Embed Size (px)
Citation preview

Pflügers Archiv, Bd. 260, S. 448—479 (1955).
Aus dem Centre d'études de Physiologie nerveuse Paris.
Analyse électrophysiologique et comparaison, chez l'Homme, du réflexe de HOFFMANN et du
réflexe myotatique.
Par
J. PAILLARD*.
Avec 12 figures dans le texte. (Eingegangen am 15 Dezember 1954.)
La technique d'évocation d'un réflexe monosynaptique (ECCLES et
PRITCHARD 1937, RENSHAW 1940), en fournissant un test direct de l'excitabilité des neurones moteurs, devait ouvrir, dans l'étude des fonctions de la moelle, un champ expérimental d'une exceptionnelle fécondité. Les progrès récemment réalisés dans cette voie de recherche, en neurophysiologie animale, donnent aujourd'hui un renouveau d'intérêt aux techniques d'investigation depuis longtemps appliquées par HOFFMANN et ses élèves à l'Homme (HOFFMANN 1934, 1952b).
L'expérimentation animale retrouve, en effet, en les précisant, de nombreuses propriétés de l'organisation des fonctions médullaires déjà implicitement contenues dans la distinction fondamentale entre « Eigenreflexe » et « Fremdreflexe » primitivement introduite par HOFFMANN en neurophysiologie humaine (HOFFMANN 1934—1953).
Les audacieuses expériences de MAGLADERY et coll. (1951a), conduites chez l'Homme jusqu'au niveau de la moelle, apportent, par l'estimation du délai central de transmission (inférieur à 1 ms), une preuve expérimentale supplémentaire de la légitimité d'une identification de l'« Eigenreflexe » de HOFFMANN avec une réponse monosynaptique.
L'identification de la réponse réflexe évoquée par un étirement brusque du muscle (réflexe myotatique de LIDDELL et SHERRINGTON 1924) avec le réflexe monosynaptique obtenu par la stimulation électri-que des fibres des bas seuils dans les racines dorsales du chat (LLOYD 1943) apportait d'autre part, plus de 25 ans après les expériences initiales de HOFFMANN, une remarquable confirmation des résultats acquis par ce dernier en expérimentant sur l'Homme. Dès 1910, en effet, HOFFMANN avait signalé la possibilité d'obtenir chez l'Homme, par stimulation électrique submaximale du nerf mixte, une contraction réflexe du muscle _______
* Herrn Professor PAUL HOFFMANN zum 70, Geburtstag als Zeichen tiefer Verehrung gewidmet.

Analyse du réflexe de HOFFMANN chez l'Homme. 449
assimilable dans ses caractéristiques essentielles à la réponse classique-ment évoquée par la percussion mécanique du tendon (HOFFMANN 1952b).
La connaissance, aujourd'hui plus claire, des origines, des destinations et des propriétés fonctionnelles des diverses catégories de fibres afférentes et efférentes qui composent le nerf mixte, ouvre le champ à de nouvelles expérimentations et fournit la base d'interprétations plus cohérentes des mécanismes de coordination motrice dans l'organisme intact.
Nous nous proposons d'aborder ici quelques problèmes méthodologi-ques posés par les conditions d'utilisation chez l'Homme d'un stimulus électrique ou d'un stimulus mécanique comme test explorateur de l'excitabilité des motoneurones. Nous analyserons ensuite les modifica-tions tardives de l'excitabilité centrale consécutives à ces deux modes de stimulation.
Techniques. L'étude a porté essentiellement sur la réponse électromyographique du triceps
sural provoquée électriquement par stimulation transcutanée du nerf sciatique poplité interne et mécaniquement par percussion du tendon d'Achille (fig. 1).
La stimulation électrique du nerf est effectuée suivant les conditions classiques préconisées par HOFFMANN. Une électrode large, humide, est fixée sur le tiers inférieur de la face antérieure de la cuisse. L'électrode de stimulation (1,5 cm de diamètre) est maintenue rigidement surremplacement convenable.
Fig. 1. Schéma du dispositif expérimental — S: Stimulation électrique monopolaire transcutanée du nerf sciatique - M: Stimulation mécanique du tendon d'Achille — R: Réception électromyo- graphique par électrodes superficielles — C1 et C2 : éléments du dispositif de contention de l'arti- culation du genou et de la cheville.
Les conditions de placement de l'électrode sont primordiales dans l'utilisation d'une stimulation trans- cutanée : une grande partie des irrégu- larités, habituellement rencontrées avec des systèmes de fixation par bandes élastiques, a pu être supprimée grâce à l'utilisation d'un système de contention rigide de la jambe. — L'électrode d'excitation est alors rendue solidaire de ce dispositif et per- met une définition relativement stable des caractéristiques de la stimulation.
Le choix des paramètres de stimulation n'est pas sans importance. Nous avons utilisé un générateur à courant rectangulaire permettant la distribution de deux chocs indépendants, décalables jusqu'à des délais de 5 à 6 secondes. — La forme du courant est respectée par le transformateur d'isolement, grâce à l'emploi d'un montage de DUBBERT et SCHMITT (1949) qui utilise, une onde porteuse à haute fréquence redressée et filtrée à la sortie du transformateur. Un courant bref tend à favoriser sélectivement les fibres motrices par rapport aux fibres sensorielles. Ce fait est en accord avec les données de ERLANGER et BLAIR (1938), SKOGLUND (1942), KUGELBERG (1944) sur les différences observables entre ces deux types de fibres quant à leurs caractéristiques d'accommodation. Il y a donc intérêt à utiliser des
450 J. PAILLARD
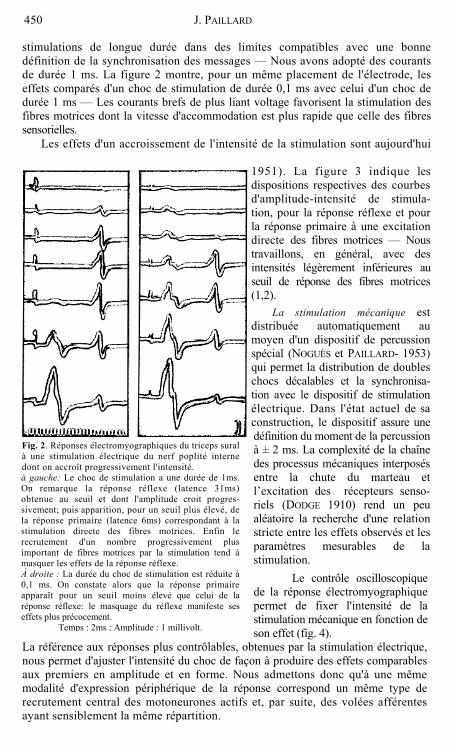
stimulations de longue durée dans des limites compatibles avec une bonne définition de la synchronisation des messages — Nous avons adopté des courants de durée 1 ms. La figure 2 montre, pour un même placement de l'électrode, les effets comparés d'un choc de stimulation de durée 0,1 ms avec celui d'un choc de durée 1 ms — Les courants brefs de plus liant voltage favorisent la stimulation des fibres motrices dont la vitesse d'accommodation est plus rapide que celle des fibres sensorielles.
Les effets d'un accroissement de l'intensité de la stimulation sont aujourd'hui classiques (HOFFMANN et KELLER 1928, SCHENCK 1951, MAGLADERY et coll.
1951). La figure 3 indique les dispositions respectives des courbes d'amplitude-intensité de stimula- tion, pour la réponse réflexe et pour la réponse primaire à une excitation directe des fibres motrices — Nous travaillons, en général, avec des intensités légèrement inférieures au seuil de réponse des fibres motrices (1,2).
La stimulation mécanique est distribuée automatiquement au moyen d'un dispositif de percussion spécial (NOGUÈS et PAILLARD- 1953) qui permet la distribution de doubles chocs décalables et la synchronisa- tion avec le dispositif de stimulation électrique. Dans l'état actuel de sa construction, le dispositif assure une définition du moment de la percussion à ± 2 ms. La complexité de la chaîne des processus mécaniques interposés entre la chute du marteau et l’excitation des récepteurs senso- riels (DODGE 1910) rend un peu aléatoire la recherche d'une relation stricte entre les effets observés et les paramètres mesurables de la stimulation.
Fig. 2. Réponses électromyographiques du triceps sural à une stimulation électrique du nerf poplité interne dont on accroît progressivement l'intensité. à gauche: Le choc de stimulation a une durée de 1ms. On remarque la réponse réflexe (latence 31ms) obtenue au seuil et dont l'amplitude croit progres- sivement; puis apparition, pour un seuil plus élevé, de la réponse primaire (latence 6ms) correspondant à la stimulation directe des fibres motrices. Enfin le recrutement d'un nombre progressivement plus important de fibres motrices par la stimulation tend à masquer les effets de la réponse réflexe. À droite : La durée du choc de stimulation est réduite à 0,1 ms. On constate alors que la réponse primaire apparaît pour un seuil moins élevé que celui de la réponse réflexe: le masquage du réflexe manifeste ses effets plus précocement.
Temps : 2ms ; Amplitude : 1 millivolt.
Le contrôle oscilloscopique de la réponse électromyographique permet de fixer l'intensité de la stimulation mécanique en fonction de son effet (fig. 4).
La référence aux réponses plus contrôlables, obtenues par la stimulation électrique, nous permet d'ajuster l'intensité du choc de façon à produire des effets comparables aux premiers en amplitude et en forme. Nous admettons donc qu'à une même modalité d'expression périphérique de la réponse correspond un même type de recrutement central des motoneurones actifs et, par suite, des volées afférentes ayant sensiblement la même répartition.
Analyse du réflexe de HOFFMANN chez l'Homme. 451
La réception électromyographique des réponses réflexes s’effectue au moyen d’électrodes de surface distantes de 3 cm, placées sur le tiers inférieur du soléaire. Elles recueillent une réponse diphasique, de forme caractéristique, qui peut, en première approximation, être considérée comme la différentielle du nombre des fibres musculaires successivement activées.
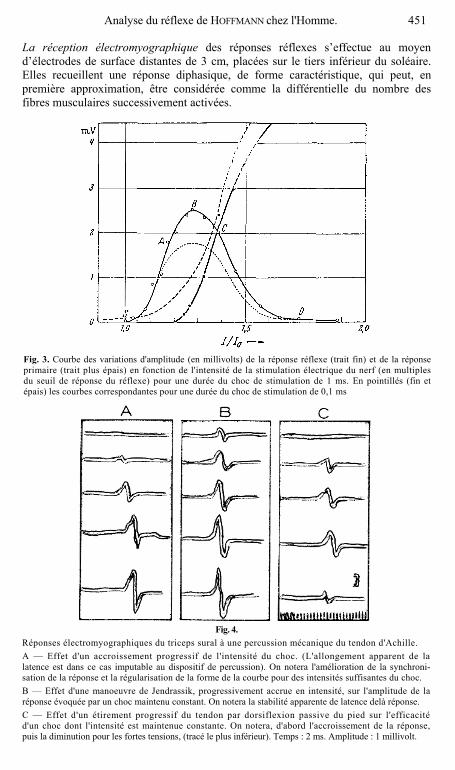
Fig. 3. Courbe des variations d'amplitude (en millivolts) de la réponse réflexe (trait fin) et de la réponse primaire (trait plus épais) en fonction de l'intensité de la stimulation électrique du nerf (en multiples du seuil de réponse du réflexe) pour une durée du choc de stimulation de 1 ms. En pointillés (fin et épais) les courbes correspondantes pour une durée du choc de stimulation de 0,1 ms
Fig. 4.
Réponses électromyographiques du triceps sural à une percussion mécanique du tendon d'Achille. A — Effet d'un accroissement progressif de l'intensité du choc. (L'allongement apparent de la latence est dans ce cas imputable au dispositif de percussion). On notera l'amélioration de la synchroni-sation de la réponse et la régularisation de la forme de la courbe pour des intensités suffisantes du choc. B — Effet d'une manoeuvre de Jendrassik, progressivement accrue en intensité, sur l'amplitude de la réponse évoquée par un choc maintenu constant. On notera la stabilité apparente de latence delà réponse. C — Effet d'un étirement progressif du tendon par dorsiflexion passive du pied sur l'efficacité d'un choc dont l'intensité est maintenue constante. On notera, d'abord l'accroissement de la réponse, puis la diminution pour les fortes tensions, (tracé le plus inférieur). Temps : 2 ms. Amplitude : 1 millivolt.

