Embed Size (px)
Citation preview

390 Biochimicu et Biophysics Acre 835 (1985) 390-401
Elsevier
BBA 51942
Apolipoprotein A-I from normal human plasma: definition of three distinct
antigenic determinants
Philip K. Weech, Ross W. Milne, Peter Milthorp and Yves L. Marcel
Laboratoire du Metabolisme des Lipoproteines, Institut de Recherches Cliniques de Montreal, I IO Avenue des Pins, ouest.
Montreal, Quebec (Canada)
(Received December 27th, 1984)
Key words: Apolipoprotein A-I; HDL; Monoclonal antibody; CNBr fingerprint; (Human plasma)
We have prepared, selected and cloned four mouse hybridomas that secreted monoclonal antibodies against human plasma apolipoprotein A-I. These antibodies are all of the IgGI subclass, and were named anti-A-I 6B8, 5G6, 3D4 and 5A6. We characterized the specificity of the antibodies, finding that all four of them reacted similarly, and with only the major proteins having the molecular weight and isoelectric focusing characteristics of apolipoprotein A-I. The antibodies reacted with all known charge-polymorphs of apolipoprotein A-I and pro apolipoprotein A-I. Thus, the polymorphs of apolipoprotein A-I are alike in that they all contain the antigenic sites of these four antibodies. In a solid-phase, antibody competition radioimmunoassay we found inhibition or enhancement of antibody binding to apolipoprotein A-I, according to the pair of antibodies tested. Antibodies 6B8, XX and 3D4 were different from one another and reacted with different antigenic determinants, but 5A6 was similar to 3D4 and reacted at the same site. We compared the reactions of the four antibodies with CNBr-cleaved fragments of apolipoprotein A-I separated by polyacrylamide gel electrophoresis. We found three different patterns of reaction with the apolipoprotein A-I fragments; 6B8,5G6 and 3D4 were different, but 5A6 resembled 3D4. Thus, the four antibodies reacted with at least three different antigenic sites in apolipoprotein A-I, which were present in different CNBr fragments of apolipoprotein A-I, but not on fragment 4 which forms the carboxy-terminal segment.
Introduction
Apolipoprotein A-I is the major apolipoprotein of normal human plasma (about 140 mg/dl) and
is found in the high-density lipoproteins (HDL),
accounting for at least 55% of the HDL protein. Apolipoprotein A-I structure and its function as a
cofactor in the reaction of lecithin : cholesterol acyltransferase are well established (for reviews see Refs. 1, 2), however its exact role in cholesterol transport remains in part speculative [l]. Serum
Abbreviations: HDL, high-density lipoproteins of density
1.063-1.21 g/ml; VLDL, very-low-density lipoproteins of den-
sity < 1.006 g/ml.
concentrations of apolipoprotein A-I and HDL cholesterol are both negative predictors for cardiovascular disease but apolipoprotein A-I ap-
pears to add to the predictive value of HDL cholesterol [3-51, a finding which has further increased the interest shown for this protein. Vari- ous immunoassays for apolipoprotein A-I have been reported which shared, to different degrees, the difficulties inherent in the determinations of serum apolipoproteins [6,7]. Self-association of apolipoprotein A-I [8,9] has made difficult the development of appropriate standards, while apolipoprotein A-I in lipoproteins does not always express all its potential antigenic sites (lO,ll], im- plying that some sites are masked by lipids in
0005-2760/85/$03.30 0 1985 Elsevier Science Publishers B.V. (Biomedical Division)

391
intact lipoproteins or that their ability to bind an antibody depends on the conformation of A-I as affected by lipids or other proteins.
A valid approach to the problems of standardiz- ing apolipoprotein A-I immunoassays is to use well-characterized monoclonal antibodies to study the expression of antigenic sites in A-I, without the confounding effects of uncharacterized antibodies from polyclonal antisera. This will enable us to develop assays that measure apolipoprotein A-I antigenic sites that are always expressed in all isomorphs of A-I [12-171, whether lipids are pre- sent or absent. Furthermore, monoclonal antibod- ies will provide precise markers to study the struc- ture-function relationship in apolipoprotein A-I. We report here the production and characteriza- tion of four monoclonal antibodies to apolipopro- tein A-I which recognize at least three distinct antigenic determinants on apolipoprotein A-I, each of which reacts with all the known polymorphic forms of apolipoprotein A-I and pro apolipopro- tein A-I.
Materials and Methods
Chemicals for electrophoresis were obtained from Bio-Rad (~chmond, CA). ~pholytes and Ultrodex for isoelectric focusing were from LKB (Bromma, Sweden), 0.45 pm nitrocellulose sheet from Millipore (Bedford, MA), Immulon II Re- movawell Strips (Dynatech Laboratories, Inc., Alexandria, VA) and all other chemicals from Fisher, Ltd (Montreal, Canada). Affinity-pu~fied rabbit anti-mouse IgG was purchased from Kirk- egaard and Perry Laboratories, Inc. (Gaithersburg, MD).
Production of monoclo~a~ antibodies Female, &week-old Balb/c mice (Jackson
Laboratories, Bar Harbor, ME) were injected in- traperitoneally with 50 pg of apolipoprotein A-I emulsified in complete Freund’s Adjuvant. Identi- cal injections were given three weeks later. After a further three weeks the mice received an in- travenous injection of 10 Erg apolipoprotein A-I, and four days later their spleens were excised for the cell fusion. Hybridomas were produced using a mixture of 30 * lo6 SP2-0 cells and 120 - lo6 cells from two spleens [ 181. The cells were distributed in
1000 microculture wells (Costar, Cambridge, MA) and growth was later seen in 300 wells. Ten wells were positive for the presence of antibodies specific for apolipoprotein A-I on screening by radioim- munoassay using a solid-phase technique with Re- movawells coated with A-I (10 pg/ml in 5 mM glycine, pH 9.2) overnight at room temperature. The cells from the positive wells were cloned by limiting dilutions in 96 well microculture plates, yielding four stable clones that were recloned three times by limiting dilutions. To identify the im- munoglobulin class and subclass, a sample of hy- bridoma supematant was concentrated by precipi- tation in 50% saturated ammonium sulphate and tested by double diffusion against antisera specific for mouse immunoglobulin (Ig) classes and sub- classes [19]. Selected subclones were injected in- traperitoneally into Balb/c mice and their ascitic fluid was used directly as the source of antibodies for most experiments.
~solafjo~ of lipoproteins and apo~~pop~otei~ A-l Either fresh, normal human serum, following an
overnight fast, or fresh plasma from the Red Cross was used to isolate HDL, by sequential ultra- centrifugation between the densities 1.125 and 1.210 g/ml for 40 h at 12°C 45 000 rpm in a 50.2 rotor and L2-65 ultracentrifuge (Beckman Instru- ments Inc., Palo Alto, CA). After dialysis against 10 mM NH,HCO,/l mM EDTA/O.OZS NaN,, the HDL, were lyophilized, then delipidated using chloroform/methanol [20]. Apolipoprotein A-I was isolated from HDL, by preparative isoelectric focusing [12] for the immunization of mice, screen- ing of hybridomas by radioimmunoassay for specific antibody secretion, and studies of the frag- ments of A-I cleaved by CNBr.
