Embed Size (px)
Citation preview

2ème partie : Neurobiologie moléculaire
Chapitre 2 – LE NEURONE
L’idée maîtresse des neurosciences modernes est que tous les comportements sont le reflet d’une fonction cérébrale. Dans ce contexte, la pensée correspond elle aussi à un type d’activité cérébrale. Cette activité ne s’exprime pas seulement à travers des comportements simples comme la marche ou le sourire mais aussi à travers des fonctions beaucoup plus complexes comme l’élaboration de sentiments, l’apprentissage, les comportement fondamentaux (alimentaire, dipsique et sexuel) ou encore l’écriture… d’un poème par exemple. En corollaire, des désordres affectifs (des émotions) ou cognitifs (de la pensée) sont le reflet de maladies névrotiques et psychotiques qui reflètent des troubles fonctionnels cérébraux..
Le cerveau est fait d’unités morphologiques et fonctionnelles, les cellules nerveuses (ou neurones) et les cellules gliales. Le propos des neurosciences est d’expliquer comment le cerveau peut disposer de ces unités pour contrôler le comportement et comment en retour ces unités en fonctionnant dans le cerveau d’un individu peuvent être influencées par le fonctionnement d’autres populations cellulaires ou encore la multitude des autres facteurs environnementaux. La neurobiologie nous permet de comprendre comment les neurones interagissent pour produire le comportement complexe de l’organisme.
Le neurobiologiste se doit donc d'expliquer: • comment chaque neurone pris individuellement acquiert ses propriétés fonctionnelles au cours du
développement; • comment chaque neurone s'intègre dans un réseau neuronal et participe à la fonction de ce réseau.
I - LE NEURONE EST L'UNITE STRUCTURALE DU SYSTEME NERVEUX
1 - MORPHOLOGIE Selon le concept proposé en 1888 par Ramon Y Cajal, chaque neurone est une cellule anatomiquement séparée et
indépendante des autres cellules n’établissant que des contacts fonctionnels spécifiques, appelés synapses: soit par l’intermédiaire de leurs prolongements dendritiques :
avec d’autres neurones : synapses neuro-neuronales ; ou des cellules sensorielles : synapses sensori-neurales;
soit par l’intermédiaire de leurs terminaisons axonales synapses axo-somatiques, axo-dendritiques, (ou encore sensori-neurales), avec
d’autres neurones (synapses neuro-neuronales); jonctions neuro-musculaires, (avec les fibres musculaires squelettiques,
cardiaques ou lisses), synapses neuro-glandulaires et synapses neuro-endocrines, (avec les cellules glandulaires).
Ainsi entre une cellule réceptrice et une cellule effectrice, plusieurs neurones forment un réseau qui transmettent -
et traitent en les transmettant- les informations dont ils assurent la propagation . Tous les neurones ont un corps cellulaire (ou soma) d’où partent deux types de prolongements, les dendrites ou
arbre dendritique d’une part, l’axone et ses prolongements (collatérales et arborisation terminale) d’autre part. Chacun d’entre eux reçoit de multiples afférences (voir la figure 1 du chapitre précédent).
Le soma est la partie du neurone qui contient le noyau cellulaire et le cytoplasme qui l’entoure (ou périkaryon) ; sa
forme est très variable (fig. 1, fig. 2): • soma pyramidaux des cellules pyramidales du cortex cérébral ou de l'hippocampe; • soma ovoïdes des cellules de Purkinje du cortex cérébelleux; • soma granulaires (en forme de grains des petites cellules multipolaires du cortex
cérébral, du cortex cérébelleux ou de l'hippocampe; • soma fusiformes du pallidum; • soma étoilés ou multipolaires des motoneurones de la moelle épinière).
Un des rôles du soma est d’assurer la synthèse d’une très grande partie des constituants nécessaires à la structure et aux fonctions du neurone. En effet c’est dans le soma que se trouve l’ensemble des organites responsables de la synthèse des macromolécules. La majorité des neurones ne se divisant plus après la naissance et étant incapables de régénérer, le corps cellulaire doit assurer l’intégrité structurale de tout le neurone pendant toute la vie de l’individu.
Les dendrites, lorsqu’elles émergent du soma, sont de simples extensions du périkaryon, de contour irrégulier et
dont le diamètre s’amenuise au fur et à mesure des branchements. Le nombre des branchements est caractéristique du type de neurone. Les dendrites ont généralement des contours irréguliers (présence "d'épines dendritiques") et par leur diamètre qui s'amenuise au fur et à mesure des branchements.