452 J. PAILLARD
Le maximum de synchronisme des fibres actives se situe approximativement au point de changement de signe de l'onde diphasique.
La réception myographique de la déformation du muscle au cours de la contrac-tion, est effectuée à l'aide d'un extensomètre à jauges de contrainte qui transcrit le phénomène avec fidélité.
Le sujet est ordinairement en position de décubitus ventral sur une table lourde, indépendante du système de percussion mécanique, pour éviter les vibrations parasites. Un système de contention rigide maintient la jambe sans contrainte excessive. Un contrôle permanent de l'état de vigilance du sujet s'est avéré utile et nous avons le plus fréquemment suivi électroencéphalographiquement les altérations de l'activité électrique corticale sur une dérivation vertex-occipital.
Les expériences relatives aux cycles d'excitabilité ont été faites sur 7 sujets apparemment normaux de 18 à 32 ans, revus un grand nombre de fois (en moyenne 15 expériences par sujet).
Les courbes d'excitabilité ont été tracées par point (moyenne de 5 à 15 mesures) ; les tests successifs étaient distribués à des cadences jamais inférieures à une stimu-lation toutes les 3 secondes.
I. Caractères généraux des réponses réflexes à un stimulus électrique (SE) et à un stimulus mécanique (SM).
On sait depuis l'expérience princeps de HOFFMANN (1918) qu'une stimulation électrique submaximale du nerf sciatique poplité interne permet de dissocier la réponse réflexe du triceps sural, provenant de la
qvarier de
stimulation des fibres sen- sorielles de bas seuils (nous l'appellerons réflexe de HOFF- MANN, ou, avec MAGLADERY et coll. 1950, réponse H), de la réponse motrice primaire évo- quée par la stimulation directe des fibres motrices (réponse M).
La réponse H, localisée au triceps sural, apparaît ré-gulièrement avec une latence
ui, suivant les sujets, peut 28 à 34 ms, avec une
forme relativement stable et une durée maximum de 8 à 12 ms (voir fig. 5).
La percussion mécanique du tendon d'Achille provoque, d'autre part, une réponse élec- tromyographique (réponse T),
tout à fait comparable (au moins pour une intensité de stimulation suffisante), en forme, en durée et quant à sa localisation, à celle du réflexe évoqué électriquement.
Fig. 5. Réponses électromyographiques du triceps sural. Ligne supérieure: à la stimulation électrique du nerf poplité interne. Le choc carré de durée 1 ms introduit l'artefact visible au début du tracé et permet le repérage précis du moment de la stimulation. Ligne inférieure: à une percussion mécanique du tendon d'Achille. Un mouvement des électrodes de réception, consécutif au choc, provoque l'artefact du début du tracé et indique approximativement le moment de la stimulation.
Temps : 2 ms. Amplitude: 1 millivolt.

Analyse du réflexe de HOFFMANN chez l'Homme. 453
La latence ne peut être appréciée rigoureusement (figure 5). En se basant sur le repère approximatif fourni par l'artéfact de mouvement de l'électrode, on peut constater un léger retard de la réponse T sur la réponse H. Les retards impliqués par les conditions d'excitation des récepteurs péri- phériques et par la plus grande longueur des voies de conduction peuvent largement en rendre compte. Il semble assez légitime d'admettre un même délai de transmission centrale pour ces deux types de réponse.
a) Nombreux sont les arguments en faveur d'une identification plus complète de la réponse H et de la réponse T (HOFFMANN 1934). Elle se trouve, en tout cas, justifiée chez l’animal, par la démonstration d'une identité des voies de conduction (Groupe I de LLOYD 1943a) pour les messages en provenance des récepteurs spiralés du fuseau neuromuscu- laire (Type A2 de MATTHEWS 1933) et pour ceux évoqués par la stimula- tion électrique des fibres dont les seuils d'excitabilité sont les plus bas et les vitesses de conduction les plus grandes (LLOYD 1943b).
L'assimilation de ces réponses à un réflexe monosynaptique, au-jourd'hui démontrée en neurophysiologie animale, paraît extrapolable à l'Homme. L'estimation, par MAGLADERY et coll. (1951), d'un délai central du réflexe de HOFFMANN inférieur à la milliseconde apporte, dans ce sens, un argument supplémentaire. L'ensemble des propriétés reconnues depuis longtemps par HOFFMANN (1934) à la catégorie des «Eigenreflexe» justifiait déjà une telle manière de voir.
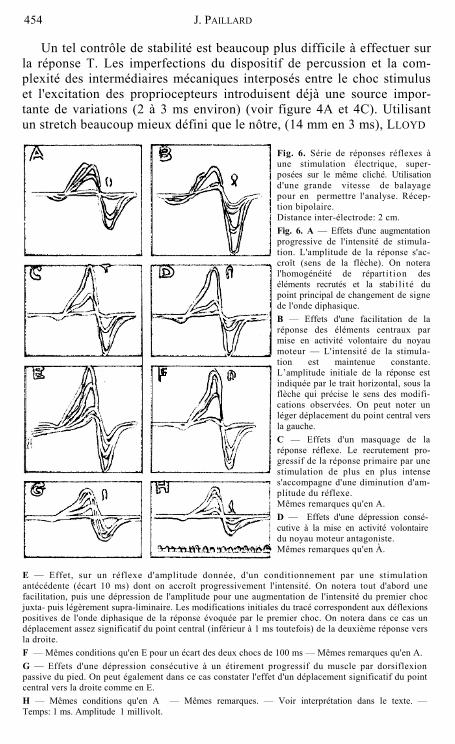
b) L'affirmation, souvent rencontrée dans les travaux anciens, d'une variabilité de la latence des réflexes tendineux a toujours été contestée par HOFFMANN (1934). Une réponse monosynaptique — il est aujourd'hui classique de l'admettre — ne peut présenter de modifications de latence que dans une marge assez étroite, inférieure à la milliseconde (LLOYD 1943a). La figure 6 présente quelques aspects d'une analyse systé- matique des réponses obtenues dans diverses conditions expérimentales. HOFFMANN avait déjà souligné les difficultés d'une appréciation rigoureuse des temps de latence. Le début de la réponse pris pour critère est beaucoup trop incertain: Le recul de l'accident initial de l'onde, surtout s'il est associé à une diminution de l'amplitude de la réponse, ne peut, en aucun cas, être significatif d'une modification des latences centrales; les fluctuations importantes (2 à 8 ms) signalées par MAGLADERY et coll. (1951a) sont probablement attribuables au choix d'un tel repère. Nous lui avons préféré la position du maximum de synchronisme des fibres actives (point d'inversion de signe de l'onde diphasique). On peut constater sa bonne stabilité dans la plupart des situations expérimentales envisagées (figure 6). Il est important de noter que lorsqu'un déplacement significatif de ce point apparaît systéma- tiquement (figure 6E et G), il ne dépasse pas 1 ms — Ce fait reste bien compatible avec l'hypothèse de la nature monosynaptique des liaisons centrales intéressées dans la réponse H.
454 J. PAILLARD
Un tel contrôle de stabilité est beaucoup plus difficile à effectuer sur la réponse T. Les imperfections du dispositif de percussion et la com-plexité des intermédiaires mécaniques interposés entre le choc stimulus et l'excitation des propriocepteurs introduisent déjà une source impor- tante de variations (2 à 3 ms environ) (voir figure 4A et 4C). Utilisant un stretch beaucoup mieux défini que le nôtre, (14 mm en 3 ms), LLOYD
Mêm
Fig. 6. Série de réponses réflexes à une stimulation électrique, super- posées sur le même cliché. Utilisation d'une grande vitesse de balayage pour en permettre l'analyse. Récep- tion bipolaire. Distance inter-électrode: 2 cm. Fig. 6. A — Effets d'une augmentation progressive de l'intensité de stimula- tion. L'amplitude de la réponse s'ac- croît (sens de la flèche). On notera l'homogénéité de répart i t ion des éléments recrutés et la stabi l i té du point principal de changement de signe de l'onde diphasique. B — Effets d'une facilitation de la réponse des éléments centraux par mise en activité volontaire du noyau moteur — L'intensité de la stimula-tion est maintenue constante. L’amplitude initiale de la réponse est indiquée par le trait horizontal, sous la flèche qui précise le sens des modifi- cations observées. On peut noter un léger déplacement du point central vers la gauche. C — Effets d'un masquage de la réponse réflexe. Le recrutement pro-gressif de la réponse primaire par une stimulation de plus en plus intense s'accompagne d'une diminution d'am- plitude du réflexe. Mêmes remarques qu'en A. D — Effets d'une dépression consé-cutive à la mise en activité volontaire du noyau moteur antagoniste.
es remarques qu'en À.
E — Effet, sur un réflexe d'amplitude donnée, d'un conditionnement par une stimulation antécédente (écart 10 ms) dont on accroît progressivement l'intensité. On notera tout d'abord une facilitation, puis une dépression de l'amplitude pour une augmentation de l'intensité du premier choc juxta- puis légèrement supra-liminaire. Les modifications initiales du tracé correspondent aux déflexions positives de l'onde diphasique de la réponse évoquée par le premier choc. On notera dans ce cas un déplacement assez significatif du point central (inférieur à 1 ms toutefois) de la deuxième réponse vers la droite. F — Mêmes conditions qu'en E pour un écart des deux chocs de 100 ms — Mêmes remarques qu'en A. G — Effets d'une dépression consécutive à un étirement progressif du muscle par dorsiflexion passive du pied. On peut également dans ce cas constater l'effet d'un déplacement significatif du point central vers la droite comme en E. H — Mêmes conditions qu'en A — Mêmes remarques. — Voir interprétation dans le texte. — Temps: 1 ms. Amplitude 1 millivolt.

Analyse du réflexe de HOFFMANN chez l'Homme. 455
(1943b) constatait néanmoins, chez le Chat, un étalement assez con-sidérable de la volée afférente contrastant avec le groupement bien synchrone de la volée évoquée par une SE de la racine dorsale. Dans l'un et l'autre cas, cependant, les réponses recueillies sur la racine ven-trale apparaissent identiques. La réponse à la SM présente simplement des fluctuations de sa latence attribuables au hasard des groupements d'afférences et aux effets de facilitation temporelle qui en résultent.
Ainsi, les complications imputables à l'emploi de la SM entraînent une variabilité des temps de réponse vraisemblablement très supérieure à celle qui peut intervenir au niveau des fonctions centrales.
Nous n'avons jamais observé de modifications de la latence aussi considérables que celles mentionnées par TEASDALL et coll. (1952). La réponse T était, dans ce cas, évoquée par l'intermédiaire d'une stimulation électrique du jambier antérieur. La contraction de ce muscle provoque un étirement brusque du triceps sural par dorsiflexion du pied. Les complications, introduites par la nature des intermédiaires mécaniques mis en jeu et par les répercussions centrales directes de la stimulation électrique du nerf antagoniste, rendent ces résultats difficilement interprétables. L'aspect polyphasique des réponses évoquées de cette manière plaide déjà en faveur de l'existence d'un étalement très considérable de la volée afférente corres-pondante.
En accord avec HOFFMANN (1934), il semble donc que l'on puisse admettre, comme pour la réponse H, une grande stabilité des délais centraux de la réponse T.
c) L'identification des réflexes H et T comme des réponses mono-synaptiques implique que la décharge des éléments centraux précède pratiquement toutes les influences centrales secondaires associées au mode de stimulation lui-même: influences qui résultent de la stimulation de diverses catégories de propriocepteurs (cas de la SM), complications inévitables associées à l'excitation d'un tronc nerveux complexe (cas de la SE), influences proprioceptives, enfin, provenant de la contraction musculaire résultante (de même nature dans le cas d'une réponse H ou d'une réponse T).
Aussi les réponses H et T peuvent donc, en première approximation, être utilisées comme expression objective d'un test direct de l'excitabilité des motoneurones.
Un facteur de complication supplémentaire peut toutefois intervenir dans l'utilisation de la SM: il concerne le fait, déjà mentionné, de la moins bonne synchronisation de la volée afférente et des retards éven-tuels qui peuvent intervenir de ce fait dans le déclenchement de la com-mande centrale (LLOYD 1943b). Les motoneurones risqueraient donc de se trouver déjà affectés de façon complexe par les premiers messages nerveux qui ont atteint les centres. En fait, cette complication ne nous a pas semblé jouer un rôle très important dans nos expériences.

