sont
augmentés en prééclampsie même si nous avons observé une diminution des F2-isoprostanes. Un
tel phénomène a été observé dans un contexte complètement différent. Mas et al. a comparé les

95
niveaux plasmatiques d’isofuranes et de F2-isoprostanes chez les patients subissant une chirurgie de
remplacement du genou sous anesthésie rachidienne ou générale [210]. Les F2-isoprostanes étaient
plus faibles et les isofuranes étaient plus abondants chez les patients ayant subi une anesthésie
générale en comparaison avec ceux ayant subi une anesthésie rachidienne.
En conclusion, j’ai développé et validé une méthode pour le dosage de sept isomères de F2-
isoprostanes par HPLC-MS/MS que j’ai utilisée afin d’étudier le stress oxydatif dans le plasma, le
sang total et les membranes érythrocytaires de femmes au troisième trimestre d’une grossesse
normale ou prééclamptique. J’ai démontré l’utilité du dosage des isomères de classe VI pour
caractérisation du stress oxydatif. De plus, j’ai lié les niveaux de certains gras trans avec les F2-
isoprostanes. Ceci constitue la première démonstration in vivo de la compétition entre la formation
des F2-isoprostanes et des gras trans.
En perspectives, ma méthode pourra maintenant être utilisée pour mesurer le stress oxydatif dans
d’autres contextes (ex. : diabète, obésité, maladies cardiovasculaires, autres pathologies de la
grossesse, etc.). Certaines améliorations pourront aussi être apportées. Par exemple, l’utilisation
d’une colonne chromatographique plus courte permettrait d’augmenter la sensibilité et la rapidité de
l’analyse. Dans ce même but, il serait également possible de modifier le traitement des échantillons.
En effet, la technique d’extraction liquide-liquide utilisée actuellement au laboratoire est laborieuse et
le rendement d’extraction pourrait être amélioré. Une méthode d’extraction en phase solide
permettrait l’automatisation du traitement des échantillons et ainsi un plus grand nombre
d’échantillons pourraient être analysés chaque jour. Enfin, il faudra inclure l’analyse des isofuranes
dans la méthode que j’ai mise au point afin de mieux caractériser le stress oxydatif en fonction des
organes et des états pathologiques étudiés.


97
Bibliographie
1. Reis, A. and C.M. Spickett, Chemistry of phospholipid oxidation. Biochim Biophys Acta, 2012. 1818(10): p. 2374-87.
2. Droge, W., Free radicals in the physiological control of cell function. Physiological reviews, 2002. 82(1): p. 47-95.
3. Finkel, T., Signal transduction by reactive oxygen species. The Journal of cell biology, 2011. 194(1): p. 7-15.
4. Robinson, J.M., Phagocytic leukocytes and reactive oxygen species. Histochemistry and cell biology, 2009. 131(4): p. 465-9.
5. Burton, G.J. and E. Jauniaux, Oxidative stress. Best practice & research. Clinical obstetrics & gynaecology, 2011. 25(3): p. 287-99.
6. Agarwal, A., S. Gupta, and S. Sikka, The role of free radicals and antioxidants in reproduction. Curr Opin Obstet Gynecol, 2006. 18(3): p. 325-32.
7. Jauniaux, E., et al., Onset of maternal arterial blood flow and placental oxidative stress. A possible factor in human early pregnancy failure. Am J Pathol, 2000. 157(6): p. 2111-22.
8. Basu, S., F2-isoprostanes in human health and diseases: from molecular mechanisms to clinical implications. Antioxid Redox Signal, 2008. 10(8): p. 1405-34.
9. Milne, G.L., et al., Isoprostane generation and function. Chem Rev, 2011. 111(10): p. 5973-96.
10. Cadenas, E. and K.J. Davies, Mitochondrial free radical generation, oxidative stress, and aging. Free Radic Biol Med, 2000. 29(3-4): p. 222-30.
11. Drose, S. and U. Brandt, Molecular mechanisms of superoxide production by the mitochondrial respiratory chain. Adv Exp Med Biol, 2012. 748: p. 145-69.
12. Muller, F.L., Y. Liu, and H. Van Remmen, Complex III releases superoxide to both sides of the inner mitochondrial membrane. The Journal of biological chemistry, 2004. 279(47): p. 49064-73.
13. Tu, B.P. and J.S. Weissman, Oxidative protein folding in eukaryotes: mechanisms and consequences. The Journal of cell biology, 2004. 164(3): p. 341-6.
14. Lambeth, J.D., NOX enzymes and the biology of reactive oxygen. Nature reviews. Immunology, 2004. 4(3): p. 181-9.
15. Schrader, M. and H.D. Fahimi, Peroxisomes and oxidative stress. Biochim Biophys Acta, 2006. 1763(12): p. 1755-66.
16. Hauptmann, N., et al., The metabolism of tyramine by monoamine oxidase A/B causes oxidative damage to mitochondrial DNA. Arch Biochem Biophys, 1996. 335(2): p. 295-304.
17. Giorgio, M., et al., Electron transfer between cytochrome c and p66Shc generates reactive oxygen species that trigger mitochondrial apoptosis. Cell, 2005. 122(2): p. 221-33.
18. Jomova, K. and M. Valko, Advances in metal-induced oxidative stress and human disease. Toxicology, 2011. 283(2-3): p. 65-87.
19. Pacher, P., J.S. Beckman, and L. Liaudet, Nitric oxide and peroxynitrite in health and disease. Physiological reviews, 2007. 87(1): p. 315-424.
20. Nordberg, J. and E.S. Arner, Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic Biol Med, 2001. 31(11): p. 1287-312.
21. Porter, N.A., S.E. Caldwell, and K.A. Mills, Mechanisms of free radical oxidation of unsaturated lipids. Lipids, 1995. 30(4): p. 277-90.