L’axone se différencie des dendrites par son aspect lisse, son diamètre uniforme le long de son trajet et son ultrastructure. Il prend généralement son origine au niveau d’une expansion conique du soma, le cône d’émergence. Après le cône d'émergence on distingue le segment initial de diamètre plus petit, puis l'axone proprement dit. Il peut se diviser en une ou plusieurs collatérales. L’axone et les collatérales peuvent être recouverts d’une gaine de myéline. La
148
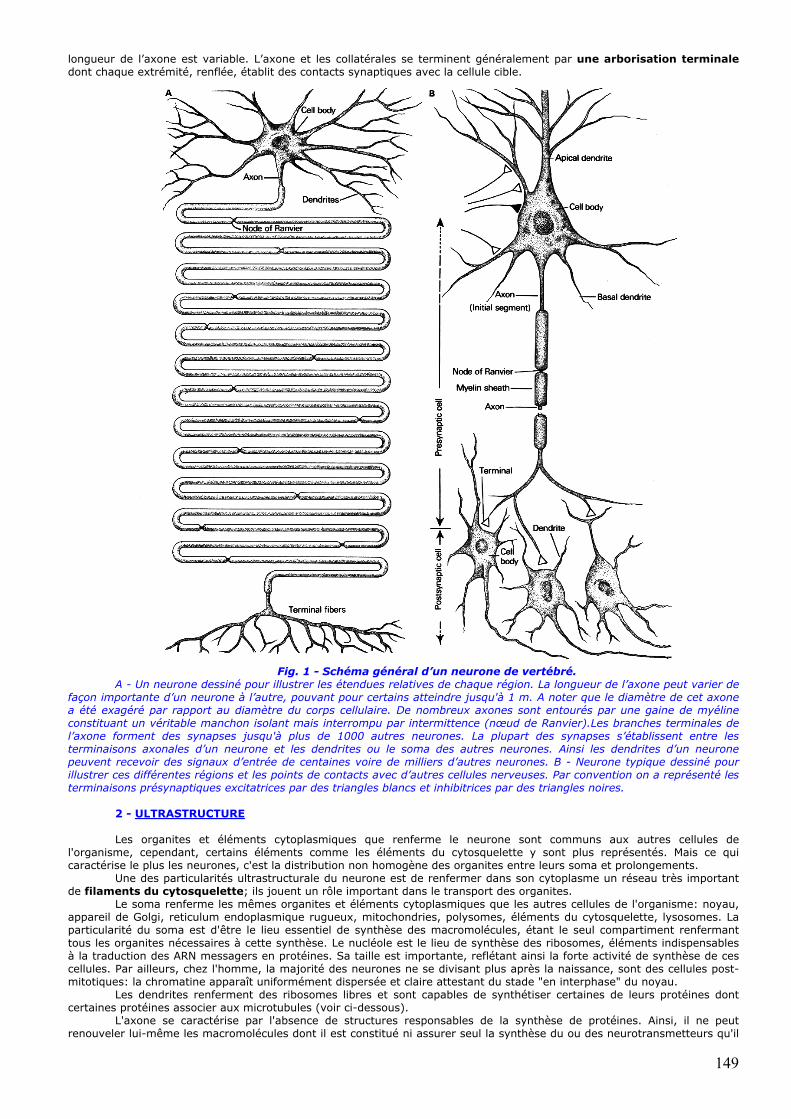
longueur de l’axone est variable. L’axone et les collatérales se terminent généralement par une arborisation terminale dont chaque extrémité, renflée, établit des contacts synaptiques avec la cellule cible.
Fig. 1 - Schéma général d’un neurone de vertébré. A - Un neurone dessiné pour illustrer les étendues relatives de chaque région. La longueur de l’axone peut varier de
façon importante d’un neurone à l’autre, pouvant pour certains atteindre jusqu'à 1 m. A noter que le diamètre de cet axone a été exagéré par rapport au diamètre du corps cellulaire. De nombreux axones sont entourés par une gaine de myéline constituant un véritable manchon isolant mais interrompu par intermittence (nœud de Ranvier).Les branches terminales de l’axone forment des synapses jusqu'à plus de 1000 autres neurones. La plupart des synapses s’établissent entre les terminaisons axonales d’un neurone et les dendrites ou le soma des autres neurones. Ainsi les dendrites d’un neurone peuvent recevoir des signaux d’entrée de centaines voire de milliers d’autres neurones. B - Neurone typique dessiné pour illustrer ces différentes régions et les points de contacts avec d’autres cellules nerveuses. Par convention on a représenté les terminaisons présynaptiques excitatrices par des triangles blancs et inhibitrices par des triangles noires.
