Embed Size (px)
Citation preview

B12 – Biologie végétale L1 Bio 2010/2011
Page 1 sur 39
Référence :
Lafon J.P. 1990
Tharaud C. Biologie plantes cultivées
Levy G. Ed lavoisier
Guignard et Dupont Botanique systématique moléculaire
Abrégés Masson
I. Organisation générale (angiospermes)
Structure de la cellule végétale :
- paroi cellulosique
- la membrane cytoplasmique
- cytoplasme
- le noyau
Anatomie de l’appareil végétatif :
- les tissus végétaux
- la racine
- la tige
- la feuille
Anatomie de l’appareil reproducteur :
- organisation de la fleur isolée
- variation de l’organisation florale
- groupement de fleurs
- répartition des sexes
- structures des organes reproducteurs
Biologie de la reproduction :
- pollinisation
- germination du pollen
- fécondation
- reproduction sans fécondation
- formation de la graine
- formation du fruit
II. Systématique
Généralité sur les grands groupes végétaux :
- les thallophytes (algues, champignons, lichens)
- les embryophytes (plantes terrestres : bryophytes, ptéridophytes, gymnospermes,
angyospermes)
Les algues
Les champignons

B12 – Biologie végétale L1 Bio 2010/2011
Page 2 sur 39
Les lichens
Les bryophytes
Les trachéophytes :
- les ptéridophytes (fougères, lycopodes, sélaginelles, prêles)
Les spermatophytes :
- les gymnospermes (ginkgo, conifères, gnétophytes)
- Les angiospermes (paléodicots, monocots, eudicots)
B12 – Biologie végétale L1 Bio 2010/2011
Page 3 sur 39
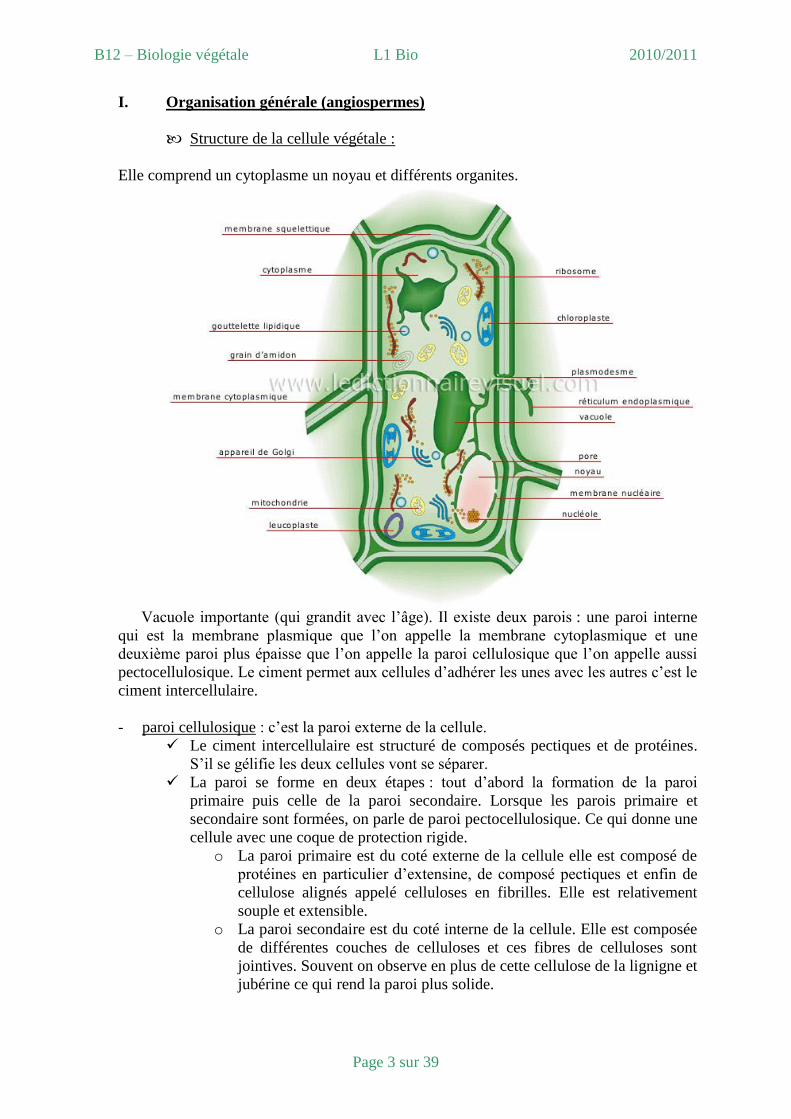
I. Organisation générale (angiospermes)
Structure de la cellule végétale :
Elle comprend un cytoplasme un noyau et différents organites.
Vacuole importante (qui grandit avec l’âge). Il existe deux parois : une paroi interne
qui est la membrane plasmique que l’on appelle la membrane cytoplasmique et une
deuxième paroi plus épaisse que l’on appelle la paroi cellulosique que l’on appelle aussi
pectocellulosique. Le ciment permet aux cellules d’adhérer les unes avec les autres c’est le
ciment intercellulaire.
- paroi cellulosique : c’est la paroi externe de la cellule.
Le ciment intercellulaire est structuré de composés pectiques et de protéines.
S’il se gélifie les deux cellules vont se séparer.
La paroi se forme en deux étapes : tout d’abord la formation de la paroi
primaire puis celle de la paroi secondaire. Lorsque les parois primaire et
secondaire sont formées, on parle de paroi pectocellulosique. Ce qui donne une
cellule avec une coque de protection rigide.
o La paroi primaire est du coté externe de la cellule elle est composé de
protéines en particulier d’extensine, de composé pectiques et enfin de
cellulose alignés appelé celluloses en fibrilles. Elle est relativement
souple et extensible.
o La paroi secondaire est du coté interne de la cellule. Elle est composée
de différentes couches de celluloses et ces fibres de celluloses sont
jointives. Souvent on observe en plus de cette cellulose de la lignigne et
jubérine ce qui rend la paroi plus solide.
B12 – Biologie végétale L1 Bio 2010/2011
Page 4 sur 39
Dans la réalité les cellules ne sont pas indépendante entre elles, il y a des connections
que l’on appelle les plasmodesmes. Le cytoplasme des cellules se joint (grâce au
plasmodesme) et ce continuum cytoplasmique prend le nom de symplasme.
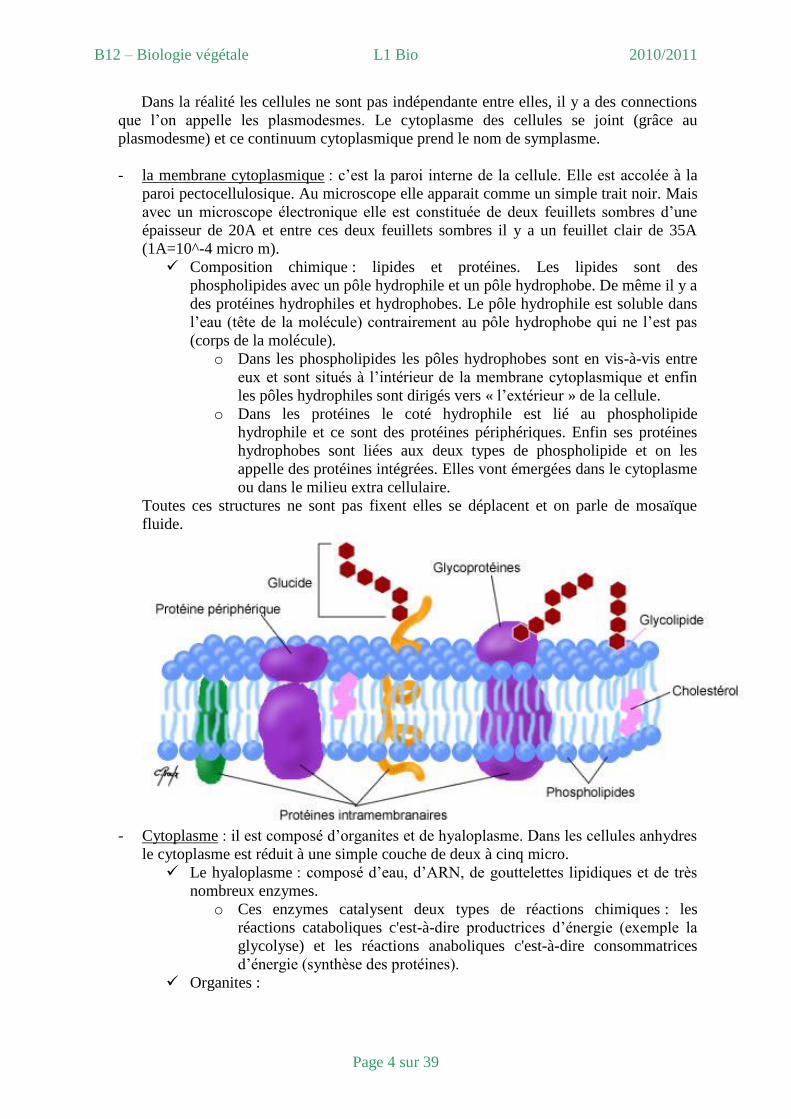
- la membrane cytoplasmique : c’est la paroi interne de la cellule. Elle est accolée à la
paroi pectocellulosique. Au microscope elle apparait comme un simple trait noir. Mais
avec un microscope électronique elle est constituée de deux feuillets sombres d’une
épaisseur de 20A et entre ces deux feuillets sombres il y a un feuillet clair de 35A
(1A=10^-4 micro m).
Composition chimique : lipides et protéines. Les lipides sont des
phospholipides avec un pôle hydrophile et un pôle hydrophobe. De même il y a
des protéines hydrophiles et hydrophobes. Le pôle hydrophile est soluble dans
l’eau (tête de la molécule) contrairement au pôle hydrophobe qui ne l’est pas
(corps de la molécule).
o Dans les phospholipides les pôles hydrophobes sont en vis-à-vis entre
eux et sont situés à l’intérieur de la membrane cytoplasmique et enfin
les pôles hydrophiles sont dirigés vers « l’extérieur » de la cellule.
o Dans les protéines le coté hydrophile est lié au phospholipide
hydrophile et ce sont des protéines périphériques. Enfin ses protéines
hydrophobes sont liées aux deux types de phospholipide et on les
appelle des protéines intégrées. Elles vont émergées dans le cytoplasme
ou dans le milieu extra cellulaire.
Toutes ces structures ne sont pas fixent elles se déplacent et on parle de mosaïque
fluide.
- Cytoplasme : il est composé d’organites et de hyaloplasme. Dans les cellules anhydres
le cytoplasme est réduit à une simple couche de deux à cinq micro.
Le hyaloplasme : composé d’eau, d’ARN, de gouttelettes lipidiques et de très
nombreux enzymes.
o Ces enzymes catalysent deux types de réactions chimiques : les
réactions cataboliques c'est-à-dire productrices d’énergie (exemple la
glycolyse) et les réactions anaboliques c'est-à-dire consommatrices
d’énergie (synthèse des protéines).
Organites :