456 J. PAILLARD
On peut donc, semble-t-il, avec une bonne approximation, admettre que les effets centraux qui affectent une réponse H d'amplitude donnée influencent une réponse T, de mêmes caractéristiques, de façon identique.
d) La valeur de la SE ou de la SM, comme test explorateur de l'excitabilité d'un centre moteur, va dépendre enfin d'une définition convenable de la frange des éléments atteints subliminairement par rapport à la population des éléments effectivement activés par la stimu-lation.
L'étendue de la population animée par la volée afférente s'accroît progressivement avec l'intensité de la stimulation. Le rapport du nombre des éléments actifs à celui des éléments atteints subliminairement déter-mine, dans une large mesure, l'aptitude de la stimulation à tester les effets de facilitation ou de dépression qui s'exercent directement sur la voie finale commune.
A ce sujet, la présence d'une réponse motrice primaire précédant la réponse H, ne doit pas, en toute rigueur, être considérée comme indifférente. La mobilisation de fibres efférentes par la SE aboutit en effet à un masquage de l'activité d'une partie de la population nerveuse centrale — Ce masquage peut aussi bien porter sur les cléments normalement activés que sur ceux de la frange disponible, dans des proportions respectives difficiles à préciser. Il n'est pas évident que l'on puisse supposer les fibres également réparties dans le contingent des éléments actifs et dans la frange subliminaire; on risque donc une certaine distorsion des indications fournies par les modifications de l'amplitude du réflexe. Notons cependant que cette cause d'erreur ne nous est pas apparue très importante et que la présence d'une réponse M peut même parfois être très utile à considérer comme signe d'instabilité du placement de l'électrode.
D'autre part, l'importance de la population activée n'est pas seule-ment dépendante de l'intensité de la volée afférente, mais elle varie également avec le niveau d'excitabilité de base de la population inter-rogée. Ce niveau peut varier dans de larges limites suivant les individus: lorsqu'on passe du normal au pathologique, parmi les normaux eux-mêmes et, chez un même sujet, suivant son état de « nervosité » ou tout simplement de vigilance.
Les seuils de stimulation sont trop dépendants de facteurs périphéri-ques, variables d'un sujet à l'autre et chez un même sujet d'une expérience à l'autre, pour fournir un critère valable d'appréciation de ce niveau de base (placement de l'électrode, résistance des tissus interposés etc. . . .).
Nous avons tenté, dans nos expériences, de tenir compte de l'accrois-sement d'amplitude de la réponse réflexe lorsqu'on lui surajoute une contraction volontaire, brève et intense, du muscle interrogé. Elle permet d'apprécier grossièrement l'étendue de la frange subliminaire touchée par la volée afférente. Les différences individuelles sont assez considé-rables sous ce rapport. Néanmoins, nous avons toujours travaillé avec des réponses nous donnant, sous l'effet de la mise en activité volontaire

Analyse du réflexe de HOFFMANN chez l'Homme. 457
du centre moteur, un accroissement de l'amplitude initiale du réflexe d'au moins 300%. (Il atteint chez certains sujets plus de 1000%.) Réciproquement, la mise en activité de la musculature antagoniste doit complètement annuler la réponse initiale (voir fig. 7A).
Le tracé des courbes Amplitude (réponses H et M). — Intensité de la stimu-lation (fig. 3) est sans doute susceptible de fournir une indication utile sur les mo-difications du niveau de base de l'excitabilité centrale. En effet, la position relative des courbes de réponse H et de réponse M dépend, dans une certaine mesure, de la plus ou moins grande facilité de recrutement des éléments centraux comparée à celui, normalement plus stable, des fibres motrices. Les indications, ainsi obtenues, deviendraient alors susceptibles de fournir la base d'une typologie du genre de celle signalée par SCHENK (1951). Les contingences propres à la stimulation transcutanée rendent cependant difficiles les appréciations rigoureuses; l'emploi d'une électrode de profondeur au contact du nerf (MAGLADERY et coll. 1950) devient alors recommandable.
Le contrôle de disponibilité d'une frange subliminaire suffisante s'avère donc nécessaire. On peut, d'autre part, compter sur une homogénéité satisfaisante de la population interrogée. La figure 6, en particulier 6A et 6C, montre l'homogénéité de recrutement des éléments progressivement actifs: la forme générale de la réponse ne se trouve pas altérée. Le test ainsi défini se présente donc avec des garanties de validité suffisantes pour légitimer son utilisation (BENSON et JEFFERSON 1952, RALL 1954).
L'utilisation de la SM introduit toutefois un important facteur de complication. L'intervention du récepteur périphérique ajoute, en effet, à la notion de frange subliminaire des éléments centraux, celle d'une frange des éléments périphériques recrutables. La sensibilité de ces derniers se trouve affectée en permanence par le jeu des influences spécifiques agissant sur la tension des myofibrilles fusoriales. Une série de travaux récents vient de préciser l'importance de ces régulations qui empruntent le circuit périphérique du fuseau neuromusculaire (HUNT et KUFFLER 1950, HUNT 1952, ELDRED et coll. 1953). Nous avons abordé ailleurs (PAILLARD 1953) l'analyse de ce type d'influence chez l'Homme.
Ainsi se trouvent intriqués, dans l'utilisation de la SM, les effets qui s'exercent au niveau du motoneurone — et qui posent alors les mêmes problèmes que ceux qui ont déjà été soulevés à propos de la SE — et les effets qui portent sur les modifications de la volée afférente attribu-ables aux fluctuations propres de la sensibilité des récepteurs périphéri-ques de tension.
Si l'on admet, et nombreux sont les arguments qui semblent actuelle-ment légitimer cette manière de voir, l'identité des éléments centraux intéressés par la SM et par la SE, l'utilisation conjointe de ces deux tests s'avère d'un très grand intérêt.
Les modifications de la réponse T qui s'accompagnent d'altérations de la réponse H dans le même sens, peuvent être rapportées à des

458 J. PAILLARD
influences qui s'exercent directement sur le centre moteur. Par contre, les modifications spécifiques à la réponse T intéressent des influences imputables à une modification de l'état de sensibilité des récepteurs périphériques (PAILLARD 1953). La comparaison systématique des résul-tats obtenus dans l'emploi de ces deux modes d'exploration fournit donc la base d'une information très utile concernant les influences spécifiques qui s'exercent directement sur la sensibilité fusoriale (système gamma, LEKSELL 1945, HOFFMANN, P. 1951).
e) Un dernier point mérite enfin d'être souligné qui rejoint la remarque précédente.
La stabilité des réponses à la SE est souvent très remarquable dans les conditions de fixation que nous utilisons; les variations excèdent rarement 10 à 20% pour les intensités de stimulation choisies. La réponse à la SM est par contre notoirement plus instable. Cette instabilité se manifeste très diversement d'un sujet à l'autre, et chez un même sujet d'une expérience à l'autre. Elle peut varier dans des limites importantes, 10 à 50% et davantage. Elle ne doit pas être mise uniquement sur le compte des imprécisions de définition de la stimulation mécanique. Nous avons pu montrer, en effet, que ces fluctuations affectaient, dans le même sens et avec un parallélisme souvent étonnant, les réponses évoquées simultanément par une SM sur le membre symétrique ou dans toute autre région du corps (PAILLARD 1953b).
Il nous est apparu légitime de les attribuer à une influence spécifique qui exerce globalement ses effets dans toute la musculature sur la tonicité des myofibrilles intra-fusoriales (PAILLARD 1953b).
En conclusion. Propagés le long des mêmes voies de conduction rapide du groupe I (Lloyd 1943), les messages provoqués par une SE du nerf ou par une SM du tendon paraissent atteindre monosynaptiquement les motoneurones et s'expriment musculairement par des réponses électromyographiques de caractéristiques identiques en forme, en durée et en latence.
S'il semble légitime d'utiliser la SE comme un test direct de l'excita-bilité des éléments centraux, par contre, l'emploi de la SM doit tenir compte des fluctuations imputables aux actions qui s'exercent directe-ment sur la sensibilité des récepteurs périphériques.
La comparaison des deux réponses peut constituer un test différentiel particulièrement précieux pour l'appréciation des propriétés spécifiques du système d'innervation des myofibrilles intra-fusoriales.
II. Effets retardés consécutifs à une stimulation électrique et à une stimulation mécanique.
On sait, depuis la découverte initiale de la « Hemmungsphase » consécutive à une réponse réflexe mécaniquement ou électriquement provoquée (HOFFMANN 1918), que les effets secondaires associés à de

Analyse du réflexe de HOFFMANN chez l'Homme. 459
telles stimulations peuvent affecter l'excitabilité des motoneurones durant une longue période après la décharge initiale. L'étude de la « période de silence » a, depuis, fait l'objet de nombreux travaux (voir bibliogra- phie dans MERTON 1951 et WESSLAU 1952) sur l'Homme et sur l'animal.
Les effets conditionnants d'un premier choc, explorés par un second choc-test distribué sur le même nerf ou sur le même tendon ont également suscité de nombreuses investigations en neurophysiologie tant animale qu'humaine (Travaux d'HOFFMANN et de ses élèves). Certains aspects en ont récemment été repris chez l'Homme par divers auteurs (SCHENK 1951, MAGLADERY et coll. 1951, PAILLARD 1951).
Utilisant alternativement soit la SE, soit la SM comme test explora-teur, nous avons repris l'étude systématique des effets retardés consécutifs à une première stimulation électrique ou mécanique dans l'espoir de préciser les divers facteurs responsables de ces influences prolongées.
a) Aspects généraux des cycles d'excitabilité. L'intensité de la stimulation électrique réflexogène étant fixée à la
valeur 1,2 subliminaire pour la réponse motrice primaire (fig. 4), nous ajustons l'intensité du choc mécanique pour obtenir une réponse T de même amplitude que la réponse H évoquée par la SE. Les courbes des variations d'amplitude de la réponse au choc explorateur en fonction des écarts séparant ce dernier du premier choc conditionnant se présentent suivant divers aspects (voir fig. 7 et 8).
l ° Avec l 'u t i l i sa t ion d 'un choc- tes t é lec tr ique. a) Nous notons l'allure très caractéristique de la courbe obtenue avec
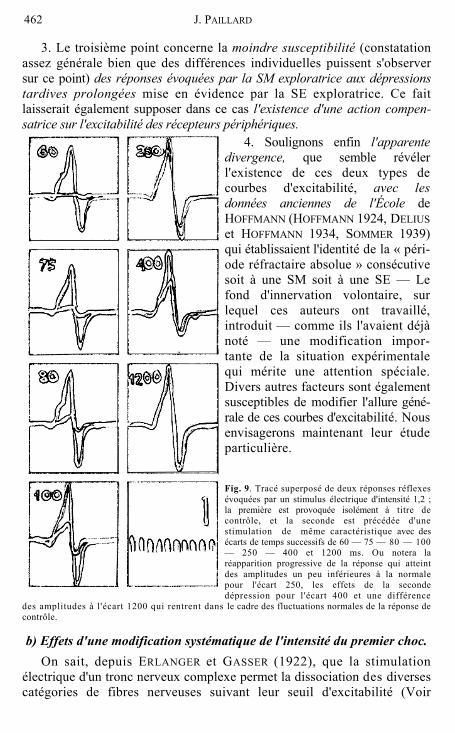
l’emploi d'un choc conditionnant de même nature (courbes SE + SE): longue période d'inhibition totale (de 55 à 120 ms suivant les sujets) suivie d'un retour progressif aux amplitudes initiales (150 à 300 ms), avec une nouvelle dépression tardive plus ou moins prolongée qui précède le retour progressif aux niveaux de l'amplitude de contrôle. (Voir fig. 9.) (Nous reviendrons ultérieurement sur les caractéristiques de ces courbes pour les écarts très courts.)
b) Avec l'emploi d'un choc conditionnant mécanique (courbes SM + SE), la courbe garde ses caractéristiques essentielles, mais avec une dominante générale d'inhibition plus ou moins marquée suivant les sujets. Le retour aux amplitudes normales exige parfois plusieurs secondes.
2° Avec l 'uti l isat ion d 'un choc-test mécanique. Quelle que soit la nature, électrique ou mécanique, du choc condi-
tionnant (courbes SM + SM et SE + SM) nous obtenons un type de courbe différent du précédent, caractérisé par une longue période de suppression (150 à 320 ms suivant les sujets) suivi d'un retour souvent
460 J. PAILLARD
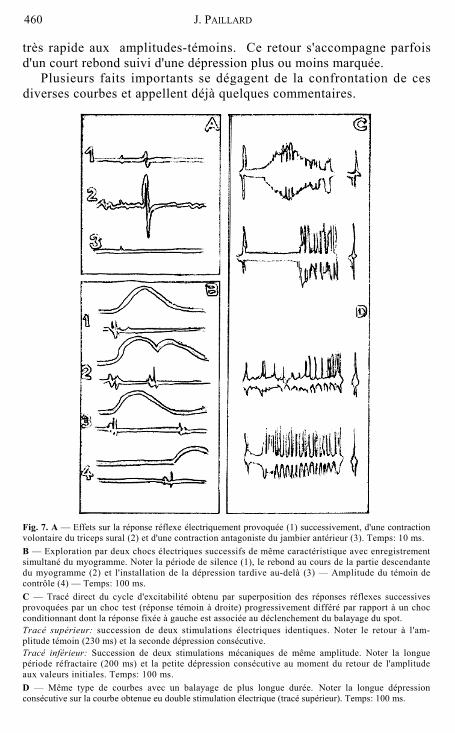
très rapide aux amplitudes-témoins. Ce retour s'accompagne parfois d'un court rebond suivi d'une dépression plus ou moins marquée.
Plusieurs faits importants se dégagent de la confrontation de ces diverses courbes et appellent déjà quelques commentaires.
Fig. 7. A — Effets sur la réponse réflexe électriquement provoquée (1) successivement, d'une contraction volontaire du triceps sural (2) et d'une contraction antagoniste du jambier antérieur (3). Temps: 10 ms. B — Exploration par deux chocs électriques successifs de même caractéristique avec enregistrement simultané du myogramme. Noter la période de silence (1), le rebond au cours de la partie descendante du myogramme (2) et l'installation de la dépression tardive au-delà (3) — Amplitude du témoin de contrôle (4) — Temps: 100 ms. C — Tracé direct du cycle d'excitabilité obtenu par superposition des réponses réflexes successives provoquées par un choc test (réponse témoin à droite) progressivement différé par rapport à un choc conditionnant dont la réponse fixée à gauche est associée au déclenchement du balayage du spot. Tracé supérieur: succession de deux stimulations électriques identiques. Noter le retour à l'am-plitude témoin (230 ms) et la seconde dépression consécutive. Tracé inférieur: Succession de deux stimulations mécaniques de même amplitude. Noter la longue période réfractaire (200 ms) et la petite dépression consécutive au moment du retour de l'amplitude aux valeurs initiales. Temps: 100 ms. D — Même type de courbes avec un balayage de plus longue durée. Noter la longue dépression consécutive sur la courbe obtenue eu double stimulation électrique (tracé supérieur). Temps: 100 ms.