98
22. Mihaljevic, B., et al., Linoleic acid peroxidation vs. isomerization: a biomimetic model of free radical reactivity in the presence of thiols. Organic & biomolecular chemistry, 2011. 9(9): p. 3541-8.
23. Yu, B.P., Cellular defenses against damage from reactive oxygen species. Physiol Rev, 1994. 74(1): p. 139-62.
24. Perkins, A.V., Endogenous anti-oxidants in pregnancy and preeclampsia. Aust N Z J Obstet Gynaecol, 2006. 46(2): p. 77-83.
25. Imai, H. and Y. Nakagawa, Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radic Biol Med, 2003. 34(2): p. 145-69.
26. Hayes, J.D., J.U. Flanagan, and I.R. Jowsey, Glutathione transferases. Annu Rev Pharmacol Toxicol, 2005. 45: p. 51-88.
27. Kamal-Eldin, A. and L.A. Appelqvist, The chemistry and antioxidant properties of tocopherols and tocotrienols. Lipids, 1996. 31(7): p. 671-701.
28. Mustacich, D.J., R.S. Bruno, and M.G. Traber, Vitamin E. Vitamin E: Vitamins and Hormones Advances in Research and Applications, 2007. 76: p. 1-21.
29. O'Byrne, D., et al., Studies of LDL oxidation following alpha-, gamma-, or delta-tocotrienyl acetate supplementation of hypercholesterolemic humans. Free Radic Biol Med, 2000. 29(9): p. 834-45.
30. Buettner, G.R., The pecking order of free radicals and antioxidants: lipid peroxidation, alpha-tocopherol, and ascorbate. Arch Biochem Biophys, 1993. 300(2): p. 535-43.
31. Traber, M.G. and J.F. Stevens, Vitamins C and E: beneficial effects from a mechanistic perspective. Free Radic Biol Med, 2011. 51(5): p. 1000-13.
32. Fahy, E., et al., Lipid classification, structures and tools. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids, 2011. 1811(11): p. 637-647.
33. Bhagavan, N.V. and C.-E. Ha, Chapter 16 - Lipids I: Fatty Acids and Eicosanoids, in Essentials of Medical Biochemistry. 2011, Academic Press: San Diego. p. 191-207.
34. Last, A.R. and S.A. Wilson, Low-carbohydrate diets. Am Fam Physician, 2006. 73(11): p. 1942-8.
35. Bhagavan, N.V. and C.-E. Ha, Chapter 11 - Gastrointestinal Digestion and Absorption, in Essentials of Medical Biochemistry. 2011, Academic Press: San Diego. p. 97-113.
36. Kindel, T., D.M. Lee, and P. Tso, The mechanism of the formation and secretion of chylomicrons. Atherosclerosis Supplements, 2010. 11(1): p. 11-16.
37. Andersson, L., B. Sternby, and A. Nilsson, Hydrolysis of phosphatidylethanolamine by human pancreatic phospholipase A2. Effect of bile salts. Scand J Gastroenterol, 1994. 29(2): p. 182-7.
38. Ponz de Leon, M., et al., Cholesterol esterase activity of human intestinal mucosa. Dig Dis Sci, 1985. 30(11): p. 1053-64.
39. Heidrich, J., et al., Inhibition of pancreatic cholesterol esterase reduces cholesterol absorption in the hamster. BMC Pharmacology, 2004. 4(1): p. 5.
40. Myers-Payne, S.C., et al., Cholesterol Esterase: A Cholesterol Transfer Protein. Biochemistry, 1995. 34(12): p. 3942-3947.
41. Bhagavan, N.V. and C.-E. Ha, Chapter 18 - Lipids III: Plasma Lipoproteins, in Essentials of Medical Biochemistry. 2011, Academic Press: San Diego. p. 225-239.
42. Jonas, A., Lipoprotein structure, in New Comprehensive Biochemistry, J.E.V. Dennis E. Vance, Editor. 2002, Elsevier. p. 483-504.