2 - ULTRASTRUCTURE Les organites et éléments cytoplasmiques que renferme le neurone sont communs aux autres cellules de
l'organisme, cependant, certains éléments comme les éléments du cytosquelette y sont plus représentés. Mais ce qui caractérise le plus les neurones, c'est la distribution non homogène des organites entre leurs soma et prolongements.
Une des particularités ultrastructurale du neurone est de renfermer dans son cytoplasme un réseau très important de filaments du cytosquelette; ils jouent un rôle important dans le transport des organites.
Le soma renferme les mêmes organites et éléments cytoplasmiques que les autres cellules de l'organisme: noyau, appareil de Golgi, reticulum endoplasmique rugueux, mitochondries, polysomes, éléments du cytosquelette, lysosomes. La particularité du soma est d'être le lieu essentiel de synthèse des macromolécules, étant le seul compartiment renfermant tous les organites nécessaires à cette synthèse. Le nucléole est le lieu de synthèse des ribosomes, éléments indispensables à la traduction des ARN messagers en protéines. Sa taille est importante, reflétant ainsi la forte activité de synthèse de ces cellules. Par ailleurs, chez l'homme, la majorité des neurones ne se divisant plus après la naissance, sont des cellules post-mitotiques: la chromatine apparaît uniformément dispersée et claire attestant du stade "en interphase" du noyau.
Les dendrites renferment des ribosomes libres et sont capables de synthétiser certaines de leurs protéines dont certaines protéines associer aux microtubules (voir ci-dessous).
L'axone se caractérise par l'absence de structures responsables de la synthèse de protéines. Ainsi, il ne peut renouveler lui-même les macromolécules dont il est constitué ni assurer seul la synthèse du ou des neurotransmetteurs qu'il
149

libère du fait de son incapacité à synthétiser les protéines. Ce problème est résolu du fait de l'existence d'un apport continuel de macromolécules du soma vers l'axone jusqu'aux terminaisons par transport axonal antérograde.
Fig. 2 - Les neurones peuvent être classés en neurones unipolaires, bipolaires ou multipolaires selon le
nombre de prolongements de leur corps cellulaire.
3 - LES TRANSPORTS AXONAUX (fig. 3) Ils permettent une communication à double sens entre le corps cellulaire et les terminaisons axonales. Des
microtubules particulièrement abondants dans les neurones jouent un rôle primordial dans ces transports. Un transport antérograde rapide permet, entre autres, le renouvellement des protéines membranaires de
l'axone. Il consiste en un déplacement de vésicules contenant les macromolécules le long des microtubules de l'axone à une vitesse de 100 à 400 mm/j. Ces vésicules sont formées dans le corps cellulaire à partir de l'appareil de Golgi. Elles apportent certaines des protéines nécessaires au renouvellement de la membrane plasmique et des membranes internes de l'axone, les enzymes de synthèse du ou des neurotransmetteurs et le précurseur du neurotransmetteur lorsque celui-ci est un peptide.
Un transport antérograde lent (0,1 à 2mm/j) permet le renouvellement du cytosquelette de l'axone
(microtubules, neurofilaments, microfilaments) ainsi que des protéines cytosolubles (enzymes du métabolisme intermédiaire, glycolyse). Dans les axones en élongation (en développement ou en régénération) ce transport lent apporte l'axoplasme nécessaire à la croissance de l'axone.
Un transport axonal des mitochondries permet le renouvellement des mitochondries de l'axone et des
terminaisons. Les mitochondries nouvellement formées dans le soma sont transportées dans l'axone jusqu'aux terminaisons. Celles qui présentent des signes de dégénérescence sont soumises à un transport rétrograde vers le soma. On ne connaît pas le mécanisme de ce transport.
Un transport axonal rétrograde permet l'élimination des déchets: il permet le retour des molécules membranaires vers le corps cellulaire afin qu'elles y soient dégradées, voire recyclées.