B12 – Biologie végétale L1 Bio 2010/2011
Page 5 sur 39
o Réticulum endoplasmique : Réseau dense de canicule infra
microscopique. Qualité unique entre elles s’il y a des ribosomes sur ce
réticulum on parlera de réticulum granuleux. Ce réticulum granuleux
est l’ergastoplasme. S’il n’y a pas de ribosomes on parlera de ribosome
lisse. Les vacuoles sont des dilations de ce réticulum. Le rôle de ce
réticulum est de conserver et canaliser les molécules dans la cellule et
la fabrication de macro molécule (protéines lipides) en association avec
l’appareil de golgi, les lysosomes et la membrane plasmique.
o Ribosomes : en moyenne on est entre 0,02 et 0,03 µm ils contiennent
40% d’acides ribonucléique et leur rôle principal : ils synthétisent les
chaines peptidiques en décodant le message porté par l’ARN messager
c’est donc en bref le lieu de synthèse des protéines. Ces ribosomes ont
les trouvent aussi avec ou dans les mitochondries. Ces chaines
peptidiques une fois synthétisés peuvent passer dans les cavités du
réticulum. (voir schéma de la synthèse de protides par les ribosomes).
o L’appareil de Golgi : il est constitué d’unités appelées dictyosomes.
Chaque dictyosomes en fait sont une pile de sac aplatis. On y retrouve
des vésicules qui sont concentrés en protéines et ces vésicules peuvent
se décrocher de l’appareil de Golgi. C’est un phénomène d’exocytose.
Enfin ces sac aplatis peuvent communiquer avec le réticulum
endoplasmique. (voir reconstitution dans l’espace d’un dictyosomes et
interprétation d’un dictyosome) La fragmentation des sacs aplatis
peuvent former des vésicules détachés.
o Lysosomes : ce sont de petites vésicules d’environs ½µm de diamètre
avec une membrane. Et leur rôle : ils sont riches en enzymes qui vont
permettre de digérer des corps étrangers.
o Mitochondries : elles sont cylindriques ou sphériques entre 1 et
quelques µm de longueur. L’enveloppe est constituée de deux
membranes qui ont la même structure que la membrane plasmique. La
membrane interne peu faire des replis et ces replis qui prennent le nom
de crêtes. En générale elles sont perpendiculaires à l’axe longitudinal
de la mitochondrie. Et le fait d’avoir ces crête augmente la surface
interne ce qui facilite l’action des enzymes. Enfin la substance interne
des mitochondries : une de base qui est la matrice et on y trouve aussi
de l’ADN des ARN des ribosomes des enzymes KREBS. Sont rôle : les
mitochondries sont le siège des réactions d’oxydation de la respiration :
ça produit de l’énergie qui est stocké sous forme d’ATP. Sur la
membrane interne on a des sphères pédicellées. Ce sont ces sphères qui
produisent l’ATP. (voir schéma reconstitution dans l’espace d’une
mitochondrie, interprétation d’une mitochondrie et membrane interne
d’une mitochondrie)
o Plastes : lorsque la cellule végétale est jeune elle va renfermer des pros
plastes. Lorsqu’elle vieilli sous l’action de la lumière on parlera de
chloroplastes. Ils sont verts et donnent la couleur verte des végétaux. La
structure de ces plastes : ils sont constitués de deux membranes. Une

B12 – Biologie végétale L1 Bio 2010/2011
Page 6 sur 39
substance interne prend le nom de stroma dans lequel on retrouve des
grains d’amidon, des lipides, de l’ADN, de l’ARN et des ribosomes
également des enzymes et en particulier des enzymes qui interviennent
dans le cycle de Calvin. Ils sont composés de différents saccules aplatis
et l’ensemble de ses saccules prend le nom de tylakoïdes. C’est au
niveau de ces tylakoïdes que se fait la photosynthèse. Enfin quand on a
un ensemble d’au moins 10 tylakoïdes, on lui donne le nom de
granuum. Les pro plastes évoluent majoritairement en chloroplastes
mais parfois ceux-ci forment des leucoplastes que l’on trouve dans les
racines et ils ont la caractéristique essentiel d’être sans tylakoïdes. On
va retrouver également des amyloplastes que l’on retrouve dans les
organes de réserves. Ils sont des leucoplastes qui synthétisent de
l’amidon à partir du saccarose des feuilles. On trouve également des
chromoplastes ils synthétisent principalement des pigments
caroténoïdes on les retrouve essentiellement dans les fleurs et les fruits.
(voir interprétation d’un chloroplaste et reconstitution dans l’espace
d’un chloroplaste).
o Peroxysomes : ils ressemblent aux mitochondries mais n’ont qu’une
seule membrane et leur rôle est de participer à la photorespiration.
o Les glyoxysomes : ils participent à l’oxydation des acides.
- le noyau : il est constitué par une enveloppe formée de deux membranes et cette
enveloppe possède des ouvertures qu’on appelle des pores. Elles peuvent être
obstruées par des molécules qui limitent les échanges entre le noyau et le cytoplasme.
Dans ce noyau on a un ou plusieurs nucléoles qui sont la synthèse des ARN
ribosomaux. Le reste c’est de la chromatine qui constitue les chromosomes. Son rôle
est d’agir dans la division cellulaire et dans la synthèse de protéines. (voir
reconstitution dans l’espace d’un noyau, interprétation d ‘une coupe transversale du
noyau et les principaux rôles des divers organites de la cellule)

B12 – Biologie végétale L1 Bio 2010/2011
Page 7 sur 39
Anatomie de l’appareil végétatif :
Les organes végétatifs assurent la nutrition et la croissance de la plante. Ils sont au nombre
de trois : la tige, la racine et les feuilles. Ce qui les relie on des tissus avec les mêmes
caractéristiques.
- les tissus végétaux :
Définition : c’est un ensemble de cellules identiques qui jouent le même rôle, ces
tissus sont donc spécialisés, mais à l’origine tous ces tissus proviennent d’un tissu
particulier non spécialisé qui prennent le nom de méristèmes qu’on appelle
également les tissus embryonnaires.
Les méristèmes : les cellules sont indifférenciées et la multiplication est rapide. On
a deux types de méristème : les méristèmes primaires et les méristèmes
secondaires. (voir mode de division des cellules cambiales : c’est toujours la
cellule génératrice qui se divise)
o Méristèmes primaires : ce sont les premiers tissus formés dans l’embryon
elles sont de petite taille et isodiamétriques. Le volume du noyau est
important, les vacuoles sont petites et les mitochondries et les plastes sont
peu structurés. (voir figure de la cellule de méristème primaire) ces
méristèmes primaires sont situés aux extrémités de la racine et de la tige.
Dans ce cas là on les appelle les méristèmes apicaux. L’évolution comporte
deux étapes : la multiplication des cellules et l’élongation et la
différenciation de ces cellules. Elle assure la croissance en longueur de la
racine et de la tige et la formation des tissus primaires. (voir localisation
des méristèmes apicaux) au niveau de la racine il y a une protection
appelée la coiffe. Le processus reste complexe.
o Méristèmes secondaires : ce sont les zones génératrices on les appelle
également les cambiums et assises annulaires. Ils apparaissent après la
formation des organes. Les cellules sont rectangulaires, le noyau est
relativement petit. Et on observe de même une grande vacuole qui occupe
la plupart du cytoplasme. Ces cambiums sont aux nombre de deux. Ils se
situent dans la racine et dans la tige. (voir couche transversale de tige ou de
racine)
La première zone est la zone subero-pmeclodermique, c’est là qu’il
y a la production de liège scientifiquement appelé suber et de
phelloderme qui va avoir un rôle de protection.
La deuxième zone est la zone libero-ligneuse où il y a la production
de deux types de tissus : le phloème et le xylème. Cette partie
produit le bois. Ils assurent la croissance en épaisseur et la
formation des tissus secondaires.
En particulier le liège le phelloderme, le phloème et le xylème secondaires.
Les tissus primaires forment la structure primaire des organes jeunes. C'est-à-dire
lorsque la croissance n’est pas achevée. Il existe 5 tissus primaires :

B12 – Biologie végétale L1 Bio 2010/2011
Page 8 sur 39
o Nutrition : on les appelle les parenchymes. Les cellules sont polygonales
ou arrondies. Les espaces entre les cellules prennent le nom de méats. Elles
sont proches de l’état mérismatique. Il y a deux types de parenchymes :
les parenchymes chlorophylliens où se fait l’activité
photosynthétique et on les retrouve dans les feuilles et la partie
externe de la tige.
Les parenchymes de réserves en particulier sous forme d’amidon
dans les amidoplastes. (voir parenchyme à chloroplastes)
o Protection : on les appelle également les épidermes. C’est une couche
continue de cellules qui recouvre les tiges et les feuilles. Les cellules ont
des plastes mais sans chlorophylles et elles sécrètent un revêtement
extérieur qui prend le nom de cuticule. Cette cuticule est composée de
cutine et de cires et également la présence de poils (voir figure 6 schéma
simplifié de la cuticule.)
o Soutien : ils assurent la solidité de la plante et dans ces tissus de soutiens
on a deux types de tissus :
le collenchyme : tissu des organes jeunes à croissance non achevé il
présente des cellules à paroi cellulosique épaisse mais qui
permettent l’allongement et enfin elles sont toujours vivantes.
Le sclérenchyme : constitué seulement de cellules mortes qui sont
riche en lignine non extensibles (voir figure 7 et 8 du collenchyme
et du sclérenchyme).
o Conducteur : deux types :
Xylème : dans les tissus primaires on appelle ces tissus le bois qui
conduit la sève brute ou montante qui est constituée d’eau et de sels
minéraux. Deux cas de figures :
Trachéides : premier type d’élément conducteur ils
proviennent de cellules dont les parois transversales sont
persistantes et dans ce cas là la sève brute circule en
« chicane » dans les régions non lignifiées.
Les vaisseaux : proviennent de cellules dont les parois
transversales ont disparu. Ce sont donc des tubes constitués
de fils de cellules et dans ce cas là la sève circule
verticalement (voir figure 9 éléments conducteurs de la sève
brute).
Dans le xylème primaire en fonction du type de croissance végétale on
va distinguer le protoxylème et le métaxylème.
Protoxylème : ce sont les premiers éléments formé avant
que l’allongement de l’organe soit terminé. Et dans ce cas là
ce sont toujours des trachéides (voir figure 10 types de
trachéides du protoxylème).
Métaxylème : il apparait au contact du protoxylème lorsque
la croissance de l’organe est achevée. Chez les
ptéridophytes et gymnospermes ce sont des trachéides
(scalariforme) et chez les angiospermes ce sont des
vaisseaux (voir figure 12 trachéides à ponctuations aréolées
et figure 13 différents vaisseaux du métaxylème).
Le phloème : c’est le phloème primaire ou liber va élaborer la sève
descendante qui est constituée d’eau et de tous les produits de
synthèses de la plante qu’elle va redistribuer à l’ensemble de la

B12 – Biologie végétale L1 Bio 2010/2011
Page 9 sur 39
plante. Les éléments conducteurs sont les tubes criblés. Ils peuvent
se fermer pendant la mauvaise saison et ils se ferment par du
callose qui bouchent les pores (figure 14 tube criblé).
o Sécréteur : hors programme.
Tissus secondaires : ils forment la structures de la plantes ils apparaissent après
que les organes soient formés et ils s’ajoutent aux tissus primaires. La croissance
en longueur est achevée et par conséquent ces tissus secondaires assurent la
croissance en épaisseur.
o Tissus conducteurs : ils sont constitués de xylèmes secondaires et de
phloèmes secondaires qui proviennent du cambium libéro-ligneux (voir
figure 6 les tissus secondaires).
Xylème secondaire : il se forme vers l’intérieur du cambium se sont
des vaisseaux ponctués avec toute une série de fibres qui sont
constituées de cellules mortes et de lignines, et de cellules
parenchymateuse (cellules vivantes riches en réserves). Son rôle est
conducteur mais avec les fibres il a également un rôle de soutien et
des éléments de réserves avec ces cellules parenchymateuses.
Phloème secondaire : il se forme vers l’extérieur du cambium, il est
constitué de tube criblé. Le fonctionnement est rythmique (phase de
fonctionnement lente et rapide en fonction des saisons) c’est ce qui
donne les cercles dans les troncs d’arbre issus à partir de ce
phloème secondaire.
o Les autres tissus sont issus du cambium subero-phellodermique avec la
formation du liège (vers l’extérieur du cambium) les cellules dans ce cas là
sont mortes et riches en subérine. C’est donc un rôle de protection. Le
phelloderme se forme vers l’intérieur du cambium et c’est du parenchyme
chlorophyllien.
- la racine
Morphologie :
o Il y a une racine principale avec des ramifications et dans ce cas là les
ramifications prennent le nom de racine de 2ème
ordre 3ème
etc. On parle de
racine latérale, ou racine adventives et dans ce cas là elles se développent à la
surface au niveau du sol et des tiges, ex : le liège.
o Il y a trois zones au niveau de la racine : la coiffe, la zone pilifère et la zone
subéreuse
La coiffe est une sorte d'étui qui protège le méristème apicale et le
renouvellement se fait par la face interne.
La zone pilifère est recouverte de poils qui absorbent l'eau et au fur
et à mesure que la racine pousse les poils supérieurs tombent et les
poils inférieurs poussent.
o Anatomie au niveau de la structure primaire : Celle-ci est constituée de deux
parties : L'écorce et le cylindre centrale.
L’écorce : En même temps que se forment les poils absorbants, les