99
43. Ordovas, J.M., LIPOPROTEINS, in Encyclopedia of Human Nutrition (Second Edition), C. Editor-in-Chief: Benjamin, Editor. 1998, Elsevier: Oxford. p. 143-150.
44. Deckelbaum, R.J., et al., Triacylglycerol and phospholipid hydrolysis in human plasma lipoproteins: role of lipoprotein and hepatic lipase. Biochemistry, 1992. 31(36): p. 8544-8551.
45. Da Costa, T.H.M., FATS | Digestion, Absorption, and Transport, in Encyclopedia of Food Sciences and Nutrition (Second Edition), C. Editor-in-Chief: Benjamin, Editor. 2003, Academic Press: Oxford. p. 2274-2278.
46. Buczynski, M.W., D.S. Dumlao, and E.A. Dennis, Thematic Review Series: Proteomics. An integrated omics analysis of eicosanoid biology. Journal of Lipid Research, 2009. 50(6): p. 1015-1038.
47. Funk, C.D., Prostaglandins and Leukotrienes: Advances in Eicosanoid Biology. Science, 2001. 294(5548): p. 1871-1875.
48. Zhou, L. and A. Nilsson, Sources of eicosanoid precursor fatty acid pools in tissues. J Lipid Res, 2001. 42(10): p. 1521-42.
49. Mann, N.J., et al., The arachidonic acid content of the Australian diet is lower than previously estimated. J Nutr, 1995. 125(10): p. 2528-35.
50. Nakamura, M.T. and T.Y. Nara, Essential fatty acid synthesis and its regulation in mammals. Prostaglandins, Leukotrienes and Essential Fatty Acids, 2003. 68(2): p. 145-150.
51. Emken, E.A., et al., Effect of dietary arachidonic acid on metabolism of deuterated linoleic acid by adult male subjects. Lipids, 1998. 33(5): p. 471-80.
52. Thies, F., et al., Unsaturated fatty acids esterified in 2-acyl-l-lysophosphatidylcholine bound to albumin are more efficiently taken up by the young rat brain than the unesterified form. J Neurochem, 1992. 59(3): p. 1110-6.
53. Hodson, L., C.M. Skeaff, and B.A. Fielding, Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Progress in Lipid Research, 2008. 47(5): p. 348-380.
54. Yamashita, A., T. Sugiura, and K. Waku, Acyltransferases and Transacylases Involved in Fatty Acid Remodeling of Phospholipids and Metabolism of Bioactive Lipids in Mammalian Cells. Journal of Biochemistry, 1997. 122(1): p. 1-16.
55. Leslie, C.C., Regulation of arachidonic acid availability for eicosanoid production. Biochem Cell Biol, 2004. 82(1): p. 1-17.
56. Perez-Chacon, G., et al., Control of free arachidonic acid levels by phospholipases A2 and lysophospholipid acyltransferases. Biochim Biophys Acta, 2009. 1791(12): p. 1103-13.
57. List, G.R., D. Kritchevsky, and W.M.N. Ratnayake, Trans Fats in Foods. 2007, AOCS Press. p. 97-126.
58. Ratnayake, W.M., et al., Trans fatty acids: current contents in Canadian foods and estimated intake levels for the Canadian population. Journal of AOAC International, 2009. 92(5): p. 1258-76.
59. Precht, D. and J. Molkentin, Trans fatty acids: Implications for health, analytical methods, incidence in edible fats and intake. Food / Nahrung, 1995. 39(5-6): p. 343-374.
60. Mendis, S., C. Cruz-Hernandez, and W.M. Ratnayake, Fatty acid profile of Canadian dairy products with special attention to the trans-octadecenoic acid and conjugated linoleic acid isomers. Journal of AOAC International, 2008. 91(4): p. 811-9.
61. Wolff, R., et al., Follow-up of the Δ4 to Δ16 trans-18:1 isomer profile and content in French processed foods containing partially hydrogenated vegetable oils during the period 1995–1999. Analytical and nutritional implications. Lipids, 2000. 35(8): p. 815-825.

100
62. Micha, R. and D. Mozaffarian, Trans fatty acids: effects on metabolic syndrome, heart disease and diabetes. Nat Rev Endocrinol, 2009. 5(6): p. 335-44.
63. Mozaffarian, D., Trans fatty acids - effects on systemic inflammation and endothelial function. Atheroscler Suppl, 2006. 7(2): p. 29-32.
64. Mozaffarian, D., A. Aro, and W.C. Willett, Health effects of trans-fatty acids: experimental and observational evidence. European journal of clinical nutrition, 2009. 63 Suppl 2: p. S5-21.
65. Mozaffarian, D., et al., Trans fatty acids and cardiovascular disease. The New England journal of medicine, 2006. 354(15): p. 1601-13.
66. Brouwer, I.A., A.J. Wanders, and M.B. Katan, Effect of animal and industrial trans fatty acids on HDL and LDL cholesterol levels in humans--a quantitative review. PloS one, 2010. 5(3): p. e9434.
67. Gebauer, S.K., T.L. Psota, and P.M. Kris-Etherton, The diversity of health effects of individual trans fatty acid isomers. Lipids, 2007. 42(9): p. 787-99.
68. Lopez-Garcia, E., et al., Consumption of trans fatty acids is related to plasma biomarkers of inflammation and endothelial dysfunction. J Nutr, 2005. 135(3): p. 562-6.
69. Smit, L.A., et al., A high intake of trans fatty acids has little effect on markers of inflammation and oxidative stress in humans. J Nutr, 2011. 141(9): p. 1673-8.
70. Kuhnt, K., et al., Dietary supplementation with 11trans- and 12trans-18:1 and oxidative stress in humans. The American journal of clinical nutrition, 2006. 84(5): p. 981-8.
71. Turpeinen, A.M., et al., Bioconversion of vaccenic acid to conjugated linoleic acid in humans. The American Journal of Clinical Nutrition, 2002. 76(3): p. 504-510.
72. Motard-Belanger, A., et al., Study of the effect of trans fatty acids from ruminants on blood lipids and other risk factors for cardiovascular disease. The American journal of clinical nutrition, 2008. 87(3): p. 593-9.
73. Lacroix, E., et al., Randomized controlled study of the effect of a butter naturally enriched in trans fatty acids on blood lipids in healthy women. The American journal of clinical nutrition, 2012. 95(2): p. 318-25.
74. Mozaffarian, D., et al., Dietary intake of trans fatty acids and systemic inflammation in women. Am J Clin Nutr, 2004. 79(4): p. 606-12.
75. Groupe d’étude sur les graisses trans. TRANSformer l’approvisionnement alimentaire. 2006 2013/01/22]; Available from: http://www.hc-sc.gc.ca/fn-an/nutrition/gras-trans-fats/tf-ge/tf-gt_rep-rap-fra.php.
76. Davies, A.G., et al., An Electron-Spin Resonance Study of Pentadienyl and Related Radicals - Homolytic Fission of Cyclobut-2-Enylmethyl Radicals. Journal of the Chemical Society-Perkin Transactions 2, 1981(4): p. 633-641.
77. Xu, L., T.A. Davis, and N.A. Porter, Rate constants for peroxidation of polyunsaturated fatty acids and sterols in solution and in liposomes. J Am Chem Soc, 2009. 131(36): p. 13037-44.
78. Zambonin, L., et al., Effect of radical stress and ageing on the occurrence of trans fatty acids in rats fed a trans-free diet. Free Radic Biol Med, 2008. 44(4): p. 594-601.
79. Ferreri, C. and C. Chatgilialoglu, Geometrical trans lipid isomers: a new target for lipidomics. Chembiochem, 2005. 6(10): p. 1722-34.
80. Jiang, H., et al., Nitrogen dioxide induces cis-trans-isomerization of arachidonic acid within cellular phospholipids. Detection of trans-arachidonic acids in vivo. J Biol Chem, 1999. 274(23): p. 16235-41.
81. Zghibeh, C.M., et al., Determination of trans-arachidonic acid isomers in human blood plasma. Analytical biochemistry, 2004. 332(1): p. 137-44.