The technique described by Brewer et al. [21], with a 500-fold excess of CNBr, was followed to prepare fragments of apolipoprotein A-I, typically using 1.65 mg lyophilized A-I redissolved in 165 pl formic acid 70% containing 9.38 mg CNBr. This was allowed to react under N,, in the dark, at about 22’C for either 2 or 24 h. In some experi- ments, the apolipoprotein A-I was treated with 5% 2-mercaptoethanol before lyophilization and CNBr treatment, and a &fold molar excess of

392
tryptophan (over apolipoprotein A-I) was added
to the CNBr solution. At the end of the CNBr treatment, the samples were diluted with water
and lyophilized.
Po!vacrylumide gel electrophoresis, isoelectric focus-
ing and transfer to nitrocellulose sheets
Polyacrylamide gels were cast as vertical slabs
(14 X 18 X 0.15 cm) for electrophoresis in the Pro- tean cell (Bio-Rad, CA). The gels were made to the recipes of (i) Kane [22] for electrophoresis in al-
kaline, urea-containing buffer, (ii) Neville [23]
using 15% acrylamide and sodium dodecyl sulphate
(SDS), or using 20% acrylamide and substituting 18% glycerol for water (v/v) in the recipe, to
improve resolution of low molecular weight poly-
peptides as recommended by Connelly and Kuksis
[24], (iii) Warnick et al. [25] for isoelectric focusing
using Ampholytes pH 4-6 or pH 3.5510. Urea-
containing gels were stained with Coomassie blue
G250 0.04% in 3.5% perchloric acid [26] and in-
tensified in 5% acetic acid [27]; SDS-containing gels were stained in Coomassie blue R250 0.25%
dissolved in 45% methanol/9% acetic acid, and destained in the same solvent; replicas of electro- phoresis on nitrocellulose sheet were stained with
Amido black 0.1% dissolved in 45% methanol/ 10% acetic acid and destained in 90% methanol/2%
acetic acid [28].
Immediately after electrophoresis. a replica of
the separated proteins was transferred onto a
nitrocellulose sheet by the method of Towbin et al. [28] using the methanol/Tris/glycine buffer for
SDS-containing gels and 0.7% acetic acid as the
electrolyte for urea-containing gels. A constant
current of 100 mA, 50-20 V, for about 16 h was
used in the Transblot cell (Bio-Rad, Richmond,
CA) to effect the transfer, at about 22’C. To detect reactions of the monoclonal antibodies with
the protein replicas, the nitrocellulose sheets were then treated as described by Theolis et al. [29].
lodinution of monoclonul antibodies Monoclonal antibodies (IgG-I) were purified by
adsorption to Protein A-Sepharose (Pharmacia, Uppsala, Sweden) at pH 8.0, and elution with 0.1 M citric acid (pH 6.0). Iodination of these affinity purified, mouse, anti-A-I monoclonal antibodies was carried out by the method of Mellman and
Unkeless [30] as described by Milne, et al. [31].
Briefly, 24 pg of antibody were labelled with 0.5 mCi of “‘1 (Amersham). Free “‘1 was removed on
an AGIX8 resin (Bio-Rad) and the labelled anti-
body (eluant) stored in phosphate-buffered saline
(0.154 M NaCl/lO mM Na2HP0,. 7H,0/5 mM NaH2P0,. H,O (pH 7.2)) containing 0.1% bovine serum albumin and 0.02% sodium azide.
Competitive antibody binding USSUJS Solubilized apolipoprotein A-I was diluted in
coating buffer (15 mM NazCO,/ 35 mM
NaHCOJ3 mM NaN, (pH 9.6)) to a final con- centration of 2 pg/ml. Aliquots (50 ~1) were distributed in Immulon 2 Removawell strips and
left refrigerated overnight to allow the binding of
the antigen to the wells. The wells were emptied.
washed one time with phosphate-buffered saline, and excess liquid was removed by inversion of the
wells on paper towelling. To saturate non-specific
binding sites, the wells were incubated with 250 ~1
of 10%’ bovine serum albumin in phosphate-
buffered saline for 1 h at room temperature, then emptied, washed with phosphate-buffered saline as described above, and drained. Serial two-fold dilu- tions of the ascites fluids were prepared in phos- phate-buffered saline containing 0.05% Tween-20,
1% bovine serum albumin, 1.5 mM MgCl Z and 2.5 mM mercaptoethanol using round-bottomed, mul-
tiwell, microtitre plates (Linbro) and a multichan- nel pipettor (Titertek). These dilutions were im-
mediately transfered with the multichannel pipet-
tor to Removawells containing apolipoprotein A-I, and incubated for 2 h at room temperature. The
wells were then emptied and washed three times
with washing buffer (phosphate-buffered saline
containing 0.05% Tween-20, 1.5 mM MgCl, and 2.5 mM mercaptoethanol). Excess liquid was re-
moved as described above. At this point we con-
tinued the experiment in one of two ways. (i) Each well was incubated with the same
amount of ‘251-labelled, goat anti-mouse im- munoglobulin, washed again, and finally the ra- dioactivity was measured. The amount of labelled. anti-immunoglobulin bound to the wells increased, with increasing monoclonal antibody concentra- tion, up to a maximum. The lowest concentration of monoclonal antibody necessary to achieve this maximum binding was the saturating concentra-

393
tion for the conditions of that experiment. In subsequent antibody competition experiments, serial, two-fold dilutions of monoclonal antibodies were made from a starting concentration that was
4-times more concentrated than this saturating
concentration. (ii) In antibody competition experiments a 50
~1 aliquot of ‘251-labelled, affinity-purified, mono-
clonal anti-A-I was added to each well. This aliquot
contained lo-20 ng of protein and from 5 . lo4 to 1 . lo5 cpm, depending on the antibody used. The
wells were then incubated for 2 h at room temper-
ature. In some experiments using combinations of the monoclonal antibodies 6B8 and 5G6, the ascites
and radiolabelled affinity-purified antibody were added to the removawells together, eliminating the washing step between the addition of the ascites
and the radiolabelled antibody. After incubation with the radiolabelled IgG, the wells were emptied,
rinsed with washing buffer three times and excess
liquid was removed as described above. The wells
were then counted in an LKB 1271 Riagamma gamma counter. The results were expressed as (B/Be) x 100, where B = 1251 counts per minute
(cpm) bound mins non-specific binding, and Bo = 125I cpm bound in the absence of competing
antibody minus non-specific binding.