150
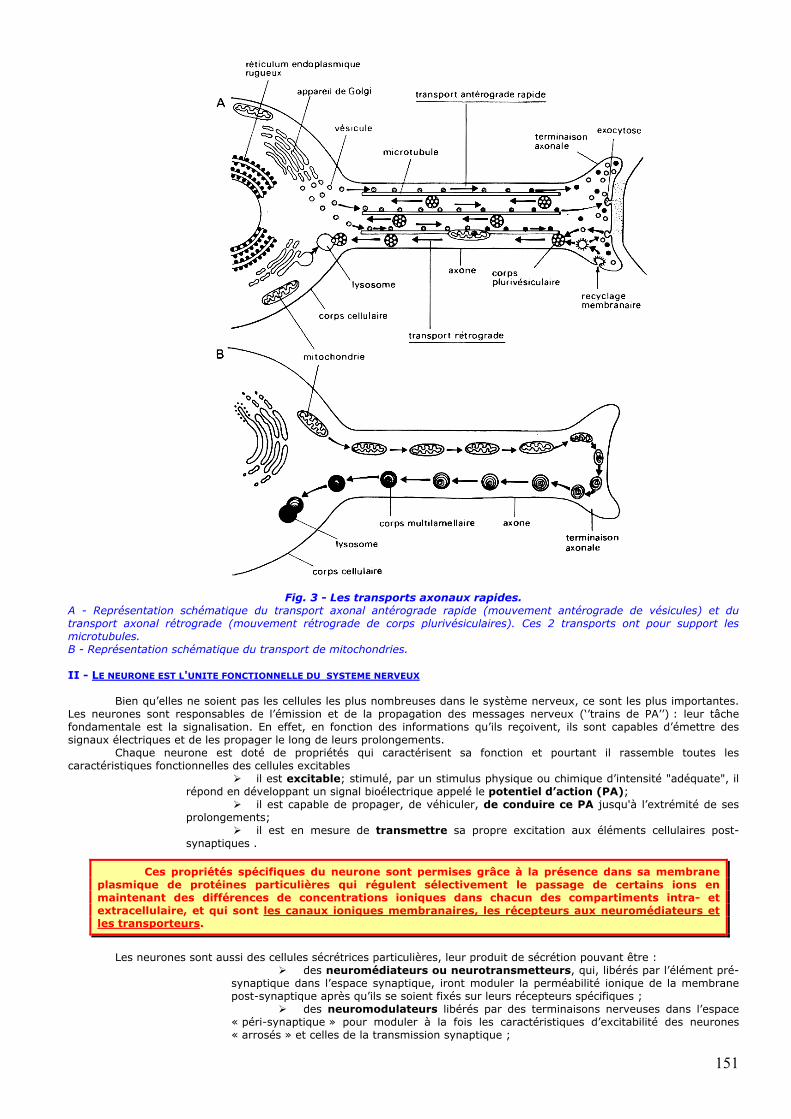
Fig. 3 - Les transports axonaux rapides. A - Représentation schématique du transport axonal antérograde rapide (mouvement antérograde de vésicules) et du transport axonal rétrograde (mouvement rétrograde de corps plurivésiculaires). Ces 2 transports ont pour support les microtubules. B - Représentation schématique du transport de mitochondries. II - LE NEURONE EST L'UNITE FONCTIONNELLE DU SYSTEME NERVEUX
Bien qu’elles ne soient pas les cellules les plus nombreuses dans le système nerveux, ce sont les plus importantes.
Les neurones sont responsables de l’émission et de la propagation des messages nerveux (‘’trains de PA’’) : leur tâche fondamentale est la signalisation. En effet, en fonction des informations qu’ils reçoivent, ils sont capables d’émettre des signaux électriques et de les propager le long de leurs prolongements.
Chaque neurone est doté de propriétés qui caractérisent sa fonction et pourtant il rassemble toutes les caractéristiques fonctionnelles des cellules excitables
il est excitable; stimulé, par un stimulus physique ou chimique d’intensité "adéquate", il répond en développant un signal bioélectrique appelé le potentiel d’action (PA);
il est capable de propager, de véhiculer, de conduire ce PA jusqu'à l’extrémité de ses prolongements;
il est en mesure de transmettre sa propre excitation aux éléments cellulaires post-synaptiques .
Ces propriétés spécifiques du neurone sont permises grâce à la présence dans sa membrane plasmique de protéines particulières qui régulent sélectivement le passage de certains ions en maintenant des différences de concentrations ioniques dans chacun des compartiments intra- et extracellulaire, et qui sont les canaux ioniques membranaires, les récepteurs aux neuromédiateurs et les transporteurs.
Les neurones sont aussi des cellules sécrétrices particulières, leur produit de sécrétion pouvant être : des neuromédiateurs ou neurotransmetteurs, qui, libérés par l’élément pré-
synaptique dans l’espace synaptique, iront moduler la perméabilité ionique de la membrane post-synaptique après qu’ils se soient fixés sur leurs récepteurs spécifiques ;
des neuromodulateurs libérés par des terminaisons nerveuses dans l’espace « péri-synaptique » pour moduler à la fois les caractéristiques d’excitabilité des neurones « arrosés » et celles de la transmission synaptique ;
151

des neurohormones libérés par les terminaisons de certains neurones de l’hypothalamus en réponse à leur excitation.