101
82. Chatgilialoglu, C. and C. Ferreri, Trans lipids: the free radical path. Acc Chem Res, 2005. 38(6): p. 441-8.
83. Chatgilialoglu, C., et al., Geometrical isomerism of monounsaturated fatty acids: thiyl radical catalysis and influence of antioxidant vitamins. Free Radic Biol Med, 2002. 33(12): p. 1681-92.
84. Knight, J.A., R.K. Pieper, and L. McClellan, Specificity of the thiobarbituric acid reaction: its use in studies of lipid peroxidation. Clin Chem, 1988. 34(12): p. 2433-8.
85. Michel, F., et al., [Biomarkers of lipid peroxidation: analytical aspects]. Ann Biol Clin (Paris), 2008. 66(6): p. 605-20.
86. Janero, D.R., Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic Biol Med, 1990. 9(6): p. 515-40.
87. Morrow, J.D., et al., A series of prostaglandin F2-like compounds are produced in vivo in humans by a non-cyclooxygenase, free radical-catalyzed mechanism. Proc Natl Acad Sci U S A, 1990. 87(23): p. 9383-7.
88. Kadiiska, M.B., et al., Biomarkers of oxidative stress study II: are oxidation products of lipids, proteins, and DNA markers of CCl4 poisoning? Free radical biology & medicine, 2005. 38(6): p. 698-710.
89. Yin, H. and N.A. Porter, New insights regarding the autoxidation of polyunsaturated fatty acids. Antioxid Redox Signal, 2005. 7(1-2): p. 170-84.
90. Morrow, J.D., et al., Non-cyclooxygenase-derived prostanoids (F2-isoprostanes) are formed in situ on phospholipids. Proc Natl Acad Sci U S A, 1992. 89(22): p. 10721-5.
91. Yin, H., et al., Formation of isoprostane bicyclic endoperoxides from the autoxidation of cholesteryl arachidonate. J Am Chem Soc, 2002. 124(26): p. 7745-54.
92. Milne, G.L., H. Yin, and J.D. Morrow, Human biochemistry of the isoprostane pathway. J Biol Chem, 2008. 283(23): p. 15533-7.
93. Yin, H., J.D. Morrow, and N.A. Porter, Identification of a novel class of endoperoxides from arachidonate autoxidation. J Biol Chem, 2004. 279(5): p. 3766-76.
94. Li, H., et al., Quantitative high performance liquid chromatography/tandem mass spectrometric analysis of the four classes of F(2)-isoprostanes in human urine. Proc Natl Acad Sci U S A, 1999. 96(23): p. 13381-6.
95. van Kuijk, F.J.G.M., et al., A new role for phospholipase A2: protection of membranes from lipid peroxidation damage. Trends in Biochemical Sciences, 1987. 12(0): p. 31-34.
96. Sevanian, A. and E. Kim, Phospholipase A2 dependent release of fatty acids from peroxidized membranes. J Free Radic Biol Med, 1985. 1(4): p. 263-71.
97. Proudfoot, J.M., et al., HDL is the major lipoprotein carrier of plasma F2-isoprostanes. J Lipid Res, 2009. 50(4): p. 716-22.
98. Roberts, L.J. and J.D. Morrow, Measurement of F(2)-isoprostanes as an index of oxidative stress in vivo. Free Radic Biol Med, 2000. 28(4): p. 505-13.
99. Roberts, L.J., 2nd, et al., Identification of the major urinary metabolite of the F2-isoprostane 8-iso-prostaglandin F2alpha in humans. J Biol Chem, 1996. 271(34): p. 20617-20.
100. Halliwell, B. and C.Y. Lee, Using isoprostanes as biomarkers of oxidative stress: some rarely considered issues. Antioxidants & redox signaling, 2010. 13(2): p. 145-56.
101. Morrow, J.D. and L.J. Roberts, 2nd, Mass spectrometric quantification of F2-isoprostanes in biological fluids and tissues as measure of oxidant stress. Methods Enzymol, 1999. 300: p. 3-12.