Results and Discussion
From the cell fusion and screening by radioim-
munoassay we selected and cloned four stable hybridomas, which were injected intraperitoneally
into mice and allowed to grow as ascites tumours. The intraperitoneal fluid from these mice served as
the source of antibodies for subsequent experi-
ments. The antibodies produced by the four clones
of cells were all of the IgG-I subclass and were
named 3D4,5A6,6B8 and 5G6. We made two kinds of experiments: (i) to iden-
tify whether the antibodies reacted with an antigen
having the known properties of apolipoprotein A-I upon polyacrylamide gel electrophoresis and iso- electric focusing, and (ii) to determine if the indi-
vidual antigenic sites, recognized by each anti- body, were close together, far apart or at least on the same molecule, based on immunoreactivities of CNBr-cleaved fragments of apolipoprotein A-I, and also on competitive cotitrations of apolipo- protein A-I with each possible pair of antibodies.
SDS-polyacrylamide gel electrophoresis After SDS-polyacrylamide gel electrophoresis
each of the antibodies reacted with the major protein band in HDL and purified apolipoprotein
A-I, and with only that band. Fig. 1 shows the
stained strips of polyacrylamide gel cut from slab
gels containing purified apolipoprotein A-I (strip
1) or HDL, (strip lo), with the autoradiographs alongside of strips cut from the nitrocellulose rep-
licas of the same gels reacted with 3D4, 5A6, 6B8 and 5G6. Even though the gels were overloaded
with both purified apolipoprotein A-I (40 pg per cm width of gel) and HDL, (40 pg per cm width
of gel), none of the minor bands seen on the stained gels were detected on the autoradiographs.
Apolipoprotein A-I is the major protein found in
HDL, and the antibody reactions and stained bands corresponded closely in electrophoretic mo-
bility with the molecular weight of apolipoprotein
A-I [21].
Polymorphism of apolipoprotein A-I by isoelectric focusing
Apolipoprotein A-I was first seen to be poly-
Fig. 1. Autoradiography of monoclonal antibody reaction with replicas of SDS-polyacrylamide gel electrophoresis. A sample of apolipoprotein A-I (40 gg/cm width of gel) was applied to one slab gel (strips 1-5) and HDL, (40 pg/cm) was applied to a second gel (strips 6-10). After electrophoresis ([23] 15% acrylamide) a slice of each gel was stained with Coomassie blue R250: apolipoprotein A-I shown in strip 1, HDL, strip 10. Replicas of the electrophoreses of the remaining gels were transferred to nitrocellulose [28] and reacted with monoclonal antibodies 5G6 (strips 2 and 6). 3D4 (strips 3, 7), 6B8 (strips 4, 8) and 5A6 (strips 5, 9). Migration was toward the anode at the bottom of the figure.

394
morphic using chromatography on DEAE-cel-
lulose [13], and since then the intraindividual het-
erogeneity has been confirmed in many laborato- ries using isoelectric focusing [12,14-171. With
ampholytes of pH 4-6, at least four forms of
apolipoprotein A-I and two forms of pro apoli-
poprotein A-I have been identified in normal
plasma and lymph [16,32]. A minor basic form of
apolipoprotein A-I was described in patients with
cerebrotendinous xanthomatosis [33] and a claim
was made, but not substantiated, for a relatively
basic form (pZ 6.5) in normal plasma HDL [14].
Fig. 2 shows two experiments to determine the specificity of our antibodies for the polymorphs of
apolipoprotein A-I in plasma HDL. A sample of delipidated HDL was applied to one isoelectric
focusing gel (36 pg protein per cm width of gel,
ampholytes pH 4-6) and a mixture of equal amounts of delipidated VLDL and HDL (18 pg
HDL protein per cm gel) was applied to a second gel. Fig. 2, strips 1 to 7, shows the replica of the
VLDL plus HDL gel; strips 8 to 10 show part of
Fig. 2. Autoradiography of monoclonal antibody reaction with
replicas of isoelectric focusing. A sample of equal amounts of
delipidated HDL (18 /.~g protein/cm width of gel) and VLDL
were applied to one slab gel (strips l-7), and twice the quantity of HDL (36 pg/cm) was applied to a second gel (strips 8-10).
Following isoelectric focusing [25] with ampholytes pH 4-6.
replicas of the two gels were transferred [28] to nitrocellulose.
Strips 1 and 10 show the nitrocellulose replicas stained with
Amido black. Strips 2-9 show autoradiography of the replicas
reacted with monoclonal anti-E 6C5 (strip 2), monoclonal
anti-E 3B7 (strip 3), monoclonal anti-A-l 5G6 (strips 4, 8). 5A6
(strips 5, 9), 6B8 (strip 6) and 3D4 (strip 7). The reactions of
the anti-A-1 are indicated by arrows, using the nomenclature of
Ghiselli et al. [16].
the replica of the HDL gel. Fig. 2, strips 1 and 10.
show the edges of the nitrocellulose replica stained with Amido black immediately after transfer. The
three major and three minor stained bands in the
top half of the replicas (Fig. 2, strip 10) are the pro
A-I and A-I isomorphs as identified by compari- son with standards and according to pl values.
The additional stained bands in the lower half of
Fig. 2, strip 10, were probably apolipoproteins A-II, C-II and C-III, visible at the higher HDL
protein concentration applied to that gel. Fig. 2 strips 4 to 7, show the autoradiograph of
the replica reacted with antibodies 5G6, 5A6, 6B8 and 3D4, respectively. Using the numbering de-
scribed by Ghiselli et al. [16] we have indicated on Fig. 2 the positions of seven, resolved, radio-
graphic bands that showed the reactions of the antibodies. In this, and in a replicate experiment,
all four antibodies reacted with all seven bands.
Bands 1 and 2 correspond in pl to pro A-I, bands
3 and 4 as the major isomorphs of apolipoprotein
A-I in plasma, and bands 4 to 7 as minor iso-
morphs of A-I. Band 7 was detected im-
munochemically but could not be seen on the stained replicas (Fig. 2, strips 1 and 10). Although
Fig. 2, strips 4 and 5 showed less intense bands than strips 6 and 7. the antibodies 5A6 did react with bands identical to those positive for 6B8 and 3D4, as seen when the film was overexposed and
also on the autoradiograph of the gel containing only HDL, at a higher protein concentration (Fig. 2. strips 8 and 9: antibodies 5A6 and 5G6, respec-
tively, on the HDL isoelectric focusing replica).
The autoradiograph of 6B8 and 3D4 reactions on the replica of this second gel (HDL only, 36 ,rtg
protein per cm) were identical to those shown in Fig. 2, strips 6 and 7. and so are not shown.