Analyse du réflexe de HOFFMANN chez l'Homme. 461
1. Les deux types de courbes sont essentiellement déterminés par la nature du choc explorateur. En admettant l'identité des éléments centraux interrogés par la SE et par la SM, il nous faudrait attribuer (voir 1ère Partie) les différences observées à une modification correspondante de l’état de sensibilité des récepteurs périphériques — Ainsi l'inefficacité de la SM exploratrice durant les premières périodes des courbes SE + SM et SM + SM devrait être mise sur le compte d'une inexcitabilité prolongée des récepteurs fusoriaux.
Fig. 8. Ensemble des courbes d'excitabilité obtenues chez 3 sujets différents par double choc de même amplitude et progressivement décalés — SE: stimulation électri-que — SM: stimulation mécanique En trait gras: courbes SE + SE dont on peut noter lapériode réfractaire absolue et le rebond suivi d'unenouvelle dépression. En traits fins: courbes SM + SE qui gardent les carac-téristiques de la précédente avec une dominantegénérale d'inhibition. En gros pointillé: courbes SM + SM avec leur longuepériode réfractaire absolue, leur pente rapide de retour àla normale et l'existence d'un petit rebond suivi d'unecourte dépression. En fin pointillé: courbes SE + SM identiques aux pré-cédentes avec accentuation du petit rebond consécutif.Les courbes hachurées indiquent l'étendue et la formede la courbe myographique consécutive au premierchoc. Latence de 10 à 15 ms, sommet autour de 100 ms etretour à la tension initiale en 200 ms environ avecparfois présence d'un renflement tardif. (voir explicationdans le texte) Amplitude en pourcent de la réponse decontrôle — Écarts en ms.
La considération du myo-gramme, résultant de la con-traction consécutive à la première stimulation, prend ici toute son importance (voir fig.7 et 8). Il évolue en général sur plus de 300 ms, après une période de latence de 10 à 15 ms et un sommet atteint en 110—130 ms environ. On sait (MATTHEWS 1933, HUNT et KUFFLER 1951) que l'excitabilité des récepteurs fusoriaux se trouve modifiée par l'état de raccourcissement du muscle. Le silence, ou le ralen- tissement de la décharge fuso- riale durant la phase ascendante du myogramme, est caractéristi-que du fonctionnement de ce type de récepteurs « en parallèle » avec les fibres musculaires — Les faits que nous observons semble- raient indiquer qu'en général le choc mécanique ne deviendrait efficace que lorsque le muscle aurait repris sa longueur initiale. Nous reviendrons ultérieurement sur ce point important à considérer pour l'interprétation de la période silencieuse.
2. Le second point digne d'être noté concerne les effets différents d'une SE et d'une SM conditionnantes. — La dominante d'inhibition associée à la seconde implique une composition différente des volées afférentes évoquées par ces deux types de stimulation.
462 J. PAILLARD
3. Le troisième point concerne la moindre susceptibilité (constatation assez générale bien que des différences individuelles puissent s'observer sur ce point) des réponses évoquées par la SM exploratrice aux dépressions tardives prolongées mise en évidence par la SE exploratrice. Ce fait laisserait également supposer dans ce cas l'existence d'une action compen-satrice sur l'excitabilité des récepteurs périphériques.
pour l'écart dépression pour l
4. Soulignons enfin l'apparente divergence, que semble révéler l'existence de ces deux types de courbes d'excitabilité, avec les données anciennes de l'École de HOFFMANN (HOFFMANN 1924, DELIUS et HOFFMANN 1934, SOMMER 1939) qui établissaient l'identité de la « péri- ode réfractaire absolue » consécutive soit à une SM soit à une SE — Le fond d'innervation volontaire, sur lequel ces auteurs ont travaillé, introduit — comme ils l'avaient déjà noté — une modification impor- tante de la situation expérimentale qui mérite une attention spéciale. Divers autres facteurs sont également susceptibles de modifier l'allure géné- rale de ces courbes d'excitabilité. Nous envisagerons maintenant leur étude particulière.
Fig. 9. Tracé superposé de deux réponses réflexes évoquées par un stimulus électrique d'intensité 1,2 ; la première est provoquée isolément à titre de contrôle, et la seconde est précédée d'une stimulation de même caractéristique avec des écarts de temps successifs de 60 — 75 — 80 — 100 — 250 — 400 et 1200 ms. Ou notera la réapparition progressive de la réponse qui atteint des amplitudes un peu inférieures à la normale
250, les effets de la seconde 'écart 400 et une différence
des amplitudes à l'écart 1200 qui rentrent dans le cadre des fluctuations normales de la réponse de contrôle.
b) Effets d'une modification systématique de l'intensité du premier choc. On sait, depuis ERLANGER et GASSER (1922), que la stimulation
électrique d'un tronc nerveux complexe permet la dissociation des diverses catégories de fibres nerveuses suivant leur seuil d'excitabilité (Voir

Analyse du réflexe de HOFFMANN chez l'Homme. 463
LLOYD 1943, BROCK et coll., 1951). Nous avons, comme pour les courbes précédemment décrites, utilisé une réponse H évoquée par une SE exploratrice d'intensité 1,2 et une réponse T équivalente. Nous avons progressivement fait varier l'intensité de stimulation du premier choc, à partir de valeurs nettement subliminaires, jusqu'aux intensités supporta-bles sans désagrément par le sujet.
Un accroissement progressif de l'intensité de la SE nous permet de décomposer la nature des volées afférentes résultantes en diverses catégories :
a) Les intensités subliminaires pour la décharge réflexe (0,75 à 1) animeront essentiellement les fibres sensorielles du groupe I (LLOYD, 1943).
b) Les stimulations associées à une décharge réflexe, mais sous-liminaires pour la stimulation directe des fibres motrices (intensité de 1 à 1,2), s'accompagneront progressivement d'un recrutement plus important des fibres du groupe I et du groupe II pour les intensités les plus fortes (LLOYD, 1943a). A la contraction résul- tante s'associeront également les afférences proprioceptives mises en jeu par l'effet du raccourcissement musculaire.
c) Avec des intensités supraliminaires pour la réponse M (1,25) interviendront, outre le recrutement plus important de fibres d'origine cutanée et de fibres du groupe II et même III, les volées antidromiques associées à la stimulation directe des fibres efférentes et les complications tardives provenant des effets réflexes de la contraction musculaire primaire.
Les effets d'un accroissement de la SM sont beaucoup moins facilement prévi-sibles. Le seuil mécanique de réponse des récepteurs tendineux do GOLGI est vrai-semblablement plus élevé que celui des terminaisons primaires et secondaires du fuseau (MATTHEWS 1933, HUNT et KUFFLER 1951).
De ce fait, on doit attendre une composition différente des volées provoquées par la SE et par la SM. Pour de faibles intensités de stimulation, la SM anime vraisemblablement davantage de fibres du groupe II (terminaisons secondaires) et moins de fibres du groupe I (GOLGI) que ne le fait la SE.
Pour la clarté de l'exposé, nous distinguerons cinq périodes successives dans le cycle d'excitabilité précédemment décrit (voir fig. 11).
Période I (0 à 2,5 ms). Un masquage progressif de la réponse H peut déjà être observé pour des intensités subliminaires d'une première SE; il est parfois presque total dès avant l'apparition d'une réponse musculaire au premier choc. Cette particularité permet de l'attribuer à une réfractorité des fibres nerveuses. La présence d'une première réponse masque ensuite ces effets, qui ne peuvent d'ailleurs que s'accentuer par l'intervention des réfractorités propres aux jonctions neuro-musculaires. Les courbes SE + SM et SM + SE présentent ce phénomène avec beaucoup moins de netteté que dans le cas de stimulations successives identiques (voir fig. 10).
Période II (2,5 à 20 ms). Pour les courbes SE + SE: déjà aux inten-sités (I = 0,60) qui donnent les premières manifestations de masquage de la phase précédente se manifeste une super-normalité qui gagne tout d'abord en importance et atteint son maximum pour des intensités 0,85 aux écarts de 3 à 8 ms (amplitude 120 à 160% du contrôle). Ces effets se
464 J. PAILLARD
trouvent ensuite en compétition avec des influences dépressives qui vont accentuer leur action jusqu'aux valeurs seuils de l'intensité. Dès l'appari-tion des premières décharges, on assiste à un effondrement de cette première « bosse » puis à une disparition complète de toute réponse pour des intensités du premier choc de 1,4 environ.
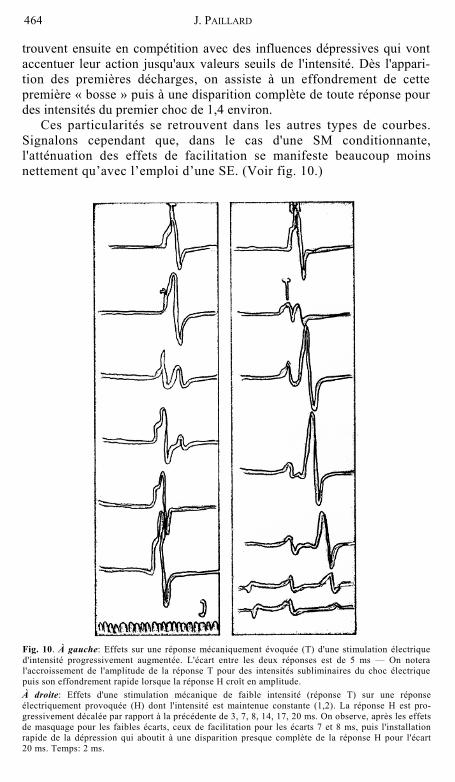
Ces particularités se retrouvent dans les autres types de courbes. Signalons cependant que, dans le cas d'une SM conditionnante, l'atténuation des effets de facilitation se manifeste beaucoup moins nettement qu’avec l’emploi d’une SE. (Voir fig. 10.)
Fig. 10. À gauche: Effets sur une réponse mécaniquement évoquée (T) d'une stimulation électrique d'intensité progressivement augmentée. L'écart entre les deux réponses est de 5 ms — On notera l'accroissement de l'amplitude de la réponse T pour des intensités subliminaires du choc électrique puis son effondrement rapide lorsque la réponse H croît en amplitude. À droite: Effets d'une stimulation mécanique de faible intensité (réponse T) sur une réponse électriquement provoquée (H) dont l'intensité est maintenue constante (1,2). La réponse H est pro-gressivement décalée par rapport à la précédente de 3, 7, 8, 14, 17, 20 ms. On observe, après les effets de masquage pour les faibles écarts, ceux de facilitation pour les écarts 7 et 8 ms, puis l'installation rapide de la dépression qui aboutit à une disparition presque complète de la réponse H pour l'écart 20 ms. Temps: 2 ms.