102
102. Wu, J.H., et al., Effects of alpha-tocopherol and mixed tocopherol supplementation on markers of oxidative stress and inflammation in type 2 diabetes. Clin Chem, 2007. 53(3): p. 511-9.
103. Milne, G.L., et al., Quantification of F2-isoprostanes in biological fluids and tissues as a measure of oxidant stress. Methods in enzymology, 2007. 433: p. 113-26.
104. Yin, H., New techniques to detect oxidative stress markers: mass spectrometry-based methods to detect isoprostanes as the gold standard for oxidative stress in vivo. Biofactors, 2008. 34(2): p. 109-24.
105. Schweer, H., et al., Improved quantification of 8-epi-prostaglandin F2 alpha and F2-isoprostanes by gas chromatography/triple-stage quadrupole mass spectrometry: partial cyclooxygenase-dependent formation of 8-epi-prostaglandin F2 alpha in humans. J Mass Spectrom, 1997. 32(12): p. 1362-70.
106. Tsikas, D., et al., Specific and rapid quantification of 8-iso-prostaglandin F2alpha in urine of healthy humans and patients with Zellweger syndrome by gas chromatography-tandem mass spectrometry. J Chromatogr B Biomed Sci Appl, 1998. 716(1-2): p. 7-17.
107. Taylor, A.W., et al., Benefits of prolonged gradient separation for high-performance liquid chromatography-tandem mass spectrometry quantitation of plasma total 15-series F-isoprostanes. Anal Biochem, 2006. 350(1): p. 41-51.
108. Zhang, B. and K. Saku, Control of matrix effects in the analysis of urinary F2-isoprostanes using novel multidimensional solid-phase extraction and LC-MS/MS. J Lipid Res, 2007. 48(3): p. 733-44.
109. Proudfoot, J., et al., Measurement of urinary F(2)-isoprostanes as markers of in vivo lipid peroxidation-A comparison of enzyme immunoassay with gas chromatography/mass spectrometry. Anal Biochem, 1999. 272(2): p. 209-15.
110. Basu, S., Radioimmunoassay of 8-iso-prostaglandin F2alpha: an index for oxidative injury via free radical catalysed lipid peroxidation. Prostaglandins Leukot Essent Fatty Acids, 1998. 58(4): p. 319-25.
111. Kinsella, B.T., D.J. O'Mahony, and G.A. Fitzgerald, The human thromboxane A2 receptor alpha isoform (TP alpha) functionally couples to the G proteins Gq and G11 in vivo and is activated by the isoprostane 8-epi prostaglandin F2 alpha. J Pharmacol Exp Ther, 1997. 281(2): p. 957-64.
112. Morrow, J.D., The isoprostanes - unique products of arachidonate peroxidation: their role as mediators of oxidant stress. Curr Pharm Des, 2006. 12(8): p. 895-902.
113. Janssen, L.J., The pulmonary biology of isoprostanes. Chem Phys Lipids, 2004. 128(1-2): p. 101-16.
114. Miggin, S.M. and B.T. Kinsella, Thromboxane A(2) receptor mediated activation of the mitogen activated protein kinase cascades in human uterine smooth muscle cells. Biochim Biophys Acta, 2001. 1539(1-2): p. 147-62.
115. Ting, H.J. and F.T. Khasawneh, Platelet function and Isoprostane biology. Should isoprostanes be the newest member of the orphan-ligand family? J Biomed Sci, 2010. 17(1): p. 24.
116. Khasawneh, F.T., et al., Characterization of isoprostane signaling: evidence for a unique coordination profile of 8-iso-PGF(2alpha) with the thromboxane A(2) receptor, and activation of a separate cAMP-dependent inhibitory pathway in human platelets. Biochem Pharmacol, 2008. 75(12): p. 2301-15.

103
117. Hou, X., et al., Augmented vasoconstriction and thromboxane formation by 15-F(2t)-isoprostane (8-iso-prostaglandin F(2alpha)) in immature pig periventricular brain microvessels. Stroke, 2000. 31(2): p. 516-24; discussion 525.
118. Marliere, S., et al., The 5-series F(2)-isoprostanes possess no vasomotor effects in the rat thoracic aorta, the human internal mammary artery and the human saphenous vein. Br J Pharmacol, 2002. 135(5): p. 1276-80.
119. Gude, N.M., et al., Growth and function of the normal human placenta. Thromb Res, 2004. 114(5-6): p. 397-407.
120. Huppertz, B., The anatomy of the normal placenta. Journal of clinical pathology, 2008. 61(12): p. 1296-302.
121. Hung, T.H. and G.J. Burton, Hypoxia and reoxygenation: a possible mechanism for placental oxidative stress in preeclampsia. Taiwan J Obstet Gynecol, 2006. 45(3): p. 189-200.
122. Redman, C.W., Preeclampsia: a multi-stress disorder. Rev Med Interne, 2011. 32 Suppl 1: p. S41-4.
123. Burton, G.J., Oxygen, the Janus gas; its effects on human placental development and function. J Anat, 2009. 215(1): p. 27-35.
124. Jauniaux, E., B. Gulbis, and G.J. Burton, The human first trimester gestational sac limits rather than facilitates oxygen transfer to the foetus--a review. Placenta, 2003. 24 Suppl A: p. S86-93.
125. Myatt, L. and X. Cui, Oxidative stress in the placenta. Histochemistry and cell biology, 2004. 122(4): p. 369-82.
126. Belo, L., et al., LDL size, total antioxidant status and oxidised LDL in normal human pregnancy: a longitudinal study. Atherosclerosis, 2004. 177(2): p. 391-9.
127. Palm, M., et al., F(2)-isoprostanes, tocopherols and normal pregnancy. Free Radic Res, 2009. 43(6): p. 546-52.
128. Redman, C.W. and I.L. Sargent, Pre-eclampsia, the placenta and the maternal systemic inflammatory response--a review. Placenta, 2003. 24 Suppl A: p. S21-7.
129. Agarwal, A., S. Gupta, and R.K. Sharma, Role of oxidative stress in female reproduction. Reprod Biol Endocrinol, 2005. 3: p. 28.
130. Powe, C.E., R.J. Levine, and S.A. Karumanchi, Preeclampsia, a disease of the maternal endothelium: the role of antiangiogenic factors and implications for later cardiovascular disease. Circulation, 2011. 123(24): p. 2856-69.
131. Helewa, M.E., et al., Report of the Canadian Hypertension Society Consensus Conference: 1. Definitions, evaluation and classification of hypertensive disorders in pregnancy. CMAJ : Canadian Medical Association journal = journal de l'Association medicale canadienne, 1997. 157(6): p. 715-25.
132. Sibai, B., G. Dekker, and M. Kupferminc, Pre-eclampsia. Lancet, 2005. 365(9461): p. 785-799.
133. Young, B.C., R.J. Levine, and S.A. Karumanchi, Pathogenesis of Preeclampsia. Annual Review of Pathology: Mechanisms of Disease, 2010. 5(1): p. 173-192.
134. MacKay, A.P., C.J. Berg, and H.K. Atrash, Pregnancy-related mortality from preeclampsia and eclampsia. Obstet Gynecol, 2001. 97(4): p. 533-8.
135. Bellamy, L., et al., Pre-eclampsia and risk of cardiovascular disease and cancer in later life: systematic review and meta-analysis. BMJ, 2007. 335(7627): p. 974.
136. McDonald, S.D., et al., Cardiovascular sequelae of preeclampsia/eclampsia: a systematic review and meta-analyses. Am Heart J, 2008. 156(5): p. 918-30.