Although there appeared to be differences in
the sensitivity with which each antibody reaction could be detected on the replica’s autoradiograph we conclude that antibodies 5G6, 5A6, 6B8 and 3D4 each reacted with all known charge poly- morphs of pro A-I and apolipoprotein A-I, and in addition with a minor form of lower pl (A-I, in Fig. 2). Further, heterogeneity was absent from the autoradiographed replica of SDS-polyacrylamide gel electrophoresis (Fig. l), which showed that the antigen had the molecular weight properties of apolipoprotein A-I. With the similarity between
the molecular weights of apolipoprotein A-I and pro A-I [32] we did not expect to separate them
easily by SDS-polyacrylamide gel electrophoresis, and show in Fig. 1 that the antibodies did not
detect either grossly lower or higher molecular
weight proteins than normal apolipoprotein A-I. In contrast to the reactions of the four anti-A-I
monoclonal antibodies (Fig. 2, strips 4 to 9) two other monoclonal antibodies, anti-apolipoprotein-
E (anti-E) 3B7 and KS previously described [18] did not react with the seven polymorphs of apoli- poprotein A-I on the replicas. Instead, both 3B7 and 6C5 (Fig. 2, strips 2 and 3) reacted similarly
with four charge isoforms of apolipoprotein E, three of which were intercalated with the iso- morphs pro apolipoprotein A-I, and pro A-I,, and
apolipoproteins A-I, and A-I,. The major band
on the autoradiograph of these anti-E, however,
was of pZ immediately higher than that of pro A-I, (Fig. 2, strips 2, 3) and could be identified
with a faintly stained band on the replica of
VLDL plus HDL isoelectric focusing (Fig. 2, strip
1). These differences between the autoradiographs of anti-A-I and anti-E reaction on isoelectric
focusing replicas indicated not only that these two sets of antibodies were each specific for their own antigen, but also the great danger that is inherent in identifying stained bands on isoelectric focusing of apolipoproteins, unless immunochemical data is simultaneously available.
Competition between antibodies for binding to
apolipoprotein A-Z
The technique for the competitive radioim-
munoassays was described under Methods. Each of the unlabelled antibodies was capable of
specifically blocking its ‘251-labelled homologue,
indicating the specificity of the binding (Figs. 3 and 4). Antibody lD7, specific for apolipoprotein
E [19], did not inhibit the binding of any of the ‘251-labelled anti-A-I antibodies (data not shown).
Antibodies 3D4 and 5A6 showed the same con-
centration-dependent inhibition as each other that was characteristic of both antibodies being the same, whether binding of labelled 3D4 (Fig. 3b) or labelled 5A6 (Fig. 3c) was tested. Thus, these two antibodies seemed to sterically hinder the binding of one another, no matter which of them bound first to apolipoprotein A-I, possibly because their
120- A
1 I I I I
256 64 16 4 I 256 64 16 4 I
REClPROCAl OF FIRST ANTIEODY DILUTION
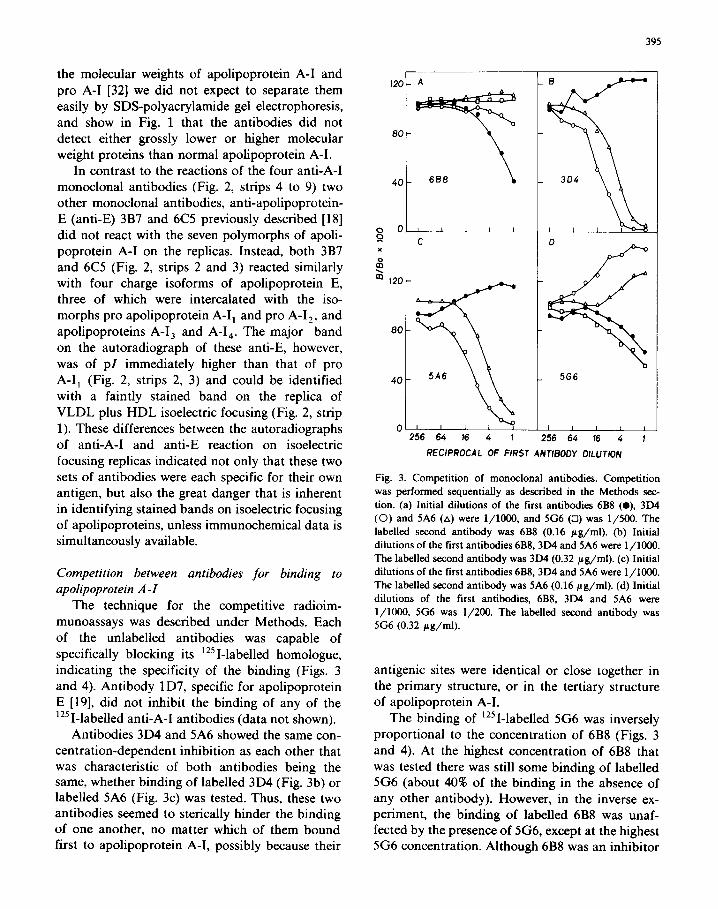
Fig. 3. Competition of monoclonal antibodies. Competition
was performed sequentially as described in the Methods sec-
tion. (a) Initial dilutions of the first antibodies 6B8 (0). 3D4
(0) and 5A6 (A) were l/1000, and 5G6 (0) was l/500. The labelled second antibody was 6B8 (0.16 ag/ml). (b) Initial
dilutions of the first antibodies 6B8,3D4 and 5A6 were l/1000.
The labelled second antibody was 3D4 (0.32 ag/ml). (c) Initial
dilutions of the first antibodies 6B8,3D4 and 5A6 were l/1000.
The labelled second antibody was 5A6 (0.16 ug/ml). (d) Initial
dilutions of the first antibodies, 6B8, 3D4 and 5A6 were
l/1000, 5G6 was l/200. The labelled second antibody was
5G6 (0.32 ug/ml).
antigenic sites were identical or close together in the primary structure, or in the tertiary structure of apolipoprotein A-I.