Leur morphologie est modelée par leurs fonctions spécifiques de réception, d’intégration et de transmission de
l’information. 1 - L’ARBRE SOMATO-DENDRITIQUE REPRESENTE LE POLE RECEPTEUR DU NEURONE. Les dendrites et le soma reçoivent de très nombreux contacts synaptiques venant d’autres neurones et constituent
de ce fait la principale surface de réception du neurone. Ce n’est pas là leur unique rôle : elles génèrent en réponse à ces messages, des signaux électriques comme les potentiels post-synaptiques ou comme les potentiels d’action calciques et intègrent ces messages afférents.
Remarque : si les dendrites sont généralement les principales zones de réception, il existe des exceptions, ainsi certaines dendrites sont connectées à d’autres dendrites et se comportent comme des régions émettrices libérant un neurotransmetteur.
2 - L’AXONE ET LES COLLATERALES AXONALES REPRESENTENT LE POLE EMETTEUR DU NEURONE.
La principale caractéristique des axones est leur capacité à conduire des potentiels d’action sur de longues
distances et ceci sans perte d’amplitude. Il est classiquement admis que c’est au niveau du segment initial que sont générés les potentiels d’action en réponse aux informations synaptiques transmises par l’arbre somato-dendritique. Le potentiel d’action se propage ensuite le long de l’axone et de ses collatérales jusqu’aux terminaisons axonales. Les terminaisons axonales, en réponse à l’arrivée d’un potentiel d’action libère le ou les neurotransmetteurs qu’elles renferment. Cette libération est localisée précisément au niveau des contacts synaptiques.
3 - SIGNALISATION NEURONALE Les neurones codent l’information par une combinaison de signaux électriques et chimiques : • Signalisation chimique Les neurones sécrètent des neurotransmetteurs et d’autres messagers chimiques à des sites de contact fonctionnel
appelés synapses chimiques. Au niveau de ces synapses, une région membranaire de la cellule présynaptique est attachée fermement et intimement à une région membranaire spécialisée de la cellule postsynaptique. Cette région contient une forte concentration de récepteurs pour le neurotransmetteur. C’est la raison pour laquelle le sens de la communication chimique de l’information d’un neurone à un autre est dirigé très spécifiquement -on dit « polarisé »- du neurone présynaptique vers le neurone postsynaptique.
Le nombre de contacts synaptiques réalisés par un même neurone peut être extraordinairement grand. Un seul motoneurone de la moelle épinière de mammifère par exemple reçoit des messages transmis par approximativement 10 000 synapses. Pour recevoir autant de contacts synaptiques, les neurones se dotent généralement de prolongements cellulaires très ramifiés spécialisés dans la réception de synapses provenant d’autres cellules et formant des arborisations complexes de dendrites. Les dendrites possèdent les récepteurs et les autres protéines nécessaires à la conversion du signal chimique en signal électrique.
• Signalisation électrique. La liaison du neurotransmetteur avec les récepteurs membranaires provoquent l’apparition d’un signal électrique
dans la membrane postsynaptique que l’on appelle le potentiel synaptique. Par rapport à un niveau de potentiel de membrane de référence et stable (potentiel de repos), le sens de la variation du potentiel synaptique peut, selon la nature de l’interaction entre le neurotransmetteur et son récepteur, se traduire par une dépolarisation ou une hyperpolarisation. Tandis qu’un potentiel synaptique hyperpolarisant le potentiel de membrane diminue l’excitabilité du neurone postsynaptique, un potentiel synaptique dépolarisant l’augmente. Ainsi, pour un neurone donné, l’ensemble de tous les potentiels synaptiques hyper- et dépolarisant qui naissent au niveau de l’arborisation dendritique sont combinés ou intégrés par sommation algébrique au niveau de la membrane du corps cellulaire du neurone. Si cette intégration se solde par une dépolarisation de la membrane somatique, elle y est traduite en d’autres signaux électriques (les potentiels d’action), reflet de l’état d’excitation du neurone. Ces signaux électriques sont immédiatement émis dans le prolongement principal du neurone appelé axone où ils se propagent à une vitesse constante comprise entre 0,5 et 40 m/s selon le type de neurone. Dans le cas contraire, aucun potentiel d’action n’est émis et la transmission de l’information est interrompue.