104
137. Osmond, C. and D.J. Barker, Fetal, infant, and childhood growth are predictors of coronary heart disease, diabetes, and hypertension in adult men and women. Environ Health Perspect, 2000. 108 Suppl 3: p. 545-53.
138. Naicker, T., et al., Quantitative analysis of trophoblast invasion in preeclampsia. Acta Obstet Gynecol Scand, 2003. 82(8): p. 722-9.
139. Granger, J.P., et al., Pathophysiology of hypertension during preeclampsia linking placental ischemia with endothelial dysfunction. Hypertension, 2001. 38(3 Pt 2): p. 718-22.
140. Gupta, S., et al., Lipid peroxidation and antioxidant status in preeclampsia: a systematic review. Obstet Gynecol Surv, 2009. 64(11): p. 750-9.
141. Redman, C.W. and I.L. Sargent, Placental stress and pre-eclampsia: a revised view. Placenta, 2009. 30 Suppl A: p. S38-42.
142. Roberts, J.M., et al., Preeclampsia: an endothelial cell disorder. Am J Obstet Gynecol, 1989. 161(5): p. 1200-4.
143. Roberts, J.M., Endothelial dysfunction in preeclampsia. Semin Reprod Endocrinol, 1998. 16(1): p. 5-15.
144. Stillman, I.E. and S.A. Karumanchi, The glomerular injury of preeclampsia. J Am Soc Nephrol, 2007. 18(8): p. 2281-4.
145. Gaber, L.W., B.H. Spargo, and M.D. Lindheimer, Renal pathology in pre-eclampsia. Baillieres Clin Obstet Gynaecol, 1994. 8(2): p. 443-68.
146. Schulz, E., T. Gori, and T. Munzel, Oxidative stress and endothelial dysfunction in hypertension. Hypertension research : official journal of the Japanese Society of Hypertension, 2011. 34(6): p. 665-73.
147. Lusis, A.J., Atherosclerosis. Nature, 2000. 407(6801): p. 233-41. 148. Raijmakers, M.T., R. Dechend, and L. Poston, Oxidative stress and preeclampsia: rationale
for antioxidant clinical trials. Hypertension, 2004. 44(4): p. 374-80. 149. Mellembakken, J.R., et al., Activation of leukocytes during the uteroplacental passage in
preeclampsia. Hypertension, 2002. 39(1): p. 155-60. 150. Cackovic, M., et al., Fractional excretion of tumor necrosis factor-alpha in women with severe
preeclampsia. Obstet Gynecol, 2008. 112(1): p. 93-100. 151. Roberts, J.M. and C.A. Hubel, The two stage model of preeclampsia: variations on the
theme. Placenta, 2009. 30 Suppl A: p. S32-7. 152. Elmarakby, A.A. and J.C. Sullivan, Relationship between oxidative stress and inflammatory
cytokines in diabetic nephropathy. Cardiovasc Ther, 2012. 30(1): p. 49-59. 153. Hung, T.H., et al., Secretion of tumor necrosis factor-alpha from human placental tissues
induced by hypoxia-reoxygenation causes endothelial cell activation in vitro: a potential mediator of the inflammatory response in preeclampsia. Am J Pathol, 2004. 164(3): p. 1049-61.
154. Silva, L.M., et al., Low socioeconomic status is a risk factor for preeclampsia: the Generation R Study. J Hypertens, 2008. 26(6): p. 1200-8.
155. Wacker, J., et al., Riboflavin deficiency and preeclampsia. Obstet Gynecol, 2000. 96(1): p. 38-44.
156. Conde-Agudelo, A. and J.M. Belizan, Risk factors for pre-eclampsia in a large cohort of Latin American and Caribbean women. BJOG, 2000. 107(1): p. 75-83.
157. Sibai, B.M., et al., Risk factors for preeclampsia in healthy nulliparous women: a prospective multicenter study. The National Institute of Child Health and Human Development Network of Maternal-Fetal Medicine Units. Am J Obstet Gynecol, 1995. 172(2 Pt 1): p. 642-8.