The binding of ‘251-labelled 5G6 was inversely proportional to the concentration of 6B8 (Figs. 3
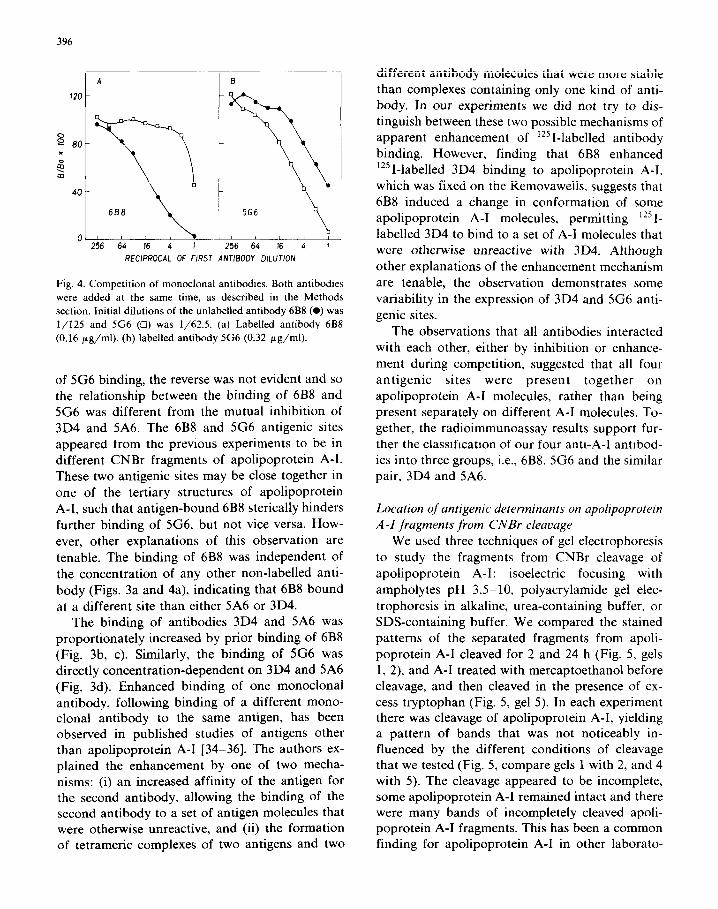
and 4). At the highest concentration of 6B8 that was tested there was still some binding of labelled 5G6 (about 40% of the binding in the absence of any other antibody). However, in the inverse ex- periment, the binding of labelled 6B8 was unaf- fected by the presence of 5G6, except at the highest 5G6 concentration. Although 6B8 was an inhibitor
396
256 64 16 4 i
RECIPROCAL OF FIRST ANTIBODY DILUTION
5G6 a
Fig. 4. Competition of monoclonal antibodies. Both antibodies
were added at the same time, as described in the Methods
section. Initial dilutions of the unlabelled antibody 6B8 (0) was
l/125 and 5G6 (0) was l/62.5. (a) Labelled antibody 6B8
(0.16 pg/ml). (b) labelled antibody 5G6 (0.32 pg/ml).
of 5G6 binding, the reverse was not evident and so the relationship between the binding of 6B8 and 5G6 was different from the mutual inhibition of 3D4 and 5A6. The 6B8 and 5G6 antigenic sites
appeared from the previous experiments to be in different CNBr fragments of apolipoprotein A-l. These two antigenic sites may be close together in
one of the tertiary structures of apolipoprotein A-l, such that antigen-bound 6B8 sterically hinders
further binding of 5G6, but not vice versa. How-
ever, other explanations of this observation are
tenable. The binding of 6B8 was independent of
the concentration of any other non-labelled anti-
body (Figs. 3a and 4a), indicating that 6B8 bound
at a different site than either 5A6 or 3D4. The binding of antibodies 3D4 and 5A6 was
proportionately increased by prior binding of 6B8 (Fig. 3b, c). Similarly, the binding of 5G6 was
directly concentration-dependent on 3D4 and 5A6 (Fig. 3d). Enhanced binding of one monoclonal antibody, following binding of a different mono- clonal antibody to the same antigen, has been observed in published studies of antigens other than apolipoprotein A-l [34-361. The authors ex-
plained the enhancement by one of two mecha- nisms: (i) an increased affinity of the antigen for the second antibody, allowing the binding of the second antibody to a set of antigen molecules that were otherwise unreactive, and (ii) the formation of tetrameric complexes of two antigens and two
different antibody molecules that were more stable
than complexes containing only one kind of anti- body. In our experiments we did not try to dis- tinguish between these two possible mechanisms of apparent enhancement of ‘2sl-labelled antibody binding. However, finding that 6B8 enhanced
‘25 I-labelled 3D4 binding to apolipoprotein A-l. which was fixed on the Removawells, suggests that
6B8 induced a change in conformation of some apolipoprotein A-l molecules, permitting 1251-
labelled 3D4 to bind to a set of A-l molecules that were otherwise unreactive with 3D4. Although
other explanations of the enhancement mechanism
are tenable, the observation demonstrates some
variability in the expression of 3D4 and 5G6 anti- genie sites.
The observations that all antibodies interacted
with each other, either by inhibition or enhance-
ment during competition, suggested that all four antigenic sites were present together on
apolipoprotein A-l molecules, rather than being present separately on different A-l molecules. To- gether, the radioimmunoassay results support fur- ther the classification of our four anti-A-l antibod- ies into three groups, i.e., 6B8, 5G6 and the similar
pair, 3D4 and 5A6.
Location of antigenic determinants on apolipoprotein
A-I fragments from CNBr cleavage
We used three techniques of gel electrophoresis
to study the fragments from CNBr cleavage of
apolipoprotein A-l: isoelectric focusing with
ampholytes pH 3.5510, polyacrylamide gel elec-
trophoresis in alkaline, urea-containing buffer, or SDS-containing buffer. We compared the stained
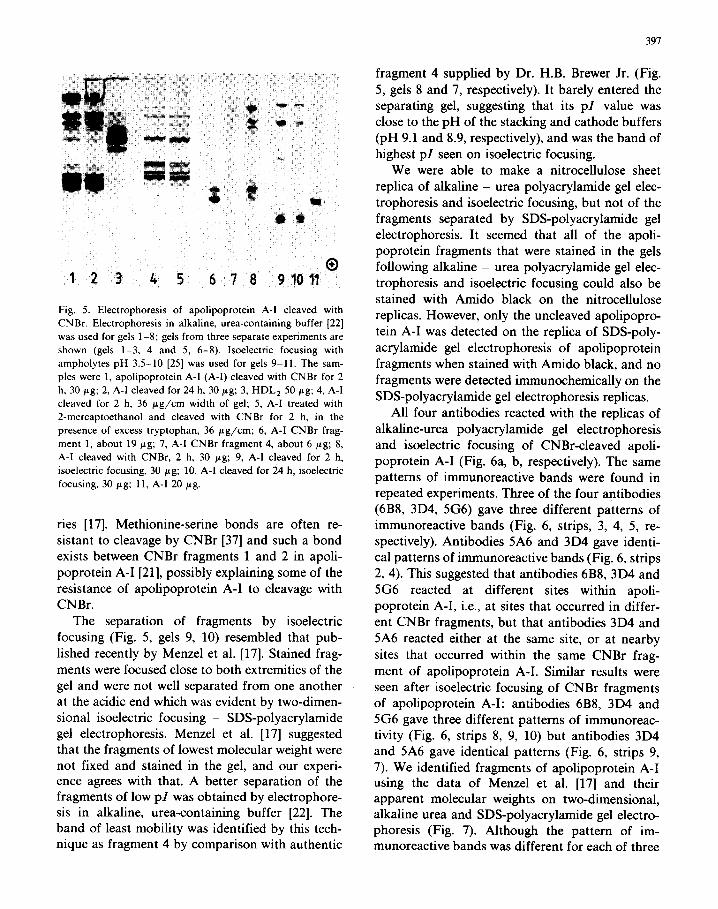
patterns of the separated fragments from apoli- poprotein A-l cleaved for 2 and 24 h (Fig. 5, gels 1, 2) and A-l treated with mercaptoethanol before cleavage, and then cleaved in the presence of ex-
cess tryptophan (Fig. 5, gel 5). In each experiment there was cleavage of apolipoprotein A-l, yielding a pattern of bands that was not noticeably in- fluenced by the different conditions of cleavage that we tested (Fig. 5, compare gels 1 with 2, and 4 with 5). The cleavage appeared to be incomplete, some apolipoprotein A-l remained intact and there were many bands of incompletely cleaved apoli- poprotein A-l fragments. This has been a common finding for apolipoprotein A-l in other laborato-
397
Fig. 5. Electrophoresis of apolipoprotein A-I cleaved with
CNBr. Electrophoresis in alkaline, urea-containing buffer [22]
was used for gels 1-8; gels from three separate experiments are
shown (gels 1-3, 4 and 5, 6-8). Isoelectric focusing with
ampholytes pH 3.5-10 [25] was used for gels 9-11. The sam-
ples were 1, apolipoprotein A-I (A-I) cleaved with CNBr for 2
h, 30 pg; 2, A-l cleaved for 24 h. 30 gg; 3, HDL, 50 pg; 4, A-I
cleaved for 2 h, 36 pg/cm width of gel; 5, A-I treated with
2-mercaptoethanol and cleaved with CNBr for 2 h, in the
presence of excess tryptophan, 36 pg/cm; 6, A-I CNBr frag-
ment 1, about 19 pg; 7, A-I CNBr fragment 4, about 6 pg; 8,
A-I cleaved with CNBr, 2 h, 30 pg; 9, A-I cleaved for 2 h,
isoelectric focusing, 30 pg; 10, A-I cleaved for 24 h, isoelectric
focusing, 30 pg; 11, A-I 20 pg.