B12 – Biologie végétale L1 Bio 2010/2011
Page 10 sur 39
autres tissus de la racine se différencient en général nettement en
écorce et cylindre central. Les cellules de l’écorce sont
parenchymateuses, normalement sans chlorophylle (elles peuvent
contenir des chloroplastes, mais uniquement dans les racines
aériennes) et constituent, surtout dans les parties âgées de la
racine, un tissus de réserve. L’assise corticale la plus interne,
l’endoderme, entoure le cylindre central, de telle sorte qu’en coupe
transversale elle apparaît sous la forme d’un anneau. L’endoderme
peut fonctionner en tant que barrière physiologique et contrôler le
passage de l’eau et des sels qui y sont dissous, de l’écorce vers le
cylindre central. Par dépôt de subérine, les épaississements
secondaires et tertiaires rendent les parois nettement
imperméables. Les cellules de passage qui restent inchangées,
prennent le relais pour assurer ce contrôle.
Au niveau de l'écorce de l’extérieur vers l’intérieur il y a une assise
pilifère recouverte de poils absorbants, ce sont des cellules
modifiées très allongées.
L’assise subéreuse : elle se subérifi et entraine la mort des poils
absorbants d'assise situés au dessus.
Le parenchyme cortical est constitué de cellule amilifère en
disposition radial.
Endoderme : ce sont des cellules profondes du parenchyme
cortical. (Attention sur les dessins : les dessins scientifiques ne
sont pas des représentations artistiques ils doivent montrer
l'essentiel et doivent être précis)
Cylindre centrale de l'extérieur vers l'intérieur on va avoir le
péricycle : Le cylindre central renferme le xylème et le phloème
disposés en un système conducteur radial. En coupe transversale,
on peut observer la différenciation des vaisseaux. Les faisceaux du
xylème se réunissent au centre ou bien des cordons de cellules
sclérenchymateuses ou parenchymateuses persistent, de telle sorte
que le xylème reste limité aux faisceaux. Les cellules
parenchymateuses du xylème situées directement près des vaisseaux
présentent la paroi, en contact avec ces vaisseaux, des
épaississements localisés caractéristiques des cellules de transfert.
Les cordons de phloème se situent entre les rayons de xylème. Ils
sont séparés de ce dernier par des rayons de tissu parenchymateux
(rayon médullaires). La différenciation du phloème a lieu avant
celle du xylème, de telle sorte que des cordons de phloème
fonctionnels peuvent amener des substances organiques vers la
zone d’élongation, condition indispensable pour que soit assurées
aussi bien les divisions des cellules que leur élongation. A la
périphérie du cylindre central se trouve le péricycle, qui peut
comporter une ou plusieurs assises cellulaires.
Ce même cycle est situé contre l'endoderme. La moelle : constituée
de cellules parenchymateuses. Le xylème primaire est du floenne

B12 – Biologie végétale L1 Bio 2010/2011
Page 11 sur 39
primaire et surtout ils sont constitués en faisceaux. On a dans la
structure primaire une disposition alterné de xylème et de floenne.
En bref chez le gymnosperme et l’eudicots: on a environ 5
faisceaux et chez les monocots on a entre dix et vingt faisceaux.
Trouver le schéma d'un poil absorbant disposition des faisceaux
primaires dans une racine.
o Au niveau de l'endoderme :
Toujours au niveau de l'anatomie mais cette fois de la structure
secondaire : elle résulte du fonctionnement des cambiums et cette
structure secondaire existe chez les gymnosperme et les eudicots.
Dans ce cas là on a un xylème et des floennes secondaires, ils se
forment en position interne par rapport au cambium et mènent à la
formation de vaisseaux, de fibres, de parenchymes verticaux et
horizontaux.
o Suber + phelloderme qui vienne du cambium subérophellodermique : Les
tissus à l'extérieur du suber vont mourir et c'est le suber qui va remplacer
l'assise subérienne et le phelloderme va remplacer le parenchyme cortical.
o La croissance : Elle se fait en longueur, dans ce cas là la croissance se fait du
méristème latéral au méristème apical situé au dessus de la coiffe
o La croissance en épaisseur se fait grâce au cambium.

B12 – Biologie végétale L1 Bio 2010/2011
Page 12 sur 39
o Dernier point : au niveau de la ramification : elles proviennent d'une hernie du
péricyte
o La racine est la partie qui est habituellement enfoncée dans le sol. Elle nourrit
la plante et la fixe plus ou moins solidement à la terre. La croissance de la
racine est subterminale, elle s'effectue près de l'extrémité. Les racines ont un
géotropisme positif, c'est à dire que peu importe la position de la plante, la
croissance de la racine se fait vers la terre, vers le bas.
o Cet organe manque chez les rhyniales, les salviniales ainsi que chez les
lycopodiacées où les organes souterrains ont une structure semblable à celle
des organes aériens.
o Les ptéridophytes n'ont pas de racine principale, la première racine est déjà
latérale. Elle meurt rapidement et est remplacée par de nouvelles racines
latérales.
o Chaque racine est protégée au bout par une coiffe qui pénètre dans le sol. La
zone pilifère est celle qui absorbe les minéraux pour nourrir la plante. La zone
subéreuse protège les tissus conducteurs.
La structure de la racine :
o Le collet est la région comprise entre la racine et la tige, il coïncide
généralement avec le niveau du sol. La zone de ramification est juste en haut
des poils absorbants. C'est l'endroit où apparaissent les nouvelles racines
latérales.
o Les poils absorbants absorbent l'eau et les minéraux dissous dans le sol.
B12 – Biologie végétale L1 Bio 2010/2011
Page 13 sur 39
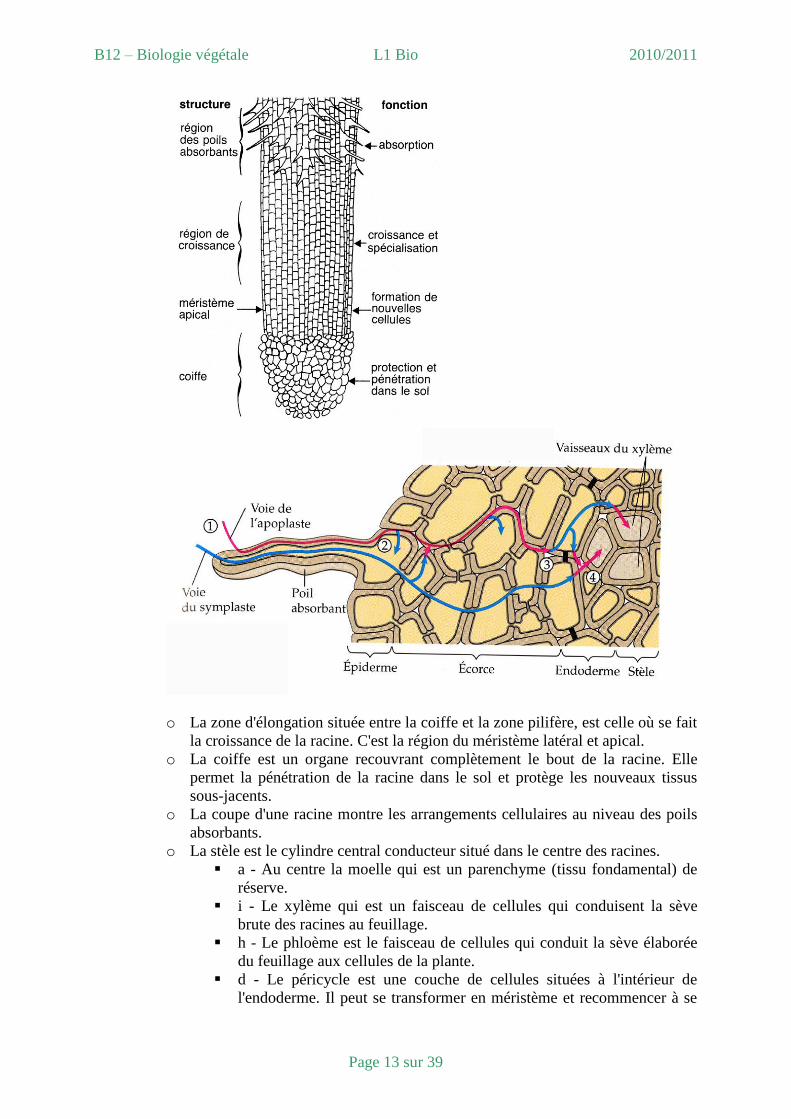
o La zone d'élongation située entre la coiffe et la zone pilifère, est celle où se fait
la croissance de la racine. C'est la région du méristème latéral et apical.
o La coiffe est un organe recouvrant complètement le bout de la racine. Elle
permet la pénétration de la racine dans le sol et protège les nouveaux tissus
sous-jacents.
o La coupe d'une racine montre les arrangements cellulaires au niveau des poils
absorbants.
o La stèle est le cylindre central conducteur situé dans le centre des racines.
a - Au centre la moelle qui est un parenchyme (tissu fondamental) de
réserve.
i - Le xylème qui est un faisceau de cellules qui conduisent la sève
brute des racines au feuillage.
h - Le phloème est le faisceau de cellules qui conduit la sève élaborée
du feuillage aux cellules de la plante.
d - Le péricycle est une couche de cellules situées à l'intérieur de
l'endoderme. Il peut se transformer en méristème et recommencer à se

B12 – Biologie végétale L1 Bio 2010/2011
Page 14 sur 39
diviser.
e - L'endoderme est une couche de cellules entre la stèle et l'écorce.
L'endoderme joue le rôle de barrière sélective qui règle le passage ces
substances provenant du sol vers les tissus conducteurs de la stèle.
Coupe transversale de racine
Coupe longitudinale de racine
o L'écorce des racines contient plusieurs rangées de cellules destinées à protéger
les tissus situés à l'intérieur de la racine.
b - L'écorce interne est formée de plusieurs couches de cellules de
protection des tissus internes.
c - L'écorce externe donne la couche extérieure qui est l'épiderme et les
poils absorbants.
g - Les poils absorbants sont des prolongements des cellules
épidermiques. Ils absorbent l'eau et les minéraux du sol et les mènent
vers les tissus conducteurs du xylème.
o Types de racines

B12 – Biologie végétale L1 Bio 2010/2011
Page 15 sur 39
Racine pivotante ; lorsque la racine primaire se développe beaucoup
plus que les racines secondaires.
Racines fasciculées lorsque la racine principale avorte et n'est pas plus
développée que les racines secondaires.
Racine pivotante Racine fasciculée
Les racines peuvent être terminales, latérales (secondaires), fasciculées
ou adventives.
Les racines peuvent aussi être un tubercule, un crampon, un suçoir.
Schéma : ramification de la racine
- la tige
Morphologie :
o C'est un axe divisé en nœuds et en entre-nœuds qui sont les points d'insertion
des feuilles.
o Les bourgeons : Terminale ou apicale, il y a aussi les bourgeons axillaires ou
latéraux situés à la base des tiges. Le bourgeon adventif apparaît lorsqu’il y a
blessure ou taille de la plante.
o Ces bourgeons évoluent en tiges feuillées ou en fleurs.
Classement :
o Consistance :
Tiges herbacées qui sont en générale verte et relativement tendre
Tige ligneuse
o Port : On parle de tige dressée ; rampantes : les fraisiers ; grimpantes; volubiles
(liseron)
o Longueur: on admet que si on est supérieur à 5 mètre on va parler d'arbre, si on
est entre 2 et 5 mètre on va parler d'arbrisseau, 1- 2 m arbustes, <1m : herbes.
Milieu : aérienne, aquatique, souterraines (tige souterraines et non des racines)
Rhizome : constitué de différentes unités (chaque unité c’est une partie de la tige
aérienne et souterraine)
Les bulbes : T plateau dans ce cas la on a une tige très courtes avec la présence de
feuilles hypertrophiées que l'on appelle les Tuniques (figure 29 : bulbes d'oignon).