105
158. Dekker, G. and B. Sibai, Primary, secondary, and tertiary prevention of pre-eclampsia. Lancet, 2001. 357(9251): p. 209-15.
159. Duckitt, K. and D. Harrington, Risk factors for pre-eclampsia at antenatal booking: systematic review of controlled studies. BMJ, 2005. 330(7491): p. 565.
160. Skjaerven, R., A.J. Wilcox, and R.T. Lie, The interval between pregnancies and the risk of preeclampsia. N Engl J Med, 2002. 346(1): p. 33-8.
161. Walker, J.J., Pre-eclampsia. Lancet, 2000. 356(9237): p. 1260-5. 162. Robillard, P.Y., et al., Association of pregnancy-induced hypertension with duration of sexual
cohabitation before conception. Lancet, 1994. 344(8928): p. 973-5. 163. Lie, R.T., et al., Fetal and maternal contributions to risk of pre-eclampsia: population based
study. BMJ, 1998. 316(7141): p. 1343-7. 164. Koga, K., et al., Elevated serum soluble fms-like tyrosine kinase 1 (sFlt1) level in women with
hydatidiform mole. Fertil Steril, 2010. 94(1): p. 305-8. 165. Siddiqui, I.A., et al., Role of oxidative stress in the pathogenesis of preeclampsia. Arch
Gynecol Obstet, 2010. 282(5): p. 469-74. 166. Barden, A., et al., Plasma and urinary 8-iso-prostane as an indicator of lipid peroxidation in
pre-eclampsia and normal pregnancy. Clin Sci (Lond), 1996. 91(6): p. 711-8. 167. Barden, A., et al., Study of plasma factors associated with neutrophil activation and lipid
peroxidation in preeclampsia. Hypertension, 2001. 38(4): p. 803-8. 168. Harsem, N.K., et al., Acute atherosis in decidual tissue: not associated with systemic
oxidative stress in preeclampsia. Placenta, 2007. 28(8-9): p. 958-64. 169. Ishihara, O., et al., Isoprostanes, prostaglandins and tocopherols in pre-eclampsia, normal
pregnancy and non-pregnancy. Free Radic Res, 2004. 38(9): p. 913-8. 170. Mandang, S., U. Manuelpillai, and E.M. Wallace, Oxidative stress increases placental and
endothelial cell activin A secretion. J Endocrinol, 2007. 192(3): p. 485-93. 171. McKinney, E.T., et al., Plasma, urinary, and salivary 8-epi-prostaglandin f2alpha levels in
normotensive and preeclamptic pregnancies. American journal of obstetrics and gynecology, 2000. 183(4): p. 874-7.
172. Morris, J.M., et al., Circulating markers of oxidative stress are raised in normal pregnancy and pre-eclampsia. British journal of obstetrics and gynaecology, 1998. 105(11): p. 1195-9.
173. Ouyang, Y.Q., et al., Interactions between inflammatory and oxidative stress in preeclampsia. Hypertens Pregnancy, 2009. 28(1): p. 56-62.
174. Ouyang, Y.Q., et al., Plasma sFlt-1-to-PlGF ratio is correlated with inflammatory but not with oxidative stress in Chinese preeclamptic women. Arch Gynecol Obstet, 2009. 280(1): p. 91-7.
175. Peter Stein, T., et al., Oxidative stress early in pregnancy and pregnancy outcome. Free Radic Res, 2008. 42(10): p. 841-8.
176. Staff, A.C., et al., Elevated level of free 8-iso-prostaglandin F2alpha in the decidua basalis of women with preeclampsia. Am J Obstet Gynecol, 1999. 181(5 Pt 1): p. 1211-5.
177. Walsh, S.W., et al., Placental isoprostane is significantly increased in preeclampsia. FASEB J, 2000. 14(10): p. 1289-96.
178. Zhang, J., et al., Placental anti-oxidant gene polymorphisms, enzyme activity, and oxidative stress in preeclampsia. Placenta, 2008. 29(5): p. 439-43.
179. Morris, J.M., et al., Circulating markers of oxidative stress are raised in normal pregnancy and pre-eclampsia. Br J Obstet Gynaecol, 1998. 105(11): p. 1195-9.
180. Regan, C.L., et al., No evidence for lipid peroxidation in severe preeclampsia. Am J Obstet Gynecol, 2001. 185(3): p. 572-8.