ries [17]. Methionine-serine bonds are often re-
sistant to cleavage by CNBr [37] and such a bond exists between CNBr fragments 1 and 2 in apoli-
poprotein A-I [21], possibly explaining some of the resistance of apolipoprotein A-I to cleavage with
CNBr.
The separation of fragments by isoelectric
focusing (Fig. 5, gels 9, 10) resembled that pub-
lished recently by Menzel et al. [17]. Stained frag- ments were focused close to both extremities of the
gel and were not well separated from one another at the acidic end which was evident by two-dimen-
sional isoelectric focusing - SDS-polyacrylamide gel electrophoresis. Menzel et al. [17] suggested that the fragments of lowest molecular weight were not fixed and stained in the gel, and our experi- ence agrees with that. A better separation of the fragments of low pZ was obtained by electrophore- sis in alkaline, urea-containing buffer [22]. The band of least mobility was identified by this tech- nique as fragment 4 by comparison with authentic
fragment 4 supplied by Dr. H.B. Brewer Jr. (Fig.
5, gels 8 and 7, respectively). It barely entered the separating gel, suggesting that its pZ value was
close to the pH of the stacking and cathode buffers (pH 9.1 and 8.9, respectively), and was the band of
highest pZ seen on isoelectric focusing. We were able to make a nitrocellulose sheet
replica of alkaline - urea polyacrylamide gel elec-
trophoresis and isoelectric focusing, but not of the fragments separated by SDS-polyacrylamide gel
electrophoresis. It seemed that all of the apoli- poprotein fragments that were stained in the gels following alkaline - urea polyacrylamide gel elec-
trophoresis and isoelectric focusing could also be stained with Amido black on the nitrocellulose
replicas. However, only the uncleaved apolipopro-
tein A-I was detected on the replica of SDS-poly-
acrylamide gel electrophoresis of apolipoprotein fragments when stained with Amido black, and no
fragments were detected immunochemically on the
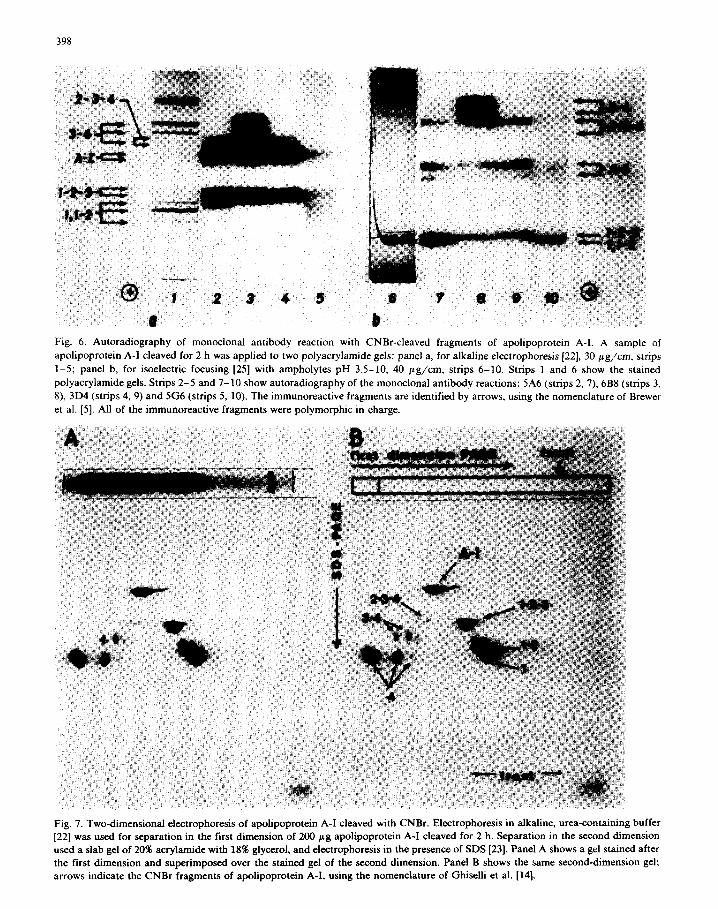
SDS-polyacrylamide gel electrophoresis replicas. All four antibodies reacted with the replicas of
alkaline-urea polyacrylamide gel electrophoresis
and isoelectric focusing of CNBr-cleaved apoli- poprotein A-I (Fig. 6a, b, respectively). The same patterns of immunoreactive bands were found in
repeated experiments. Three of the four antibodies (6B8, 3D4, 5G6) gave three different patterns of immunoreactive bands (Fig. 6, strips, 3, 4, 5, re- spectively). Antibodies 5A6 and 3D4 gave identi-
cal patterns of immunoreactive bands (Fig. 6, strips
2, 4). This suggested that antibodies 6B8, 3D4 and 5G6 reacted at different sites within apoli-
poprotein A-I, i.e., at sites that occurred in differ-
ent CNBr fragments, but that antibodies 3D4 and
5A6 reacted either at the same site, or at nearby
sites that occurred within the same CNBr frag-
ment of apolipoprotein A-I. Similar results were
seen after isoelectric focusing of CNBr fragments of apolipoprotein A-I: antibodies 6B8, 3D4 and 5G6 gave three different patterns of immunoreac-
tivity (Fig. 6, strips 8, 9, 10) but antibodies 3D4 and 5A6 gave identical patterns (Fig. 6, strips 9, 7). We identified fragments of apolipoprotein A-I using the data of Menzel et al. [17] and their apparent molecular weights on two-dimensional, alkaline urea and SDS-polyacrylamide gel electro- phoresis (Fig. 7). Although the pattern of im- munoreactive bands was different for each of three
398
Fig. 6. Autoradiography of monoclonal antibody reaction with CNBr-cleaved fragments of apolipoprotein A-l. A sample of
apolipoprotein A-I cleaved for 2 h was applied to two polyacrylamide gels: panel a, for alkaline electrophoresis 1221, 30 pg/cm, strips
1-5; panel b, for isoelectric focusing [25] with ampholytes pH 3.5-10, 40 pg/cm, strips 6-10. Strips 1 and 6 show the stained
polyacrylamide gels. Strips 2-5 and 7-10 show autoradiography of the monoclonal antibody reactions: 5A6 (strips 2, 7). 6B8 (strips 3,
8). 3D4 (strips 4, 9) and 5G6 (strips 5, 10). The immunoreactive fragments are identified by arrows, using the nomenclature of Brewer
et al. [5]. All of the immunoreactive fragments were polymorphic in charge.