III - LE NEURONE EST UNE CELLULE EXCITABLE ET SECRETRICE QUI PRESENTE UNE EXTREME REGIONALISATION DE SES FONCTIONS
1 - REGIONALISATION DES FONCTIONS METABOLIQUES L'essentiel de l'activité de synthèse du neurone est localisé dans le corps cellulaire, les dendrites ne synthétisant
que quelques-unes de ses protéines et l'axone étant incapable de synthétiser ses propres protéines. Dans cette cellule où le volume de l'axone peut représenter jusqu'à 1000 fois celui du corps cellulaire, on conçoit aisément que le maintien de la structure et du fonctionnement de l'axone et de ses terminaisons nécessite un apport important et permanent de macromolécules. Cet apport est assuré par les transports axonaux antérogrades, transports qui s'effectuent du corps cellulaire vers les terminaisons axonales. En ce qui concerne les dendrites, il a été mis en évidence un transport d'ARN du corps cellulaire vers les polysomes, ceci pouvant leur permettre de synthétiser certaines de leurs protéines mais un transport de macromolécules du corps cellulaire vers les dendrites équivalent au transport axonal, n'a pu être mis en évidence jusqu'à présent.
La dégradation d'éléments non neuronaux venant du milieu extérieur et captés par les terminaisons axonales par un processus d'endocytose (virus, certaines toxines comme la toxine tétanique ou la toxine du choléra par exemple), ainsi que les déchets du métabolisme cellulaire, a lieu dans le corps cellulaire, au niveau des lysosomes. A noter que certaines de ces toxines avant d'être dégradées ont le temps d'exercer leur toxicité sur le corps cellulaire. Néanmoins, Il est nécessaire pour le neurone d'acheminer la plupart de ces constituants des terminaisons axonales vers le corps cellulaire. Ceci est réalisé par le transport axonal rétrograde. Enfin pour qu'il y ait une coordination entre l'activité de synthèse au niveau du
152
corps cellulaire et les besoins des terminaisons axonales, l'existence d'un système d'information rétrograde (des terminaisons vers le corps cellulaire) paraît nécessaire. Ceci aurait lieu via le transport axonal rétrograde.
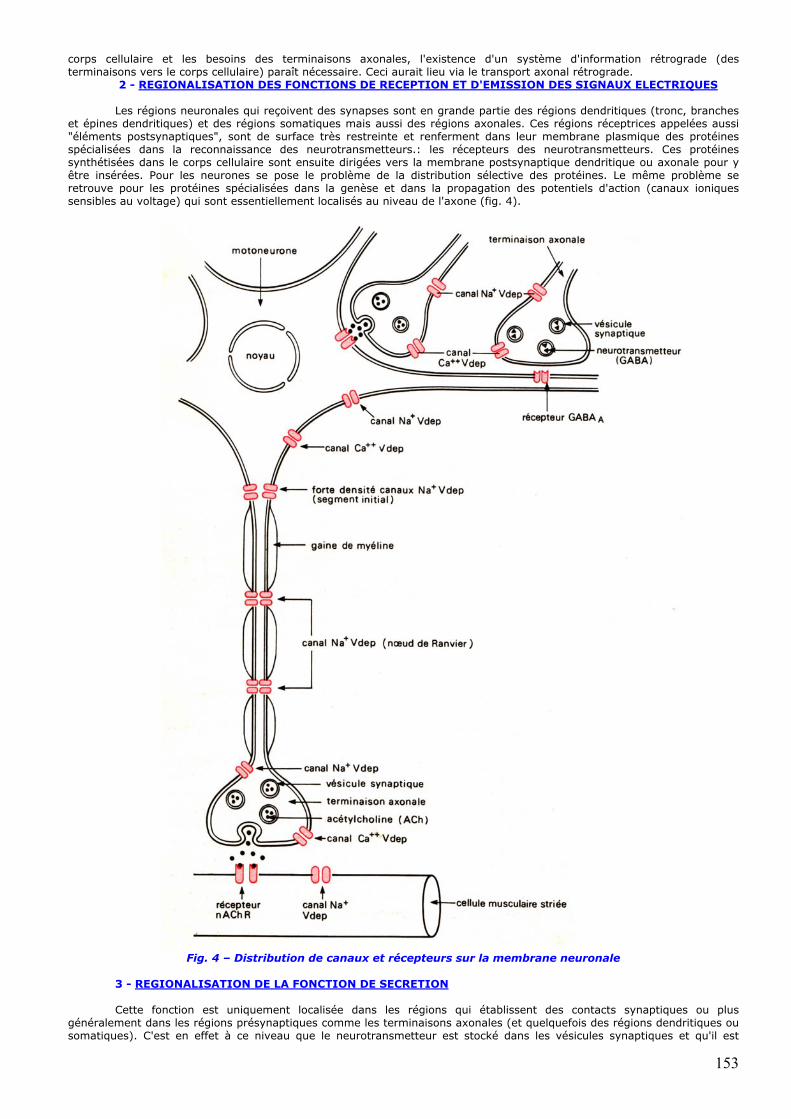
2 - REGIONALISATION DES FONCTIONS DE RECEPTION ET D'EMISSION DES SIGNAUX ELECTRIQUES Les régions neuronales qui reçoivent des synapses sont en grande partie des régions dendritiques (tronc, branches
et épines dendritiques) et des régions somatiques mais aussi des régions axonales. Ces régions réceptrices appelées aussi "éléments postsynaptiques", sont de surface très restreinte et renferment dans leur membrane plasmique des protéines spécialisées dans la reconnaissance des neurotransmetteurs.: les récepteurs des neurotransmetteurs. Ces protéines synthétisées dans le corps cellulaire sont ensuite dirigées vers la membrane postsynaptique dendritique ou axonale pour y être insérées. Pour les neurones se pose le problème de la distribution sélective des protéines. Le même problème se retrouve pour les protéines spécialisées dans la genèse et dans la propagation des potentiels d'action (canaux ioniques sensibles au voltage) qui sont essentiellement localisés au niveau de l'axone (fig. 4).