B12 – Biologie végétale L1 Bio 2010/2011
Page 16 sur 39
Un autre exemple de tige souterraine : tubercules: feuilles réduites à des écailles, ex
pomme de terre etc.
Le cornu: on dans ce cas là une hypertrophie de la tige.
Au niveau de l’anatomie il y a trois régions principales :
o L'épiderme: est constitué de cuticule et de stomate
o L'écorce: même structure que la racine mais beaucoup plus réduite /
parenchyme/méats et limité par l’intérieur par l’endoderme, plus précisément
pour le parenchyme : on observe des éléments de soutien
Cellule oxalate de calcium
Cellule sécrétrice
o Le cylindre central, constitué de xylème primaire (faisceau superposé) et de
moelle (Schéma 30 dicotylédone)
Anatomie de la structure secondaire : on a du xylème secondaire et du phloème
secondaire.
o Le cambium produit également des cellules parenchymateuses
Plantes herbacées : ce cambium ne fonctionne qu’au niveau des
faisceaux primaires et l'activité de ce cambium est rythmique, ce
cambium va donc fonctionner au maximum au printemps.
Au printemps on a donc un bois riche en faisceaux et cette couche de
bois est claire et tendre, on va parler de bois de printemps.
En automne ces faisceaux diminuent de diamètre, présences de
nombreuses fibres, dans ce cas la on a un bois foncé et dur, c'est ce que
l’on appelle le bois d'automne.
o On a la présence de suber et de phelloderme qui apparaissent après les
cambiums libéroligneux. On va avoir différents cambiums qui peuvent se
former successivement. A la suite de la formation de suber les cellules externes
vont mourir et former le rhytidome. (Voir figure 31 Tige âgée de
dicotylédone).
o Croissance en longueur, elle se fait au niveau des entre-nœuds proches de
l'apex grâce à des méristèmes intercalaires. Il existe différent type
d'allongement :
Monopedial
Sympodial
Dichasial
(Voir figure 32 Type d'allongement)
La croissance en épaisseur se fait grâce au cambium.
- la feuille
C’est une structure très complexe mais en simplifiant on peut dire que c’est une expansion
latérale de la tige. Elle est presque toujours verte et c’est le lieu de la photosynthèse.
1. Morphologie.

B12 – Biologie végétale L1 Bio 2010/2011
Page 17 sur 39
Cette feuille est constituée de trois parties :
o La première c’est le limbe, c’est une lame plate à symétrie bilatérale soutenue
par des nervures. Au niveau de ce limbe il y a différent type de feuilles (voir
figure 33 différents types de feuilles). Ce limbe peut être considéré en fonction
de la disposition des feuilles. L’étude de la disposition des feuilles sur la tige
s’appelle la phyllotaxie. On parle de feuilles alternées lorsqu’il y a une feuille
par nœud, on parle de feuilles opposées lorsqu’il y a deux feuilles par nœud et
on parle de feuilles verticillées lorsqu’il y a trois ou plus feuilles par nœud.
o La deuxième partie est le pétiole, c’est l’axe foliaire sans appendice et il a
toujours une section triangulaire ou arrondie.
o La troisième partie est la base foliaire, c’est la partie arrondie de la base du
pétiole
2. Anatomie.
Structure primaire du limbe qui est constituée d’un épiderme (voir figure 34. épiderme du
limbe), du xylème primaire et du phloème primaire. Ils sont disposés en faisceaux (voir figure
35. feuille de topinambour (dicotylédone)) et sont superposés.
3. Formation.
Elle se fait à partir du méristème apical de la tige (voir figure 36. méristème apical de la tige
et figure 37. formation des feuilles). Du méristème apical des cellules se spécialisent, donnent
un deuxième type de méristème, le méristème terminal qui va croitre et former la feuille.
C’est une zone latérale de l’anneau qui se soulève et forme la feuille.
De ce même méristème apical on a un méristème marginal qui va assurer la croissance de la
feuille. Accroissement longitudinal grâce au méristème terminal (voir tableau E comparatif
des caractères anatomiques de la plantes).

B12 – Biologie végétale L1 Bio 2010/2011
Page 18 sur 39
Anatomie de l’appareil reproducteur :
- organisation de la fleur isolée
Les pièces florales sont disposées selon quatre cycles qu’on appelle verticilles. De l’extérieur
vers l’intérieur on a deux gros types de structures :
Le périanthe : deux verticilles, les pièces de ces verticilles sont stériles.
On a donc le calice qui est formé par l’ensemble des sépales, ce
sont des pièces verdâtres d’aspect foliacé avec du parenchyme
chlorophyllien.
On a ensuite la corolle qui est l’ensemble des pétales. Ces pièces
sont sans chlorophylle, elles sont riches en pigments colorés et ces
pigments on les retrouve essentiellement dans les vacuoles. A la
base de chaque pétale on a une glande sécrétrice qui va sécrétée une
substance sucrée qui est le nectar.
Les organes reproducteurs : deux verticilles, de l’extérieur vers l’intérieur.
On parle de l’androcée qui est formé par l’ensemble des étamines
qui sont constituées à leur tour de filet (partie fine et allongée) et de
l’anthère qui est la partie supérieur et élargie. En bref c’est la
structure mâle de l’appareil reproducteur.
Gynécée ou encore pistil qui est formé par l’ensemble des carpelles
qui sont de petites structures vertes et à leur tour chaque carpelle
est composé d’une partie renflée qu’on nomme ovaire, d’une partie
supérieur à l’ovaire qu’on appelle le style et au dessus du style le
stigmate avec des papilles.
(Voir figure 1. coupe longitudinale d’une fleur de renoncule)
(Voir figure 2. coupe longitudinale d’un carpelle)
(Voir figure 4. coupe transversale d’ovaire)
- variation de l’organisation florale
Principales variations on les observe à différents niveau :
Premièrement au niveau de la symétrie de la fleur :
Fleur régulière (qu’on appelle fleur actinomorphe). Dans ce cas là
les pièces du périanthe sont disposées autours d’un axe et sont
identiques. Il existe deux cas de figures : si les pétales sont libres
(non soudées entre elles) on parle de fleurs dialypétales (exemple
les Rosacées et les Renoncules). Si les pétales sont partiellement ou
totalement soudées on parle de fleurs gamopétales (exemple les
Primulacées).
Fleur irrégulière (on parle de fleur zygomorphe). Dans ce cas là les
pièces du périanthe ne sont pas toutes identiques entre elles et elles
sont disposées de part et d’autre du plan de symétrie. Si les pétales
sont libres on parle de dialypétales (exemple les Orchidacées) et si
les pétales sont partiellement ou totalement unie on parle de
gamopétales.
Deuxièmement au niveau du nombre d’étamines. Selon les espèces on
observe de une à cinquante étamines. Elles peuvent être libre entre elles
(cas des Rosacées) ou bien soudées par leur filet (c’est le cas des

B12 – Biologie végétale L1 Bio 2010/2011
Page 19 sur 39
Papilionacées) ou encore soudées par leur anthère (c’est le cas des
Composées).
Troisièmement au niveau des carpelles. Soit on a des carpelles
indépendant, dans ce cas là on a un nombre élevé de carpelles, soit les
carpelles sont soudés entre eux.
Soit les feuilles carpellaires sont fermées et réunies entre elles et
dans ce cas on a autant de loges que de carpelles.
Soit les feuilles carpellaires ne sont pas fermées mais réunies entre
elles, dans ce cas là on a une seule loge carpellaire sans cloison.
Quatrièmement au niveau de la placentation. C’est la disposition des
placentas donc des ovules dans l’ovaire et cette disposition correspond au
carpelle. Il existe plusieurs situations :
Placentation pariétale, lorsque les ovaires sont formées de deux ou
plusieurs carpelles ouverts et soudés.
Placentation axile, lorsque les ovaires sont formés par plusieurs
carpelles fermés et soudés.
Placentation centrale, lorsque les ovules sont directement fixées sur
le réceptacle floral.
(Voir figure 5. placentation axile)
(Voir figure 6. placentation pariétale)
(Voir figure 7. placentation centrale)
Cinquièmement au niveau de l’ovaire et du réceptacle floral
o Fleur ovaire supère c’est lorsque le réceptacle est bombé et l’ovaire
est situé au dessus du niveau d’insertion des autres pièces florales
(exemple orangé) (voir figure 8. ovaire supère).
o Fleur à ovaire infère c’est lorsque le réceptacle est en forme de
coupe et le pistil est situé au fond (voir figures 9 et 10 ovaire infère
libre et ovaire infère adhérent).
Sixièmement au niveau des fleurs à périanthe incomplet
o Il peut être réduit au simple sépale, on parle de fleur sans pétale soit
apétale
o Lorsque le périanthe est rudimentaire (Graminées)
- groupement de fleurs
Dans certain cas la fleur est unie mais en général elles sont regroupées on parle alors
d’inflorescence.
Toute d’abord au niveau des inflorescences on a deux types fondamentaux
d’inflorescence.
Le premier prend le nom de grappe lorsque le méristème est situé à l’extrémité, il y a donc
sur le coté la formation périodique de bractées d’où naissent les fleurs. Croissance avec un
seul axe soit un seul méristème.
Deuxième type on parle de cyme c’est lorsqu’il y a plusieurs méristèmes à l’extrémité de
plusieurs axes.
On peu également parler d’inflorescence simple si on est au niveau de type grappe on peu
avoir plusieurs catégories de figure : grappe simple, épi, corymbe, ombelle et capitule. Ce
sont toutes des inflorescences simples.