Analyse du réflexe de HOFFMANN chez l'Homme. 465
Cette supernormalité précoce a déjà été signalée par ECCLES (1946), LLOYD (1946), et GRANIT (1950) chez l'animal. MAGLADERY et coll. (1951) ont également mentionné sa présence chez l'Homme. BROOKS et coll. (1950) en ont discuté l'origine. Les résultats obtenus chez l'Homme s'avèrent sur ce point parfaitement identi-fiables à ceux qui ont été rapportés chez l'animal dans des conditions de stimulation homosynaptique. Avec HOFFMANN et RIOTTE (1941) nous pensons qu'il est difficile d'envisager un pur effet de sommation se manifestant à des écarts aussi grands que 15 à 20 ms. Les faits observés semblent bien rendre compte de cette compétition des influences facilitatrices et inhibitrices conduites par les voies du groupe I et exerçant leurs effets très précocement sur les motoneurones (LAPORTE et LLOYD 1952). La différence observée à ce sujet entre les effets de la SE et de la SM pourrait rendre compte d'une richesse moins grande de la volée afférente, évoquée par la SM, en fibres inhibitrices du groupe I (en provenance des récepteurs tendineux de GOLGI dont le seuil mécanique de réponse est plus élevé).
Dès l'apparition de la décharge réflexe intervient la réfractorité des éléments activés, associée à la subnormalité mise en évidence par GASSER et GRAHAM (1933) et aujourd'hui classique. Elle masque peu à peu le jeu des autres influences. Dès que l’amplitude de la réponse au choc conditionnant dépasse celle de la réponse au test explorateur on observe une disparition complète de cette dernière. La sub- normalité consécutive à la première décharge peut alors entièrement rendre compte de cette extinction.
Période III (15—120 ms). Avec les premières manifestations d'activité des fibres de bas seuils apparaissent les signes d'une dépression entre 50 et 100 ms qui s'accuse de plus en plus lorsqu'on accroît l'intensité de la SE (voir fig. 12).
Dès le franchissement du seuil de la réponse réflexe on assiste, du côté des faibles écarts, à un effondrement rapide des amplitudes allant jusqu'à la disparition complète de ces dernières pour dos intensités supérieures à 1,25 environ.
Du côté des écarts les plus longs, on observe une extension progressive de l'inhibition qui se stabilise finalement à des valeurs pouvant encore varier de 90 à 130 ms suivant les sujets.
Cette longue dépression a souvent retenu l'attention des chercheurs. Elle constitue, en effet, la manifestation de cette « période de silence » dont, depuis la découverte de Hoffmann (1918), on a souvent discuté le mécanisme. L'analyse ci-dessus permet, dans une certaine mesure, de décomposer l'intervention des divers facteurs mis en jeu.
Pour un premier choc subliminaire, l'existence de cette première dépression indique, sans aucun doute, des effets retardés inhibiteurs, attribuables aux actions de messages conduits dans les fibres du groupe I. Les analyses de l'expérimentation animale nous donnent peu d'éclaircissements sur ce point.
Au-delà du seuil vont progressivement intervenir divers éléments susceptibles de contribuer à cette dépression:
1° La dépression due à la subnormalité consécutive à la décharge des éléments activés par la première réponse et dont on sait que les effets peuvent influencer l'excitabilité des motoneurones durant une longue période (Hughes et Casser 1934). Elle contribue pour une large part, du côté des faibles écarts, au

466 J. PAILLARD
creusement de la «vallée» dont SCHENK (1951) avait analysé le premier la nature chez l'Homme et pour une part moindre, à la dépression observable avec les écarts plus longs. Elle a longtemps été invoquée comme l'interprétation exclusive de l'origine de la période de silence (HOFFMANN 1934, HOFFMANN et coll. 1934, GASSER 1939, BREMER et coll. 1942, BERNHARD 1947). Elle en constitue, sans aucun doute, un facteur essentiel.
2e Les effets retardés inhibiteurs de la volée afférente elle-même sont moins bien connus. Les premières manifestations d'inhibition, attribuables à des fibres du groupe I, accentuent vraisemblablement leurs effets aux fortes intensités du côté des grands écarts. Nous manquons d'informations sur les modalités d'action des fibres appartenant aux autres groupes.
3° Les effets réflexes introduits par la contraction musculaire. Il est à noter que le myogramme de cette contraction apparaît avec une latence de 10 à 15 ms et que sa phase ascendante couvre approximativement toute la période que nous avons actuellement à considérer. Son sommet se situe en effet suivant les sujets entre 100 et 130 ms.
Les effets centraux de cette contraction ont fait l'objet de nombreuses études (GRANIT 1950, HAGBARTH et NAESS 1950), les analyses de MATTHEWS (1933) et de KUFFLER et HUNT (1950) nous permettent de prévoir durant cotte phase ascendante:
a) une décharge précoce de propriocepteurs, qui, compte tenu des délais de transmission, se situerait vraisemblablement autour de 20 à 30 ms. Nous ne pouvons en apercevoir les effets étant donné la présence de la subnormalité maximum dans cette région.
b) une accélération de la décharge des récepteurs du groupe B de MATTHEWS (1933) durant toute la phase ascendante et dont les manifestations centrales sont celles d'une inhibition (GRANIT et SUURSOET 1949, GRANIT 1950, HUNT 1953). On pourrait attribuer à cet élément le creusement de la « vallée » du côté des écarts les plus larges.
c) une cessation ou un ralentissement de la décharge des messages activa- teurs en provenance des fuseaux, au moins pour les faibles tensions initiales.
Ces faits, sans aucun doute, permettent de rendre compte du prolongement de la dépression vers les longs délais et de son accentuation avec l'accroissement de l'intensité de stimulation du premier choc. Les observations récentes de MERTON (1951) sur les rapports de la durée de la contraction musculaire avec l'extension de la période de silence plaident également en faveur d'une telle interprétation.
4° A un certain niveau de la stimulation, les fibres motrices se trouvent directement stimulées. Les effets du blocage antidromique résultant s'ajoutent à ceux de la subnormalité des éléments activés suivant un décours temporel approximativement identique (ECCLES et PRITCHARD 1937, BROCK et coll. 1951), les manifestations plus prolongées (au-delà de 100 ms) étant sans doute moins importantes que celles de la subnormalité. Ces facteurs de dépression ne semblent pas intervenir de façon très déterminante sur l'allure de nos courbes; ils interviennent à un stade où, pratiquement, la dépression est déjà bien installée sous l'influence d'autres facteurs.
Il ressort de cette analyse qu'une interprétation cohérente de la période de silence doit, à notre sens, obligatoirement tenir compte de toutes ces influences simultanément présentes. Aucune interprétation, basée sur l'influence exclusive de l'un ou de l'autre des facteurs mis en jeu, ne saurait être complète.
Période IV (130 à 300 ms). Cette phase correspond à ce qu'on avait pu improprement appeler «la période réfractaire relative», car elle se manifeste
Analyse du réflexe de HOFFMANN chez l'Homme. 467
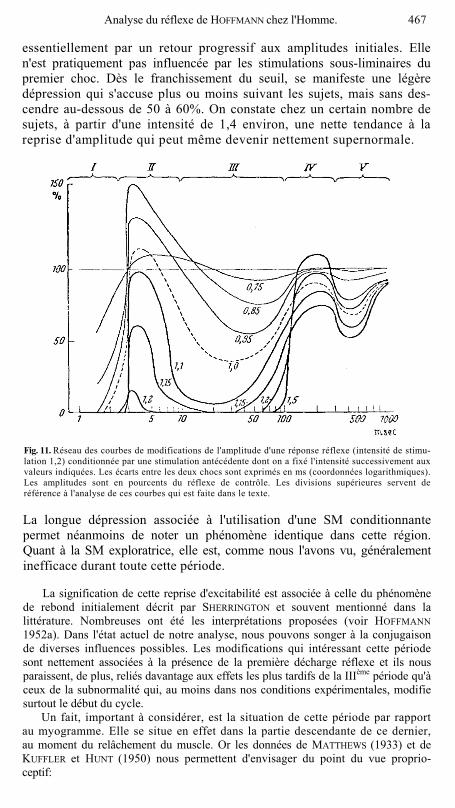
essentiellement par un retour progressif aux amplitudes initiales. Elle n'est pratiquement pas influencée par les stimulations sous-liminaires du premier choc. Dès le franchissement du seuil, se manifeste une légère dépression qui s'accuse plus ou moins suivant les sujets, mais sans des-cendre au-dessous de 50 à 60%. On constate chez un certain nombre de sujets, à partir d'une intensité de 1,4 environ, une nette tendance à la reprise d'amplitude qui peut même devenir nettement supernormale.
Fig. 11. Réseau des courbes de modifications de l'amplitude d'une réponse réflexe (intensité de stimu-lation 1,2) conditionnée par une stimulation antécédente dont on a fixé l'intensité successivement aux valeurs indiquées. Les écarts entre les deux chocs sont exprimés en ms (coordonnées logarithmiques). Les amplitudes sont en pourcents du réflexe de contrôle. Les divisions supérieures servent de référence à l'analyse de ces courbes qui est faite dans le texte.
La longue dépression associée à l'utilisation d'une SM conditionnante permet néanmoins de noter un phénomène identique dans cette région. Quant à la SM exploratrice, elle est, comme nous l'avons vu, généralement inefficace durant toute cette période.
La signification de cette reprise d'excitabilité est associée à celle du phénomène de rebond initialement décrit par SHERRINGTON et souvent mentionné dans la littérature. Nombreuses ont été les interprétations proposées (voir HOFFMANN 1952a). Dans l'état actuel de notre analyse, nous pouvons songer à la conjugaison de diverses influences possibles. Les modifications qui intéressant cette période sont nettement associées à la présence de la première décharge réflexe et ils nous paraissent, de plus, reliés davantage aux effets les plus tardifs de la IIIème période qu'à ceux de la subnormalité qui, au moins dans nos conditions expérimentales, modifie surtout le début du cycle.
Un fait, important à considérer, est la situation de cette période par rapport au myogramme. Elle se situe en effet dans la partie descendante de ce dernier, au moment du relâchement du muscle. Or les données de MATTHEWS (1933) et de KUFFLER et HUNT (1950) nous permettent d'envisager du point du vue proprio-ceptif:

468 J. PAILLARD
a) Un arrêt de la décharge des récepteurs du groupe B: la suspension de ces influences inhibitrices pourrait à elle seule être responsable d'un certain rebond (CREED et coll. 1932).
b) Une reprise de la décharge des fuseaux et par suite un relèvement de l'ex-citabilité centrale: c'est cette dernière interprétation qui est le plus souvent invoquée par les tenants de la théorie périphérique de la période de silence (voir MERTON 1951).
L'inefficacité de la SM durant cette période apparaît quelque peu contra-dictoire avec cette dernière interprétation. Il faudrait en effet supposer qu'un
Fig. 12. Effets d'une augmentation progressive de l'intensité du choc électrique conditionnant sur l'amplitude de la réponse réflexe exploratrice évoquée par une stimulation électrique maintenue constante (intensité 1,2). Chaque courbe correspond à un écart déterminé entre les deux chocs. Ces écarts successifs ont été replacés à gauche de la figure par rapport à la courbe type amplitude-écarts obtenue avec deux stimulations de caractéristiques identiques: On notera les altérations caractéristiques des diverses courbes pour les valeurs subliminaires et supraliminaires de l'intensité du premier choc. Explication dans le texte. Intensité en multiples du seuil de la réponse réflexe au premier choc — Amplitude en pour cent de la réponse réflexe exploratrice de contrôle. Chaque point de la courba constitue la moyenne de 15 à 30 mesures.
étirement du muscle deviendrait incapable d'évoquer le réflexe dans une période où l'excitabilité des récepteurs fusoriaux se trouverait augmentée.
D'autres éléments de notre étude viendront ultérieurement éclairer ce point.
Période V. (au-dessus de 300 ms). Avec la SE exploratrice, on constate une dépression plus ou moins prolongée suivant les sujets. La position du minimum peut varier dans d'assez larges limites. Nous aurions tendance (d'après le contingent de sujets que nous avons examiné) à le situer plus tôt que ne le signalent MAGLADERY et coll. (1952) : 450 ms au lieu de 600 ms. On peut même constater chez certains sujets un creusement initial beaucoup plus abrupt et précoce (autour de 300 ms).
Les prémisses de cette dépression tardive apparaissent déjà pour les valeurs subliminaires de la stimulation. Elle se creuse ensuite progressive-ment et tend, avec les intensités plus fortes, à déplacer son minimum vers des délais plus courts et parfois aussi à relever son niveau général.