Fig. 4 – Distribution de canaux et récepteurs sur la membrane neuronale
3 - REGIONALISATION DE LA FONCTION DE SECRETION Cette fonction est uniquement localisée dans les régions qui établissent des contacts synaptiques ou plus
généralement dans les régions présynaptiques comme les terminaisons axonales (et quelquefois des régions dendritiques ou somatiques). C'est en effet à ce niveau que le neurotransmetteur est stocké dans les vésicules synaptiques et qu'il est
153

libéré. La fonction de sécrétion implique la présence dans la région présynaptique de molécules et organites particuliers: enzyme de synthèse du neurotransmetteur, vésicules synaptiques, microtubules et protéines associées etc...Là encore, la distribution sélective des organites et des protéines a une grande importance. IV - LES MECANISMES DE SIGNALISATION SONT COMMUNS A TOUS LES NEURONES
La plupart des neurones indépendamment de leur taille, leur forme ou encore la biochimie de leurs
neurotransmetteurs et de leurs fonctions comportementales peuvent être décrits en termes de «modèle généralisé de neurone» (fig. 5).
Fig. 5 - Les neurones sensitifs (ou sensoriels ) et moteurs, les interneurones et les neurones endocrines ont en commun 4 composantes fonctionnelles : une composante d’entrée, une composante d’intégration, une composante de conduction et une composante de sortie. Sur la base de ces observations, un modèle de neurone résumant cette organisation fonctionnelle peut être construit.
Si la plupart des neurones sont construits selon ce modèle, de petits détails peuvent les différencier les uns des autres. Ces différences peuvent avoir des répercussions lorsqu’elles sont affectées, par exemple, déclenchant alors des maladies : la sclérose amyotrophique latérale et la polyomyélite affecte les motoneurones, le tabes dorsalis affecte les neurones sensoriels, le diabète insipide touche seulement certains neurones endocrines. Néanmoins, les propriétés de signalisation électrique sont pour tous les neurones remarquablement similaires et globalement stéréotypées. Par exemple, le PA d’un motoneurone ressemble et est du aux mêmes causes que le PA d’un neurone sensoriel. Par ailleurs, la fréquence des PA d’un interneurone ou d’un neurone sensoriel (fig. 6) constituant un message nerveux dépend du niveau de dépolarisation de leur composante d’intégration (potentiel récepteur de la figure 6).
154

Fig. 6 – Message sensoriel Pour conclure, il est bon de répéter que le neurobiologiste se doit donc d’expliquer: comment chaque neurone se développe depuis l’œuf jusque chez l’adulte pour atteindre sa complète maturité
et comment différents neurones se mettent en place dans les réseaux; c'est l'objet de la neuroanatomie; comment ils fonctionnent individuellement ou en réseau à chacune des étapes de leur développement et
ce jusqu'à l'âge adulte et comment ils peuvent être impliqués dans tel ou tel comportement normal et/ou pathologique (fig. 7); c'est l'objet de la neurophysiologie et de la neurobiologie cellulaires et moléculaires.