Fig. 7. Two-dimensional electrophoresis of apolipoprotein A-I cleaved with CNBr. Electrophoresis in alkaline, urea-containing buffer
[22] was used for separation in the first dimension of 200 pg apolipoprotein A-I cleaved for 2 h. Separation in the second dimension
used a slab gel of 20% acrylamide with 18% glycerol, and electrophoresis in the presence of SDS [23]. Panel A shows a gel stained after
the first dimension and superimposed over the stained gel of the second dimension. Panel B shows the same second-dimension gel; arrows indicate the CNBr fragments of apolipoprotein A-I, using the nomenclature of Ghiselli et al. [14].

antibodies (Fig. 6, strips 2-5) some fragments or bands reacted with all four of the antibodies, e.g., uncleaved apolipoprotein A-I and fragment l-2-3.
A pair of bands (fragment 2-3-4) reacted with antibodies 6B8, 5A6 and 3D4 (Fig. 6, strips 3, 2, 4), thus containing two different antigenic sites (6B8 and 5A6-3D4). Antibody 6B8 was the only antibody that reacted with three bands (fragment 3-4) which migrated between fragment 4 and apolipoprotein A-I. Thus, fragment 3-4 contained the antigenic site of only 6B8, but fragment 2-3-4 contained the site of 3D4-5A6 as well, and frag- ment l-2-3 contained all three antigenic sites (6B8, 5G6, 3D4-5A6). Antibody 5G6 was the only antibody which reacted with a pair of bands (frag- ment l-2 or 1; Fig. 6, strip 5) which we tentatively identified as fragment l-2 or 1, by comparison with authentic fragment 1 from Dr. H.B. Brewer Jr. and from the data of Menzel et al. [17] and apparent molecular weights (Fig. 7). Fig. 8 sum- marizes the CNBr cleavage sites of apolipoprotein A-I, and Table I lists the patterns of reactions of the antibodies and fragments. For each antibody we could see that the immunoreactive fragments contained a specific part of the apolipoprotein A-I structure, and that fragments missing that struc- ture did not react with the antibody. The results therefore indicate that the antigenic site for 6B8 is in fragment 3, and that the 3D4-5A6 site is in fragment 2. The 5G6 reactions were not as intense on autoradiography as those of the other antibod- ies. Nevertheless, 5G6 gave its strongest reactions when fragments 1 and 2 were present, and we suggest that the 5G6 site is either in fragment 1, or spans the CNBr cleavage site between fragments 1
TABLE I
399
“2fJm l Coon
RESIDUE I 86 112 14.9 243
AV Met Met Met Gill ‘--k
CNBr I CNEr 2 CNBr 3 CNBr 4
Mr 9880 3190 4250 10700
Fig. 8. Simplified diagram of the primary structure of apoli- poprotein A-I. The position of the methionine residues at
which CNBr can potentially cleave the peptide chain are shown.
The fragments are indicated, as CNBr 1 to CNBr 4, using the
nomenclature of Brewer et al. (51, and their calculated molecu-
lar weights are shown.
and 2, or requires the presence of intact fragments 1 and. 2 for a protein conformation that expresses antigenic activity.
All of the charge polymorphs of apoli- poproteins A-I reacted with all antibodies (Fig. 2, strips 4-9), indicating that the charge heterogene- ity of apolipoprotein A-I is qualitatively unrelated to the presence of antigenic sites recognized by these four antibodies. It appeared that the CNBr fragments of apolipoprotein A-I were themselves polymorphic in charge, both from their patterns of immunoreaction (Fig. 6) and from the series of two or three spots of identical molecular weight but of a different charge than we saw on two-di- mensional polyacrylamide gel electrophoresis (Fig. 7). The charge heterogeneity is most evident for fragments 4 and 3-4, but is also present for frag- ments 1, 1-2, and l-2-3 (Fig. 7). Thus, it may be that several parts of apolipoprotein A-I are poly- morphic in charge, and that the polymorphism of whole A-I is the average of these potentially varia- ble sites within the molecule. This charge polymor- phism was seen in both fragments 1 and 4 sup- plied by Dr. H.B. Brewer Jr. (Fig. 5, gels 6, 7).
IMMUNOREACTIVITY OF CNBKLEAVED FRAGMENTS OF APOLIPOPROTEIN A-I
Reactions were found on replicas of alkaline electrophoresis or isoelectric focusing (Fig. 6).
Apolipoprotein
Common fragments
Fragment
A-I 2-3-4
3-4 l-2-3 l-2orl
Antibodies
3D4 5A6 6B8 5G6
+ + + + + + + - - + -
+ + + + - _ - + 2 or 2-3 2 or 2-3 3 I,2 or 1-2

400
In conclusion, we have characterized four monoclonal antibodies which react with a protein which has the molecular weight and charge char- acteristics of apolipoprotein A-I and also with purified apo~poprotein A-I. Noteworthy was the observation that these antibodies react with all known charge polymorphs of apolipoprotein A-I and pro A-I and that therefore the antigenic de- terminants recognized by these antibodies are un- affected by this polymo~~sm. The finding that different CNBr-fragments of apolipoprotein A-I are also heterogeneous in charge is important. since it indicates that apolipoprotein A-I charge polymorphism is the sum of several charge varia- tions at different sites within the molecule. Com- petitive radioi~unoassays using different pairs of antibodies have shown that the antibodies re- cognized antigenic sites present together on apolipoprotein A-I molecules rather than on alter- nate molecules and that three different sites are expressed and identified by 6B8, 5G6 and by the pair, 3D4 and 5A6. The location of these sites on the apolipoprotein‘A-I molecule has been further defined by the CNBr-cleavage experiments which indicate that 5G6 is on fragment 1 or l-2,3D4-5A6 on fragment 2, and 6B8 on fragment 3. While these antibodies recognize antigenic deter~nants that are far apart on apolipoprotein A-I molecule, it is interesting to note that none of them could inhibit the apolipoprotein A-I-mediated activation of lecithin : cholesterol acyltransferase reaction in a system using a purified enzyme and an apoli- poprotein A-I-phosphatidylcholine-cholesterol substrate (unpublished results). These results are consistent with the data of several groups, showing that the apolipoprotein A-I fragment responsible for activation of the enzyme is located within CNBr fragment 4 [2], a fragment not recognized by our antibodies. Further experiments with these and other antibodies specific for different sites will be needed to help elucidate the mechanism which mediates the apolipoprotein A-I activation of lecithin : cholesterol acyltransferase. Finally, we have observed that the four ~tibodies described here are specific for sites on apolipoprotein A-I which are expressed equally well in native as in delipidated HDL and which should thus be appli- cable to quantitative immunoassays of apolipopro- tein A-I.