B12 – Biologie végétale L1 Bio 2010/2011
Page 20 sur 39
On va parler de type cyme bipare lorsqu’elles poussent à droite et à gauche, on peut aussi
parler de cyme unipare hélicoïde ou unipare scorpioïde.
On peut aussi trouver des inflorescences composées ce sont des groupements de plusieurs
inflorescences.
- répartition des sexes
Hermaphroditisme : lorsque la fleur contient à la fois les organes mâles (étamines) et les
organes femelles (carpelles) c’est la plupart des cas.
On a quelques exceptions :
Monœcies : espèces monoïques, chaque pied va porter deux types de fleurs unisexuées
(fleurs mâles et à coté fleurs femelles, exemple : noisetier et maïs).
Dioécie : ce sont des espèces dioïques et dans ce cas là on a des pieds mâles et sur une
autre plantes des pieds femelles (exemple le saule).
- structures des organes reproducteurs
Etamines, pollen :
Etamines : l’anthère est constituée de deux loges avec des grains de pollen à l’intérieur
(voir figure 5 – étamine et figure 21)
Pollen : structure sphérique et jaune d’une taille qui varie entre 200 et 300 µm et chaque
grain de pollen contient deux cellules, une première de grande taille qu’on appelle la
cellule végétative (elle possède toutes les substances de réserves) et une deuxième cellule
de petite taille qui est la cellule génératrice, cette cellule va donner plus tard les deux
gamètes mâles ou spermatozoïdes (voir figure 22 et 8 grain de pollen bicellulaire).
Formation du pollen, plusieurs étapes chronologiques :
A chacun des 4 angles d’une jeune anthère, les cellules sous-épidermiques se
différencient en grandes cellules.
Chacune de ces cellules se divise en deux cellules (par mitose) : une cellule
externe et une cellule interne.

B12 – Biologie végétale L1 Bio 2010/2011
Page 21 sur 39
La cellule externe se divise plusieurs fois et donne les différentes couches de la
paroi de l’anthère.
La cellule interne se divise également et donne à chaque angle de l’anthère un
massif de cellules : ce sont les cellules-mères du pollen.
Chaque cellule-mère (2n chromosomes) subit une méiose et donne quatre petites
cellules (à n chromosome soient haploïdes).
Ces petites cellules sont appelées des microspores et restent dans la cellule-mère.
Chaque microspore va élaborer une double paroi (exine et intine) et se divise
inégalement dans la paroi.
Ce qui donne une petite cellule (cellule génératrice) dans une grande cellule
(cellule végétative). C’est un grain de pollen.
Les grains de pollen se déshydratent.
La paroi qui unissait les quatre microspores disparait.
Résorption des couches cellulaires sous l’assise mécanique. Les grains de pollen
sont prêts à être disséminés.
(Voir figure 6 Formation du pollen de Lis).
(Voir schéma formation du pollen)
Formation de l’ovule :
Quand on parle de funicule, c’est un cordon fin qui fixe l’ovule au placenta. Sur cet
ovule on a également le micropyle, ce sont deux téguments qui présentent une étroite
couverture. On a également le nucelle qui est le massif parenchymateux interne. La région
qui prend le nom de chalaze est lorsqu’on a un faisceau libéro ligneux qui se termine dans
le nucelle. Enfin dans cet ovule il y a le sac embryonnaire qui est constitué de sept cellules
dont la centrale avec deux noyaux. Orthotrope lorsque l’ouverture du micropyle est vers le
haut.
Sac embryonnaire contient sept cellules :
Au pôle micropylaire, deux cellules synergides et une cellule oosphère. Il y a une
cellule centrale qui possède deux noyaux qu’on appelle noyaux polaires. Sur le pôle
opposé on a trois cellules antipodes.
(Voir figure 14 et schéma de la figure 27 et 28).

B12 – Biologie végétale L1 Bio 2010/2011
Page 22 sur 39
Biologie de la reproduction :
- Pollinisation
Définition : c’est le transport du pollen émis par une fleur sur le stigmate de la même fleur
ou d’une autre fleur.
a. Agents du pollen
Le vent : les fleurs sont dites anémophiles, les grains de pollen sont
abondants, très léger et le stigmate présente une grande surface réceptrice
importante, environs 10% (exemple : les Graminées).
Insectes : on parle de fleurs entomophiles, 90% des végétaux actuels.
L’insecte va secouer l’étamine et le pollen tombe sur le stigmate de la même
fleur ou alors le pollen va s’accrocher aux insectes qui vont aller sur d’autres
fleurs. Les fleurs qui sont pollinisées par les insectes ont des caractéristiques
communes : le périanthe est vivement coloré et dégage une forte odeur. Les
grains de pollen sont de grande taille et ils présentent des systèmes
d’accrochages, l’exile présente des crochets. En général, les glandes qui
produisent le nectar, les nectaires, sont situées au fond des fleurs ce qui oblige
l’insecte à plonger à l’intérieur de la fleur et par conséquent favoriser la
pollinisation. On parle de coévolution entre l’insecte et les fleurs (exemple : les
abeilles dans les vergers).
b. Deux modes
La pollinisation directe : autogamie. C’est lorsque le pollen des fleurs
pollinisent le stigmate de la même fleur ou d’une fleur de même pied. En
général ce sont les fleurs cultivées (exemple : les Graminées, les haricots, les
poids, les violettes).
La pollinisation indirecte : allogamie. C’est lorsque le pollen d’une fleur
pollinise une fleur sur un autre pied, c’est le cas le plus fréquent, cas de toutes
les plantes sauvages, cela permet de maintenir l’hétérogénéité du génotype.
- germination du pollen
Elle se fait en quatre étapes :
1 – Germination du pollen : le stigmate va absorber de l’eau de l’extérieur, cette eau va
passer à l’intérieur du stigmate et le grain de pollen va gonfler. L’intine va se détendre et
le cytoplasme va sortir par un pore de l’exine. C’est le début de la formation du tube
pollinique. Cette hernie (ce tube) annonce le début de la germination.

B12 – Biologie végétale L1 Bio 2010/2011
Page 23 sur 39
2 – Croissance du tube pollinique : On a le noyau de la cellule végétative qui est dans le
tube pollinique et la cellule génératrice va passer dans ce tube pollinique. La croissance
est rapide et se fait en plusieurs millimètres par heure et dure entre cinq et huit heures, il
prend plusieurs centimètres.
3 – Le tube pollinique va rentrer dans le pistil, il va s’insinuer entre les cellules du
stigmate et ensuite dans l’ovaire. Et il arrive au contact de l’ovule par micropyle (voir
schéma trajet du tube pollinique dans le pistil).
4 – Formation des spermatozoïdes mais c’est au cours de la traversé du style que la cellule
génératrice se divise en deux spermatozoïdes.
- Fécondation
Au contact de l’ovule, le tube pollinique aborde la partie supérieure du sac embryonnaire.
Il pénètre dans une synergide et déverse ses deux spermatozoïdes à l’intérieur. Le noyau
végétatif dégénère.
Après la traversée de la synergide, les spermatozoïdes sont réduits à leur noyau.
L’un pénètre dans l’oosphère et s’unit à son noyau : un zygote principal diploïde est
constitué.
L’autre s’unit aux deux noyaux polaires : un deuxième zygote accessoire est formé mais il
est triploïde.
C’est une double fécondation, caractéristique des angiospermes.
(Voir schéma de la fécondation).
- reproduction sans fécondation
1. Formation d’un embryon haploïde :
Dans ce cas, l’embryon peut se former à partir de l’oosphère ou d’une autre cellule
haploïde du sac embryonnaire (cas très rare et peu viable).
2. Formation d’un embryon diploïde :
Embryon issu à partir de l’oosphère à 2n chromosome qui n’avait pas subit de réduction
chromatique.

B12 – Biologie végétale L1 Bio 2010/2011
Page 24 sur 39
Embryon issu d’une autre cellule diploïde (2n).
- formation de la graine
Les deux zygotes issus de la double fécondation se développent mais avec deux destinés
différentes.
Le premier zygote principal, c'est-à-dire celui qui provient de l’oosphère va donner
l’embryon (1) de la graine.
Le deuxième zygote ou zygote accessoire est triploïde et va donner un tissu qui est
l’albumen (2).
(1) L’embryon :
Il va se diviser et donner deux cellules :
a. La cellule basale, qui va continuer de se diviser en donnant une file de cellule
provenant d’un suspenseur. Il va donner la radicale qui est l’ébauche de la racine.
b. La cellule terminale, qui va donner les cotylédons (un ou deux) qui vont par la suite
donner la plantule.
(Voir schéma développement de l’embryon)
(2) L’albumen :
L’albumen est un tissu nourricier pour l’embryon et on observe un développement
différent entre l’albumen et l’embryon.
Dans le cadre de ce développement on a une formation des téguments et de substances de
réserve tels que des protéines, des lipides et de l’amidon.
(Voir figure 26 formation de la graine et figure 36 différents types de graines).
- Formation du fruit
1. Croissance du fruit : après la fécondation, l’ovaire se développe en un fruit qui
contient une ou plusieurs graines. En bref, la croissance du fruit est en fait la
croissance de l’ovaire et qui est du à une hormone de croissance qu’on nomme
l’auxine (moteur de base, en vrai il y en a bien plus).
2. Principaux types de fruits :
a. Fruits simples : à partir de l’ovaire unique d’une fleur dont chaque fleur donne
un seul fruit (exemple : groseille, tomate, cerise).
b. Fruits multiples : ils sont issus de fleurs dont le gynécée est constitué de
plusieurs carpelles indépendantes donc chaque fleur va donner plusieurs fruits
(exemple : le mûre).
c. Fruits composés : ils sont issus du gynécée d’une fleur auxquels ce sont
associés d’autre partie de la fleur (exemple : les fraises, les pommes).
d. Fruits charnus (exemple tomate et cerise).
e. Fruits secs indéhiscents (exemple les Akènes).
f. Fruits secs déhiscents (ce sont tous les autres).
BILAN : Voir figure 44 cycle de développement d’une angiosperme à fleurs bisexuées et
figure 23 de la fleur au fruit.
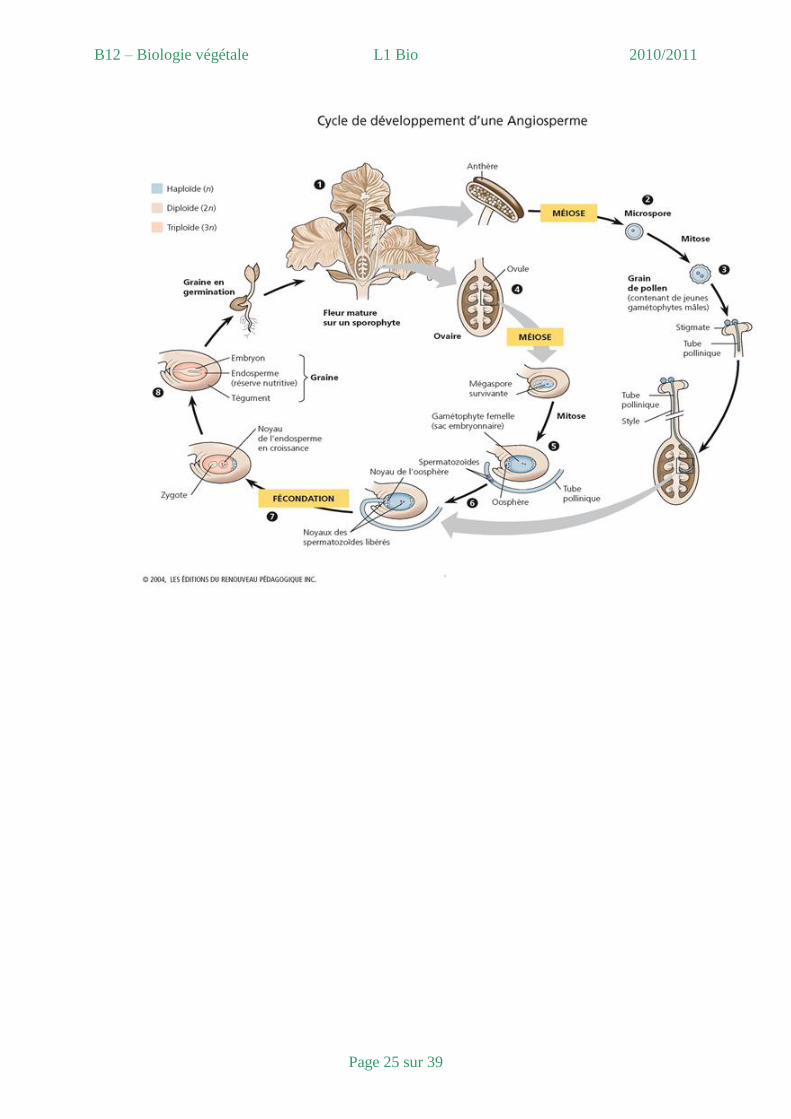
B12 – Biologie végétale L1 Bio 2010/2011
Page 25 sur 39
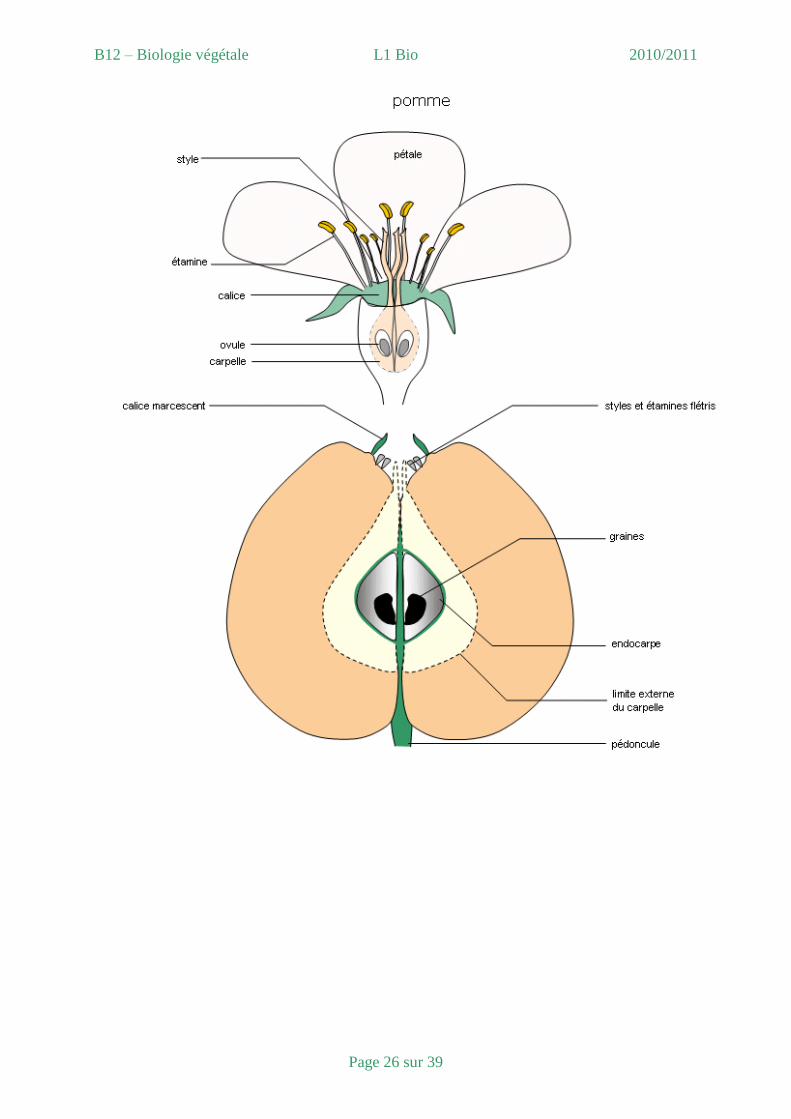
B12 – Biologie végétale L1 Bio 2010/2011
Page 26 sur 39