155
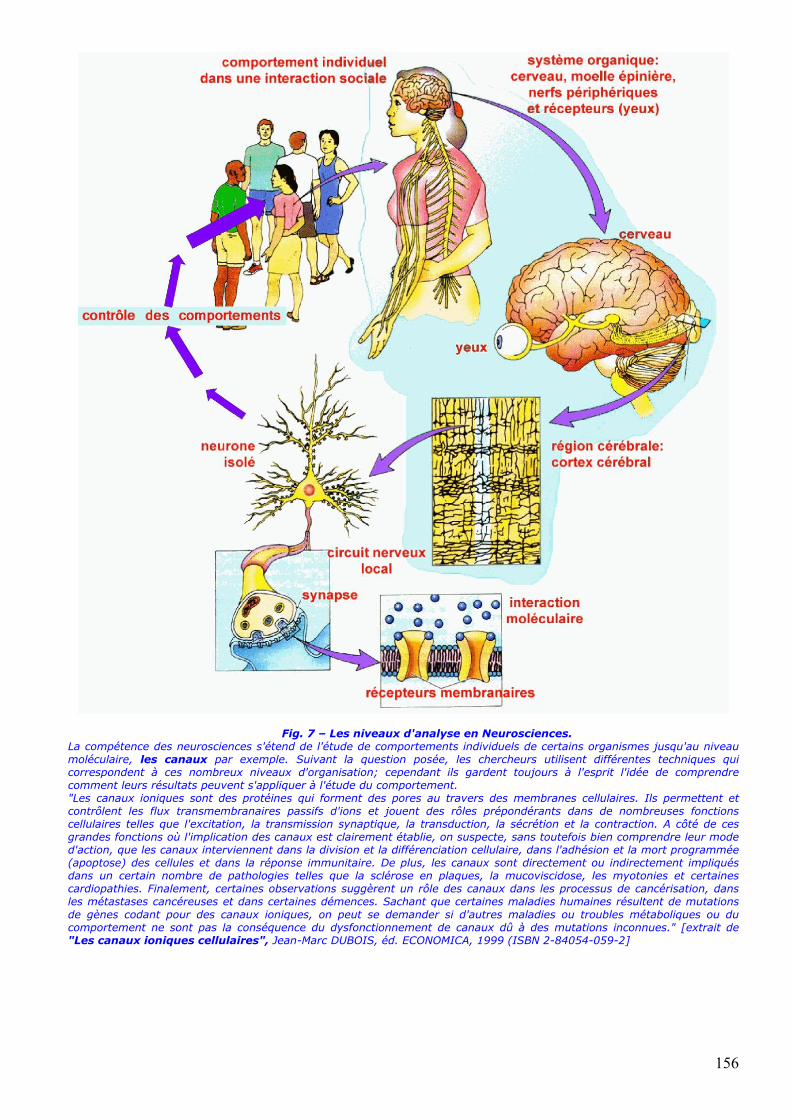
Fig. 7 – Les niveaux d'analyse en Neurosciences. La compétence des neurosciences s'étend de l'étude de comportements individuels de certains organismes jusqu'au niveau moléculaire, les canaux par exemple. Suivant la question posée, les chercheurs utilisent différentes techniques qui correspondent à ces nombreux niveaux d'organisation; cependant ils gardent toujours à l'esprit l'idée de comprendre comment leurs résultats peuvent s'appliquer à l'étude du comportement. "Les canaux ioniques sont des protéines qui forment des pores au travers des membranes cellulaires. Ils permettent et contrôlent les flux transmembranaires passifs d'ions et jouent des rôles prépondérants dans de nombreuses fonctions cellulaires telles que l'excitation, la transmission synaptique, la transduction, la sécrétion et la contraction. A côté de ces grandes fonctions où l'implication des canaux est clairement établie, on suspecte, sans toutefois bien comprendre leur mode d'action, que les canaux interviennent dans la division et la différenciation cellulaire, dans l'adhésion et la mort programmée (apoptose) des cellules et dans la réponse immunitaire. De plus, les canaux sont directement ou indirectement impliqués dans un certain nombre de pathologies telles que la sclérose en plaques, la mucoviscidose, les myotonies et certaines cardiopathies. Finalement, certaines observations suggèrent un rôle des canaux dans les processus de cancérisation, dans les métastases cancéreuses et dans certaines démences. Sachant que certaines maladies humaines résultent de mutations de gènes codant pour des canaux ioniques, on peut se demander si d'autres maladies ou troubles métaboliques ou du comportement ne sont pas la conséquence du dysfonctionnement de canaux dû à des mutations inconnues." [extrait de "Les canaux ioniques cellulaires", Jean-Marc DUBOIS, éd. ECONOMICA, 1999 (ISBN 2-84054-059-2]
156