Analyse du réflexe de HOFFMANN chez l'Homme. 469
Avec la SM exploratrice, on assiste le plus généralement à un retour rapide aux amplitudes initiales. Pour les intensités plus fortes du premier choc et, en particulier, avec l'utilisation d'une SE conditionnante, on voit s'amorcer un rebond suivi d'une dépression.
Cette période est donc caractérisée par la « late depression » décrite par MAGLADERY et coll. (1952) et que PAILLARD et TURNER (1953) signalèrent indépendamment un peu plus tard. Son origine reste obscure.
a) Les effets prolongés de la subnormalité initiale pourraient être invoqués. Cette hypothèse semble toutefois exclue par les observations de MAGLADERY et coll. (1952) dans des cas pathologiques. Elles indiquent en effet la possibilité d'une disparition de ces effets dépressifs sans que la subnormalité associée à la décharge des éléments activés puisse apparemment être mise en cause.
b) La possibilité de répercussions tardives associées à des circuits centraux plus complexes a été discutée par LANGUTH et coll. (1952) mais reste hypothétique. La participation des messages conduits par les fibres du groupe I semble en tous cas confirmée par notre analyse.
c) La contraction antécédente ne peut intervenir que par ses propres effets retardés et elle mettrait en jeu des processus relevant de ceux déjà envisagés dans le précédent paragraphe.
d) La dépression plus profonde et plus précoce qui semble suivre les rebonds importants fait vraisemblablement intervenir des effets de subnormalité associés aux éléments activés durant cette période. Le myogramme présente le plus souvent, dans ce cas, un renflement caractéristique (« myotatic appendage » FULTON 1926, NEZ DE FUNKE) qui semble justifier la présence d'une décharge secondaire.
e) L'élévation générale du niveau de la courbe, parfois constatée pour les fortes intensités du choc conditionnant, dépend de certains processus d' « after-discharge » prolongée sur lesquels nous n'avons encore que peu d'information. On peut sans doute rattacher à ces phénomènes, la « late discharge » décrite par MAGLADERY et coll. (1950) et dont nous avons nous-même fréquemment noté la présence chez certains sujets.
f) Pour la dépression généralisée consécutive à une SM conditionnante, on peut sans doute invoquer la proportion importante d'éléments du groupe II dans la volée afférente qui en résulte. La participation, aujourd'hui bien démontrée (LLOYD 1943, HUNT 1953), de ces fibres au réflexe de flexion, les destine à un effet dépresseur du noyau du muscle extenseur sur lequel nous travaillons.
g) Quant à l'inaptitude de la SM exploratrice à tester ces influences dépressives prolongées, elle nous semble devoir être mise sur le compte d'une action com-pensatrice sur la sensibilité fusoriale dont la nature reste à préciser.
c) Effets d'une modification systématique de l'intensité de la stimulation exploratrice.
D'une façon générale, les courbes ne subissent pas d'altérations très importantes lorsqu'on fait varier le niveau du test explorateur.
Nous signalerons cependant que l'emploi d'un test légèrement suprali-minaire, comme dans les derniers travaux de MAGLADERY et coll. (1952), tend à favoriser les influences dépressives du type de celles que nous avons observées dans les périodes III et V avec des premiers chocs subliminaires.

470 J. PAILLARD
Ce fait explique, sans doute, la longue durée des périodes silencieuses rapportées par ces auteurs et éventuellement l'étalement de la dépression tardive autour de 600 ms.
D'autre part, les manifestations de rebond et les effets tardifs de relèvement de l'excitabilité sont d'autant plus nets que l'on travaille avec une intensité de choc plus élevée.
Aux très fortes intensités, interviennent les complications inhérentes à la présence de la réponse motrice primaire dont nous avons déjà parlé (voir première Partie). Les courbes ne subissent cependant pas d'altéra-tions très notables.
L'ensemble de ces données démontre l'existence d'une certaine homogénéité fonctionnelle de l'ensemble de la population du noyau moteur. Les courbes reflètent un état d'équilibre entre les éléments interrogés par le premier choc et ceux qui répondent au second. L'emploi d'une population d'étendue restreinte (faibles intensités de stimulation), tend à diminuer l'étendue de la frange subliminaire et favorisera la traduction des influences dépressives consécutives à la stimulation des fibres de bas seuils. Une stimulation exploratrice d'intensité plus grande favorise, par contre, l'étude des répercussions centrales de la contraction musculaire consécu-tive au premier choc.
d) Effets d'une mise en activité volontaire du centre moteur (Bahnung).
Une innervation volontaire du muscle tend à modifier les aspects du cycle d'excitabilité comme le montre bien l'ensemble des résultats de travaux de l'École de HOFFMANN et en particulier l'étude de SCHOEN (1951) sur la longueur de la période dite « réfractaire absolue » (période III de notre cycle). Il est spécialement instructif, dans ce cas, de tenir compte des modifications introduites dans l'allure du myogramme par l'état de tension permanente du muscle dont l'activité est entretenue volontaire-ment.
Pour une innervation moyenne constante, un accroissement de l'intensité du premier choc tend à donner progressivement son amplitude maximum au myogramme de la réponse réflexe; on constate un allongement correspondant de la période silencieuse. Ensuite s'installe un compromis temporaire entre le myogramme de la réponse primaire et celui de la réponse H. L'extinction progressive de cette dernière (voir fig. 2) amène un recul du myogramme résultant de 20 à 25 ms. La période silencieuse tend également à se raccourcir puis à se stabiliser enfin aux valeurs imposées par l'étendue du myogramme de la seule réponse primaire. C'est exac-tement ce que semble montrer l'analyse des courbes de SCHOEN (1951) concernant les modifications de la durée de la période silencieuse, (voir également J. PAILLARD 1955).
A intensité de choc constant, l'étendue et l'amplitude du myogramme correspondant vont dépendre du degré de la tension volontaire imposée au muscle. Il tend progressivement à diminuer d'amplitude au fur et à

Analyse du réflexe de HOFFMANN chez l'Homme. 471
mesure que s'accroît l'effort volontaire de tension isométrique. Nous signalerons même la possibilité d'obtenir une inversion de la déformation musculaire correspondante. Pour de fortes tensions initiales, en effet, le résultat d'un choc électrique n'est plus celui d'une contraction avec raccourcissement du muscle, mais l'effet prédominant devient alors celui du relâchement de la tension qui accompagne le brusque silence des éléments synchroniquement activés par la stimulation ou simultanément envahis par la décharge antidromique. On peut dans ce cas constater la diminution progressive de la durée de la période silencieuse qui se stabilise finalement à des valeurs voisines de celles de la période de subnormalité vraie des éléments centraux (voir courbes de SCHOEN 1951).
Les faits ainsi analysés donnent finalement entièrement raison aux inter-prétations initiales de HOFFMANN concernant la période de silence observée dans ces conditions de travail.
Ils rendent également compte, en les confirmant, des faits rapportés par MERTON (1951) concernant la corrélation entre le myogramme et retendue de la période de silence.
Ils établissent, dans une certaine mesure, la double origine de la dépression de la période III de notre cycle. La première serait associée à la subnormalité consécutive à la décharge des éléments centraux, la seconde dans nos conditions expérimentales tout au moins, à une influence dépressive d'origine périphérique provenant de la réponse des propriocepteurs musculaires à la déformation mécanique du muscle. Cette manière de voir rejoint d'ailleurs une conception de la période de silence aujourd'hui de plus en plus répandue (WESSLAU 1952, HOFFMANN 1952, LUSCHNAT 1954).
Ils expliquent également, notamment en ce qui concerne les cycles d'excitabilité relatifs aux SM, les divergences de nos résultats, avec les courbes de SOMMER (1939) par exemple. Outre les effets de la tension volontaire sur la déformation mécanique consécutive à une contraction du muscle, il nous faut, dans ce cas, tenir également compte d'une éventuelle mobilisation du système de fibres gamma, lors de l'innerva-tion volontaire du centre moteur. Les récepteurs fusoriaux se trouvent alors sensi-bilisés au cours de la déformation mécanique du muscle et ne subissent plus cette longue période d'inexcitabilité si caractéristique de nos courbes utilisant une SM exploratrice sur le tendon du muscle au repos.
e) Effets d'une modification passive de la tension musculaire. 1. Un accroissement de la tension du muscle par dorsiflexion passive du
pied tend, en général, à diminuer progressivement l'amplitude de la réponse H, souvent jusqu'à extinction totale (MAGLADERY et coll. 1950, PAILLARD 1953, ENGELHORN et HOFFMANN 1953).
La réponse T subit tout d'abord un accroissement d'amplitude (attri-buable aux modifications exogènes de la tension fusoriale) puis finalement une diminution (voir fig. 4c) (PAILLARD 1953).
Ces faits expriment l'existence d'une profonde dépression de l'exci-tabilité centrale associée à cette manoeuvre. Nous avons entrepris le tracé des courbes dans ces nouvelles conditions. La modification la plus caractéristique des cycles porte sur la période IV et sur le début de la

472 J. PAILLARD
période V. On observe en effet un rebond souvent considérable (variations d'amplitude de 130 à 150%), suivi d'une dépression immédiatement consécutive, parfois très accusée mais principalement dans le cas où la présence d'une post-décharge électromyographique se trouve révélée par la modification correspondante du myogramme. Ces manœuvres de flexion favorisent d'ailleurs, chez certains sujets, l'apparition de rebonds spontanés de l'activité électromyographique et la présence de post-décharges prolongées du type des « late-discharge » de MAGLADERY et coll. (1950). Plus l'intensité de la stimulation conditionnante est forte et plus ces effets consécutifs sont accentués.
2. Une diminution de la tension du muscle, par abaissement passif du pied sur la jambe a, en général, une répercussion beaucoup moins im-portante sur l'amplitude de la réponse au choc explorateur que la précé-dente. L'effet, bien que variable suivant les sujets, est généralement dans le sens d'un affaiblissement de la réponse. La modification du cycle est par contre toujours dans le sens d'une atténuation du rebond de la période IV. La courbe marque alors un simple palier qui se fond ensuite avec la dépression tardive.
L'ensemble do ces phénomènes relève des faits initialement rapportés par SHERRINGTON, concernant les manifestations inhibitrices de la « lengthening reaction ». L'importance de l' « inhibition autogénique » dans les coordinations réflexes a été soulignée, récemment encore, dans une série de travaux de l'École Scandinave (GRANIT 1950, HAGBARTH et NAESS 1950). La fonction, aujourd'hui bien admise, des récepteurs tendineux de GOLGI les désigne comme les acteurs principaux de ces effets d'inhibition observables au niveau du centre moteur.
La volée afférente restant constante (dans le cas de la SE tout au moins) il faut admettre une origine périphérique aux effets constatés dans ce type d'expé-rience. La position moyenne du pied, à laquelle nous nous référons dans toute cette étude, définit un certain état d'équilibre dans la compétition des influences facilita-trices et inhibitrices provenant du fonctionnement permanent des propriocepteurs musculaires (GRANIT 1950).
L'exaltation des phénomènes de rebond, dans les conditions expérimentales qui favorisent les influences inhibitrices d'origine proprioceptive, souligne encore l'importance vraisemblable de leur rôle dans la genèse de ces manifestations.
L'influence, enfin, de ces effets des manœuvres passives sur la 4ème période du cycle démontre le rôle prépondérant des facteurs périphériques dans la déter-mination de cette partie de la courbe. Ce fait suffit vraisemblablement à expliquer la similitude d'allure des courbes SE + SE et SM + SE, similitude qui tient essentiellement, outre l'identité des périodes initiales, à la présence de cette 4ème période de rebond modulée par la contraction musculaire réflexe. Nous avons, en effet, souligné précédemment l'identité morphologique des expressions musculaires de la réponse H et de la réponse T.
f) Effets des modifications du niveau de vigilance du sujet. Nous noterons, enfin, un effet particulier des altérations du niveau de
vigilance du sujet sur les courbes d'excitabilité. Nous avons suivi systématiquement une grande partie de nos expé-
riences avec contrôle électroencéphalographique de la vigilance du sujet.