106
181. Redman, C.W., Current topic: pre-eclampsia and the placenta. Placenta, 1991. 12(4): p. 301-8.
182. Roland, L., et al., Existence of compensatory defense mechanisms against oxidative stress and hypertension in preeclampsia. Hypertension in pregnancy : official journal of the International Society for the Study of Hypertension in Pregnancy, 2010. 29(1): p. 21-37.
183. Harsem, N.K., K. Braekke, and A.C. Staff, Augmented oxidative stress as well as antioxidant capacity in maternal circulation in preeclampsia. Eur J Obstet Gynecol Reprod Biol, 2006. 128(1-2): p. 209-15.
184. Mori, T.A., et al., An improved method for the measurement of urinary and plasma F2-isoprostanes using gas chromatography-mass spectrometry. Anal Biochem, 1999. 268(1): p. 117-25.
185. Gritti, F. and G. Guiochon, Facts and Legends on Columns Packed with Sub-3-µm Core-Shell Particles. LCGC North America, 2012. 30(7): p. 586-595.
186. Korecka, M., et al., Simultaneous HPLC-MS-MS quantification of 8-iso-PGF(2alpha) and 8,12-iso-iPF(2alpha) in CSF and brain tissue samples with on-line cleanup. J Chromatogr B Analyt Technol Biomed Life Sci, 2010. 878(24): p. 2209-16.
187. Haschke, M., et al., HPLC-atmospheric pressure chemical ionization MS/MS for quantification of 15-F2t-isoprostane in human urine and plasma. Clin Chem, 2007. 53(3): p. 489-97.
188. Yin, H., N.A. Porter, and J.D. Morrow, Separation and identification of F2-isoprostane regioisomers and diastereomers by novel liquid chromatographic/mass spectrometric methods. J Chromatogr B Analyt Technol Biomed Life Sci, 2005. 827(1): p. 157-64.
189. Bohnstedt, K.C., et al., Porous graphitic carbon chromatography-tandem mass spectrometry for the study of isoprostanes in human cerebrospinal fluid. J Chromatogr B Analyt Technol Biomed Life Sci, 2005. 827(1): p. 39-43.
190. Williams, M.A., et al., Risk of preeclampsia in relation to elaidic acid (trans fatty acid) in maternal erythrocytes. Gynecol Obstet Invest, 1998. 46(2): p. 84-7.
191. Oken, E., et al., Diet during pregnancy and risk of preeclampsia or gestational hypertension. Ann Epidemiol, 2007. 17(9): p. 663-8.
192. Radesky, J.S., et al., Diet during early pregnancy and development of gestational diabetes. Paediatr Perinat Epidemiol, 2008. 22(1): p. 47-59.
193. Bowers, K., et al., A prospective study of prepregnancy dietary fat intake and risk of gestational diabetes. Am J Clin Nutr, 2012. 95(2): p. 446-53.
194. Elias, S.L. and S.M. Innis, Infant plasma trans, n-6, and n-3 fatty acids and conjugated linoleic acids are related to maternal plasma fatty acids, length of gestation, and birth weight and length. Am J Clin Nutr, 2001. 73(4): p. 807-14.
195. Dirix, C.E., A.D. Kester, and G. Hornstra, Associations between term birth dimensions and prenatal exposure to essential and trans fatty acids. Early Hum Dev, 2009. 85(8): p. 525-30.
196. van Eijsden, M., et al., Maternal n-3, n-6, and trans fatty acid profile early in pregnancy and term birth weight: a prospective cohort study. Am J Clin Nutr, 2008. 87(4): p. 887-95.
197. Martyn, C.N., Fetal and infant origins of cardiovascular disease. Midwifery, 1994. 10(2): p. 61-6.
198. Martyn, C.N. and D.J. Barker, Reduced fetal growth increases risk of cardiovascular disease. Health Rep, 1994. 6(1): p. 45-53.
199. Oken, E. and M.W. Gillman, Fetal origins of obesity. Obes Res, 2003. 11(4): p. 496-506. 200. Shiell, A.W., et al., Diet in late pregnancy and glucose-insulin metabolism of the offspring 40
years later. BJOG, 2000. 107(7): p. 890-5.

107
201. Barker, D.J., Fetal origins of coronary heart disease. BMJ, 1995. 311(6998): p. 171-4. 202. Decsi, T., et al., Inverse association between trans isomeric and long-chain polyunsaturated
fatty acids in pregnant women and their newborns: data from three European countries. Ann Nutr Metab, 2011. 59(2-4): p. 107-16.
203. Sugano, M. and I. Ikeda, Metabolic interactions between essential and trans-fatty acids. Curr Opin Lipidol, 1996. 7(1): p. 38-42.
204. Innis, S.M., Fatty acids and early human development. Early Hum Dev, 2007. 83(12): p. 761-6.
205. Innis, S.M., Dietary (n-3) fatty acids and brain development. J Nutr, 2007. 137(4): p. 855-9. 206. Bouwstra, H., et al., Neurologic condition of healthy term infants at 18 months: positive
association with venous umbilical DHA status and negative association with umbilical trans-fatty acids. Pediatr Res, 2006. 60(3): p. 334-9.
207. Kadiiska, M.B., et al., Biomarkers of oxidative stress study III. Effects of the nonsteroidal anti-inflammatory agents indomethacin and meclofenamic acid on measurements of oxidative products of lipids in CCl4 poisoning. Free Radic Biol Med, 2005. 38(6): p. 711-8.
208. Fessel, J.P., et al., Discovery of lipid peroxidation products formed in vivo with a substituted tetrahydrofuran ring (isofurans) that are favored by increased oxygen tension. Proc Natl Acad Sci U S A, 2002. 99(26): p. 16713-8.
209. Roberts, L.J. and J.P. Fessel, The biochemistry of the isoprostane, neuroprostane, and isofuran pathways of lipid peroxidation. Chemistry and Physics of Lipids, 2004. 128(1-2): p. 173-186.
210. Mas, E., et al., Effects of spinal or general anesthesia on F(2)-isoprostanes and isofurans during ischemia/reperfusion of the leg in patients undergoing knee replacement surgery. Free Radic Biol Med, 2011. 50(9): p. 1171-6.
211. Barden, A.E., et al., Is there a role for isofurans and neuroprostanes in pre-eclampsia and normal pregnancy? Antioxid Redox Signal, 2012. 16(2): p. 165-9.


109
Annexe 1
The polyol pathway in the bovine oviduct


111

112

113

114

115

116

117

118

119

120