B12 – Biologie végétale L1 Bio 2010/2011
Page 27 sur 39
II. Systématique et grands groupes végétaux
Il y a plusieurs divisions du vivant en fonction des auteurs :
(1) schéma classique qui divise le monde en deux règnes : animaux et végétaux. Dans ce
cas les bactéries et les champignons faisaient partie des végétaux. Cependant on
pouvait se demander : que fait-on des virus, des procaryotes et des eucaryotes ?
(2) Autre possibilité est de diviser le monde en cinq règnes : celui des bactéries, des
protistes, des champignons, des végétaux et enfin des animaux (un classement fait par
Whittaber en 1969 et que nous approfondirons en biologie animale). Procaryotes (qui
englobent les eubactéries et les archéobactéries) et les eucaryotes sont des super-
règnes.
Généralité sur les grands groupes végétaux :
- les thallophytes (algues, champignons, lichens) :
Ce sont les algues, les champignons et les lichens et ils sont unicellulaires ou pluricellulaires.
Ce sont toujours des plantes aquatiques ou bien vivant dans des zones humides. La
différenciation cellulaire est peu poussée et on observe une très grande diversité dans les
cycles de développement. Il y a également une très grande diversité dans les cycles de la
reproduction sexuée.
Reproduction :
Gamétophyte : c’est le stade haploïde à n chromosome, soit les gamètes.
Sporophyte : c’est un stade diploïde à 2n chromosome et il débute par la formation
de l’œuf.
Ils sont aussi caractérisés par une variabilité de l’alternance des générations :
Chez les algues et les champignons inférieurs (moisissures) c’est le gamétophyte
qui est dominant alors que la phase à 2n chromosome est très limité.
Chez les laminaires et les champignons supérieurs, on a une alternance de
génération équivalente entre gamétophyte et sporophyte.
Chez le fucus (fucales) c’est le sporophyte qui est dominant alors que les gamètes
sont très limitées.
Forme variable des gamètes :
Lorsque les gamètes mâles ont la même forme que les gamètes femelles on parle
d’isogamie.
Lorsque les gamètes mâles n’ont pas la même forme que les gamètes femelles on
parle d’hétérogamies.
Lorsque le gamète femelle est immobile et que ce sont les gamètes mâles qui se
déplacent on parle de oogamie.
On parle de cystogamie lorsque les gamètes restent à l’intérieur du thallophyte et
dans les cellules sexuelles qui les ont créées. Dans ce cas là il y a la formation d’un
tube de conjugaison entre ces structures et il y a une fusion et une fécondation par
la suite.
- les embryophytes (plantes terrestres : bryophytes, ptéridophytes, gymnospermes,
angiospermes)
Anthéridies et archégones :
Les gamètes et les méiospores ne sont plus formés dans les cellules au contacte de l’eau,
ils sont formés dans les cellules entourées d’une enveloppe de cellule protectrice : les

B12 – Biologie végétale L1 Bio 2010/2011
Page 28 sur 39
gamétanges (sporanges). Dans ces gamétanges la cellule à l’origine des gamètes donnent
d’abord naissance à des cellules d’enveloppe qui protègent les gamètes du milieu externe.
Là où les cellules mères produisent les gamètes. Le gamétange mâle est l’anthéridie et le
gamétange femelle est l’archégone.
(Voir schéma embryon et cycle de développement).
Le zygote à 2n chromosomes va donner naissance à quatre méiospores (tétraspores)
qu’après un certain temps, la méiose est ainsi retardée, ce retard permet au zygote de se
développer en un ensemble tissulaire à 2n chromosomes.
Cet ensemble tissulaire est l’embryon, d’où le nom de taxon des embryophytes. Au cours
de l’évolution le retard va devenir de plus en plus grand.
On observe une alternance d’une génération diploïde (le sporophyte débute par la
formation de l’embryon et se termine par la production des méiospores) avec une
génération haploïde (le gamétophyte assure la formation des gamètes) et devient alors une
caractéristique du cycle de développement des végétaux terrestres. Le jeune embryon à 2n
chromosomes se développe en parasite sur le gamétophyte qui est à n chromosomes.
(Voir schéma des embranchements).

B12 – Biologie végétale L1 Bio 2010/2011
Page 29 sur 39
Les algues
Protocaryotes (cyanobactéries) et eucaryotes.
1. Embranchement Cyanophycota ou Schizophytes (Protocaryotes), il n’y a qu’une seule
classe : les cyanobactéries.
A. Les cyanobactéries.
a. Caractéristique
Absence d’un vrai noyau, pas de reproduction sexuée et la multiplication se fait par division
végétative et production de spores : les alkinètes. Ces cyanobactéries sont les premiers être
vivants apparus sur terre, elles sont très cosmopolites, on les trouve depuis les eaux thermales
jusque dans les glaces de l’Arctique. Elle forme donc des pronuages dans les lacs et en mer on
les appelle les fleurs d’eau ou de bloom. Elles sont responsables du processus
d’entrophisation des eaux et sont capables de produire des toxines mortelles.
2. Embranchement chlorophytes ou algues vertes, il existe quatre classes : les
Euchlorophycées, les ulothricophycées, les zygophycées et les charophycées.
A. Caractéristiques générales
Présence d’un vrai noyau, plusieurs types de pigments à l’intérieur des plastes. Les plus
abondants sont les plastes d’un vert pur. Il y a présence d’amidon dans ces plastes.
a. Euchlorophycées
Elles sont sous forme solitaire ou coloniale mais ne sont pas ni filamenteuses ni thalloïdes.
b. Ulothricophycées
Elles sont filamenteuses et thalloïdes.
c. Zygophycées
Elles sont pluricellulaires ou filamenteuse.
d. Charophycées
Avec un seul ordre : les charales. Elles sont apparues au Dévonien et elles poussent
essentiellement dans les eaux ?
Elles sont pluricellulaires et ressembles à des plantes supérieures comme les prêles.
3. Embranchement Chromophytes ou algues brunes, il existe cinq classes :
Euglénophycées, les Chrysophycées, les Xanthophycées, les Bacillariophycées et les
Phéophycées (ou grandes algues brunes) dans trois sous-embranchement.
A. Sous embranchement des Euglénophytes : une seule classe les Euglénophycées
B. Sous embranchement des Chrysophytes ou Chromophytes
a. Classe des Chrysophycées
b. Classe des Xanthophycées
c. Classe des Bacillariophycées ou encore Diatomées
Ce sont les algues essentielles dans les lacs et les océans et sont à la base de tous les réseaux
trophiques. Elles sont constituées de deux coques : épivalve et hypovalve qui s’emboitent
avec un ruban autours qui constitue la ceinture connective qui les maintiennent. Elles sont

B12 – Biologie végétale L1 Bio 2010/2011
Page 30 sur 39
constituées essentiellement de silice. Elles sont capables de former des spores de résistances :
les auxospores, elles constituent l’essentiel du phytoplancton marin et assurent l’essentiel de
toute la photosynthèse des océans. Elles concentrent le DMS-diméthylsulfide qui va être à
l’origine des noyaux de condensations qui vont pousser dans l’atmosphère et qui vont être à
l’origine de la formation des nuages.
Il existe deux sous classes :
- Les centrophycidées (symétrie centrale)
- Les pennatophycées (symétrie axiale)
C. Sous embranchement des Pyrrophytes
d. Classe cryptophycées
Elles sont unicellulaires avec des flagelles.
e. Classe Dynophycées
C’est le deuxième grand groupe avec les bacillariophycées qui forme le plancton marin et ?
Elles sont indicatrices d’un certain processus d’Eutrophisation et elles sont à l’origine d’une
production importante de toxines marines. Par exemple les Dynophysis ou encore les
Ceratium qui intervient dans le processus d’eutrophisation.
f. Classe Phéophycées
Généralité
Grandes algues brunes, pluricellulaire de structure complexe, la plupart sont marines et
abondantes sur les côtes océaniques des régions tempérées et froides. La couleur est très
variée, elle passe du jaune au vert grâce à des plastes riches en chlorophylles a et c et en bêta-
carotène. Les parois des cellules sont riches en cellulose et en polysaccharides, en alginate
(gélifiant) et en différentes géloses. Ces gélifiants permettent à la plante de gonfler dans l’eau,
on obtient ainsi une texture visqueuse qui protège les thalles de la plante dans les vagues.
Evolution continue et parallèle entre trois caractères : 1er
organisation thalle, 2nd
le mode de
fécondation, 3ème
différents équilibres dans les cycles. Ils sont riches en iode et avec tous les
oligo-minéraux que l’on peut trouver et surtout ils sont sans racines, sans tiges et sans feuilles.
Un thalle qui est composé d’un crampon qui est analogue aux les racines, d’un stipe analogue
à la tige et de frondes qui sont analogues aux feuilles. On a la même structure pour les algues
rouges.
Souvent la présence de vésicules aérifères qui vont maintenir les frondes en surface.
On a environs une douzaine d’ordres pour environs 12 000 espèces.
Systématique
o Ordre des Ectocarpales
Algues filamenteuses dont le cycle ressemble à celui de
Chadophora.
Formation direct d’un nouvel individu diploïde à partir de
croisement mâle et femelles.
o Ordre des Cutlériales
Thalles en lames (laitue de mer)
Cycle haplophasique et diplophasique
Parfois individus de l’haplophase différents des individus de
la diplophase.
Gamètes anisogamiques (mâles petits, femelles gros).
o Ordre des Dictyotales
Comme l’ordre précédent