Analyse du réflexe de HOFFMANN chez l'Homme. 473
L'assoupissement entraîne, en général, une chute du niveau d'excitabilité de base; un certain relèvement de l'intensité de stimulation est alors nécessaire.
L'aspect le plus caractéristique de cette situation expérimentale nous a semblé être la dominante générale d'inhibition qui caractérise les effets du choc conditionnant. La courbe s'effondre dans toutes ses phases, la phase IV (rebond) marque une petite bosse à peine perceptible suivie d'un retour très progressif aux amplitudes témoins. Notons que, dans ces con-ditions, une augmentation de l'intensité du premier choc ne fait qu'accen-tuer cet effondrement; l'étirement passif du muscle garde par contre la possibilité de créer un rebond important de l'activité dans la période IV du cycle.
Ces résultats peuvent d'ailleurs dépendre des sujets et du niveau de profondeur du sommeil. Certains sujets présentent ces modifications dès les premiers symptômes de somnolence. D'autres montrent, dans les premières phases du sommeil, un net accroissement de l'excitabilité de base et ne présentent ces manifestations qu'à des niveaux de profondeur beaucoup plus avancés.
Le fait que cet abaissement de l'excitabilité de base va de pair avec un relâche-ment de la musculature fusoriale (comme le montre l'affaissement des réponses mécaniquement provoquées) peut apporter un nouvel élément d'interprétation des données précédemment acquises: La persistance des effets d'étirement passif sur le rebond de la 4ème période du cycle paraît favoriser l'hypothèse d'une intervention des récepteurs tendineux dans sa genèse. Leur seuil d'excitabilité en effet, ne semble pas dépendre, au moins dans la même proportion que celles des récepteurs fusoriaux, d'une diminution de tonus des myofibrilles intrafusoriales (HUNT et KUFFLER 1952).
Commentaire final.
Il nous a paru légitime de faire appel aux données de la Neurophysio-logie animale pour interpréter un certain nombre de faits et émettre certaines hypothèses explicatives.
1° Les éléments de début de nos courbes d'excitabilité semblent trouver une explication satisfaisante dans les données classiques de l'expérimentation animale: réfractorité des fibres nerveuses à laquelle nos conditions de travail viennent ajouter celle des jonctions neuro-muscu-laires (période I); facilitation consécutive à des stimulations condition-nantes subliminaires (période II); subnormalité prolongée associée à la décharge des éléments actifs (période III); blocage dû aux décharges antidromiques dans les fibres motrices atteintes par la stimulation condi-tionnante. Ces effets dominent toutes les premières phases de l'explora-tion et viennent masquer les manifestations centrales précoces consécu-tives à la volée incidente. Nous retrouvons là les difficultés qui se trou-vent attachées à l'emploi de stimulations successives de nature homo-synaptique.

474 J. PAILLARD
2° Les effets directs de la volée incidente, en dehors de ses manifestations immédiates (Voir 1°), soulèvent par contre des problèmes beaucoup plus discutables. Ils ne sont d'ailleurs interprétables que dans la mesure où la présence d'une réponse réflexe ne vient pas y ajouter les complications dues aux afférences proprioceptives associées à la contraction musculaire correspondante.
Autant que le permet notre analyse (effets d'une stimulation con-ditionnante subliminaire pour la décharge réflexe) nous pensons pouvoir attribuer : aux messages des fibres du Groupe I, outre la facilitation rési-duelle et les effets précoces d'inhibition, dont on trouve les équivalents chez l'animal, un effet d'inhibition dont le maximum se situerait entre 60 et 100 ms et un effet de dépression plus prolongée situant son maximum entre 400 et 600 ms. Les fibres d'origine cutanée et celles du Groupe II (en supposant légitime d'invoquer la réponse des terminaisons secondaires du fuseau à la SM: HUNT 1954) seraient responsables d'une longue dépression se manifestant sur toute l'étendue du cycle. Ce fait semble confirmé par l'apparition d'un effet identique de dépression générale, qui se manifeste lorsqu'on accroît l'intensité de la SE; nous avons vu, par contre, que, pour des intensités encore plus fortes, on retrouve parfois un relèvement de toute la courbe — l'éventuelle participation des fibres du Groupe II à ces phénomènes reste hypothétique — Nous utilisons, en tout cas, des intensités de stimulation très largement inférieures au seuil de douleur et nous sommes d'accord avec SCHENCK (1951) et MAGLADERY et coll. (1951) pour écarter l'influence d'effets nociceptifs dans les phéno-mènes observés.
3° Les effets autogéniques de la contraction musculaire apparaissent jouer un grand rôle dans la détermination de la forme générale des courbes observées.
Avec MERTON (1951) nous pensons que l'étendue du myogramme peut, dans certaines conditions, déterminer pour une large part la longueur de la période de silence. La subnormalité consécutive à la décharge centrale doit, par contre, être rendue responsable de la non-réactivité initiale des éléments interrogés. Suivant les cas, l'un ou l'autre de ces facteurs peut devenir prépondérant (J. PAILLARD 1955). A l'encontre de MERTON (et de ceux qui, avant lui, ont adopté cette explication) nous ne croyons pas que le seul silence de la décharge fusoriale au cours de la contraction puisse suffire à rendre compte des faits observés. On pourrait déjà invoquer le fait que certaines données relatives aux effets de la manœuvre de JENDRASSIK (Sommer 1939), HOFFMANN F. A. 1951, PAILLARD. J. 1953) semblent contester l'existence d'une décharge per-manente des récepteurs fusoriaux dans le muscle au repos chez un grand nombre de sujets normaux.

Analyse du réflexe de HOFFMANN chez l'Homme. 475
Il est bien démontré, d'autre part, que la période de silence, mise en évidence dans l'électromyogramme du muscle mis sous tension volon-taire, coïncide avec la période III de nos courbes SE + SE et SM + SE, obtenues sur un muscle au repos. Or la longue période d'inexcitabilité des récepteurs fusoriaux, révélée par le tracé des courbes SE + SM et SM + SM, situerait la reprise d'activité de ces derniers largement au delà de la période de silence (en accord avec certains résultats de DENNY-BROWN 1928).
Ce dernier fait semble également exclure l'intervention de ce processus dans la genèse du rebond d'excitabilité qui marque la fin de la période de silence.
Nombreux sont les éléments de nos résultats (dans le triceps sural) qui semblent, par contre, plaider en faveur d'une intervention prépondé-rante des récepteurs tendineux de GOLGI dans ces manifestations (Effets des manœuvres passives qui, à volonté, permettent d'exalter ou d'atténuer les effets de l'inhibition autogénique — Influence du sommeil, qui s'accom-pagne d'une dépression de la tonicité de la musculature fusoriale). Les arguments invoques par MERTON (voir discussion de la communication de MERTON 1953): seuil mécanique trop élevé des récepteurs de GOLGI, inertie trop grande de leur « rate of change », semblent assez fragiles à la lumière des travaux de KUFFLER et HUNT (1950).
On peut, semble-t-il, rejeter l'éventualité d'une influence de l'exci-tation des terminaisons secondaires du fuseau dans ces phénomènes sur la base des évidences récemment acquises de leur relation avec le réflexe de flexion (HUNT 1953). La période de silence présente, en effet, les mêmes caractères dans un muscle extenseur et dans un muscle fléchisseur.
Nous sommes donc d'accord avec les résultats de l'école scandinave (GRANIT 1950, GRANIT et STRÖM 1951, HAGBARTH et NAESS 1950) pour accorder à l'inhibition autogénique un rôle possible dans la genèse de la période de silence au moins dans le triceps sural. Le rebond con-sécutif pourrait même être envisagé comme une manifestation post-inhibitrice centrale de la décharge des récepteurs de GOLGI.
En fait, de nombreux types de rebonds peuvent sans doute coexister. Toute suppression brusque d'une influence dépressive peut en effet s'accompagner d'une reprise d'activité supernormale, suivant un principe bien souvent vérifié — (CREED et coll. 1932).
d) Les systèmes dépresseurs de l'activité spinale semblent, de fait, jouer un rôle très important dans les manifestations tardives de nos cycles d'excitabilité. Nos conclusions sur l'intervention des fibres de bas seuils dans la genèse de ces phénomènes rejoignent les données de MAGLADERY et coll. (1952) acquises sur des cas pathologiques.
Les données de l'expérimentation animale, sur ce point, sont encore beaucoup trop fragmentaires pour fournir une explication satisfaisante

476 J. PAILLARD
des faits observés (voir discussion dans LANGUTH et coll. 1952, voir également discussion de la communication de JEFFERSON et SCHLAPP 1953, de LUSCHNAT 1954).
La Neurophysiologie a établi l'existence de nombreux circuits dans le système nerveux dont la signification fonctionnelle peut, aujourd'hui, se trouver utilement éclairée par un rapprochement avec les principes de fonctionnement des servomécanismes (RUCH 1951). Que l'on parle avec VON HOLTZ (1951) d'un principe de réafférence, ou avec les cybernéticiens d'un système de feedbacks négatifs, on se retrouve devant des problèmes de contrôle des commandes et de leur asservissement aux effets qu'elles réalisent — Le rôle des propriocepteurs, directement associés au fonction-nement des fibres musculaires contractiles, apparaît évidemment fonda-mental comme détecteurs de cette information périphérique susceptible de réagir, soit directement par les voies les plus rapides, soit secondaire-ment par des circuits plus complexes, sur le niveau d'excitabilité de la « voie finale commune » des commandes motrices. Sans doute les courbes d'excitabilité, du type de celles que nous obtenons, révèlent-elles les oscillations transitoires de ces systèmes d'autorégulation brusquement investis par une masse d'influx synchrones, que représente la volée sensorielle résultant du choc brutal évoquée par les types artificiels de stimulation employés. En ce sens la SM n'est guère moins « inadéquate » que la SE. L'une et l'autre constituent des modes d'action artificielle qui limitent grandement la portée des renseignements qu'ils sont sus-ceptibles de nous fournir (voir à ce sujet ALVORD et FUORTES, 1953).
e) Le problème de la fonction des fibres gamma dissimule encore bien des inconnues. Le fuseau neuro-musculaire réalise, comme MATTHEWS l'avait pressenti, le prototype de ce que la Cybernétique appelle aujourd'hui un détecteur d'écarts différentiel. Si son fonctionnement élémentaire a pu faire l'objet d'analyses précises (HUNT 1951, 1952), les modalités de son intégration dans l'ensemble des circuits de régulation sensitivo-motrice restent encore à préciser (HOFFMANN 1951, ROBERTS 1952, MERTON 1953, PAILLARD 1953).
L'une des données les plus curieuses de notre analyse concerne la remarquable différence d'aspect de nos courbes selon que SM ou SE constituent le test explorateur. Elle nous amène à envisager un accroisse-ment de la sensibilité des récepteurs périphériques venant compenser la dépression de l'excitabilité des motoneurones que nous révèle dans l'autre type de courbes, une exploration directe au moyen de la SE. Ce fait impliquerait un élargissement des fonctions régulatrices de ce circuit périphérique et son intégration à des systèmes plus complexes. La possi-bilité d'une action spécifique de certaines influences centrales hautes sur les motoneurones d'origine des fibres gamma ressort des travaux récents de GRANIT et KAADA (1952) et de ELDRED et coll. (1953).