Acknowledgements
We are grateful to Dr. H.B. Brewer Jr., Molecu- lar Disease Branch, National Heart, Lung and Blood Institute, National Institutes of Health, U.S.A. for the generous gift of purified A-I CNBr fragments 1 and 4. Louise Lalonde gave excellent secretarial help. We are indebted to C. Vezina and T.D. N’guyen for their assistance in several parts of this work. The studies were supported by grants from the Medical Research Council of Canada (PG-27), the Quebec Heart Foundation and the Fondation de Recherche en Sante du Quebec. R.W.M. is a scholar of the Fondation de Re- cherche en Sante du Quebec. P.M. is a recipient of an Industrial Research Fellowship {Natural Scien- ces and Engineering Research Council of Canada) and under contract with A-B Biological Supplies Inc., Hamilton, Ontario.
References
I
2
3
4
5
6
7
8
9
10
II
12
13
14
15
16
Eisenberg, S. (1984) J. Lipid Res. 25, 1017-1058
Marcel, Y.L. (1982) Adv. Lipid Res. 19. 85-136
Avogaro. P., Cazzolato, G., Bittolo Bon, G. and Quinzi.
G.B. (1979) Lancet i. 901-903
Weisweiler, P., Sperland. B. and Schwandt, P. (1981) Clin.
Chem. 27, 348
Maciejko, J.J., Holmes, D.R., Kottke, B.A., Zinsmeister,
A.R., Dinh, D.M. and Mao, S.J.T. (1983) N. Engl. J. Me.d
309, 385-389
Lippel, K. (ed.) (1983) Proceedings of the Workshop on
apolipoproteinquantification. In U.S. Department of Health
and Human Services, National Institutes of Health. NIH
Publication 83-1266, pp. 7-22. 443-449. 450-458
Steinberg, K.K., Cooper, G.R.. Graiser. S.R. and Rosseneu.
M. (1983) Chin. Chem. 29, 415-426
Vittelo, L.B. and Scanu. A. (1976) J. Biol. Chem. 251.
1131.-1136
Formisano, S., Brewer. H.B., Jr. and Osborne, J.C. (1978) J.
Biol. Chem. 253, 354-359
Schonfeld. G., Bradshaw, R.A. and Chen. J.S. (1976) J.
Biol. Chem. 251, 3921-3926
Mao, S.J.T.. Miller, J.P., Gotto, A.M.. Jr. and Sparrow. J.T. (1980) J. Biol. Chem. 255. 3448-3453
Nestruck. A.C., Suzue, G. and Marcel. Y.L. (1980) Biochim.
Biophys. Acta 677, 110-l 21
Shore. G. and Shore, V. (1969) Biochemistry 8, 4510-4516
Zannis, V.I.. Breslow, J.L. and Katz. A.J. (1980) J. Biol.
Chem. 255. 8612-8617
Utermann, G.. Feussner, G.. Franceschini, G., Haas. J. and
Steinmetz, A. (1982) J. Biol. Chem. 257, Sol-507
Ghiselli, G., Schaefer, E.J., Light, J.A. and Brewer, H.B., Jr.
(1983) J. Lipid Res. 24, 731-736

401
17 Menzel, H.J., Assman, G., Rall, SC., Weisgraber, K.H. and
Mahley, R.W. (1984) J. Biol. Chem. 259, 3070-3076
18 Milne, R.W., Douste-Blazy, p., Retegui, L. and Marcel,
Y.L. (1981) J. Clin. Invest. 68, 111-117
19 Chaion, M.-P., Milne, R.W. and Vaerman, J.-P. (1979) Eur.
J. Immunol. 9, 747-751
20 Olofsson, S.-O., McConathy, W.J. and Alaupovic, P. (1978)
Biochemistry 17, 1032-1036
21 Brewer, H.B., Jr., Fairwell, T., LaRue, A., Ronan, R.,
Houser, A. and Bronzert, T.J. (1978) B&hem. Biophys.
Res. Commun. 80, 623-630
22 Kane, J.P. (1973) Anal. B&hem. 53, 350-364
23 Neviile, D.M. (1971) J. Biol. Chem. 246, 6328-6334
24 Connelly, P.W. and Kuksis, A. (1982) B&him. Biophys.
Acta 711, 245-251
25 Wamick, G.R., Mayfield, C., Albers, J.J. and Hazzard,
W.R. (1979) Clin. Chem. 25, 279-284
26 Reisner, A.H., Nemes, P. and Bucholtz, C. (1975) Anal.
B&hem. 64, 509-516
27 Holbrook, LB. and Leaver, A.G. (1976) Anal. Biochem. 75,
634-636
28 Towbin, H., Staehelin, T. and Gordon, J. (1979) Proc. Nat].
Acad. Sci. USA 76, 4350-4354
29 Theolis, R., Jr., Weech, P.K., Marcel, Y.L. and Mime, R.W.
(1984) Arteriosclerosis 4, 498-509
30 Mellman, I.S. and Unkeless, J.C. (1980) J. Exp. Med. 152,
1048-1069
31 Milne, R.W., Theohs, R., Jr., Verdery, R.B. and Marcel,
Y.L. (1983) Arteriosclerosis 3, 23-30
32 Brewer, H.B., Jr., Fairwell, T., Kay, L., Meng, M., Ronan,
R., Law, S. and Light, J.A. (1983) Biochem. Biophys. Res.
Commun. 113, 626-632
33 Shore, V., Salen, G., Cheng, F.W., Forte, T., Shefer, S.,
Tint, G.S. and Lindgren, F.T. (1981) J. Chn. Invest. 68,
1295-1304
34 Tosi, R., Tanigaki, N., Sorrentino, R., Accolla, R. and
Corte, G. (1981) Eur. J. Immunol. 11, 721-726
35 Ehrhch, P.H., Moyle, W.R., Moustafa, Z.A. and Canfield,
R.E. (1982) J. Immunol. 128, 2709-2713
36 Holmes, N.J. and Parham, P. (1983) J. Biol. Chem. 258,
1580-1586
37 Schroeder, W.A. Shelton, J.B. and Shelton, J.R. (1969)
Arch. Biochem. Biophys. 130, 551-556





