B12 – Biologie végétale L1 Bio 2010/2011
Page 31 sur 39
Gamètes oogamiques (mâles mobiles, femelles immobiles)
Exemple : Padina pavonia
o Ordre des Laminariales
Ces sont des algues de grande voir très grande taille
Le thalle est en forme de ruban
Exemple : Goëmon, Kombu, Macrocystis (5m de long),
Leissonia (10m de long), Nereocystis (40m de long).
Reproduction : ce sont des cycles complexes avec une
alternance de génération. L’individu diploïde c’est le
sporophyte (2n) et il va produire des cellules reproductrices
qui sont appelées spores. L’individu haploïde est le
gamétophyte (n) et il va produire des gamètes.
On appelle ses algues des hétérothalliques, souvent les
gamètes mâles et femelles sont produits par des individus
différents.
(voir cycle des algues)
o Ordre des Fucales
Les organismes de ces ordres là on les même caractéristique que les laminaires mais avec une
structure plus complexe. Exemple : Fucus, Sargassum.
4. Embranchement Rodhophytes ou Rodobiontes ou algues rouges, une seule classe : les
Rhodophycées.
A. Classes Rhodophycées
Pluricellulaires en majorité marine mais quelques espèces en eau douce. Elles ont un pigment
rouge en commun qui est la phycoerythrine. Il y a de nombreux pigments accessoires qui
donnent des couleurs noires en eaux profondes, rouge vif à des profondeurs moyennes et une
tendance vers de vers dans les eaux en surfaces. Les spores et les gamètes ne sont pas
flagellés. On a un amidon modifié qui ressemble au glycogène et ce sont des algues très riches
en substances gélifiantes, exemple : Agar-agar, carréginine, porphyra.
Les cycles présentent des alternances de générations sporophytes, gamétophytes comme chez
les algues brunes.

B12 – Biologie végétale L1 Bio 2010/2011
Page 32 sur 39
Les champignons
Le carpophore est la partie non-souterraine qui s’organise en pied et chapeau. Sous le chapeau
on observe soit des lamelles soit des tubes. Ce sont les organes de la reproduction.
On les appelle les mycètes ou les Fongi, ils sont sans chlorophylles, ni chloroplastes et sans
amyloplastes. Ils vivent en saprophytes c'est-à-dire au dépend de la matière organique
(parasite). Ils se développent également dans les animaux ou dans les végétaux. Le pouvoir de
synthèse est important car ils utilisent l’azote des composés minéraux.
Appareil végétatif : mycélium qui est composé de filaments très ramifiés, on parle d’hyphes.
Reproduction sexuée et multiplication asexuée avec des spores qui se forment par voie
végétative. Plusieurs champignons sont producteurs d’antibiotiques.
Définition : dans ce grand règne on a les champignons et les moisissures, ils décomposent la
matière organique, c’est un règne à part entière, ils sont toujours eucaryotes et souvent
pluricellulaire.
Caractéristique :
Nutrition : ils sont hétérotrophes, l’alimentation se fait par absorption et non pas par ingestion
comme les autres hétérotrophes. Lorsqu’on absorbe des petites molécules organiques du
milieu et la digestion de la nourriture se fait à l’extérieur du corps par hydrolyse grâce à des
enzymes, on passe de molécules complexes à des molécules plus simples.
Saprophytes lorsqu’ils absorbent des nutriments de la matière organique non vivant.
Parasites lorsqu’ils absorbent des nutriments à partir de cellules vivantes ou hôte.
Symbiontes mutualistes c’est une coévolution avec une autre plantes, ils prennent les
nutriments de cette plantes mais favorisent l’absorption des sels minéraux à cette même
plantes.
Habitats : ils sont tous terrestres, mais la plupart nécessite de la présence d’eau voir la
possibilité de vivre dans les milieux aquatiques.
Symbiose avec les algues : le lichen.
Structure : éléments de bases que l’on appelle des hyphes. En bref ce sont plusieurs cellules
qui présentent une membrane plasmique, ces hyphes se mettent en réseaux, ces réseaux sont
des filaments ramifiés que l’on appelle mycélium.
Ces hyphes sont divisés en cellules par des cloisons et ses cloisons peuvent avoir des
ouvertures, des pores, qui permettent la circulation des ribosomes, mitochondries et noyaux.
La paroi cellulaire est différente de celle des végétaux et cette paroi est riche en chitine.
Lorsque les hyphes sont sans cloisons on parle de cénocytes ou de syphons.
Quand on a 10cm3 on a 1km d’hyphes.
Croissances et reproduction : croissance très rapide, d’abord en longueur puis en épaisseur, on
a la libération de spores et ces spores peuvent être produite soit de manière sexué soit de
manière asexué. La colonisation de ces spores se fait par différentes manières, par exemple
par le vent.
Définitions :
Syngamie : lorsque c’est l’union de deux cellules sexuelles qui proviennent de deux
organismes différents. Elle se déroule en deux étapes : la 1ère
étape la plasmogamie avec
fusion des cytoplasmes, la 2ème
étape qui est la caryogamie avec la fusion des noyaux.
Cytoplasmes fusionnée mais pas les noyaux on parle de dicaryons. Ce stade peut passer
plusieurs mois voir plusieurs années avant de passer à la caryogamie. Puis après fusion il est
diploïde.
Systématique :
1er
critère : les structures participant à la plasmogamie.
2ème
critère : la durée du stade dicaryotique.

B12 – Biologie végétale L1 Bio 2010/2011
Page 33 sur 39
3ème
critère : Le site, l’endroit où se déroule la caryogamie.
4ème
critère : Nom de chaque embranchement fait référence aux cellules sexuelles dans
lesquelles se produit la caryogamie.
1. Embranchement Zygomycète.
600 espèces (toutes des moisissures, toutes terrestre sur sol,
2. Embranchement Ascomycète.
3. Embranchement Basidiomycète.
4. Embranchement Bryophytes.

B12 – Biologie végétale L1 Bio 2010/2011
Page 34 sur 39
Les lichens
Ne sont pas un taxon de la systématique. Au niveau de leur développement c’est une
symbiose très précise entre une algue et un champignon (photobionte et mycobionte) l’algue
assure l’alimentation carbonée par ces pigments assimilateurs et le champignon par le
feutrage de ses filaments assure un support et évite la déshydratation.
Les embryophytes
1. Généralités, l’habitat terrestre se traduit par :
Des rameaux dressés : cet habitat terrestre produit des structures rigides peu
ramifiées et feuillées qui vont favoriser la capture de la lumière et des échanges
gazeux. Tiges, feuilles, racines avec un appareil vasculaire qui permet la
circulation des liquides.
De nouvelles molécules protectrices : Pour éviter une rapide dessiccation les
épidermes vont se recouvrir d’une cuticule riche en cutines. Ce sont des molécules
hydrophobes en sporopollénines qui vont se mettre dans la paroi des cellules et
d’anthocyanes qui vont s’insérer dans les épidermes et protéger la plante contre les
ultra-violets.
De nouveaux organes de l’appareil reproducteur et cycle de développement
Les trachéophytes :
Ce sont les plantes vasculaires.
Généralité : terme qui provient de Trakheia qui veut dire conduit raboteux en grecque,
apparition environs 400 million d’année, la phase sporophyte est dominante et ce
sporophyte représente la plante feuillée qui est indépendant du gamétophyte.
C’est le sporophyte qui va porter à maturité les sporanges, ce sporophyte possède un
appareil conducteur et cet appareil conducteur assure le transport des liquides ou des sèves
(brute et élaborée). On a les vaisseaux du xylème qui sont réunis en faisceaux vasculaires.
Dans ce cas là ils assurent la circulation des parties souterraines vers le haut de la plante,
c’est la sève brute.
Phloème réunis en faisceaux et assurent la production de la sève élaborée des feuilles vers
toutes les autres régions de la plantes. Tous ces vaisseaux vont présentés des
épaississements qui sont du à la présence de lignine, c’est donc une étape fondamentale de
l’évolution.
Conséquence de ces appareils conducteurs :
On a la présence de racines. Pour les groupes précédents, c’était de simples fil de cellules
ou des rhizoïdes. Ces trachéophytes se développent dans des conditions, dans ce cas là ces
trachéophytes développent une symbiose avec des champignons et on a donc une
association racine filament mycéliens qu’on appelle endomycorhize.
On a la présence de feuille proprement dite. Ce sont des expansions aplaties avec un
appareil conducteur. Sur la face inférieure on a la présence de stomates. Et ces stomates
favorisent l’évaporation de l’eau qui assure par différence de pression la montée de la sève
brute.
Organisation :
Elle est rendue possible par des zones de croissances spécialisées qui sont des méristèmes.
On a des méristèmes primaires ou apicaux, qui sont situés aux extrémités des racines et
des tiges et assurent la croissance en longueur. Méristèmes secondaires ce sont des
manchons circulaires qui assurent la croissance en épaisseur des arbres et des arbustes. On
a le cambium qui est une zone génératrice libéro-ligneuse à l’origine du bois, méristème

B12 – Biologie végétale L1 Bio 2010/2011
Page 35 sur 39
secondaire, et à l’origine du liber qui est le phloème secondaire. Et on a également du
phellogène qui donne naissance au liège, avec des propriétés protectrice et isotherme (voir
figure 10 Méristème).
- Embranchement les ptéridophytes (fougères, lycopodes, sélaginelles, prêles)
Il était appelé également les cryptogames vasculaires et on a donc les fougères et un autre
grand groupe qu’on appelle Alliées qui comprennent les lycopodes, les sélaginelles et les
prêles.
Caractère généraux :
Ce sont généralement de petites plantes, elles ont une tige souterraine horizontale qui
prend le nom de rhizome. Ils vivent en général dans des milieux ombragés et humides qui
implique la présence nécessaire de l’eau. On les trouve des régions tempérés jusque dans
les forêts tropicales. Ce qui les rallie à cet embranchement c’est leur cycle de
développement. Présence encore plus importante de lignine elles ont donc une forme
herbacée, c’est ce qui leur permet de mieux capter la lumière, facteur déterminant dans
leur croissance.
Caractères fondamentaux de ce cycle de développement :
1. A l’inverse des mousses, le sporophytes est très développé, et correspond à la plante
feuillée. Elles sont les première plantes terrestres à avoir tout le cycle de
développement constitué par des organes à 2n chromosomes. Le gamétophyte lui est a
n chromosomes est réduit à un petit organe rudimentaire et transitoire qui prend le
nom de prothalle. C’est une conquête fondamentale de l’évolution, par la suite les
gymnospermes et angiospermes, perfectionneront cette alternance de génération ce qui
leur permettra d’atteindre des tailles importantes et une forte diversité. Ils sont les
premiers végétaux avec des racines des vaisseaux et des stomates. La forme est encore
archaïque par rapport aux végétaux supérieurs et la principale évolution c’est qu’elles
peuvent capter l’eau à une certaine profondeur dans le sol ce que les bryophytes ne
peuvent pas faire.
2. Les ptéridophytes comme les bryophytes conservent encore des anthérozoïdes, par
conséquent la fécondation a encore besoin de la présence d’eau. Il existe de nos jours
15 000 espèces.
(Voir le cycle de développement des fougères mâle et femelle.)
Variantes évolutives :
Du cycle de développement chez les ptéridophytes.
Les spermatophytes :
- les gymnospermes (ginkgo, conifères, gnétophytes) : conifères
- Les angiospermes (paléodicots, monocots, eudicots) : plantes à fleurs
Généralité :
Au niveau de la reproduction on a deux innovations : le gamétophyte reste dans la tétraspore.
Dans ce cadre là le prothalle se développe dans la spore et son alimentation dépend du
sporophyte. Cet endoprothallie est à l’origine du grain de pollen et de l’ovule. Et enfin après
la fécondation il se transforme en graine.
La fécondation ne dépend plus de l’eau extérieur et la plante s’affranchie de l’eau extérieur
par l’apparition d’un processus interne avec une nouvelle structure qui est le tube pollinique.