Analyse du réflexe de HOFFMANN chez l'Homme. 477
f) Sans doute devrait-on également tenir compte de divers autres facteurs de complication possibles introduits par nos conditions de travail. Les effets secondaires d'une éventuelle participation des centres moteurs synergiques et antagonistes des éléments interrogés sont, par exemple, susceptibles d'influencer nos résultats. Les expériences de blocage de la conduction dans le tronc nerveux antagoniste (MAGLA-DERY et coll. 1951) ne semblent pas montrer de modification de l'allure des courbes.
Dans une analyse exhaustive, il faudrait tenir compte également des influences extra-synaptiques associées à l'emploi de la stimulation artifi-cielle. Elles constituent, en effet, autant d'artefacts possibles dont on a déjà souligné l'importance lors de l'utilisation de volées synchrones (LLOYD, 1944): interactions électriques entre fibres nerveuses adjacentes (KATZ et SCHMITT, 1950); effets stimulants de l'activité musculaire sur les terminaisons nerveuses (LLOYD, 1942); effets antidromiques sur les organes sensoriels périphériques (MATTHEWS 1933); décharges réflexes rétrogrades dans les fibres afférentes (TOENNIES 1938). Rien ne semble indiquer dans notre analyse que ces facteurs puissent intervenir de façon notable dans la genèse des phénomènes observés.
Résumé.
1° Une identification du réflexe de HOFFMANN (stimulation électrique du nerf poplité interne) avec le réflexe myotatique, obtenu par étirement mécanique du muscle (percussion du tendon d'Achille), apparaît légitime en expérimentation humaine. La démonstration de la stabilité des latences apporte un argument supplémentaire à l'hypothèse de la nature mono-synaptique de ces réponses.
2° Les conditions d'utilisation de la stimulation électrique du nerf comme test direct de l'excitabilité du centre moteur ont été définies. L'emploi de la stimulation mécanique doit tenir compte des fluctuations imputables aux actions qui s'exercent directement sur la sensibilité des récepteurs de tension intra-musculaire. La confrontation des résultats obtenus à partir de ces deux modes de stimulation des mêmes voies centrales peut constituer un test différentiel utilisable pour l'appréciation des influences spécifiques empruntant les voies efférentes du type gamma.
3° L'analyse des cycles d'excitabilité, au moyen de la technique du double choc, révèle l'existence de deux types de courbes suivant que l'on utilise la stimulation électrique ou la stimulation mécanique comme choc explorateur. Ces différences paraissent impliquer, d'une part une inexcitabilité des récepteurs fusoriaux se prolongeant largement au delà des limites de la classique période de silence, d'autre part un ajustement de la tension des myofibrilles fusoriales susceptibles de compenser les effets de dépression qui influencent tardivement les motoneurones.

478 J. PAILLARD
4° Les divers éléments de ces courbes ont été analysés dans leur rapport avec la composition de la volée afférente, avec le niveau d'excitabilité centrale de base, avec la forme et la durée de la déformation du muscle au cours de la contraction et avec l'état de tension mécanique initiale.
5° La confrontation avec les données actuellement acquises de là neurophysiologie animale n'autorise qu'une interprétation partielle des résultats obtenus. Les fibres afférentes les plus rapides du Groupe I, outre leur rôle essentiel dans le déclenchement de la réponse réflexe, semblent responsables d'une succession d'effets dépresseurs se manifestant aux intervalles allant respectivement des délais les plus courts jusqu'à 15 ms, de 70 à 100 ms, et de 400 à 600 ms. Les messages des fibres d'origine cutanée et des fibres du Groupe II ont sur l'excitabilité du centre moteur du triceps sural un effet d'inhibition pouvant se prolonger sur plusieurs secondes. Les effets autogéniques de la contraction musculaire du triceps sural et, en particulier, les influences inhibitrices de l'activité des récepteurs tendineux de GOLGI peuvent, au même titre que d'autres facteurs d'influences (Subnormalité postréactionnelle, suspension d'acti-vité fusoriale), être invoqués dans la genèse de la période de silence et du rebond qui lui fait suite et, dans certaines conditions expérimentales, devenir prépondérant.
Bibliographie .
— ALVORD, E. C, and M. F. G. FUORTES: J. of Physiol. 122, 302 (1953). — BENSON, A. J., and A. A. JEFFERSON: J. of Physiol. 118 (1952). — BERNHARD, C.G.: Acta physiol. scand. (Stockh.) 14, supp. 47 (1947). — BREMER, F., V. BONNET et J. MOLDAVER: Ard. int. Physiol. 52, 1 (1942). — BROCK, L. G., J. C. ECCLES and W. RALL: Proc. Roy. Soc. 138 B (1951). — BROOKS, C., McC., C. B. B. DOWNMAN and J. C. ECCLES: J. of Neurophysiol. 13,
157 (1950); 13, 9 (1950). — COOPER, S., D. E. DENNY-BROWN and C. S. SHERRINCTON: Proc. Roy. Soc. B
100, 448 (1926). — CREED, R. S., D. DENNY-BROWN, J. C. ECCLES, E. G. T. LIDDEL and C. S.
SHERRINGTON: Reflex activity of the spinal cord. Oxford. The Clarendon Press, 183 pp (1932).
— DELIUS, H., u. P. HOFFMANN: Z. Biol. 95, 229 (1934). — DENNY-BROWN, D. E.: Proc. Roy. Soc. B 103, 321 (1928); B 104, 252 (1929). — DODGE, R,: Z. allg. Physiol. (Jena) 12, 1 (1910). — DUBBERT, D. R., and O. H. SCHMITT: Rev. Sc. Instrum. 26, 145 (1949). — ECCLES, J. C., and J. J. PRITCHARD: J. of Physiol. 89, 43 P (1937). — ECCLES, J. C.: J. of Neurophysiol. 9, 87 (1946). — ECCLES, J. C., P. FATT, S. LANDGREN and G. J. WINSBURY: J. of Physiol.
125, 590 (1954). — ENGELHORN, R., u. F. A. HOFFMANN: Dtsch. Z. Nervenheilk. 171, 161 (1953). — ELDRED, E., R. GRANIT and P. A. MERTON: J. of Physiol. 122, 498 (1953). — ERLANGER, J., and E. A. BLAIR: Amer. J. Physiol. 121, 431 (1938). — FULTON, J. F.: Muscular contraction and the reflex control of movement. 644 pp.
Baltimore: Waverly press 1926. — GASSER, H. S., and E. ERLANGER: Amer. J. Physiol. 62, 496 (1922). — GASSER, H. S., and H. T. GRAHAM: Amer. J. Physiol. 103, 303 (1933). — GASSER, H. S.: J. of Neurophysiol. 2, 361 (1939) (période silence). — GRANIT, R., and V. SUURSOET: Nature (Lond.) 164, 270 (1949). — GRANIT, R.: J. of Neurophysiol. 13, 351 (1950). — GRANIT, R., and B. R. KAADA: Acta physiol. scand. (Stockh.) 27, 130
(1952). — GRANIT, R., and G. STRÖM: J. of Neurophysiol. 14, 113 (1951).

Analyse du réflexe de HOFFMANN chez l'Homme. 479
— HAGBARTH, K. E., and K. NAESS: Acta physiol. scand (Stockh.) 21, 41 (1950). — HOFF, H. E., E. C. HOFF, P. C. BUCY and J. PI-SUNDER: Amer. J. Physiol. 109,
123 (1934). — HOFFMANN, F. A.: Dtsch. Z. Nervenheilk. 167, 310 (1952). — HOFFMANN, P.: Untersuchung über die Eigenreflexe (Sehnenreflexe) menschlicher
Muskeln. 106 pp. Berlin: Springer 1922. — Z. Biol. 81, 37 (1924). — HOFFMANN, P., u. J. KELLER: Z. Biol. 87, 327 (1928). — HOFFMANN, P.: Erg. Physiol. 36, 15 (1934). — Dtsch. Z. Nervenheilk. 166, 60
(1951). — Schweiz. med. Wschr. 82, 57 (1952a). — Experientia (Basel) 8, 371 (1952b). — Nervenarzt 24, 19 (1953).
— HUGHES, J., and H. S. GASSER: Amer. J. Physiol. 108, 295 (1934). — HUNT, C. C., and S. W. KUFFLER: J. of Physiol. 113, 298 (1951). — HUNT, C. C.: J. of Physiol. 117, 359 (1952). — Abstr. XIX. Int. Physiol. Congr.
p. 485 (1953). — J. Gen. Physiol. 38, 117 (1954). — JOB, C.: Pflügers Arch. 256, 391 (1953). — JEFFERSON, A. A., and W. SCHLAPP: p. 99 in The spinal cord. Cyba svmpos.
London: Churchill 300 pp. (1953). — KATZ, B., and O. H. SCHMITT: J. of Physiol. 97, 471 (1940). — KUFFLER, S. W., and C. C. HUNT: Proc. Ass. Res. Nerv. Ment. Dis. 30, 24
(1950). — KUGELBERG, E.: Acta physiol. scand (Stockh.) 8, supp. 24 (1944). — LANGUTH, H. W., R. D. TEASDALL and J. W. MAGLADERY: Bull. Johns Hopkins
Hosp. 91, 257 (1952). — LAPORTE, Y., and D. P. C. LLOYD: Amer. J. Physiol. 169, 609 (1952). — LEKSELL, L.: Acta physiol. scand (Stockh.) 10, supp. 31 (1945). — LIDDELL, E. G. T., and C. S. SHERRINGTON: Proc. Roy. Soc. 96 B, 212 (1924). — LLOYD, D. P. C.: J. of Neurophysiol. 5, 153 (1942); 6, 213 (1943a); 6, 316
(1943b). — Physiologie. Rev. 24, 1 (1944). — J. of Neurophysiol. 9, 421 (1946). — Proc. Ass. Res. Nerv. Ment. Dis. 30, 48 (1950).
— LLOYD, D. P. C., and A. K. MCINTYRE: J. of Neurophysiol. 13, 39 (1950). — LUSCHNAT, K. D.: Plügers Arch. 258, 431 (1954). — MAGLADERY, J. W., and D. B. MCDOUGAL Ir.: Bull. Johns Hopkins Hosp. 86,
265 (1950). — MAGLADERY, J. W., W. E. PORTER, A. M. PARK and R. D. TEASDALL: Bull.
Johns Hopkins Hosp. 88, 499 (1051a). — MAGLADERY, J. W., R, D. TEASDALL, A. M. PARK and W. E. PORTER: Bull. Johns
Hopkins Hosp. 88, 520 (1951b). — MAGLADERY, J. W., R, D. TEASDALL, A. M. PARK and H. W. LANGUTH: Bull.
Johns Hopkins Hosp. 91, 219 (1952). — MAGOUN, H. W., and R. RHINES: J. of Neurophysiol. 9, 165 (1946). — MATTHEWS, B. M. C.: J. of Neurophysiol. 78, 1 (1933). — MERTON, P. A.: J. of Physiol. 114, 183 (1951). — p. 247 in The Spinal cord - Ciba
sympos. London: Churchill 300 pp. (1953). — NOGUÈS, P., et J. PAILLARD: C. r. Soc. Biol. (Paris) 147, 583 (1953). — PAILLARD, J., et M. TURNER: Electroencephalogr. Clin. Neurophysiol. 5, 467
(1953). — PAILLARD, J.: J. de Physiol. 45, 206 (1953a) — C. r. Acad. Sci (Paris) 236, 1505
(1953b); 236, 1701 (1953c). — J. de Physiol.46, 484 (1954); (1955) sous presse. — RALL, W.: J. de Physiol. 125 n° 1, 30 P (1954). — RENSHAW, B.: J. of Neurophysiol. 3, 373 (1940). — ROBERTS, T. D. M.: J. of Physiol. 118, 8 P (1952). — RUCH, T. C: p. 154 in Stewens. SS. Handbook of experimental Psychol. London:
Chapman et Hall, 1436pp. (1951). — SCHOEN, H.: Pflügers Arch. 253, 285 (1951). — SCHENK, E.: Pflügers Arch. 253, 286 (1951). — SKOGLUND, C. R.: Acta physiol. scand (Stockh.) 4, supp. 12 (1942). — SOMMER, J.: Z. Biol. 99, 514 (1939). — TEASDALL, R. D., H. W. LANGUTH and J. W. MAGLADERY: Bull. Johns
Hopkins Hosp. 91, 267 (1952). — TÖNNIES, J. F., u. R, JUNG: Pflügers Arch. 250, 667 (1948). — V. HOLST, E., u. H. MITTELSTEDT: Nturwissenschaften 20, 464 (1950). — WESSLAU, W.: Pflügers Arch. 255, 273 (1952).
Dr. J. PAILLARD, Laboratoire de Neurophysiologie Générale 4, Avenue Gordon Bennett Paris (16ème)



![SYSTÈME NERVEUX CENTRAL, PÉRIPHÉRIQUE ET … · électrode de surface, ... [O, 9] Exploration électrophysiologique d'une polyneuropathie Formation : spécifique à cet acte en](https://img.pdfslide.fr/doc/110x75/5b96144509d3f2c2678d0ecd/systeme-nerveux-central-peripherique-et-electrode-de-surface-o-9.jpg)

