B12 – Biologie végétale L1 Bio 2010/2011
Page 36 sur 39
Le grain de pollen est donc le gamétophyte mâle, il a une taille réduite et il est constitué
seulement de quelque cellule prothalienne et de deux anthérozoïdes. Les microsporanges sont
donc appelées sac pollinique et les microsporophyles sont les étamines. Au niveau de l’ovule
c’est l’addition du macrosporange et de la macrospore prothalisée. La cellule mère des spores
du macrosporange vont donner quatre macrospores après réduction chromatique. Trois vont
avortés, une va rester et cette quatrième va donner le prothalle femelle. Ce dernier va se
développer dans les tissus du macrosporange qu’on appelle nucelle. Les écailles du
macrosporange se soudent on obtient alors une enveloppe protectrice c’est le tégument avec le
micropyle. Le prothalle reste en parasite sur la plante mère et il accumule les réserves
nécessaires au développement de l’embryon (voir figure macrosporange des cycadales).
La graine est l’ensemble macrosporange macrospore prothalisée jeune embryon. Elle résulte
de la transformation de l’ovule après la fécondation. Les téguments vont se lignifier,
l’oosphère fécondée donne le zygote puis l’embryon.
Pollinisation et fécondation : les grains de pollen sont retenus sur l’ovule par des sécrétions
collantes et sucrées. Ce pollen germe au sommet du micropyle qui forme un tube pollinique
qui va s’enraciner et libérer les anthérozoïdes. Ces derniers vont gagner les oosphères à
travers le tissu et il y aura fécondation. Ce processus s’est affranchi de l’eau extérieur.
(Voir fécondation chez les cycadales figure 23) : il reste cependant un peu d’eau, un liquide,
qui permet aux anthérozoïdes d’accéder aux oosphères à l’intérieur de l’ovule, on s’affranchi
de l’eau externe mais pas interne. Légende : micropyle, grain de pollen, anthérozoïdes,
tégument, oosphères, prothalle femelle, nucelle liquéfié.
1. Embranchement gymnosperme
Généralité : on les appelle également les plantes à ovule nue (d’où le nom). Ces ovules sont
portées par une écaille qui est appelé écaille ovulaire ou séminale. Les grains de pollen quand
à eux sont transportés par le vent, passe par le micropyle et la germination se fait sur le
nucelle. On a un tissu de réserve qui est le prothalle femelle qui est appelée également
endosperme. Les gymnospermes sont toutes des plantes ligneuses avec une reproduction qui
s’étale sur plusieurs années et à titre d’information ils ont une longévité élevée. On connait
une espèce Pinus longaeva qui peut vivre jusqu’à 4000 ans.
On a trois sous embranchements :
a. Cycadophytes et Ginkgophytes
Ce sont des plantes relictuelles et la fécondation se fait comme les ptéridophytes en milieu
liquide mais sans eaux externe. On dit qu’ils sont ovipares soit que les ovules ont déjà l’aspect
de la graine avant la fécondation. Toutes les plantes Cycas à l’heure actuelle il en existe
environs 185 espèces différentes toutes des pays chauds, les ovules sont gros comme des œufs
de pigeons et dans ce groupe là les plantes classiques sont les fougères arborescentes. Dans
les Ginkgo ce sont des espèces fossiles sauf une qui vie actuellement qui est Ginkgo biloba.
Ils ont eu leur apogée au Permien de l’ère primaire et ils étaient très répandus au tertiaire.
b. Pinophytes ou Conifères ou Résineux
Ce sont tous les pins et les sapins, des plantes ligneuses en cône avec une racine profonde on
parle de racine en pivot. Les tissus sécrètent des oléorésines, les feuilles sont de petites tailles
en forme d’écailles pour les cyprès et en forme d’aiguilles pour les pins et les sapins. Plus
précisément pour les pins on a deux catégories de feuilles : sur les rameaux longs on a des
écailles sans chlorophylle et sur les rameaux courts des aiguilles coupées par deux à cinq
pièces.

B12 – Biologie végétale L1 Bio 2010/2011
Page 37 sur 39
Ils peuvent vivre plusieurs années et sont toujours verts même en hiver sauf le mélèze et le
cyprès chauve qui perdent leur aiguilles, ces feuilles sont adaptées à la sécheresse et au gèle et
sont capables d’assurer des échanges gazeux même à de très basse température.
Au niveau des organes sexuels ce sont toujours des cônes unisexués. Soit mâle soit femelle
mais portés par un même pied.
Etamines ou feuilles sporangifères avec deux sacs polliniques.
Voir cycle des gymnospermes.
c. Sous embranchement chlamydosperme
C’est un groupe charnière entre les gymnospermes et les angiospermes. On y trouve les
Ephesdra dans les régions méditerranéennes. Gnetum, la plus grosse ùajorité des lianes et
proto lianes. Welwitschia qui sont la plupart des espèces endémique, spécifique au Kalahari.
En bref dans la systématique on trouve sur l’ensemble de l’embranchement les Pins, les
Sapins, les Mélèzes, les Cèdres, les Douglas, Sequoias, Génévrier, Ifs.
2. Embranchements angiospermes
a. généralité
Groupe immense de 250 000 espèces réunis environs en 500 familles. La morphologie est très
variable, on a des formes herbacées, arbustives et arborescentes. Trois caractéristiques de
bases fondamentales :
Les écailles ovulifères vont entourer complètement les ovules.
Après fécondation ces carpelles se transforment en fruits.
Les organes reproducteurs se regroupent en fleurs bisexuées. Double fécondation.
b. Carpelles
On peut avoir des ovaires simples ou composés avec un seul ou plusieurs carpelles, libres ou
soudés et fermés ou ouverts.
c. fruits
Fruits indéhiscents : l’ovaire qui se transforme en fruit et cette ovaire se
lignifie. Dans cette catégorie il y a deux grands groupes :
i. Akènes : fruits secs et uni-séminés. Dispersion qui se fait soit par le
vent soit par les animaux. Donc possibilité d’y avoir des fruits avec des
ailes ou des crochets. Exemple : les noisettes.
ii. Drupes : la dispersion de ces fruits se fait par la gourmandise des
animaux. Il y a donc un processus de coévolution. Seulement la partie
interne de l’ovaire qui se lignifie, on va parler de noyaux. Exemple :
olive, cerise.
Fruits déhiscents : ils sont multi-séminés. Ces graines doivent s’échapper
rapidement du fruit pour suivre le cycle. Elles ont une paroi épaisse. Et on a
deux grosses catégories :
i. Capsules, Folliculus, Gousses, Siliques.
ii. Baies : exemple : raisin.

B12 – Biologie végétale L1 Bio 2010/2011
Page 38 sur 39
d. Fleurs
Elles se regroupent en inflorescences qui sont de type grappes ou cymes. Pollinisation par le
vent ou par les insectes. Colonisation antofile par les insectes.
e. Autres caractéristiques des angiospermes.
Miniaturisation extrême des gamétophytes. Le prothalle mâle est réduit à deux cellules :
végétatives et reproductives. C’est le grain de pollen. Le prothalle femelle composé de 7sept
cellules c’est le sac embryonnaire. Appareil végétatif très perfectionné en particulier tout
système de vaisseaux rayé, réticulé, ponctué, etc. et l’accroissement en largeur qui se fait par
des formations secondaires.
Une diversification extrême, plante rugueuse, captiforme, trompe, certain de plus de 100m de
hauteur, forme herbacés, forme de lianes, flottant, etc.
f. Cycle de développement
Il y a des apports fondamentaux des angiospermes à l’évolution qui sont :
La fleur.
L’ovaire et le fruit.
L’albumen qui est un tissu de réserve polyploïde.
g. Classification
On a trois classes :
Classe des Paléodicots, qui sont Monoaperturés et Dicotylés.
Généralité : groupe archaïque, plus ou moins 9000 espèces en général présent
dans les régions chaudes. Les feuilles sont généralement simples et sans stipules.
Les fleurs sont souvent de grandes tailles. Exemple le Magnolia et les Nénuphars.
Les pièces florales sont disposées en hélices donc il n’y a pas de différenciation
nette entre sépales et pétales. Il n’y a donc pas de verticilles. Pour les plus évolués
on voit apparaitre les premiers verticilles et dans ce cas là on va avoir une
cyclisation de la fleur sur un mode trimère. Exemple : 3S + 3S + 3E + 3E + 3E +
3E + 1C. Exemple : Canneliers, Lauriers rose, Poivrier.
Classe des Monocots, qui sont Monoaperturés et Monocotylés.
Un seul cotylédon, les feuilles sont toujours simples, elles sont réduites au
pétiole et il n’y a pas de formation secondaire. Il y a environs 55 000 espèces pour
une centaines de familles. La tige étant toujours en position latérale. La racine
principale avorte très rapidement et est remplacée par des racine infantile. Des
tiges sont toujours feuillée et de type herbacés. Il n’y a pas de port type
arborescent. Il n’y a pas de « vrai » tronc avec de « vrai » branche. Fleurs toujours
simple et énervation parallèle. Fleurs cinq cycles : 3S + 3S + 3E + 3E + 3C mais
les plus évolué comme la tulipe, le lys et le muguet on a des sépales identiques aux
pétales qu’on appelle des tépales (3+3)T + (3+3)E + 3C.
Classe des Eudicots, qui sont Triaperturés et Dicotylés.
La racine principale n’avorte pas au contraire elle s’allonge en racine principale
avec des racines secondaires. La tige est ramifiée et pour les plus évoluée avec le

B12 – Biologie végétale L1 Bio 2010/2011
Page 39 sur 39
méristème secondaire ça va donner de véritable trompe. La feuille est composée
d’un limbe à nervation pennée ou palmée et le limbe peut se diviser en folioles. Au
niveau de l’appareil reproducteur on a une vraie pentamérisation de la fleur : 5 !
Apparition des vrais pétales avec odeurs, couleurs, parfums etc. en général la
première forme florale est à 5cycles : 5S + 5P + (5+5)E + 5C. En revanche pour
les plus évolué de ces fleurs on a des formes 4 cycles : 5S + 5P + 5E + 5C.